







































































































































































































































































































































































































































































































































































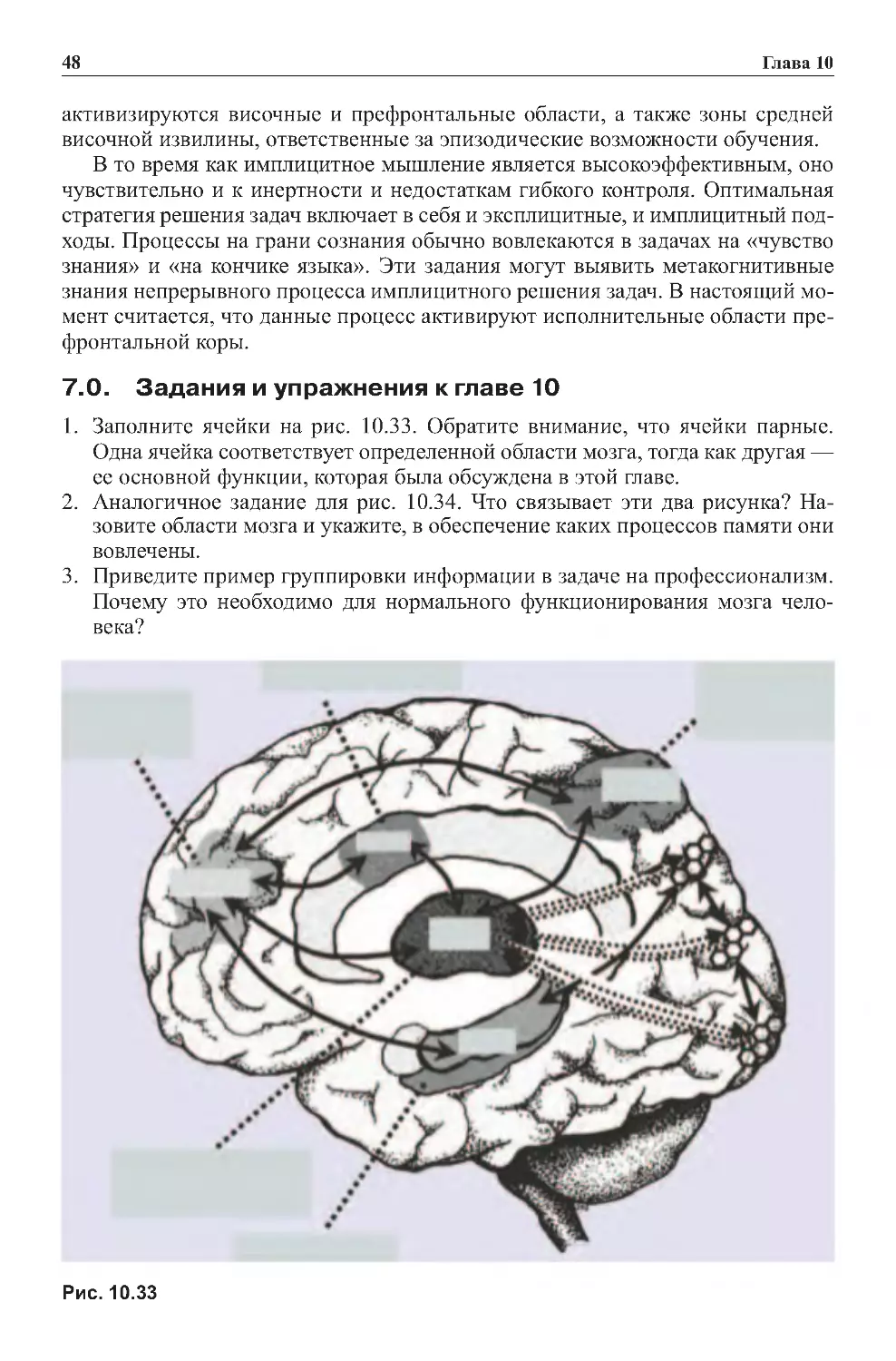













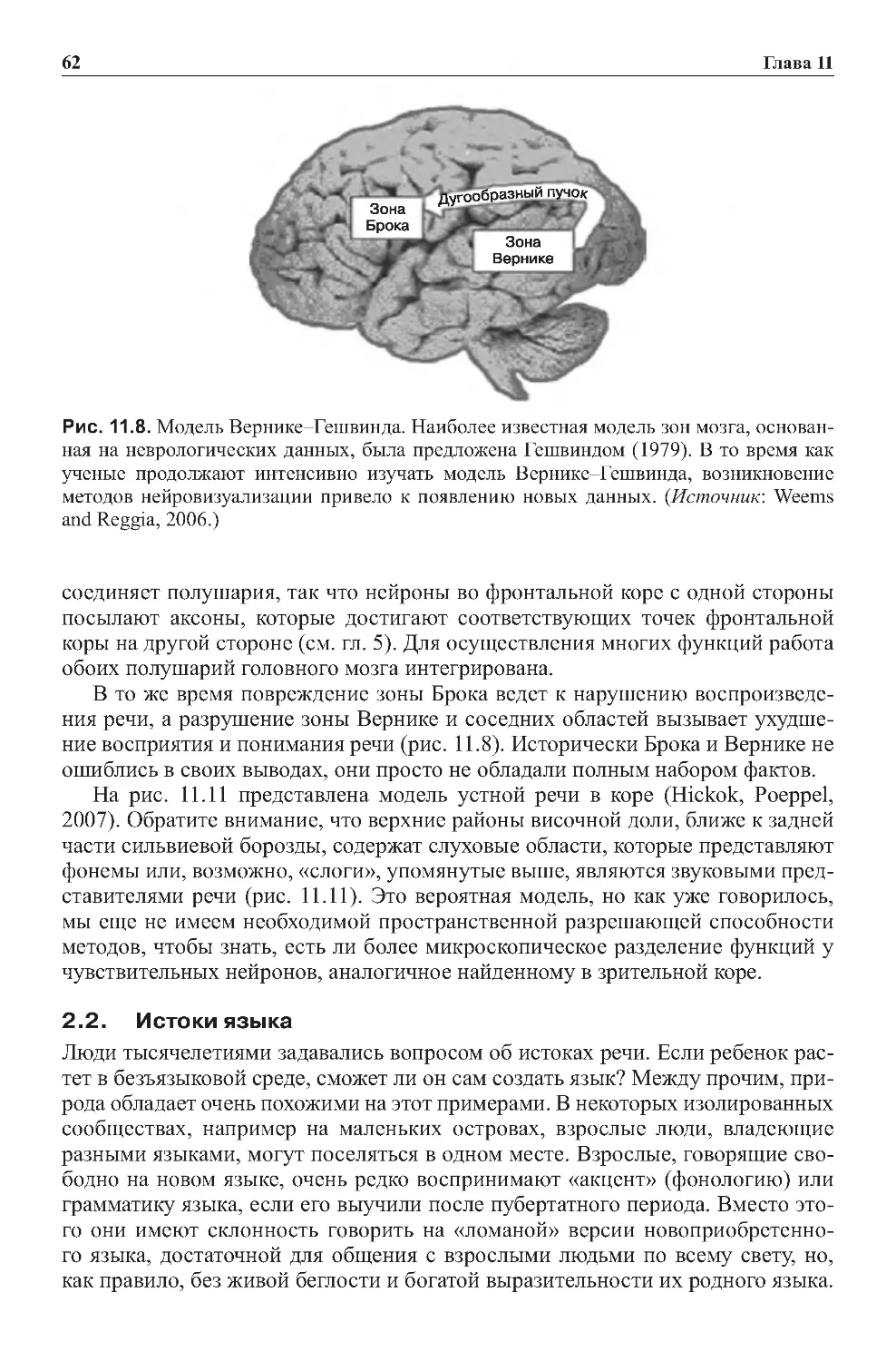








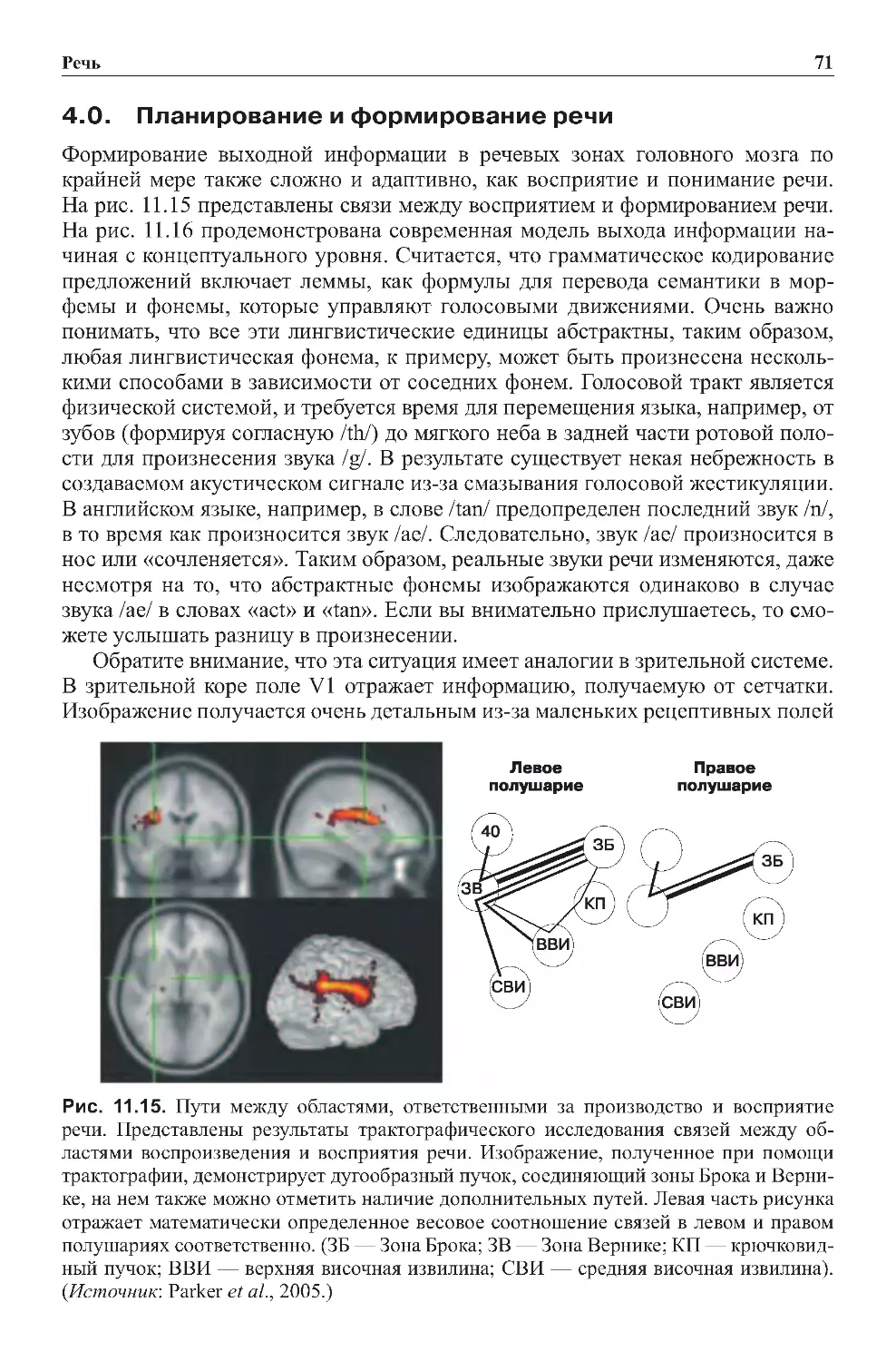




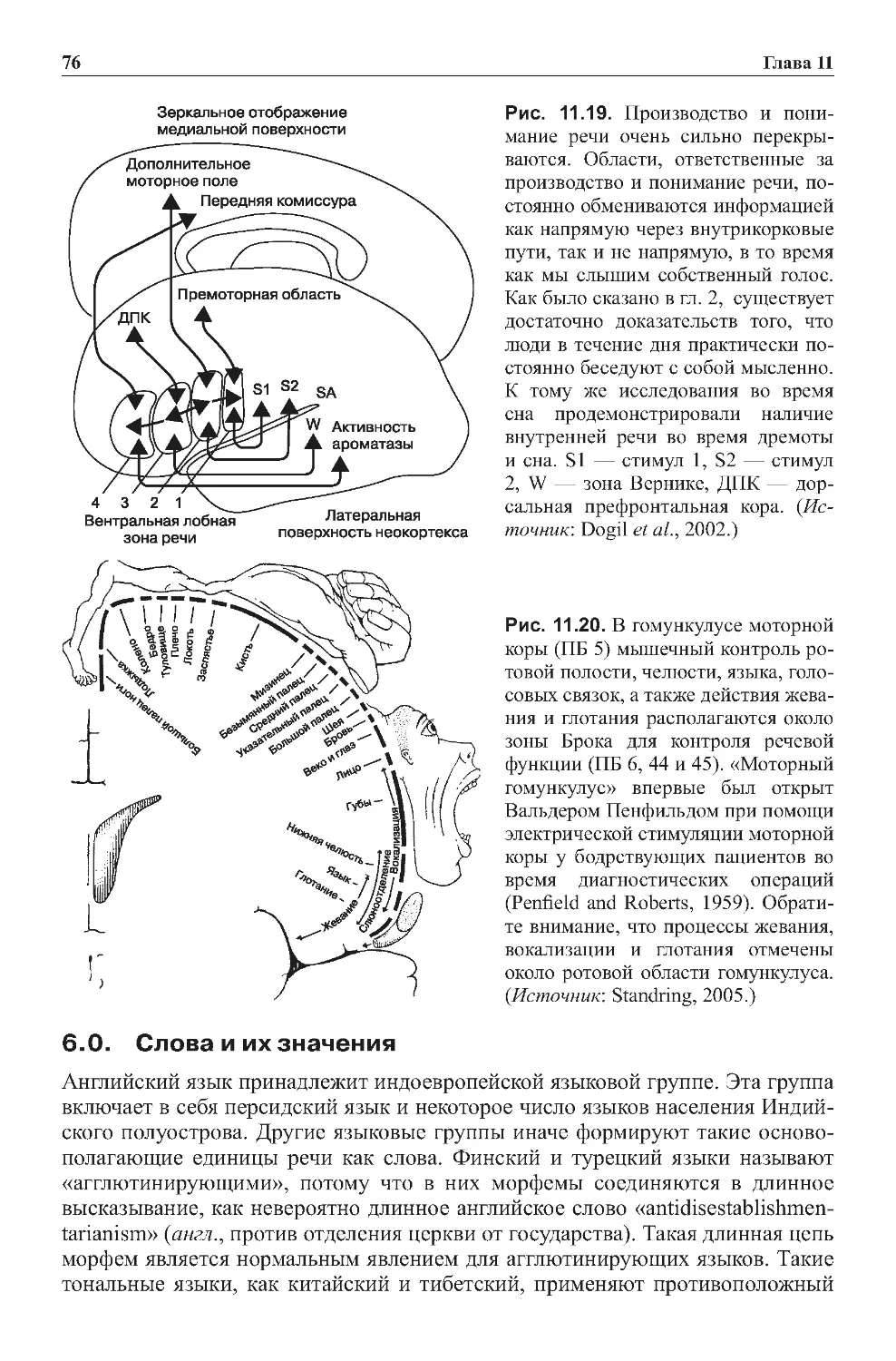



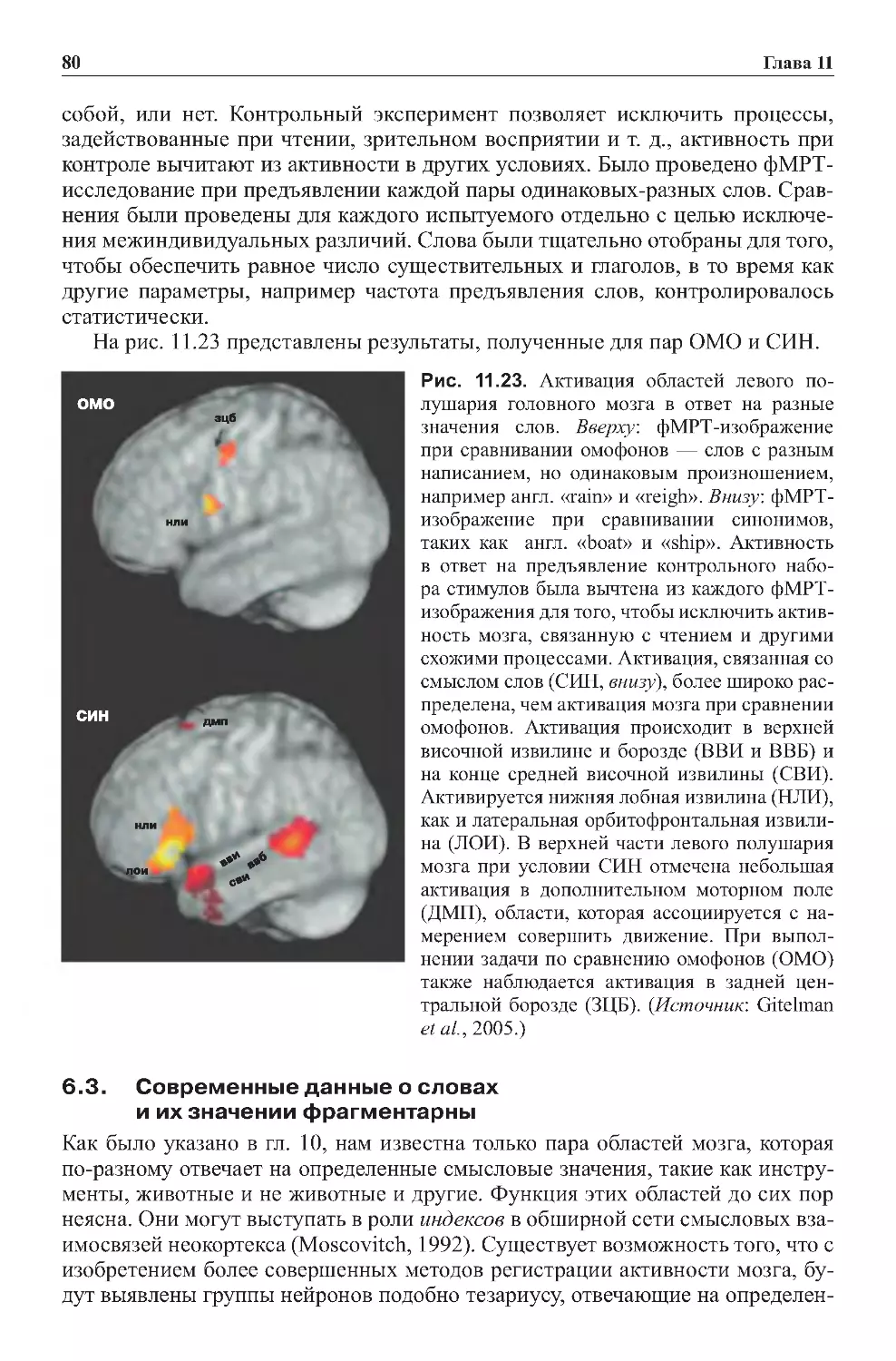

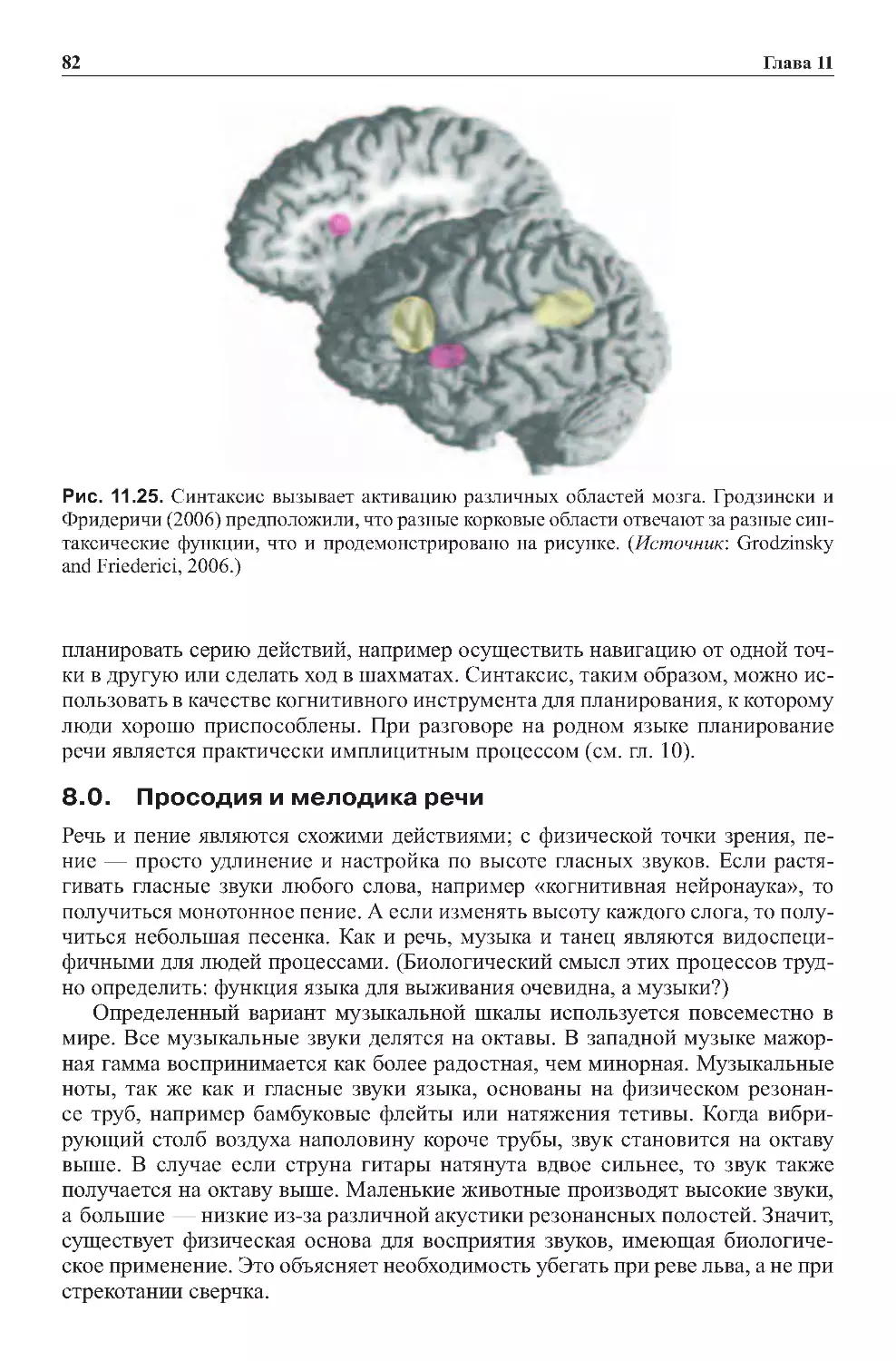










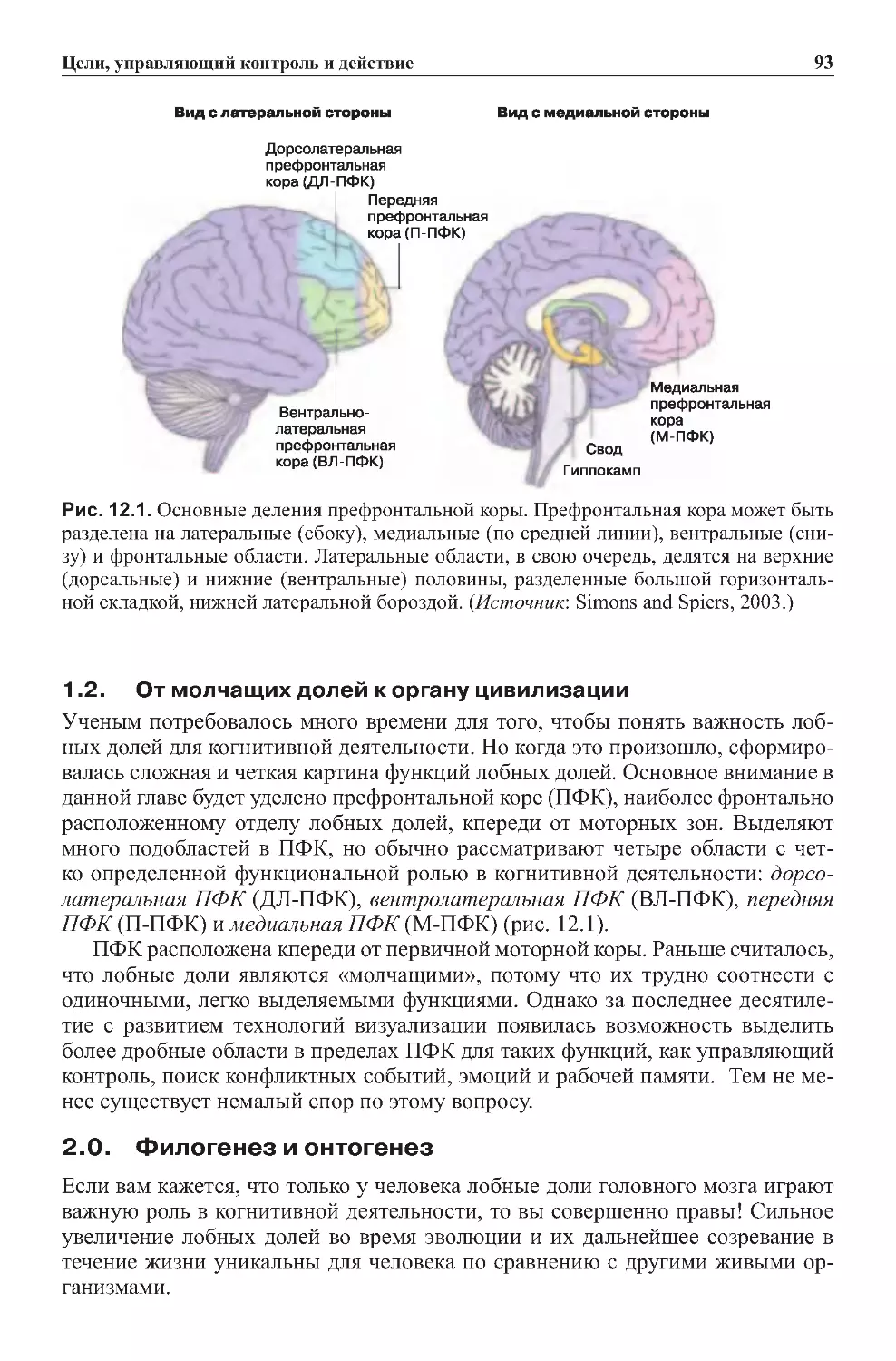
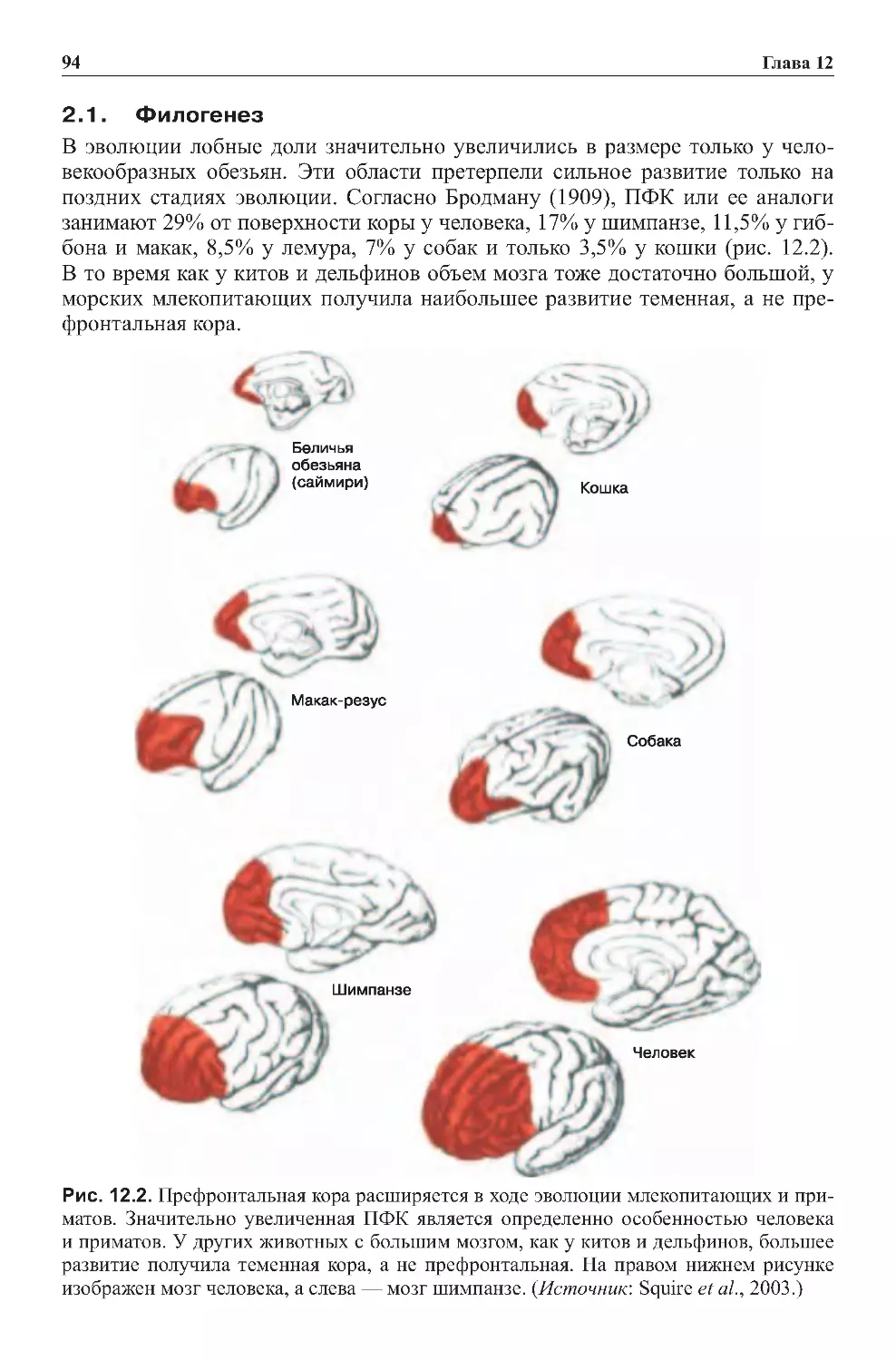























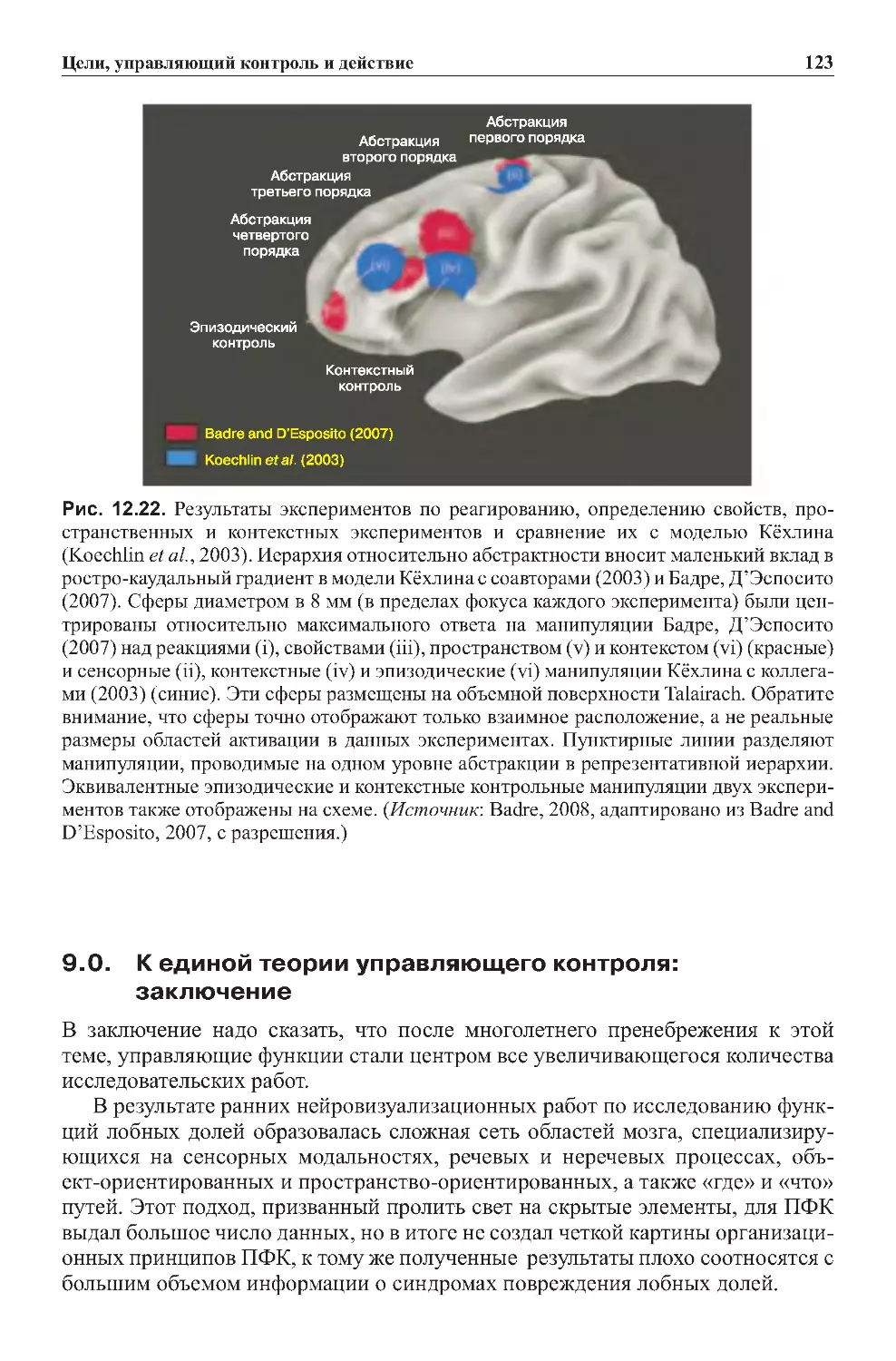




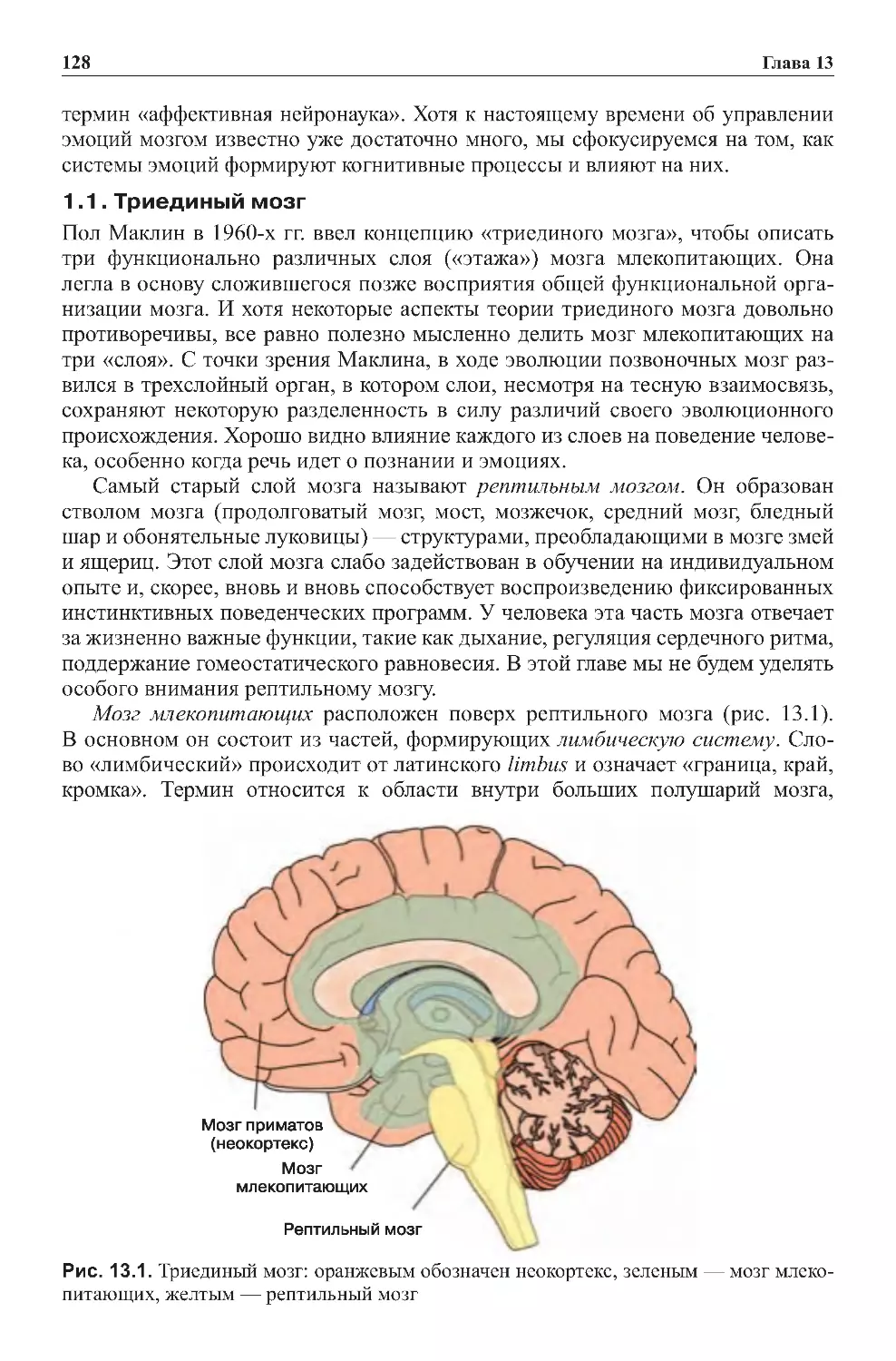









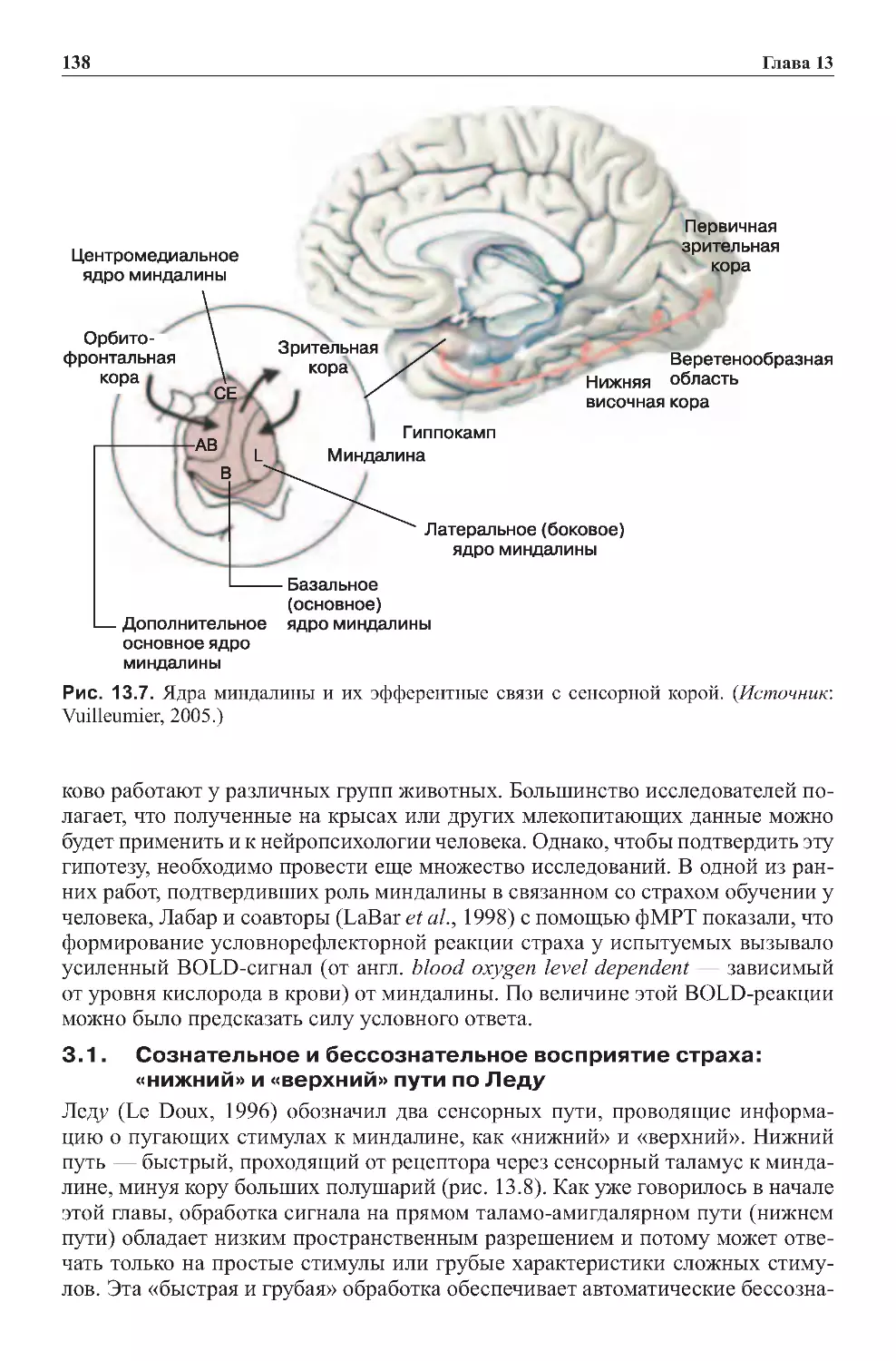
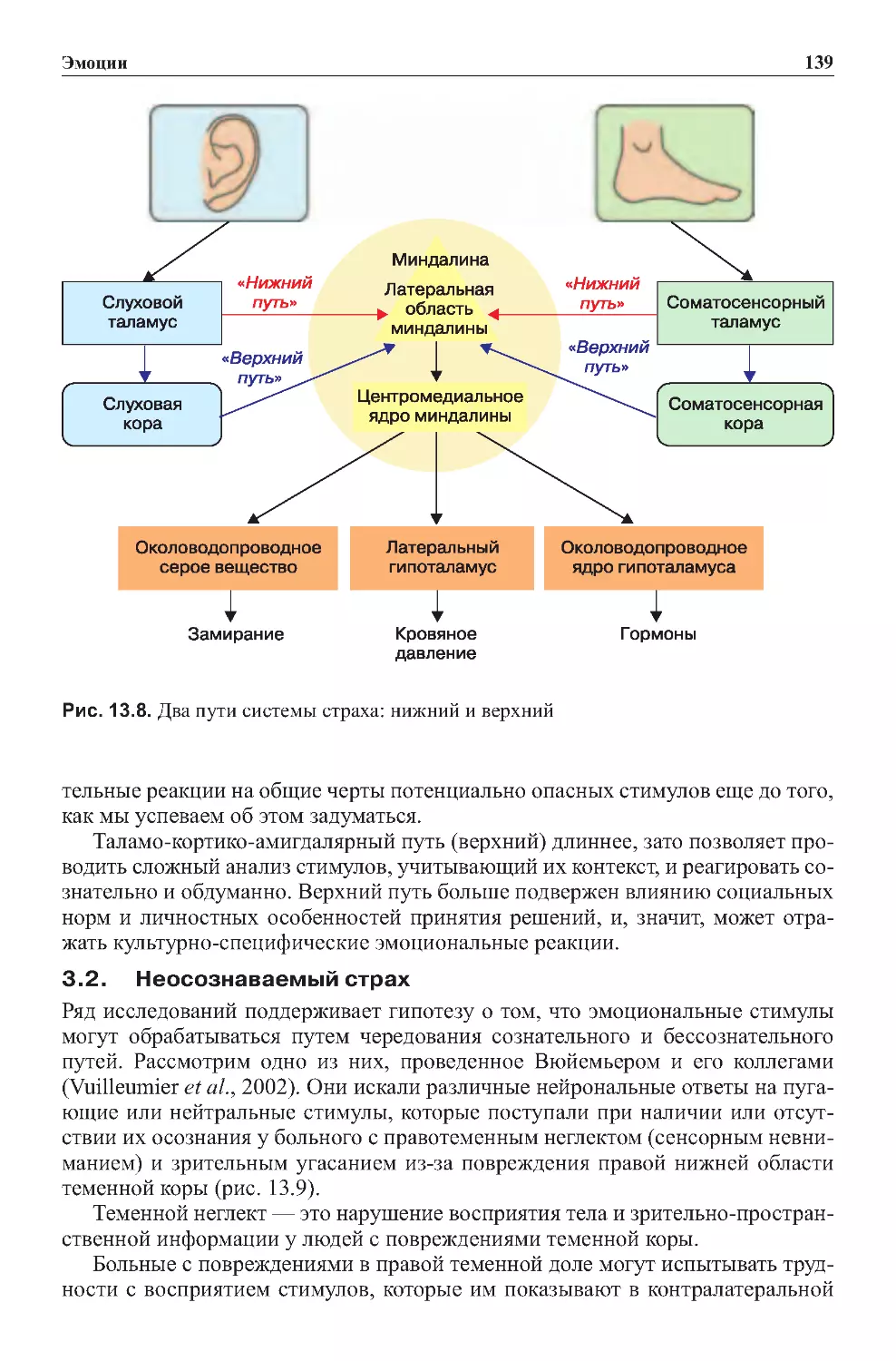


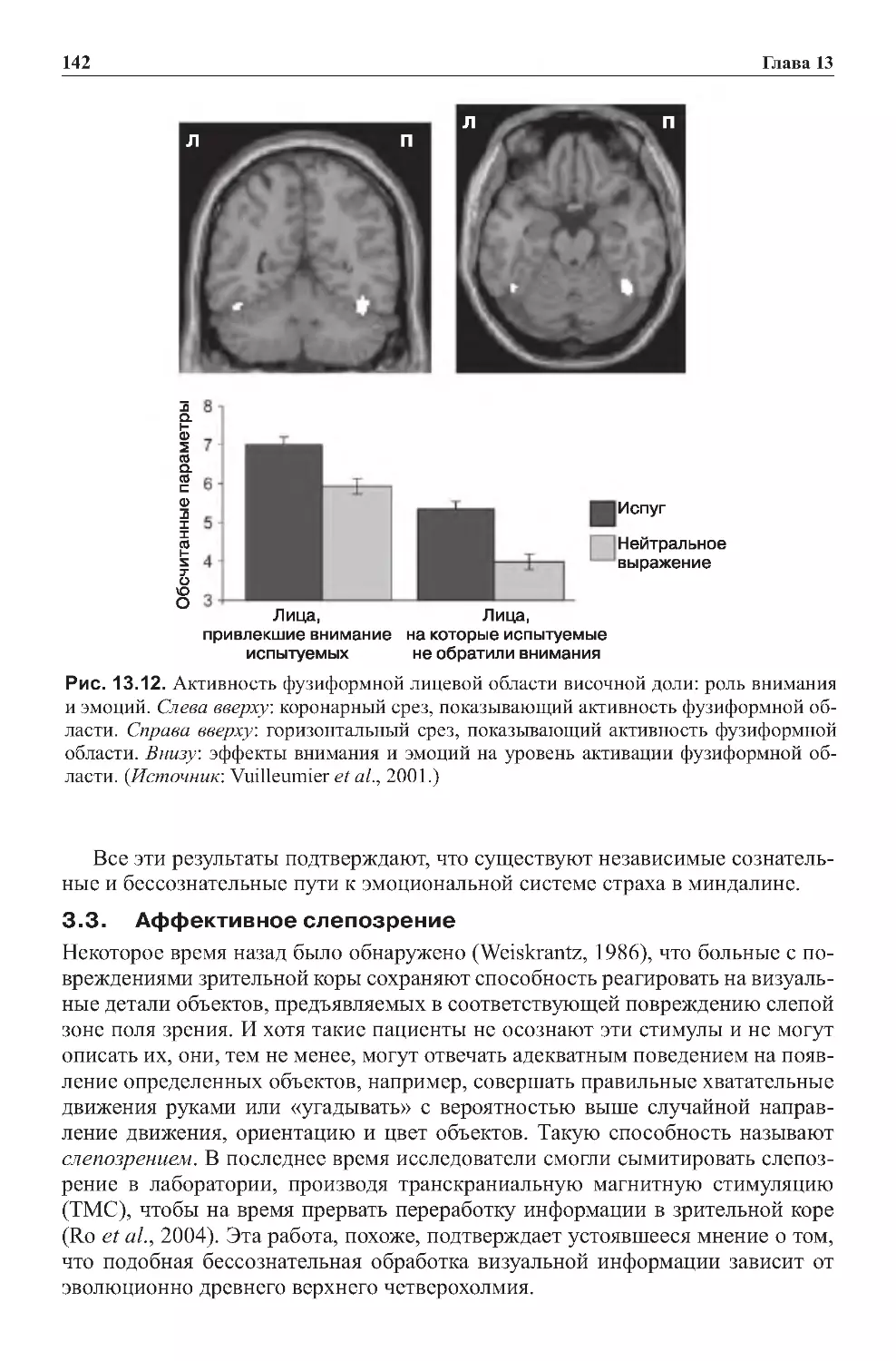


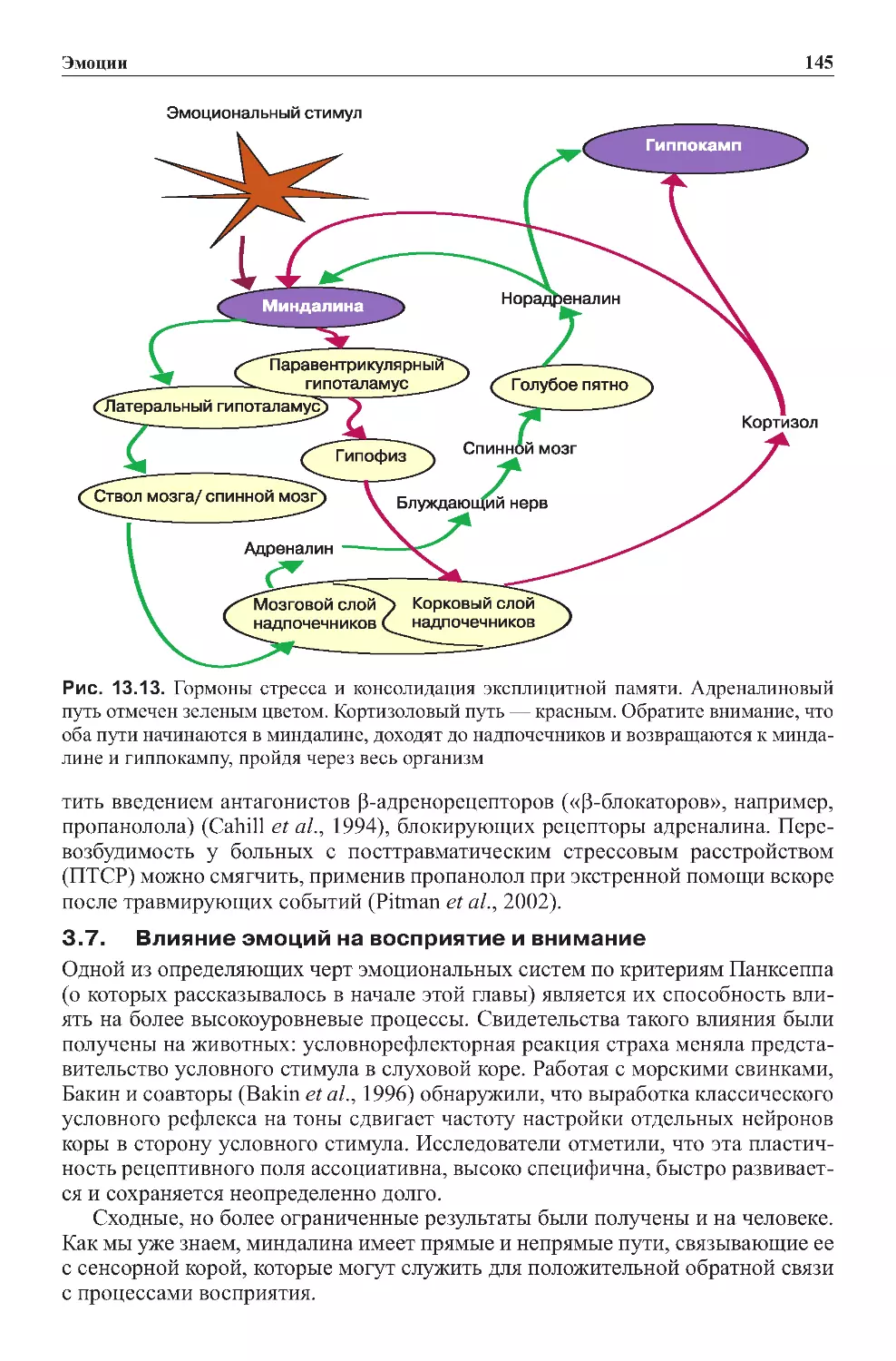

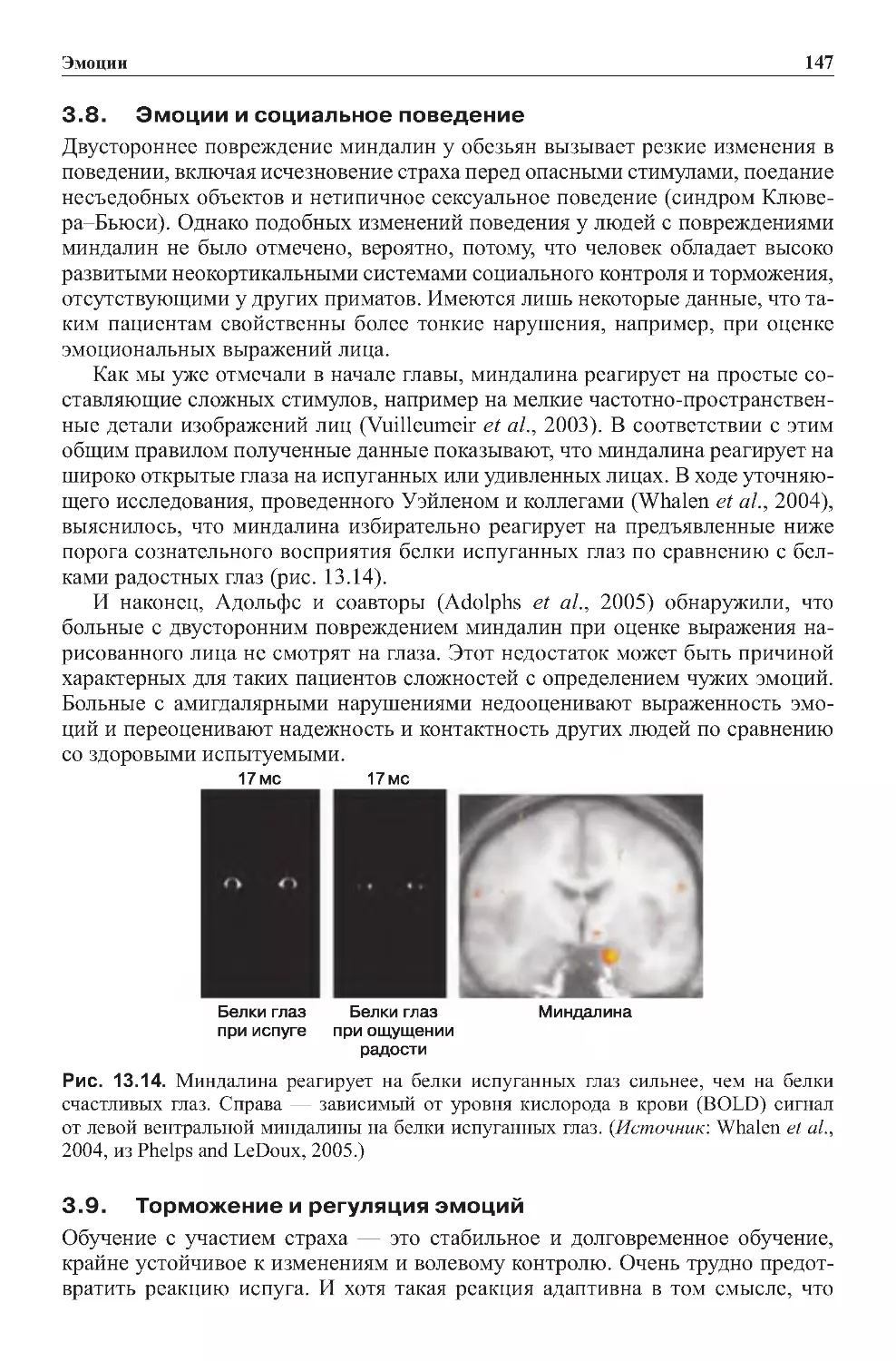



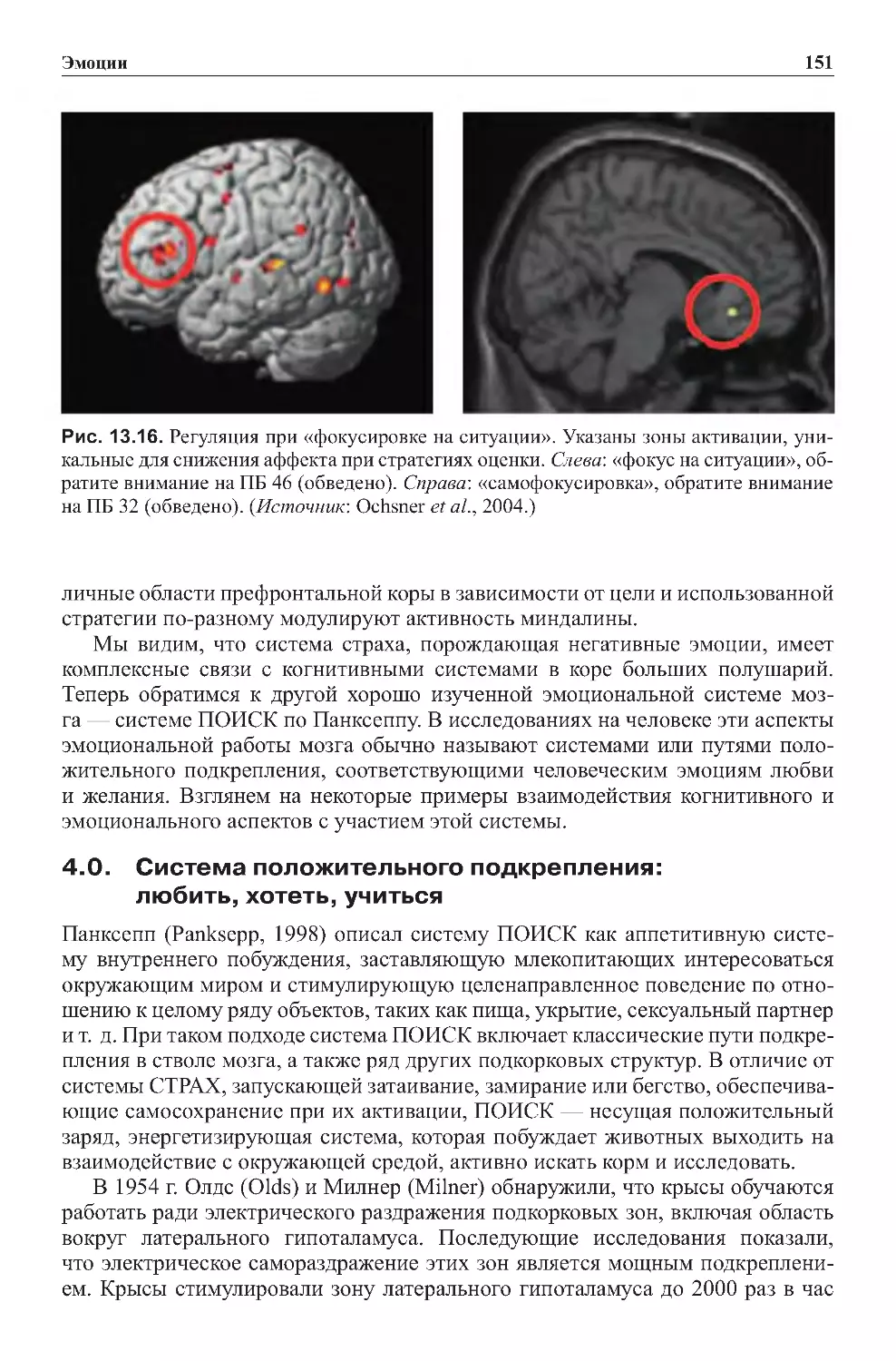
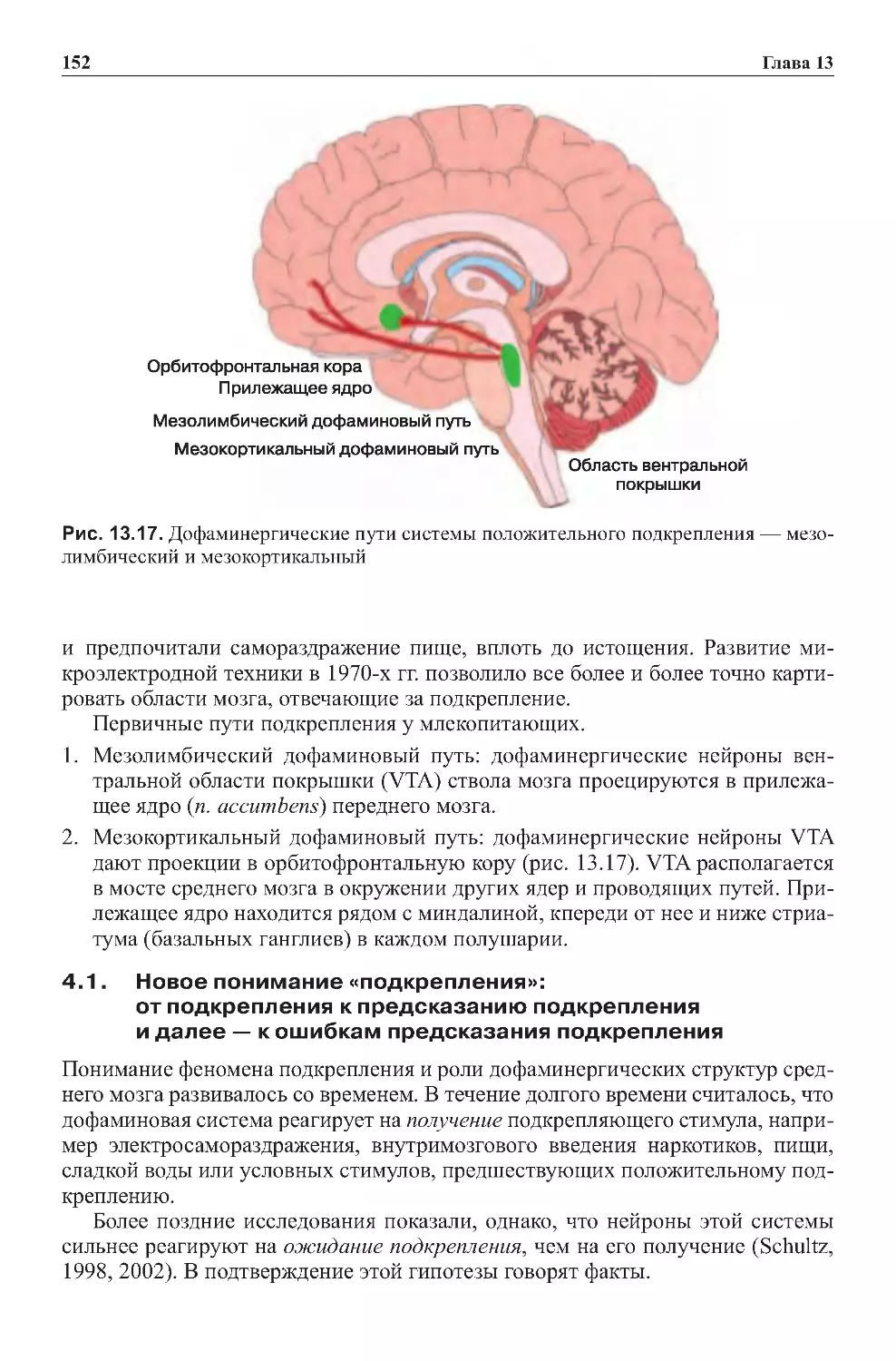

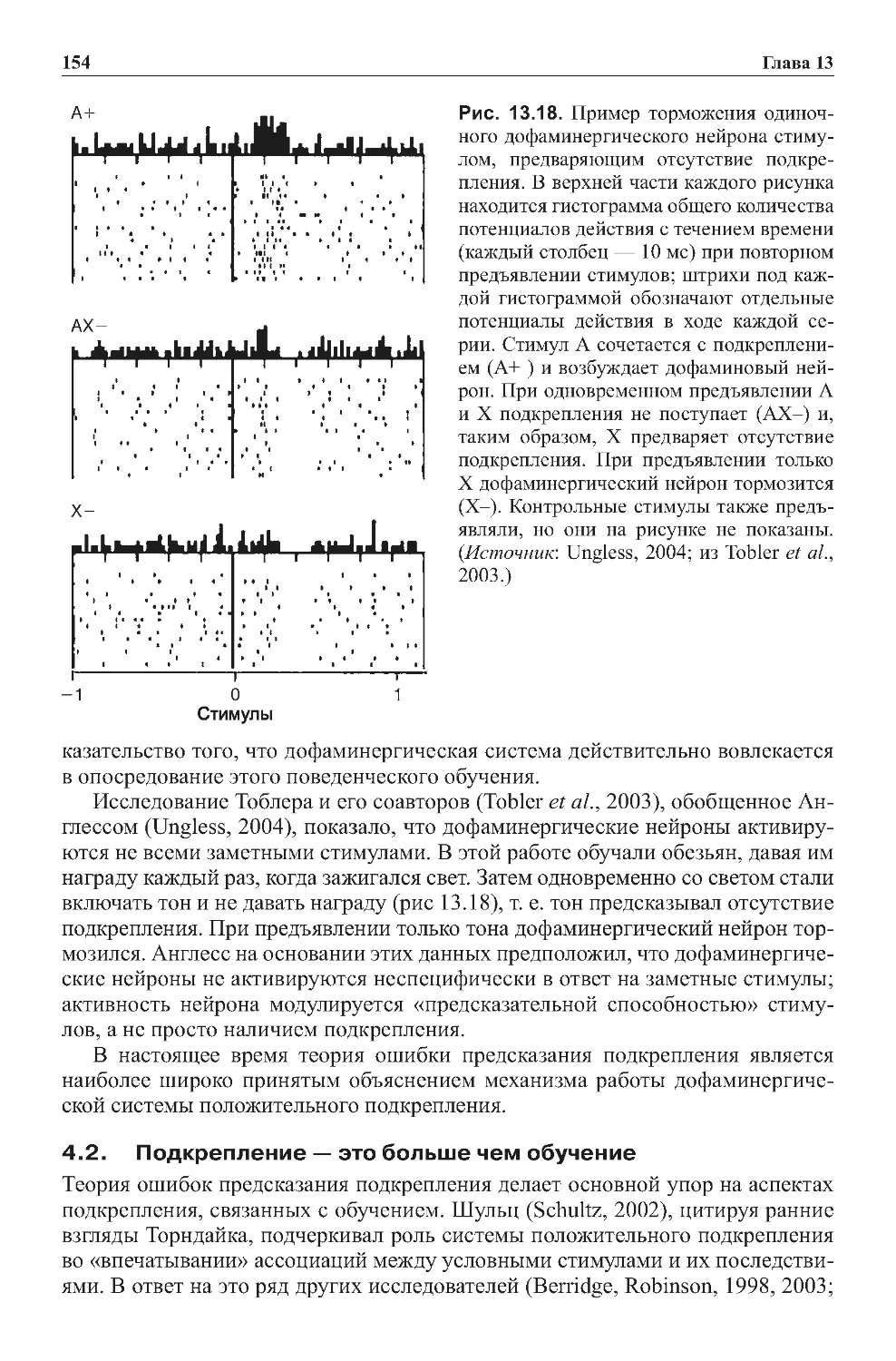








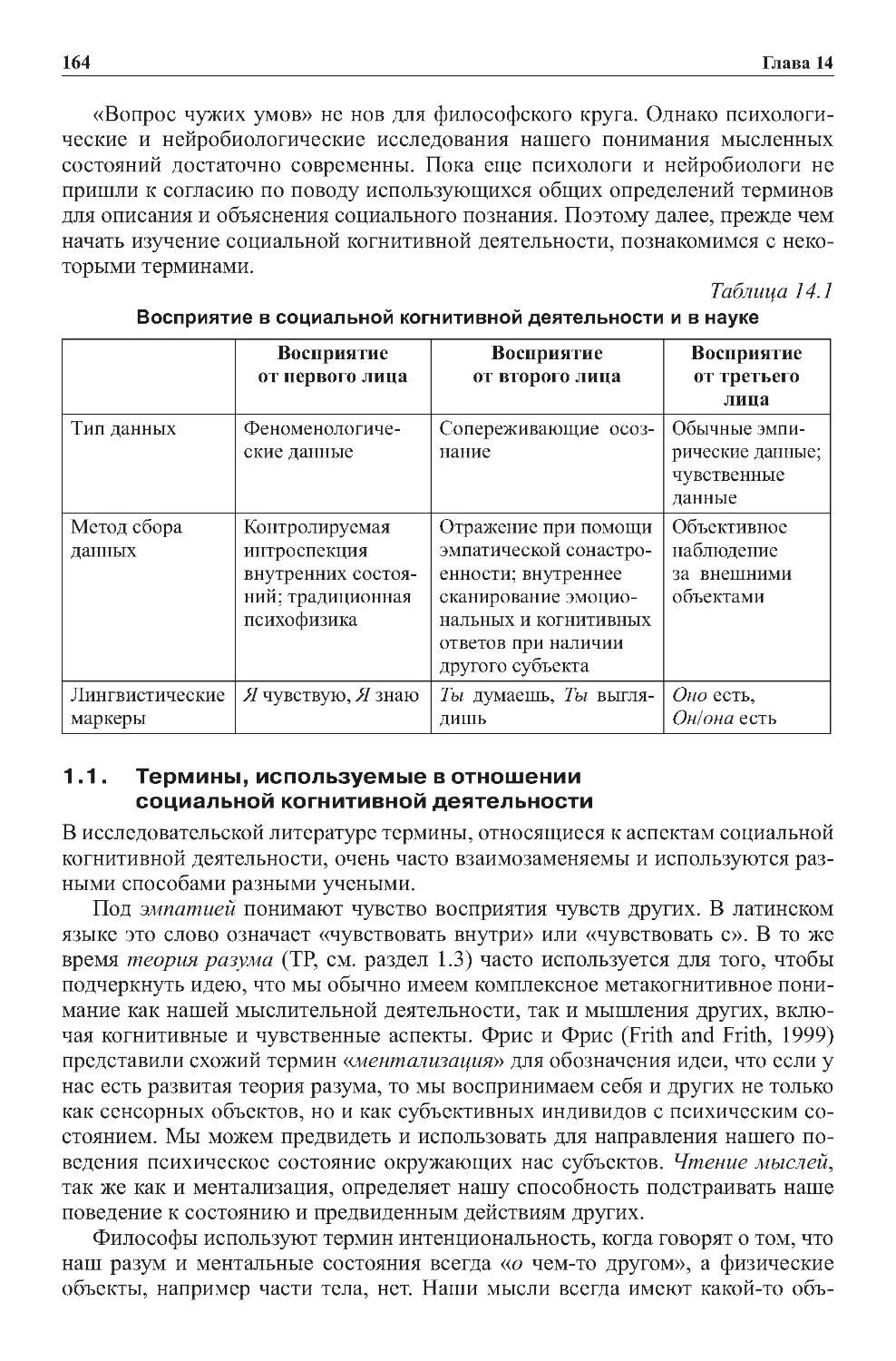














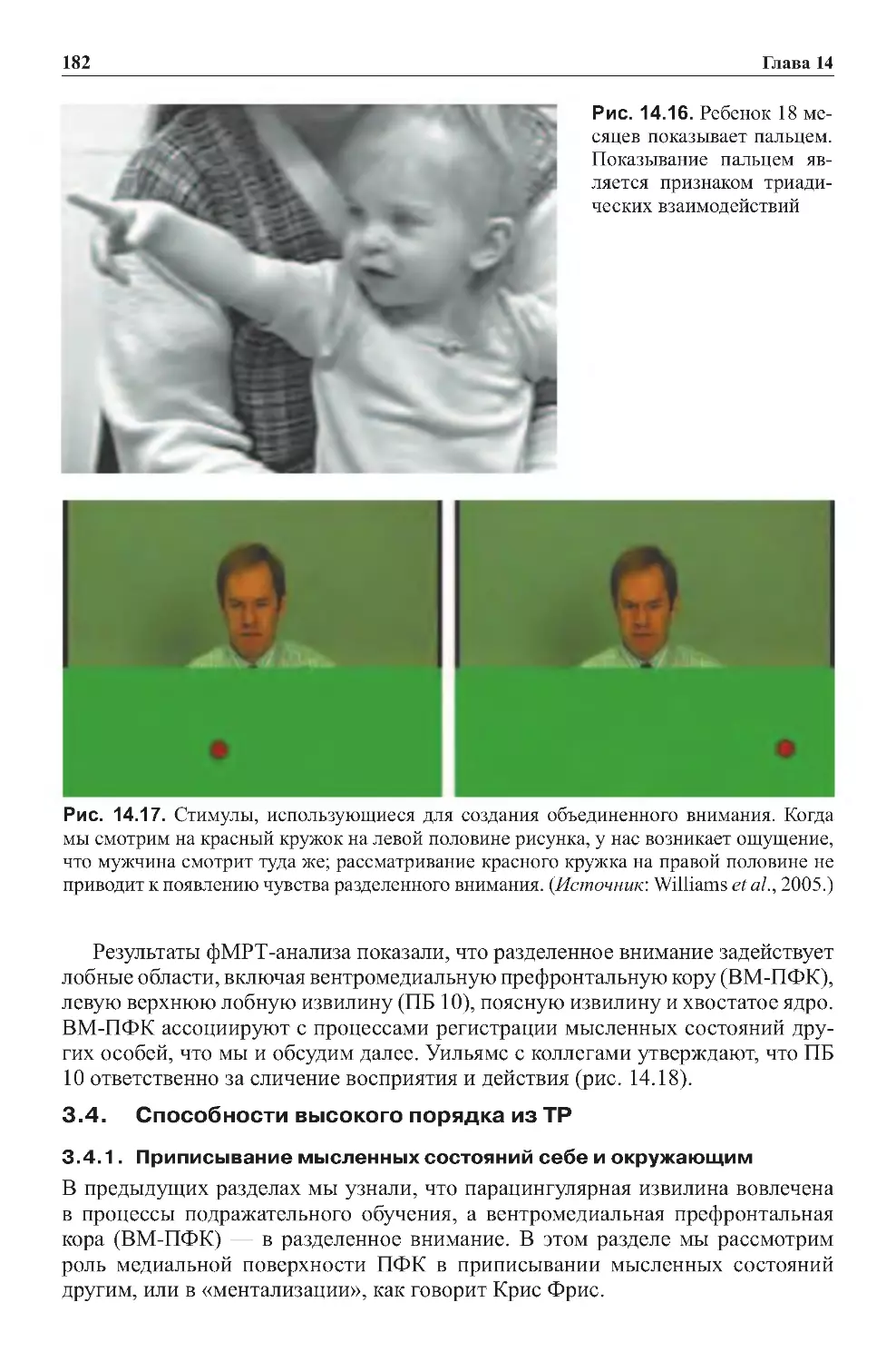























































































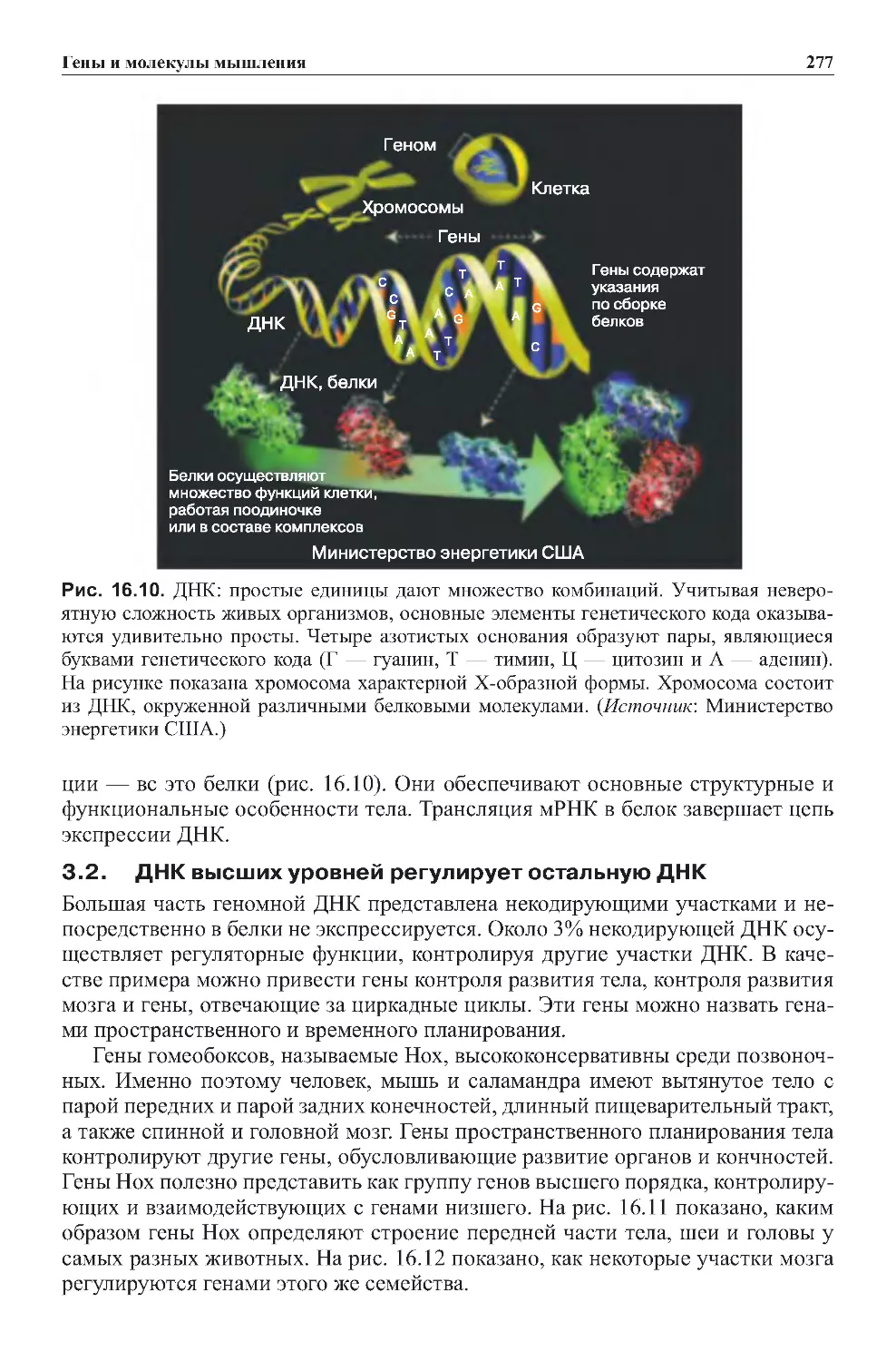



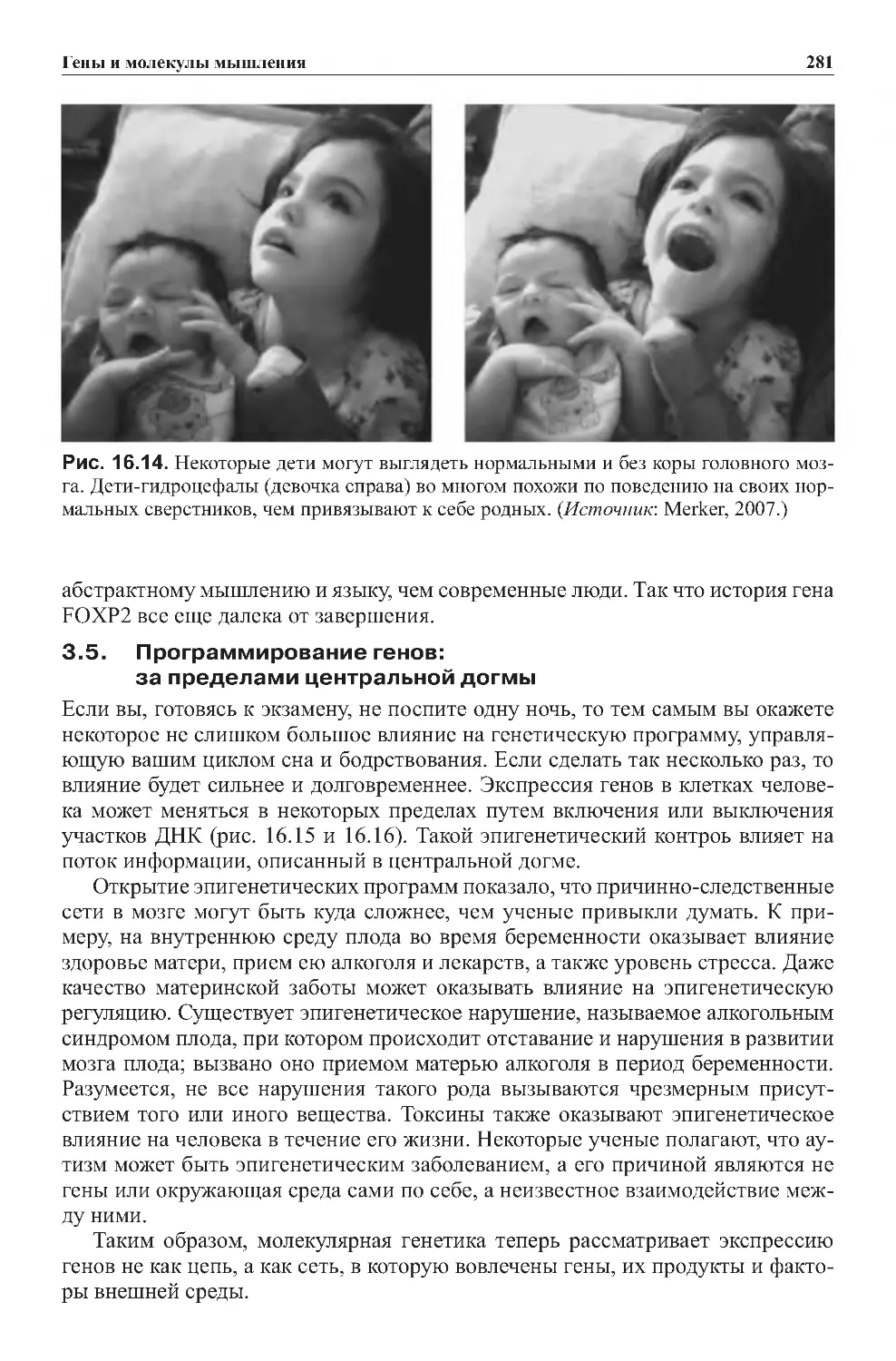
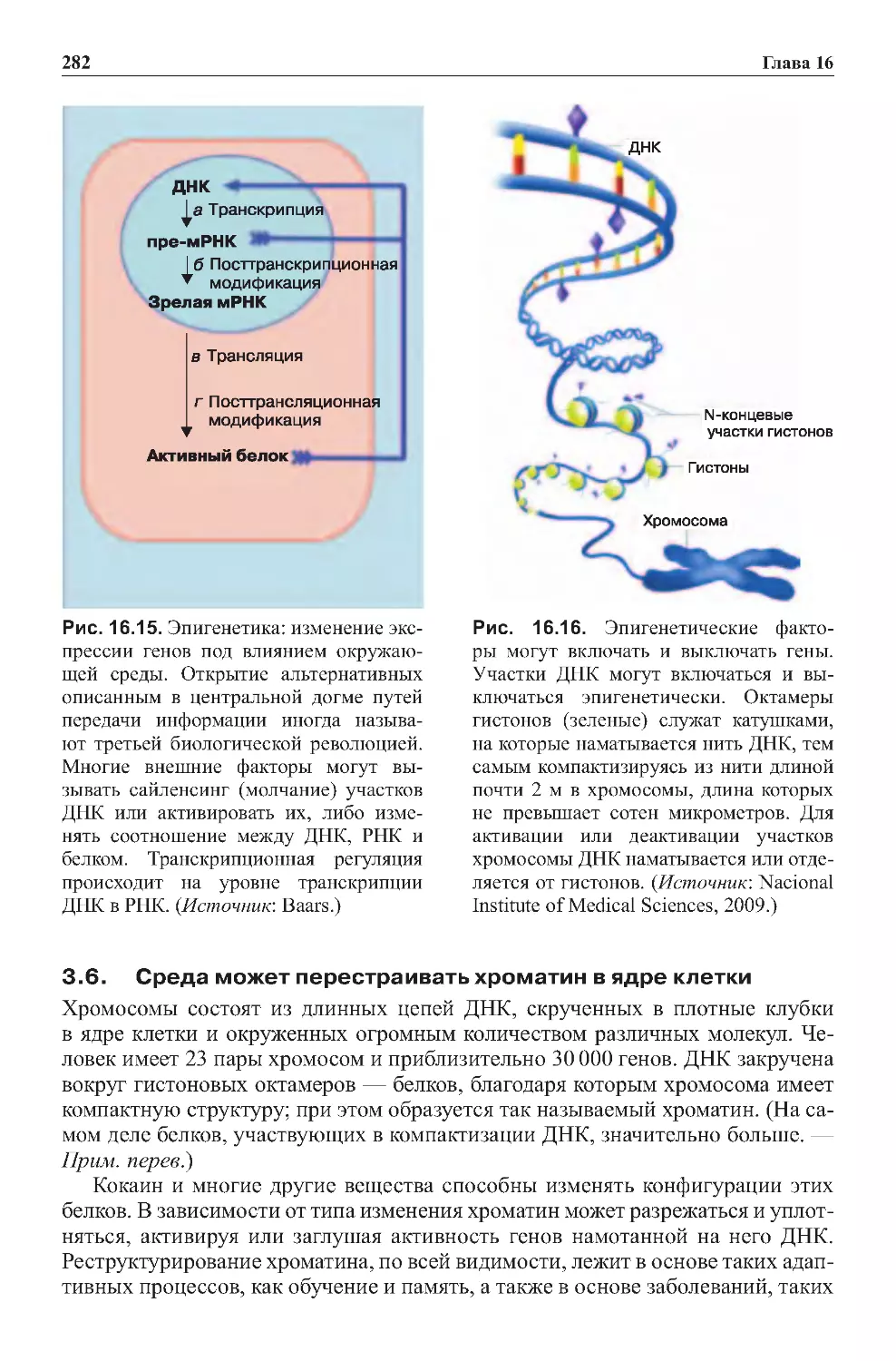


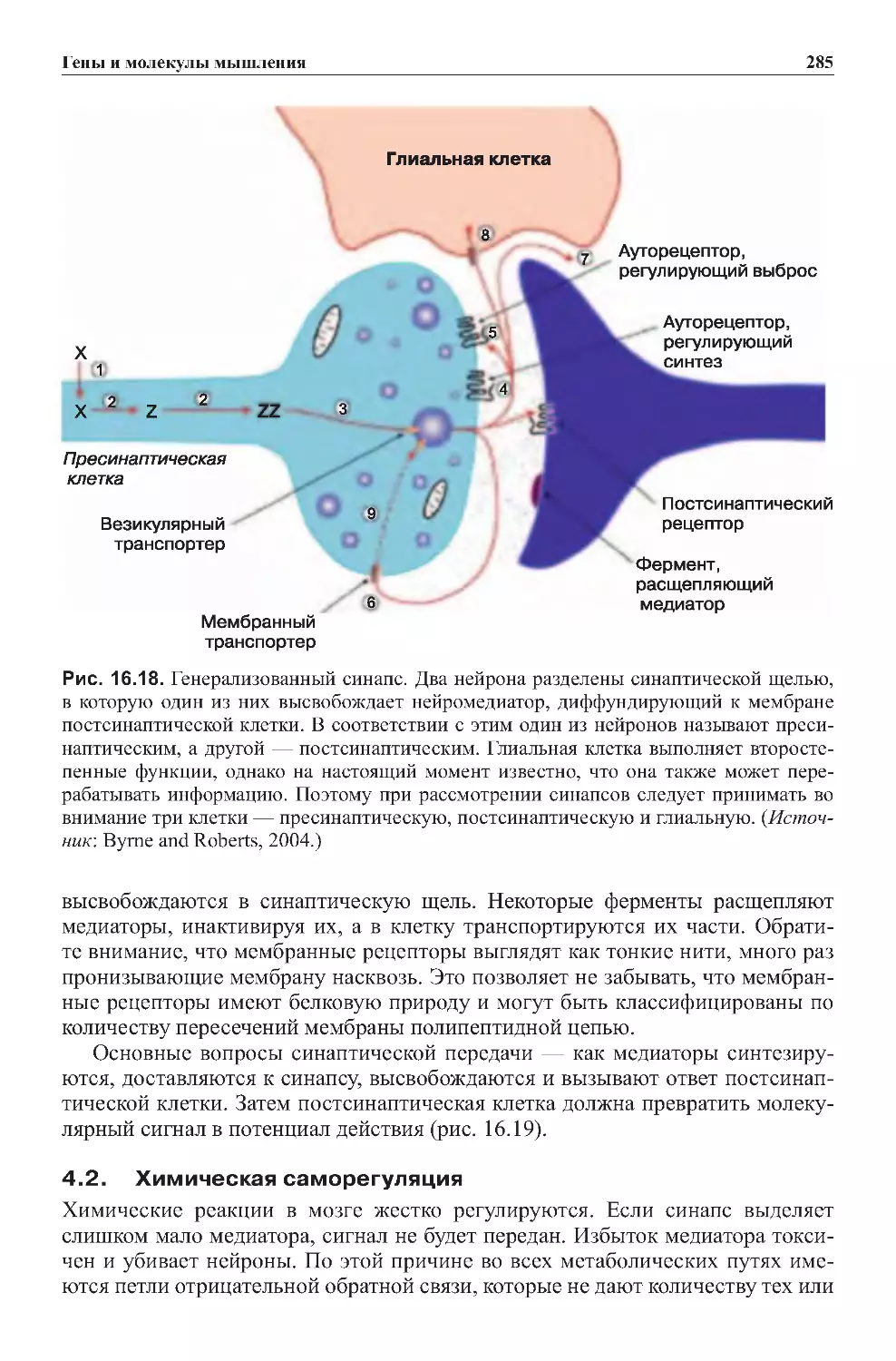
















































































































































































Теги: физиология сравнительная физиология антропология психология нейропсихология мозг человека
ISBN: 978-5-9963-2352-4
Год: 2014




Текст
МОЗГ
ПОЗНАНИЕ
РАЗУМ
COGNITION,
BRAIN,
AND CONSCIOUSNESS
INTRODUCTION TO COGNITIVE
NEUROSCIENCE
Second Edition
Bernard J. Baars
Nicole M. Gage
AMSTERDAM • BOSTON • HEIDELBERG • LONDON • NEW YORK • OXFORD
PARIS • SAN DIEGO • SAN FRANCISCO • SINGAPORE • SYDNEY • TOKYO
Academic Press is an imprint of Elsevier
Москва
БИНОМ. Лаборатория знаний
2014
МОЗГ
ПОЗНАНИЕ
РАЗУМ
ВВЕДЕНИЕ В КОГНИТИВНЫЕ
НЕЙРОНАУКИ
Перевод 2
го английского издания
под общей редакцией
профессора, д
ра биол. наук
В. В . Шульговского
Б. Баарс, Н. Гейдж
ЛУЧШИЙ ЗАРУБЕЖНЫЙ УЧЕБНИК
ЛУЧШИЙ ЗАРУБЕЖНЫЙ УЧЕБНИК
В двух томах
1
ЭЛЕКТРОННОЕ ИЗДАНИЕ
УДК 612
ББК 28.707
М74
Серия основана в2006 г.
Переводчики:
В. Н. Егорова (гл. 6, 8), М. А. Каменская (гл. 1, 2),
В. М. Ковальзон (гл. 5, 7, 9, 13), А. В. Любителев (гл. 3, 16),
О. Б. Мацелепа (гл. 4, 10–12, 14, 15)
М74
Мозг, познание, разум: введение в когнитивные нейронауки
[Электронный ресурс] : в 2 ч. Ч. 1 / под ред. Б. Баарса, Н. Гейдж ;
пер. с англ. под ред. проф. В . В . Шульговского. — Эл. изд. — Элек-
трон. текстовые дан. (1 файл pdf : 552 с.) . — М . : БИНОМ. Лабо-
ратория знаний, 2014. — (Лучший зарубежный учебник). — Систем.
требования: Adobe Reader XI ; экран 10".
ISBN 978-5-9963-2352-4 (Ч. 1)
ISBN 978-5-9963-1452-2
В книге изложены ре зультаты новейших исследований связи строения
мозга и способно сти человека к познанию. В наглядной и до ступной фор-
ме представлена взаимо связь строения мозга, психологических функций,
эффектов во сприятия и умственных спо собно стей человека. Выводы о сно-
ваны на данных, полученных в психологии, биологии, медицине, биохимии
и физике. Показано, что когнитивные механизмы и механизмы во сприя-
тия, которые изучались на уровне поведения, в настоящее время могут
наблюдаться непо средственно на уровне работы с амого мозга благодаря
использованию новейших методов визуализации мозговых процессов.
Для студентов и преподавателей биологических, психологиче ских и ме-
дицинских вузов, педагогов, всех интере сующихся когнитивной нейронаукой
и работающих в этой области.
УДК 612
ББК 28.707
Деривативное электронное издание на основе печатного аналога: Мозг,
познание, разум: введение в когнитивные нейронауки : в 2 ч. Ч . 1 / под ред.
Б. Баарса, Н. Гейдж ; пер. с англ. под ред. проф. В . В . Шульговского. —
М. : БИНОМ. Лаборатория знаний, 2014. — 541 с. : ил ., [8] с. цв. вкл. —
(Лучший зарубежный учебник).
В соответствии со ст. 1299 и 1301 ГК РФ при устранении ограничений, уст ановленных
техническими средствами защиты авторских прав, правообладатель вправе требовать
от нарушителя возмещения убытков или выплаты компенсации
ISBN 978-5-9963-2352-4 (Ч. 1)
ISBN 978-5-9963-1452-2
c
○ 2010 Elsevier Ltd. All rights reserved.
This edition of COGNITION, BRAIN, AND
CONSCIOUSNESS, INTRODUCTION TO COGNITIVE
NEUROSCIENCE by Bernard Baars and Nicole Gage
is published by arrangement with ELSEVIER LIMITED
of The Boulevard, Langford Lane, Kidlington, Oxford,
OX5 1GB, UK. ISBN 978-0 -12-375070-9 .
Книга «Мозг, познание, разум: введение
в когнитивные нейронауки» под ред. Б. Баарса,
Н. Гейдж опубликована с разрешения ELSEVIER
LIMITED, The Boulevard, Langford Lane, Kidlington,
Oxford, OX5 1GB, UK
c
○ Перевод и оформление. БИНОМ. Лаборатория
знаний, 2014
4
Предисловие..............................................14
Глава1.Психикаимозг.....................................23
1.0.Введение......................................................23
2.0.Приглашаемвнейропсихологию..................................24
3.0.Некоторыеисходныеположения..................................25
3.1.Расстояния:семьпорядковразмерностей..........................25
3.2.Время:десятьпорядковразмерностей............................28
3.3. Умозаключения — выход за рамки эмпирических данных . . . . . . . . . . . 30
3.4.Согласуемостьданных.........................................34
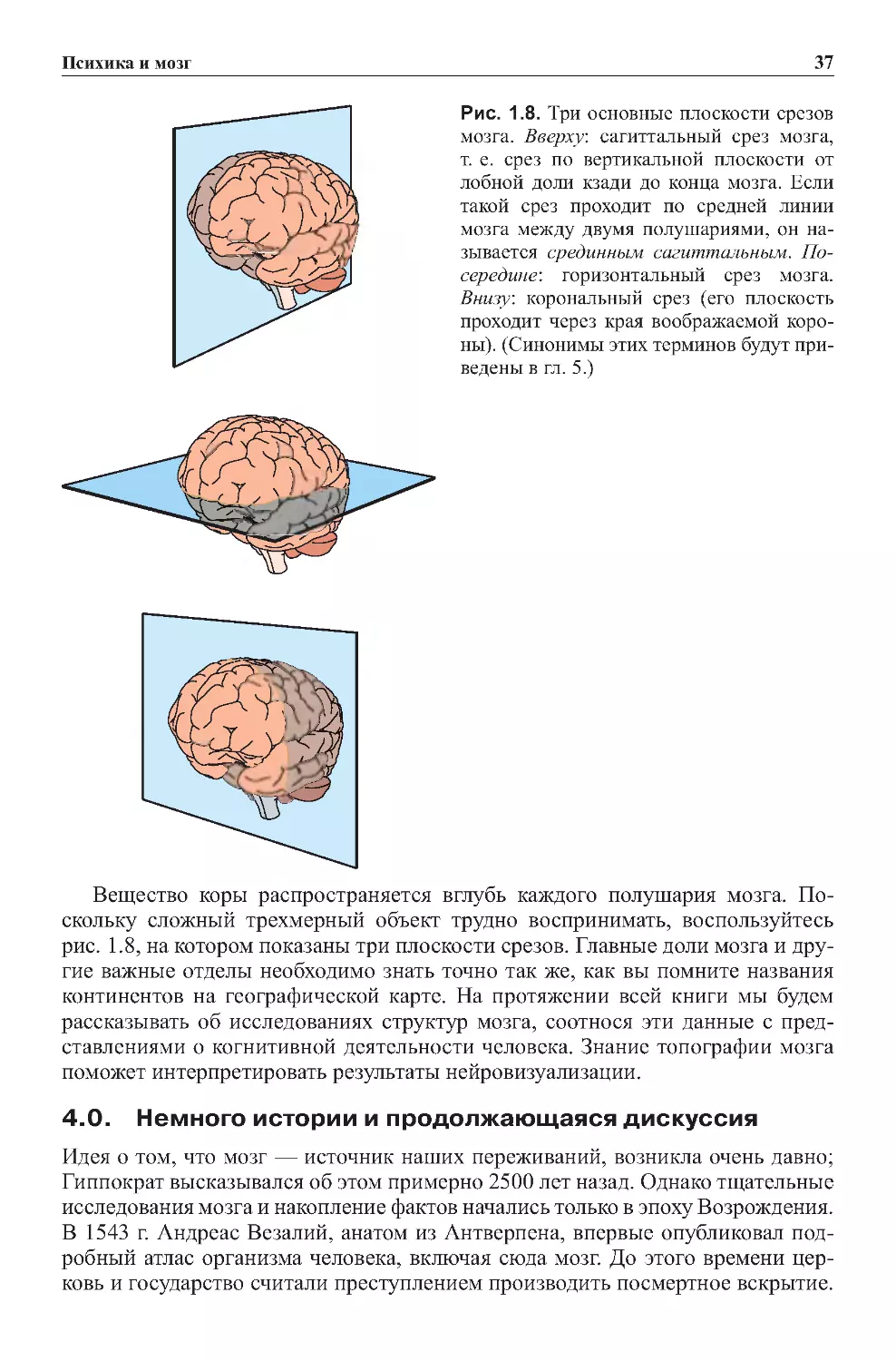
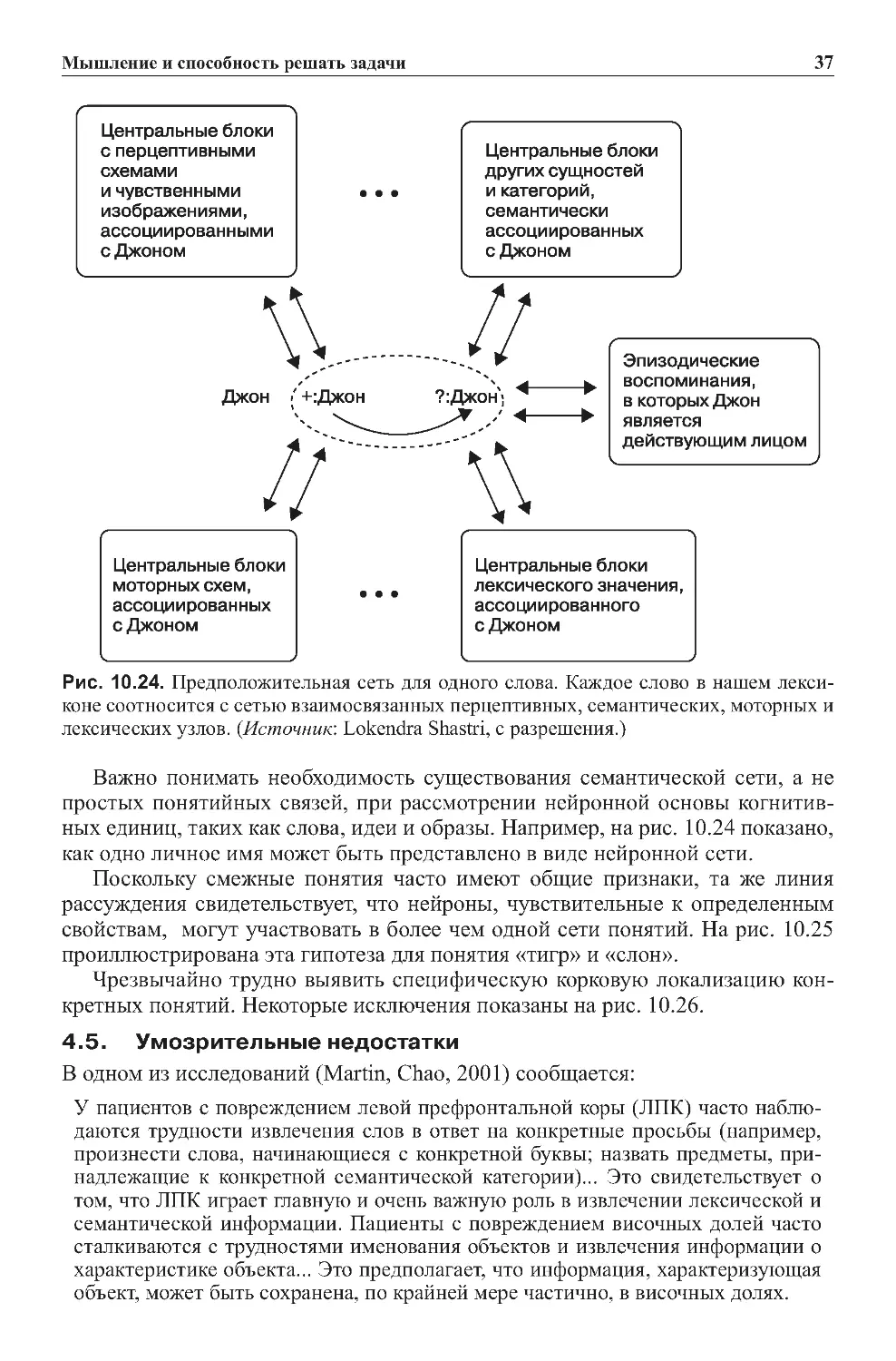
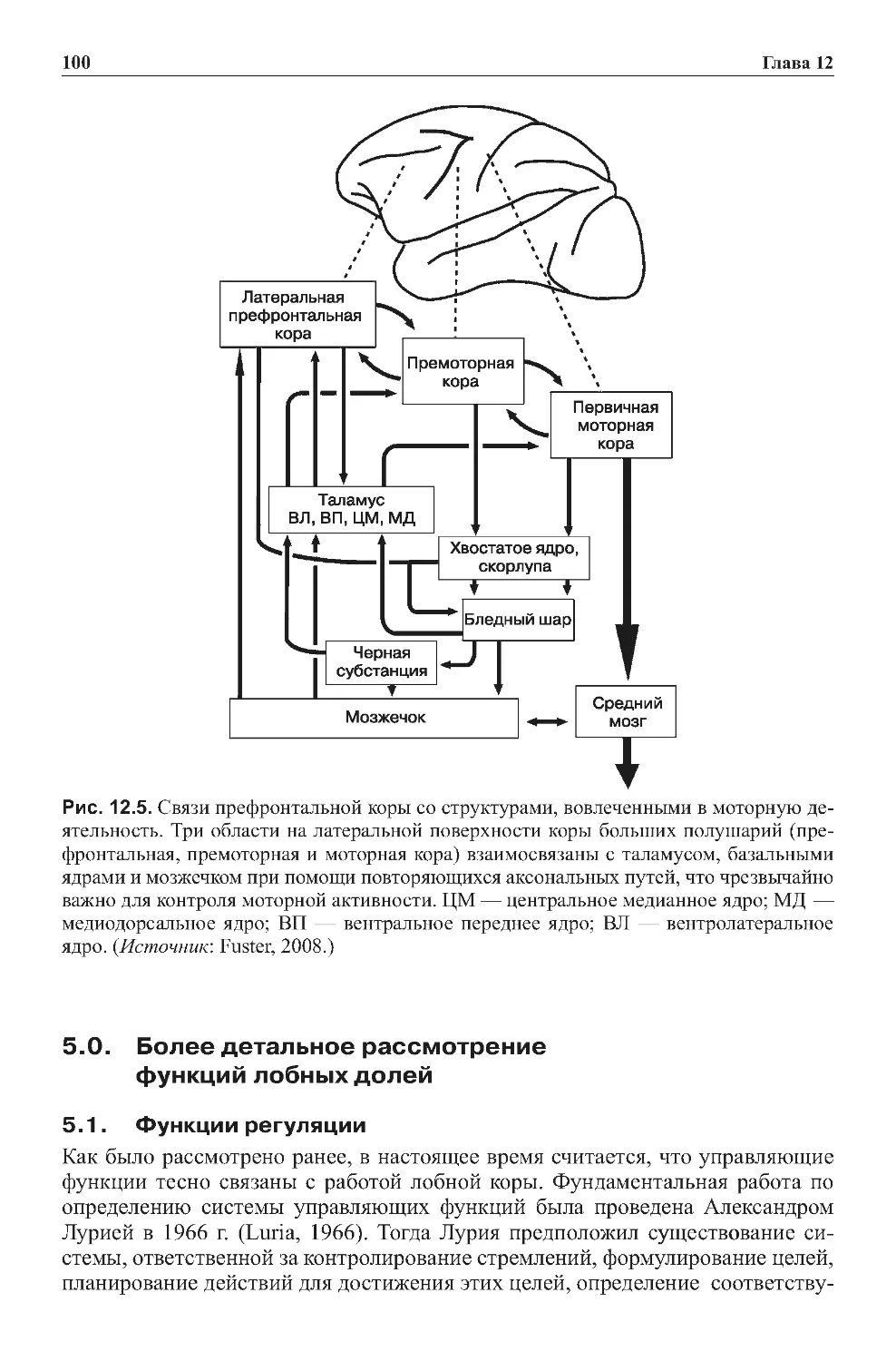
3.5.Главныеориентирынаизображенияхмозга.......................35
4.0.Немногоисторииипродолжающаясядискуссия.....................37
4.1.Психикаимозг...............................................40
4.2. Биологическая природа когнитивной деятельности и эмоций . . . . . . . . . 41
4.3. Нейронная доктрина Рамон-и-Кахала: рабочая гипотеза
наукиомозге.................................................44
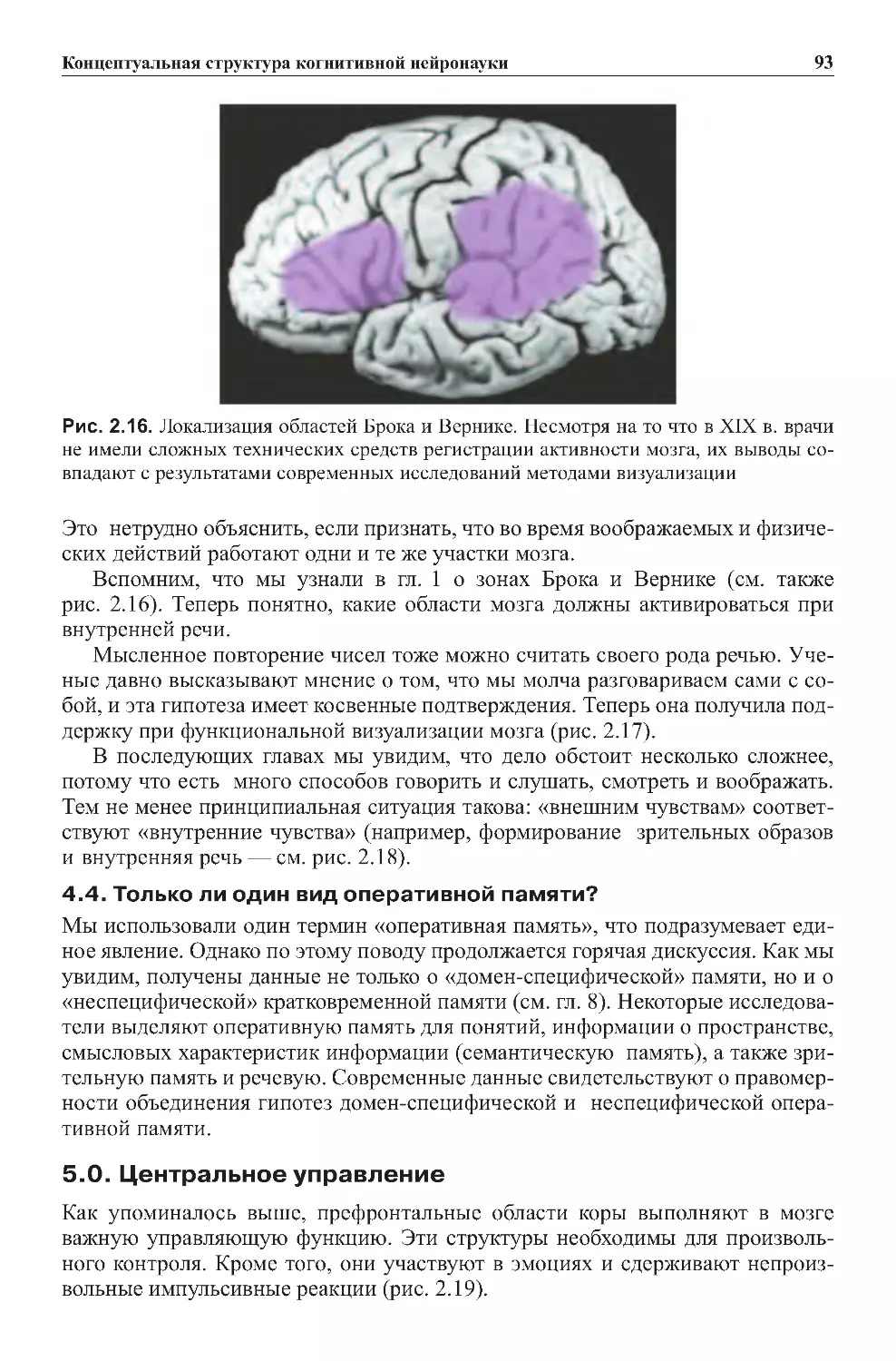
4.4.Пьер-ПольБрока:локализацияцентраречи.......................47
4.5. Осознаваемые и неосознаваемые психические явления . . . . . . . . . . . . . . 54
5.0.Наукавозвращаетсякпредставлениямосознании...................59
5.1. Современные методы исследования осознаваемых
инеосознаваемыхпроцессоввмозге.............................61
5.2.Историческоеразвитиенеостановилось..........................62
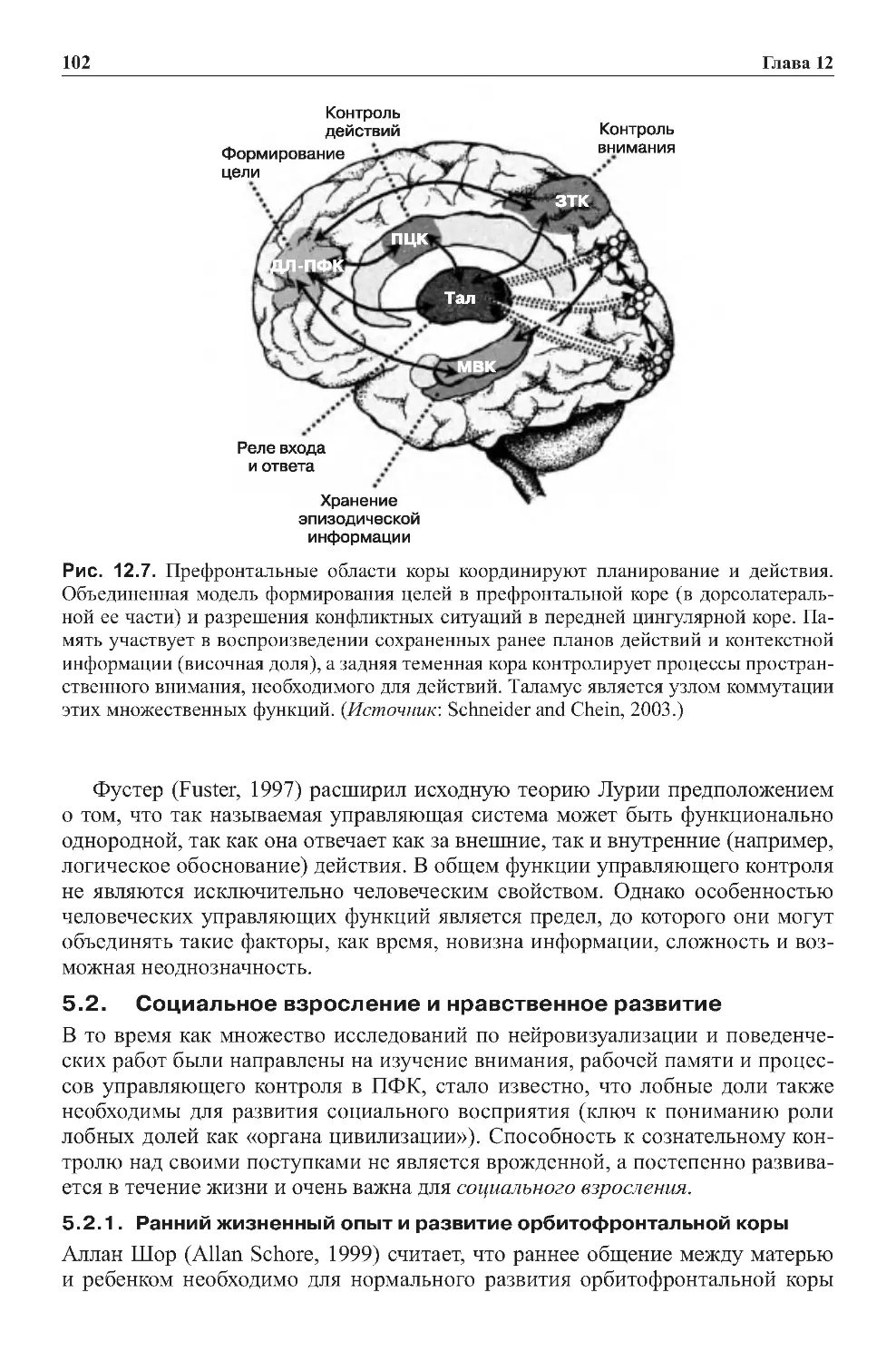
6.0.Заключение....................................................64
7.0.Заданияиупражнениякглаве1...................................66
7.1.Контрольныезадания..........................................66
7.2.Графическиеупражнения.......................................67
Оглавление
6
Оглавление
Глава 2. Концептуальная структура когнитивной нейронауки . . . . 69
1.0.Введение......................................................69
2.0.Классическоепонятиеоперативнойпамяти.........................71
2.1.«Внутренниечувства».........................................72
2.2.Выходныефункции............................................74
2.3.Одинкраткиймиг...............................................74
2.4. Случай Клайва Уэринга с позиций функциональной схемы . . . . . . . . . . 78
2.5.Рольмгновеннойпамяти.......................................79
3.0.Ограниченностьфункциональныхспособностей....................81
3.1.Границывыполнениядвойныхзадач.............................81
3.2. Некоторые функциональные способности мозга имеют
широкиеграницы.............................................84
3.3. Почему функциональные способности могут быть
стольограниченными?.........................................85
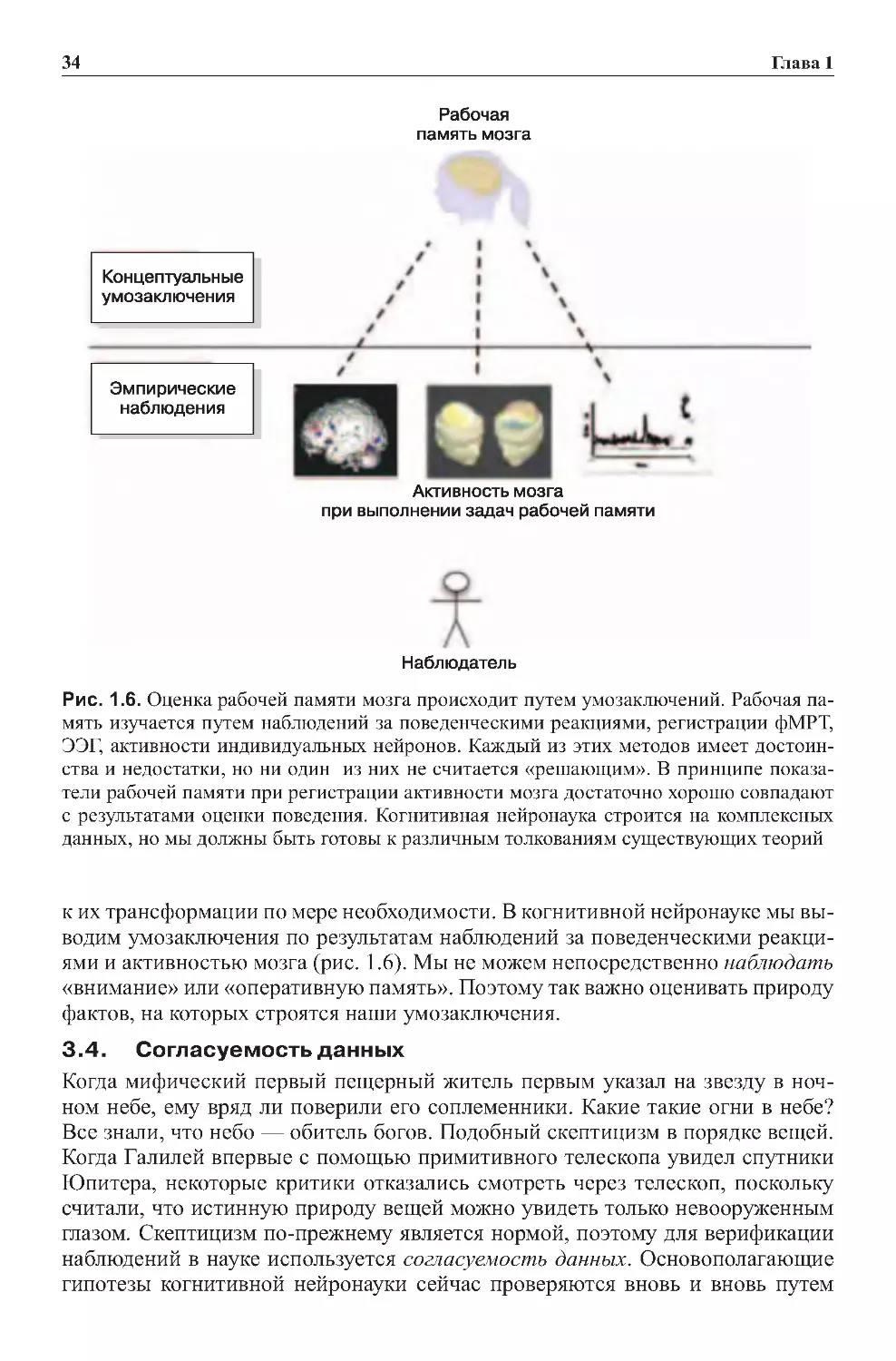
3.4.Оценкаоперативнойпамяти....................................86
4.0.Внутренниеивнешниечувства ..................................89
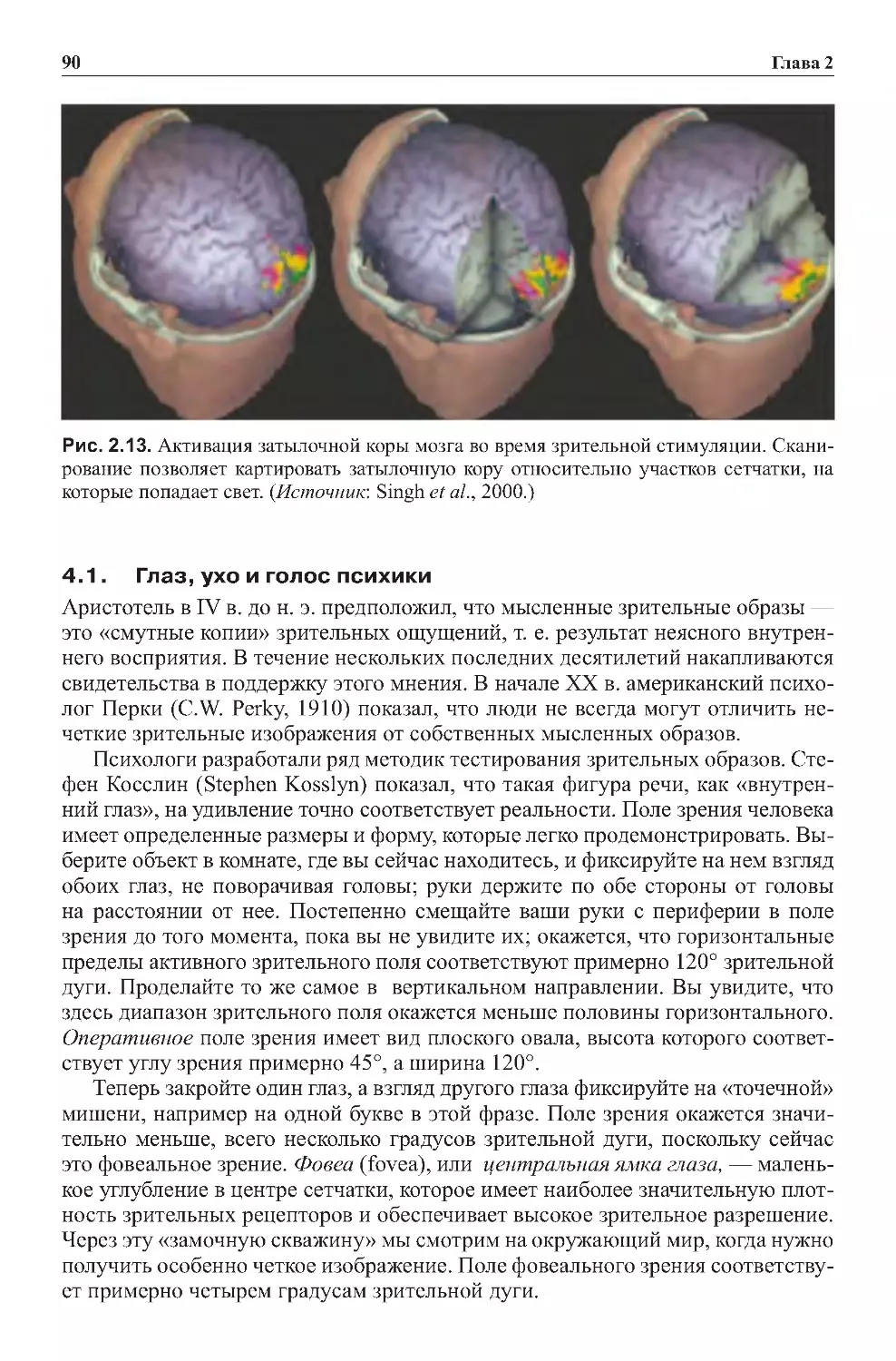
4.1.Глаз,ухоиголоспсихики.......................................90
4.2. Психические образы формируются в зрительных областях
корымозга...................................................92
4.3.Идентичналивнутренняяречьвнешнейречи?.....................92
4.4.Тольколиодинвидоперативнойпамяти? .........................93
5.0.Центральноеуправление.........................................93
5.1.Исполнительнаяреакцияиавтоматизм...........................96
5.2.Управляемоеиспонтанноевнимание ............................98
6.0.Действие.....................................................100
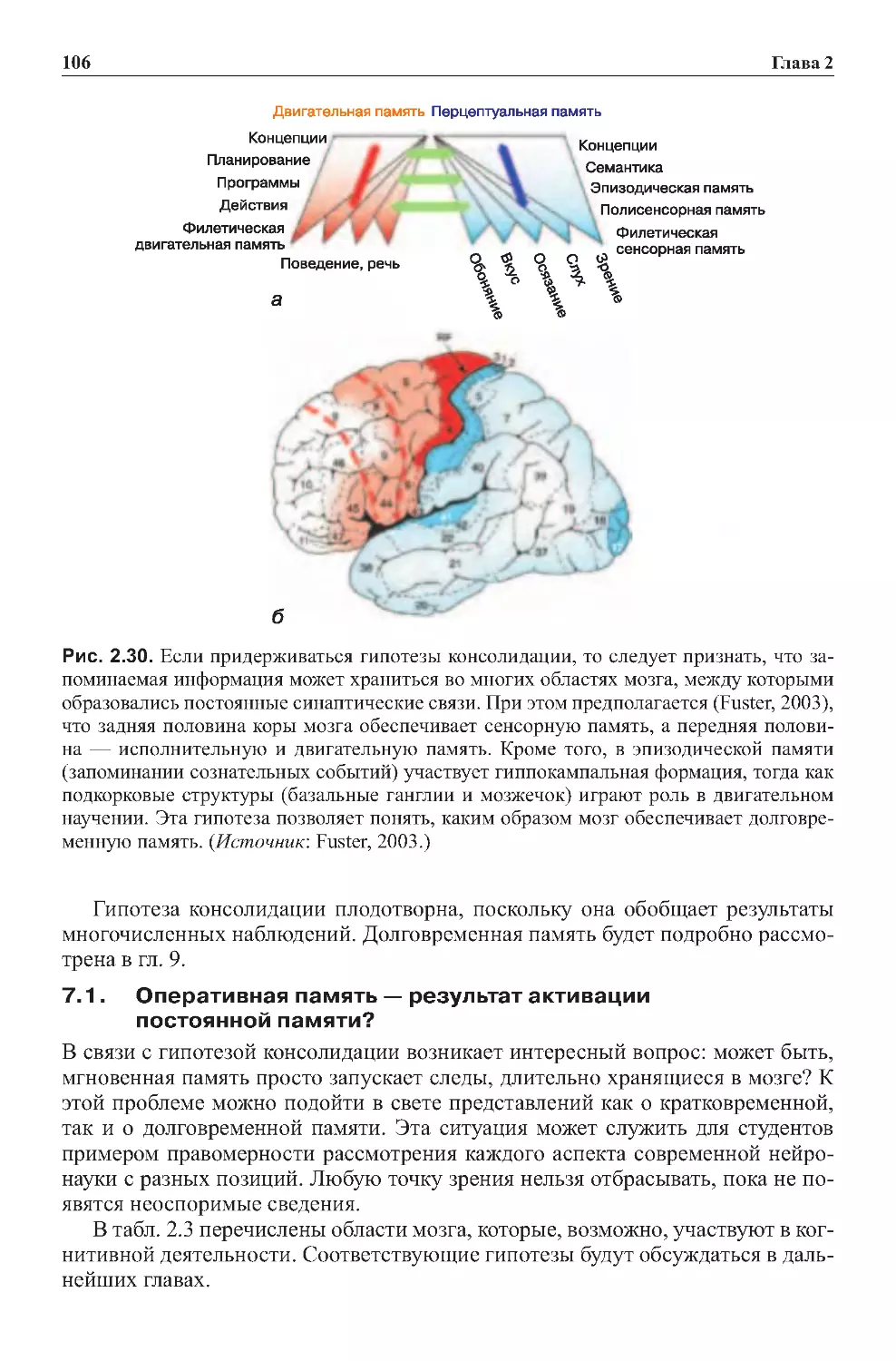
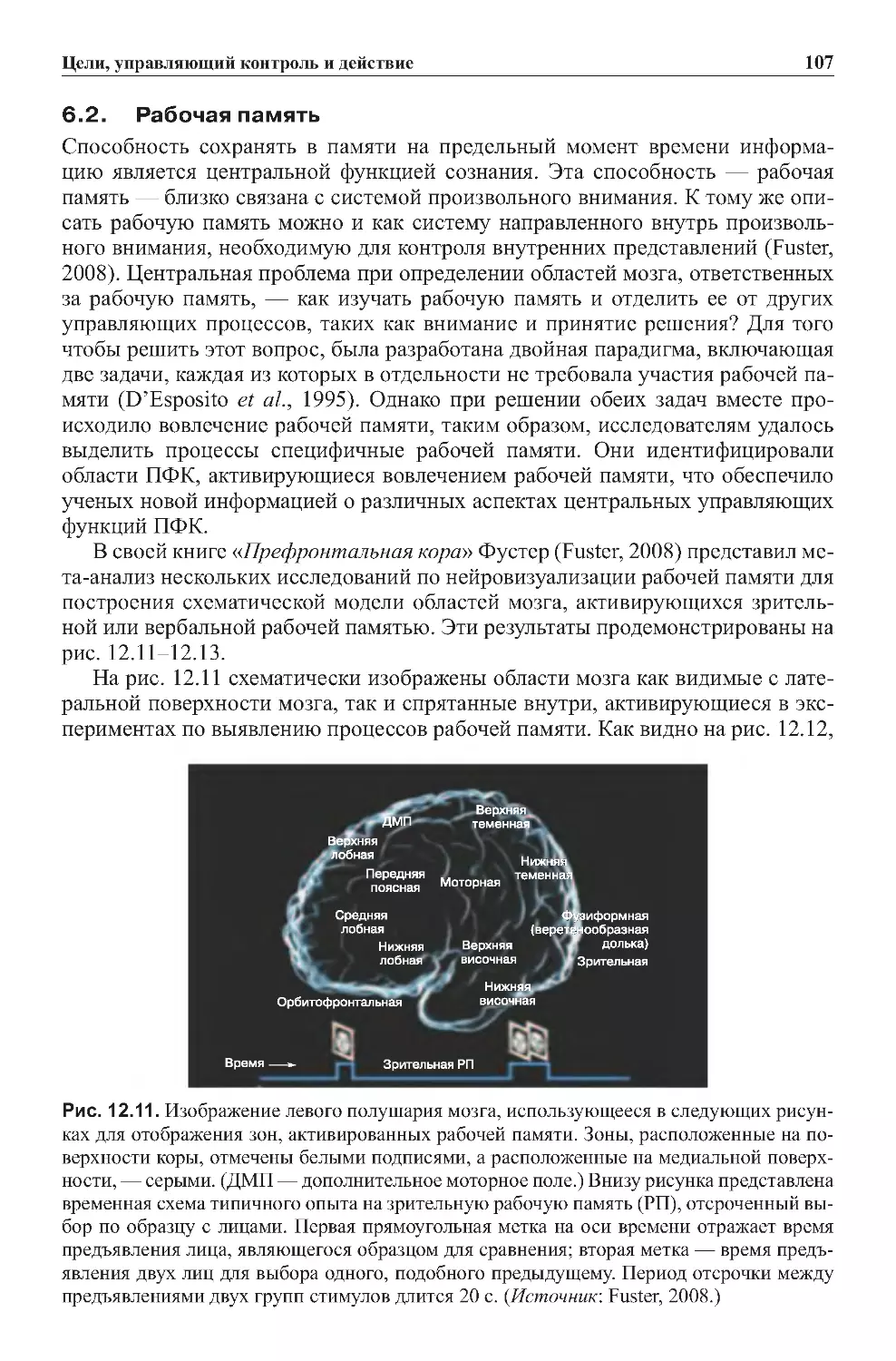
7.0. Консолидация кратковременных событий в долговременную память . .104
7.1. Оперативная память — результат активации постоянной памяти? . . . . 106
8.0.Заключение...................................................107
9.0.Заданияиупражнениякглаве2..................................108
9.1.Контрольныезадания.........................................108
9.2.Графическиеупражнения......................................109
Глава3.Нейроныисвязимеждуними.......................111
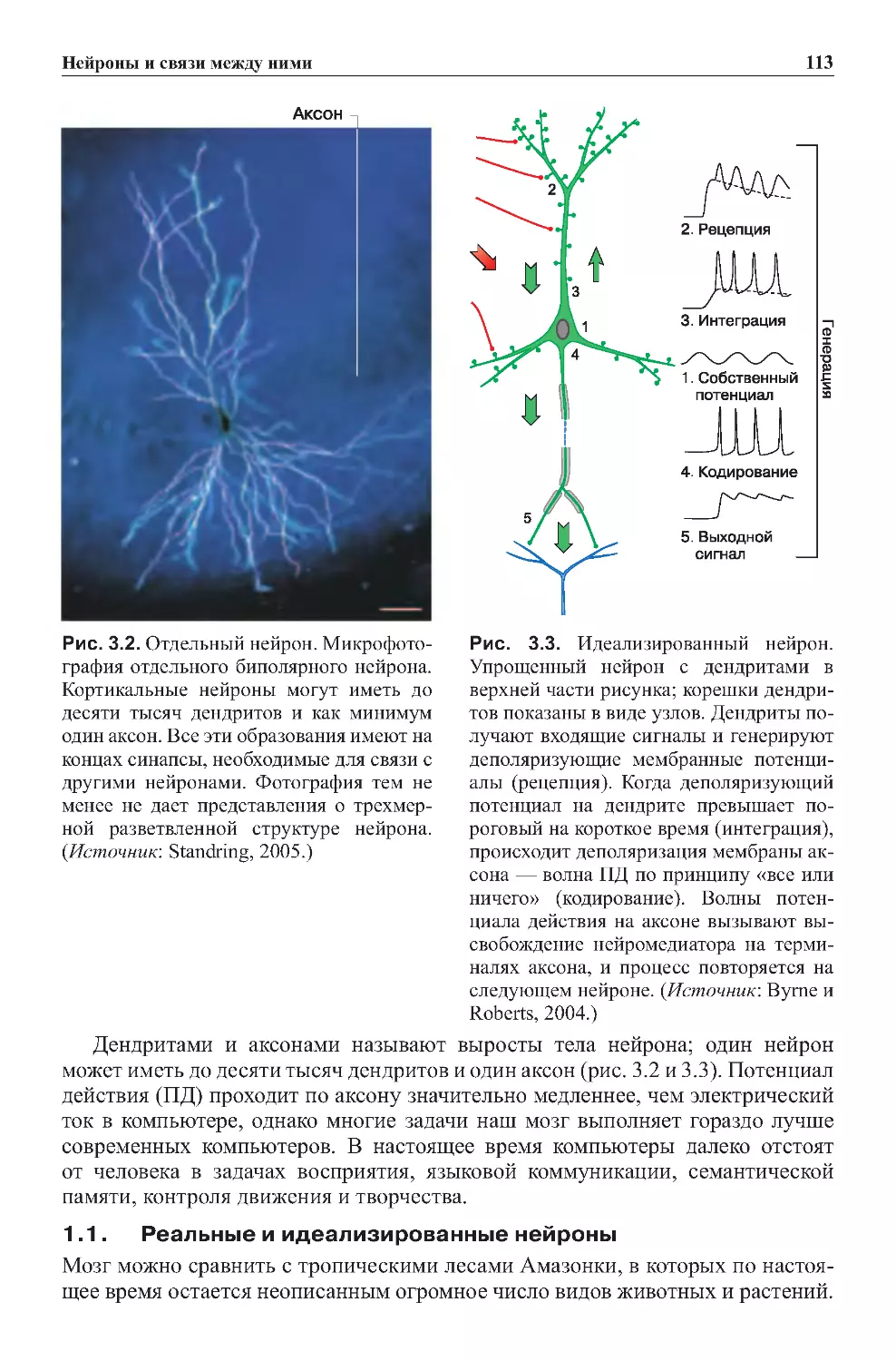
1.0.Введение....................................................111
1.1.Реальныеиидеализированныенейроны.........................113
1.2.Возбуждениеиторможение...................................115
1.3.Обработкаинформациинейронами.............................117
2.0.Рабочиегипотезы.............................................118
2.1.Упрощенныйслучай:рецепторы,путииконтуры..................119
Оглавление
7
3.0.Массивыикарты..............................................124
3.1.Картыпереходятвдругиекарты................................127
3.2. Массивы нейронов чаще всего
имеютсвязидвухтипов.......................................127
3.3.Сенсорныеимоторныесистемыработаютсовместно..............127
3.4. Временная кодировка:
рисункиспайковиритмымозга................................129
3.5.Точкивыборавпотокахинформации............................133
3.6.Обработка,обусловленнаяожиданием...........................135
4.0.Адаптацияиобучениемассивовнейронов.........................138
4.1. Обучение по Хэббу: «Активирующиеся одновременно нейроны
образуютсоединениямеждусобой».............................139
4.2. Дарвинистский подход в нервной системе:
выживают клетки и синапсы,
наиболееприспособленныекданнойзадаче......................143
4.3. Обработка символьной информации и нейронные сети . . . . . . . . . . . . . 145
5.0.Координацияработынейрональныхсетей.........................146
5.1.Функциональнаяизбыточность.................................148
6.0.Заключение...................................................149
7.0.Заданияиупражнениякглаве3..................................150
7.1.Контрольныезадания.........................................150
7.2.Графическиеупражнения......................................150
Глава4.Методы:визуализацияживогомозга.................153
1.0.Введение....................................................153
1.1. Регистрация работы мозга: более и менее прямые
методыизмерения............................................155
1.2. Соотношение пространственного и временного
разрешенийметодов..........................................155
2.0 . Ряд полезных инструментов для измерения электрической
имагнитнойактивности........................................159
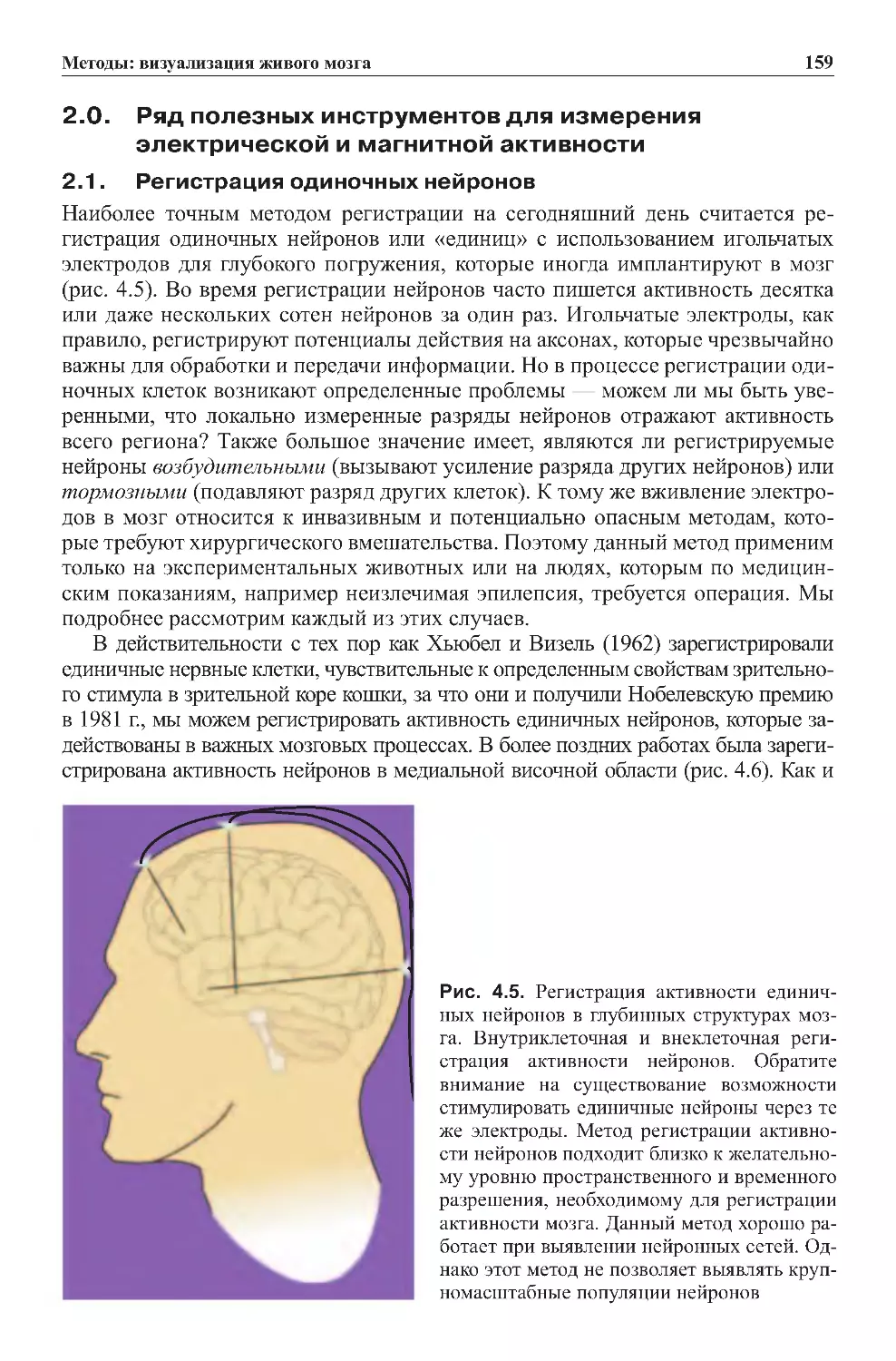
2.1.Регистрацияодиночныхнейронов..............................159
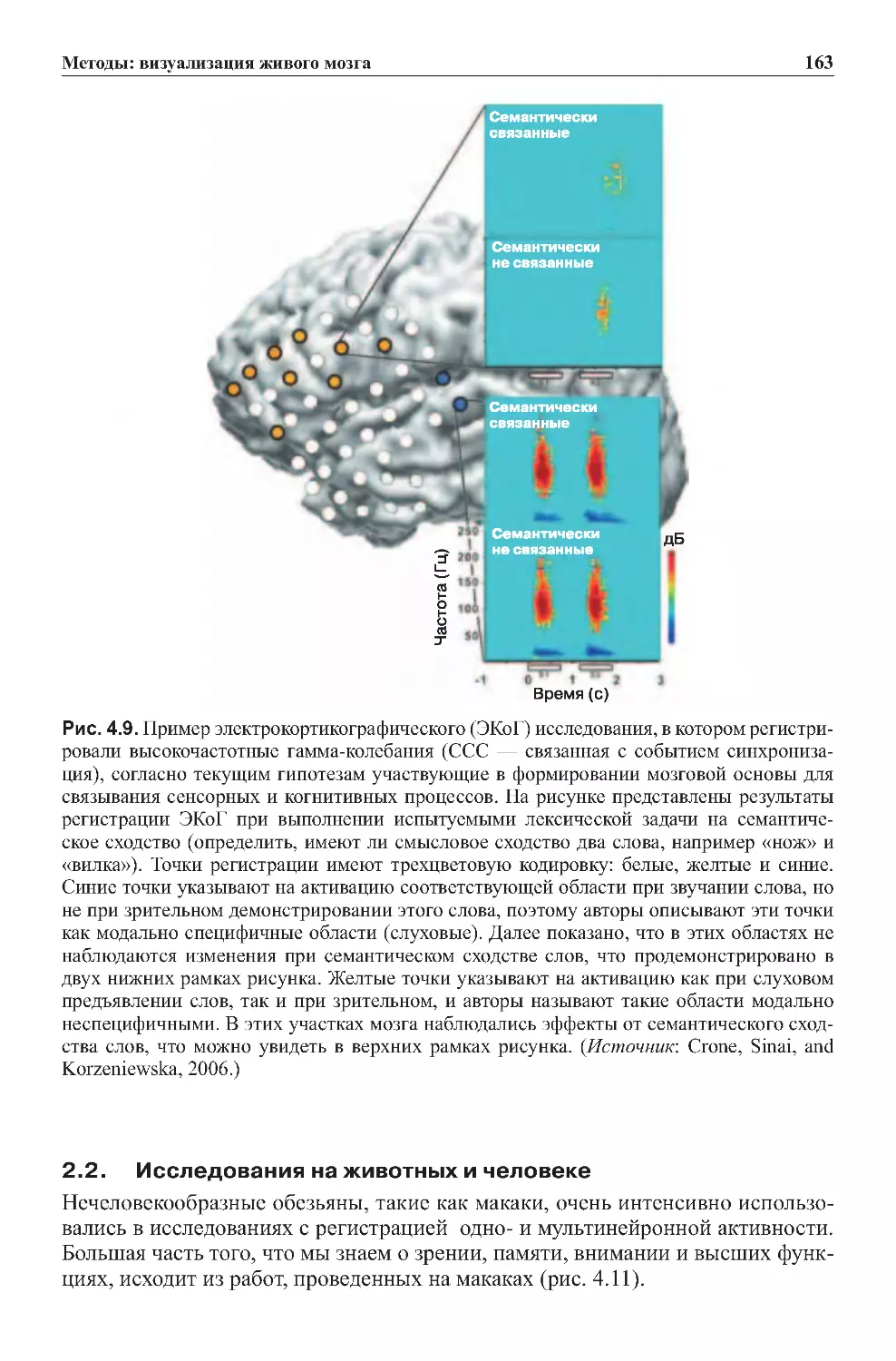
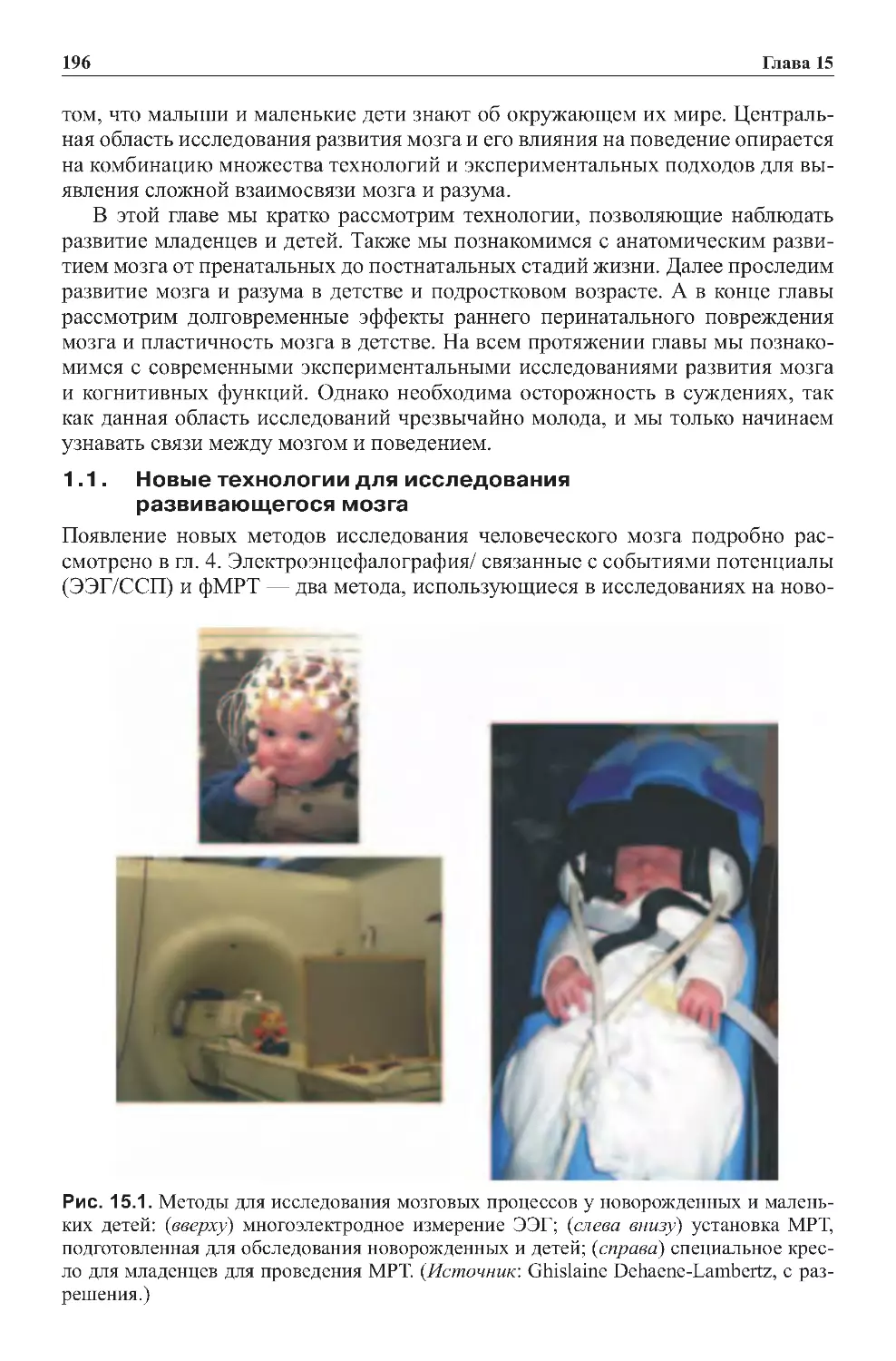
2.2.Исследованиянаживотныхичеловеке..........................163
2.3.Электроэнцефалография(ЭЭГ).................................165
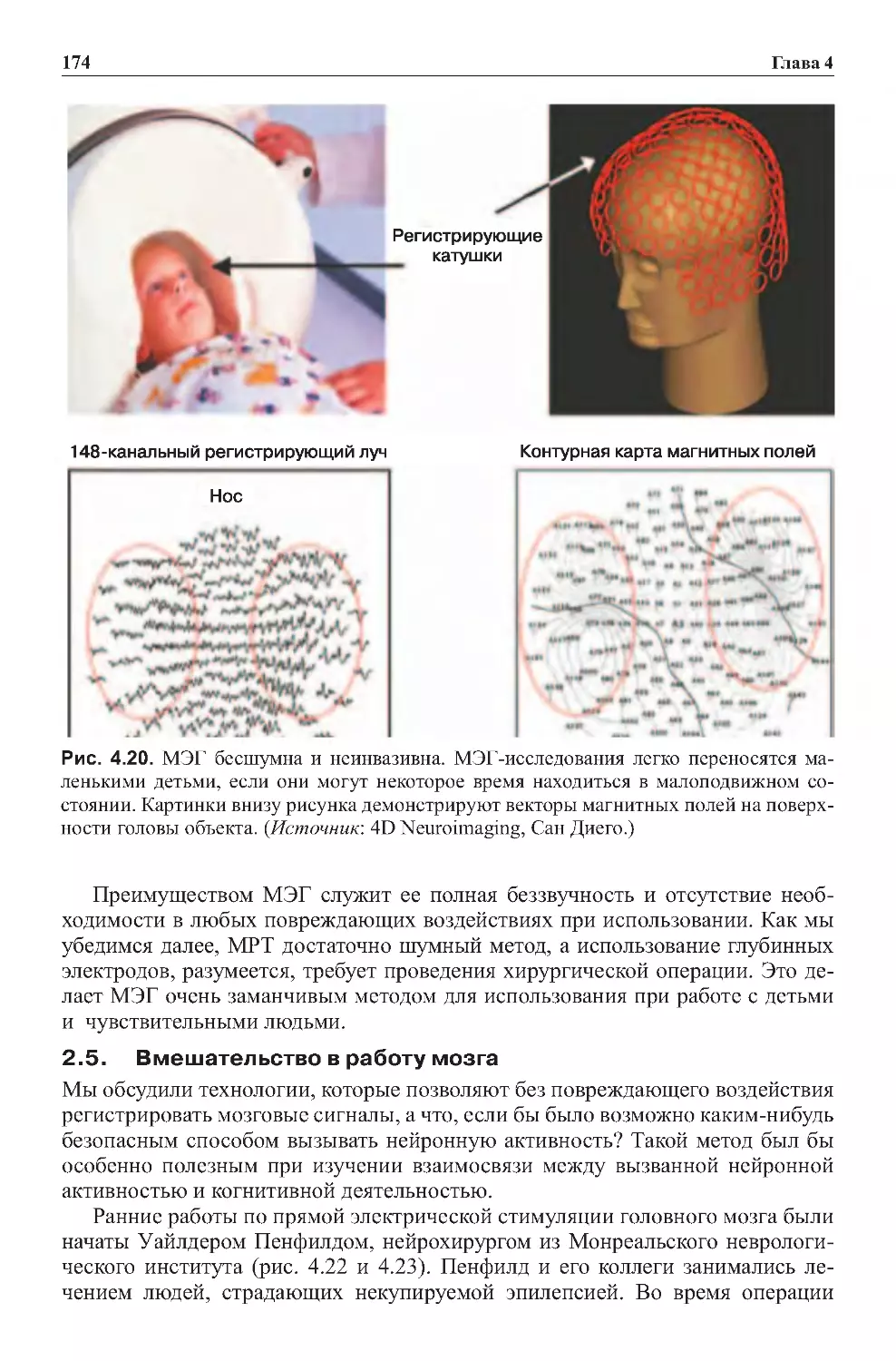
2.4.Магнитоэнцефалография......................................173
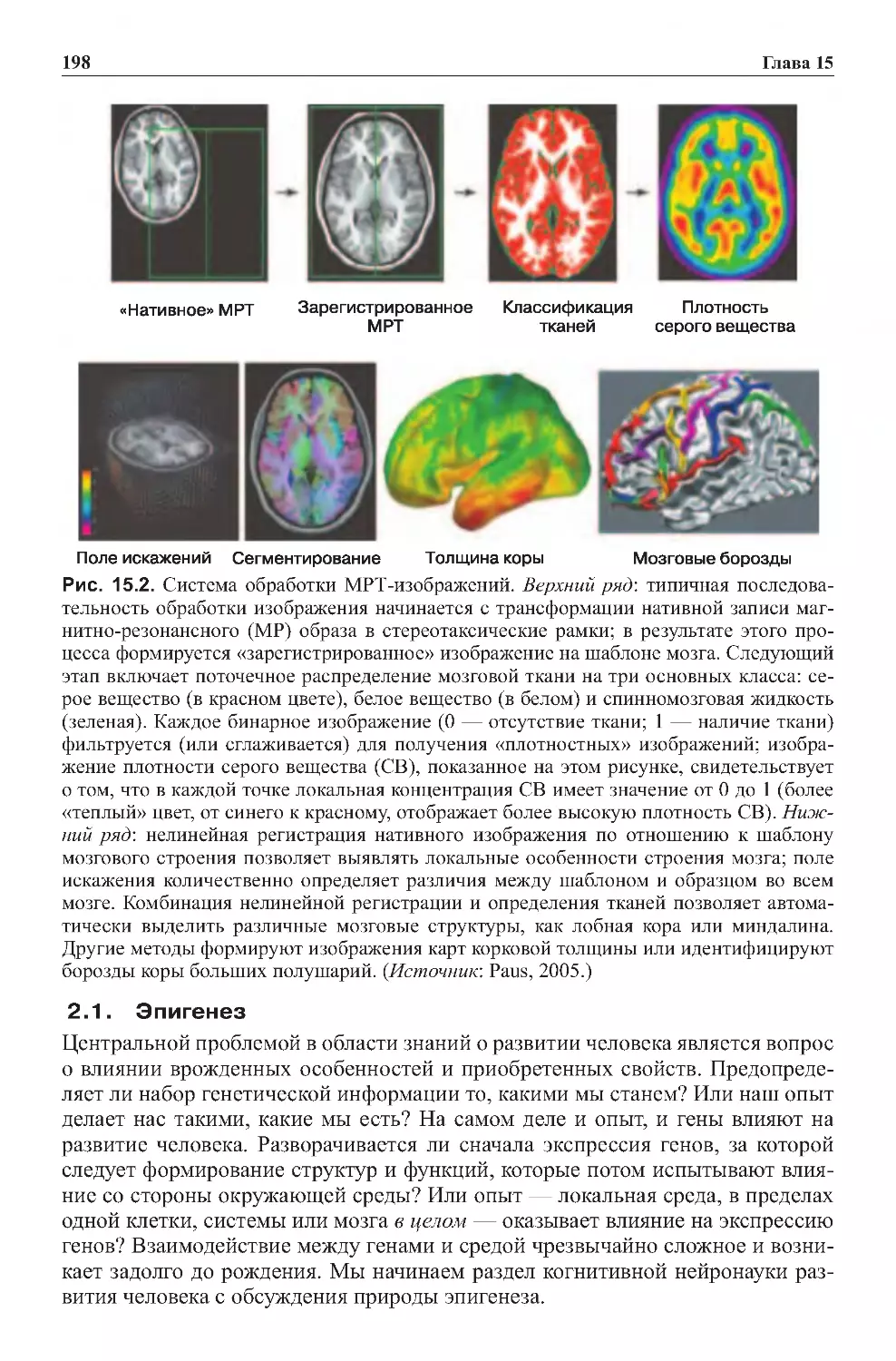
2.5.Вмешательствовработумозга.................................174
3.0. фМРТ И ПЭТ: непрямое измерение нейронной активности . . . . . . . . . .180
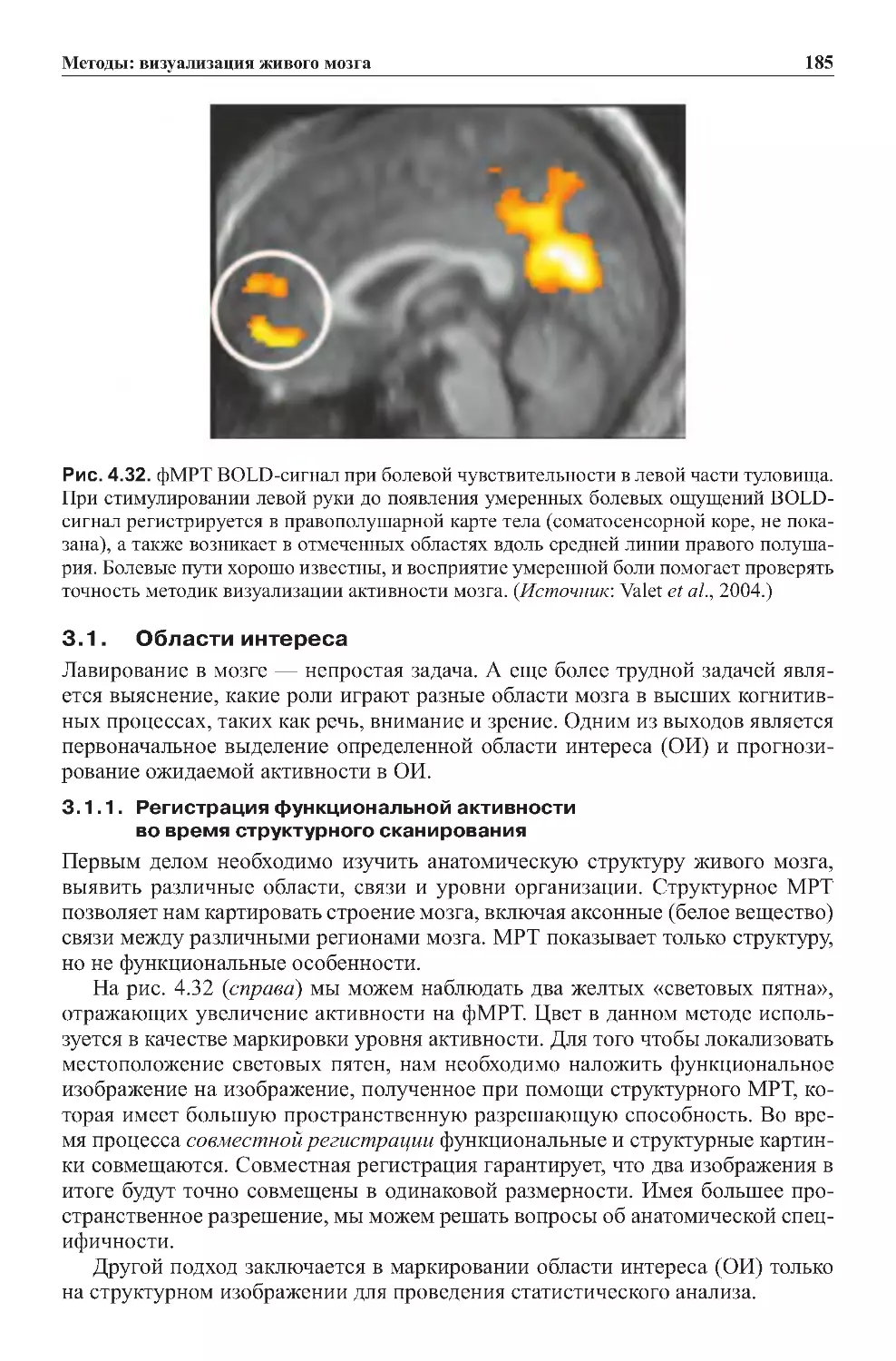
3.1.Областиинтереса............................................185
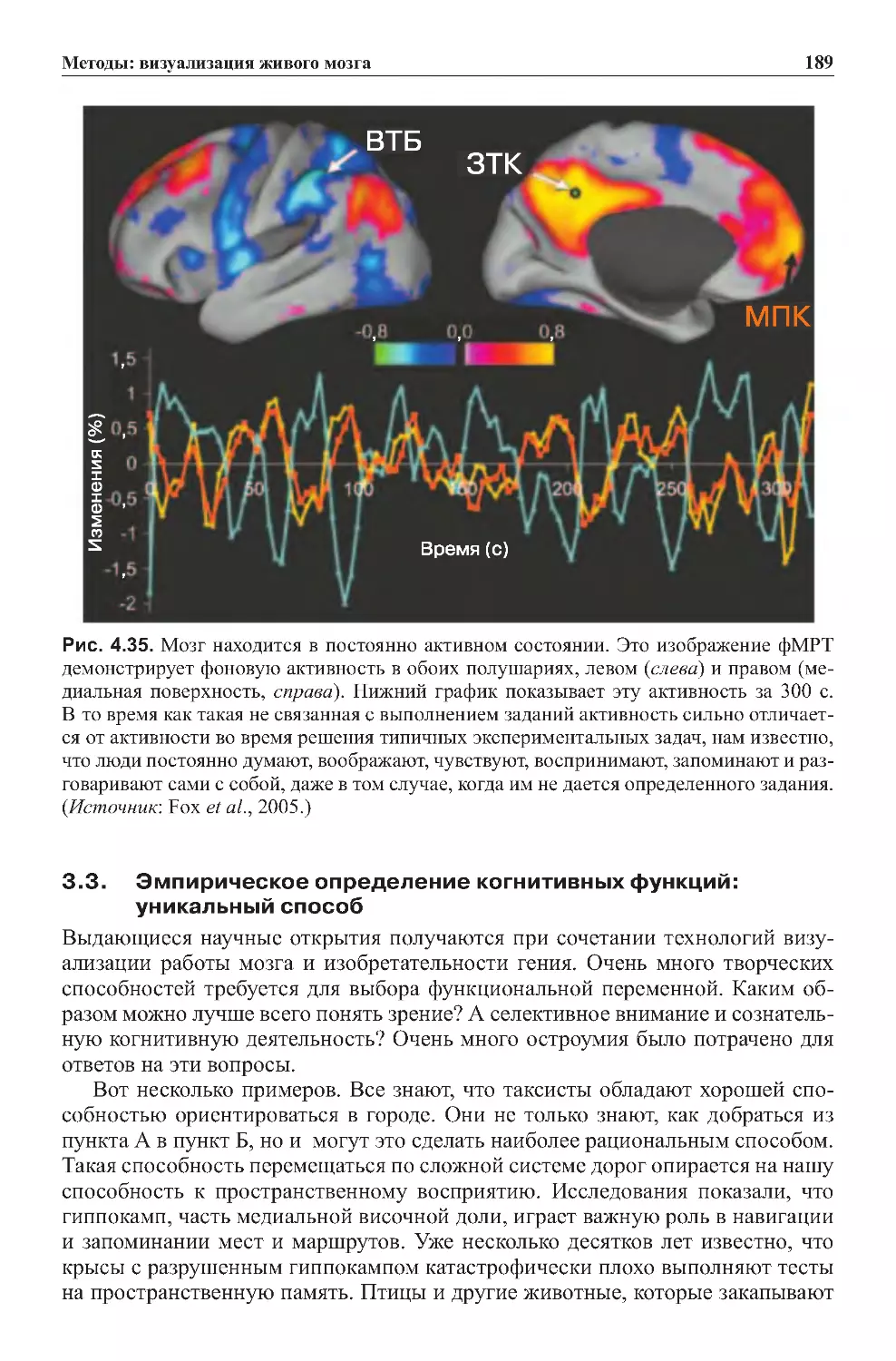
3.2. Мозг в состоянии покоя не молчит:
внутренниепроцессымозга....................................188
8
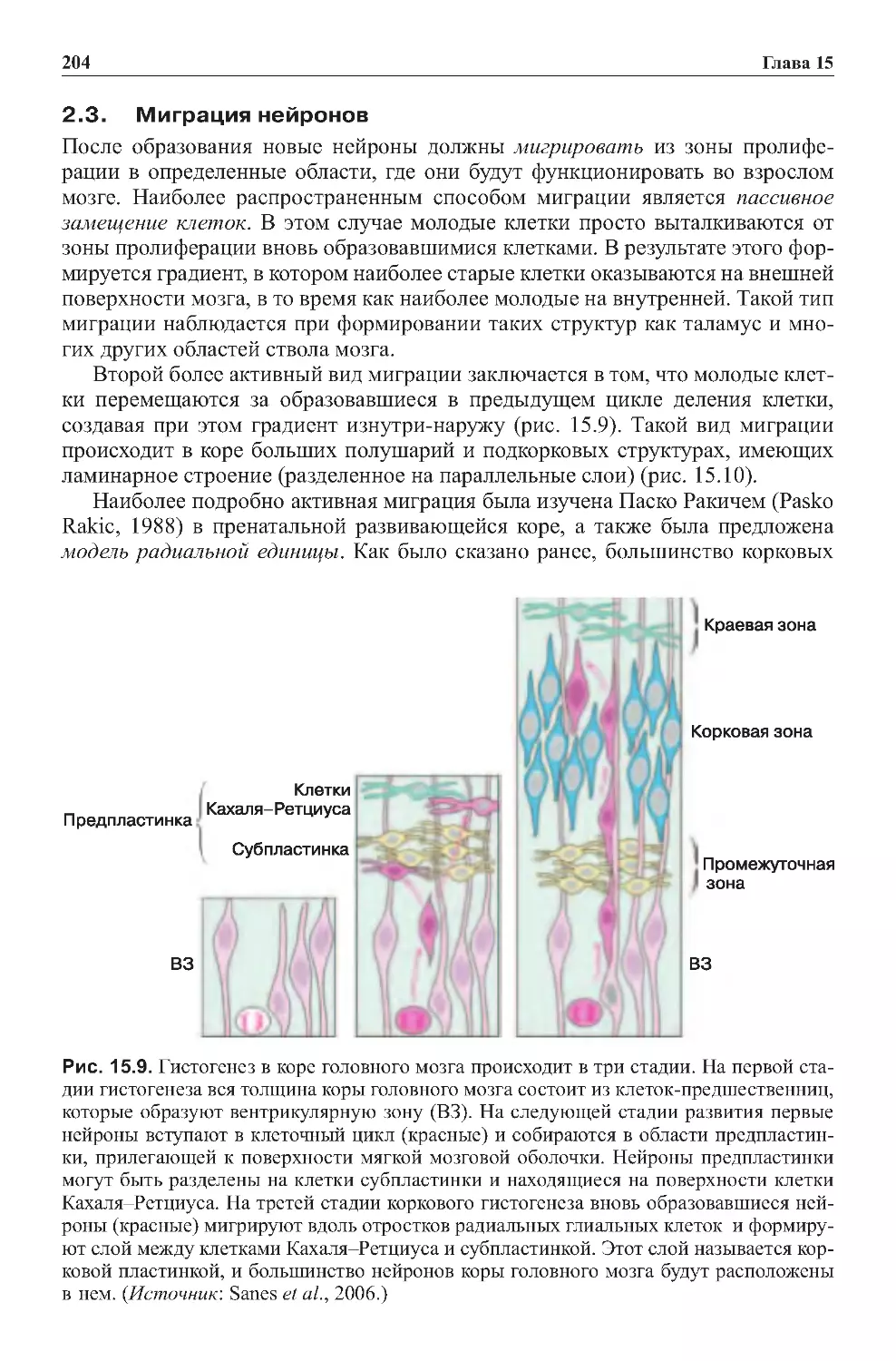
Оглавление
3.3. Эмпирическое определение когнитивных функций:
уникальныйспособ...........................................189
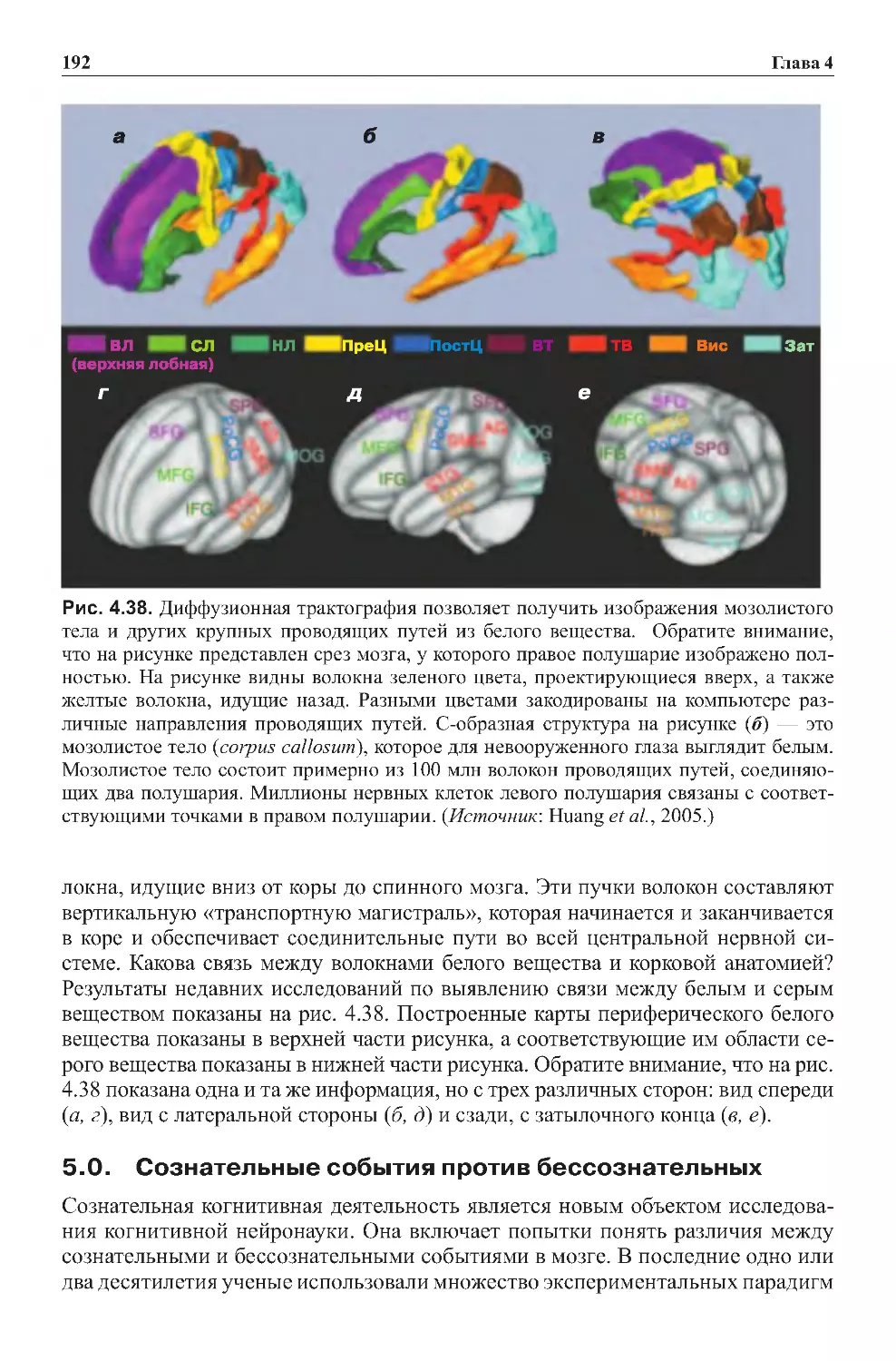
4.0. Новые способы определения взаимосвязи областей мозга:
визуализацияраспределениятензора.............................191
5.0.Сознательныесобытияпротивбессознательных....................192
6.0.Корреляцияипричиннаясвязь..................................193
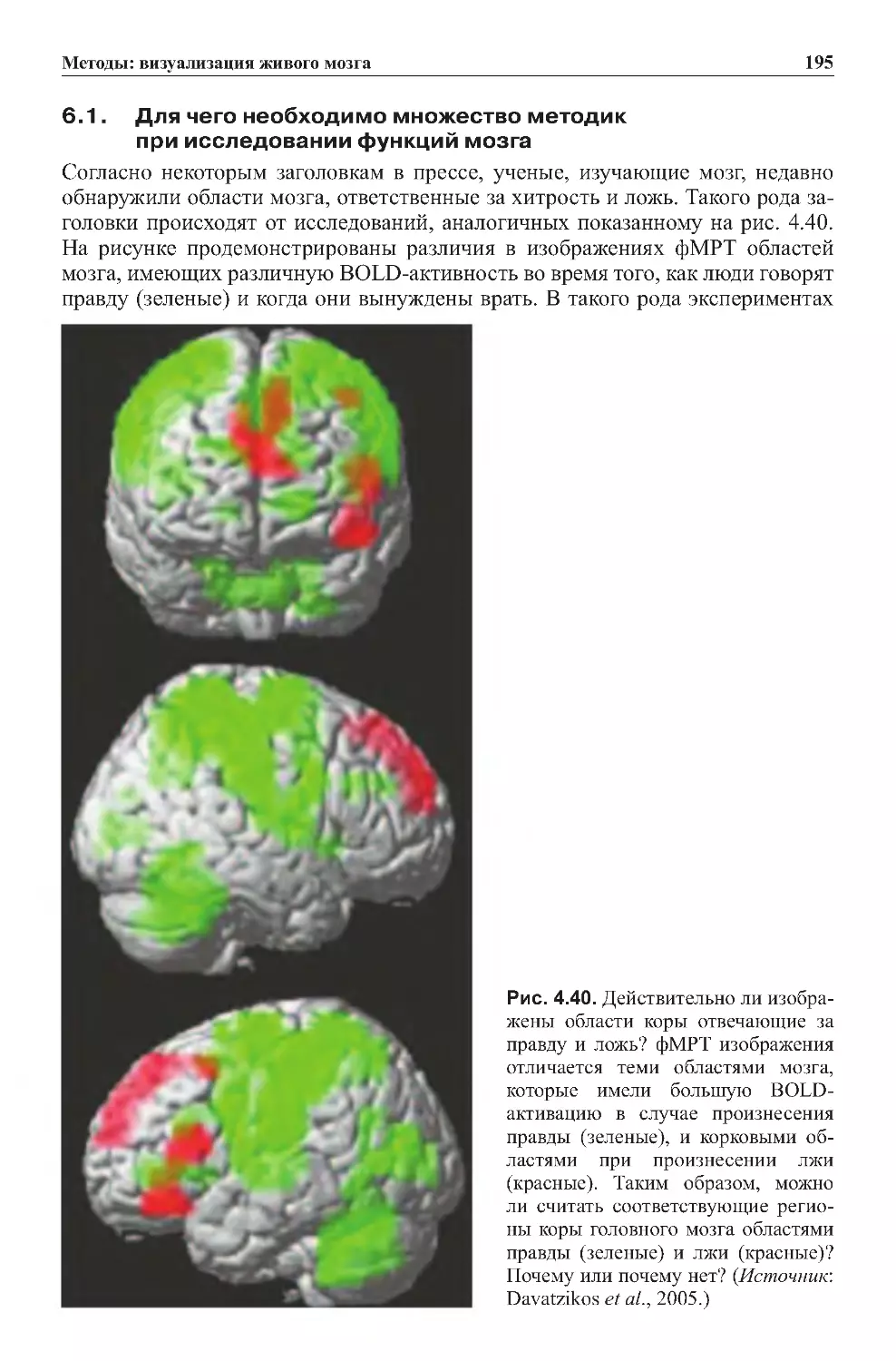
6.1. Для чего необходимо множество методик
приисследованиифункциймозга...............................195
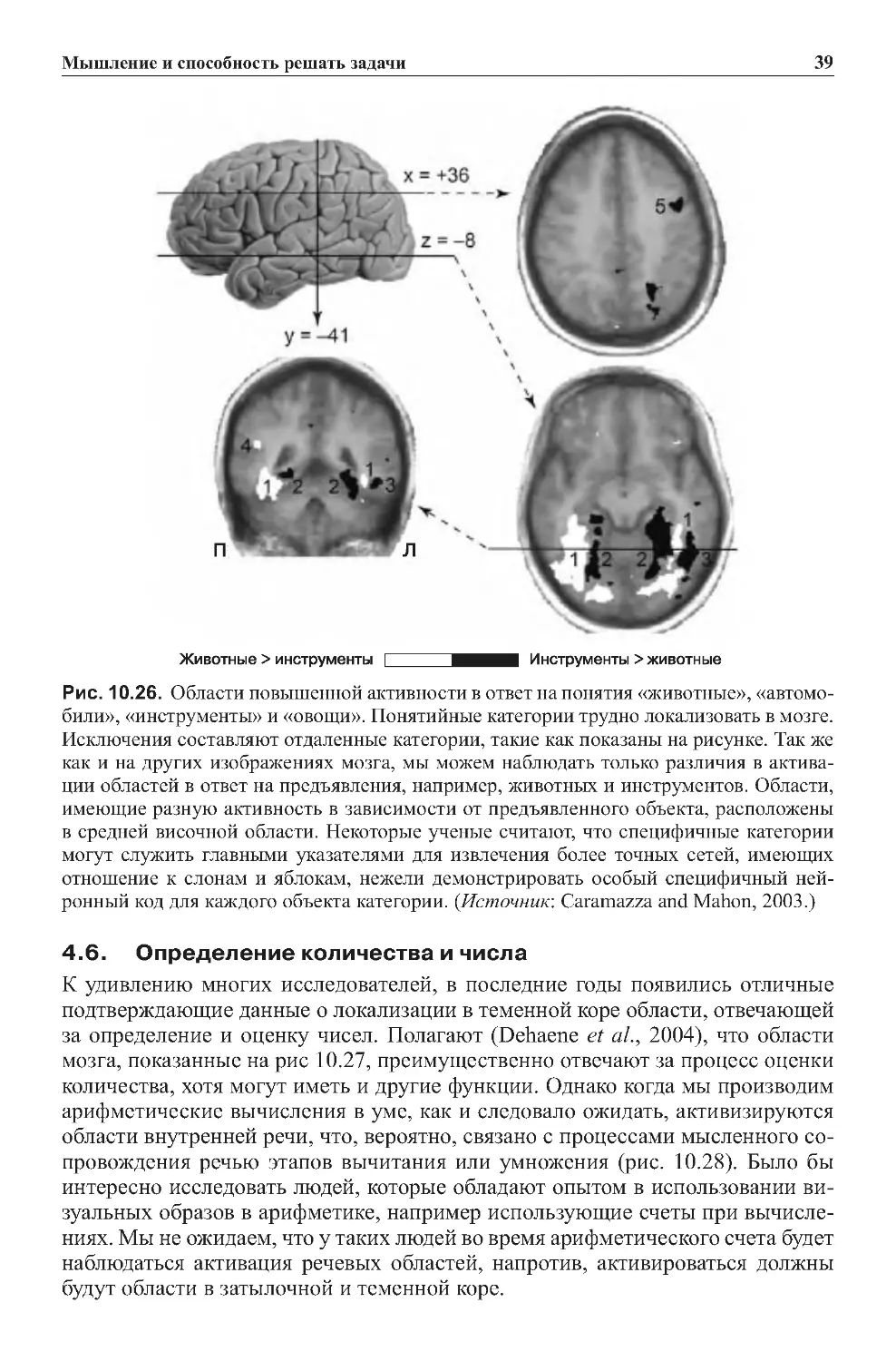
6.2.Повреждениямозгаиследствияэтихповреждений................196
7.0.Заключение...................................................198
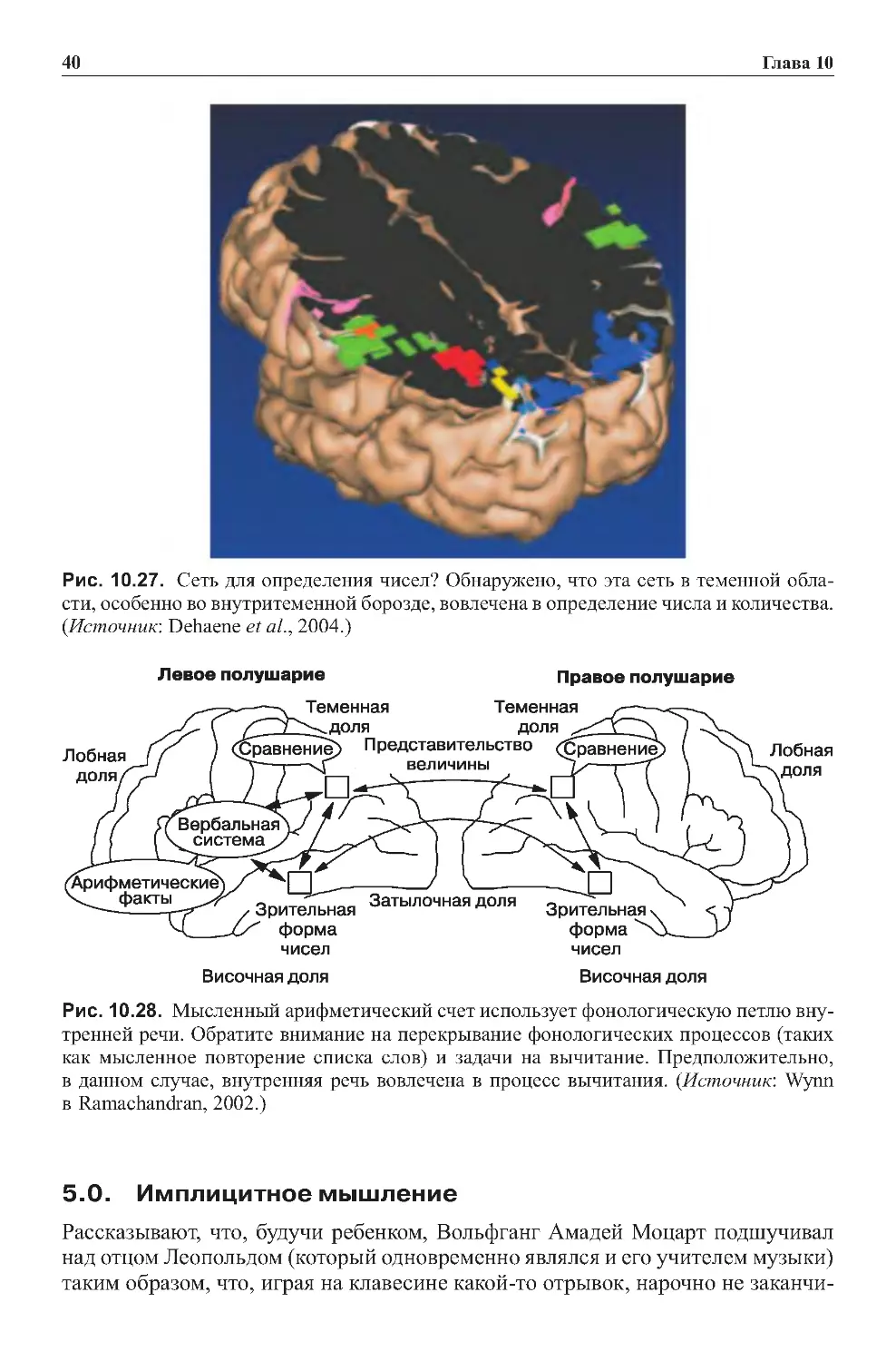
8.0.Заданияиупражнениякглаве4 .................................198
8.1.Контрольныезаданияиграфическиеупражнения.................198
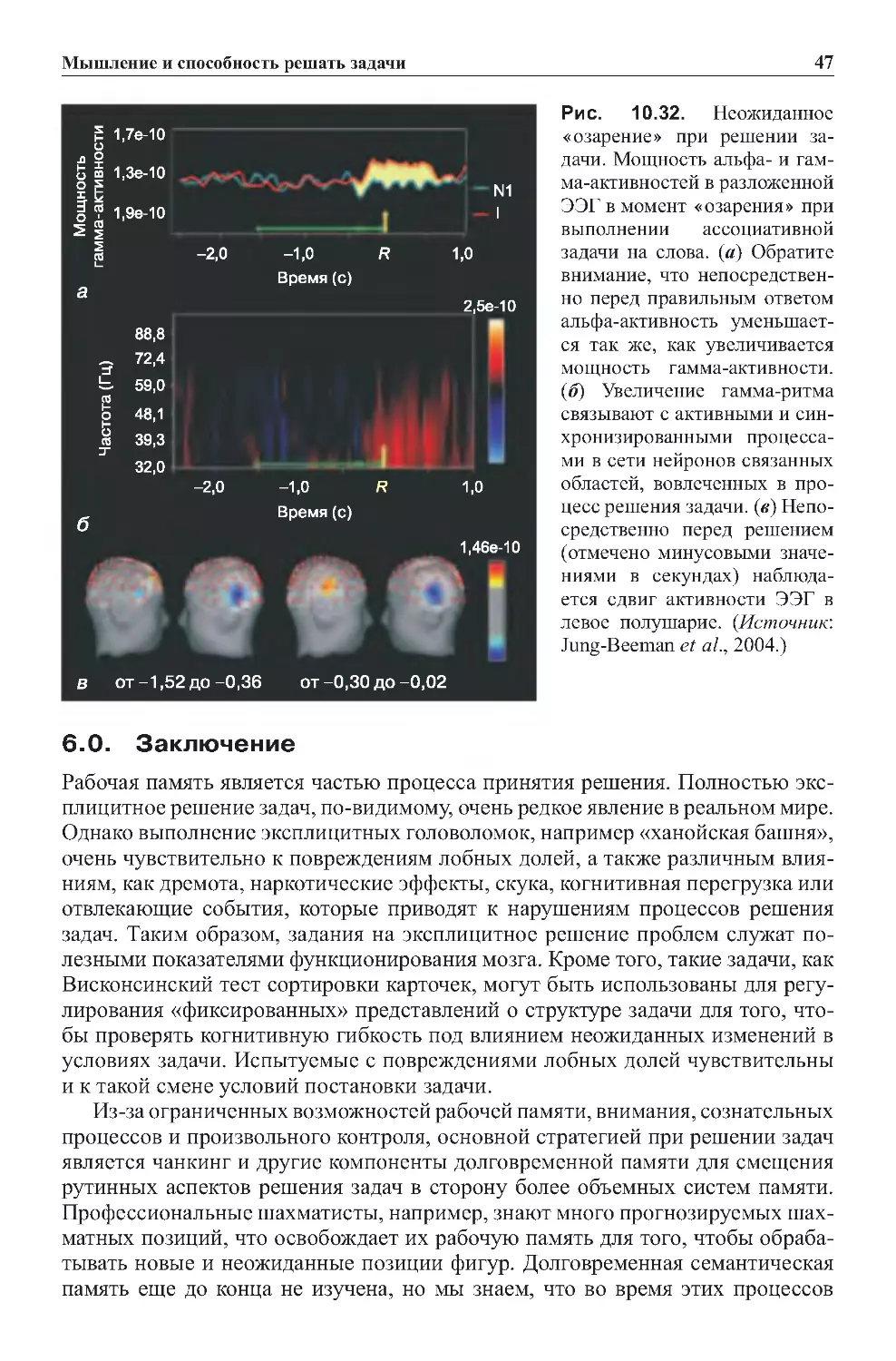
Глава5.Мозг..............................................201
1.0.Введение.....................................................201
1.1.Нервнаясистема.............................................202
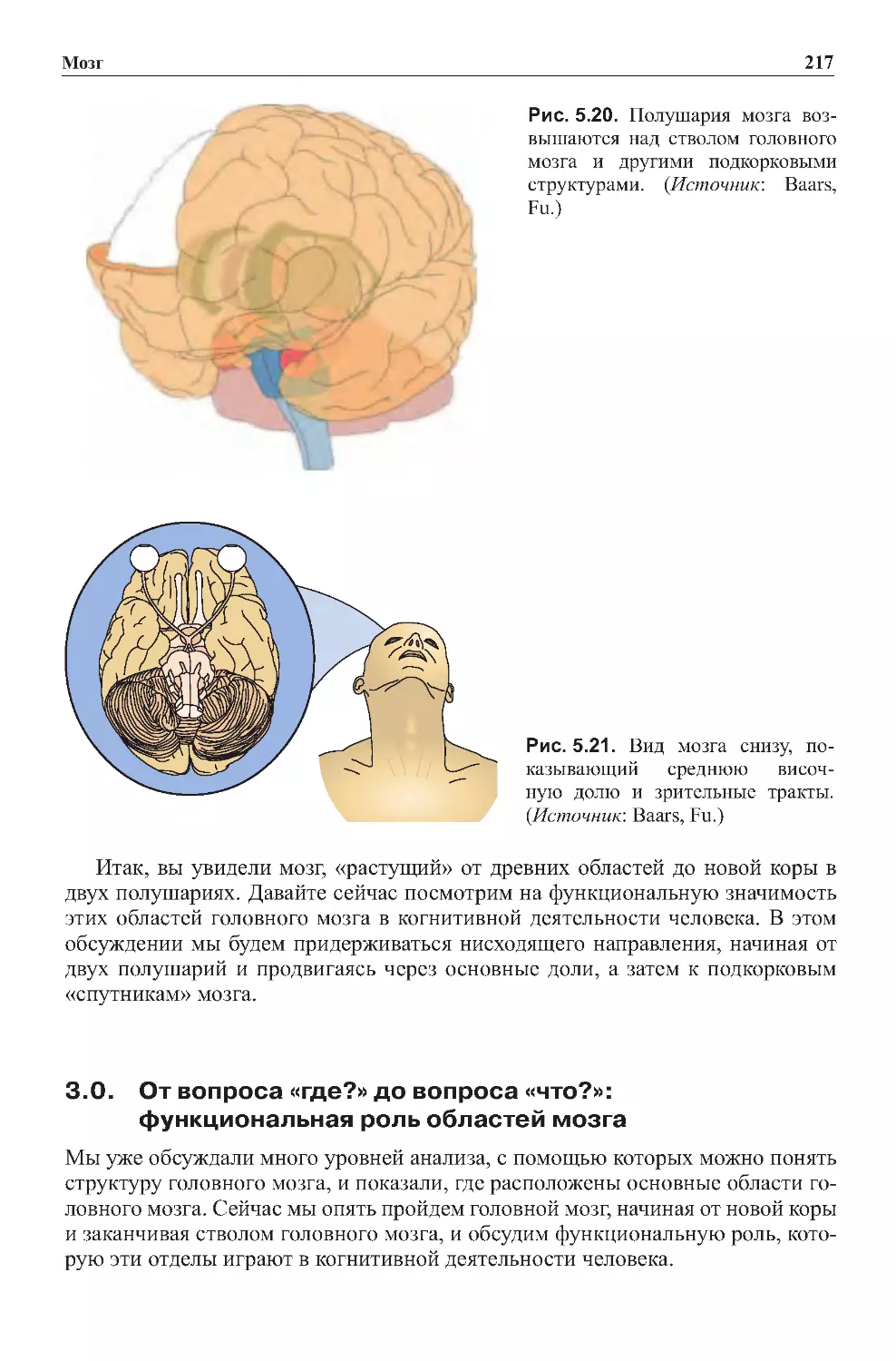
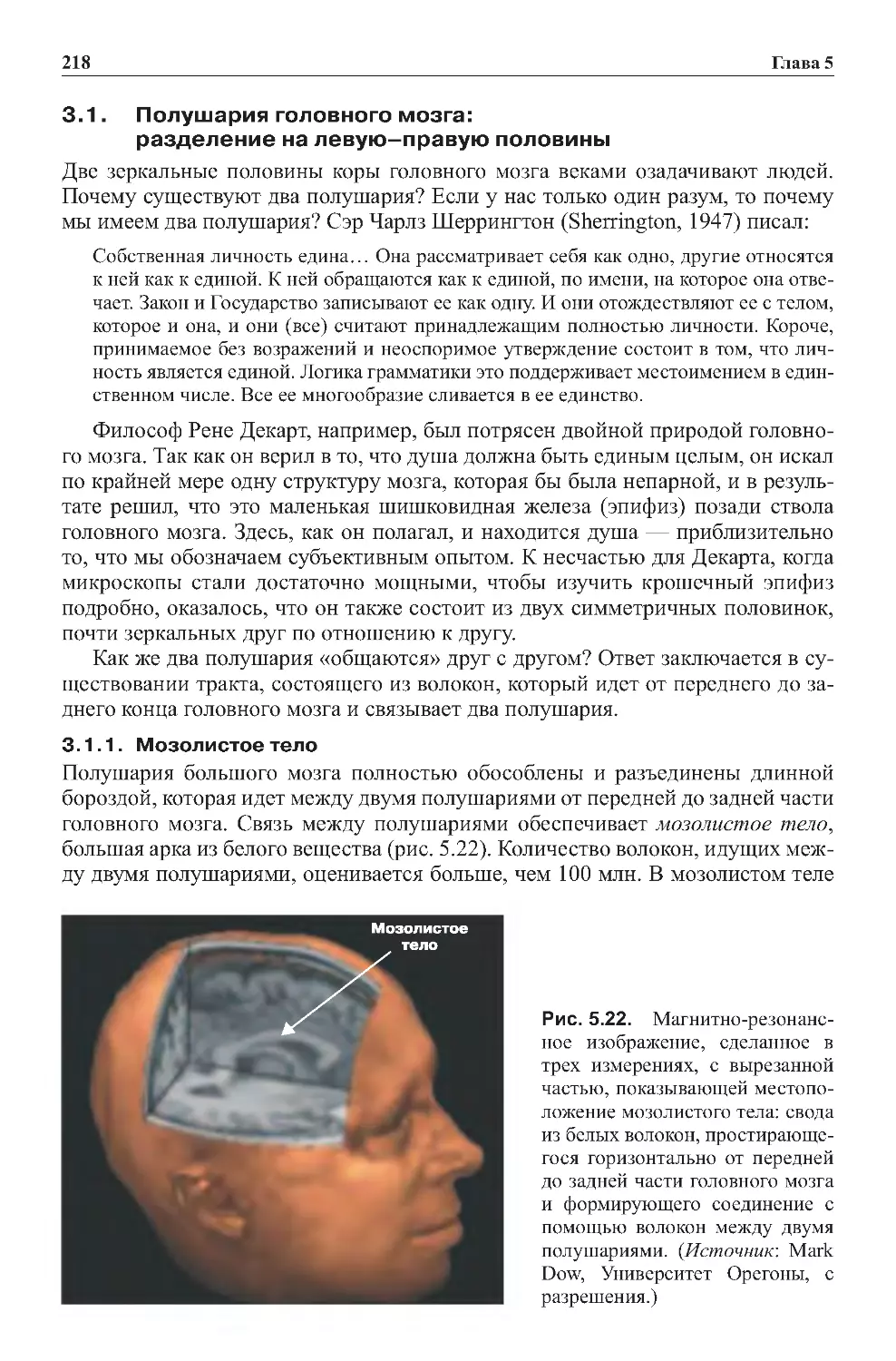
1.2.Географиямозга.............................................203
2.0.Развитиемозганачинаетсяснижнихотделов......................210
2.1. В строении мозга находят отражение эволюция
ииндивидуальноеразвитие ...................................210
2.2.Развитиемозгаотоснованиядовершины........................210
3.0. От вопроса «гд е?» до вопроса «что?»: функциональная роль
областеймозга................................................217
3.1. Полушария головного мозга:
разделениеналевую–правуюполовины..........................218
3.2. Исходящие и входящие сигналы:
подразделениевнаправлениисперединазад......................222
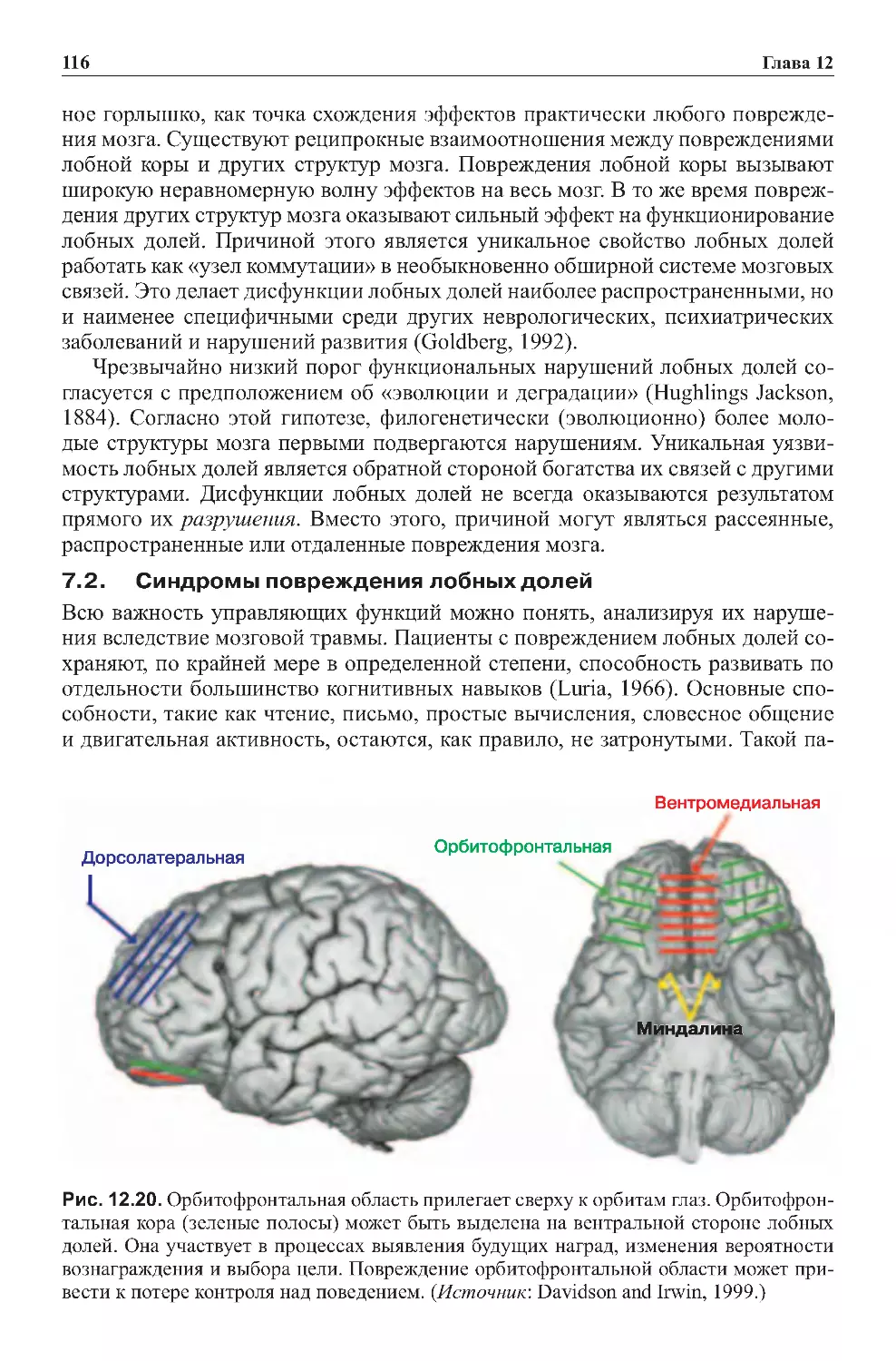
3.3.Основныедоли:видимыеискрытыеучастки.....................226
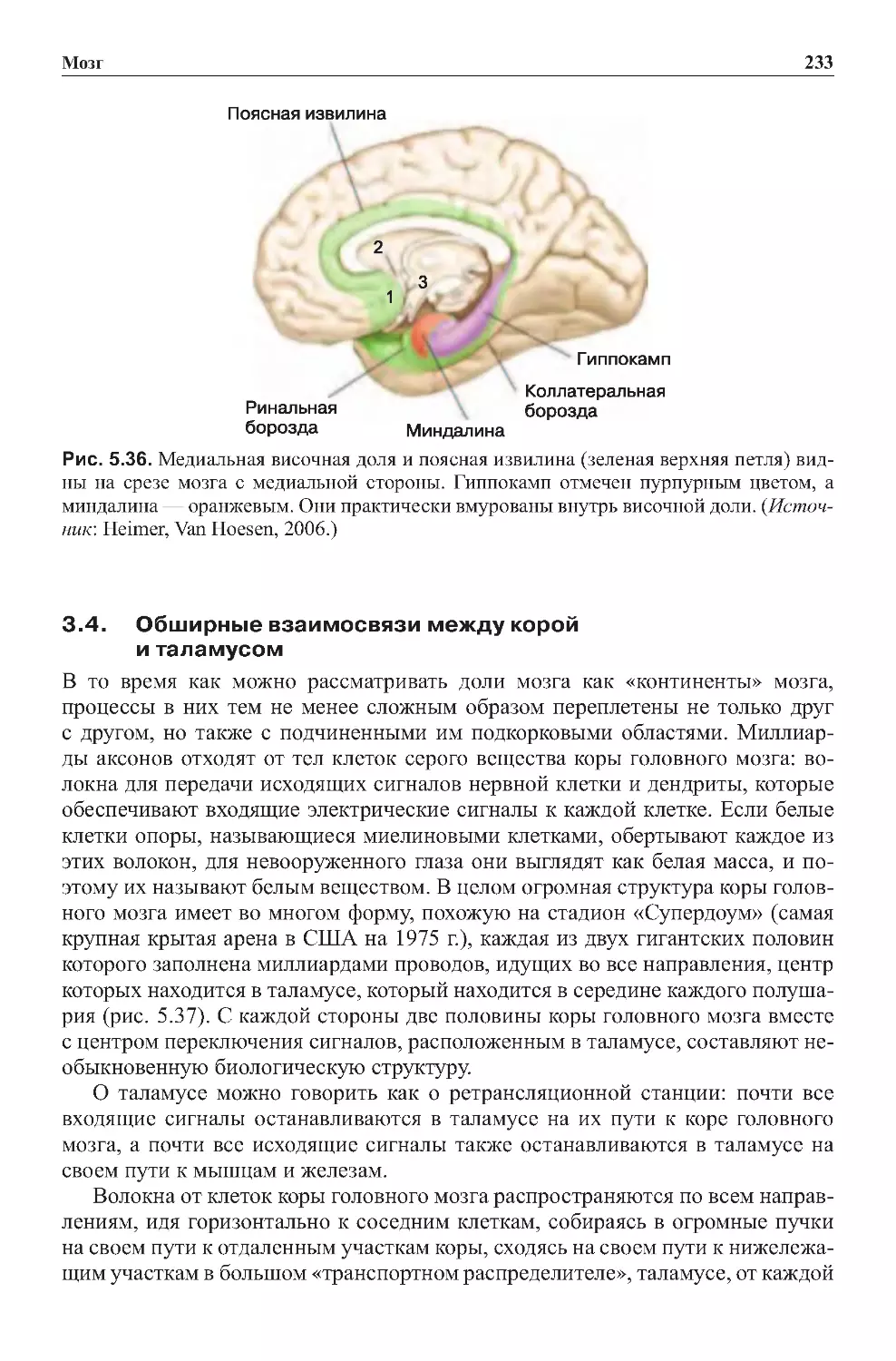
3.4.Обширныевзаимосвязимеждукоройиталамусом ................233
3.5.Подчиненныеучасткиподкорковыхслоев........................236
4.0.Заключение...................................................239
5.0.Заданияиупражнениякглаве5..................................240
5.1.Контрольныезадания.........................................240
5.2.Графическиеупражнения .....................................240
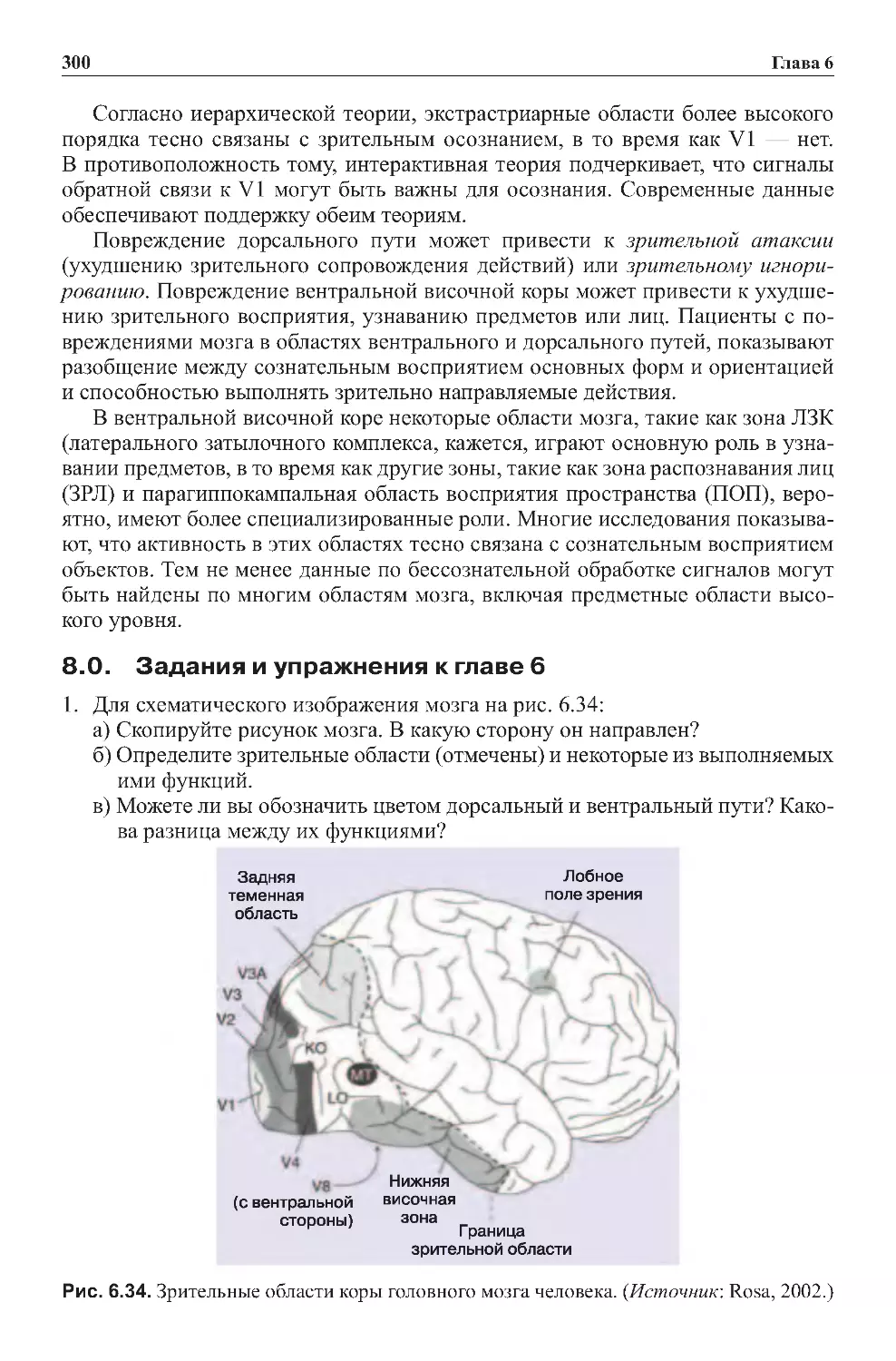
Глава6.Зрение...........................................243
1.0.Введение.....................................................244
1.1.Тайнаопытазрительныхвпечатлений...........................244
1.2.Цельзрения:знаниетого,чтогденаходится......................245
Оглавление
9
1.3. Знание об объекте:
восприятиехарактерныхчерт,группиобъектов..................245
1.4.Знаниетого,гденаходятсяобъекты.............................248
2.0. Функциональная организация зрительной системы . . . . . . . . . . . . . . . . .248
2.1.Сетчатка....................................................248
2.2.Латеральноеколенчатоетело..................................252
2.3.Первичная,илистриарная,зрительнаякора......................254
2.4. Экстрастриарные зрительные зоны — за пределами зоны V1 . . . . . . . 258
2.5.ПолеMT....................................................258
2.6. Вентральныйидорсальныйпути:знания«что»и«где»............258
2.7.Зоны,вовлеченныевраспознаваниеобъектов....................262
2.8.Латеральныйзатылочныйкомплекс ............................262
2.9.Зонараспознаваниялиц ......................................264
2.10. Парагиппокампальная область восприятия пространства . . . . . . . . . . 264
3.0. Теории зрительного осознания: где оно происходит? . . . . . . . . . . . . . . . .264
3.1.Иерархическаяиинтерактивнаятеориизрения...................268
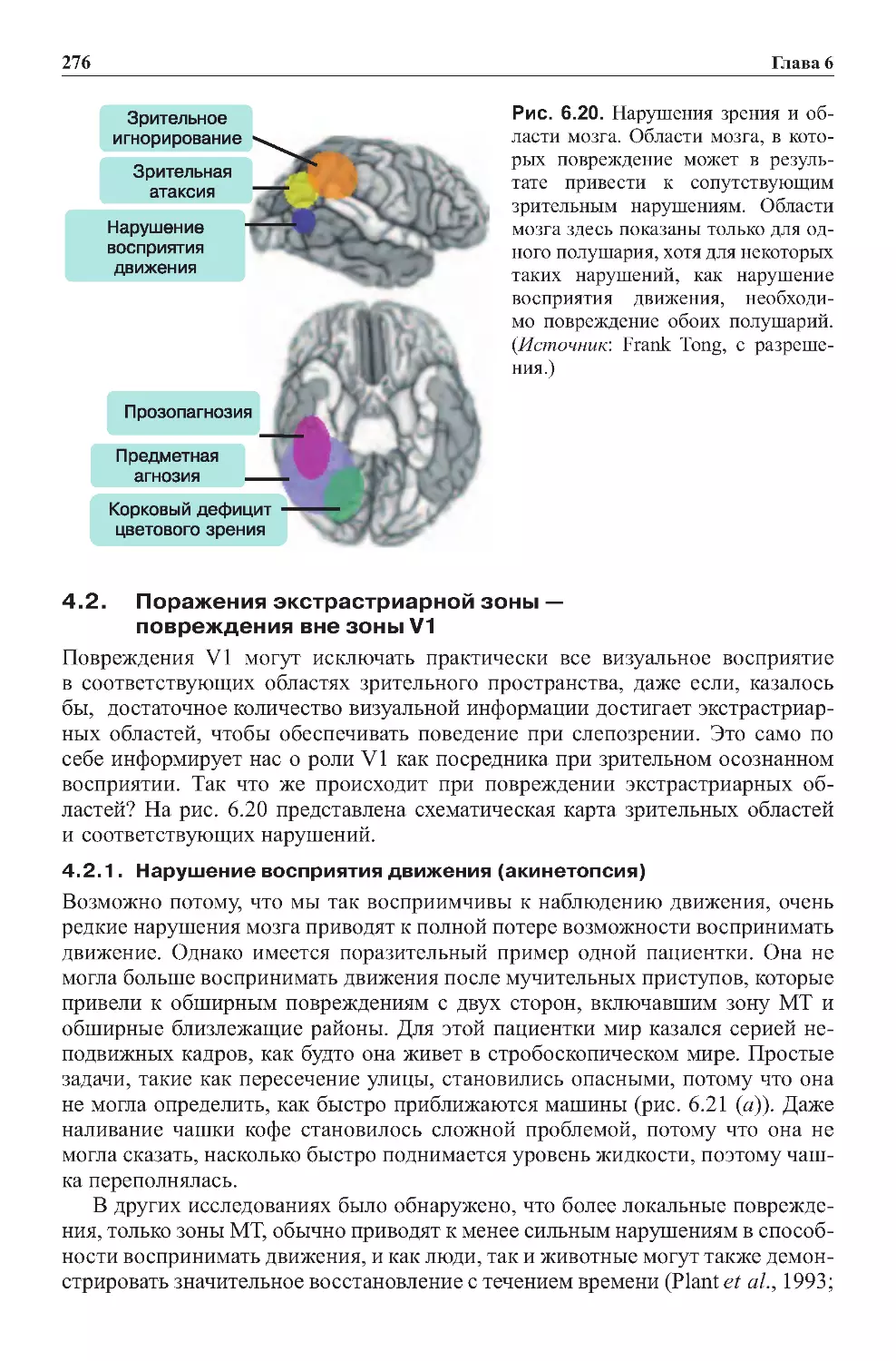
4.0. Области мозга, необходимые для визуального восприятия:
исследованияповреждений.....................................273
4.1. Последствия повреждений первичных зрительных зон . . . . . . . . . . . . . 273
4.2. Поражения экстрастриарной зоны —
повреждениявнезоныV1.....................................276
4.3. Повреждение вентральных областей,
воспринимающихпредметы...................................277
4.4. Повреждениедорсальныхобластейтеменнойдоли................282
5.0.Связьдеятельностимозгаизрительногоопыта.....................283
5.1. Восприятиесомногимиустойчивымисостояниями...............284
5.2. Бинокулярная конкуренция: то, что вы видите, —
этото,чтостимулируетвас....................................285
5.3.Зрительноеобнаружение:виделиливыэто?.....................288
5.4. Созидательное восприятие: видеть больше, чем доступно глазу... . . . 289
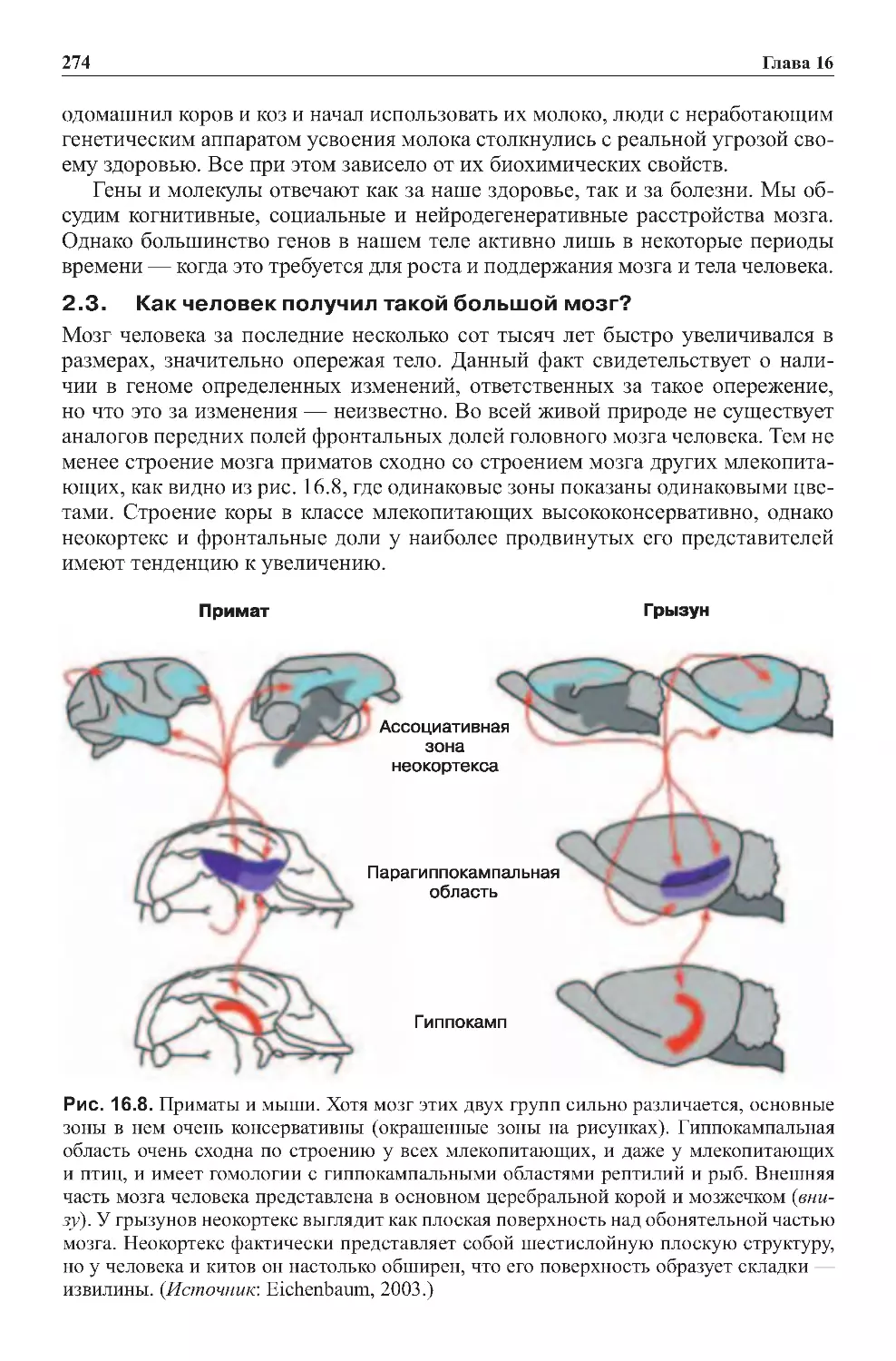
5.5.Нервныекорреляциисраспознаваниемобъектов..................291
6.0.Управлениезрительнымосознанием..............................292
6.1.Транскраниальнаямагнитнаястимуляция .......................293
6.2.Бессознательноевосприятие...................................296
7.0.Заключение...................................................299
8.0.Заданияиупражнениякглаве6 .................................300
Глава7.Слухиречь.......................................303
1.0.Введение.....................................................303
1.1.Модельобработкиинформации.................................304
10
Оглавление
1.2.Основызвукаислуховоговосприятия...........................307
2.0.Центральнаяслуховаясистема..................................312
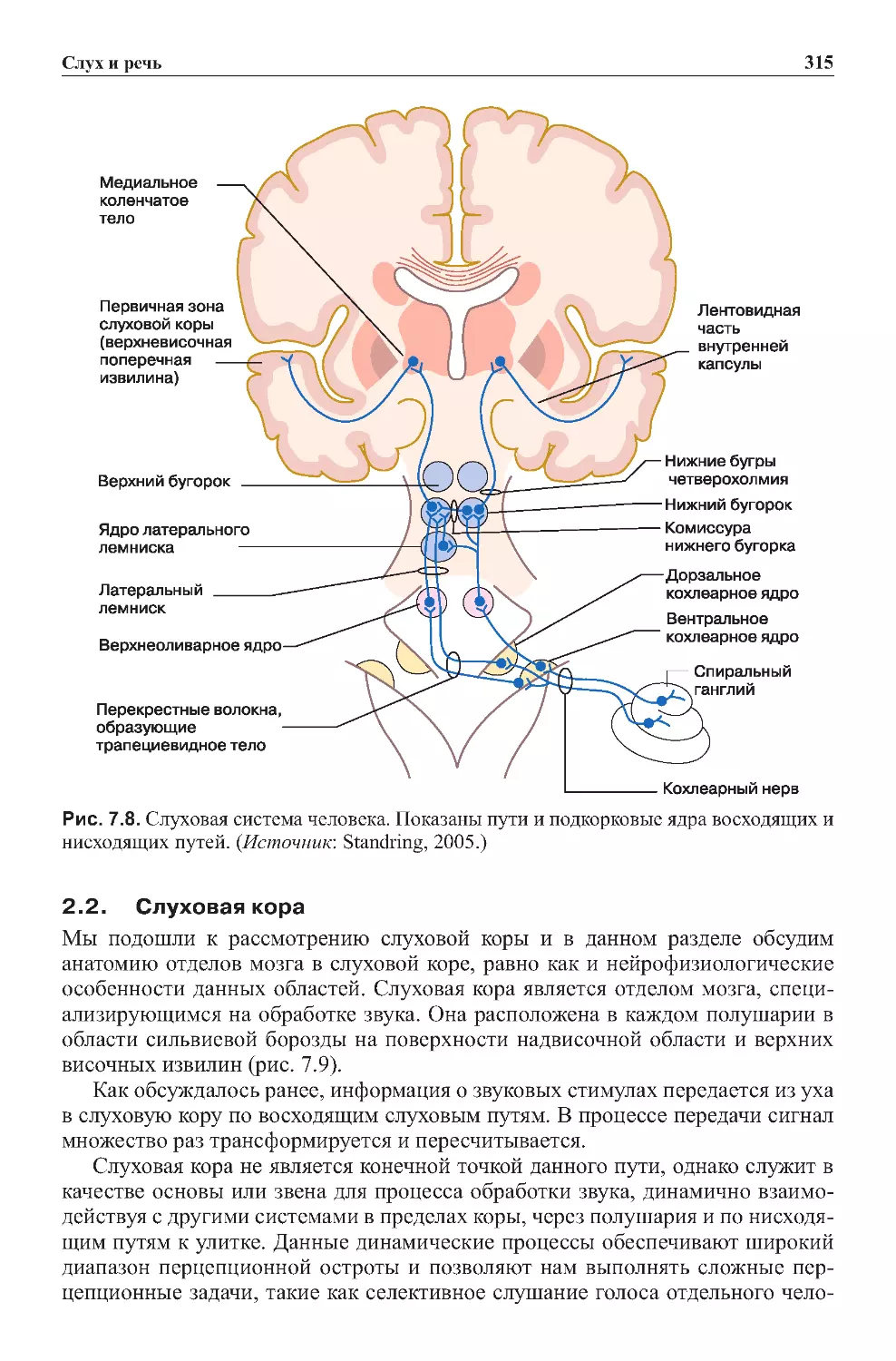
2.1.Путипередачислуховойинформации...........................312
2.2.Слуховаякора...............................................315
3.0. Функциональное картирование обработки
слуховойинформации..........................................323
3.1.Первичнаяслуховаякора......................................323
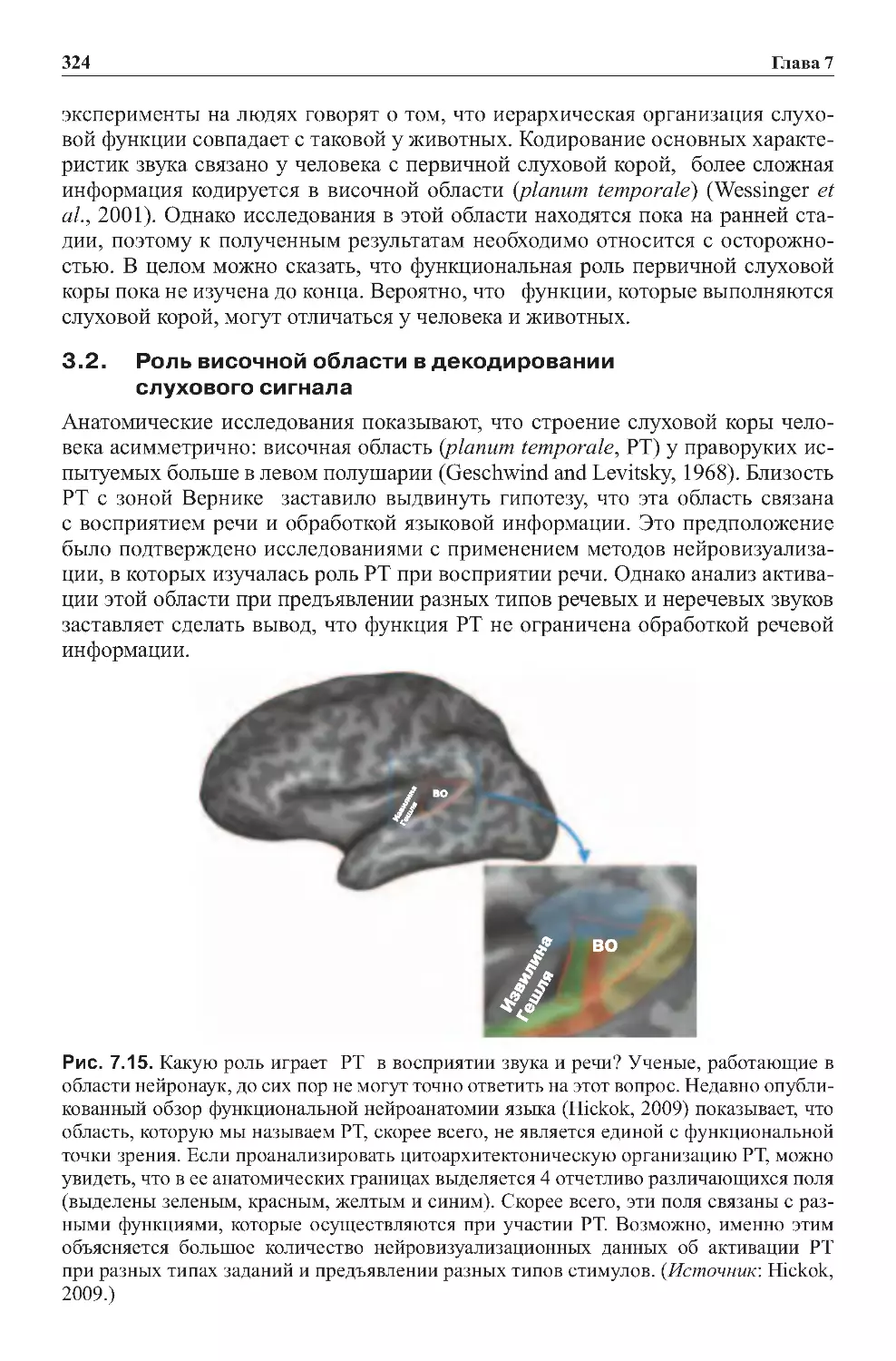
3.2. Роль височной области в декодировании
слуховогосигнала............................................324
3.3.Корковыесистемы«что»и«где»................................325
4.0.Восприятиеречи..............................................336
4.1.Историявопроса.............................................337
4.2.Ранниетеориивосприятияречи................................340
4.3. Функциональное картирование специфических
речевыхпроцессов...........................................342
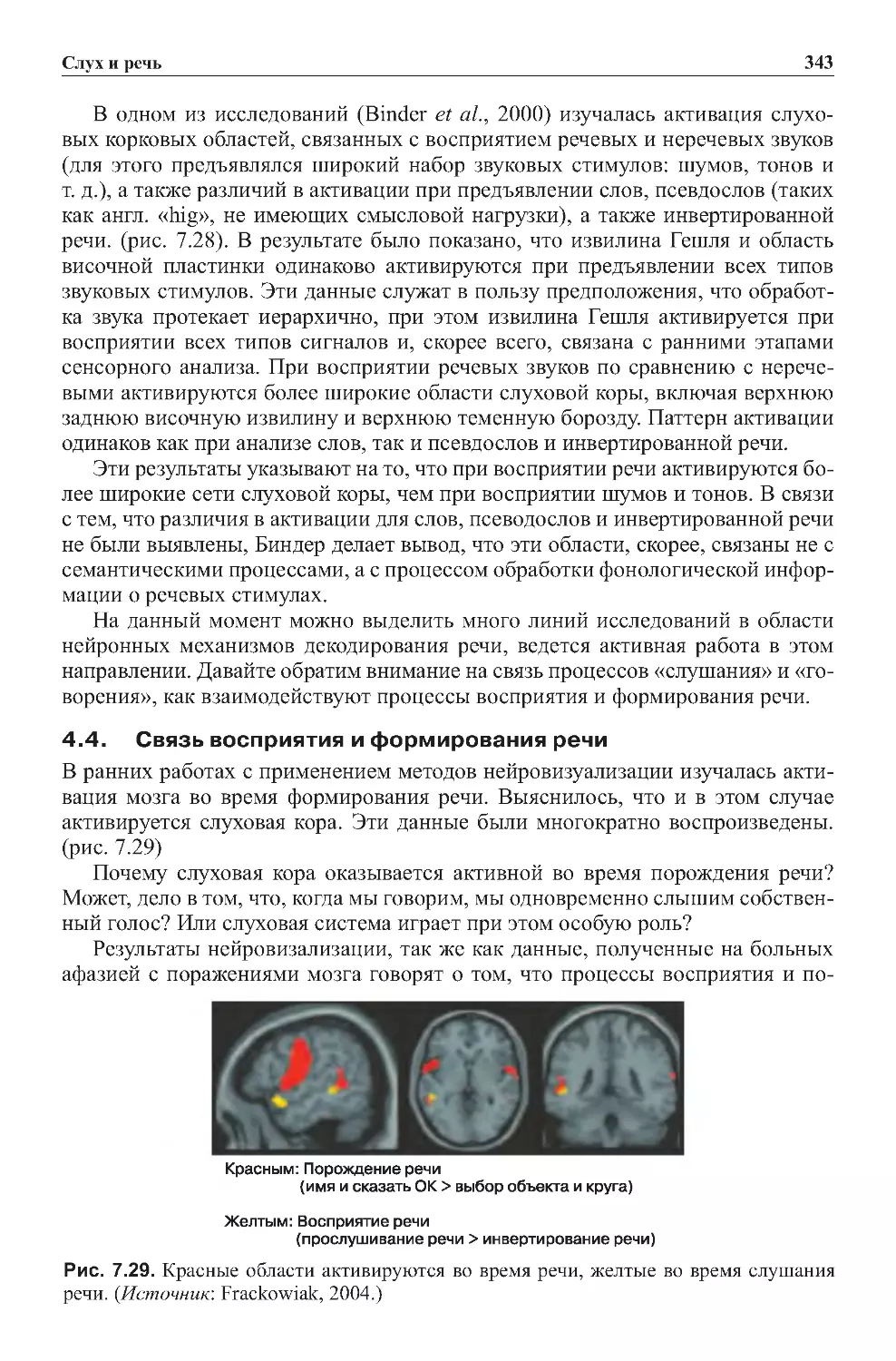
4.4.Связьвосприятияиформированияречи.........................343
4.5.Повреждениясистемвосприятияречи...........................344
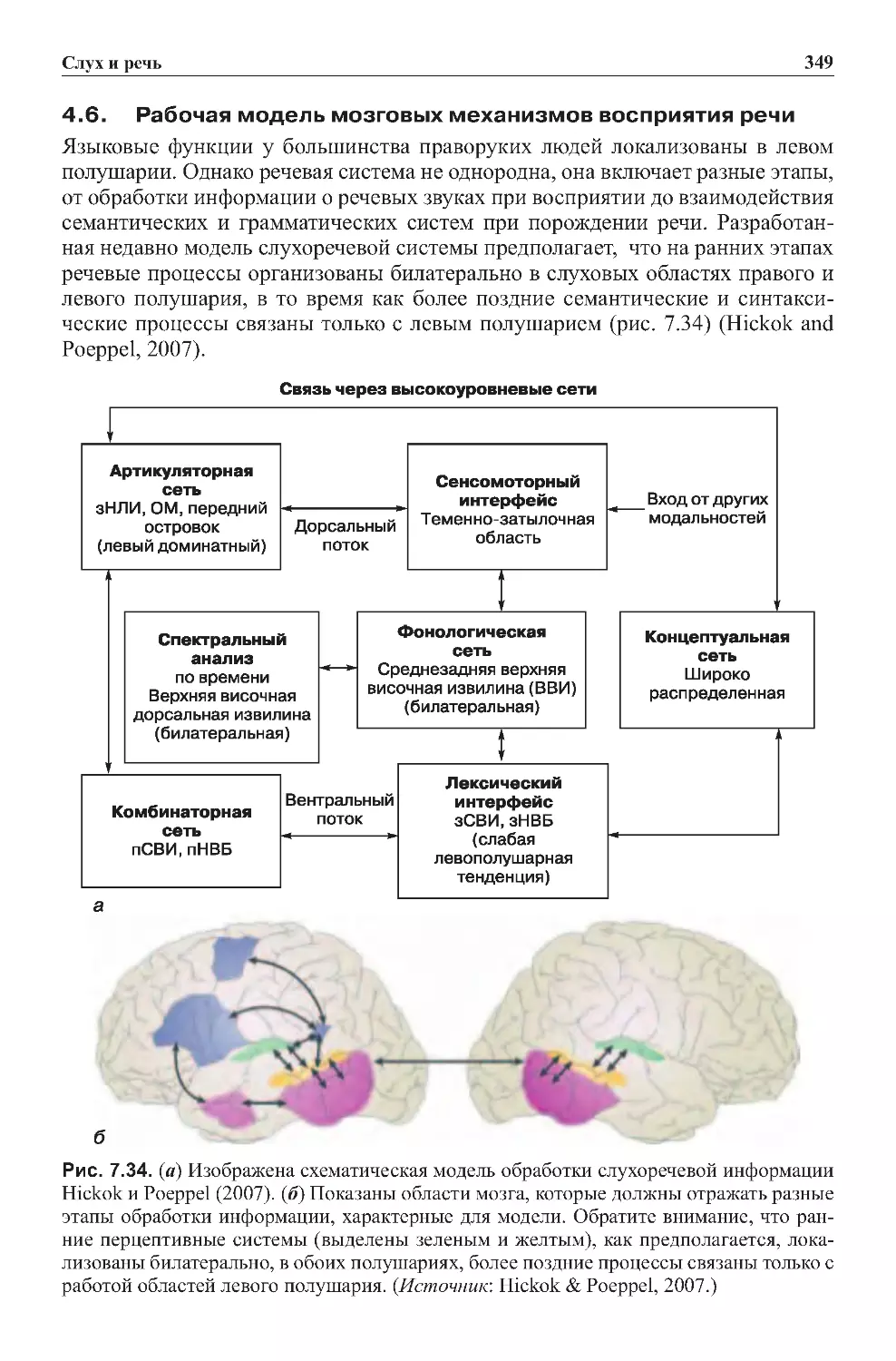
4.6. Рабочая модель мозговых механизмов восприятия речи . . . . . . . . . . . . 349
5.0.Восприятиемузыки............................................352
5.1.Этапыслуховойобработкимузыки..............................352
5.2.Отдельнаясистемадлямузыкальноговосприятия.................353
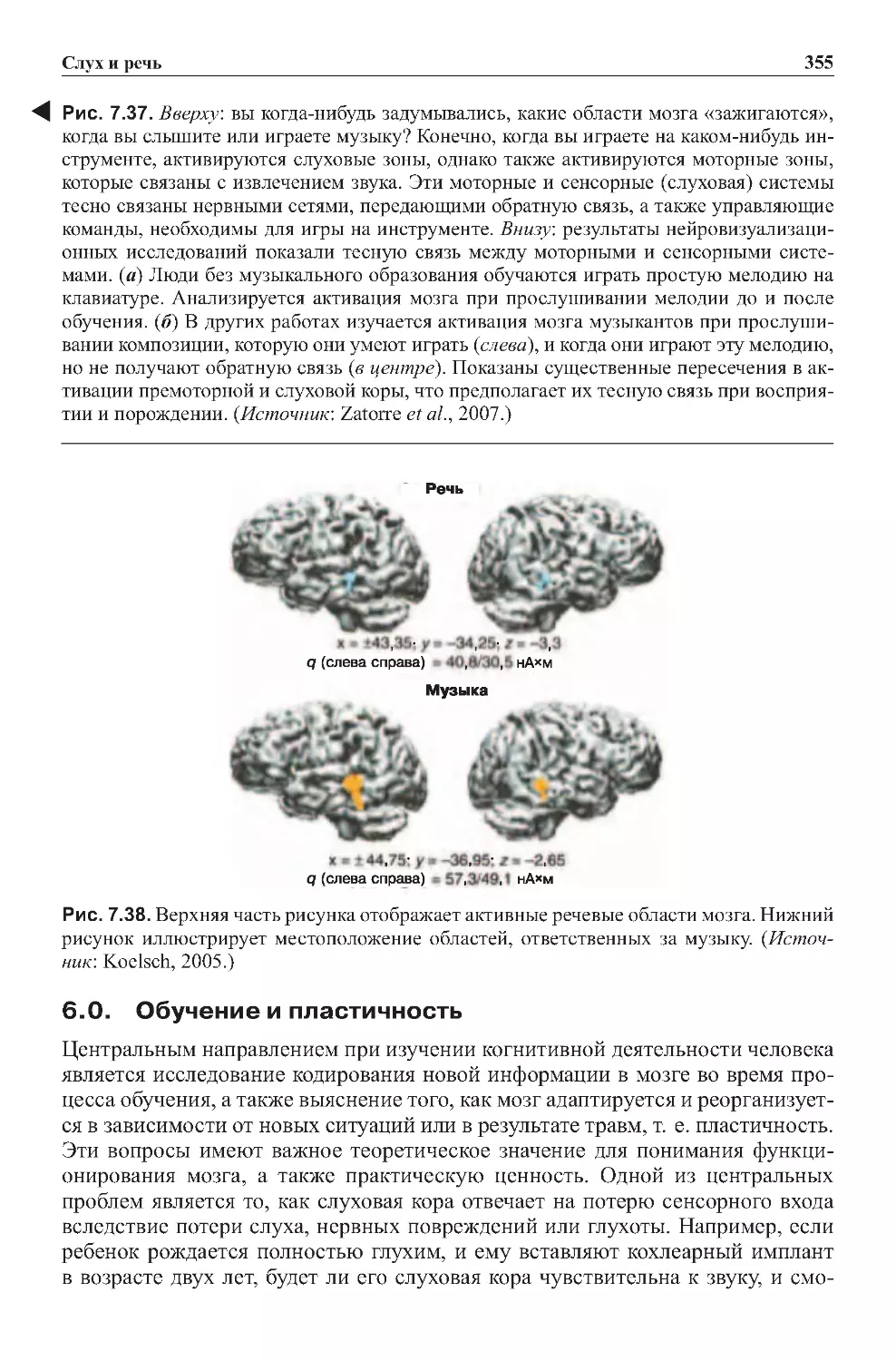
6.0.Обучениеипластичность.......................................355
6.1.Пластичность,связаннаясдепривацией.........................356
6.2.Пластичность,связаннаясобучением...........................356
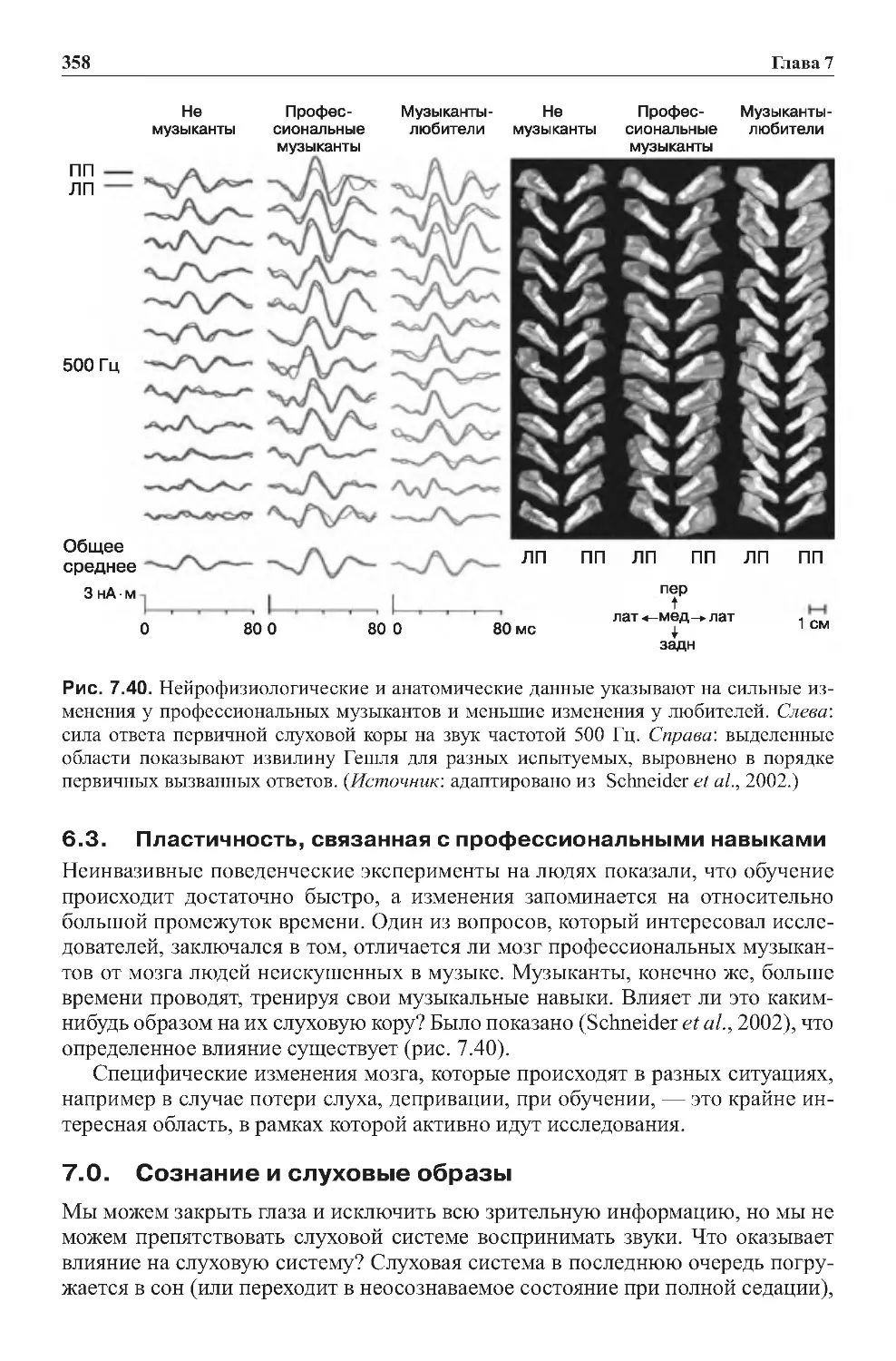
6.3. Пластичность, связанная с профессиональными навыками . . . . . . . . . . 358
7.0.Сознаниеислуховыеобразы....................................358
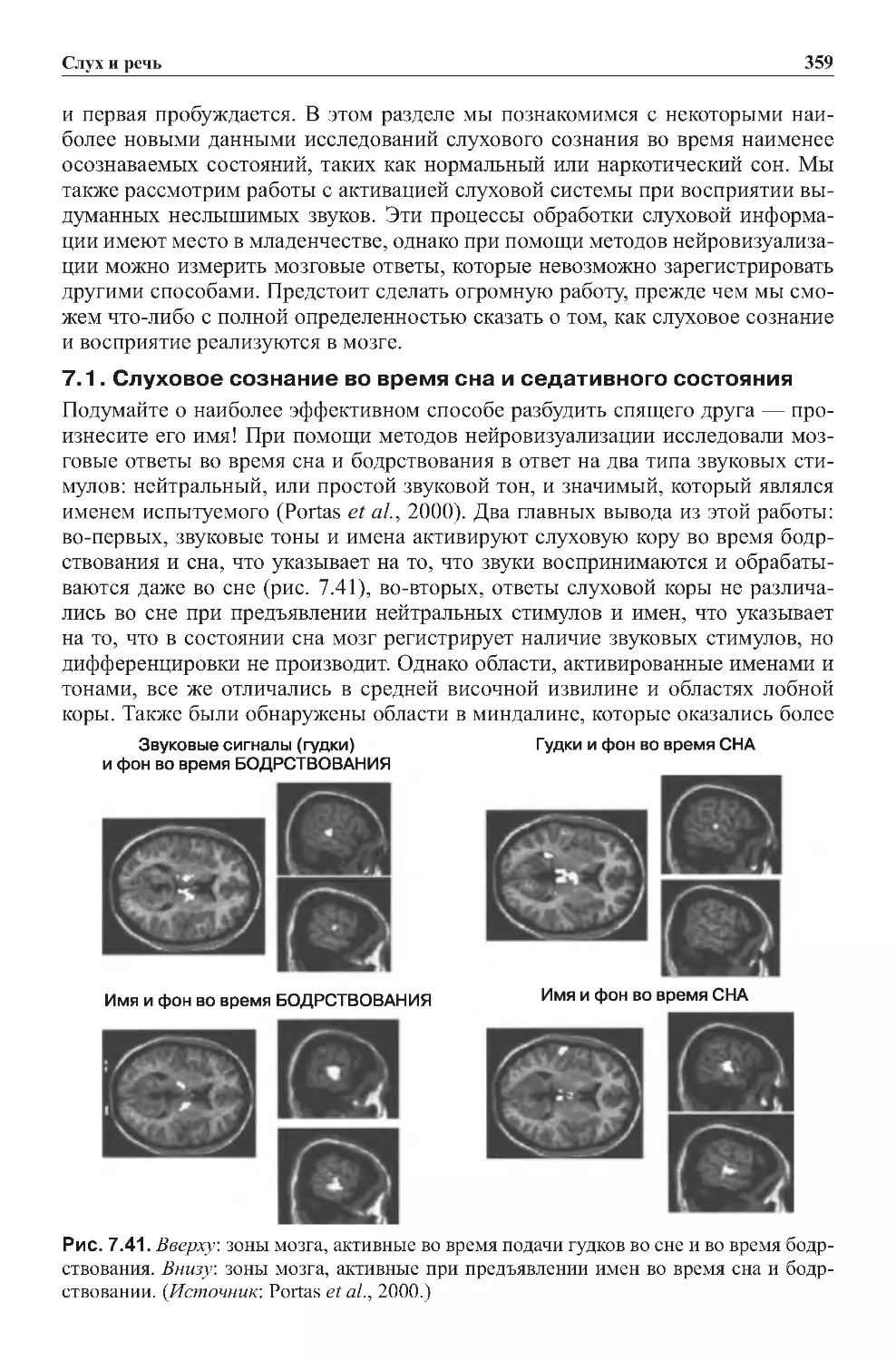
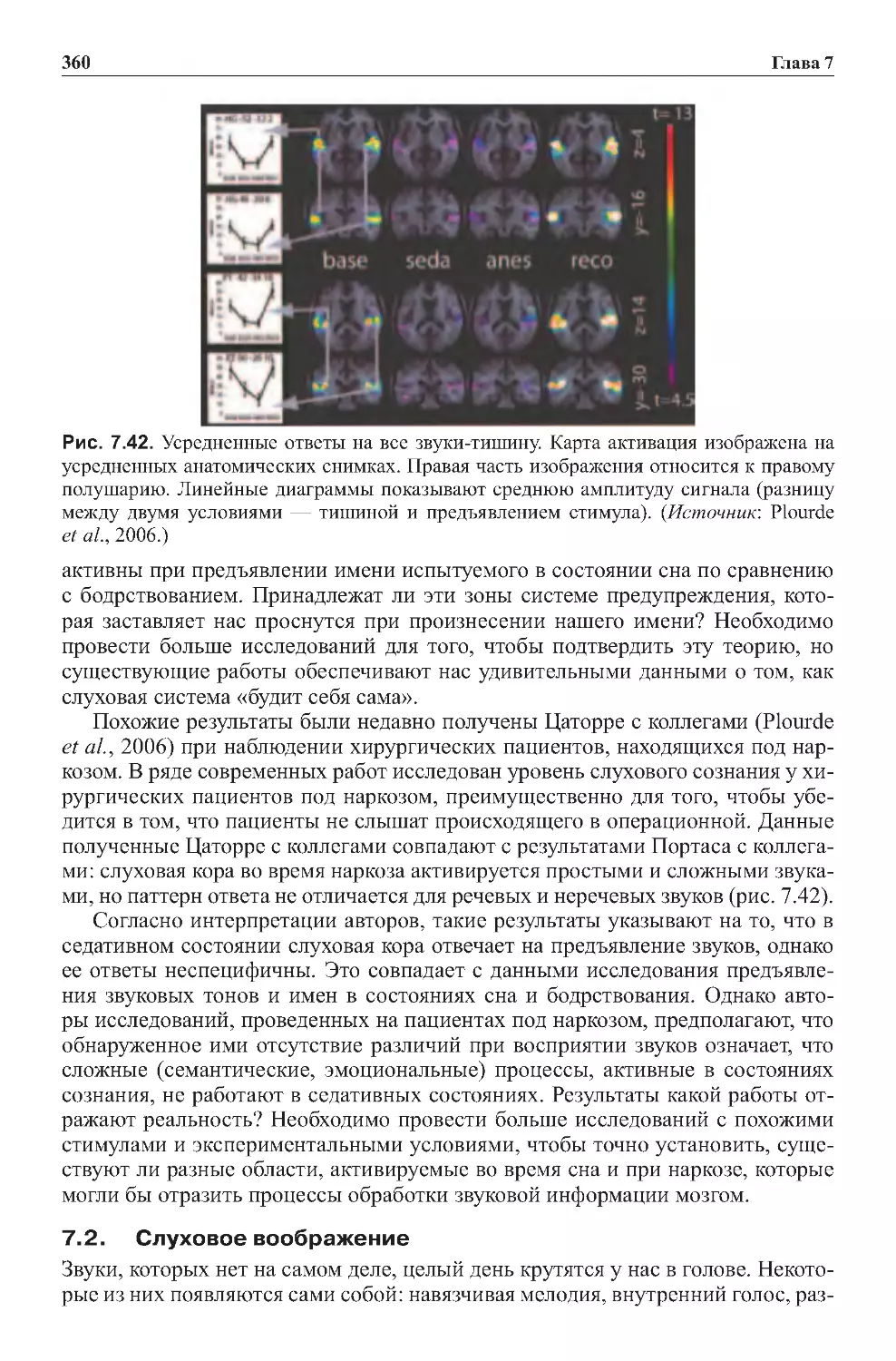
7.1. Слуховое сознание во время сна и седативного состояния . . . . . . . . . . 359
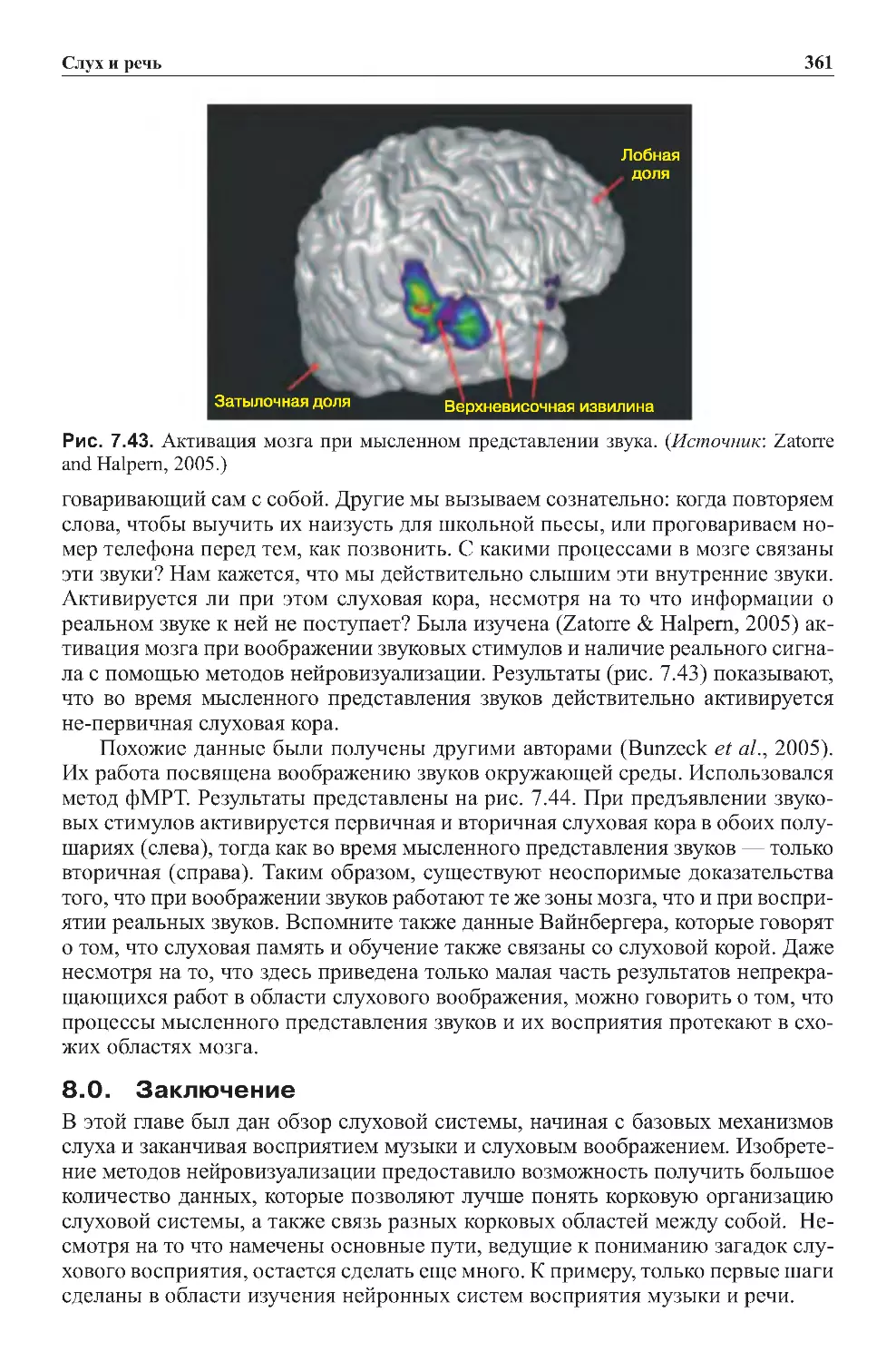
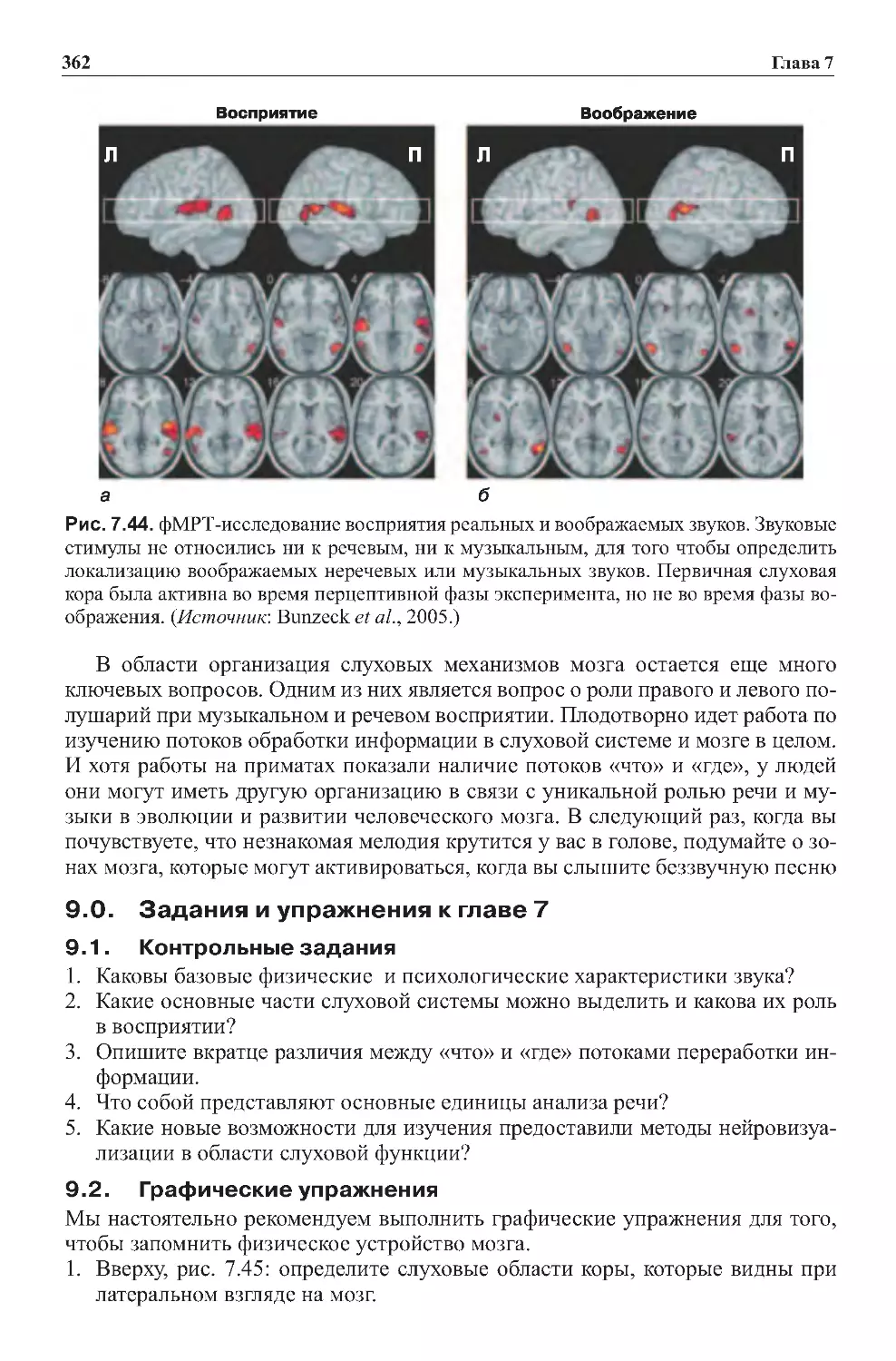
7.2.Слуховоевоображение........................................360
8.0.Заключение...................................................361
9.0.Заданияиупражнениякглаве7..................................362
9.1.Контрольныезадания.........................................362
9.2.Графическиеупражнения.....................................362
9.3.Литература.................................................363
Глава8.Сознаниеивнимание..............................365
1.0.Введение.....................................................366
1.1. Бодрствующее состояние опосредовано сознанием . . . . . . . . . . . . . . . . 370
1.2. Состояния бодрствования, сна и сновидений должны быть
регулярныповремени........................................371
Оглавление
11
1.3.Общиеритмысуточныхсостояний.............................373
1.4. Состояние нахождения в сознании
имеет типичную таламокортикальную активность. . . . . . . . . . . . . . . . . 378
1.5. Таламокортикальный центр:
плотновзаимосвязанныйиоченьактивный......................381
1.6.Картыиритмы..............................................385
1.7.Двусторонниесвязи..........................................385
1.8.Каксинхронизируютсянейроны................................387
1.9.Синхронностьдляполученияконтроля..........................390
2.0.Бодрствование................................................394
2.1.Практическоемышление......................................394
2.2.Ориентацияпоместу,времениилюдям.........................396
2.3. Бодрствование для обучения;
сонделаетвозможнымзакреплениепамяти......................398
2.4. Внимание и сознание в общем
делаютвозможнымобучение..................................398
2.5.Потеряосознанияпредсказуемыхсобытий.......................401
2.6.Скрытоеобучениетакжетребуетсознания.......................404
2.7. Быстрые ритмы координируют задачи в состоянии
бодрствования...............................................404
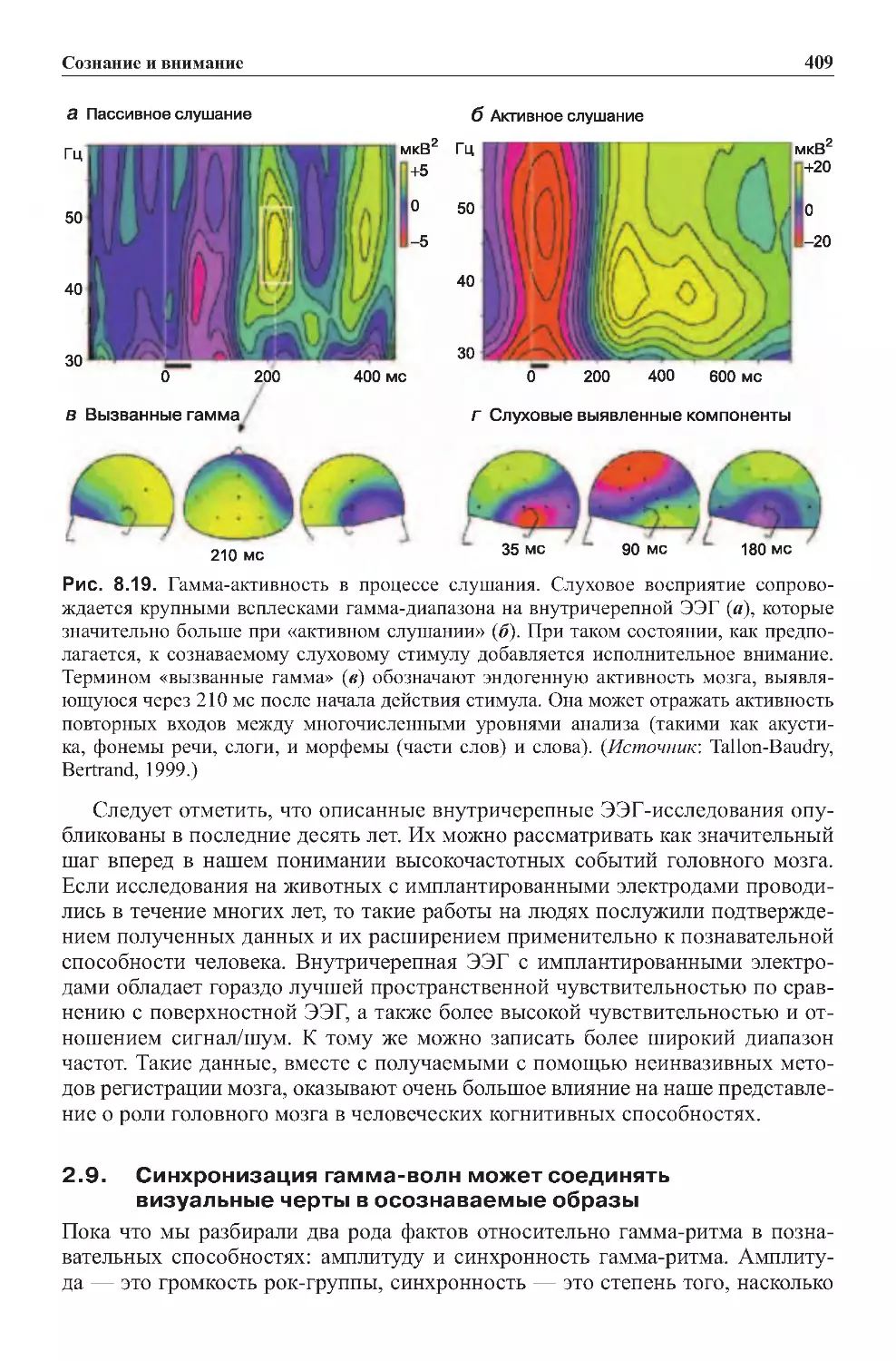
2.8.Гамма-активностьиграетмножестворолей......................406
2.9. Синхронизация гамма-волн может соединять визуальные черты
восознаваемыеобразы.......................................409
2.10.Тета-ритмыиграютмножестворолей...........................415
2.11.Альфа-ритмы...............................................417
3.0. Внимание улучшает восприятие,
познавательнуюспособностьиобучение..........................420
3.1.ФланговаязадачаПознера.....................................420
3.2.Модельвнимания............................................427
3.3.Опытыповниманиюиопытысознания.........................433
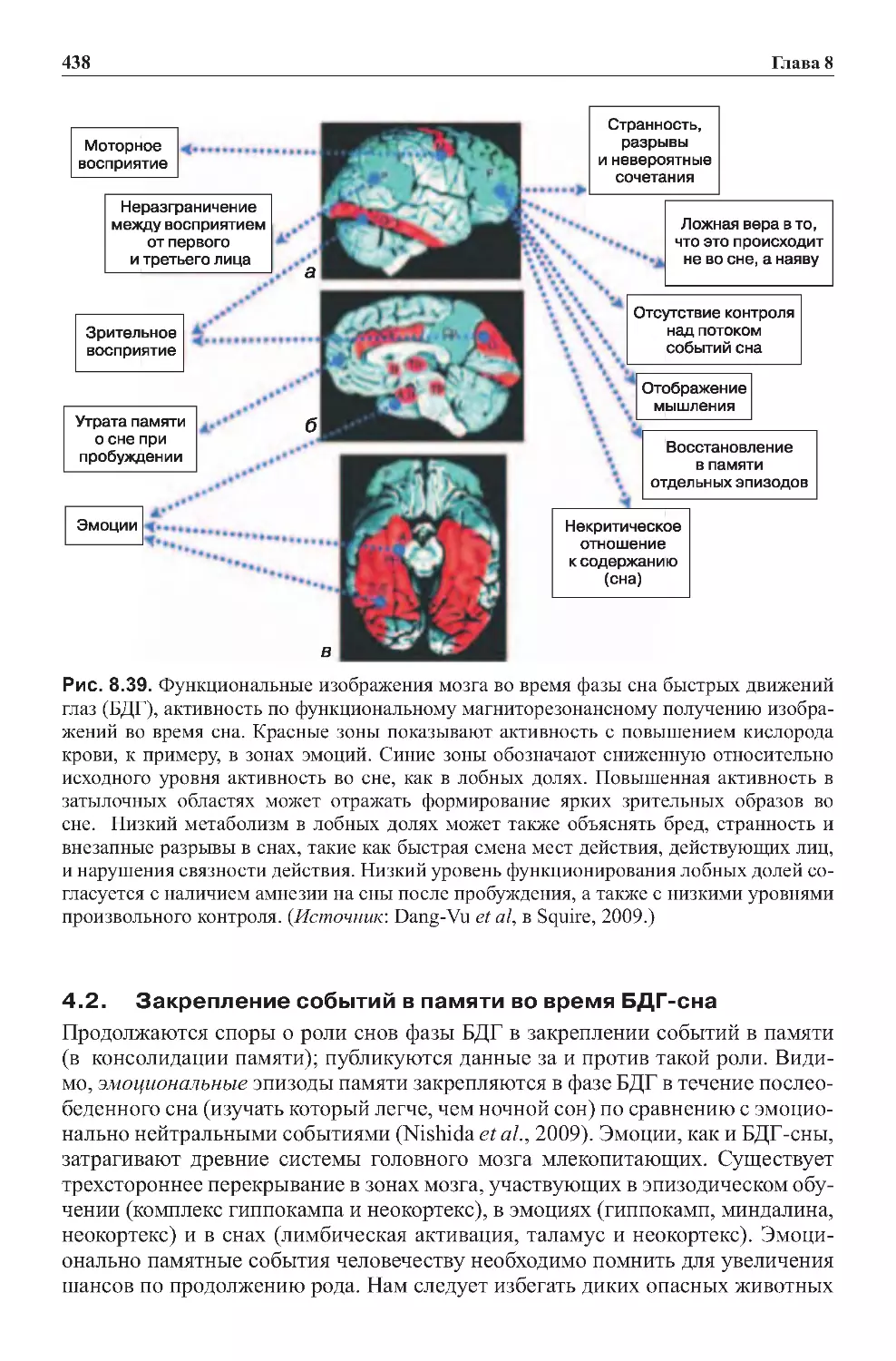
4.0.Сонфазыбыстрыхдвиженийглаз...............................435
4.1.Сныкаксостояниесознания...................................437
4.2.ЗакреплениесобытийвпамятивовремяБДГ-сна..................438
5.0.Глубокийсон:подъемыиспады.................................441
5.1. Некоторая умственная активность происходит даже во время
медленноволновогосна.......................................442
5.2.Порогпробужденияизменяетсянапротяжениисна................442
5.3. Повторное «проигрывание» эпизодов в памяти
иконсолидация..............................................443
6.0.Общеесопоставление..........................................447
6.1. Отражает ли сознание функцию глобального
рабочегопространствавголовноммозге?........................447
12
Оглавление
6.2. Передача сигналов повторным входом и степень интеграции
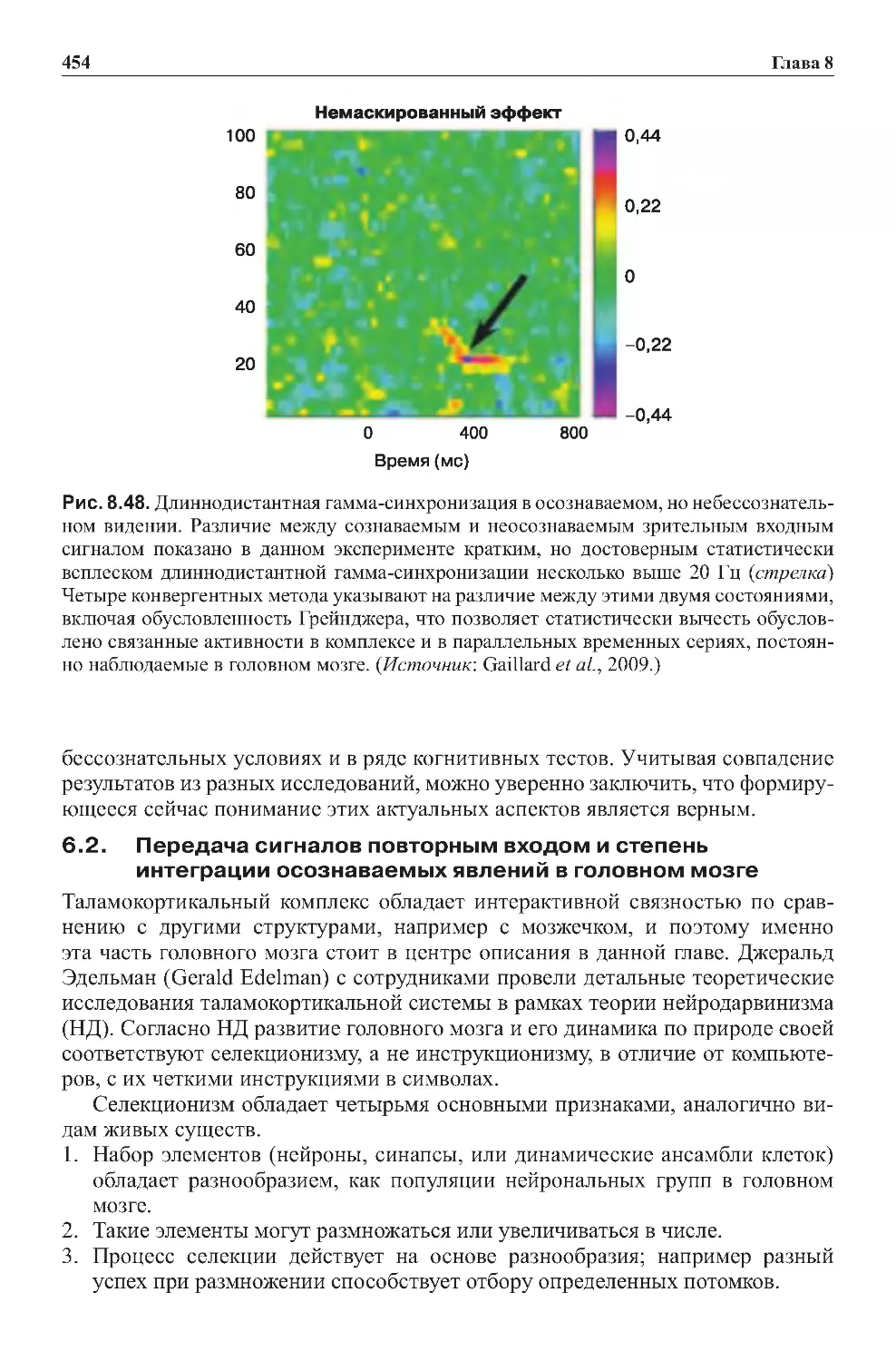
осознаваемыхявленийвголовноммозге.........................454
6.3.Требуетсялидлясознанияличноепознание?.....................457
6.4. Почему сознаваемые события подлежат регистрации?. . . . . . . . . . . . . . 459
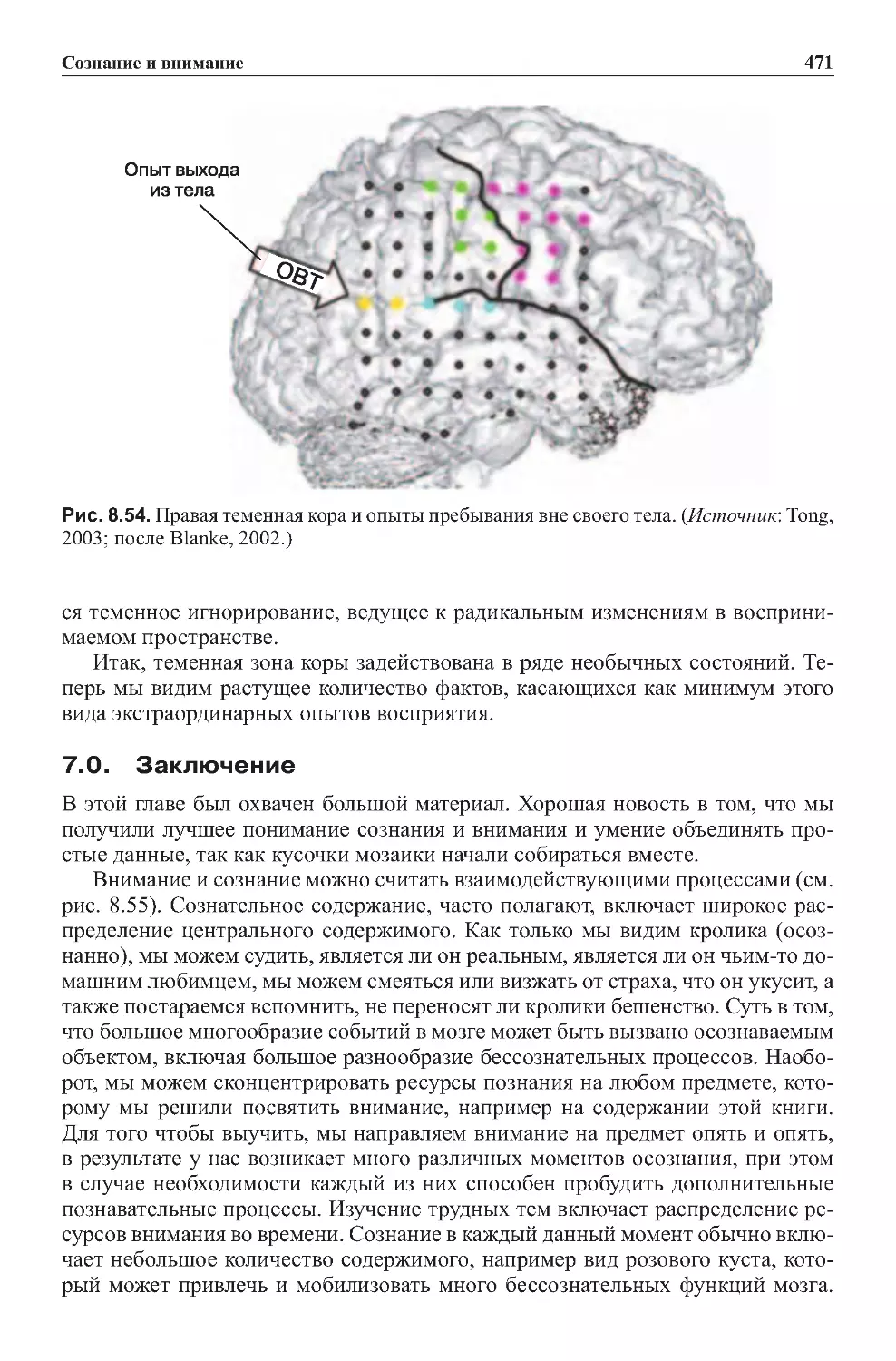
6.5.Фактыпонеобычнымсостояниям..............................460
7.0.Заключение...................................................471
8.0Заданияиупражнениякглаве8 .................................473
Глава9.Обучениеипамять.................................479
1.0.Введение.....................................................480
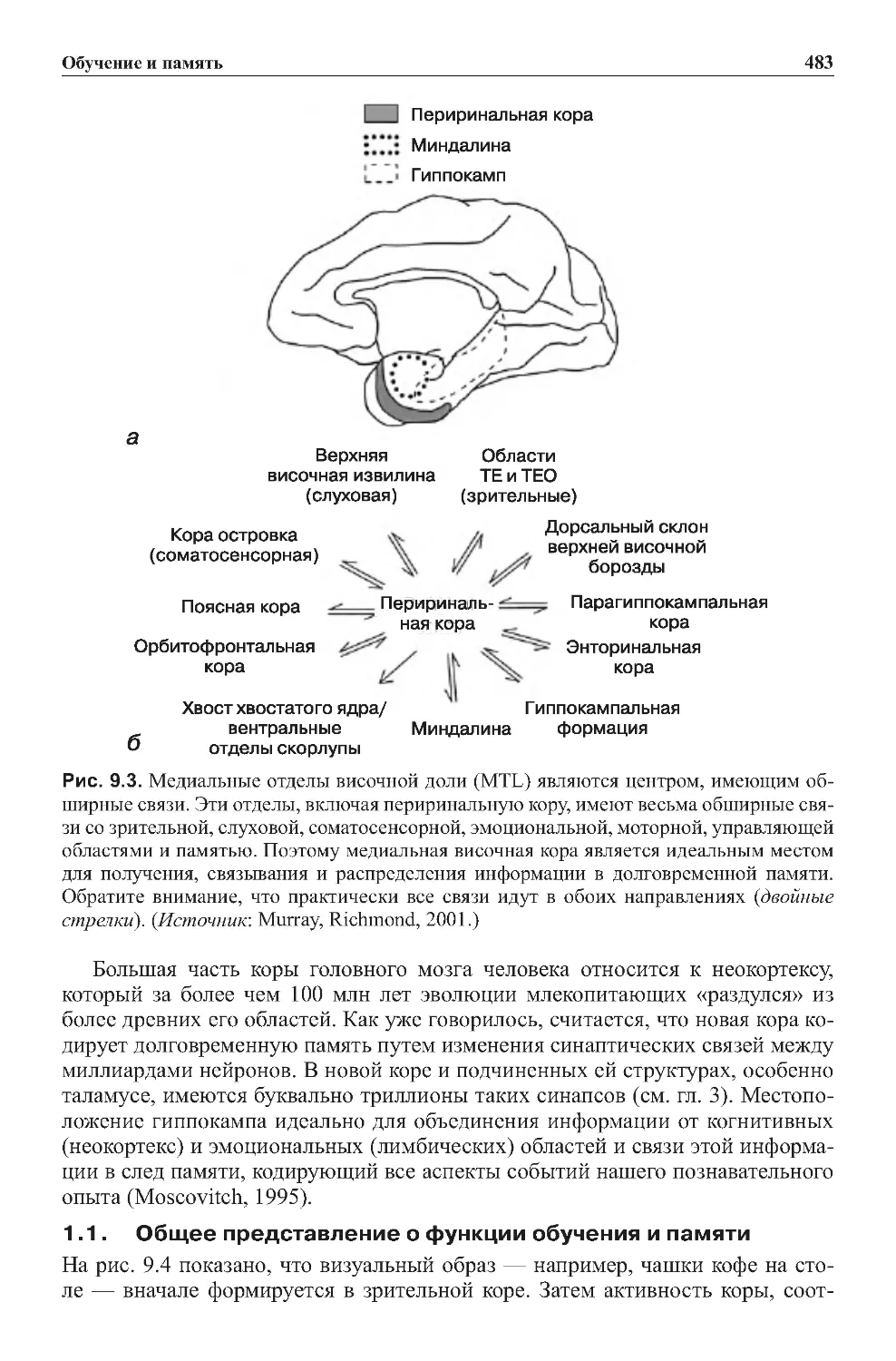
1.1.Общеепредставлениеофункцииобученияипамяти ..............483
1.2. Обучение и память на функциональной схеме работы мозга . . . . . . . . . 485
1.3.Имплицитнаяиэксплицитнаяпамять............................488
2.0.Амнезия.....................................................489
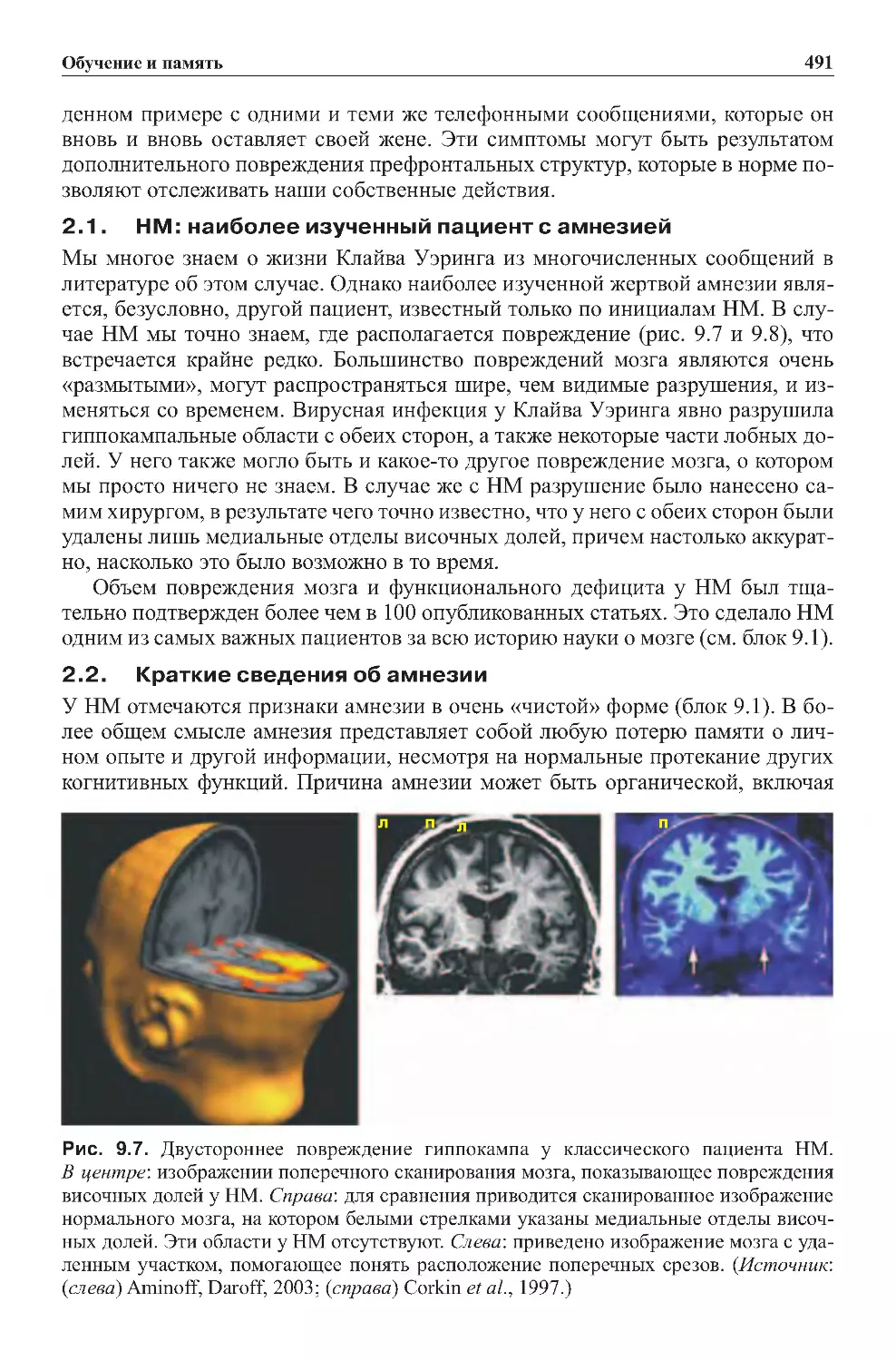
2.1.HM:наиболееизученныйпациентсамнезией....................491
2.2.Краткиесведенияобамнезии..................................491
2.3. Сохраняющиеся функции при амнезии: имплицитная
ипроцедурнаяпамять.........................................494
2.4.Сохраненноеимплицитноеобучение............................496
3.0.Какформируютсявоспоминания.................................500
3.1. Электрически вызванные автобиографические воспоминания . . . . . . . 501
3.2. Длительная потенциация и длительная депрессия:
возбуждающиеитормозныеследыпамяти.......................504
3.3. Консолидация: от временного хранения до постоянного . . . . . . . . . . . . 507
3.4. Быстрая консолидация: синаптические механизмы,
транскрипциягеновисинтезбелка..............................508
3.5 . Системная консолидация: взаимодействие между
медиальными отделами височных долей и новой корой. . . . . . . . . . . . . 510
4.0.Разновидностипамяти.........................................512
4.1. Эпизодическая и семантическая память:
«припоминание»против«знания»..............................512
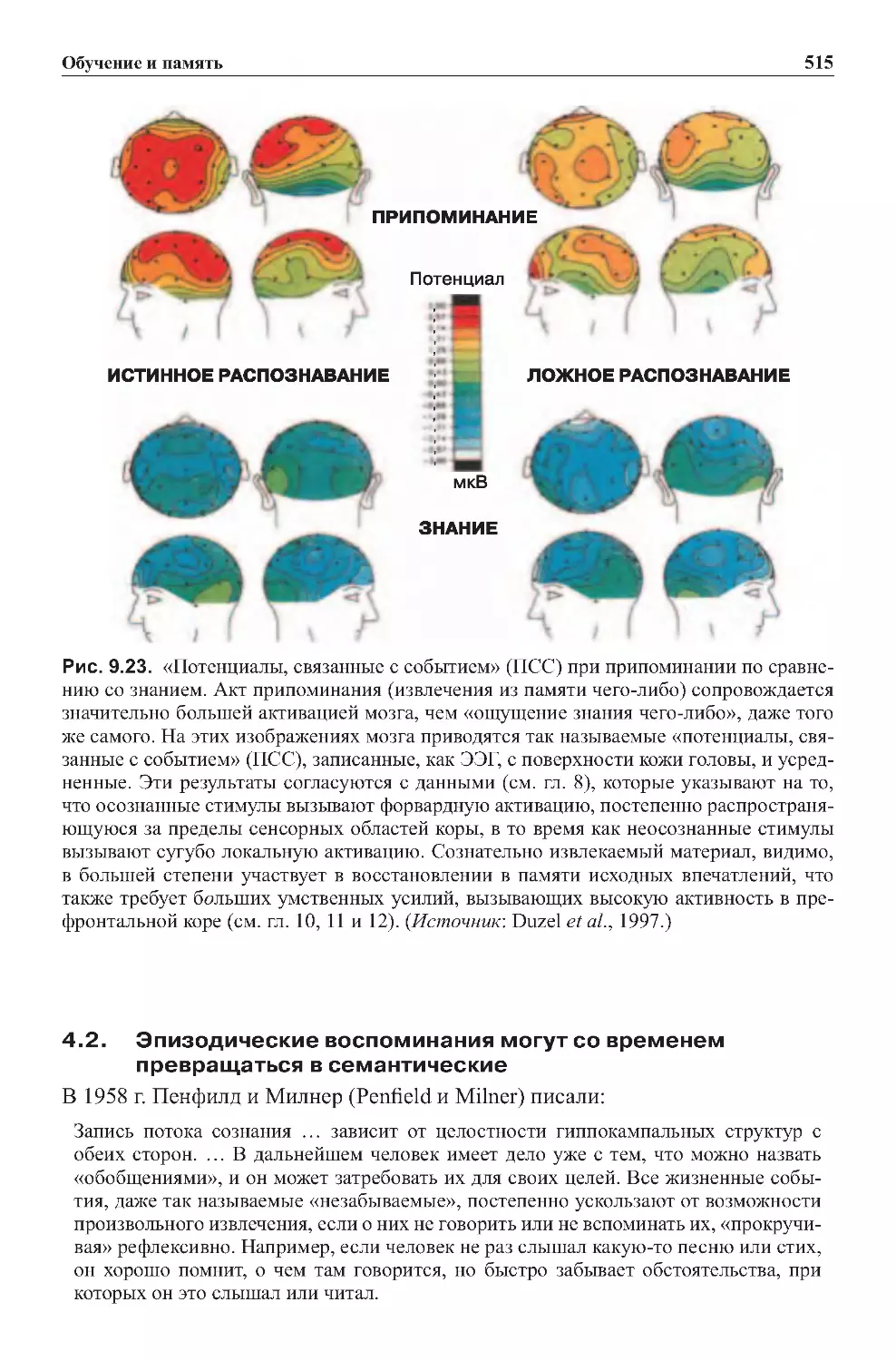
4.2. Эпизодические воспоминания могут со временем
превращатьсявсемантические.................................515
4.3. Эпизодическая память и семантическая память
частокомбинируются.........................................517
5.0. Медиальная височная кора в эксплицитном научении и памяти . . . . . . .518
5.1.Отвлечениевниманиямешаетобучению.........................519
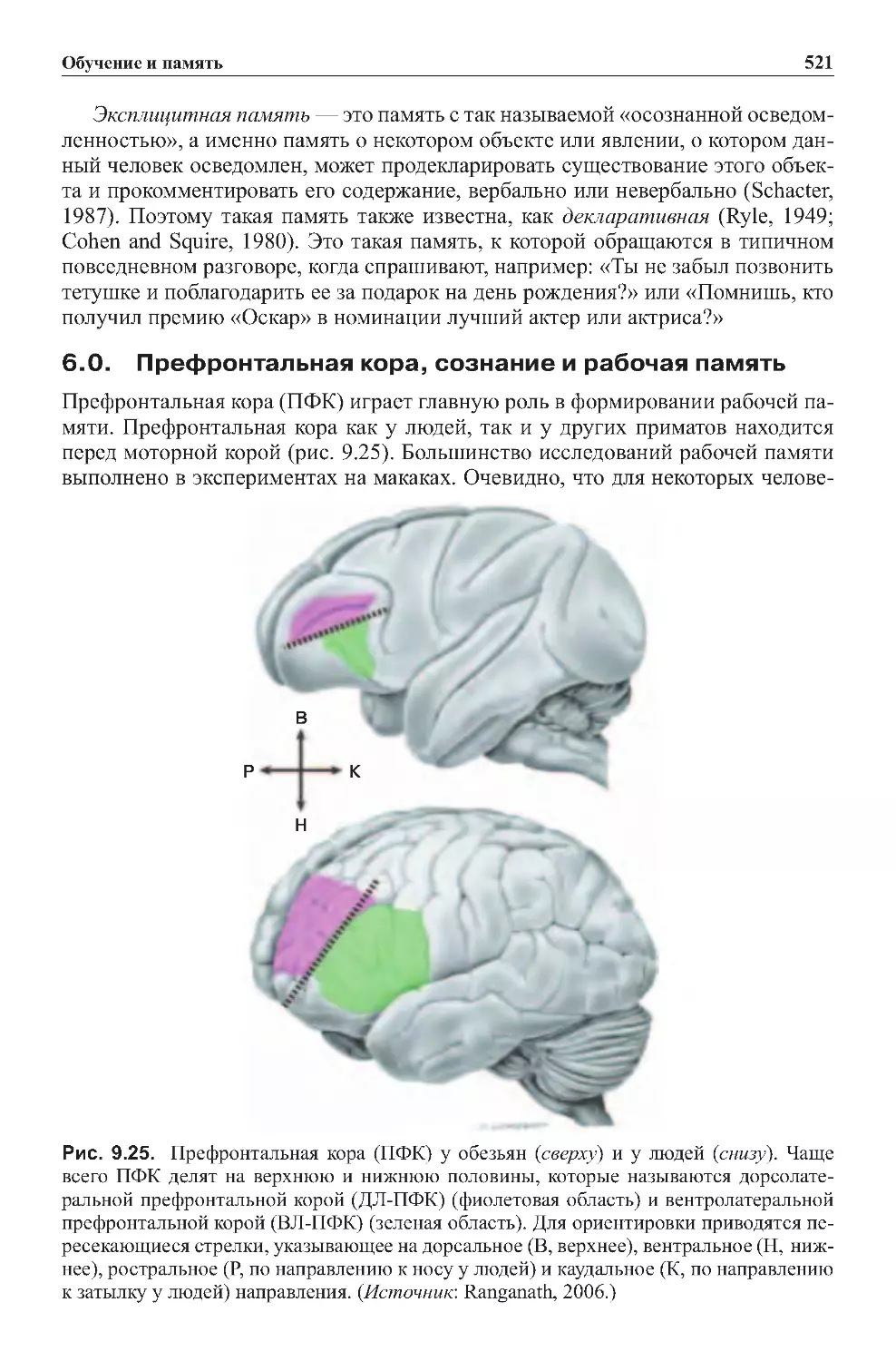
6.0.Префронтальнаякора,сознаниеирабочаяпамять..................521
6.1. Работа с памятью: лобная доля целенаправленно
работаетспамятью...........................................525
Оглавление
13
6.2. Префронтальная кора в эксплицитном (осознанном)
иимплицитном(неосознанном)обученииипамяти................526
6.3.Различныетипырабочейпамяти................................528
6.4. Префронтальная кора — хранение информации
илиуправлениепроцессом?....................................530
6.5. Объединение префронтальных и медиальных
височных областей для обеспечения процессов
рабочейпамяти..............................................531
7.0.Извлечениеинформацииизпамятииметапознание.................533
7.1.Извлечениеизпамятиложнойинформации......................534
7.2. Межполушарная латерализация
приизвлеченииинформации ..................................534
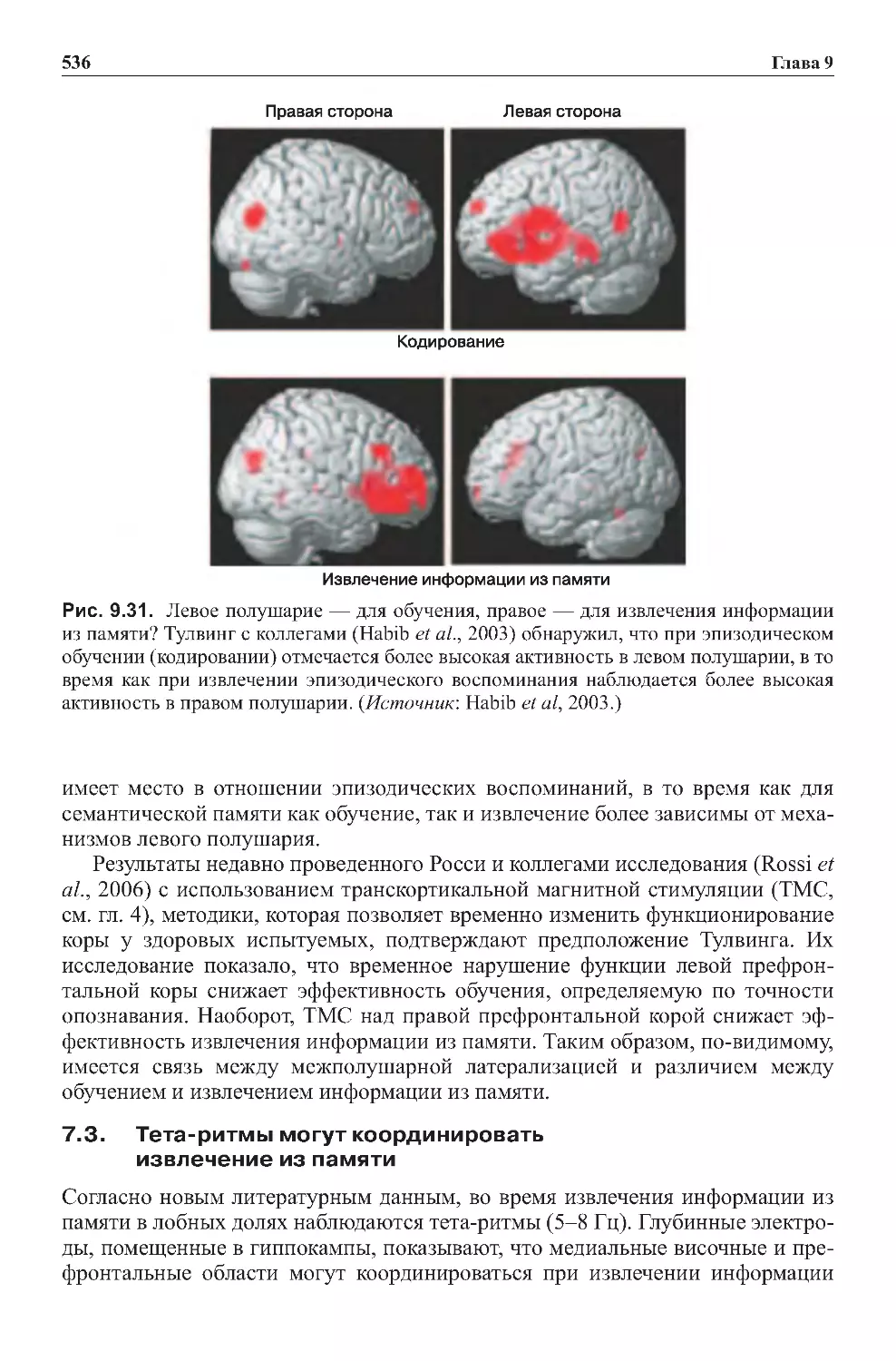
7.3. Тет а -ритмы могут координировать извлечение из памяти . . . . . . . . . . . 536
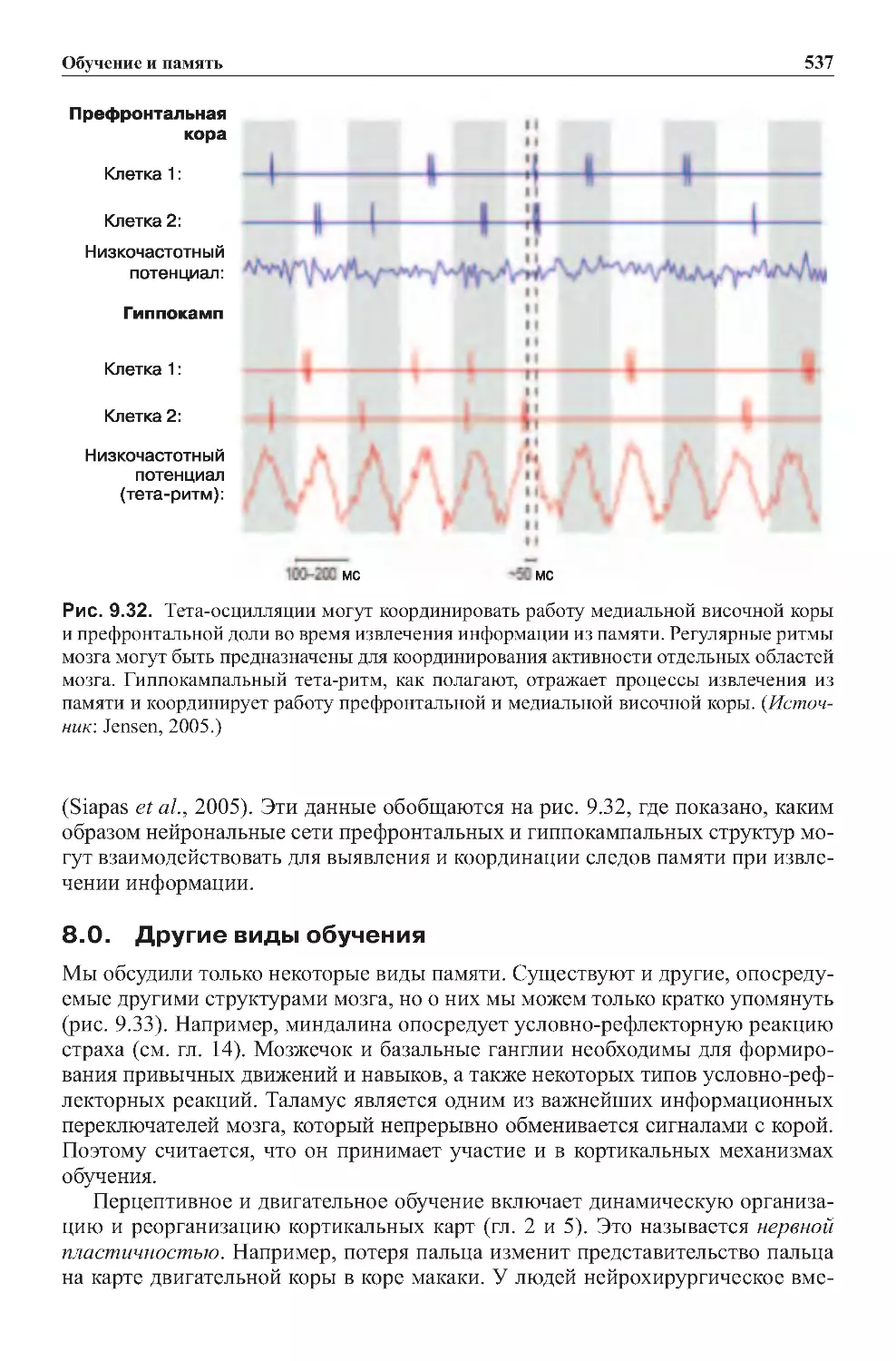
8.0.Другиевидыобучения.........................................537
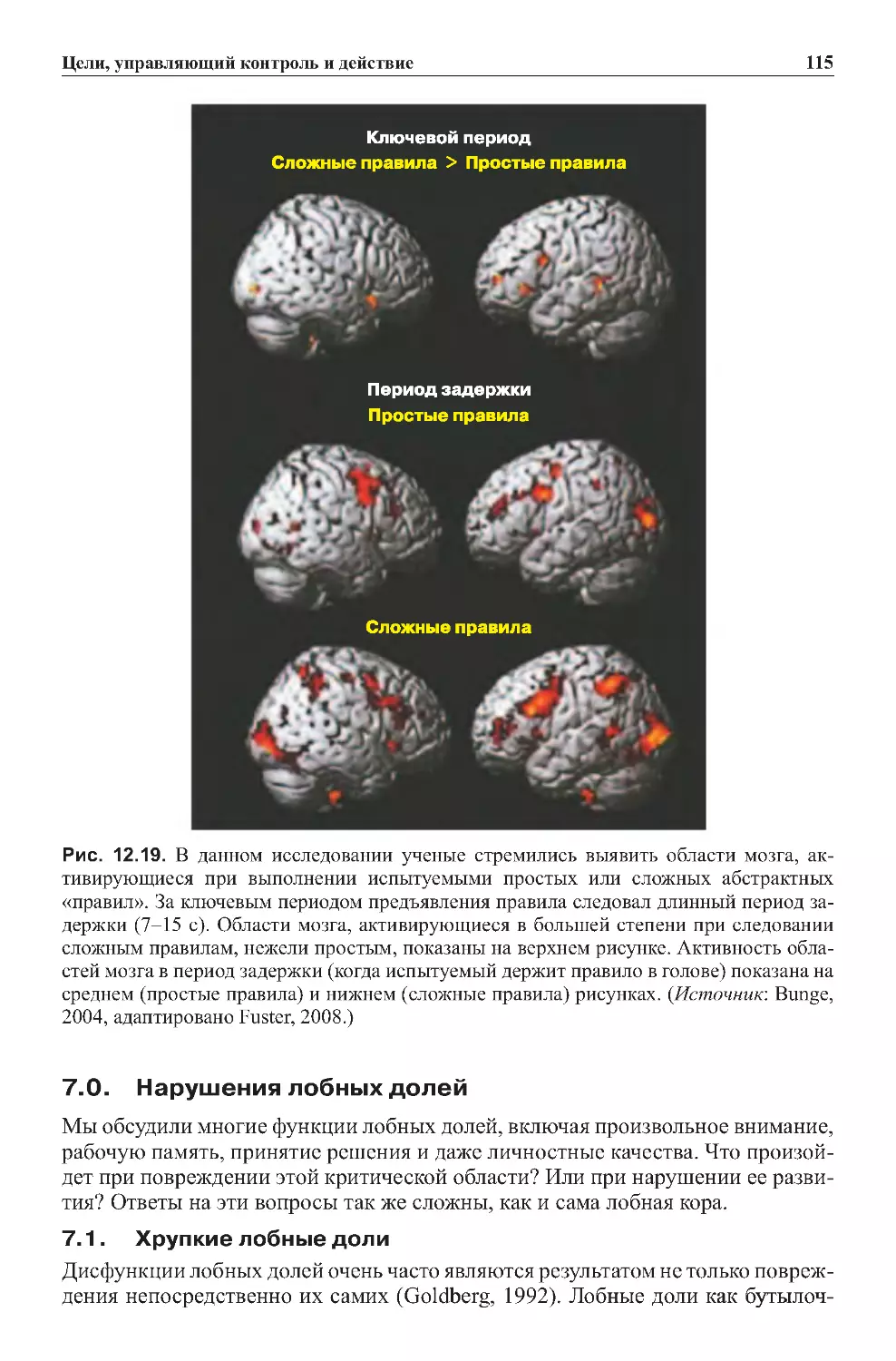
9.0.Заключение...................................................539
10.0.Заданияиупражнениякглаве9.................................540
Стремление идти в ногу со временем при изучении когнитивной нейробиологии
очень схоже со стремлением оседлать на серфинге Большую волну около пляжа
Вайкики. От постоянного появления новых фактов в этой области становится
захватывающе интересно и немного страшно. И все же мы продолжаем взби-
раться на наши умственные доски для серфинга, чтобы поймать набегающую
волну этого все усложняющегося научного знания. Данная книга создавалась
с целью формирования общего представления о еще только зарождающейся
науке. Особый стиль изложения основного материала делает ее доступной как
для студентов, так и для всех интересующихся читателей любого уровня.
Второе издание книги претерпело ряд усовершенствований.
1. Добавлена новая глава Гены и молекулы мышления (гл. 16), которая знако-
мит читателя с современными разработками в области молекулярных основ
сознания. Известно, что нейроны формируют связи с другими нейронами,
экспрессируя белки под контролем генетического (и эпигенетического) ап-
парата клетки. Таким образом, знание молекулярного уровня необходимо
для понимания таких функций, как обучение, речь, восприятие и другие.
2. С учетом данных, полученных за последние десять лет, полностью пере-
смотрена глава о Сознании и внимании (гл. 8). Невероятный прорыв в данной
области позволил совершить открытие новых методов регистрации деятель-
ности мозга. Например, было показано, что мозговые ритмы несут информа-
цию как о сознательных, так и о бессознательных процессах.
3. Была произведена большая ревизия гл. 12 Цели, управляющий контроль и
действия, в которой с позиции современных данных о строении префрон-
тальной коры проясняются многие загадки о значении лобных долей — «ор-
ганов цивилизации», длительное время считавшихся непостижимыми.
4. Добавлены новые текстовые вставки Перспективы когнитивной нейрона-
уки. Свои точки зрения на важные направления и открытия, сделанные не-
давно в рамках когнитивной нейронауки, представляют ведущие ученые со
всего мира:
• Нельсон Кован (Nelson Cowan, PhD), Университет Миссури
• Давид Иглман (David Eagleman, PhD), Медицинский колледж Бэйлора
• Джеральд Эдельман (Gerald Edelman, MD), Институт нейронаук
• Пол Флетчер (Paul Fletcher, PhD), Кембриджский университет
• Анжела Фридеричи (Angela Friederici, PhD), Институт человеческой ког�
, PhD), Институт человеческой ког�
), Институт человеческой ког-
нитологии и науки о мозге общества Макса Планка
Предисловие
Предисловие
15
• Кристофер Фрис (Christopher Frith, PhD), Центр исследования мозга
«Вэлкам Траст»
• Кристоф Кох (Christof Koch, PhD), факультет биологии Калифорнийского
технологического института
• Стефан Л. Макник (�tephen �. Mac�ni�, PhD), Нейрологический инсти�
�tephen �. Mac�ni�, PhD), Нейрологический инсти�
tephen �. Mac�ni�, PhD), Нейрологический инсти�
�. Mac�ni�, PhD), Нейрологический инсти�
. Mac�ni�, PhD), Нейрологический инсти�
Mac�ni�, PhD), Нейрологический инсти�
), Нейрологический инсти-
тут Барроу
• Сюзана Мартинез�Конде (�usana Martinez�Conde, PhD), Нейрологиче�
, PhD), Нейрологиче�
), Нейрологиче-
ский институт Барроу
• Анирудх Пател (Aniruddh Patel, PhD), Институт нейронаук
• Чаран Ранганах (Charan Ranganath, PhD), Университет Калифорнии,
Дейвис
• Майкл Рагг (Michael Rugg, PhD), Университет Калифорнии, Ирвин
• Дженни Саффран (�enn� �affran, PhD), Университет Висконсина, Мэди�
�enn� �affran, PhD), Университет Висконсина, Мэди�
�affran, PhD), Университет Висконсина, Мэди�
�affran, PhD), Университет Висконсина, Мэди�
), Университет Висконсина, Мэди-
сон
• Лари Скваер (�arr� �quire, MD), Университет Калифорнии, Медицинская
школа Сан Диего
5. Во второе издание включен словарь терминов. Так как освоение терминоло-
гии является одним из наиболее сложных этапов изучения когнитивной ней-
ронауки, мы надеемся, что это дополнение будет хорошим вспомогательным
средством для студентов и преподавателей.
6. В начало книги помещен новый мини�атлас человеческого мозга, знание
строения которого служит первым шагом к пониманию когнитивных наук.
Так как мозг невероятно сложен по своему топографическому устройству, в
первом издании мы использовали иллюстрации из анатомии Грея и других
источников издательства Elsevier/Academic Press. Для большей помощи но-
вичкам при изучении «ландшафта мозга» во второе издание добавлен мини�
атлас — компас и карта «территории».
7. И наконец, приложение по методам визуализации мозга было полностью
обновлено экспертом в данной области, доктором Томасом Рамсой из Уни-
верситета Копенгагена.
Как писали Кристофер Фрис, Михаель Познер и другие, в последние не-
сколько десятилетий мы наблюдаем заключение союза между когнитивной на-
укой и наукой о мозге, который строится на взаимовыгодных исходных пре-
имуществах каждой из них. Когнитивные механизмы и механизмы восприятия,
изучавшиеся на уровне поведения, в настоящее время могут наблюдаться непо-
средственно на уровне работы самого мозга благодаря использованию новей-
ших методов визуализации мозговых процессов. Впервые в реальном времени
мы можем наблюдать деятельность живого мозга, которая формировалась в
процессе эволюции более сотни миллионов лет. Результаты этого исследования
невероятно богаты и соединяют в себе новейшие данные психологии и биоло-
гии, медицины, биохимии и физики. В то же время наибольшее число научных
разработок строится на базе хорошо устоявшихся психологических концепций
и методик. В результате мы можем наблюдать, как психология и наука о мозге
16
Предисловие
удивительным образом дополняют друг друга. Когнитивная нейронаука стано-
вится обязательной составляющей образовательных программ по психологии,
биологии, педагогике и медицине.
Сложно охватить науку о мышлении за один курс. Многие лекторы замеча-
ют, что большая часть учебного времени тратится на изучение строения самого
мозга, и очень мало времени остается для освещения сложных интегративных
процессов. Несмотря на то, что знание устройства мозга необходимо, фокуси-
рование исключительно на анатомии может помешать достижению целей пре-
подавания.
В данной книге к решению этой проблемы авторы подходят с нескольких
сторон. Во�первых, текст в своей основе придерживается гибкой обучающей
линии, которая включает стандартные разделы, такие как сенсорное восприя-
тие в зрительной и слуховой системах, рабочая память, внимание и сознание,
высшие нервные функции, язык и восприятие, когнитивная деятельность, эмо-
ции, социальное познание и развитие. Во�вторых, знакомство с деятельностью
мозга происходит постепенно, шаг за шагом, с поэтапным усложнением мате-
риала. Для облегчения восприятия информации в книге используется особая
функциональная основа. Этот широко распространенный способ позволяет со-
брать все главные разделы в единую схему, которая углубляется и разрастается
деталями по ходу изложения. Функциональную основу книги можно рассма-
тривать с разных сторон. Например, процесс хранения информации можно из-
учать исходя из понятий активной рабочей памяти, а восприятие, когнитивную
и высшую нервную деятельность — на фоне постоянного сохранения в мозге
текущей информации (гл. 2). Функциональная основа помогает разобраться в
обоих случаях.
После прохождения бесплатной регистрации для преподавателей и студен-
тов становится доступной информация на сайте http://textboo�s.elsevier.com.
Дополнительные материалы для старших наставников включают все рисунки
книги и подписи к ним в формате PowerPoint, а также видеоинструкции и муль-
тимедийные файлы. Студенты могут ознакомиться с краткими обзорами глав,
тестами, рисунками и видео. Сайт будет постоянно обновляться и дополняться
в зависимости от поступления новых данных, и авторы будут рады учесть ваши
предложения и идеи по поводу его содержимого.
Материал глав предлагается изучать в любой последовательности, удобной
для преподавания. Для студентов, продолжающих обучение, содержание глав
4 и 5 можно подать в кратком изложении. Для начальных курсов эти темы яв-
ляются основными и могут быть дополнены информацией из приложения (ав-
торы — Томас Рамсой с коллегами). Приложения также удобно использовать в
качестве справочного материала.
В книге рассматриваются все возможные нарушения мозговой деятельности,
начиная с описания случаев повреждения памяти у Генри Молайсона (Henr�
Gustav Molaison, HM) и возникновения антероградной амнезии в результате
вирусной инфекции у Клайва Веринга (гл. 2 и 9) и заканчивая слепым зрением,
гемиагнозией, прозопагнозией и другими расстройствами зрительной систе-
Предисловие
17
мы. В гл. 11, посвященной высшим нервным функциям, описаны расстройства,
вызванные чрезмерным или недостаточным центральным управлением. Так, в
случае некоторых расстройств не наблюдаются нарушения двигательной систе-
мы и когнитивных функций: кажется, что пациенты просто не желают выпол-
нять движение. Однако иногда они спонтанно начинают копировать поведение
другого человека и не могут остановиться. Например, импульсивно поднима-
ются, если осматривающий их врач встает со стула. Подобные расстройства,
связанные с чрезмерным или недостаточным центральным управлением, по-
зволяют выявить основы высших нервных функций человека.
Ряд нарушений имеет психологические аналоги. Профессиональные музы-
канты, такие как Ван Клиберн (Van Cliburn), иногда не в состоянии подавить
желание подпевать во время игры на инструменте, а высококвалифициро-
ванные специалисты могут потерять высший контроль над автоматическими
действиями, особенно при наличии одновременной мыслительной нагрузки.
Наоборот, классическим симптомом глубокой депрессии, например, является
невозможность инициировать и выполнить движение. Области мозга, задей-
ствованные в такие «чисто психологические» недостатки, как правило, связаны
с органическими нарушениями. Здесь мы наблюдаем еще один пример значи-
тельного упрощения информации, что позволяет читателям получить общую
картину быстрее, чем разрозненные факты.
В научных исследованиях психологическая тематика обычно сводится к рас-
смотрению данных о работе мозга. Например, на сегодняшний день считается,
что вербальное проявление классической рабочей памяти — возможность мыс-
ленно повторять последовательности чисел и наборы слов — является частью
нормального проявления речевой функции. Однако открытие возможности ак-
тивации областей мозга, ответственных за речевые функции, при мысленном
повторении слов (Baddele�, 2003), доказывает, что «фонологическая петля» ра-
бочей памяти, больше не является отдельной когнитивной функцией, а рассма-
тривается в виде беззвучной активности речевой зоны коры. Похожие резуль-
таты (Kossl�n et al., 2004) показали, что при создании мысленных зрительных
образов участвует совокупность областей коры головного мозга, вовлеченных в
нормальную зрительную перцепцию. Более удивителен тот факт, что внимание
к зрительным стимулам очень тесно связано с контролем движений глаз. Спор-
тсмены и музыканты используют сенсомоторные области коры головного мозга
для осуществления мысленных тренировок. Таким образом, во «внутренние» и
«внешние» процессы вовлечены перекрывающиеся области мозга, что значи-
тельно облегчает понимание вышеизложенных фактов.
В когнитивной нейробиологии далеко не всегда существует возможность
упрощенно изложить факты. Авторы смогли организовать данный текст на
основе повторения тем, благодаря чему материал стал легче для понимания и
преподавания и позволяет нам исследовать широкий круг основных проблем,
включающих эмоции, социальный разум и развитие.
Для углубления знаний студентов разработаны дополнительные материалы.
Они представлены с помощью наглядных пособий, изображений и обучающих
18
Предисловие
схем, а также в виде презентаций формата PowerPoint и видеоклипов. Наилуч-
шим образом проиллюстрировать ряд феноменов можно на примере экспери-
ментов и с помощью видеоматериала. Например, можно показать пациента с
синдромом окружения (псевдокома), который способен общаться с окружаю-
щим миром только при помощи движения глаз по виртуальной клавиатуре. Для
сравнения можно продемонстрировать пациента, который выглядит аналогич-
но, но находится в настоящей коме.
Для облегчения процесса обучения в конце каждой главы приводятся во-
просы для повторения и графические упражнения. Мы особенно рекомендуем
пользоваться такими упражнениями, так как самостоятельное создание слож-
ных трехмерных моделей организации мозга способствует лучшему их пони-
манию.
В книге представлен ряд актуальных тем о постоянно меняющихся пер-
спективах когнитивной нейронауки. Одной из них является взаимоотношение
«разума», такого как мы его осознаем, и «мозга», такого как мы его наблюда-
ем, т. е. исторически сложившаяся проблематика о сознании и его мозговых
коррелятах. Как недавно сказал Алан Бэдли (Alan Baddele�), «пожалуй, самым
значительным изменением за последние двадцать лет в области психологии и
когнитивной науки...можно назвать принятие того, что сознание является за-
конной и разрешимой научной проблемой».
Новый взгляд на проблему сознания внес изменения в исследования процес-
сов восприятия, памяти и внимания, которые можно наблюдать в пионерских
работах Енделя Тульвинга (Endel Tulving), Дэниела Шахтера (Daniel �hachter),
Джеральда Эдельмана (Gerald Edelman), Френсиса Крика (Francis Cric�), Кри-
стофа Коха (Chrisnof Koch) и других. В то время как во многих других учебни-
ках имеется только одна отдельная глава про сознание, мы считаем, что к этой
широкой теме надо постоянно возвращаться по ходу всего учебника. Как было
написано в юбилейной 125�й статье в журнале �cience, проблема сознания счи-
тается в настоящее время одной из самых главных, неразрешенных проблем
биологии. Несмотря на то что многое в этом вопросе еще необходимо понять,
психологи уже давно изучают процессы сознания в контексте таких проблем,
как «эксплицитное сознание» и «фокусное внимание». Эти понятия оценивают-
ся исходя из точного поведенческого ответа и с самого начала развития психо-
физики, т. е. более двух сотен лет назад, сигнализируют о наличии когнитивных
процессов в мозге. Таким образом, термин «сознание» может быть использован
как ярлык, аналогичный «памяти» и «восприятию», с определенным числом
подтем, таких как подсознательное восприятие, автобиографическая память и
селективное внимание.
Произвольный контроль функций и деятельности тоже перемещается в ис-
следованиях на передний план, но иногда под названиями «стратегический
контроль» и «высшие психические функции». В мозге произвольные и непро-
извольные функции могут быть четко разделены на основании данных анато-
мии и физиологии. Значительные различия возникают также при визуализации
функций мозга и поведения. И наконец, изучение высших психических функ-
Предисловие
19
ций открывает новое понимание «личности» в повседневной жизни, что изуча-
ется социальной и личностной психологией.
Все эти разделы показывают поразительную взаимозависимость данных о
поведении и работе мозга.
На сегодняшний день механизмы возникновения эмоций и социального по-
ведения также находятся в развитии. «Зеркальные нейроны» связывают с воз-
можностью воспринимать намерения других; бессознательные «лица угрозы»
могут стимулировать миндалину; и противоположные проявления самокон-
троля, вероятно, приводят к возникновению конкурирующих импульсов в пре-
фронтальной коре.
Когнитивная нейробиология расположена на передовом крае науки и пред-
ставляет интерес для многих ученых. Наградой студентам за старания при ее
изучении будет более глубокое понимание человеческой природы, которая ни-
когда не была более понятной и убедительной, чем сейчас.
Авторы выражают огромную благодарность доктору Йоханесу Мензелу
(�ohannes Menzel), редактору издательства Elsevier, за предложенные решения и
помощь в разработке структуры книги. Мы очень признательны Кларе Каруана
(Clare Caruana), редактору по развитию отдела книг по биологическим наукам,
Academic Press, Elsevier, за ее руководство и поддержку на протяжении работы
над вторым изданием. Во время подготовки текста Йоханес и Клара занялись
новыми и более сложными проектами, и мы приняли в наш коллектив Май-
ка Хейли (Mica Hale�), старшего рецензента издательства, и Мелиссу Тернер
(Melissa Turner), редактора по развитию. Майк и Мелисса начали работу над
вторым изданием и с легкостью провели нас через сложные стадии подготовки
этого огромного текста. Мы ценим их бесконечную доброту, советы и поддерж-
ку, оказанные на всем протяжении этого трудного периода. Более того, данная
работа не была бы закончена без использования обширных архивных ресурсов
издательства.
Мы благодарны многим нашим сотрудникам, которые поделились своими
знаниями и опытом. Среди них:
Даниэла Бальслев (Daniela Balslev)
Датский исследовательский центр магнитного резонанса, Госпиталь Видовре,
Дания
Дмитрий Боугаков (Dmitri Bouga�ov)
Факультет неврологии, Медицинский университет Нью�Йорка, Нью�Йорк,
Нью�Йорк, США
Джейсон М. Чейн (�ason M. Chein)
Факультет психологии, Темпл Университет, Филадельфия, Пенсильвания, США
Мелани Кун (Melanie Cohn)
Факультет психологии, университет Торонто, Онтарио, Канада
Элконон Голдберг (El�honon Goldberg)
20
Предисловие
Факультет неврологии, Медицинский университет Нью�Йорка, Нью�Йорк,
Нью�Йорк, США
Марк Х. Джонсон (Mar� H. �ohnson)
Центр мозга и когнитивного развития, факультет психологии, Биркбек колледж,
Лондон, Великобритания
Катарина МакГоверн (Katharine McGovern)
Калифорнийский институт интегративных исследований, Сан Франциско, Ка-
лифорния, США
Морис Московитч (Morris Moscovitch)
Факультет психологии, университет Торонто, Торонто и Бейкрест�центр гериа-
трической помощи, исследовательский институт Ротмана, Северный Йорк, Он-
тарио, Канада
Олаф Паулсон (Olaf Paulson)
Датский исследовательский центр магнитного резонанса, Госпиталь Видовре,
Дания
Джоэл Пирсон (�oel Pearson)
Факультет психологии, Университет Вандербилта, Нашвилль, Теннеси, США
Томас Рамсой (Thomas Ramso�)
Датский исследовательский центр магнитного резонанса, Госпиталь Видовре,
Дания
Дебора Талми (Deborah Talmi)
Факультет психологии, университет Торонто, Торонто, Онтарио, Канада
Франк Тонг (Fran� Tong)
Факультет психологи, Университет Вандербилта, Нашвилль, Теннеси, США
Первый редактор выражает огромную благодарность доктору Джеральду
Эдельману (Gerald Edelman) и его коллегам из Института Нейронауки в Сан�
Диего, так как они являются уникальным источником вдохновения и приме-
ром для ученых во многих областях нейронауки. Автор очень признателен
сотрудникам Фонда наук о разуме в г. Сан�Антонио штата Техас за поддерж-
ку первопроходческих исследований в области когнитивной нейронауки и
сознания. Глава и исполнительный директор Фонда Джозеф Дайел (�oseph
Dial) особенно активно участвует в работе по объединению поведенческих
подходов и методик исследования мозга для изучения сознания (http://www.
mindscience.org).
Обрести более глубокое понимание работы мозга нам помогали коллеги
и друзья. Их слишком много, чтобы перечислять здесь всех, но мы хотим,
чтобы они знали о нашей признательности. Стэн Франклин (�tan Fran�lin),
Уолтер Фримен (Walter Freeman), Уильям П. Бэнкс (William P. Ban�s), Е. Р. Джон
(E.R. �ohn), Кристоф Кох (Christof Koch), Фрэнсис Крик (Francis Cric�), Карл
Предисловие
21
Прайбрэм (Karl Pribram), Дэн Дэннет (Dan Dennett), Патриция Ч�рчленд (Patri�
Karl Pribram), Дэн Дэннет (Dan Dennett), Патриция Ч�рчленд (Patri�
Pribram), Дэн Дэннет (Dan Dennett), Патриция Ч�рчленд (Patri�
Pribram), Дэн Дэннет (Dan Dennett), Патриция Ч�рчленд (Patri�
, Дэн Дэннет (Dan Dennett), Патриция Ч�рчленд (Patri�
Dan Dennett), Патриция Ч�рчленд (Patri�
Dennett), Патриция Ч�рчленд (Patri�
Dennett), Патриция Ч�рчленд (Patri�
, Патриция Ч�рчленд (Patri�
Patri-
cia Churchland), Патрик Уилкен (Patric� Wil�en), Джерейнт Рис (Geraint Rees),
Крис Фрит (Chris Frith),Стэн Дехэйн (�tan Dehaene), Бь�рн Меркер (Bjorn Mer�
�er), Йаак Панксэпп (�aa� Pan�sepp), Сту Хаммерофф (�tu Hammeroff), Томас
Рамсой (Thomas Ramsø�), Антти Ревонсуо (Antti Revonsuo), Анри Монтандон
(Henr� Montandon), Марри Шанахан (Murra� �hanahan) и многие другие очень
помогли нам при подготовке глав. Мы не всегда соглашались с их замечания-
ми, но всегда были благодарны за неугасающий интерес этих людей к работе и
ценные замечания.
Нам очень повезло: эта книга многое потеряла бы без глубоких знаний на-
ших замечательных соавторов и их желания поделиться ими со всеми. В содер-
жании книги отражен вклад в работу каждого из них.
В более широком смысле данная книга обязана своим выходом всему сооб-
ществу преподавателей и ученых, которых мы цитировали по ходу изложения.
Первый редактор хочет выразить признательность своим близким за их тер-
пение, проявленное во время нашей работы над этим всепоглощающим про-
ектом. Его родители, ныне покойные, до сих пор являются для него источником
вдохновения. Он также благодарит доктора Барбару Колтон (Barbara Colton) за
отзывчивость и поддержку.
На последних этапах написания книги неизмеримо большой оказалась по-
мощь Блейра Дэвиса (Blair Davis), а также Шона Фу (�hawn Fu), любезно пре-
доставившего нам значительное число великолепных иллюстраций мозга.
Второй редактор выражает особую благодарность первому редактору за то,
что он щедро делился своими представлениями и энтузиазмом по поводу этого
проекта, сотрудничество и работа над которым начались несколько лет назад
во время трехчасовой беседы в Чикаго и переросли потом в крепкую дружбу и
полноценный проект. Она также признательна всем своим коллегам и друзьям,
с которыми изучала процессы формирования речи в мозге. Их так много, что
трудно перечислить, но она признательна им всем: Грегу Хикоку (Greg Hic�o�),
Дэвиду П�пплу (David Poeppel), Ларри Кахиллу (�arr� Cahill), Норму Вайнбер�
�arr� Cahill), Норму Вайнбер�
Cahill), Норму Вайнбер�
Cahill), Норму Вайнбер�
, Норму Вайнбер-
геру (Norm Weinberger), Брине Сигал (Br�na �iegal), Анне Спэнс (Anne �pance)
и Коурошу Сабери (Kourosh �aberi) — за замечательные беседы, продолжи-
тельные дискуссии и вдумчивые обсуждения в течение многих лет.
Второй редактор выражает глубокую признательность всем членам своей
семьи за их постоянную поддержку во время работы над этим проектом. И на-
конец, она благодарит Кима за его понимание и любовь.
. . . мо зг, и только мозг порождает у нас удовольствие, радость, смех и шут-
ки, так же как печаль, страдание, горе и слезы. Посредством мозга мы ду-
маем, видим, слышим и отличаем безобразное от прекрасного, плохое от хо-
рошего, приятное от неприятного... когда мозг молчит, человек способен как
следует размышлять.
Приписывается Гиппократу, V век до н.э .
(цит. по: Kandel et.al., 1991).
Наверху слева: тело человека и основные плоскости сечений. Внизу слева:
обычный вид мозга с левой стороны. Левое полушарие «смотрит» влево. Го-
лубая область в задней части мозга является затылочной долей. Диаграмма в
нижней правой части рисунка иллюстрирует «иерархию нейронов», упрощен-
ный способ демонстрации нейрональных связей в коре. А на верхней правой
части рисунка вы можете рассмотреть «круговую иерархию». Желтая стрелка
в центре круга обозначает наиболее распространенную точку зрения на роль
перцептивного сознания в мозге. (Источник: Drake et al., 2005.)
Содержание
1.0.Введение........................................................23
2.0.Приглашаемвнейропсихологию.....................................24
3.0.Некоторыеисходныеположения.....................................25
3.1.Расстояния:семьпорядковразмерностей..........................25
3.2.Время:десятьпорядковразмерностей.............................28
3.3. Умозаключения — выход за рамки эмпирических данных . . . . . . . . . . . . 30
3.4.Согласуемостьданных .........................................34
3.5.Главныеориентирынаизображенияхмозга........................35
4.0.Немногоисторииипродолжающаясядискуссия........................37
4.1.Психикаимозг................................................40
4.2 . Биологическая природа когнитивной деятельности и эмоций . . . . . . . . . 41
4.3 . Нейронная доктрина Рамон-и-Кахала:
рабочаягипотезанаукиомозге...................................44
4.4.Пьер-ПольБрока:локализацияцентраречи........................47
4.5. Осознаваемые и неосознаваемые психические явления . . . . . . . . . . . . . . 54
5.0.Наукавозвращаетсякпредставлениямосознании......................59
5.1 . Современные методы исследования осознаваемых и неосознаваемых
процессоввмозге .............................................61
5.2.Историческоеразвитиенеостановилось...........................62
6.0.Заключение......................................................64
7.0.Заданияиупражнениякглаве1.....................................66
7.1.Контрольныезадания...........................................66
7.2.Графическиеупражнения.......................................67
1.0 . Введение
Эта глава дает общие представления о когнитивной нейронауке, объединя-
ющей психику и мозг. С точки зрения современных знаний о Вселенной,
мозг — самая сложная структура, с десятками миллиардов нервных клеток,
триллионами межнейронных связей. Мозг реагирует на поступление различ-
ных молекул (допустим, бокал вина) или на внешние стимулы (например,
важная новость). Некоторые процессы в нервной системе протекают за ты-
сячные доли секунды, тогда как другие продолжаются в течение десятиле-
тий. Несмотря на разнообразие рабочих ситуаций, о системе психика–мозг
накопилось много несложных для восприятия базовых фактов, которые будут
рассмотрены в этой книге.
Глава 1
Психика и мозг
24
Глава 1
2.0 .
Приглашаем в нейропсихологию
Когда речь заходит о последней декаде развития когнитивной нейронауки,
трудно избежать определений «впечатляющее», «революционное». Результа-
ты, накопленные за столетие исследований поведения организмов и устройства
мозга, получили убедительное подкрепление благодаря новым технологиям ви-
зуализации мозга, возможности прижизненных наблюдений в реальном време-
ни. Разумеется, это не просто подтверждение уже известного. Регистрация де-
ятельности живого мозга позволила получить новые факты, выдвинуть новые
идеи, поставить новые вопросы. Многие ученые почувствовали, что через про-
пасть между исследованиями психики и мозга переброшен мост. Для проблем,
которые издавна признавались запутанными, теперь удается найти остроумные
решения.
Одновременно с ощущением прогресса значительно расширились наши
знания. Всего лишь десять лет назад специалисты в области поведения вряд
ли усматривали связь между представлениями о когнитивной деятельности
человека, выдающимися достижениями генетики, данными о молекулярном
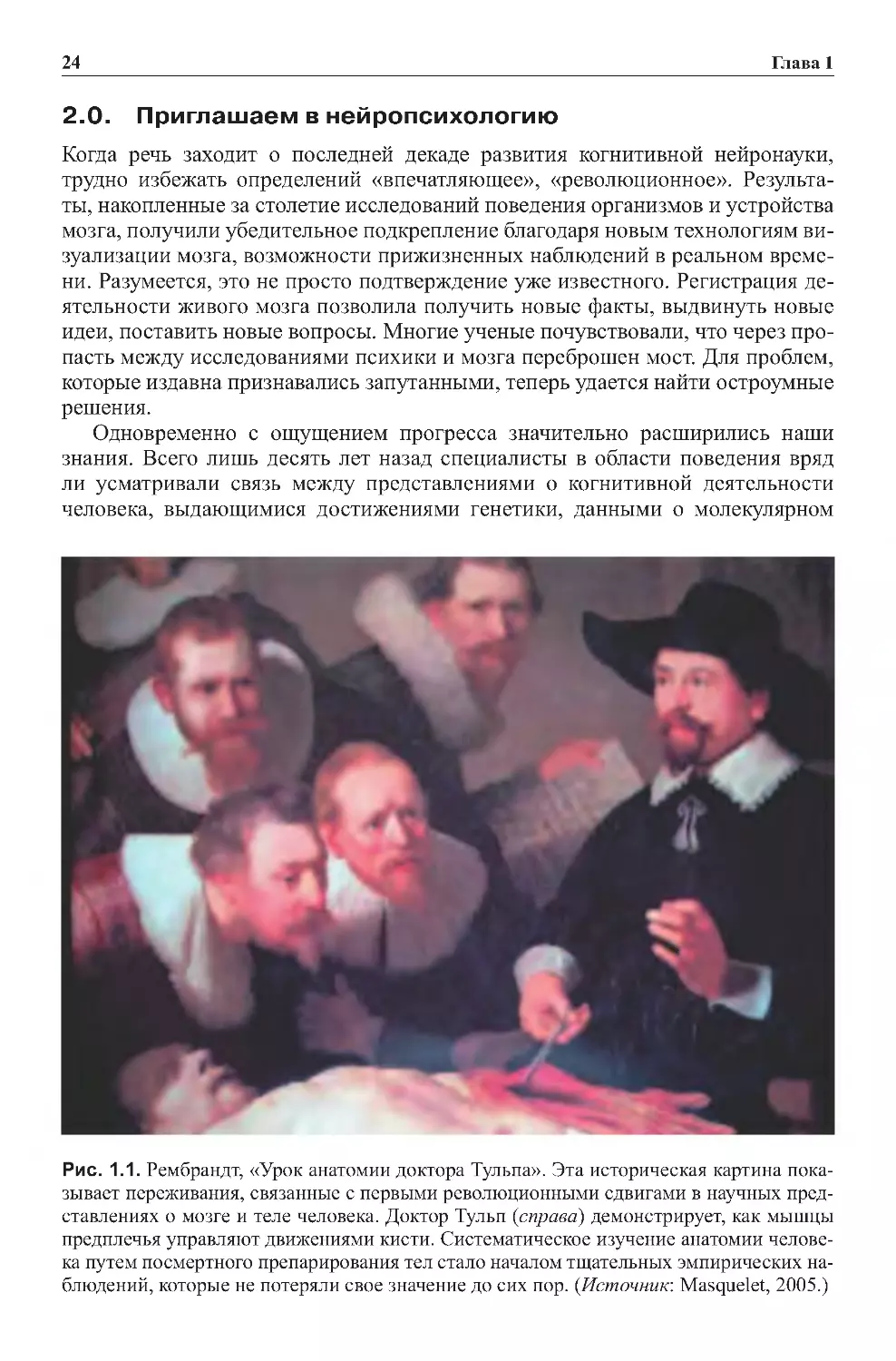
Рис. 1.1. Рембрандт, «Урок анатомии доктора Тульпа». Эта историческая картина пока-
зывает переживания, связанные с первыми революционными сдвигами в научных пред-
ставлениях о мозге и теле человека. Доктор Тульп (справа) демонстрирует, как мышцы
предплечья управляют движениями кисти. Систематическое изучение анатомии челове-
ка путем посмертного препарирования тел стало началом тщательных эмпирических на-
блюдений, которые не потеряли свое значение до сих пор. (Источник: Masquelet, 2005.)
Психика и мозг
25
составе мозга или математическими моделями нейронных сетей. Сейчас эти
направления составляют континуум целой области знания. В мозге выявлены
вероятные корреляты таких явлений, как сознательный опыт, неосознаваемые
процессы, мысленные образы, произвольное управление, которые раньше оста-
вались обойденными, — интуиция, эмоции и даже «Я» (самость). Некоторые
загадки оказались более трудными, чем можно было предполагать, например
природа формирования постоянной памяти. Неразрывность психологического
и нейробиологического подходов к исследованию восприятия, памяти и языка
стала более очевидной, чем когда-либо прежде.
В некоторых случаях результаты изучения мозга помогают найти ключ к во-
просам, над которыми психологи безуспешно трудились в течение десятилетий.
В частности, шли споры вокруг проблемы «раннего» и «позднего» выбора при
восприятии информации. Давая описание такого предмета, как кофейная чаш-
ка, наблюдатель может обратить внимание прежде всего на зрительные при-
знаки «нижнего» уровня — цвет, форму, местоположение. Или же он выделит
характеристики более высокого уровня: «предмет служит для потребления го-
рячих напитков». Можно найти данные в пользу приоритета как первого, так и
второго принципа выбора. К настоящему времени, после многолетних споров,
нейробиологи показали, что в выборе воспринимаемых признаков участвуют
нейроны практически каждого уровня зрительной системы. Отсюда ответ, об-
суждаемый многими психологами: возможен как ранний, так и поздний выбор.
В целом ряде подобных случаев мы обнаруживаем удивительную конверген-
цию данных о деятельности мозга и о поведении.
3.0 .
Некоторые исходные положения
3.1.
Расстояния: семь порядков размерностей
Чтобы рассуждать о соотношении между психикой и мозгом, полезно узнать, с
какими порядками размерностей здесь приходится иметь дело. Длина мозга от
начала до конца составляет около одной седьмой метра. Для сравнения — шаг
человека равен примерно одному метру. Если перейти к размерностям следую-
щего порядка, т. е. 10 м, то это приблизительная длина классной комнаты. Сто
метров — стандартная спринтерская дистанция, а 1000 метров (1 км) — длина
обычной городской улицы. Когда мы дойдем до 107 м (10 000 км), это уже будет
расстояние от восточного до западного побережья Северной Америки или от
Парижа до линии экватора в Европе и Африке. Такое расстояние соответствует
десяти миллионам шагов. Чтобы представить себе размерности структур мозга,
нам придется следовать в обратном направлении — от 1 м до 10–7
м (табл. 1 .1).
С точки зрения размеров структур и сложности организации, подобный диа-
пазон поистине смущает.
Зрительно воспринимаемые действия человека измеряются расстояниями
примерно от одного сантиметра и более. Передвижение пальца при нажатии на
клавишу компьютера составляет не более нескольких сантиметров. Во время
речи наш язык смещается на 1–2 см. Длина одного шага — примерно 1 м. Рост
большинства людей не превышает двух метров, а протяженность самых длинных
отростков нервных клеток в организме человека может составлять около 1 м.
26
Глава 1
Таблица 1.1
Расстояния: от 10
–7
м до 1 м
1. Алкоголь вызывает в мозге изменения; размер молекул этилового спирта составляет
~10
–7
м (0,0000001) (рис. 1 .2).
2. После приема таблетки транквилизатора изменяется концентрация химического
нейромедиатора в синаптической щели некоторых синапсов мозга; ширина синап-
тической щели равна ~ 10
– 6 м (одной миллионной метра, т. е. одному микрону).
3. Тело нервной клетки примерно в 100 раз больше (~ 10
–4
), чем ширина синаптиче-
ской щели.
4. Учтем, что нейроны работают не поодиночке, а группами, размеры которых иногда
в десятки раз больше, чем размеры одного нейрона. Диаметр небольших колонок
нейронов в коре мозга составляет ~ 10
– 3 м (1 мм). Таким образом, мы подошли к
размерности одна тысячная метра — подобные структуры мы еще можем разли-
чить невооруженным глазом.
5. Размеры карт мозга могут быть достаточно велики (например, карта зрительной
системы). Первая зрительная область коры мозга, V1, сравнима по площади с кре-
дитной карточкой, т. е. 10
–2м2.
6. Зрительные области мозга работают не в изоляции друг от друга. Они входят в
состав обширных нейронных сетей, которые активируются при постоянном ди-
намическом взаимодействии. Размер активированной нейронной сети составляет
~10
– 1 см; такой участок можно различить невооруженным глазом благодаря мето-
дам прижизненных наблюдений. Современные методы позволяют наблюдать не-
прерывно изменяющуюся активность мозга, направляя луч лазера через оголенную
поверхность головы крысы или других мелких млекопитающих. В активном участ-
ке мозга заметен усиленный ток крови в сосудах, так же как это происходит в коже
человека во время быстрого бега.
3.1.1. Молекулярный уровень: нейрохимические соединения
Молекулы нейромедиаторов, имеющие различные размеры, проникают путем
диффузии через синаптическую щель (ее ширина от 25 нм до 100 мкм) между
нейронами (Iversen, 2004). В большинстве случаев влияние химических со-
единений на мозг обусловлено тем, что они могут усиливать или ослаблять
межнейронные молекулярные коммуникации. В бесконечный перечень ней-
роактивных веществ входят никотин, алкоголь, кислород воздуха, токсичные
газы (например, монооксид углерода), поступающая из печени глюкоза, саха-
роза пищевых продуктов, шоколад, кофе, нейротоксины (например, свинец) и
огромный список лекарственных средств (рис. 1 .3). Роль таких молекул в по-
вседневной жизни нельзя недооценивать.
Молекулярные посредники (мессенджеры) в мозге подразделяются на две
большие группы. К первой группе относятся нейромодуляторы. Подобные
соединения продуцируются мелкими скоплениями нейронов, находящихся в
основании мозга. Аксоны этих нервных клеток образуют диффузные проек-
ции большой протяженности, так что вещества, выделяемые из их окончаний,
распространяются по обширным участкам переднего мозга. Благодаря такому
«распылению» нейрохимических соединений осуществляется модуляция круп-
ных отделов центральной нервной системы. Наряду с диффузным влиянием
нейромодуляторы могут быть и посредниками местных эффектов в результате
взаимодействия со специфическими молекулярными рецепторами. Например,
Психика и мозг
27
дофамин широко распространяется в мозге, однако его рецепторы типа D1/D2
в лобной коре обеспечивают местное влияние, связанное с оперативной памя-
тью (����s and D��s��s�t�, 2006). Таким образом, нейромодулятор дофамин мо-
(����s and D��s��s�t�, 2006). Таким образом, нейромодулятор дофамин мо-
Таким образом, нейромодулятор дофамин мо-
жет оказывать не только диффузное воздействие, но и местные эффекты, когда
он поступает непосредственно к специфическим рецепторам в определенной
области мозга.
Вторую группу составляют нейромедиаторы, обладающие более «адрес-
ным», местным действием. Среди них есть пептиды — небольшие субъедини-
цы белковых молекул, секретируемые в синаптическую щель. К настоящему
времени в мозге выявлено более 40 типов пептидов. Однако особенно хорошо
известны два вещества непептидной природы — глутамат (возбуждающий ней-
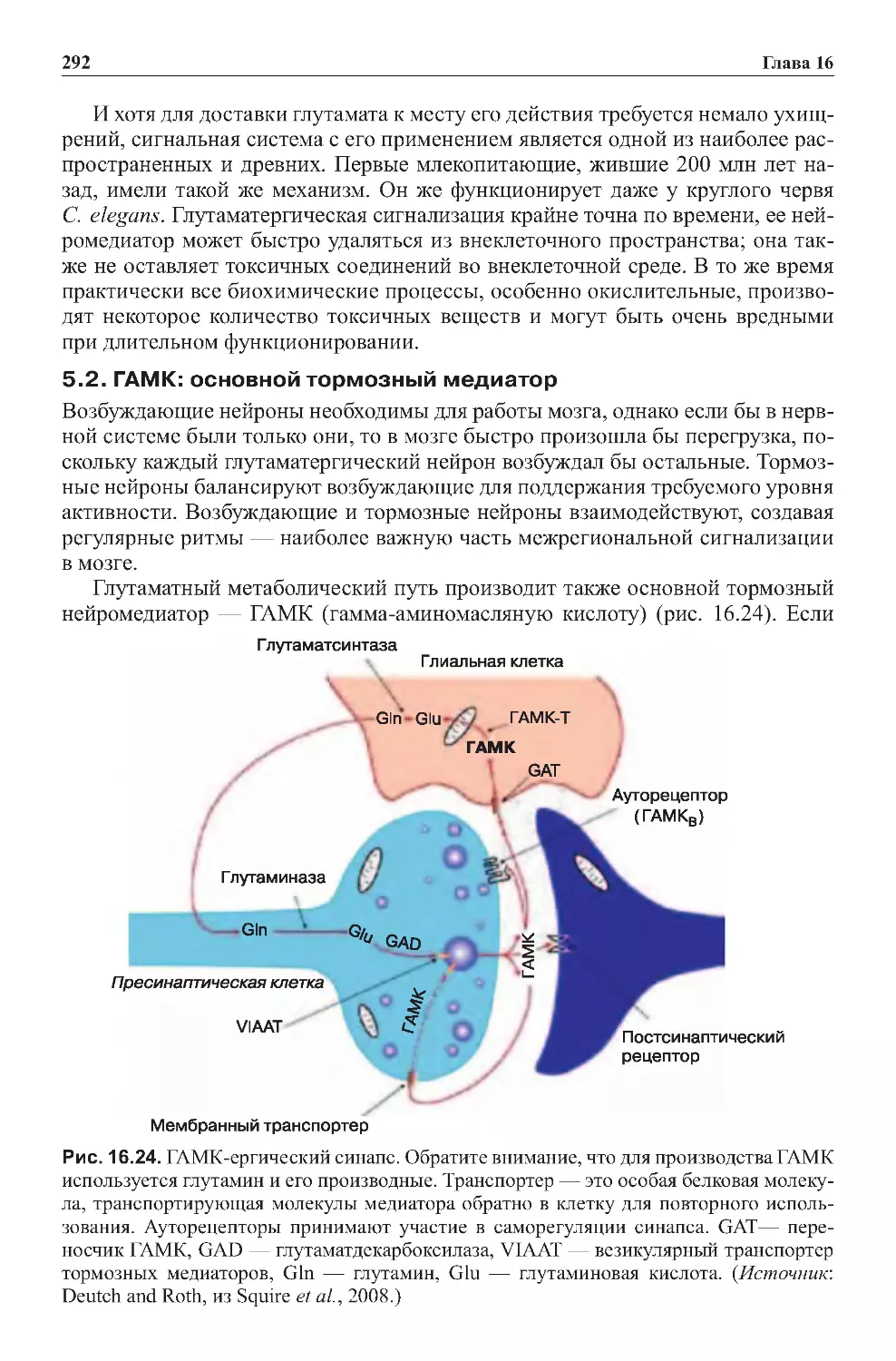
ромедиатор, характерный для коры мозга) и ГАМК (наиболее распространен-
ный тормозный нейромедиатор).
Рис. 1 .2. Порядки пространственных размерностей. (а) Изображение мозга человека
при интенсивной стимуляции зрительной коры (затылочной области мозга) путем вра-
щения черно-белых фигур. (б) Изображение среза вдоль средней линии мозга. Черным
обозначена область V1 (от англ. vision) — первый участок коры, куда приходит зритель-
ный путь. Эта область по размерам примерно соответствует кредитной карточке. (в) Го -
лова плодовой мушки. Мозг этого насекомого содержит ~ 1 000 000 нейронов. (г) Ин-
дивидуальный нейрон. Размеры нейронов различны, но все они очень малы. В мозге
человека находятся десятки миллиардов нервных клеток. (д) Структура молекулы до-
фамина. Дофамин необходим для рабочей памяти, ощущения удовольствия, управления
мышцами. Одним из последствий недостаточности дофамина является болезнь Паркин-
сона. На пути от (а) до (д) размеры компонентов мозга изменяются примерно на семь
порядков (в 107 раз)
28
Глава 1
Научные успехи определяются новыми техническими достижениями, в том
числе расширением размерностей для наблюдения. В последнее время мы
стали свидетелями череды открытий, связанных с новыми методами прижиз-
ненных исследований мозга. Благодаря тому что изображения объектов теперь
возможно увеличивать примерно на семь порядков, для нейропсихологии от-
крылись новые горизонты.
3.2 .
Время: десять порядков размерностей
Шкала времени для функций человеческого организма имеет громадный диа-
пазон (табл. 1 .2). Что касается поведения, то счет идет на десятые доли секунды
(100 мс). Время самой быстрой (простой) реакции на стимул равно ~ 100 мс,
тогда как время, которое требуется, чтобы сенсорный стимул достиг сознания,
обычно составляет уже несколько сотен миллисекунд. Это допустимо в той сре-
де, где возник и развивался человеческий вид. Если же при встрече с голодным
хищником ваша реакция продлится несколько секунд, вы обеспечите ему вкус-
ную, богатую белком трапезу и потеряете шанс на биологическое воспроиз-
ведение. Однако, постаравшись отреагировать в течение 10 мс (т. е. за одну
сотую долю секунды), вы бы заставили свой мозг работать с такой высокой
скоростью, при которой он не успевает интегрировать зрительные и слуховые
сигналы о приближении тигра. Следовательно, вы не смогли бы определить,
с какой стороны появится хищник. Итак, реальная средняя скорость реакции —
это 100 мс.
Рис. 1.3 . Низкомолекулярные вещества оказывают влияние на мозг. Маленькая моле-
кула закиси азота (N2O) способна изменить специфические функции мозга. Поведение
людей в такой ситуации оказалось большой неожиданностью для врачей в начале XIX
века (приведенный здесь рисунок был сделан студентом-медиком в 1808 г.). Подобные
факты продолжают удивлять и до сих пор. Оксид азота (NO), при вдыхании которо-
го развивается токсический эффект, в чрезвычайно малых количествах присутствует
в организме и является важным нейромедиатором. Препарат Виагра, применяемый в
качестве стимулятора эректильной функции, улучшает опосредуемую NO передачу сиг-
налов в стенках кровеносных сосудов пениса. (Источник: Adelman and Sm�th, 2004.)
Психика и мозг
29
Таблица 1.2
Время: десять порядков размерностей, от нескольких лет до миллисекунд
1. Когда вы слышите высокий музыкальный тон, частота импульсов в слуховом нерве
соответствует частоте звуковых волн, вызывающих вибрацию барабанной перепон-
ки уха — до 1000 волн в секунду, или 1 волна в миллисекунду (напомним: 1 цикл в
секунду = 1 Гц).
2. Частота импульсных нейронных разрядов может достигать 1000 Гц, хотя большин-
ство нейронов коры мозга генерируют импульсы с частотой ~ 10 в секунду.
3. Слуховой нерв — это пучок аксонов, каждый из которых дает разряд с частотой
ниже 1000 импульсов в секунду, но в совокупности они могут обеспечивать ско-
рость реакции в пределах миллисекунд. Некоторые сенсорные явления протекают
с еще большей скоростью. На пределе своих возможностей наш мозг способен
дифференцировать звуки, поступающие к одному и к другому уху с интервалом
до нескольких микросекунд (то есть миллионных долей секунды). Это в 1000 раз
быстрее, чем самая высокая частота разряда нейрона (см. гл . 7). Однако в контексте
нашего изложения наиболее употребительными единицами измерения малых зна-
чений на шкале времени являются миллисекунды.
4. Для сравнения: если вы будете считать про себя от одного до десяти, это займет
время порядка секунд или десятков секунд. Изучение этой главы потребует у вас
один час или дольше — более 3000 с (сожалею, но это так!).
5. Вернемся к шкале времени. Если во время кинофильма фонограмма задержится на
доли секунды по отношению к видеоряду, вы заметите расхождение между звуками
и мимикой актеров. Асинхронность между звуком и изображением появляется при
разрыве между ними примерно на 100 мс, т. е. на одну десятую секунды. Это еще
одна причина, чтобы выделить десятую долю секунды как особый порядок размер-
ности.
6. Кроме того, 100 мс — это практически самая быстрая реакция на явление, ожидае-
мое в скором времени, например запуск автомобиля при смене сигнала светофора
(время простой реакции). Десятой доле секунды соответствует время одной альфа-
или тета-волны в мозге. (Диапазон частот альфа-волн составляет 8–12 Гц, тета-волн
—
5–7 Гц.) К такому же порядку размерностей относится период времени, необхо-
димый для сознательного восприятия сенсорного стимула.
В мозге протекают параллельно процессы разных временных и простран-
ственных размерностей, подобно элементам симфонии — нотам, фразам,
целым фрагментам. Слушая песню, вы улавливаете в каждый момент лишь
несколько нот, но в процессе их поступления складываются более крупные
когнитивные структуры, так что вы воспринимаете мелодию. Во время показа
фильма частота кадров должна составлять 24 изображения в секунду, т. е. при-
мерно 40 мс на кинокадр — тогда движения будут слитными. При снижении
частоты кадров ниже 24 Гц получаются прерывистые движения, как в ранних
немых фильмах. Однако сюжет развивается в течение минут или часов. Разо-
блачение преступника в конце детективного фильма теряет смысл, если зри-
тель не помнит начало преступления. Следовательно, для восприятия сюжета
фильма необходима интеграция когнитивных процессов на протяжении многих
минут, так что приходится каким-то образом сочетать разные шкалы времени.
Каждый из этих процессов имеет свою структуру и диапазон размерностей вре-
мени. Мозг параллельно отслеживает процессы различной размерности.
30
Глава 1
Что касается более длительных периодов времени, то для овладения слож-
ными навыками (например, лыжным спортом или игрой на гитаре) могут по-
требоваться годы. Дети обучаются говорить за несколько лет, а у взрослых
структура базовых характеристик личности консервативна и поддерживается
десятилетиями. Такая продолжительность зависит от событий, протекающих в
мозге в течение периодов порядка 100 мс. Итак, что касается времени, мы име-
ем дело примерно с десятью порядками размерностей, от одной тысячной доли
секунды (т. е. одной миллисекунды) для разряда индивидуального нейрона до
100 000 с (дни) и десяти миллионов секунд (годы).
3.3 .
Умозаключения — вы ход за рамки
эмпирических данных
Наука складывается как непрерывный ряд умозаключений, переход от эм-
пирических наблюдений к концепциям. Тысячи лет назад люди начали заду-
мываться, что представляют собой небесные светила (солнце, луна, звезды);
некоторые из них перемещались предсказуемо, другие — нет. Греки назвали
«странников» ночного неба планетами. Эти странствующие светила очаровы-
вали и изумляли. Только в XVII в. люди стали понимать и предсказывать пути
движения небесных светил. Решение загадки заключалось в осознании того,
что планеты — это гигантские шарообразные объекты наподобие Земли, ко-
торые вращаются по орбите вокруг самого большого из них, Солнца. Споры
и наблюдения продолжались сотни лет, прежде чем удалось прийти к такому
выводу. Лишь исчисление бесконечно малых величин, разработанное Исааком
Ньютоном, позволило свести споры к простому уравнению. А именно орбиты
планет можно предсказывать исходя из того, что гравитационная сила притя-
жения пропорциональна массе планеты и ее ускорению. Обратим внимание:
понятия «солнце», «планета», «сила», «гравитация» представляют собой умо-
заключения. Эти понятия далеко ушли от первых наблюдений за небесными
светилами (рис. 1 .4), однако именно с помощью таких концепций можно объ-
яснить эмпирические факты.
Вся наука строится на тщательных наблюдениях и концептуальных умоза-
ключениях. Схема объяснения, называемая «номологической сетью», отражает
категориальные концепции и соотношения между ними. В итоге мы понимаем,
в чем состоит природа явления. Кстати, научные выводы позволяют делать бо-
лее точные прогнозы, а применение полученного знания иногда кардинально
меняет жизнь. И все начинается с точных фактов и умозаключений, построен-
ных на этой основе.
Эти базовые идеи имеют непосредственное отношение к когнитивной ней-
ронауке. Когда мы говорим о когнитивных (познавательных) процессах — речи,
научении, зрении — мы тоже используем концептуальные умозаключения, кото-
рые должны быть прочно связаны с надежными наблюдениями. Например, объ-
ем минимальной кратковременной памяти («мгновенной», «непосредственной»,
«сенсорной» — информация, которую мы в состоянии сразу мысленно повто-
рить) равна примерно семи плюс/минус два, как указал Джордж Миллер в зна-
менитой статье «Магическое число семь плюс/минус два» (1956). Очевидно, это
число применимо ко многим элементам: цвету, количеству предметов, коротким
словам, музыкальным нотам, единицам оценочных шкал и т. д. Согласно более
Психика и мозг
31
новым сведениям, действительный объем мгновенной памяти даже меньше семи,
примерно четыре разных элемента (C�wan, 2001). Однако особенно важным яв-
ляется необычайное постоянство фактов. Попробуйте, например, перечислить
десять различных продуктов из списка, составленного вами для посещения ма-
газина; вы вспомните только примерно семь продуктов, а если одновременно вы
еще заняты другими мыслями, названное вами число уменьшится до четырех.
Это удивительно узкие границы, если учесть достаточно большие размеры мозга.
Существует лишь несколько основных условий для оценки объема опера-
тивной памяти. Во-первых, каждый элемент информации должен быть предъ-
явлен на очень короткий период (порядка нескольких секунд), иначе он пере-
йдет в постоянную память. Во-вторых, элементы информации должны быть
непредсказуемыми с позиций уже имеющегося знания. Если мы попросим че-
ловека запомнить упорядоченную последовательность чисел, например 0, 5, 10,
15, 20, 25..., ему будет достаточно запомнить правило формирования последо-
вательности, и тогда покажется, что объем оперативной памяти безграничен.
Такие понятия когнитивной науки, как «оперативная память», созданы в ходе
Рис. 1 .4 . Как строятся умозаключения о светилах в ночном небе? Для наблюдателя,
находящегося на Земле, планеты выглядят как источники света, которые перемещаются
в ночном небе. После многих лет тщательных астрономических наблюдений Исаак
Ньютон и другие ученые пришли к достаточно простым объяснениям путей движения
планет. Переход от эмпирических наблюдений к концептуальным умозаключениям –
решающий этап построения научного знания
32
Глава 1
многолетних экспериментальных наблюдений, результаты которых были на-
столько убедительными, что их удалось обобщить в виде единой концепции
(рис. 1 .5).
Понятия, подобные концепции оперативной памяти, оказались полезными,
однако вполне возможно, что в ближайшем будущем для тех же явлений бу-
дут найдены более подходящие объяснения. Все концептуальные умозаключе-
ния носят в известной степени предположительный характер. Представления
Ньютона о силе тяжести господствовали в физике в течение трех веков, а затем
Эйнштейн предложил иной взгляд на те же факты. Научные концепции — не
метафизические догмы. Они могут быть пересмотрены в любой момент.
Когнитивная нейронаука тоже основывается на умозаключениях по резуль-
татам эмпирических наблюдений. При сканировании мозга получаются изобра-
жения, похожие на физические объекты, которые можно видеть и трогать, поэ-
тому мы полагаем, что именно такова реальность. Однако это лишь заманчивое
заблуждение. Результаты электроэнцефалографии (ЭЭГ) представляют собой
умозрительную оценку активности мозга, так же как и данные функциональной
магнитно-резонансной томографии (фМРТ), позитронной эмиссионной томо-
графии (ПЭТ) и других исследований современными высокотехничными мето-
дами (блок 1.1). Даже при регистрации активности индивидуальных нейронов
мы получаем крошечный участок разряда единственной клетки из десятков
Рис. 1 .5 . Когнитивные концепции основаны на систематических наблюдениях за по-
ведением. Например, концепция «рабочей памяти» не возникла в готовом виде. Такого
рода идеи созревают в результате многолетних исследований, когда умозаключение мо-
жет стать объяснением многих фактов. Концепция рабочей памяти была предложена
в 1974 г. после двух десятилетий изучения кратковременной памяти. Сейчас это пред-
ставление расширилось и охватывает зрительное, вербальное и другие виды восприятия
Психика и мозг
33
миллиардов других. Нейроны образуют десятки тысяч связей; есть сведения,
что обработка информации может осуществляться уже на уровне входных кле-
точных отростков нейрона (дендритов) (Alle and �e�ger, 2006). Таким образом,
электрическая активность индивидуальных нейронов — это всего лишь очень
маленький компонент сложнейшего динамического ансамбля перемещений мо-
лекул и электромагнитных потоков. Новые методы визуализации чрезвычайно
полезны, но картина работы мозга, которая строится на их основе, тоже содер-
жит умозаключения.
Однако нам все равно приходится прибегать к некоторым допущениям —
таков процесс развития науки. Эти моменты важно учитывать и быть готовыми
Блок 1 .1 . П рижизненные исследования мозга
Десять–двадцать лет назад сама мысль о возможности наблюдать работу живого моз-
га казалась невероятной. Для решения труднейшей задачи регистрации активности
мозга были привлечены методы физики и молекулярной биологии. В идеале метод
должен мониторировать десятки миллиардов нейронов, регистрируя активность
каждой клетки тысячи раз в секунду. Кроме того, необходимо отслеживать непре-
рывное динамическое взаимодействие мелких и крупных групп нейронов, в котором
участвуют триллионы межнейронных связей. По аналогии — идеальный спутник-
шпион, который вращается по околоземной орбите, должен был бы видеть каждого
человека и регистрировать динамику отношений между отдельными личностями и
группами, от семей до целых наций.
Такого идеального метода нет. Наши знания о мозге — это своего рода коллаж из
множества фрагментов головоломки, собранных в правдоподобную целостную кар-
тину. Однако обдумывание совершенного метода наблюдения помогает понять, к ка-
ким параметрам нам следует стремиться.
Технология визуализации мозга обеспечила прорыв в когнитивной нейронауке благо-
даря добавлению новых фактов к результатам, полученным ранее методами когни-
тивной психологии, психофизики, поведенческого обусловливания, фундаментальной
нейробиологии. До появления этой технологии наши знания основывались на резуль-
татах экспериментов на животных и отрывочных клинических наблюдениях за людь-
ми с поражениями мозга. Однако наблюдения за больными с травмой мозга не позво-
ляют получить систематических данных; даже простое описание повреждений мозга
обычно основано на посмертном патологоанатомическом исследовании. После травмы
в мозге часто развиваются компенсаторные изменения, со временем клетки погибают
и происходит адаптация, так что данные аутопсии не всегда отражают характер по-
вреждения на момент установления диагноза. Результаты экспериментов на животных
подразумевают гомологию — межвидовое сходство, признаваемое далеко не всеми
учеными. Ни один живой организм, кроме человека, не обладает речью, а также други-
ми особенностями. Таким образом, деятельность мозга очень трудно изучать.
Многие проблемы были разрешены, когда появилась возможность прижизненной ре-
гистрации активности мозга человека, вначале с помощью электроэнцефалографии
(ЭЭГ), затем рентгенографии, позднее посредством компьютерной томографии («по-
слойной» рентгенографической регистрации, от греческого слова «томос» — слой).
В настоящее время мы располагаем более чем десятком методов, которые быстро
совершенствуются, обеспечивая повышение точности и расширение области приме-
нения. Наиболее активно используются ЭЭГ, позитронная эмиссионная томография
(ПЭТ), магнитно-резонансная томография (МРТ), функциональная МРТ (фМРТ) и
магнитная энцефалография (МЭГ). Эти и другие новые методы исследования рабо-
тающего мозга человека подробно описаны в гл. 4 .
34
Глава 1
к их трансформации по мере необходимости. В когнитивной нейронауке мы вы-
водим умозаключения по результатам наблюдений за поведенческими реакци-
ями и активностью мозга (рис. 1 .6). Мы не можем непосредственно наблюдать
«внимание» или «оперативную память». Поэтому так важно оценивать природу
фактов, на которых строятся наши умозаключения.
3.4 .
Согласуемость данных
Когда мифический первый пещерный житель первым указал на звезду в ноч-
ном небе, ему вряд ли поверили его соплеменники. Какие такие огни в небе?
Все знали, что небо — обитель богов. Подобный скептицизм в порядке вещей.
Когда Галилей впервые с помощью примитивного телескопа увидел спутники
Юпитера, некоторые критики отказались смотреть через телескоп, поскольку
считали, что истинную природу вещей можно увидеть только невооруженным
глазом. Скептицизм по-прежнему является нормой, поэтому для верификации
наблюдений в науке используется согласуемость данных. Основополагающие
гипотезы когнитивной нейронауки сейчас проверяются вновь и вновь путем
Рис. 1 .6 . Оценка рабочей памяти мозга происходит путем умозаключений. Рабочая па-
мять изучается путем наблюдений за поведенческими реакциями, регистрации фМРТ,
ЭЭГ, активности индивидуальных нейронов. Каждый из этих методов имеет достоин-
ства и недостатки, но ни один из них не считается «решающим». В принципе показа-
тели рабочей памяти при регистрации активности мозга достаточно хорошо совпадают
с результатами оценки поведения. Когнитивная нейронаука строится на комплексных
данных, но мы должны быть готовы к различным толкованиям существующих теорий
Психика и мозг
35
регистрации активности индивидуальных нейронов, экспериментов на живот-
ных, ЭЭГ, фМРТ, МЭГ, оценок поведения в виде вербальных сообщений и вре-
мени реакции. Никакое единичное исследование не может служить достаточ-
ным обоснованием гипотезы. Любое важное утверждение требует поддержки
многими фактами.
Часть обсуждения всегда касается природы оцениваемого показателя. Такой
вопрос возникает применительно к любому новому методу. Сейчас очень по-
пулярен метод фМРТ. Однако, как мы увидим далее, продолжается дискуссия
о том, что именно оценивают эти методы. То же самое можно сказать и о по-
веденческой оценке оперативной памяти, о регистрации активности индиви-
дуальных нейронов, ЭЭГ и т. д .
3.5 .
Главные ориентиры на изображениях мозга
Как соотносятся активность мозга и когнитивные процессы? Чтобы пояснить
результаты исследований, о которых пойдет речь, будут приведены функци-
ональные изображения мозга. Однако представления о деятельности мозга
всегда основываются на знаниях анатомии, поэтому сначала рассмотрим топо-
графию мозга. На рис. 1 .7 (внизу) показано левое полушарие, вид сбоку. Ввер-
ху — правое полушарие на сагиттальном срезе мозга по средней линии.
На изображениях мозга следует различать главные ориентиры. К особенно
важным среди них относятся крупные складки коры больших полушарий —
поверхностной структуры мозга. Самая большая складка, расположенная по
средней линии между правым и левым полушариями, называется продольной
щелью (fissura longitudinalis). Следующая большая складка проходит по боко-
вой части мозга под углом к средней линии и называется латеральной бороздой
(sulcus lateralis). Она отделяет височную долю от остальной части коры мозга.
Поскольку височная доля всегда «указывает» в направлении глаза, по ней легче
всего определить, в какую сторону обращен мозг. Когда вы смотрите на изобра-
жение мозга, сразу ищите височную долю.
Еще один важный ориентир — мозолистое тело (cоrpus callosum), плотный
пучок нервных волокон в виде моста, соединяющего правое и левое полуша-
рия. На рис. 1 .7 (вверху) мозолистое тело выглядит как изогнутая структура,
которая начинается под лобной корой, поворачивает кверху, потом к затылку
и заканчивается непосредственно перед мозжечком. На срезе мозолистое тело
имеет для невооруженного глаза белый цвет, поскольку состоит из миелинизи-
рованных аксонов, т. е. нервных волокон, покрытых белой липидной миели-
новой оболочкой. Мозолистое тело (т. е. «плотное», «твердое») получило свое
название благодаря тому впечатлению, которое возникло у первых анатомов,
обозначивших основные структуры мозга. Это образование было открыто на
достаточно раннем этапе, потому что его легко обнаружить, осторожно раздви-
нув два полушария. На рис. 1 .9 показан классический рисунок великого худож-
ника эпохи Возрождения Тициана, иллюстратора первой подробной книги по
анатомии, которую написал Андреас Везалий.
И наконец, последний ориентир — центральная борозда (sulcus centralis)
между задней половиной мозга и лобной долей. Задняя область коры больших
полушарий обеспечивает преимущественно сенсорные функции и содержит
центры зрительной, пространственной, слуховой, соматической чувствитель-
36
Глава 1
ности, тогда как в лобной коре находятся двигательные и когнитивные центры.
Центральная борозда служит четкой границей между входными и выходными
областями коры.
Чтобы сориентироваться, нужно найти на изображении мозга прежде всего
эти три главные складки.
Рис. 1 .7 . Вид мозга сбоку и на сагиттальном срезе. Вверху: сагиттальный срез по сред-
ней линии мозга, правое полушарие. Внизу: вид сбоку, левое полушарие. Слева — пе-
редняя часть мозга, справа — задняя. Показаны префронтальная, височная, теменная,
затылочная доли и мозжечок. (Источник: Drake et al., 2005.)
Психика и мозг
37
Вещество коры распространяется вглубь каждого полушария мозга. По-
скольку сложный трехмерный объект трудно воспринимать, воспользуйтесь
рис. 1 .8, на котором показаны три плоскости срезов. Главные доли мозга и дру-
гие важные отделы необходимо знать точно так же, как вы помните названия
континентов на географической карте. На протяжении всей книги мы будем
рассказывать об исследованиях структур мозга, соотнося эти данные с пред-
ставлениями о когнитивной деятельности человека. Знание топографии мозга
поможет интерпретировать результаты нейровизуализации.
4.0 .
Немного истории и продолжающаяся дискуссия
Идея о том, что мозг — источник наших переживаний, возникла очень давно;
Гиппократ высказывался об этом примерно 2500 лет назад. Однако тщательные
исследования мозга и накопление фактов начались только в эпоху Возрождения.
В 1543 г. Андреас Везалий, анатом из Антверпена, впервые опубликовал под-
робный атлас организма человека, включая сюда мозг. До этого времени цер-
ковь и государство считали преступлением производить посмертное вскрытие.
Рис. 1 .8 . Три основные плоскости срезов
мозга. Вверху: сагиттальный срез мозга,
т. е . срез по вертикальной плоскости от
лобной доли кзади до конца мозга. Если
такой срез проходит по средней линии
мозга между двумя полушариями, он на-
зывается срединным сагиттальным. По-
середине: горизонтальный срез мозга.
Внизу: корональный срез (его плоскость
проходит через края воображаемой коро-
ны). (Синонимы этих терминов будут при-
ведены в гл. 5 .)
38
Глава 1
Рис. 1 .9 . Вверху: Андреас Везалий демонстрирует препарат руки (предплечье и кисть).
Бельгийский врач Везалий (1514–1564) совершил переворот в традиционном препо-
давании анатомии, производя посмертные вскрытия человеческих тел. На рисунке он
изображен показывающим препарат кисти и предплечья. Для него такая демонстрация
была особенно важна в связи с представлением о роли божественной руки в мирских де-
лах. Везалий опроверг основанный на библейском сказании об Адаме и Еве миф о том,
что у женщины на одно ребро меньше, чем у мужчины. Вскрытия человеческих тел
в то время не производились, и точные зарисовки были редкостью. В 1543 г. Везалий
опубликовал свою книгу о новых данных по анатомии под названием «О строении че-
ловеческого тела». В том же году появилась книга Коперника «Обращение небесных
тел», которая по-новому представила Солнечную систему. Труды обоих ученых полу-
чили широкую известность, горячо обсуждались и явились вехами в истории науки.
Источник: Masquelet, 1986. Внизу: чернильные зарисовки открытой поверхности моз-
га; авторство приписывается Тициану. Эти иллюстрации в классической книге Везалия
по анатомии объединяют искусство и науку. Обратим внимание, что на правом рисунке
два полушария раздвинуты и между ними видно мозолистое тело — образование, кото-
рое соединяет два полушария и состоит примерно из 100 млн нервных волокон. Первые
анатомы характеризовали эту структуру как плотную, или мозолистую, ткань — отсюда
латинское название corpus callosum (мозолистое тело). (Источник: Squ�re et аl., 2003.)
Психика и мозг
39
Спустя столетие знаменитая картина Рембрандта «Урок анатомии доктора Туль-
па» показывает, с каким изумлением врачи рассматривают подробности стро-
ения человеческого тела (рис. 1 .1). Примерно тогда же (1490–1510) Леонардо
да Винчи сделал зарисовки черепа и тела человека. Людям эпохи Возрождения
был свойствен интерес ко всем проявлениям человеческой природы; в общей
атмосфере любознательности попытки понять устройство мозга становились
более настойчивыми.
Современные исследования уходят своими корнями глубоко в историю. На-
ука о поведении возникла в XIX в. Однако начало подробного изучения мозга
совпадает с истоками современной науки в эпоху Европейского Возрождения,
с появлением трудов Галилея, Коперника, Ньютона и Декарта. В 1665 г. Ньютон
проводит свои эксперименты с призмами, желая понять природу восприятия
цвета. В 1600-е гг. Левенгук и другие ученые изобретают световой микроскоп,
благодаря чему в XIX–XX в. Сантьяго Рамон-и -Кахал сделал свои открытия от-
носительно свойств нервных клеток.
История науки показывает, что чем больше мы знаем, тем яснее нам видны
закономерности полученных фактов. Такого рода обобщения будут представле-
ны на протяжении всей книги.
Связь возникновения науки о мозге с эпохой Возрождения очевидна и в наше
время. В основе терминов по анатомии мозга лежат понятия на латинском язы-
ке — международном языке науки. Мы до сих пор обозначаем доли (lobеs) коры
мозга (cortex cerebri) латинскими названиями: lobus occipitalis, l. temporalis,
l. parietalis, l. frontalis (затылочная, височная, теменная, лобная доли соответ-
ственно). Поскольку ранние исследования проводились невооруженным глазом
или с помощью простых увеличительных конструкций, большинство струк-
тур получали название исходя из первого зрительного впечатления. Отсюда
возникли такие термины, как thalamus — «чертог», amygdala — «миндаль»,
cortex — «наружный слой» и т. д . В названиях практически всех структур мозга
используются обыденные латинские слова. Мы будем упоминать происхожде-
ние каждого нового термина, поскольку это помогает усвоить понятия.
Мозг человека развивался, начиная с его зачатков у древних млекопитаю-
щих, на протяжении более чем 200 млн лет. Он устроен очень сложно, в том
числе его части, которые видны невооруженным глазом. Многие поколения
ученых внесли свой вклад в его изучение. Например, Рене Декарт (рис. 1 .10),
который теперь известен главным образом как математик и философ, вместе с
тем был внимательным исследователем мозга. Одно из своих замечательных
наблюдений он сделал, когда взял глаз быка, обнажил поверхность плотной бе-
лой оболочки (склеры), удалив ткань с внутренней стороны глазного яблока,
и показал, что свет, проходящий через хрусталик глаза, проецируется на белую
склеру, как на экран; приблизив глазное яблоко к освещенному объекту, можно
было увидеть проекцию изображения. Многие люди удивлялись, что зритель-
ный стимул проецировался на заднюю стенку глаза в перевернутом положении
(верх и низ менялись местами). Декарт сумел доказать, что это объясняется
оптическими свойствами хрусталика*
.
* До сих пор многие люди не понимают, почему мы воспринимаем зрительные объекты не в пере-
вернутом виде, т. е . не так, как изображение проецируется на сетчатку. Как бы вы ответили на этот
вопрос?
40
Глава 1
4.1.
Психика и мозг
Декарт часто считается основателем современных философских взглядов на
соотношение между психическим и телесным (соматическим). Главный во-
прос кажется простым: каков мир по своей природе — психический или фи-
зический? Или, используя современные понятия, можно ли объяснить наш со-
знательный опыт деятельностью нервных клеток? Может, и сами-то нервные
клетки существуют лишь в воображении ученых? Мозг как базис сознания —
таково сейчас главное направление когнитивной нейронауки (�delman, 1989;
Palmer, 1999; K�ch, 1996; Tulv�ng, 2002; Baars et al., 2003a). В течение послед-
них 15 лет опубликованы многочисленные статьи на эту тему. В июле 2005 г.
журнал «Sc�ence» отметил проблему «биологические основы сознания» в числе
приоритетов современной науки. Нобелевские лауреаты Френсис Крик, Дже-
ральд Эдельман, Герберт Саймон посвятили этому вопросу годы своей жизни.
В своем повседневном опыте мы постоянно совершаем переходы между
языком в терминах психики и в терминах мозга. Мы принимаем аспирин (фи-
зическая сущность), чтобы облегчить головную боль (психическая сущность).
Мы идем к холодильнику (физическая сущность), потому что испытываем же-
лание (психическая сущность) съесть мороженое. Как определить истинную
последовательность событий — наше сознательное переживание «вызывает»
Рис.1 .10 . Рене Декарт — философ, математик, исследователь мозга. Справа приведен
сделанный Декартом рисунок оптической системы глаза. Поскольку Декарт был убеж-
ден, что душа (психика) уникальна, он отвергал идею о том, что носителем души мо-
гут быть парные структуры мозга. Однако почти все макроструктуры мозга являются
парными: два полушария, два глаза и два уха, две половины подкорковых образований,
два полушария мозжечка. Поэтому Декарт решил, что крошечная шишковидная железа
(пинеальная железа, или эпифиз), которая для невооруженного глаза выглядит как одна
точка, должна быть связующим звеном между божественной душой и бренным земным
телом. К несчастью для Декарта, после XVII в. микроскопические исследования пока-
зали, что шишковидная железа тоже характеризуется билатеральной симметрией, как и
весь мозг. (Источник: Bennett, 1989.)
Психика и мозг
41
физические действия, или же наоборот? С точки зрения здравого смысла, это
безразлично. Мы свободно переходим от понятий «психических сущностей» к
понятиям «телесных сущностей», и наоборот. Однако дело осложняется, если
мы попытаемся мыслить глубже. Мы хорошо представляем себе обычные яв-
ления, связанные с приемом аспирина и доставанием мороженого из холодиль-
ника. Мороженое тает на солнце, а аспирин растворяется в воде. Это проис-
ходит согласно законам физики и химии. Однако психические явления зависят
от целей, эмоций и мыслей, подчиняющихся иным законам. Мороженое тает не
потому, что оно хочет этого, тогда как человек ест мороженое, чтобы удовлет-
ворить свое желание. В человеческом языке есть тысячи слов для обозначения
желаний и переживаний, но эти слова неприменимы к физическим объектам.
Как мы увидим, накоплено много эмпирических данных по проблеме созна-
тельной когнитивной деятельности. Признано, что некоторые области мозга,
например вентральный зрительный путь, участвуют в сознательном воспри-
ятии зрительных образов, которые человек без труда может описать. Вместе
с тем показано, что далеко не все области мозга обеспечивают сознательное
восприятие, в результате которого человек способен пересказать события. Так,
дорсальный зрительный путь задействован в зрительной координации движе-
ния руки к предмету, однако соответствующие области мозга не могут сфор-
мировать сознательное восприятие сами по себе, без участия других структур
(���dale et al., 1991). Все больше ученых поддерживают мнение о том, что
проблема соотношения между психикой и мозгом должна решаться опытным
путем, в доступных для проверки экспериментах.
4.2 .
Биологическая природа когнитивной деятельности
и эмоций
Биологические корни человеческих эмоций заложены в древних структурах
мозга млекопитающих и регулируются более поздними слоями новой коры.
Неудивительно, что история эволюции мозга человека, уходящая в глубь при-
мерно двухсот миллионов лет к первым представителям млекопитающих, вы-
зывает много важных вопросов. Процессы, составляющие основу материнской
любви и привязанности потомков, протекают в древней структуре мозга мле-
копитающих, так называемом околоводопроводном сером веществе. Звуки, вы-
ражающие горе или призыв, по-видимому, связаны в ходе эволюции с возник-
новением речевой просодии (тональности и мелодики языка) и даже с музыкой.
Таким образом, здесь обнаруживается биологическая природа во многих ее
проявлениях.
Чарлз Дарвин (рис. 1 .11) в большей мере, чем кто-либо другой, помог опре-
делить биологический контекст действий человека. Ему принадлежат много-
численные наблюдения относительно того, каким образом животные и люди
выражают свои эмоции. В течение сотен лет люди не могли не замечать сход-
ства таких проявлений, однако в Европе и других регионах считалось абсо-
лютно недопустимым признавать это. Теперь убедительно доказано сходство
эмоциональных центров мозга у человека и других млекопитающих (Pankse��,
2005). Во времена Дарвина такая идея была бы совершенно одиозной. Сейчас
книга Дарвина «Выражение эмоций у человека и животных» считается класси-
ческой (�kman, 2003).
42
Глава 1
Дарвин никогда не собирался отрицать роль культуры и среды обитания. Че-
ловек — самый адаптирующийся вид из всех известных, наиболее способный к
обучению путем культуральной передачи. Противоречие между «врожденным
и приобретенным» перестало быть предметом споров. Большинство ученых
рассматривает поведение человека как результат объединенного воздействия
биологических, культуральных факторов и среды (см. гл . 15).
В XIX в. великим ученым в области физиологии сенсорных процессов
и психологии был Герман Гельмгольц (рис. 1 .12), труды которого о природе
зрения и слуха не потеряли значения до сих пор. Особую известность получи-
ли опыты, в которых Гельмгольц продемонстрировал физический закон сохра-
нения энергии при электрическом раздражении препарата задних конечностей
лягушки. Представление о том, что электричество играет роль в активности
нервной системы, привело в начале ХХ в. к открытию электрических потен-
циалов нерва и мозга. В наше время благодаря регистрации электромагнитной
активности мозга возник целый ряд методов визуализации: регистрация ЭЭГ,
вызванных потенциалов, активности индивидуальных нейронов, метод магнит-
ной энцефалографии (МЭГ) и даже транскраниальной магнитной стимуляции
(ТМС) (гл. 4). Все методы визуализации мозга основаны на фундаментальных
законах физики и химии.
Рис. 1.11. Чарлз Дарвин внес вклад в биологическое обоснование психики. После Рене
Декарта многие люди стали размышлять о мышлении и его связи с мозгом. В их числе
был и Чарлз Дарвин, который написал книгу «Выражение эмоций у человека и жи-
вотных». По мнению Дарвина, эмоции человека имеют биологическую природу; при
этом он отнюдь не забывал о влиянии факторов культуры и о роли индивидуальных
особенностей. На фотографии Дарвин показан в молодом возрасте, примерно в пери-
од его исторического путешествия через Тихий океан на корабле «Бигль». (Источник:
F�nkelste�n, 2000.)
Психика и мозг
43
В эпоху Гельмгольца было хорошо известно, что электрический стимул вы-
зывает сокращение конечностей лягушки — такое же, как во время прыжка.
Представление о том, что электричество пронизывает мозг и все тело, стало
модным и выразилось в таких популярных идеях, как «животный магнетизм».
Эксперимент Гельмгольца на препарате конечностей лягушки стал важней-
шим шагом на пути к натуралистической концепции соотношения мозга и тела
(рис. 1 .13).
Рис. 1 .12 . Герман Гельмгольц не был профессиональным ученым, но внес историче-
ский вклад в науку. Он одним из первых предположил, что в зрительной системе фор-
мируются «бессознательные умозаключения», которые выходят далеко за рамки про-
стого потока сигналов от глаз. Идея была встречена такими возражениями, что автору
пришлось отказаться от нее, однако в современной науке она стала базовым понятием.
(Источник: Bennett, 1999.)
44
Глава 1
4.3.
Нейронная доктрина Рамон-и -Кахала:
рабочая гипотеза науки о мозге
Сантьяго Рамон-и-Кахалу (рис. 1 .14) принадлежит заслуга разработки нейрон-
ной доктрины — одной из основополагающих теорий науки о мозге, соглас-
но которой «нервная система состоит из многочисленных нервных элементов
(нейронов), анатомически и генетически независимых друг от друга». Важно
отметить, что идея о клеточном строении организма относится только к 1839 г.,
когда с помощью светового микроскопа и методов окрашивания тканей были
впервые в истории человечества выявлены живые клетки.
Возникло предположение, что из клеток состоят все ткани организма, кроме
нервной ткани. Такое исключение было обусловлено тем, что нейроны имеют
множество отростков, их аксоны и дендриты густо переплетаются, образуя тес-
ные контакты с другими нервными клетками; при этом не удавалось различить,
соприкасаются они друг с другом или нет. Так появились разногласия относи-
тельно устройства нервной системы — либо это скопление миллиардов клеток,
либо одна огромная непрерывная сеть.
Работая в Мадриде, Сантьяго Рамон-и-Кахал выделил с помощью метода
окрашивания по Гольджи небольшую группу нейронов на срезе ткани таким
образом, что можно было рассмотреть форму клеток (рис. 1 .15). Кроме того,
Кахал показал, что у аксонов свободные, неприкрепленные окончания и, следо-
вательно, нейроны являются самостоятельными клетками.
Рис. 1 .13 . Схема препарата, который был использован Гельмгольцем для оценки скоро-
сти распространения нервного импульса, вызывающего сокращение мышц конечности
лягушки. (Источник: Bennett, 1999.)
Психика и мозг
45
Рис. 1 .14 . Слева: Сантьяго Рамон-и-Кахал, основоположник науки о мозге, которого,
вероятно, можно считать самым выдающимся среди первых нейробиологов. Применив
метод окрашивания по Гольджи, Кахал получил с помощью светового микроскопа ба-
зовые сведения по микроанатомии нервных клеток. Справа: зарисовки нейронов, сде-
ланные Кахалом с помощью микроскопа. Современные исследования микроструктуры
нейронов являются развитием представлений Кахала. (Источник: DeFel��e, 2002.)
Рис. 1 .15. Первый рисунок Рамон-и-Кахала, показывающий микроскопическую карти-
ну среза мозжечка курицы. Кахал применил метод окрашивания по Гольджи, что позво-
лило отличить нервные клетки от окружающей их ткани. (Источник: DeFel��e, 2002.)
46
Глава 1
Нервный импульс — электрохимический «спайк», который распространя-
ется по нервному волокну по направлению к его терминалям, был впервые за-
регистрирован в гигантском аксоне кальмара. Поскольку нервные клетки вы-
явлены у всех животных, а их базовые свойства оказались сходными у многих
организмов, аксон кальмара стал важным стандартным объектом для изучения
других классов нейронов. Это открытие было сделано в 1939 г., через сто лет
после провозглашения клеточной доктрины. Спустя десять лет было обнару-
жено, что ионы натрия и калия (Nа+ и К+) создают быстрые входящие и выхо-
дящие потоки через мембрану аксонов, обеспечивая распространение спайка.
В 1952 г. Ходжкин и Хаксли разработали модель потенциала действия, которая
признана до сих пор (рис. 1 .16). Примерно в тот же период благодаря электрон-
ному микроскопу с разрешением до нескольких сотен микронов удалось выде-
лить еще более мелкие образования — синапсы.
Однако завершение истории об открытии Кахала несет в себе долю иронии.
Великий спор о том, разделены ли нейроны, продолжается поныне. Казалось
бы, Кахал окончательно закрыл этот вопрос, обнаружив свободные окончания
аксонов. Тем не менее, согласно более поздним данным, многие пункты пере-
дачи сигналов между нервными клетками представляют собой не химические
Рис. 1 .16 . Современное изображение клетки, впервые зарисованной Кахалом. Микро-
рентгенограмма нейрона после химического окрашивания среза ткани. (Источник:
Standr�ng, 2005.)
Психика и мозг
47
синапсы, а непрерывные электрические соединения — так называемые щеле-
вые контакты. Есть нейроны, которые прилегают друг к другу без классической
синаптической щели. Таким образом, некоторые нейроны действительно об-
разуют непрерывную структуру, хотя нет сомнений и в существовании трил-
лионов химических синапсов. Подобно многим другим научным спорам, дис-
куссия продолжается на более детальном уровне.
4.4 .
Пьер-Поль Брока: локализация центра речи
В развитие науки о соотношении психики и мозга вносят свой вклад медицин-
ские наблюдения. Французский врач Пьер-Поль Брока (рис. 1 .17) обнаружил
в левом полушарии область, связанную с высшей психической функцией —
речью. Поэтому «речевая» зона левого полушария получила название в честь
Брока.
По поводу локализации в мозге специфических функциональных центров
высказывались противоречивые мнения. Брока первым неоспоримо доказал
в 1861 г., что в мозге имеются специализированные области. Открытие было
сделано в результате наблюдений за больным эпилепсией, который полностью
лишился речи, кроме единственного слога «tan». Когда пациент умер, Брока
получил возможность провести патолого-анатомическое исследование и уви-
дел «повреждение в задней области третьей лобной извилины левого полу-
шария». Препарат мозга этого пациента был сохранен, и в нем можно видеть
поврежденный участок лобной части левого полушария (рис. 1 .18). Брока сде-
лал вывод, что эта часть лобной доли необходима для продукции речи. Спустя
шесть месяцев Брока встретился с аналогичным клиническим случаем, и снова
было выявлено повреждение того же участка левой лобной доли. Несмотря на
возражения, что в этих случаях были затронуты и другие области мозга и что
Рис. 1 .17. Пьер-Поль Брока ввел определение экспрессивной афазии. Он первым описал
клинический случай, который доказывал специфическую локализацию в коре больших
полушарий конкретной специфической функции. (Источник: Am�n�ff and Dar�ff, 2003.)
48
Глава 1
у некоторых больных с аналогичными расстройствами отсутствовали повреж-
дения лобных долей коры мозга, открытие Брока получило признание благода-
ря последующим убедительным фактам. Теперь центр Брока в левом полуша-
рии широко признается в качестве критически необходимого участка речевой
зоны (Am�n�ff and Dar�ff, 2003).
Многое из того, что мы знаем сейчас о мозге человека, стало впервые извест-
но в результате тщательного изучения определенных патологических состоя-
ний. Такого рода исследования продолжаются. Важное наблюдение, касающе-
еся механизмов речи, принадлежит Карлу Вернике (рис. 1 .19). В монографии,
опубликованной в 1874 г., он представил модель речи на основе своих невроло-
гических наблюдений; эта концепция поныне остается ключевой в когнитивной
нейронауке. В настоящее время зона Вернике в левой верхней части височной
доли рассматривается как область, ответственная за восприятие речи. До сих
пор больные, у которых нарушено понимание речи вследствие поражения этой
области, получают диагноз «афазия Вернике». Вместе с тем по мере развития
методов визуализации мозга оказалось, что организация соответствующих об-
ластей мозга является более сложной и динамичной, чем считалось раньше.
В современной публикации (Dr�nkers and Ogar, 2003) приведен образец речи
больного с рецептивной афазией при поражении зоны Вернике. Больной гово-
рит гораздо более связно, чем в описанном Брока в 1861 г. классическом слу-
чае, когда пациент произносил только один слог. Однако у больного нарушена
способность продуцировать и воспринимать осмысленную речь. В настоящее
Рис. 1 .18 . Препарат мозга одного из первых пациентов Брока. В лобной доле левого по-
лушария видно большое отверстие в «области Брока». (Источник: Ramachandran, 2002.)
Психика и мозг
49
время стандартно применяется визуализационное определение пораженного
участка, так что диагноз уже не зависит только от неврологических заключений
и данных аутопсии.
В XIX в. было хорошо известно, что основную массу коры больших полуша-
рий составляет белое вещество — миллиарды аксонов, отростков серых клеточ-
ных тел, находящихся в поверхностном слое коры. Аксоны покрыты защитной
и поддерживающей миелиновой оболочкой белого цвета, богатой молекулами
липидов. Поэтому невооруженный глаз видит белую массу. Белое вещество со-
стоит преимущественно из крупных пучков нервных волокон, соединяющих
области мозга друг с другом наподобие гигантской дорожной системы. В ре-
зультате тонкой анатомической препаровки были прослежены нервные пучки
между центрами Брока и Вернике левого полушария. Они называются аркуат-
ными (дугообразными) пучками. (Еще один пример термина латинского проис-
хождения.)
Рис. 1 .19 . Немецкий врач Карл Вернике через несколько лет после работ Брока по экс-
прессии речи (процессам лингвистического выхода) обнаружил отдел мозга, участву-
ющий в восприятии и понимании речи (процессах лингвистического входа). Вернике
исследовал ряд пациентов с афазией (термин «афазия» означает «нет речи») и пришел
к выводу, что в зависимости от локализации повреждения коры мозга возникают раз-
личные расстройства речи. Он отделил семантические формы афазии от рецептивных и
продуктивных форм. Справа: модель «домика» — схема процесса формирования речи в
мозге согласно представлениям в XIX в. А — область слухового восприятия речи, В —
двигательная область продукции речи, С — распределенные представительства речи в
мозге. (Источник: Wern�cke and K�ehler, 2003.)
50
Глава 1
Исходя из этих данных, Вернике в 1874 г. высказался о вероятном существо-
вании еще одного расстройства речи — коннекционной афазии (нарушения свя-
зи). Он рассуждал следующим образом: если повреждена связь между центрами
Брока и Вернике, у больного должна страдать способность к повторению ус-
лышанной речи, поскольку нарушается передача информации от рецептивной
области (центра Вернике) к экспрессивной области (центру Брока). Прогноз
подтвердился (см. рис. 1 .20). Считается, что существует ряд подобных синдро-
мов дисконнекции, симптомы которых различны в зависимости от вовлечения
конкретных групп нейронов коры. Предполагаются и другие формы афазии.
Заметим, что эти речевые зоны обычно находятся в левом полушарии
(рис. 1 .21). Согласно современным сведениям, правая половина мозга участву-
ет в понимании языка, но не в вокализации, т. е . не в речевом выходе. Считается,
что правое полушарие восприимчиво к эмоциональному содержанию речи —
юмору, иронии. Доминантность левого полушария по отношению к речи харак-
терна примерно для 90% населения, тогда как примерно у 10% нормальных
людей доминантно правое полушарие.
Уильям Джеймс (рис. 1 .22) известен сейчас прежде всего как психолог и
философ, однако он обучался также медицине и рисованию. Вначале Джеймс
преподавал в Гарвардском университете анатомию мозга, и опубликованные
им в 1890 г. описания хорошо согласуются с современными представлениями.
Обратим внимание на его зарисовки зон Брока и Вернике, основанные на кли-
нических наблюдениях при афазии (рис. 1 .23).
Книга Джеймса «Принципы психологии» часто упоминается как лучшее в
XIX в. англоязычное обобщение в области психологии и науке о мозге. Наря-
Рис. 1.20 . Проводниковая афазия. В результате аккуратной анатомической препаровки
был обнаружен пучок нервных волокон, соединяющий в левом полушарии центр Брока
(B) и центр Вернике (W). Это позволило Вернике предположить (1874 г.) существова-
ние еще не известного расстройства речи — дисконнекционной афазии. По мнению
Вернике, при повреждении этого пучка нарушается передача информации от рецеп-
тивной (воспринимающей) зоны к продуктивной (моторной) зоне. Прогноз Вернике
подтвердился, и такая форма расстройства получила название проводниковой афазии.
(Источник: Dr�nkers and Ogar, 2003.)
Психика и мозг
51
ду со многими другими фактами Джеймс обсуждает результаты исследований
доктора Моссо, обнаружившего усиление кровотока в активной области мозга
(рис. 1 .24). Это открытие стало прообразом современных методов визуализа-
ции мозга, которые основаны на регистрации гемодинамики (кровотока), —
таких как ПЭТ и МРТ (рис. 1.25).
Рис. 1 .21 . Речевые центры левого полушария, исследование с помощью фМРТ (функ-
циональной магнитно-резонансной томографии). Красные зоны участвуют в продукции
речи, желтые — в восприятии и понимании речи. Классические зоны Брока и Вернике
обведены кружками. Отметим близкое сходство между современными представления-
ми о локализации центров речи и объяснениями, существовавшими в XIX в. (ПСК —
первичная слуховая кора, область коры мозга, где начинается вторичная обработка сиг-
налов в процессе формирования речи; пВВБ — передняя верхняя височная борозда,
передняя часть верхней складки височной доли). (Источник: Frack�w�ak, 2004.)
52
Глава 1
Рис. 1.22 . Уильям Джеймс преподавал анатомию в Гарвардском университете. Теперь
он известен прежде всего как психолог и философ, однако имел также медицинское
и художественное образование. Опубликованное им в 1890 г. описание мозга хорошо
согласуется с современными представлениями. Фотография любезно предоставлена
Национальной медицинской библиотекой США
Рис.1 .23 . У. Джеймс обозначил зоны Брока и Вернике, основываясь на медицинских
данных, которые были получены при исследовании больных с поражениями мозга.
(Источник: James, 1890.)
Психика и мозг
53
Рис. 1 .24 . У. Джеймс (1890) описал резуль-
таты исследований доктора Моссо, который
открыл способ измерить давление крови в
мозге человека во время умственной работы.
Исследование Моссо явилось прообразом
технологии регистрации мозгового кровото-
ка в наше время, например фМРТ. (Источ-
ник: James, 1890.)
Рис. 1.25. Регистрация из-
менений
регионального
мозгового кровотока с помо-
щью фМРТ. Метод основан
на том, что в тех областях
мозга, где усиливается кро-
воток, возрастает потребле-
ние кислорода и глюкозы.
Исследование можно рас-
сматривать как современ-
ную версию опыта доктора
Моссо в XIX в. (Источник:
Th�mas Ramsöy.)
54
Глава 1
[В опытах Моссо] «испытуемый лежал на горизонтальном стенде, находившемся в
неустойчивом равновесии, так что передний или задний конец опускался при уве-
личении веса соответственно головы или ног испытуемого. Когда испытуемый ис-
пытывал эмоциональное или интеллектуальное напряжение, головной конец стенда
опускался, потому что в организме происходило перераспределение крови» (Уильям
Джеймс, Принципы психологии (1890)).
Итак, в XIX в. было доказано, что определенные области мозга отвечают
за функцию речи. При этом ученые интересовались не только классическими
психологическими проявлениями, но и их зависимостью от мозга.
4.5 .
Осознаваемые и неосознаваемые
психические явления
В XIX в. врачи полагали, что сознательная деятельность человека зависит от
коры больших полушарий — наружного слоя мозга. Джеймс писал:
«Однако можно ли считать, что осознаваемые процессы, которые зависят от
активности коры мозга, — единственная форма сознания у человека? Или низ-
шие центры мозга тоже участвуют в сознательной деятельности? ...единствен-
ный орган, обеспечивающий сознание у человека, — это кора мозга».
Результаты, полученные с помощью современных методов нейровизуали-
зации, поддерживают, скорее, точку зрения Джеймса, хотя некоторые ученые
высказываются и в пользу участия подкорковых структур (см. гл . 2 и 8). Вопрос
о роли сознания в восприятии и в когнитивной (познавательной) деятельности
вновь оказался в центре исследований, как мы увидим далее.
Проблема сознания глубоко интересовала ученых XIX в. Рамон-и -Кахал
выдвинул идею о «нейронах с психической функцией» в коре мозга, обеспе-
чивающих сознательные состояния. Возникновение психофизики в начале
XIX в. было в существенной мере связано со стремлением разрешить загадку
соотношения между психическими и телесными функциями. Основоположник
психофизики Густав Фехнер объявил, что решение найдено: он вывел матема-
тическую зависимость интенсивности субъективного ощущения от силы физи-
ческого стимула. В конце XIX в. Уильям Джеймс написал: «Психология — это
наука об умственной жизни»; при этом он имел в виду сознательные умствен-
ные действия (James, 1980/1983). Джеймс не был одинок в своем мнении.
Но примерно в то же время появились возражения. Такие ученые, как Гельм-
гольц, Лёб и Павлов, придерживались более физикалистской точки зрения. По-
сле 1900 г. получили известность опыты И. П. Павлова с выработкой класси-
ческого условного рефлекса у собак. После этого многие психологи пришли к
убеждению, что все формы поведения можно объяснить через элементарные
поведенческие акты на основе рефлексов (рис. 1 .26). В США Джон Уотсон ре-
шительно провозгласил радикальный бихевиоризм, заявив, что всякое упоми-
нание о сознании неправомерно, поскольку наблюдать в явном виде мы можем
только реальное поведение и мозг как физическое образование. Знаменитый
лозунг Уотсона таков: «Сознание — это не более чем воплощение теологии»
и, следовательно, не является научным понятием. В XX в. ученые длительное
время обходили вопрос о сознании человека. Некоторые считали, что он слиш-
Психика и мозг
55
ком отягощен философскими аспектами, трудно поддается экспериментальной
проверке или имеет слишком субъективный характер, чтобы подойти к нему с
научных позиций.
Для таких рассуждений есть практические причины. Во многих случаях
бывает трудно судить о состоянии другого человека, поэтому сложно воспро-
изводить эксперименты в соответствии с научными требованиями к обоснован-
ности фактов.
Биологическая теория сознания
Все мы постоянно испытываем осознаваемые психические явления — внутреннюю
речь, мысли и ощущения, осознание своего «Я». Осознание себя и окружающего
мира формируется в течение первых нескольких лет жизни. Это нечто такое, в чем
мы сходны с миллионами других людей, но в то же время — глубоко личное; никто
другой не видит, не чувствует, не понимает в точности так же, как мы. Осознан-
ность присуща человеку как биологическому виду, но вместе с тем — это частное
свойство каждого. Известно, однако, что некоторые отделы мозга, например мозже-
чок, не дают начало сознательным переживаниям. В чем же заключается разница
между отделами мозга, обеспечивающими сознательную деятельность, и теми, ко-
торые не участвуют в ней? (см. гл. 8).
Нейробиологические основы сознания составляют центральный раздел нейропси-
хологии. Один из лидеров этого направления — Джеральд Эдельман, лауреат Нобе-
левской премии, директор Института нейронаук и руководитель кафедры нейроби-
ологии научно-исследовательского института Скриппса Университета Ла Джолла
(штат Калифорния). Интересуется классической музыкой и театром. Джеральд
Рис. 1 .26. И . П . Павлов вместе с сотрудниками оперирует под наркозом собаку; от-
дел физиологии Института экспериментальной медицины в Санкт-Петербурге, 1902 г.
(Источник: Tansey, 2006.)
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
56
Глава 1
Эдельман выдвинул биологическую теорию нейродарвинизма, центральное место
в которой принадлежит представлению об осознаваемых процессах, развитие кото-
рых продолжается в течение жизни (см. гл . 8).
Нейродарвинизм — фундаментальная теория, согласно которой развитие и дина-
мическая деятельность мозга имеют селекционистскую природу, в отличие от ин-
струкционизма цифровых компьютеров, выполняющих символьные, т. е . заданные
в явном виде, инструкции (рис. 1 .28).
Селекционистские системы характеризуются четырьмя особенностями.
1. Наличие спектра разнообразных элементов (например, особей биологического
вида, антител иммунной системы, нейронов мозга).
2. Эти элементы могут репродуцироваться или амплифицироваться (повышать
силу).
3. Среди разнообразных элементов происходит отбор, благодаря которому в ходе
эволюции часть потомства остается, а часть исчезает. В иммунной системе се-
лекция антител определяется успешностью их взаимодействия с антигенами. Что
касается мозга, то он обладает двумя селекционными репертуарами. Первичный
репертуар (набор нейронов) образуется в процессе нейрогенеза на ранних этапах
развития. Вторичный репертуар (набор нейронных сетей) создается в результате
избирательного отсеивания функционально неэффективных синаптических свя-
зей при формировании зрелого мозга. Вторичный репертуар нейронных связей
продолжает действовать в течение всей нашей жизни, обеспечивая адаптацию и
развитие.
4. Система является избыточной (по Эдельману — «вырожденной»). Это означает,
что одну и ту же функцию могут осуществлять различные наборы ее элементов
(�delman, �ally, 2001). Избыточность проявляется на всех уровнях живого. В на-
�delman, �ally, 2001). Избыточность проявляется на всех уровнях живого. В на-
, �ally, 2001). Избыточность проявляется на всех уровнях живого. В на-
�ally, 2001). Избыточность проявляется на всех уровнях живого. В на-
, 2001). Избыточность проявляется на всех уровнях живого. В на-
шем организме два легких, две половины мозга, множество проводящих путей
на каждом уровне организации. Благодаря избыточности компенсируются по-
следствия травмы или болезни, поскольку начинают использоваться альтерна-
тивные проводящие пути и органы. Таким образом, организмы могут выживать
в периоды кризисов.
Рис. 1.27. Джеральд М. Эдельман, директор
Института нейронаук в Сан-Диего, Кали-
форния, США
Психика и мозг
57
Сознательный опыт
По Эдельману, сознательный опыт — это результат обратных связей между нейро-
нами таламокортикальной системы, которую можно назвать центральной частью
мозга млекопитающих. Повторный вход (reentry) поддерживает постоянную актив-
reentry) поддерживает постоянную актив-
) поддерживает постоянную актив-
ность между парами нейронных ансамблей, что приводит к модификации связей
между ними, а также к отбору самих ансамблей. Это в высокой степени адаптивный
процесс, его можно сравнить с социальными сетями в Интернете, которые расши-
ряются и развиваются в зависимости от контактов между пользователями. С точки
зрения этой аналогии, можно сказать, что таламокортикальные нейроны взаимо-
действуют в составе большого «динамического центра». Динамический центр как
единое образование может чрезвычайно быстро изменяться, адаптируясь к новым
условиям, извлекая сохраняемую в памяти информацию, выполняя целевые уста-
новки. Первичное сознание относится к миру восприятия и памяти; более высокие
уровни сознания распространяются за рамки первичного сознания, в их сферу вхо-
дят речь, абстрактные понятия, мышление.
Рис. 1 .28 . Три основных положения теории нейродарвинизма. Селекция на ран-
них стадиях онтогенеза обеспечивает формирование разнообразных наборов ней-
ронных сетей в ходе деления клеток, клеточной смерти и распространения отрост-
ков (дендритов и аксонов), которые достигают нейронов-мишеней и образуют с
ними синаптические соединения. На следующем этапе селекция в процессе опыта
(научения при взаимодействии со средой) приводит к изменениям синаптической
силы, в результате которых одни проводящие пути получают преимущество перед
другими. Рекуррентные нейронные сети обеспечивают координацию нейронной
активности разных отделов мозга. (Черными точками обозначены усилившиеся си-
напсы.) (Источник: Squ�re, 2004.)
58
Глава 1
Содержательное наполнение сознания (его контент) многоаспектно — это сенсор-
ное восприятие, зрительные образы, эмоциональные состояния, внутренняя речь,
абстрактные концепции, идеи, связанные с действием, и многое другое. Такой ши-
рокий диапазон содержимого сознания возможен благодаря вовлечению множества
областей мозга. Как известно, в осознаваемых процессах восприятия участвует
сенсорная кора; однако есть данные о том, что при некоторых сенсорных явлениях
(примером могут служить процессы в мозге человека при виде чашки кофе) во-
влекаются и другие области, в частности лобная и теменная кора. Целостное пред-
ставление о сознании должно также учитывать взаимодействие между областями
мозга, которое происходит во время сознательного извлечения информации из па-
мяти, сознательного управления движениями и так далее. Эдельман с сотрудниками
полагают, что динамический центр осуществляет контроль очень широкого спектра
функций мозга. В этот общий контур включены функциональные биологические
модели таламокортикальной системы, гиппокампа и других областей мозга. Неко-
торые из них воспроизведены в реальных условиях, чтобы выяснить, имитируют ли
они процессы научения у млекопитающих.
Согласно предположению Эдельмана и Тон�ни (2000; �delman, T�n�n�, 1998), со-
�delman, T�n�n�, 1998), со-
, T�n�n�, 1998), со-
T�n�n�, 1998), со-
, 1998), со-
1998), со-
1998), со-
держательное наполнение любого осознаваемого процесса не является простым
следствием активности соответствующей области коры. Ощущение красного цвета
зависит не только от активации зрительной коры, хотя эта активация необходима.
Такое ощущение определяется состоянием всего динамического центра. На фор-
мальном языке каждый сознательный процесс можно представить в виде одиночной
точки в N-мерном пространстве, где N — число нейронных групп, составляющих
часть динамического центра в каждый момент времени. Таким образом, восприятие
красного цвета связано с высокой степенью пространственной дискриминации —
не только с распознаванием определенного цвета, а с выбором в пределах гораздо
более обширного спектра возможных состояний динамического центра.
В соответствии с расширенной теорией нейродарвинизма разнообразие контен-
тов сознания обусловлено способами воздействия различных нейронных групп на
N-мерное пространство. Несмотря на удивительную гистологическую однород-
-мерное пространство. Несмотря на удивительную гистологическую однород-
ность значительной части коры мозга млекопитающих, разные области коры полу-
чают различные входы*
. Например, зрительный вход имеет совершенно иные ста-
тистические характеристики, чем проприоцептивный вход. Следовательно, группы
нейронов, чувствительных к цвету, могут организовать пространство иначе, чем
проприоцептивные нейроны. Итак, каждая сенсорная модальность влияет на со-
стояние динамического центра только свойственным ей образом, и это объясняет,
почему содержание осознаваемого явления, в котором доминирует зрительное вос-
приятие, будет иным, чем при доминировании звука, осязания или вкуса.
Литература
1. �delman �.M . (1993). Neural darw�n�sm: Select��n and reentrant s�gnal�ng �n h�gher
�ra�n funct��n. Neuron, 10(2), 115–125.
2. T�n�n� �. & �delman �.M . (1998). C�nsc��usness and c�m�lex�ty. Sc�ence, 282
(5395), 1846–1851.
3. Kr�chmar J.L. & �delman �.M. (2005). Bra�n-�ased dev�ces f�r the study �f nerv�us
systems and the devel��ment �f �ntell�gent mach�nes. Artif. Life, 11(1–2), 63–77 .
4. Seth A.K ., Izh�kev�ch �., Reeke �.N . & �delman �.M. (2006). The�r�es and measures
�f c�nsc��usness: An extended framew�rk. Proceedings of the National Academy of
Sciences of the United States of America, 11 ;103(28), 1079–1084.
** Это не совсем так. Например, в новой коре поля, отвечающие за двигательную функцию (4, 6, 10,
8 и др.), лишены IV слоя, т. н . гранулярная кора. Кора гиппокампа формируется как трехслойная
кора. См. атласы по цитоархитектонике. — Прим. перев.
Психика и мозг
59
5. �delman �.M. & T�n�n� �. (2001). A Universe of Consciousness: How Matter Be-
comes Imagination. Bas�c B��ks. New Y�rk.
6. �delman �.M. (2007). Second Nature: Brain Science and Human Knowledge. Yale
Un�vers�ty Press, New Haven.
7. �delman �.M. (2005). Wider than the Sky: The Phenomenal Gift of Consciousness.
Yale Un�vers�ty Press, New Haven.
8. Seth A.K . & Baars B.J. (2005). Neural darw�n�sm and c�nsc��usness. Consciousness
and Cognition, 14(2005), 140–168.
5.0 . Наука возвращается к представлениям о сознании
В 1970-е гг. многие психологи разочаровались в бихевиоризме и стали искать
иной путь. Когнитивные психологи не только проводили оценки поведения, но
и стремились к умозаключениям. Мы рассмотрим в этой главе примечатель-
ный пример, касающийся концепции оперативной памяти. Есть много и других
примеров. Когнитивные психологи вновь обратились к зрительным образам,
разным формам памяти, неосознаваемой (неявной, имплицитной) когнитивной
деятельности и другим понятиям. Однако эти концепции всегда проверяются
путем исследования поведенческих реакций в строго контролируемых лабо-
раторных условиях. Теории оперативной памяти, образов и т. д. выводятся из
результатов наблюдений за поведением. Это теоретические заключения напо-
добие представлений об электроне в физике. Никто никогда не видел электрон,
но соответствующая теория позволяет объяснить множество эмпирических на-
блюдений (Baars, 1986).
«Возможно, важнейший сдвиг в когнитивной психологии и вообще в когнитивной
науке за последние 20 лет заключается в том, что понятие сознания приобрело ста-
тус признанной, изучаемой научной проблемы. В течение значительного периода в
ХХ в. эта тема была погружена в дебри философических рассуждений о соотноше-
нии между психическим и телесным, зажата эмпирикой интроспекционизма. Затем
постепенно выяснилось, что эти ограничения в действительности не являются кар-
динальным препятствием, которого вначале опасались. Кроме того, были известны
феномены слепого зрения и имплицитной (процедурной) памяти, когда восприятие
и вспоминание осуществляются иначе, чем при сознательных действиях того, кто
ощущает или вспоминает; необходимость объяснить эти явления настоятельно по-
буждала вернуться к эмпирическому подходу в психологии сознательного восприя-
тия.» (Baddeley, личное сообщение)
В последние годы предубежденность к исследованиям сознания стала ос-
лабевать. Многие когнитивные психологи заняты изучением как эксплицит-
ных (осознаваемых) процессов, так и имплицитных (неосознаваемых). Как мы
увидим, накопилось большое количество данных в пользу того, что наше вос-
приятие окружающего мира происходит отчасти неосознаваемо, но конечный
результат процесса восприятия осознаваем. Многие аспекты памяти имеют не-
осознаваемый характер, хотя в эпизодической памяти регистрируются осозна-
ваемые события прошлого. Методы функциональной визуализации позволяют
сравнивать процессы в мозге во время сознательной и бессознательной когни-
тивной деятельности. Сейчас каждый год публикуется примерно 5000 статей,
в которых фигурирует понятие сознания или его синонимы — «осведомлен-
ность» (awareness); «эксплицитный познавательный процесс», или «сознатель-
60
Глава 1
ный когнитивный процесс» (ex�l�c�t c�gn�t��n); «эпизодическое вспоминание»
(e��s�d�c recall); «фокусное внимание» (f�cal attent��n). Терминология отраба-
тывается экспериментально путем оценки «точного сообщения». Мы просим
человека описать восприятие или припоминание какого-нибудь события, а за-
тем стараемся оценить точность сообщения. В этом смысле оценка синонимов
для обозначения осознаваемых явлений является единообразной и должна обе-
спечить одинаковый смысл терминов. Мы остановимся на термине «осознавае-
мые» процессы, а также будем пользоваться языком специалистов (Baars et al.,
2003).
В XIX в. такие ученые, как Зигмунд Фрейд и Уильям Джеймс, проявили
глубокий интерес к соотношениям между психическими функциями и работой
мозга (блок 1.2). Фрейд начинал свою карьеру в медицине как невролог, ему
даже принадлежит одна из первых моделей нервной сети. В начале своей дея-
тельности он нашел новый химический краситель — хлорид золота, позволя-
ющий выявлять под микроскопом определенные нейроны. Впервые краситель
подобного назначения открыл Камилло Гольджи в 1873 г. Это принципиально
улучшило возможности выявления нервных клеток с помощью светового ми-
кроскопа (см. htt�://n��el�r�ze.�rg/med�c�ne/art�cles/g�lg�/).
Блок 1 .2 . Современная дискуссия о соотношении между психикой и мозгом
Философы продолжают спор о том, можно ли объяснить процессы сознания с пози-
ций науки о мозге. Многие считают, что этот вопрос сейчас опять приобрел научную
значимость. К видным философам, которые вновь обращаются к натуралистическо-
му взгляду на проблему психика–мозг, принадлежит Патрисия Чёрчланд. По ее сло-
вам, «нейробиологические данные имеют прямое отношение к давним, традицион-
ным аспектам философии, таким как природа сознания, самость («Я»), свобода воли,
знание, этика и научение».
Рис. 1.29 . Патрисия Чёрчланд принадлежит к числу видных философов, вернувших-
ся к натуралистическому подходу в проблеме соотношения психики и мозга. (Источ-
ник: Patr�c�a Churchland, UCSD.)
Психика и мозг
61
5.1.
Современные методы исследования осознаваемых
и неосознаваемых процессов в мозге
Психологи и нейробиологи разработали обширный набор методов исследова-
ния осознаваемых и неосознаваемых процессов в мозге. В табл. 1 .3 перечис-
лены аспекты психической деятельности, изучаемые современными методами.
Разумеется, каждый метод имеет свои возможности. Некоторые из них помо-
гают оценить эпизодическую память, другие показывают, что, когда человек
протягивает руку к увиденному им предмету, это происходит в существенной
мере неосознанно. Остается много нерешенных вопросов: считать ли «осозна-
ваемый» (или «сознательный») когнитивный процесс оптимальным термином,
должно ли избирательное внимание предшествовать сознательному опыту. Все
эти споры являются здравыми и нормальными. Важнейшее преимущество со-
стоит в том, что теперь мы располагаем целым арсеналом надежных методов
изучения осознаваемых и неосознаваемых процессов в мозге (рис. 1 .30). Мы
можем отличить характеристики активности мозга при восприятии реальных
событий и во время образов событий, а также от реакций при бессознательном
восприятии стимулов (см. табл. 1.3).
С научной точки зрения, очень важно учитывать, что сознательная когни-
тивная деятельность имеет много общего с оперативной памятью или внима-
нием. Такое умозаключение следует из результатов наблюдений за поведением
(Mandler, 2003) и, как правило, согласуется с нашим личным опытом. Например,
вы сможете сказать, каким шрифтом было напечатано только что прочитанное
вами слово или даже что вы ели сегодня на завтрак. Как мы увидим в последу-
ющих главах, эта проблема фигурирует в ряде направлений когнитивной нейро-
науки, таких как память, зрительное восприятие, последствия поражений мозга.
Психологи изучают сенсорные процессы с начала 1800-х гг. Мы воспринимаем мир
через посредство своих ощущений: мы чувствуем, что система психика–мозг каким-
то сложным образом участвует во взаимодействии между физическим миром и на-
шим мозгом. Мы стремимся улучшить свои способности к восприятию с помощью
очков или слуховых аппаратов, разрабатываемых специалистами. Мы совершенству-
ем аппаратуру (например, автомобильные аудиосистемы), которая обеспечивает нам
сенсорные входы. Однако степень соответствия между нашим зрительным или слу-
ховым восприятием окружающего мира и его реальными физическими свойствами
до сих пор не вполне выяснена. Разум и мозг, несомненно, имеют представление о
том, как выглядят и какие звуки издают объекты, среди которых мы находимся, но
механизм перехода от физического к психическому остается загадкой.
Вероятно, понятия «психика» и «мозг» относятся к различным источникам инфор-
мации. Если мы смотрим на мир с помощью нашего мозга (как мы всегда делаем),
перед нами предстает совсем иная картина, чем изображения мозга, которые мы по-
лучаем, например, при томографическом сканировании.
Когда компьютер показывает нам объем своей памяти, мы не думаем, что он рас-
крывает перед нами некую новую реальность. Аналогичным образом, это не проис-
ходит, когда люди говорят нам о своем сенсорном опыте. Вместе с тем они сообщают
нам нечто важное. Обсуждение проблемы психика–мозг продолжается на страницах
философских журналов, однако до сих пор ученым приходится оперировать эмпи-
рическими понятиями. Сейчас расширяется наше знание по обоим направлениям —
психологическому и нейробиологическому; при тщательном исследовании начинает
вырисовываться перспектива их сближения.
62
Глава 1
Таблица 1.3
Активно исследуемые осознаваемые и неосознаваемые явления в мозге
Осознаваемое
Неосознаваемое
1. Эксплицитные когнитивные явления
Имплицитные когнитивные явления
2. Кратковременная память
Долговременная память
3. Новые, информативные, значимые
события
Повседневные, прогнозируемые, незна-
чимые события
4. Информация от центра внимания
Информация на периферии внимания
5. Фокальный контент
Периферический контент (т. е. ранее зна-
комый)
6. Декларативная (эксплицитная)
память (факты и т. д.)
Процедурная (имплицитная) память
(навыки и т. д.)
7. Надпороговые стимулы
Подпороговые стимулы
8. Управляемые задачи
Спонтанные/автоматические задачи
9. Вспоминание
Узнавание
10. Доступные воспоминания
Недоступные воспоминания
11. Стратегическое управление
Автоматическое управление
12. Последовательности грамматик
Имплицитные грамматики
13. Интактное состояние ретикулярной
формации и билатеральных интрала-
минарных ядер таламуса
Нарушенное состояние ретикулярной
формации или билатеральных интрала-
минарных ядер таламуса
14. Повторяемые элементы оперативной
памяти
Неповторяемые элементы
15. Бодрствование и сновидения (актив-
ное состояние коры мозга)
Глубокий сон, кома, синдром седации
(медленные волны)
16. Эксплицитные умозаключения
Автоматические умозаключения
17. Эпизодическая (автобиографическая)
память
Семантическая память (концептуальное
знание)
18. Автоматическая память
Ноэтическая память
19. Интенциональное научение
Случайное научение
20. Нормальное зрение
«Слепое» зрение (кортикальная слепота)
5.2 .
Историческое развитие не остановилось
Практически все вопросы, которые обсуждались на протяжении истории разви-
тия науки, актуальны до сих пор, причем нередко выступают под новым углом
зрения (см. табл. 1 .4 и блок 1.3). Расширяется практическое приложение ког-
нитивной нейронауки. В современной неврологии активно применяются мето-
ды визуализации мозга и оценки когнитивной деятельности для диагностики
скрытых форм нарушений, которые не обнаруживаются при сканировании. С
помощью новых методов удалось идентифицировать такие дефекты, как эмо-
циональная слепота или неспособность пациента использовать определенные
существительные или глаголы. Эти трудно определяемые признаки могут ука-
зывать на серьезные патологические изменения мозга, которые иначе не вы-
являются.
Психика и мозг
63
Таблица 1.4
Проблемы, по которым продолжается дискуссия
Локализованные и распределенные функции в мозге
Нейронная доктрина
Сознание
Неосознаваемые умозаключения при зрительном восприятии
Пределы мощности мозга
Кратковременная и долговременная формы памяти: насколько они разделены?
Биологические основы эмоций
Соотношение врожденного и приобретенного, роль наследственности и внешней среды
Рис. 1 .30. Осознаваемое и неосознаваемое зрительное восприятие. В одном и том же
эксперименте проводилось сопоставление осознаваемых и неосознаваемых явлений.
Метод обратного зрительного маскирования позволяет сравнивать фМРТ-активность у
испытуемого при осознаваемом и неосознаваемом зрительном восприятии слов. Рису-
нок (Dehaene et al., 2001) показывает, что неосознаваемое восприятие слова сопрово-
ждается местной активностью в зрительной области коры; при осознаваемом восприя-
тии того же слова область фМРТ-активности расширяется на теменную и лобную доли.
В обеих экспериментальных ситуациях к сетчатке поступает одинаковый зрительный
стимул, следовательно, различия должны зависеть от более высоких уровней мозга. Ре-
зультаты были воспроизведены и другими авторами. Эти данные убедили многих уче-
ных в том, что правильно разработанная структура эксперимента позволяет исследовать
осознаваемые когнитивные процессы. (Источник: Baars et al., 2003.)
64
Глава 1
6.0.
Заключение
В этой главе мы затронули ряд вопросов, которые пытается решать когнитивная
нейронаука. Над некоторыми из них люди задумывались издавна (например,
связь психического и телесного), тогда как другие возникли в результате со-
временных экспериментов. В центре внимания в нашей книге находится про-
блема научного подхода к соотношению психики и мозга. Появление методов
визуализации мозга побудило заново обратиться к многим вопросам (блок 1.4).
Когнитивная нейронаука сочетает психологию, нейронауку и биологию.
Исторические корни этой дисциплины уходят в глубь веков к ранним докумен-
там, отражающим восточные и западные традиции. Современные исследования
Блок 1 .3 . Адаптивная или символьная система?
Еще одно направление продолжающейся дискуссии посвящено тому, какой системой
является мозг — адаптивной или символьной. Суть спора в том, как соотнести ког-
нитивные процессы с картой мозга. Предпосылки вкратце таковы.
Читая это предложение, вы осуществляете символьную интерпретацию последова-
тельности черных знаков на окрашенном фоне. Лингвисты и психологи рассуждают
о языке в понятиях символов: фонемы, буквы, морфемы, слова, фразы, предложе-
ния. Эти символьные единицы комбинируются исходя из норм грамматики и правил
применения. Человек, владеющий разговорным английским языком, понимает около
100 000 слов, некоторые из них более чем в одном значении; это приблизительно
соответствует объему хорошего словаря. Что касается более высокого уровня, то мы
мыслим предложениями, в которых заложены абстрактные концепции и утвержде-
ния; из них строятся структуры еще более высокого порядка — объяснения, беседы,
рассказы. Даже экспрессивные аспекты языка (эмоциональные интонации фраз, вер-
бальное утверждение негодования или любви) можно передать с помощью символов:
в языке много слов, обозначающих эмоций. Итак, мы искусно пользуемся символа-
ми. Они необходимы в любом явлении культуры — в науке, литературе, музыке, изо-
бразительном искусстве. Даже компьютерные программы представляют собой сим-
вольные системы. Символы окружают нас, поэтому естественно размышлять о мозге
как о процессоре, оперирующем символами.
Однако биологический мозг содержит только нейроны, которые связаны друг с дру-
гом разнообразными способами. Как мы должны представлять себе взаимосвязь пси-
хики и мозга — в виде устройства для обработки символов или как сложнейшую
сеть нейронов? Для каждого из этих подходов есть аргументы за и против. Оперируя
символами, удобно рассуждать о понятиях смысла и цели. Символьные блок-схемы
(см. гл . 2) помогают ответить на вопросы, которые нельзя проверить в лаборатории.
Однако искусственные нейронные сети позволяют моделировать простые задачи по
научению лучше, чем это удается с помощью символьных систем (см. гл . 2). Таким
образом, мы имеем набор средств, сочетающих символы и нейроны, — блок-схемы,
искусственные нейросети, семантические сети, схемы с ветвящейся структурой, мо-
дели. При внимательном рассмотрении мы обнаруживаем, что даже в нейросетях
присутствуют узлы и связи с символическими именами.
Вокруг этих вопросов постоянно идут споры. Когнитивная нейронаука бурно раз-
вивается, новые факты и теории часто заставляют нас усомниться в справедливости
прежних представлений. Это увлекательная, захватывающая область, и мы надеемся,
что в последующих главах вы узнаете о новых удивительных открытиях, связанных
с психикой и мозгом человека.
Психика и мозг
65
Блок 1 .4 Как изучать этот курс
Мы настоятельно рекомендуем вам при изучении карт мозга делать зарисовки и
раскрашивать области мозга. Как известно, обучение требует активного участия.
Это очень важно для данного курса. Необходимо внимательно читать текст, раз-
мышлять о прочитанном, задавать вопросы и обдумывать ответы. Помогают со-
вместные занятия. Поскольку картина поверхности мозга очень сложна, особенно
полезно делать зарисовки. Материал для графических упражнений есть в конце
большинства глав.
Традиционный метод обучения включает препаровку мозга овцы и лягушки. Это
эффективно, потому что студент имеет дело с реальным мозгом. Постарайтесь по-
дойти к изучению текста и графического материала книги так, как будто перед вами
находится настоящий мозг, только без повреждения живого организма. На наших ри-
сунках показаны разные анатомические уровни и иллюстрируются наиболее важные
эксперименты.
Мозг — это емкое трехмерное образование, которое в результате миллионов лет эво-
люции сконцентрировалось до малых размеров. Он получает данные о самом себе и
внешней среде подобно тому, как мы узнаем устройство города, совершая прогулки
и поездки, запоминая основные ориентиры, посещая дальние переулки и отдельные
дома. Эта книга послужит для вас путеводителем, сообщая об ориентирах и о жизни
обитателей.
Главные пункты, существенные для обучения, сопровождаются иллюстрациями, схе-
мами или экспериментальными данными. В последующих главах вы найдете описа-
ния клинических случаев поражения мозга, а также получите сведения о границах
возможностей нормального мозга.
Наконец, следует разобраться в терминологии. Изучение анатомии мозга началось
четыре столетия назад, когда международным языком науки был латинский. До сих
пор названия многих структур мозга основаны на латинских и греческих словах. На-
пример, в этой главе представлены виды коры больших полушарий сбоку («lateral»),
по средней линии среза («med�al»), снизу («�nfer��r») и сверху («su�er��r»). Эти четы-
ре английских слова происходят от латинских «lateral�s, med�al�s, �nfer��r, su�er��r» —
боковой (латеральный), срединный (медиальный), нижний, верхний. Англичанам
повезло в том отношении, что значительная часть словарного состава их языка осно-
вана на латыни, так что нетрудно находить ассоциации со знакомыми словами.
Вы легче выучите новые слова, если будете не просто повторять их, но и проду-
мывать смысл. Тыльную часть мозга принято называть дорсальной, поскольку по-
латыни «d�rsum» — спина. Посмотрите на собаку или кошку — тыльная часть их
головы является продолжением спины. Как мы уже отмечали выше, боковая сторона
называется латеральной; следовательно, структуры, находящиеся на тыльной сторо-
не и в то же время сбоку, обозначаются как дорсолатеральные. Мы будем объяснять
составные термины по мере их упоминания.
При изучении анатомии студенты часто используют техники запоминания. Может
быть, вы попробуете их и выберете самую подходящую для себя. Студенты-медики
обычно зарисовывают изображения мозга, раскрашивая отдельные части рисунка.
В принципе помогают любые активные ассоциации — подбирать рифмы для слов,
создавать зрительные образы, находить аналогии. Может быть, вы вообразите мозг
в виде автомобиля, ассоциируя определенные структуры с колесами или капотом.
Пусть вас не смущает, если ассоциации окажутся нелепыми, лишь бы у вас возникла
четкая мысленная связь между тем, что вам нужно запомнить, и тем, что вам уже
известно. Так же как нет неподобающих вопросов, не бывает и глупых методик. Все
дело в том, помогает вам это или нет.
66
Глава 1
посвящены темам, которые были недоступны всего лишь несколько лет назад.
Для студентов открываются новые профессиональные направления. Возникают
практические приложения в сферах образования, медицины, психотерапии. Са-
мые сложные задачи не решены, но чувствуется волнующая атмосфера ожида-
ния. Сведения о новых открытиях поступают быстрее, чем когда-либо прежде.
В этой главе мы хотели дать общее представление о комплексном изучении
психических явлений и мозга. Сейчас уже есть результаты визуализационных
исследований мозга в связи со зрением и слухом, научением и памятью, осозна-
ваемыми и неосознаваемыми процессами, зрительными образами, внутренней
речью. Наука получила поистине исключительные возможности. В большин-
стве визуализационных исследований мозга используются стандартные когни-
тивные задачи, что способствует интеграции поведенческих и нейрологических
данных. Практический выход для сведений о деятельности мозга в нормальных
и в патологических условиях чрезвычайно велик.
Когнитивная наука стремится осмыслить поток новых фактов. В этой главе
даны общие исходные сведения. Наша цель — показать, что объединение пред-
ставлений о психических и нейрологических процессах часто приводит к не-
ожиданным и ценным результатам.
7.0 .
Задания и упражнения к главе 1
7.1.
Контрольные задания
1. Назовите три явления в мозге, для которых характерны малые простран-
ственные размерности.
2. Назовите три явления в мозге, для которых характерны малые временные
размерности.
3. Какие представления о природе психики преобладали в XIX в.? В начале
ХХ в.?
4. Каковы главные различия между бихевиоризмом и когнитивной психоло-
гией?
5. Какое влияние оказали на развитие психологии такие физиологи, как
И. П . Павлов?
6. Расскажите о возможностях взаимодействия между психологией и наукой о
мозге.
Наконец, необходимый компонент обучения — это самопроверка, как при игре на
музыкальном инструменте. Иначе мы склонны переоценивать себя. Имейте в виду,
что способность узнавать слово еще не означает, что вы сможете вспомнить его,
когда это понадобится. Если вам предстоит проверка знания терминов, нужно по-
тренироваться, вспоминая их.
Полезно поискать в сети Интернет данные когнитивных экспериментов и визуали-
зации мозга. Национальная медицинская библиотека США (NLM) размещает в от-
крытом доступе базу данных, которая содержит рефераты десятки миллионов публи-
каций. PubMed даст ответы на любые вопросы научного и клинического характера.
Существуют прекрасные сайты по анатомии мозга, а также много сайтов, демонстри-
рующих фундаментальные когнитивные и перцептивные явления.
Психика и мозг
67
7. В чем трудности научного исследования поражений мозга?
8. Каково соотношение между подходом с позиций «психического» и «сомати-
ческого»? Что представляет собой философский натурализм?
9. В своей повседневной жизни отдаете ли вы себе отчет в том, что пользуетесь
внутренней речью? А зрительными образами? Если да, то в каких случаях?
Если нет, то в каких ситуациях подобные явления все же могли бы произой-
ти? (Учтите существенные индивидуальные различия зрительного вообра-
жения; спонтанные зрительные образы возникают не у каждого человека.)
7.2 .
Графические упражнения
Мы настоятельно рекомендуем использовать зарисовки и раскрашивание как
методику, помогающую запоминать местоположение мозговых структур.
Представленная здесь схема поможет вам, когда вы будете рисовать и рас-
крашивать области мозга, перечисленные далее. Многие считают полезным
закрашивать одну и ту же область постоянным цветом, это способствует об-
разованию ассоциаций между определенной структурой и цветом. Старайтесь
включать все виды памяти.
1. Основные доли мозга (внешний вид справа и слева).
2. Названия основных долей.
3. Где расположена зона Брока? Зона Вернике? Их главные функции?
4. Какая анатомическая структура соединяет эти две зоны?
5. Нарисуйте схему мозга, вид слева, обозначьте основные доли.
6. Какие области ассоциируются со зрением? С речью? С управлением?
По-видимому, наш мозг должен сначала независимо сформировать образы
признаков, прежде чем мы сможем найти их в реальных объектах... Знание
не может возникнуть на основе одного лишь опыта, оно создается только
в процессе сопоставления результата интеллектуальной деятельности с на-
блюдаемыми фактами.
Альберт Эйнштейн (1949)
Функциональная схема, используемая на протяжении всей книги, сочетает две классиче-
ские модели когнитивного (познавательного) процесса (Baddeley, Hitch, 1974; Atkinson,
Shiffrin, 1968). Две желтые стрелки символизируют произвольное внимание (реализу-
емое по принципу «сверху вниз») и спонтанное внимание («снизу вверх»). В нижней
части рисунка представлены долговременная память, знания, навыки. Центральное ме-
сто на схеме принадлежит рабочей памяти согласно модели Баддели. В гл. 8 добавится
понятие «сознательная когнитивная деятельность» для обозначения явлений, которые
происходят в мозге, например при осознаваемом восприятии текста на лежащей перед
вами странице (Baddeley, 2002). Хотя схема имеет самый общий характер, ее достоин-
ство в простоте и широком подходе. Она дает первоначальное представление о функци-
ях психики и мозга.
Содержание
1.0.Введение........................................................69
2.0.Классическоепонятиеоперативнойпамяти............................71
2.1.«Внутренниечувства» .........................................72
2.2.Выходныефункции............................................74
2.3.Одинкраткиймиг...............................................74
2.4 . Случай Клайва Уэринга с позиций функциональной схемы . . . . . . . . . . 78
2.5.Рольмгновеннойпамяти........................................79
3.0.Ограниченностьфункциональныхспособностей........................81
3.1.Границывыполнениядвойныхзадач..............................81
3.2 . Некоторые функциональные способности мозга имеют
широкиеграницы..............................................84
3.3 . Почему функциональные способности могут быть
стольограниченными?..........................................85
3.4.Оценкаоперативнойпамяти.....................................86
4.0.Внутренниеивнешниечувства......................................89
4.1.Глаз,ухоиголоспсихики.......................................90
4.2 . Психические образы формируются в зрительных областях коры мозга . 92
4.3.Идентичналивнутренняяречьвнешнейречи?.....................92
4.4.Тольколиодинвидоперативнойпамяти?..........................93
5.0.Центральноеуправление...........................................93
5.1.Исполнительнаяреакцияиавтоматизм............................96
5.2.Управляемоеиспонтанноевнимание.............................98
6.0.Действие........................................................100
7.0 . Консолидация кратковременных событий в долговременную память . . . . . . 104
7.1 . Оперативная память — результат активации постоянной памяти? . . . . 106
8.0.Заключение.....................................................107
9.0.Заданияиупражнениякглаве2....................................108
9.1.Контрольныезадания..........................................108
9.2.Графическиеупражнения......................................109
1.0 . Введение
В этой главе рассматривается концептуальная схема, которая существенно об-
легчит структурирование когнитивной нейронауки. Хотя согласие достигнуто
не по всем вопросам, схема позволяет объединить значительную часть нейро-
биологических и психологических данных. С учетом сложности когнитивной
нейронауки схема будет полезна в процессе усвоения материала.
Глава 2
Концептуальная структура
когнитивной нейронауки
70
Глава 2
На схеме (с. 68), а также на рис. 2 .1 показана ассоциация некоторых обла-
стей коры больших полушарий с определенными функциями. Как мы увидим
далее, каждый компонент нашей схемы (с. 68) основывается на комплексных
данных — психологических и нейробиологических. Однако мы не настаиваем
на том, что приведенная схема является единственным вариантом толкования
фактов. По тем пунктам, где существуют разногласия, мы приведем доводы за
и против.
В этой главе мы коснемся всех аспектов функциональной схемы. Более под-
робное обсуждение каждого ее компонента вы найдете в последующих главах.
На левой части схемы (с. 68) обозначены сенсорные системы. Каждая из
них начинается миллионами периферических рецепторных клеток (например,
нейроны сетчатки в глубине глаза). Все сенсорные пути заканчиваются в зад-
ней половине коры больших полушарий (рис. 2 .1). Считается, что информация,
Рис. 2 .1 . Основные функции коры больших полушарий человека. В верхней части ри-
сунка представлен вид коры снаружи сбоку (латеральный), внизу — на сагиттальном
срезе по средней линии. Запомните основные функциональные зоны (они окрашены на
рисунке в различные цвета), чтобы легче понимать последующий текст книги. Вверху
и внизу справа показаны сенсорные области коры: задняя кора, включая затылочную
(зрительные зоны) и часть верхней височной коры (слуховые зоны). Представительства
соматической чувствительности находятся непосредственно позади центральной бороз-
ды (светло-зеленый цвет). Слева от центральной борозды расположены двигательные
отделы (розовый цвет), а перед моторной корой – предфронтальная кора, выполняю-
щая функции управления. Таким образом, кору мозга удобно подразделить (по крайней
мере, в первом приближении) на заднюю половину, где содержатся входные области, и
переднюю половину с выходными областями. (На этом рисунке не показаны подкорко-
вые области, а также проводящие пути между областями входа и областями выхода.)
(См. гл . 5 .) (Источник: рисунок Shawn Fu.)
Концептуальная структура когнитивной нейронауки
71
соответствующая каждой из классических сенсорных модальностей, способна
в течение короткого времени сохраняться — так называемый сенсорный бу-
фер. (В русскоязычной литературе термин «буфер» иногда заменяют на «ре-
гистр» — Прим. ред. перев.) Изучение зрения и слуха продолжается более чем
два столетия, так что об этих органах чувств известно многое. Достаточно хо-
рошо исследованы и такие виды соматической чувствительности, как осязание,
боль и проприоцепция. Однако хотя обоняние и тактильная чувствительность —
эволюционно наиболее древние сенсорные модальности, сведения о них менее
подробны. До сих пор обнаруживаются прежде неизвестные формы чувстви-
тельности, в частности связанные с циркадианными ритмами, с пищеварением,
даже с половой и репродуктивной функциями.
Восприятие сенсорных стимулов может усиливаться при избирательном
внимании (см. схему на с. 68, две желтые стрелки). Как показывает схема, вклю-
чение внимания может произойти по принципу «снизу вверх» («bottom-up»),
когда сенсорный сигнал поступает на вход внезапно — при вспышке света,
неожиданном звуке или невольном взгляде на чье-то лицо. Мы можем также
переключать внимание произвольно, по принципу «сверху вниз» («top-down»),
например, если вам предложат посмотреть или послушать что-то интересное.
Хотя продолжается дискуссия относительно участия конкретных областей моз-
га в избирательном внимании, появились сведения, что области коры, обеспе-
чивающие зрительное внимание, в существенной мере перекрываются с зона-
ми, которые управляют движениями глаз (см. гл . 8).
На упомянутой схеме не обозначены осознаваемые действия, однако, как мы
увидим в гл. 8, между избирательным вниманием и сознательным восприятием
существует тесная связь.
2.0 .
Классическое понятие оперативной памяти
В средней части схемы (с. 68) показаны классические компоненты оперативной
(рабочей) памяти (Baddeley, Hitch, 1974; Baddeley, 2002), которые подробно из-
учаются в течение трех последних десятилетий.
В высших отделах мозга находится центральное управление оперативной
памятью — предмет текущих исследований. Вначале центральное управление
рассматривалось применительно к задаче научения. Однако по мере углубления
сведений о лобных долях коры выяснилось, что их управляющее влияние рас-
пространяется на все аспекты произвольной деятельности (например, данные
Luria, 1976; Goldberg, 2001; cм. гл . 8 и 11). В описаниях функций этих цен-
тров коры нередко фигурирует метафора «генеральный директор». Обратимся
к примеру.
В чем состоит роль управляющего крупной компании, который контролирует всю
ее деятельность? Он определяет цели и планы, организует работу компании, на-
правленную на достижение этих целей, устанавливает сроки начала и завершения
конкретных действий, время перехода к другим действиям или прекращения всякой
деятельности.
Руководитель ориентирован на будущее и на достижение цели; чтобы добиться ре-
зультата, он должен обладать гибкостью и адаптивностью. Именно эти характери-
стики личности зависят от префронтальной области коры больших полушарий чело-
века (Mateer et al., 2005; курсив автора книги).
72
Глава 2
Префронтальная зона соответствует передним участкам лобных (фронталь-
ных) долей коры и находится перед моторными зонами (которые на рис. 2 .1
обозначены ярко-красным цветом). На рис. 2 .1 префронтальные области окра-
шены в бежевый и желтый цвета (вверху — вид полушария мозга снаружи сбо-
ку, внизу — вид изнутри). Обратим внимание, что центр Брока (впервые обна-
руженный в XIX в.) морфологически входит в состав префронтальной коры.
Некоторые управляющие функции принадлежат также моторным зонам в боко-
вой части каждого полушария (розовый цвет на рис. 2 .1), хотя их роль сужена
по сравнению с той, которая свойственна классической префронтальной зоне.
Обозначенные границы нельзя считать абсолютными, но они удобны в каче-
стве первого приближения. В дальнейшем тексте (см. гл . 12) управляющая роль
коры мозга будет рассмотрена подробнее.
Под центральным управляющим блоком на схеме (с. 68) расположен эле-
мент оперативного хранения информации. Считается, что функцию оператив-
ной памяти обеспечивают медиальная височная область и префронтальная
кора мозга. Оперативное хранение информации динамично, поскольку в нем
участвуют активные популяции нейронов — так называемые клеточные ан-
самбли, при консолидации активности которых формируется долговременная
память. Вследствие динамичности хранения информации оперативная память
более уязвима, чем постоянная.
2.1.
«Внутренние чувства»
Ниже блока оперативного хранения информации на схеме находятся блоки
двух видов «внутренних чувств» — вербального повторения и обработки зри-
тельно-пространственной информации. Они постоянно взаимодействуют с
блоками длительного хранения информации (показаны серым цветом внизу).
Сейчас считается, что термин вербальное повторение — это синоним понятия
внутренняя речь. А именно в период бодрствования человек почти все вре-
мя разговаривает сам с собой (Luria, 1976). Внутренняя речь нужна не про-
Luria, 1976). Внутренняя речь нужна не про-
, 1976). Внутренняя речь нужна не про-
1976). Внутренняя речь нужна не про-
1976). Внутренняя речь нужна не про-
сто для повторения и запоминания информации, мы таким образом постоянно
обсуждаем наши «текущие заботы». При этом функция голосового тракта за-
торможена и внутренняя речь не озвучивается (Singer, 1993). Поскольку в этот
процесс вовлечены богатые языковые возможности человека, внутренняя речь
непосредственно связана с лингвистическим и с семантическим компонентами
длительного хранения информации (блоки в нижней части схемы).
Основные особенности вербального повторения легко продемонстрировать
на задачах мгновенной памяти, например когда человек хочет запомнить номер
телефона или список предстоящих покупок. Как можно ожидать из материала
гл. 1, внутренняя речь происходит с участием центров Брока и Вернике (см.
рис. 2 .1, желтые и оранжевые области). Однако области, которые связаны с ре-
чью, выходят далеко за пределы традиционных зон Брока и Вернике. Как мы
увидим далее, в обеспечение семантики при формулировании абстрактных кон-
цепций вовлекаются также височные и лобные доли.
Система обработки зрительно-пространственной информации имеет отноше-
ние к нашей способности временно сохранять зрительные и пространственные об-
разы, например место парковки автомобиля или путь от дома до бакалейного мага-
зина. Зрительные образы нетрудно продемонстрировать, если, скажем, попросить
человека, чтобы он представил себе входную дверь в его дом, а потом спросить его,
Концептуальная структура когнитивной нейронауки
73
с какой стороны расположена дверная ручка. Даже человек, не наделенный ярким
воображением, обычно «видит» ручку на правой или левой стороне своей вооб-
ражаемой входной двери. Косслин и другие выявили много нейробиологических и
поведенческих черт сходства между зрительными образами и зрительным восприя-
тием (например, Kozhevnikov et al., 2005). По мнению этих авторов, зрительные об-
разы в принципе зависят от совокупности центров зрительного восприятия (синие
области на рис. 2 .1).
Вместе с тем в формирование зрительно-пространственной картины также
вовлекается более абстрактная и перекрестно-модальная (связанная с различ-
ными видами чувствительности) информация. Мы можем с закрытыми глазами
на ощупь идентифицировать предмет, даже если мы никогда раньше не при-
касались к нему. Очевидно, между центрами зрительного и тактильного вос-
приятия существует перекрестная передача информации. Такой перекрестный
поток информации ассоциируется с теменной корой (на рис. 2 .1 показана зе-
леным цветом), которая выполняет функцию «куда направить», в отличие от
функции «что направить», осуществляемой зрительными центрами нижнего
уровня. Слуховая чувствительность тоже имеет пространственный компонент:
мы достаточно точно можем с закрытыми глазами определить, откуда посту-
пают звуки, особенно если в них есть быстрые высокие составляющие (напри-
мер, чириканье воробья). Итак, каждая сенсорная система начинается с домен-
специфического восприятия зрительных, слуховых, тактильных стимулов, но
вскоре входит в состав общего полимодального пространства — среды, окру-
жающей наш организм.
В обработке зрительно-пространственной информации и вербальном по-
вторении участвуют умственные способности, благодаря которым мы запо-
минаем новые слова, новые лица, пространственные характеристики (напри-
мер, на каком перекрестке следует повернуть направо) и т. д. Эти повседневно
реализуемые способности возникли на основе эволюции языка и восприятия
пространства. Вероятно, «внутренние чувства» участвуют также в обонянии и
вкусе, соматической чувствительности, боли и удовольствии и т. д . Показано,
например, что при ожидании боли активируются те же области мозга, которые
активны при реальных болевых ощущениях. Однако наиболее подробно изуче-
ны вербальная и зрительно-пространственная «внутренняя чувствительность».
На них мы и сосредоточимся.
В нижней части схемы (с. 68) представлены блоки долговременной памяти.
Здесь хранятся автобиографические эпизоды, разнообразные знания, привыч-
ки. Такая информация после ее перехода в долговременную память становится
бессознательной. Однако она постоянно взаимодействует с текущей деятель-
ностью.
Все части системы работают в сотрудничестве с другими, но иногда могут
и конкурировать между собой. Например, между зрительными и слуховыми
стимулами возникает конкуренция, если они не интегрируются в единое пере-
живание. Хороший пример — нарушение синхронности между зрительным
изображением и звуком при показе кинофильма. Если несовпадение между зри-
тельным и звуковым треками превышает одну десятую долю секунды, воспри-
ятие каждого из них нарушается. При синхронизации треков они объединяются
на уровне восприятия (McGurk аnd MacDonald, 1976). Мы относим речь к тому
персонажу, артикуляция которого соответствует издаваемым звукам.
74
Глава 2
2.2 .
Выходные функции
На правой стороне схемы представлены компоненты выхода: центральное
управление, планирование действия и двигательный выход. Наиболее вырази-
тельный пример — произвольные двигательные функции, особенно управление
скелетными мышцами, т. е. мышцами туловища, головы, лица. Мы не можем
произвольно контролировать сокращения гладких мышц пищеварительного
тракта, кровеносных сосудов или желез, выделяющих жизненно важные гормо-
ны. Давно известно, что система регуляции этих физиологических функций не
зависит от произвольного контроля, поэтому она называется автономной (или
вегетативной) нервной системой. Автономная нервная система анатомически
отделена от системы произвольного контроля движений, центры управления
которой находятся в лобных долях коры мозга.
Человек произвольно контролирует и некоторые психические функции.
Примеры — вербальное повторение и мысленное переворачивание зритель-
ных образов. Если вы представите себе кухонный табурет, вы можете в своем
воображении перевернуть его сиденьем вниз. Во время таких воображаемых
действий активируются моторная, пространственная (теменная) и зритель-
ная области коры мозга. Поскольку эти задачи осуществляются произвольно,
управление принадлежит, по-видимому, префронтальным областям.
Далее мы увидим, как можно использовать функциональную схему при рас-
смотрении трагического случая поражения мозга, когда человек лишился спо-
собности переводить мгновенную информацию в долговременную память.
2.3 .
Один краткий миг.. .
Это случилось в Великобритании в 1985 г. Молодой перспективный музыкант
Клайв Уэринг вдруг забыл, как зовут его жену. В тот же день он не смог вспом-
нить имена своих двух детей (рис. 2 .2). Незадолго до этого состоялось офици-
альное бракосочетание Клайва и Деборы, однако он не помнил и этого. Его со-
стояние, которое было перманентным, возникло неожиданно после сильнейшей
головной боли и высокой температуры, продолжавшихся два дня. За это время
вирусная инфекция нарушила области мозга, необходимые для формирования
памяти. Уэринг был поражен хронической амнезией с полной потерей эпизоди-
ческой памяти (о событиях прошлого) и частичной утратой семантической па-
мяти (способности понимать смысл некоторых понятий) (Wilson et al., 1995).
Что особенно важно — он перестал усваивать новую информацию. Его жизнь,
как позднее выразилась Дебора, внезапно сузилась до «одного краткого мига»,
без прошлого и будущего.
Клайв Уэринг стал на 20 лет объектом сообщений, исследований, доку-
ментальных телевизионных передач. Этот случай особенно драматичен, если
учесть, что Уэринг не изменился как личность. Он полностью осознает окружа-
ющее, читает и пишет, поддерживает беседы на сиюминутные темы. Он даже
дирижирует своим небольшим хором при исполнении тех классических про-
изведений, которые знает с прежних времен. Это человек с сильными пере-
живаниями, может быть, даже более эмоциональный, чем прежде, особенно в
проявлении чувств к своей жене Деборе.
Клайв Уэринг существует в вечно настоящем времени. В течение первых
восьми лет он каждый день в больничной палате заполнял дневник, пытаясь
Концептуальная структура когнитивной нейронауки
75
вернуть то, что он называет своим сознанием. Через каждые несколько минут
он отмечал время суток, восклицая: «Вот теперь я, наконец, пришел в созна-
ние!» Вскоре он зачеркивал предыдущую запись, полагая, что на самом деле
не находился в сознании, поскольку не может вспомнить тот момент. Все это
повторялось тысячи раз, с заполнением множества дневников. Когда его наве-
щали жена или друзья, он обращался к ним так, как если бы никогда не видел
их раньше: эти люди были знакомы ему, но он не мог назвать их. Если они вы-
ходили из комнаты на несколько минут, Уэринг уже не помнил, что недавно они
были здесь. Даже теперь, встречаясь с Деборой, он считает, что давно не видел
ее (синдром известен как синдром Корсакова. — Прим. ред. перев.) . Историю
Клайва и Деборы Уэринг можно прочитать в книге, которую написала сама Де-
бора (Wearing, 2005).
О Клайве Уэринге в личностном отношении известно больше, чем о других
людях с аналогичным поражением мозга. Что касается научных исследований,
то наиболее подробно они были проведены в случае пациента, известного под
инициалами ХМ (Scoville and Milner, 1957). В 1950-е гг. было мало лекарствен-
ных средств от эпилептических припадков. При тяжелой неконтролируемой
эпилепсии применялось в качестве крайней меры хирургическое удаление ча-
сти височной доли. У пациента ХМ были удалены с обеих сторон гиппокамп и
часть окружающей его ткани медиальной височной доли (рис. 2 .3). Подробное
наблюдение в течение нескольких десятилетий показало, что ХМ не запоми-
нал новые автобиографические эпизоды, т. е. потерял память об осознаваемых
событиях. Вместе с тем ХМ сохранил процедурную память — способность
приобретать новые сенсомоторные навыки. Так же как Клайв Уэринг, ХМ не
утратил возможность понимать, что говорят и что происходит вокруг него. Сле-
довательно, у него не было серьезного повреждения семантической памяти —
способности понимать смысл явлений.
Рис. 2.2 . Клайв Уэринг (и его жена Дебора),
портрет на обложке книги Деборы Уэринг. По-
сле двустороннего поражения гиппокампа (а
также частичного поражения лобных долей)
Клайв Уэринг сохранил способность играть на
фортепьяно и дирижировать исполнением му-
зыкальных произведений, которые он знал до
своего заболевания. При этом он не мог запо-
минать новые эпизодические (сознаваемые) со-
бытия. Сохранение осознаваемой информации
продолжалось у него в течение 10–20 с, следо-
вательно, мгновенная память, по-видимому, не
была нарушена. Однако катастрофически по-
страдало эпизодическое научение, т. е. переход
осознаваемой информации в долговременную
эпизодическую память. (Источник: Wearing,
2005.)
76
Глава 2
Представление о кардинальных различиях между автобиографической (эпи-
зодической), процедурной и семантической памятью возникло в результате
многолетних исследований, в которых участвовали пациенты с поражениями
мозга и здоровые люди, а также проводились эксперименты с повреждения-
ми мозга у животных. Наряду с этими видами памяти наш мозг обладает об-
ширной перцептуальной памятью, т. е . человеку свойственны длительные
изменения способности воспринимать окружающий мир. Например, новые
возможности перцептуальной памяти могут быть задействованы, когда чело-
век обучается различать музыкальные инструменты во время прослушивания
новой песни. По мере того как ребенок растет, он начинает воспринимать звуки
речи, постоянные зрительные объекты, различать лица и голоса — все это про-
исходит благодаря перманентной перцептуальной памяти. У Клайва Уэринга
не пострадала развитая у него до болезни способность воспринимать музыку и
Рис. 2.3 . Повреждение гиппокампа у пациента ХМ (результаты томографического ис-
следования). Слева: корональный срез мозга ХМ, от одного уха до другого. Справа: срез
нормального мозга для сравнения; две стрелки указывают местоположение гиппокампа
в одном и в другом полушарии. Гиппокамп трудно визуализировать на поперечных сре-
зах, так как он образует загиб и скрыт внутри височной доли (см. рис. 2 .4). В мозге ХМ
на месте удаленных гиппокампов образовались полости с жидкостью; на томограммах
они выглядят как темные участки. (Источник: Hodges and Graham, 2001.)
Рис.
2.4. «Виртуальное изображение» гип-
покампа правой и левой височных долей при
«взгляде насквозь» через кору мозга. Красным
цветом обозначены миндалины; они играют
принципиально важную роль в эмоциях. Сосед-
ние области медиальной височной доли необхо-
димы для обеспечения памяти (см. рис. 2 .5).
Концептуальная структура когнитивной нейронауки
77
получать от нее удовольствие. (Опытные музыканты, у которых натренирована
перцептуальная память по отношению к музыке, воспринимают больше аспек-
тов музыкального произведения, чем новички.)
Существуют и другие долговременные способности. Люди обладают об-
ширными знаниями о своем родном языке, культуре и окружающем их мире.
Образованный носитель английского языка понимает примерно 100 000 слов,
каждое из которых связано с сетью ассоциированных знаний. Мы обладаем на-
выками грамотного письма, изложения своих мыслей, межличностного обще-
ния. По большей части это знание является бессознательным в любой конкрет-
ный момент времени (Bargh et al., 2006). По имеющимся сведениям, у Клайва
Уэринга лингвистическое и семантическое знания не изменились, даже при
таком серьезном поражении мозга.
Человек обитает в богатом визуальном мире, о котором нам известно многое.
Так, мы знаем, что яйцо может упасть со стола и разбиться, но немыслимо, что-
бы стол падал с яйца. Нелепость второго высказывания соответствует нашему
бессознательному знанию о зрительно воспринимаемом мире. У нас нет причин
для вывода о нарушении зрительного мира Клайва Уэринга и пациента ХМ.
Наконец, существует декларативное знание, т. е. способность человека
вспоминать факты и мнения. Сюда относится то, что мы выучили в школьные
годы и «заявляем» как неоспоримое утверждение: «Китай — большая страна»,
«Кит — морское млекопитающее» и т. д. Такое ранее приобретенное деклара-
тивное знание тоже не страдает при повреждении гиппокампа. Судя по всему, у
пациентов, подобных ХМ и Клайву Уэрингу, сохраняются предсуществующие
знания и память. И тем не менее, как пишет Дебора Уэринг, жизнь ее мужа
была разбита. Величайшим лишением стала утрата способности сохранять в
памяти повседневную информацию и извлекать ее.
Несмотря на продолжающуюся дискуссию относительно функции самого
гиппокампа, нет сомнений относительно важной роли гиппокампальной фор-
мации, расположенной в медиальной височной доле (рис. 2 .4 и 2.5). Теперь
«медиальная височная доля» и «гиппокампальная формация» считаются еди-
ной структурой. Отметим, что каждый из двух гиппокампов (в правой и левой
височных долях) представляет собой сложное образование в виде двойной пет-
ли; два гиппокампа соединены поперек средней линии мозга.
Рис. 2 .5 . Вид мозга снизу; показано, какие участ-
ки были хирургически удалены у пациента ХМ.
Сейчас считается, что для минимального срока
сохранения информации необходим не только гип-
покамп, но, по-видимому, вся область медиальной
височной доли (на рисунке окрашена). Термин «ме-
диальная» относится к средней линии мозга, иду-
щей спереди назад
78
Глава 2
Медиальную височную долю удобнее различить, если смотреть на основа-
ние мозга снизу — это еще один вариант изображения мозга, где нужно уметь
ориентироваться. Мозг — высокоорганизованный орган, эволюционное раз-
витие которого продолжалось сотни миллионов лет, причем он представляет
собой очень компактное образование. Учитывая это, важно научиться рас-
познавать такую структуру, как гиппокамп, при взгляде с двух-трех позиций
(рис. 2 .5).
Подобно пациенту с расстройством речи, исследованному Брока в 1860-е гг.,
пациент ХМ являет собой один из классических случаев в истории науки; ему
посвящено более сотни научных публикаций. Случаи ХМ и других пациентов
чрезвычайно интересны в научном плане, тогда как ситуация Клайва Уэринга
дает богатый материал в личностном контексте. Дебора Уэринг провела мно-
голетнюю работу, благодаря которой о последствиях поражения гиппокампа
узнал широкий круг людей (Wearing, 2005). Обстоятельства личной трагедии
таких пациентов дают нам принципиально важные сведения о психике и мозге.
Каждый из двух гиппокампов (по одному в правом и в левом полушариях)
представляет собой вытянутую петлю в глубине височной доли. Гиппокамп и
соседние структуры медиальной височной доли не видны при взгляде на мозг
снаружи, но они совершенно необходимы для запоминания информации, ее об-
работки и извлечения из памяти.
2.4 .
Случай Клайва Уэринга с позиций функциональной схемы
Пациент ХМ и Клайв Уэринг лишились способности переносить в память си-
юминутно поступающую информацию. На структурной схеме (рис. 2.6) пред-
ставлена гипотеза, объясняющая такого рода поражение. Прежде всего рассмо-
трим, какие функции остались интактными после поражения гиппокампа в
обоих полушариях. У Клайва Уэринга не пострадали сенсорные функции, в
том числе восприятие музыки; сохранились сознательное восприятие и цен-
тральное управление, а также двигательное управление (хотя возникли некото-
рые нарушения двигательной сферы, по-видимому, не связанные с поражением
гиппокампа); не были затронуты некоторые виды долговременной памяти, за
исключением полной потери способности запоминать и извлекать из памяти
сведения о событиях собственной жизни; даже присутствует мгновенная па-
мять в течение примерно 10 с. Уэринг прекрасно говорит и понимает речь.
В большинстве тестов на уровень интеллекта он получает достаточно высокие
баллы, кроме тестов, касающихся запоминания и извлечения автобиографиче-
ской информации.
Клайв Уэринг тяжело переживает свое состояние, т. е . он понимает, что
произошло нечто очень серьезное. Для него также характерна высокая эмоци-
ональность в отношениях с другими людьми. Именно то обстоятельство, что
Клайв Уэринг остался нормальной личностью, делает его ситуацию незауряд-
ной и вместе с тем бесконечно трагической.
Итак, главные дефекты Клайва Уэринга не относятся к большинству блоков
функциональной схемы (рис. на с. 68). По-видимому, в его случае произошел
разрыв прежде всего между переходом информации из мгновенной памяти в
долговременную и обратно, т. е . между кодированием информации об осозна-
ваемых событиях и ее извлечением (рис. 2 .6).
Концептуальная структура когнитивной нейронауки
79
2.5 .
Роль мгновенной памяти
Мгновенная память необходима даже при выполнении самых простых дей-
ствий. Если вы не запомнили начало этого предложения, вы не сможете по-
нять его конец. Чтобы прочитать предложение, нужно несколько секунд,
следовательно, вы должны сохранить информацию в течение этого времени.
Аналогичным образом, если информация, полученная при фиксации взора на
одной точке, не сохранится в вашем мозге в течение всего периода сканирова-
ния остальной части зрительного поля, вы не сможете объединить отдельные
элементы картины перед вашими глазами (рис. 2 .7). Наконец, если вам нужно
поесть, но вы не можете удержать в уме эту задачу в течение времени, достаточ-
ного для необходимых действий, вы можете остаться голодным. Исследования,
проведенные в течение последних 50 лет, показывают, что мгновенная память
участвует во всех сенсорных, двигательных и когнитивных функциях*
.
Мы многое узнали из описаний случаев Клайва Уэринга и пациента ХМ. Изучение
гиппокампальной формации остается актуальным направлением. Поскольку меди-
альная височная доля относится к филогенетически древнему обонятельному отделу
мозга, составляющему значительную часть мозга первых рептилий и млекопитаю-
щих, она выполняет целый ряд разнообразных функций. Кроме того, в медиальной
височной доле происходит конвергенция разных сенсорных модальностей.
Как мы увидим, долговременная память может локализоваться во многих от-
делах мозга. Возможно, это вся кора и даже подкорковые структуры — базаль-
* Есть данные также о существовании промежуточной памяти, продолжительность которой состав-
ляет несколько часов или дней, но не дольше: человек помнит, где он оставил свой велосипед, какие
продукты предстоит купить на обед, как ориентироваться в новой среде, с кем и как он недавно об-
щался (Delaney et al., 2004).
Рис. 2 .6 . Функциональная схема применительно к ситуации поражения медиальной
височной доли. Отметим, что в классических случаях поражения медиальной височ-
ной доли большинство когнитивных функций сохраняется. Однако пациенты теряют
способность преобразовывать для длительного хранения (запоминать) информацию о
сознательном опыте и впоследствии извлекать ее (вспоминать).
80
Глава 2
ные ганглии и мозжечок (см. далее). Однако почти все участки, которые показа-
ны на рис. 2.1, являются эволюционно более новыми, это так называемая новая
кора, неокортекс (противопоставляемая древней коре мозга рептилий и низших
млекопитающих). В ходе филогенеза млекопитающих новая кора существенно
увеличилась в размерах, особенно за последние три миллиона лет, в течение
которых происходила эволюция приматов и человекообразных обезьян. Область
гиппокампа называется «древней корой» — палеокортексом. В то время как в
неокортексе четко различаются шесть клеточных слоев, палеокортекс подраз-
деляется только на четыре или пять слоев. У человека и других млекопитающих
гиппокампальная формация постоянно сотрудничает с неокортексом, осущест-
вляя по мере потребности кодирование, сохранение и извлечение информации.
Роль гиппокампальной формации может служить примером непрерывного
взаимодействия функциональных способностей мозга, имеющих узкие или ши-
рокие границы. «Узкие функциональные способности» характерны для созна-
тельных когнитивных процессов, избирательного внимания, кратковременной
памяти и произвольного управления, а «широкие функциональные способно-
сти» — для долговременной памяти, часто практикуемых навыков, словарного
запаса нашего языка. Далее эта тема обсуждается более подробно.
Рис. 2 .7. Мгновенная память необходима, чтобы локальные изображения, фиксируемые
в области центральной ямки глазного дна, интегрировались в единый осознаваемый об-
раз. Мгновенная память участвует во всех когнитивных функциях. Глаз непрерывно
совершает быстрые скачкообразные движения от одной точки к другой (саккады), оста-
навливаясь лишь в течение долей секунды на особенно важных участках поля зрения.
В каждой точке наиболее чувствительный участок сетчатки (центральная ямка) фикси-
рует маленький фрагмент информации, как это показывает классическая иллюстрация
Ярбуса (Yarbus, 1967; модификацию выполнил Марк Дубин; воспроизведено с его со-
гласия). Нижний рисунок отражает саккадические движения глаз и точки фиксации
взора при рассматривании портрета девочки. Считается, что мозг запоминает каждый
«снимок», сделанный центральной ямкой, планирует следующий быстрый скачок взора
и объединяет все мелкие «снимки» в общую связную картину. Таким образом, для ин-
теграции множества фрагментарных данных в единый сознательный образ требуется
буфер мгновенной зрительной памяти. (Источник: M. Dubin, с разрешения.)
Концептуальная структура когнитивной нейронауки
81
3.0 .
Ограниченность функциональных способностей
Несмотря на то что мозг человека содержит десятки миллиардов нервных кле-
ток, его функциональные способности в некоторых отношениях существенно
ограничены. Емкость кратковременной памяти (то, что мы способны мыслен-
но повторить) равна примерно «семи элементам плюс-минус два» (этот зна-
менитый вывод сделал Джордж Миллер в 1956 г.). Такое число применимо к
самым разнообразным элементам — цвету, количеству, коротким словам, му-
зыкальным нотам, оценочным баллам и т. д . Однако есть несколько условий.
Во-первых, сознательное восприятие каждого элемента должно происходить
однократно; если это совершается много раз, информация переходит в долго-
временную память. Во-вторых, характеристики элемента не должны прогно-
зироваться исходя из уже имеющегося знания. Если мы предложим человеку
запомнить принцип, например, формирования последовательности чисел 1, 5,
10, 15, 20, 25... (и т. д .), он сможет удержать в кратковременной памяти гораздо
больший объем информации, так как ему достаточно помнить только правило.
Если у нас нет возможности мысленно повторять элементы в кратковремен-
ной памяти, ее емкость падает с примерно семи элементов до менее четырех
(Cowan, 2001).
Еще одна ограниченная функциональная способность — избирательное
внимание. Первые исследования были проведены группой Дональда Бродбента
с помощью метода дихотического слухового входа (рис. 2 .8). В этом экспери-
менте испытуемый получал через наушники одновременно два канала речевой
информации, которая различалась для одного и другого уха. Когда испытуемого
просили «игнорировать» один из каналов и повторять поступающий текст без
остановок, он слышал только другой канал. То же самое происходит, если, на-
пример, слушать новости сразу по двум радиоприемникам. При этом человек
воспринимает текст только по одному каналу, хотя может улавливать звуки и по
другому каналу.
Если учитывать размеры мозга, такие функциональные границы кажутся
слишком узкими. Они удивительно малы, например, по сравнению с возмож-
ностями цифрового компьютера. Однако подобные ограниченные диапазоны
свойственны не только кратковременной памяти или избирательному внима-
нию. В табл. 2 .1 перечислены и другие функциональные явления с узкими гра-
ницами возможностей.
3.1.
Границы выполнения двойных задач
Современные персональные технические устройства в принципе позволяют че-
ловеку в одно и то же время обучаться, слушать любимую музыку и говорить
по сотовому телефону. К сожалению, при попытках выполнять одновременно
несколько дел эффективность снижается тем быстрее, чем больше число задач.
Как правило, мы не способны параллельно осуществлять даже две сознатель-
ные задачи, например активно беседовать и управлять транспортным средством
при интенсивном уличном движении. Если нет необходимости серьезно вни-
кать в тему разговора, человек может в это же время вести автомобиль. Однако
чем глубже наше сознание вовлечено в каждую из этих задач, тем значительнее
конкуренция между их выполнением. Поэтому метод двойных задач нередко
используется, чтобы оценить, в какой мере наши ограниченные способности
82
Глава 2
задействованы в выполнении задачи. При повышении уровня когнитивных за-
просов в связи с одной из задач происходит соответствующее снижение эф-
фективности выполнения второй задачи. Таким образом, метод двойных задач
очень важен в когнитивных исследованиях.
Как мы справляемся с проблемой ограниченности своих функциональных
способностей? Есть по крайней мере два пути. Первый — образование блоков
информации, сжатие большого количества информации в единый организован-
ный элемент. Такими блоками являются многие слова. Представьте себе, к ка-
кому объему информации открывают доступ такие слова, как «мать», «школа»,
«любовь» и т. д. Существует много способов сжатия информации в блок. Будем
надеяться, что при изучении материала этой главы вы сконцентрируете его по
главным пунктам. Это хорошая стратегия, поскольку вы можете обобщить ты-
сячи слов в виде нескольких десятков фраз.
Второй способ преодолеть ограниченность наших функциональных способ-
ностей — это практика, тренировка. Как известно, человек выполняет привыч-
ные задачи гораздо лучше, чем новые. Мозг решает новые проблемы довольно
медленно; он делает много ошибок, выполняет поэтапно действия, которые
можно было бы сделать в один прием. При выработке навыка выполнение за-
дачи выходит за пределы области сознательного (Raaijmakers and Shiffrin, 1992;
Рис. 2 .8. Дональд Бродбент, исследователь избирательного внимания. Интерес к из-
бирательному вниманию возобновился в 1950-е гг., когда Д.А . Бродбент начал свои
впечатляющие эксперименты. Вверху: предложенный Бродбентом образ «воронки»
демонстрирует функциональное ограничение емкости, подчеркивая тот удивительный
факт, что наш относительно большой по объему мозг имеет узкие пределы избиратель-
ного внимания, сознательного восприятия и произвольного контроля. Предложенная
Бродбентом задача избирательного слухового восприятия, когда одно и другое ухо одно-
временно получают разное сообщение, является классическим методом двойной зада-
чи для исследования функций с ограниченными емкостями. (Источник: фотоснимок
предоставлен Д. А . Бродбентом с разрешением для публикации.)
Концептуальная структура когнитивной нейронауки
83
Chein and Schneider, 2005). Например, человек с достаточным опытом видеоигр
может одновременно разговаривать, тогда как новичку необходимо сосредото-
читься на игре. Практический опыт позволяет снизить функциональные затра-
ты на выполнение задачи.
Однако существуют и функциональные способности с широкими граница-
ми. Таковы некоторые когнитивные возможности (табл. 2 .2).
Таблица 2.1
Функциональные способности с узкими границами
1. Ограниченность восприятия при двойном входе. Согласно результатам исследова-
ний, проводившихся примерно в течение 50 лет, человек не способен сознательно
воспринимать два разных одновременных потока информации.
а) Избирательное слуховое восприятие: одновременное поступление двух потоков
слуховой информации.
б) Слепота вследствие дефицита внимания: подача на экран сразу двух конкуриру-
ющих изображений.
в) Бинокулярная конкуренция и ее варианты: разный зрительный вход для одного
и другого глаза.
2. Ограниченность кратковременной памяти, в том числе способности удерживать
вспоминаемую информацию.
3. Неоднозначные стимулы и понятия, например кубик Некера, слова с двояким тол-
кованием и другие ситуации, когда вход можно интерпретировать по-разному.
4. Конкуренция между разными признаками одного объекта, например цветовой тест
Струпа (см. гл. 3).
5. Поиск сочетанных признаков: например, нужно найти в сложной экспозиции объ-
екты, одновременно обладающие заданными цветом и формой.
6. Конкуренция между трудновыполнимыми задачами. Чем труднее две задачи для
испытуемого, тем в большей степени выполнение каждой из них мешает выполне-
нию другой. Максимальное число трудных задач, которые человек может выпол-
нить в течение одного дня, является ограниченным (Muraven аnd Baumeister, 2000).
7. Конкуренция между реакциями, т. е . между двумя разными планами осуществле-
ния реакции или реальными действиями (Pashler, 1989).
8. Ограниченность временной интеграции.
а) Пробел внимания: при просмотре быстро сменяющихся букв возможен «сле-
пой» период в течение 200–500 мс после буквы-мишени.
б) Слепота в отношении смены изображения: если две визуальные картины раз-
делены краткой вспышкой белого света, их смена может быть незаметной. В ре-
зультате ограничивается восприятие последовательных событий.
9. Ограниченность поиска в долговременной памяти, например при подборе нужного
слова.
10. Концептуальная несовместимость и функциональная фиксированность. Человек
затрудняется или не может понять «очевидную» проблему с непривычной для него
точки зрения.
11. Домен-специфические ограничения. Специфические модальности входа и выхода
могут быть ограничены определенным участком тела. Примеры: (а) объекты вос-
принимаются как очень мелкие при высокой остроте зрения (если изображение
фокусируется на область центральной ямки); (б) трудно действовать сразу двумя
руками — допустим, похлопывать себя по голове и потирать область живота.
84
Глава 2
Таблица 2.2
Функциональные способности с широкими границами
1. Различные виды долговременной памяти.
а) Емкость эпизодической и автобиографической памяти составляет 109 бит
(Landauer, 1986). Человек может вспоминать события, происходившие 50 и бо-
лее лет назад: сцены из старых фильмов, случаи из детства, лица одноклассников
ит.д.
б) Большая емкость семантической памяти на факты и понятия.
в) Значительная емкость процедурной памяти (на часто практикуемые навыки).
2. Словарный запас языка: образованный носитель английского языка распознает при-
мерно 105 слов, с каждым из которых связана сложная сеть понятий, звуков, пись-
менных обозначений.
3. Высокая сложность организации сенсорных и двигательных процессов.
4. Колоссальное число нервных клеток, нервных связей и функциональных сетей мозга.
3.2 .
Некоторые функциональные способности мозга
имеют широкие границы
На клеточном уровне кора мозга представляет собой грандиозную структу-
ру — согласно недавним подсчетам, она содержит от 10 до 100 млрд нейронов.
Снаружи мозг выглядит как сложное складчатое образование с множеством вы-
ступов и углублений, занимающее верхнюю половину полости черепа. Если
бы удалось аккуратно развернуть обширную мантию, которую представляет
собой кора мозга, получился бы покров площадью около трех квадратных фу-
тов (1 м2), состоящий из шести слоев. В каждом слое находится бесчисленное
множество нейронов с ветвящимися отростками; нейроны окружены вспомога-
тельными клетками. Этот слоистый покров мозга подразделяется на миллионы
вертикальных колонок. Таким образом, мы имеем дело с колоссальной шестис-
лойной трехмерной системой. Каждый слой коры мозга выполняет специали-
зированную функцию — осуществляет вход, выход или внутренние коммуни-
кации в пределах коры (см. гл . 3 и 5).
Нейроны коры соединены аксонами — нервными волокнами, окруженными
белой миелиновой оболочкой. На вертикальном срезе через кору мы видим в
основном белое вещество, что позволяет судить об огромном количестве со-
единительных нервных волокон. Для невооруженного глаза кора представля-
ет собой сферу, покрытую снаружи тонким слоем серого вещества — телами
нервных клеток. Белое вещество образовано бесчисленными аксонами, кото-
рые идут от серых клеточных тел на большие расстояния и могут оканчиваться
в коре противоположного полушария. Два полушария соединены, наподобие
моста, массивным пучком волокон — мозолистым телом. По современным све-
дениям, оно состоит примерно из 200 млн волокон. Каждое из них передает
электрохимические сигналы с частотой примерно 10 в 1 с, так что получается
поток информации порядка 2 млрд сообщений в 1 с.
Большое число аксонов также входит в состав пучков, которые образуют
в подкорковых структурах петлю и возвращаются в то же полушарие, откуда
они вышли. Итак, между двумя полушариями и в пределах каждого из них идет
огромный поток сообщений.
Концептуальная структура когнитивной нейронауки
85
Горизонтальные волокна слоя I, верхнего из шести упомянутых слоев, густо
переплетаются, наподобие войлока, накрывающего остальные слои коры. В то
время как большинство дистантных коммуникаций коры осуществляется по-
средством длинных вертикальных выходных аксонов, волокна верхнего слоя об-
разуют многочисленные горизонтальные связи; высказывается мнение, что для
этого слоя характерно интенсивное распространение электрической активности.
Нейроны коры мозга связаны не только друг с другом. От них направляют-
ся длинные пучки волокон также к нервным центрам, расположенным под ко-
рой наподобие яиц в птичьем гнезде. Сюда относится таламус — важнейший
транспортный узел, стратегический пункт управления, где происходит пере-
ключение почти всех сенсорных сигналов, идущих в кору. Массивные пучки
волокон проецируются от коры к базальным ганглиям и мозжечку — структу-
рам, которые необходимы для нормальных двигательных функций.
Когда человек утром просыпается, мозг переходит в состояние бодрство-
вания. В его отделах возрастает скорость передачи сигналов и расширяются
области активации, т. е. сигналы генерируются и устремляются к местам на-
значений. Когда наше внимание привлечено чем-то новым или неожиданным,
в мозге спустя десятые доли секунды возникает волна электрического возбуж-
дения. Это так называемый «вызванный потенциал, связанный с событием»
(см. гл . 4 и Приложение Б).
Есть достаточно данных в пользу высокой степени функциональ-
ной специализации анатомических областей мозга (например, Luria, 1976;
Geschwind, 1979a). Многие ученые представляют организацию коры мозга в
виде совокупности дискретных функциональных единиц — «узловых моду-
лей» (Mountcastle, 1978; Edelman and Mountcastle, 1978). Розин (Rozin, 1976)
рассматривал эволюцию интеллекта как развитие способности реализовать эти
специализированные функции в конкретных жизненных ситуациях. Например,
новорожденный не умеет читать. Чтобы научиться этому, нужны годы, хотя у
младенца спонтанно вырабатывается способность понимать речь других лю-
дей и говорить. Вместе с тем мы удивительно легко осваиваем замену слухо-
вого восприятия языка на зрительное, а также передачу языка не фонетически,
а письменно. Это согласуется с результатами обучения языку жестов и чтению
по азбуке Брайля. По-видимому, способность читать зависит от биологической
преадаптации (имеются в виду «врожденные грамматики» Н. Хомского.
—
Прим. ред. перев.).
По мнению Розина и других авторов, функциональные способности моз-
га возникают в качестве специализированных приспособительных изменений.
В то же время предполагается, что в течение жизни человека специализирован-
ные функции адаптируются к новым задачам.
3.3 .
Почему функциональные способности
могут быть столь ограниченными?
Хорошо бы было выполнять сразу несколько дел. Почему же мозг с его десят-
ками миллиардов нервных клеток так ограничен в некоторых своих функциях?
Это нельзя объяснить только за счет исполнительных органов: один рот, чтобы
говорить, две руки, чтобы держать предметы. Ограничения проявляются уже на
уровне системы ввода, восприятия, так что проблема не в количестве устройств
на выходе. Ограниченность демонстрируется на примере неоднозначно толкуе-
86
Глава 2
мых изображений (кубик Некера): в конкретный момент человек способен вос-
принять объект только в одной интерпретации.
Итак, ограничения обусловлены не тем, что эффекторная система может
одномоментно осуществлять только одну функцию. И проблема не в том, что
мозг не обладает достаточными резервами для хранения и обработки информа-
ции — соответствующие возможности нервной системы таковы, что мы просто
не в состоянии их описать. Некоторые ученые полагают, что ограниченность
функциональных способностей заложена в том механизме, посредством кото-
рого сознание интегрирует деятельность многочисленных компонентов мозга
в единое целое (Baars, 1988, 2002b; Edelman, 1989; Llinas and Pare, 1991). Функ-
ции с узкими границами непосредственно ассоциируются с сознательными
действиями, тогда как функции с широкими границами обычно осуществля-
ются на уровне бессознательного (см. табл. 2 .1 и 2.2). Однако до сих пор не
выяснено, почему происходит именно так.
3.4 .
Оценка оперативной памяти
На схеме с. 68 отражен блок классической оперативной памяти (Baddeley and
Hitch, 1974; Burgess and Hitch, 1999; Baddeley, 2000). Концепция оперативной
памяти возникла в когнитивной психологии несколько десятилетий назад, когда
ученые начали понимать, как важна кратковременная память. Эта концепция
способствует развитию представлений о когнитивной деятельности мозга. Этой
тематике посвящено более десятка тысяч публикаций. Концепция оперативной
памяти помогает раскрывать природу других явлений, таких как мысленные
образы, язык, внутренняя речь, управляющие программы. Об оперативной па-
мяти накопилось много психологических данных, а в течение последних деся-
ти лет быстро расширяются сведения, составляющие ее нейробиологическую
основу. Таким образом, изучение оперативной памяти открывает путь к тому,
чтобы приблизиться к осмыслению сложнейших соотношений между психи-
кой и мозгом.
По словам Cowan et al. (2005), «оперативная память — это комплекс пси-
хических процессов, посредством которых ограниченный объем информации
удерживается в состоянии временной доступности для обслуживания когни-
тивной деятельности». Такое широкое определение оправданно. Оно отвечает
представлению о сохранении в памяти временного доступа к ранее восприня-
тым данным нашего разнообразного опыта. Смысл понятия оперативной па-
мяти существенно расширился с тех пор, как была предложена несколько иная
формулировка (Atkinson, Shiffrin, 1968; Baddeley and Hitch, 1974). Однако об-
щий смысл понятия оперативной памяти остается.
Заметим, что приведенное определение применимо к ситуациям не только
вербального повторения или зрительно-пространственного восприятия, но так-
же, например, мысленного воспроизведения песенного мотива. Если мы пере-
даем мелодию правильно и это подтверждается эмпирически, можем считать,
что в оперативную память добавлен компонент «внутренней музыки». В мозге
опытных музыкантов существует, очевидно, много приемов обработки нот и
мелодий. Мысленно воспроизводить свои действия могут спортсмены, и это
способствует улучшению их реальных результатов. Сейчас рассматриваются
различные типы оперативной памяти — для движений глаз, эмоций и, может
быть, даже для снов и фантазий. В самом общем смысле можно сказать, что
Концептуальная структура когнитивной нейронауки
87
оперативная память относится к разнообразным сенсорным модальностям.
В пользу более широкого толкования базовых идей получено много нейробио-
логических и поведенческих данных.
Считается, что при избирательном внимании сенсорная информация по-
ступает в оперативную память. Согласно приведенному выше определению
(Cowan et al., 2005), данные о запахе или вкусе «становятся временно доступ-
ными для когнитивной обработки». Если вы задержались на этом определении
примерно в течение десяти секунд, оно находится в оперативной памяти. Не-
трудно заметить, что вас отвлекают раздражители, в том числе ваши собствен-
ные мысли и ощущения, например прочитывание вот этой фразы. Таким об-
разом, ваша способность думать на любую тему имеет ограничения. Поэтому
можно говорить о «мыслительной оперативной памяти» («концептуальной опе-
ративной памяти»), а также о зрительной или вербальной оперативной памяти.
В науке очень важны количественные оценки. Оперативная память измеря-
ется с помощью разнообразных методов. На рис. 2 .9 показано, как можно оце-
нить зрительную оперативную память путем последовательного предъявления
на доли секунды слайдов с определенными фигурами. Спустя несколько секунд
любой из этих слайдов будет показан снова и экспериментатор попросит испы-
туемого вспомнить исходную фигуру или узнать ее. Показателем оперативной
памяти служит правильность вспоминания или узнавания, а также скорость от-
вета (время реакции).
Такие несложные, но эффективные методики существуют в разных вариан-
тах. Широко применяется методика отсроченного сопоставления с образцом
(delayed match to sample, DMTS), когда испытуемый должен вспомнить, если
предъявляемая ему фигура совпала с недавно показанным образцом. Методику
можно использовать в экспериментах не только с взрослыми людьми, но также
с младенцами или с животными.
Рис. 2 .9 . Один из способов оценки оперативной зрительной памяти. Испытуемому по-
следовательно предъявляют на нескольких сотен миллисекунд слайды с определенны-
ми фигурами. Спустя несколько секунд любой из этих слайдов может быть показан
снова и испытуемый должен вспомнить или узнать исходную фигуру
88
Глава 2
Как оценить состояние мозга во время процесса оперативной памяти? На
рис. 2.10 проиллюстрована задача «n ходов назад» — одна из часто применя-
емых методик оценки оперативной памяти. Задача позволяет варьировать ум-
ственную нагрузку, и эти изменения отражаются на состоянии мозга. Задача
требует усилий, хотя ее идея проста. В каждой серии стимулов испытуемый
должен запомнить фигуру на последнем слайде — это очень легко. Однако по-
сле этого экспериментатор просит его вспомнить слово на втором слайде с
конца, затем на третьем и т. д. Для этого испытуемому придется мысленно по-
вторить n последних стимулов и выделить фигуру «n ходов назад» среди других
стимулов, которые должны храниться в его оперативной памяти.
Задача становится все труднее, когда значение «n» увеличивается вместо
одного до трех или четырех. При этом с помощью фМРТ (см. гл . 4) регистри-
руется содержание кислорода в крови сосудов основания мозга. На рис. 2 .11
мы видим, что по мере усложнения задачи область активации мозга заметно
расширяется. В основании височной доли содержатся нейроны, специализи-
Рис. 2 .10 . Задачи «n ходов назад» для те-
стирования оперативной памяти. Испыту-
емый должен вспомнить фигуру, которая
была показана после первого, второго или
третьего слайда. Эта задача требует сосре-
доточенного внимания. С помощью фМРТ
было обнаружено повышение активности
в коре мозга по мере увеличения числа об-
ратных ходов (см. рис. 2 .11).
Рис. 2 .11. Усиление активности мозга по
мере возрастания нагрузки на оператив-
ную память. На томограммах при выполне-
нии задачи «n ходов назад» прослеживает-
ся значительное расширение вовлеченной
области коры мозга по мере увеличения n
от 0 до 3. В ситуации «0 ходов назад» ис-
пытуемый должен был просто назвать фи-
гуру на показанном ему слайде. На томо-
граммах более яркий цвет соответствует
повышению уровня активности. (Источ-
ник: Smith and Jonides, 1997.)
Концептуальная структура когнитивной нейронауки
89
рующиеся на восприятии зрительных объектов. Кроме того, эта область сосед-
ствует с медиальной височной долей, где находится гиппокамп. При высоком
уровне сложности предлагаемой когнитивной задачи (этот уровень может быть
сопоставим с таковым при одновременном жонглированием дюжиной мячей)
активность повышается также в префронтальных областях мозга.
4.0 .
Внутренние и внешние чувства
Получены убедительные сведения, что эндогенные (генерируемые внутри моз-
га) процессы являются в определенной мере имитацией «внешних чувств»
(рис. 2.12). При этом следует напомнить, что затылочная половина коры боль-
ших полушарий участвует главным образом в сенсорных функциях (рис. 2 .13).
Хотя наши глаза, уши, нос и рот расположены на передней части головы (об-
ращены в ту сторону, куда мы двигаемся), все сенсорные пути оканчиваются в
задней части коры мозга, где обеспечивается высший уровень анализа сенсор-
ных стимулов.
Каждый сенсорный путь имеет в коре свою собственную первичную про-
екционную область. Области первичных проекций в коре обозначаются по
следующей системе: V1 (первичная зрительная проекция), А1 (первичная слу-
ховая проекция), S1 (первичная соматосенсорная кора — карта тела). V1 — это
точная карта входа сигналов от сетчатки. Проекции V2, V3 и т. д. на более вы-
соких уровнях зрительной коры обеспечивают все более и более обобщенные
зрительные образы (см. гл . 6). При поступлении в теменную кору зрительные
сигналы интегрируются со слуховыми и тактильными сигналами, сигналами
положения тела в пространстве, с двигательным управлением.
Рис. 2 .12 . Функциональная схема; обратим внимание на сенсорные системы. Входные
сигналы поступают от рецепторов, преобразующих физическую энергию стимула в раз-
ряды нервных импульсов. Для областей сенсорной коры, обеспечивающих зрение и со-
матическую чувствительность, существуют карты проекций от участков тела, которые
получают соответствующие стимулы
90
Глава 2
4.1.
Глаз, ухо и голос психики
Аристотель в IV в. до н. э . предположил, что мысленные зрительные образы —
это «смутные копии» зрительных ощущений, т. е. результат неясного внутрен-
него восприятия. В течение нескольких последних десятилетий накапливаются
свидетельства в поддержку этого мнения. В начале ХХ в. американский психо-
лог Перки (C.W. Perky, 1910) показал, что люди не всегда могут отличить не-
четкие зрительные изображения от собственных мысленных образов.
Психологи разработали ряд методик тестирования зрительных образов. Сте-
фен Косслин (Stephen Kosslyn) показал, что такая фигура речи, как «внутрен-
ний глаз», на удивление точно соответствует реальности. Поле зрения человека
имеет определенные размеры и форму, которые легко продемонстрировать. Вы-
берите объект в комнате, где вы сейчас находитесь, и фиксируйте на нем взгляд
обоих глаз, не поворачивая головы; руки держите по обе стороны от головы
на расстоянии от нее. Постепенно смещайте ваши руки с периферии в поле
зрения до того момента, пока вы не увидите их; окажется, что горизонтальные
пределы активного зрительного поля соответствуют примерно 120° зрительной
дуги. Проделайте то же самое в вертикальном направлении. Вы увидите, что
здесь диапазон зрительного поля окажется меньше половины горизонтального.
Оперативное поле зрения имеет вид плоского овала, высота которого соответ-
ствует углу зрения примерно 45°, а ширина 120°.
Теперь закройте один глаз, а взгляд другого глаза фиксируйте на «точечной»
мишени, например на одной букве в этой фразе. Поле зрения окажется значи-
тельно меньше, всего несколько градусов зрительной дуги, поскольку сейчас
это фовеальное зрение. Фовеа (fovea), или центральная ямка глаза, — малень-
кое углубление в центре сетчатки, которое имеет наиболее значительную плот-
ность зрительных рецепторов и обеспечивает высокое зрительное разрешение.
Через эту «замочную скважину» мы смотрим на окружающий мир, когда нужно
получить особенно четкое изображение. Поле фовеального зрения соответству-
ет примерно четырем градусам зрительной дуги.
Рис. 2 .13. Активация затылочной коры мозга во время зрительной стимуляции. Скани-
рование позволяет картировать затылочную кору относительно участков сетчатки, на
которые попадает свет. (Источник: Singh et al., 2000.)
Концептуальная структура когнитивной нейронауки
91
Поле зрения своего «внутреннего глаза» вы можете измерить аналогично
тому, как вы оценивали свое реальное поле зрения; разумеется, вы будете делать
это только в своем воображении. Закрыв глаза, представьте себе, что сдвигаете
руки по обе стороны от своего «внутреннего глаза»; запишите горизонтальный
размер поля зрения. Затем повторите то же самое по вертикали. Обычно шири-
на поля зрения составляет менее 120° горизонтальной дуги, а высота пример-
но 45°. В подобных экспериментах обнаружено примечательное совпадение
между физически определяемым полем зрения и его психическим аналогом.
В последние годы начаты исследования с целью объяснить это. Как показали
Стефен Косслин, Марта Фара и другие авторы, мысленные зрительные образы
сопровождаются активацией участков зрительной коры. «Формирование зри-
тельных образов и восприятие внешних зрительных сигналов обеспечиваются
в принципе одними и теми же нервными механизмами».
Вместе с тем существуют различные способы видения мира и множество
форм пространственных образов. В зависимости от условий эксперимента раз-
личается характер (паттерн) активности в зрительной коре. Создавая условия
максимального сходства изображений при восприятии внешних зрительных
сигналов и при формировании зрительных образов, Гэнис с соавторами (Ganis
et al., 2004) наблюдали в той и другой ситуациях практически идентичную ак-
тивность мозга. Рис. 2 .14 показывает, что при вычитании (последовательно
этап за этапом) одного паттерна активности из другого паттерна между ними
практически не остается различий — поразительный результат.
Косслин (1994) отмечает, что мысленный образ «нельзя рассматривать как
единый процесс, он складывается из множества специализированных способ-
ностей». Например, получившая известность задача мысленного поворачива-
ния предмета (Shephard, Cooper, 1982) требует, видимо, участия зрительных,
пространственных, двигательных, а также управляющих областей (рис. 2 .15).
Учитывая точное совпадение карт области V1 и сетчатки, Косслин писал: «Воз-
можно, она (область V1) активируется только при таких задачах, когда форми-
рование образов требует высокой степени разрешения.»
Рис. 2 .14 . Во время восприятия зрительного образа могут активироваться участки зри-
тельной коры. На томограммах, полученных Ganis et al. (2004), области мозга, активи-
руемые при зрительном восприятии и при зрительном образе, практически идентичны;
это выявлено путем вычитания одной томограммы из другой (см. томограмму вторую
справа). Как показывает томограмма крайняя справа, виртуальные срезы проходят че-
рез затылочную и теменную области. Три первые слева томограммы соответствуют ко-
рональным срезам; внизу виден мозжечок. (Источник: Ganis et al., 2004.)
92
Глава 2
4.2 .
Психические образы формируются
в зрительных областях коры мозга
Вспомните ситуацию, когда вы первый раз увидели эту книгу. Какие зритель-
ные образы возникли в вашем мозге? Можете ли вы представить себе, где и как
была расположена книга? Лежала на горизонтальной поверхности или стояла
на полке? Видно ли было название на ее обложке? Живость спонтанных мыс-
ленных образов не одинакова у разных людей, но большинство из них способ-
ны представить себе сказанное выше. В каком участке мозга все это происхо-
дит? На рис. 2 .14 мы видим, что в восприятии внешних зрительных стимулов
и в формировании мысленных образов участвуют практически одни и те же
области мозга.
Обратим внимание, что в упомянутых примерах совпадают результаты пси-
хологических и нейробиологических исследований. Восприятие внешних зри-
тельных стимулов осуществляется в затылочных, височных и теменных обла-
стях коры мозга — там же, где формируются (при строгом соблюдении условий
эксперимента) зрительные образы, т. е. «внутреннее зрение».
4.3 .
Идентична ли внутренняя речь внешней речи?
Большинство людей часто разговаривают сами с собой, и многие охотно пере-
сказывают свои мысленные монологи. По просьбе ученых люди записывали
свою внутреннюю речь, как только она возникала; в результате был собран
очень полезный материал. Если мы отнесем внутреннюю речь к категории вну-
тренних чувств, то обнаружим сходство между внутренней и внешней артику-
ляцией слов.
Специалист по психологии речи Гэри Делл показал, что при мысленном
произнесении скороговорок человек страдает таким же косноязычием, как и
при их озвучивании (Dell and Sullivan, 2004). Попробуйте, например, повторить
про себя какую-нибудь скороговорку как можно быстрее. Вы заметили дефек-
ты мысленной речи? Но ведь вам не приходится двигать своим «мысленным
языком»! Мысленные образы могут служить очень эффективной тренировкой.
Рис. 2 .15. Задачи на зрительные образы. (а) Классическая задача с мысленным поворо-
том фигуры, по Shephard and Cooper (1982). Испытуемого спрашивают, одинаковы две
фигуры или различны. Чтобы ответить на этот вопрос, испытуемый мысленно повора-
чивает одну из фигур. (б) Классическая задача построения «башни» путем мысленного
перекатывания разноцветных шаров из одного гнезда в другое. Каким образом можно
превратить верхнюю фигуру в нижнюю? Испытуемый снова должен использовать зри-
тельный образ, однако задача отличается от мысленного поворота. (Источник: Heslow,
2002.)
Концептуальная структура когнитивной нейронауки
93
Это нетрудно объяснить, если признать, что во время воображаемых и физиче-
ских действий работают одни и те же участки мозга.
Вспомним, что мы узнали в гл. 1 о зонах Брока и Вернике (см. также
рис. 2.16). Теперь понятно, какие области мозга должны активироваться при
внутренней речи.
Мысленное повторение чисел тоже можно считать своего рода речью. Уче-
ные давно высказывают мнение о том, что мы молча разговариваем сами с со-
бой, и эта гипотеза имеет косвенные подтверждения. Теперь она получила под-
держку при функциональной визуализации мозга (рис. 2 .17).
В последующих главах мы увидим, что дело обстоит несколько сложнее,
потому что есть много способов говорить и слушать, смотреть и воображать.
Тем не менее принципиальная ситуация такова: «внешним чувствам» соответ-
ствуют «внутренние чувства» (например, формирование зрительных образов
и внутренняя речь — см. рис. 2 .18).
4.4 . Только ли один вид оперативной памяти?
Мы использовали один термин «оперативная память», что подразумевает еди-
ное явление. Однако по этому поводу продолжается горячая дискуссия. Как мы
увидим, получены данные не только о «домен-специфической» памяти, но и о
«неспецифической» кратковременной памяти (см. гл . 8). Некоторые исследова-
тели выделяют оперативную память для понятий, информации о пространстве,
смысловых характеристик информации (семантическую память), а также зри-
тельную память и речевую. Современные данные свидетельствуют о правомер-
ности объединения гипотез домен-специфической и неспецифической опера-
тивной памяти.
5.0 . Центральное управление
Как упоминалось выше, префронтальные области коры выполняют в мозге
важную управляющую функцию. Эти структуры необходимы для произволь-
ного контроля. Кроме того, они участвуют в эмоциях и сдерживают непроиз-
вольные импульсивные реакции (рис. 2 .19).
Рис. 2 .16. Локализация областей Брока и Вернике. Несмотря на то что в XIX в. врачи
не имели сложных технических средств регистрации активности мозга, их выводы со-
впадают с результатами современных исследований методами визуализации
94
Глава 2
Вот что пишет невролог Оливер Сакс (цит. по: Goldberg, 2001b):
«Лобные доли коры — самое позднее приобретение в эволюции нервной системы.
Такого высокого уровня развития они достигают только у человека (и в меньшей мере
у высших обезьян). .... Для них не свойственны простые, легко идентифицируемые
функции более примитивных областей коры мозга — сенсорных и двигательных.
. . . Однако их роль чрезвычайно важна. Они необходимы для высокоорганизованно-
го целенаправленного поведения — определения цели, составления стратегической
программы, планирования этапов достижения цели, обеспечения средств реализации
плана, монторинга достигаемых результатов и оценки их соответствия намерениям.
... Без развития в человеческом мозге лобных долей (и речевых областей) никогда бы
не возникла цивилизация.
Получила известность история железнодорожного мастера Финеаса Гейджа. В 1848 г.
во время взрывных работ в его лобные доли вонзился 60-сантиметровый железный
болт. Гейдж сохранил интеллект и способность ходить, говорить, видеть. Однако он
серьезно изменился в других отношениях — стал беспечным, расточительным, им-
пульсивным, несдержанным в выражениях, не задумывался о будущем, не строил
никаких планов. Для тех, кто знал его раньше, это был «другой Гейдж». Он лишился
себя, главной части своей личности, причем не осознавал этой потери (так же как
и другие пациенты с серьезными повреждениями лобных долей).
Рис. 2 .17. Внутреннюю речь можно
считать нормальной речью (а), но
при торможении фонации. В отсут-
ствие фонации (б, в) активируются
классические речевые зоны левого
полушария. B — центр Брока; W —
центр Вернике; M — двигательная
кора; A — слуховая кора. (Источ-
ник: Heslow, 2002.)
Концептуальная структура когнитивной нейронауки
95
Тестирование поражения лобных долей обычно проводится с помощью ме-
тода Струпа. Принцип теста заключается в том, что для испытуемого созда-
ется конфликт между смыслом слова и цветом обозначающих его букв. На-
пример, слово «синий» может быть напечатано зелеными буквами. При этом
испытуемый должен быстро назвать только цвет (рис. 2 .20). Грамотный человек
Рис. 2 .18 . Мысленное музицирование? «Внутренние чувства» не ограничиваются вер-
бальными и зрительно-пространственными образами. Мы можем ощущать себя «мыс-
ленным музыкантом» или «мысленным спортсменом». Результаты томографии мозга
показывают, что, когда музыкант-любитель реально играет на инструменте, у него про-
исходит более значительная активация двигательной и соматосенсорной коры, чем при
«внутреннем музицировании». Однако в обеих ситуациях задействованы одни и те же
области мозга. (Источник: Lotze et al., 2000.)
Рис. 2 .19 . Компьютерная реконструкция
повреждения мозга у Финеаса Гейджа, вы-
полненная на основании посмертной ма-
ски этого пациента (Damasio et al., 1994).
В то время, когда Гейдж получил травму
(1848 г.), большинство людей в подобных
ситуациях умирали вследствие нарушения
деятельности мозга и кровопотери либо в
результате инфекции. Гейдж выжил, и у
него восстановилось большинство функ-
ций, но он потерял способность к целена-
правленной деятельности и перестал кон-
тролировать свои импульсивные реакции.
Эти изменения были настолько выражен-
ными, что указывают на изменения лич-
ности. Аналогичные явления наблюдались
и в других случаях повреждения лобных
долей (см. гл . 12). (Источник: Caplan and
Gould; цит. по: Squire et al., 2003.)
96
Глава 2
автоматически пытается прочитать слово, прежде чем назвать его цвет. Сле-
довательно, требования теста противоречат многолетней привычке испытуемо-
го. В префронтальной коре возникает конфликт, но здоровый человек обычно
справляется с тестом, хотя по сравнению с контролем у него замедляется время
реакции и он делает ошибки. Пациенты с поражением лобных долей выпол-
няют задачу гораздо хуже: больше ошибок, продолжительнее время реакции,
сильнее субъективное ощущение невозможности справиться с задачей, фру-
страция. Тест Струпа служит для выявления относительно слабых изменений в
лобных долях, не замеченных при сканировании мозга.
5.1.
Исполнительная реакция и автоматизм
В экспериментах с визуализацией мозга сделано примечательное наблюдение:
при выполнении многих разнообразных задач, требующих умственного напря-
жения, всегда «включаются» две области переднего мозга. Во-первых, это дор-
солатеральная префронтальная область коры* в боковой части каждой лобной
доли. Важную роль в управлении выполняет также цингулярная кора — перед-
няя поясная область коры, расположенная в каждом полушарии вдоль средней
* Такие термины, как «дорсолатеральный префронтальный», на первый взгляд смущают, одна-
ко понять их нетрудно. Их можно сравнить с обозначениями на компасе: Север, Юг, Восток, За-
пад, Северо-Запад и т. д . Применительно к мозгу человека «дорсальная часть» — «верхняя», «ла-
теральная» — боковая, «медиальная» — расположенная по средней линии, «inferior» — нижний,
«superior» — верхний. Отсюда понятно, что означают сложные слова: «дорсолатеральная» кора —
это «верхняя боковая» часть коры, «вентромедиальная» кора — «нижняя срединная» часть. Если
это трудно представить себе, попытайтесь обозначить соответствующие направления на каком-либо
видимом объекте, например на изображении ботинка или автомобиля. Некоторые студенты предпо-
читают изображение пары ботинок по аналогии с левой и правой половинами коры мозга. У этих
объектов четко различаются передняя, задняя, левая и правая стороны. Если вы используете вместо
латинских терминов такие слова, как «передний, задний, верхний, нижний, срединный, боковой», вы
без труда справитесь с анатомическими названиями, разделяя их на составные части.
Рис. 2 .20 . Словесно-цветовой тест Стру-
па применяется для оценки функций цен-
трального управления. Попробуйте на-
зывать цвет напечатанных слов в верхней
половине рисунка; наверняка это окажется
трудным. В нижней половине рисунка, где
слова нечитаемы (буквы перевернуты), вы
будете называть цвет легче, быстрее и пра-
вильнее. (Источник: Miller and Wallis; цит.
по: Squire et al., 2003.)
Концептуальная структура когнитивной нейронауки
97
линии мозга. Обе эти области активируются при выполнении теста Струпа,
когда у испытуемого возникает конфликт между выработанным навыком (при-
вычкой прочитывать слова) и новым требованием (называть цвет букв) (Duncan
and Owen, 2000; Frackowiak, 2004).
Произвольные движения приобретают автоматизм в процессе практики (Shiffrin
and Schneider, 1977). По мере выработки навыков ослабевает их исполнительный
контроль (см., например, Langer and Imber, 1979). Отсутствие управления по от-
Langer and Imber, 1979). Отсутствие управления по от-
and Imber, 1979). Отсутствие управления по от-
and Imber, 1979). Отсутствие управления по от-
, 1979). Отсутствие управления по от-
ношению к привычным стандартным действиям означает, что наше сознание пре-
кратило доступ к этим процессам (Schneider, 1995). На томограммах мозга мы на-
блюдаем значительное ослабление активности коры, если ожидаемое произвольное
действие доведено до автоматизма благодаря частому его повторению. Есть сведе-
ния, что управление рутинными произвольными действиями частично переходит к
подкорковым структурам — базальным ганглиям и мозжечку.
Однако между произвольными и автоматическими действиями нельзя про-
вести границу по принципу «все или ничего». Автоматические действия мо-
гут вновь оказаться под произвольным контролем при потере предсказуемости
их выполнения, например, если мы пытаемся нормально ходить после пере-
лома ноги (Sacks, 1984). Большинство нашей повседневной деятельности на-
ходится под смешанным произвольным и автоматическим управлением. Наи-
более часто практикуемые ее компоненты приобретают автоматизм, тогда как
по отношению к новым и неожиданным действиям сохраняется произвольный
контроль. Например, оказавшись на незнакомом перекрестке, мы обдуманно
принимаем решение, куда пойти, однако затем, повернув направо, уже не от-
даем себе отчета в каждом своем шаге. Та же закономерность относится к речи,
чтению, движениям глаз и т. д. Мозг постоянно переходит от автоматического
управления к произвольному и наоборот.
Интересно, что в мозге существуют системы двойного управления. Напри-
мер, улыбка бывает как намеренной, так и спонтанной (рис. 2.21). Одни и те же
движения мышц могут инициироваться различными отделами мозга, но в про-
извольных мышечных сокращениях больше участвует кора, а именно лобные
доли.
Аналогичным образом, мы можем сделать глубокий вдох «по своей воле»,
но обычно дышим в спонтанном автоматическом ритме. Контроль движений
глаз тоже может быть как произвольным, так и автоматическим. Улыбка, дыха-
ние, движения глаз чаще контролируются автоматически, независимо от коры
мозга. Как мы увидим далее, двойственный контроль характерен и для избира-
тельного внимания: выбор объекта внимания может происходить произвольно
(управляемо) или спонтанно.
Такого рода двойное управление — обычная стратегия мозга, однако она не-
применима для жизненных функций, например ритмических сокращений серд-
ца. Естественно, что сердечный ритм сердца задается автоматически; останав-
ливать и запускать сердце по собственному желанию было бы катастрофой. То
же самое справедливо и по отношению к другим автономным (вегетативным)
функциям. Повреждения мозга иногда приводят к разобщению деятельности
центров произвольного и автоматического управления. Примером служит
поражение мозга, при котором искажена только произвольная улыбка, но не
спонтанная (рис. 2 .21). При других повреждениях могут произойти нарушения
98
Глава 2
противоположного характера: поражение ствола мозга может сопровождаться
потерей автоматического контроля при сохранении произвольного.
Продолжается дискуссия о том, соответствует ли произвольное управление
сознательным действиям (Shiffrin, 1995; Schneider, 1995). Это трудный вопрос.
Вместе с тем практически нет разногласий по поводу тесной взаимосвязи про-
извольного контроля и сознательной когнитивной деятельности.
Недавно опубликованные сведения позволяют полагать, что система управ-
ления соответствует собственному «Я» человека в его повседневной жизни
(Vogeley et al., 1999; Baars, 2002b). Мы рассмотрим эту проблему в последую-
щих главах применительно к исполнительным функциям и эмоциям.
5.2 .
Управляемое и спонтанное внимание
Вы можете специально обратить внимание на конкретное слово в этом пред-
ложении, но ваш взгляд может также время от времени на мгновение останав-
ливаться на том же слове независимо от вашего намерения. Как показал Майкл
Познер, центральная система, управляющая вниманием, включает префрон-
тальные области, а также теменные — в ситуациях пространственного выбора
(рис. 2 .22).
Пример спонтанного внимания — эффект зрительного «выхватывания»
(рис. 2.23). Некоторые зрительные стимулы достигают сознания независимо от
контролируемого произвольного поиска. Но если тот же стимул находится сре-
ди других аналогичных стимулов, эффект выхватывания не работает и нужен
произвольный поиск.
Рис. 2 .21 . Избирательное нарушение произвольного, но не спонтанного сокращения
мимических мышц. На левой фотографии у пациентки асимметричная улыбка, тогда
как на правой фотографии она улыбается совершенно нормально. Левый снимок был
сделан, когда пациентка специально старалась улыбнуться, а на правом снимке она улы-
бается непреднамеренно. Поражение двигательных областей лобной доли не влияет на
мимику, поскольку она обеспечивается подкорковыми структурами. У других пациен-
тов возможны нарушения противоположного характера: изменена спонтанная улыбка,
а не произвольная. Независимое поражение двух функций мозга называется двойствен-
ной диссоциацией. (Источник: Paxinos and Mai, 2004.)
Концептуальная структура когнитивной нейронауки
99
Часто полагают, что внимание обязательно участвует в ситуациях сознатель-
ного опыта. Такая гипотеза отражена на функциональной схеме (рис. 2 .24). Как
будет показано в гл. 8, это не всегда верно, хотя в повседневной жизни мы за-
мечаем, что направленное внимание чаще ведет к сознательному опыту. Роль
сознания в когнитивной деятельности стала темой интенсивных исследований,
так что сейчас накоплено достаточно данных, помогающих прояснить вопрос
(см. гл . 8). Тем не менее многое неизвестно и проблема сознания остается акту-
альным направлением научных исследований.
Рис. 2 .22 . Модель избирательного внимания, предложенная Познером. Блок избира-
тельного внимания входит в состав функциональной схемы и означает способность вы-
бирать между двумя и более альтернативными событиями. На схеме (первая справа),
предложенной группой Познера (Fan et al., 2005), представлена шкала управления вни-
манием. Отметим, что наиболее значительна активация двух долей коры — префрон-
тальной и теменной. При избирательном внимании может изменяться активность мно-
гих областей мозга. (Источник: Holstege et al., 2004.)
Рис. 2.23 . Спонтанный «захват» внимания. Испытуемый должен найти горизонталь-
ную зеленую черту-мишень в правой и в левой частях рисунка. (а) В левой части ри-
сунка мишень «выскакивает» спонтанно, без особого усилия. (б) Обнаружение мишени
в правой части рисунка требует специального усилия, в котором должны участвовать
управляющие области мозга. Зрительное «выскакивание» проявляется также по отно-
шению к биологически или индивидуально значимым стимулам, например лицам или
фигурам людей, к физически интенсивным стимулам. Таким образом, существуют два
различных типа избирательного внимания: произвольное, целенаправленное, управляе-
мое внимание и спонтанный, по принципу «снизу вверх», «захват» внимания значимы-
ми стимулами. (Источник: Squire et al., 2003.)
100
Глава 2
6.0.
Действие
Последний элемент функциональной схемы — это выход: управление произ-
вольными движениями. Примечательно, что между восприятием и действием
существует параллелизм (Fuster, 2003). Как вход, так и выход представляют
собой иерархическую систему процессов обработки информации. Иерархия
обработки зрительных сигналов начинается в первичной зрительной коре (об-
ласти V1), получающей от сетчатки «пиксельные» изображения; затем сигналы
поступают в специализированные области, отвечающие за восприятие цвета,
движения и за распознавание объекта (см. рис. 6.11). На выходе двигательная
иерархия начинается с общих целей, на реализацию которых влияет эмоцио-
нальный и мотивационный вход от лимбических структур. Цели самого обще-
го характера представлены в префронтальных областях, сигналы от которых
спускаются в соответствии с иерархией к ассоциативным и премоторным об-
ластям, где инициируются намерение и побуждение к действию (Penfield and
Roberts, 1959). Первичная моторная кора (область М1) непосредственно запу-
Первичная моторная кора (область М1) непосредственно запу-
скает движения скелетных мышц.
На рис. 2 .25 показано, как происходит активация мозга при нажатии на
кнопку пальцем правой руки. Внизу справа — шкала времени в секундах. Опе-
режающая активность мозга начинается за несколько секунд до нажатия паль-
цем. Отметим, что активация происходит в моторной коре левого полушария,
Рис. 2 .24 . Функциональная схема отражает гипотетические отношения между избира-
тельным вниманием и осознаваемыми событиями. На практике осознаваемые события
можно определить как такие, которые человек может точно описать. По мнению ряда
ученых, сознательный сенсорный опыт зависит от избирательного внимания. Однако
некоторые наблюдения противоречат такому выводу. Сейчас по этой проблеме опубли-
кованы новые экспериментальные данные
Концептуальная структура когнитивной нейронауки
101
т. е. на стороне, противоположной (контралатеральной) той руке, которая по-
лучает двигательную команду. Однако моторная кора посылает команду на ту
же (ипсилатеральную) сторону мозжечка, где находится рука, совершающая
движение; наконец, палец нажимает на кнопку — это нулевая точка на шкале
времени.
Память человека: исследования последних десятилетий
Когнитивная нейронаука — это новая увлекательная область знания, которая объ-
единила прежде раздельные направления биологии и психологии, чтобы понять,
как работают психика и мозг человека. Видная роль в осуществлении этой цели
принадлежит Ларри Скуайру, заслуженному профессору психиатрии, нейронаук
и психологии Медицинского факультета Калифорнийского университета в Сан-
Диего, посвятившему многие годы объяснению нервных процессов, которые со-
ставляют основу долговременной памяти.
В беседе с Л. Скуайром мы обсуждали, как изменились в течение последних деся-
тилетий представления о механизмах человеческой памяти. Вот о чем мы говорили.
Рис. 2 .25. Что делает мозг, чтобы палец нажал на кнопку? Hulsmann и соавторы (2003)
обнаружили, что произвольная задача формируется на несколько секунд раньше, чем
произойдет спонтанное действие. Внизу справа обозначено целевое время (0), когда
палец нажмет на кнопку. На двух томограммах в левой части рисунка видна актива-
ция двигательных областей левого полушария, управляющего мышцами правой руки.
Вместе с тем правое верхнее изображение свидетельствует, что активируется и правая
половина мозжечка, обеспечивающая тонкие движения — в данном случае движение
пальца. Такая активность контралатеральных областей соответствует анатомическим
данным об управлении движениями. (На приведенных здесь томограммах отсутствуют
другие области, которые участвуют в регуляции движений — базальные ганглии, мо-
торные проводящие пути.) (Источник: Hulsmann et al., 2003.)
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
102
Глава 2
Людей издавна занимает проблема памяти — как она формируется, почему воспо-
минания о некоторых давних событиях живо сохраняются на долгие годы, почему
они могут искажаться и исчезать. Современный этап в изучении памяти начался в
1957 г. с публикации Бренды Милнер (Scoville, Milner, 1957).
Это произошло, когда Бренда Милнер и известный нейрохирург Уайлдер Пенфильд
исследовали двух пациентов после одностороннего удаления структур медиаль-
ной височной доли с целью устранить эпилептические припадки, которые иначе
не удавалось купировать (Penfield, Milner, 1958). Неожиданным результатом опе-
Penfield, Milner, 1958). Неожиданным результатом опе-
, Milner, 1958). Неожиданным результатом опе-
Milner, 1958). Неожиданным результатом опе-
, 1958). Неожиданным результатом опе-
рации оказалась тяжелая амнезия: пациенты потеряли память. Сообщение об этих
клинических случаях было сделано на научной конференции в 1955 г., и вскоре
У. Пенфильд получил известие от Уильяма Сковилла из Хартфорда, штат Коннек-
тикут. Этот нейрохирург обнаружил аналогичный феномен амнезии у пациента по-
сле двустороннего удаления структур медиальной височной доли. Б . Милнер отпра-
вилась в Хартфорд, чтобы исследовать этого пациента, молодого человека 29 лет,
страдавшего эпилепсией с детства после падения с велосипеда. В 1953 г. припадки
были настолько сильными, что доктор Сковилл решился на хирургическое удаление
структур медиальной височной доли, в том числе гиппокампа. Судорожные припад-
ки прекратились, но молодой человек (в публикациях он известен под инициалами
ХМ) навсегда изменился: он потерял память. Этот клинический случай стал вехой
в истории развития представлений о человеческой памяти и роли гиппокампа в ней.
После смерти пациента в 2008 г. ученые смогли опубликовать его имя и подробно-
сти этого исторического случая (Squire, 2009).
Сообщения о нарушениях памяти у пациента ХМ после хирургической операции
послужили стимулом к многочисленным исследованиям роли медиальной височ-
ной доли и, в частности, гиппокампа; эта работа продолжается поныне. Случай
пациента ХМ позволил выявить три фундаментальных принципа. (1) Память —
особая функция мозга, отличная от когнитивной деятельности мозга и от речи, если
учесть, что ХМ мог говорить и понимал речь. (2) Медиальная височная доля не
является необходимой для кратковременной памяти, поскольку эта функция сохра-
нилась у ХМ. (3) Область медиальной височной доли — не окончательное место
хранения памяти, так как у пациента ХМ после удаления этих структур, на фоне
глубокой амнезии, которая охватила многие прожитые им годы, все же остались
воспоминания о детстве. Еще одно ключевое наблюдение состояло в том, что па-
мять — это не монолитная система. Хотя память существует независимо от общей
когнитивной функции, она складывается из подкатегорий, или подсистем. Об этом
Рис. 2 .26 . Ларри Скуайр (Larry R. Squire, PhD), за-
служенный профессор Калифорнийского универ-
ситета и Медицинского центра ветеранов в Сан-
Диего, США, штат Калифорния
Концептуальная структура когнитивной нейронауки
103
свидетельствует способность пациента ХМ обучаться выполнению новых задач и
действий. Тщательное изучение того, какие функции у ХМ оказались нарушенны-
ми, а какие сохранились, позволило Л. Скуайру и другим ученым сформулировать
теории раздельных компонентов памяти.
В течение последующих четырех лет появились обширные сведения о мозге и па-
мяти. Более ранние исследования были посвящены анализу поражений медиальной
височной доли мозга у пациентов, а также результатам экспериментов с удалением
соответствующих структур у лабораторных животных. В 1990-е гг. благодаря по-
явлению современных методов молекулярной генетики и функциональной визуа-
лизации мозга открылись новые перспективы выяснения генетических и нервных
механизмов памяти.
Л. Скуайру принадлежит видная роль в этих исследованиях. Ученый реализует ме-
тадисциплинарный подход, объединяя данные, которые получены в клинических
исследованиях и при экспериментальном удалении структур мозга у приматов и
других млекопитающих, с результататами генетических исследований, анализа по-
ведения, функциональной визуализации мозга. Недавно получены новые сведения,
проясняющие участие гиппокампа в памяти. Обобщая, можно полагать, что гиппо-
камп исходно вовлечен вместе с корой мозга в обеспечение долговременной памя-
ти, однако эта его функция ослабевает с возрастом. Таким образом, представления о
гиппокампе как окончательном месте хранения долговременной памяти постепен-
но меняются, согласуясь с более ранними наблюдениями, которые показали, что у
пациента ХМ, несмотря на глубокую амнезию, сохранились детские воспоминания.
Кроме того, новые данные поддерживают идею о роли сна и об участии гиппокампа
в консолидации памяти. А именно во время сна между гиппокампом и корой мозга
ведется диалог, в процессе которого информация переходит из кратковременной па-
мяти в долговременную и постоянную (Squire, в печати).
На каком этапе мы находимся сейчас? Л. Скуайр разработал общую теорию иерар-
хических систем долговременной памяти и структур мозга, обеспечивающих эти
системы. Теория очень важна для объяснения человеческой памяти (рис. 2.27; Kan-
Kan-
del, Squire, 2000; Squire, 2004). Что ожидает нас завтра? Никто не знает, но весьма
вероятно, что новые методы и средства исследования обеспечат быстрый прогресс
наших знаний о механизмах памяти.
Рис. 2 .27. Схема иерархической организации систем долговременной памяти моз-
га млекопитающих. (Источник: Squire, 2004.)
104
Глава 2
Литература
1. Scoville W.B ., Milner B. (1957). Loss of recent memory after bilateral hippocampal
lesions. J . Neurol. Neurosurg. Psychiatry, 20 (1), 11–21 .
2. Penfield W., Milner B. (1958). Memory deficit produced by bilateral lesions in the hip-
pocampal zone. AMA Arch. Neurol. Psychiatry, 79 (5), 475–497.
3. Squire L.R. (2009). The legacy of patient H.M. for neuroscience. Neuron, 61, 6–9 .
4. Squire L.R. (2009). Memory and brain systems: 1969–2009. In press. Journal of Neu-
roscience.
5. Kandel E.R., Squire, L.R. (2000). Neuroscience: breaking down scientific barriers to
the study of brain and mind. Science, 290 (5494), 1041–1248 .
6. Squire L.R . (2004). Memory systems of the brain: A brief history and current perspec-
tive. Neurobiology of Learning and Memory, 82, 171–177 .
7.0 .
Консолидация кратковременных событий
в долговременную память
Клайв Уэринг был способен воспринимать текущее событие, но не мог сохра-
нять этот опыт в долговременной автобиографической памяти. Виды долго-
временного хранения информации (от перцептуальной памяти до привычек)
отражены внизу функциональной схемы (рис. 2 .28).
Проблема локализации долговременной памяти в мозге остается неразре-
шенной. Вместе с тем накопились косвенные данные в пользу гипотезы кон-
солидации, согласно которой хранение информации обеспечивается теми же
областями мозга, которые управляют соответствующими функциями в конкрет-
ный момент. Оперативная память изменчива, поскольку она зависит от дина-
Рис. 2 .28 . Функциональная схема; в нижней части рисунка отражены виды долговре-
менной памяти (от перцептуальной памяти до сформированных навыков). До тех пор
пока не активированы структуры, сохраняющие эти виды памяти, их содержание не
осознается
Концептуальная структура когнитивной нейронауки
105
мических электрохимических процессов в крупных популяциях нейронов и в
синапсах. Однако считается, что при повторении события повышается эффек-
тивность и увеличивается продолжительность существования синаптических
соединений, участвующих в запоминании. На рис. 2.29 отражена гипотеза о том,
что задействованные синапсы становятся стационарными соединениями, в них
синтезируются белки и РНК. Гипотеза консолидации гласит: «Между нейрона-
ми, которые одновременно активны, формируется проводная связь» (см. гл . 3).
По мнению ряда ученых, разрабатывающих теории памяти, следы долго-
временной памяти сохраняются в тех же участках мозга, которые непосред-
ственно участвуют в соответствующей функции. Рис. 2 .30 иллюстрирует эту
точку зрения применительно к коре больших полушарий (Fuster, 2003). Задняя
половина коры (голубой цвет) задействована главным образом в сенсорных
функциях и, следовательно, является хранилищем сенсорной и перцептуальной
памяти. Фронтальная часть коры (см. выше), которая играет исполнительную
роль, управляет движениями, включает центр Брока и зоны других когнитив-
ных функций, должна, по-видимому, обеспечивать консолидацию памяти при-
менительно к этим функциям (см. гл . 5).
Рис. 2 .29 . Консолидация памяти. Происходит интерференция эпизодов кратковре-
менной памяти, по-видимому, в результате участия активированных нейронных ан-
самблей. Если информация сохраняется от нескольких секунд до часов, она может
стать перманентной, или «консолидированной». Стало известно, что консолида-
ции памяти способствует крепкий ночной сон. Считается, что для долговременной
памяти требуется синтез белков, благодаря которым повышается эффективность
синаптических связей. (Источник: Nader, 2003.)
106
Глава 2
Гипотеза консолидации плодотворна, поскольку она обобщает результаты
многочисленных наблюдений. Долговременная память будет подробно рассмо-
трена в гл. 9.
7.1.
Оперативная память — результат активации
постоянной памяти?
В связи с гипотезой консолидации возникает интересный вопрос: может быть,
мгновенная память просто запускает следы, длительно хранящиеся в мозге? К
этой проблеме можно подойти в свете представлений как о кратковременной,
так и о долговременной памяти. Эта ситуация может служить для студентов
примером правомерности рассмотрения каждого аспекта современной нейро-
науки с разных позиций. Любую точку зрения нельзя отбрасывать, пока не по-
явятся неоспоримые сведения.
В табл. 2.3 перечислены области мозга, которые, возможно, участвуют в ког-
нитивной деятельности. Соответствующие гипотезы будут обсуждаться в даль-
нейших главах.
Рис. 2 .30 . Если придерживаться гипотезы консолидации, то следует признать, что за-
поминаемая информация может храниться во многих областях мозга, между которыми
образовались постоянные синаптические связи. При этом предполагается (Fuster, 2003),
что задняя половина коры мозга обеспечивает сенсорную память, а передняя полови-
на — исполнительную и двигательную память. Кроме того, в эпизодической памяти
(запоминании сознательных событий) участвует гиппокампальная формация, тогда как
подкорковые структуры (базальные ганглии и мозжечок) играют роль в двигательном
научении. Эта гипотеза позволяет понять, каким образом мозг обеспечивает долговре-
менную память. (Источник: Fuster, 2003.)
Концептуальная структура когнитивной нейронауки
107
Таблица 2.3
Гипотетическая роль некоторых структур мозга
в рамках функциональной схемы
Сенсорный вход и хранение сенсор-
ной информации
Сенсорная кора (задняя половина коры), сен-
сорные пути и релейные ядра таламуса
Произвольное избирательное вни-
мание
Префронтальная и теменная области могут
осуществлять в сенсорной коре модуляцию,
направленную на отбор определенных сен-
сорных входов
Спонтанное избирательное внима-
ние
Сенсорные области могут запускать избира-
тельную обработку сигналов
Вербальная оперативная память
Расширенные зоны Брока и Вернике — пре-
фронтальная кора и медиальная височная доля
Системы ввода и обработки зритель-
но-пространственной информации
Зрительная кора, в том числе теменная доля и
префронтальные области
Системы выхода
Префронтальная и моторная кора, базальные
ганглии, мозжечок, моторные тракты
Кратковременное хранение опера-
тивной памяти
Медиальная височная доля, взаимодействую-
щая с новой корой
Системы долговременной памяти
Долго сохраняющиеся изменения межнейрон-
ных связей в коре
8.0 .
Заключение
В этой главе представлена обобщенная функциональная схема когнитивной
нейронауки на основе признанных идей. Показаны ассоциации между некото-
рыми структурами мозга и конкретными функциями.
В гл. 1 рассматривались исследования Брока и Вернике, благодаря которым
были выявлены центры речи в соответствующем полушарии мозга; эти центры
участвуют и во внутренней речи (вербальном повторении). Аналогичным об-
разом в формирование зрительных образов вовлечена зрительная кора, а при
«внутреннем звучании музыки» — области мозга, которые задействованы при
реальном прослушивании музыки и игре на инструментах. Следовательно, для
построения упрощенной схемы мы можем проводить параллели между явными
и скрытыми когнитивными функциями. Такая гипотеза должна помочь в пони-
мании материала последующих глав.
Мгновенная память зависит, по-видимому, от медиальной височной доли, в
состав которой входят гиппокамп и окружающие его структуры. При повреж-
дении этих областей нарушается способность к переходу текущей информации
на длительное хранение. Согласно гипотезе консолидации, долговременная па-
мять обеспечивается благодаря прочному усилению активных межнейронных
связей, которые образуются в результате текущей информации, удерживаемой
в оперативной памяти.
Задняя половина коры мозга участвует в сенсорных процессах и, вероятно, в
сенсорной (перцептивной) памяти. Фронтальная кора обеспечивает моторную
и управляющую функции, а также, видимо, долговременную память, необходи-
108
Глава 2
мую для этих процессов. Однако мгновенную память можно рассматривать и с
иной точки зрения — как извлечение информации из долговременной памяти в
результате входа сигналов.
Изучение избирательного внимания продолжается примерно 50 лет, и в по-
следние 15 лет на передний план снова вышел традиционный вопрос о созна-
тельном опыте. Как мы увидим, в этих исследованиях применяется множество
разнообразных экспериментальных методов. Сознательная когнитивная дея-
тельность отражена наряду с другими характеристиками на функциональной
схеме (см. рис. 2 .24).
Эти идеи важно учитывать в размышлениях на тему когнитивной нейронау-
ки. Более подробно они будут обсуждаться в последующих главах.
Продолжается обсуждение терминов «оперативная память», «внимание»
и «сознательный опыт». Подобные споры в науке обычны. Сейчас достигнут
приемлемый рабочий консенсус относительно эмпирических определений —
о том, какого рода явлениям соответствует конкретное понятие. На практике
понятие «оперативная память» относится к любому феномену кратковремен-
ной памяти продолжительностью порядка десятков секунд — период, в течение
которого мы успеваем удержать информацию и обработать ее. Такому опреде-
лению соответствуют удержание в памяти номера телефона в течение перио-
да его набора, мысленные арифметические действия или выбор оптимального
пути от одного места до другого. В первых экспериментах по оценке опера-
тивной памяти испытуемые получали список не связанных по смыслу слов
или чисел и должны были вспомнить их через 10–30 мин. Другая популярная
методика, «отсроченное сопоставление с образцом» (delayed match to sample),
заключается в том, что испытуемому показывают образец, а затем просят от-
ветить «да» или «нет», когда он снова увидит некую фигуру.
Метод отсроченного сопоставления с образцом полезен в экспериментах на
животных, поскольку крыса или обезьяна не могут рассказать нам, что они ду-
мают. Утвердительным ответом здесь можно считать совпадение реакции на
стимул с той, которая была получена ранее. Эксперименты на животных явля-
ются важным источником информации. Например, именно таким образом была
подтверждена роль гиппокампа в обучении. На практике показателем «опера-
тивной памяти» считается способность удерживать в памяти информацию о
сигнале в течение нескольких секунд. Такое объяснение применимо к опера-
тивной памяти для запахов и даже для преднамеренных движений глаз.
О когнитивной деятельности и об участии мозга человека получены об-
ширные новые знания. Сложная задача состоит в том, как экспериментально
разделить психические и нервные аспекты мышления и действий. Несмотря
на достигнутый прогресс в понимании механизмов взаимодействия этих сфер,
нам еще далеко до ясных представлений о динамических интегративных и ин-
терактивных процессах, которые составляют существо человека.
9.0.
Задания и упражнения к главе 2
9.1.
Контрольные задания
1. Что мы узнали о памяти при рассмотрении случаев Клайва Уэринга и паци-
ента ХМ?
Концептуальная структура когнитивной нейронауки
109
2. Какие области мозга участвуют в оперативной и в долговременной памяти?
В зрительных образах и в воспроизведении пространственного плана мест-
ности?
3. Что такое внутренняя речь и как она связана с обычным познавательным
процессом?
4. Какие ориентиры на изображениях структур мозга можно использовать для
объяснения, что вы видите? Где находится затылочная часть? Как выглядит
левая сторона? Правая сторона?
5. Сформулируйте понятия оперативной памяти, избирательного внимания,
разных видов долговременной памяти.
9.2 .
Графические упражнения
1. Проставьте обозначения на схеме, рис. 2 .31.
2. Определите участки головного мозга по рис. 2 .32.
Рис. 2 .31
Рис. 2 .32
Возможно, наиболее важной задачей науки является попытка понять, ка-
кие эволюционные события привели к развитию у мозга способностей к осоз-
нанию разных частей картины мира: цвета, звука, боли, удовольствия и много-
гранного мысленного опыта.
Роджер Сперри (1976)
Упрощенная схема организации нейронов. Полезно представлять себе нейроны коры
(как и многих других структур) в виде иерархической системы. В коре сигнал способен
идти в любом направлении. На рисунке показан пример циклической иерархической
структуры, в котором возможна коммуникация между зрительной, слуховой и другими
областями коры. (Источник: Friston, 2003, с изменениями.)
Содержание
1.0.Введение........................................................111
1.1.Реальныеиидеализированныенейроны..........................113
1.2.Возбуждениеиторможение....................................115
1.3.Обработкаинформациинейронами..............................117
2.0.Рабочиегипотезы.................................................118
2.1.Упрощенныйслучай:рецепторы,путииконтуры..................119
3.0.Массивыикарты.................................................124
3.1.Картыпереходятвдругиекарты.................................127
3.2.Массивынейроновчащевсегоимеютсвязидвухтипов............. 127
3.3. Сенсорные и моторные системы работают совместно. . . . . . . . . . . . . . . 127
3.4. Временная кодировка: рисунки спайков и ритмы мозга . . . . . . . . . . . . . 129
3.5.Точкивыборавпотокахинформации............................133
3.6.Обработка,обусловленнаяожиданием...........................135
4.0.Адаптацияиобучениемассивовнейронов............................138
4.1 . Обучение по Хэббу: «Активирующиеся одновременно нейроны
образуютсоединениямеждусобой».............................139
4.2 . Дарвинистский подход в нервной системе: выживают клетки
и синапсы, наиболее приспособленные к данной задаче . . . . . . . . . . . . . 143
4.3. Обработка символьной информации и нейронные сети . . . . . . . . . . . . . 145
5.0.Координацияработынейрональныхсетей............................146
5.1.Функциональнаяизбыточность.................................148
6.0.Заключение.....................................................149
7.0.Заданияиупражнениякглаве3....................................150
7.1.Контрольныезадания..........................................150
7.2.Графическиеупражнения......................................150
1.0 . Введение
В гл . 2 был дан краткий обзор функций мозга, таких как восприятие сигналов,
рабочая память, внимание и исполнительный контроль. Их изучение значитель-
но упростилось после внедрения томографических методов. Однако что мы
знаем о процессах, происходящих на уровне нейронов? Можем ли мы сейчас
построить непротиворечивую теорию относительно событий на этом уровне?
Мы начнем с рассмотрения некоторых основных сведений относительно
нейронов и связей между ними. В гл . 1 мы подчеркивали, что наука не только
собирает данные, но и строит на их основе теорию, объясняющую наблюдения.
Глава 3
Нейроны и связи между ними
112
Глава 3
Люди изучают нейроны вот уже два века, и те упрощения, которые мы будем
здесь использовать, основаны на огромном количестве информации. Однако,
как и любое другое упрощение, оно может в какой-то момент перестать
нас удовлетворять. В этом случае мы будем вынуждены внести в модель
соответствующие изменения.
Основными клетками мозга являются нейроны, высококонсервативные с
эволюционной точки зрения. Они сохранялись в относительно неизменном виде в
течение многих сотен миллионов лет, и даже очень разные виды животных имеют
одинаковые типы нейронов. Со многих точек зрения нейроны не отличаются от
остальных клеток, но есть то, что выделяет их среди остальных: специализация на
электрохимической сигнализации, благодаря которой они способны принимать
входящий сигнал на дендритах и посылать электрохимический сигнал вдоль
аксона. Весь мозг можно рассматривать как сверхсложную сруктуру, состоящую
из связанных между собой нейронов (рис. 3.1).
Рис. 3 .1 . Нейроны, сети и мозг. Три уровня организации — от нейрона до больших
объединений сетей. Сенсорная и моторная кора часто рассматривается в виде иерархи-
ческой структуры (Friston, 2003). В этом примере показано, как в зрительной коре за-
тылочной доли происходит распознавание все более и более сложных объектов, а также
влияние на височные и теменную доли. Схема в верхней части рисунка представляет
архитектурную организацию всей коры. (Источник: Friston, 2003.)
Нейроны и связи между ними
113
Дендритами и аксонами называют выросты тела нейрона; один нейрон
может иметь до десяти тысяч дендритов и один аксон (рис. 3.2 и 3.3). Потенциал
действия (ПД) проходит по аксону значительно медленнее, чем электрический
ток в компьютере, однако многие задачи наш мозг выполняет гораздо лучше
современных компьютеров. В настоящее время компьютеры далеко отстоят
от человека в задачах восприятия, языковой коммуникации, семантической
памяти, контроля движения и творчества.
1.1 .
Реальные и идеализированные нейроны
Мозг можно сравнить с тропическими лесами Амазонки, в которых по настоя-
щее время остается неописанным огромное число видов животных и растений.
Рис. 3.2 . Отдельный нейрон. Микрофото-
графия отдельного биполярного нейрона.
Кортикальные нейроны могут иметь до
десяти тысяч дендритов и как минимум
один аксон. Все эти образования имеют на
концах синапсы, необходимые для связи с
другими нейронами. Фотография тем не
менее не дает представления о трехмер-
ной разветвленной структуре нейрона.
(Источник: Standring, 2005.)
Рис. 3 .3 . Идеализированный нейрон.
Упрощенный нейрон с дендритами в
верхней части рисунка; корешки дендри-
тов показаны в виде узлов. Дендриты по-
лучают входящие сигналы и генерируют
деполяризующие мембранные потенци-
алы (рецепция). Когда деполяризующий
потенциал на дендрите превышает по-
роговый на короткое время (интеграция),
происходит деполяризация мембраны ак-
сона — волна ПД по принципу «все или
ничего» (кодирование). Волны потен-
циала действия на аксоне вызывают вы-
свобождение нейромедиатора на терми-
налях аксона, и процесс повторяется на
следующем нейроне. (Источник: Byrne и
Roberts, 2004.)
114
Глава 3
Для начала мы познакомимся с генерализованным нейроном, который тем не
менее далеко не всегда удовлетворительно отражает реальное положение дел.
На самом деле существует огромное количество разнообразных нейронов, ней-
роактивных веществ и потенциальных механизмов обработки информации.
Первым нашим упрощением будет рассмотрение только интегративного ней-
рона (см. рис. 3.3). Этот классический нейрон принимает сигналы от других
нервных клеток при помощи возникающих на дендритах деполяризующих
потенциалов. Если суммарный потенциал дендрита превышает –50 мВ, то по
аксону нейрона проходит быстрораспространяющаяся волна потенциала дей-
ствия (см. рис. 3 .3 и 3.5).
Классический нейрон передает сигнал путем проведения по аксону волны
потенциала действия от тела клетки к аксонным терминалям. На терминалях
происходит выделение нейромедиатора в синаптическую щель. Нейромедиатор
вызывает деполяризацию постсинаптической мембраны. Этот процесс схема-
тически представлен на рис. 3 .4 и 3.5 . Нейробиология концентрирует внимание
на связи и взаимодействии нейронов. Рассмотрение таких связей удобно начи-
нать как раз с генерализованного нейрона.
Несмотря на всю потрясающую сложность мозга, идеи, помогающие нам в
его исследовании, относительно просты. Но простые составные части способ-
ны образовывать крупные системы. Существует огромное количество способов
объединения нейронов в каскады, контуры и сети.
Рис. 3 .4 . Два нейрона соединены при помощи синапса. Помните, что некоторые синап-
сы могут работать в обе стороны. (Источник: Byrne и Roberts, 2004.)
Нейроны и связи между ними
115
1.2.
Возбуждение и торможение
Классические нейроны соединяются при помощи синапсов (рис. 3 .6), которые
могут быть возбуждающими и тормозящими. С их помощью вероятность про-
хождения ПД по аксону постсинаптического нейрона может быть как увели-
чена, так и уменьшена. Нейрон коры головного мозга может иметь несколько
десятков тысяч входящих и около десяти исходящих синапсов. Активность ней-
рона опосредована десятками факторов — циклом сна и бодрствования, до-
ступностью предшественников нейромедиаторов и многими другими. Все эти
факторы влияют на вероятность прохождения сигнала между двумя нейронами
и могут быть представлены в виде синаптических весов. Таким образом, все
разнообразие нейронов можно с успехом представить в виде интегративного
нейрона, а все способы межнейронной коммуникации — в форме вероятности
прохождения сигнала между нейронами.
Нейроны крайне разнообразны по форме, характеру ветвления и типам
синапсов. Существует по меньшей мере шесть основных нейромедиаторов и
не менее тридцати «менее важных», в основном являющихся нейропептидами.
Также в нервной системе имеются электрические синапсы, в которых передача
сигнала происходит без помощи медиатора; они, как оказалось, распростране-
ны гораздо шире, чем полагали ранее. Даже дендриты отдельной клетки, по
всей видимости, способны к обработке информации. Имеются также данные,
что способна принимать участие в обработке информации и нейроглия — под-
держивающая ткань нервной системы. И это далеко не полные данные относи-
Рис. 3 .5 . Прохождение сигнала по аксону. (а) Прохождение сигнала по аксону обеспе-
чивается за счет прохождения ионов через соседние участки мембраны. На рисунке об-
ласть 2 деполяризуется, в то время как область 3 уже сгенерировала потенциал действия
и в настоящее время гиперполяризована. Потенциал действия пойдет дальше за счет
деполяризации области 1. (б) Миелинизированные аксоны обернуты специализирован-
ными шванновскими клетками. Мембрана аксона открыта только в области перехватов
Ранвье. (в) Потенциал действия миелинизированного аксона регенерируется в области
перехватов. Миелинизированные аксоны обладают повышенной по сравнению с немие-
линизированными скоростью проведения. (Источник: Ramachandran, 2002.)
116
Глава 3
Рис. 3 .6 . Типичный синапс. Клетки, образующие синапс, называют пресинаптической
(голубая) и постсинаптической (синяя). Прохождение ПД по аксону пресинаптической
клетки вызывает высвобождение нейромедиатора, диффундирующего в синаптиче-
скую щель и понижающего мембранный потенциал постсинаптической клетки. Неко-
торые элементы этой биологической системы показаны на этом рисунке. (Источник:
Standring, 2005.)
Рис. 3 .7. Микрофотография единичного нейрона базального ганглия. Обратите внима-
ние на выросты дендритов, часто несущие синапсы. Длина линейки 50 мкм. (Источник:
Paxinos, 2004.)
Нейроны и связи между ними
117
тельно функционирования нервной системы. К примеру, более ста лет счита-
лось, что после рождения новых нейронов не образуется. На настоящий момент
известно о существовании в некоторых частях взрослого головного мозга ство-
ловых клеток. Образование новых синапсов идет в течение всей жизни; для
образования новых синапсов отростки дендритов способны образовываться за
несколько минут (рис. 3 .7). Таким образом, очевидно, что генерализованный
нейрон отражает лишь часть общей картины. И новые доказательства этому
утверждению будут в ближайшие годы только прибывать.
В 2004 г. Шеперд (Shepherd) писал:
Представление о нервной клетке как о независимой единице ... продолжает быть
основой наших представлений о нервной системе. Тем не менее на данный момент
очевидно, что нейрон представляет собой только один из уровней организации ней-
рональных контуров и систем. В настоящий момент мы осознаем, что функциони-
рование нервной системы имеет множество уровней организации, некоторые из
которых лежат внутри нейрона, а некоторые обусловлены межнейронными взаимо-
действиями.
1.3.
Обработка информации нейронами
Так что же позволяет нам ограничиваться изучением упрощенных нейронов?
Возможно, самым весомым аргументом в их пользу является успех моделей
нейронных сетей, на протяжении нескольких последних десятилетий использу-
ющих именно упрощенные нейроны. Искусственные нейронные сети использо-
вались для моделирования многих функций мозга — распознавания элементов
изображений, управления роботами, обучения и улучшения функционирования
на основе опыта. Большинство моделей, основанных на искусственных ней-
ронных сетях, относительно небольшого масштаба и ограничены по функциям.
Ни одна из них даже не приближается по сложности к мозгу. Однако во многих
случаях такие сети выполняли задачи лучше, чем компьютерные программы,
основанные на логике и математике. Важно помнить о том, что искусствен-
ные нейронные сети, как указано в их названии, — творение человеческих рук.
И тем не менее они помогают нам понять принципы работы реальных нейрон-
ных сетей в мозге.
В истории науки немало примеров того, как новые математические методы
помогали решить дотоле неразрешимые проблемы. Так же и нейросети помога-
ют нам понять работу нервной системы. Математическая основа нейросетей с
момента их создания значительно улучшилась. По всей видимости, существует
довольно ограниченный набор базовых архитектур, аналогичных сетям мозга.
Нейробиологов в первую очередь интересуют те нейросети, которые осно-
ваны на известных свойствах групп нейронов и связей между ними. Так искус-
ственные нейросети могут служить моделями для изучения реальных структур
в мозге.
Помимо упрощенного нейрона, изображенного на рис. 3.3, мы также будем
использовать упрощенный синапс. Как уже указывалось выше, мы ограничимся
рассмотрением синапсов только двух типов — возбуждающего (повышающего
вероятность прохождения ПД на постсинаптическом нейроне) и тормозящего
(понижающего такую вероятность). Глутамат — наиболее распространенный
медиатор в ЦНС — является возбуждающим. ГАМК (гаммааминомасляная
118
Глава 3
кислота) является наиболее распространенным тормозным медиатором. Таким
образом, наша упрощенная модель все же отражает определенную часть реаль-
ной картины.
2.0 .
Рабочие гипотезы
Мы обсуждали использование упрощенных и идеализированных нейронов для
описания основ возбуждения и связи нейронов. Далее мы будем использовать
эти предположения в качестве рабочих гипотез при рассмотрении нейронов и
их связей.
1. Интегративный нейрон. Мы примем, что нейроны работают путем после-
довательного добавления деполяризующих потенциалов к мембранному
до превышения некоторого порогового значения (в настоящих нейронах —
приблизительно –50 мВ). Когда это происходит, по аксону проходит волна
потенциала действия по принципу «все или ничего».
2. Синапсы могут быть как возбуждающими, так и тормозными. Таким
образом, мы можем выразить вклад синапса в вероятность возбуждения
нейрона В нейроном А в виде одного числа. Это число называется весом
соединения, или синаптическим весом. Значения синаптического веса
варьируют в пределах от –1 до + 1. Синаптический вес + 1 означает 100%-ю
вероятность того, что постсинаптический нейрон сгенерирует аксональный
ПД; синаптический вес –1 означает 100%-ю вероятность предотвращения
генерации аксонального ПД. Могут встречаться и более сложные случаи,
однако это хорошее первое приближение. Широкий набор нейромедиаторов
можно достаточно успешно заменить значением вероятности прохождения
сигнала — весом соединения.
3. Второй причиной использования упрощенных нейронов и синапсов при мо-
делировании является то, что сравнительно простой математический аппарат
нейросети с большим успехом моделирует многие функции нервной системы.
Нейросети способны осуществлять распознавание рисунков, как это делают
сенсорные системы, хотя такие модели и значительно проще, чем любая из
известных на сегодняшний день структур мозга. Нейросети способны учить-
ся и хранить информацию способами, сходными с теми, которые, как пола-
гают, имеют место в настоящих нейронах. С момента своего появления не-
сколько десятилетий назад искусственные нейросети применялись в самых
различных областях, начиная с компьютерного распознавания лиц и заканчивая
предсказанием обстановки на рынке ценных бумаг. Такое применение также
важно с научной точки зрения, поскольку дает нам представления для
выяснения механизмов, позволяющих мозгу делать то же самое. Мы должны
быть готовы к тому, что механизм, работающий в мозге, будет далек от модели
нейросетей, но она все же может служит отправной точкой.
4. Нейроны способны образовывать однонаправленные пути, подобные зри-
тельному тракту, оканчивающемуся в таламусе (латеральном коленчатом
теле). Тем не менее однонаправленные пути весьма редки. Гораздо обшир-
нее распространены двухсторонние пути, в которых информация движется
в обе стороны, и сети, в которых активность точки А обусловливает актив-
ность точки В, и наоборот. Это обстоятельство часто именуют возвратными
соединениями (Edelman, 1989).
Нейроны и связи между ними
119
5. Как мы впоследствии увидим, в нервной системе распространены масси-
вы нейронов, часто именуемые картами. Кора больших полушарий пред-
ставляет собой огромный шестислойный массив, состоящий примерно из
10 миллиардов нейронов с триллионами синапсов. Сетчатка глаза также
представляет собой массив, на этот раз трехслойный (см. гл . 6). Практиче-
ски любая сенсорная поверхность представляет собой массив рецепторов и
нейронов, осуществляющих первичную обработку и передачу информации.
Массивы также обнаружены в сенсорных ядрах таламуса и в четверохол-
мии. Распространенность массивов в мозге также можно принять в качестве
полезной рабочей гипотезы.
6. Хэббовские совокупности клеток. Когда нейроны образуют связи, полу-
ченный в результате паттерн активности может быть как стабильным, так и
нестабильным. Нестабильные паттерны, как правило, отмирают, тогда как
стабильные сохраняются в течение довольно долгого времени. Такие ста-
бильные паттерны часто называют совокупностями клеток; введение это-
го термина часто приписывают Дональду Хэббу. Хэббовские совокупности
клеток могут состоять как из близко расположенных клеток, так и из клеток,
отстоящих друг от друга на значительное расстояние. Совокупности клеток,
включающие как возбуждающие, так и тормозные синапсы, как правило, от-
личаются повышенной стабильностью. Короткоживущие соединения в моз-
ге имеют в основном электрохимическую природу, тогда как долгоживущие
требуют синтеза белка.
2.1.
Упрощенный случай: рецепторы, пути и контуры
Хотя рефлекторные дуги активируются внешним воздействием, они тем не ме-
нее полностью интегрированы в выполнение произвольных движений. К при-
меру, вы поворачиваете голову из стороны в сторону, когда читаете эту статью.
Таким образом, вы обеспечиваете выполнение произвольной задачи, а ваши
глаза тем временем обеспечивают фокусировку. Это — одно из важных свойств
сенсомоторной адаптации, причем осуществляющееся без участия произволь-
ных механизмов. Окуломоторная координация сложнее, чем обычная рефлек-
торная дуга.
Произвольные действия, подчиненные внешним целям, у людей ассоции-
рованы с корой. Сложные субкортикальные системы также задействованы в
планировании и исполнении различных действий. Спинномозговые центры
способны выполнять команды высших структур, используя сенсомоторные
рефлексы; также они посылают в мозг сигналы обратной связи. На всех уров-
нях контроля движения имеется как эндогенный, так и экзогенный (сенсорный)
входы, как осознанные, так и неосознанные (Goodale and Milner, 1992). Таким
образом, хотя и существует некоторое количество простых рефлексов, таких
как знаменитый коленный рефлекс, показанный на рис. 3 .8, рефлексы редко
работают изолированно. В норме они служат для выполнения обусловленных
корой задач.
Рефлексы являются врожденными механизмами. Они образуются в процес-
се развития; некоторые из них впоследствии исчезают, как, например, младен-
ческий хватательный рефлекс. Однако приобретенные непроизвольные про-
цессы рефлексами не являются. Они представляют собой сложные процессы,
120
Глава 3
называемые автоматизированными навыками, и зависят от практики. Так, вы,
читая эту книгу, прекрасно отличаете друг от друга буквы b и р, тогда как для
человека, незнакомого с алфавитом, сделать это куда сложнее. При разговоре и
письме контроль мышечных сокращений очень точный и большей частью бес-
сознательный. Человек использует множество автоматизированных навыков
для исполнения повседневных задач и нужд. Подобные навыки чаще всего бес-
сознательные, не требуют значительных усилий и не требуют детального созна-
тельного контроля. Их не следует путать с рефлексами. Автоматизированные
навыки изначально находятся под контролем коры, однако после интенсивной
практики они постепенно переходят под контроль субкортикальных структур.
(Haier et al., 1992; Chein and Schneider, 2005).
Рис. 3 .8. Простая рефлекторная
дуга. Если человека, положившего
ногу на ногу, несильно ударить под
коленную чашечку, то его верх-
няя нога подпрыгнет вверх. Это и
есть знаменитый коленный реф-
лекс, номенклатурно называемый
пателлярным. Этот рефлекс явля-
ется классическим спинномозго-
вым рефлексом, обусловленным
простейшей рефлекторной дугой.
Афферентный нейрон реагирует
на механическую стимуляцию и
преобразует ее в сигнал, идущий
в спинной мозг. Там вставочный
нейрон передает сигнал нейрону
моторному, который заставляет
мышцы сократиться. (Источник:
Standring, 2005.)
Рис. 3 .9 . Рецепторы преобразуют
внешнее воздействие в нейро-
нальный сигнал. Хотя показан-
ные здесь рецепторы принадле-
жат разным сенсорным системам,
они сходны как по структуре, так
и по функциям. Все они кодируют
внешнее физическое воздействие
в форму нейрональных сигналов.
(Источник: Squire et al., 2003.)
Нейроны и связи между ними
121
Каждый сенсорный нерв может содержать несколько параллельных кана-
лов, каждый из которых проводит несколько различающуюся информацию.
Так, зрительный тракт имеет канал передачи цвета, называемый мелкоклеточ-
ным, и канал передачи формы и размеров объекта, называемый крупноклеточ-
ным (рис. 3.9). Точно так же соматосенсорные пути сочетают каналы передачи
прикосновения, давления, боли и некоторые другие.
Большинство сенсорных волокон оканчиваются в таламусе, где они переда-
ют сигнал нейронам, оканчивающимся в коре (рис. 3 .10 и 3.11). Таламические
ядра обеспечивают соединения. Зрение и осязание организованы топографиче-
ски, сенсорные поля первичных рецепторов переходят в поля более высокого
уровня. Слух организован тонотопически; массивы нейронов при этом соответ-
ствуют определенной частоте звука.
На своем пути от периферии к коре большая часть сенсорных путей пересе-
кает плоскость симметрии тела. Эволюционное значение этого явления все еще
не выяснено, но тем не менее оно чрезвычайно распространено в организмах
как человека, так и других млекопитающих.
а
б
Рис. 3 .10 . Сходства сенсорных путей — осязание, зрение и слух. Все сенсорные си-
стемы начинаются с массивов рецепторов, таких как слои рецепторов прикосновения в
коже или сетчатка глаза. После первичной обработки на месте сенсорная информация
направляется для дальнейшей обработки в кору. Обратите внимание на то, что все три
указанных сенсорных пути оканчиваются в ядрах таламуса (зеленые). Все эти пути так-
же разделяются на две части, одна из которых оканчивается в противоположном полу-
шарии. Это явление носит название перекреста. (Источник: Herdy et al., цит. по Squire
et al., 2003, Elsevier.)
122
Глава 3
Хотя мы привыкли считать, что по различным путям сигналы идут только в
одну сторону, на самом деле это редко является правдой. Так, сигналы от сет-
чатки в таламус идут в одном направлении, тогда как исходящий сигнал оттуда
двунаправленный. До 90% нейронов, соединяющих таламус и зону V1, переда-
ют сигнал в обратном направлении — из зоны V1 в таламус. В слуховом анали-
заторе обратный поток сигналов идет непосредственно к рецепторам (см. гл . 5).
Таким образом, в большинстве сигнальных путей существуют петли об-
ратной связи — такие, как в нейросети с двумя или более слоями (рис. 3 .11).
Идельман с соавторами придавали обратной связи особое значение, считая ее
одним из основных свойств мозга (Edelman et al., 1989). С этой точки зрения
мозг представляется системой воздействующих друг на друга массивов и сетей.
2.1.1. Рецептивные поля и латеральные взаимодействия
Впервые гипотеза латерального ингибирования была предложена немецким
физиком Эрнстом Махом в 1860-х гг. на основе того, что целостные визуаль-
ные градиенты воспринимались как разорванные. Эта гипотеза блестяще под-
твердилась прямыми экспериментами (рис. 3 .12). Латеральное ингибирование
имеет место во многих системах мозга (рис. 3 .13). В сетчатке соседние клетки
ингибируют друг друга, благодаря чему яркая точка четко воспринимается на
темном фоне (рис. 3 .14). При прикосновении у соседних кожных рецепторов
также наблюдается латеральное ингибирование. На высших уровнях органи-
зации мозга сходные семантические конструкции ингибируют друг друга,
Рис. 3 .11 . Сенсорные органы взаимодействуют с ядрами таламуса. Таламус часто назы-
вают узловым центром мозга. Однако вполне возможно, что именно он является важней-
шей его частью. Обратите внимание на то, что зрительная, слуховая и соматосенсорная
информация, прежде чем попасть в кору, достигает таламуса. Вместе с тем из коры также
поступают сигналы в таламус, что означает наличие непрерывного потока информации
между корой и многочисленными таламическими ядрами. В некоторых случаях таламус
усиливает активность коры, тогда как в других — тормозит или блокирует ее. Обратите
внимание на сходство кортикального входа и выхода всех трех систем. ЛКТ — латераль-
ное коленчатое тело, РЯ — ретикулярные ядра, МКТ — медиальное коленчатое тело,
ВБ — вентробазальный комплекс ядер таламуса. (Источник: Alitto and Ursey, 2003.)
Нейроны и связи между ними
123
Рис. 3 .12 . Латеральные взаимодействия. Часто одна и та же стратегия используется в
различных областях мозга. Слева показано рецептивное поле типа «Центр-периферия»
латерального коленчатого тела. Свет падает в область красного пятна в центре, но не в
окружающее кольцо. Нейроны реагируют на оба этих факта и усиливают контраст за
счет взаимного ингибирования в одном и том же слое, называемого латеральным инги-
бированием. Сходный механизм используется в рецептивном поле в центре, а также в
колонках коры крысы, каждая из которых обрабатывает сенсорный вход одной вибрис-
сы. По этой причине колонки часто выступают объектом исследования при изучении
подобных процессов. Соседние вибриссы оказывают друг на друга ингибирующее вли-
яние, имеющее место также в областях мозга человека, обрабатывающих звуковую и
тактильную информацию, а также в области, контролирующей внимание. (Источник:
Alitto and Ursey, 2003.)
Рис. 3 .13 . Наглядная демонстрация латерального ингибирования. Впервые механизм
латерального ингибирования был предложен немецким физиком Эрнстом Махом в
1860-х гг. на основе наблюдения восприятия непрерывного градиента как прерывисто-
го. Гипотеза с того времени нашла неоднократное блестящее подтверждение. Заметьте,
что влияние латерального ингибирования заметно при рассматривании близколежащих
черных квадратов, восприятии противоположных цветов, а также при рассмотрении па-
раллельных линий, пересеченных многочисленными наклонными отрезками. (Источ-
ник: Eagleman, 2001.)
124
Глава 3
благодаря чему мы не путаем слова «астрономия» и «астрология». Как и мно-
гие другие нейрональные механизмы, обсуждаемые в этой главе, этот механизм
появился рано в эволюционной истории и затем неоднократно повторялся во
многих системах.
3.0 .
Массивы и карты
Как мы уже указывали ранее, массивы нейронов широко распространены в
мозге. Массив представляет собой двумерную сеть нейронов. Когда массивы
соответствуют пространственной организации той или иной структуры, их на-
зывают картами. Пространственные карты являются одной из форм простран-
ственного кодирования в нервной системе. В мозге имеет место как временное,
так и пространственное кодирование, наряду со многими другими способами
кодирования и обработки информации. Пространственные карты являются
наиболее наглядной формой пространственного кодирования (рис. 3 .15 и 3.16).
Сетчатку часто представляют в форме трехслойной карты световых рецеп-
торов, включая ганглиозные клетки, аксоны которых простираются от сетчатки
до таламуса (точнее, его латерального коленчатого тела) (см. гл . 6). Клетки та-
ламуса затем передают сигнал в первичную зону зрительной коры — зону V1
затылочной доли. Зона V1 также является картой зрительного входа — разные
четверти поля зрения проецируются в разные же области этой зоны. Из зоны
V1 зрительная информация передается в другие карты, некоторые из которых
расположены в пределах зрительной коры, тогда как другие лежат вне ее. Раз-
решение после прохождения зоны V1 падает, поскольку зрительные карты выс-
ших уровней реагируют на более абстрактные стимулы, такие как лица и объ-
екты, а также связаны с сетями, контролирующими внимание, эмоции и даже,
Рис. 3 .14 . Рецептивные поля вида «центр-периферия» в сенсорной коре. Картирова-
ние соматосенсорных (тактильных), зрительных и слуховых сигналов осуществляется
нейронами с рецептивными полями вида «центр-периферия», реагирующими на те или
иные свойства сенсорного стимула, такие как расположение, цвет или частота. З
—
зеленый; К — красный; С —синий; Ж — желтый
Нейроны и связи между ними
125
возможно, принятие решение. И хотя эта схема сильно упрощена, она весьма
точно отражает реальное положение дел.
Такая соматосенсорная информация, как информация о прикосновении и
боли, также обрабатывается кортикальными картами. Другие чувства, такие
как слух, вкус и обоняние, гораздо меньше ассоциированы с положением ре-
цепторов, однако в слуховой коре имеется картоподобный клиновидный реги-
он. Таким образом, даже информация от не ассоциированных с пространством
органов чувств обрабатывается массивами и картами нейронов. После обработ-
ки информация от сенсорных систем комбинируется в теменной области коры
с использованием массивов, объединяющих тактильную, слуховую и зритель-
ную информацию в комплексные карты. Они могут быть привязаны к телу (эго-
центрическая карта) или ко внешнему объекту (аллоцентрическая карта). Та-
ким образом, наш мозг организует огромные количества входящей информации
так, чтобы отражать положение окружающих объектов. Моторная кора, как вы
можете догадаться, также выглядит как непропорциональная карта скелетных
мышц тела (см. гл . 5).
Рис. 3 .15. Зрительный вход проециру-
ется в соответствующие области коры.
Tootell с соавторами (1996) показали,
что окружность вокруг точки фиксации
зрения вызывает соответствующую ак-
тивность в зоне V1 зрительной коры.
Авторы воспользовались топографиче-
ским представлением зрительного вхо-
да в области V1 и симметрией окруж-
ности относительно точки фиксации
зрения. На рисунке показана уплощен-
ная проекция зоны V1. (Источник:
Tootell et al., 1996.)
Рис. 3 .16 . Соотношение четвертей
поля зрения с областями зрительной
коры. Заметьте, что в зоне V1 разные
четверти поля зрения проецируют-
ся в разные области. Нижняя правая
четверть проецируется в верхнюю по-
ловину зоны V1 левого полушария.
Первичные зрительные поля сосредо-
точены в затылочной области коры во-
круг зрительной борозды. (Источник:
Zeki, 2003.)
126
Глава 3
Очень привлекательно думать, что все многочисленные карты сенсорной
коры централизованно управляются некой структурой. Однако на самом деле
это неверно. Слабым местом идеи о контролирующем центре представляется ее
неспособность что-либо объяснить — она лишь передвигает вопрос на уровень
выше, ведь и этот центр также нуждается в регуляции (рис. 3 .17).
Рис. 3 .17. Зрительные карты макаки. Зрительный анализатор макака-резуса часто из-
учается по причине его сходства с таковым у человека. Слева показаны основные зри-
тельные пути коры макаки. Каждый из них ведет от одной карты к другой. Начиная с
зоны V1 импульс направляется в последующие зоны с сохранением топографии сиг-
нала. Верхний путь чувствителен к положению, и по этой причине его часто именуют
путем «где». Нижний путь проводит информацию относительно цвета, формы, кон-
траста и природы объекта; по этой причине он носит название пути «что». Полагают,
что активность этого пути прямо связана с осознанными зрительными ощущениями,
берущими начало, по всей видимости, в области опознания объекта. Путь «где» опре-
деляет область действия пути «что». Этот процесс не находится под прямым контролем
сознания, однако обусловливает воспринимаемую локализацию объектов. (Источник:
Lamme and Roelfsma, 200.)
Рис. 3 .18 . Двусторонняя передача
информации между массивами. Дву-
сторонняя передача информации —
распространенное в мозге явление.
Поэтому может быть полезно предста-
вить зрительные области в виде слоев
двунаправленной сети, а не в виде од-
нонаправленных путей. Заметьте, что
низшие слои чувствительны к простым
стимулам, тогда как высшие — к слож-
ным объектам, таким как лица. Суще-
ствует тем не менее постоянный нисхо-
дящий поток информации. (Источник:
Ahissar and Hochstein, 2004.)
Нейроны и связи между ними
127
Таким образом, главным вопросом относительно сенсорики на сегодняш-
ний день яляется вопрос о том, как осуществляется высокоуровневая обработка
воспринятой информации. И модель нейросетей предоставляет один из воз-
можных ответов.
3.1.
Карты переходят в другие карты
В нервной системе нередки случаи организации многих слоев нейронов в еще
более сложные сети. Так, вся кора представляет собой шестислойную сеть, со-
держащую около 10 млрд нейронов. Иерархические структуры мозга представ-
ляют собой соединенные массивы нервных клеток.
3.2 .
Массивы нейронов чаще всего
имеют связи двух типов
В процессе исследования зрительного анализатора был установлен один весьма
неожиданный факт: большая часть аксонов, соединяющих таламические ядра с
корой, направлены не от таламических ядер, а к ним. Более 90% аксонов между
латеральным коленчатым телом и зоной V1 проводят информацию к латераль-
ному коленчатому телу. На высших уровнях также существует двунаправлен-
ный поток информации, как между зонами V1 и V2 или V2 и V3. В свете идеи
о иерархическом устройстве анализатора такой нисходящий поток информации
может показаться странным. Однако далее мы убедимся, что двунаправленная
передача информации — это норма, а не исключение (рис. 3 .18). Однонаправ-
ленных потоков в коре очень мало. Этот факт заставляет рассматривать нейро-
нальную сигнализацию как нечто куда более сложное, чем кажется изначально.
Далее мы более детально рассмотрим двунаправленную сигнализацию. Рецеп-
тивные поля сенсорных нейронов по мере продвижения вверх по иерархиче-
ской лестнице усложняются.
3.3 .
Сенсорные и моторные системы работают совместно
Хотя существует четкое анатомическое разделение между сенсорными и мотор-
ными путями, они тем не менее взаимодействуют между собой. Когда мы гово-
рим, мы можем слышать себя. Когда мы идем, изображение на нашей сетчатке
меняется. Видеоигры, представляющие собой по сути серию быстроменяю-
щихся картинок, дают весьма достоверное ощущение движения. Мозг постоян-
но корректирует работу моторных систем на основании сенсорной информации
и адаптирует сенсорные системы при помощи моторной активности. Класси-
ческая схема коры Фустера резко акцентирует на этом внимание (рис. 3 .19).
Информация в нервной системе непрерывно циркулирует между моторной и
сенсорной системами, что необходимо для их синхронизации (Fuster, 2004;
Kandel et al., 2004).
Fuster (2004) предполагает, что сенсорные системы можно представить
в виде иерархических систем, состоящих из иерархических систем низшего
порядка, начиная с рецепторов и постепенно переходя ко все более сложным
объектам (см. рис. 3 .19). Моторные системы могут быть представлены в виде
иерархической структуры противоположного направления, заканчивающиеся
на мотонейронах. Тем не менее, как видно из диаграммы, идет непрерывный
обмен информацией между двумя системами в процессе цикла от восприятия
128
Глава 3
Рис. 3.19 . Сенсорные и моторные иерархические системы сами организованы в иерар-
хические системы. Фустер (2004) предположил, что вся кора может быть представлена
в виде взаимосвязанных иерархических систем, расположенных в задней (сенсорной)
и передней (моторной) частях коры. Информация из сенсорной коры направляется в
унимодальную ассоциативную зону, что, как полагают, вызывает «побуждение» к дей-
ствию. Выше полимодальные ассоциативные зоны комбинируют тактильные, слуховые
и зрительные сигналы, а также взаимодействуют с передней частью префронтальной
коры. Также не забывайте о постоянном притоке информации извне. Благодаря этому
мы можем слышать собственную речь и видеть смену окружения при ходьбе. (Источ-
ник: Fuster, 2004.)
Рис. 3 .20 . Ступенчатая пирамида
как иерархическая система. Полез-
но представлять себе иерархическое
устройство мозга в виде ступенчатой
пирамиды, подобной пирамиде инков
Мачу-Пикчу. Уровни располагаются
один над другим, но тем не менее сиг-
нал может идти в различных направле-
ниях. Так, люди могут передвигаться
по пирамиде вверх, вниз и в сторону
Нейроны и связи между ними
129
до действия, начиная с наинизшего и заканчивая высшими уровнями планиро-
вания, мышления и анализа возможного развития событий (на рис. 3 .20 показа-
на иерархическая система из области архитектуры).
Friston построил очень полезную схему иерархической системы обработки
информации (рис. 3.21), которую мы принимаем. В этой схеме каждый массив
нейронов назван картой; карты существуют на разных уровнях, и сигнал может
идти вверх, вниз и к другой карте того же уровня. Примеры подобного мы уви-
дим в последующих главах.
Обратите внимание на то, что модель нейросетей часто использует разные
термины для обозначения одних и тех же понятий. Так, термин «снизу вверх»
аналогичен термину «прямой поток информации». Передача «сверху вниз» так-
же называется «обратной связью» или «обратным потоком информации». Что-
бы не путаться, просто поверните мысленно иерархическую пирамиду набок,
и тогда направление «снизу вверх» станет для вас направлением «прямо».
3.4 .
Временная кодировка:
рисунки спайков и ритмы мозга
Массивы, карты и иерархические структуры являются основой пространствен-
ного кодирования в мозге. Тем не менее в мозге имеет место также и временное
кодирование. На рис. 3 .22 показаны примеры двух различных рисунков спай-
ков одного и того же нейрона. Время прохождения спайка относительно других
Рис. 3 .21 . Абстрактная иерархическая система. Заметьте, что, подобно ступенчатой пи-
рамиде, в иерархической системе поток информации возможен во всех направлениях.
Такое строение типично для сенсорных и моторных иерархических систем. (Источник:
Friston, 2003, с изменениями.)
130
Глава 3
нейронов также содержит информацию, так же как положение стрелки на часах
показывает текущее время относительно 24-часового цикла. Нейроны, подоб-
ные этому, могут выступать в роли водителей ритма для крупных популяций
кортикальных нейронов (рис. 3 .23). Другая гипотеза говорит о том, что быстрое
прохождение серии спайков таламических нейронов опосредует пробуждение
при значительном сигнале, таком как звук плача младенца. Эта гипотеза не ли-
шена смысла, поскольку древесные обезьяны, к примеру, должны реагировать
очень быстро, когда ветка дерева ломается.
Отдельные нейроны также имеют свои рисунки спайков. Так, слуховой нерв
осуществляет частотное кодирование (см. гл . 5). При вибрации барабанной пе-
репонки механическое ее движение передается слуховыми косточками на улит-
ку, в которой вибрирующая жидкость стимулирует волосковые клетки.
Волосковые клетки и являются слуховыми рецепторами, отвечающими ак-
сонным ПД на механическую стимуляцию. Их аксоны образуют слуховой нерв;
при сравнительно низкой частоте звука частота прохождения сигналов по нему
соответствует частоте колебаний барабанной перепонки. В этом случае имеет
место кодировка простым преобразованием одного сигнала в другой. Это —
временная, а не пространственная кодировка (см. разд. 3 .5).
Рис. 3 .22 . Нейроны имеют различные рисунки спайков. Пространственное кодирова-
ние в мозге очевидно, однако также имеет место и изменение сигналов с течением вре-
мени. Здесь представлены результаты отведения мембранного потенциала от стимули-
руемых таламических нейронов. Эти нейроны имеют два различных рисунка спайков.
(McCormack and Huguenard, 1992.) (Источник: White, 2002, из Ramachandran, 2002.)
Рис. 3 .23 . Регистрация активности
единичного нейрона животного. Иголь-
чатый электрод вводится безболез-
ненно, поскольку в самом мозге от-
сутствуют болевые рецепторы. После
имплантации электрод никак не пре-
пятствует свободным движениям кош-
ки. Электрод принимает спайки еди-
ничного нейрона, показанные вверху
справа. Поведение нейрона во времени
отображается на экране компьютера.
(Источник: Tsodyks et al., 1999.)
Нейроны и связи между ними
131
При рассмотрении электрической активности десятков миллиардов нейро-
нов мозг поневоле начинает казаться огромным оркестром, а не одним инстру-
ментом. За сотни миллионов лет эволюции в мозге появились нейроны с самы-
ми разными видами временного и пространственного кодирования (блок 3.1).
Блок 3.1 . Некоторые способы визуализации нейрональной коммуникации
Нейрональные коммуникации можно визуализировать с использованием стандарт-
ных графиков.
На рис. 3.24(а) показана зависимость активности одного нейрона от активности
другого. Регистрировались спайки целевого нейрона (постсинаптического) при про-
хождении спайка на пресинаптическом нейроне. Это позволяет оценивать эффектив-
ность синаптической передачи. Регистрировалась активность нейронов в такой зоне
мозга, в которой четко прослеживается зависимость активности одного нейрона от
активности другого. Пресинаптическая клетка принадлежит зрительному таламусу
и связана с нейронами первичной зрительной коры — зоны V1 (см. гл . 6).
По вертикальной оси отложено количество спайков пресинаптического нейрона в
единицу времени. Большинство нейронов имеет постоянную фоновую спайковую
активность, поэтому интерес представляет только активность заметно больше фо-
новой. В этом примере активность двух нейронов очень тесно связана. Постсинап-
тическая клетка генерирует аксональный ПД спустя несколько миллисекунд после
прохождения спайка по пресинаптическому нейрону. Таким образом, имеет место
очень точная синаптическая передача сигнала через синапс.
На рис. 3 .24(а) показан простейший пример взаимодействия двух нейронов. Однако
гораздо чаще взаимодействия между ними носят гораздо более сложный характер.
Зависимость активности одного нейрона от другого можно построить для двух лю-
бых связанных нейронов.
На рис. 3.24(б) показана корреляция на уровне целого мозга. На диаграмме кора че-
ловека показана как круг, причем глаза и нос находятся в верхней части рисунка.
Черные линии показывают синхронный гамма-ритм по данным отведения внутри-
черепных электродов. (При помощи интракраниальных электродов обследуют боль-
ных эпилепсией перед хирургическим вмешательством.) Гамма-ритмом называют
мозговой ритм на энцефалограмме в области от 25 до 120 Гц.
Больным эпилепсией демонстрировали вспышки света, находившиеся в левой и
правой точках фиксации зрения (Doesburg et al., 2007). На рис. 3 .24(б) показан ответ
Рис. 3 .24 (а). Диаграмма зависимо-
сти активности нейрона зрительной
коры от активности таламического
нейрона. (Источник: Sherman and
Guillery, 1998, с изменениями.)
132
Глава 3
Рис. 3 .24 (б). Схематическое изображение активности головного мозга в ответ на
визуальную стимуляцию левого поля зрения, вид сверху. В левой части рисунка по-
казан путь, пройденный сигналом, возникшим в ответ на визуальную стимуляцию.
Черные линии на схеме головы в правой части рисунка обозначают синхронные гам-
ма-ритмы, возникшие в зрительной коре и других частях коры, находящихся как в
левом, так и в правом полушарии. Синхронные гамма-ритмы, как полагают, играют
важную роль в передаче сигнала между областями коры, имеющей место при мно-
гих когнитивных задачах. (Источник: Doesburg et al., 2007, с изменениями.)
Рис. 3 .24 (в). Зависимость частоты от времени, используемая для отображения ре-
зультатов исследования нейрональной сигнализации в мозге. На таком графике ясно
виден частотный характер электромагнитной активности в некоторый период вре-
мени после стимуляции, а также ее интенсивность. На рисунке показан масштабный
выброс высокой интенсивности в гамма-области (около 50–150 Гц), начинающийся
спустя 100 мс после стимуляции и резко идущий на убыль спустя 500 мс. Черные
полосы в нижней части графика соответствуют снижению интенсивности сигнала
в тета- и альфа-областях с частотой 4–12 Гц, длящемуся порядка одной секунды.
Горизонтальная черная полоса под графиком соответствует длительности стимула
(см. гл . 8). (Источник: Lachaux, J. P. et al., 2007.)
Нейроны и связи между ними
133
3.5 .
Точки выбора в потоках информации
Для нас было бы большим сюрпризом, если бы вдруг обнаружилось, что ин-
формация в мозге двигается по неразветвленным путям. Напротив, в путях
следования информации имеется множество точек выбора, с которых она мо-
жет быть направлена по нескольким различным путям или быть передана на
уровень выше или ниже. Если вернуться к ступенчатой пирамиде на рис. 3.20,
то такой разветвленный путь похож на путь человека к вершине: он может до-
стигнуть ее прямым или окружным путем, а то и вовсе пританцовывая по пути.
По каким же признакам мы можем установить наличие точек выбора? Хотя
анатомических доказательств в целом вполне достаточно, существует еще одно,
дополнительное доказательство: человек и животные в своей жизни встречают-
ся с целым рядом объектов, трактуемых неоднозначно. Большая часть слов раз-
говорного языка имеет более одного значения (сознательно мы редко отмечаем
этот факт, однако в этом легко убедиться, заглянув в словарь). На самом деле
словарь недооценивает разнообразие смыслов слов, поскольку имеет значение
мозга на демонстрацию вспышки в левой точке фиксации зрения. Из-за перекре-
ста зрительного пути стимуляция левой точки фиксации приводит к возбуждению
правой части зрительной коры. При возбуждении затылочная зона коры генерирует
гамма-ритмы, которые вызывают ответы во многих других частях коры, о чем можно
судить по высокой синхронности соответствующих гамма-ритмов. Подобная связан-
ная с задачами синхронность обнаружена во многих частях мозга (см. гл . 8).
Наконец, на рис. 3 .24(в) приведена зависимость частоты от времени для активности
мозга после зрительной стимуляции; приведенные данные получены Lachaux с соав-
торами (2007) с использованием магнитоэнцефалографа и транкраниальных электро-
дов в латеральной затылочной извилине, которая входит в состав зрительной коры.
На графике по вертикальной оси отложена частота сигналов энцефалограммы, а по
горизонтальной — время в миллисекундах, прошедшее после интересующего экспе-
риментатора события, которым может быть, к примеру, демонстрация лица. Цвет на
цветовой шкале отражает интенсивность электрической активности; желтый и белый
цвета соответствуют наибольшей интенсивности.
Как вы можете видеть, имеет место взрывообразное повышение активности непо-
средственно после t = 0 . Частота импульсов высокой интенсивности при этом со-
ставляет 50–150 Гц и длится примерно 100–500 мс после стимуляции. Графики та-
кого типа широко распространены в когнитивной нейронауке, и важно понимать их.
Черные полосы в нижней части рис. 3 .24(в) соответствуют альфа-ритмам частотой
8–12 Гц, характерным для зрительных задач..
Литература
1. Sherman, S.M., Guillery, R.W. (1998). On the actions that one nerve cell can have on
another. Distinguishing ‘drivers’ from ‘modulators’. Proceedings of the National Academy
of Sciences USA, 95(12), 7121–7126.
2. Lachaux, J-P., Baillet, S., Adam, C., Ducorps, A., Jerbi, K., Bertrand, O., Garnero, L.,
Martinerie, J. (2007). A simultaneous MEG and intracranial EEG study of task-related
brain oscillations. International Congress Series, 1320, 421–424 .
3. Doesburg, S.M., Roggeveen, A.B ., Kitajo, K., & Ward, L.M . (2007). Large-Scale Gamma-
Band Phase Synchronization and Selective Attention Cerebral Cortex doi:10.1093/cercor/
bhm073.
134
Глава 3
не только само слово, но и, скажем, интонация, с которой оно было произне-
сено. Существует множество таких параметров, которые составитель словаря
неспособен учесть, — в отличие от нашей нервной системы.
Зрительная картина мира подвержена постоянным изменениям, на которых
наше сознание практически не фиксирует внимания. Солнечный свет изменяет
свой цвет на закате. В лесу свет достигает поверхности, пройдя через свето-
фильтры листьев деревьев. Стекла солнечных очков и даже обычные оконные
также работают как светофильтры. Такие различия редко фиксируются созна-
нием, поскольку зрительная система стремится к поддержанию постоянства
цветов, полагаясь в этом на память. Однако мозг тем не менее ведет обработку
таких изменений и отделяет их от изменений цвета, которые должны достичь
сознания. Животное не может позволить себе не заметить хищника, прячуще-
гося в траве, только потому, что сейчас закат, или потому, что на него падает
тень от близлежащих деревьев, равно как и по причине утреннего тумана. Для
того чтобы выжить, мы должны иметь превосходную зрительную систему.
На рис. 3 .25(б) показан знаменитый куб Неккера. На рис. 3 .25(а) показана
широко известная картинка, в которой можно увидеть и два человеческих лица,
и вазу.
Вы можете сказать, что эти примеры искусственны и в реальности ничего
подобного вам встретиться не может, однако вы будете неправы (рис. 3 .26).
Рис. 3 .25. Неоднозначные стимулы
заставляют нервную систему делать
выбор. Два общеизвестных неодно-
значных рисунка: (а) Иллюзия «ли-
цо-ваза» и (б) куб Неккера. Углы куба
будут казаться направленными то
внутрь, то наружу, если их рассма-
тривать, не видя окружения. (Источ-
ник: Kim And Blake, 2005.)
Рис. 3.26 . Неоднозначно трактуемый
объект. Что это такое? Форму подчас
трудно бывает выделить и идентифи-
цировать даже в естественной среде:
вы можете убедиться в этом, попы-
тавшись рассмотреть животное, пря-
чущееся в кустах, птицу, летящую в
тумане, или машину, приближающу-
юся со стороны солнца
Нейроны и связи между ними
135
Любой угол комнаты на самом деле представляет собой часть куба Неккера.
(Чтобы убедиться в этом, достаточно свернуть в трубку лист бумаги и
посмотреть через него на любой из углов комнаты. При этом его вершина будет
восприниматься то как направленная к наблюдателю, то как направленная от
него.)
Кошка, выслеживающая жертву, может осматривать дерево только одним
глазом, тогда как другой глаз остается пассивным. Это приводит к явлению
бинокулярной конкуренции — конкуренции между зрительными входами от
разных глаз. Многие животные получают от разных глаз совершенно разные
входы — такие животные, как кролики и олени, вообще не имеют участков
перекрытия полей зрения, поэтому для них явление бинокулярной конкуренции
невозможно.
Лица очень важны с биологической точки зрения для людей и прочих при-
матов. Импринтинг образа матери основан на опознании лица, так же как и
конфронтации, половые взаимоотношения и многое другое. Однако лица также
содержат неоднозначности. Взгляните на рис. 3 .27 . В какую сторону смотрит
Хамфри Богарт? Какие эмоции выражает его лицо? На какой из фотографий он
выглядит более скептическим или подозрительным?
3.6 .
Обработка, обусловленная ожиданием
Мозг постоянно имеет некоторые ожидания относительно встречаемых им
внешних условий. Спускаясь по лестнице в темноте, мы ожидаем, что под ногой
будет ступенька. При анализе неоднозначно трактуемых объектов ожидание об-
условливает выбор наиболее приемлемого варианта трактовки. Многие слова в
языке имеют больше одного значения, поэтому, даже читая это, вы вынуждены
иметь дело с неоднозначностями. Мозг опирается не только на входящую ин-
формацию — он имеет множество причин выбора того или иного варианта, ос-
нованных на предсказании результата и ожидании. Латеральные процессы, как
мы уже знаем, также играют важную роль при определении границ стимулов,
как, например, при анализе зрительной информации. Как мы увидим позже,
селективное внимание позволяет нам динамически изменять свои сенсорные
предпочтения (см. гл . 8), а долговременная память увеличивает силу синапсов,
ответственных за точное восприятие.
Карты и слои не следует рассматривать как единственные функциональные
единицы мозга. Многие области коры связаны между собой, в результате чего
возбуждение одной области приводит к возбуждению других. По этой причине
Рис. 3 .27. В какую сторону смотрит
Хамфри Богарт? Лица — одни из
самых важных среди окружающих
нас объектов. Способность опреде-
лять направление взгляда и выра-
жение лица — одни из важнейших
социальных навыков. (Источник:
Wexler, 2001.)
136
Глава 3
многие ученые считают, что всю кору целиком, наряду с сопутствующими об-
ластями, такими как таламус, следует рассматривать как одну функциональную
единицу. Ее часто называют таламокортикальной системой. Тем не менее для
того, чтобы понять, как эта сложная система работает, следует подробнее из-
учать ее отдельные части.
Пластичный мозг
В течение многих лет исследование обучения у человека и животных было связано
с экспериментальной психологией. Причины этого вполне понятны: одним из ос-
новных свойств поведения животных является способность адаптироваться — вы-
яснять, какие места или действия ведут к получению удовольствия, какие — к боли,
а какие — несущественны. Помимо этого, следует помнить, что внешние условия
непостоянны во времени. Условия или действия, ранее приводившие к полезным
результатам, могут оказаться вредными, и животное должно адаптироваться к это-
му независимо от того, сколько раз до этого оно получало пользу от этого действия.
Основным свойством мозга, таким образом, является приспособляемость.
Однако какие изменения в структуре самого мозга приводят к такой приспособляе-
мости? Подход, которым пользуется нейропсихология, а именно изучение поврежде-
ний мозга, для исследования приспособляемости не совсем удачен, поскольку про-
цессы, лежащие в ее основе, динамичны и обусловлены пластичностью практически
всего мозга, а не только отдельных его областей. Для этих целей гораздо лучше под-
ходят методы визуализации структур мозга, получившие интенсивное развитие в два
прошедших десятилетия. Хотя большая часть методов визуализации ориентирована
на конкретную область, подчеркивая тем самым функциональное разделение, а не
интеграцию, были предприняты попытки изучения обучения как системного про-
цесса, включающего глобальные изменения структуры и функций мозга.
С точки зрения функциональности, мозг представляет собой постоянно изменяю-
щуюся систему, активность отдельных областей которой изменяется под воздей-
ствием опыта. Тем не менее подобное локальное изменение активности — лишь
часть общей картины, поэтому более поздние методы визуализации разрабатыва-
лись с упором на исследование взаимного влияния областей, проявляющегося в
усилении или ослаблении влияния одной области на другую. Недавние исследо-
вания показали, что человеческий мозг крайне чувствителен к побочным случай-
ным взаимосвязям между слуховыми и зрительными стимулами (den Ouden et al.,
2009). Интересно отметить, что хотя добровольцы, участвовавшие в эксперименте,
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
Рис. 3 .28. Пол Флетчер (Paul Fletcher, PhD),
Институт поведенческих и клинических ней-
ронаук Кембриджского университета (Вели-
кобритания)
Нейроны и связи между ними
137
и не были осведомлены о наличии таких взаимосвязей, наблюдалось усиление свя-
зи между первичной зрительной и слуховой зонами при воздействии на них по-
добных стимулов; при этом сами добровольцы даже не догадывались о том, что их
мозг сравнивает стимулы. Возможно, эти данные помогут понять, каким образом
некоторые люди способны при определенных обстоятельствах действовать как «ин-
туитивные статистики» (Shanks, 1995).
Хотя изучение функциональных связей выглядит привлекательным, структурные
основы таких связей остаются неясными. Тем не менее развитие технологии маг-
нитно-резонансной томографии позволило начать изучение изменений структур-
ных компонентов связи — трактов белого вещества — под влиянием обучения. Как
было показано (Scholz et al., 2009), обучение жонглированию вызывает изменения
как в сером веществе, так и в упомянутых выше трактах (рис. 3 .29). Это означает,
что в результате обучения моторному навыку происходят изменения как в сером,
так и в белом веществе мозга. Данные результаты стали поистине революционны-
ми, поскольку долгие годы считалось, что структура мозга неизменна. Подобные
открытия, позволяющие рассматривать мозг как функционально и структурно ла-
бильный орган, вне всякого сомнения являются шагом вперед в нашем понимании
процесса обучения.
Макроанатомические изменения, сходные с описанными, непременно должны со-
провождаться также изменениями на уровне синапсов, изучение которых предоста-
вит гораздо более полное понимание приспособляемости мозга к изменяющимся
Рис. 3 .29 . Изменения белого вещества в мозге в результате беговых тренировок.
(а) Цветные точки отражают кластеры (скорректированы относительно p < 0,05)
достоверного увеличения ФА (фракционная анизотропия) последних тренировок
по сравнению с более ранними, наложенные на усредненную карту ФА. (б) Среднее
изменение изображения ФА кластера, указанного в (а), по сравнению со сканом 1.
Планки погрешности отображают стандартные ошибки среднего (* — достоверное
отличие по отношению к базовой линии, p < 0,05; НТБ — нижняя теменная бороз-
да, ТЗБ — теменно-затылочная борозда). (Источник: Scholz et al., 2009.)
138
Глава 3
внешним условиям. Такое понимание само по себе послужит основой для изучения
отклонений в процессах обучения, которые могут быть причиной многих психиче-
ских заболеваний, таких как шизофрения (Fletcher, Frith, 2009).
Литература
1. den Ouden, H.E ., Friston, K.J., Daw, N.D ., McIntosh, A.R., & Stephan, K.E . (2009
May). A dual role for prediction error in associative learning. Cerebral Cortex, 19(5),
1175–1185.
2. Shanks, D.R . (1995). The Psychology of Associative Learning. Cambridge: Cambridge
University Press.
3. Scholz, J., Klein, M.C., Behrens, T.E.J., Johansen-Berg, H. (2009). Training induces
changes in white matter architecture. Nature Neuroscience, 12, 1370–1371 .
4. Fletcher, P.C., & Frith, C.D. (2009). Perceiveing is believing: A Bayesian approach to
explaining the positive symptoms of schyzophrenia. Nature Reviews Neuroscience,
10(1), 48–58.
4.0 .
Адаптация и обучение массивов нейронов
Процесс обучения интенсивно изучался на слоях стимулируемых нейронов.
После периода относительно слабого прогресса, в 1980-х гг. в этой области
имел место скачкообразный рост (Rumelhart и McClelland, 1986). Теоретиче-
ской основой подобных исследований служил коннекционизм. Большая часть
этой главы опирается на результаты именно коннекционистского подхода к об-
учению (см. далее раздел «Дарвинистский подход в нервной системе»). И хотя
первые исследования нейросетей имели место в 1950-х гг., ранние работы в
этой области сталкивались со множеством сложностей. К 1980-м годам стало
ясно, что добавление еще одного, скрытого слоя способно устранить многие из
этих сложностей, как и рассмотрение обратной связи. Именно эти дополнения
и дали начало коннекционистскому движению.
Рис. 3 .30. Дональд Хэбб. Дональд Хэбб был
одним из теоретиков, оказавших наиболь-
шее влияние на нейронауку. Он дал четкое
определение совокупности клеток и предло-
жил наиболее известное правило обучения
нейросетей, выражаемое в лозунге «neurons
that fire together, wire together». (Источник:
Brown and Milner, 2003.)
Нейроны и связи между ними
139
4.1.
Обучение по Хэббу: «Активирующиеся одновременно
нейроны образуют соединения между собой»
Дональд Хэбб (см. рис. 3 .30) в 1949 г. постулировал, что ассамблеи нейронов
способны обучаться благодаря усилению связей между нейронами, активиру-
ющимися при стимуляции одновременно. Эта идея заключена в высказывании
«Активирующиеся одновременно нейроны образуют соединения между со-
бой». Этот способ обучения оказался очень полезен для нейросетей, и на на-
стоящий момент имеются свидетельства в пользу того, что обучение в нервной
системе происходит сходным образом.
Ключевой идеей обучения по Хэббу является положение о том, что в основе
обучения и памяти лежит эффективность синаптической связи. Как показано
на рис. 3 .31, существует множество способов воздействия на эффективность
синаптической передачи. Так, два нейрона могут образовать больше синапсов,
в самих синапсах может вырабатываться больше нейромедиатора, рецепторы
постсинаптического нейрона могут стать эффективнее, и т. д.
На настоящий момент полагают, что в обучении задействовано два типа
изменений; их можно рассматривать как усиленное возбуждение и усиленное
торможение. Долговременное усиление возбудимости одного нейрона называ-
ют долговременой потенциацией. Долговременное понижение же — долговре-
менной депрессией. Существуют данные, свидетельствующие в пользу того,
что оба события имеют место в гиппокампе.
Полезно не упускать из виду промежуток времени, потребовавшийся для
развития современного мозга, — мозг млекопитающих эволюционировал в те-
чение 200 млн лет, но даже раньше мозг низших позвоночных был способен
Рис. 3.31 . С инапс Хэбба и долговременное потенцирование. Может существовать
несколько способов увеличения эффективности синаптической передачи.
(Источник: Burne, цит. по Squire et al., 2003.)
140
Глава 3
на выработку сложных взаимоотношений с окружающей средой. Поэтому, не-
смотря на убедительные доказательства обучения мозга по Хэббу, не стоит от-
вергать возможность существования и других механизмов обучения.
Визуально обучение по Хэббу можно представить в виде утолщения линий
между узлами сети, как в простой совокупности клеток на рис. 3 .32. Здесь зри-
тельный цикл активирует соответствующие клетки модели ранней зрительной
коры. Узлы цикла соответствуют небольшим областям поля зрения, и связь
между ними усиливается при их одновременной активации. Так утверждает
теория. На рис. 3.33 приведены экспериментальные данные относительно уве-
личения вероятности совместной активации областей гиппокампа кошки, под-
тверждающие эту модель. Тем не менее мозг — крупная и сложная структура,
и в силу этого нельзя говорить о справедливости модели для всего мозга после
подтверждения этого для отдельной области. Однако этот результат не может
не обнадеживать.
На рис. 3 .34 представлена простая сеть, которая может использоваться для
распознавания входящих сигналов. В ней имеются двусторонние вертикальные
связи и латеральное ингибирование на каждом из слоев. Иногда подобные
сети называют локальными, поскольку в ней не задействовано обусловленное
обучением усиление связей. Тем не менее такая сеть полезна при наглядном
объяснении некоторых основных когнитивных процессов (Rumelhart and
McClelland, 1986b).
Рис. 3 .32 . Хэббовское обучение сово-
купности клеток. Нейроны представле-
ны в виде кругов, а связи между ними —
в виде линий. Более интенсивный
красный отображает более активные
клетки, а более толстые линии — более
сильные соединения. В момент времени
t1 совокупность клеток кодирует опре-
деленный ответ. Этот ответ сохраня-
ется в моменты времени t2 и t3. Более
реалистичные модели включают посте-
пенное забывание, предотвращаемое пе-
риодической стимуляцией. (Источник:
Abraham and Robins, 2005.)
Рис. 3 .33 . Экспериментально наблюдавшееся обучение по Хэббу. Усиление нейрональ-
ных связей непосредственно наблюдалось для нейронов гиппокампа кошки. (Источник:
Sutherland and McNaughton, 2000.)
Нейроны и связи между ними
141
На рис. 3 .35 показана новая модель сети. Ранние модели были разработаны
в 1950-х гг. (Rosenblatt et al., 1962), были одно- или двухслойными и встречали
логические преграды при их применении к анализу основных функций мозга,
таких как обучение или распознавание паттернов. Некоторое время модели
нейросетей пребывали в забвении, пока около двадцати лет назад не были
разработаны модели с третьим, скрытым слоем, который позволяет нейросети
изменять силу соединений. На рис. 3 .35 представлена классическая трехслойная
прямая сеть со скрытым слоем и настраиваемой силой взаимодействий. Эта
сеть может эффективно обучаться путем сопоставления выхода нейросети с
желаемым выходом и подстройки силы соединений для достижения желаемого
результата. Процесс носит название обратного распространения ошибки
обучения и во многом подобен отрицательной обратной связи. Сети такого типа
на сегодняшний день наиболее распространены.
На рис. 3.36 показана самоорганизующаяся аутоассоциативная сеть, в ко-
торой выход ставится в соответствие входу. Такая стратегия полезна при рас-
познавании паттернов, таких как звук знакомого голоса. Самоорганизующиеся
Рис. 3.34 . Простейшая сеть. Обратите
внимание на сочетания возбуждаю-
щих и тормозных связей. (Источник:
Palmer-Brown et al., 2002.)
Рис. 3 .35. Классическая трехслойная
сеть. Скрытый слой делает сеть зна-
чительно более лабильной. Обратная
передача сигнала об ошибке позволя-
ет сети сопоставлять выход с обуча-
ющим стимулом и изменять силу свя-
зей в сети в соответствии с ним. Так
сеть способна учиться достижению
своей цели на основе обратной связи.
(Источник: Abraham and Robins, 2005.)
142
Глава 3
системы используются в природе для решения многих задач. Сами организмы
и их нервные системы можно рассматривать как самоорганизующиеся систе-
мы. В человеческой культуре главенствующее место занимает обучение. Начи-
ная с момента рождения человек учится у других людей. Это позволяет выйти
за рамки самоорганизующихся обучаемых систем и развить знания и навыки,
получаемые в результате обучения, что соответствует сетям, показанным на
рис. 3.35 и 3.36.
На рис. 3 .37 показано, как самоорганизующаяся сеть способна справляться
с фундаментальой проблемой распознавания человеческих лиц. Человек учит-
ся реагировать на нормальные, недеформированные лица в очень ранний пери-
од жизни и вскоре становится способен отличать знакомые лица от незнакомых.
Задача, решаемая сетью, гораздо проще решаемой человеком, поскольку в мо-
дели происходит только формирование цепи. Сеть способна обучаться пред-
угадывать расположение рта в нижней части рисунка и двух глаз — в верхней.
Однако она неспособна работать с трехмерными объектами, с объектами раз-
ной освещенности, движущимися и другими, еще более сложными объектами.
Тем не менее распознающая лица сеть, приведенная на рис. 3 .37, позволяет нам
попытаться понять, каким образом мозг справляется с этой жизненно важной
задачей (см. гл . 6).
Рис. 3 .36 . Сеть распознавания паттернов. Аутоассоциативная сеть сопоставляет выход
со входом, что полезно для распознавания паттернов, таких как лица или автомобили.
На правом рисунке часть нейронов окрашена в красный, что обозначает усиление связи
после обучения. (Источник: Burne, из Squire et al., 2003.)
Нейроны и связи между ними
143
4.2 .
Дарвинистский подход в нервной системе:
выживают клетки и синапсы,
наиболее приспособленные к данной задаче
Джеральд Эдельман постулировал, что мозг представляет собой сложный се-
лекционистский орган. Теория Эдельмана известна как нейральный дарвинизм,
поскольку в ней предполагается, что нейроны развиваются и соединяются друг
с другом в соответствии с дарвинистскими принципами. Бологическая эволю-
ция происходит за счет размножения, мутаций, ведущих к изменениям, и от-
бору полезных изменений. С течением времени этот процесс дает разные виды
животных, приспособленные к определенным экологическим нишам. Сходный
процесс идет в иммунной системе, позволяя организму синтезировать антитела
к чужеродным веществам и не реагировать на свои собственные. Так, клетки,
успешно отвечающие на проникновение чужеродных агентов, размножаются,
тогда как неспособные это делать — уменьшаются в количестве. Таким спосо-
бом иммунная система учится распознавать и бороться с новыми угрозами. Это
подтверждает эффективность селекционизма как способа адаптации.
Согласно представлениям Эдельмана (1989), мозг имеет две стадии селек-
ционистской адаптации. Первая начинается спустя короткое время после опло-
дотворения, во время дифференцировки, размножения и соединения нейронов.
Сохранившиеся после этой стадии нервные клетки составляют мозг. Вторая
стадия начинается до завершения первой, во время образования первых соеди-
нений между нейронами. Адаптированные соединения сохраняются, тогда как
остальные — пропадают. Таким образом, подобие естественного отбора дей-
ствует как во время дифференцировки, так и во время обучения (рис. 3 .38). Дву-
сторонние связи между группами нейронов делают возможной двустороннюю
обработку информации. Если такая обработка стабильна, то она сохраняется.
Если же нет, то со временем она исчезает.
Если вспомнить принцип обучения по Хэббу, гласящий, что «активирую-
щиеся одновременно нейроны образуют соединения между собой», то полу-
чится, что нейроны в стабильной группе должны образовывать более сильные
Рис. 3 .37. Сеть распозна-
вания лиц. Распознавание
лиц — одна из базовых про-
блем зрительной системы.
Здесь распознавание проис-
ходит при помощи рисования
простой линии. Однако если
лицо находится в естест-
венных условиях, задача
становится гораздо сложнее
и приобретает больше изме-
няющихся параметров. Обра-
тите внимание на то, что эта
сеть — трехслойная. (Источ-
ник: Luckman et al., 1995.)
144
Глава 3
соединения между собой. Таким образом, отбор нейронов будет приводить к
образованию долгоживущих нейронных совокупностей, выполняющих задачи
адаптации, обучения, разпознавания паттернов и им подобные.
Эти принципы были улучшены при помощи результатов изучения мозга и
создания компьютерных моделей. Была создана серия роботов, использующих
селекционистские приемы для симуляции работы определенных областей моз-
га, к примеру гиппокампа. На рис. 3 .39(а) показан робот, имитирующий поведе-
ние крысы в водном лабиринте Морриса, в котором животное, плавающее в не-
прозрачной жидкости, должно найти платформу, расположенную на небольшой
глубине и неразличимую с поверхности. На рис. 3 .39(б) показан робот, исполь-
Рис. 3 .38 . Пример выполнения теории Дарвина при обучении. На этом рисунке приве-
дены стадии кодирования паттерна активации нейронов, происходящего тогда, когда ди-
намическая активность синапсов способствует установлению долговременных связей,
позволяя тем самым памяти храниться там же, где образовались исходные соединения.
(Источник: Riberio et al., 2006.)
Рис. 3 .39 . (а) Модель, имитирующая поведение крысы в водном лабиринте Морри-
са, — крыса плавает до тех пор, пока не найдет платформу, на которую можно встать.
Модель создает нейрональную карту лабиринта, позволяющую обучаться нахождению
платформы. (б) Робот, использующий дарвинистский подход для обучения игре в фут-
бол. (Источник: Neuroscience Institute, Krichmar, 2006.)
Нейроны и связи между ними
145
зующий селекционизм для обучения игре в футбол. И хотя эти задачи сильно
различаются, основные принципы при обучении их выполнению сходные.
4.3 .
Обработка символьной информации и нейронные сети
Стандартная компьютерная программа пишется при помощи символов. Ней-
росети можно описать в математических терминах, при этом оказывается, что
они отличаются высоким уровнем параллельности (что означает способность
производить много разных вычислений одновременно) и распределенности
(способности обрабатывать информацию в разных местах с использованием
разных механизмов). Это говорит о большей близости нейросетей к биологи-
ческим способам обработки информации, чем к стандартной алгебре и логике.
Тем не менее человек создал математику и логику, а язык позволяет нам хоро-
шо понимать символические выражения. Большинство используемых нами слов
являются символами, обозначающими то или иное явление. Более того, нейрон-
ные сети довольно просто перевести на язык математических выражений.
Как же свести воедино эти две разные философии обработки данных? Один
из предложенных методов (McClelland, Rogers, 2003) показан на рис. 3.40,
Рис. 3 .40 . Нейросети способны обрабатывать символьную информацию. Эта нейросеть
представляет набор простых утверждений, таких как «Малиновка — это птица» или
«У розы есть лепестки». (Источник: McClelland and Rogers, 2003.)
146
Глава 3
изображающем нейросеть, разработанную для сопоставления символов (та-
ких как «рыба» и «расти») с их логическими взаимоотношениями (такими как
«имеет свойство»). С точки зрения человека, это — сложный способ для выра-
жения простого утверждения о том, что «рыба может расти». Однако для боль-
ших нейросетей, содержащих миллионы единиц и триллионы связей, таких как
наш мозг, эта модель может оказаться вполне жизнеспособной.
Таким образом, нейросети способны обрабатывать символьную информа-
цию, а символы могут быть переведены в нейросети. Это означает, что символь-
ный и нейросетевой способы обработки информации не являются взаимоисклю-
чающими. Главный вопрос будет в результате заключаться в том, какая форма
обработки будет наиболее подходящей для той или иной задачи. Если стоит за-
дача понимания функционирования мозга на уровне нейронов, то здесь логич-
но использовать модель нейросетей. Если же нам требуется понять грамматику
языка, то символы, вне всякого сомнения, подойдут для этой задачи лучше.
Адаптация и представление являются двумя параллельными способами рас-
смотрения мозга. Ниже мы увидим доказательства того, что мозг специализи-
рован для познания мира. Также будут приведены доказательства, говорящие
об адаптивных процессах в мозге, позволяющих приспосабливаться к меняю-
щимся условиям. Обе эти идеи, по всей видимости, фундаментальны. Неко-
торые результаты будут свидетельствовать об адаптивной стороне мозга, не-
которые — о репрезентативной. Мы увидим, что обе точки зрения могут быть
полезны.
5.0 .
Координация работы нейрональных сетей
Мозг часто называют параллельным органом, поскольку нейроны работают до
определенной степени независимо друг от друга. Не существует единого ко-
мандного центра, диктующего нейронам, что делать.
Несмотря на это, существует масса способов координации работы нейронов.
Одним из них являются масштабные ритмы, координирующие работу больших
групп нейронов так же, как дирижер координирует игру симфонического ор-
кестра. Если большая масса нейронов активируется одновременно, то их ак-
тивность, как правило, суммируется. Тем не менее существуют ограничения
такой суммации. Долгое время считалось, что эпилептические припадки вызва-
ны нейральной шрамовой тканью, обозначавшейся как эпилептический фокус,
которая, как считалось, генерирует регулярные, периодичные и интенсивные
волны возбуждения, активирующие другие группы нейронов. Результатом это-
го являются потеря сознания и неконтролируемая двигательная активность.
Имеется очевидная необходимость регуляции между координацией и вы-
полнением нейронами и их группами локальных функций. Должен быть баланс
между интеграцией и дифференциацией (Edelman and Tononi, 2000).
На рис. 3 .41 приведена усредненная корреляция между областями зритель-
ной коры во время просмотра подопытным фильма. Как вы можете видеть,
одни и те же области в разных полушариях (обозначены как «l» и «r») активи-
руются одновременно, что показано желтыми и красными линиями. Поскольку
оба полушария получают практически одинаковый вход от глаз, этот результат
выглядит вполне логичным. Также внутри каждого полушария наблюдается
строгая корреляция между областью раннего зрительного анализа и областью
Нейроны и связи между ними
147
визуального восприятия объекта. Тем не менее поскольку в этом исследовании
использовался метод функционального магнитного резонанса, данные которо-
го, полученные в течение длительного периода времени, затем усреднялись,
быстрые корреляционные взаимодействия остались не видны. Над преодоле-
нием этого методологического ограничения в настоящее время ведутся интен-
сивные работы. Современные данные говорят в пользу гораздо более быстрых
гамма- и тета-корреляций на тех частотах, на которых мозг выполняет большую
часть этой работы (Fries, 2005).
Ритмы энцефалограммы на сегодняшний день считаются сигнализирую-
щими о разных, но скоординированных процессах. К примеру, гамма-ритмы
высокой плотности считаются ассоциированными с осознанным зрительным
восприятием и процессом решения простй проблемы эквивалентности. Аль-
фа-ритмы традиционно ассоциируются с отсутствием задач, требующих фоку-
сировки внимания, тогда как тета-ритмы, как полагают на настоящий момент,
контролируют гиппокампальную область и фронтальную кору в процессе об-
ращения к долговременной памяти. Дельта-ритмы — сигналы глубокого сна —
группируют быструю нейрональную активность с целью консолидации полу-
ченных данных (Kemp et al., 2004).
Рис. 3 .41 . Корреляция между активностью областей мозга в процессе просмотра филь-
ма. Наглядная иллюстрация коррелированной активации зрительной коры в процессе
просмотра подопытным фильма. Верхняя часть центрального изображения соответ-
ствует левому полушарию, нижняя часть — правому. Области V с одинаковыми номера-
ми сооветствуют левой и правой сторонам, которые обычно соединены через середину.
Более теплые цвета и более толстые линии обозначают более тесную корреляцию меж-
ду областями. Обратите внимание на то, что наиболее сильная корреляция наблюдает-
ся между активностями одинаковых областей разных полушарий. LOp — задняя часть
латерального затылочного комплекса. LOI — латеральная часть латерального затылоч-
ного комплекса. (l) — левое полушарие, (r) — правое полушарие. Слева представлены
данные фМРТ. (Источник: Bartels and Zeki, 2005.)
148
Глава 3
На рис. 3 .42 представлена простая и довольно убедительная гипотеза от-
носительно механизма координации регулярными мозговыми ритмами актив-
ности миллионов отдельных нейронов. Нейроны, по которым проходит волна
возбуждения на пике, к примеру альфа-волны, добавляют небольшую часть
электрохимической активности к общей. Нейроны, активные в минимуме вол-
ны, свою активность вычитают. В результате этого синхронизированные нейро-
ны подкрепляются положительной обратной связью от нейронов, поддержива-
ющих ритм, тогда как десинхронизированные — подавляются. Такой механизм
усиливает ритмичность работы нейронов.
Как мы уже подчеркивали, простой синхронной активности в большинстве
случаев недостаточно. Разные группы нейронов должны быть способны гене-
рировать разный ответ на вход, который может соперничать с ответами, гене-
рируемыми другими группами. Такая модель показана на рис. 3 .38 и включает
дарвинистскую конкуренцию между разными популяциями клеток.
5.1.
Функциональная избыточность
При проектировании самолета инженеры закладывают в его конструкцию не-
которую функциональную избыточность на случай выхода из строя важнейших
систем. Так, если откажет один двигатель, то большая часть самолетов будет
способна дотянуть до взлетно-посадочной полосы на оставшихся. Человек
и животные также обладают определенной функциональной избыточностью.
Рис. 3 .42. Сеть распознавания паттернов. Желтая звезда на левом верхнем рисунке
представляет собой исходный вход для предварительно обученной нейросети. После-
дующие рисунки отображают текущее состояние сети, показанное в виде окрашенных
точек. Обратите внимание на то, как цвет и форма желтой звезды постепенно проявля-
ются по мере распознавания сетью входа и отсечения ею альтернативных вариантов.
(Источник: M. Shannahan, Imperial College London, с разрешения.)
Нейроны и связи между ними
149
Мозга это правило тоже касается. Даже потеря значительной части коры не яв-
ляется невосполнимой, если произошла в раннем детстве. Мозг способен рабо-
тать даже после получения весьма значительных повреждений.
6.0.
Заключение
Основным вопросом когнитивной нейронауки является вопрос о том, каким об-
разом взаимодействие нервных клеток делает возможным восприятие, память
и действия. Эта проблема еще очень далека от полного решения, однако опре-
деленные успехи на этом пути уже имеются. Достигнуто достаточно полное
понимание функционирования интегративного нейрона в результате его ин-
тенсивного изучения. Однако в мозге существует множество типов нейронов,
взаимодействий между ними и нейромедиаторов, играющих важные роли в его
работе. Интегративный нейрон был выбран в качестве модели в силу того, что
он достаточно полно изучен и может быть упрощен, что позволяет строить раз-
личные модели на его основе. Для этих целей полезны как полностью искус-
ственные, так и построенные на основе биологических нейросетей.
В некоторых случаях нейроны образуют простые циклы, такие как рефлек-
торная дуга коленного рефлекса. Гораздо чаще они являются частью больших
двумерных сетей, которые, в свою очередь, организованы в иерархические
структуры. Все сенсорные и моторные комплексы можно рассматривать в фор-
ме таких структур. Для зрения, осязания и двигательного контроля эти нейроны
представляют собой топографические карты окружения. Зрительную систему,
к примеру, можно рассматривать как иерархию таких карт.
Иерархические структуры не являются жестко детерминированными путя-
ми. Сигнал в них способен двигаться вверх, вниз и латерально относительно
структуры. Основной функцией обратного потока сигналов является разреше-
ние неоднозначностей в трактовке входящей информации. Неоднозначности
часто встречаются в зрительном входе, но имеются и в языке, а также в других
типах воспринимаемой нами информации. В моторных системах обратный по-
ток сигналов помогает принимать решения относительно моторных команд.
Латеральное торможение является распространенной стратегией для вы-
деления различий между двумя однородными областями сигнала, такими как
темные пятна на светлом фоне. Клетки сенсорных систем имеют так называе-
мые рецептивные поля, настроенные на определенные параметры входа, такие
как ориентация линии, цвет, движение, форма и тип объекта. При повышении
уровня визуальных карт их разрешение падает, в то время как способность к
интеграции информации растет. Поскольку сенсорные и моторные системы
изучают отдельно друг от друга, мозг представляется нам огромным сенсор-
но-моторным органом, делающим возможным непрерывные высокоуровневые
взаимодействия между входом и выходом.
Пространственные массивы нейронов делают возможным пространственное
кодирование, однако не стоит забывать о том, что в нервной системе имеется
еще и кодирование временное. Основные ритмы энцефалограммы, как
полагают, отвечают за временную координацию активности больших групп
нейронов. Последние исследования позволяют предположить, что гамма-ритм
ответственен за интеграцию сенсорной информации осознанные ощущения,
а тета-ритм — за извлечение информации из долговременной памяти.
150
Глава 3
7.0 .
Задания и упражнения к главе 3
7.1.
Контрольные задания
1. Опишите основные функции интегративного нейрона.
2. Что такое латеральное торможение и какую роль оно играет в сенсорных
системах?
3. Каким образом сенсорные и моторные системы можно рассматривать в фор-
ме иерархических структур?
4. Опишите роль двусторонних взаимодействий в функционировании мозга.
5. Что такое дарвинистский подход к нервной системе и какие аспекты про-
цессов, происходящих в мозге, он затрагивает?
6. Назовите три наиболее общих свойства сенсорных систем.
7.2 .
Графические упражнения
1. Обозначьте части, отмеченные цифрами на рис. 3 .43.
2. Нарисуйте обучение совокупности клеток по Хэббу в зрительном цикле.
Какой фразой описывается обучение по Хэббу?
Рис. 3 .43 . Идеализированный нейрон
Различные изображения «системы внутренних магистралей» коры — проводящих во-
локон, идущих внутри мозга, — построенные при помощи метода под названием диф-
фузионная трактография. Все волокна на представленных изображениях идут крест-
накрест через мозолистое тело, а затем загибаются вверх наподобие фонтанов в обоих
полушариях. (а) Вид сбоку, отображающий волокна, возносящиеся вверх в левом по-
лушарии мозга. Зеленые волокна расположены в передней области (префронтальной),
голубые волокна берут начало и оканчиваются в премоторной коре, темно-синие во-
локна восходят из моторной коры, оранжевые — из теменной, а желтые проводящие
волокна идут от затылочной коры одного полушария в затылочную кору другого. Пучки
волокон фиолетового цвета, расположенные вдоль среза, связывают височные доли моз-
га. (б) Показаны пучки проводящих волокон в обоих полушариях мозга, направленные
назад (желтые) и вперед (зеленые). Обратите внимание на ориентационный компас,
расположенный в правом нижнем углу рисунка, указывающий направление проекций:
П — передний; Н — нижний; Л — левый; З — задний; Пр — правый; В — верхний.
(Источник: Hofer and Frahm, 2006.)
Я верю, что изучение нейровизуализации подтвердило предположение о лока-
лизации умственных процессов в пределах головного мозга человека.
Михаэль И. Познер (2003)
Содержание
1.0.Введение.......................................................153
1.1 . Регистрация работы мозга: более и менее прямые методы измерения. . 155
1.2 . Соотношение пространственного и временного разрешений методов . 155
2.0 . Ряд полезных инструментов для измерения электрической и магнитной
активности......................................................159
2.1.Регистрацияодиночныхнейронов...............................159
2.2.Исследованиянаживотныхичеловеке...........................163
2.3.Электроэнцефалография ......................................165
2.4.Магнитоэнцефалография......................................173
2.5.Вмешательствовработумозга.................................174
3.0.фМРТи ПЭТ: непрямое измерение нейронной активности. . . . . . . . . . . . . . . 180
3.1.Областиинтереса................................................185
3.2. Мозг в состоянии покоя не молчит: внутренние процессы мозга . . . . . . . . 188
3.3. Эмпирическое определение когнитивных функций:
уникальныйспособ..................................................189
4.0 . Новые способы определения взаимосвязи областей мозга:
визуализацияраспределениятензора ...............................191
5.0.Сознательныесобытияпротивбессознательных ......................192
6.0.Корреляцияипричиннаясвязь......................................193
6.1 . Для чего необходимо множество методик при исследовании
функциймозга...............................................195
6.2.Повреждениямозгаиследствияэтихповреждений ................196
7.0.Заключение.....................................................198
8.0.Заданияиупражнениякглаве4....................................198
8.1.Контрольныезаданияиграфическиеупражнения .................198
1.0 . Введение
В этой главе представлены различные методы измерения активности мозга
с точки зрения вдумчивого исследователя, который стремится разобраться в
том, как работает мозг. В приложении эти методы рассмотрены более подробно.
Если бы существовал идеальный прибор для регистрации деятельности
мозга, то можно было бы проследить за работой десятка миллиардов нейронов
и зарегистрировать активность каждой клетки с частотой в тысячу раз за секун-
ду. Использование такого идеального устройства позволило бы проследить за
постоянно меняющимся взаимодействием между маленькими и более больши-
ми группами нейронов, которые формируют между собой триллионы всевоз-
Глава 4
Методы:
визуализация живого мозга
154
Глава 4
можных связей. Можно привести в качестве аналогии такой пример — очень
хороший и мощный спутник-шпион был бы способен из космоса видеть как
каждого отдельного человека, так и проследить изменения во взаимоотноше-
ниях между индивидуумами и группами, от отдельной семьи до целой нации.
Однако такого идеального прибора для наблюдения не существует. И наше
понимание работы мозга — это коллаж из множества отдельных фрагментов,
собранных вместе таким образом, чтобы получилась приемлемая цельная кар-
тина. Но мечта об идеальном устройстве для наблюдения за работой мозга дает
нам те параметры, к которым мы должны стремиться.
Методы нейроимиджинга мозга — это прорыв для когнитивной нейронауки,
которая многие десятилетия формировалась на основе когнитивной психологии,
инструментальных условных рефлексов, психофизики и науки о мозге. До того мо-
мента, как инструменты визуализации деятельности мозга стали более совершенны-
ми, наше знание о мозге базировалось на исследованиях, проводимых на животных,
и на данных, получаемых при случайных травмах у людей. Однако данные о функ-
ционировании мозга, получаемые при его травмах, чрезвычайно неточны, и часто,
чтобы определить точное место повреждения, неврологи должны прибегнуть к по-
смертному обследованию мозга пациентов — что и было в случае пациентов Брока
и Вернике, как было описано в гл. 1 . Во многих случаях мозг способен скомпенси-
ровать повреждение, поэтому характер поражения меняется с течением времени, по
мере того как некоторые клетки отмирают, и возникает адаптация. Следовательно,
посмертное обследование необязательно отразит характер травмы, каков он был на
момент постановки диагноза. Исследования, проводимые на животных, основыва-
ются на предполагаемых гомологиях — сходствах между видами — которые, однако,
не всегда являются убедительными. Никакой другой вид не обладает речью и други-
ми чисто человеческими особенностями, поэтому очень трудно понять, как соотне-
сти функционирование мозга животных и человеческое сознание.
Многие проблемы были решены с появлением технологии визуализации
работы живого мозга, сначала при помощи электроэнцефалографии (ЭЭГ), за-
тем рентгеновских лучей и в конце концов компьютерной томографии (КТ, из-
учение срезов — от греч. tomos — срез) на основе рентгеновских лучей, пози-
тронно-эмиссионной томографии (ПЭТ) и магнитно-резонансной томографии
(МРТ). В настоящее время у нас в арсенале более дюжины технологий, которые
очень быстро совершенствуются. Потребности медицины часто являются дви-
жущей силой для развития этой очень дорогостоящей технологии, так как она
позволяет обследовать многие органы организма. И в результате мы обладаем
способами для исследования распространения миллиардов нейрохимических
рецепторов в мозге, толщину коры, большую магистральную систему пучков
белых волокон и, что самое важное для когнитивной нейронауки, функциональ-
ную активность мозга — основу адаптивной способности. Новые усовершен-
ствования дают ученым возможность изучать не только функциональную ак-
тивность в определенных областях мозга, но и измерять динамичный паттерн
связей между областями. Наиболее массивная часть «проводки» мозга изобра-
жена на рисунке в начале главы, но, как и в случае всемирной компьютерной
паутины, это является лишь небольшой частью: существует множество посто-
янно меняющихся функциональных связей между популяциями нейронов, ко-
торые способны преобразовываться в течение долей секунды.
Методы: визуализация живого мозга
155
1.1 .
Регистрация работы мозга:
более и менее прямые методы измерения
Некоторые способы нейровизуализации способны регистрировать нейронную ак-
тивность напрямую. Если мы поместим электрод рядом с нейроном (или внутрь
его), мы можем записать его электрическую активность, а именно потенциалы дей-
ствия на аксоне. Этот одноэлектродный метод можно считать прямым измере-
нием электрической активности нейрона. Как уже говорилось в главе 3, нейро-
ны не только разряжаются спайками. Дендриты — ветви, несущие информацию
к нейрону, — также вовлечены в важные процессы. Регистрируя разные части
нейрона, мы получаем несколько различные показания активности нейрона.
Это очень хорошо проиллюстрировано в недавних исследованиях авто-
ров из Калифорнийского технологического института (Quiroga et al., 2005)
(рис. 4 .1). Эта группа исследователей обнаружила, что один нейрон в медиаль-
ной височной коре селективно активировался при демонстрации фотографии
актрисы Дженнифер Энистон. Этот нейрон не реагировал при предъявлении
изображений других актрис или актеров, а также всевозможных картинок до-
мов, пейзажей и объектов. Исследователи также нашли другие нервные клетки
с аналогичной высокоселективной активацией, в том числе один нейрон, чув-
ствительный к предъявлению изображения актрисы Холли Берри, и другой — к
изображению Сиднейского оперного театра. Разумеется, необходимо все время
помнить о том, что мы регистрируем только небольшое количество из милли-
арда нейронов. В любом случае, это очень сложная сеть взаимосвязанных ней-
ронов, и только счастливый случай может выявить фаната Дженнифер Энистон
или Сиднейского оперного театра.
Исследователи в качестве показателя нейронной активности использовали
частоту аксональных разрядов. Значит ли это, что данный единичный нейрон
«знает» Дженнифер Энистон? Отнюдь нет. Более правильно было бы говорить
о том, что активность этой клетки отражает большую и протяженную сеть ней-
ронов, которые чувствительны к одному определенному лицу. Если единичный
нейрон потерян, человек был бы все равно в состоянии узнать Дженнифер Эни-
стон. Мозг как единое целое не претерпел бы никаких заметных изменений.
Регистрация единичных нейронов у человека является чрезвычайно редкой
практикой. По этическим соображениям это возможно только у пациентов с
фармакологически неизлечимой эпилепсией или раком головного мозга, ког-
да единственным шансом является хирургическое удаление поврежденного
участка. Электроды погружают глубоко в мозг для идентификации расположе-
ния необходимых для когнитивной деятельности областей, таких как центры
речи. Если такое хирургическое вмешательство необходимо, то ученые могут в
течение очень короткого времени, пока электроды погружены в мозг, провести
некоторые тесты.
1.2.
Соотношение пространственного
и временного разрешений методов
Наиболее часто применяемые на сегодняшний день методы показаны на
рис. 4 .2 . Несложно заметить, что ни один метод не обладает необходимым про-
странственно-временным разрешением для прослеживания единичных нейро-
нов и даже небольших групп нервных клеток, например колонок в коре. Одна-
156
Глава 4
ко можно с уверенностью сказать, что эти методы обретают силу. Считается,
что технологии как фМРТ, которая позволяет регистрировать физиологические
изменения, включая насыщение крови кислородом, обладают хорошей про-
странственной разрешающей способностью, но довольно слабой временной
разрешающей способностью. Время ответа фМРТ составляет около 6 с, так как
для изменения локального кровотока требуется некоторое время — это очень
Рис. 4 .1 . Нейрон Дженнифер Энистон. Обнаружен нейрон в области левого гиппокам-
па, селективно реагирующий на различные изображения актрисы Дженнифер Энистон
(Quiroga et al., 2005). (а) Ответы на 30 из 87 предъявленных фотографий, показаны на
рисунке. (б) Показаны нейронные спайки в ответ на предъявление 30 фотографий из 87.
Числа указывают на номер изображения; каждый столбик отображает количество за-
регистрированных нейронных спайков. Единичные нейроны, вероятно, отражают боль-
шие нейронные сети с разными чувствительностью и предпочтением
Методы: визуализация живого мозга
157
медленно для непосредственной регистрации единичных нейронов и их попу-
ляций. Тем не менее недавние исследования показывают, что фМРТ может от-
ражать разряды нейронов, произошедшие 6 с ранее.
По сравнению с ЭЭГ и магнитоэнцефалографией (МЭГ), которые исполь-
зуют электрические и магнитные сигналы соответственно, фМРТ имеет очень
хорошую пространственную разрешающую способность, поэтому ее очень
часто применяют для локализации функций мозга. Но ЭЭГ и МЭГ обладают
очень высокой временной разрешающей способностью — почти мгновенны —
и относительно низкой пространственной точностью. С помощью этих методов
можно регистрировать усиление и затихание разрядов в популяциях нейронов
за десятки и сотни миллисекунд, но при этом невозможно точно сказать, какая
группа нейронов вызвала этот сигнал. Именно поэтому в некоторых исследова-
ниях для достижения оптимального пространственного и временного разреше-
ния одновременно применяют ЭЭГ и фМРТ. Объединение двух регистрирую-
щих технологий позволило узнать много нового о работе мозга.
Мы приводим несколько МРТ-изображений для демонстрации возможности
этой технологии картировать области мозга (рис. 4 .3). Можно получить все-
стороннее изображение строения мозга, используя разные варианты срезов, та-
ких как фронтальный (рис. 4 .4), горизонтальный или центральный саггитальный
(рис. 4.3), или воссоздать трехмерное изображение мозга (рис. 4.4).
Рис. 4.2 . За и против различных методов визуализации мозга. Различные методы ре-
гистрации активности мозга имеют свои плюсы и минусы относительно их точности.
В то время как у некоторых методов очень высокое временное разрешение, но низкая
пространственная разрешающая способность, у других методов эти значение противо-
положны
158
Глава 4
Рис. 4 .3 . Программа навигации по мозгу. Программное обеспечение для определе-
ния местоположения в мозге позволяет пользователям переводить точные координаты
(в формате x, y, z) в определенные мозговые структуры. Обратите внимание на ориента-
цию стандартного среза. Координаты x, y, z известны под названием системы Талайраха
(Talairach и Tournoux, 1988). Анатомические ориентиры, такие как мозолистое тело, мо-
гут быть увидены на верхнем левом дисплее. (Источник: Mai and Thomas, 2006.)
Рис. 4.4 . Коронарный срез
мозга. Трехмерный кадр из
программы Brain Navigator.
Желтым и красным цветом
обозначены области ме-
диальной височной коры
в правом и левом полуша-
риях. (Источник: Mai and
Thomas, 2006.)
Методы: визуализация живого мозга
159
2.0 .
Ряд полезных инструментов для измерения
электрической и магнитной активности
2.1.
Регистрация одиночных нейронов
Наиболее точным методом регистрации на сегодняшний день считается ре-
гистрация одиночных нейронов или «единиц» с использованием игольчатых
электродов для глубокого погружения, которые иногда имплантируют в мозг
(рис. 4 .5). Во время регистрации нейронов часто пишется активность десятка
или даже нескольких сотен нейронов за один раз. Игольчатые электроды, как
правило, регистрируют потенциалы действия на аксонах, которые чрезвычайно
важны для обработки и передачи информации. Но в процессе регистрации оди-
ночных клеток возникают определенные проблемы — можем ли мы быть уве-
ренными, что локально измеренные разряды нейронов отражают активность
всего региона? Также большое значение имеет, являются ли регистрируемые
нейроны возбудительными (вызывают усиление разряда других нейронов) или
тормозными (подавляют разряд других клеток). К тому же вживление электро-
дов в мозг относится к инвазивным и потенциально опасным методам, кото-
рые требуют хирургического вмешательства. Поэтому данный метод применим
только на экспериментальных животных или на людях, которым по медицин-
ским показаниям, например неизлечимая эпилепсия, требуется операция. Мы
подробнее рассмотрим каждый из этих случаев.
В действительности с тех пор как Хьюбел и Визель (1962) зарегистрировали
единичные нервные клетки, чувствительные к определенным свойствам зрительно-
го стимула в зрительной коре кошки, за что они и получили Нобелевскую премию
в 1981 г., мы можем регистрировать активность единичных нейронов, которые за-
действованы в важных мозговых процессах. В более поздних работах была зареги-
стрирована активность нейронов в медиальной височной области (рис. 4 .6). Как и
Рис. 4 .5 . Регистрация активности единич-
ных нейронов в глубинных структурах моз-
га. Внутриклеточная и внеклеточная реги-
страция активности нейронов. Обратите
внимание на существование возможности
стимулировать единичные нейроны через те
же электроды. Метод регистрации активно-
сти нейронов подходит близко к желательно-
му уровню пространственного и временного
разрешения, необходимому для регистрации
активности мозга. Данный метод хорошо ра-
ботает при выявлении нейронных сетей. Од-
нако этот метод не позволяет выявлять круп-
номасштабные популяции нейронов
160
Глава 4
любой другой метод, регистрация электрической активности аксонов имеет свои
ограничения, однако все равно он остается одним из главных источников информа-
ции. Частота разрядов нейронов достигает 1000 Гц, но для корковых нейронов сред-
няя частота разряда составляет около 10 Гц. Ученые не обладают «универсальным
знанием» о кортикальных нейронах, поэтому очень сложно судить о том, насколько
показательны данные, полученные от небольших групп исследуемых клеток.
Во многих странах регистрация нейронной активности при помощи глубин-
ных электродов разрешена на приматах, в частности на макаках, при условии
соблюдения биоэтических норм. Мозг макаки имеет удивительное сходство с
мозгом человека. Регистрация одиночных нейронов в префронтальной коре у
макаки может продемонстрировать феномен рабочей памяти. Во время такого
эксперимента макаку обучают фиксировать свой взор на изображении креста на
экране компьютера и выполнять отсроченные реакции на появляющиеся зри-
тельные стимулы. Животное обучается выжидать определенное время, прежде
чем перевести взгляд с точки фиксации на вновь появившийся стимул или в
противоположную от стимула сторону. При выполнении такой задачи мы можем
регистрировать активность единичного нейрона префронтальной коры в трех
состояниях: (1) во время предъявления зрительного стимула; (2) в тот момент,
когда обезьяна держит в рабочей памяти информацию о месте положения зри-
тельного стимула; и (3) во время реакции перемещения взора в ту точку, в кото-
рой находился уже исчезнувший стимул или в противоположную сторону. Этот
эксперимент проиллюстрирован на рис. 4.7 .
Рис. 4 .6 . Регистрация единичных нейронов в гиппокампе. Иглы микроскопического
размера могут быть введены внутрь нейронов или с их помощью можно регистрировать
внеклеточные электрические потенциалы. Эти же электроды могут быть использованы
для стимуляции определенных клеток. Гиппокамп содержит регулярные сети нейронов,
обладающие определенными функциями. (Источник: Squire et al. , 2 0 0 3.)
Методы: визуализация живого мозга
161
Рис. 4 .7 . Рабочая память в активности единичного нейрона. Макака выполняет задачу
на рабочую память под названием отсроченный выбор по образцу (ОВПО). «Стимулом-
образцом» в данном случае является синяя точка на экране (а). Обратите внимание,
что обезьяна успешно нажимает на синюю кнопку по прошествии 20 с. Таким обра-
зом, животное может совершить ‘выбор по образцу’ даже в случае отсутствия образца,
что подразумевает сохранение его в памяти на короткий промежуток времени. Рисунок
(б) показывает растр нейронных разрядов для каждого предъявления. Каждый распо-
ложенный горизонтально набор маленьких черточек обозначает одно предъявление.
Мы можем видеть, что нейроны разряжаются быстрее во время периода запоминания
(задержки) по сравнению с предшествующим и последующим интервалами времени.
На (в) показана импульсная гистограмма нейрона, расположенного в височной доле.
Обратите внимание, что красные столбики во время периода задержки указывают на
непрерывность разряда «нейрона памяти» даже при отсутствии стимула, если он сохра-
няется в рабочей памяти. Данный нейрон имеет фоновую частоту разряда, показанную
синим цветом, равную 5–10 Гц, что является типичным для корковых нейронов. Однако
между предъявлением стимула-образца и успешным выбором по образцу, нейрон вдвое
увеличивает частоту разряда, что показано красным цветом. Считается, что такие ней-
роны вовлечены в процесс временного хранения информации в рамках рабочей памяти.
Данные нейроны обнаруживаются в височных и префронтальных областях. (Источник:
Fuster, 1997.)
162
Глава 4
Глубинные электроды также используются и на людях. Как правило, такие
электроды вживляют перед хирургической операцией тем пациентам, у кото-
рых наблюдается ничем более не излечимая эпилепсия. Вживленные электроды
помогают определить локализацию эпилептического очага при возникновении
приступа и критических для жизнедеятельности регионов мозга, которые нель-
зя повредить (рис. 4 .8). В дополнение к локализации эпилептического очага,
глубинные электроды позволяют получить доступ к областям мозга, отвечаю-
щим за обработку информации, например семантическое сходство, что показа-
но на рис. 4.9.
Как же соотносятся регистрация в одиночном нейроне и человеческое вос-
приятие? В то время как одна клетка ничего не может нам сообщить о чело-
веческом сознании, современные исследования снабжают нас удивительными
фактами, относящимися к осознанному и бессознательному зрительному вос-
приятию (рис. 4 .10).
Разряжающийся нейрон считается наиболее вероятной единицей в мозговой
активности, но существует ряд альтернативных точек зрения. Некоторые ученые
убеждены, что градуированные дендритические токи в каждом нейроне могут
участвовать в важных процессах обработки информации; некоторые склоняют-
ся к мысли о важности внутриклеточных процессов; другие выделяют неклас-
сические нервные клетки и синапсы, которые встречаются в нервной системе
значительно чаще, чем предполагалось ранее; другие считают, что глиальные
клетки тоже участвуют в процессе обработки информации; и очень многие уче-
ные считают, что о реальных процессах в мозге можно судить, только находясь
на уровне популяций нейронов. Следовательно, регистрация аксональных по-
тенциалов действия необходима, но не является единственно важным событием,
происходящим в нервной системе. Очевидно, что очень рискованно переносить
знания, полученные на единичной клетке, на более 10-миллиардную популяцию
нейронов в мозге.
Рис. 4 .8 . Глубинные электроды в мозге человека. В то время как большинство экспери-
ментов с записью активности одиночных нейронов осуществляется на животных, иссле-
дования на людях с вживлением внутримозговых электродов проводятся только по ме-
дицинским показаниям. Стрелки указывают на местоположение электродов в височной
коре. Если внимательно всмотреться в левое МРТ-изображение, то можно разглядеть
треки электродов и маленькие отверстия в затылочной части черепа, через которые они
были погружены. Нейрохирургическая операция такого типа, как правило, безопасна и
безболезненна, так как мозг не имеет болевых рецепторов. (Источник: Dietl et al., 2005.)
Методы: визуализация живого мозга
163
2.2 .
Исследования на животных и человеке
Нечеловекообразные обезьяны, такие как макаки, очень интенсивно использо-
вались в исследованиях с регистрацией одно- и мультинейронной активности.
Большая часть того, что мы знаем о зрении, памяти, внимании и высших функ-
циях, исходит из работ, проведенных на макаках (рис. 4 .11).
Рис. 4.9 . Пример электрокортикографического (ЭКоГ) исследования, в котором регистри-
ровали высокочастотные гамма-колебания (ССС — связанная с событием синхрониза-
ция), согласно текущим гипотезам участвующие в формировании мозговой основы для
связывания сенсорных и когнитивных процессов. На рисунке представлены результаты
регистрации ЭКоГ при выполнении испытуемыми лексической задачи на семантиче-
ское сходство (определить, имеют ли смысловое сходство два слова, например «нож» и
«вилка»). Точки регистрации имеют трехцветовую кодировку: белые, желтые и синие.
Синие точки указывают на активацию соответствующей области при звучании слова, но
не при зрительном демонстрировании этого слова, поэтому авторы описывают эти точки
как модально специфичные области (слуховые). Далее показано, что в этих областях не
наблюдаются изменения при семантическом сходстве слов, что продемонстрировано в
двух нижних рамках рисунка. Желтые точки указывают на активацию как при слуховом
предъявлении слов, так и при зрительном, и авторы называют такие области модально
неспецифичными. В этих участках мозга наблюдались эффекты от семантического сход-
ства слов, что можно увидеть в верхних рамках рисунка. (Источник: Crone, Sinai, and
Korzeniewska, 2006.)
164
Глава 4
Каким образом области мозга макаки можно соотнести с областями мозга
человека? В то время как существуют значительные анатомические различия,
особенно во фронтальных и теменных областях мозга, есть достаточно сильное
сходство мозга макаки и человека (рис. 4 .12).
Рис. 4 .10 . Регистрация активности одиночных нейронов у человека и сознательное вос-
приятие. Удивительный эксперимент, в котором осознаваемые и неосознаваемые сти-
мулы предъявляются одновременно в разные глаза, с применением вариации методики
проверки несовпадения изображения в глазах (см. гл . 6). В верхнем ряду (б) изображе-
ние женского лица осознается, тогда как рисунок мяча нет; в нижнем ряду (г) наоборот.
Максимальные частоты разрядов нейронов, красные в периоды, обозначенные зелены-
ми горизонтальными полосками, когда испытуемый утверждает, что видит лицо жен-
щины. Выглядит так, как будто мозг определяет, какой из стимулов будет осознан, когда
сигнал о нем достигнет определенных областей коры. Локализация электродов показана
на сканах мозга в левой части рисунка (а, в). (Источник: Rees et al., 2002.)
Рис. 4 .11 . Обезьяны и люди удивительным
образом похожи. Макаки интенсивно исполь-
зуются в исследованиях из-за наличия явных
гомологий в строении мозга (биологического
сходства) с человеком. Зрительная кора макак
является наилучшим проводником при иссле-
довании зрительной системы у человека вплоть
до настоящего времени, когда стало возможным
изучать строение зрительной коры человека бо-
лее прямыми методами. Макаки также имеют
сходство с людьми в эмоциональных процессах,
близкую связь между родителями и потомством,
и существует даже наличие префронтальных
областей, которые имеют сходство с префрон-
тальной корой человека. Разумеется, у них от-
сутствуют речь и другие видоспецифичные осо-
бенности человека. (Источник: Биологический
журнал открытого доступа PLOS.)
Методы: визуализация живого мозга
165
Регистрация одиночных нейронов у макаки не только способствует карти-
рованию сенсорных и относящихся к восприятию областей, но также может
включать и более когнитивные аспекты, например внимание.
Опыты с регистрацией одиночных нейронов обеспечили нас изобилием ин-
формации относительно кодирующих свойств нейронов коры головного моз-
га. Однако нейроны реагируют как единое целое, со сложной системой связей
клеток, имеющих разнообразное местоположение. Как же тогда мы можем за-
регистрировать подобный динамический ответ мозга? Один из ранних методов,
который позволяет получить данную информацию, представлен в следующем
разделе: электроэнцефалография.
2.3 .
Электроэнцефалография
Полномасштабная электрическая активность мозга может быть зарегистриро-
вана с черепа или от поверхности коры (блок 4.1). Вместо того чтобы улавли-
вать электрическую активность непосредственно от нейронов, которые можно
представить в виде крохотных электрических батареек, электроэнцефалограм-
Рис. 4 .12 . Гомологии в строении мозга у человека и макак. На верхнем рисунке изо-
бражен мозг человека, на нижнем — макаки . Желтым цветом отмечены области, ответ-
ственные за зрительное распознавание объектов у обоих видов. Изображения справа де-
монстрируют вид на правое полушарие снизу, при этом фронтальные отделы обращены
на рисунке наверх. Вид с вентральной стороны демонстрирует нижнюю поверхность
правого полушария. (Источник: Squire et al., 2003.)
166
Глава 4
Блок 4.1 . ЭКоГ: прямое измерение активности мозга
Даже при быстром развитии технологий нейровизуализации наиболее прямые дан-
ные о работе живого мозга до сих пор получают во время внутричерепной регистра-
ции электрической активности (рис. 4 .13). Одной из причин является то, что электри-
ческий сигнал, измеренный непосредственно от тканей мозга, значительно сильнее,
чем при регистрации от поверхности головы (порядок сигнала меняется от сотен
милливольтов до микровольтов соответственно). Также ЭЭГ с поверхности головы
изменяется благодаря прохождению сигнала через водосодержащие ткани мозга,
кожу и мышцы, и сама электрическая активность оказывает влияние на сигнал. На-
пример, когда вы хмуритесь, мышцы над вашими глазами сокращаются, в это же
время тонкий мышечный слой черепа растягивается и приспосабливается. Даже дви-
жения глаз оказывают значительное влияние на ЭЭГ, записанную с головы. Таким
образом, поверхностная ЭЭГ несет в себе электрическую активность различного ге-
неза, и к тому же она профильтрована многими слоями тканей. ЭКоГ (электрокорти-
кография) имеет улучшенное соотношение сигнал–шум (рис. 4 .14). Самый большой
недостаток метода заключается в необходимости хирургической операции. В связи
с этим на людях он применим только при явных медицинских показаниях. Но не-
смотря на это, большая часть базовых знаний о работе мозга получены в результате
многочисленных исследований на животных с помощью метода ЭКоГ или глубин-
ных электродов.
Рис. 4 .13 . (а) Хирургически об-
наженная поверхность коры с
расположенной непосредствен-
но на ней сеткой электродов
8 × 8. Электроды (белые кружки)
вмонтированы в прозрачную ква-
дратную пластиковую пластину,
размещенную над левым полу-
шарием. Обратите внимание, что
пластина покрывает и височную
долю, которая является частым
источником судорожной актив-
ности. Также считывается инфор-
мация и с воспринимающей рече-
вой области (зона Вернике), и от
зоны Брока (левая нижне-лобная
извилина), которая отвечает за
производство речи. Левое полу-
шарие является для большинства
людей доминантным для речевой
функции, поэтому электроды рас-
полагают над наиболее критиче-
скими областями. (б) Рентгенов-
ский снимок того же пациента,
совмещенный со структурной
МРТ мозга. (Источник: Dalal et
al., 2008.)
Методы: визуализация живого мозга
167
Уайлдер Пенфилд с коллегами в 1950-х гг. были первопроходцами в области электро-
кортикографии у людей. У больных эпилепсией с неконтролируемыми приступами
после хирургического удаления очагов возникновения приступа испытывали облег-
чение своего состояния. ЭКоГ может указать на месторасположения такого «эпилеп-
тогенного очага». В дополнение оперирующий хирург должен знать, повреждения
каких областей мозга жизненно необходимы для нормального восприятия и речевой
функции, во время операции ему следует избегать. Исследования с ЭКоГ достаточно
безопасны и фактически предшествуют операции. Зондирование корковой поверх-
ности в основном безболезненная процедура, так как там отсутствуют болевые ре-
цепторы. Научная польза от данной работы чрезвычайно большая (см. гл . 8). В то
время как исследования на животных дали основу для прямой регистрации актив-
ности мозга, работы с регистрацией ЭКоГ на бодрствующих людях открыли нейрон-
ную основу речи, сознательного восприятия и произвольного управления.
Рис. 4 .14 . Результаты ЭКоГ исследования пациента с эпилепсией, наложенные на
МРТ изображение латеральной поверхности мозга. В трех маленьких рамках на
рисунке показано быстрое увеличение гамма-активности (80–100 Гц) сразу после
предъявления изображения предмета, который необходимо назвать. Пациента просят
называть предметы на рисунках, при этом усредняют электрическую активность по
всем предъявлениям. Цветные полоски указывают, какие пары электродов использо-
вались для корковой стимуляции, которая обычно препятствует функционированию
подлежащей корковой области. Красные и фиолетовые полоски отмечают области,
в которых электрическая стимуляция мешала называнию предметов с картинок, дви-
жению рта и другим речевым процессам. Зелеными полосками отмечены области,
стимуляция которых не повлияла на выполнение задания. (Источник: Crone, Sinai
and Korzeniewska, 2006.)
168
Глава 4
ма регистрирует целое электрическое поле. В результате получается запись, ко-
торую называют электроэнцефалограммой (ЭЭГ), или записью электрической
активности мозга. ЭЭГ была открыта в 1929 г. Хансом Бергером (рис. 4 .15). Так
как электромагнитные волны распространяются фактически мгновенно, метод
ЭЭГ имеет очень высокое временное разрешение. Однако ЭЭГ достаточно се-
лективно, так как в большей степени воспринимает активность нейронов, нахо-
дящихся на поверхности, по сравнению с глубокорасположенными нервными
клетками. ЭЭГ регистрируется через множество слоев влажной ткани, поэтому
исходная электрическая активность ослабляется и искажается формой и прово-
дящими свойствами клеток, находящихся на пути сигнала. Некоторые иссле-
дователи считают, что ЭЭГ в большей степени отражает активность первого
слоя коры, который в основном состоит из сплетения нервных волокон, тесно
переплетенных дендритов корковых нейронов (Freeman, 2004).
Легче всего регистрировать ЭЭГ от поверхности кожи головы, однако ино-
гда электрическую активность можно записать от электродов, расположенных
прямо на коре головного мозга. Так же как и при регистрации одиночных кле-
ток, ЭЭГ является в определенной степени прямым методом измерения элек-
трической активности мозга. Но когда десятки миллиардов корковых нейронов
разряжаются с частотой 10 Гц, получается несколько триллионов событий в
секунду. Именно поэтому необработанные записи ЭЭГ очень трудно понимать,
и до разработки мощного компьютерного анализа их было невероятно сложно
интерпретировать.
Однако если записи ЭЭГ усреднить по результатам нескольких эксперимен-
тов и привязать к определенной точке отсчета, например моменту подачи сти-
мула, такая усредненная электрическая активность будет состоять из изящных
и четких волн. Эти потенциалы, связанные с событиями (ПСС), чувствительны
к активности большой нейронной популяции, которая характеризует зритель-
ные, слуховые и даже семантические процессы.
2.3.1 . Исследование нейронных популяций
Активность крупных популяций клеток и их связей составляет следующий немало
важный уровень исследования (Freeman, 2004; �ohn, 2004). Спонтанная ЭЭГ де-
Freeman, 2004; �ohn, 2004). Спонтанная ЭЭГ де-
, 2004; �ohn, 2004). Спонтанная ЭЭГ де-
�ohn, 2004). Спонтанная ЭЭГ де-
, 2004). Спонтанная ЭЭГ де-
монстрирует разные паттерны активации. Мозг может работать с множествами
разных уровней интерактивности. Например, во время глубокого сна в нативной
записи ЭЭГ преобладают высокие медленные волны. Это свидетельствует о том,
что значительные группы нейронов синхронизированы в большой области голов-
ного мозга. Когда человек просыпается, такая медленная активность замещается
маленькими, частыми электрическими колебаниями, указывающими на быстрые
и пластичные механизмы взаимодействия информационных образов. В настоящее
Рис. 4 .15. Первая запись ЭЭГ. Этот исторически важный ЭЭГ сигнал был зарегистри-
рован Гансом Бергером в 1929 г. при помощи одной пары электродов, помещенных
на поверхность головы. Верхняя запись ЭЭГ была записана с головы сына Бергера и
демонстрирует альфа-активность (регулярная синусоидальная волна на верхней запи-
си). Нижняя запись является электрической синусоидой масштаба времени с частотой
в 10 Гц. (Источник: Gottesmann, 1999.)
Методы: визуализация живого мозга
169
время считается, что частота разряда нейронов во время сна и бодрствования не
различается, но тем не менее при бодрствовании ЭЭГ демонстрирует увеличение
информационных процессов, происходящих в мозге. Таким образом, сон не явля-
ется пассивной стадией, а лишь другим вариантом работы мозга, имеющим свои
особые функции. К ним, например, относят консолидацию в памяти событий, про-
изошедших во время бодрствования (Hobson and Stickgold, 1995).
Стандартным методом для обработки комплексной ЭЭГ является Фурье-ана -
лиз, названный так в честь французского математика Пьера Фурье, который пока-
зал, что любой сложный сигнал может быть разложен на синусоидальные волны с
различной мощностью (плотностью распределения) на каждой частоте (рис. 4.16).
Нативная (необработанная) запись ЭЭГ имеет различные, зрительно различи-
мые волны, которые подразделяют по частоте колебания на альфа-, бета-, гамма-,
тета- и дельта-диапазоны. Однако исходная запись ЭЭГ на самом деле является
комбинацией нескольких видов перечисленных активностей. Используя матема-
тический анализ, мы можем разложить комплексную и зашумленную запись на
частотные составляющие. Это весьма напоминает процесс разложения шумных
радиопомех по диапазонам их частот, что, собственно, мы и делаем, когда настраи-
Рис. 4 .16. Регулярные ритмы в разных областях мозга. Анализ Фурье — метод, позво-
ляющий нам разлагать на составные части плотность (или мощность) регулярных волн,
которые не видны в шумных записях ЭЭГ (см. табл. 4 .1). Графики демонстрируют ре-
зультирующие кривые мощности. Согласно данному источнику, наибольшая мощность
альфа-активности сосредоточена в затылочной коре, тогда как наибольшая мощность
тета-ритма наблюдается в лобной коре. Считается, что тета-активность отражает фрон-
тально-гиппокампальные взаимодействия во время извлечения информации из долго-
временной памяти. Гамма-активность распространена повсеместно в мозге и отражает
функциональные взаимодействия между различными регионами мозга во время ак-
тивного бодрствования. Цвета отражают различные частотные диапазоны. (Источник:
Zoran �osipovich, с разрешения.)
170
Глава 4
ваем для прослушивания определенную радиостанцию. Обработанная таким обра-
зом ЭЭГ может демонстрировать, что определенные задачи могут сопровождаться
появлением специфического ритма, например альфа- или гамма -, в определенных
областях мозга. Это проиллюстрировано на рис. 4 .16 и 4.17. Типичные частотные
ритмы представлены в табл. 4 .1 .
Классические ЭЭГ ритмы можно выявить и в исходной или необработанной за-
писи. Необходимо заметить, что в бодрствующем сознательном состоянии в ЭЭГ
широко распространена низкоамплитудная бета- и гамма-активности. Обратная
картина наблюдается в состоянии глубокого сна, когда в ЭЭГ преобладают медлен-
ные, высокоамплитудные и в большей степени синхронизированные волны. Во вре-
мя других бессознательных состояниях также отмечают некоторое сходство (Baars
et al., 2004). Альфа-активность регистрируется в затылочных областях головного
мозга в расслабленном, но бодрствующем состоянии.
Рис. 4 .17. Регулярные волнообразные колебания в разных областях мозга. Одним из
трех важных ритмов, отражающих важные когнитивные функции, является альфа-ак-
тивность, которую классически ассоциируют с расслабленными состояниями и состоя-
ниями готовности и, как правило, регистрируют в затылочных отведениях. В настоящее
время считается, что тета-активность в лобных областях мозга отражает взаимодей-
ствия между гиппокампом и корой. И наконец, гамма-активность, участвующая в со-
знательном восприятии и других когнитивных процессах, отражает работу временных
крупномасштабных нейронных объединений. «Спектральная мощность» — это об-
ласть, расположенная под кривой на частотной диаграмме и отражающая физическую
энергию каждой частотной полосы. (Источник: Ward, 2002.)
Методы: визуализация живого мозга
171
Таблица 4.1
Частотные диапазоны ЭЭГ и функции, которые с ними ассоциированы
Название
и пример
Описание
Дельта
Дельта-ритм — медленноволновая активность, характерная для глу-
бокого, бессознательного сна. Частота ее меньше 4 Гц, и похожие
частотные составляющие ЭЭГ присутствуют при эпилептических
припадках и потери сознания, а также в некоторых коматозных состо-
яниях. Именно поэтому считается, что дельта-ритм отражает актив-
ность мозга человека, находящегося в бессознательном состоянии.
Активность дельта-диапазона имеет тенденцию быть самой высо-
коамплитудной и низкочастотной активностью мозга. Дельта-вол-
ны усиливаются одновременно с уменьшением осознания происхо-
дящего в окружающей действительности.
Тета
Тета-активность имеет частоту 3,5–7,5 Гц.
Предположительно тета-волны возникают в результате синхронной
активности большого числа нейронов. Тета-ритм наблюдается во
время некоторых стадий сна и в состояниях спокойной концентра-
ции, например медитации. Также отмечено его появление при вы-
полнении некоторых задач на кратковременную память и во время
извлечения информации из памяти.
Тета-волны, вероятно, участвуют во взаимодействии гиппокампа и
коры в процессах запоминания и извлечения памятного следа.
Альфа
Частота альфа-волн составляет 7,5–13 Гц и возникает в результа-
те синхронной и когерентной (по фазе) электрической активности
большой группы нейронов в головном мозге человека. Их также на-
зывают ритмом Бергера в память об открывателе ЭЭГ.
Альфа-активность преимущественно возникает в затылочных об-
ластях мозга во время спокойного бодрствования с закрытыми гла-
зами. И наоборот, альфа-активность подавляется при открывании
глаз, а также при дремотных состояниях и сне.
Бета
Бета-ритм — «быстрая» нерегулярная низкоамплитудная электри-
ческая активность мозга (12–25 Гц).
Бета-волны, как правило, ассоциируют с нормальным сознатель-
ным бодрствованием, часто активным, работающим состоянием
или усиленным мышлением и активной концентрацией внимания.
Ритмическая бета-активность с доминированием определенной ча-
стоты может свидетельствовать о различных патологиях и наркоти-
ческих эффектах.
Бета-ритм обычно регистрируется в обоих полушариях, симме-
трично распространен и наиболее явно выделяется во фронтальных
отделах головного мозга. Он может отсутствовать или его амплиту-
да может быть снижена в поврежденных корковых областях.
Гамма
Гамма-активность в целом имеет частотный диапазон 26–70 Гц
с пиком мощности около 40 Гц.
Считается, что гамма-волны отражают процессы активного обмена
информацией между корковыми и подкорковыми областями. Они
регистрируются в состоянии активного бодрствования и во время
БДГ-сна (сна с быстрыми движениями глаз). Обратите внимание,
что гамма- и бета-активности могут перекрываться.
172
Глава 4
Как было уже упомянуто, временная разрешающая способность ЭЭГ имеет
порядок миллисекунд, однако его пространственная разрешающая способность
довольно мала. Достаточно сложно локализовать электрический источник ЭЭГ
сигнала. Данная задача немного упрощается при увеличении числа электродов и
использовании сложных аналитических методов. Однако, несмотря на это, ЭЭГ
дает нам очень мало информации о мозговых структурах, расположенных глубоко
под корой головного мозга. А так как такие подкорковые структуры, как таламус
чрезвычайно важны, у ЭЭГ существуют некие ограничения.
Исходно ЭЭГ использовали для определения мозговых ответов во время сна
и дремотного состояния (рис. 4 .15). Такое новое усовершенствование, как вы-
сокоплотное размещение электродов, позволяет ученым более детально изме-
рять деятельность мозга.
Важным моментов регистрации ЭЭГ является измерение вызванных потен-
циалов (ВП), которые возникают в ответ на предъявление специфического сти-
мула. Для улучшения соотношения сигнала к шуму эти ответы усредняются по
многим пробам.
Рис. 4 .18 . Вызванные потенциалы часто изображают на топографических картах с ус-
реднением напряженности по поверхности кожи головы. Красные цвета отображают
более высокое напряжение. В этом случае эксплицитное извлечение сознательной ин-
формации из памяти вызывает более выраженные ПСС, чем «ощущение знания» для
того же воспоминания. Активность мозга не может быть четко локализована при по-
мощи ППС, по сравнению с результатами ПЭТ и фМРТ. (Источник: Düzel et al., 1997.)
Методы: визуализация живого мозга
173
Например, зрительные ВП регистрируют для исследования динамики обра-
ботки зрительного стимула (рис. 4 .18). ВП зависят от многих факторов, имею-
щих большое значение для когнитивной деятельности человека, такой как спо-
собность слышать свое имя или восприятие музыки (рис. 4 .19). При помощи
ЭЭГ можно выявить определенные паттерны, характерные для стадий сна и
бодрствования, патологические паттерны заболеваний, например эпилепсии, и
даже выявить область мозга, которые активируются в ответ на прослушивание
музыки. Современная технология, магнитоэнцефалография (МЭГ), имеет боль-
шое сходство с ЭЭГ и позволяет по-новому визуализировать процессы, проис-
ходящие в мозге.
2.4 .
Магнитоэнцефалография
Магнитоэнцефалография (МЭГ) позволяет измерить магнитное поле, которое
возникает в результате электрической активности в мозге (рис. 4 .20). Простран-
ственное разрешение этого метода для некоторых областей коры головного
мозга в настоящее время достигает нескольких миллиметров, в то время как
его временное разрешение составляет миллисекунды (рис. 4 .21).
Из-за физических свойств магнитного поля МЭГ высокочувствительна к
дендритным входам, которые расположены под прямыми углами к поверхно-
стям извилин (складкам коры), и намного меньше зависит от нижележащих
структур. МЭГ обладает великолепным временным разрешением, а ее про-
странственная разрешающая способность лучше, чем у ЭЭГ.
Как и при использовании других методов измерения активности мозга, ре-
зультаты МЭГ должны быть совмещены со структурным изображением живого
мозга. В МЭГ используется процесс определения источника магнитных полей
(MSI, от англ. magnetic source imaging), позволяющий при регистрации со-
вмещать источники магнитной активности мозга со снимками анатомических
структур, полученных при помощи МРТ. Таким образом, метод определения
источника магнитных полей имеет высокую пространственную разрешающую
способность МРТ и высокое временное разрешение МЭГ. Его используют пе-
ред операциями на головном мозге для точного определения жизненно важных
областей мозга, которые нельзя затрагивать во время хирургического вмеша-
тельства.
Рис. 4 .19. Вызванные потенциалы на музыку. Регулярная музыкальная последователь-
ность воспринимается как более приятная и демонстрирует более выраженные вызван-
ные потенциалы (см. стрелку на рис. (в)). Нерегулярная музыкальная последователь-
ность вызывает совершенно другой ответ. (Источник: Koelsch and Siebel, 2005.)
174
Глава 4
Преимуществом МЭГ служит ее полная беззвучность и отсутствие необ-
ходимости в любых повреждающих воздействиях при использовании. Как мы
убедимся далее, МРТ достаточно шумный метод, а использование глубинных
электродов, разумеется, требует проведения хирургической операции. Это де-
лает МЭГ очень заманчивым методом для использования при работе с детьми
и чувствительными людьми.
2.5 .
Вмешательство в работу мозга
Мы обсудили технологии, которые позволяют без повреждающего воздействия
регистрировать мозговые сигналы, а что, если бы было возможно каким-нибудь
безопасным способом вызывать нейронную активность? Такой метод был бы
особенно полезным при изучении взаимосвязи между вызванной нейронной
активностью и когнитивной деятельностью.
Ранние работы по прямой электрической стимуляции головного мозга были
начаты Уайлдером Пенфилдом, нейрохирургом из Монреальского неврологи-
ческого института (рис. 4 .22 и 4.23). Пенфилд и его коллеги занимались ле-
чением людей, страдающих некупируемой эпилепсией. Во время операции
Рис. 4 .20 . МЭГ бесшумна и неинвазивна. МЭГ-исследования легко переносятся ма-
ленькими детьми, если они могут некоторое время находиться в малоподвижном со-
стоянии. Картинки внизу рисунка демонстрируют векторы магнитных полей на поверх-
ности головы объекта. (Источник: 4D Neuroimaging, Сан Диего.)
Методы: визуализация живого мозга
175
Рис. 4 .21 . Пример применения вейвлет-анализа для данных МЭГ при предъявлении ре-
чевых и неречевых звуков. Неречевые звуки состоят из стационарных тонов, тональный
контур (ТК) которых включает динамические изменения; речевые же звуки включают
стационарные гласные и согласные звуки в динамическом состоянии. Используя вейв-
лет-анализ и откладывая по оси х время, а по оси y — частоту и кодируя уровень актив-
ности цветом, авторы исследовали нейронную активность на различающиеся стимулы и
задания (АУД, испытуемым необходимо определить, являются ли предъявленные звуки
идентичными; КАТ, испытуемым надо определить, принадлежат ли два звука одной ка-
тегории). Результаты: высокий уровень активности в альфа-диапазоне (8–13 Гц) показан
красным цветом на вейвлет-диаграммах.
Рис. 4 .22 . Уайлдер Пенфилд (Wilder Penfield). Пен-
филд с коллегами разработали в 1950-х гг. нейрохирур-
гическую операцию на открытом мозге для неподда-
ющейся лечению эпилепсии. (Источник: Adelman and
Smith, 2004.)
176
Глава 4
на открытом мозге пациенты оставались в бодрствующем состоянии и были
способны реагировать на происходящие события, так как при данном хирур-
гическом вмешательстве применяется только локальная анестезия. В самом
мозге отсутствуют болевые рецепторы, поэтому кору головного мозга можно
стимулировать и проводить на ней операции без общего наркоза. Это жизненно
необходимое условие операции, потому что хирургам требуется картировать
функционально важные области мозга. И очаг эпилептической активности,
вызывающий генерализованные судороги, таким образом может быть удален
с минимальными побочными эффектами.
2.5.1 . Безопасный способ вмешательства в работу мозга:
транскраниальная магнитная стимуляция (ТМС)
В настоящее время существует возможность моделировать повреждения мозга
на здоровых объектах. Без оперативного вмешательства в мозг испытуемого мы
можем очень локально изменять уровень мозговой активности. Кратковремен-
ные магнитные импульсы, приложенные к черепу, могут либо подавить, либо
возбудить активность в маленькой области коры. Например, если мы будем сти-
мулировать представительство руки в двигательной области коры, рука испыту-
емого неожиданно будет двигаться и поворачиваться. Прикладывание тормоз-
ных импульсов к этой же области вызовет у испытуемого трудности в движении
рукой. Этот метод носит название транскраниальной магнитной стимуляции
(ТМС) или, как называет этот метод его ведущий разработчик, «вмешательство
в работу мозга» (Cowe� and Walsh, 2001). Метод ТМС (рис. 4.26) кажется впол-
Cowe� and Walsh, 2001). Метод ТМС (рис. 4.26) кажется впол-
and Walsh, 2001). Метод ТМС (рис. 4.26) кажется впол-
and Walsh, 2001). Метод ТМС (рис. 4.26) кажется впол-
Walsh, 2001). Метод ТМС (рис. 4.26) кажется впол-
Walsh, 2001). Метод ТМС (рис. 4.26) кажется впол-
, 2001). Метод ТМС (рис. 4.26) кажется впол-
не безопасным. Благодаря применению ТМС мы теперь можем проверять ги-
потезы о вкладе определенных областей головного мозга в высшие когнитив-
ные процессы. А так как ТМС работает с миллисекундной шкалой времени,
становится возможным изучать, как разворачиваются процессы переработки
информации.
Ряд авторов (Overgaard et al., 2004) использовали ТМС для исследования
зрительного восприятия. Испытуемому предъявляли простые геометриче-
ские фигуры: треугольники, квадраты и круги — трех разных цветов (крас-
ные, зеленые и синие). Зрительные стимулы предъявляли в одном из трех
возможных местоположений на экране. Время экспозиции фигур было до-
статочное для восприятия изображений всеми испытуемыми. Исследователи
прикладывали при помощи ТМС тормозный импульс к средневисочной обла-
сти головного мозга обоих полушарий, что приблизительно соответствует на
черепе точке немного фронтальнее уха. Если импульс подавали через 120 мс
Рис. 4 .23 . Принадлежащая Пенфилду
карта областей мозга, электрическая сти-
муляция которых вызывает нарушения
речи. Пенфилд и Робертс обнаружили,
что электрическое раздражение указан-
ных областей нарушает воспроизведение
или понимание речи. Обратите внимание,
насколько точно соотносятся эти области
мозга, с классическими областями Брока и
Вернике, которые были открыты веком ра-
нее. (Источник: Adelman and Smith, 2004.)
Методы: визуализация живого мозга
177
после предъявления стимула, то у испытуемых наблюдалось значительное
снижение восприятия стимула. Люди не могли четко воспринимать изобра-
женные фигуры, вместо этого у них было только смутное ощущение о том,
было что-то показано или нет.
Интерес вызывает тот факт, что, если испытуемых просили угадать предъ-
являемые фигуры, все продемонстрировали нормальные результаты. Однако
никто не давал отчета о наличии стимула. Несмотря на это, восприятие стиму-
ла происходило (бессознательно), что подтверждается точным обнаружением.
Все же о субъективном восприятии слов ничего не было сообщено.
А теперь давайте обратимся к методам изучения мозга, которые используют
технологии пространственного картирования для одновременного исследова-
ния анатомии и функций мозга.
Беседа с Майклом Д. Раггом: Корреляты сознания,
выявляемые с помощью нейровизуализации
Развивающееся направление когнитивной нейронауки предоставляет нам потрясаю-
щие новые и постоянно меняющиеся способы исследования нейронных коррелятов
человеческого мышления и когнитивной деятельности (рис. 4 .25). Ведущим лиде-
ром в этой области является доктор Майкл Рагг, профессор кафедры нейробиологии
и поведения, а также директор центра нейробиологии обучения и памяти в универ-
ситете Калифорнии в Ирвине. Мы побеседовали с доктором Раггом и узнали о том,
как его подход к изучению нейронных коррелятов памяти у человека изменился за
последнее десятилетие и в каком направлении, по его мнению, он изменится в буду-
щем. Ниже приведены основные моменты этого интервью. (МПР — мозг, познание
разум: редакторы Бернард Дж. Баарс и Николь М. Гейдж; МР: Майкл Рагг).
МПР: Давайте начнем с истории: как, будучи когнитивным нейробиологом, Вы из-
менили ваш подход к исследованиям за последние десять лет?
МР: Для меня, как мне кажется, он совпадает с возможностями методов нейрови-
зуализации, которые позволяют напрямую идентифицировать нейронные популя-
ции и нейронные цепи, возникающие во время разных когнитивных задач. Когда я
впервые стал работать в направлении, которое сейчас носит название когнитивной
нейронауки, оно развивалось в основном людьми, стремящимися с использованием
методов измерения активности мозга разделить когнитивные процессы. В то время
единственным доступным методом было измерение ЭЭГ, поэтому был достаточно
большой уклон в сторону простого использования полученных измерений нейрон-
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
Рис. 4 .24 . Майкл Д. Рагг (Michael D.
Rugg, PhD), кафедра нейробиологии
и поведения и кафедра когнитивной
нейронауки, университет Калифор-
нии, Ирвин
178
Глава 4
Методы: визуализация живого мозга
179
ной активности для выделения различных когнитивных процессов. Логика такова:
если у вас есть два экспериментальных условия, которые вызывают сильно раз-
личающуюся активность мозга, имеющую к тому же разную мозговую основу, то
это убедительное доказательство при определенных допущениях того, что вы раз-
делили две различные когнитивные деятельности.
И после появления сначала ПЭТ, затем фМРТ стало возможным рассмотреть все
это логически, но сейчас есть гораздо больше информации о том, где в головном
мозге расположены участки, вызывающие разные паттерны активации. И поэтому
ученые больше начинают размышлять о роли разных областей мозга в том или
ином гипотетическом когнитивном процессе: вместо того чтобы использовать
данные об активности мозга как (более или менее подходящую) замену или допол-
нение к времени реакции или количеству ошибок или другим поведенческим из-
мерениям, эти данные сейчас стали действительно самостоятельным источником
информации. Вы сейчас можете спросить: почему некоторые варианты информа-
ционно-поисковых операций вовлекают разные части теменной коры, ответствен-
ные за другие процессы? Какую роль выполняет теменная кора, что она оказыва-
ется задействованной только в определенных процессах, и какую теоретическую
информацию это дает нам о строении нейронных сетей, вовлеченных в различные
когнитивные процессы?
МПР: Исследование таких сложных нервных процессов, как память, ставит перед
ученым сложную задачу разработки экспериментов, которые позволили бы разо-
брать на еле уловимые, но очень важные составляющие сложную систему. Каков
ваш подход к этой проблеме?
МР: Мне кажется, одно стало четко понятно: нельзя изучать память или нейрон-
ные корреляты памяти без логической, четко сформулированной предэксперимен-
тальной теории об организации памяти и без представления, как ваш эксперимент
повлияет на разные процессы памяти. В других областях исследования когни-
тивной деятельности все примерно так же, но при изучении памяти становится
очевидно, что нельзя засунуть испытуемого в сканнер и сказать: «Я буду изучать
Рис. 4 .25. Пример экспериментальной схемы, использовавшейся в фМРТ-
экспериментах доктора Рагга по изучению памяти. В этих экспериментах иссле-
довали роль внимания к цвету и местоположению объектов во время задачи на
память. Верхний рисунок: экспериментальная парадигма. Слева: материалы для
запоминания с применением цветовых подсказок и подсказок местоположения во
время сканирования испытуемых в обучающей фазе эксперимента. Справа: мате-
риалы для запоминания, использовавшиеся в тестовой фазе, когда были убраны
указатели цвета и местоположения. Средний рисунок: эффекты на память, вытека-
ющие из свойство-специфичности. Сл ева: эффекты на память специфичные к цве-
ту (отмечены желтым) показаны на стандартизованном изображении мозга. Часть
этих эффектов дополнительно модулировалась вниманием к цвету (наложение зе-
леного). Справа: эффекты на память специфичные к местоположению (отмечены
желтым), а также показаны области мозга, модулируемые вниманием к местополо-
жению (зеленые). Нижний рисунок: средняя височная доля кодирует воспринятую
контекстную информацию. Слева: область правого гиппокампа демонстрирует эф-
фекты памяти на контекстную информацию, которая была селективно воспринята
(независимо от свойства), и отсутствие значимых эффектов в ответ на невосприня-
тую информацию. Справа: на графиках указаны различия в опытах с восприятием
и не восприятием информации, а также графики с разделением на цвет и местопо-
ложение. (Источник: Uncapher and Rugg, 2009.)
180
Глава 4
память». Необходимо иметь четко сформулированную логическую когнитивную
или функциональную теорию памяти, которая бы направляла ваши эксперименты и
помогала интерпретации. Один из самых значительных споров в области когнитив-
ной нейронауки памяти — вовсе не спор о данных, так как с полученными данными
соглашаются все. Дело в интерпретации этих данных, и интерпретации дают люди,
имеющие разные предэкспериментальные положения о строении памяти, о методах
исследования, и каковы должны быть критерии для разделения разных процессов
памяти с точки зрения поведения. Несколько лабораторий могут иметь одинаковый
набор экспериментальных данных, но интерпретация их будет значительно разли-
чаться: таким образом, разные интерпретации основаны не на нейронауке, а на экс-
периментальной психологии.
МПР: Давайте заглянем теперь в будущее. Какие вы видите изменения в подходах
по изучению когнитивной нейронауки, а именно человеческой памяти?
МР: Мне кажется, все идет к усложнению поведенческих задач, а также к более
глубокому осмыслению теоретических посылок различия обнаруженных данных и
попыткам рассмотреть их с нескольких точек зрения. Еще стало совершенно оче-
видно, и это исходит с методологической стороны, что более не полезно пытаться
охарактеризовать активность, связанную с памятью, в таких обширным терминах,
как «гиппокамп» и «не гиппокамп», «энторинальная кора» и «парагиппокампаль-
ная кора» и другие. Причина этого высокая дифференциация этих структур и, ве-
роятно, было бы огромным упрощением думать, что если две задачи затрагивают,
например, гиппокамп, они должны в некотором роде иметь какой-то общий когни-
тивный процесс низкого уровня. Сейчас у нас есть возможность с применением
МРТ получать с высоким разрешением функциональную нейровизуализацию гип-
покампа на уровне паттернов активности гиппокампальных полей. Например, есть
люди, которые утверждают, что разделяют активность зубчатой извилины и поля
СА1 против СА3. Можно ли утверждать, что при этом вовлекается гиппокамп и,
следовательно, выполняется одна и та же процедура? Другими словами, если мож-
но так выразиться, уровень разрешающей способности для разделения нейронных
коррелятов когнитивной деятельности все более повышается. Именно это изменит
представление людей о «нейронных нарушениях взаимосвязи» и «когнитивных
разобщениях».
3.0 .
фМРТ И ПЭТ: непрямое измерение
нейронной активности
При помощи ЭЭГ и МЭГ активность мозга измеряют почти напрямую. Другие
методы нейровизуализации используют непрямое измерение активности мозга,
например кровоток или локальный уровень кислорода. В настоящее время наи-
более широко используемым методом является фМРТ (функциональная маг-
нитно-резонансная томография) (рис. 4 .27), и в особенности тот ее вид, кото-
рый позволяет измерять уровень кислорода в локальном кровотоке, — BOLD
(от англ. blood-oxygen level dependent activity), рис. 4 .28 и 4.29.
Когда нейроны разряжаются, они поглощают кислород и глюкозу и выде-
ляют метаболические продукты обмена. Активная область мозга использует
запас кислорода в локальном кровотоке, и по мере того, как кислород погло-
щается, мы регистрируем маленькое уменьшение сигнала BOLD. В течение
долей секунды данное снижение локального уровня кислорода запускает при-
ток богатой кислородом крови в эту область. В этот момент мы регистрируем
Методы: визуализация живого мозга
181
Рис. 4 .26 . Магнитная стимуляция мозга осуществляется без хирургического вмеша-
тельства. Стимуляция мозга может быть проведена без операции. В данном проил-
люстрированном примере ее применяют относительно зон Брока и Вернике в левом
полушарии головного мозга испытуемого. ТМС является безопасной процедурой при
средних значениях интенсивности воздействия и частоты. Она позволяет проверять
причинно-следственные гипотезы в экспериментах на мозге, что является значитель-
ным методологическим преимуществом. (Источник: Andoh et al., 2006.)
восстановление сигнала. Однако компенсаторный механизм зашкаливает,
вызывая гораздо больший прилив кислорода, чем требуется, поэтому мы на-
блюдаем увеличение сигнала выше начального уровня. И наконец, когда неис-
пользованный кислород вымывается из исследуемой области, мы регистрируем
возвращение сигнала BOLD к начальному уровню.
182
Глава 4
Таким образом, по мере того как меняется BOLD-сигнал вслед за измене-
нием уровня кислорода в кровотоке, мы можем непрямым способом регистри-
ровать активацию нервной ткани. Время возникновения сигнала BOLD равно
примерно 6 с после предъявления стимула. Взаимосвязь нейронной активации
и BOLD фМРТ продемонстрирована на рис. 4 .28 и 4.29.
Позитронно-эмиссионная томография (ПЭТ) была разработана намного
раньше, чем МРТ и фМРТ, и обеспечивает измерение метаболической активно-
сти мозга (рис. 4 .30). ПЭТ в настоящее время применяют намного реже, так как
данная методика является очень дорогостоящей и требует наличия циклотро-
на. Так же при использовании этого метода испытуемому необходимо вводить
Рис. 4 .28 . Основа функциональной
МРТ (фМРТ). Одним из самых попу-
лярных методов визуализации работы
мозга, вероятнее всего, является фМРТ,
которая менее дорогостояща по срав-
нению с ПЭТ и обеспечивает хорошую
локализацию активности мозга. фМРТ
является методом непрямой регистра-
ции локальной нейронной активности,
что и показано на рисунке. Повышение
активности нейронов ведет к увеличе-
нию потребления кислорода нейронами
и окружающими клетками, что приво-
дит к дополнительному притоку крови,
несущей кислород, к этому региону. Это
можно измерить при помощи методики
BOLD — измерения активности, зави-
сящей от уровня кислорода в крови. Так
как нейроны начинают разряжаться не-
сколькими секундами ранее, чем возрас-
тет активность BOLD, существует вре-
менная задержка у фМРТ. (Источник:
Dogil et al., 2002.)
Рис. 4 .27. Вид установки для МРТ. Большин-
ство фМРТ регистрируют у лежачего испы-
туемого. Даже в настоящее время МРТ созда-
ет достаточно большое количество шума при
включении и выключении электромагнитной
катушки. Небольшой экран может применяться
для предъявления зрительных стимулов или на-
ушники для предъявления слуховых стимулов.
Из-за того что устройство генерирует мощное
магнитное поле, в экспериментальную комнату
нельзя вносить никакие металлические предме-
ты, например шариковые ручки и даже скрепки.
(Источник: Sharma and Sharma, 2004.)
Методы: визуализация живого мозга
183
радиоактивную метку. В области немедицинского исследования МРТ и фМРТ
применяются намного шире (рис. 4 .31). Несмотря на это, ПЭТ все равно имеет
большое значение, так как разные метки могут быть присоединены к разным
молекулам. Таким образом, может быть определен нейрохимический состав ре-
цепторов.
На сегодняшний момент невозможно одновременно иметь и высокую про-
странственную разрешающую способность, и высокое временное разрешение
при использовании только одного метода. Существует компромисс. Такие ме-
тоды, как фМРТ и ПЭТ, указывают нам, где в мозге происходят определенные
процессы. ЭЭГ, МЭГ и регистрация одиночных нейронов показывают милли-
секундные изменения активности. В любом исследовании встает важный во-
прос, почему авторы использовали именно этот метод и что они с помощью
его наблюдали — или пропустили. В идеале необходимо использовать разные
методы, которые показывают перекрывающиеся результаты.
Рис. 4 .29 . Обычный BOLD-ответ в нейронной популяции. Верхняя линия демонстри-
рует вспышку разрядов группы нейронов в какой-то области мозга. Через несколько
секунд активный участок мозга полностью израсходует свой непосредственный запас
питательных веществ —– кислорода и глюкозы. Таким образом, кривая BOLD прогиба-
ется для отражения потери кислорода. В следующий момент времени изгиб вверх кри-
вой BOLD свидетельствует о новой волне приносимых кровью питательных элементов
к активированному региону мозга. Эта волна пришедшего кислорода быстро исполь-
зуется нейронами, постоянно требующими энергии, и поэтому кривая снова падает к
минимальному уровню. В конце концов, обеспечение питательными веществами этого
участка мозга приходит в равновесие с их потреблением, и кривая выходит на исходный
уровень. BOLD-ответ также называют «гемодинамическим», так как он отражает быст-
рые и точные изменения локального кровотока
184
Глава 4
Рис. 4 .31 . Как происходит циклическое включение и выключение BOLD-сигнала. Физи-
ческие характеристики метода измерения BOLD требуют периодического включения и
выключения магнитного поля для регистрации радиочастотных колебаний в связи с из-
менением спина у атомов кислорода. Так как когнитивные задачи требуют секунд или до-
лей секунды, стандартным методом является чередование состояний покоя и выполнения
задания с периодом примерно полминуты или около того (в данном случае фаза длится
40 с). Это обеспечивает наличие контрольных данных. Слева функциональный сигнал
(BOLD) наложен на структурное изображение мозга (полученное с помощью МРТ). Изо-
бражение мозга получено при совмещении активностей мозга при выполнении задачи и
усреднении относительно фоновой активности. (Источник: Robinson, 2004.)
Рис. 4 .30 . Классическое ПЭТ-исследование: сравнение активности зрительных и слу-
ховых областей мозга. Ранние изображения, полученные при помощи ПЭТ, показывают
различия при произнесении речи, чтении, слуховом восприятии и внутреннем воспро-
изведением слов (Posner и Raichle, 1994). Обратите внимание на активацию зрительных,
слуховых и соматосенсорных областей мозга. Однако очертания мозга (белая линия)
нарисованы приблизительно. На более современных изображениях функциональная ак-
тивность мозга накладывается на структурные МРТ-изображения мозга испытуемого.
(Источник: Posner and Raichle, 1994.)
Методы: визуализация живого мозга
185
3.1.
Области интереса
Лавирование в мозге — непростая задача. А еще более трудной задачей явля-
ется выяснение, какие роли играют разные области мозга в высших когнитив-
ных процессах, таких как речь, внимание и зрение. Одним из выходов является
первоначальное выделение определенной области интереса (ОИ) и прогнози-
рование ожидаемой активности в ОИ.
3.1.1. Регистрация функциональной активности
во время структурного сканирования
Первым делом необходимо изучить анатомическую структуру живого мозга,
выявить различные области, связи и уровни организации. Структурное МРТ
позволяет нам картировать строение мозга, включая аксонные (белое вещество)
связи между различными регионами мозга. МРТ показывает только структуру,
но не функциональные особенности.
На рис. 4 .32 (справа) мы можем наблюдать два желтых «световых пятна»,
отражающих увеличение активности на фМРТ. Цвет в данном методе исполь-
зуется в качестве маркировки уровня активности. Для того чтобы локализовать
местоположение световых пятен, нам необходимо наложить функциональное
изображение на изображение, полученное при помощи структурного МРТ, ко-
торая имеет большую пространственную разрешающую способность. Во вре-
мя процесса совместной регистрации функциональные и структурные картин-
ки совмещаются. Совместная регистрация гарантирует, что два изображения в
итоге будут точно совмещены в одинаковой размерности. Имея большее про-
странственное разрешение, мы можем решать вопросы об анатомической спец-
ифичности.
Другой подход заключается в маркировании области интереса (ОИ) только
на структурном изображении для проведения статистического анализа.
Рис. 4 .32 . фМРТ BOLD-сигнал при болевой чувствительности в левой части туловища.
При стимулировании левой руки до появления умеренных болевых ощущений BOLD-
сигнал регистрируется в правополушарной карте тела (соматосенсорной коре, не пока-
зана), а также возникает в отмеченных областях вдоль средней линии правого полуша-
рия. Болевые пути хорошо известны, и восприятие умеренной боли помогает проверять
точность методик визуализации активности мозга. (Источник: Valet et al., 2004.)
186
Глава 4
Современные МРТ-машины (рис. 4 .27) с более мощным магнитным по-
лем позволяют исследовать клеточную организацию живого мозга и сравни-
вать мозг разных групп людей (например, людей, страдающих шизофренией, и
здоровых испытуемых). Различные уровни коры имеют либо локальные, либо
отдаленные связи, поэтому данный вид информации очень полезен для понима-
ния взаимодействия разных областей мозга.
Из-за того что мозг постоянно активен, до сих пор составляет значительную
трудность выделение мимолетной, связанной с заданием активности. Одним из
широко распространенных методов является вычитание между двумя фМРТ
изображениями, различающимися по условиям: например, сравнение мозговых
ответов в задаче на понимание арабских чисел (1, 2, 3, ...) и ответов в такой же
задаче, но в которой числа были прописными (один, два, три, ...) (рис. 4 .33).
Вычитание используется потому, что оно позволяет исключить большую часть
«нерелевантной» мозговой активности, которая в противном случае заглушила
Рис. 4 .33 . Метод вычитания данных ПЭТ и фМРТ. Мозг постоянно находится в со-
стоянии динамической активности. Для того чтобы избавиться от фоновой активности,
BOLD- или ПЭТ-сигналы при выполнении задач вычитаются, шаг за шагом, из актив-
ности мозга при наиболее сходной контрольной ситуации. Индивидуальные различия в
полученных изображениях затем усредняют для получения среднегрупповых. (Источ-
ник: Posner and Raichle, 1997.)
Методы: визуализация живого мозга
187
бы интересующий нас сигнал. Это очень похоже на сравнение неспокойной по-
верхности моря до и после того, как проплыл кит. Если необходимо увидеть
волны, вызванные движением кита, можно произвести вычитание одной за-
писи волн из другой. Именно разница между двумя состояниями важна (см.
рис. 4.33, 4.34).
Рис. 4 .34. Типичный зрительный эксперимент с использованием фМРТ. Обратите
внимание на то, что предъявляемые лица очень точно подобраны с точки зрения по-
хожести. Предъявляемые лица сравнивают с другими зрительными стимулами той же
пространственной ориентации. В исследовании используется методика сличения, по-
этому испытуемым необходимо концентрировать внимание и нажимать на кнопку при
появлении совпадающего стимула. Результаты демонстрируют более высокий BOLD-
сигнал в красной области в ответ на предъявление лиц, выше в зеленой области для
пространственного положения и высокий в светло-зеленой области при сличении одно-
временно лица и пространственной ориентации по сравнению с контролем. (Источник:
Squire et al., 2003.)
188
Глава 4
Вычитание двух условий может, однако, иметь и нежелательное послед-
ствие. Во время этих состояний могут происходить другие важные процессы.
В дополнение различия между экспериментальными и контрольными усло-
виями могут варьировать, может существовать перекрестное взаимодействие
между независимыми переменными и т. д . Следовательно, другим подходом
могут служить параметрические вариации, при которых изменение каждого
важного параметра и его взаимодействия могут быть разделены статистически.
Например, если мы изучаем рабочую память (РП) вместо того, чтобы вычитать
мозговую активность во время процессов рабочей памяти из активности без
нее, можно исследовать, как постепенное увеличение загрузки рабочей памяти
отражается на нейронной активации. А так как статистические проверки долж-
ны быть выполнены для каждой области интереса в каждый момент времени,
получается очень большой объем данных.
3.2 .
Мозг в состоянии покоя не молчит:
внутренние процессы мозга
Когда ученые стали изучать разные когнитивные функции, основным подходом
при исследовании был сравнительный, разностной подход. В данных работах
нервная активация во время выполнения когнитивной задачи, такой как произ-
несение речи, сравнивалась с состоянием, при котором испытуемых просили
расслабиться и «ничего не делать». Такое сопоставление активности с отдыхом
выявляло значительные основные эффекты. Сотни сравнительных исследова-
ний были проведены для картирования областей мозга, ответственных за ког-
нитивные функции.
И все же существует скрытое допущение в этих работах. Что бы вы делали,
если бы вас попросили спокойно лежать и «отдыхать»? Будет ли ваш разум как
белый чистый лист? Очень большое число фактов указывает на то, что люди
в таких ситуациях начинают возвращаться к своим повседневным мыслям, об-
разам и чувствам. В это время вы можете думать о том, что произошло днем
ранее; или о том, что надо будет сделать позже; или даже немного помечтать.
У вас, скорее всего, будет внутренняя речь, зрительные образы, эпизодические
воспоминания и тому подобное. Для мозга это не будет являться «отдыхом».
Вместо этого экспериментальное сравнение будет происходить между двумя со-
стояниями активности. Одно из этих состояний будет вызываться требованием
экспериментальной задачи, тогда как другое будет отражать собственные мыс-
ли, надежды, чувства, образы, внутреннюю речь и т. д. испытуемого. В некото-
рых случаях спонтанная активность может рассказать гораздо больше о есте-
ственном состоянии человеческой когнитивной деятельности, чем специальные
экспериментальные задания. Хотя, конечно, оба варианта чрезвычайно важны.
В специальной литературе теперь возросло количество работ, посвященных
изучению состояния «покоя», т. е. состояния, при котором испытуемого не про-
сят выполнять определенной деятельности (рис. 4 .35).
Использование МРТ как для получения точных анатомических изображе-
ний, так и для функционального картирования областей мозга без преувели-
чений внесло революционные изменения в область когнитивной нейробиоло-
гии. Современные и улучшенные способы использования фМРТ представлены
в следующем разделе.
Методы: визуализация живого мозга
189
3.3 .
Эмпирическое определение когнитивных функций:
уникальный способ
Выдающиеся научные открытия получаются при сочетании технологий визу-
ализации работы мозга и изобретательности гения. Очень много творческих
способностей требуется для выбора функциональной переменной. Каким об-
разом можно лучше всего понять зрение? А селективное внимание и сознатель-
ную когнитивную деятельность? Очень много остроумия было потрачено для
ответов на эти вопросы.
Вот несколько примеров. Все знают, что таксисты обладают хорошей спо-
собностью ориентироваться в городе. Они не только знают, как добраться из
пункта А в пункт Б, но и могут это сделать наиболее рациональным способом.
Такая способность перемещаться по сложной системе дорог опирается на нашу
способность к пространственному восприятию. Исследования показали, что
гиппокамп, часть медиальной височной доли, играет важную роль в навигации
и запоминании мест и маршрутов. Уже несколько десятков лет известно, что
крысы с разрушенным гиппокампом катастрофически плохо выполняют тесты
на пространственную память. Птицы и другие животные, которые закапывают
Рис. 4 .35. Мозг находится в постоянно активном состоянии. Это изображение фМРТ
демонстрирует фоновую активность в обоих полушариях, левом (слева) и правом (ме-
диальная поверхность, справа). Нижний график показывает эту активность за 300 с.
В то время как такая не связанная с выполнением заданий активность сильно отличает-
ся от активности во время решения типичных экспериментальных задач, нам известно,
что люди постоянно думают, воображают, чувствуют, воспринимают, запоминают и раз-
говаривают сами с собой, даже в том случае, когда им не дается определенного задания.
(Источник: Fox et al., 2005.)
190
Глава 4
или прячут пищу, имеют более развитый гиппокамп по сравнению с животны-
ми, которые этого не делают. Вследствие этого, когда мы думаем о таксистах,
возникает один вопрос: имеют ли таксисты более развитые участки мозга, от-
вечающие за пространственное ориентирование, по сравнению с обычными
людьми? В действительности было обнаружено, что задняя часть гиппокам-
па у таксистов больше, чем аналогичная часть у людей с другой профессией
(рис. 4 .36). Хорошо, вы можете спросить, а что если люди с большим гиппокам-
пом выбирают себе профессию таксиста, а не наоборот? Имеется исследование,
показывающее, что размер гиппокампа зависит от того, как долго человек про-
работал таксистом. Другими словами, чем дольше человек работает в качестве
водителя такси (и постоянно использует свои способности к ориентированию в
пространстве), тем больше становится соответственная часть гиппокампа.
Обратите внимание, насколько творческим было исследование, проведен-
ное на таксистах. Как правило, намного легче отобрать случайную группу ис-
пытуемых (обычно студентов!), которые бы отображали всю человеческую
популяцию. Но факт остается фактом, что существуют разница в возрасте, ин-
дивидуальные способности и таланты и другие когнитивные различия между
«усредненной» группой испытуемых. Лондонские таксисты являются выда-
ющимися профессионалами (от них требуется пройти экзамен по географии
Рис. 4 .36 . Размер гиппо-
кампа у таксистов Лондона.
Лондонские таксисты имеют
значительно отличающуюся
в размере область мозга, от-
вечающую за пространствен-
ную ориентацию, т. е. гип-
покамп. (Источник: Maguire
et al., 2000.)
Методы: визуализация живого мозга
191
Рис. 4 .37. Изображение проводящих пу-
тей белого вещества, которые напоминают
по своему виду фонтан. Эти великолепные
пучки нервных трактов получены с помо-
щью технологии магнитной визуализации,
названной диффузионной трактографией,
которая позволяет получить изображение
трактов белого (миелинизированного) ве-
щества. Это вертикальные проводящие
пути, которые идут от коры головного мозга
(особенно расположенные фронтально, они
обозначены синим цветом) вниз к спинному
мозгу, а также вверх от спинного мозга в за-
днюю половину коры головного мозга (обо-
значены желтым и зеленым цветом). Данные
массивные тяжи белого вещества занимают
большую часть объема головного мозга.
(Источник: Maria Lazar, с разрешения.)
города), и их можно сравнивать с любой контрольной группой. Одним важным
дополнением служит то, что абсолютный размер структур мозга может изме-
няться в результате определенного опыта (Maguire et al., 2000). Это заявление
в настоящее время подтверждено и для других областей мозга. Таким образом,
исследование, проведенное на таксистах, служит великолепным примером изо-
бретательного подбора сравниваемых условий, что привело к получению ново-
го понимания.
4.0 .
Новые способы определения взаимосвязи областей
мозга: визуализация распределения тензора
Пучки волокон белого вещества составляют обширную внутреннюю систему
проводящих путей коры. Можно выявить эти волокна, используя метод МРТ,
называемый визуализация распределения тензора (ВРТ). ВРТ использует по-
ток жидкости в белом веществе для определения относительного направления
трактов белого вещества. Эта информация позволяет нам изучать паттерны
взаимосвязи между двумя полушариями и между четырьмя долями мозга (для
более подробной информации об этом методе см. Приложение). ВРТ помогает
нам понять систему связей здорового мозга, а также выявить эти паттерны у
пациентов с заболеваниями нервной системы, которые вызывают повреждения
белого вещества, например множественный рассеянный склероз (МС).
Самым большим пучком проводящих волокон в мозге является мозолистое
тело (см. гл . 5), которое соединяет два полушария, но имеются также и другие
волокна, соединяющие области мозга внутри полушарий. Посмотрите на ри-
сунок в начале главы. На нем изображено распространение цветных волокон,
которые расходятся от мозолистого тела во все области коры. Ученые, которые
выполнили данный рисунок, закодировали в цвете волокна, соединяющие раз-
ные области коры, от зеленых волокон в лобных отделах до желтых — в заты-
лочной доли. Другое изображение обширного пучка волокон белого вещества
показано на рис. 4 .37, в средне-саггитальном разрезе (по линии центра) изобра-
жены волокна, направляющиеся вверх от спинного мозга до коры, а также во-
192
Глава 4
локна, идущие вниз от коры до спинного мозга. Эти пучки волокон составляют
вертикальную «транспортную магистраль», которая начинается и заканчивается
в коре и обеспечивает соединительные пути во всей центральной нервной си-
стеме. Какова связь между волокнами белого вещества и корковой анатомией?
Результаты недавних исследований по выявлению связи между белым и серым
веществом показаны на рис. 4 .38. Построенные карты периферического белого
вещества показаны в верхней части рисунка, а соответствующие им области се-
рого вещества показаны в нижней части рисунка. Обратите внимание, что на рис.
4.38 показана одна и та же информация, но с трех различных сторон: вид спереди
(а, г), вид с латеральной стороны (б, д) и сзади, с затылочного конца (в, е).
5.0 .
Сознательные события против бессознательных
Сознательная когнитивная деятельность является новым объектом исследова-
ния когнитивной нейронауки. Она включает попытки понять различия между
сознательными и бессознательными событиями в мозге. В последние одно или
два десятилетия ученые использовали множество экспериментальных парадигм
Рис. 4 .38 . Диффузионная трактография позволяет получить изображения мозолистого
тела и других крупных проводящих путей из белого вещества. Обратите внимание,
что на рисунке представлен срез мозга, у которого правое полушарие изображено пол-
ностью. На рисунке видны волокна зеленого цвета, проектирующиеся вверх, а также
желтые волокна, идущие назад. Разными цветами закодированы на компьютере раз-
личные направления проводящих путей. С-образная структура на рисунке (б) — это
мозолистое тело (corpus callosum), которое для невооруженного глаза выглядит белым.
Мозолистое тело состоит примерно из 100 млн волокон проводящих путей, соединяю-
щих два полушария. Миллионы нервных клеток левого полушария связаны с соответ-
ствующими точками в правом полушарии. (Источник: Huang et al., 2005.)
Методы: визуализация живого мозга
193
для того, чтобы сравнить эти два состояния. Одним из важных примеров этого
является бинокулярное соревнование, когда поток из двух разных зрительных
стимулов подается в оба глаза таким образом, что только один из поступающих
стимулов осознается. Поведенчески человек может дать отчет только об одном
зрительном потоке (см. гл. 6). Тесты на бинокулярное соревнование схожим
образом работают и на макаках (Logothetis, 1998), и волна текущего интереса
по поводу осознания зрительной информации может быть перенаправлена к
классическим работам о бинокулярном соревновании у макак с использовани-
ем регистрации одиночных нейронов на разных уровнях зрительной обработки
(Sheinberg and Logothetis, 1997); о бинокулярном соревновании читайте также
гл. 6.
6.0.
Корреляция и причинная связь
Как правило, мы соотносим данные о работе мозга с наблюдениями поведенче-
ских проявлений. Мы можем предъявлять зрительные изображения на экране
и просить испытуемого либо вслух читать, либо медитировать. Таким образом,
мы обычно наблюдаем корреляцию между поведением и мозговой активностью.
При использовании методов с высоким пространственным разрешением, таких
как фМРТ, при выполнении разных задач наблюдается усиление или уменьше-
ние сигнала BOLD, свидетельствующего о том, что некоторые области мозга
находятся в более активном состоянии, чем другие (см. рис. 4 .31).
Мы можем для примера рассмотреть «счетную задачу Струпа», когда испы-
туемых просят считать количество слов, предъявляемых на экране компьюте-
ра. В некоторых случаях слово, например «собака», демонстрируется три раза.
В этом случае испытуемый должен сказать «три», и это не доставляет трудно-
стей. Однако мы можем создать конфликт между числом изображенных слов
и собственно словами. Если мы будем предъявлять слово «один» четыре раза,
существует автоматическая тенденция у постоянно читающих людей (таких
как студенты колледжа) сказать «один», в то время как правильным ответом
было бы «четыре». Это очень похоже на цветовую задачу Струпа (см. гл . 2). Ис-
пытуемым требуется больше времени для того, чтобы правильно ответить, так
как им требуется подавить автоматическое стремление просто считывать слова
с экрана.
Была зарегистрирована BOLD-активность (Zheng, Rajapakse, 2006) во вре-
мя двух вариантов подсчета слов (рис. 4 .39). В то время как многие области
мозга были активированы во время обоих условий, фронтальные отделы мозга
были более активны при конфликтных условиях. Аналогичные результаты мо-
гут быть многократно получены в настоящее время в таких конфликтных за-
дачах. Одной из главных функций префронтальной коры является разрешение
конфликтных склонностей, таких как автоматическая тенденция просто читать
слова, в противоположность стремлению выполнять указания экспериментато-
ра. Следовательно, у нас имеются корреляции между (а) активацией фронталь-
ных отделов, (б) более длительными латентными периодами реакций, (в) субъ-
ективным восприятием затраченных усилий, (г) увеличением числа ошибок
при конфликтных условиях. Эти результаты очень значительны, так как суще-
ствует большое число жизненных ситуаций, в которых требуется регулировать
противоречивые действия.
194
Глава 4
Однако в настоящее время мы не имеем возможности проверять причинно-
следственные предположения. Например, нам известно, что решение задачи
требует зрительного распознавания слов, подготовки ответной реакции с вы-
бором между двумя возможными вариантами ответа, возможно обнаружение
конфликта, блокировки произнесения неправильного ответа, выбора правиль-
ного ответа и т. д . Подход, названный динамическим причинно-следственным
моделированием (ДПСМ), применяется для анализа причинно-следственных
связей. Был применен метод ДПСМ к мозговой активности, обнаруженной в
обоих вариантах задачи на подсчет слов (Zheng, Rajapakse, 2006). Как можно
видеть из рис. 4 .39, по результатам ДПСМ каждая задача сопровождается раз-
личным активационным паттерном. И хотя многие области мозга активны во
время выполнения обеих задач, их относительные связи и вклад различались.
Особенно интересен тот факт, что конфликтные условия требуют более об-
ширной активации головного мозга по сравнению с контрольными условиями.
Этот факт является стандартным для условий, требующих мысленных усилий
(Duncan and Owen, 2000).
Рис. 4 .39 . Причинная взаимос-
вязь между областями мозга,
участвующими в тесте Струпа.
Активность мозга при простом
подсчете без интерференции по-
казана на верхнем рисунке (а),
а активность счета с интерфе-
ренцией
продемонстрирована
на нижнем рисунке (б). Каждый
круг отражает область мозга. Как
видно из рисунка (б), интерфе-
ренция ведет к вовлечению бо-
лее обширной области, чем для
контрольной ситуации (а). Такое
изменение в причинной взаимо-
связи между областями мозга ре-
гистрируется несмотря на то, что
многие из этих областей задей-
ствованы в выполнении обeих
задач. Обратите внимание, что
некоторые связи исчезают при
переходе от выполнения кон-
трольной задачи к задаче с ин-
терференцией. (Источник: Zheng
and Rajapakse, 2006.)
Методы: визуализация живого мозга
195
Рис. 4 .40 . Действительно ли изобра-
жены области коры отвечающие за
правду и ложь? фМРТ изображения
отличается теми областями мозга,
которые имели большую BOLD-
активацию в случае произнесения
правды (зеленые), и корковыми об-
ластями при произнесении лжи
(красные). Таким образом, можно
ли считать соответствующие регио-
ны коры головного мозга областями
правды (зеленые) и лжи (красные)?
Почему или почему нет? (Источник:
Davatzikos et al., 2005.)
6.1.
Для чего необходимо множество методик
при исследовании функций мозга
Согласно некоторым заголовкам в прессе, ученые, изучающие мозг, недавно
обнаружили области мозга, ответственные за хитрость и ложь. Такого рода за-
головки происходят от исследований, аналогичных показанному на рис. 4.40.
На рисунке продемонстрированы различия в изображениях фМРТ областей
мозга, имеющих различную BOLD-активность во время того, как люди говорят
правду (зеленые) и когда они вынуждены врать. В такого рода экспериментах
196
Глава 4
обычно используют игральные карты, и испытуемых просят «лгать» относи-
тельно того, какая карта находится у них в руках.
Одной из основных целей когнитивной нейронауки является соотнесение
функций со структурами, т. е. определение того, выполняет ли какая-нибудь об-
ласть мозга специфическую для нас работу. Однако во время такого исследова-
ния мы должны быть более тщательными относительно того, какие выводы де-
лаем из полученных результатов. Популярная пресса может быть не настолько
аккуратна в выводах, как того требуется от настоящих ученых. Вы верите, что
области мозга зеленого цвета на рис. 4 .40 действительно являются областями
«говорящими правду»?
6.2 .
Повреждения мозга и следствия этих повреждений
Мозговые повреждения могут снабдить нас информацией об областях, которые
необходимы для определенных когнитивных процессов. Исторически Поль
Брока обнаружил пациентов, которые не могут говорить и у которых одновре-
менно с этим наблюдались повреждения левой лобной доли. Однако их воз-
можность понимать речь была относительно сохранной. В это же время Карл
Вернике сделал открытие, что повреждения различных областей левого полу-
шария связаны с возможностью понимать речь, тогда как способность говорить
оставалась нетронутой. Сегодня данную пару взаимодополняющих симптомов
мы называем двойным расстройством.
Повреждения мозга можно рассматривать только с точки зрения корреляции
их с нарушением функций, так как мы не можем воспроизвести определенные
повреждения и исследовать их последствия на людях. Несмотря на это, после
многочисленных дискуссий и споров, сегодня нет сомнения в том, что одной
из основных функций поля Брока является воспроизведение речи, а главной
функцией поля Вернике служит понимание речи (рис. 4 .41). Разрушения в об-
ласти поля Брока могут привести к дизартрии, состоянию, когда контроль над
движениями рта и языка нарушен, а воспроизведение речи не страдает. Таким
образом, мы имеем три вида повреждений, которые ведут соответственно к
трем нарушениям речевой функции: одна область мозга важна для понимания
смысла речи; другая необходима для воспроизведения речи; а третья требуется
для управления моторными функциями при произнесении слов.
Травмы головного мозга у людей возникают по множеству причин, от авто-
мобильных аварий до инсультов. Случайные повреждения, как правило, не сле-
дуют определенным границам мозговых областей. Для того чтобы соотнести
структуры мозга и их функции, необходимо иметь более точную информацию о
том, где возникло повреждение. Для этого проводятся различные исследования
на животных, в основном на крысах и обезьянах. Однако так как речь явля-
ется чисто человеческим свойством, невозможно провести прямые параллели
с повреждениями мозга у других видов. (Хотя, как уже было написано выше,
в настоящий момент существует способ безопасного вмешательства в работу
определенных областей мозга, использующий ТМС.)
Значительные результаты принесли многолетние исследования локаль-
ных разрушений областей мозга у обезьян и крыс. Например, производились
очень локальные разрушения в разных областях медиальной височной доли
(МВД) у макак (Buckle�, Gaffan, 2006). Очень специфическое повреждение
Методы: визуализация живого мозга
197
периринальной коры (расположенной около «обонятельной коры») вызывало у
обезьян увеличение числа ошибок в задачах на дискриминацию зрительных
стимулов. При разрушении соседних областей такого эффекта не наблюдалось.
Чем более сложной была дискриминационная задача, чем более похожими
были предъявляемые стимулы, тем больше ошибок совершали животные с опи-
санным повреждением. Однако с простыми задачами на различение зритель-
ных стимулов, когда требовалось различать цвета, формы и ориентацию, такие
животные справлялись нормально. Из этого можно сделать вывод, что перири-
нальная кора выполняет причинную роль в обработке сложных зрительных сти-
мулов, таких как лица. Большое число исследований, проведенных на людях,
подтверждают данное положение. Исследования на животных очень часто ис-
полняют роль пионерных исследований, которые позволяют ученым находить
новые идеи для дальнейших исследований, которые впоследствии проверяются
и подтверждаются в работах на людях.
Рис. 4 .41 . Разрушение речевых зон. Разрушение либо зоны Брока, либо зоны Верни-
ке вызывает очень серьезное нарушение речевой функции. Обратите внимание, что в
правом полушарии отсутствуют зоны Брока и Вернике, несмотря на то что традиционно
считается, что оно также вовлечено в восприятие речи и языка. (Источник: Standring
et al., 2005.)
198
Глава 4
Сравнение поврежденного и здорового головного мозга является мощным
источником фактов для понимания основополагающих нервных процессов.
7.0 .
Заключение
Технологии для исследования мозга позволяют измерять как активность еди-
ничных нейронов, так и больших корковых областей, а также структуру го-
ловного мозга, динамику активности и связей. Изобретение методов нейрови-
зуализации работы мозга изменило технологию исследования человеческого
сознания. Постоянно идет изобретение новых и усовершенствование старых
методов. Одним из современных достижений является регистрация нескольких
параметров с помощью разных методов в одном исследовании, что позволяет
дополнять результаты электромагнитных измерений данными от непрямого из-
мерения активности мозга.
8.0 .
Задания и упражнения к главе 4
8.1.
Контрольные задания и графические упражнения
1. Назовите отличия между изображенными на рис. 4.42 сканами мозга и при-
ведите объяснение применения метода вычитания для каждого изображе-
ния.
2. Дайте определение термину BOLD-ответ. Расшифруйте аббревиатуру
BOLD.
3. Сколько составляет задержка во времени между нейронной активностью и
BOLD-ответом? А между нейронной активностью и ЭЭГ-ответом?
4. Перечислите все за и против использования методики единичной регистра-
ции нейронов в головном мозге.
5. Какие проблемы могут возникнуть при сравнении активности головного
мозга при выполнении когнитивной задачи с активностью в покое?
6. Какую информацию о лжи и областях коры головного мозга мы можем полу-
чить из рис. 4 .40?
7. Какой объем нервной ткани, на основе данных диффузионной трактогра-
фии, вовлечен в формирование связей между корковыми нейронами (при-
близительно)?
8. Опишите четыре основных типа ритмов электрической активности головно-
го мозга. С какими функциями эти ритмы обычно соотносят (см. табл. 4 .1)?
Методы: визуализация живого мозга
199
Рис. 4 .42 . Воспроизведение рис. 4 .33 без подписей для использования в графических
упражнениях
Мозг заключен внутри черепной коробки и представляет собой переднюю,
наиболее развитую часть центральной нервной системы.
Генри Грей (1918)
Если посмотреть сквозь серый внешний слой коры головного мозга, можно увидеть
скопление белого вещества. В центре расположена группа больших ядер, включающая
базальные ганглии, гиппокамп, миндалину и две яйцевидные структуры в самом центре,
едва видимые на этом рисунке, — таламус. Таламус покоится на нижележащем стволе
головного мозга (темно- и светло-голубом). Вы также можете видеть питуитарную же-
лезу (гипофиз) впереди (бежевую) и мозжечок позади головного мозга (розовый).
В этой главе мы последовательно разберем и реконструируем эти структуры по отдель-
ности, начиная с нижних отделов мозга.
Содержание
1.0.Введение.......................................................201
1.1.Нервнаясистема..............................................202
1.2.Географиямозга..............................................203
2.0.Развитиемозганачинаетсяснижнихотделов.........................210
2.1 . В строении мозга находят отражение эволюция и индивидуальное
развитие.....................................................210
2.2.Развитиемозгаотоснованиядовершины.........................210
3.0 . От вопроса «где?» до вопроса «что?»: функциональная роль
областеймозга...................................................217
3.1 . Полушария головного мозга: разделение на левую–правую
половины....................................................218
3.2 . Исходящие и входящие сигналы: подразделение в направлении
сперединазад................................................222
3.3.Основныедоли:видимыеискрытыеучастки......................226
3.4.Обширныевзаимосвязимеждукоройиталамусом.................233
3.5.Подчиненныеучасткиподкорковыхслоев........................236
4.0.Заключение.....................................................239
5.0.Заданияиупражнениякглаве5....................................240
5.1.Контрольныезадания..........................................240
5.2.Графическиеупражнения......................................240
1.0 . Введение
Наш мозг является самым большим нашим эволюционным достижением. Дру-
гие млекопитающие большого размера имеют большую мышечную массу или
развивают более высокую скорость, а люди имеют исключительно большой и
пластичный мозг, приспособленный для отличных зрения и слуха, речевых и
социальных навыков, для управления моторикой рук и для гибкого контроля
выполнения действий. Человеческий мозг делает возможным развитие культу-
ры и техники.
В этой главе мы рассмотрим строение мозга, а в следующих осветим его
функции, то, как он, вероятно, работает. Важно понимать, что анатомия мозга
не является статичной и неизменной областью знаний; постоянно обнаружива-
ются новые и важные факты. На микроскопическом и наноскопическом уров-
нях были найдены целые новые классы нейронов, синапсов, систем связи и
нейромедиаторов. Несмотря на то что знания о мозге постоянно расширяются,
мы сконцентрируемся на основах.
Когнитивная нейробиология неизбежно сосредоточивает свое внимание
на коре головного мозга, функции которой, как полагают, включают «высший
Глава 5
Мозг
202
Глава 5
уровень» обработки информации. Кора является только внешней и видимой
частью огромного мозга, который развивался в процессе эволюции в течение
сотен миллионов лет. Слово «кора» пришло в голову первым анатомам по ана-
логии с корой дерева. В то время как кора больших полушарий необходима для
выполнения познавательных функций, она постоянно взаимодействует с ос-
новными «подчиненными» органами, в частности с таламусом, базальными
ганглиями, мозжечком, гиппокампом и лимбической областью. Самые тесные
связи образованы между корой и таламусом, и поэтому их часто объединяют
в таламокортикальную систему. В этой центральной системе мозга движение
сигналов может легко осуществляться по всем направлениям, как полет воз-
душных судов над Землей.
Основные доли коры сравнимы с континентами Земли, каждый со своими
популяционными центрами, естественными ресурсами и торговыми отноше-
ниями с другими регионами. Несмотря на то что области коры часто специали-
зированы, они также тесно объединены с другими областями через похожие на
паутину связи, которые простираются по всей коре, и связанными с ней струк-
турами. Этот внешний слой называется серым веществом из-за того, что так
выглядит. Он является внешней «кожей» для белого вещества больших полу-
шарий, которое заполняет их, как мякоть плод. Однако это только видимость. В
действительности серое вещество содержит клеточные тела десятков миллиар-
дов нейронов, которые посылают даже большее количество аксонов во все на-
правления, и покрыты поддерживающими миелиновыми клетками, заполнен-
ными белыми молекулами липидов. Эта белая миелиновая оболочка отростков
нейронов коры такая плотная, что из-за нее связи между корковыми нейронами
окрашены в белый цвет для невооруженного глаза.
1.1 .
Нервная система
Мозг является частью нервной системы, пронизывающей тело человека. Две
ее главные части — это центральная нервная система (ЦНС), которая включает
головной и спинной мозг, и периферическая нервная система (ПНС), которая
содержит автономную и периферическую чувствительную и двигательную си-
стемы (рис. 5 .1). ЦНС и ПНС совместно образуют динамичную и обширную
систему передачи информации через все части организма, с центром в голов-
ном мозге, который получает данные через спинной мозг. В этой главе мы будем
говорить об одной части ЦНС — о головном мозге (рис. 5 .2).
В данной главе мы сконцентрируемся на двух сенсорных системах в моз-
ге, обеспечивающих вход сигналов, — зрительной и слуховой системах. Хотя
существуют и другие сенсорные системы, такие как обонятельная (запах) и со-
матосенсорная (осязание), зрительная и слуховая системы наиболее изучены из
всех систем в головном мозге человека. Мы также сконцентрируемся на двух
системах исходящих сигналов: речи и управлении моторикой, в связи с тем
что они были предметом большинства исследований. В этой главе о мозге мы
опишем анатомию мозга в целом и его областей в частности, а также выделим
функции, которые они выполняют. Начнем мы с обсуждения многоуровневого
анализа, на который можем опираться при описании мозга: от обширных об-
ластей, таких как полушария головного мозга и доли коры, до систем меньшего
масштаба, таких как топография корковых слоев.
Мозг
203
1.2.
География мозга
Давайте начнем с рассмотрения крупных областей мозга и в процессе работы
перейдем к более подробному анализу. Для начала укажем достаточно очевид-
ную вещь, что мозг находится в голове человека, как изображено на рис. 5 .3 .
Мы можем рассматривать мозг в терминах различных географических уров-
ней — от континентов до стран, областей и городов. Таким образом, мы имеем
несколько уровней детализации. Первые выделенные географические обла-
сти — это два полушария головного мозга, которые полностью разделены, но
соединены посредством сложной связывающей структуры, которая называется
мозолистым телом. Позже в этой главе мы опишем большие полушария голов-
ного мозга более подробно: вопрос, почему у нас в головном мозге имеется два
обособленных полушария, давно интересовал ученых, а также философов.
Рис. 5 .1 . Центральная и периферическая
нервная система. (Источник: Standring,
2005.)
Рис. 5 .2 . Центральная нервная систе-
ма включает спинной мозг и головной
мозг. (Источник: Standring, 2005.)
204
Глава 5
На следующем уровне находятся доли коры головного мозга (рис. 5 .4):
в каждом полушарии имеется четыре доли. Спереди, в передней части мозга
(расположенной слева на рис. 5 .4), можно увидеть лобную долю. Непосред-
ственно за лобной долей, на верхней стороне мозга выделяется теменная доля.
Внизу от теменной доли и по соседству с лобной долей находится височная
Рис. 5 .3 . Местоположение головного
мозга, правое полушарие показано с ме-
диальной стороны
Рис. 5.4 . При взгляде на головной мозг с латеральной стороны мы можем увидеть четы-
ре основные доли коры. На данном рисунке показан вид левого полушария: лобная доля
расположена в передней части головного мозга; теменная доля позади от лобной доли
на верхней стороне мозга; височная доля позади от лобной доли и снизу от теменной
доли; затылочная доля позади как теменной, так и височной долей. Прямо под затылоч-
ной долей находится мозжечок, который не является частью коры, но видим при взгляде
на мозг с большинства сторон. (Источник: Squire et al., 2003.)
Мозг
205
доля. Сзади располагается затылочная доля. Позднее в этой главе мы опишем
анатомические особенности и когнитивные функции этих долей. Мы можем
различить невооруженным глазом основные доли с поверхностью, покрытой
выпуклостями и вогнутостями, извилинами и бороздами. Некоторые из них на-
столько важны, что мы назовем их опознавательными знаками — нам нужно
их знать, чтобы понять все остальное. На рис. 5 .5 показаны некоторые главные
ориентиры, которыми давно пользовались ученые, изучавшие мозг, для иденти-
фикации его областей. Эти ориентиры широко используются в настоящее время
при обсуждении результатов исследований мозга.
Цитоархитектонические поля Бродмана — более детальный уровень опи-
сания головного мозга. При тщательном изучении с помощью микроскопа по-
верхностных слоев коры головного мозга можно увидеть локальные различия
во внешнем виде клеток в различных слоях и их связях. Такие тонкие отличия
были впервые описаны Корбинианом Бродманом в 1909 г. и в связи с этим из-
вестны как поля Бродмана, каждое из которых имеет свой собственный номер
(на рис. 5 .6 показан вид сбоку цитоархитектонических полей Бродмана лево-
го полушария, на рис. 5 .7 медиальный вид (медиальный сагиттальный) полей
Бродмана правого полушария). Сейчас выделяют около 100 цитоархитектони-
ческих полей Бродмана, и таким образом, можно принять эту цифру как грубую
оценку количества специализированных областей коры головного мозга. Поля
Бродмана предопределяют функциональную специализацию коры, например
зрительные и слуховые поля, двигательную кору и поля, отвечающие за рече-
вые и познавательные функции. По существу, они являются почтовыми индек-
сами мозга. Их размеры колеблются от нескольких квадратных сантиметров
(первичная зрительная кора, например, имеет размер кредитной карточки) до
небольшого участка поля Бродмана 5 на вершине соматосенсорной зоны коры.
Заметьте, что на рис. 5 .6, где мозг повернут влево, соседние поля Бродмана
окрашены для того, чтобы показать их основные функции, включая зритель-
ную, слуховую, обонятельную, двигательную, центр Брока (центр речи) и центр
Вернике (восприятие речи и понимание). На этот рисунок мы будем ссылаться
на протяжении всей книги.
Мы можем сосредоточиться и на более конкретных деталях, исследуя ги-
перколонки, колонки и отдельные нейроны. На этом детальном уровне разре-
шения в настоящее время эталоном являются координаты Талайраха (Talairach,
Tournoux, 1988), которые используются при в функциональной нейровизуа-
лизации мозга. Систему Талайраха можно сравнить с координатами на карте
мира, которые появляются на GPS-устройстве (устройстве по выводу коорди-
нат глобальной системы позиционирования). Они указывают «адреса улиц»
в головном мозге.
Обратимся к рис. 4 .3 . Тонкие красные линии показывают оси трехмерной
системы координат. В верхнем левом углу мы видим изображение медиальной
стороны правого полушария, направленного влево (ориентир — небольшой
рисунок головы). На этом изображении горизонтальная красная линия всегда
проходит между эпифизом (невидимым нам) и небольшим поперечным срезом
передней комиссуры — одним из крошечных мостов из белых волокон, который
проходит между двумя частями мозга. Нулевая отметка трехмерной системы
координат (0, 0, 0) всегда находится впереди этих двух точек. Это позволяет с
206
Глава 5
большой надежностью определять все три координаты. Можно отметить три
стандартных проекции мозга: медиальная (сагиттальная), горизонтальная или
осевая, и коронарная (венечная). Такой способ демонстрации позволяет любой
точке головного мозга иметь точное обозначение в виде координат трех изме-
рений. Человеческий мозг, так же как и человеческие лица, очень изменчив,
система Талайраха позволяет математически «втиснуть в обувную коробку»
индивидуальные различия в строении мозга, так что их можно сопоставить по
единой схеме.
Рис. 5 .5 . Некоторые важные ориентиры головного мозга на левом полушарии, вид сбо-
ку (слева) и вид с медиальной стороны (справа). (Источник: Standring, 2005.)
1 — верхняя лобная извилина (gyrus frontalis superior); 2 — верхняя лобная борозда
(sulcus frontalis superior); 3 — средняя лобная извилина (gyrus frontalis medius); 4 —
нижняя лобная извилина (gyrus frontalis inferior); 5 — восходящая ветвь латеральной
борозды (ramus ascendence); 6 — треугольная часть (pars triangularis); 7 — глазнич-
ная часть (pars orbitalis); 8 — покрышечная часть (pars opercularis); 9 — латеральная
борозда (sulcus lateralis); 10 — верхняя височная извилина (gyrus temporalis superior);
11 — верхняя височная борозда (sulcus temporalis superior); 12 — средняя височная
извилина (gyrus temporalis medius); 13 — нижняя височная борозда (sulcus temporalis
inferior); 14 — нижняя височная извилина (gyrus temporalis inferior); 15 — задняя
ветвь (ramus posterior); 16 — полулунная борозда (sulcus lunatus); 17 — боковая за-
тылочная борозда (sulcus occipitalis lateralis); 18 — угловая извилина (gyrus angularis);
19 — надкраевая извилина (gyrus supramarginalis); 20 — теменно-затылочная борозда
(sulcus parieto-occipitalis); 21 — нижняя теменная долька (lobulus parietalis inferior);
22 — верхняя теменная долька (lobulus parietalis superior); 23 — внутритеменная бо-
розда (sulcus intraparietalis); 24 — постцентральная борозда (sulcus postcentralis);
25 — постцентральная извилина (gyrus postcentralis); 26 — центральная борозда (sulcus
centralis); 27 — предцентральная извилина (gyrus precentralis); 28 — предцентраль-
ная борозда (sulcus precentralis); 29 — клин (cuneus); 30 — шпорная борозда (sulcus
calcarinus); 31 — язычная извилина (gyrus lingualis); 32 — перешеек поясной извилины
(isthmus gyri cinguli); 33 — коллатеральная борозда (sulcus collateralis); 34 — боковая
затылочно-височная извилина (gyrus occipitotemporalis lateralis); 35 — затылочно-ви-
сочная борозда (sulcus occipitotemporalis); 36 — медиальная затылочно-височная из-
вилина (gyrus occipitotemporalis medialis); 37 — парагиппокампальная извилина (gyrus
parahippocampalis); 38 — носовая борозда (sulcus rhinalis); 39 — крючок (uncus);
40 — подмозолистое поле (area subcallosa); 41 — средняя лобная извилина (gyrus
frontalis medius); 42 — поясная борозда (sulcus cinguli); 43 — мозолистое тело (corpus
callosum); 44 — поясная извилина (gyrus cinguli); 45 — парацентральная долька (lobulus
paracentralis); 46 — предклинье (precuneus)
Мозг
207
Продолжим наше описание строения мозга, обратив внимание на тонкую
структуру коры головного мозга. Видимая наружная поверхность головного
мозга покрыта обширным тонким «плащом» толщиной только в шесть клеточ-
ных слоев (рис. 5 .8), который называется корой головного мозга (аналогично,
кора дерева). Этот слой называется серым веществом из-за того, как мы его
видим невооруженным глазом. Не любая кора имеет шесть слоев, а только раз-
витая кора головного мозга млекопитающих, поэтому она иногда называется
«неокортексом». Еще раньше возникшие области коры также можно найти у
рептилий, у ящерицы, например, такие как лимбическая кора, о которой мы
будем говорить позднее в этой главе. В этой области коры головного мозга име-
ется 5 слоев, и иногда ее упоминают как «древнюю кору» (палеокортекс).
Шесть горизонтальных слоев коры головного мозга образуют кортикальные
колонки, вертикальные цилиндрические срезы (рис. 5 .8). Они часто содержат
тесно связанные нейроны, такие как зрительные клетки, которые реагируют на
определенную ориентацию световой полоски только в одной части зрительного
поля. Колонки можно объединить в гиперколонки, которые могут быть частью
еще больших скоплений. Таким образом, кора головного мозга имеет как гори-
зонтальную структуру, образованную шестью слоями клеток, так и вертикаль-
Рис. 5 .6 . Распределение полей
Бродмана в левом полушарии,
вид сбоку. Поля 41 и 52 указаны
линиями. Некоторые области,
такие как островок и слуховая
область, спрятаны за височной
долей
Рис. 5 .7 . Распределение полей
Бродмана в правом полушарии,
сагиттальный срез, вид с ме-
диальной стороны
208
Глава 5
ную — колонки, гиперколонки и в конечном счете целые специализированные
области. В качестве модельной при изучении зрения на животных часто ис-
пользуют зрительную кору макаки. Зрительная кора человека выглядит доволь-
но схоже. Нужно заметить, что эти шесть слоев нумеруются (римскими цифра-
ми), начиная сверху, где находится слой I, и дальше вниз до слоя VI (рис. 5 .9).
Аналогия с географией полезна, но мозг, как и мир, динамичен. Строятся
новые улицы, а старые передвигаются или перестраиваются. Дома и их жители
появляются и исчезают. Примерно до последнего десятилетия существовало
мнение, что во взрослом мозге нейроны не изменяются, но сейчас мы знаем
множество способов того, как нейроны продолжают расти, мигрировать, об-
разовывать или разрывать связи, умирать даже в здоровом мозге взрослого че-
ловека. Мозг никогда не застывает в статичном, похожем на камень, состоянии.
Эти стороны вопроса динамики мозга можно увидеть даже на уровне шести
слоев коры головного мозга. Давайте еще раз посмотрим на шесть слоев коры
головного мозга, но, с другой точки зрения, при этом используем схематиче-
ский рисунок слоев, а также их входящих и исходящих путей (рис. 5 .9). Заметь-
те, что некоторые нейроны коры посылают свои аксоны к таламусу, в то время
как другие получают входящий сигнал от нейронов таламуса. Миллионы нерв-
Рис. 5 .8. Шесть основных слоев коры
головного мозга, поперечный разрез.
На рисунке показаны три колонки
поля 17, также называемого V1, пер-
вичным зрительным проекционным
полем коры. (Источник: Squire et al.,
2003.)
Мозг
209
ных клеток коры идут к противоположному контралатеральному полушарию,
в то время как множество других посылают свои аксоны в то же полушарие
(ипсилатеральная сторона). Однако самые тесные связи существуют с соседни-
ми нейронами. I слой коры головного мозга состоит в основном из дендритов
(входящие волокна), которые так тесно уложены и соединены между собой, что
этот слой иногда называют «фиброзная сеть», слой переплетенных дендритов.
Нейроны на этом рисунке (рис. 5 .9) названы «пирамидными», потому что их
тела выглядят как микроскопические пирамиды. Они заключены в матрикс из
глиальных клеток, который здесь не показан. Эта структура связей в коре голов-
ного мозга претерпевает наибольшие изменения во время развития человека, а
также в течение всей жизни (см. также обсуждение этого в гл. 15).
Рис. 5 .9 . Схематический рисунок шести слоев серого вещества коры головного мозга.
Заметьте, что некоторые корковые нейроны посылают свои аксоны к таламусу, в то вре-
мя как другие получают входящий сигнал от нейронов таламуса. Ипсилатеральный —
с той же стороны коры (того же полушария); контралатеральный — с противоположной
стороны (противоположного полушария)
210
Глава 5
2.0 .
Развитие мозга начинается с нижних отделов
2.1.
В строении мозга находят отражение эволюция
и индивидуальное развитие
Обычно мы рассматриваем мозг с внешней стороны, так что кора головного
мозга является наиболее заметной структурой. Однако мозг растет и изменяет-
ся по направлению изнутри наружу, как дерево, которое берет начало из одного
семени, далее превращаясь в тоненький росток, а затем быстро вырастая в трех
направлениях: вверх, вперед и наружу от оси роста. Эта точка зрения приложи-
ма как для филогенеза — о развитии видов, так и для онтогенеза — о развитии
мозга человека, начиная от стадии плода и далее.
Взрослый мозг воспроизводит эту схему роста и развития. Например, это
означает, что более нижележащие области, такие как стволовая часть мозга,
в основном более древние, чем вышележащие области, такие как лобная кора.
Основные жизненно важные функции, такие как дыхание, контролируются
нервными центрами в нижележащих частях ствола головного мозга, в то время
как обширная префронтальная кора человека является более поздним добавле-
нием к основной схеме мозга млекопитающего. Она расположена дальше вверх
и вперед от оси мозга (рис. 5 .11). Таким образом, частичное повреждение пре-
фронтальной коры имеет небольшое влияние на основные функции жизнедея-
тельности (рис. 5 .12), но оно может ухудшить такие сложные способности, как
принятие решений, самоконтроль, и даже влиять на личность.
2.2 .
Развитие мозга от основания до вершины
Так как в процессе сотен миллионов лет эволюции мозг развивается путем на-
ложения новых слоев на поверхность более старых, таким образом более мо-
лодые слои скрывают более древние. Особенно это справедливо в отношении
быстро развивающегося неокортекса приматов и человека, который называют
«новой корой» из-за того, что он возник позже в эволюции и имеет шесть слоев
по сравнению с четырьмя или пятью слоями мозга рептилий и ранних млеко-
питающих. Таким образом, развитие мозга в эволюции напоминает рост гриба.
Осевая часть нервной системы, спинной и головной мозг, образуется из кро-
шечных скоплений клеток, затем вырастает в тонкую цилиндрическую трубоч-
ку, затем утолщается по направлению к центру и образует спинной мозг, покры-
тый сверху образованием, напоминающим по форме гриб. Вначале головной
мозг эмбриона при развитии принимает S-образную форму, а затем новая кора
покрывает более ранние области.
Мы можем проследить структуру головного мозга от основания до верши-
ны, чтобы показать то, что в норме спрятано под «шляпкой гриба». Рекомен-
дуем вам зарисовать эти последовательные уровни огромного «здания» мозга
(рис. 5 .10).
В отличие от большинства других млекопитающих люди встали вертикаль-
но и, таким образом, повернули свои глаза и кору под прямым углом вперед по
отношению к телу. Вот почему верхнюю поверхность головного мозга челове-
ка называют как «дорсальной», что означает «по направлению к спине», так и
«верхней», что означает по направлению вверх. Противоположная поверхность
называется «вентральной», «по направлению к животу», а также «нижней».
Мозг
211
Рис. 5 .10 . Рост мозга от основания до вершины. Если вы сможете запомнить эти ос-
новные структуры, у вас будет твердый фундамент для понимания строения головного
мозга. Обратите внимание, как мозг достраивается над стволом мозга, на вершине кото-
рого лежат таламусы, основные центры переключения входящих сигналов. Гиппокамп
и миндалины на самом деле располагаются внутри каждой из височных долей. Свет-
ло-голубые желудочки с цереброспинальной жидкостью не имеют нейронов, но обе-
спечивают собственную циркуляторную систему головного мозга. Базальные ганглии
можно рассматривать как центр исходящих сигналов. Огромное количество сигналов
идет также обратно к коре головного мозга
Рис. 5 .11 . Схема эволюции мозга млекопитаю-
щих. Передний мозг развивается и расширяется
вдоль линий трех основных нервных структур,
которые анатомически и биохимически отра-
жают общность с рептилиями, ранними мле-
копитающими и поздними млекопитающими.
(Источник: MacLean, 1967.)
212
Глава 5
Для того чтобы избежать путаницы, необходимо освоить двойную термино-
логию, которой пользуются при описании головного мозга человека. В этом
разделе мы будем «выращивать» мозг, начиная от основания, более ранних об-
ластей мозга, и наслаивая один слой за другим до тех пор, пока не достигнем
самой новой части головного мозга — неокортекса. Начнем со ствола головно-
го мозга и моста (рис. 5 .16), которые являются основанием или «старейшими»
частями мозга.
Ствол головного мозга (рис. 5 .13) является продолжением спинного мозга.
Его верхняя часть, мост, имеет нервные волокна, которые соединяют две по-
ловины мозжечка. Ствол головного мозга и мост формируют основной путь от
спинного мозга к головному. Здесь регулируются некоторые основные функ-
ции, такие как контроль частоты дыхания и сокращений сердца.
Далее идет таламус, на самом деле это таламусы, так как их имеется два —
по одному с каждой стороны (рис. 5 .14). Два яйцевидных таламуса образуют
верхнюю часть ствола головного мозга. Таламус является основным центром
передачи сигналов в головном мозге. Он также тесно связан с каждым полу-
шарием головного мозга.
Непосредственно снизу и впереди от таламуса находится группа ядер, ко-
торая называется гипоталамусом. Он связан с гипофизом, который часто на-
зывают «главной железой» организма (рис. 5.15). Вместе гипоталамус и гипо-
физ образуют чрезвычайно важный нейрогуморальный комплекс. Гипоталамус
контролирует физиологический гомеостаз. Когда нейроны гипоталамуса реги-
стрируют отклонение от нормального уровня кислорода в крови, они запускают
усиление дыхания, вдох, который вы можете сделать после напряженного чте-
ния при сидении в зажатом положении. Гипоталамус также выполняет ключе-
вые функции в эмоциональной сфере.
На вершине таламуса находятся, как всадник на лошади, два гиппокампа,
по одному с каждой стороны (рис. 5 .10, 5.17). Каждый гиппокамп, как мы уви-
дим позже, удобно устроился внутри височной доли с каждой стороны. Важно
отметить двойную структуру двух гиппокампов. Как мы уже говорили, гиппо-
Рис. 5 .12 . Действительно ли нам необходима
кора? Сканированное изображение структуры
мозга (МРТ) семилетней девочки, которой уда-
лили хирургическим путем левое полушарие
головного мозга в возрасте 3 лет по причине
энцефалита Расмуссена. Такие хирургические
операции могут спасти жизнь детей, если они
выполнены достаточно рано. Так как мозг в та-
ком возрасте очень пластичен, способность к
речи переместилась в правое полушарие. Одна-
ко заметьте, что ее ствол головного мозга и тала-
мус были не затронуты. Ствол головного мозга
ответственен за жизненно важные функции и
не может быть удален. Девочка может играть
и говорить и имеет легкие нарушения двига-
тельной функции справа. (Источник: Borgstein,
Grotendorst, 2002.)
Мозг
213
камп играет важную роль в передаче опытной информации в долговременную
память, а также в извлечении эпизодических воспоминаний. Он также имеет
отношение к пространственной ориентации.
Рис. 5 .13 . Детальная анатомия ствола головного мозга и моста. Заметим, что здесь рас-
положены все основные входящие и исходящие пути головного мозга, как идущие вниз
к спинному мозгу, так и выходящие через узкие отверстия в черепе. Зрение, слух, обо-
няние и вкус используют черепно-мозговые нервы как основные пути. То же делают
болевая и осязательная чувствительность в пределах головы. Мозговой ствол также
контролирует такие жизненно важные функции, как дыхание и скорость сердечных со-
кращений. (Афферентный — входящий в кору; эфферентный — исходящий из коры.)
(Источник: Standring, 2005.)
214
Глава 5
Недалеко от каждой петли гиппокампа находится миндалевидная структура,
называемая миндалиной, которая играет главную роль в эмоциях и эмоциональ-
ных ассоциациях (рис. 5.18).
Следующий уровень сверху вводит нас в заблуждение. Эта структура вы-
глядит как относящаяся к нервной системе, но на самом деле ею не являет-
ся. Это четыре желудочка, из которых можно выделить правый и левый (рис.
5.10). Желудочки представляют собой небольшие полости, содержащие цир-
кулирующую жидкость, которая отделена от кровотока. Эта специализирован-
ная циркуляторная система мозга опускается в спинной мозг (имеется в виду
в продолговатый мозг. — Прим. перев.) через крошечный канал, называемый
Рис. 5 .16. Мы начинаем «растить» мозг
от ствола головного мозга и моста. (Ис-
точник: Baars, Fu.)
Рис. 5 .17. Схематический рисунок гип-
покампа. (Источник: Baars, Fu.)
Рис. 5 .14 . Таламус, просвечивающий
в центре каждого полушария, а также
базальные ганглии, выглядящие как
«щит и петля», с наружной стороны от
каждой половины таламуса. (Источ-
ник: Ohye, 2002.)
Мозг
215
Р
и
с
.
5
.
1
5
.
Г
и
п
о
т
а
л
а
м
у
с
(
в
ы
д
е
л
е
н
т
е
м
н
о
-
г
о
л
у
б
о
й
л
и
н
и
е
й
)
о
б
р
а
з
о
в
а
н
г
р
у
п
п
о
й
я
д
е
р
,
р
а
с
п
о
-
л
о
ж
е
н
н
ы
х
к
а
к
р
а
з
н
и
ж
е
и
в
п
е
р
е
д
и
о
т
к
а
ж
д
о
й
и
з
п
о
л
о
в
и
н
т
а
л
а
м
у
с
а
.
Г
и
п
о
т
а
л
а
м
у
с
в
а
ж
е
н
д
л
я
р
е
г
у
л
и
р
о
в
а
н
и
я
ф
и
з
и
о
л
о
г
и
ч
е
с
к
и
х
и
э
м
о
ц
и
о
н
а
л
ь
н
ы
х
п
р
о
ц
е
с
с
о
в
,
т
е
с
н
о
с
в
я
з
а
н
с
г
и
п
о
ф
и
з
о
м
.
(
И
с
т
о
ч
н
и
к
:
S
t
a
n
d
r
i
n
g
e
t
a
l
.
,
G
r
a
y
’
s
A
n
a
t
o
m
y
,
2
0
0
8
,
р
и
с
.
2
1
.
1
1
.
)
216
Глава 5
сильвиевым водопроводом, а жидкость системы желудочков соответственно
называется цереброспинальной (спинномозговой) жидкостью. Недавно, к боль-
шому удивлению многих ученых, было обнаружено, что в стенках желудочка
находятся стволовые клетки нейронов. Долгое время полагали, что нейроны
во время жизни не могут замещаться новыми клетками, но определенные об-
ласти, такие как гиппокамп и обонятельная поверхность, заменяют свои клетки
так же, как это делают остальные части тела. Полагают, что стволовые клетки
желудочков являются источником таких новых нейронов.
Следующими идут базальные ганглии, если говорить дословно — группа
ядер, находящаяся в основании мозга (рис. 5 .19). Они находятся снаружи от
каждой половины таламуса. Изящная щитовидная структура с каналами, расхо-
дящимися наружу, называется скорлупой. Около каждой из них образует петлю
другое искусное образование, которое называется хвостатым ядром. Эта струк-
тура из «щита и петли» в целом важна для моторного контроля и познания.
Обратите внимание, что базальные ганглии расположены снаружи от таламуса.
Наконец, поверх нижележащих уровней ствола головного мозга — таламу-
са, гиппокампа и миндалин, желудочков и базальных ганглиев — мы можем
водрузить два полушария (рис. 5 .20). Поэтому не нужно удивляться, что при
удалении коры вы увидите глубоко утопленные более древние структуры го-
ловного мозга, появившиеся в образовавшемся углублении.
Как заключение процесса «выращивания» мозга мы покажем вид основа-
ния мозга, чтобы продемонстрировать некоторые области мозга, которые в про-
тивном случае будет трудно увидеть (рис. 5 .21). Здесь вы заметите, например,
зрительный нерв, соединяющий глаза с корой головного мозга.
Рис. 5.18. Миндалины находят-
ся как раз впереди кончика каж-
дой половины гиппокампа. (Ис-
точник: Baars, Fu.)
Рис. 5 .19. Вид базальных ганглиев сбоку с «щи-
том и петлей», образованных скорлупой и хвоста-
тым ядром соответственно. (Источник: Standring,
2005.)
Мозг
217
Итак, вы увидели мозг, «растущий» от древних областей до новой коры в
двух полушариях. Давайте сейчас посмотрим на функциональную значимость
этих областей головного мозга в когнитивной деятельности человека. В этом
обсуждении мы будем придерживаться нисходящего направления, начиная от
двух полушарий и продвигаясь через основные доли, а затем к подкорковым
«спутникам» мозга.
3.0 .
От вопроса «где?» до вопроса «что?»:
функциональная роль областей мозга
Мы уже обсуждали много уровней анализа, с помощью которых можно понять
структуру головного мозга, и показали, где расположены основные области го-
ловного мозга. Сейчас мы опять пройдем головной мозг, начиная от новой коры
и заканчивая стволом головного мозга, и обсудим функциональную роль, кото-
рую эти отделы играют в когнитивной деятельности человека.
Рис. 5 .20 . Полушария мозга воз-
вышаются над стволом головного
мозга и другими подкорковыми
структурами. (Источник: Baars,
Fu.)
Рис. 5 .21 . Вид мозга снизу, по-
казывающий среднюю височ-
ную долю и зрительные тракты.
(Источник: Baars, Fu.)
218
Глава 5
3.1.
Полушария головного мозга:
разделение на левую–правую половины
Две зеркальные половины коры головного мозга веками озадачивают людей.
Почему существуют два полушария? Если у нас только один разум, то почему
мы имеем два полушария? Сэр Чарлз Шеррингтон (Sherrington, 1947) писал:
Собственная личность едина... Она рассматривает себя как одно, другие относятся
к ней как к единой. К ней обращаются как к единой, по имени, на которое она отве-
чает. Закон и Государство записывают ее как одну. И они отождествляют ее с телом,
которое и она, и они (все) считают принадлежащим полностью личности. Короче,
принимаемое без возражений и неоспоримое утверждение состоит в том, что лич-
ность является единой. Логика грамматики это поддерживает местоимением в един-
ственном числе. Все ее многообразие сливается в ее единство.
Философ Рене Декарт, например, был потрясен двойной природой головно-
го мозга. Так как он верил в то, что душа должна быть единым целым, он искал
по крайней мере одну структуру мозга, которая бы была непарной, и в резуль-
тате решил, что это маленькая шишковидная железа (эпифиз) позади ствола
головного мозга. Здесь, как он полагал, и находится душа — приблизительно
то, что мы обозначаем субъективным опытом. К несчастью для Декарта, когда
микроскопы стали достаточно мощными, чтобы изучить крошечный эпифиз
подробно, оказалось, что он также состоит из двух симметричных половинок,
почти зеркальных друг по отношению к другу.
Как же два полушария «общаются» друг с другом? Ответ заключается в су-
ществовании тракта, состоящего из волокон, который идет от переднего до за-
днего конца головного мозга и связывает два полушария.
3.1.1. Мозолистое тело
Полушария большого мозга полностью обособлены и разъединены длинной
бороздой, которая идет между двумя полушариями от передней до задней части
головного мозга. Связь между полушариями обеспечивает мозолистое тело,
большая арка из белого вещества (рис. 5 .22). Количество волокон, идущих меж-
ду двумя полушариями, оценивается больше, чем 100 млн. В мозолистом теле
Рис. 5 .22 . Магнитно-резонанс-
ное изображение, сделанное в
трех измерениях, с вырезанной
частью, показывающей местопо-
ложение мозолистого тела: свода
из белых волокон, простирающе-
гося горизонтально от передней
до задней части головного мозга
и формирующего соединение с
помощью волокон между двумя
полушариями. (Источник: Mark
Dow, Университет Орегоны, с
разрешения.)
Мозг
219
находятся волокна, которые в определенном порядке проходят между полуша-
риями: области передней части соединяются с подобными областями головного
мозга в лобной доле, а области в задней части соединяются со сходными обла-
стями головного мозга в затылочной доле.
Роль двух полушарий в когнитивной, умственной деятельности была объек-
том интенсивного изучения, и мы до сих пор обнаруживаем тонкие и не такие
тонкие отличия в том, какое участие зеркальные полушария принимают в вос-
приятии, речи, мышлении и сознании. Имеются некоторые довольно хорошо
известные различия в двух полушариях, такие как перекрест волокон. Многие
виды обработки сенсорных и двигательных сигналов требуют перехода входя-
щей (чувствительной) и исходящей (двигательной) информации с левой сторо-
ны на правую, и наоборот (рис. 5 .23).
Например, каждый зрительный нерв, приходя из сетчатки, расщепляется
на медиальную половину (по каждую сторону от носа), которая переходит на
противоположную сторону головного мозга, и латеральную (височную) поло-
вину, которая направляется к той же стороне (ипсилатерально). Только обоня-
тельный нерв, который является очень древней сенсорной системой, по пути к
коре остается на той же стороне мозга. Пути исходящего коркового контроля
моторики рук также перекрещиваются, при этом левое полушарие управляет
Рис. 5 .23. Вид двух полушарий сверху. Схематический рисунок двух половин коры
мозга, показывающий некоторые основные функции правого и левого полушарий. Об-
ратите внимание на массивный мост мозолистого тела, соединяющий две стороны. Гла-
за наверху направляются вдоль сходящихся линий для того, чтобы сделать стереоскопи-
ческое восприятие глубины возможным. (Источник: Standring, 2005.)
220
Глава 5
правой рукой, а правое контролирует левую руку (рис. 5 .24). В то время как
левое и правое полушария имеют несколько различающиеся функции, мозоли-
стое тело имеет около 100 млн волокон, постоянно переходящих туда и обратно
для интегрирования информации с обеих сторон. Промежуток времени, за ко-
торый два полушария обработают одну и ту же задачу, может быть очень корот-
ким — 10 мс или одна сотая доля секунды (Handy et al., 2003). Таким образом,
когда великий информационный тракт, мозолистое тело, не поврежден, разница
в функционировании полушарий не очень очевидна. Но когда он перерезан и
используется надлежащий экспериментальный контроль для разделения вхо-
дящих сигналов от правой и левой половин зрительного поля каждого глаза,
неожиданно становятся заметны основные различия между полушариями.
Вопрос о воспринимаемом единстве мира продолжает волновать ученых.
Наиболее замечательным открытием в этом отношении было обнаружение
того, что мозолистое тело может быть разрезано у людей без изменения вос-
принимаемого единства мира и самого себя. Действительно, многие годы та-
кие каллозэктомии (разделение или рассечение мозолистого тела) проводи-
лись, чтобы улучшить состояние при неконтролируемой эпилепсии (рис. 5 .26).
Полное разделение мозолистого тела называется каллозотомией; в настоящее
время более часто проводят частичное рассечение только тех областей двух по-
лушарий, от которых при приступе эпилепсии распространяется активность.
Такое частичное иссечение называется каллозэктомией. Доктора и их пациенты
верили, что рассечение около 100 млн волокон в мозолистом теле не приведет
вообще к заметному эффекту! Это яркая иллюстрация способности мозга при-
спосабливаться к довольно серьезным повреждениям — дополнять отсутству-
ющие детали мира, который мы ощущаем, например, посредством движений
глаз. Однако более глубокие исследования показали, что полное рассечение
мозолистого тела имеет неуловимые, однако длительные эффекты, и таким об-
разом, предпочтительным считается частичное рассечение мозолистого тела.
Рис. 5 .24. Модель контроля коры над областями тела. Обратите внимание, что сенсор-
ные пути и корковые пути контроля двигательных функций пересекаются в головном
мозге. Простые рефлексы не пересекаются, а координация требует взаимодействия
между обеими сторонами. (Источник: Standring, 2005.)
Мозг
221
Синестезия
Представьте мир пурпурных вторников, вкусов, которые имеют форму, и волни-
стых симфоний зеленого цвета. Из сотни людей один, во всем остальном обыч-
ный человек ощущает мир так. Такое состояние называется синестезия — слияние
различных форм чувствительного опыта. Например, голос или звук в музыке не
только слышится, но также видится, имеет вкус или ощущается как прикосновение.
Синестезия — это слияние различных сенсорных ощущений: прикосновение к на-
ждачной бумаге может вызвать ощущение звука фа-диез в музыке, симфония может
слышаться в синих и золотых красках, а представление о феврале будет восприни-
маться как расположение над чьим-то правым плечом. Большинство синестетов
(людей с синестезией) не знают, что их ощущения в любом случае необычны.
Существует много вариантов синестезии, и если вы имеете один из них, у вас вы-
сокая вероятность иметь также другой или третий вид. Восприятие дней недели в
цвете является наиболее распространенным типом синестезии, за которым следу-
ют цветные буквы и цифры. Другие нередко встречающиеся типы также включают
ощущения вкуса слов, цветное восприятие на слух, линии чисел, воспринимаемые
в трех измерениях и персонификацию букв и цифр.
Синестетическое восприятие непроизвольно, автоматично и обычно постоянно во
времени. Более того, при таком восприятии, часто врожденном, появляется ощуще-
ние простого цвета, формы или структуры, а не результата мыслительной ассоци-
ации. Синестетики не говорят: «Эта музыка заставляет меня чувствовать себя как
ваза цветов на ресторанном столике». Это с ними как раз происходит.
Вероятно, синестезия является результатом увеличенных перекрестных связей
между сенсорными областями мозга, подобно соседним странам с проницаемыми
границами. Синестезия завораживала как непрофессионалов, так и ученых своим
богатством объединения чувств, но только в настоящее время стало, наконец, по-
нятно функционирование мозга при изучении этих удивительных людей.
Хотя синестезия исследовалась в экспериментах по поведению и с помощью об-
работки нейроизображений, ее генетическая основа остается неивестной. Наша ла-
бораторная группа выяснила, что синестезия является идеальным состоянием для
генетического анализа по трем причинам: (1) синестезия встречается в семейных
группах и, по-видимому, наследуется; (2) синестетическое восприятие является
результатом возрастающего количества перекрестного обмена сигналами между
областями нейронов, что предполагает набор генов-кандидатов; и (3) комплекс те-
стов, разработанных в нашей лаборатории, позволяет уверенно выявлять действи-
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
Рис. 5 .25. Дэвид Иглмэн (David
Eagleman, PhD), факультет невроло-
гии, медицинский колледж Бэйлора,
Хьюстон, штат Техас, США
222
Глава 5
тельную синестезию для проведения
исследований, а не только людей со
свободными ассоциациями.
Таким образом, мы проводим широко-
масштабное исследование, называе-
мое анализом семейных связей, чтобы
картировать гены, которые связаны с
цветовой синестезией. К настоящему
времени разработана система тестов,
которые безусловно определяют сине-
стетов и отличают их от контрольной
группы. Эти тесты выложены открыто
для научного сообщества (www.synes-
thete.org). Несколько семей с много-
кратными синестезиями предоставили
свои родословные, и мы получили об-
разцы ДНК от более чем 100 людей в
этих семьях. Полногеномное исследо-
вание выявляет наиболее вероятную
генетическую область, ответственную
за синестезию в этих семьях. Понима-
ние генетических основ синестезии
вносит вклад в понимание, каким об-
разом в норме обеспечивается связь в
головном мозге. Это показывает, что
возможны более чем один тип мозга и
один тип мышления.
Синестезия присутствует у одного из
нескольких сотен людей, что делает ее
намного более распространенной, чем
обычно полагают, и намного более
важной с научной точки зрения, неже-
ли простое любопытство. Другие факты свидетельствуют, что мы в какой-то степе-
ни все можем быть синестетиками, но большинство из нас не осознает сращение
сенсорных сигналов, которые имеются в наших головах.
3.2 .
Исходящие и входящие сигналы:
подразделение в направлении спереди назад
Кора является складчатым слоем серого вещества, который в расправленном
виде имеет приблизительный размер 60 × 60 см. Чтобы уместиться внутри
черепа, кора головного мозга собрана в складки (извилины и борозды). Кора
имеет четыре доли, которые видны снаружи, и две большие области, которые
не видны. Перед тем как обсуждать функции этих областей, взглянем на дру-
гой основной способ подразделения мозга: разделение коры головного мозга
в лобно-затылочном направлении. Для того чтобы понять такое подразделе-
ние, вам понадобится умение определять местонахождение в головном мозге
некоторых ориентиров. Попробуйте найти на рис. 5 .27 центральную борозду,
которая идет вертикально между лобной и височной долями. Для того чтобы
Рис. 5 .26 . При обычной форме синесте-
зии месяцы и дни месяца могут иметь
как цвета, так и специфическое размеще-
ние в пространстве. (Источник: Cytowic,
Eagleman, 2009: Wednesday is Indigo
Glue: Discovering the Brain of Synesthesia.
Cambridge: MIT Press)
Мозг
223
определить ее местонахождение, найдите область, помеченную как «первичная
соматосенсорная область». Центральная борозда как раз находится впереди от
нее. Также нужно найти второй ориентир, Сильвиеву борозду (в русском языке
более употребителен термин «латеральная борозда». — Прим. перев.) . Она идет
более или менее горизонтально от лобной доли по направлению назад, отделяя
височную долю от теменной и лобной долей.
Сенсорные, или входящие, области коры расположены позади от централь-
ной и латеральной борозд в теменной, височной и затылочной долях. Эти доли
содержат зрительную кору, слуховую кору и соматосенсорную кору, в которых
обрабатывается информация, приходящая от глаз, ушей и тела. Например, зри-
тельная кора начинается в затылочной доле, но переходит на теменную и височ-
ную доли. Слуховая кора расположена в височной доле, но также простирается
до теменной доли. Соматосенсорная область находится в теменной доле. Поля,
отвечающие за вкус и обоняние, расположены в основании теменных долей.
Эта обширная область, расположенная в задней части мозга, охватывающая три
доли коры головного мозга, — не просто место обработки сенсорной информа-
ции. Она также является областью для обработки ассоциативных данных, где
информация от разных чувств «связывается вместе» для обработки высшего
порядка. Задумайтесь о том, как мы смотрим фильмы, — эти ассоциативные
зоны помогут вам понять, как соотнести то, что вы слышите, с тем, что вы ви-
дите на экране. Большая часть процессов такого рода происходит в теменной
Рис. 5 .27. Изображение функциональных областей некоторых сенсорных полей коры.
Центральная борозда отделяет лобную долю от теменной. Сразу сзади от центральной
борозды находится первичная соматосенсорная область. Сильвиева борозда также на-
зывается латеральной бороздой. (Источник: Squire et al., 2008.)
224
Глава 5
доле, и мы будем обсуждать эту важную структуру более подробно в следу-
ющем разделе. Самые большие ассоциативные области имеются у приматов,
и самые большие из них — у людей.
Двигательные, или исходящие, области коры расположены в лобной доле,
впереди от центральной и латеральной борозд. Взгляните опять на рис. 5 .27
и найдите область, обозначенную «первичная соматосенсорная область» поза-
ди центральной борозды. Хотя она и не отмечена на рисунке, первичная двига-
тельная область находится в лобной доле по ту сторону центральной борозды,
как раз впереди от соматосенсорной области в теменной доле. Близкие физиче-
ские связи между соматосенсорной и двигательной корой обусловливают тес-
ное взаимодействие между чувствами осязания, давления и боли и двигатель-
ной или моторной системой. В сущности имеется сложная карта тела, которая
построена похожим образом в соматосенсорной области, расположенной как
раз позади от центральной борозды, и в соответствующей двигательной обла-
сти, расположенной точно впереди. Такая сложная карта является изображе-
нием частей тела: разные области тела в этих областях коры представлены не-
одинаковым образом; некоторые области, такие как лицо и кисть руки, имеют
довольно непропорционально большое представительство, а другие области,
такие как центр спины, — непропорционально маленькое. Представьте, на-
сколько чувствительнее ваши подушечки пальцев к прикосновению, давлению
и боли, чем, например, поясница. Такая карта представлений в коре отражает
эту отличающуюся чувствительность. Имеется две карты тела: одна в сомато-
сенсорной коре, а очень похожая — в двигательной коре (рис. 5 .28 и 5.29).
Эти две карты тела или гомункулуса («маленького человечка») были впер-
вые открыты первым нейрохирургом Уайлдером Пенфилдом в монреальском
университете в 1950–60-е гг. Группа Пенфилда первой стимулировала кору
бодрствующих пациентов, что стало возможным потому, что поверхность коры
головного мозга не содержит болевых рецепторов. Таким образом, местной
анестезии, примененной в месте надреза, было достаточно, чтобы приглушить
боль от удаления кожи головы, и хирурги могли электрически стимулировать
обнаженную поверхность коры и спрашивать бодрствующих пациентов о том,
что они чувствуют в результате стимуляции. Их открытия в значительной
степени выдержали проверку временем. Исследования путем электрической
стимуляции были необходимы по медицинским показаниям для того, чтобы
узнать, в каком месте оперировать мозг, чтобы минимизировать повреждения
функциональных областей пациентов. В случае соматосенсорного гомункулу-
са (соматической чувствительности) местная стимуляция пробуждает чувства
прикосновения в соответствующей части тела. Стимуляция двигательного го-
мункулуса вызывает определенные движения тела, но интересно, что пациен-
ты отрицают то, что они совершали эти движения сами. Когда Пенфилд спра-
шивал: «Двигаете ли вы своей рукой», в то время как стимулировал область
руки, пациент мог сказать: «Нет, доктор, вы двигаете моей рукой». Однако если
хирург передвигался, например, на сантиметр вперед к премоторному участ-
ку, стимуляция вызывала намерение движения собственным телом, сообщен-
ное пациентом, чувства внешнего контроля движений при этом не возникало.
В этом заключается существенная разница, к которой мы вернемся позднее.
Мозг
225
Рис. 5 .28. Изображение
соматосенсорного гомун-
кулуса, демонстрирую-
щее представительство
областей тела в коре го-
ловного мозга. Обратите
внимание, что некоторые
области тела, такие как
лицо, имеют диспро-
порционально большее
представительство, чем
другие области, такие
как туловище. (Источ-
ник: Standring, 2005.)
Рис. 5 .29 . Изображение
двигательного гомунку-
луса, демонстрирующее
представительство
об-
ластей тела в моторной
коре. Заметьте, что не-
которые области, такие
как лицо, имеют диспро-
порционально большее
представительство, чем
другие области, такие как
туловище.
(Источник:
Standring, 2005.)
226
Глава 5
Необходимо подчеркнуть, что центральная борозда является важным ори-
ентиром, который нужно знать. Она не только разделяет соматосенсорный и
двигательный гомункулусы (можно сказать, проекции тела на чувствительную
и двигательную область коры головного мозга), но, в более широком смысле,
центральная борозда отделяет сенсорную половину коры (сзади) от лобной по-
ловины (впереди). Задняя часть коры содержит области проекций основных ор-
ганов чувств: зрения, слуха, осязания, обоняния и вкуса. Напротив, передняя
часть включает регуляцию действий, планирование, некоторые функции рабочей
памяти, речь и им подобное. В известной степени задняя половина имеет дело с
воспринимаемой действительностью, в то время как передняя половина пытает-
ся предсказать и контролировать будущее.
3.3 .
Основные доли: видимые и скрытые участки
Мы уже использовали для мозга аналогию с географией. В таком контексте ос-
новные доли головного мозга можно рассматривать в терминах географии как
большие континенты. В то время как каждая из долей отделена от другой и име-
ет свои частные функции и анатомические особенности, каждая является также
частью целого головного мозга и, таким образом, объединена и тесно связана с
общей функцией головного мозга. Четыре «континента» мозга показаны на рис.
5.30 и включают лобную, теменную, височную и затылочную доли. В этом раз-
деле обсуждаются их функциональные роли в когнитивной деятельности. Две
других важных области, невидимые при осмотре мозга снаружи, также играют
важную роль в познании, и мы их тоже опишем.
3.3.1 . Лобная доля
Массивная лобная доля — место планирования действий и исходящих двига-
тельных сигналов. Как уже упоминалось, двигательные области тесно связаны
с соматосенсорной областью, которая имеет схожий гомункулус. Такие двига-
тельные функции, которые присутствуют в мозге человека, подобным же об-
разом организованы в мозге большинства млекопитающих. Но лобная доля у
человека намного обширнее, чем у остальных приматов или любого другого
существа. Какие еще функции выполняет лобная доля и каким образом ее роль
является уникальной для человека?
Лобную долю назвали «органом цивилизованности» (Luria, 1966). Области
лобной доли, которые заслужили это название, находятся в основном в пре-
фронтальной коре. Префронтальная кора расположена на медиальной, лате-
ральной и орбитальной поверхностях наиболее передней части лобной доли
(рис. 5 .31).
Префронтальная кора не является двигательной частью лобной коры. Об -
ратите внимание, что префронтальная кора — наиболее передняя часть лобной
коры. Термин «префронтальный» немного вводит в заблуждение, так как оз-
начает «впереди лобной коры». Явных маркеров границ для префронтальной
области не существует, границы ее установлены по местоположению выростов
таламуса. Тем не менее префронтальная доля, возможно, является наиболее
«познавательной» частью мозга. Префронтальная кора — большая область
коры, занимающая примерно одну треть от поверхности целой коры. В чем же
специализируется префронтальная кора и почему эта часть мозга человека так
уникальна?
Мозг
227
Рис. 5 .30 . Основные направления в мозге. Так как человеческий мозг повернут на 90°
по отношению к спинному мозгу (что отличает нас от большинства млекопитающих и
рептилий), у него имеется два способа разметки. Дорсальное направление также назы-
вается верхним, вентральное также называется нижним, ростральное приблизительно
значит то же самое, что передний, а каудальное иногда называется задним. Для упро-
щения используйте обычные термины, такие как передний, задний, верхний и нижний.
(Источник: Standring, 2005.)
Рис. 5 .31 . Слева: карта активности, построенная на трехмерном магнитно-резонансном
изображении, показывающая области медиальной префронтальной коры. Справа: как
отыскать префронтальную кору. В целом лобная кора находится впереди от централь-
ной борозды, вертикальной складки, которая идет от вершины коры вниз до височной
доли. Найдите центральную извилину на этом рисунке. Две пурпурных извилины сразу
впереди от центральной борозды называются моторной и премоторной корой. Крас-
новато-пурпурное пятно впереди от них называется добавочной двигательной корой.
Однако три закрашенных желтым цветом пятна в передней трети всей коры называют-
ся префронтальной корой, и ее часто считают наиболее «когнитивной» частью мозга.
(Источник: Harenski, Hamann, 2006.)
228
Глава 5
Префронтальная кора особенно необходима для:
• инициации движений;
• планирования;
• удержания необходимой информации в состоянии готовности для ис-
пользования (одна из функций рабочей памяти);
• перестройки мышления от одной линии мышления к другой;
• отслеживания эффективности своих действий;
• определения и разрешения конфликтных планов действий;
• подавления неэффективных или обреченных на провал планов и дей-
ствий.
Список показывает, какой важной для человеческой когнитивной деятель-
ности является префронтальная кора. Многие анатомы полагают, что префрон-
тальная кора является самой большой у человека и, таким образом, отличает
наш вид от остальных приматов. К тому же префронтальная кора имеет обла-
сти, отвечающие за эмоциональные и личностные действия, так же как и за со-
циальное познание — знания, как себя нужно вести, например. На латеральной
извилине, вклиниваясь между дорсолатеральной стороной префронтальной и
вентральной частями премоторной коры, находится центр Брока. Эта область
служит посредником, связующим звеном абстрактных словесных выражений
в языке, уникальной функции человека.
Лобная доля намного больше у человека, чем у других приматов, и она при-
обрела много новых функций и процессов, которые связаны с такими чело-
веческими действиями, как речь, мышление и исполнительный контроль над
процессами более высокого порядка. Вторая доля, которая также развивается
намного больше у людей, — это теменная доля. Какие функции она выполняет,
мы увидим в следующем разделе.
3.3.2. Теменная доля
Как мы отмечали ранее, передняя область теменной доли содержит соматосен-
сорную кору. Однако у людей теменная доля — это не только саматосенсорная
область, почти как лобная доля — это не только двигательная часть. Одними из
важных функций теменной доли являются многочисленные карты простран-
ства тела. Что точно означают слова «карта тела»? Вообразите, что вы сидите
на стуле за столом и глядите вниз на свои руки. Ваши глаза приносят сенсорные
входящие сигналы к вашему мозгу о том, где находятся ваши руки по отноше-
нию к вашему телу, но имеются и другие сигналы, также показывающие вам,
где находятся руки (вот поэтому вы знаете, где находятся ваши руки, даже если
ваши глаза закрыты). Ваше воображаемое местоположение рук будет вашим
собственным соответствием или эгоцентрическим соответствием. Теперь вооб-
разите друга, сидящего напротив вас, и представьте, где находятся ваши руки
с его точки зрения. Как вы это делаете? Это легко выполнять, и области те-
менной доли — именно то место, где происходит этот вид обработки сигналов
(рис. 5 .32).
Позади и внизу от соматосенсорнй области находится зона, которую назы-
вают нижней теменной областью, или НТО. Ее функциональное значение до
сих пор изучается, однако полагают, что это место интеграции информации от
органов чувств.
Мозг
229
3.3.3 . Височная доля
Височная доля — область, где обрабатываются звуковые сигналы, и неудиви-
тельно, что это также область, где расположены системы восприятия речи на
слух и понимания речи. Слуховая кора расположена на верхних участках ви-
сочной области и внутри сильвиевой борозды. Как раз позади слуховой коры
находится центр Вернике, центр понимания речи. Но височная доля не только
область обработки звуковых сигналов и речи. Считается, что средние части ви-
сочной доли содержат абстрактные образы для семантических знаний. Более
нижние и задние части височной доли приводят в более тонкое соответствие
зрительные объекты и включают веретенообразную извилину, ответственную
за узнавание лиц людей.
3.3.4. Затылочная доля
В затылочной доле, самой задней области коры головного мозга, находится
зрительная кора. Большая часть зрительной коры спрятана внутри шпорной
борозды (sulcus calcarinus). Система зрительного анализатора занимает боль-
шую зону внутри затылочной доли, которая простирается вперед до теменной
Рис. 5.32 . Схематическое изображение некоторых мультисенсорных функций темен-
ной коры (функций теменной коры, затрагивающих несколько органов чувств). Вид и
звук колокольчика объединяются нейронами теменной коры, используя «карту» про-
странства, окружающего тело (эгоцентрическое пространство). (Источник: Beauchamp,
2005.)
230
Глава 5
и височной долей. Новые технологии предоставляют возможность сделать эти
области коры плоскими, чтобы сдвинуть эти поля и дать возможность увидеть
функциональные зрительные поля, которые в норме спрятаны внутри шпорной
борозды и которые трудно увидеть на томограмме.
3.3.5. Островок и сильвиева борозда
Как большое дерево, кора головного мозга выросла и накрыла сверху обшир-
ные части самой себя, как мы можем увидеть, если математически выпрямим
кору и растянем ее в плоскую поверхность. Две области, которые скрываются
обширной корой, особенно важны: островок и сильвиева борозда. Если височ-
ную долю осторожно отделить от остальной коры, появляется новый спрятан-
ный «мир». Эта область называется «центральная доля», или «островок», пото-
му что она выглядит как отдельный островок коры (рис. 5 .33 и 5.34). Островок
не всегда описан в популярных работах о головном мозге, но он включает сот-
ни миллионов нейронов и довольно обширное пространство поверхности коры
головного мозга. Неврологические данные предполагают, что она включена в
такие «инстинктивные чувства», как сознательные чувства отвращения и омер-
зения. Но островок настолько обширен, что, вероятно, у него множество функ-
ций. Вполне допустимо, что интерорецепция — ощущение внутренних орга-
нов, может быть одной из основных ролей этого тайного островка коры.
Один из исследователей предполагал, что у человека правый передний
островок может служить основанием формирования субъективной картины ма-
териальной личности как чувствующего (сознательного) существа, т. е. эмоци-
онального понимания» (Craig, 2005).
Сильвиева борозда — очень большая борозда, которая идет в приблизитель-
но горизонтальном направлении от лобной доли между теменной и височной
долями, и заканчивается около места объединения теменной, височной и за-
тылочной долей. Анатомия борозды в значительной степени различается у раз-
ных людей, а также в разных полушариях. Внутри сильвиевой борозды на верх-
них стенках верхней височной извилины спрятана верхневисочная плоскость.
Рис. 5 .33 . Настоящий человеческий мозг с выделенным островком, который обычно
спрятан внутри височной доли. (Источник: Standring, 2005.)
Мозг
231
Эта область называется плоскостью, потому что это отчасти плоский край
коры, который вдается с боковой поверхности в срединные области. Верхневи-
сочная плоскость является местоположением первичной и вторичной слуховой
коры, а также части поля Вернике, важного для понимания речи. В более верх-
них областях сильвиевой борозды, расположенных рядом с теменной долей
и противоположных верхневисочной плоскости, находится соматосенсорная
кора, которая заворачивается вокруг и под верхней частью борозды.
3.3.6 . Медиальная височная доля
Медиальная височная доля (МВД) является в сущности частью височной доли,
но ее строение и функции поразительно отличаются, и обычно к ней относятся
как к отдельной структуре. В МВД находятся гиппокамп и родственные ему
области, которые связаны с функциями памяти (рис. 5 .35). В МВД находятся
многие области, включая часть, называющуюся лимбической областью. Слово
«лимбический» обозначает «граница», и соответственно ее названию ведется
много споров о точных границах этой области. Время от времени можно ви-
деть название «лимбическая система» для комплекса гиппокампа, миндалины
и лимбической коры. Все эти термины имеют свое применение, и очень важно
знать, что именно имеется в виду.
Верхняя дуга (рис. 5 .36) называется поясной извилиной (слово «cingulum»
обозначает пояс или кушак, как в «подпруге»). Передняя половина этой области
обычно активируется во время решения задач, которые включают конфликтую-
щие стимулы или ответы, что является очень важной стороной функции управ-
ления. В классическом опыте Струпа (Stroope), например, имеется противоре-
Stroope), например, имеется противоре-
), например, имеется противоре-
чие между цветом слов и значением тех же самых слов. Передняя половина
Рис. 5 .34 . Вид левого полушария с удаленным срезом, открывающим островок, кото-
рый невидим при взгляде сбоку. «Островок» означает «остров» из-за его появления,
когда головной мозг разрезан. (Источник: Standring, 2005.)
232
Глава 5
поясной извилины каким-то образом вовлечена в определение и разрешение
таких конфликтующих стимулов.
Нижняя арка лимбической доли первоначально является частью обонятель-
ного мозга, обонятельной корой, и поэтому называется периринальной корой
(peri означает вокруг или около, а rhinal — обонятельный). Вспомните, что
мы ранее установили, что не вся кора имеет шесть слоев, а только развитая
кора млекопитающих, которая поэтому называется «новая кора», или неокор-
текс (новая кора, потому что она возникла только 200 млн лет назад). Более
древние области коры имеются у рептилий, у ящериц, например такие, как
лимбическая кора. Эта область имеет пять слоев, и иногда на нее ссылают-
ся как на древнюю кору («палеокортекс»). Она часто связана с эмоциями и
памятью, а в случае верхней арки лимбической системы — с принятием ре-
шений и анализом конкурирующих импульсов. В дополнение к тому, лимби-
ческая кора тесно связана с гиппокампом и миндалиной, которые спрятаны
внутри височной доли и, таким образом, невидимы при взгляде с медиальной
стороны. Недавнее исследование показало очень тесную взаимосвязь между
этими древними областями коры и кратковременной памятью, т. е . памятью
об опыте, находящемся под контролем сознания. Это древний мозг рептилий,
который, однако, до сих пор остается жизненно важным центром деятельности
человека и других млекопитающих.
Рис. 5.35. Медиальная височная доля (МВД) — область, находящаяся по средней линии
и видимая со стороны основания головного мозга. Это древний «обонятельный мозг»,
над которым сейчас располагается массивная «новая» кора высших млекопитающих.
Таким образом, его трудно увидеть снаружи, но он до сих пор сохраняет многие необ-
ходимые функции, включая кодирование сознательных событий в эпизодические вос-
поминания. (Источник: Buckley, Gaffen, 2006.)
Мозг
233
Рис. 5 .36 . Медиальная височная доля и поясная извилина (зеленая верхняя петля) вид-
ны на срезе мозга с медиальной стороны. Гиппокамп отмечен пурпурным цветом, а
миндалина — оранжевым. Они практически вмурованы внутрь височной доли. (Источ-
ник: Heimer, Van Hoesen, 2006.)
3.4 .
Обширные взаимосвязи между корой
и таламусом
В то время как можно рассматривать доли мозга как «континенты» мозга,
процессы в них тем не менее сложным образом переплетены не только друг
с другом, но также с подчиненными им подкорковыми областями. Миллиар-
ды аксонов отходят от тел клеток серого вещества коры головного мозга: во-
локна для передачи исходящих сигналов нервной клетки и дендриты, которые
обеспечивают входящие электрические сигналы к каждой клетке. Если белые
клетки опоры, называющиеся миелиновыми клетками, обертывают каждое из
этих волокон, для невооруженного глаза они выглядят как белая масса, и по-
этому их называют белым веществом. В целом огромная структура коры голов-
ного мозга имеет во многом форму, похожую на стадион «Супердоум» (самая
крупная крытая арена в США на 1975 г.), каждая из двух гигантских половин
которого заполнена миллиардами проводов, идущих во все направления, центр
которых находится в таламусе, который находится в середине каждого полуша-
рия (рис. 5 .37). С каждой стороны две половины коры головного мозга вместе
с центром переключения сигналов, расположенным в таламусе, составляют не-
обыкновенную биологическую структуру.
О таламусе можно говорить как о ретрансляционной станции: почти все
входящие сигналы останавливаются в таламусе на их пути к коре головного
мозга, а почти все исходящие сигналы также останавливаются в таламусе на
своем пути к мышцам и железам.
Волокна от клеток коры головного мозга распространяются по всем направ-
лениям, идя горизонтально к соседним клеткам, собираясь в огромные пучки
на своем пути к отдаленным участкам коры, сходясь на своем пути к нижележа-
щим участкам в большом «транспортном распределителе», таламусе, от каждой
234
Глава 5
половины коры головного мозга. В дополнение к этому, сотни миллионов аксо-
нов идут в поперечном направлении от одного полушария до другого, создавая
белые аксонные мосты, называемые комиссурами (рис. 5 .38). Самое большое
соединение из поперечных волокон называется мозолистым телом (corpus cal-
corpus cal-
cal-
cal-
losum). Если разрезать головной мозг прямо по средней линии, можно увидеть
мозолистое тело в форме изогнутой белой дуги. Белый цвет, как говорилось
раньше, появляется из-за миелина, окружающего корковые аксоны, которые об-
разуют большой мост, соединяющий два полушария.
Рис. 5 .37. Кора головного мозга и таламус: исключительно единая система. Схематиче-
ский рисунок, показывающий карту связей с цветовым кодом от таламуса до областей
коры. (Источник: Standring, 2005.)
Мозг
235
В конце концов корковые сенсорные и моторные проводящие пути состав-
ляют основные входящие и исходящие пути головного мозга (рис. 5 .39). Все
эти пути идут от основания мозга. Сенсорные и двигательные пути можно раз-
делить на две группы. Одна группа идет через небольшие отверстия в череп-
ной коробке, верхней части черепа, и поэтому называется черепно-мозговыми
Рис. 5 .39 . Белые пучки миелинизированных аксонов идут по всем направлениям по
сводам больших полушарий. (Источник: Mario Lazar, с разрешения.)
Рис. 5 .38 . Схематический рисунок связей в головном мозге, показывающий основные
системы волокон. (Источник: Standring, 2005.)
236
Глава 5
нервами. Они включают зрительный нерв, идущий от задней части глазниц,
слуховой, обонятельный и языко-глоточный нервы, так же как нервы, прово-
дящие осязание и болевое ощущение от области лица и ротовой полости; и
двигательные нервы, обслуживающие выражение нашего лица, голосовой ап-
парат, рот, язык и так далее. Вторая группа путей идет в спинной мозг и кон-
тролирует функции нашего тела, как сознательные — движения туловища, рук
и ног, так и вегетативные — кровяное давление и потоотделение. Со стороны
входящих сигналов чувствительные нервы от тела дают нам всю информацию,
как сознательную, так и бессознательную, которую мы получаем от тела. Так
как эти проводящие пути являются сложными при детализации, их обзор по-
мещен далее.
Традиционно между названиями соединяющихся участков головного мозга
ставится буква «о» (это не общепринято для английского языка в отличие от
русского), поэтому нужно говорить о таламо-кортикальных связях. Путь, иду-
щий от коры к таламусу, называется кортико-таламическим, и верите вы или
нет, движение по нейронам может даже быть кортико-таламо-кортикальным.
Это покажется немного менее сложным, если вы будете думать об этом как
о транспортном потоке в большом городе или даже как о всемирной паутине,
соединяющей миллионы компьютеров с помощью крупных узловых центров и
магистралей.
3.5 .
Подчиненные участки подкорковых слоев
Из-за того что кора человека так огромна, она покрывает важные подкорковые
органы, которые действуют в качестве сателлитов (подчиненных участков) по
отношению к коре, постоянно взаимодействуя с ней. Эти подкорковые струк-
туры совсем не выглядят так, как обычно представляют мозг: это гигантские
скопления нейронов, часто называемые «ганглии», или «ядра». Подкорковые
структуры часто имеют удивительно изящную форму: петли, рога и яйцевид-
ного овала.
Структуры-сателлиты особенно важны в когнитивной нейробиологии. Тала-
мус, часто называемый воротами к коре, был описан выше; две части таламуса
находятся в самом центре мозга, по обе стороны от средней линии (поэтому
их нельзя в действительности увидеть на поперечном срезе). Таламус также
связан с разными корковыми областями, поэтому имеются важные кортико-та -
ламо-кортикальные замкнутые цепи, которые, как было показано, играют роль
в процессе внимания и других когнитивных функциях более высокого порядка.
Таламус находится над стволом мозга и как раз под корой, что является отлич-
ным местоположением, чтобы выполнять свою функцию ретрансляционной
станции головного мозга.
Комплекс гиппокампа (см. рис. 5 .17) необходим для запоминания сознатель-
ного опыта и имеет форму двух маленьких «колбасок», замурованных в височ-
ную кору с каждой стороны. Однако в настоящее время известно, что области,
соседние к «колбаскам» гиппокампа, также нужны для эпизодической памяти.
Исходя из этого мы, скорее, будем говорить о комплексе гиппокампа в целом,
а не только о гиппокампе.
На самом переднем конце гиппокампа с каждой стороны находится мин-
далина, amygdala (см. рис. 5 .18). Она имеет небольшую шаровидную форму,
Мозг
237
похожа на орех, и очень важна для таких эмоций, как страх или гнев, так
же как для процессов обучения, которые затрагивают эти эмоции. В заклю-
чение упомянем базальные ганглии (см. рис. 5 .19) — сложные структуры,
имеющие форму диска и петли, находящиеся снаружи от каждой полови-
ны таламуса, а также мозжечок (или малый мозг), находящийся сверху над
всей верхней частью ствола головного мозга и таламусом. Базальные ган-
глии включены в функции планирования и бессознательные познавательные
действия. Однако новые данные связывают базальные ганглии с когнитив-
ной деятельностью высшего порядка, такой как расшифровка грамматики,
синтаксиса и языка.
Мозжечок находится позади нижележащей части ствола головного мозга.
Он является сам по себе очень большой структурой. У многих млекопитающих
мозжечок имеет столько же нейронов, сколько и кора, хотя с более короткими
аксонами. Большинство нейронов мозжечка соединены локально в небольшие
группы. Ранее мозжечок считался в основном вовлеченным в процесс контро-
ля тонких движений, таких как движений пальцев машинистки или музыкан-
та. Сейчас также известно, что он необходим также для когнитивных функ-
ций. В действительности функциональная томография показала, что мозжечок
«подсвечен» при почти любой познавательной задаче. Причина этого еще не
полностью понятна.
В заключение необходимо упомянуть некоторое количество крошечных
ядер ствола головного мозга и основания переднего мозга, которые широко
рассылают клеточные волокна по всему переднему мозгу. Эти нейромодули-
рующие ядра иногда неформально называются «брызгалками», потому что они
распыляют нейромедиаторы из окончаний своих аксонов так, что эти вещества
широко распространяются. Они могут содержать только несколько тысяч ней-
ронов, но являются ключевыми для нормального функционирования мозга. Ос-
новные нарушения, характеризующиеся тремором мышц, при котором человек
становится инвалидом, такие как болезнь Паркинсона, являются результатом
нехватки таких нейромодуляторов. Они также контролируют суточный цикл
сон-бодрствование.
Мы завершаем этот раздел описанием ретикулярной формации, располо-
женной в центре мозга (рис. 5 .40). Это особенно увлекательная область голов-
ного мозга с точки зрения ее роли в сознательном опыте человека. Ретикулярная
формация называется «ретикулярной» (т. е . похожей на сеть), так как аксоны
нейронов этой системы особенно короткие, что предполагает особо тесное вза-
имодействие между соседними нейронами. Кроме того, она получает входящие
сигналы от всех сенсорных и двигательных систем, так же как от других ос-
новных структур головного мозга. Посредством своих связей с таламусом она
может посылать и получать информацию от всех участков коры.
Что это говорит о роли ретикулярной формации в сознательном опыте? Име-
ются нейрофизиологические данные, что специальные системы в головном моз-
ге могут сотрудничать и конкурировать за доступ к центральной интегративной
«информационной доске». Имеется основание полагать, что расширенная ре-
тикулярно-таламическая система (РРТС) (видимо, наиболее распространенный
русский аналог — активирующая система ретикулярной формации — Прим.
перев.) соответствует такой «информационной доске».
238
Глава 5
Не новость, что «чувство общего» Аристотеля было предложено для обо-
значения области интеграции между различными чувствами. В действитель-
ности анатомы, которые исследовали ретикулярную формацию, указали на ее
сходство с этим понятием у Аристотеля. Шайбель и Шайбель (Scheibel, Schei-
(Scheibel, Schei-
Schei-
Schei-
bel, 1965) писали: «Анатомические исследования Конштамма (Kohnstamm) и
Куэнсела (�uensel), которые предполагали объединение некоторого числа аф-
�uensel), которые предполагали объединение некоторого числа аф-
, которые предполагали объединение некоторого числа аф-
ферентных и эфферентных систем в коре ретикулярной формации, привели
их к предположению, что эта область является «рецепторным центром 2», или
«чувствительным центром», — общей совокупностью чувствительных нейро-
нов осевой части ЦНС».
Более того, эти же авторы заметили, что «... центральная часть ретикулярной
формации является посредником между специфическим предотвращением рас-
пространения сигналов от центра и согласованным подавлением тех сенсорных
входящих сигналов, которые на время снизили свою сенсорную роль». Подоб-
ным образом, Гасто (Gastaut, 1958) описывает ретикулярную формацию ствола
головного мозга как «область конвергенции..., в которой сигналы собираются
перед тем, как будут перераспределены другим образом по пути в кору». Таким
образом, различные по содержанию сенсорные сигналы могут подавлять друг
друга, как мы и можем в действительности ожидать от входящих сигналов во
всеобщее рабочее пространство. Это предполагает, что различные специализи-
рованные процессы могут конкурировать за доступ к РРТС.
Рис. 5 .40 . Восходящая активирующая ретикулярная система находится в стволе голов-
ного мозга и таламусе и посылает проекции по всей коре. Ретикулярная формация (РФ)
считается необходимой для нормального бодрствующего состояния сознания. (Источ-
ник: Filley, 2002.)
Мозг
239
Как это понятие об «информационной доске» в действительности работает
на языке терминов нейронных процессов и как распространяются послания?
По одной возможной гипотезе, входящий сигнал в РРТС дает одна сенсор-
ная проекционная область коры. Если этот входящий сигнал преобладает над
другими конкурирующими входящими сигналами, он становится всеобщим
сообщением, который широко распространяется по другим областям мозга,
включая оставшуюся часть коры головного мозга. Таким образом, один избран-
ный входящий сигнал в РРТС усиливается и распространяется за счет других.
Следовательно, РРТС лежит в основе функции «всеобщего распространения»
сознания, в то время как выбранное «обрабатывающее устройство» (processor)
восприятия в коре задает специфическое содержимое сознания, которое долж-
но быть распространено.
Какова роль РРТС в сознательном мышлении? Это может быть как раз тем
случаем, в котором любая деятельность коры должна инициировать «под-
держку» РРТС в циркулирующем потоке информации перед тем, как она будет
распространена по всему объему и станет сознательной (например, Scheibel,
Sceibel, 1965; Shevrin, Dickman, 1980). Диксон (Dixon, 1971) также показал, что
движущийся по кругу поток информации между ретикулярной формацией и
сенсорными зонами коры головного мозга необходим перед тем, как входящий
сенсорный сигнал перейдет в сознание.
Возможная роль РРТС в сознательном опыте захватывающа! Возникает ин-
туитивное чувство, что должен иметься какой-то тип передающей (распростра-
няющей) системы в головном мозге, которая позволяет всем формам сенсорных
процессов, зрению, слуху, осязанию, сочетаться с сознательным мышлением и
опытом для того, чтобы концентрироваться на некоторых входящих сигналах и
подавлять другие. Ясно, что РРТС не работает отдельно от этих видов функци-
онирования мозга. Таламус и области префронтальной коры, весьма вероятно,
тесно связаны друг с другом процессами, похожими на деятельность РРТС. Тем
не менее РРТС, кажется, играет ключевую роль в сознательном опыте человека.
4.0 .
Заключение
Попытка очень кратко описать динамическую и сложную биологическую струк-
туру, какой является головной мозг, является действительно трудной задачей.
Нашей целью в данной главе было представить вам основные структуры и обла-
сти головного мозга и их функции в человеческом познании. Необходимо запом-
нить некоторые важные положения: мозг развивался и изменялся во времени и
поэтому некоторые области мозга «старше», чем другие. Кора головного мозга,
или новая кора, представляет собой недавнее образование у человека, а лобная
и теменная доли чрезвычайно расширили свою территорию по сравнению с не-
человекообразными приматами. В то время как имеются различные области и
части головного мозга, такие как два полушария и четыре основные доли, они
тесно взаимосвязаны в мозге с помощью огромной системы нервных волокон,
которая объединяет полушария, доли и обеспечивает путь в подкорковые об-
ласти.
Некоторые важные вопросы о структуре и функциях головного мозга оста-
ются для нас загадкой. Почему мы наблюдаем так много фактов о двойствен-
ности мозга, например, имеющего два полушария, два таламуса, в то время
240
Глава 5
как у нас имеется только один разум? Какую роль играют зеркально отобра-
жающие друг друга области мозга в человеческом познании? В то время как
некоторые «более новые» области головного мозга, такие как префронтальная
кора и нижняя теменная доли, кажется, являются местоположением ассоциа-
тивной познавательной деятельности высшего порядка, также имеются некото-
рые древние области, такие как ретикулярная формация, которые могут играть
ключевую роль в сознании. Многие области головного мозга, как новые, так и
древние, объединяются и создают динамическую и интригующую биологиче-
скую структуру, которая содержит намного больше загадок для ученых, чтобы
продолжить их разгадывать.
5.0 .
Задания и упражнения к главе 5
5.1.
Контрольные задания
1. Почему иногда кору головного мозга называют «новой корой»?
2. Каковы четыре основные доли головного мозга и каковы некоторые их глав-
ные функции в когнитивной деятельности человека?
3. Где расположена медиальная височная доля? Каковы ее основные струк-
туры?
4. Кратко опишите роль таламуса в функционировании мозга.
5. Как связаны полушария головного мозга? Есть ли какие-нибудь различия в
том, как они функционируют?
6. Что такое ретикулярная формация и какую роль она может играть в созна-
тельном мышлении?
5.2 .
Графические упражнения
Покажите местоположения и названия основных опознавательных точек голов-
ного мозга, используя рис. 5 .41.
Мозг
241
Рис. 5 .41. Построение схемы головного мозга для графического упражнения
Вопрос заключается не в том, на что вы смотрите, а в том, что вы видите.
Генри Дэвид Торо
Схематический рисунок, показывающий вращающееся шахматное поле с чередующи-
мися черными и белыми клетками, которое сильно активирует первичные зрительные
поля, расположенные в затылочной доле (обозначены голубым). Вид медиальной сторо-
ны правого полушария. (Источник: Buckley, Gaffan, 2006.)
Содержание
1.0.Введение.......................................................244
1.1.Тайнаопытазрительныхвпечатлений...........................244
1.2.Цельзрения:знаниетого,чтогденаходится......................245
1.3 . Знание об объекте: восприятие характерных черт,
группиобъектов............................................245
1.4.Знаниетого,гденаходятсяобъекты.............................248
2.0.Функциональнаяорганизациязрительнойсистемы.....................248
2.1.Сетчатка...................................................248
2.2.Латеральноеколенчатоетело..................................252
2.3.Первичная,илистриарная,зрительнаякора......................254
2.4 . Экстрастриарные зрительные зоны — за пределами зоны V1 . . . . . . . 258
2.5. ПолеMT...................................................258
2.6. Вентральныйидорсальныйпути:знания«что»и«где»............ 258
2.7. Зоны,вовлеченныевраспознаваниеобъектов....................262
2.8.Латеральныйзатылочныйкомплекс.............................262
2.9.Зонараспознаваниялиц......................................264
2.10. Парагиппокампальная область восприятия пространства . . . . . . . . . . 264
3.0.Теориизрительногоосознания:гдеонопроисходит?...................264
3.1. Иерархическаяиинтерактивнаятеориизрения...................268
4.0 . Области мозга, необходимые для визуального восприятия:
исследованияповреждений........................................273
4.1 . Последствия повреждений первичных зрительных зон. . . . . . . . . . . . . 273
4.2 . Поражения экстрастриарной зоны — повреждения вне зоны V1 . . . . . 276
4.3 . Повреждение вентральных областей, воспринимающих предметы . . . 277
4.4. Повреждение дорсальных областей теменной доли. . . . . . . . . . . . . . . . 282
5.0.Связьдеятельностимозгаизрительногоопыта........................283
5.1. Восприятиесомногимиустойчивымисостояниями...............284
5.2 . Бинокулярная конкуренция: то, что вы видите, —
этото,чтостимулируетвас....................................285
5.3. Зрительноеобнаружение:виделиливыэто?.....................288
5.4 . Созидательное восприятие: видеть больше, чем доступно глазу... . . . 289
5.5. Нервныекорреляциисраспознаваниемобъектов.................291
6.0.Управлениезрительнымосознанием................................292
6.1. Транскраниальнаямагнитнаястимуляция .......................293
6.2.Бессознательноевосприятие...................................296
7.0.Заключение.....................................................299
8.0.Заданияиупражнениякглаве6....................................300
Глава 6
Зрение
244
Глава 6
1.0 . Введение
1.1 . Тайна опыта зрительных впечатлений
Вспомните, когда вы последний раз стояли наверху высокой смотровой пло-
щадки, смотря на мир внизу. Возможно, вы были на самой вершине и могли
видеть широкое пространство горизонта, прорезанного повторяющимися вер-
шинами гор, лесами, долинами и реками внизу; или, возможно, вы были на вер-
шине высокого здания, в то время как на улицах происходил цветной суетливый
водоворот движения в час пик машин, людей. Стоит остановиться на секунду,
чтобы подумать, как такие яркие и мощные впечатления могут быть простым
результатом работы группы нейронов, возбуждающихся в головном мозге. Как
может совместная электрическая активность миллиардов нейронов быть ответ-
ственна за все, что вы видите и чувствуете в каждый конкретный момент? Это
некоторые из трудных, но приводящих в восторг вопросов, которые появляют-
ся, когда мы обсуждаем отношения между деятельностью мозга и субъектив-
ным зрительным восприятием.
Большинство людей интуитивно думают, что человеческое зрение работает
очень похоже на фотоаппарат (рис. 6 .1 (а)). Если мы говорим о нашей повсед-
невной деятельности, легко поверить, что мы видим весь мир в четких цветах
с полным разрешением, в большинстве похожим на фотографию, полученную
с высоким разрешением. Однако оказывается, что это очень далеко от прав-
ды. Частично благодаря строению наших глаз (которое мы кратко повторим),
наши зрительные ощущения полноцветны и имеют высокое разрешение только
Рис. 6 .1 . Зрительный опыт. (а) Только одна из миллионов картин, которые вы можете
воспринимать с помощью зрительной системы. В отличие от фотоаппарата вы в дей-
ствительности ощущаете картинку; вы знаете, на что вы сейчас смотрите. (Фотография
получена благодаря любезности сотрудников лаборатории Тонга.) (б) Другой пример
способностей к детализации и выделению множественных признаков вашей зритель-
ной системы. Вы можете различать множество разных направлений, цветов и форм.
(Источник: Frank Tong, с разрешения.)
Зрение
245
в центре взгляда. В это трудно поверить, так как вы сейчас бросаете быстрые
взгляды по всей комнате; и иллюзия того, что вы ощущаете полную зритель-
ную картину в полном цвете и четкости изображения, кажется убедительной.
Однако если вы зафиксируете свой взгляд на определенной точке пространства,
например на уголке подоконника или этом слове на странице, и попросите кого-
нибудь показать вам несколько пальцев на краю поля зрения, вы обнаружите,
что, не двигая глазами, невероятно трудно посчитать количество пальцев, кото-
рые вам показывают. Также вы можете заметить, что цвета по вашим ощущени-
ям немного тусклее, чем в реальности.
Имеется несколько причин, почему имеет смысл ограничить зрение с высо-
ким разрешением только небольшой частью нашего зрительного поля: это толь-
ко один из многих методов, которые использует головной мозг, чтобы помочь
представить специфические особенности и объекты, которые мы видим наи-
более эффективным и действенным путем из возможных. Это доказательство
способности мозга представить внешний мир без резких переходов, так что мы
обычно не обращаем внимания на этот поразительный факт.
1.2 . Цель зрения: знание того, что где находится
Для большинства людей зрение может быть наиболее важным из пяти чувств
для ориентирования в повседневной жизни и для выполнения дел. Какова цель
зрения? Дэвид Марр, ученый, занимающийся первичным зрительным восприя-
тием, и компьютерный эксперт, сделал обманчиво простое замечание, что це-
лью зрения является «знать, где что находится». Например, если вы идете или
едете в университет, чтобы найти аудиторию в новом здании, важно знать: где
находятся другие машины и пешеходы, светофор горит красным или зеленым,
куда поедет та машина, как быстро она приближается, должен ли я повер-
нуть за этот угол улицы, это ли здание я ищу?
Принимая во внимание цель зрения, начинаем понимать, что зрительное
восприятие намного более сложное, чем простое фотографирование цифровым
фотоаппаратом (рис. 6 .1 (а)). Цифровой фотоаппарат может зафиксировать кар-
тинку, перенесенную из окружающей обстановки, и сохранить ее в виде набо-
ров пикселей. Однако в действительности фотоаппарат ничего не делает с этим
изображением и не имеет никакого знания о том, что сохранено в фотографии,
так же как какие объекты находятся в изображении или где они находятся. Са-
мый простой путь выяснить, что находится на изображении, это чтобы кто-то
взглянул на него и объяснил ее с помощью своего мозга. Зрительное воспри-
ятие — это то, что происходит после того, как картинка достигнет глаз: изо-
бражение формирует структуру распространения возбуждения совокупности
рецепторов глаза, а детализированная структура анализируется зрительными
центрами мозга, выявляя посредством этого, что где находится.
1.3 . Знание об объекте: восприятие характерных черт,
групп и объектов
Каким образом мозг устанавливает, как что-то является чем-то? Изучение
зрительного восприятия человека и неврология наводят на мысль, что имеет-
ся много уровней восприятия. Выявлено, что на наиболее фундаментальном
уровне человеческий мозг обрабатывает основные зрительные характеристи-
246
Глава 6
ки, такие как цвет, ориентация, движение, детализация поверхности (тек-
стура) и стереоскопическая глубина. Например, при взгляде на фотографию
цветка, показанного на рис. 6.1 (б), мы можем различить, что центр цветка
желтый, лист, находящийся прямо под ним, зеленый, а каждый из двух стеблей
находится под разным углом. Мы очень хорошо воспринимаем малейшую раз-
ницу в ориентации (1–2° угла наклона), неуловимые различия в цвете (напри-
мер, красный цвет у розы и красный цвет у клубники) и очень слабые следы
движения.
Как мы увидим в следующем разделе, большинство нейронов в первичных
зрительных полях мозга точно настроены на восприятие специфических осо-
бенностей: некоторые могут очень сильно реагировать на линию, нарисован-
ную под определенным углом, другие — на определенный цвет или на опре-
деленное направление движения. Эти нейроны отвечают за очень небольшие
области зрительного поля (т. е . вашего поля зрения в данный момент), которые
колеблются от такой небольшой величины, как 1°, до таких, как несколько гра-
дусов угла зрения. (Если вы держите свой большой палец на расстоянии вы-
тянутой руки, его ширина составляет, вероятно, около 2° угла зрения (O’Shea,
1991).) (Диаметр Луны, когда на нее смотрят с Земли, составляет около 0,5°.)
Если деятельность каждого из этих нейронов формирует представление
только о небольшой части зрительного поля (например, содержит ли малень-
кое пятно зрительного поля вертикальную или горизонтальную линию, крас-
ная она или синяя, находится в движении или представляет собой что-то не-
подвижное), то каким образом мозг способен объединять эту информацию,
полученную от многих нейронов? Так или иначе мозг способен объединить
эти основные элементы особенностей объектов в упорядоченные группы вос-
приятия. Гештальт-психологи предполагали, что восприятие не может быть
понято путем простого изучения основных элементов восприятия (Wertheimer,
1912; Koffka, 1935). Немецкое слово гештальт трудно перевести прямо, оно
выражает идею, что целое больше, чем сумма его частей. Эти психологи пред-
лагали гештальт-законы организации восприятия, такие как законы схоже-
сти, близости, подходящего продолжения, общей судьбы и т. д . (рис. 6.2). Эти
законы предполагают, что элементы, более схожие по цвету или форме, будут
более вероятно восприниматься как группа. Подобным образом, если элемен-
ты в группе расположены так, что между ними наименьшие промежутки на-
ходятся в рядах или столбцах, это определит, будут элементы восприниматься
как ряды или как столбцы.
Почему группирование так важно в восприятии? Оно помогает нам понять,
какие характерные особенности принадлежат возможному объекту, и помогает
нам отличить объект от фона. Представьте, например, что вы видите золотисто-
го ретривера, лежащего в густой траве. Группирование по схожести может по-
мочь нам увидеть, что собака отделена от травы, а лежащий рядом виляющийся
хвост может быть распознан как часть собаки, даже если виден только кончик
хвоста. Если собака побежит к кусту, яростно лая, то кот, который спрятался за
кустом, может быть незаметен до тех пор, пока не начнет двигаться. Но так как
движение кота приводит к группированию по общей судьбе, это может вызвать
ясное впечатление о завершенной форме животного, а не о частицах и фрагмен-
тах шкуры, разделенной ветками и листьями.
Зрение
247
В конце концов мы можем постигать форму объектов как целого и сопостав-
лять это представление о форме тем объектам, которые мы знали по предше-
ствующему опыту. Для восприятия объекта мозг должен пройти через многие
стадии обработки визуальной информации, от обработки характерных элемен-
тов объекта, сборки элементов вместе в связанные группы и наконец, вычисле-
ния, как эти элементы образуют согласованную и организованную форму. Это
восприятие формы объекта должно затем быть сопоставлено с соответствую-
щим представлением об объекте, сохраненным до этого в памяти. При условии,
что имеются тысячи объектов и то двухмерное изображение любого объекта,
которое проецируется на заднюю часть глазного яблока, может отличаться в за-
висимости от изменения точек наблюдения, освещения и условий видимости,
задача узнавания объектов становится особенно проблематичной. Каким-то об-
разом головной мозг должен обобщать устойчивые или неизменные свойства
объекта и одновременно игнорировать все кажущиеся особенности, по кото-
рым может различаться двухмерное изображение объекта.
Позже в этой главе мы обсудим, как мозг обрабатывает различные типы ха-
рактерных черт и объектов, воспринимаемых визуально.
Рис. 6 .2. Гештальт-организация.
(а) Группирование по схожести: бе-
лые точки объединяются с другими
белыми точками. Справа квадраты
объединяются с квадратами. Объ-
екты со схожими особенностями
мы стремимся сгруппировать вме-
сте. (б) Здесь, несмотря на то что
все объекты — круги, благодаря их
сгруппированному месторасполо-
жению или близости друг к другу
мы их воспринимаем как две от-
дельные группы точек. (в) Группи-
рование путем хорошего продолже-
ния. Слева мы воспринимаем один
объект. Справа нарисованы точно
такие же линии, но они разделены
так, что нет продолжения цело-
го объекта. Во всех этих случаях
совокупность объектов образует
группы или более обширные гло-
бальные объекты, которые являют-
ся большим, нежели простая сум-
ма частей. (Источник: Frank Tong,
с разрешения.)
248
Глава 6
1.4 . Знание того, где находятся объекты
Как мы понимаем, где расположены объекты во внешнем мире? Когда мы смо-
трим на мир вокруг нас, картина, которая отпечатывается на задней поверхности
наших глаз, будет по существу двухмерной, подобной изображению, которое
мы бы получили с помощью фотоаппарата. Эта двухмерная карта мира, соз-
данная в глазах, сохраняется в первичной зрительной области коры головного
мозга, которая вырабатывает схему того, где объекты расположены относитель-
но центра взгляда. Головной мозг также может разгадать потерянное третье из-
мерение и вычислить, как далеко объекты находятся в пространстве. В то время
как области первичной зрительной коры представляют положение объектов от-
носительно центра взгляда, вероятно, более высшие участки головного мозга в
теменной, височной или лобной доле представляют положение объектов более
обобщенным (менее относящимся к зрению) образом: по отношению к поло-
жению тела человека или относительно окружающего пространства в общем.
2.0 . Функциональная организация
зрительной системы
Когда свет, пришедший от объекта, достигает наших глаз, он запускает каскад
событий в нервной системе, при котором зрительный образ преобразуется в
нервные импульсы, которые перемещаются по зрительной системе от одного
участка головного мозга до другого. На протяжении серий процессов, проис-
ходящих в нейронах во многих участках головного мозга, деятельность этих
нейронов в многочисленных областях мозга каким-то образом приводит к зри-
тельным ощущениям, распознаванию объекта и многочисленных особенностей
его составных частей. Давайте проследим последовательность событий, чтобы
понять, как головной мозг обрабатывает зрительную информацию на каждом
уровне зрительного процесса. Это поможет нам понять, какой вклад различные
зрительные области мозга вносят в зрительное восприятие.
2.1. Сетчатка
Существуют два типа фоторецепторов: колбочки и палочки (рис. 6.3). Колбочки
избирательно реагируют на цвет, менее чувствительны к тусклому свету, чем
палочки, и важны для детального цветового зрения при дневном свете. Каждая
колбочка содержит один из трех видов фотопигментов, специализированных
белков, чувствительных к различным длинам волн света. Эти длины волн при-
близительно соответствуют нашей способности различать красный, зеленый и
синий. Когда свет попадает на молекулу фотопигмента, энергия света погло-
щается, и молекула затем преобразует свою форму таким образом, что в этом
фоторецепторном нейроне изменяется движение электрического тока. Колбоч-
ки плотно упакованы в желтом пятне (макуле), центральной части сетчатки,
которое мы используем при прямом взгляде на объекты, чтобы получить пред-
ставление об их мельчайших деталях. На периферии колбочки более рассре-
доточены и рассеяны, это является причиной того, что объекты по периферии
зрения кажутся более расплывчатыми, а их цвет менее ярким.
Палочки содержат другой фотопигмент, который намного более чувстви-
телен к низкому уровню освещенности. Палочки важны для ночного зрения,
Зрение
249
наша способность видеть зависит от палочек, когда наши глаза адаптировались
к темноте (темновая адаптация). Интересно, что в желтом пятне нет палочек,
а только колбочки, доля палочек увеличивается по направлению к периферии.
Вот почему вы, возможно, замечали, глядя на ночное небо, что очень тусклую
звезду легче увидеть, если смотреть немного вбок.
Сигналы от фоторецепторов обрабатываются совокупностью промежуточ-
ных нейронов: биполярными клетками, горизонтальными клетками и амакри-
новыми клетками, затем они достигают ганглиозных клеток, конечного уровня
обработки в сетчатке перед тем, как сигнал покинет глаз. Фактически клеточ-
ные тела ганглиозных клеток находятся в сетчатке, но эти клетки имеют длин-
ные аксоны, которые покидают сетчатку в слепом пятне, образуя зрительный
тракт. Каждая ганглиозная клетка получает возбуждающие сигналы от груп-
пы палочек и колбочек: эта квинтэссенция информации образует рецептивное
пол е, понятие, к которому мы вскоре вернемся. Ганглиозные клетки в желтом
пятне получают информацию только от небольшого числа колбочек, в то время
как ганглиозные клетки на периферии получают входящие сигналы от большо-
го количества палочек (иногда тысяч). Такое количество палочек формирует
Рис. 6 .3 . Глаз. (а) Рисунок, показывающий, как объекты окружающего мира физиче-
ски отображаются на задней стенке глаза — сетчатке. (б) Глаз и поперечное сечение
сетчатки. Показаны как колбочки, так и палочки. Они реагируют на разные виды света.
Нервные импульсы попадают через биполярные клетки на ганглиозные клетки. Аксоны
ганглиозных клеток выводят информацию от нейронов по направлению от глаза назад,
к коре головного мозга. (Источник: Squire et al., 2003.)
250
Глава 6
суммарный входящий сигнал на единственную ганглиозную клетку, если хотя
бы одна из таких палочек была активирована фотонами света, эта активация
может возбудить ганглиозную клетку, что повышает вероятность возможности
заметить тусклый рассеянный свет. Однако это увеличение в чувствительности
к тусклому свету достигается за счет более слабого разрешения: палочки обе-
спечивают большую чувствительность, но также более расплывчатое изобра-
жение, чем резкая картина при дневном свете, которую дает зрение с помощью
колбочек.
Ганглиозные клетки сетчатки получают как возбуждающие, так и тормозные
входящие сигналы от биполярных нейронов, и пространственное расположе-
ние этих входящих сигналов определяет рецептивное поле клетки (рис. 6 .4 (а)).
Рецептивное поле нейрона относится к части зрительного поля, которая мо-
жет активировать или сильно тормозить ответ этой клетки. Ганглиозные клетки
сетчатки имеют рецептивные поля, организованные по принципу «центр–пери-
ферия». Например, рецептивное поле клетки с on-центром (зоной включения)
и off-периферией (зоной выключения) будет сильно реагировать, если пятно
света будет находиться в центре рецептивного поля. Если пятно света увели-
чивается, ответ будет усиливаться, пока свет не начнет выходить за пределы
области on-центра. После этого ответ ганглиозной клетки начинает ослабевать
по мере того, как пятно света становится шире и стимулирует все больше и
больше окружающей центр off-зоны. (Рецептивное поле клетки с off-центром и
Рис. 6 .4 . Рецептивные поля, организованные по принципу «центр–периферия». (а)
Схематический пример ответа клеток, организованного по принципу «центр–перифе-
рия», на пятна света, различные по размерам. Заметьте, что наибольший пиковый от-
вет (показанный линиями справа) имеется для центрального светового пятна среднего
размера. Пятно света должно быть точно соответствующего размера, чтобы получить
максимальный исходящий сигнал данного конкретного нейрона. (б) Модель того, как
может быть организовано рецептивное поле по принципу «центр–периферия» путем
совместной работы и конкуренции между различными соединенными друг с другом
нейронами в сетчатке. (Источник: Frank Tong, с разрешения.)
Зрение
251
on-периферией будет больше всего реагировать на темное пятно, находящееся
в центре рецептивного поля.)
Как можно понять поведение ганглиозных клеток сетчатки? Ведущей иде-
ей является принцип латерального торможения (Kuffler, 1953). Латеральное
торможение означает, что активность нейрона может быть заторможена вхо-
дящими сигналами, приходящими от нейронов, которые отвечают за соседние
области зрительного поля. Например, ганглиозные клетки на рис. 6 .4 (б) полу-
чают возбуждающие сигналы от клеток, соответствующих области on-центра,
а тормозные сигналы из области off-зоны. Силы этих возбуждающих и тормоз-
off-зоны. Силы этих возбуждающих и тормоз-
- зоны. Силы этих возбуждающих и тормоз-
ных сигналов обычно уравновешены таким образом, что, если свет как на on-,
так и на off-зонах одинаков, нейрон не будет реагировать на это равномерное
освещение.
Почему рецептивные поля, организованные по принципу «центр–перифе-
рия», и латеральное торможение так важны? Латеральное торможение важно
для усиления представительства нейронов краев (периферии), области изобра-
жения, где интенсивность света резко изменяется. Эти внезапные изменения
служат признаком наличия возможных контуров, структур, теней или объектов
в любой видимой обстановке, в то время как однородные части картины не
особенно информативны или интересны. На рис. 6 .5 изображена лиса в исход-
ном виде и после того, как компьютер отфильтровал только границы (правая
картинка) так, что области, выделенные черным, показывают, в каком месте
ганглиозные клетки наиболее сильно реагировали бы на изображение. Лате-
ральное торможение также приводит к более эффективному представитель-
ству границ на уровне нейронов, потому что будут сильно возбуждены только
нейроны, соответствующие именно границам стимулов; другие нейроны, чьи
рецептивные поля располагаются в неизменных условиях, не будут активиро-
ваться. Так как при возбуждении нейроны расходуют большое количество ме-
таболической энергии, это намного эффективнее. Таков пример эффективного
нейронного кодирования: быть активными в любой момент времени требуется
Рис. 6 .5 . Границы содержат больше всего информации. Пример того, как наибольшая
часть информации на фотографии содержится в границах объектов. Фотография сле-
ва — исходная, рисунок справа содержит только информацию о границах, он получен
с помощью специального компьютерного алгоритма. (Источник: Frank Tong, с разре-
шения.)
252
Глава 6
только небольшому количеству нейронов, чтобы представить детальный зри-
тельный стимул.
Латеральное торможение также помогает обеспечивать похожие ответы го-
ловного мозга на объект или видимую обстановку как в тусклый серый, так и в
яркий день. Изменения в абсолютном уровне яркости совсем немного повлия-
ют на структуру активности на сетчатке: наибольшее значение имеет только от-
носительная яркость. И, наконец, латеральное торможение на многочисленных
уровнях обработки зрительной информации, включающих сетчатку, латераль-
ные коленчатые тела (ЛКТ) и зрительную кору, может привести к интересным
зрительным иллюзиям, таким как иллюзия «решетки Германна» (рис. 6 .6).
2.2 . Латеральное коленчатое тело
От глаза ганглиозные клетки сетчатки посылают свои аксоны к структуре в та-
ламусе, которая называется латеральное коленчатое тело (ЛКТ). В частности,
левая половина каждой сетчатки проецируется на левое ЛКТ, правая половина
сетчатки — на правое ЛКТ. Чтобы это произошло, входящие сигналы от но-
совой порции каждой сетчатки должны перекрещиваться в месте зрительно-
Рис. 6 .6 . Иллюзия «решетки Германна». Взгляните внимательно на совокупность чер-
ных квадратов на рисунке. Замечаете ли вы что-то необычное? Создалось ли у вас впе-
чатление, что вы видите маленькие темные круги между черными квадратами на их
периферии? Не беспокойтесь, это вполне обычно. Это прекрасный пример работы ре-
цептивных полей и латерального торможения (Hermann, 1870). Возможные рецептив-
ные поля показаны в правой части таблицы из квадратов. Рецептивное поле, которое на-
ходится между углами четырех черных квадратов, будет получать больше тормозящих
сигналов из своей периферии, которая стимулируется белыми частями решетки, чем
рецептивное поле, которое находится только между двумя темными квадратами. В ре-
зультате нейроны с рецептивными полями, расположенными между четырьмя темны-
ми квадратами, будут более слабо возбуждены, что приводит к ощущению небольших
темных пятен в этих местах пересечения. В желтом пятне рецептивные поля настолько
более малы, что эта иллюзия будет ощущаться только на периферии зрения. (Источник:
Frank Tong, с разрешения.)
Зрение
253
го перекреста и идти к противоположному ЛКТ (рис. 6 .7). В результате левое
ЛКТ получает входящие сигналы от правого зрительного поля, а правое ЛКТ
получает входящие сигналы от левого зрительного поля, так что каждое ЛКТ
служит представителем контралатерального (противоположного) зрительно-
го поля. Заметьте, что входящие сигналы от каждого глаза идут к отдельным
слоям ЛКТ, которые служат для представительства только одного глаза (моно-
кулярны), поэтому сигналы от двух глаз идут отдельно до тех пор, пока не до-
стигнут первичной зрительной коры, в которой эти сигналы объединяются.
Рис. 6 .7 . Зрительный путь от сетчатки к коре. (а) Пример среза мозга, полученного
методом функциональной магнитно-резонансной томографии (фМРТ), показывающий
латеральное коленчатое тело (ЛКТ) и первичную зрительную область на задней сторо-
не мозга (затылочную кору). Два различных цвета отмечают два больших полушария
головного мозга. (б) Схематический рисунок, показывающий зрительные пути от сет-
чатки глаза до первичной зрительной коры на задней стороне мозга. Здесь вы можете
увидеть, что информация от нейронов с носовой или с внутренней сторон глаз пере-
секается в месте зрительного перекреста и потом обрабатывается на противоположной
(контралатеральной) стороне мозга. Левое зрительное поле, окрашенное голубым, об-
рабатывается правой зрительной корой (также окрашена голубым). ЛКТ, выделенное
зеленым, передает зрительную информацию к первичным зрительным областям коры
головного мозга. (Источник: Squire et al., 2003.)
254
Глава 6
На что похожи рецептивные поля ЛКТ? Они имеют ту же форму и основные
свойства ганглиозных клеток сетчатки с рецептивными полями, организован-
ными по принципу «центр–периферия». Таламус часто считается промежуточ-
ной остановкой для сигналов, которые в конце концов должны достичь коры
головного мозга, в которой нейроны начинают реагировать уже очень различ-
ным образом.
Итак, как же будут отвечать нейроны ЛКТ или ганглиозных клеток сетчатки
на обычный контур дома, показанный на рис. 6 .10? Клетки с рецептивным по-
лем с on-центром, располагающиеся на любом из контуров дома, будут сильно
возбуждены. Заметьте, что эти клетки возбудятся одинаково сильно как для вер-
тикальных, так и для горизонтальных и наклонных линий. Несмотря на то что
небольшая часть периферической зоны с тормозящими сигналами также будет
стимулироваться этими линиями, в целом будут возбуждаться все области on-
центров, что обусловит более сильную, чем торможение, активацию и в резуль-
тате приведет к постоянному уровню возбуждения.
2.3 . Первичная, или стриарная, зрительная кора
От ЛКТ нейроны посылают свои импульсы в первичную зрительную кору, ино-
гда называемую V1, потому что эта зона является первой зрительной областью
коры. V1 часто называют «стриарной» корой, потому что на срезах она имеет
явно полосатый (от лат. striatus — полосатый) вид. Около 90% выходящих из
сетчатки сигналов достигают сначала ЛКТ, а затем идут далее до V1. Левое
ЛКТ посылает сигналы к V1 в левом полушарии, правое ЛКТ — к правой V1
(рис. 6 .7). В V1 все еще сохраняется пространственное расположение входя-
V1 все еще сохраняется пространственное расположение входя-
1 все еще сохраняется пространственное расположение входя-
щих сигналов от сетчатки. Левая V1 получает через таламус упорядоченный
ряд сигналов от левых половин обеих сетчаток. Часть зрительного поля от цен-
тральной ямки представлена в задней части затылочной доли около затылочно-
го полюса, а более близкие к периферии части зрительного поля представлены
в более передней части. Левая V1, таким образом, содержит ретинотопиче-
скую карту правого зрительного поля в целом, в то время как правая V1 содер-
V1 содер-
1 содер-
жит карту целого левого зрительного поля. Эта ретинотопическая организация
преобладает в первичных зрительных зонах (от V1 до V4), где нейроны имеют
небольшие рецептивные поля, но становится более слабо выраженной и менее
упорядоченной в высших зрительных зонах вне затылочной доли.
Нейроны в V1 чувствительны к целой совокупности зрительных особенно-
V1 чувствительны к целой совокупности зрительных особенно-
1 чувствительны к целой совокупности зрительных особенно-
стей, не обнаруживаемых в ЛКТ. Одной из наиболее важных зрительных харак-
теристик является ориентация (Hubel, Wiesel, 1962, 1968). Некоторые нейроны
V1 сильнее реагируют на вертикальные линии, другие на линии под углом в
20°, другие на горизонтальные линии и так далее. Каким образом эти нейроны
приобретают новые свойства или рецептивные поля? На рис. 6 .8 представлена
модель избирательности нейронов V1 к ориентации. Если нейрон V1 получа-
V1 к ориентации. Если нейрон V1 получа-
1 к ориентации. Если нейрон V1 получа-
V1 получа-
1 получа-
ет возбуждающий входящий сигнал от трех нейронов ЛКТ с выстроенными в
линию рецептивными полями, организованными по принципу «центр–перифе-
рия», то в этом случае нейрон V1 будет наиболее сильно отвечать на соответ-
V1 будет наиболее сильно отвечать на соответ-
1 будет наиболее сильно отвечать на соответ-
ствующе расположенную линию. Например, если представлена вертикальная
полоска, нейрон, показанный на рисунке, будет наиболее сильно реагировать,
потому что будет стимулироваться вся возбуждающая зона, в то время как тор-
мозящая зона периферии не будет возбуждаться. Если полоска как-то отклоня-
Зрение
255
ется от горизонтали, нейрон будет отвечать слабее, потому что теперь будет воз-
буждаться часть периферического окружения, а часть возбуждающего центра
не будет. В конце концов, при горизонтальной полосе нейрон не будет вообще
возбуждаться, так как равные пропорции зон, которые получают стимуляцию,
в центре и периферии приводят к равновесию в силе входящих возбуждающих
и тормозящих входящих сигналов. Такая форма рецептивных полей, организо-
ванных по принципу «центр–периферия», может объяснить избирательность к
ориентации у нейронов V1.
Нейроны V1 кроме ориентации также чувствительны ко многим другим
зрительным особенностям (Hubel, Wiesel, 1998). Некоторые нейроны сильнее
отвечают на определенное направление движения, такое как движение вперед,
влево или вниз. Другие нейроны более чувствительны к определенным цветам
или различиям в цвете (например, красный в отличие зеленого, желтый — от
синего), хотя некоторые основные типы чувствительных к цвету нейронов мож-
но также найти в сетчатке и ЛКТ. В заключение скажем, что некоторые ней-
роны наиболее сильно отвечают на определенные бинокулярные расхождения
(бинокулярную диспарантность) (Barleo et al., 1967; Cumming, 2002), которые
относятся к степени совмещения между изображениями в двух глазах. Именно
Рис. 6 .8 . Избирательность V1 к ориентации объекта. Пример того, как совокупность
рецептивных полей, организованных по принципу «центр–периферия», может привести
к избирательности в работе нейронов V1. Перекрывающиеся круги слева показывают ре-
цептивные поля, работающие по принципу «центр–периферия. Если полоска света рас-
положена вертикально, она активирует все on-центры (+ ) каждого рецептивного поля.
В то время как если ее расположение изменяется (полоска становится наклонной), воз-
буждается меньшее количество центров и большее число периферийных зон, в резуль-
тате получается более слабый нейронный ответ (меньшее количество пиков на графике).
Таким образом, вертикальная полоса вызывает больший ответ нейронов, а когда стимул
повернут, величина ответа V1 уменьшается. Это составляет избирательность по ориен-
тации, которая наблюдается только в коре. (Источник: Frank Tong, с разрешения.)
256
Глава 6
Рис. 6.9 . Пути зрительной системы от глаза к V1. Схематический рисунок путей би-
нокулярного зрения, показывающий зрительные входящие сигналы от левого и право-
го зрительных полей (вверху), проходящие через зрительный нерв и зрительный тракт
(в ц ентре), далее через ЛКТ и вход в кору (внизу). (Источник: Standring, 2005.)
Зрение
257
небольшие сдвиги между изображениями в обоих глазах позволяют нам полу-
чить стереоскопическую глубину, когда мы смотрим на объекты обоими откры-
тыми глазами. (Попробуйте закрыть один глаз, вытянув руки на полную длину,
и попытайтесь быстро переместить ваши два указательных пальца так, чтобы
они встретили друг друга. А теперь опять попытайтесь это сделать, открыв оба
глаза. Если вы имеете нормальное бинокулярное зрение, сделать это должно
быть намного легче, когда оба глаза открыты, потому что вы можете лучше оце-
нивать расстояние до ваших двух пальцев.) Смотрите на рис. 6 .9 схематическое
отображение путей в бинокулярной зрительной системе.
Каким же образом нейроны V1 отвечают на контур домика? Нейрон V1, со-
V1 отвечают на контур домика? Нейрон V1, со-
1 отвечают на контур домика? Нейрон V1, со-
V1, со-
1, со-
ответствующий линиям с наклоном 45° и имеющий расположение рецептивно-
го поля вдоль крыши, может сильно возбуждаться на наклонную крышу. Ней-
рон V1, который сильнее возбуждается на вертикали, поможет подать сигнал
о наличии вертикальной стены, а горизонтальный нейрон будет отвечать на
потолок или пол дома. В этом смысле можно легко видеть, что нейроны V1
делают намного больше, чем отвечают на простые точки света, как делает ЛКТ.
Нейроны V1 обеспечивают нейронное отображение ориентации зрительных
признаков, которое составляет контуры и формы объектов. На рис. 6 .10 дано
Рис. 6 .10. Иерархия обработки зрительных сигналов. Демонстрация иерархических
свойств ответа зрительной системы на простые и сложные стимулы. В левой колонке
показаны стимулы от дома, а что собой представляют рецептивные поля каждой зритель-
ной области, мы увидим в облачках. В каждой визуальной зоне увеличиваются не только
размеры рецептивного поля, но также сложность форм, на которые они отвечают. Правая
колонка показывает оценку того, где находится в мозге каждая из зон. Вы можете видеть,
что первичные зрительные зоны отвечают за простые особенности, в то время как при
движении по направлению обработки информации, зоны отвечают за все более сложные
формы и объекты. Это твердо установленное основное направление работы зрительной
системы. (Источник: Frank Tong, с разрешения.)
258
Глава 6
заключение по иерархии обработки зрительной информации. От ЛКТ, V1, V4
до вентральной височной коры вы можете видеть, что нейроны от одной обла-
сти к другой постепенно отвечают на все более сложные стимулы.
Итак, подведем итог: V1 важна при анализе зрительных особенностей на
тонком уровне детализации. Эти нейроны имеют маленькие рецептивные поля,
которые чувствительны к ориентации, цвету, движению или бинокулярной дис-
парантности. После того как зрительные сигналы проанализированы в V1, они
посылаются в высшие зрительные зоны для дальнейшей обработки.
2.4 . Экстрастриарные зрительные зоны — за пределами зоны V1
V1 посылает прямые положительные сигналы ко многим высшим зрительным
зонам, если назвать несколько, то это такие поля, как V2, V3, V4 и чувстви-
тельное к движению поле МТ (рис. 6 .11) (Felleman, Va n Essen, 1991). Поле V4
известно тем, что особенно важно для восприятия цвета (�eki, 1977), а некото-
�eki, 1977), а некото-
, 1977), а некото-
рые нейроны этого поля хорошо отвечают на более сложные признаки или ком-
бинации признаков (Pasupathy, Connor, 2002). Например, некоторые нейроны
V4 чувствительны к изгибам или к двум линиям, встречающимся под опреде-
4 чувствительны к изгибам или к двум линиям, встречающимся под опреде-
ленным углом. Эти нейроны могут сигнализировать о присутствии или кривого
контура, или угла. В нашем примере о доме — нейрон V4 мог наиболее сильно
возбуждаться при пересечении двух линий, которые образуют вершину крыши
или другой угол дома.
Каким образом в дальнейшем эти разные кусочки и части дома, которые
представлены простым расположением линий и углов, в конце концов пред-
стают перед нами как целый объект? Поле V4 посылает много исходящих сиг-
V4 посылает много исходящих сиг-
4 посылает много исходящих сиг-
налов в высшие зрительные зоны по вентральному зрительному пути, который
важен для узнавания объектов (�ngerleider, Mishkin, 1982). Передняя часть вен-
�ngerleider, Mishkin, 1982). Передняя часть вен-
, Mishkin, 1982). Передняя часть вен-
Mishkin, 1982). Передняя часть вен-
, 1982). Передняя часть вен-
трального зрительного пути состоит из вентральной височной коры, которая
особенно важна для узнавания объектов.
2.5 . Поле MT
Средняя височная область — ее обычно называют поле MT (middle-temporal)
(см. рис. 6 .11) — важна для восприятия движения. Почти все нейроны поля
MT селективны по направлению: они выборочно отвечают на определенный
ряд направлений движения и не отвечают на направления за пределами это-
го ряда (�eki, 1974; Albright, 1984). Более того, некоторые из этих нейронов
хорошо возбуждаются на структуру движения (Albright, 1992), что означает,
что эти нейроны могут объединять много различных направлений движения
и вычислять, каким может быть общее направление объекта. Как мы увидим,
активность этой области, вероятно, тесно связана с восприятием движений, и
при нарушении активности этой зоны восприятие движений может быть серьез-
но нарушено.
2.6 . Вентральный и дорсальный пути: знания «что» и «где»
Проекции от V1 к высшим зонам коры головного мозга можно приблизительно
разделить по двум основным параллельным путям: вентральный путь, веду-
щий от V1 к височной доле, который важен для представления того, «чем» яв-
ляются объекты, и дорсальный путь, идущий от V1 к теменной доле, который
важен для представления, «где» расположены объекты (рис. 6 .12).
Зрение
259
Рис. 6 .11 . Зрительные зоны мозга. Функционально различающиеся зрительные об-
ласти человеческого мозга, показанные на реконструкциях со сканированных изобра-
жений анатомических структур головного мозга. (а) Уплощенное изображение коры
зрительной системы человека. Уплощенная карта была создана путем развертывания
извилистой поверхности коры головного мозга, включая разрез через шпорную борозду,
которая разделяет поле V1 на дорсальную и вентральную половины, а также уплоще-
ния поверхности коры, используя компьютерные методы. Это позволяет показать все
зрительные поля одного полушария, включая части, погруженные в борозды, на одном
изображении. Верхняя половина показывает дорсальную часть зрительных полей V1,
V2, V3, а также V3a. Нижняя половина показывает вентральные части зрительных по-
лей V1, V2, V3 и V4, а также парагиппокампальную область восприятия пространства
(ПОП) и зону распознавания лиц (ЗРЛ). Также показаны области латерального заты-
лочного комплекса (ЛЗК) и MT. (б) Вид с вентральной стороны одного полушария, по-
казывающий местоположение этих областей в коре. (в) Вид с латеральной стороны того
же полушария, показывающий позиции других зрительных полей. (г) Вид с медиаль-
ной стороны этого полушария, в основном показывающий V1. (Источник: Squire et al.,
2003.)
260
Глава 6
Эти различия между вентральным и дорсальным путями, иногда упомина-
емыми как пути «что» и «где» соответственно, являются важным организа-
ционным принципом зрительной системы, предложенным Ангерлейдером и
Мишкиным (�ngerleider, Mishkin, 1982) (рис. 6 .13).
Доказательствами этого разграничения являются открытия об анатомиче-
ских связях между зрительными зонами в этих двух путях, особенности от-
ветов зрительных зон каждого из путей и виды нарушения зрения в результате
поражения в теменной или височной долях.
В дорсальном пути сигналы от V1 идут в дорсальные зоны экстрастриар-
V1 идут в дорсальные зоны экстрастриар-
1 идут в дорсальные зоны экстрастриар-
ной коры, такие как MT или V3A, которые затем посылают основные отростки
во многие области теменной коры. Дорсальный путь важен для представления
о месторасположении объектов таким образом, чтобы зрительная система мог-
ла бы управлять действиями по отношению к этим объектам (�oodale, Hum-
�oodale, Hum-
, Hum-
Hum-
prey, 1998). Рассмотрите, что включает достижение любого объекта, такого как
кружка кофе, находящаяся на столе: этот тип зрения требует детальной инфор-
мации о точном положении, размере и ориентации предмета. Без такой деталь-
ной информации вы можете не достигнуть нужного места, ваша ладонь может
не подойти по размеру к ручке кружки, или ваша рука может быть направлена
не под тем углом для захвата ручки. Зоны MT и V3A важны для обработки зри-
MT и V3A важны для обработки зри-
и V3A важны для обработки зри-
V3A важны для обработки зри-
3A важны для обработки зри-
A важны для обработки зри-
важны для обработки зри-
тельной информации по движению и стереоглубине, в то время как определен-
ные области в теменной доле специализируются на управлении движениями
глаз или движений руки по направлению к определенным месторасположениям
в зрительном пространстве.
По вентральному пути многие сигналы от V1 идут к вентральным экс-
V1 идут к вентральным экс-
1 идут к вентральным экс-
трастриарным полям V2, V3 и V4 и далее ко многим полям височной доли.
Вентральный путь, или путь «что», важен для обработки информации о цве-
те, форме и опознании зрительных объектов, обработка, которая подчеркивает
устойчивые, неизменные черты объектов. Например, вентральный путь имеет
меньшее отношение к точным размерам, ориентации и расположении круж-
ки кофе; вместо этого его целью является возможность идентификации такого
объекта где бы то ни было в зрительном поле, а также возможность отличить ее
от других похожих предметов (например, чашек, мисок, чайников). Мы пого-
ворим о свойствах вентрального пути гораздо более детально, особенно о раз-
личных полях височной доли.
Рис. 6 .12 . Пути «что» и «где». Путь
«где» обычно называется дорсальным
путем, потому что он включает дорсаль-
ные зоны, такие как MT, и теменную
кору, которая расположена на верхней
поверхности мозга. Путь «что» вклю-
чает вентральные зоны, такие как V4,
ЛЗК и НВК (нижнюю височную кору,
IT), поэтому он известен как вентраль-
ный путь обработки информации. Эти
два пути — прекрасный способ пред-
ставить наглядно поток различной кор-
ковой информации. (Источник: Spuire
et al., 2003.)
Зрение
261
Рис. 6 .13 . Дорсальный и вентральный зрительные пути. Из новаторской работы Ан-
герлейдера и Мишкина (�ngerleider, Mishkin, 1982), схематическое изображение дор-
сального и вентрального путей, основанное на исследовании обезьян. (а) Вид мозга
обезьяны сбоку, показывающий расположение первичной зрительной коры (V1), экс-
трастриарной коры (V2–V4) и других ключевых зрительных зон, таких как MT. (б) Схе-
ма, показывающая зрительные поля, вовлеченные в дорсальный и вентральный пути
обработки зрительной информации. (Источник: Squire et al., 2003.)
262
Глава 6
Хотя это разграничение на дорсальный и вентральный пути полезно для
объединения зон головного мозга, а также для понимания того, сколько инфор-
мации передается туда и обратно между зрительными зонами, его нельзя при-
нимать за полное разделение. Имеется множество взаимных влияний между
двумя путями. Кроме того, теменная и височные доли посылают отростки в
некоторые общие поля в префронтальной коре, где информация каждого пути
также может воссоединиться.
2.7. Зоны, вовлеченные в распознавание объектов
Записи с одного нейрона у обезьян показали, что многие нейроны вентральной
височной коры лучше всего возбуждаются на контуры, простые фигуры или
сложные объекты. Например, некоторые нейроны этой области могут лучше
всего отвечать на квадрат или круг, другие на треугольники и т. д . до еще бо-
лее сложных фигур, таких как контур дома (см. рис. 6.10), который выглядит
как треугольник, лежащий на вершине квадрата. Некоторые нейроны этой зоны
высокоизбирательны и отвечают только на определенный тип объектов, таких
как рука, лицо, показанное с определенного ракурса, определенное животное,
легко узнаваемая игрушка, или предмет, который обезьяна научилась узнавать
и т. д. (Desimone et al., 1984; �ross, 1992; Logothetis et al., 1995; Tanaka, 1996).
Изучение человека путем сканирования мозга выявило много областей го-
ловного мозга, включенных в обработку объектов. Эти чувствительные к объ-
ектам поля, которые лежат как раз впереди от первичных зрительных полей
V1–V4, отвечают наиболее сильно на логически связанные формы и объекты
по сравнению с запутанными, бессмысленными стимулами. В этой главе мы
сконцентрируемся на трех таких областях головного мозга (см. рис. 6 .10). Ла-
теральный затылочный комплекс (ЛЗК) находится на боковой поверхности
затылочной доли, непосредственно позади зоны MT. Так как эта зона сильно
вовлечена в узнавание объектов, мы будем считать ее частью вентрального
пути, несмотря на то что она располагается выше (дорсальнее) по сравнению
с другими полями, отвечающими за объекты. Зона распознавания лиц лежит
на латеральной затылочно-височной извилине (веретенообразной извилине)
на вентральной поверхности задней височной доли. Парагиппокампальная об-
ласть узнавания пространства находится на парагиппокампальной извилине,
которая лежит как раз медиальнее от веретенообразной извилины на вентраль-
ной поверхности височной доли.
2.8 . Латеральный затылочный комплекс
Латеральный затылочный комплекс, вероятно, играет главную роль в распозна-
вании объектов и сильно возбуждается на множество форм и объектов (Malack
et al., 1995). На рис. 6 .14 показан пример нейронной активности в ЛЗК по срав-
нению с V1. Картинка становится все более и более перемешанной, V1 продол-
V1. Картинка становится все более и более перемешанной, V1 продол-
1. Картинка становится все более и более перемешанной, V1 продол-
V1 продол-
1 продол-
жает возбуждаться и даже дает более сильный ответ, а активность ЛЗК умень-
шается. Это показывает, что ЛЗК предпочитает целостные формы и предметы
перепутанным зрительным признакам.
Эта область, вероятно, отображает конкретные формы объектов (блок 6.1).
Предположительно, различные нейроны этой зоны отвечают лучше всего на
различные типы объектов. Так как сканирование мозга человека не имеет до-
статочного разрешения, чтобы измерить предпочтения в объектах отдельных
Зрение
263
нейронов, имеется другой метод тестирования избирательности к объектам —
измерить нейронную адаптацию к определенной форме. Этот метод включает в
себя представление двух объектов друг за другом. Если в этой группе показыва-
ют одну и ту же форму объекта дважды, то область с избирательностью к объ-
ектам должна адаптироваться к повторяющейся форме и менее возбуждаться во
второй раз. Именно это и происходит в ЛЗК, даже если повторяющийся объект
показывают расположенным по-новому или имеющим другой вид, и поэтому
адаптация зрительных полей с ретинотопической организацией не происходит
(�rill-Spector et al., 1999; Kourtzi, Kanwisher, 2001).
Рис. 6 .14. Ответ нейронов от низших по уровню до высших зон. Ответ первичной зри-
тельной коры (V1) и латеральной затылочной коры (ЛЗК) на фотографию котенка при
различном уровне связанности. При приведенной в беспорядок фотографии V1 про-
должает отвечать, она, как ни удивительно, практически увеличивает ответ, пока фото-
графия беспорядочна. Это показывает, что в V1 ответ зависит не от фотографии котенка,
а от ограниченных пятен света и их расположения. И наоборот, активность латеральной
затылочной коры показывает большее возбуждение, вызванное котенком, и наоборот,
в то время как картинка перемешана, активность ЛЗК резко падает. Это показывает, что
в отличие от V1 активность ЛЗК является ответом на предъявление фотографии котен-
ка. (Источник: Frank Tong, с разрешения.)
264
Глава 6
2.9 . Зона распознавания лиц
Нейрофизиологические исследования показали, что нейроны, избирательные
к лицам, можно найти во многих частях височной доли обезьян. Некоторые из
этих клеток, реагирующих на лица, демонстрируют замечательную точность
ответа на стимул и могут лучше всего отвечать на предъявление лица с опре-
деленными особенностями, выражения лица или на лица с определенной точки
рассматривания (например, правый профиль). Обычно эти нейроны лежат меж-
ду нейронов, отвечающих на различные типы объектов, что привело ученых
к предположению: отдельной зоны головного мозга, специализирующейся на
обработке лиц, не существует.
Однако исследования по сканированию нейронов головного мозга челове-
ка показали, что имеется зона в веретенообразной извилине, названная зоной
распознавания лиц (ЗРЛ), которая наиболее сильно отвечает на лица, чем на
любую другую категорию объектов (Kanwisher et al., 1997). Эта область отвеча-
ет больше на лица человека, животных или на рисунки лиц, чем на множество
нелицеподобных стимулов, включающих руки, туловища, глаза, показанные
отдельно, головы, изображенные сзади, цветы, здания и неодушевленные объ-
екты (Kanwisher et al., 1997; McCarthy et al., 1997; Tong et al., 2000; Schwarzlose
et al., 2005). В недавней работе исследователи пытались сканировать мозг обе-
зьян, чтобы выяснить, не может ли он также иметь зону в вентральной височ-
ной коре, избирательную к лицам, и оказалось, что он тоже ее имеет (Tsao et
al., 2006). Затем исследователи также смогли записать активность отдельных
нейронов в этой зоне распознавания лиц и обнаружили, что 97% нейронов в
этой зоне больше отвечают на лица, чем на другие типы объектов. Более того,
эти нейроны очень хорошо распознавали разные характерные особенности объ-
ектов, что предполагает, что они играют важную роль в узнавании и различении
разных лиц. Так как эта зона очень мала, такую сильно избирательную к лицам
область, которая почти целиком состоит из клеток, реагирующих на лица, до
этого никогда не открывали. Как мы увидим, эта область, вероятно, важна для
осознанного восприятия лиц.
2.10. Парагиппокампальная область
восприятия пространства
Парагиппокампальная область восприятия пространства (ПОП) является дру-
гой высокоизбирательной областью, которая лучше всего распознает дома, ори-
ентиры местности, места действия внутри помещений и снаружи (Epstein, Kan-
Epstein, Kan-
, Kan-
Kan-
wisher, 1998). По сравнению с другими областями, эта зона мозга более слабо
отвечает на другие типы стимулов, такие как лица, тела или неживые объекты.
Так как эта зона отвечает на очень разные стимулы, в отличие от зоны распозна-
вания лиц, многие исследования воспользовались различными способностями
к возбуждению ЗРЛ и ПОП для исследования нервных связей при получении
зрительной информации, как мы увидим позже.
3.0 . Теории зрительного осознания: где оно происходит?
Итак, возможно ли сказать, в каком месте этого потока нервной активности дей-
ствительно происходит осознание? Возможно ли указать на определенную груп-
пу нейронов или определенную область мозга и сказать: «это... то место в моем
головном мозге, где я ощущаю зримый мир в моем разуме».
Зрение
265
Блок 6.1 . Люди воспринимают людей: нахождение связей нейронами
в случае движения биологических объектов
Узнавали ли вы когда-либо друга в толпе просто потому, что вы что-то узнали в его
или ее манере двигаться, идти или жестикулировать? Имеется ли что-то особенное в
том, как человек воспринимает движение, специфичное для нашего вида, в отличие
от других видов движений, таких как волна, разбивающаяся о линию берега, флаг,
развевающийся на флагштоке, или самолет, движущийся в небе?
Исследователи зрения долгое время задавали эти и близкие к ним вопросы о зри-
тельном восприятии человека. Первой задачей, на которую нужно было обратить
внимание, было исследование зрительного восприятия, которое характерно для дви-
жения человека как биологического объекта и не относится к любой другой стороне
зрительного восприятия. Шведский ученый разработал новый путь, как выяснить
именно это. В 1973 г. Гуннар Йоханссон (�unnar Johansson)* опубликовал труд, оза-
главленный «Зрительное восприятие движения биологических объектов и модель его
анализа», который вызвал десятилетия ис-
следований по теме восприятия движения
биологических объектов. Д -р Йоханссон
разработал метод прикрепления небольших
точек света на части тела его научных ас-
систентов, одетых в темное: плечи, локти,
кисти рук, колени и ступни. Он снимал сво-
их помощников на камеру в то время, как
те двигались по тускло освещенной лабора-
тории, тем самым создавая набор стимулов,
которые составляли только двигающиеся
точки света (рис. 6.15). Результаты этих ран-
них и проведенных с тех пор многих других
исследований движений точек света показа-
ли, что, да, мы, люди, довольно искусны в
восприятии движений друг друга. Интригу-
ющей стороной этих исследований являет-
ся то, что ни одно отдельное пятно света не
может дать нам информацию, как двигается
человек: она требует наличия всех точек
света (или, по крайней мере, большую их
часть). Последующие работы предоставили
данные о том, что при использовании набо-
ра стимулов из точек света воспринимается
не только движение биологических объек-
тов: наблюдатели могли описать пол (Ko-
Ko-
zlowski, Cutting, 1977; Mather, Murdock,
1994; Tro�e, 2002), эмоциональное состо-
Tro�e, 2002), эмоциональное состо-
, 2002), эмоциональное состо-
яние (Dittrich, Troscianko, Lea, Morgan,
1996) и даже размеры (Jokisch, Tro�e, 2003)
«человека, помеченного точками света».
Воспринимается ли движение биологических объектов теми же самыми областями
мозга, где обрабатываются другие типы движений? Хотя имеется много областей
мозга, активных при зрительном восприятии движения, из нескольких лабораторий
пришли данные о том, что движение биологических объектов возбуждает область в
задней верхней височной борозде (STSp), которая является одной зоной, вероятно
Рис. 6 .15. Исходный рисунок из рабо-
ты Йоханссона (Johannson, 1973), пока-
зывающий расположение точек света
на человеке (а) и как они выглядят при
удалении внешних очертаний человека
(б). (Источник: Johannson, 1973.)
*
Подобные исследования в 1920-х гг. проводил Н. А. Бернштейн. — Прим. ред.
266
Глава 6
Зрение
267
особенно чувствительной к движению живых объектов (Bonda, Petrides, Ostry, Evans,
1996; Howard, Brammer, Wright et al., 1996; Vaina et al., 2001).
Исследования профессора Эмили Гроссман (Emily �rossman) из Калифорнийского
университета в Ирвайне четко выявили области мозга и нервные сети, которые лежат
в основе восприятия движений биологических объектов (�rossman, Battelli, Pascual-
Leone, 2005; �rossman, Blake, 2001, 2002a, 2002б; �rossman, Blake, Kim, 2004; �ross-
, 2005; �rossman, Blake, 2001, 2002a, 2002б; �rossman, Blake, Kim, 2004; �ross-
�rossman, Blake, 2001, 2002a, 2002б; �rossman, Blake, Kim, 2004; �ross-
, Blake, 2001, 2002a, 2002б; �rossman, Blake, Kim, 2004; �ross-
Blake, 2001, 2002a, 2002б; �rossman, Blake, Kim, 2004; �ross-
, 2001, 2002a, 2002б; �rossman, Blake, Kim, 2004; �ross-
a, 2002б; �rossman, Blake, Kim, 2004; �ross-
, 2002б; �rossman, Blake, Kim, 2004; �ross-
�rossman, Blake, Kim, 2004; �ross-
, Blake, Kim, 2004; �ross-
Blake, Kim, 2004; �ross-
, Kim, 2004; �ross-
Kim, 2004; �ross-
, 2004; �ross-
�ross-
man, Donnelly, Price et al., 2000).
Но специализируется ли STSp на обработке движений именно человека? Гроссман со
своими сотрудниками изучила этот вопрос более детально и недавно сообщила, что
STSp была более восприимчива к движению человека, обозначенного точками света,
по сравнению с движением точек света, создаваемым многозвенными, но не похо-
жими на человека «созданиями» (рис. 6.16) (Pyles, �arcia, Hoffman, �rossman, 2007).
Таким образом, они сделали вывод, что STSp действительно является областью, спе-
STSp действительно является областью, спе-
действительно является областью, спе-
циализирующейся на кодировании движений человека, а не на только сложных дви-
жениях или динамическом изображении.
Литература
1. Bonda E., Petrides M., Ostry D., Evans A. (1996). Specific involvement of human pari-
etal systems and the amygdala in the perception of biological motion. Journal of neuro-
science, 16 (11). 3737 –3744 .
2. �rossman E., Battelli L., Pascual-Leone A., 2005. TMS over STSp disrupts perception of
biological motion. Vision research, 45 (22). 2847 –2853.
3. �rossman E., Blake R., 2002a. Brain areas active during visual perception of biologial
motion. Neuron, 35 (6). 1157–1165.
4. �rossman E., Blake R., 2002б. An investigation of neural activity associated with viewing
point-light animal, face and hand movements. Journal of vision, 1 (13). Abstract 359.
5. �rossman E., Donnelly M., Price R., Pickens D., Morgan V., Neighbor �. et al.,
2000. Brain areas involved in perception of biological motion. Journal of cognitive
neuroscience, 12 (5). 711 –720.
6. �rossman E.D ., Blake R., 2001. Brain activity evoked by inverted and imagined biologi-
cal motion. Vision research, 41. 1475–1482 .
7. �rossman E.D., Kim C.Y., Blake R., 2004. Learning to see bilolgical motion: brain activ-
ity parallels begavior. Journal of cognitive neuroscience, 16 (9). 1 –11 .
8. Howard R.J., Brammer M., Wright I., Woodruff P.W., Bullmore E.T., �eki S., 1996.
A direct demonstration of functional specialization within motion-related visual and
auditory cortex of the human brain. Current biology, 6. 1015–1019.
9. Johannson �., 1973. Visual perception of biological motion and a model for its analysis.
Perception and psychophysics, 14. 201 –211 .
10. Jokisch D., Tro�e N.F., 2003. Biological motion as a cue for the perception of size.
Journal of vision, V. 3, No 4 (1). 252–264 .
11. Vaina L.M., Solomon J., Chowdhury S., Sinha P., Belliveau J.W., 2001. Functional neu-
roanatomy of biological motion perception in humans. Proceedings of the national acad-
emy of sciences of the United States of America, 98 (20). 11656–11661 .
12. Pyles J.A., �arcia J.O ., Hoffman D.D ., �rossman E.D., 2007. Visual perception and neu-
ral correlates of novel «biological» motion. Vision research, 47. 2786 –2797.
Рис. 6 .16. (а) Показаны три полностью снабженных источниками света «создания»
и изображение их движения из точек света. (б) Полученные результаты работы моз-
га для исследования движения «созданий» и человека. Заметьте, что область STSp
сверху панели более активна в случае движений человека, чем в случае беспорядоч-
ного движения. (Источник: Pyles et al., 2007.)
268
Глава 6
Выяснилось, что ответ не такой простой, но ученые собирают важные
ключевые факты. Даже если глаза человека закрыты или человек больше не
может видеть из-за повреждения глаз или ЛКТ, он продолжает «чувствовать
способность видеть», если применить электрическую стимуляцию первичной
зрительной коры. Другими словами, возможно обойти возбуждение сетчат-
ки и ЛКТ и вызывать зрительные впечатления стимуляцией непосредственно
зоны V1.
Возможно ли обойти первичную зрительную кору и вызвать четкие зритель-
ные впечатления? Мы еще не знаем определенный ответ, но и на настоящий мо-
мент ответ, вероятно, «нет». Первичная зрительная кора кажется необходимой
для способности осознанно воспринимать любую визуальную особенность,
в то время как высшие зрительные поля могут играть более тонкую роль в вос-
приятии определенных зрительных признаков или объектов (Tong, 2003). Как
мы увидим в оставшейся части главы, различные корковые зрительные зоны,
вероятно, играют разные роли в нашем сознательном зрительном восприятии.
Сейчас развивается точка зрения, что одни и те же области мозга и нейроны, во-
влеченные в обработку специфических типов зрительных стимулов, таких как
ориентация, движение, лица или объекты, также включены в представлении
этих типов стимулов в сознании. Многие нейроны более активны, когда чело-
век осознает то, что видит какой-либо стимул, чем когда стимул показывается,
но не может достичь сознания. Итак, вероятно, нет какой-либо определенной
области мозга, которая является единственной ответственной за осознание. На-
оборот, кажется, что многие области головного мозга работают вместе, чтобы
достичь этого замечательного искусства. Области мозга, ответственные за вни-
мание, также важны для способности воспринимать и отвечать на зрительные
и другие сенсорные сигналы; эта тема будет раскрыта в гл. 8 .
3.1. Иерархическая и интерактивная теории зрения
Согласно одной из теорий зрительного осознания, называемой иерархической
теорией (Crick, Koch, 1995; Rees et al., 2002), сознание организовано в иерар-
хической манере, при этом чем выше уровень зрительной области, тем более
тесно она связана с нашим внутренним сознательным опытом (рис. 6 .17 (а)).
Эта теория согласуется с представлением о том, что высшие зрительные поля
отвечают на более сложные стимулы, такие как целые объекты, и могут инте-
грировать информацию о многих зрительных признаках, которые обрабатыва-
ются в более ранних зрительных областях. Однако в этом случае как быть с тем
фактом, что мы можем осознавать характерные зрительные признаки или очень
тонкие детали о положении в пространстве: ту информацию, которая наиболее
представлена в первичных зрительных полях, таких как V1?
Интерактивная теория зрительного осознания подчеркивает другую точ-
ку зрения. Оказалось, что сигналы, входящие в мозг, не просто идут вверх по
иерархической структуре зрительных полей: высшие зрительные поля посыла-
ют сигналы обратной связи обратно вниз к первичным зрительным областям,
особенно зоне V1 (Bullier, 2001). В зрительной системе имеется столько же про-
екций обратной связи: нейронов, которые идут от высших зрительных областей
к более низким, сколько имеется прямых проекций. Согласно интерактивной
Зрение
269
теории, в то время как предъявляется стимул, сигналы прямой связи идут по
зрительной иерархической структуре, возбуждая многие нейроны по своему
пути, но этой активности недостаточно для осознания. Вместо этого, области
высшего уровня должны посылать сигналы обратной связи назад к областям
более низкого уровня, откуда выходят сигналы прямой связи, таким образом,
что нервные сигналы, возвращаясь, делают полный круг, образуя нервный цикл
(рис. 6.17 (б)) (Pollen, 1999; Lamme, Roelfsema, 2000). Почему эта совокупность
прямых и обратных сигналов может быть важной для осознания? Возможно,
потому, что высшим областям необходимо проверять сигналы низших областей
и подтверждать, что они получают правильные сообщения, или, возможно, для
того, чтобы связывать нервное представление об объекте с характерными осо-
бенностями, которые составляют объект.
До настоящего времени неясно, достоверность какой из теорий будет дока-
зана, и имеется вероятность, что обе теории охватили некоторые части большей
картины. Возникла точка зрения, что наш сознательный опыт может быть ото-
бражением принципа распределения деятельности мозга, включающего многие
зрительные области, вид диалога между нейронами в первичных зрительных
областях, включая V1, и высшими зрительными областями, такими как вен-
V1, и высшими зрительными областями, такими как вен-
1, и высшими зрительными областями, такими как вен-
тральная височная кора и теменная доля. Это говорит о том, что вклад каждой
области может быть до некоторой степени различным и в некоторых случаях
единственным. В следующих частях этой главы мы опишем наиболее яркие
стороны данного утверждения.
Рис. 6 .17. Иерархическая и интерактивная модели зрительного сознания. (а) В иерар-
хической модели с каждым дальнейшим шагом в обработке зрительной информации
все более вероятно возникнет понимание. (б) В интерактивной модели для достижения
понимания необходимы сигналы обратной связи от позднее обрабатывающих информа-
цию полей к первичным. В настоящее время неясно, какая из двух моделей наилучшим
образом описывает путь, при котором в результате деятельности мозга возникает пони-
мание зрительных образов. (Источник: Frank Tong, с разрешения.)
а
б
270
Глава 6
Движения глаз и зрительное исчезновение
Зрительное исчезновение — явление, при котором неизменяемые зрительные сти-
мулы исчезают из восприятия. Нервной основой зрительного затухания является
привыкание (адаптация) нейронов, при котором зрительные нейроны уменьшают
или прекращают ответ на неизменяемые зрительные стимулы.
Зрительное затухание и адаптация нейронов
Нервная система возникла, чтобы замечать изменения в окружающей среде. Мы мо-
жем замечать неподвижные объекты только потому, что изображения, которые про-
ецируются на нашу сетчатку, никогда не бывают постоянными длительное время:
даже во время зрительной фиксации наши глаза никогда не бывают неподвижными.
Чтобы провести опыты по зрительному исчезновению, мы должны устранить все
эффекты движений глаз таким образом, чтобы визуальные изображения неизменяе-
мых объектов остались на сетчатке постоянными, что приведет к адаптации ответов
нейронов.
Зрительное исчезновение в условиях постоянного изображения на сетчатке
Чтобы достичь полной стабилизации изображения на сетчатке, мы должны исклю-
чить все движения глаз. Это сложно, потому что наши глаза находятся в постоянном
движении (рис. 6 .19 (а)). Даже когда мы фиксируем взгляд точно на объекте внима-
ния мы тем не менее постоянно совершаем небольшие непроизвольные движения
глаз, которые называют фиксационными движениями глаз (рис. 6 .19 (б)). Эти кро-
шечные движения глаз обычно эффективны для предотвращения нервной адапта-
ции и зрительного исчезновения.
Физическим образом удерживать глаза человека в неподвижном состоянии (напри-
мер, с помощью парализующих лекарств) технически очень сложно и потенциально
опасно для здоровья испытуемого. Как правило, для исследования на неподвижной
сетчатке обычно используют альтернативный подход: перемещение зрительного
стимула происходит таким образом, чтобы все движения глаз были эффективно
скомпенсированы. То есть зрительный стимул должен двигаться в том же направле-
Рис. 6 .18 . (а) Сюзанна Мартинес-Конде (Susana Martinez-Conde), лаборатория зри-
тельной нейробиологии, Нейрологический институт Барроу, Финикс, штат Аризо-
на, США. (б) Стефан Л. Мэкник (Stephen L. Macknik), лаборатория нейрофизиоло-
гии поведения, Нейрологический институт Барроу, Финикс, штат Аризона, США
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
Зрение
271
Рис. 6.19. Движения глаз и зрительное исчезновение. (а) Наблюдатель видит изо-
бражение (слева), при этом контролируется положение глаз (справа). Глаза пры-
гают, как кажется, фиксируются или остаются неподвижными на мгновение, что
образует небольшую точку на записи, затем прыгают в новую область привлечения
внимания. Большие скачки в положении глаз, какие мы привели в данном примере,
называются саккадами. Однако даже во время фиксации, или времени «отдыха»,
глаза никогда не остаются неподвижными, они постоянно производят фиксацион-
ные движения: дрейф, тремор и микросаккады. (Источник: Yarbus, 1967) (б) Упро-
щенное изображение фиксационных движений глаз у человека и приматов. Микро-
саккады (прямые и быстрые движения), дрейф (волнистые медленные движения)
и тремор (колебания, наложенные на дрейф) сдвигают зрительное изображение
вдоль мозаики фоторецепторов на сетчатке. (Источник: Martinez-Conde, Macknik,
2007.) (в) Исчезновение Трокслера. В 1804 г. швейцарский философ Игнац Пауль
Виталис Трокслер (Ignaz Paul Vital Troxler) обнаружил, что сознательная фикса-
ция на чем-либо вызывает исчезновение окружающих неподвижных изображений.
Чтобы выявить это, пристально смотрите на центральную точку, одновременно
удерживая внимание на окружающем ее бледном круге. Вскоре круг пропадет, и
будет казаться, что центральная точка находится на белом фоне. Переведите глаза,
и он опять появится в поле зрения. (Источник: Martinez-Conde, Macknik, Hubel,
2004, с изменениями.) (г) Микросаккады служат причиной зрительного восприятия
при фиксации людьми взгляда. Человек пристально смотрит на маленькую точку в
центре компьютерного экрана, что приводит к тому, что неподвижная цель на пе-
риферии исчезает из поля зрения и затем опять появляется. Как раз перед тем, как
цель исчезает, микросаккады наблюдателя становятся редкими, а незадолго до того,
как она появится снова, эти движения глаз становятся более частыми. (Источник:
Martinez-Conde, Macknik, 2006.)
272
Глава 6
нии, с теми же скоростью и амплитудой, как и глазное яблоко, таким образом изо-
бражение на сетчатке остается неподвижным, несмотря на движения глаз.
Первые эксперименты по неподвижности глаз выявили, что изображения исчезают
только после нескольких секунд неподвижности. Однако Коппола и Первис (Cop-
Cop-
pola, Purves, 1996) ранее показали, что изображения тени сосудов сетчатки (которые
чрезвычайно стабильны по отношению к глазу) исчезают меньше, чем за 80 мс.
Предполагают, что визуальная обработка в норме использует очень быстрый меха-
низм создания и уничтожения изображения.
Зрительное исчезновение без стабилизации изображения на сетчатке
Хотя идеальная неподвижность сетчатки наиболее легко достигается в лабора-
торных условиях, исчезновение неподвижных объектов, особенно на зрительной
периферии, довольно часто происходит при повседневном использовании зрения.
В 1804 г. швейцарский философ Игнац Пауль Виталис Трокслер (Ignaz Paul Vital
Troxler) заметил, что сознательная фокусировка взгляда на чем-то служит причиной
того, что неподвижные изображения в близлежащей области постепенно исчезают
(рис. 6.19 (в)).
Мартинес-Конде с коллегами (Martinez-Conde et al., 2006) показали, что исчезнове-
ние Трокслера происходит благодаря подавлению микросаккад (наиболее крупный
и быстрый тип фиксационных движений глаз) во время четкой и продолжительной
фиксации. Образование микросаккад уменьшается как раз перед исчезновением
зрительной цели и служит признаком того, что отсутствие микросаккад приводит к
адаптации нейронов и исчезновению восприятия. И наоборот, образование микро-
саккад достигает максимума непосредственно перед обратным появлением цели.
Эти результаты впервые показали, что микросаккады играют решающую роль в
стимуляции зрительного восприятия во время фиксации (рис. 6.19 (г).
Последствия для осознанного восприятия зрительных объектов
Фиксационные движения глаз препятствуют зрительному исчезновению непод-
вижных объектов и стимулируют их зрительное восприятие во время фиксации
взгляда. Таким образом, фиксационные движения глаз могут помочь удерживать
пространственно-временные характеристики осознанно видимых стимулов. Более
того, ответы нейронов, которые инициируют фиксационные движения глаз вдоль
зрительного пути, должны помочь сформировать нервный код для восприятия зри-
тельного мира.
Литература
1. Coppola D., Purves D., 1996. The extraordinarily rapid disappearance of entopic images.
Proceedings of the National academy of sciences of the United States of America, 93,
8001–8004 .
2. Martinez-Conde S., Macknik S.L., 2007. Windows on the mind. Scientific American,
297, 56–63 .
3. Martinez-Conde S., Macknik S.L., Hubel D.H., 2004. The role of fixational eye
movements in visual perception. Nature reviews neuroscience, 5, 229–240 .
4. Martinez-Conde S., Macknik S.L ., Troncoso X., Dyar T.A., 2006. Microsaccades
counteract visual fading during fixation. Neuron, 49, 297–305.
5. Yarbus A.L., 1967. Eye movements and vision. N .- Y.: Plenum press. 222 p. (Впервые
издана на русском языке: Ярбус А.Л ., 1965. Роль движений глаз в проц ессе зрения.
М.: Наука. 166 с.)
Зрение
273
4.0 . Области мозга, необходимые для визуального
восприятия: исследования повреж дений
В предыдущем разделе мы узнали о том, как нейроны различных зрительных
зон отвечают на определенные визуальные признаки, объекты или информацию
о положении в пространстве. Такие данные с записей от одного нейрона или
полученные при нейросканировании мозга человека могут указывать на то, что
отдельная область мозга играет роль в обработке определенного типа стимулов,
но не могут показать, является ли эта область мозга действительно необходи-
мой для восприятия данного стимула. Возможно, эту область мозга можно бы
было удалить без причинения каких-либо затруднений для восприятия. Напри-
мер, если многочисленные области мозга вовлечены в обработку определенно-
го визуального признака (например, движения), то повреждение любой одной
области мозга может не снизить возможность воспринимать этот признак, так
как другие незатронутые области смогут компенсировать эту утрату.
Исследования повреждений мозга важны для понимания, какие области
мозга могут быть необходимыми для определенных типов зрительного пони-
мания — восприятия цвета, движения, лиц, объектов или способности осоз-
навать то, что ты видишь что-нибудь вообще! Повреждения мозга могут быть
воспроизведены в экспериментальных условиях при исследовании животных
или могут изучаться на людях, которые пострадали от неудачного поврежде-
ния определенных частей мозга, что может случиться в результате ударов, опу-
холей, травм или нейродегенеративных заболеваний. Ухудшение зрения вслед-
ствие повреждений определенных частей зрительной системы может очень
значительно снижать здоровье. Однако при изучении таких пациентов можно
понять невральные причины ухудшения их состояния, что может дать ученым
информацию о функционировании мозга и в итоге привести к новым возмож-
ностям в лечении таких нарушений.
4.1. Последствия повреждений первичных зрительных зон
Различные ухудшения зрения могут возникнуть в результате повреждений
нервной системы на различных уровнях иерархического порядка обработки
зрительных сигналов. Повреждения сетчатки или зрительного нерва одного
глаза могут привести к монокулярной слепоте — потери зрения одного глаза.
Повреждения ЛКТ, зрительной лучистости, идущей к V1, или самой V1 могут
привести к потере зрения в контралатеральном (находящемся с противополож-
ной стороны) зрительном поле (см. рис. 6 .7). Повреждение небольшой части V1
может привести к явно определяющемуся дефекту поля зрения, области зри-
тельного поля, в которой отсутствует восприятие. Первые ретинотопические
карты V1 были фактически составлены путем картирования траектории пуле-
V1 были фактически составлены путем картирования траектории пуле-
1 были фактически составлены путем картирования траектории пуле-
вых ран солдат, раненых в Русско-японской войне и в Первой мировой войне.
Имелась явная связь между расположением каждого слепого пятна в поле зре-
ния и поврежденной частью V1 (Inouye, 1909; Holmes, 1918).
274
Глава 6
4.1.1. V1 и «слепозрение»
Приводят ли повреждения V1 к полной потере зрения? В 1965 г. исследователи
изучали этот вопрос на макаке-резусе, которую звали Хелен, после того как у
нее была удалена большая часть зрительной коры. В первые 19 месяцев Хелен
демонстрировала поведение, которое предполагало, что она была полностью
слепа. Однако оказалось, что ее зрение постепенно возвращается. Со временем
обезьяна могла передвигаться между объектами в переполненной комнате и
даже протягивать руку, чтобы схватить пролетающую муху. Вначале она могла
только видеть и достигать объектов, которые двигались. Затем по прошествии
времени она отвечала на вспышки света, затем на постоянный источник све-
та и, наконец, на неподвижный темный объект, находящийся на светлом фоне
(Humphrey, Weiskrantz, 1967; Humphrey, 1974). Это прекрасный пример того,
как может происходить восстановление функций после повреждения мозга. Хе-
лен была способна определять местонахождение хорошо заметных объектов,
но, казалось, была неспособна узнавать их.
Осознавала ли Хелен то, что она видит, или она была способна производить
действия, регулируемые зрением, несмотря на потерю понимания зрительных
стимулов? Трудно спросить животное, осознает оно что-то или нет, но недав-
нее исследование обезьян с односторонним повреждением V1 предположило,
что они могли не осознавать того, что видят. В этом исследовании обезьяны
могли безошибочно передать расположение объектов в их «слепой» половине
зрительного поля, если их заставляли сделать выбор между двумя возможными
вариантами. Однако если им давали задание сообщить, имеется объект или нет,
а объект иногда располагался в здоровой половине зрительного поля, в слепой
половине зрительного поля или вообще отсутствовал, они были неспособны
передать, что объекты находятся на слепой половине зрительного поля (Cowey,
Stoerig, 1995; Stoerig et al., 2002), т. е . вели себя так, как если бы эти объекты
были «невидимы».
Интересно, что люди с повреждениями V1 могут демонстрировать результат,
превышающий вероятностный, даже если они сообщают о потере зрительного
восприятия в своей слепой половине зрительного поля. Пациент ДБ страдал
от чрезвычайно сильных мигреней из-за венозной опухоли, расположенной в
коре его правой шпорной борозды (V1). Хирургическое удаление этой опухоли
привело к потере у него правой первичной зрительной коры. Как вы можете
ожидать, эта процедура сделала его практически слепым в левом зрительном
поле. Однако интересно, что, когда систематически тестировали его слепую по-
ловину зрительного поля, поведение пациента предполагало иное.
Вейскранц с сотрудниками (Weiskrantz et al., 1974) доложили, что, когда ДБ
просили указать цель в его слепом зрительном поле, тот утверждал, что он не
может вообще ничего видеть, однако он мог указать достаточно точно на распо-
ложение источника света. Также он безошибочно говорил, была ли палка накло-
нена вертикально или горизонтально. Эта способность выполнять визуальные
задания на уровне выше теории вероятности, вопреки заявлению пациентов о
потере любых зрительных ощущений, известна как «слепозрение» (blindsight).
В других экспериментах задачей ДБ было различение Х и О. Его действия были
Зрение
275
практически безошибочными и улучшались в зависимости от размера и про-
должительности предъявления стимулов. Во время всех этих экспериментов
ДБ настаивал, что он ничего не видит. Интересно, что, когда на него оказали
давление, он описал, что иногда имеет ощущение «гладкости» и «зубчатости»
при различении Х и О, но отрицал, что эти ощущения были связаны с каким-
либо зрительным впечатлением. Каким же образом это возможно? Как может
ДБ указывать или различать зрительные стимулы без возможности видеть их?
Эти данные предполагают, что между обработкой зрительных сигналов в
мозге и субъективным сознанием человека может быть разобщение: зрительной
системы ДБ достигает достаточная информация для того, чтобы позволить ему
различать стимулы в эксперименте с несвободным выбором, но этой инфор-
мации недостаточно, чтобы обеспечить понимание (Weiskrantz, 1986). Однако
появляется вопрос, полностью ли утеряно опознание зрительных стимулов в
случае слепозрения. Пациентов можно вынудить сообщать о слабых зритель-
ных впечатлениях, которые, однако, являются достаточными для процесса раз-
личения в экспериментах с несвободным выбором. Подобные эффекты могут
происходить со здоровыми людьми под воздействием условий, близких к по-
роговой чувствительности. Некоторые пациенты со слепозрением сообщают об
остаточных впечатлениях от хорошо заметных движущихся стимулов, которые
они описывают как «движение черных объектов на черном», но, как правило,
они говорят, что ничего не видят, когда им показывают неподвижные стимулы.
Зрение при «слепозрении» серьезно нарушено, но не всегда полностью отсут-
ствует.
Неповрежденность экстрастриарной коры может быть решающим факто-
ром для слепозрения, так как пациенты, которым удалили целое полушарие, де-
монстрировали мало признаков наличия остаточных зрительных способностей
(Fqubert et al., 1999). Хотя путь от сетчатки через ЛКТ до V1 обеспечивает по-
V1 обеспечивает по-
1 обеспечивает по-
давляющее большинство зрительных входящих сигналов в кору, несколько ре-
зервных подкорковых путей идут к экстрастриарной области, минуя V1. Записи
от одного нейрона у обезьян показали, что зрительная информация может все
еще достигать экстрастриарной области после повреждения V1. Хотя уровень
возбуждения снижен, существенная часть нейронов в двигательной средней
височной зоне МТ и V3 все еще выборочно реагируют на зрительные стимулы
(Rodman et al., 1989; �irard et al., 1991). Недавние исследования по нейроска-
нированию показывают, что неосознаваемые стимулы, находящиеся в слепом
поле зрения, при исследовании путем функционального магнитно-резонанс-
ного отображения (фМРТ) все еще вызывают сильный ответ чувствительной
к движению зоны МТ, цветочувствительной зоны V4 и областей, вовлеченных
в восприятие объекта (�oevel et al., 2001). Таким образом, значительная избира-
тельность к стимулам поддерживается в экстрастриарной коре, хотя эта актив-
ность оказывается недостаточной, чтобы поддерживать осознание человеком
при отсутствии V1. Это согласуется с предсказаниями интерактивной модели.
Однако возможно, что экстрастриарные сигналы слишком неустойчивы или
слабы, чтобы поддерживать осознанное восприятие, но достаточны, чтобы
обеспечивать различение в экспериментах с несвободным выбором.
276
Глава 6
4.2 . Поражения экстрастриарной зоны —
повреждения вне зоны V1
Повреждения V1 могут исключать практически все визуальное восприятие
в соответствующих областях зрительного пространства, даже если, казалось
бы, достаточное количество визуальной информации достигает экстрастриар-
ных областей, чтобы обеспечивать поведение при слепозрении. Это само по
себе информирует нас о роли V1 как посредника при зрительном осознанном
восприятии. Так что же происходит при повреждении экстрастриарных об-
ластей? На рис. 6 .20 представлена схематическая карта зрительных областей
и соответствующих нарушений.
4.2.1 . Нарушение восприятия движения (акинетопсия)
Возможно потому, что мы так восприимчивы к наблюдению движения, очень
редкие нарушения мозга приводят к полной потере возможности воспринимать
движение. Однако имеется поразительный пример одной пациентки. Она не
могла больше воспринимать движения после мучительных приступов, которые
привели к обширным повреждениям с двух сторон, включавшим зону МТ и
обширные близлежащие районы. Для этой пациентки мир казался серией не-
подвижных кадров, как будто она живет в стробоскопическом мире. Простые
задачи, такие как пересечение улицы, становились опасными, потому что она
не могла определить, как быстро приближаются машины (рис. 6 .21 (а)). Даже
наливание чашки кофе становилось сложной проблемой, потому что она не
могла сказать, насколько быстро поднимается уровень жидкости, поэтому чаш-
ка переполнялась.
В других исследованиях было обнаружено, что более локальные поврежде-
ния, только зоны МТ, обычно приводят к менее сильным нарушениям в способ-
ности воспринимать движения, и как люди, так и животные могут также демон-
стрировать значительное восстановление с течением времени (Plant et al., 1993;
Рис. 6 .20 . Нарушения зрения и об-
ласти мозга. Области мозга, в кото-
рых повреждение может в резуль-
тате привести к сопутствующим
зрительным нарушениям. Области
мозга здесь показаны только для од-
ного полушария, хотя для некоторых
таких нарушений, как нарушение
восприятия движения, необходи-
мо повреждение обоих полушарий.
(Источник: Frank Tong, с разреше-
ния.)
Зрение
277
Pasternak, Merigan, 1994). Итак, очевидно, что зона МТ является очень важной
для восприятия движений, но другие зрительные поля могут способствовать
восприятию движений, даже если МТ повреждена.
4.2.2. Корковый дефицит цветового зрения
Повреждение вентральной области V4 может привести к корковому дефициту
цветового зрения или к тому, что иногда называют ахроматопсией (Meadows,
1974 a; Bouvier, Engel, 2006). Пациенты описывали, что, по их мнению, из мира
убрали краски, и он состоит из серых теней, что, вероятно, похоже на фото-
графию на рис. 6 .21 (б). Эти люди все еще могут воспринимать границы меж-
ду цветами, но имеют трудность с различением самих цветов. Ахроматопсия
обычно связана с повреждениями, которые затрагивают зону V4 и, возможно,
области как раз впереди от зоны V4. Повреждение одного полушария может
даже привести к выборочной потере восприятия цвета в противоположном зри-
тельном поле.
4.3 . Повреждение вентральных областей,
воспринимающих предметы
4.3.1 . Зрительная агнозия
Пациенты с зрительной агнозией имеют трудности с распознаванием предме-
тов из-за нарушений в обработке основных сенсорных сигналов либо в процес-
сах распознавания высшего уровня. Такие пациенты все же могут распознавать
предметы, используя другие чувства, такие как осязание, слух или запах, поэто-
му потеря этой функции связана только со зрением. Слово агнозия может быть
переведено с греческого как «потеря знания о», поэтому зрительная агнозия
предполагает потерю сведений, получаемых с помощью зрения.
Здесь мы обсудим три типа зрительной агнозии: апперцептивная агнозия,
ассоциативная агнозия и прозопагнозия. Пациенты с апперцептивной агнозией
могут по-прежнему замечать появление визуально представленных отдельных
Рис. 6 .21. Нарушения восприятия движения и цвета. (а) Повреждение воспринимаю-
щей движение зоны МТ в обоих полушариях может привести к потере восприятия дви-
жения: акинетопсии. Пациенты описывают то, что они видят многочисленные непод-
вижные кадры вместо плавного движения. Это может сделать простые задачи, такие как
пересечение дороги, сложными и опасными. (б) Повреждение зон, воспринимающих
цвет, в коре только одного полушария может привести к потере восприятия цвета только
с одной стороны визуального пространства. Корковый дефицит цветового зрения назы-
вается ахроматопсией. (Источник: Frank Tong, с разрешения.)
278
Глава 6
элементов, но имеют трудности с восприятием их формы и не могут опознать
или назвать их. Эти пациенты обычно неспособны пройти тесты на повторе-
ние формы и имеют трудности с копированием очень простых форм, таких как
круг, квадрат или даже, возможно, единственной наклонной линии. Отравление
угарным газом является частой причиной апперцептивной агнозии, так как оно
может привести к массивному повреждению всей площади затылочной доли.
Замечательно, что некоторые больные апперцептивной агнозией показыва-
ют признаки неосознанной обработки данных по визуальным признакам, кото-
рые они не могут воспринимать осознанно. Гудейл (�oodale et al., 1991) иссле-
довал пациентку ДФ, больную апперцептивной агнозией, которой было трудно
сообщить об ориентации простых линий или материальных предметов. Когда
ее просили сообщить о расположении узкой щели, вырезанной в поверхности
барабана, она не могла сообщить об угле, который образовывала эта щель, и де-
лала много ошибок (рис. 6.22, вверху; барабан поворачивали в новом направле-
нии для каждой попытки). Однако когда ее просили опустить открытку через
щель, она могла это делать с замечательной точностью (рис. 6 .22 внизу) и даже
не ожидала, что сама может выполнять это. Удивительно, что, когда ее попро-
сили только держать у себя письмо и поворачивать его, чтобы оно соответство-
вало углу щели, ее результаты были опять слабыми и она сообщала, что щель
кажется «менее ясной», чем тогда, когда ей позволяли опускать открытку. Что
может объяснить такую разницу в поведении, как способность сообщить об
угле щели и выполнить визуально управляемое действие?
Рис. 6 .22 . Пациентка может это сделать,
хотя и не может об этом сообщить. Резуль-
таты исследуемой ДФ. Каждая линия пред-
ставляет одну из попыток ДФ, при которой
она либо подбирала соответствие ориента-
ции почтовой щели, либо действительно
опускала в него письмо. В верхней части
показаны попытки ДФ найти соответствие
ориентации щели по сравнению с открыт-
кой (никаких действий не выполнялось).
В этом случае линии распределены по кру-
гу, что наводит на мысль, что у ДФ имеют-
ся трудности с восприятием ориентации.
Однако нижняя часть показывает точность
ДФ в опускании открытки в щель с задан-
ным направлением, при этом правильное
направление на схеме каждый раз показано
вертикально. Так как все линии сгруппиро-
ваны около вертикали, это дает основание
предполагать, что ДФ точна при выполне-
нии действия опускания открытки в щель.
Пациентка ДФ может опускать письма в по-
чтовый ящик без всяких трудностей: ее рука
знает, что нужно делать. Но если ДФ попро-
сить найти соответствие ориентации щели,
она выполняет это очень плохо. (Источник:
Frank Tong, с разрешения.)
Зрение
279
Пациентка ДФ предоставляет веские доказательства для предположения,
что существуют отдельные пути обработки информации о том, «чем» является
объект и «где» он находится по отношению к выполнению визуально управля-
емого действия. Согласно Гудейлу и Милнеру, пациентка ДФ имеет поврежде-
ния вентрального зрительного пути, но незатронутой область обработки ин-
формации в дорсальном пути, что недавно было подтверждено исследованиями
по сканированию мозга (James et al., 2003). Они предполагают, что дорсальная
система ответственна не только за обработку того, «где» находятся объекты,
но также за то, «как» действия могут быть выполнены по отношению к опре-
деленному объекту, такие как указать на объект или достать его. Явно, что об-
работка визуальной информации в дорсальной системе недоступна сознанию:
пациент не может сообщить об ориентации щели, однако дорсальная система
может управлять правильным действием. Дополнительные исследования паци-
ентов со зрительной атаксией показали противоположный пример нарушений
зрения, указывая на двойное разобщение между сознательным зрительным вос-
приятием и действием, регулируемым зрительно. Зрительная атаксия обычно
возникает в результате повреждения теменной доли, которая является частью
дорсального пути. Такие пациенты с помощью зрения могут воспринимать
ориентацию и хорошо узнавать объекты, но имеют большие трудности с вы-
полнением действий, управляемых с помощью зрительных сигналов.
Ассоциативная агнозия обозначает неспособность узнавать объекты, несмо-
тря на явно незатронутое восприятие объектов. Например, если попросить по-
вторить простой рисунок, то пациенты с ассоциативной агнознией сумеют сде-
лать приемлемую работу, особенно если им дать достаточно времени. Пример
для сравнения: это задание будет практически невозможно выполнить страда-
ющим апперцептивной агнозией.
Доктор Оливер Сакс описал пациента, который в результате повреждения
мозга, «принял свою жену за шляпу». У пациента были огромные трудности с
идентификацией объектов, несмотря даже на то, что его зрение было во всем
остальном нормальным и он мог описывать признаки того, что он видел. Ког-
да ему показали розу с лацкана доктора Сакса, пациент описал ее как «закру-
ченная спиралью красная структура с линейными зелеными прикрепленными
деталями», но, только нюхая ее, он узнал, что это роза. Когда его жена пришла
встретиться с ним в кабинете доктора, он нечаянно потянулся к ее голове, когда
захотел найти свою шляпу на вешалке (Sacks, 1985).
Ассоциативная агнозия обычно возникает в результате повреждения вен-
тральной височной коры. Обычно люди с ассоциативной агнозией имеют труд-
ности с распознаванием множества объектов, особенно объектов, относящихся
к одному классу, например таких, как лица. Однако как мы увидим ниже, это
частый, но не всегда единственный случай.
Хотя большинство больных зрительной агнозией будут с трудом различать
как лица, так и предметы, были сообщения о нескольких удивительных исклю-
чениях. Пациенты с прозопагнозией все еще способны хорошо узнавать пред-
меты, но имеют огромные трудности с узнаванием или различением именно
лиц (Bodamer, 1947; Meadows, 1974б). Нарушения могут быть серьезными:
некоторые больные прозопагнозией больше не могут узнавать близких членов
своей семьи или друзей, а вместо этого должны опираться на другую инфор-
280
Глава 6
Блок 6.2 . Как лица и предметы представляются мозгом
Имеется две основных теории о том, как зрительная система управляет обработкой
и узнаванием предметов и лиц. Мозг может это делать либо модульным образом, с
разными блоками (модулями) для обработки лиц или обработка может выполняться
раздельными путями среди многочисленных областей вентральной височной коры.
Согласно модульной точке зрения, восприятие объектов разбито по нейронным бло-
кам, специфическим зонам мозга, которые специализируются на обработке опреде-
ленной категории объектов. Исследования предполагают, что зона распознавания
лиц (ЗРЛ) может быть специализированным модулем для обработки и распознавания
правильно повернутых лиц (Kanwisher et al., 1997; Tsao et al., 2006). Кроме того, зона,
которая отвечает за представление о месте (например, доме), кажется, также является
специализированным блоком (Epstein, Kanwisher, 1998). Эта зона стала известна как
парагиппокампальная область восприятия пространства (ПОП). Склонность к мо-
дульному представлению объектов не распространяется на каждый объект; фактиче-
ски она в основном наблюдается только в двух вышеупомянутых случаях. Например,
в человеческом мозге нет зон восприятия бананов или туфель.
Интересным поворотом модульной гипотезы является гипотеза квалификации (ги-
потеза экспертной оценки), которая предполагает, что так называемая зона распоз-
навания лиц в действительности специализирована для распознавания объектов на
высококвалифицированном уровне (�authier et al., 2000). Вероятно, что больше всего
времени мы проводим, вглядываясь в лица, чем в любой другой объект (особенно
если включить рассматривание лиц в телевизоре). Остановитесь только на секунду
Рис. 6 .23. Эффект переворачивания лиц. Демонстрация того, как мы плохо распознаем
лица, перевернутые вниз. Взгляните на эти два изображения Барака Обамы. Замечаете
ли вы что-либо странное? А теперь переверните страницу сверху вниз и посмотрите
опять. Теперь вы должны увидеть, что одна из этих фотографий была серьезно иска-
жена. Этот эффект, называемый эффектом переворачивания лица, показывает, как наша
зрительная система приспособлена к обработке лиц, повернутых вверх
Зрение
281
мацию, такую как голос или одежда, чтобы узнать этого человека. Некоторые
пациенты больше не могли узнавать собственные лица на фотографии или даже
отражение в зеркале.
Возникает ли прозопагнозия действительно благодаря специфическому
ухудшению в узнавании лиц? Может быть, узнавание лиц просто более затруд-
нено, чем другие формы узнавания предметов? В конце концов, лица в зритель-
ном плане очень похожи одно на другое: каждое лицо имеет ту же самую основ-
ную форму и топологию признаков, в то время как другие предметы намного
сильнее отличаются друг от друга (см. блок 6.2).
Исследования выявили несколько пациентов, которые могли находить от-
личия при малейшей разнице в предметах, но больше не могли различать лица.
Например, фермер с прозопагнозией стал очень слабо различать лица людей,
но все еще мог узнавать разных овец в своем стаде (McNeil, Warrington, 1993).
Другой пациент больше не мог точно узнавать лица, повернутые вверх, но был,
как ни странно, более удачлив в узнавании лиц, перевернутых вниз, что точно
и подумайте о том, как много информации мы можем получить ото всех малейших
изменений чьего-либо лица, когда мы говорим с этим человеком, и вы вскоре пойме-
те, что вы в действительности являетесь экспертом по лицам. Было предположено,
что ЗРЛ ответственна за процесс распознавания любого объекта, по которому мы яв-
ляемся «экспертами». Исследования показали, что, разглядывая изображения птиц,
специалисты по птицам показывают в некоторой степени большую активность в ЗРЛ,
чем не специалисты по птицам (�authier et al., 2000). Является ли ЗРЛ специфиче-
ской зоной распознавания лиц или любого объекта на высококвалифицированном
уровне? Оба случая говорят о специализированных структурах, которые отличны от
областей вентральной височной коры и вовлечены в процесс обработки информации
о предметах. Посмотрите на два лица на рис. 6 .23, замечаете ли вы что-то странное?
А теперь переверните книгу вверх тормашками и взгляните опять. Видите ли вы это
сейчас? Это пример воздействия переворота лиц. Когда лица перевернуты вниз, мы
крайне плохо идентифицируем их. Вот пример того, как мы приспособлены к вос-
приятию лиц.
По другой гипотезе, мозг обрабатывает лица и предметы по раздельному пути сре-
ди многочисленных областей вентральной височной коры. Исследование Хаксби
с сотрудниками показало, что области вне зоны ЗРЛ тем не менее показывают воз-
буждение в ответ на лица, отличающееся по сравнению с ответом на другие типы
стимулов (Haxby et al., 2001). Даже если разница в возбуждении в ответ на лица и
различные предметы довольно мала во многих из этих областей, имеется достаточ-
но информации по всем таким зонам вне зоны ЗРЛ, чтобы утверждать о наличии
различий между лицами и предметами. Однако, согласно этой точке зрения, трудно
объяснить нейропсихологические данные по двойному разобщению между узнава-
нием лиц и предметов. Одним из возможных выводов является тот, что активность
высокоспециализированных по отношению к лицам нейронов в таких зонах, как
ЗРЛ, может быть важна для различения слабой разницы между лицами отдельных
людей. Но более распределенные формы активности вне этих высокоспециализи-
рованных по отношению к лицам областей достаточны для нахождения основных
различий между лицами и предметами или, возможно, для более заметных различий
между лицами (например, мужское лицо по отношению к женскому, молодое по
сравнению со старым).
282
Глава 6
противоположно нормальным действиям человека (Farah et al., 1995). Что мог-
ло бы быть причиной этому? Согласно одной гипотезе, мозг может иметь спе-
циализированные механизмы для обработки лиц, повернутых прямо вверх, в то
время как лица, повернутые вниз, обрабатываются общей системой узнавания
объектов (перевернутые лица продемонстрированы на рис. 6 .23). Итак, если
система узнавания лиц повреждена у пациента с прозопагнозией, то эта систе-
ма может привести к автоматическим ошибкам, когда пациенту предъявляются
лица в нормальном положении. Однако лица, перевернутые вниз, не будут ав-
томатически обрабатываться этой системой, позволяя, таким образом, системе
распознавания предметов выполнить эту задачу.
Возможно, самые веские данные для некоторого разделения систем по рас-
познаванию лиц и предметов получены из исследования очень необычного па-
циента СК с предметной агнозией, у которого были серьезные нарушения в
распознавании предметов, но который мог узнавать лица, повернутые вверх,
так же, как обычных людей (Moscovitch et al., 1997). Удивительно, что, если
лица были повернуты вниз, СК становился медлительным, выполняя задание
в шесть раз хуже, чем обычные участники в тех же условиях. Очевидно, что
СК имеет неповрежденную систему обработки нормально повернутых лиц,
а система, ответственная за обработку предметов и перевернутых вниз лиц,
серьезно повреждена. Взятые вместе данные о пациенте СК и больных про-
зопагнозией свидетельствуют о двойной связи между визуальной обработкой
лиц, направленных вверх, по сравнению с обработкой предметов и лиц, пере-
вернутых вниз.
Какой вид повреждений мозга приводит к прозопагнозии? Прозопагнозия
может возникнуть в результате двустороннего повреждения областей около бо-
ковой затылочной коры, нижней височной коры и веретенообразной извилины
(Meadows, 1974 б; Bouvier, Engel, 2006). В некоторых случаях одностороннее
повреждение правого полушария может привести к этому нарушению. Из-за
того что повреждения обычно довольно обширны и могут привести к повреж-
дению нервных волокон, идущих к необходимой области мозга, трудно иденти-
фицировать точное место. Тем не менее места повреждения мозга, связанные с
прозопагнозией, кажется, охватывают зону распознавания лиц и простираются
намного далее назад.
4.4 . Повреждение дорсальных областей теменной доли
Повреждение дорсальной области теменной доли (или верхней теменной из-
вилины) может привести к необычному изменению общего зрительного по-
нимания, которое называют игнорирование и при котором пациент полностью
игнорирует или не отвечает на объекты, находящиеся в противоположной по-
ловине зрительного поля (Driver, Mattingley, 1998). Пациенты с повреждением
правой теменной доли могут игнорировать левую половину зрительного поля,
есть только половину еды на своей тарелке или делать макияж только половины
своего лица. Они также могут игнорировать звуки или прикосновения, прихо-
дящие слева.
Этот синдром может напоминать нарушения зрительного восприятия. Од-
нако игнорирование встречается при отсутствии повреждения зрительной си-
стемы и может включать комбинированные нарушения, включая нарушения
Зрение
283
двигательной и тактильной систем. Более того, если пациентам дали указания
обращать внимание на игнорируемое поле, они могут иногда отвечать на эти
стимулы (Posner, 1980). Поэтому данный синдром лучше отнести к дефициту
внимания, чем к неспособности воспринимать стимулы. Без подачи специаль-
ных сигналов пациентам трудно воспринимать объекты в своем игнорируемом
поле. Мы не будем уделять много времени тщательному исследованию многих
интересных граней зрительного внимания, так как этому посвящена целая гла-
ва (см. гл. 8).
Двусторонние нарушения теменных областей могут привести к намного
более глубокому нарушению, которое называют синдром Балинта, который в
основном является нарушением пространственного внимания. Он может быть
охарактеризован тремя основными нарушениями: 1) оптической атаксией, не-
способностью указать на цель; 2) апраксией взора, неспособностью по своему
желанию переключать взгляд; и 3) симультанной агнозией, неспособностью
воспринимать более чем один объект в зрительном поле одновременно, даже
если объекты занимают большую часть пространства. Люди с синдромом Ба-
линта часто кажутся слепыми, так как в действительности они могут концен-
трироваться только на одной части зрительного пространства или одном объ-
екте одновременно и находят трудным по собственному желанию переключать
внимание на другие области. Эти данные опять выдвигают на первый план важ-
ность областей мозга, отвечающих за внимание, для поддержки зрительного
осознания.
5.0 . Связь деятельности мозга и зрительного опыта
В каждый конкретный момент в зрительной части вашего мозга возбуждены
миллионы нейронов. Активность некоторых из этих нейронов, вероятно, тесно
связана с тем, что вы сознательно воспринимаете здесь и сейчас, в то время
как активность других нейронов может быть сильно удалена от имеющихся в
настоящий момент ощущений. Если бы ученые могли как-то изолировать ней-
роны, которые точно отражают сознательный опыт человека, показало бы это,
какие нейроны или области мозга являются главными для сознания, а какие
нет? Данное исследование проводится для нейронов, активность которых кор-
релирует с изменениями в понимании и осознании того, что воспринимаешь,
таким образом, его назвали исследованием нейробиологического соответствия
сознательному состоянию (Krick, Koch, 1995).
Первая проблема включает разделение деятельности, запускаемой стиму-
лами, и деятельности, связанной с сознательным восприятием. Не все ответы
на зрительные стимулы являются сознательными, что можно вынести из того
факта, что многие зрительные зоны хорошо отвечают на стимулы, даже если
животное находится под наркозом.
Как тогда возможно измерить деятельность, связанную с осознанием и отде-
лить ее от любой возможной бессознательной активности, которая запускается
самими зрительными стимулами? Для этой цели ученые могут использовать
много методов. В следующем разделе мы обсудим некоторые из наиболее ши-
роко распространенных и полезных методов для изоляции нейронов, чья актив-
ность коррелирует с изменениями в зрительном осознании.
284
Глава 6
5.1. Восприятие со многими устойчивыми состояниями
Вы когда-нибудь в темноте, возможно, лежа в кровати, вглядывались в какие-то
странные очертания предмета на противоположной стороне комнаты? В один
момент они могут выглядеть как человек, затем как некоторое странное непод-
вижное животное, затем как статуя из камня. После того как ваш мозг напряга-
ется, чтобы опознать этот таинственный объект, вы в конце концов включаете
свет и — подумать только, это всего лишь тусклый свет, освещающий ваш пид-
жак на кресле. В этом примере, когда зрение затруднено и нечетко, восприятие
делает нечто необычное: оно меняется или колеблется между разными объекта-
ми. Это пример восприятия со многими устойчивыми состояниями: пиджак на
стуле (физический стимул) не изменяется, в то время как ваше восприятие —
да! Такой тип состояния является ценным инструментом, так как дает возмож-
ность ученым изучать изменения в зрительном осознании, независимо от лю-
бых изменений в зрительном стимуле.
Имеется много примеров моделей со многими состояниями или двусмыс-
ленных рисунков, которые ученые могут использовать, чтобы исследовать эти
нервные связи сознания. Рисунки, которые первоначально имеют только две
основные интерпретации, называются изображеними с двумя устойчивыми со-
стояниями (рис. 6 .24). Попытайтесь увидеть, можете ли вы воспринимать обе
интерпретации каждого из двусмысленных рисунков: лицо и вазу и два вида
куба Неккера. Восприятие с двумя устойчивыми состояниями также может на-
блюдаться, если вы зафиксируете глаза на единственной точке в центре стиму-
ла. В этом случае изображение на сетчатке и, таким образом, активность, вызы-
Рис. 6.24. Рисунки с двумя устойчивыми состояниями. (а) После того как вы неко-
торое время посмотрите на изображение, вы заметите, что имеется два его истолко-
вания. Одно — это ваза, стоящая в центре, второе — один контур, обрисовывающий
два лица, смотрящих друг на друга. Этот рисунок имеет два устойчивых состояния:
пока вы смотрите на него, ваше восприятие будет колебаться между вазой и лицами.
(б) Куб из проволочной рамки, обычно известный как куб Неккера, имеет две равно ве-
роятных интерпретации расположения в пространстве. Восприятие имеет склонность
к колебаниям между очертаниями самой близкой стороны, которая выступает вверх, и
самой близкой стороной, которая выступает вниз. Подобно вазе и силуэтам, упомяну-
тым выше, этот рисунок имеет два устойчивых состояния. Такая бистабильность по-
зволяет разделить слабую стимуляцию и осознание. Физически образец не изменяется,
но осознание — да!
Зрение
285
ваемая стимулом, является почти постоянной во времени, в то время как ваше
восприятие изменяется. Наблюдая за областями мозга, которые показывают из-
менения активности, взаимосвязанные с изменениями в восприятии, ученые
могут установить нейробиологические взаимосвязи сознания.
5.2 . Бинокулярная конкуренция: то, что вы видите, —
это то, что стимулирует вас
Одно из наиболее сильных и самых изученных примеров восприятия с двумя
устойчивыми состояниями — явление, называемое бинокулярной конкуренци-
ей. Если показывать два очень разных образца, по одному каждому глазу, то
из-за того, что они такие разные, мозг не может связать их вместе, как он обыч-
но делает. То, что происходит затем, довольно поразительно: осознание одного
образца длится несколько секунд, затем другой образец появляется и, по ощу-
щению, магическим образом стирает изображение, которое было видно до это-
го. Кажется, что два образца борются в мозге за ваше осознанное восприятие!
Если вы возьмете очки с зеленым и красным фильтрами, то вы сможете узнать
на опыте, что такое бинокулярная конкуренция, глядя на рис. 6 .25 (а). В другом
примере попытайтесь объединить два образа на рис. 6 .25 (б). (Чтобы их объеди-
нить, попытайтесь направить свои глаза так, чтобы два изображения казались
наложенными друг на друга. Точки в центре и окружающий их квадрат сольют-
ся. Если эти демонстрационные рисунки эффективны, то вы должны увидеть
один образец, в то время как другой будет полностью невидим, почти так, как
если бы вы закрывали один глаз, а затем другой.)
Что происходит в мозге во время бинокулярной конкуренции? Тонг с сотруд-
никами (Tong et al., 1998) пытался найти решение этой проблемы, концентри-
руясь на двух избирательных к категориям областях в вентральной височной
доле: ЗРЛ и ПОП. Они использовали очки с красным и зеленым фильтрами,
чтобы предъявить изображение лица одному глазу, и дома — другому, при этом
активность областей мозга измеряли с помощью функционального магнитно-
резонансного томографа (рис. 6 .25 (в)). В этом исследовании участники могли
вначале воспринимать дом, затем быстро переключиться на восприятие лица,
а затем обратно — что обычно для бинокулярной конкуренции (рис. 6 .26 (а)).
Интересно, что ЗРЛ была активна только тогда, когда испытуемые сообщали
о том, что они видят лицо. Подобным образом, ПОП была активной только тог-
да, когда участники говорили, что они видят изображение дома (рис. 6 .26 (б)).
Затем исследователи проверяли физические чередования двух изображений,
включая одно изображение в то время, как другое выключали. Получающиеся
в результате, регулируемые этими стимулами ответы в ЗРЛ и ПОП были при-
мерно такой же силы, как и те, которые были измерены во время бинокулярной
конкуренции и, к удивлению, не сильнее. Кажется, что активность в этих об-
ластях мозга близко отображает то, что наблюдатель воспринимает во время
конкуренции и не отражает временно подавляемый стимул, все еще возбужда-
ющий сетчатку.
Другие исследования мозга с помощью нейровизуализации выявили, что
активность на более ранних стадиях обработки зрительного сигнала тесно свя-
зана с тем, что воспринимает наблюдатель во время бинокулярной конкурен-
ции. Например, сильные, связанные с осознанием, модуляции были найдены
в V1 человека (Polonsky et al., 2000; Tong, Engel, 2001) и даже в латеральном
286
Глава 6
коленчатом теле (Haynes et al., 2005; Wunderlich et al., 2005). Эти результаты
поддерживают идею о том, что активность в первичных зрительных областях
может быть важной для осознания, как предполагается по интерактивной мо-
дели. Однако ученые все еще пытаются обнаружить, какая из моделей осозна-
ния (интерактивная или иерархическая) является лучшей для описания того,
как работает мозг.
Другим подходом является обучение обезьян отвечать на доминантный об-
раз во время бинокулярной конкуренции (рис. 6 .27). Это требует некоторой тре-
нировки, но может быть достигнуто, если обеспечить подкрепление обезьянам
в виде фруктового сока. Пока обезьяны сообщают о своем восприятии во время
конкуренции, исследователи могут измерить активность отдельных нейронов в
разных частях мозга. Эти исследования обнаружили сильные изменения актив-
ности в вентральной височной коре, соответствовавшие восприятию, о котором
сообщала обезьяна во время конкуренции (Sheinberg, Logothetis, 1997). Фак-
Sheinberg, Logothetis, 1997). Фак-
, Logothetis, 1997). Фак-
Logothetis, 1997). Фак-
, 1997). Фак-
тически около 85% нейронов, записываемых в височной доле, показали такие
Рис. 6 .25. Бинокулярная конкуренция. (а) Если у вас есть очки с красным и зеленым
фильтрами, этот рисунок должен создать бинокулярную конкуренцию. Ваше сознание
будет колебаться туда и сюда между образами, имеющими вертикальные и горизонталь-
ные полосы. (б) Если у вас нет очков с красным и зеленым фильтрами, попытайтесь
объединить эти два образца. Чтобы их объединить, вам нужно расположить свои глаза
таким образом, чтобы два образа выстроились так, чтобы они казались раположенными
друг над другом. Окружающие их квадраты могут помочь вам сделать это. Построй-
те контуры квадратов и точку с концентрической окружностью в середине. (в) Схема
того, как Тонг с сотрудниками (Tong et al., 1998) использовал очки с красным и зеленым
фильтрами для того, чтобы достигнуть бинокулярной конкуренции в фМРТ, а также
области ЗРЛ и ПОП, где была найдена активность, коррелирующая с осознанным вос-
приятием. (Источник: Frank Tong, с разрешения.)
Зрение
287
Рис. 6 .26 . Стимулы и данные из публикации Тонга (Tong et al., 1998). (а) Верхний
блок показывает состояние бинокулярной конкуренции. Человек вначале воспринима-
ет лицо, затем дом, затем лицо и т. д. Нижний блок показывает контрольное состоя-
ние без бинокулярной конкуренции: изображения появлялись или исчезали на экране.
(б) Верхний блок показывает колебания активности как ЗРЛ (синий график), так и ПОП
(красный график) во время бинокулярной конкуренции. Во время того, как каждое изо-
бражение становится доминирующим, активность в соответствующей зоне возрастает.
На нижнем блоке показан нейронный ответ в тех же зонах при контрольном состоянии
без бинокулярной конкуренции. Колебания активности в обоих случаях были прибли-
зительно одинакового размера. (Источник: Tong et al., 1998.)
288
Глава 6
модуляции. Хотя похожие работы показали более слабые эффекты конкуренции
в первичных зрительных полях обезьян (Leopold, Logothetis, 1996), исследова-
Leopold, Logothetis, 1996), исследова-
, Logothetis, 1996), исследова-
Logothetis, 1996), исследова-
, 1996), исследова-
ния, описанные выше, доказали, что действительно возможно изучать нейро-
биологические соответствия сознательному состоянию как у обезьян, так и у
людей.
5.3 . Зрительное обнаружение: видели ли вы это?
Другим путем для разделения физической стимуляции и осознания восприятия
является задача на зрительное обнаружение. В данном случае человек должен
обнаружить и сообщить, когда он или она видит определенный образ. Иссле-
дователь делает образец все более трудным и трудным для рассмотрения, по-
этому в разных случаях образец может быть легко обнаружен, в то время как
в другой раз это почти невозможно, а иногда его там вообще не будет. Так как
эта задача становится трудной, люди ее иногда выполняют неверно. Имеются
случаи, когда кто-то сообщает о том, что видит образец, когда он даже не при-
сутствует, а в другом случае испытуемые не замечают образец и говорят, что
ничего не видят. Если такой тип экспериментов проводить с использованием
фМРТ, мы сможем увидеть шаблон, по которому активность головного мозга
соответствует зрительному образу. Точно так же, если не предъявлен ни один
образец и испытуемые сообщают, что они не видят изображение (истинный
отрицательный ответ), мы не увидим тип активности мозга, соответствую-
щий рисунку. Однако, как вы думаете, что происходит, когда кто-то ошибется?
Это тот случай, когда никакого образца в действительности не предъявлено, но
человек думает, что он что-то видит и сообщает: «Да, образец был там», (л ож-
ный положительный ответ). Интересно, что активность в зонах V1, V2 и V3
непосредственно соответствует тому, что, как мы думаем, мы видели. Другими
Рис. 6 .27. Иллюстрация того,
как обезьяна выполняет задание
на бинокулярную конкуренцию
в восприятии, в то время как ис-
следователь делает запись от
нейронов в мозге обезьяны, как
в работе Шейнберга и Логотети-
са (Sheinberg, Logothetis, 1997).
(Источник: Blake, Logothetis,
2002.)
Зрение
289
словами, в тех испытаниях, где представлялся слабый стимул, но испытуемый
не мог его обнаружить, активность в этих зонах была намного слабее (Ress,
Heeger, 2003). Однако если не было представлено стимула, но испытуемый
ошибочно думал, что слабый стимул присутствовал, оказалось, что активность
V1, V2 и V3 в этих опытах была сильнее.
Как объяснить такие колебания в уровне активности от испытания к испы-
танию? Это объяснялось непостоянством нервной активности от опыта к опыту
или нервному шуму в системе. Ваша память об образце может «сформировать»
произвольную нервную активность, которая существует в сенсорных областях,
что оказывается решающим фактором и заставляет вас думать о том, что вы ви-
дели образец. Это другой пример того, как активность головного мозга может
тесно отображать необыкновенное ощущение того, что вы что-то видели, в то
время когда ничего даже не было фактически представлено. Это интересно по-
тому, что демонстрирует, что не имеет значения то, какой физический образец
был предъявлен человеку; единственное, что имеет значение, — то, что проис-
ходит в мозге!
5.4 . Созидательное восприятие:
видеть больше, чем доступно глазу...
Если вы водите машину, то, вероятно, знаете, что такое слепое пятно: для во-
дителя это область позади машины, которую невозможно увидеть ни в боко-
вых, ни в заднем зеркалах. Наши глаза тоже имеют слепое пятно сетчатки, где
аксоны ганглиозных клеток сетчатки собираются и образуют зрительный нерв,
выходящий из глазного яблока (см. рис. 6 .3 (б)). В слепом пятне нет фоторецеп-
торов, и поэтому в этой области мы не видим.
Показать слепое пятно легко. Посмотрите на схему на рис. 6 .28 (а). Закройте
левый глаз, посмотрите прямо на крест правым глазом и поднесите книгу близ-
ко к своему носу. Затем медленно отодвигайте ее обратно от своего лица, про-
должая фиксировать взгляд на кресте. На определенном расстоянии, которое
должно быть около 30 см от страницы, вы должны заметить, как точка исчезает.
В то время как изображение этого пятна на сетчатке передвинулось к местора-
сположению слепого пятна, оно исчезает!
Будем надеяться, эта демонстрация оказала на вас влияние, и, возможно, вы
теперь думаете: «...минуточку, если все время в моем поле зрения имеется сле-
Рис. 6.28. Демонстрация слепого пятна. (а) Закройте левый глаз, посмотрите пря-
мо на крест правым глазом и поднесите книгу близко к своему носу. Затем медленно
отодвигайте ее обратно от своего лица, продолжая фиксировать взгляд на кресте. На
определенном расстоянии, которое должно быть около 30 см от страницы, вы долж-
ны заметить, как красная точка исчезает. В то время как изображение пятна на вашей
сетчатке передвигается к месту слепого пятна, в котором нет фоторецепторов, оно ис-
чезает! (б) Подобным же образом, заметьте, черные полоски теперь заполняют место
красного пятна: они соединяются, а красное пятно исчезает. (Источник: Frank Tong,
с разрешения.)
б
а
290
Глава 6
пое пятно, то почему я не вижу дыру, когда я оглядываю все вокруг с одним
закрытым глазом?» В действительности мозг делает нечто замечательное: он
фактически заполняет восприятие в слепом пятне. Мозг использует зритель-
ную информацию из окружения слепого пятна, чтобы сделать заключение о
том, что должно быть в слепом пятне, и создает наше знание из того, что он
«считает» там должно быть. Заполнение в месте слепого пятна является при-
мером созидательного восприятия или заполнения восприятия.
Заполнение восприятия имеется не только в слепом пятне, но также проис-
ходит в других частях зрительного поля. Заметьте, как область между цветными
линиями на рис. 6.29 (а) тоже каким-то образом становится цветной. Однако
тщательное исследование показывает, что эта область фона в действительно-
сти только белая — никакого цвета. Эта иллюзия называется неоновым распро-
странением цвета, и впечатление окрашенности здесь, кажется, происходит
из-за созидательных механизмов заполнения при работе зрительной коры. По-
следние исследования по сканированию мозга обнаружили, что активность в
V1 больше в соответствующих областях, в которых испытуемые воспринимали
распространение неонового цвета (Sasaki, Watanabe, 2004). Это предполагает,
что уже нейроны в V1 могут быть важны для заполнения восприятия и нашего
впечатления о цвете, даже иллюзорного.
Во время другого типа заполнения, известного как зрительные фантомы
(рис. 6 .29 (б)), наша зрительная система может также заполнять пробелы между
двумя структурами. На рис. 6 .29 (б) у вас может создаться ощущение темных
полос, продолжающихся вдоль пустого пространства, даже если там вообще
не представлено никаких стимулов. При использовании движущихся стимулов
можно в дальнейшем усилить иллюзию зрительного фантома. Когда у нас на-
блюдается такой тип заполнения, корковые области V1 и V2 отвечают таким об-
V1 и V2 отвечают таким об-
1 и V2 отвечают таким об-
V2 отвечают таким об-
2 отвечают таким об-
Рис. 6 .29 . Демонстрация за-
полнения восприятия. (а) В этих
трех примерах фон белый – он не
имеет цвета. Однако вы можете
заметить, как красный и голубой
стремятся заполнить его, прида-
вая фону белый цвет. (б) Светлые
и темные пятна сверху и снизу
создают впечатление светлых и
темных участков вдоль централь-
ной полоски, несмотря на то что
центральная полоска является
однородно серой. Эта иллюзия
намного сильнее проявляется
при движении. (Источник: Frank
Tong, с разрешения.)
Зрение
291
разом, как будто в пространстве была представлена действительная структура
(Meng et al., 2005).
Мозг может дополнять не только цвета или шаблоны, но также движение.
Если вы создадите пятно света в одном месте, а затем за ним последует похожая
вспышка в другом месте через определенное время, у наблюдателей сложится
впечатление об иллюзии движения — кажущееся движение. Подобный фокус
все время используется на высоко висящих вывесках магазинов, а также лежит
в основе того, почему кинофильмы кажутся такими плавными и реальными.
Если вы посмотрите фильм, показанный с помощью проектора, вы обнаружи-
те, что имеется серия кадров, которые высвечиваются на экран один за другим:
впечатление плавного движения — просто иллюзия. Недавние исследования
обнаружили, что воспринимаемый путь кажущегося движения отображается
нейронной активностью V1 (Jancke et al., 2004; Muckli et al., 2005). Как и в
случае предыдущих примеров с цветом и шаблоном, представляется, что мозг
дополняет информацию о кажущемся движении как с точки зрения восприятия,
так и с точки зрения нервного пути, даже если не происходит никакого физиче-
ского движения.
Из приведенных выше примеров должно быть ясно, что мозг может активно
составлять воспринимаемые образы, даже если какой-то физический стимул
в определенной части зрительного поля не представлен. Создается впечатле-
ние, что заполнение происходит в первичных зрительных зонах на такой ран-
ней стадии, как V1. Это указывает на то, что первичные зрительные зоны могут
обеспечивать основу для созидательных зрительных впечатлений.
5.5 . Нервные корреляции с распознаванием объектов
Какие области мозга важны для распознавания объектов? Как мы видели при
изучении бинокулярной конкуренции, активность зоны распознавания лиц и
парагиппокампальной области восприятия пространства тесно связана с осоз-
нанием испытуемым лиц и домов. Подобным образом, если человеку пока-
зать двусмысленную фигуру, такую, какая показана на рис. 6 .24 (а), области
вентральной височной коры демонстрируют более сильную активность, когда
узнавание переключается на другую интерпретацию. Например, при рассма-
тривании изображения Рубина «лицо–ваза» переключение на восприятие лица
приводит к увеличению активности в зоне распознавания лиц.
Другие исследования изучали осознание визуально скрытых объектов, кото-
рые предъявлялись на такое короткое время, что только-только могли быть рас-
познанными. Если наблюдатели могут успешно распознать объект, высвечен-
ный короткое время, то активность во многих областях вентральной височной
коры больше, чем тогда, когда распознавание не было успешным (�rill-Spector
et al., 2000; Bar et al., 2001). То же происходит, если наблюдателям показывать
двусмысленное черно-белое изображение, такое как лунное лицо, показанное
на рис. 6 .30 справа, которое трудно распознать; если его показать в другой ори-
ентации, активность в вентральных височных областях намного сильнее, когда
человек осознает скрытый объект (Dolan et al., 1997; McKeef, Tong, 2006). Ин-
McKeef, Tong, 2006). Ин-
, Tong, 2006). Ин-
Tong, 2006). Ин-
, 2006). Ин-
тересно, что в большинстве исследований первичные зрительные области не
показывают разницы между условиями, когда наблюдатель воспринимает объ-
ект или нет. Еще более удивительно, что когда человека просто просят предста-
292
Глава 6
вить лицо, это может привести к активации зоны распознавания лиц (O’Craven,
Kanwisher, 2000). Подобно этому, воображение места или объекта местности
может активировать парагиппокампальную область восприятия пространства.
Эти исследования предполагают, что простое извлечение зрительной памяти об
определенном объекте может активировать области восприятия объектов.
6.0. Управление зрительным осознанием
В предыдущем разделе мы обсуждали корреляционные методы изучения осоз-
нанного восприятия, которые опираются на обнаружение нейробиологических
соответствий сознательному состоянию. Знание, что активность в области А
связана с восприятием Б (например, наблюдение движения влево), предостав-
ляет данные о корреляции: область А вовлечена в отображающее его восприя-
тие Б. Однако что-то может быть связано с определенной деятельностью, но не
выражать связь причины и следствия. Например, мотор машины ревет всякий
раз, когда она напряженно работает, но звук ревущего двигателя — это не то,
что заставляет колеса вращаться. Глушение звука машины не остановит дви-
жение машины, а повреждение поршней — да. Чтобы предоставить дальней-
шие данные о роли области мозга в сознании, мы должны обеспечить данные о
причинно-следственных связях, показывающие, что воздействие на активность
в области А может создать или нарушить восприятие Б. В нейробиологии
имеется много методов обнаружения причинно-следственных связей, которые
включают возбуждающую и тормозящую активность в определенных областях
мозга, как на временной, так и постоянной основе (например, исследования при
повреждениях). Следующий параграф осветит некоторые открытия, получен-
ные в результате манипуляций нервной активностью и зрительным осознанием.
Электрическая стимуляция затылочной доли может вызвать субъективное
зрительное восприятие или то, что называется фосфеном. Например, слепые
пациенты, у которых имеются вживленные электроды в первичной зрительной
коре, сообщали о том, что когда эта область стимулировалась, у них впервые за
долгие годы возникло зрительное впечатление. Когда стимулировались неболь-
шие участки, испытуемые сообщали о том, что видят фосфены, которые вы-
глядели как «маленькие зернышки риса, которые удерживались на расстоянии
длины руки».
Рис. 6 .30 . Эти изображения на-
зываются лунными лицами. Они
вначале были сделаны из фото-
графий реальных лиц. Так как эти
изображения лиц можно с трудом
распознать, они полезны при из-
учении распознавания объектов.
Когда, в конце концов, люди рас-
познают, что это лицо, вы увидите
увеличение нервной активности в
зоне распознавания лиц. (Источ-
ник: Frank Tong, с разрешения.)
Зрение
293
Это кажется само собой разумеющимся, если мы вспомним, что язык мозга
в действительности является электрическим. Нейроны сообщаются друг с дру-
гом через слабые электрические токи, поэтому, когда ток подводится к группе
нейронов, неудивительно, что человек испытывает некоторое чувство. Имеет-
ся много путей сделать так: нейроны могут электрически стимулироваться во
время хирургической операции, или в большем масштабе электрический ток
может проходить сквозь черепную коробку. Однако более современный и менее
инвазивный метод — это транскраниальная магнитная стимуляция (ТМС).
6.1. Транскраниальная магнитная стимуляция
Транскраниальная магнитная стимуляция, или ТМС, включает быстро гене-
рируемое магнитное поле, внешнее по отношению к голове, которое индуци-
рует электрическую активность на поверхности коры головного мозга (Walsh,
Cowey, 2000). Обычно это делается с использованием портативной ТМС катуш-
ки, которая может быть помещена на поверхности кожи головы. Когда вклю-
чается электрический ток, возбуждается магнитное поле, которое инициирует
нервную активность на поверхности коры головного мозга (рис. 6 .31 (а)). Такая
индуцированная активность может либо действовать как разновидность «зри-
тельного стимула», либо нарушать происходящую в настоящее время зритель-
ную активность.
6.1.1. Фосфены и стимуляция V1
Если ТМС применяют к первичным зрительным областям, получают два основ-
ных результата в восприятии. В одном случае, когда у людей закрытые глаза,
они часто чувствуют слабую вспышку света, фосфен. Как упоминалось выше,
это относится к активации зрительных нейронов. Во втором случае вы можете
воспринимать визуальную дыру или кратковременную слепую точку — вре-
менную скотому (временная область, в которой зрение понижено или утеряно)
при зрительной стимуляции как раз после ТМС (Kamitani, Shimo�o, 1999).
Интересно то, что тип фосфенов испытуемых соответствует области тка-
ни коры головного мозга, стимулируемой ТМС. Например, когда стимулируют
V1, люди сообщают о маленьких неподвижных фосфенах. Когда стимулиру-
ют область MT (область движений), люди сообщают о движущихся фосфенах
(Pascual-Leone, Walsh, 2001)!
Такие открытия, взятые отдельно, могут рассматриваться как данные о том,
что эти области ответственны за обработку соответствующих зрительных при-
знаков (предметы, движение и т. п.) . Однако мы знаем, что большинство зон
мозга связаны с большинством других областей, а нервное возбуждение может
очень быстро перемещаться с одного места на другое. Следовательно, верно то,
что когда вы стимулируете одну область, возбуждение распространяется за ее
пределы на множество других областей. Итак, что если мы используем стиму-
ляцию ТМС во многих расположениях и в разное время? Может ли это дать нам
данные о нервной передаче и обратной связи между нейронами?
6.1.2. ТМС и корковая обратная связь
Недавнее исследование предоставило новые данные в поддержку роли соеди-
нений по принципу обратной связи при зрительном осознании (Pascual-Leone,
294
Глава 6
Walsh, 2001). Двигательные фосфены достоверно вызывались при приложении
ТМС к области MT/V5 (двигательные зоны), при этом второй импульс ТМС
давался либо на V1, либо в область MT через различный временной промежу-
ток перед первым импульсом или после него. Восприятие движущихся фосфе-
нов выборочно уменьшалось, когда стимуляция V1 проводилась через короткое
время после, но не перед стимуляцией зоны MT (позднее на 10–40 мс). Други-
ми словами, исследователи должны были первым дать импульс в область MT,
которая выше в иерархии обработки информации. Затем, позже на заданный
промежуток времени, они должны были давать импульс на зону V1, которая на-
ходится раньше в потоке обработки информации. Если это проводить позднее
этого определенного временного промежутка, испытуемые больше не воспри-
нимали фосфены. Предполагается, что проекции по принципу обратной связи
от MT к V1 могут быть необходимы для сознательного восприятия фосфенов.
Авторы исследования сделали заключение, что возбуждение только области
MT может быть недостаточно для поддержания восприятия движения, и что
возбуждение, идущее по принципу обратной связи к зоне V1, может быть не-
обходимо для зрительного осознания. Эти полученные данные обеспечили под-
держку интерактивной теории зрительного осознания.
Исследование с помощью ТМС слепозрячего пациента ГИ (помните, что па-
циенты со слепозрением обычно имеют большие поврежденные участки V1)
обеспечило некоторые данные в пользу модели обратного возвращения сиг-
налов по принципу обратной связи при зрительном осознании (Cowey, Walsh,
2000). У ГИ стимуляция зоны MT вызывала движущиеся фосфены только при
прикладывании к полушарию, содержащему неповрежденную зону V1, а не к
стороне с поврежденной корой V1. Итак, даже если зона MT была абсолют-
но неповрежденной в коре обоих полушарий, стимуляция MT с той же сто-
роны головы, что и поврежденная V1, не приводила к восприятию фосфенов.
Наоборот, движущиеся фосфены успешно вызывались у всех исследованных
здоровых испытуемых так же, как и у слепых из-за нарушений сетчатки паци-
ентов. Дальнейшие тесты интерактивной модели должны рассмотреть, может
ли прямая стимуляция коры в экстрастриарных областях вызывать фосфены у
пациентов с повреждением V1.
Другой замечательный пример модуляции зрительного осознания пришел
из исследования, в котором ученые стимулировали непосредственно нейроны в
мозге обезьян (Salzman et al., 1990). Как мы упоминали выше, нейроны в обла-
сти коры MT избирательно отвечают на определенные направления движения,
а нейроны, которые предпочитают сходные направления, имеют тенденцию
собираться в кортикальные колонки. Другими словами, только определенная
группа нейронов в MT может быть высоко активна при предъявлении обезьяне
объекта, движущегося направо. В этом эксперименте исследователи непосред-
ственно стимулировали некоторые из избирательных к направлению нейронов
в зоне MT обезьян. Они делали это в то время, когда животное рассматривало
неопределенные (имеется в виду по направлению движения) точки, двигающи-
еся по многим возможным направлениям при проведении задачи на распозна-
вание движения. Эта микростимуляция влияла на решение животного так, что
оно выбирало то направление движения, за которое типично отвечали стимули-
руемые нейроны. Другими словами, все происходило так, как будто стимулиро-
Зрение
295
вание таких нейронов приводило к более сильному восприятию этого опреде-
ленного направления движения, достаточному для того, чтобы четко изменить
впечатление обезьяны о движении, если движение в стимуле было довольно
неопределенным.
Намного более сложные воспринимаемые впечатления могут быть вызваны
нервной стимуляцией, например те, которые можно назвать впечатлениями на-
хождения за пределами тела. Пациентка, больная эпилепсией (из-за серьезно-
сти ее состояния с электродами, временно имплантированными в ее мозг), со-
общала о таких впечатлениях. Когда стимулировали два конкретных электрода
над соединением височной и теменной долей (см. рис. 6.31 (б)), она говорила о
новом чувстве падения или полета (Blanke et al., 2002), описывая чувство как
«падение с высоты». Более сильная стимуляция приводила к тому, что пациент-
ка сообщала о явном впечатлении нахождения вне тела. Более правильно, что
она заявляла на самом деле о впечатлении того, что она видит свое тело сверху,
при этом лежа на постели. Однако она не видела своего лица, только туловище
внизу. Это открытие важно, потому что угол, с которого по ее сообщениям она
видела свое тело, отличался от того, с которого она обычно его видела. Предпо-
лагается, что обычные ощущения (ее нормальный взгляд на тело) могут быть
объединены, чтобы образовать неизвестные ощущения видения собственного
тела с разных углов. Эта идея имеет захватывающие последствия для сенсорных
впечатлений, так как она предполагает, что опыт внутреннего происхождения не
просто должен быть повторением предыдущих воспринимаемых событий.
Рис. 6 .31 . Стимуляция мозга.
(а) Катушка транскраниальной
магнитной стимуляции (ТМС)
над мозгом. Пунктирные линии
показывают магнитное поле
и область ткани, на которую в
основном оно действует. ТМС
является ценным инструмен-
том изучения для исследования
причинно-следственной связи
между активностью мозга и
зрительным осознанием. (б) Во
время стимуляции этой группы
электродов пациентка сообщи-
ла о том, что у нее создалось
впечатление того, что она на-
ходится выше собственного
тела — «впечатление нахожде-
ния вне своего тела». (Источ-
ник: Blake, Logothetis, 2002.)
296
Глава 6
Исследования с помощью стимуляции позволяют ученым точно определить
причинно-следственные взаимосвязи и, таким образом, обнаружить, какие об-
ласти мозга ответственны за то, что мы видим. По существу, это похоже на
установление контроля над действиями мозга. Прямой стимуляцией коры в
обход чувствующих органов, таких как сетчатка, возможно будет определить,
какие участки мозга непосредственно связаны с воспринимаемым зрительным
опытом.
6.2 . Бессознательное восприятие
Как что-то может быть одновременно бессознательным и все еще называться
восприятием? Не предполагает ли факт, что вы не знаете чего-то, то, что ваш
мозг не смог обработать или идентифицировать этот стимул? В настоящем кон-
тексте мы используем термин бессознательное восприятие по отношению к
ситуациям, когда люди сообщают о том, что они не видят подаваемый стимул,
но их поведение или активность мозга дают основание предполагать, что опре-
деленная информация о невоспринимаемом стимуле была в действительно-
сти обработана мозгом. Нервная активность не всегда приводит к осознанию.
Нейроны могут возбуждаться по пути специфическому для данного стимула на
многих уровнях мозга, но эта активность бывает недостаточно сильной, длит-
ся недостаточно долго или не включает достаточное количество нейронов или
участков мозга, чтобы это привело к осознанию. Один из лучших примеров
имеется при записи нейронной активности животных под наркозом; зритель-
ные нейроны во многих областях мозга могут все еще показывать высокие из-
бирательные ответы на стимул. В этом разделе мы узнаем, что даже когда чело-
век полностью пробужден и находится в сознании, бессознательная обработка
зрительных сигналов также может происходить во многих случаях.
В предыдущем разделе вы узнали о том, как действует принцип «то, что ты
видишь, — это то, что стимулирует тебя» во время бинокулярной конкуренции.
Другими словами, если вы осознаете определенный стимул, то нейроны ваше-
го мозга, которые представляют этот шаблон, становятся высоко активными.
Однако здесь мы узнаем, что противоположное утверждение не всегда является
правдой. Если у вас есть некоторая активность в данной области мозга, это не
означает, что вы обязательно воспринимаете этот стимул. Без достаточной ак-
тивности в определенной области мозга осознание может быть недостаточным,
и в результате произойдет бессознательное восприятие.
Когда два разных стимула мгновенно появляются на достаточно короткое
время в быстрой последовательности, зрительная система не сможет больше
разделить эти два стимула. Вместо этого то, что воспринимают люди, явля-
ется смешением или слиянием двух изображений. Например, если мы долж-
ны предъявить вам красный квадрат, а затем за ним быстро следует зеленый
квадрат, то, что вы можете воспринимать, является комбинацией этих двух:
желтый квадрат. Этот метод может быть использован, чтобы представить не-
видимые образы, такие как лицо или дом, путем вспышек красных контуров
на зеленом фоне для одного глаза и противоположных цветов для другого глаза
(рис. 6 .32). При предъявлении испытуемым таких изображений и при помо-
щи магнитно-резонансного сканера область, ответственная за узнавание лиц,
наиболее сильно отвечала на лица, а парагиппокампальная область восприятия
пространства больше отвечала на дома, даже если испытуемые не осознавали
Зрение
297
эти изображения (Moutoussis, �eki, 2002). Это пример бессознательного вос-
Moutoussis, �eki, 2002). Это пример бессознательного вос-
, �eki, 2002). Это пример бессознательного вос-
�eki, 2002). Это пример бессознательного вос-
, 2002). Это пример бессознательного вос-
приятия. Активность в вентральных височных частях мозга могла быть разной
до определенной степени в зависимости от того, предъявлялось лицо или дом,
даже хотя испытуемый не мог на словах определить разницу. Поэтому из-за
того, что область имеет некоторую активность, нельзя делать вывод, что мы
осознаем то, что отображают области мозга. Возможно, этот уровень актив-
ности еще недостаточен. Более сильный ответ был обнаружен в этих областях
мозга, когда единственное красно-зеленое изображение показывалось так, что
было ясно видно.
Зона MT, область мозга, специализирующаяся на обработке движения, так-
MT, область мозга, специализирующаяся на обработке движения, так-
, область мозга, специализирующаяся на обработке движения, так-
же отвечает на невоспринимаемое движение. Когда движущийся стимул предъ-
является далеко на периферии зрительного поля и поле наполнено другими
стимулами, область MT все еще отвечает на движение, даже если испытуемые
не осознают это движение (Moutoussis, �eki, 2006). В этом эксперименте дви-
Moutoussis, �eki, 2006). В этом эксперименте дви-
, �eki, 2006). В этом эксперименте дви-
�eki, 2006). В этом эксперименте дви-
, 2006). В этом эксперименте дви-
жущийся стимул был окружен с двух сторон мерцающими стимулами, что де-
лало зрительное пространство таким заполненным и загроможденным, что ис-
пытуемые не могли видеть движение на экране. Хотя MT отвечала в некоторой
степени на воспринимаемый движущийся стимул, она возбуждалась намного
сильнее, когда находящиеся вокруг мерцающие сигналы были удалены, а дви-
жущийся стимул ясно воспринимался.
В других исследованиях ученые использовали бинокулярную конкуренцию
(которую мы обсуждали в предыдущем разделе), чтобы делать невидимыми
изображения испуганного лица (рис. 6 .33 (а)). Выражения лиц содержали эмо-
циональную составляющую, хотя испытуемые никогда не осознавали то, что
они видят лица. Миндалина, область мозга в медиальной височной доле, кото-
рая в норме отвечает на эмоциональные стимулы, также отвечала на невиди-
мые испуганные лица (Pasley et al., 2004). На рис. 6 .33 (б) изображен график
активности в миндалине: красная линия представляет собой ответ миндалины
на изображения эмоционального лица.
Ясно, что многие области мозга могут продолжать демонстрировать актив-
ность на специфические стимулы, несмотря на тот факт, что мы не осознаем их.
Это подтверждает гипотезу, что даже если область мозга обрабатывает стимул,
это не означает, что мы будем воспринимать этот стимул. Если нервная актив-
ность в данной области недостаточна, чтобы в результате появилось осознание,
то тогда что это? В большинстве таких исследований большая активность на-
блюдалась, когда испытуемые осознавали стимул, чем когда нет. Следователь-
но, минимальный уровень активности может быть необходим, чтобы различать
Рис. 6 .32 . Красный + зеленый
= желтый. Красный и зеленый
цвета соединяются и образуют
квадрат желтого цвета. Таким
образом, изображение дома или
лица становится невидимым
при его коротком показе по
одному каждому глазу. Однако
мозг все еще отвечает на эти
невидимые образы. (Источник:
Moutoussis, �eki, 2002).
298
Глава 6
Рис. 6 .33 . Эмоция все еще воспринимается, но не изображение лица. (а) Схематическое
изображение стимула, используемого в исследовании Пэсли (Pasley et al., 2004). Крас-
ное здание, которое предъявлялось левому глазу, скрывало от осознания лицо, предъ-
являемое правому глазу, как в случае бинокулярной конкуренции. Лица имели эмо-
циональные выражения. (б) Этот график показывает активность миндалины (область
эмоционального ответа мозга). Красный график показывает, что активность миндалины
увеличивается, когда предъявляются эмоциональные лица, даже если они находятся за
пределами восприятия. Мозг не может видеть лицо, но он может определять эмоцию.
(Источник: Pasley et al., 2004.)
Зрение
299
осознание и его отсутствие. Низкий уровень нервной активности, ниже опре-
деленной пороговой величины, может быть недостаточным, чтобы в результате
произошло осознание стимула.
Эта идея о нервном пороге для зрительного осознания прекрасно подходит
как для иерархической, так и для интерактивной моделей зрительного осоз-
нания, которые мы обсуждали ранее. Для иерархической модели подходящий
уровень нервной активности должен поддерживаться в зрительных областях
высшего порядка для осуществления процесса осознания. В интерактивной
модели должны осуществляться сигналы обратной связи, чтобы поддерживать
нервную активность на каждом уровне обработки, что приводит к активности,
которая превышает пороговый уровень для осознания зрительных образов в
многочисленных областях мозга. Подводя итог, мы можем заключить, что, по
крайней мере, две вещи необходимы для зрительного осознания: 1) активность
в надлежащих нейронах и областях мозга и 2) активность, которая превышает
критический порог. Дальнейшие исследования смогут показать, какая из этих
моделей лучше описывает основы зрительного осознания в головном мозге.
7.0 . Заключение
Зрение, возможно, один из самых важных типов ощущений. Конечно, именно
ему была посвящена большая часть исследований. За последнее десятилетие
или около того, ученые изучили огромное количество информации о нервных
корреляциях сознательного и бессознательного восприятия, и каким образом
повреждения различных областей головного мозга могут нарушить специфи-
ческие стороны зрительного сознания. Согласующимся с этим открытием яв-
ляется то, что первичная зрительная кора, кажется, является важной для спо-
собности воспринимать любую зрительную информацию вообще, в то время
как высшие области мозга могут быть важными для восприятия определенных
зрительных признаков или предметов. Современные данные поддерживают как
иерархическую, так и интерактивную теории зрительного осознания. Будущие
исследования улучшат наше понимание того, каким образом головной мозг об-
условливает наши субъективные зрительные ощущения.
В этой главе мы проследили функциональные свойства нейронов, то, как
зрительные сигналы распространяются от сетчатки к первичной зрительной
коре и далее к высшим областям по дорсальному и вентральному зрительным
путям. При продвижении вверх по зрительному пути рецептивные поля посте-
пенно становятся больше и отвечают на более сложные стимулы, следуя иерар-
хической организации зрительной системы.
V1 выборочно реагирует на многие зрительные особенности, включая ори-
1 выборочно реагирует на многие зрительные особенности, включая ори-
ентацию, движение и бинокулярное различие. Повреждение V1 может серьезно
снизить или уничтожить осознанное зрение (осознанные зрительные стимулы),
хотя оставшаяся активность в экстрастриарных областях может поддерживать
способность определять зрительные события даже без того, что они осознают-
ся: состояние, называемое слепозрение. Экстрастриарные зрительные области
(находящиеся вне V1) кажутся важными для восприятия специфических зри-
V1) кажутся важными для восприятия специфических зри-
1) кажутся важными для восприятия специфических зри-
тельных особенностей: зона V4 важна для восприятия цвета, а зона MT для вос-
V4 важна для восприятия цвета, а зона MT для вос-
4 важна для восприятия цвета, а зона MT для вос-
MT для вос-
для вос-
приятия движения. Повреждение данных зон может привести к выборочному
ухудшению в восприятии этих особенностей более высокого уровня в визуаль-
ном мире.
300
Глава 6
Согласно иерархической теории, экстрастриарные области более высокого
порядка тесно связаны с зрительным осознанием, в то время как V1 — нет.
В противоположность тому, интерактивная теория подчеркивает, что сигналы
обратной связи к V1 могут быть важны для осознания. Современные данные
обеспечивают поддержку обеим теориям.
Повреждение дорсального пути может привести к зрительной атаксии
(ухудшению зрительного сопровождения действий) или зрительному игнори-
рованию. Повреждение вентральной височной коры может привести к ухудше-
нию зрительного восприятия, узнаванию предметов или лиц. Пациенты с по-
вреждениями мозга в областях вентрального и дорсального путей, показывают
разобщение между сознательным восприятием основных форм и ориентацией
и способностью выполнять зрительно направляемые действия.
В вентральной височной коре некоторые области мозга, такие как зона ЛЗК
(латерального затылочного комплекса, кажется, играют основную роль в узна-
вании предметов, в то время как другие зоны, такие как зона распознавания лиц
(ЗРЛ) и парагиппокампальная область восприятия пространства (ПОП), веро-
ятно, имеют более специализированные роли. Многие исследования показыва-
ют, что активность в этих областях тесно связана с сознательным восприятием
объектов. Тем не менее данные по бессознательной обработке сигналов могут
быть найдены по многим областям мозга, включая предметные области высо-
кого уровня.
8.0 . Задания и упражнения к главе 6
1. Для схематического изображения мозга на рис. 6.34:
а) Скопируйте рисунок мозга. В какую сторону он направлен?
б) Определите зрительные области (отмечены) и некоторые из выполняемых
ими функций.
в) Можете ли вы обозначить цветом дорсальный и вентральный пути? Како-
ва разница между их функциями?
Рис. 6.34 . Зрительные области коры головного мозга человека. (Источник: Rosa, 2002.)
Зрение
301
2. Можете ли вы описать своими словами, что происходит на рис. 6 .35? Что ви-
дит в данное время кот? Какие зрительные области, по всей вероятности, во-
влечены в этот процесс? Какие нервные механизмы улучшают способность
кота воспринимать контраст и границы?
3. Для рис. 6 .36:
а) Нарисуйте каждый рисунок в цвете.
б) Что мы можем узнать из (а)? Какие части зрительной коры, по всей веро-
ятности, вовлечены в процесс?
в) Как насчет (б)? Вероятно ли, что вовлечены другие зоны зрительной
коры, чем в (а)?
г) Для третьего изображения на рисунке, что воспринимает испытуемый в
опыте? Почему цвета выглядят так, как будто их смешали? Какова цель
эксперимента и каковы результаты?
4. Вопрос для обдумывания: как мы можем сравнить осознаваемый и бессоз-
нательный зрительные стимулы?
Рис. 6 .36. Зрительная неопределенность. (Источник: Kim, Blake, 2005.)
Рис. 6 .35. Запись с мозга кота. (Источник: Rosa, 2002.)
Счастливым случаем был тот факт, что уже во времена Гельмгольца вели-
кие анатомические открытия, сделанные Корти и другими, дали ясно понять,
что наиболее важной вибрирующей тканью для слуха является базилярная
мембрана внутреннего уха, так как клетки, на которых оканчиваются нерв-
ные волокна, расположены на этой мембране... вопрос о том, как мы слышим,
был в значительной степени сведен к механической проблеме: каким образом
вибрирует базилярная мембрана, когда барабанная перепонка подвергается
воздействию синусоидальным тоном?
Бекеши, Лекция на вручении Нобелевской премии
Слуховая система на самом деле начинается глубоко внутри слухового канала на бара-
банной перепонке. Колебания воздуха, являющиеся быстрыми сжатиями и расширени-
ями воздуха, заставляют вибрировать барабанную перепонку, которая передает меха-
нические колебания дальше, что заканчивается возбуждением рецепторов (волосковых
клеток), расположенных на базилярной мембране. Нервные окончания, иннервирую-
щие волосковые клетки, собираются вместе, образуя слуховой нерв, который проходит
через несколько структур, прежде чем достичь коры. Слуховая система чрезвычайно
чувствительна, способна зафиксировать даже колебание молекул воздуха в очень тихой
обстановке. (Источники: (вверху) Standring, 2005; (внизу) Ramachandran, 2002.)
Содержание
1.0.Введение.......................................................303
1.1.Модельобработкиинформации.................................304
1.2.Основызвукаислуховоговосприятия............................307
2.0.Центральнаяслуховаясистема.....................................312
2.1.Путипередачислуховойинформации............................312
2.2.Слуховаякора................................................315
3.0 . Функциональное картирование обработки слуховой информации . . . . . . . . . 323
3.1.Первичнаяслуховаякора.......................................323
3.2 . Роль височной области в декодировании слухового сигнала. . . . . . . . . . 324
3.3.Корковыесистемы«что»и«где»................................325
4.0.Восприятиеречи.................................................336
4.1.Историявопроса..............................................337
4.2.Ранниетеориивосприятияречи.................................340
4.3 . Функциональное картирование специфических речевых процессов . . . 342
4.4.Связьвосприятияиформированияречи..........................343
4.5.Повреждениясистемвосприятияречи...........................344
4.6. Рабочая модель мозговых механизмов восприятия речи . . . . . . . . . . . . . 349
5.0.Восприятиемузыки...............................................352
5.1.Этапыслуховойобработкимузыки..............................352
5.2.Отдельнаясистемадлямузыкальноговосприятия.................. 353
6.0.Обучениеипластичность..........................................355
6.1.Пластичность,связаннаясдепривацией..........................356
6.2.Пластичность,связаннаясобучением............................356
6.3 . Пластичность, связанная с профессиональными навыками . . . . . . . . . . 358
7.0.Сознаниеислуховыеобразы.......................................358
7.1. Слуховое сознание во время сна и седативного состояния . . . . . . . . . . . 359
7.2.Слуховоевоображение........................................360
8.0.Заключение.....................................................361
9.0.Заданияиупражнениякглаве7....................................362
9.1.Контрольныезадания..........................................362
9.2.Графическиеупражнения......................................362
9.3.Литература..................................................363
1.0 . Введение
В этой главе речь пойдет о том, как мы слышим — от самых простых звуков
и до сложноорганизованной речи и музыкальных симфоний. Мы начнем с ос-
новной информации о том, как происходит обработка звукового сигнала: от уха
Глава 7
Слух и речь
304
Глава 7
через восходящие слуховые пути к слуховой коре головного мозга. Затем мы
отдельно обсудим обработку таких особых типов акустических сигналов, как
речь и музыка. Как вы можете представить, особенности восприятия звуков
изменяются у человека по ходу жизни. Это может быть заметно на примере
звуков речи, восприятие которых преобразуется по мере того, как в детстве мы
учимся говорить. Вы также можете предположить — и будете правы! — что
нейрональные системы, которые лежат в основе обработки звуковой инфор-
мации у профессиональных музыкантов или лингвистов, знающих несколь-
ко языков, могут отличаться от тех, которые мы находим у обычных людей. В
связи с этим мы обсудим, как на мозговую организацию слухового восприятия
влияет обучение, как она изменяется с течением жизни и какими могут быть ее
индивидуальные особенности у разных людей. Более того, обработка звуковой
информации не может существовать изолированно от всего остального — то,
что мы слышим, связано с тем, что мы видим и воспринимаем на ощупь, и все
это вместе связано с тем, что хранится у нас в памяти.
1.1 .
Модель обработки информации
Окружающий нас мир полон шума, разные типы звуков достигают наших ушей
в одно и то же время. Представьте себе большую аудиторию перед тем, как нач-
нется лекция: раздаются голоса, скрипят стулья, двери открываются и закрыва-
ются, шумят застежки-молнии от студенческих рюкзаков, учебники с грохотом
опускаются на столы. Все эти звуки попадают к нам в уши одновременно, од-
нако для нас не составляет труда воспринимать их по отдельности. Такое раз-
ложение всей слуховой информации на отдельные акустические «объекты» на-
зывается слуховой анализ сцены (ASA, auditory scene analysis), и именно оно
лежит в основе понимания того, как слуховая система декодирует сложноор-
ганизованное звуковое окружение (Bregman, 1990). В этой главе мы обсудим,
как расшифровывается информация о таком типе звуковой сцены. Мы начнем
с рассмотрения процессов, протекающих внутри слуховой системы, а также их
взаимодействия с другими корковыми и подкорковыми системами.
1.1.1 . Рабочий диапазон воспринимаемых звуков
В гл. 2 была рассмотрена общая модальностная модель работы мозга (modal
model). Для описания принципов работы слуховой системы можно использовать
те же основания: существуют сенсорные (звуковые) стимулы, которые поступа-
ют в систему и помещаются в специальное хранилище (эхоическая память), где
содержатся короткое время (рис. 7.1). Процессы произвольного внимания по-
зволяют выбрать из всех поступивших стимулов какую-то часть, которая затем
подвергнется дальнейшей обработке. На этой стадии информация о только что
поступивших сенсорных стимулах взаимодействует с уже имеющимся опытом
и знаниями, а также с информацией от других сенсорных систем. Кроме того,
важную роль играет конечная цель — действие, которое предстоит исполнить
в связи с этой информацией, — она оказывает определенное воздействие на то,
как информация кодируется и как она будет храниться. Важно отметить, что
описанная модель предполагает не односторонний процесс, в котором инфор-
мация о звуке поступает в систему, декодируется, «понимается», а потом запи-
сывается в долговременной памяти. В течение всего времени анализа стимулов
Слух и речь
305
в слуховой системе происходит взаимодействие как между ее собственными
частями, так и между другими сенсорными, когнитивными, моторными систе-
мами и системами памяти. Анатомия и морфология слуховой системы, которая
включает в себя несколько последовательных этапов анализа, восходящие пути
от уха к коре больших полушарий, нисходящие пути, по которым информация
поступает назад к периферической нервной системе, а также большое количе-
ство параллельных путей обработки информации, — все это отражает разноо-
бразие и сложность связей внутри и вне слуховой системы.
1.1.2. Ограниченные и имеющие большую емкость возможности
Как уже обсуждалось в гл. 1, мозговые процессы, с одной стороны, достаточно
ограничены, а с другой, имеют очень большие возможности. Это действительно
и для слуховой системы. Существуют определенные нарушения в декодирова-
нии звуковых стимулов. Например, если отрывок речи предъявляется через на-
ушники, можно легко различать каждое отдельно произносимое слово. Однако,
если два разных речевых потока предъявляются в два уха одновременно, стано-
вится очень трудно прислушиваться к каждому. На самом деле мы выборочно
слушаем или один речевой поток, или другой (Broadbent, 1982). Таким образом,
возможности слуховой системы ограниченны для восприятия сложных звуко-
вых стимулов, и определенную роль в этом процессе играют центральные моз-
говые механизмы, которые могут направлять внимание на те или иные аспек-
ты звукового окружения. Но наша способность к изучению новых звуков или
Рис. 7 .1. Рабочая схема слуховой обработки на основе общей рабочей схемы, пред-
ставленной в гл. 2 . Входящие сенсорные сигналы, такие как звук голоса или звонок
мобильного телефона, поступают в систему (см. красные стрелки слева). Есть ранние
влияния восходящих и нисходящих процессов, относящихся к вниманию. Сенсорные
входы связаны с рабочей и долговременной памятью, а также с двигательной системой.
Важно помнить, что процессы, обеспечивающие функцию слуха, — это высокоинтерак-
тивные и интегративные процессы с прямыми и обратными связями
306
Глава 7
акустических объектов (таких как слова, например) неизменна на протяжении
жизни и представляется практически неограниченной: средний словарный за-
пас взрослого человека составляет более 100 000 слов. То же самое справедливо
и в том, что касается узнавания новых мелодий, а также голосов друзей и зна-
комых. Получается, что, несмотря на существование ограничения восприятия
звуков, как только они восприняты, наша возможность их запомнить оказыва-
ется безграничной.
1.1.3. Порядок величин и уровни анализа
Как и в других мозговых системах, в слуховой системе обработка информации
идет на разных уровнях: от индивидуальных волосковых клеток на перифери-
ческом уровне, через отдельные нейроны слуховой коры к распределенным
нейронным сетям, включающим разные отделы мозга. Слуховая система из-
учается на каждом из этих уровней как на человеке, так и на животных. В этой
главе мы рассмотрим те факты, которые относятся к разным уровням. Однако
наибольшее внимание мы уделим данным, которые связаны со слуховой систе-
мой в целом. Эти данные получены с помощью методов нейровизуализации:
ПЭТ (позитронно-эмиссионной томографии), яМРТ (ядерной-магнитно-ре-
зонансной томографии), фМРТ (функциональной магнитно-резонансной то-
мографии), МЭГ (магнитоэнцефалографии) и ЭЭГ (электроэнцефалографии).
Изобретение неинвазивных методов исследований произвело революцию в об-
ласти когнитивных нейронаук и психологии. До этого данные получали в экс-
периментах на животных, поведенческих и психофизических экспериментах
на здоровых людях, а также анализировали особенности восприятия звуков
и речи у больных, мозг которых по той или иной причине поврежден (меха-
нические повреждения, опухоли, удары). Возможность исследовать мозговые
процессы у здоровых людей позволила получить большое количество новых
данных. Кроме того, появилась возможность изучать системы, которые широко
распределены по всему мозгу, такие как, например, речевая система.
1.1.4. Время
Время — это один из ключевых аспектов для обработки слуховой информации:
в отличие от зрительной системы все процессы в слуховой системе растянуты
во времени. Самым сложным для обработки типом сигнала по праву считается
речь: различия между фонемами /b/ и /�/ заметны только на промежутке време-
b/ и /�/ заметны только на промежутке време-
/ и /�/ заметны только на промежутке време-
�/ заметны только на промежутке време-
/ заметны только на промежутке време-
ни около 20–30 мс. Тем не менее слуховая система справляется с такой задачей
даже в относительно зашумленных условиях (Gage et al ., 1998, 2002).
Таким образом, система распознавания звуков речи имеет высокое разреше-
ние в области быстрых и тонких изменений на уровне фонем. Однако для рече-
вой системы в целом необходимо также анализировать информацию на более
длительных временных промежутках: на уровне слогов и ударений (например,
различия в ударениях между словами) изменение происходят со временем поряд-
ка 200 мс; интонационные изменения (такие как изменение интонации на вопро-
сительную), которые важны для восприятия целых предложений — со временем
около 1–2 с. Каждое из этих временных окон — 20, 200, 2000 мс — оказывается
критичным для анализа информации о речевых звуках. Результат такого анализа
необходим для целого комплекса процессов, которые лежат в основе преобразо-
вания звуков речи в единицы осмысленной информации (рис. 7.2).
Слух и речь
307
Перед обсуждением того, как мозг анализирует информацию о своем звуко-
вом окружении, состоящем из человеческих голосов, звуков природы и музыки,
нам надо разобраться в базовых характеристиках звука и слуха. Мы начнем с
физических характеристик звуковых стимулов и того, как они связаны с харак-
теристиками психологическими. После этого мы перейдем к процессам, про-
текающим на разных стадиях периферического слуха, и подкорковым механиз-
мам извлечения ключевых для слуховой системы признаков.
1.2.
Основы звука и слухового восприятия
1.2.1. Физические свойства звука
Каким образом слуховая система человека превращает набор звуков в понятную
речь или узнаваемые мелодии? Давайте вспомним, каким образом мы воспри-
нимаем простые звуки на уровне уха. С точки зрения физики, звук представляет
из себя колебания, которые вызваны движением объекта в пространстве. Мы
слышим не сами колебания, а то, как они распространяются в пространстве в
виде звуковых волн, а затем вступают в контакт с нашими ушами.
Базовым кирпичиком, из которого строятся все остальные звуки, является
синусоидальная волна. Три ее основных физических аспекта — частота, ин-
тенсивность и время. Частота звука — это характеристика звуковых колебаний,
она измеряется как количество колебаний в секунду, единица частоты — 1 Гц.
Человеческое ухо способно воспринимать звуки в широком диапазоне частот,
в среднем от 20 до 20 000 Гц. Интенсивность синусоидального звука отражает
амплитуду (или смещение) звуковой волны за один цикл (или период) колеба-
ний. На рис. 7 .3 (слева) показан синусоидальный тон с частотой 1000 Гц, на оси
абсцисс отложено время, на оси ординат — интенсивность. На том же рисунке
справа — тот же тон с частотой по оси ординат и временем по оси абсцисс.
Обратите внимание, что спектральная энергия синусоидального тона огра-
ничена узкой полосой только в области частот, равной частоте тона. В связи
Рис. 7 .2 . Спектрограмма — это ото-
бражение связанных со звуком осо-
бенностей речи. По оси х отложено
время, по оси у — частота. Более тем-
ные участки соответствуют большей
интенсивности. Речь включает в себя
гармоническую компоненту (форман-
ту) в специфических областях в спек-
тральном (основанном на частоте)
аспекте спектрограммы. Здесь пред-
ставлена спектрограмма, показываю-
щая три наиболее существенных для
расшифровки речи временных мас-
штаба. Вверху сл ева: детали помех,
возникающих при произнесении со-
гласных, масштаб 20 мс. Вверху спра-
ва: детали форманты, возникающей
при произнесении слогов, масштаб
200 мс. Внизу: предложение, масштаб
2000 мс
308
Глава 7
с этим такие тона часто называют «чистыми» тонами. Большинство звуков,
которые мы слышим в реальной жизни, конечно, не являются чистыми тона-
ми. Фортепианный аккорд, гудок автомобиля, человеческий голос — все они
имеют гораздо более сложную структуру. Каким образом мы можем описать
сложные звуковые стимулы в терминах таких трех физических параметров, как
частота, интенсивность и время? Джозеф Фурье (1768–1830), французский ис-
следователь, живший в эпоху Наполеона, доказал несколько теорем, которые
позволили раскладывать сложные сигналы на набор из более простых. Такая
процедура получила название Фурье-анализ (Фурье, 1822). Работу Фурье про-
должил Георг Ом (1789–1854), который предположил, что такую же процедуру
разложения сложного сигнала на более простые синусоидальные производит
человеческое ухо при восприятии звука.
Кроме уже описанных частоты, интенсивности и времени звук имеет и дру-
гие характеристики. Когда ноту До (261 Гц) сначала сыграют на фортепиано, а
потом на гобое, сможете ли вы отличить одну от другой, даже несмотря на то,
что они имеют одинаковую частоту? Да, причем сделать это можно с легко-
стью, а следовательно звук должен иметь среди своих характеристик не только
частоту. В приведенном примере отличить две ноты можно благодаря тому, что
они имеют разный тембр звучания. Разный тембр также помогает нам разли-
чать человеческие голоса.
1.2.2. Масштаб измерения интенсивности звука
Динамический диапазон слуховой системы человека очень широк: мы можем
слышать едва уловимые звуки слабой очень маленькой интенсивности и очень
громкие звуки, которые иногда даже вызывают болевые ощущения. Интен-
сивность в таком диапазоне колеблется от 1 ед. до 1 000 000 000 000 000 (1015).
Поэтому для того, чтобы измерять интенсивность звука, удобно пользоваться
логарифмической шкалой децибел (дБ). Шкала децибел относительна (а не аб-
солютна), она основана на отношении пары величин: собственно интенсивно-
сти звука и либо уровня звукового давления в воздухе (тогда измеряется дБ по
Рис. 7 .3 . Слева: изображение синусоидального тона. По оси х – время, по оси у – ам-
плитуда. Частота синусоиды связана с количеством циклов в секунду, т. е. тон с частотой
1000 Гц соответствует 1000 циклам в секунду. Справа: изображение синусоидального
тона с частотой, сфокусированной на частоте 1000 Гц. Синусоидальные тоны имеют
единственную частоту, поэтому их рассматривают как «чистые» тоны. Большинство
звуков, которые мы слышим, характеризуются сочетанием множества частот. (Источ-
ники: (слева) Brown, 2003; (справа) Boatman, 2006.)
Слух и речь
309
УЗД, уровню звукового давления), либо уровня, который соответсвует слухо-
вому порогу испытуемого (дБ СП). (Заметьте, что это только два чаще всего
используемых варианта измерения интенсивности, существуют и другие.) Че-
ловеческое ухо способно воспринимать звук интенсивностью от 1 дБ до 150 дБ
УЗД (рис. 7 .4).
1.2.3. Психологические характеристики звука
Звук имеет физические характеристики, которые мы можем измерить с до-
статочно большой точностью. Однако как нам узнать как он воспринимается?
Частота звука связана с психологическим параметром высоты. Человеческий
голос или нота, сыгранная на пианино, могут звучать «высоко» или «низко».
Когда мы говорим о восприятии звука, нам необходимо различать его физи-
ческие и психологические характеристики. Так, мы точно знаем частоту зву-
ка, который мы предъявляем испытуемому, но не знаем, звук какой высоты он
на самом деле слышит. То, как воспринимает различия между звуками разной
высоты опытный оперный певец, может сильно отличаться от того, как те же
различия воспринимает обычный человек даже при условии, что оба они име-
ют нормальный слух. То же самое касается и интенсивности, которая связа-
на с психологической громкостью звука. Разные люди сильно различаются по
тому, как они воспринимают громкость. Факторы, которые могут влиять на этот
Рис. 7 .4 . Порог слышимости и границы слуха у человека. Показаны также частотные
характеристики и уровни звукового давления распространенных звуков окружающей
среды, в том числе человеческой речи. Наиболее интенсивные звуки могут повреждать
рецепторный аппарат внутреннего уха. Также показаны слуховые чувствительности
у кошки, лабораторного животного, обычно используемого в изучении центрального
и периферического отделов слуховой системы. (Источник: Brugge and Howard, 2002,
с изменениями с разрешения.)
310
Глава 7
процесс, также разнообразны, от нарушений слуха до индивидуальных пред-
почтений и привычек. Таким образом, когда мы описываем звуковые стимулы
надо хорошо различать, говорим ли об измеренных физических или о субъек-
тивных психологических характеристиках.
1.2.4. От барабанной перепонки к слуховому нерву
Как мы уже говорили выше, мы будем идти постепенно, затрагивая разные
уровни переработки слуховой информации и извлечения ключевых признаков
звука в той же последовательности, которую звук проходит на пути к слуховой
коре. Вы увидите, что даже при анализе простых звуковых стимулов в тихом
окружении необходимо задействовать комплексные сложные механизмы. Да-
вайте начнем с рассмотрения того, как звук через барабанную перепонку воз-
действует на жидкость внутреннего уха для того, чтобы иннервировать волокна
слухового нерва (рис. 7 .5).
Колебание объекта вызывает звуковые волны, которые распространяются в
воздухе. Звуковые волны достигают барабанной перепонки, механически за-
действуют три слуховых косточки среднего уха: молоточек, стремечко и нако-
Рис. 7 .5 . Рисунок периферического отдела слуховой системы человека. Указаны на-
ружное ухо (ушная раковина и наружный слуховой проход) и среднее ухо (мембрана
барабанной перепонки и три слуховые косточки: молоточек, наковальня и стремечко).
Также показано внутреннее ухо, включающее улитку слуховой системы и полукружные
каналы вестибулярной. Два слуховых окна: круглое и овальное. Через овальное окно
стремечко передает звуковые колебания жидкости, заполняющей полость внутреннего
уха. (Источник: Brown, 2003.)
Слух и речь
311
вальню. Колебания звука таким образом усиливаются и достигают органа слу-
ха — улитки, расположенного во внутреннем ухе (рис. 7 .5) (более подробно см.
Moore, 1995).
На уровне улитки происходит декодирование физических характеристик
звука (рис. 7 .6, вверху, представлена форма базилярной мембраны улитки). Бе-
гущая волна звука движется по базилярной мембране от основания к вершине
(рис. 7.6, внизу). Базилярная мембрана имеет частотно-специфическую топо-
графическую (тонотопическую) организацию, звуки высокой частоты кодиру-
ются у основания мембраны, низкой — у вершины.
Каким образом бегущая волна преобразуется в нейронный код и передается
мозгу?
Внутри улитки находится около 16 000 рецепторных клеток, которые назы-
ваются волосковыми. Движение бегущей волны вдоль базилярной мембраны
приводит в движение крохотные волосковые клетки. В том месте, где бегущая
волна имеет максимальную амплитуду, происходит максимальное смещение
Рис. 7 .6 . Вверху: передача звука с изображением улитки в таком разрезе, что видна
базилярная мембрана. Стоит отметить, что красные стрелки показывают направление
передачи звука и действуют в двух направлениях. Внизу: изображение движения звуко-
вой волны через базилярную мембрану. Волна показана в застывший момент времени и
увеличена, чтобы проиллюстрировать движение звука по мембране. (Источники: (ввер-
ху) Drake, 2005, (внизу) Javel, 2003.)
312
Глава 7
волосковых клеток. Они, в свою очередь, расположены в определенных местах
на базилярной мембране, таким образом кодируя частоту стимула. Такой прин-
цип кодирования называется принципом места. Он основан на предположении,
что мозг декодирует информацию о частоте стимула на основе максимальной
активации волосковых клеток в определенном локусе на базилярной мембране.
На этом этапе обработки информации движение волосковых клеток, вызван-
ное бегущей волной, преобразуется в электрический сигнал внутри волокон
слухового нерва (Kelly et al., 1996). Отдельные волосковые клетки связаны с
конкретными нервными волокнами. Эти волокна имеют свою характеристи-
ческую частоту, к которой они наиболее чувствительны.
Связь волосковых клеток с определенными волокнами слухового нерва по-
зволяет сохранить информацию о частотных характеристиках звука на всем
пути от колебаний барабанной перепонки через движение жидкости во вну-
треннем ухе и бегущую волну в улитке к нейронному коду внутри слухового
нерва. На рис. 7 .7 вверху схематически изображен путь слуховой информации
в мозге, внизу — нервные волокна, которые показывают наилучший ответ на
определенной частоте.
Таким образом, звук проходит путь от наружного слухового прохода к сред-
нему и внутреннему уху, а оттуда в слуховую систему мозга. На этом этапе
информация, закодированная в электрических импульсах волокон слухового
нерва, поступает в слуховую кору. Вы можете заметить, что до того, как ин-
формация достигает коры, много раз происходит ее преобразование и переко-
дирование. Эти преобразования являются ключом к пониманию того, как мозг
расшифровывает сложную звуковую картину окружающего мира, подобную
ситуации в аудитории, которая была описана в начале этой главы.
2.0 .
Центральная слуховая система
Информация о звуковых стимулах многократно изменяется в процессе пере-
дачи в слуховую кору головного мозга. В данном разделе мы рассмотрим ос-
новные анатомические и нейрофизиологические аспекты центральной слухо-
вой системы. В слуховой системе можно выделить различные этапы обработки
информации, начиная от уха и заканчивая стволом головного мозга, подкорко-
выми ядрами и корой. Тремя основными отделами слуховой системы являются
периферическая система, которая уже обсуждалась нами ранее, пути передачи
(восходящие пути к коре, нисходящие пути от коры и параллельные пути между
корковыми областями) и центральная (корковая) система. Хотя на каждом этапе
происходит обработка различных аспектов звуковой информации, важно рас-
сматривать слуховую систему как единое целое в связи со сложным характером
взаимодействия как между ее составляющими, так и внутри них.
2.1.
Пути передачи слуховой информации
Как упоминалось ранее, процесс обработки звуков растянут во времени. От-
личительной чертой слуховой системы является способность к восприятию
информации с высоким временным разрешением (Gage, Roberts, 2000; Gage
et al., 2006). Важным аспектом временной разрешающей способности слуховой
системы является быстрая и точная передача поступившей звуковой информа-
ции. Кроме того, необходима не только быстрая передача сигнала от среднего
Слух и речь
313
Рис. 7.7 . Вверху: изображение афферентной и эфферентной иннервации клеток Кор-
тиева органа, расположенных внутри улитки (см. рис. 7 .6). Афферентная иннервация
обеспечена ганглиозными клетками спирального ганглия улитки, которые, в свою оче-
редь, иннервируются волокнами слухового нерва. Изображены два типа чувствитель-
ных нейронов: 1) нейроны типа I, получающие информацию от внутренних волосковых
клеток, и 2) нейроны типа II, получающие информацию от внешних волосковых клеток.
Аксоны ганглиозных клеток направляются в центральный отдел слуховой системы и
формируют слуховой нерв. Эфферентная иннервация обеспечивается субпопуляцией
нейронов ядер верхней оливы, которые посылают аксоны в улитку и потому называют-
ся оливокохлеарные (ОК) нейроны. Существует два типа ОК нейронов: 1) латеральные
ОК нейроны иннервируют внутренние волосковые клетки, 2) медиальные ОК нейроны
иннервируют наружные волосковые клетки. Латеральные ОК нейроны расположены в
основном ипсилатерально по отношению к иннервируемой области улитки, в то время
как медиальные — билатерально, из них две трети контралатерально и треть ипсилате-
рально. Внизу: кривые настройки слухового нерва. На левой кривой нервное волокно
настроено на 2000 Гц, на правой на 15 000 Гц; частота по оси х. По оси у пороговые
значения уровня слухового давления в децибеллах. (Источники: (вверху) Brown, 2003;
(внизу) Javel, 2003.)
314
Глава 7
уха (барабанной перепонки) к коре, которая необходима для составных частей
сложных звуков, например согласных звуков в речи или элементов музыкаль-
ной фразы, важно также то, что информация, получаемая от обоих ушей, долж-
на быть объединена и передана в надлежащем виде. Давайте обсудим, где и
каким образом это происходит в процессе обработки звука. Восходящие (аф-
ферентные) пути передают информацию о поступающих звуках от периферии
к коре. Данные пути включают множество этапов обработки: они не является
простой системой передачи сигналов, так как в процессе прохождения сигнал
подвергается многочисленному кодированию и расшифровке. Нервный сигнал
идет от слухового нерва в нижнее (вентральное) кохлеарное ядро. Кохлеарное
ядро имеет тонотопическую структуру. От ядра сигнал далее по восходящему
пути проходит через латеральную петлю, нижние бугры и таламус к слуховой
коре (рис. 7 .8). Данный путь передачи сложен и включает множество этапов
обработки и процессов комбинирования звуков, поступающих от обоих ушей.
Ключевой функцией восходящего пути является оценка информации, поступа-
ющей от обоих ушей, с целью определения местонахождения источника звука
в пространстве — позже мы остановимся на этом более подробно. Нисходящие
(эфферентные) пути проходят от различных областей корковой и подкорковой
слуховых систем коры к периферии под прямым или непрямым контролем
коры. Недавно проведенные исследования говорят о том, что данный контроль
наблюдается на протяжении всего пути вплоть до волосковых клеток улитки!
Важной функцией нисходящего пути является проведение информации, позво-
ляющей избирательно обрабатывать отдельные звуки из массы всех поступаю-
щих звуков. Механизм, с помощью которого происходит обработка звуков при
нисходящем пути у человека, не изучен до конца. Однако вот пример некоторых
аспектов слушания, в которых ключевую роль играют нисходящие пути. Пред-
ставьте̧ что у вас проходит очень важная беседа с близким другом снаружи
классной комнаты. Вы сосредоточены на разговоре, но в это время на соседней
стоянке паркуется мотоцикл, над головой пролетает вертолет, а садовник косит
близлежащую лужайку мощной газонокосилкой. Вы стараетесь услышать сво-
его друга, но при этом еще игнорируете большинство конкурирующих звуков.
Неожиданно дверь классной комнаты открывается и оттуда выбегает тол-
па переговаривающихся и смеющихся учащихся. Это может положить конец
вашей беседе на некоторое время, однако в процессе слушания голоса вашего
друга среди всего этого шума ваши слуховые пути находятся в работе, направ-
ленной на то, чтобы помочь вам обоим сконцентрироваться на беседе и выде-
лить голоса друг друга на фоне других звуков, идущих отовсюду.
Слуховые пути не только восходят к коре или исходят от нее, но и сущест-
вует множество важных связей между слуховой корой левого и правого полу-
шарий, чье сообщение происходит через мозолистое тело. Данные связи между
полушариями имеют тонотопическую организацию. Существуют также корти-
ко-кортикальные пути, обеспечивающие связь между процессами обработки
слуховой информации и другими сенсорными системами, равно как и с процес-
сами работы и долговременной памятью. Совместно с восходящими и нисходя-
щими путями кортикальные пути представляют собой паттерны связи, являю-
щиеся необходимыми не только для обработки звука, но и для интегрирования
информации в другие отделы головного мозга.
Слух и речь
315
2.2 .
Слуховая кора
Мы подошли к рассмотрению слуховой коры и в данном разделе обсудим
анатомию отделов мозга в слуховой коре, равно как и нейрофизиологические
особенности данных областей. Слуховая кора является отделом мозга, специ-
ализирующимся на обработке звука. Она расположена в каждом полушарии в
области сильвиевой борозды на поверхности надвисочной области и верхних
височных извилин (рис. 7 .9).
Как обсуждалось ранее, информация о звуковых стимулах передается из уха
в слуховую кору по восходящим слуховым путям. В процессе передачи сигнал
множество раз трансформируется и пересчитывается.
Слуховая кора не является конечной точкой данного пути, однако служит в
качестве основы или звена для процесса обработки звука, динамично взаимо-
действуя с другими системами в пределах коры, через полушария и по нисходя-
щим путям к улитке. Данные динамические процессы обеспечивают широкий
диапазон перцепционной остроты и позволяют нам выполнять сложные пер-
цепционные задачи, такие как селективное слушание голоса отдельного чело-
Рис. 7 .8 . Слуховая система человека. Показаны пути и подкорковые ядра восходящих и
нисходящих путей. (Источник: Standring, 2005.)
316
Глава 7
века в шумной комнате, полной людей, или узнавание мелодии, даже если она
сыграна в другой тональности или в другом темпе.
Данные перцепционные задачи настолько сложны, что мы полностью не по-
нимаем, каким образом их решает слуховая система. Однако мы проделываем
их каждый день без осознания или с лишь незначительным осознанием нашего
в них участия. Слуховая кора не является одиночной областью головного моз-
га, она представлена несколькими структурными (анатомическими) областями,
различающимися по их роли в декодировании звука. Ранние описания этих об-
ластей в пределах слуховой коры, например цитоархитектоническая классифи-
кация, были сделаны на основании структур, в частности извилин в корковой
области или нейрофизиологических черт. Несмотря на то что роль каждой об-
ласти в пределах слуховой коры человека до конца не изучена, работа в этом на-
правлении ведет к выявлению границ этих областей и пониманию их функций
в перцепции. Ниже представлено обсуждение текущих знаний о слуховой коре
человека с описанием структуры или анатомического строения, сопровожда-
ющееся детальным описанием их клеточной организации и свойств отклика,
а также нейрофизиологических свойств.
Рис. 7 .9. Вверху: вид с латеральной стороны головного мозга человека. Внизу: вид с ме-
диальной стороны. Раскраска полей коры головного мозга из Бродмана (1909). Слухо-
вые и рецептивные языковые области включают поля 22, 41, 42, и 52 по Бродману
Слух и речь
317
2.2.1 . Анатомия слуховой коры
Большинство из того, что мы знаем об анатомии слуховой коры, происходит из
работ по изучению обезьян (Galaburda and Pandya, 1983). У макак основными
отделами слуховой коры являются ядра, пояс и парапояс (рис. 7 .10) (в русской
анатомической традиции такие зоны не выделяют, а говорят о участках ви-
сочной коры; поясная извилина находится в другом месте.
—
Прим. перев.).
Данные отделы известны своими цитоархитектоническими, физиологически-
ми и соединительными свойствами. Ядро принимает сигналы от таламуса и,
в свою очередь, передает его в латеральные поясные области (Kaas et al., 1999).
Эта анатомическая и связующая структура привела исследователей к предполо-
жению, что слуховая кора имеет иерархическую организацию с подкорковыми
проекциями на ядро или первичную слуховую область (А1). Предполагается,
Рис. 7 .10 . Вверху: изображены уровни и области обработки слуховой информации в
коре макак. Изображенные области располагаются в верхней части височной извилины
и в глубине височной борозды, которая раскрыта, чтобы показать расположение коры,
связанной со слухом внутри борозды. ДБ — дугообразная борозда; ЦБ — центральная
борозда; НТБ — нижняя теменная борозда; СБ — серповидная борозда. Внизу: анало-
гичные области у человека, с разрезом сильвиевой (латеральной) борозды, чтобы по-
казать слуховую кору в суправисочных областях
318
Глава 7
что А1 декодирует базовые характеристики и свойства звуков. От А1 исходят
проекции на поясные области, где происходят процессы дальнейшей расшиф-
ровки более сложных компонентов звука (Kaas et al., 1999). Области пояс и
парапояс, окружающие А1, специализируются на обработке звука и, таким об-
разом, являются однородной слуховой корой. В слуховой коре человека (см.
рис. 7 .10, правая панель) имеются те же области, сильвиева борозда раскрыта,
чтобы показать области слуховой коры, соответствующие областям, имеющим-
ся у макак (см. рис. 7 .10, слева).
У человека первичная слуховая кора расположена в извилине Гешля
(рис. 7.11, слева) и практически полностью аналогична корковым областям,
описанным у обезьян. Извилина Гешля соответствует полю 41 (Бродманн,
1909). Как правило, первичная слуховая кора является составной частью из-
вилины Гешля. Наблюдается широкое разнообразие в анатомии этой структуры
как между обоими полушариями, так и между отдельными индивидуумами: в
правом полушарии извилина Гешля обычно расположена немного более впе-
реди (на 6 мм), чем в левом, также у некоторых индивидуумов имеется более
одной извилины Гешля. Структурное разнообразие в извилине Гешля имеет
важное значение при интерпретации результатов нейровизуализации, так как
реальный размер и расположение извилины Гешля сильно различаются между
индивидумами (рис. 7 .11, справа; Frakowiak, 2004).
Слуховая кора располагается в направлении сверху вниз в извилине Гешля,
а также в направлении назад и сверху вдоль надвисочной области и верхнего
края верхней височной извилины. Второй анатомически важной областью слу-
ховой коры человека является височная область, расположенная позади изви-
лины Гешля. В строении височной области наблюдаются различия как между
полушариями, так и между индивидуумами. Однако, в отличие от извилины
Гешля, эти отличия проявляются в том, что височная область, как правило, на-
много крупнее в левом полушарии по сравнению с правым. В действительно-
сти височная область в левом полушарии может быть до десяти раз больше у
праворуких индивидуумов (см. рис. 7 .11, слева). Асимметрии височной области
Рис. 7 .11 . Сл ева: области слуховой коры, которые отражают типичную асимметрию
правого и левого полушария. Справа: МРТ-визуализация обработки звука в этих же об-
ластях. (Источник: (слева) Standring, 2003; (справа) Frackowiak, 2004.)
Слух и речь
319
были описаны в серии анатомических исследований, проведенных Гешвиндом
и коллегами (Geschwind and Galaburda, 1985a, b, c). Учеными был отмечен тот
факт, что речевая функция преобладает в левом полушарии. Предполагается,
что большие размеры височной области, расположенной в левом полушарии,
отражают ее роль в декодировании звуков речи. Последующие недавние иссле-
дования, однако, дали возможность специфически проанализировать данную
гипотезу, что, в свою очередь, породило различные взгляды на роль височной
области в обработке звука. Впереди извилины Гешля располагается planum p��
p��
p�-
lare. Эта область не являлась объектом многочисленных исследований у чело-
века, в связи с чем данных о ее роли в перцепции звука крайне мало. Позади
planum p�lare и однородных слуховых областей располагается поле Бродман-
на 22. Карл Вернике выдвинул гипотезу о важности данной области в пони-
мании речи (Wernicke, 1874/1977). В соответствии с гипотезой Вернике эта
область не является слуховой по существу, но образует языковую область, не-
обходимую для процессов понимания речи, которые тесно связаны (физически
и функционально) с процессами обработки звука. Данная область, как правило,
рассматривается как область Вернике.
2.2.2. Нейрофизиология
Некоторые руководящие принципы организации слуховой коры были установ-
лены в исследованиях, проведенных на кошках и обезьянах. Основными орга-
низационными единицами слуховой коры, как и в других корковых сенсорных
зонах, являются нейроны, колонки неокортекса и нейронные сети.
В слуховой системе наблюдается несколько различных видов нейронов
(рис. 7 .12). Различные свойства ответов данных нейронов кодируют частоту,
интенсивность и временные характеристики звуков, а также кодируют про-
странственную информацию, необходимую для локализации звуков в про-
странстве. Большинство корковых нейронов откликаются на сигналы от обоих
ушей, демонстрируя тем самым важность корковых процессов декодирования
бинауральной информации для локализации звука, а также других сложных
слуховых процессов. Сообща все типы нейронов формируют динамическую
сеть, кодирующую временные свойства звуков. Значительно меньше извест-
но о типах нейронов в слуховой коре человека. Однако крупные пирамидные
клетки, распределенные вдоль надвисочной области, вероятнее всего, играют
ключевую роль в обработке звуков.
Определение местоположения рецептивных полей в нейронах слуховой
коры являлось предметом многочисленных исследований на животных. Боль-
шая часть нейронов слуховой коры откликаются на сигналы от обоих ушей.
Однако оба уха представлены по-разному в пределах одного полушария. В слу-
ховой коре левого полушария правое ухо, т. е . противоположное, представле-
но намного значительнее или интенсивнее, чем левое̧ размещенное на той же
стороне.
Тот же образец схожести и различия наблюдаются в слуховой коре правого
полушария, левое ухо представлено значительнее, чем правое. Данная асимме-
трия в представлении обоих ушей при бинауральном слушании справедливо
также и для человека (Woldorff et al., 1999). Важным аспектом декодирования
характеристик звука является определение местоположения звука относитель-
но слушателя. На рис. 7 .13 представлены рецептивные свойства двух нейронов
320
Глава 7
слуховой коры в виде функции, в которой ось х представлена определением
местоположения звука в пространстве относительно головы, а ось y — гром-
костью звука. Рецептивные свойства, представленные на верхней панели, де-
монстрируют, что нейрон способен воспринимать широкий набор оттенков
поступающего звука от противоположного уха, а также различать их интенсив-
ность. Эти свойства показывают, что данный нейрон неспецифичен в отноше-
нии звуковой информации, передаваемой через уровни громкости от противо-
положного уха. На нижней панели у нейрона наблюдается чувствительность,
сфокусированная в большей степени и представленная диапазоном громкости
30–60 дБ, исходящих от ипсилатерального уха. Нейрон специфичен. Специфич-
ные и неспецифичные нейроны играют разную роль в обработке звука. Неспе-
цифичный нейрон не способен предоставить детализированную информацию о
точном местоположении звука в пространстве или о его точной громкости. Од-
нако данный нейрон будет высокочувствителен при определении какого-либо
звука в широком диапазоне громкости, исходящего от противоположного уха.
Рис. 7 .12 . Схематичное изображение основных анатомических типов клеток кохлеар-
ных ядер, а также их постстимульные (ПСТ) гистограммы. Слева: волокно слухового
нерва и его типичный ответ, первичная ПСТ. В центре: разветвление слухового нерва
для иннервации разных типов клеток кохлеарного ядра. Справа: ПСТ-гистограммы для
каждого типа клеток
Слух и речь
321
Таким образом, этот нейрон необходим для обнаружения новых появляющихся
звуков и предоставлении общей информации о том, от какого уха исходит этот
звук. Специфичный нейрон предоставляет более специфическую информацию
о местоположении звука, т. е . не только от какого уха он исходит, но и о поло-
жении головы относительно звука, а также специфическую информацию о его
громкости. Таким образом, этот нейрон важен для дискриминации детальной
информации о звуке.
Аналогично другим корковым сенсорным областям, слуховая кора имеет
колончатую структуру, т. е . нейроны слуховой коры расположены в виде коло-
нок, проходящих через все шесть корковых слоев (см. гл. 3). В пределах одной
колонки нейроны проявляют одинаковые свойства ответов. Главная особен-
ность колончатой структуры слуховой коры состоит в том, что нейроны, от-
кликающиеся на бинауральные сигналы, образуют перемежающиеся колонны,
Рис. 7 .13 . Рецептивные поля двух нейронов слуховой коры, изображенные как функ-
ция звукового давления с азимутом во фронтальной полуобласти. В качестве стимулов
использовались шумовые посылки. Небольшими ромбами показаны области макси-
мальной активности, уменьшение яркости цвета показывает уменьшение активности.
Нулевой азимут относится к направлению прямо перед головой, положительный —
к контралатеральной полуобласти. УЗД — уровень звукового давления. (Источник:
Brown, 2003.)
322
Глава 7
играющие различные роли в обработке звука, порождая как возбуждающий
(суммация), так и ингибирующий (супрессия) эффект на информацию, идущую
от обоих ушей (Brugge and Merzenich, 1973). Эти комплексные взаимодействия
суммации и супрессии информации, идущей от обоих ушей, вероятно, лежат в
основе перцепционных функций, в частности избирательного восприятия ин-
формации, идущей от одного уха.
Рис. 7 .14 . Слуховые поля в височной коре кошки. (a) Латеральный срез. (б) «Развертка»
латерального среза для изображения областей, которые обычно скрыты внутри извили-
ны (выделены оранжевым), а также связаны с низко- и высокочастотными ограничени-
ями полей с тонотопической организацией. Выделяют четыре тонотопических области:
передняя (A, anterior), первичная (AI), задняя (P, �osterior), задняя вентральная (VP).
Обратите внимание, что на границах тонотопических областей направление инверти-
ровано таким образом, что прилегающие поля имеют «зеркальную» тонотопическую
организацию. В других областях коры тонотопическая организация выражена меньше.
Это вторичная (AII), височная (T) и заднедорсальная (DP, dorso�osterior) области. Также
изображены надсильвиева борозда (sss) и передняя и задние эктосильвиевы борозды
(aes, �es). (Источник: Brown, 2003.)
Слух и речь
323
Основной главенствующий принцип слуховой коры обезьян заключается в
тонотопической организации. К примеру, в слуховой коре кошек рецептивные
области нейронов демонстрируют тонотопическую организацию первичного
поля (А1), которые, в свою очередь, имеют зеркальные проекции в смежных
(впереди и позади) корковых областях (рис. 7 .14).
В то время как основные нейрофизиологические аспекты, вероятнее всего,
аналогичны для человека, уникальность восприятия человеческой речи, языка
и музыки, а также значительно большая доля всей коры, отданная слуховой
коре, возможно, означает, что существуют нейроны и сети нейронов, специ-
ализирующиеся на этих сложных процессах и характерных только для чело-
веческой слуховой коры. К примеру, ряд областей слуховой коры у обезьян
имеет тонотопическую организацию, наличие тонотопии было менее очевид-
но в исследованиях на людях, поскольку исследование ограничено первичной
слуховой корой (Wessinger et al ., 2001). Исследование принципов организации
человеческой слуховой коры ведется и на данный момент, причем существен-
ную помощь оказывает открытие новых технических возможностей для этих
исследований, в частности транскраниальной магнитной стимуляции (ТМС)
или функциональной магнитно-резонансной томографии (фМРТ).
3.0 .
Функциональное картирование обработки
слуховой информации
Существуют ли внутри слуховой коры отдельные области, которые специфи-
чески связаны с восприятием отдельных тонов или речевых или музыкальных
звуков? Или вся кора целиком включена в процесс обработки информации це-
ликом, вне зависимости от типа стимуляции? Одинаковы ли процессы обра-
ботки слуховой информации в правом и левом полушариях или между ними
существуют тонкие различия? Ученые, которые работают в области слухового
восприятия, до сих пор ищут ответы на эти вопросы. Изобретение методов ней-
ровизуализации открыло новые пути исследования зон мозга, связанных с об-
работкой слуховой информации. Большинство работ в этой области направлены
на изучение областей, которые ответственны за расшифровку речевых сигна-
лов. В то время как нейропсихологические исследования больных с поражени-
ями мозга принесли большое количество данных, которые касаются языковых
нарушений и их связи с конкретными областями мозга, методы нейровизали-
зации позволяют лучше понять, как протекают речевые процессы у здоровых
испытуемых. Эти методы позволяют также изучить те аспекты функционирова-
ния слуховой системы, которые были недоступны для исследователей до этого,
например отличия мозговых зон, которые задействованы при реальном воспри-
ятии стимула и воображаемом. Другой аспект новых исследований — изучение
того, что происходит со слуховой корой, пока мы спим.
3.1.
Первичная слуховая кора
Мы уже обсудили иерархическую модель обработки слуховой информации, ко-
торая была разработана в экспериментах на животных. Суть этой модели сво-
дится к тому, что нейроны первичной слуховой коры настроены на извлечение
физических характеристик звука, нейроны более высоких областей слуховой
коры — на извлечение более сложных, комплексных характеристик. Недавние
324
Глава 7
эксперименты на людях говорят о том, что иерархическая организация слухо-
вой функции совпадает с таковой у животных. Кодирование основных характе-
ристик звука связано у человека с первичной слуховой корой, более сложная
информация кодируется в височной области (planum temp�rale) (Wessinger et
al., 2001). Однако исследования в этой области находятся пока на ранней ста-
дии, поэтому к полученным результатам необходимо относится с осторожно-
стью. В целом можно сказать, что функциональная роль первичной слуховой
коры пока не изучена до конца. Вероятно, что функции, которые выполняются
слуховой корой, могут отличаться у человека и животных.
3.2 .
Роль височной области в декодировании
слухового сигнала
Анатомические исследования показывают, что строение слуховой коры чело-
века асимметрично: височная область (planum temp�rale, PT) у праворуких ис-
PT) у праворуких ис-
) у праворуких ис-
пытуемых больше в левом полушарии (Geschwind and Levitsky, 1968). Близость
РТ с зоной Вернике заставило выдвинуть гипотезу, что эта область связана
с восприятием речи и обработкой языковой информации. Это предположение
было подтверждено исследованиями с применением методов нейровизуализа-
ции, в которых изучалась роль РТ при восприятии речи. Однако анализ актива-
ции этой области при предъявлении разных типов речевых и неречевых звуков
заставляет сделать вывод, что функция РТ не ограничена обработкой речевой
информации.
Рис. 7.15. Какую роль играет PT в восприятии звука и речи? Ученые, работающие в
области нейронаук, до сих пор не могут точно ответить на этот вопрос. Недавно опубли-
кованный обзор функциональной нейроанатомии языка (Hickok, 2009) показывает, что
область, которую мы называем PT, скорее всего, не является единой с функциональной
точки зрения. Если проанализировать цитоархитектоническую организацию РТ, можно
увидеть, что в ее анатомических границах выделяется 4 отчетливо различающихся поля
(выделены зеленым, красным, желтым и синим). Скорее всего, эти поля связаны с раз-
ными функциями, которые осуществляются при участии РТ. Возможно, именно этим
объясняется большое количество нейровизуализационных данных об активации РТ
при разных типах заданий и предъявлении разных типов стимулов. (Источник: Hickok,
2009.)
Слух и речь
325
Эти данные привели к изменению взглядов на то, какую роль играет РТ в
восприятии звука.
В недавнем обзоре Хикок с коллегами (2004) указывают на то, что наличие
ответа РТ на такое количество разнообразных акустических стимулов не долж-
но удивлять, если рассматривать эту область с точки зрения ее цитоархитек-
тонической организации. На рис. 7 .15 видно, что внутри РТ можно выделить
четыре различающихся поля. Разные поля, вероятно, связаны с различными
функциями, которые осуществляются в этой области. Таким образом, можно
объяснить данные нейровизуализационных исследований, которые говорят об
активации РТ при предъявлении большого количества стимулов разных типов.
Для того чтобы уточнить роль разных областей слуховой коры, таких как РТ,
необходимо проделать еще много работы.
3.3 .
Корковые системы «что» и «где»
Основная функция слуховой системы заключается в том, чтобы вычленить из
слухового окружения конкретную информацию о том, что происходит вокруг
нас.
Давайте опять представим аудиторию в колледже, в которой слышно, как
открываются и закрываются двери, разговаривают студенты, расстегиваются
молнии рюкзаков и учебники со стуком кладутся на парты. Все эти звуки по-
являются одновременно, а их частоты и интенсивность пересекаются. Каким
образом слуховая система распознает отдельные слуховые «объекты», такие
как голос друга, стук двери или звонок телефона? Для того чтобы можно было
справиться с этой задачей, необходимо сохранять информацию о том, где в про-
странстве и когда во времени произошло событие. Из этих аспектов слуховой
информации формируется слуховая сцена восприятия для того, чтобы можно
было отличить разные слуховые объекты друг от друга. Конечно, такие перцеп-
тивные задачи лежат не перед слуховой системой в отдельности. При воспри-
ятии в одно целое объединяется информация о том, что мы видим, чувствуем и
ощущаем обонянием. Кроме того, такие задачи также требуют информации об
уже хранящихся в памяти слуховых объектах.
3.3.1 . Система «где» — локализация звука
Важным аспектом восприятия слуховой сцены является информация о том,
откуда идет звук. Представьте себе выдачу багажа в аэропорту, где вы ждете
друга, который вот-вот должен приехать. В помещении постоянно звучит речь,
часто появляются общие объявления по громкой связи: вся эта картина пред-
ставляет из себя сложную слуховую сцену! В конце концов, ваш друг находит
вас в толпе и кричит вам: «Я здесь!». Вам эта информация не очень помогает,
ведь вы не знаете, где это «здесь». Крик «Я немного справа и в 3 метрах позади
от тебя », выглядел бы гораздо более полезным. Несмотря на информационную
ограниченность фразы «Я здесь», вы, скорее всего, обернетесь и быстро уста-
новите с этим человеком визуальный контакт. В этом примере слуховая система
делает то, что умеет лучше всего: локализовывает звук в пространстве, даже
при наличии конкурирующих звуковых стимулов.
Локализация звука — это один из фундаментальных для слуховой системы
процессов. Знать, откуда идет звук, достаточно важно для расшифровки инфор-
326
Глава 7
мации о слуховой сцене. Но, кроме того, это крайне важно и для выживания,
потому что позволяет нам, например, отскочить от приближающейся машины,
когда мы слышим ее шум. Каким образом мозг локализует информацию в про-
странстве? Вы можете догадаться, что в некоторых отношениях это гораздо бо-
лее сложный процесс, чем локализация стимула зрительной системой (см. гл .
6). Таким его делает, например, то, что сами звуки изменяются во время своего
звучания.
Система локализации работает следующим образом: когда мы слышим звук,
его источник, скорее всего, находится по одну или другую сторону от вас. Кро-
ме того, он может находиться и позади. Для того чтобы понять, где находится
источник звука, слуховая система должна очень быстро сравнить сигналы, при-
шедшие к разным ушам. Она оценивает два основных показателя: первый —
это различия во времени прихода стимулов к разным ушам, второй — это раз-
личия в интенсивности стимулов. Различия в интенсивности возникают из-за
того, что когда звук идет по направлению к голове, она может создать так на-
зываемую «звуковую тень», и звук, который попадает в то ухо, которое рас-
положено дальше, оказывается несколько тише. Абсолютные различия между
интенсивностью звуков, которые поступают на разные уши, также зависят от
частоты звука (рис. 7 .16).
Таким образом, процессы локализации звука в основном связаны с тем, что
если источник звука находится слева от вас, то звуковая волна чуть раньше при-
ведет в движение барабанную перепонку левого уха, так же как интенсивность
звука слева окажется чуть больше, чем справа. Действительные вычислитель-
Рис. 7.16 . На рисунке схематично проиллюстрированы две характеристики, необходи-
мые для бинауральной локализации звука. Источник звука изображен в виде черной
точки справа от кошачьей головы. Концентрические линии представляют звуковые вол-
ны. (а) Интерауральные различия во времени прихода звука возникают из-за того, что
звуковым волнам требуется разное время, чтобы пройти расстояние от источника звука
до разных ушей. (б) Интерауральные различия в уровне громкости возникают из-за
«звуковой тени» от головы, которые снижают громкость звука, поступающего в более
далекое от источника ухо. (Источник: Brown and Santos-Sacchi in Squire et al., 2008.)
Слух и речь
327
ные механизмы, которые используются для того, чтобы определить положение
источника звука в пространстве, связаны с вычислением головных передаточ-
ных функций. Нейронные механизмы, которые лежат в основе этих вычислений
до конца неясны. Один из ключевых вопросов касается того, что при взросле-
нии размер головы человека существенным образом изменяется. Следователь-
но, нейронный код, связанный с вычислением передаточных функций, должен
каким-то образом учитывать возрастные изменения.
Недавние исследования показали, что эти вычисления протекают как в под-
корковых, так и в корковых слуховых областях мозга. Однако характер взаи-
модействия разных зон остается дискуссионным. Получены данные (Tiitinen
et al., 2006), которые говорят в пользу того, что вклад правого полушария в де-
кодирование информации о положении источника звука более значителен (рис.
7.17). Значит ли это,что правое полушарие специализируется на локализации?
Скорее всего, нет: в то время как для правого полушария более характерно об-
рабатывать информацию о положении источника звука, необходимо более под-
робно исследовать, как изменяющаяся во времени комплексная информация о
пространственных характеристиках звука декодируется в слуховой коре. Тем не
менее все исследователи согласны, что интерауральные (межушные) различия
во времени прихода звука и его интенсивности крайне малы (порядка долей
миллисекунд) и требуют системы, которая способна расшифровать крошечные
Рис. 7 .17. Пики P1m, N1m, N2m и P2m как индикаторы корковой активности, связан-
ной с обработкой информации о направлении источника звука. Реалистичные звуко-
вые стимулы предъявлялись с восьми пространственно эквивалентных направлений.
(а) Общие усредненные МЭГ ответы. Источники звука для каждого из восьми направ-
лений вызывали сложный тип ответа, содержащий пики P1m, N1m и P2m. Пики P1m,
N1m правого полушария имеют более короткий латентный период для звуков контра-
латерального полушария. Амплитуды P1m и N1m для каждого из полушарий и P2m
для правого полушария изменяются в зависимости от направления источника звука.
В целом звук с источником в контралатеральном направлении вызывали ответы с боль-
шей амплитудой, при этом амплитуды ответов в правом полушарии были больше, чем
в левом. (б) Усредненные минимальные текущие оценки (МТО), полученные на латент-
ном периоде пиков P1m, N1m, N2m и P2m, при предъявлении объемного звука с источ-
ником в направлении под 90°. Активация в окрестностях слуховой коры была проана-
лизирована на всех латентных периодах. (Источник: Tiitinen et al., 2006, с разрешения.)
328
Глава 7
различия между стимулами с удивительной точностью. Фактически система,
которая локализует источник звука, должна лежать в основе таких сложных си-
стем, как системы восприятия речи и музыки.
Система «где» и слуховое внимание
Несмотря на то что нейроны слуховой коры имеют входы от обоих ушей, ин-
формация в нейроны разных полушарий поступает асимметрично, преобладает
сигнал от контрлатеральной стороны тела. Сохраняется ли такой же эффект для
разных участков слухового пространства? Исследования (Hillyard et al., 1973)
дают основания полагать, что да. Используя методику потенциалов, связанных
с событием (ССП), он показал, что компонент N1 потенциала имеет большую
выраженность в зонах коры, расположенных контралатерально предъявляемым
стимулам (Hillyard et al., 1973). Последние работы с использованием методов
нейровизуализации, также направленные на изучение особенностей восприя-
тия слухового пространства, подтвердили эти результаты в рамках другой ме-
тодологии (Tzourio et al., 1997; Woldorff et al., 1999). С помощью методики ПЭТ
было показано, что слуховая кора правого полушария активируется сильнее
в случае, когда испытуемый должен обращать внимание на то, что находится
слева, а слуховая кора левого полушария — наоборот (рис. 7 .18) (Tzourio et al.,
1997). Эти результаты представляют из себя пример обработки информации
«сверху вниз»: экспериментаторы не просто предъявляли звуковые стимулы
справа или слева, а просили испытуемых сосредоточиться на том, что проис-
ходит в той или иной области. Таким образом, можно видеть то, какой вклад
оказывают процессы внимания на функционирование слуховой системы. Кро-
ме того, учитывая, что различия между полушариями могут быть связаны с
установкой внимания на разных областях слухового восприятия, эти данные
также заставляют более тщательно относиться к планированию эксперимен-
Рис. 7 .18 . Пример ПЭТ-изображения
активации, полученного в области
извилины Гешля (сверху) и верхней
височной извилины (снизу) у одного
испытуемого. В левой части рисунка
изображен ответ на отклоняющиеся
звуки слева, в правой части — от-
вет на отклоняющиеся звуки справа.
(Источник: Tzourio et al., 1997.)
Слух и речь
329
тальных процедур. К примеру, изучая роль разных полушарий при восприятии
инструментальной музыки и музыки с вокалом, важно обращать внимание на
то, чтобы испытуемые не концентрировались на информации, которая поступа-
ет только слева или только справа.
Роль тренировки в процессах слухового внимания
Несмотря на то что роль процессов внимания при обработке информации о
пространстве изучается на протяжении многих лет, мы до сих пор остаемся
в неведении относительно того, каким образом наш мозг справляется с теми
трудностями, которые перед ним возникают. Одна из недавних работ была на-
правлена на изучение того, как люди используют внимание для того, чтобы из-
влекать пространственные характеристики стимулов (Munte et al., 2001). Хотя
в этой области исследования находятся только на ранних стадиях, работа Мунте
с соавторами заслуживает упоминания. Они изучали, как воспринимают харак-
теристики пространства дирижеры, пианисты и контрольная группа людей без
музыкального образования. Гипотеза заключалась в том, что в связи с особен-
ностями работы дирижеров, когда необходимо воспринимать множество зву-
чащих инструментов одновременно, у них должно быть более развито слухо-
вое пространственное внимание. Было показано, что дирижеры лучше других
групп воспринимают звуки, которые предъявляются на периферии (рис. 7 .19).
Эти данные достаточно интересны, тем не менее необходимы дополнительные
исследования перед тем, как мы сможем сказать, какую роль индивидуальные
особенности играют в процессах слухового внимания.
3.3.2. Система «что» — распознавание звуковых объектов
и анализ слуховой сцены
Наши знания о звуках и о том, что они значат, начинают формироваться еще до
рождения (см. гл . 15) и продолжают на протяжении всей жизни, пока мы ана-
лизируем сложное слуховое окружение. Можно представить, что нейронные
процессы, которые участвуют в расшифровке информации о таком слуховом
окружении, как аудитория перед началом лекции, формируется не за одну ночь,
а на протяжении многих лет. В этом разделе мы обсудим процессы обучения и
формирования мысленных образов как слуховых объектов, так и сложноустро-
енного слухового окружения. Начнем мы с слуховых объектов.
Распознавание слуховых объектов
Знать, где находится источник звука, — это важный аспект анализа слуховой
сцены, который важен для выживания. Следующим шагом должно стать понима-
ние того, что ты слышишь. Для того чтобы справиться с восприятием звуковых
событый или звуковых объектов, слуховая система должна в реальном времени
декодировать информацию о звуках в том порядке, в котором они возникают.
Новые объекты появляются, когда мы растем, они изменяются в зависимости
от того опыта, который мы приобретаем. Так же как объекты зрительные, слу-
ховые объекты могут принимать разные формы. Они могут отличаться по слож-
ности, начиная с простых звуков и заканчивая человеческой речью и звучанием
симфонии. Кажется, что мозг обладает безграничным возможностями хранения
и воспроизведения слуховых объектов. Объекты организованы в определенные
категории, такие как, например, человеческие голоса, музыкальные инструмен-
330
Глава 7
Рис. 7.19 . Эффекты звукового внимания у дирижеров, пианистов и контрольной груп-
пы. (а) Экспериментальная установка; громкоговорители расположены в шести разных
направлениях. Изображены усредненные по группе ВП (фронтальное центральное от-
ведение) для группы дирижеров. Синей линией около каждого из громкоговорителей
изображен ответ на стимул от этого громкоговорителя при концентрации внимания на
громкоговоритель C1, красной линией — P1. Направление внимания на стимул связа-
но с усилением негативности с латентным периодом 60 мс. ВП, связанные с прилега-
ющими громкоговорителями, показывают уменьшающийся градиент. (б) Разностные
волны, полученные путем вычитания ответов с ненаправленным вниманием из ответов
с направленным вниманием. У всех испытуемых для центральных локализаций обнару-
жен градиент ВП, градиент для периферических звуков обнаружен только у дирижеров.
(в) Верхний ряд: электрофизиологические эффекты внимания (фронтальное центральное
отведение, средняя амплитуда, 180–220 мс; отношение C1/P1 установлено 100%). Для
центральных локализаций не обнаружено межгрупповых различий. Для группы дири-
жеров характерен более крутой градиент для периферических звуков. Нижний ряд: на-
жатие кнопки в ответ на предъявление малочастотного стимула. У дирижеров снижено
количество ложных тревог для звуков из соседних областей при периферической лока-
лизации стимулов. (г) Спин-интерполированные карты мозга для эффектов внимания
при центральном расположении стимулов (временное окно 180–220 мс): для всех групп
характерна одинаковая топографическая локализация. (Источник: Munte et al., 2001.)
Слух и речь
331
ты, звуки животных. Категоризация помогает декодировать информацию о но-
вых объектах и встраивать их в уже существующие схемы. С течением времени
между различным объектами начинают укрепляться ассоциативные связи, все
вместе они объединяются с информацией от других сенсорных систем. Так фор-
мируется общая концептуальная репрезентативная система.
Первой работой, в которой описывалось, как формируются и комбиниру-
ются концептуальные знания, была работа Вернике (Wernicke, 1874/1977). Он
говорил о том, что когда у человека уже есть достаточно опыта, если он слышит
звук колокольчика, у него возникают зрительные, тактильные и т. д. ощущения.
В связи с тем что способность говорить — это уникальное человеческое свой-
ство, неудивительно, что было предпринято много попыток изучить то, каким
образом мы воспринимаем звучащую речь. Эту тему мы обсудим позже. Сейчас
сосредоточимся на том, как декодируется информация о неречевых звуках, та-
ких как, например, звуки окружающей среды. На рис. 7 .20 видны результаты ра-
боты Биндера и его соавторов (Lewis et al., 2004), направленной на изучение вос-
приятия звуков окружающего мира. Основным выводом можно считать то, что
при восприятии как опознаваемых, так и неопознаваемых звуков привычного
окружения активируется область передней височной извилины. При восприятии
опознаваемых звуков при этом также активируется передняя височная борозда
и срединная височная извилина (СВИ) как правого, так и левого полушария.
Эти результаты интересны в связи с более ранними исследованиями, в которых
изучались функции СВИ. Активировавшиеся в подобных условиях области ча-
стично пересеклись с областями, связанными с семантическими процессами и
восприятием зрительных объектов как конкретных орудий.
Когда мы запоминаем звук из нашего привычного окружения, очень вероятно,
что мы воспринимаем его одновременно с каким-то видимым действием. Так,
стук молотка обычно воспринимается вместе со зрительным образом самого мо-
лотка. Биндер с соавторами (Lewis et al., 2004) предположили, что и в том, и в
другом случае СВИ активируется при распознавании конкретного объекта. Их
результаты хорошо согласуются с данным, полученными у пациентов с пораже-
ниями мозга, которые страдали слуховой агнозией, невозможностью распознать
слуховые объекты. При этом ухудшение распознавания только звуков окружаю-
щего мира возникало достаточно редко. Изучение таких редких случаев пока-
зало, что локализация поражений может оказываться очень сложной, включая
отдельно правое, отдельно левое или оба полушария одновременно (Clarke et al.,
2002). Данные показывают, что распознавание слуховых объектов включает раз-
нообразную активность в связанных между собой областях коры. Это доказы-
вает, что слуховое восприятие объектов окружающего мира включает сложные
механизмы, которые реализуются в разных областях мозга, в обоих полушариях.
Приведенная работа служит примером того, каким образом исследования, про-
веденные на больных с поражениями мозга дополняют результаты, полученные с
помощью методов нейровизуалиции.
Эффект вечеринки с коктейлем
Выше мы обсудили, каким образом звук воспринимается слуховой системой
и как он становится слуховым объектом, который необходимо опознать или вы-
учить заново. Такой механизм характерен для ситуации, когда одиночный звук
предъявляется в тишине. Каким образом перцептивная задача решается, если
Рис. 7 .20. Области мозга, вовлеченные в распознавания звуков окружающей среды. Жел-
тым цветом окрашены усредненные по группе активные области и темно-синим цветом
показано относительное уменьшение активности, вызванное (а) узнаваемыми, прямы-
ми звуками по сравнению с тишиной, или (б) соответствующие неузнаваемые обратные
звуки по сравнению с тишиной. (в) Вычитание данных (б) из (а) выявило области, пре-
имущественно вовлеченные в узнавание звуков (желтые) по сравнению с соответствую-
щими неузнаваемыми звуками, проигранными в обратном направлении (голубой); изо-
бражение получено относительно активности во время тишины. (г) Развернутые карты,
отображающие данные с (в). Левая верхняя височная борозда отмечена серым цветом
для четкости представления данных. (д) Аксиальные срезы данных с (в), наложенные на
изображение мозга одного испытуемого. (Источник: Lewis et al., 2004.)
Слух и речь
333
вокруг много посторонних шумов, источники которых, возможно, находятся в
одинаковых точках пространства? Каким образом слуховая система разделяет
эти шумы на отдельные слуховые объекты? Перцептивная задача такого рода по-
лучила название феномена «вечеринки с коктейлем» (Cherry, 1953). Этот фено-
Cherry, 1953). Этот фено-
, 1953). Этот фено-
мен связан с большим количеством исследований. С теоретической точки зрения,
интерес в данном случае представляет то, как слуховая система вычленяет из
сложного звукового окружения отдельные сигналы, с практической — эти иссле-
дования перспективны для разработки систем распознавания речи.
Брегман (Bregman, 1990) разработал модель, которая позволяет описать, как
происходит разделение различных сигналов в зашумленном окружении. Мо-
дель включает четыре элемента.
1. Источник.
2. Поток.
3. Группировка.
4. Разделение потока.
Источник — это сам звуковой сигнал. Поток — перцептивный образ, ко-
торый связан со звуком. Такое различие между физической составляющей сиг-
нала и его восприятием схож с различием между частотой звука и высотой,
которую воспринимает слушающий. Источник описывается в терминах физи-
ческих характеристик, а поток отражает психологические аспекты восприятия,
которые могут быть различны для разных людей.
Группировка относится к тому, как комбинируется сигнал для идентифика-
ции и удержания внимания к определенным аспектам слуховой сцены (таким
как выделения знакомого голоса в толпе). Процессы перцептивной группи-
ровки создают поток. Существует два типа группировок: первый тип — си-
мультанная (одновременная) группировка, когда вместе группируются два или
более стимула, которые начинают звучать одновременно. Представьте себе ор-
кестр или хор: вы скорее будете воспринимать отдельный звуковой поток, а не
разделять его на отдельные голоса или инструменты. (Но, если вы осознанно
направите на них свое внимание, вы сможете их услышать. Это отличный при-
мер того, какие возможности имеет слуховая система!) Второй тип — последо-
вательная группировка, когда вместе группируются стимулы, которые в разные
промежутки времени имеют одни и те же характеристики. Примером такого
рода процессов может служить ситуация, когда вы слушаете лекцию, и вдруг
перед вами кто-то начинает кашлять. Звуковой поток, который идет от лектора,
прерывается кашлем, однако это вряд ли окажет воздействие на ваше восприя-
тие того, что говорится. Слуховая система распознает голос лектора как отдель-
ный поток, несмотря на короткую помеху от кашля.
Разделение потоков — это процесс выделения отдельных слуховых объек-
тов или событий, их разделение на разные потоки.
Ниже представлены основные принципы группировки, разработанные в
рамках модели Брегмана.
1. Близость: звуки, которые воспринимаются близко по времени и имеют об-
щие качества и характеристики, могут быть сгруппированы вместе.
2. Родство: звуки, которые принадлежат одному и тому же объекту, группи-
руются вместе. В примере с лекцией — кашель не «принадлежит» потоку,
который идет от лектора.
334
Глава 7
3. Хорошее продолжение: звуки, переход между которым оказывается глад-
ким, скорее всего, группируются вместе (этот принцип схож с принципом
близости)
4. Общая судьба: вместе группируются звуки от одного источника или совпа-
дающие во времени (как в примере с оркестром).
5. Эксклюзивное распределение ресурсов: этот важный принцип для воспри-
Эксклюзивное распределение ресурсов: этот важный принцип для воспри-
ятия речи и состояний, когда необходимо распределять внимание на два
разных потока, которые невозможно воспринять одновременно. По другому
этот процесс иногда называют селективным слушанием. Когда чей-то го-
лос предъявляется в наушники, проблем в распознавании речи не возникает.
Однако если предъявить два разных речевых потока в разные уши, то вос-
принимается смысл только одного из них. При этом остается возможность
переключаться между разными потоками.
Каким образом мозг осуществляет анализ слуховой сцены? Исследования
нейронных механизмов перцептивной организации в слуховой системе приве-
ли к возникновению нескольких теорий того, где и как происходит декодиро-
вание информации о слуховом потоке. Одна из точек зрения предполагает, что
разделение разных потоков происходит при участии первичной слуховой коры,
а механизмы разделения включают подавление информации, не относящейся
к определенному потоку (Fishman et al., 2001). Вторая точка зрения состоит
в том, что в этом процессе участвует корковый механизм детекции измене-
ний, который отслеживает изменения, не относящиеся к одиночному потоку
(Sussman, 2005). Согласно этой точке зрения, индивидуальный слуховой по-
Sussman, 2005). Согласно этой точке зрения, индивидуальный слуховой по-
, 2005). Согласно этой точке зрения, индивидуальный слуховой по-
ток формируется на основе акустических аспектов сигнала, таких как частота
и расположение в пространстве. Как только эти характеристики сформирова-
ны в нейронную модель потока, входные сигналы, которые с этими характе-
Рис. 7 .21 . Зоны коры, отвечающие
за анализ слуховых потоков: область
(выделена салатовым) в межтеменной
борозде (МТБ), где два слуховых по-
тока воспринимаются в противопо-
ложность одному. МТБ рассматрива-
ется в качестве области, отвечающей
за организацию (совмещение) чув-
ствительности к мультимодальной ин-
формации. (Источник: Cusack, 2005.)
Слух и речь
335
ристиками не совпадают, детектируются с помощью специальных корковых
механизмов. Третье представление о механизмах разделения потоков состоит
в том, что процессы перцептивной организации протекают в областях коры, в
которых происходит связывание зрительной и соматосенсорной информации,
а конкретно — во внутритеменной борозде (Cusack, 2005). Эта точка зрения
предполагает, что перцептивная организация сложных звуковых стимулов про-
текает вне слуховой коры, в мультимодальных ассоциативных областях коры
(рис. 7 .21)
Разделение слуховой сцены на составляющие части или потоки — это слож-
ная, комплексная проблема для слуховой системы, и, скорее всего, в решении
этой проблемы также участвуют комплексные мозговые механизмы. Таким об-
разом, неудивительно, что нейронный субстрат процессов анализа слуховой
сцены до сих пор остается сложной темой для анализа в рамках когнитивной
нейронауки.
3.3.3 . Потоки обработки информации «что» и «где»
Мы рассмотрели важные аспекты слухового восприятия, такие как декодирова-
ния бинауральных характеристик того, где звук локализуется в пространстве, и
каким образом анализируются временные и спектральные характеристики для
того, чтобы определить, что за звук мы слышим. Потоки информации «что»
и «где» активно изучались в большом количестве инвазивных исследований
на обезьянах для зрительной и для слуховой модальности. Если обобщить ре-
зультаты этих работ, можно сделать вывод, что существуют четко оформленные
и разделенные потоки информации о том, «что» за стимул воспринимается и
«где» это происходит. (рис. 7 .22)
Существует много данных, которые говорят о том, что кортикальные сети,
которые участвуют в процессе декодирования этих аспектов звуковой инфор-
мации, разделены (хотя и активно взаимодействуют между собой) и в человече-
ском мозге. Предполагается, что височная пластинка выступает как некоторый
узел связи между потоками «что» и «где» на ранних корковых этапах обработки
Рис. 7.22 . Слуховые обла-
сти и потоки в мозге при-
матов. (а) Латеральная по-
верхность мозга макаки,
выделены области зритель-
ной (розовым цветом) и
слуховой (синим) чувстви-
тельности. (б) Два широких
«потока» обработки в слу-
ховой системе. (Источник:
Scott, 2005.)
336
Глава 7
информации (Grif�ths and Warren, 2002). Тем не менее роль височной пластин-
Grif�ths and Warren, 2002). Тем не менее роль височной пластин-
and Warren, 2002). Тем не менее роль височной пластин-
and Warren, 2002). Тем не менее роль височной пластин-
Warren, 2002). Тем не менее роль височной пластин-
Warren, 2002). Тем не менее роль височной пластин-
, 2002). Тем не менее роль височной пластин-
ки остается не до конца выясненной.
Последние исследования процессов обработки «что» и «где» информации
с применением методов нейровизуализации показали, что при декодировании
разных типов информации можно выделить разные паттерны активации. Были
обобщены имеющиеся на данный момент результаты исследований и выдви-
нута гипотеза о том, какие области человеческого мозга связаны с «что», «где»
и «как» потоками (Scott, 2005) (рис. 7 .23). Однако картирование областей, ко-
Scott, 2005) (рис. 7 .23). Однако картирование областей, ко-
2005) (рис. 7 .23). Однако картирование областей, ко-
торые связаны с обработкой звуковой информации, до сих пор остается пред-
метом активного изучения.
4.0 .
Восприятие речи
Давайте теперь обратим свое внимание на такую важную часть функций слуха
человека, как декодирование речевых звуков. Так как язык — это уникальная
способность человека, вполне вероятно, что наша слуховая система может рез-
ко отличаться от слуховой системы приматов. Обсуждение способов распоз-
навания речи мы начнем с краткого обсуждения того, какие элементы анализа
можно в этой области выделить.
Рис. 7 .23 . Функциональные ответы на речь и возможные потоки обработки информа-
ции в мозге человека. (а) Латеральная поверхность человеческого мозга: раскрашенные
области указывают на какой тип акустического сигнала отвечает каждая часть височ-
ной области (соответствующие части теменной и лобной областей). Синие участки от-
вечают на специфичные, связанные с языком фонологические структуры. Сиреневые
участки отвечают на стимулы с фонетическими сигналами и характеристиками речи, в
то время как фиолетовые участки реагируют в ответ на разумную речь. Розовые участки
отвечают на вербальные, связанные с кратковременной памятью, и артикуляционные
проявления речи. Зеленые участки реагируют при возникновении пространственных
слуховых задач. (б) Предполагаемые направления потоков обработки «что», «где» и
«как» в человеческом мозге. Источник: Scott, 2005
Слух и речь
337
Основная задача речевой системы — из группы звуков получить некоторую
осмысленную информацию. Такой процесс представляется достаточно пря-
молинейным: когда мы слышим речевой звук, например «д», его физические
характеристики связываются с абстрактным представлением этого звука как
фонемы. Два основных типа фонем — это согласные (например, «д») и гласные
(например, «и»). В то время как произносится слово (допустим, слово «дом»),
отдельные фонемы сохраняются в эхоической памяти. Для того чтобы это сло-
во было декодировано, необходимо, чтобы была последовательно декодирована
информация о фонемах «д», «о» и «м», которая напрямую связана с физическим
звучанием слова. Результатом работы такой системы становится активация сло-
ва «дом» в концептуальной/семантической системе знаний. К сожалению, эта
весьма разумная схема не отражает того, как на самом деле работает система
распознавания речи. Фактически между исследователями в области восприятия
речи нет согласия даже относительно простейших «блоков» речи: действитель-
но ли одиночные фонемы являются элементами анализа для речевой системы?
Или на эту роль больше подходят слоги? Позже мы еще вернемся к поставлен-
ному вопросу. Сейчас же давайте обратим внимание на то, что речевая система
должна не только декодировать смысл того, что говорится, но и «кто» информа-
цию о говорящем, а также «когда» информацию для анализа порядка звучания
фонем, слогов, слов и предложений. Как уже отмечалось выше, анализ речи
идет в разных временных промежутках (20, 200, 2000 мс, см. рис. 7.2). Эта ин-
формация должна быть точно воспринята вне зависимости от индивиуальных
характеристик говорящего, таких как высокий ли это голос ребенка или низкий
голос мужчины, говорим ли громко или шепотом, быстро или медленно.
Очевидно, что речевая система не просто вычленяет смысл из звука, она не
может опираться только на физические характеристики речи, так эти характе-
ристики могут сильно варьировать для разных людей. Несмотря на сложность
процессов восприятия речи, они происходят без участия внимания или види-
мых усилий с нашей стороны. Давайте начнем наше обсуждение с истории
того, как развивались науки о речи.
4.1.
История вопроса
Исследования в области декодирования речевой информации ведутся сравни-
тельно недавно. Несмотря на то что связь языка и мозга давно стала предметом
изучения, а анализ физических характеристик звука ведется с начала XIX в.,
специфика того, каким образом из акустических сигналов рождается смысл, —
это относительно новая область (блок 7.1). Исследования восприятия речи на-
чались в период перед Второй мировой войной. Несколько событий выдвинули
эту тему на передовую науки. Во-первых, перед и во время войны существовала
необходимость разработки систем распознавания речи для передчи секретной,
скрытой военной информации. Первой такой системой стал вокодер (от англ.
v�ice c�der — кодировщик голоса), разработанный Дадли из Bell Tele�hone Lab-
Bell Tele�hone Lab-
Tele�hone Lab-
Tele�hone Lab-
Lab-
Lab-
oratories (Dudley, 1936). Вокодер предоставлял возможность передавать речевые
сигналы через телефонные узлы при помощи анализа и рекодирования речевых
сигналов в более простые сигналы, которые содержали меньше информации,
чем человеческая речь. На выходе получался гораздо более простой речевой
сигнал, который тем не менее был узнаваем. Это служило доказательством
338
Глава 7
того, что речь содержит множество избыточных характеристик. Полученные
результаты запустили серии экспериментов, целью которых было определить
минимальное количество информации, необходимой для понимания речи.
Следующим изобретением, которое было создано во время Второй мировой
войны, однако оставалось засекреченным до тех пор, пока война не кончилась,
был спектрограф. Он также был разработан специалистами из Bell Labs на не-
Bell Labs на не-
Labs на не-
Labs на не-
на не-
которых принципах, сформулированных Дадли во время разработки вокодера.
Спектрограф анализировал звуковой сигнал и выдавал изображение видимой
речи, которое называлось спектрограммой (рис.7 .25) (Potter et al., 1947).
В спектрограмме по оси ординат расположена частота сигнала, по оси аб-
сцисс — время. Затенение относится к третьему измерению, интенсивности
(или энергии) сигнала. Интенсивность представлена в двух видах — во вре-
мени (по оси х) и по частоте (по оси у). В такой ситуации количество энер-
гии (интенсивности) может быть представлено и как функция от длительности
предложения, и как функция от частотного диапазона внутри слогов или слов.
Белые области спектрограммы по оси х указывают, что в этой точке была тиши-
на. Белые области по оси у говорят о том, что интенсивность на данной частоте
равна нулю. Соответственно, чем темнее область по той же оси, тем больше
Блок 7.1 . От вокодера к «бионическому уху»
Технологии, которые лежат в основе ранних вокодеров, по-прежнему активно ис-
пользуются и составляют основу, которая позволяет слуховым имплантантам стиму-
лировать слуховую систему для коррекции слуха у людей с некоторыми конкретны-
ми типами дефектов слуха (рис. 7 .24).
Рис. 7 .24. Источник: Brown, 2003
Слух и речь
339
интенсивность в этом частотном диапазоне. На рис. 7 .25 можно видеть, что су-
ществуют темные области в достаточно узком диапазоне частот: эти области
называются формантами. Форманты располагаются на относительно постоян-
ных интервалах и связаны с потоками воздуха, идущего через голосовой тракт.
Форманты различаются у разных людей, что связано с размерами голосового
тракта. Спектрограф радикально изменил способы изучения восприятия речи
и дал совершенно новый метод изучения структурных элементов речевого сиг-
нала. Из ранних исследований с помощью спектрографа следует два основных
вывода: во-первых, промежутки или моменты тишины в предложениях не со-
впадают с границами слов, но часто оказываются внутри самих слов; во-вторых,
формантная структура фонем, таких как «д», сильно различается в зависимости
от того, какая за ними следует гласная (рис. 7 .26).
Эти данные оказали существенное влияние на то, на какие типы моделей
разделялись представления о декодировании речи. Во-первых, промежутки,
пустоты в речевом потоке не предоставляют речевой системе информацию о
начале и конце слов. Тогда ясно, что требуются другие указатели для опознания
разных слов. Во-вторых, физические характеристики отдельных фонем доста-
точно сильно различаются в зависимости от предшествующих и последующих
фонем. Отсутствие инвариантов указывает на то, что в мозге не может суще-
ствовать звуковых шаблонов, которые один к одному соотносятся с фонемами.
Несмотря на открывшуюся сложность декодирования информации, спек-
трограммы позволили исследователям описать важные характеристики фонем,
Рис. 7 .25. Спектрограмма индивидуально произнесенных слов. По оси х время, по оси
у частота. Более темные участки соответствуют большей амплитуде, в основном распо-
ложены в области формант каждого звука речи. (Источник: Binder et al., 2000.)
Рис. 7.26 . Схематическое направление и частота начала формант в слогах /ди/ и /ду/
демонстрируют, что, хотя восприятие двух слогов начинающихся со звука «д» будет
отображено в области одной и той же фонемы /д/, физические реализации начальной /д/
различаются. (Источник: Carroll, 1999, цитируется из Liberman, 1970.)
340
Глава 7
такие как место артикуляции (где в артикуляторной системе они производятся)
и длительность звучания голоса. Согласные могут быть категоризованы отно-
сительно этих отличительных характеристик (рис. 7.27).
4.2 .
Ранние теории восприятия речи
Важным выводом, который был на ранних этапах изучения речи, стало понима-
ние того, что физические характеристики сигнала или отдельные фонемы не со-
держат инвариантной информации для декодирования. Вспомните, что работы
Фурье и Ома предоставили основу для разложения сложных звуковых сигналов
на ряд простых синусоидальных составляющих. Данные об отсутствии инвари-
антов в речевых звуках указали на то, что система анализа речевой информации
должна отличаться от анализа других типов звуков.
Если восприятие речи не связано с разложением сигнала на более простые
части (а это так, потому что физические характеристики фонем могут сильно
различаться), то каким образом протекает этот процесс? Одна из теорий пред-
полагает, что нейронная система декодирования речи специализирована и не
является частью общей слуховой системы. В строгой формулировке эта теория
гласит, что речевые звуки воспринимаются специальной системой уже на уров-
не уха (Liberman et al., 1967). Отсутствие инвариантов в речи привело Либер-
мана и его соавторов к разработке моторной теории восприятия речи (Liber-
Liber-
man and Mattingly, 1985). Согласно этой теории, восприятие речи тесно связано
с порождением речи, особенно с процессами артикуляции. В то время как для
фонем не удается выделить инвариатные характеристики, моторная теория гла-
сит, что артикуляторные процессы обладают такими инвариантами и при вос-
приятии речи есть доступ к нейронным механизмам ее порождения.
Рис. 7 .27. Схематическая иллюстрация образцов формант, имеющих отличительные
особенности различных классов звуков речи. (Источник: Brown, 2003.)
Слух и речь
341
С того момента, как была сформулирована теория моторного восприятия,
появились и новые представления о том, как происходит восприятие речи. Об -
щим для всех теоретических взглядов было представление о том, что речевая
система должна иметь возможность сохранять константность восприятия вне
зависимости от широкого спектра физических характеристик фонем и слов. Ос-
новные дискуссии в этой области науки разразились между 1950-ми и 1960-ми,
после выхода работы Либермана, в которой постулировалась специфичность
речи и существование отдельной речевой системы. Альтернативой этой точке
зрения было представление о том, что язык и речь опираются на общие позна-
вательные системы мозга. Работы Tallal показали, что латерализованная об-
Tallal показали, что латерализованная об-
показали, что латерализованная об-
работка речевой информации связана скорее со спецификой работы слуховой
системы в левом полушарии (обработка быстро изменяющихся характеристик
звука, свойственных, в том числе, звукам речи), а не с выделением отдельной
речевой системы (Tallal, 2001).
Разделение на две подобные противоречащие друг другу точки зрения суще-
ствует до сих пор, однозначного ответа в пользу того или иного взгляда пока не
существует. Тем не менее наиболее вероятной представляется ситуация, когда
в восприятии речи участвует как специфическая речевая система, так и более
общие познавательные механизмы.
4.2.1 . Элементы анализа — с оставные части речевой системы
Что служит базовыми элементами для системы распознавания речи? Фонемы?
Слоги? Данные об отсутствии инвариантных характеристик у фонем служат
серьезным доводом против представления о фонемах как о таких базовых эле-
ментах, хотя они и являются критически важными для восприятия речи. Од-
нако в звуках речи содержится большое количество информации, так что не
удивительно, что прямое сопоставление фонем и слов не позволяет полностью
описать процессы распознавания речи (Scott and Wise, 2004). Обратите внима-
Scott and Wise, 2004). Обратите внима-
and Wise, 2004). Обратите внима-
and Wise, 2004). Обратите внима-
Wise, 2004). Обратите внима-
Wise, 2004). Обратите внима-
, 2004). Обратите внима-
ние, что слуховая система имеет сложные обратные связи на всем протяжении
обработки информации от улитки до коры, существуют параллельные связи
внутри слуховой коры, между слуховой системой правого и левого полушария,
а также связи с другими сенсорными и моторными системами. Такая комплекс-
ная обработка информации, естественно, помогает при декодировании сигнала.
Слог интуитивно представляется адекватным в качестве основного элемен-
та речи, в том числе из-за важности силлабического ударения для понимания
смысла. К примеру, в словах мелодия и мелодический набор фонем кажется до-
статочно схожим. Основной характеристикой, которая помогает в различении
этих двух слов, является различие в ударениях. Несмотря ни на что, у исследо-
вателей не существует единой точки зрения на базовые элементы, необходимые
для понимания речи.
4.2.2. Минимальное количество информации,
которое необходимо для декодирования слухового сигнала
Если речевые сигналы оказываются настолько сложными, возможно, более про-
дуктивным для понимания процессов декодирования окажется исследовать ми-
нимальное количество информации, которая необходима для понимания речи?
Такой подход использовали Шеннон с соавторами (Shannon et al., 1998). Основ-
342
Глава 7
ное предположение заключалось в том, что минимальная информация, которая
необходима для распознавания речевого сигнала содержится в его форме, т. е.
в подъемах и падениях или временной огибающей сигнала. Shannon предъявлял
речевые звуки, в которых была устранена информация о частотных составля-
ющих и варьировались временные характеристики сигнала. Испытуемые с по-
разительной точностью распознавали речь. Эти данные указывают на то, что
хотя частотная составляющая сигнала безусловно важна для декодирования
речевых стимулов, ее наличие не обязательно. Было показано, что временная
огибающая речи может нести больше информации, необходимой для распозна-
вания, чем более тонкие характеристики, связанные с фонемами.
Тот факт, что исследователи не могут прийти к согласию относительно ос-
новных элементов речи, делает исследования мозговых механизмов речевого
восприятия крайне непростой. Однако существует много работ, которые на-
правлены на изучения того как в мозге происходит анализ речевых и некоторых
других типов звуков. Мы рассмотрим часть из них в следующем разделе.
4.3 .
Функциональное картирование
специфических речевых процессов
В работе Биндера (Binder, 1997) с применением методов нейровизуализации
были изучены различия в активации слуховой коры, связанные с восприятием
речевых сигналов (слов), тонов или шума. Было показано, что при восприя-
тии слов сильнее активируется верхняя теменная извилина и верхняя темен-
ная борозда (superi�r temp�ral gyrus и superi�r temp�ral sulcus) по сравнению
с восприятием тонов или шума. Хотя эти результаты можно трактовать как до-
казательство в пользу существования в мозге специфической речевой системы,
в этом исследовании остаются неучтенные факторы. Тоны и шум отличаются
от речевых звуков не только тем, что относятся к языку или нет. Они также от-
личаются по сложности своего устройства. Таким образом, разные паттерны
активации мозга могут быть связаны не только с наличием функциональных ре-
чевых зон, но и с обработкой комплексных характеристик звуковых стимулов.
Рис. 7 .28 . Сравнение трех вариантов различий Речь–Тон: Речь–Тон (Р–Т), Псевдоречь–
Тон (П–Т) и Инвертированная речь–Тон (И–Т). Области, отвечающие больше на Тоны,
очень похожи во всех случаях. (Источник: Binder et al., 2000.)
Слух и речь
343
В одном из исследований (Binder et al., 2000) изучалась активация слухо-
вых корковых областей, связанных с восприятием речевых и неречевых звуков
(для этого предъявлялся широкий набор звуковых стимулов: шумов, тонов и
т. д .), а также различий в активации при предъявлении слов, псевдослов (таких
как англ. «hig», не имеющих смысловой нагрузки), а также инвертированной
речи. (рис. 7 .28). В результате было показано, что извилина Гешля и область
височной пластинки одинаково активируются при предъявлении всех типов
звуковых стимулов. Эти данные служат в пользу предположения, что обработ-
ка звука протекает иерархично, при этом извилина Гешля активируется при
восприятии всех типов сигналов и, скорее всего, связана с ранними этапами
сенсорного анализа. При восприятии речевых звуков по сравнению с нерече-
выми активируются более широкие области слуховой коры, включая верхнюю
заднюю височную извилину и верхнюю теменную борозду. Паттерн активации
одинаков как при анализе слов, так и псевдослов и инвертированной речи.
Эти результаты указывают на то, что при восприятии речи активируются бо-
лее широкие сети слуховой коры, чем при восприятии шумов и тонов. В связи
с тем, что различия в активации для слов, псеводослов и инвертированной речи
не были выявлены, Биндер делает вывод, что эти области, скорее, связаны не с
семантическими процессами, а с процессом обработки фонологической инфор-
мации о речевых стимулах.
На данный момент можно выделить много линий исследований в области
нейронных механизмов декодирования речи, ведется активная работа в этом
направлении. Давайте обратим внимание на связь процессов «слушания» и «го-
ворения», как взаимодействуют процессы восприятия и формирования речи.
4.4 .
Связь восприятия и формирования речи
В ранних работах с применением методов нейровизуализации изучалась акти-
вация мозга во время формирования речи. Выяснилось, что и в этом случае
активируется слуховая кора. Эти данные были многократно воспроизведены.
(рис. 7 .29)
Почему слуховая кора оказывается активной во время порождения речи?
Может, дело в том, что, когда мы говорим, мы одновременно слышим собствен-
ный голос? Или слуховая система играет при этом особую роль?
Результаты нейровизализации, так же как данные, полученные на больных
афазией с поражениями мозга говорят о том, что процессы восприятия и по-
Рис. 7 .29 . Красные области активируются во время речи, желтые во время слушания
речи. (Источник: Frackowiak, 2004.)
344
Глава 7
рождения речи тесно связаны. С раннего детства, когда человек овладевает язы-
ком, существует взаимодействие между слышимой и произносимой речью. Это
взаимодействие во многом определяет развитие языковых навыков. Карл Вер-
нике в конце XIX в. предложил модель, которая включала связь перцептивных
речевых областей мозга с моторными. Он предположил, что «звуковой образ»
слова необходим для коррекции произносимых слов. Эта модель используется
по сей день. Она содержит важные теоретические предпосылки к изучению
речевой системы даже сейчас, несмотря на то, что наши знания о протекающих
в мозге процессах существенно шире, чем во времена Вернике.
4.4.1 . Внутренняя речь
Какие процессы протекают в мозге, когда мы разговариваем сами с собой? Ана-
лиз мозговых механизмов внутренней речи раскрывает перед исследователями
возможность исследовать сознание человека. Предложена упрощенная модель
мозговой активности, связанной с внутренней речью (Hesslow, 2002) (рис. 7 .30).
Взаимосвязь между процессами декодирования речевой информации и по-
рождения речи сначала была предсказана теоретически в моторных теориях
речевого восприятия. Наличие этой связи было доказано экспериментально и
на сегодняшний день признается всеми. Однако точно природа интегративных
процессов и мозговых областей, которые с ними связаны, до сих пор неясна.
В этой области требуются дополнительные исследования.
4.5 .
Повреждения систем восприятия речи
До изобретения методов нейровизуализации основным источником информа-
ции о системах мозга, которые связаны с декодированием речи, было изучение
больных с поражениями мозга. Чаще всего это были больные, перенесшие ин-
сульт, так как в этом случае повреждения более локальны, чем при травмах
головы. Тем не менее при инсульте может быть затронут очень разный объем
мозга. В результате инсульта происходит блокада кровяного потока, которая вы-
зывает массовую гибель нейронов в определенной области мозга. Повреждения
специфической области вызывают поведенческие нарушения. Когда поврежде-
ния касаются речевых зон, у человека развивается афазия. Симптомы афазии
могут быть разнообразны в зависимости от локализации и размера затронутых
при инсульте областей мозга (блок 7.2). Две основные классификации афазий
связаны с нейроанатомическими исследованиями XIX в. Поль Брока обнару-
XIX в. Поль Брока обнару-
в. Поль Брока обнару-
жил зону во фронтальных отделах мозга, которая связана с порождением речи,
Карл Вернике — зону в височной доле, связанную с восприятием речи. По-
вреждения этих зон вызывают два разных типа афазий: когда в качестве основ-
ного симптома выступают проблемы с порождением речи, говорят, что больной
страдает от афазии Брока, когда нарушается понимание речи — афазии Верни-
ке. (рис. 7 .31).
Блюмштайн (Blumstein, 1997) пытался с помощью тестирования определить,
какие аспекты понимания речи нарушены при афазии. В одной из серий экспе-
риментов проверялась гипотеза о том, связаны ли проблемы с пониманием речи
с перцептивными процессами (нарушено восприятие абстрактных фонем, фо-
нематический дефицит) или страдают процессы распознавания слов и переход
от фонем к смысловым характеристикам стимулов (семантический дефицит).
Слух и речь
345
Рис. 7.30 . Внутренняя симуляция раз-
говора. (а) Можно отвечать на вопрос
бессознательно. Вербальный сигнал
поступает в слуховую кору, а затем в
зону Вернике (В). Это вызывает фор-
мирование ответа в зоне Брока (Б)
и первичной моторной коре (М). (б)
Можно слушать и отвечать на соб-
ственную речь, используя те же самые
области мозга. (в) Если подготовка
вербального ответа может непосред-
ственно быть связана (красная стрел-
ка) с сигналами из слуховой коры или
зоны Вернике, мы также можем гово-
рить молча сами с собой, используя в
основном те же механизмы. (Источ-
ник: Hesslow, 2002.)
Рис. 7 .31 . Слева: классические области, связанные с языком, с изменениями из работ
нейроанатомов XIX в. Справа: современные взгляды на классические модели. (Источ-
ник: Frackowiak, 2004.)
346
Глава 7
Для того чтобы оценить нарушения при афазии, автором использовалась мето-
дика сопоставления слов и картинок. Суть этой методики заключается в том,
что человеку на слух предъявляется слово, и он должен подобрать к этому сло-
ву картинку. Для каждого слова испытуемый мог выбрать одну из четырех кар-
тинок: один правильный вариант и три ошибочных. Среди неподходящих кар-
тинок одна изображала предмет, название которого совпадало с предъявленным
словом по звучанию, на другой был предмет с похожим по смыслу названием,
третий предмет не подходил ни по смыслу, ни по звучанию. На рис. 7.33 пред-
ставлен этот эксперимент: на левой панели изображены ключи («keys») и бобы
(«�eas»). На правой панели также изображена морковь и калькулятор. Испы-
�eas»). На правой панели также изображена морковь и калькулятор. Испы-
»). На правой панели также изображена морковь и калькулятор. Испы-
туемому на слух предъявляется слово «�eas» (бобы). В этом случае картинка
с ключами — это пример ошибки по звучанию, морковь — семантической
ошибки, калькулятор — никак не связанного со стимулом предмета. Характер
ошибок, сделанных пациентами, позволяет проанализировать особенности на-
рушений в процессах декодирования информации. Если ошибки в основном
связаны со звуковым сходством, можно сделать вывод, что нарушены процессы
анализа акустических характеристик речевых звуков. Если пациенты делают в
основном семантические ошибки, значит анализ акустических характеристик
сохранен, а трудности в распознавании речи связаны с декодированием смысла
предложений.
В результате проведенных исследований было сделано два вывода. Во-
первых, характер ошибок, которые делают пациенты, существенным образом
Блок 7.2 . Новый взгляд на классическую модель:
реконструкция языковых путей
Недавние открытия в области нейровизуализации позволили найти новые способы
рассмотреть работу мозга. Одной из главных нервных «магистралей» мозга, долгое
время привлекающей интерес исследователей языка, является дугообразный пучок.
Этот большой тракт соединяет анатомические области, функционально соотнося-
щиеся с зонами Брока и Вернике в левом полушарии. Разрушение этого тракта, как
считается, лежит в основе одного из типов приобретенного слухового поражения
(афазии), называемого проводниковой афазией. Однако после тщательного обсле-
дования больных с диагнозом проводниковая афазия был выявлен широкий спектр
симптомов, поднимающий вопрос о том, одно ли и то же повреждение мозга вызыва-
ет эти симптомы. Исследователи Катани с коллегами использовали метод диффузной
тензорной трактографии, чтобы реконструировать трехмерную картину проводящих
путей, соединяющих зоны Брока и Вернике (Catani et al., 2005). Они нашли доказа-
тельства того, что волокна белого вещества, называемые дугообразным пучком, на-
прямую соединяют зоны Брока и Вернике, как показано красным на рис. 7 .32. Тем
не менее были также обнаружены непрямые пути, которые идут латерально по на-
правлению к переднему сегменту (показано зеленым на рис. 7 .32) и соединяют зону
Брока с участком нижней теменной коры около извилины Гешвинда. Задний сегмент
соединяет область около извилины Гешвинда с зоной Вернике. Несмотря на то, что
эти новые открытия ожидают подтверждения, они уже могут говорить о том, почему
у пациентов с проводниковой афазией так сильно варьируют симптомы: если прямые
и непрямые пути параллельно соединяют зоны Брока и Вернике, тогда нарушения
речи будут сильно различаться в зависимости от того, в каком именно месте прово-
дящего пути возникло повреждение.
Слух и речь
347
Рис. 7 .32 . Внизу: проводящие пути, соединяющие функциональные участки зоны
Вернике в височной доле и зоны Брока в лобной доле в левом полушарии. Пути
реконструированы с использованием методов трактографии и МРТ. Прямой путь
между зонами Брока и Вернике, показанный красным, соответствует классическому
описанию дугообразного пучка, который долгое время считался главной «магистра-
лью», соединяющей эти важные области, связанные с языком. Обнаружены также
непрямые пути: передний сегмент соединяет зону Брока с областью в нижней те-
менной доле рядом с извилиной Гешвинда, а задний сегмент соединяет эту область
с зоной Вернике (показано желтым). Вверху: показана связь этих путей с анатоми-
ческими структурами левого полушария. (Источник: Catani et al., 2005, с измене-
ниями с разрешения.)
348
Глава 7
не связан с тем, какой тип афазии у них диагностирован. Можно предположить,
что больные с афазией Вернике должны делать больше фонематических оши-
бок, чем больные с афазией Брока. Однако полученные результаты не соответ-
ствуют этой гипотезе. Во-вторых, больные как с афазией Брока, так и с афа-
зией Вернике в принципе достаточно хорошо справлялись с заданием такого
рода. Следовательно, речевое восприятие, способность связывать акустические
характеристики звука со смыслом слов при афазии нарушено не так сильно.
Таким образом, сложности при понимании речи не связаны со способностью
анализировать звуковые сигналы.
Эта и другие работы в области изучения мозговых механизмов речевого вос-
приятия говорят о том, что хотя языковые функции у праворуких людей ло-
кализованы в левом полушарии, перцептивные процессы, связанные с речью
могут быть организованы билатерально, т. е. при участии обоих полушарий.
Последующие исследования в этой области предоставили данные в пользу та-
кого вывода: грубые нарушения восприятия речи («полная глухота на слова»)
проявляются только в том случае, когда повреждения затрагивают оба полуша-
рии или повреждения в левом полушарии затрагивают также мозолистое тело,
и информация не может быть передана в правое полушарие (Poe��el, 2001).
Рис. 7.33. Слева: минимальные пары слов, у которых начальные согласные отличают-
ся только по одному признаку. В таком случае начальные /k/ и /�/ отличаются областью
артикуляции (задненебный против губно-губного), но сопоставимы генерацией звука
голосовыми связками (беззвучно) и манерой (взрывной согласный). Справа: стимулы,
используемые в одиночном слуховом испытании называния картинки словом. Испы-
туемый слышит слово, в данном случае «�eas», и должен указать на картинку, соот-
ветствующую слову. Сверху сл ева: семантический профиль, сверху справа: независи-
мый профиль, снизу сл ева: корректный (целевой) ответ, снизу справа: фонематический
профиль
Слух и речь
349
4.6 .
Рабочая модель мозговых механизмов восприятия речи
Языковые функции у большинства праворуких людей локализованы в левом
полушарии. Однако речевая система не однородна, она включает разные этапы,
от обработки информации о речевых звуках при восприятии до взаимодействия
семантических и грамматических систем при порождении речи. Разработан-
ная недавно модель слухоречевой системы предполагает, что на ранних этапах
речевые процессы организованы билатерально в слуховых областях правого и
левого полушария, в то время как более поздние семантические и синтакси-
ческие процессы связаны только с левым полушарием (рис. 7 .34) (Hickok and
Poe��el, 2007).
Рис. 7.34 . (a) Изображена схематическая модель обработки слухоречевой информации
Hickok и Poe��el (2007). (б) Показаны области мозга, которые должны отражать разные
этапы обработки информации, характерные для модели. Обратите внимание, что ран-
ние перцептивные системы (выделены зеленым и желтым), как предполагается, лока-
лизованы билатерально, в обоих полушариях, более поздние процессы связаны только с
работой областей левого полушария. (Источник: Hickok & Poe��el, 2007.)
350
Глава 7
Хотя благодаря методам нейровизуализации и изучению больных с пора-
жениями мозга на сегодняшний день имеется достаточно много данных о том,
как организованы речевые процессы в мозге, наше понимание неполно. Это
не кажется таким удивительным, так как исследователи до сих пор не могут
договориться о базовых элементах анализа системы восприятия речи. Новые
методы, например транскраниальная магнитная стимуляция (ТМС), а также ин-
новационные экспериментальные процедуры предоставляют новые данные о
процессах декодирования информации и локализации речевой системы в мозге.
Музыка и мозг
Музыка, как и язык, является достоянием любой че-
ловеческой культуры. Ее появление уходит корнями
глубоко в прошлое. Самые старые из найденных му-
зыкальных инструментов, флейты из костей птиц и
бивней мамонтов, были сделаны более 35 000 лет
назад (Conard et al., в печати). Несмотря на то что
сила влияния музыки на человека поражала мысли-
телей со времен Платона, связь музыки и мозга на-
чала активно изучаться недавно (Peretz & Zatorre,
2003).
Несколько фактов на сегодняшний день уже очевид-
ны. Во-первых, изучение пациентов с поражениями
мозга с нарушенным слухом показало, что воспри-
ятие музыки — это высокоорганизованная когни-
тивная структура с большим количеством разных
компонентов (Stewart et al., 2006). Во-вторых, иссле-
дования с применением нейровизуализационных ме-
тодов на нормальных испытуемых дало понять, что
при прослушивании музыки задействовано большое
количество разных зон мозга (связанных не только
со слухом) даже у людей без музыкального образова-
ния (Koelsch et al., 2005). В -третьих, обучение игре
на музыкальных инструментах влияет на мозговые
структуры и их функции как на корковом, так и на подкорковом уровне и повыша-
ет немузыкальные когнитивные способности (Schellenberg, 2005; Wong et al. 2007;
Hyde et al., 2009). В -четвертых, восприятие музыки показывает тесную взаимосвязь
между когнитивными и эмоциональными процессами (Huron, 2006; Steinbeis et al.,
2006).
Как и в других областях науки, в исследованиях связи музыки и мозга много про-
тиворечий и споров. Я участвовал в одном из таких споров, касавшихся проблемы
«модульности», т. е. связи восприятия музыки со специально отведенным участком
мозга или задействования других когнитивных систем, таких как языковая, напри-
мер (Peretz, 2006). Музыка, конечно, связана с определенным уровнем мозговой
специализации, но работы некоторых моих коллег и мои собственные указывают
на то, что обработка музыкальной и речевой информации имеет множество пере-
сечений (Patel, 2008). Эти данные могут иметь значение для ряда проблем, включая,
например, изучение и разработку методов лечения речевых нарушений (Patel et al.,
2008; Schlaug et al., 2008).
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
Рис. 7 .35. Амируд Д. Па-
тель (Aniruddh D. Patel,
PhD), Институт нейронаук,
Сан-Диего, США. (Источ-
ник: Tom Cogill.)
Слух и речь
351
Другой важный спор касается эволюции. Появилась ли музыка как биологическая
адаптация, которая помогла нашим предкам выжить? Или это человеческое изо-
бретение, которое основано на мозговых механизмах, связанных с другими функ-
циями? Эти вопросы могут легко увести к спекуляциям на тему, однако в последнее
время наметились тенденции решать их лабораторно, изучая восприятие музыки у
разных видов животных. В моей лаборатории, например, было показано, что живот-
ные могут двигаться под музыкальной ритм (Patel et al., 2009; см. рис. 7 .36). Таким
образом, как минимум один фундаментальный аспект музыкального восприятия не
требует специально отобранных в эволюции мозговых механизмов.
Основной вопрос для будущих исследований: почему музыка оказывает такое силь-
ное эмоциональное воздействие на людей? Биологические основы эмоционального
восприятия музыки только начинают изучаться (Juslin & Västfjäll, 2008) и в буду-
щем нас ждут волнующие открытия в этой области.
Литература
1. Conard, N. J ., Malina, M., Münzel (in �ress). New flutes document the earliest musical
tradition in southwestern Germany. Nature .
2. Juslin, P. N ., & Västfjäll, D. (2008). Emotional res�onses to music: The need to
consider underlying mechanisms. The Behavi�ral and Brain Sciences, 31, 559–621.
3. Hauser, M. D., & McDermott, J. (2003). The evolution of the music faculty: A
com�arative �ers�ective. Nature Neur�science, 6, 663–668 .
4. Huron, D. (2006). Sweet Anticipati�n: Music and the Psych�l�gy �f Expectati�n.
Cambridge, MA: MIT Press.
5. Hyde, K. L., Lerch, J., Norton, A., Forgeard, M., Winner, E., Evans, A. C, & Schlaug,
G. (2009). Musical training sha�es structural brain develo�ment. The J�urnal �f
Neur�science, 29, 3019–3025.
6. Koelsch, S., Fritz, T., Schulze, K., Also�, D., & Schlaug, G. (2005). Adults and
children �rocessing music: An fMRI study. Neur�Image, 25, 1068–1976.
7. Patel, A. D . (2008). Music, Language, and the Brain. New York: Oxford Univ. Press.
8. Patel, A. D., Iversen, J. R ., Wassenaar, M., & Hagoort, P. (2008). Musical syntactic
�rocessing in agrammatic Broca’s a�hasia. Aphasi�l�gy, 22, 776–789.
9. Patel, A. D ., Iversen, J. R ., Bregman, M. R., & Schulz, I. (2009). Ex�erimental
evidence for synchronization to a musical beat in a nonhuman animal. Current
Bi�l�gy, 19, 827–830 .
10. Peretz, I. (2006). The nature of music from a biological �ers�ective. C�gniti�n, 100,
1–2 .
11. Peretz, I., & Zatorre, R. (2003). The C�gnitive Neur�science �f Music. New York:
Oxford Univ. Press.
Рис. 7 .36 . Снежок, желтохохлый какаду (Cacatua
galerita ele�n�ra), который синхронизировал свое
поведение с музыкальным ритмом. (Источник:
Irena Schulz.)
352
Глава 7
12. Schellenberg, E. G. (2005). Music and cognitive abilities. Current Directi�ns in
Psych�l�gical Science, 14, 317–320 .
13. Schlaug, G., Marchina, S., & Norton, A. (2008). From singing to s�eaking: Why
singing may lead to recovery of ex�ressive language function in �atients with Broca’s
a�hasia. Music Percepti�n, 25, 315–323 .
14. Steinbeis, N., Koelsch, S., & Sloboda, J. A . (2006). The role of harmonic ex�ectancy
violations in musical emotions: Evidence from subjective, �hysiological, and neural
res�onses. J�urnal �f C�gnitive Neur�science, 18, 1380–1393.
15. Stewart, L., von Kriegstein, K., Warren, J. D., & Grif�ths, T. D . (2006). Music and the
brain: Disorders of musical listening. Brain, 129, 2533–3553.
16. Wong, P. C. M., Skoe, E., Russo, N. M ., Dees, T., & Kraus, N. (2007). Musical
ex�erience sha�es human brainstem encoding of linguistic �itch �atterns. Nature
Neur�science, 10, 420–422 .
5.0 .
Восприятие музыки
Как и в случае с восприятием речи, для человека характерно и восприятие му-
зыки. Существует множество сходств в восприятии речи и музыки: музыка
имеет сложную фразовую структуру и ее восприятие предполагает преображе-
ние звуков в какое-либо значение (и эмоцию). В процессе восприятия музыки
возможно опознавание мелодии, даже если она сыграна другим инструментом,
в другой тональности или темпе; таким образом, данная система не является
оценивающей абсолютные высоты звука, но должна воспринимать относи-
тельные. В связи с вышесказанным, очевидно, что системы восприятия музы-
ки должны иметь возможность поддерживать перцептуальное постоянство в
отображении музыки. Основное различие между восприятием речи и музыки
состоит в том, что все нормально развивающиеся люди научаются восприни-
мать речь. Мы не только способны к восприятию речи, мы — мастера! Однако
это не справедливо для восприятия музыки: существует множество градаций
способности к восприятию музыки и значительно больше усилий необходимо
приложить для развития остроты восприятия музыки. Различия в способностях
к восприятию музыки вкупе со множеством степеней музыкальных навыков
весьма осложнили процесс исследования восприятия музыки в связи с унасле-
дованными индивидуальными различиями. Эти сложности, однако, дают воз-
можность понимания эффектов обучаемости и пластичности областей головно-
го мозга, декодирующих музыку.
5.1.
Этапы слуховой обработки музыки
Восприятие элементов музыки включает множество этапов обработки как в
пределах слуховой системы, так и между различными областями головного
мозга (рис. 7 .37) (Zatorre, Chen, and Penhune, 2007). Процесс обработки должен
включать в себя системы обратной и прямой связи, взаимодействие с памятью
и эмоциями. Восприятие музыки несколько отличается от восприятия речи тем,
что множество музыкальных звуковых сигналов не содержит никакого текста.
Таким образом, процесс восприятия музыки, вероятно, имеет более абстракт-
ную (нелингвистическую) репрезентативную основу.
Слух и речь
353
В то время как музыкальный сигнал, как и всякий другой звуковой сигнал,
является сложным, музыка в общем состоит из основных физических элемен-
тов: частоты, интенсивности и продолжительности. Физиологические аспек-
ты частоты и продолжительности музыки соответствуют высотной (мелодия)
и временной (ритм) структурам. Обычно мелодические и временные аспекты
музыки изучаются по отдельности, т. е . как различные аспекты восприятия му-
зыки. Однако, судя по всему, они не являются полностью независимыми. В то
время как ученые, исследующие речь, утверждают, что обработка речи в мозге
происходит в специальных областях, ответственных непосредственно за речь,
ученые, исследующие музыку, утверждают, что могут существовать системы
нейронов, специализирующихся на музыке. Факты, подтверждающие гипоте-
зу о музыкально-специализированных системах нейронов, обнаруживаются в
нейрофизиологических исследованиях с участием пациентов, страдающих по-
ражением мозга. Проведена серия исследований индивидуумов с поражением
головного мозга, показавших, что у некоторых из них восприятие мелодии мо-
жет быть нарушено частично, а система временного восприятия остается не-
поврежденной, в то время как у других система временного восприятия мо-
жет быть нарушена, а восприятие мелодии неповреждена. Данные открытия
привели к развитию модели организации системы музыкального восприятия в
головном мозге (Peretz and Zatorre, 2005), при которой мелодические элементы
музыки обрабатываются преимущественно в правом полушарии, и которая мо-
жет избирательно повреждаться при поражениях правого полушария головного
мозга, в то время как временная структура музыкальной ткани подвергается
декодированию в обоих полушариях.
5.2 .
Отдельная система для музыкального восприятия
Является ли восприятие музыки отдельным аспектом обработки звука? В то
время как работа Peretz и коллег предоставляет веские доводы в пользу это-
Peretz и коллег предоставляет веские доводы в пользу это-
и коллег предоставляет веские доводы в пользу это-
го утверждения, недавно опубликованный обзор (Koelsch, 2005) исследований
восприятия музыки с применением нейровизуализации представляет увели-
чивающуюся совокупность данных, говорящих в пользу того, что некоторые
аспекты музыкального восприятия, в особенности музыкальной структуры или
синтаксиса, музыкальных значений и семантики, функционируют совместно
в областях головного мозга, отвечающих за обработку речи (рис. 7 .38).
Хотя наблюдается значительное число подтверждений того, что восприятие
музыки может быть избирательно (Peretz и Zatorre, 2005) и дифференциально
нарушено при поражениях левого или правого полушария, обзор исследований
(Koelsch, 2005) приводит веский довод в пользу того, что по крайней мере не-
Koelsch, 2005) приводит веский довод в пользу того, что по крайней мере не-
2005) приводит веский довод в пользу того, что по крайней мере не-
которые системы обработки музыки и речи функционируют совместно. Речь и
музыка являются специфичными для человека и в значительной степени ком-
плексными сигналами со множеством измерений по спектральной и временной
осям для более полного понимания их основной и детализированной структу-
ры. Возможно, не существует единой области в головном мозге, отвечающей
как за речь, так и за музыку: весьма вероятно, что речь и музыка имеют неко-
торые области нейронов, специфичных для их обработки, но также и области
совместного функционирования. Усложнение уже сложных задач является от-
личительной особенностью музыкального образования и возможности оцени-
вания среди людей, музыкантов и немузыкантов.
354
Глава 7
Слух и речь
355
6.0.
Обучение и пластичность
Центральным направлением при изучении когнитивной деятельности человека
является исследование кодирования новой информации в мозге во время про-
цесса обучения, а также выяснение того, как мозг адаптируется и реорганизует-
ся в зависимости от новых ситуаций или в результате травм, т. е . пластичность.
Эти вопросы имеют важное теоретическое значение для понимания функци-
онирования мозга, а также практическую ценность. Одной из центральных
проблем является то, как слуховая кора отвечает на потерю сенсорного входа
вследствие потери слуха, нервных повреждений или глухоты. Например, если
ребенок рождается полностью глухим, и ему вставляют кохлеарный имплант
в возрасте двух лет, будет ли его слуховая кора чувствительна к звуку, и смо-
Рис. 7.38 . Верхняя часть рисунка отображает активные речевые области мозга. Нижний
рисунок иллюстрирует местоположение областей, ответственных за музыку. (Источ-
ник: Koelsch, 2005.)
Рис. 7 .37. Вверху: вы когда-нибудь задумывались, какие области мозга «зажигаются»,
когда вы слышите или играете музыку? Конечно, когда вы играете на каком-нибудь ин-
струменте, активируются слуховые зоны, однако также активируются моторные зоны,
которые связаны с извлечением звука. Эти моторные и сенсорные (слуховая) системы
тесно связаны нервными сетями, передающими обратную связь, а также управляющие
команды, необходимы для игры на инструменте. Внизу: результаты нейровизуализаци-
онных исследований показали тесную связь между моторными и сенсорными систе-
мами. (а) Люди без музыкального образования обучаются играть простую мелодию на
клавиатуре. Анализируется активация мозга при прослушивании мелодии до и после
обучения. (б) В других работах изучается активация мозга музыкантов при прослуши-
вании композиции, которую они умеют играть (слева), и когда они играют эту мелодию,
но не получают обратную связь (в ц ентре). Показаны существенные пересечения в ак-
тивации премоторной и слуховой коры, что предполагает их тесную связь при восприя-
тии и порождении. (Источник: Zatorre et al., 2007.)
t
356
Глава 7
жет ли ребенок нормально слышать? Или эти два года без звуковой стимуля-
ции ограничат эффективность применения импланта? Эти и другие вопросы,
касательно корковой пластичности, в настоящее время интенсивно изучаются
учеными.
6.1.
Пластичность, связанная с депривацией
Многое из того, что мы знаем о пластичности слуховой системы, связанной с
депривацией, исходит от работ на животных. Вспомним, что улитка и слуховые
ядра ствола мозга рганизованы тонотопически, и то, что тонотопия присутству-
ет в слуховой коре. При изучении нейрональной пластичности после депри-
вации у животных были повреждены специфические участки улитки и ствола
так, что диапазон частот больше не мог быть закодирован и передан в слуховую
кору. После повреждения изучали организацию слуховой коры для того, чтобы
определить, были ли какие-либо специфические частотные изменения, отра-
жающие нейрональную пластичность. Ирвин и коллеги провели многочислен-
ные исследования, используя общие подходы, и нашли доказательства того, что
корковые карты частот действительно подвергаются изменениям после трав-
мы (Rajan et al., 1993). Мы не можем по этическим соображениям повреждать
улитку или ствол мозга человека, так что наше изучение пластичности, связан-
ной с депривацией, у человека может выполняться только неинвазивными ме-
тодами. Хотя эти исследования находятся пока на начальном этапе, существуют
доказательства того, что у взрослых людей с резким проявлением потери высо-
кочастотного слуха происходят некоторые изменения в популяциях нейронов в
слуховой коре, допуская, что организационные изменения коры, возникающие
в результате потери слуха у людей, сходны с таковыми у животных (Dietrich et
al., 2001). Тем не менее в данном случае идет разговор о резкой потере слуха,
о том, что происходит при постепенной, могущей занимать много лет потере
слуха или при ее легких формах, известно меньше. А что насчет детей, рожда-
ющихся с частичной или полной глухотой? Если их снабдить слуховыми аппа-
ратами или сделать улитковые имплантаты, разовьется ли у них нормальный
слух? Эггермонт и коллеги исследовали ответы слуховой коры у нормальных
детей и детей с потерей слуха, у которых были улитковые имплантаты (Ponton
et al., 1996a, b). У детей с имплантатами было показано отставание в развитии
по сравнению с контролем. Однако в дальнейшем их слуховая система разви-
валась типичным образом, что свидетельствует о пластичности. Эти результа-
ты вдохновляют, так как демонстрируют, что слуховая кора может развиваться
нормально даже после сенсорной депривации в раннем возрасте.
6.2 .
Пластичность, связанная с обучением
Наша слуховая система постоянно встречается с новой сенсорной информаци-
ей, которая должна быть декодирована для того, чтобы мы могли интерпрети-
ровать процессы, протекающие в нашем слуховом окружении. Каждый день мы
выучиваем новые голоса, музыку, звуки окружающей среды. Что представляют
из себя мозговые механизмы обучения новым звукам? Каким образом форми-
руются новые сенсорные воспоминания, и где они хранятся в мозге? Все эти
вопросы были предметом исследований на модели обучения и пластичности
слуховой системы животных.
Слух и речь
357
Некоторые ученые считают, что сенсорные воспоминания хранятся там же,
где и остальные типы воспоминаний, в центральных системах памяти. Дру-
гие считают, что специфическая сенсорная память связана с теми областями,
которые ответственны за выучивание новой сенсорной информации. Вайнбер-
гер с коллегами (Rutkowski and Weinberger, 2005) разработали модель обучения
слуховой системы, которая предполагает, что изменение нейронных механиз-
мов для восприятия новых звуков происходит в первичной слуховой коре (А1)
практически мгновенно, во время первых же тренировочных проб. Согласно
этой точке зрения подстройка нейронов зоны A1 связана с восприятием ре-
A1 связана с восприятием ре-
1 связана с восприятием ре-
левантных в данных условиях характеристик звука. Вайнбергер использовал
классическую методику выработки условных рефлексов, предъявляя разные
тоны одновременно со слабым электрическим током.
После обусловливания карта нейронов зоны А1 реорганизовывалась та-
ким образом, что большие области коры отводились для кодирования частоты
предъявленных тонов (рис. 7 .39).
Рис. 7 .39 . Примеры эффектов тренировки на репрезентационные области. Организа-
ция центральных частот (ЦЧ) в зоне А1 (первичная слуховая кора) для наивных (а)
и тренированных (экспериментальная группа) (б) крыс. Каждый «×» указывает на ме-
сто проникновения электрода, цветом выделены оцениваемые зоны А1 для каждой ЦЧ
(соотношение цветов и ЦЧ изображено под рисунком); Д — вид сзади (дорсальный),
П — спереди. Зоны коры, связывающие ЦЧ с условными стимулами внутри октавной
полосы 4,1–8 кГц, выделены подчеркиванием и вертикальной штриховкой. (в) Кривые
обучения для трех животных с низким (синие квадраты), средним (черные ромбы) и
высоким (красные треугольники) уровнем мотивации. (г) Распределение репрезента-
ционных зон (процент от общей площади А1) для каждого животного, и средние зоны
у необученных животных (серая линия). (Источник: Rutkowski and Weinberger, 2005.)
358
Глава 7
6.3 .
Пластичность, связанная с профессиональными навыками
Неинвазивные поведенческие эксперименты на людях показали, что обучение
происходит достаточно быстро, а изменения запоминается на относительно
большой промежуток времени. Один из вопросов, который интересовал иссле-
дователей, заключался в том, отличается ли мозг профессиональных музыкан-
тов от мозга людей неискушенных в музыке. Музыканты, конечно же, больше
времени проводят, тренируя свои музыкальные навыки. Влияет ли это каким-
нибудь образом на их слуховую кору? Было показано (Schneider et al., 2002), что
определенное влияние существует (рис. 7 .40).
Специфические изменения мозга, которые происходят в разных ситуациях,
например в случае потери слуха, депривации, при обучении, — это крайне ин-
тересная область, в рамках которой активно идут исследования.
7.0 .
Сознание и слуховые образы
Мы можем закрыть глаза и исключить всю зрительную информацию, но мы не
можем препятствовать слуховой системе воспринимать звуки. Что оказывает
влияние на слуховую систему? Слуховая система в последнюю очередь погру-
жается в сон (или переходит в неосознаваемое состояние при полной седации),
Рис. 7 .40 . Нейрофизиологические и анатомические данные указывают на сильные из-
менения у профессиональных музыкантов и меньшие изменения у любителей. Слева:
сила ответа первичной слуховой коры на звук частотой 500 Гц. Справа: выделенные
области показывают извилину Гешля для разных испытуемых, выровнено в порядке
первичных вызванных ответов. (Источник: адаптировано из Schneider et al., 2002.)
Слух и речь
359
и первая пробуждается. В этом разделе мы познакомимся с некоторыми наи-
более новыми данными исследований слухового сознания во время наименее
осознаваемых состояний, таких как нормальный или наркотический сон. Мы
также рассмотрим работы с активацией слуховой системы при восприятии вы-
думанных неслышимых звуков. Эти процессы обработки слуховой информа-
ции имеют место в младенчестве, однако при помощи методов нейровизуализа-
ции можно измерить мозговые ответы, которые невозможно зарегистрировать
другими способами. Предстоит сделать огромную работу, прежде чем мы смо-
жем что-либо с полной определенностью сказать о том, как слуховое сознание
и восприятие реализуются в мозге.
7.1. Слуховое сознание во время сна и седативного состояния
Подумайте о наиболее эффективном способе разбудить спящего друга — про-
изнесите его имя! При помощи методов нейровизуализации исследовали моз-
говые ответы во время сна и бодрствования в ответ на два типа звуковых сти-
мулов: нейтральный, или простой звуковой тон, и значимый, который являлся
именем испытуемого (Portas et al., 2000). Два главных вывода из этой работы:
во-первых, звуковые тоны и имена активируют слуховую кору во время бодр-
ствования и сна, что указывает на то, что звуки воспринимаются и обрабаты-
ваются даже во сне (рис. 7 .41), во-вторых, ответы слуховой коры не различа-
лись во сне при предъявлении нейтральных стимулов и имен, что указывает
на то, что в состоянии сна мозг регистрирует наличие звуковых стимулов, но
дифференцировки не производит. Однако области, активированные именами и
тонами, все же отличались в средней височной извилине и областях лобной
коры. Также были обнаружены области в миндалине, которые оказались более
Рис. 7 .41. Вверху: зоны мозга, активные во время подачи гудков во сне и во время бодр-
ствования. Внизу: зоны мозга, активные при предъявлении имен во время сна и бодр-
ствовании. (Источник: Portas et al., 2000.)
360
Глава 7
активны при предъявлении имени испытуемого в состоянии сна по сравнению
с бодрствованием. Принадлежат ли эти зоны системе предупреждения, кото-
рая заставляет нас проснутся при произнесении нашего имени? Необходимо
провести больше исследований для того, чтобы подтвердить эту теорию, но
существующие работы обеспечивают нас удивительными данными о том, как
слуховая система «будит себя сама».
Похожие результаты были недавно получены Цаторре с коллегами (Plourde
et al., 2006) при наблюдении хирургических пациентов, находящихся под нар-
козом. В ряде современных работ исследован уровень слухового сознания у хи-
рургических пациентов под наркозом, преимущественно для того, чтобы убе-
дится в том, что пациенты не слышат происходящего в операционной. Данные
полученные Цаторре с коллегами совпадают с результатами Портаса с коллега-
ми: слуховая кора во время наркоза активируется простыми и сложными звука-
ми, но паттерн ответа не отличается для речевых и неречевых звуков (рис. 7 .42).
Согласно интерпретации авторов, такие результаты указывают на то, что в
седативном состоянии слуховая кора отвечает на предъявление звуков, однако
ее ответы неспецифичны. Это совпадает с данными исследования предъявле-
ния звуковых тонов и имен в состояниях сна и бодрствования. Однако авто-
ры исследований, проведенных на пациентах под наркозом, предполагают, что
обнаруженное ими отсутствие различий при восприятии звуков означает, что
сложные (семантические, эмоциональные) процессы, активные в состояниях
сознания, не работают в седативных состояниях. Результаты какой работы от-
ражают реальность? Необходимо провести больше исследований с похожими
стимулами и экспериментальными условиями, чтобы точно установить, суще-
ствуют ли разные области, активируемые во время сна и при наркозе, которые
могли бы отразить процессы обработки звуковой информации мозгом.
7.2 .
Слуховое воображение
Звуки, которых нет на самом деле, целый день крутятся у нас в голове. Некото-
рые из них появляются сами собой: навязчивая мелодия, внутренний голос, раз-
Рис. 7 .42 . Усредненные ответы на все звуки-тишину. Карта активация изображена на
усредненных анатомических снимках. Правая часть изображения относится к правому
полушарию. Линейные диаграммы показывают среднюю амплитуду сигнала (разницу
между двумя условиями — тишиной и предъявлением стимула). (Источник: Plourde
et al., 2006.)
Слух и речь
361
говаривающий сам с собой. Другие мы вызываем сознательно: когда повторяем
слова, чтобы выучить их наизусть для школьной пьесы, или проговариваем но-
мер телефона перед тем, как позвонить. С какими процессами в мозге связаны
эти звуки? Нам кажется, что мы действительно слышим эти внутренние звуки.
Активируется ли при этом слуховая кора, несмотря на то что информации о
реальном звуке к ней не поступает? Была изучена (Zatorre & Hal�ern, 2005) ак-
Zatorre & Hal�ern, 2005) ак-
& Hal�ern, 2005) ак-
Hal�ern, 2005) ак-
, 2005) ак-
тивация мозга при воображении звуковых стимулов и наличие реального сигна-
ла с помощью методов нейровизуализации. Результаты (рис. 7 .43) показывают,
что во время мысленного представления звуков действительно активируется
не-первичная слуховая кора.
Похожие данные были получены другими авторами (Bunzeck et al., 2005).
Их работа посвящена воображению звуков окружающей среды. Использовался
метод фМРТ. Результаты представлены на рис. 7 .44 . При предъявлении звуко-
вых стимулов активируется первичная и вторичная слуховая кора в обоих полу-
шариях (слева), тогда как во время мысленного представления звуков — только
вторичная (справа). Таким образом, существуют неоспоримые доказательства
того, что при воображении звуков работают те же зоны мозга, что и при воспри-
ятии реальных звуков. Вспомните также данные Вайнбергера, которые говорят
о том, что слуховая память и обучение также связаны со слуховой корой. Даже
несмотря на то, что здесь приведена только малая часть результатов непрекра-
щающихся работ в области слухового воображения, можно говорить о том, что
процессы мысленного представления звуков и их восприятия протекают в схо-
жих областях мозга.
8.0 .
Заключение
В этой главе был дан обзор слуховой системы, начиная с базовых механизмов
слуха и заканчивая восприятием музыки и слуховым воображением. Изобрете-
ние методов нейровизуализации предоставило возможность получить большое
количество данных, которые позволяют лучше понять корковую организацию
слуховой системы, а также связь разных корковых областей между собой. Не-
смотря на то что намечены основные пути, ведущие к пониманию загадок слу-
хового восприятия, остается сделать еще много. К примеру, только первые шаги
сделаны в области изучения нейронных систем восприятия музыки и речи.
Рис. 7 .43. Активация мозга при мысленном представлении звука. (Источник: Zatorre
and Hal�ern, 2005.)
362
Глава 7
В области организация слуховых механизмов мозга остается еще много
ключевых вопросов. Одним из них является вопрос о роли правого и левого по-
лушарий при музыкальном и речевом восприятии. Плодотворно идет работа по
изучению потоков обработки информации в слуховой системе и мозге в целом.
И хотя работы на приматах показали наличие потоков «что» и «где», у людей
они могут иметь другую организацию в связи с уникальной ролью речи и му-
зыки в эволюции и развитии человеческого мозга. В следующий раз, когда вы
почувствуете, что незнакомая мелодия крутится у вас в голове, подумайте о зо-
нах мозга, которые могут активироваться, когда вы слышите беззвучную песню
9.0 .
Задания и упражнения к главе 7
9.1.
Контрольные задания
1. Каковы базовые физические и психологические характеристики звука?
2. Какие основные части слуховой системы можно выделить и какова их роль
в восприятии?
3. Опишите вкратце различия между «что» и «где» потоками переработки ин-
формации.
4. Что собой представляют основные единицы анализа речи?
5. Какие новые возможности для изучения предоставили методы нейровизуа-
лизации в области слуховой функции?
9.2 .
Графические упражнения
Мы настоятельно рекомендуем выполнить графические упражнения для того,
чтобы запомнить физическое устройство мозга.
1. Вверху, рис. 7 .45: определите слуховые области коры, которые видны при
латеральном взгляде на мозг.
Рис. 7 .44 . фМРТ-исследование восприятия реальных и воображаемых звуков. Звуковые
стимулы не относились ни к речевым, ни к музыкальным, для того чтобы определить
локализацию воображаемых неречевых или музыкальных звуков. Первичная слуховая
кора была активна во время перцептивной фазы эксперимента, но не во время фазы во-
ображения. (Источник: Bunzeck et al., 2005.)
Слух и речь
363
2. Внизу, рис. 7 .45: определите слуховые области коры, выделенные синим,
розовым и коричневым.
9.3 .
Литература
Рекомендуемый материал для чтения и интернет-источники:
Blumstein, S. E. (1997). A �ers�ective on the neurobiology of language. Brain and Language.
60 (3), 335–346 .
Hickok, G., & Poe��el, D. (2004). Dorsal and ventral streams: A framework for understanding
as�ects of the functional anatomy for language. C�gniti�n. 92, 67–99.
Moore, B. C. (2002). Psychoacoustics of normal and im�aired hearing. British Medical
Bulletin, 63, 121–134 .
Peretz, I., & Zatorre, R. J. (2005). Brain organization for music �rocessing. Annual review of
Psych�l�gy, 56, 89–114 .
NIH websiteon theauditory cortex< htt�://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid= neurosci.
section.919>.
NIH website for searching the literature on auditory �rocesses Entrez Pubmad — <htt�://
www.ncbi.nlm.nih.gov/entrez>.
You can enter names of scientists mentioned in this cha�ter into a search engine like Google.
Many have laboratory websites that are interesting to ex�lore.< htt�://www.google.com>.
Рис. 7 .45. Аксиальный (горизонтальный) срез мозга. Показана анатомическая структу-
ра слуховой коры левого и правого полушарий.
Центральная сцена знаменитой картины Диего Веласкеса (1599–1660) «Менины» или
«Фрейлины», которые были королевскими спутницами инфанты (маленькая девочка
в центре), принцессы Испании и дочери короля Филиппа IV и его жены (показаны на
затуманенном изображении в задней части комнаты). Большинство предполагает, что
эта картина является размышлением о самосознании, одной из тем этой главы. Обра-
тите внимание, что почти все на этом полотне, кажется, уделяют внимание себе или
зеркальному отображению себя, начиная с самого Веласкеса (слева), который смотрит-
ся в большое зеркало для того, чтобы изобразить всю сцену. Мы, наблюдатели, таким
образом, являемся частью этой сцены, так как она нарисована с точки зрения зрителей.
Саму инфанту, кажется, тоже интересует, выглядит ли она красивой. У детей развива-
ется социальное самосознание в возрасте около четырех лет. Это крайне важная сторо-
на человеческого разума и нашей социальной жизни. (Источник: Wikimedia. Оригинал
картины хранится в музее Прадо в Мадриде.)
Миллионы вещей... присутствуют в моих чувствах, которые никогда,
строго говоря, не входят в мои сознательные впечатления. Почему? Потому
что они не интересны для меня. Мои впечатления — это то, чему я согласился
уделить внимание... Каждый из нас буквально выбирает, используя свой путь
внимательно следить за предметами, в какой вселенной он окажется сам и
будет ее заполнять.
Уильям Джеймс (1890), «Принципы психологии»
Содержание
1.0.Введение.......................................................366
1.1. Бодрствующее состояние опосредовано сознанием. . . . . . . . . . . . . . . . 370
1.2 . Состояния бодрствования, сна и сновидений должны быть
регулярныповремени........................................371
1.3.Общиеритмысуточныхсостояний.............................373
1.4 . Состояние нахождения в сознании имеет типичную
таламокортикальнуюактивность...............................378
1.5 . Таламокортикальный центр: плотно взаимосвязанный
иоченьактивный............................................381
1.6.Картыиритмы..............................................385
1.7.Двусторонниесвязи..........................................385
1.8.Каксинхронизируютсянейроны...............................387
1.9.Синхронностьдляполученияконтроля..........................390
2.0.Бодрствование...................................................394
2.1.Практическоемышление......................................394
2.2.Ориентацияпоместу,времениилюдям.........................396
2.3 . Бодрствование для обучения; сон делает возможным
закреплениепамяти..........................................398
2.4 . Внимание и сознание в общем делают возможным обучение. . . . . . . . 398
2.5. Потеряосознанияпредсказуемыхсобытий.......................401
2.6.Скрытоеобучениетакжетребуетсознания.......................404
2.7 . Быстрые ритмы координируют задачи в состоянии бодрствования . . . 404
2.8. Гамма-активностьиграетмножестворолей......................406
2.9 . Синхронизация гамма-волн может соединять визуальные черты
восознаваемыеобразы.......................................409
2.10.Тета-ритмыиграютмножестворолей...........................415
2.11.Альфа-ритмы...............................................417
3.0 . Внимание улучшает восприятие, познавательную способность и обучение . 420
3.1.ФланговаязадачаПознера.....................................420
3.2.Модельвнимания............................................427
3.3.Опытыповниманиюиопытысознания.........................433
4.0.Сонфазыбыстрыхдвиженийглаз ..................................435
4.1.Сныкаксостояниесознания...................................437
4.2. ЗакреплениесобытийвпамятивовремяБДГ-сна.................438
5.0.Глубокийсон:подъемыиспады.....................................441
5.1 . Некоторая умственная активность происходит
дажевовремямедленноволновогосна..........................442
5.2. Порогпробужденияизменяетсянапротяжениисна...............442
5.3 . Повторное «проигрывание» эпизодов в памяти и консолидация . . . . . 443
Глава 8
Сознание и внимание
366
Глава 8
6.0.Общеесопоставление.............................................447
6.1 . Отражает ли сознание функцию глобального рабочего пространства
вголовноммозге?.............................................447
6.2 . Передача сигналов повторным входом и степень интеграции
осознаваемыхявленийвголовноммозге..........................454
6.3.Требуетсялидлясознанияличноепознание?......................457
6.4 . Почему сознаваемые события подлежат регистрации? . . . . . . . . . . . . . . 459
6.5.Фактыпонеобычнымсостояниям...............................460
7.0.Заключение.....................................................471
8.0.Заданияиупражнениякглаве8....................................473
1.0 . Введение
Мы просыпаемся каждое утро и окунаемся в непрерывный поток осознаваемых
событий. Как только мы открываем глаза, зрительный мир опять начинает дей-
ствовать. Поток связанной с этим внутренней речи также возникает, как только
мы просыпаемся: мы начинаем говорить сами с собой. Содержание сознания
начинает кодироваться как воспоминания о событиях, так что мы можем уз-
навать и вызывать заново свои впечатления (см. гл . 9). По сравнению с бодр-
ствующим состоянием, мы обычно плохо вспоминаем события во сне, даже в
тех снах, которые были непосредственно перед просыпанием. Помимо этого,
мы видим сны от 90 до 120 мин каждую ночь. Таким образом, мы никогда не
вспомним большинство снов.
Ко гд а мы приходим в сознание каждое утро, мы помним наши наиболее
важные текущие цели. Это означает, что мы можем проявлять сознательное
внимание к информации, которая придет в наше сознание. Уильям Джеймс пи-
сал ( выше): «Мой опыт это то, чему я разрешу присутствовать... Каждый из нас
буквально выбирал, своим собственным образом уделяя внимание событиям, в
каком типе вселенной он окажется сам, чтобы там жить». Джеймс не объясняет
самопроизвольное внимание — вещам, которым мы уделяем внимание, даже
если они не по своему выбору пришли на ум. Но он объясняет, как с течением
времени предметы, которые мы выбрали, чтобы уделить им внимание, прихо-
дят, чтобы оказывать преобладающее влияние на наши сознательные периоды
бодрствования.
После пробуждения мы опять можем контролировать свои скелетные мыш-
цы — в отличие от того, что происходит во время сна, ко гда они почти пара-
лизованы, — поэтому мы можем встать, исследовать мир, ходить на охоту или
собрания, говорить друг с другом. Каждая из этих возможностей набирает в
головном мозге различные цепи событий, которые можно исследовать, мобили-
зовать и контролировать только в бодрствующем состоянии. В бодрствующем
состоянии мы имеем широкий набор возможностей, которые недоступны в дру-
гом состоянии.
Наша функциональная схема (рис. 8.1) показывает общее представление о
«внимании» в виде стрелки, которая идет от областей, находящихся под ко н -
тролем внимания, к познавательным процессам, которые улучшаются внима-
нием. Сознательное внимание показано как стрелка, идущая от центральной
исполнительной системы (приблизительно лобные доли) к разным типам дея-
тельности мозга, которые усиливаются с помощью внимания. Сенсорная кора
Сознание и внимание
367
также может быть усилена с помощью событий, не находящихся под контролем
сознания. Если кто-то громко закричит, неожиданно залает большая собака на
расстоянии в несколько футов от нас или грузовик зловеще вырастет в вашем
поле зрения, внимание будет привлечено этим стимулом. Внимание иницииру-
ют многие биологически значимые события, такие как запах пищи, когда вы
голодны. Мы также самопроизвольно выбираем стимулы, значимые лично для
нас, такие как звуки наших собственных имен. Мы можем изобразить внима-
ние, привлеченное стимулами, в виде стрелки, выходящей от сенсорных отде-
лов диаграммы, чтобы навести на мысль, что некоторые стимулы по своей сути
могут захватить внимание.
Сенсорные области задней коры особенно чувствительны к усилению рабо-
ты путем привлечения внимания. Однако на моторные области, зону Брока и
семантическую память также может выборочно влиять внимание.
Если мы принимаем то, что сознательный опыт во многих случаях является
результатом избирательного внимания, это также может быть легко отражено
на диаграмме. Хотя ведутся научные дискуссии, является ли весь сознатель-
ный опыт результатом выбора с помощью внимания, никто не сомневается, что
многое зависит именно от этого. Каждый, кто преподает, привлекал внимание
класса к воспринимаемым сознанием событиям, так же мы начинаем любое
Рис. 8 .1 . Функциональная структура для внимания и сознательных событий. Каким
образом осознание и внимание появляются в функциональной структуре? Заметьте,
что сознание может находиться под произвольным контролем центральной исполни-
тельной системы, так как людей можно попросить обратить внимание на много разноо-
бразных событий. Однако захват внимания происходит тогда, когда сенсорные события
интенсивны, удивительны или достаточно важны, чтобы заставить нас обратить на них
внимание. Привлечение внимания обычно приводит к особому сознательному опыту.
(Источник: Bernard J. Basars, 2009.)
368
Глава 8
групповое собрание. Эксперименты над человеком и животными во многом
представляют то же самое. Связь между вниманием и сознанием глубока и со-
ставляет большую часть нашей каждодневной психологии.
Путем тщательных исследований мы можем отделить явления внимания и
сознания. Чтобы сконцентрироваться на событиях сознания «такое как», мы
обычно изучаем их экспериментально в отличие от близко лежащих бессоз-
нательных событий, как мы уже видели в предыдущих главах (см. гл . 1, 3, 6
и 7). В экспериментах на внимание обычно просят людей выбрать один из двух
альтернативных стимулов. «Внимание», таким образом, связано с процессом
выбора и сознания с собственно сообщаемым опытом.
Некоторые ключевые вопросы для познавательной нейробиологии состоят
в следующем: что характерно для сознательных событий в мозге? Что действи-
тельно значит для кого-то быть сознательным? И как находящиеся в мозге кри-
терии выбора на основе внимания соотносятся с нашим личным сознательным
постижением мира?
Границы познавательной нейробиологии
Как мы изучаем сознание? Как связи между субъективными ощущениями, на кото-
рые философы ссылаются как as qualia (элементы, составляющие любой опыт че-
ловека, независимо от того, является ли это ощущение видения красного, обоняния
коровьих фекалий или нахождения в раздраженном состоянии), и головным мозгом
могут быть поняты объективным, количественно измеряемым и эмпирическим пу-
тем? Было много противников идеи о том, что кто-то может изучать сознание — как
у человека, так и у близкородственных животных, таких как мыши или обезьяны —
научным путем. «Оставьте эту работу нобелевским лауреатам, находящимся на пен-
сии, философам, художникам, литераторам и толпе, образующей финансовую пи-
рамиду» — таковой была общая реакция. Однако сознание является неотъемлемым
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
Рис. 8.2 . Кристоф Кох (слева) является хорошо известным нейробиологом и ис-
следователем основ сознания и их связи с головным мозгом. Кох работал многие
годы с Франсом Х.С . Криком (справа), нобелевским лауреатом и сооткрывателем
ДНК. Крик (1916–2004) посвятил последние десятилетия своей жизни этой теме и
вдохновлял других ученых следовать за ним
Сознание и внимание
369
свойством природы. И конечно, единственным путем, которым я или кто-либо еще
познает вселенную и узнает, что он существует. Декарт сделал наиболее известный
среди западных мыслителей логический вывод: «Je pense, donc je suis». Если мы же-
. Если мы же-
лаем видеть всеобъемлющую научную точку зрения на вселенную и все в ней, тогда
сознание должно играть заметную роль в конечном описании всего, что есть. И за
последние 20 лет мы полагаем, что сделали выдающийся эмпирический прогресс в
зарождающейся науке о сознании. Богатые плоды принес подход, который Фрэнсис
Крик и я поддерживаем в нашей первой статье по данной теме с 1990 г., концентри-
руясь сейчас на нейронных соотношениях с сознанием: минимальные условия со-
стоят в одновременной работе нейронов, необходимых для любого специфического
сознательного ощущения.
Но когда мы в конце концов идентифицировали неуловимые НКС (нервные корре-
ляции сознания), например для зрительного восприятия взрослого человека, нам
нужно будет понять, почему именно эти особенные НКС дают начало сознанию,
а не другие. И нам нужно будет понять, кто из созданий, не относящихся к людям,
обладает сознанием. Это легко сделать для близкородственных организмов, таких
как обезьяны и мыши и, вероятно, для всех млекопитающих, но становится намного
сложнее для случаев, которые отклоняются от данного условия. Обладает ли плод
сознанием? Как насчет одного из тысяч пациентов, находящихся в длительном ко-
матозном состоянии? Пчела? Кальмар? Чтобы ответить на эти или другие вопросы
определенно, нам нужна фундаментальная теория сознания. И языком такой теории
должна стать математика теории информации, так как информация может выражать
причинно-следственные связи между многими частями любой системы, независи-
мо от структурного вещества этой системы, независимо от того, будут ли то нейро-
ны, передающие импульсы, или кремниевые транзисторы.
По моему мнению, из фундаментальных теорий сознания вызывает доверие гипо-
теза Джулио Тонони, называемая Интегральной теорией (ИТ) сознания. Она пред-
лагает захватывающий новый путь изучения сознания, использующий строгий на-
учный подход (Tononi, Edelman, 1998; Balduzzi, Tononi, 2008; Tononi, 2008).
Структура ИТ построена на представлении о том, что сознание является последо-
вательностью систем с огромным количеством различной информации, которая
также в них высокоинтегрирована. В этом контексте интегрированная информация
означает информацию, которая приложима ко всей системе как целое. Например,
iPhone имеет 16 Гб объема памяти. Она представляет около 16 млрд обособленных
различных состояний. Но они совсем не интегрированы. Для iPhone каждый пик-
iPhone каждый пик-
каждый пик-
сель в каждой картинке подобен любой другой информации, например, календарю.
Он не воспринимает, например, что различные фотографии молодой девушки на
моем iPhone, которая превращается в уверенную в себе женщину, — это фотогра-
iPhone, которая превращается в уверенную в себе женщину, — это фотогра-
, которая превращается в уверенную в себе женщину, — это фотогра-
фии моей дочери, чьи данные есть в моей телефонной книге, и что она является
родственницей улыбающемуся уверенному мужчине, ее брату, в следующей груп-
пе фотографий. Для последнего поколения iPhone (и других компьютеров) все эти
пиксели равно значимы (он даже не знает, что изображения по своему существу
двумерны). ИТ предоставляет точный путь для вычисления размера спектра (на-
бора) сознания, состояние сознательности в данной ситуации использует число, из-
меренное в единице информации, которая обозначена Φ.
ИТ Тонони предоставляет важный инструмент для знакомства с основами сознания
в природе в количественной мере. Согласно Тонони, количество интегрированной
информации, которые вы или я имеем в мозге, соответствует нашему набору воз-
можных сознательных состояний. Многие интересные вопросы возникают с появ-
лением такой теории. Один из особенно изумляющих моментов — это механизм
дизайна (я не только профессор биологии, но также являюсь им в технических на-
уках). Могут ли искусственные объекты, сделанные из кремния, медных проводов
370
Глава 8
и даже стали быть наделенными сознанием? Могут ли они чувствовать мир (позна-
вать мир на собственном опыте)? ИТ ясно говорит «да», доказывая, что они обла-
дают как высокой дифференциацией, так и интегрированной информацией (Koch,
Tononi, 2008).
Больше информации о нейробиологических аспектах сознания можно найти в моей
книге «Поиск сознания: нейробиологический подход» (The Quest for Consciousness:
A �euro�iolo�ical A��roach). Исследование, которое мы проводили в моей лабора-
�euro�iolo�ical A��roach). Исследование, которое мы проводили в моей лабора-
�euro�iolo�ical A��roach). Исследование, которое мы проводили в моей лабора-
A��roach). Исследование, которое мы проводили в моей лабора-
A��roach). Исследование, которое мы проводили в моей лабора-
). Исследование, которое мы проводили в моей лабора-
тории, детально описано на htt�://www.kla�.caltech.edu/.
Литература
1. Tononi G., Edelman G.M . (1998). Consciousness and com�lexity 4. Science, 282
(5395), 1846–1851.
2. Balduzzi D., Tononi G. (June 2008). Inte�rated information in discrete Dynamical
‘systems: motivation and theoretical framework. PLoS Computational biology, 4 (6).
3. Tononi G. (Decem�er 2008). Consciousness as inte�rated information: a �rovisional
manifesto. Biological bulletin, 215 (3), 216–242 .
4. Koch C., Tononi G. (June 2008). Can machines �e conscious? IEEE Spectrum, 45 (6),
55–59.
5. Koch C. Division of �iolo�y. PhD. California institute of technolo�y, Pasadena, CA,
USA.
1.1 . Бодрствующее состояние опосредовано сознанием
Бодрствующее состояние содержит нескончаемый ряд адаптивных функций.
Во время бодрствующего сознания мы учим новые действия, чувства, слова,
идеи, умения, цели и даже новые виды сенсорных событий, такие как новый
путь понимания музыки или зрительного искусства; каждое из них включает
характерные шаблоны деятельности мозга. Ряд функций в бодрствующем со-
стоянии, таким образом, является открытым рядом, который может продолжать
расширяться столь же долго, пока мозг жив. Наиболее основные обсуждаются
по всей этой книге: зрение, слух, память, эмоции, исполнительный контроль,
социальное распознавание и т. д .
Каждая задача в бодрствующем состоянии имеет как сознательный, так и
подсознательный компоненты. Например, нижний вентральный путь зрения
предоставляет передаваемые на словах осознаваемые образы объектов, такие
как зрительные образы чашек кофе или кухонного стула. Верхний дорсальный
путь зрительных зон мозга контролирует действия, такие как «достигнуть ру-
кой объекта», но об их компонентах нельзя говорить как о сознательных (Mil-
Mil-
ner, Goodale, 2007). Как дорсальный, так и вентральный пути требуют состоя-
ния бодрствующего сознания, чтобы работать. Почти все когнитивные задачи,
которые мы знаем, выполняются во время бодрствующего состояния и имеют
как сознательные (о которых можно сообщить), так и подсознательные (не под-
лежащие сообщению) компоненты.
Таким образом, большинство когнитивных задач, которые нам известны,
опосредованы сознанием. Рабочая память, например, имеет как сознательные,
так и подсознательные составные элементы. Если вы повторите семь чисел, вы
заметите, что некоторые являются осознанными в любой момент, а другие —
нет. Инструкции для повторения и запоминания явно сознательны, и таким же
Сознание и внимание
371
является ряд пунктов, если вы видите или слышите их. Но мы не осознаем не-
повторяемые элементы в любой момент, так же как и важную роль базальных
ганглиев в контролировании внутренней речи или автоматические, например,
элементы любой задачи. Насколько мы знаем, нет полностью осознаваемых
когнитивных задач, а также может не быть полностью неосознаваемых задач
(Baars, Franklin, 2003); профессор Стэн Франклин предложил термин «опосре-
дованный сознанием» для когнитивных задач, которые имеют сознательный
компонент, но во всем остальном неосознаваемые.
Базальные ганглии и мозжечок являются очень большими структурами моз-
га, которые, как полагают, функционируют без поддержки в каждый момент
содержимым сознания даже в бодрствующем состоянии. Мозжечок может быть
фактически поврежден с двух сторон, но люди и животные будут продолжать
вести себя во многом так же, как перед тем, но без возможности контролиро-
вать тонкие моторные движения. У людей эти структуры играют много других
когнитивных ролей, но не приводят прямо к сознательному содержимому.
Что мы можем делать полностью бессознательно? Мы все еще не знаем
ответ, потому что трудно сделать точные исследования по хождению во сне,
двигательным нарушениям во время сна, эпилептическому «автоматичному
поведению» и другим «состояниям зомби» (Crick, Koch, 2003). Существует
много сведений по автоматическому поведению от людей с нарушениями сна и
эпилепсией. Чтобы подтвердить эти сообщения, нам необходимы записи с моз-
га, которые трудно получить от свободно двигающегося человека. Например,
также возможно, что поведенческие автоматизмы при эпилепсии отражают
моментальные «вспышки» сознания, о которых известно, что они существуют
(Kranczioch et al., 2006). Таким образом, трудно протестировать, существует
ли сложные, но полностью бессознательные акты поведения, отчасти потому,
что мы просто все еще не знаем характерные корреляции сознания в головном
мозге (Gaillard et al., 2009; Revonsuo, 2008). Нам пока неизвестно, какая разница
включает сознание по вентральному, но не по дорсальному пути зрительной
коры. Однако делаются непрерывные усилия по достижению успехов в этих
вопросах (Laureys, Tononi, 2008).
1.2 . Состояния бодрствования, сна и сновидений
должны быть регулярны по времени
Крысы, постоянно лишенные сна, умирают в течение трех недель. Таким обра-
зом, мы узнали, что сон необходим для выживания млекопитающих, но его точ-
ные функции еще неясны. Спящие животные более уязвимы для хищников. Сле-
довательно, наши восемь часов потери восприятия нашего окружения каждую
ночь должны обладать некоторыми компенсирующими это преимуществами. Мы
знаем, что сон и сновидения, кажется, делают возможным объединение памя-
ти, преобразуя новые и нестабильные воспоминания в долговременную память.
Сон, сновидения и бодрствование являются сложными состояниями с длинной
эволюционной историей. Как другие важные адаптации, они, вероятно, приоб-
ретали многие биологические функции. Известно, что большое число генов на-
чинают работать (экспрессируются) во время сна, а многие гены изменяют свою
экспрессию из-за нарушений сна. Множество биологических параметров, веро-
ятно, включаются или выключаются благодаря сну или его потере.
372
Глава 8
Продолжительность сна, вероятно, находится под контролем гомеостаза,
так как мы имеем склонность возмещать недостаток сна более продолжитель-
ным временем отдыха. У людей, лишенных сна, возникают моменты «микро-
сна», во время которых они «засыпают», что может серьезно влиять на такие
действия, как вождение автомобиля. Вы можете наблюдать моменты засыпа-
ния непосредственно во время учебы поздно ночью. Когда ваш мозг быстро
переключается с бодрствования на сон, корректирующие химические вещества
головного и спинного мозга переключаются достаточно быстро (рис. 8 .3). На
секунду или две мускулы вашей шеи могут потерять тонус, и ваша голова бу-
дет стремиться упасть вперед. Моторные спинно-мозговые и черепно-мозговые
нейроны переключаются на торможение, и это время, чтобы получить некото-
рый отдых. У нас имеется некоторый контроль коры головного мозга над под-
держанием бодрствующего состояния, но в конце концов биологические часы
добьются успеха.
К удивлению, единственная ночь лишения сна может улучшить состояние
при депрессии, даже если другие средства лечения не помогли. Таким образом,
Рис. 8 .3 . Три измерения бодрствования, сна и сновидений. Многие явления, относящи-
еся к нашему состоянию, могут быть описаны по этим трем измерениям (Ho�son et al.,
2000; Giocomo, Hassem�lo, 2007). «Быстрая активация коры» относится к обширной,
быстрой, нерегулярной активности с низкой амплитудой на ЭЭГ. «Источники входящих
сигналов» либо внутренние, как при сновидении, либо внешние, как в бодрствующем
состоянии. «Нейромодуляция» относится к четырем широко распространенным нейро-
медиаторам, которые включаются или выключаются базальными ганглиями головного
мозга. Это ацетилхолин (Ach), норадреналин (�E), серотонин (5-HT, 5-гидрокситрип-
тамин) и глутаминовая кислота (Glut). Эти молекулы играют жизненно важные роли в
восприятии, двигательном контроле и обучении. Медленноволновый сон (МВС) харак-
теризуется дельта-волнами в нативной ЭЭГ, имеет самый высокий порог пробуждения в
циркадианном цикле. БДГ — сон с быстрыми движениями глаз. Во время БДГ уровень
ацетилхолина может быть высоким. (Источник: Ho�son, 2000, с изменениями.)
Сознание и внимание
373
было предложено не ложиться спать в качестве безопасного, недорогого и эф-
фективного средства лечения депрессии. Но хроническое лишение сна является
стрессовым и ухудшает нормальное функционирование сознания.
Цикл бодрствование–сон контролируется биологическими часами, запуска-
емыми частично началом и длительностью светового дня. Небольшая группа
рецепторов света на сетчатке определяет начало светового дня, подавая сигнал
супрахиазматическому ядру, эпифизу, гипоталамусу и глубоким ядрам головно-
го мозга, для высвобождения молекул, специфических для данного состояния.
Основные нейромодуляторы имеют действие, специфичное для этого состоя-
ния (рис. 8 .3). Изменения уровня серотонина, норадреналина и ацетилхолина
были хорошо описаны для запуска переходов основных состояний головного
мозга (см. гл . 16).
1.3 . Общие ритмы суточных состояний
В момент пробуждения от сна миллиарды нейронов изменяют функционирова-
ние с медленноволнового сна с синхронизированными периодами «работы —
паузы» на более дифференцированную и специфическую для задач передачу
сигналов. Разница между сном и бодрствованием не только субъективна и про-
является в поведении. На рис. 8.4 показаны основные рисунки записей, которые
мы обнаружили на нативной ЭЭГ в трех основных состояниях.
Хотя снятая со скальпа ЭЭГ очень полезна, она относительно груба, так
как показывает суммирование результатов миллиардов быстрых электриче-
ских сигналов от огромной совокупности клеток, прошедших через плотные
слои тканей и костей. Большие усилия были направлены на выяснение основ
этих состояний в мозге, с использованием глубоких электродов и томографии.
Большую часть того, что мы знаем о таламо-кортикальной системе, получи-
ли при исследованиях других млекопитающих и людей-пациентов. Все мле-
копитающие имеют основную таламо-кортикальную систему. Нативная ЭЭГ
кажется относительно грубой, но даже такая ЭЭГ дает фундаментально важ-
ную информацию о сне, бодрствовании и некоторых нарушениях, таких как
эпилепсия.
На рис. 8 .4 представлена разница по поверхностной ЭЭГ между бодрство-
ванием, сном и БДГ-сновидениями. Каждое состояние ЭЭГ также сопровожда-
ется типичными субъективными впечатлениями. Удивительно, но медленно-
волновый сон также показывает некоторую умственную активность; однако во
время глубокого сна статистически менее вероятны данные о наличии менталь-
ной активности. Глубокий сон это наименее сознательное состояние нормаль-
ного суточного цикла, если измерять по способности к поведению при про-
буждении: вероятность того, что животное в ответ на стимул сядет, настроит
рецепторы и покажет ЭЭГ бодрствования, очень мала.
Стандартная поверхностная ЭЭГ образуется отростками кортикальных ней-
ронов, расположенных в верхних слоях: обычно это дендриты пирамидных кле-
ток, клетки, имеющие форму пирамиды с длинными аксонами, которые идут
вниз от поверхности коры. Их длинные аксоны соединяются с другими частями
коры или таламуса. Пирамидные клетки возбуждающие, используют глутами-
новую кислоту как основной нейромедиатор, но они окружены небольшими
тормозными интернейронами, которые используют в качестве нейромедиаото-
374
Глава 8
Рис. 8 .4 . Поверхностные ЭЭГ основных состояний. Головной мозг образует электро-
магнитное поле, которое можно записать с кожи головы. Каждая линия представляет
собой колебания поля, возникающего в результате суммирования невысокого электри-
ческого напряжения миллиардов нейронов около поверхности коры, прошедших через
слои тканей. Линия ЭЭГ отражает сложное смешение многих дифференцированных
волн, но каждое состояние показывает доминирующую форму волн, которая доволь-
но характерна. Заметьте сходство между ЭЭГ в бодрствующем состоянии (вверху) и
БДГ-сновидения (внизу). Судя по электрической активности головного мозга, БДГ-
сновидения похожи на состояние нахождения в сознании — то, что мы действитель-
но ощущаем во время сновидений. Когда мы засыпаем и переходим на более глубокие
стадии сна (стадии 1–4), появляются сонные веретена (веретенообразные всплески),
К-комплексы и медленные дельта-волны. (Дельта-волны определяются как имеющие
частоту менее 3 Гц.) В последнее время были изучены медленные колебания, умень-
шающиеся до 0,01 Гц. При глубоком сне в основном видны дельта-волны, отражающие
одновременные возбуждения и паузы миллиардов нейронов, которые возбуждаются и
останавливаются вместе. Сейчас известно, что очень медленные колебания происходят
даже во время состояния бодрствования. Ц/с — циклов в секунду. (Источник: Squire,
2008.)
Сознание и внимание
375
ра ГАМК. Возбуждающие и тормозные нейроны работают вместе в одном и том
же локальном участке коры. Нейроны коры складываются в мини-колонки из
шести слоев (см. гл . 5), которые постоянно взаимодействуют с клетками тала-
муса, поэтому на ЭЭГ с поверхности головы мы видим массовые суммирован-
ные колебания таламокортикальной системы как целого. Эта система является
мозговой основой состояния сознания.
Было трудно получить «чистые» сигналы головного мозга, используя ЭЭГ
с поверхности головы, хотя в течение нескольких лет наблюдался заметный
прогресс. Как вы знаете из гл. 4, МЭГ (магнитоэнцефалография) позволяет по-
лучить лучшую локализацию участка, чем поверхностная ЭЭГ, хотя она реги-
стрирует магнитную составляющую. Локальные потенциалы поля могут быть
записаны от мозга с помощью глубоких электродов. Тем не менее ЭЭГ является
наиболее широко используемой методикой записей активности мозга, потому
что она относительно дешева, улавливает жизненно важные медицинские яв-
ления (такие как эпилепсия и нарушения сна), а тщательный математический
анализ помогает «очистить» сигнал.
ЭЭГ напоминает тип информации, которую инопланетяне могут получить из
радиосигналов, приходящих от Земли, без большого понимания человеческой
речи и только со смутной идеей того, что могли сказать люди. Тем не менее здесь
был заметный прогресс в понимании (Buzsaki, 2002). Интракраниальные записи
более аккуратны и дают наименьшее количество шума, но главным недостатком
является их инвазивный характер. Безусловно, огромное количество глубоких за-
писей было выполнено на животных, но интракраниальная ЭЭГ человека (иЭЭГ)
в настоящее время начинает выступать как основной источник научных данных.
Об иЭЭГ упоминал еще первый нейрохирург Уайлдер Пенфилд около 50 лет
тому назад. Сила сигнала внутри головного мозга до 1000 раз больше, чем на
коже головы, и может быть записан более широкий спектр частот. К тому же
иЭЭГ не имеет некоторых экспериментальных артефактов, таких как отдельные
потенциалы, происходящие от движений глаз и мышц кожи головы. иЭЭГ чело-
века выполняется только по оправданным с точки зрения медицины обстоятель-
ствам, но она, как мы увидим, добавила очень много к нашим знаниям.
Обширные области мозга вне таломокортикальной системы могут иметь
небольшой прямой эффект на осознанное состояние. Например, люди и жи-
вотные с двусторонней потерей мозжечка показывают ухудшенный контроль
тонких движений (движений пальцев и рук), но они сохраняют бодрствующее
осознание мира и самих себя. Последние исследования показали, что мозже-
чок выполняет когнитивные функции, вероятно, путем взаимодействия с корой.
Мозжечок содержит примерно столько же нейронов, сколько и кора, и этот ос-
новной факт, вероятно, доказывает, что сознание является не только результа-
том действия большого количества нейронов, а требуется что-то еще.
Эделман и Тонони (Edelman, Tononi, 2000) предположили, что именно вы-
сокая степень взаимодействия между корой и таламусом позволяет выполнять
обширный набор задач и впечатлений, опосредованные сознанием. На рис. 8 .5
и 8.6 показаны анатомические основы этой эластичной способности, основан-
ной на взаимодействии. В сравнении с этим, мозжечок намного больше поде-
лен на ячейки, в нем имеются отдельные группы нейроны, которые, вероятно,
действуют параллельн друг с другом. Всемирная сеть является естественным
376
Глава 8
аналогом для таламокортикальной системы, так как любой компьютер в Интер-
нете может соединяться с любым другим.
Глубокий сон является наиболее бессознательным состоянием суточно-
го цикла, для которого характерны медленные высоковольтные формы волн,
включая скоординированные между очень обширными группами клеток «со-
стояния подъема» и «спада». ЭЭГ глубокого сна имеют некоторую схожесть
с картиной при потери сознания во время эпилепсии, так же как с состояниями
комы и общей анестезии (рис. 8 .5).
Эпилепсия может уменьшать сознание благодаря гиперсинхронии большо-
го количества нейронов коры и таламуса. На рис. 8 .5 показана ЭЭГ, сделан-
ная во время эпилептического припадка 7-летней девочки с потерей созна-
ния (Blumenfeld, 2004). Обратите внимание на медленные сигналы, имеющие
большую амплитуду, особенно в лобных долях. Такие гиперсинхронные волны
часто распространяются путем «рекрутирования» (вовлечения) других частей
коры. фМРТ и ПЭТ (позитронно-эмиссионная томография) показывают па-
дение метаболической активности во время припадков в лобной и теменной
долях. Заметьте некоторую схожесть между естественным медленноволновым
сном (рис. 8 .4) и потерей сознания при эпилепсии. Обе ЭЭГ показывают мед-
ленную, синхронную активность в больших частях таламокортикального цен-
Рис. 8 .5. Медленные гиперсинхронные потенциалы при потере сознания при эпилеп-
сии. Поверхностное ЭЭГ 7-летней девочки во время эпилептического приступа с по-
терей сознания. Потеря сознания при эпилепсии показывает медленное гиперсинхро-
низированное всеобъемлющее ЭЭГ, похожее на глубокий сон. Другие бессознательные
состояния, такие как кома и общая анестезия, также демонстрируют медленные син-
хронизированные волны с большой амплитудой, распространяющиеся по коре, что
предполагает: сама по себе синхронность не является маркером сознания. (Источник:
Blemendfled, 2005.)
Сознание и внимание
377
тра. Однако волны при эпилепсии намного более зубчатые, чем дельта-волны
при глубоком сне*
.
Имеется явный парадокс в обширной гиперсинхронизации в ЭЭГ при бес-
сознательных состояниях. С одной стороны, синхронизированные ритмы
головного мозга позволяют широко разделенным областям мозга работать
вместе. С другой стороны, электрические возбуждения глобальной гиперсин-
хронии препятствуют выполнению обычных функций мозга. Из этого явствует,
что нормальная деятельность для выживания не может выполняться во время
эпизодов обширной гиперсинхронизации.
* Пациенты в коме, так же как пациенты под общей анестезией, могут показывать ЭЭГ с высоки-
ми и длинными волнами. Вероятно, что бессознательные состояния обычно имеют высокие, мед-
ленные, координированные волны. Однако в наиболее тяжелых случаях комы ЭЭГ уменьшится до
очень низких амплитуд. Клиническая смерть мозга часто определяется как нулевое напряжение на
ЭЭГ. Однако недавние исследования показали, что обычная ЭЭГ с кожи головы может не улавливать
остаточную активность мозга в случаях комы. Медицинские исследователи тогда предложили новую
диагностическую категорию — «минимально сознательное состояние» , которая может имитировать
настоящую кому, но все еще делает возможными моменты сознания. Большой процент пациентов,
которым до этого диагностировали необратимую кому, сейчас считаются находящимися в минималь-
но сознательном состоянии с эпизодами перемежающегося сознания (Laureys, Tononi, 2008).
Рис. 8 .6 . Три нейрона в таламокортикальной петле. Основной водитель ритма в моз-
ге. Бодрствование, сон и сновидения регулируются таламокортикальными колеба-
ниями. Ядра таламуса тесно взаимодействуют с соответствующими областями коры.
Этот центр мозга является общим с другими млекопитающими и птицами. (Источник:
Steriade, 2006, с изменениями.)
378
Глава 8
Один из возможных ответов — то, что нормальное познавательная способ-
ность требует избирательной локальной одновременной деятельности между
областями мозга, и скорость этой деятельности при работе мозга больше, чем
волны с частотой примерно 1 Гц глубокого сна и эпилептических припадков.
Задачи при бодрствовании используют синхронизацю тета-, альфа-, бета- и
гамма-волн, что охватывает ряд частот приблизительно от 4 Гц до 200 Гц или
даже больше. Во время бодрствования наблюдались даже внезапные вспышки
в 1000 Гц. Колебания во время бордствования крайне структурированы и диф-
ференцированы так, что синхронные дейстия, асинхронные и непериодичные
«единичные» волны постоянно появляются и исчезают (Tononi, 2004).
Сознание во время бодрствования напоминает транспортный поток в ожив-
ленном большом городе. Ночью уличное движение может уменьшаться до не-
скольких машин, но в час пик машины постоянно едут из одной точки в дру-
гую. Именно способность передвигаться в любом направлении, руководствуясь
ограниченными целями, делает транспортные средства весьма полезными. По-
добным образом, задачи во время бодрствования обеспечивают выполнение
огромного разнообразия функций с большой гибкостью. В отличие от этого во
время медленной ЭЭГ гиперсинхронизации кажется, что все светофоры загора-
ются зеленым на несколько секунд, затем красным, затем опять зеленым, опять
и опять. Таким образом, транспортный поток заставляют тормозить, чтобы
одновременно остановиться, затем опять стартовать, опять остановиться и так
далее. Нормальный поток сигналов мозга постоянно нарушается.
1.4 . Состояние нахождения в сознании
имеет типичную таламокортикальную активность
На рис. 8 .6 показано основное объединение таламокортикальной системы, со-
стоящей из трех соединенных нейронов в коре, таламусе и ретикулярном ядре
таламуса. Корковый нейрон на рисунке является пирамидной клеткой, кото-
рая названа так из-за того, что ее тело имеет форму пирамиды. Они являются
нейронами, передающими возбуждение на большие расстояния в коре, и часто
окружены более мелкими клетками. Помните, что таламус считается «ворота-
ми к коре» и что большинство таламических ядер проецируются в соответству-
ющие области коры, с которыми они взаимодействуют. Некоторые таламиче-
ские ядра, такие как ЛКТ (латеральное коленчатое ядро) (см. гл . 6), включают
нейроны, переключающие сигналы от сенсорной области к области коры. Дру-
гие являются кортико-кортикальными, которые позволяют системам концен-
трации внимания в теменной и лобной долях усиливать обработку сенсорной
информации в задней коре (через медиодорсальные (срединно-задние) ядра и
ядра подушки таламуса).
Все вместе нейроны на рис. 8 .6 образуют круг, который определяет основ-
ные состояния суточного (циркадианного) цикла. Четвертая клетка глубоко в
мозге может включать и выключать этот контур, используя нейромодулирую-
щие химические вещества на всем протяжении обширных частей переднего
мозга. Имеется несколько миллионов таких цепей, но основная схема проста
и сответствует таламокортикальной петле (рис. 8.8).
Мы можем думать о такой петле как о круге колебаний, состоящем из трех
колеблющихся единиц. Как дети на двух качелях на площадке, которые подве-
шены на одной перекладине (см. гл . 3), каждая единица может быть рассмотре-
Сознание и внимание
379
на как генератор колебаний. Но качели как целое также являются колебатель-
ной системой. Отдельные нейроны представляют собой быстро заряжающиеся
и разряжающиеся генераторы электрических волн. Цепи нейронов имеют коле-
бания по более сложной схеме. Таламокортикальную петлю на рис. 8 .6 можно
считать основным водителем ритма головного мозга, работающим по-разному
в основных состояниях мозга: бодрствования, медленно-волнового сна и сна с
быстрыми движениями глаз.
На рис. 8 .7 показано, как во время бодрствования миллионы отдельных ней-
ронов возбуждаются по большей части независимо друг от друга. Даже когда
группы нейронов возбуждаются синхронно во время состояния бодрствова-
ния, их взаимодействие относительно быстрое (возможно 4–200 Гц) и распро-
странено местно; быстрая синхронность, вероятно, связана с выполняемыми
задачами, и она изменяется во время продолжения мыслительной деятельно-
Рис. 8 .7 . ЭЭГ и активность одного нейрона в бодрствующем состоянии и при глубоком
сне. Обычная ЭЭГ измеряет электрическое поле мозга поверх кожи головы. Каждая ли-
ния ЭЭГ на рисунке представляет собой сложную сумму активности нижележащих ней-
ронов в верхнем слое коры. ЭЭГ медленноволнового сна отражает синхронные «шумы»
и «паузы» миллиардов нейронов, также называемые состояниями «вверх» и «вниз».
Так как в бодрствующем состоянии нейроны не возбуждаются и не останавливаются
синхронно, их напряжение не суммируется в большие волны. ЭЭГ в бодрствующем со-
стоянии выглядит маленькой, нерегулярной и более быстрой, чем медленноволновый
сон. Вероятно, что таким образом бодрствование (и БДГ-сон) включает больше раз-
личной обработки информации, подобно тому, как стадион, полный говорящих людей,
способствует переработке большей информации, чем те же люди, все поющие в унисон.
Согласованное пение более избыточно (вы можете предсказать, что поет толпа, всего
по одному человеку), поэтому содержание информации в нем ниже (см. раздел 6.2).
(Источник: Steriade, 2006.)
380
Глава 8
сти. В результате возбуждение нейронов при бодрствовании не суммируется
до высокоамплитудных форм волн в отличие от медленноволнового сна (МВС)
(рис. 8 .7, внизу). При медленноволновом сне огромное количество нейронов
таламокортикальной системы формируют одновременные вспышки и паузы
примерно каждую секунду, суммируясь в медленные волны на общей ЭЭГ. Ус-
ловно считают, что такие дельта-волны варьируют от 0,5 до 3 Гц, но недавно
было обнаружено, что они также распространяются на уровень более медлен-
ных колебаний (0,01–0,5 Гц) (рис. 8 .9).
Другой полезной аналогией служит то, что медленноволновый сон напоми-
нает большой футбольный стадион, на котором все фанаты поют и молчат в
унисон каждую секунду или около того. Когда они кричат вместе, суммарная
амплитуда звука очень высока. Когда они вместе делают паузу, общий уровень
Рис. 8 .8. Переключатели состояний головного мозга. Цикл светового дня восприни-
мается специальными рецепторами в сетчатке, которые связаны с СОЯ (супраоптиче-
ским ядром), расположенным как раз над «перекрестом», имеющим Х-образную форму,
местом пересечения зрительных нервов. СОЯ является частью циркадианного круга,
который выполняет много функций, связанных как с мозгом, так и с телом в целом. Из-
менение состояния запускается небольшими ядрами ствола головного мозга и базаль-
ными ядрами, посылающими нейромодулирующие волокна (см. гл . 16), которые изме-
няют рисунок возбуждения многих миллиардов других нейронов. Система, восходящая
от ствола головного мозга к коре, участвует в переключении с медленноволнового сна
(МВС) ко сну с быстрыми движениями глаз (БДГ). Гипоталамус, эндокринный центр
мозга, вовлечен в переключение сон–бодрствование. Таламус связан со всеми частями
коры и является станцией на пути почти всех сенсорных нейронов, идущих к коре. При
МВС образуются сонные веретена (см. рис. 8 .6). Совместно эти области контролируют
дневной цикл основных состояний мозга. Они являются системой, контролирующей
движение для десятков миллиардов других нейронов и триллионов синаптических свя-
зей. (Источник: Bacon et al., 2007.)
Сознание и внимание
381
звука очень низок. Но когда отдельные зрители просто болтают со своими со-
седями, звук нерегулярен, относительно низок по амплитуде и намного более
сложен. С этой точки зрения, именно состояние бодрствования содержит наи-
большее количество полезной информации (Tononi, 2004; см. раздел 7.0).
Настоящие данные, основанные на этих источниках, показывают жизненно
важную роль синхронизированных и взаимодействующих электрических коле-
баний мозга. Полагают (Canolty et al., 2006), что они делают возможными:
• внимание, восприятие и сознание;
• обучение, нейропластичность и извлечение памяти;
• долгосрочное взаимодействие между областями мозга, включая взаимо-
действие между сенсорными и двигательными областями.
Мозговые волны часто взаимодействуют друг с другом, при этом медленные
ритмы стремятся объединиться с быстрыми. Взаимодействия часто кратковре-
менны и относятся к специфичным задачам. Играют роль как синхронность так
и асинхронность.
1.5 . Таламокортикальный центр:
плотно взаимосвязанный и очень активный
Наиболее легко понять схему соединения таламуса и коры, используя анало-
гии с простой цепью, как представлено на рис. 8 .10 (а) — точно так же легче
разобраться в автомобильном движении в большом городе в терминах един-
ственной машины, едущей из пункта А в пункт Б. Также важно помнить про
огромный размер всей столичной области — таламокортикальной системы и
ее постоянный поток движения со многими событиями, которые происходят в
одно и то же время. Ижикевич с соавторами разработали широкомасштабные
имитационные модели этой системы, которые помогают нам понять, как систе-
ма работает (Izhikevich et al., 2004, 2008). Рисунки в этом разделе показывают
разные виды головного мозга с активной корой.
Рис. 8 .9 . Ритмы головного мозга можно выстроить по нескольким последовательно-
стям величин. Широкий порядок функциональных колебаний мозга. Слева очень мед-
ленная активность проявляется как опускание основной линии напряженности. Очень
медленные колебания происходят при частоте меньше 1 Гц, и их сопровождают тета-,
альфа-, бета- и гамма -волны (θ-, α-, β-, γ-волны). Эти метки являются до некоторой
степени случайными. События с высокой частотой были названы «пульсациями» («ря-
бью»). С правой стороны отдельные нейроны и группировки, насчитывающие несколь-
ко единиц, показывают вспышки до 10 кГц. Порядок от 80 до 150 Гц иногда называют
диапазоном «высоких гамма». (Источник: Bazhenov, Timofeev, 2006.)
382
Глава 8
Большая часть видимой массы внутренней стороны мозга состоит из трак-
тов, состоящих из нервных волокон, покрытых белым веществом: миелинизи-
рованных нейронов и их аксонов. Сейчас показано, что эти магистрали с вы-
соким уровнем движения подчиняются математическим правилам «небольших
мировых сетей», таких как Всемирная сеть в Интернете (World Wide We � ),
которая дает возможности для наиболее эффективного потока сигнала с лю-
бой стартовой точки мозга к любому месту назначения (Achard et al., 2007).
В основном небольшая мировая сеть состоит из каналов информации с высо-
ким уровнем движения и переключательных узлов, наряду с многочисленны-
ми улочками и аллеями для намного более ограниченного локального потока.
Рис. 8 .10 (а). Таламокортикальные нервные тракты. Огромного размера тракты из ак-
сонов нейронов между таламусом и слоями коры. Так как аксоны, выходящие из каждо-
го тела клетки, завернуты в глиальные клетки, заполненные молекулами липидов, они
кажутся белыми для невооруженного глаза, и таким образом, традиционно анатомы их
назвали «белым веществом». Эти тракты были искусственно окрашены на данном изо-
бражении, сформированном с помощью компьютера, чтобы показать различные основ-
ные пути. Все нервные тракты, как кажется здесь, берут начало в левом таламусе, но, по
крайней мере, такое же количество аксонов идет от тел клеток из коры к соответствую-
щим ядрам левого таламуса. Поток в таламокортикальной системе всегда двусторонний
или «возвратный» — основной принцип, который требует объяснений. Заметьте, что на
этом изображении нет потока из противоположного полушария. На следующем рисунке
показаны в равной степени огромные кортико-кортикальные магистрали, соединяющие
полушария друг с другом, в большинстве идущие через мозолистое тело. Как показано
в гл. 3, возможно лишиться целого полушария без потери сознания, если сохранилось
второе полушарие, а также ствол мозга. (Источник: Izhikevich, Edelman, 2008.)
Сознание и внимание
383
Многие сети по своей природе следуют такому оптимально эффективному рас-
пределению связей с высоким и низким уровнями движения.
На рис. 8 .10 (а–в) показаны анатомические основы огромных супермаги-
стральных сетей мозга. На первом рисунке показана таламокортикальная си-
стема в левом полушарии, большое число двусторонних улиц, соединяющих
клетки левого таламуса с клетками левой половины коры. Именно таламокор-
тикальная система, как мы можем видеть, изменяет крупномасштабную актив-
ность при основных состояниях мозга: бодрствовании, сне и сне с быстрыми
движениями глаз. Эти состояния контролируются небольшими ядрами в осно-
вании мозга, как было указано ранее, которые направляют нейромодулирую-
щие аксоны к обширным областям мозга, включая кору.
На втором рисунке показана кортико-кортикальная магистральная система,
в которой движение сигнала идет от одной точки в коре к другой. Имеется в
основном дви типа кортико-кортикальных связей: либо внутри того же полу-
шария, либо из одного полушария в противоположное. Два полушария име-
ют изящный параллелизм в своей структуре связей, так что соответствующие
Рис. 8 .10 (б). Кортико-кортикальные нервные тракты. Огромная взаимосвязность коры.
Как Всемирная сеть (World Wide We�), кора показывает математические методы «не-
большой всемирной связуемости» с магистралями с большой проходимостью и узлов
для переключения направления движения, которые проводят большую часть сигналов,
а также многочисленные местные улицы и аллеи с низким уровнем движения. Большая
часть сигналов, идущих на дальние расстояния, используют супермагистрали, что явля-
ется наиболее эффективным путем для функционирования широкой и плотной сетевой
работы. Заметьте, что два полушария в значительной степени соединены перекрест-
ными связями, в основном через мозолистое тело. (Источник: Human Brain Ma��in�
Meetin�, 2005.)
384
Глава 8
точки от задней части до передней связываются с теми же самыми точками на
другой стороне. Хотя имеются важные функциональные различия между двумя
половинами коры, такие как функционирование языка (обычно поддержива-
ется левой полушарием), наиболее поразительной особенностью с анатомиче-
ской точки зрения является глубокая симметрия двух полушарий.
Имеются другие транспортные системы головного мозга, особенно корти-
коспинальные тракты, которые несут информацию по направлению и из тела
через спинной мозг, а также черепные чувствительные и двигательные нервы.
Зрительная и слуховая системы, обсуждаемые в предыдущих главах, функци-
онируют с помощью черепных нервов, в то время как соматосенсорная и со-
матодвигательная системы идут через спинной мозг. Однако нам нужно только
предоставить наибольшие проводящие пути, чтобы получить хорошее пред-
ставление об обширности карты этого огромного города — головного мозга,
а также его постоянном транспортном движении.
Рис. 8 .10 (в). Спонтанная самоорганизация совокупности клеток в коре. Широко-
масштабная действующая модель огромного количества нейронов и синапсов в коре
и таламусе. Слева показаны только 10% клеток, которые спонтанно появляются в ши-
рокомасштабной модели 100 000 нейронов коры и 8,5 млрд синаптических контактов.
Восемьдесят процентов синапсов были возбуждающими, 20% связей были тормозны-
ми. Возбуждение нейронов воспроизведено с точностью в миллисекунду. Нейроны
были связаны «возвратно» с двусторонним потоком активности, которая обычна для
таламокортикальной передачи сигналов. Импульсы аксонов распространяются за ко-
роткое время, которое зависит от длины аксонов. В модель была встроена задержка при
распространении сигнала, а также было приложено правило Хебба о том, что «нейро-
ны, которые вместе возбуждаются, связаны друг с другом» (гл. 3). Собрания клеток в
модели показали самоорганизацию. Стабильные собрания клеток возникали спонтанно
(помечены цветом слева) вместе с регулярными ритмами групп клеток и активностью,
ограниченной по времени. Модель не имитировала внешние сенсорные входящие сиг-
налы или двигательные исходящие сигналы, но даже чисто внутреннее движение сиг-
налов становилось причиной реалистичных видов активности, похожих на активность
мозга, возникающую спонтанно. (Источник: Iahikevich, Gally, Edelman, 2004.)
Сознание и внимание
385
Помните также, что таламус является основным переключающим узлом (ха-
бом) входящих сигналов для коры, а также основным распределителем движе-
ния сигналов от коры к коре, как огромный аэропорт, который может обслужи-
вать как внутренний, так и международный транспорт. Базальные ганглии тем
не менее действуют как основной распределитель исходящих сигналов для дви-
гательного контроля и исполнительных функций. У мозга есть многочисленные
центры распределения, так же как и многочисленные супербольшие главные
линии связи. Это помогает сконцентрироваться только на самых больших, по-
тому что основные принципы строения в основном одинаковы. Но эволюция
млекопитающих имела 200 млн лет для формирования таламуса и коры над
наиболее древними структурами, такими как ствол головного мозга, спинного
мозга и обонятельного мозга. Имеется огромная избыточность системы, так же
как есть альтернативные пути транспорта в любой большой центральной об-
ласти.
1.6 . Карты и ритмы
Ритмы головного мозга во многом похожи на хронометры и часы. Они под-
держивают способ кодирования времени в головном мозге. Мы уже обсуждали
топографические карты зрения и других чувств, которые обеспечивают способ
кодирования пространства. Таким образом, можно наблюдать как временнóе,
так и пространственное кодирование событий головного мозга, что обеспечи-
вает очень богатый словарь шифрования для мозга.
Сетчатка может рассматриваться как плоская пиксельная карта зрительного
поля (см. гл. 6). Когда лучи света, отраженные от чашки кофе, сходятся в жел-
том пятне — наиболее плотной группе фоторецепторов сетчатки, полагают, что
эта световая модель отражается в многочисленных пространственно-зритель-
ных картах в зрительном отделе мозга. Возможно, существует дюжина разных
пространственно-зрительных карт, каждая из которых представляет зрительное
поле на различном уровне анализа. Мы можем думать об этих картах как об от-
ражающих друг друга, начиная с сетчатки, затем зрительный таламус и область
для первичного отображения зрительных сигналов — зона �1. В гл. 6 указы-
�1. В гл. 6 указы-
1. В гл. 6 указы-
вается, что зрительная кора разделяется на два отдельных потока, называемые
дорсальный и вентральный пути. Однако для наших целей мы можем просто
думать о зрительном мозге как о стопке карт. Это станет особенно важным, ког-
да мы будем обсуждать позже тему внимания (см. рис. 8.11 и рис. 8 .31).
1.7. Двусторонние связи
Большинство магистралей в мозге направлены в две стороны. Мы обычно ду-
маем о зрительной системе как об одностороннем потоке входящих сигналов,
начинающихся от сетчатки, затем идущих в ЛКТ, а затем в первичную зритель-
ную кору (V1) и т. д. В действительности около 90% волокон между ЛКТ и V1
служат двусторонними путями. Это является преобладающей особенности го-
ловного мозга, и понять, как работают двусторонние связи, — сложная задача.
Когда одна и та же топографическая точка активирована в двух или более
пространственно-зрительных картах, вероятно, мы получаем повторно входя-
щий сигнал, как будто много людей со своими собственными картами города
по телефону передают один и тот же набор координат друг другу. Повторная
386
Глава 8
подача сигналов иногда называется «резонансной активностью», и она широко
совместима с повсеместностью ритмов мозга, связывающих топографические
карты. Постоянные ритмы и топографическое картирование могут быть двумя
сторонами одной монеты — биологическим действующим способом, которые
позволяет сигналам переходить между подобными точками на многих слоях
Рис. 8 .11 . Кора как группа топографических карт. Представительства зрения, ощуще-
ний тела, областей исходящих двигательных сигналов, и даже слуха в коре, можно рас-
сматривать как соподчиненность карт (см. гл. 3). Они топографически связаны либо с
сенсорными поверхностями, такими как сетчатка, и/или исходящими массивами, таки-
ми как двигательный гомункулюс коры головного мозга (см. гл. 5). Пространственное
картирование позволяет нейронам, представляющим разные стимулы или характери-
стики исходящих сигналов на различных уровнях обобщения, гарантировать, что они
состоят в близких связях друг с другом, если они относятся к той же точке пространства
или к той же части тела. Такое пространственное картирование дополняется времен-
ными ритмами, которые добавляют способность осуществлять координацию соответ-
ствующих областей и потока событий, происходящих в точно определенные моменты
времени. Совместно топографические карты и кодирование колебаний делают возмож-
ным многие виды пространственно-временного кодирования мозга. 1 — боковые дву-
сторонние связи, 2 — обратные связи, 3 — прямые связи. (Источник: Friston, 2003,
с изменениями.)
Сознание и внимание
387
карт. Ощущения тела и двигательные сигналы, вероятно, работают во многом
по тому же пути, используя топографические карты тела и окружающего про-
странства (см. гл. 5).
Петли повторно входящих сигналов между похожими картами могут усили-
вать или подавлять друг друга. Если все, кто смотрят на карту, в нашей метафо-
ре сходятся на координатах местоположения, ваша уверенность в их сообще-
ниях возросла бы. Каждое подтверждающее сообщение усиливает совместный
сигнал и уменьшает ошибки и шум. Но если люди с картами не сходятся в поло-
жении координат, их взаимная информация будет ослабляться. Активные ней-
ронные карты могут вовлекаться как в тормозящие взаимодействия, так же как
и в возбуждающие. Около 90% нейронов коры являются возбуждающими, но
они не могли бы функционировать без многочисленных тормозящих нейронов
для регулирования уровня своей активности, для того чтобы наложить регуля-
цию ритма на возбуждающую активность и чтобы создать границы тормозящих
участков между островками активности.
Теория нейронного дарвинизма (НД) согласуется с этим представлением
с добавлением пункта, что в НД топографические карты развиваются во вре-
мени до большего и большего соответствия входящих или двигательных исхо-
дящих сигналов (Edelman, 1989). По НД нейроны и синапсы усиливаются, если
между ними образуются успешные связи, и ослабляются, когда они не смогли
этого сделать. Та же точка зрения применима к возбуждающим и тормозящим
связям между конкурирующими группами нейронов. Как и в биологической
эволюции, НД позволяет более «соответствующим» картам выигрывать по
сравнению с менее «подходящими» картами.
Как упоминалось, карты мозга плюс ритмические сигналы составляют про-
странственно-временной код. Как мы видели, телевизионная реклама приме-
няет пространственно-временной код. Если вы посылаете свою фотографию по
сотовому телефону, вы также используете пространственно-временной код. Го-
ловной мозг должен иметь дело с теми же инженерными задачами, с которыми
люди научились справляться, используя компьютеры и телефоны.
1.8 . Как синхронизируются нейроны
Синхронность является всеобъемлющей особенностью головного мозга, не-
сомненно, для того чтобы координировать группы нейронов, находящихся в
разных местах. Действительно, синхронность появляется, даже когда нейроны
культурируют в чашке Петри или когда тонкий срез таламокортикального цен-
тра поддерживается в жизнеспособном состоянии факторами роста. Синхрон-
ная активность, вероятно, является самоорганизующимся свойством нейронов
в своей естественной среде. Однако в мозге синхронность выполняет много-
численные сигнальные функции и, вероятно, находится под гомеостатическим
контролем, как все другие психологические параметры. Детальные обработки
ритмов головного мозга все еще находятся на стадии исследования.
Когда две области головного мозга возбуждаются синхронно с периодом
запаздывания, термин фазовая синхронизация является более точным, чем
синхронность. Звуковые волны, отдающиеся эхом в каньоне, имеют период
запаздывания, но не синхронны, потому что они отражаются назад и вперед
с небольшим периодом запаздывания. Так как нейронам также требуется вре-
388
Глава 8
Рис. 8 .12 . Синхронность в восприятии, внимании и памяти. (а) Показано, как несколько
нейронов, возбуждающихся в одно и то же время, могут суммироваться до гамма-волн в
напряжении электрического поля. Синхронность гамма-волн позволяет добиться более
тонкого временного разрешения, чем альфа-волны. (б) Показано, как могут работать
механизмы внимания путем выборочного усиления групп нейронов, которые управля-
ют целевыми нейронами с частотой гамма-волн. Если вы смотрите на красное лицо на
листе белой бумаги, то те нейроны, которые чувствительны к разнице между красным
и белым, те, которые возбуждаются при высоком зрительном контрасте, и те, которые
отвечают за лица, будут иметь тенденцию синхронно возбуждаться, чтобы сделать воз-
можным расшифровку красного лица головным мозгом. Если вы затем решите обратить
внимание на красное лицо, ваш мозг может увеличить активность тех групп сенсор-
ных нейронов путем возбуждения дополнительных клеток, работающих синхронно с
группами, воспринимающими красное лицо. Голубое лицо не получит этих добавочных
импульсов активности, совмещенных по времени с нейронами, воспринимающими этот
признак, и, таким образом, усилия не будет. (в) Показано, как синхронизированные ней-
роны могут сохранять некоторое время динамическую память о зрительных стимулах
(секунды или минуты). Лицо на образце запускает группы нейронов, чувствительных
к признакам лица, которые возбуждаются синхронно друг другу. Если лицо на образце
было удалено, некоторые их этих нейронов, относящихся к чертам лица, продолжают
возбуждаться несколько секунд или минут. Когда образец для узнавания представляют,
соответствующие нейроны уже возбуждены, чтобы отвечать синхронно узнаваемому
стимулу. Имеются прямые данные для таких механизмов кратковременной памяти. (Ис-
точник: Jansen et al., 2007.)
Сознание и внимание
389
мя, чтобы посылать по своим аксонам импульсы, имеется период запаздыва-
ния в распространении импульса по связанным областям мозга, что приводит,
скорее, к закреплению периода, чем к синхронности. Как синхронность, так и
фазовая синхронизация обычно наблюдаются в головном мозге.
Возбуждающие нейроны могут синхронизировать свое возбуждение, когда
они подчиняются общей тормозящей клетке. Полагают, что синхронизирован-
ные колебания исполняют многие роли в мозге, в восприятии, внимании и памя-
ти (рис. 8 .12 и 8.13). Синхронизированные группы нейронов, возбуждающиеся
на уровне гамма-волн, стремятся вовлечь дополнительные нейроны, возбужда-
ющиеся с той же частотой (Llinas et al., 2005). Более быстрые ритмы предпо-
чтительнее в этом отношении, потому что они делают возможным лучшую точ-
ность во времени: бета- и гамма -волны обеспечиваются нейронными «часами»,
которые могут обнаруживать небольшую разницу во времени по сравнению с
более медленными ритмами. Синхронизированные колебания групп нейронов
отражают путь, по которому действуют отдельные нейроны. Отдельные нейро-
ны могут собирать входящие сигналы от других клеток примерно 10 мс перед
тем, чем образуется потенциал действия. Если два нейрона посылают свои
сигналы к нейрону-цели, который расположен дальше, чем 10 мс, их действие
Рис. 8 .13. Тормозящие клетки синхронизируют возбуждающие нейроны. Как тормо-
зящие нейроны управляют синхронностью возбуждающих клеток. (а) Кодирование
синхронности. Если две возбуждающие клетки (черные) формируют произвольные по-
следовательности импульсов, они редко будут возбуждать общую нижележащую клетку
(серая). В противоположность этому, при синхронизации тормозящим вставочным ней-
роном (красный), эти нейроны, возбуждающиеся с одинаковой частотой, сейчас с боль-
шей вероятностью активируют нижележащий нейрон. (б) Кодирование фаз. Если пять
пирамидальных нейронов (черные) возбуждаются при различных частотах, введение
ритмически действующей тормозящей клетки преобразует частоту в период для перво-
го импульса каждого цикла. Находящаяся под углом красная линия указывает линию
кодирования периода, которая делает возможным этот механизм. (Источник: Mann,
Paulson, 2007.)
390
Глава 8
просто ослабевает, перед тем как добавиться к новому импульсу нейрона-це-
ли (см. гл . 3). Полагают, что этот основной факт определяет верхнюю границу
уровня передачи сигнала от нейрона к нейрону. Относительно быстрые ритмы
бодрствующего состояния, такие как тета-, альфа-, бета- и гамма-волны, нахо-
дятся на том уровне частоты, который набирает добавочные нейроны.
Регулярные ритмы групп нейронов, такие как мы видим на ЭЭГ, вероятно,
требуют как возбуждающие, так и тормозящие нейроны. Пирамидальные (воз-
буждающие) нейроны коры окружены многими тормозящими вставочными
нейронами, которые необходимы, чтобы сформировать постоянные ритмиче-
ские формы волн. На рис. 8 .12 (а) показано, как возбуждение нейронов сум-
мируется в синхронность гамма-волн на ЭЭГ или в локальном электрическом
поле. Синхронность альфа-волн более медленная и, таким образом, имеет
меньшую временную точность.
В принципе, области контроля внимания в мозге могут синхронизировать
свои выходящие сигналы с областями своего воздействия; например, сеть вни-
мания, обнаруженная Познером (Posner) с коллегами, может действововать с
таким же ритмом, как и зрительные карты, воспринимающие лица, в височной
доле (рис. 8 .31). На рис. 8 .12 (б) показано, что увеличение общей синхронности
усиливает сигналы от красного лица и уменьшает активность голубого лица
путем разрушения его синхронной активности во входящих сигналах. Наконец,
на рис. 8.12 (в) синхронная группа нейронов может отвечать на стимул и может
поддерживать действие на несколько секунд или минут после окончания стиму-
ла. Синхронная волновая активность, таким образом, может служить запасом
временной памяти, но она может ослабевать довольно быстро и также может
быть восприимчивой к вмешательству от других стимулов.
Это только некоторые из возможностей кодирования синхронных нейронов
и их групп. Ясно, что это гипотезы, которые требуют, как мы убедимся, под-
тверждающих данных.
1.9 . Синхронность для получения контроля
Синхронность гамма-волн может увеличивать амплитуду импульсов нейрон-
ных групп, так как синхронизированные нейроны могут усиливать актив-
ность друг друга. Синхронность стремится увеличить размер сигнала во мно-
гом так же, как микрофон, который собирает звуки от внешних источников.
В биологических системах такой обратной связи, увеличивающей саму себя,
не обнаружено. Аудиосистемы имеют регулирующие цепи, чтобы предотвра-
тить усиление обратной связь до уровня выше уровня говорящего, не говоря
об ушах слушателей. Эпилептические приступы могут действительно пред-
ставлять собой неправильный тип самоусиления медленных ритмов, которые
вмешиваются в нормальное функционирование мозга и даже приводят к по-
тере сознания.
У нас не должно оставаться ощущения, что бодрствующее состояние полнос-
тью синхронизировано на высоких частотах. Наоборот, синхронность появля-
ется в ограниченных, связанных с задачей, участках мозга, окруженных несин-
хронизированными группами нейронов. Если такая задача, как зрение, требует
согласовать многочисленные зрительные карты, синхронность гамма-волн мо-
жет появиться на многочисленных участках зрительной коры (см. рис. 8 .23).
Сознание и внимание
391
Или, как мы увидим позднее, если находящегося в сознании пациента во вре-
мя хирургической операции на мозге попросят назвать зрительный объект, мы
сможем увидеть кратковременную синхронность гамма-волн между зонами уз-
навания объекта и образования речи.
Необработанная ЭЭГ показывает только волны на поверхности глубокого и
бурного океана. Под видимой ЭЭГ находятся многочисленные потоки колеба-
ний, взаимодействующие по широкому ряду частот (0,01–1000 Гц). Некоторые
из этих волн замкнуты в синхронность друг с другом, некоторые из них имеют
фазовую синхронизацию, многие кратковременно осуществляют согласованные
действия, другие показывают соединение по перекрещивающимся частотам.
Метафора с бушующим океаном полезна для первого приближения, но в отли-
чие от океана в биологическом мозге имеется много функциональной деятельно-
сти, продолжающейся все время. Знания о роли ритмов головного мозга растут
с поражающей скоростью.
На рис. 8.14 целый зрительный стимул (а) может в действительности быть
увиден как подсвечник или как одно лицо, смотрящее на нас (б), или как два
лица, смотрящих друг на друга (в). Таким образом, один и тот же физический
входящий сигнал имеет три трактовки. Каждый воспринимаемый объект вклю-
чает различные способы группировки фона рисунка. Если нейроны, чувстви-
тельные к объекту, уловили подсвечник (объект 1), они могут заставить другие
нейроны синхронизироваться с ним и подавляют другие объекты. Объединение
синхронизированных карт, которые возбуждают наибольший сигнал, может
либо перевесить при голосовании невыделенные объекты, либо активно тор-
мозить их (En�el et al., 2001). Мы увидим ниже прямые записанные с мозга
данные синхронизированного возбуждения нейронов, чувствительных к зри-
тельным характеристикам (рис. 8 .15 и 8.16).
Некоторые авторы полагают, что все колебания мозга — бодрствование,
сновидение и сон — усложнены очень медленными колебаниями, имеющие
период меньше, чем 0,5 Гц (Steriade, 2000). Как мы увидим, имеются прямые
данные для наличия усложняющих гамма-волн на тета-волнах или, возможно,
также гамма- на альфа-волнах. Так как диапазон гамма-волн расширяется до
120 Гц, 200 Гц или даже 1000 Гц, вероятно, что мы увидим намного больше
типов усложнения волн в мозге в процессе улучшения методов записи.
Основным преимуществом усложнения является снижение порогов возбуж-
дения нижележащих нейронов путем уменьшения полярности их мембран (см.
гл. 3). Таким образом, тета-волны могут распространяться до некоторой группы
нейронов-мишеней и делать их слегка более чувствительными, после чего до-
бавленные гамма-волны могут подтолкнуть их выше порога возбуждения. Если
нейроны-мишени способны следовать за входящим сигналом на уровне гамма-
волн, они могут затем производить свои собственные синхронизированные
гамма-импульсы, которые могут, в свою очередь, «зажигать» большие группы
нейронов. Например, показано (�anRullen, Koch, 2003), что зрительные сигна-
�anRullen, Koch, 2003), что зрительные сигна-
, Koch, 2003), что зрительные сигна-
Koch, 2003), что зрительные сигна-
, 2003), что зрительные сигна-
лы могут быть лучше замечены, если они происходят на вершине эндогенной
альфа-волны. Одним из правдоподобных объяснений является то, что многие
области зрительного мозга имеют колебания на частоте альфа-волн во время
состояния отдыха и что основной уровень напряжения большого количества
нейронов спокойно поднимается и опускается с частотой от 12 до 14 Гц. Эта
392
Глава 8
точка зрения на медленные колебания предлагается на обсуждение Стериэйдом
(Steriade, 2000) и другими.
Кажется, что кора имеет некоторое количество микроцепей, которые могут
выполнять более сложные вычисления, такие как определение точного совпа-
дения и даже элементарные логические и арифметические операции, такие как
те, что встроены в основные компьютерные чипы (Ga��ian et al., 2002). Насто-
ящее обсуждение сконцентрировано только на основных сторонах колебаний
нейронных расчетов.
Сейчас мы рассмотрим немного подробнее то, как работает таламокорти-
кальное ядро головного мозга в трех основных состояниях.
Рис. 8 .14. Как синхронизированные нейроны могут связывать сенсорные признаки.
Ритмы мозга могут «связывать» многочисленные особенности зрительных стимулов
в один логически связанный зрительный объект. Мы знаем, что зрительная система
расщепляет физические стимулы на части и преобразует их в характеристики состав-
ных элементов: местоположение, цвет, детали на уровне точек, контрастность краев и
ориентация и, в конечном счете, идентифицирует объект (см. гл . 6). «Тождество объ-
ектов», такое как видение чашки кофе как целого, объединяет все эти характеристики в
единственную сущность целостной формы. На основании этого представление об объ-
екте «связывает» многочисленные зрительные характеристики. Иногда это называют
«проблемой связывания». Она связана с вопросом о зрительном осознании, так как мы
стремимся осознать скорее целые объекты и события, чем отдельные черты, парящие в
пространстве. (Источник: En�el et al., 2001.)
Сознание и внимание
393
Рис. 8 .15. Более быстрые волны усложняют структуру медленных волн. Быстрые вол-
ны могут «усложнять» вершину более медленных волн подобно амплитудной модуля-
ции радиосигналов. Это известно под несколькими другими названиями. И, вероятно,
является общей линией для кодирования сигналов в мозге. (Источник: �anRullen, Koch,
2003.)
Рис. 8.16 . Бодрствование: стимуло-специфичное отражение во время раннего обуче-
ния. Динамичная нейронная цепь для первой фазы обучения. Во время бодрствующего
состояния предполагается, что рабочая память поддерживается повторяющейся цепью,
включающей кору, таламус и базальные ганглии (стриатум), усиленной нейромодуля-
цией, начинающейся в черной субстанции (ЧС) и бледном шарее (БШ). В то время как
зрительный входящий сигнал поступает в кору в затылочную долю, полагают, что объ-
единение в одно целое зрительных объектов происходит в нижней височной доле (назы-
ваемой зоной TE в мозге макаки, показанном здесь). Включение теменной доли требует-
ся для пространственной локализации зрительных событий на пространстве тела и для
того, чтобы протянуть руку и манипулировать ей. Включение предлобной зоны необхо-
димо для всеобщей интеграции, сознательного внимания и двигательного контроля (ис-
полнительные функции). Заметьте, что глутаминовая кислота и гамма-аминомасляная
кислота (ГАМК) показаны двумя основными нейромедиаторами. Стабилизация этой
цепи может потребовать �-метил-D-аспартатных (НМДА) рецепторов для увеличения
эффективности активных синапсов. Комплекс гиппокампа (не показан) требуется для
случайного обучения и, по прошествии времени, для семантической памяти. Укрепле-
ние памяти во время медленноволнового сна необходимо для становления постоянной
памяти путем синтеза белков, которым и создаются новые синапсы и другие клеточные
структуры, необходимые для поддержания долговременной памяти. (Источник: Wan�,
2001.)
394
Глава 8
2.0 . Бодрствование
Содержание нашего бодрствующего сознания включает непосредственный мир
восприятий, внутреннюю речь и совокупность зрительных мысленных образов,
мимолетное настоящее и его исчезающие следы в текущей памяти, ощущения
тела, таких как удовольствие, боль и восхищение, автобиографическая память,
когда вспоминают кого-то, ясные и непосредственные намерения, ожидания и
действия, определенные верования о себе и о мире, а также идеи абстрактные,
но находящиеся в центре мышления. Очевидно, что мы осознаем намного боль-
ше, чем можем рассказать в любое время, но люди могут отвечать на вопросы
о поразительном диапазоне сознательных событий — о виде одинокой звезды
в темной ночи, разнице между звуками «ба» и «па», а также о цели подготовки
к экзаменам. Все сенсорные системы производят обработку множества бессоз-
нательной информации, что приводит к осознанным зрительным образам, зву-
кам и чувствам, которые можно передать другим.
Активизация автобиографической памяти также приводит к осознанному
опыту, такому как память о том, что эту книгу вы видите в первый раз. Внутрен-
няя речь и совокупность мысленных образов имеют сознательные стороны.
Просьба к кому-либо перечислить список слов обычно приводит к осознанной
внутренней речи. Яркость зрительных образов людей различается, например,
традиционно сообщается, что сны имеют яркую совокупность зрительных об-
разов (Stick�old et al., 2001). Наконец, планирование действий может иметь
сознательные компоненты, особенно когда нам нужно сделать выбор между
действиями (Lau et al., 2004 а, 2000 б). Обычно мы не должны много думать
о ходьбе, но если мы сломаем ногу, то даже процесс вставания и ходьба могут
стать сознательно планируемыми действиями (Sacks, 1984).
Есть надежные данные, что у нас имеется самопроизвольный внутренний
поток содержимого сознания, который борется с воспринимаемым миром
(Sin�er, 1994; Mazoyer et al., 2001; Fox et al., 2005). Таким образом, в повседнев-
ной жизни имеется продолжающаяся борьба между внутренними и внешними
событиями, между воспринимаемым и ответом на него, а также между одно-
временными внутренними потоками. Выбор с помощью внимания, кажется,
действует в каждый момент бодрствования.
2.1. Практическое мышление
Общеизвестно, что сны нереалистичны, но бодрствующее сознание долго счи-
талось областью логического, зрелого, ориентированного на реальность мыш-
ления. В этом есть значительная правда, хотя мы также обычно увлекаемся
фантазиями наяву, мечтаниями, мыслями, направляемыми эмоциями, и блуж-
данием по памяти. Иногда трудно полностью сконцентрироваться только на
одной задаче, и для этого могут быть хорошие биологические причины. Люди
не эвлюционировали в мире школьных классов и путем изучения книг. Охот-
ники и собиратели палеолита должны были постоянно находиться в состоянии
повышенной готовности к неожиданным опасностям и возможностям; концен-
трация полностью на одном деле и исключение всех других могли быть опас-
ны. Люди, скорее, имеют склонность слегка исследовать чувствительный мир,
чем упрямо следовать единственной цели или единственной цепочке мыслей.
В то же время наш большой мозг дает нам гигантскую приспособляемость.
Сознание и внимание
395
Мы учимся контролировать наше внимание, по крайней мере в некоторые пе-
риоды времени, а традиционные культуры обучают детей при помощи задач,
требующих познания, таких как охота, нахождение пути, распознавание сотен
растений, путешествие на длинные расстояния, делание инструментов, приго-
товление пищи и соблюдение многочисленных социальных обычаев. Эти при-
вычки требуют обучения и внимания при исполнении. Они также дают награду
за реалистичное понимание мира. Однако основы «реализма» мозга до сих пор
не были эффективно исследованы. В результате мы не понимаем, как мы можем
смотреть фильм ужасов без испуга (потому что мы знаем, что это иллюзия) и
сравниваем его с действительным опытом. С теми инструментами, которые мы
имеем сегодня, к этому вопросу можно обращаться на основе опыта.
В шатком мире требуется, чтобы самоконтроль не позволял таким чувствам,
как страх, голод, одиночество или гнев, преобладать. Дети в собирательно-
охотничьих культурах часто обучаются выдерживать нужду и эмоциональный
стресс. Наш мозг развивается, чтобы справляться с такими условиями. Только
очень недавно люди стали жить более спокойной жизнью, с надежными ис-
точниками пищи, социальной поддержки и защиты от опасности и неудобств.
Бодрствующий мозг был описан как «уравновешенный на точке перехода
между упорядоченным и случайным поведением (табл. 8 .1). Это тонкое состо-
яние «оптимизирует передачу и запасание информации ... (включая) как про-
должительные периоды фазовой синхронизации, так и присутствие больших
быстрых изменений в состоянии всеобщей синхронизации ...» (Kitz�ichler
et al., 2009). Бодрствующее таламокортикальное состояние оптимизировано
Таблица 8.1
Основные черты сознательного бодрствующего состояния
1. Очень широкий диапазон содержания сознания
2. Широкораспространенные влияния мозга за пределами специфического
содержания сознания
3. Информативное содержание сознание
4. Быстрая приспособляемость и быстротечность
5. Внутренняя логичность
6. Ограниченная вместимость и периодичность
7. Связь с сенсорной системой
8. Власть над собой
9. Аккуратная регистрируемость
10. Субъективность
11. Структура центр-периферия
12. Сознание облегчает обучение
13. Стабильность сознательного содержания
14. Центрированный на объектах и явлениях
15. Сознательность полезна для осуществления произвольных решений
16. Вовлечение таламокортикального центра
(Источник: Seth, Baars, 2005.)
396
Глава 8
для объединения таких возможностей. Синхронность ритмов и фазовая син-
хронизация позволяют мозгу соотносить друг с другом различные временные
шкалы. Таким образом, пикколо в оркестре головного мозга может играть со-
гласованно с басовым барабаном.
Последнее десятилетие принесло лавину открытий о ритмах мозга в сенсор-
ной и двигательной системах, в системе памяти гиппокампа, выборочном вни-
мании и исполнительном контроле. Эти открытия были сделаны при помощи
прямых записей активности мозга животных и человека, из ЭЭГ и МЭГ, а так-
же от объединения этих методов электромагнитной записи с пространствен-
ными преимуществами функционального магнитно-резонансного томографа.
Сцепленные ритмы наблюдались в отдельных нейронах, в нейронных цепях
и множествах, и даже в целых лобных долях. Ритмическая активность в коре
головного мозга является наиболее ясным признаком различных состояний со-
знания, а недавние исследования предполагают, что отбор с помощью внима-
ния действует также путем синхронизации ритмов головного мозга. Состояние
глубокого сна с медленными колебаниями, видимо, посвящено укреплению
воспоминаний сознательного опыта в предшествующий день. Как мы увидим,
даже сон, возможно, использует быстрые ритмы во время пика медленной дель-
та-волны, чтобы закрепить сознательный опыт предшествующего дня.
В табл. 8 .2 представлены различные типы познания, опосредованного со-
знанием — в большей степени все процессы, которые мы изучаем. Обратите
внимание на наличие бессознательных сторон у этих частично сознательных
задач. Как сознательный, так и бессознательный компоненты, вероятно, явля-
ются ключевыми для нормального, гибкого и приспосабливающегося позна-
ния. Однако подсознательное восприятие и познание, например, могут присут-
ствовать для высоко значимых и стереотипных стимулов.
2.2 . Ориентация по месту, времени и людям
Наши основные знания о сознательном опыте получены из того, что люди могут
рассказать нам (см. гл . 2). Когда клинически исследуют пациента с возможны-
ми повреждениями мозга, нейропсихолог начинает с проведения «мини-иссле-
дования психического статуса». Оно разъясняет, что мы ожидаем от человека,
находящегося в сознании, со здоровым мозгом (McDou�all, 1990).
1. Ориентация во времени, месте и личности: «Какой день недели, дата или
время года?» «Где вы находитесь сейчас?» «Кто вы?» «Кто я?»
2. Простая рабочая память: «Запомните эти три предмета...» .
3. Внимание и вычисления: «Посчитайте в порядке убывания через 7, начиная
от 100» (100, 93, 86, 79, 72...).
4. Вызов промежуточной памяти: «Какие три предмета я упоминал перед
этим?»
5. Язык: Попросите пациента назвать наручные часы и карандаш, повторить
предложение, выполнить три простых команды, прочитать и понять простое
предложение, а также записать свое предложение.
6. Основные зрительно-моторные навыки: повторение рисунка контура пяти-
угольника.
Сознание и внимание
397
Таблица 8.2
Познание, опосредованное сознанием
Задача
Сознательные аспекты
Бессознательные аспекты
Рабочая память
Сенсорные входящие сиг-
налы, планирование ответа
и исходящие сигналы; кра-
тковременный повторяе-
мый в уме элемент
Предвосприятие, неповторя-
емые элементы, базальные
ганглии
Самопроизвольное
решение проблемы
Определение
проблемы,
реализация решения (опыт
«Ага»!)
Скрытый период существова-
ния проблемы
Объединение сенсор-
ных признаков, задняя
кора
Результаты связывания
Детальный процесс интегра-
ции
Интеграция сенсорных
и
пространственных
сигналов,
теменная
доля
То же самое
То же самое
Сенсорно-двигатель-
ные умения (например,
речь)
Акустическая фаза и вне-
сение изменений в обыч-
ные задачи
Предсказуемые компоненты
после интенсивной практики
Сравнение сознательно-
го опыта, который про-
исходил в разное время
Новые воспоминания и но-
вые связи
Хорошо установленные вос-
поминания и порядок осу-
ществления действий
Произвольное внима-
ние
Контроль внимания, тре-
бующий усилий, или под-
держание внимания во-
преки противоположным
устремлениям, таким как
усталость, дремота, или
более
привлекательным
объектам внимания
Повседневное,
автоматиче-
ское, лишенное конкуренции
и высоко предсказуемый кон-
троль внимания
Автоматическое внима-
ние
Результаты внимания
обычно осознанные
(но не всегда)
Детальный контроль автома-
тического внимания
Скрытое обучение
Сознательные сигналы к
скрытым закономерностям
или заключениям
Автоматическая или скрытая
информация, которую изу-
чают
Обратите внимение, что нормальный произвольный контроль подразумева-
ет все эти задачи. Люди с нарушениями лобной доли могут иметь трудности
с их выполнением из-за ослабленных исполнительных функций. Так как эти
вопросы кажутся фундаментальными, они покрывают сенсорное восприятие,
произвольный контроль, рабочую память, мгновенную и промежуточную па-
мять, извлечение из памяти через длительный промежуток времени, мыслен-
ное усилие, речь, внимание к повседневным событиям, постоянное обновление
памяти с течением времени, мысленные образы, пространственные навыки и
398
Глава 8
многое другое. Все эти задачи требуют бодрствующего состояния (живость по-
нимания) и познания, опосредованного сознанием. Они снижаются при дре-
моте, отклонении внимания, медленноволновом сне и сне с быстрыми движе-
ниями глаз, а также при применении лекарств и нарушениях мозга, которые
ухудшают сознание.
В норме бодрствующее состояние является необходимым условием для всех
процессов познания, которые мы обсуждаем в этой книге.
2.3 . Бодрствование для обучения;
сон делает возможным закрепление памяти
Хотя бодрствование безоговорочно является лучшим временем для обучения,
сон и сновидения с быстрыми движениями глаз усиливают закрепление па-
мяти. Предполагается, что воспоминания нестабильны и уязвимы к помехам
в первые часы после обучения. Полагают, что ранняя память зависит от ак-
тивных, резонирующих нейронных цепей (см. рис. 8 .16). Однако некоторые
исследователи полагают, что бодрствование может также позволить некоторое
закрепление памяти. Примерно через день выученный материал оказывается
«закрепленным» — процесс, который, как полагают, требует синтеза белков.
Устойчивые воспоминания включают структурные изменения в синапсах и
клетках, и иногда в сети головного мозга в целом. Сон, по-видимому, усиливает
укрепление памяти с помощью синтеза белков, что требует экспрессии генов
в многочисленных клетках мозга.
Хотя все состояния сознания имеют многочисленные функции, осознанное
бодрствование является безоговорочно наиболее адаптивным. Как мы увидим в
гл. 9, каждая область мозга имеет свою собственную способность к приспосо-
блению и к обучению. Таким образом, имеется много типов обучения. Но для
изучения новых типов информации и передачи ее памяти мы постоянно делаем
вещи осознанными. В повседневной жизни это то, что мы подразумеваем под
словами «обращать внимание». Исследователи памяти показали, что большая
часть обучения является «случайной», т. е. оно происходит как выгода от об-
ращения внимания (Krist��nsson, 2006). Мы обучаемся во время большей ча-
Krist��nsson, 2006). Мы обучаемся во время большей ча-
�nsson, 2006). Мы обучаемся во время большей ча-
nsson, 2006). Мы обучаемся во время большей ча-
, 2006). Мы обучаемся во время большей ча-
сти бодрствования. Это можно показать, используя чувствительные измерения
эпизодической (эмпирической) памяти. Но, конечно, мы не преднамеренно за-
поминаем вещи в каждый момент бодрствования. Запоминание — только один
путь, чтобы произошло обучение. Большую часть времени, по-видимому, мы
запоминаем вещи, только обращая внимание на них, — это означает, что они
для нас становятся осознанными. Исследователи памяти со времен Германа Эб-
бингауза (Hermann E��in�haus) указали на центральную роль, которую играет
осознанное внимание при нормальном обучении. Хотя имеются некоторые дан-
ные для бессознательного приобретения информации, мы сконцентрируемся на
значительной роли осознанного внимания в мобилизирующих гиппокампаль-
ных и кортикальных механизмах обучения (см. гл. 9).
2.4 . Внимание и сознание в общем
делают возможным обучение
Как происходит, что вы можете выучить материал этой книги? Человек в дей-
ствительности не знает. То, что мы делаем в повседневной жизни, — это просто
обращение внимания на новые и интересные вещи, исследование их с помощью
Сознание и внимание
399
наших чувств, мысленное их повторение и опять направление нашего внима-
ния на них. Магическим образом, кажется, происходит обучение. На следую-
щий день мы внезапно осознаем, что вчерашняя новая информация кажется
более знакомой. Должно быть, мы ее выучили. Ориентирующие задачи важны,
чтобы сделать возможным изучение. «Простого предоставления» информации
часто достаточно для запуска памяти при опознании. В терминах когнитивной
науки, большая часть нашего ежедневного познания является случайной.
Что мы в общем делаем — это просто обращаем внимание на новый матери-
ал, даже если он кажется трудным для понимания. Самой большой проблемой
является обратить внимание на новую и трудную информацию, а также быть
достаточно терпеливым, чтобы позволить нашему мозгу изумляться, задавать
вопросы и, в конце концов, осмыслить любой новый материал. Лишь только мы
восприняли и осмыслили что-то новое, изучение произошло. Данные по иссле-
дованию мозга указывают, что самопроизвольное обращение внимания на не-
которое сенсорное содержимое также активирует гиппокампальный комплекс
(Stark, Okado, 2003).
Мы можем направлять наше внимание произвольно, хотя иногда это требует
заметного усилия (см. гл. 12). Выборочное внимание в общем случае приводит
к сознательному опыту (ранее) и кажется действием, которое делает предметы
осознанными, что обусловливает изучение. Действительно, «эпизодическое и
объявленное изучение» определяется как «изучение осознанных событий», и
мы знаем, что гиппокамп, вероятно, особенно приспособлен к этому (Squire
et al., 2008; см. гл . 9). Хотя имеются надежные данные для подсознательного
восприятия и изучения, мы ограничим это обсуждение эпизодической и декла-
ративной памятью сознательных событий.
Научные работники, занимающиеся познавательной нейробиологией, пола-
гают, что декларативное и случайное изучение в общем случае происходит как-
то так (Seitz, Watana�e, 2005): мы обращаем внимание на новую информацию,
так что она стремится стать осознанной. Как только мы узнаем на опыте новую
информацию с достаточной ясностью, наш мозг становится в состоянии запа-
сти ее. Повторное внимание к новому и трудному материалу часто необходимо
перед тем, как у нас возникнет чувство ясности. Используя чувствительные ме-
тодики измерения эпизодической и декларативной (сознательной) памяти, та-
кие как измерение распознавания объекта, мы можем показать, что люди очень
быстро учатся после того, как они получили четкий сознательный опыт новой
информации. Имеются надежные данные подсознательного обучения, но здесь
мы сконцентрируемся на изучении сознательных событий или на материале,
стимулом которого являются осознанные события.
Это не значит, что мы можем вызывать все воспоминания по сигналу. Это
означает, что мы можем распознавать события сознательного опыта, которые
случились давно. Тесты на распознавание легкие, потому что они просто вос-
станавливают тот же самый опыт, который мы уже получили. Мы совсем не
обязаны по своему выбору искать наши воспоминания. Все, что мы должны
сделать, — это сказать: «Да, я видел это раньше» или «Нет, я этого не видел».
Простой сознательный опыт материала, который мы должны опознать, запу-
400
Глава 8
стит автоматический поиск воспоминаний. Решение да/нет делается с исполь-
зованием осведомленности о предмете.
Например, вы можете быть удивлены, когда узнаете сцену в фильме, кото-
рый смотрели 10 лет назад. Для вас не было необходимо запоминать эту сцену.
Все, что вы должны были сделать, — это просмотреть ее однажды — осоз -
нанно, а затем посмотреть ее опять 10 лет спустя. Осознание события кажется
достаточным, чтобы оставить его в памяти. Во многом это правильно для рас-
познавания фотографий ваших школьных товарищей из альбома выпускного
класса, новых фотографий, заголовков спустя годы и всего подобного.
К несчастью для студентов, учебные экзамены редко связаны с тестами на
распознавание. Скорее, экзамены проверяют воспоминания по ассоциациям пу-
тем задавания вопросов, таких как «Какой город является столицей Франции?».
Экзамены в учебном заведении были бы намного проще, если бы на них дава-
лась часть ответа, такое как частичное сообщение для распознавания: «Пар...
является столицей Франции».
Задачи по вызову воспоминаний по ассоциациям дают намного более низ-
кие оценки точных воспоминаний, чем тесты на опознание. Вот почему учеб-
ные экзамены трудны: проверяемые знания не являются нашими отложенными
воспоминаниями, которые имеют недостатки, но экзамены — это наша спо-
собность находить их по требованию. По аналогии, вы можете хранить книгу
случайным образом в огромной библиотеке. Книга (память) находится где-то
там, но вы не можете найти ее по требованию до тех пор, пока вы не будете
иметь очень хорошую систему нахождения, такую как механизм для поиска
или каталог карточек. В противном случае книга может быть также потеряна
навсегда.
Заметьте, что простое обращение внимания не всегда делает вещи осознава-
емыми. Например, мы можем обращать внимание на двусмысленную картинку,
но не видеть все ее интерпретации (см. гл. 1). В результате мы можем никогда не
узнать бессознательную интерпретацию картины в качестве сознательного слу-
чайного воспоминания. Изучение сложного объекта, такого как наука о головном
мозге, часто подобно изучению двусмысленного стимула и никогда нельзя уви-
деть интерпретацию, к которой у вас не было доступа. Вначале новый материал
может казаться неясным, трудным для понимания или сбивающим с толку. Затем
мы можем провести некоторое время, обдумывая его, обращая на него внимание,
пытаясь нарисовать его от руки или отвечая на вопросы по нему. Со временем
будет приходить чувство более ясного понимания. После того как мы стали ясно
осознавать требуемую информацию, быстро последует изучение. Большая часть
усилий в изучении нового материала, таким образом, посвящена задаче осмысле-
ния. Как только мы ясно поймем материал, начнется запоминание.
Итак, обращение внимания может быть средством к осознанной четкости,
что в свою очередь делает возможным случайное и декларативное изучение.
Полагают, что гиппокампальный комплекс превращает сознательный опыт в
память. Вначале гиппокампальная память, вероятно, является нестабильной
или лабильной. Закрепление способствует образованию долговременных из-
менений в гиппокампе и переводу познаний в новую кору. Полагают, что тета-
колебания играют важную роль как в локальных функциях гиппокампа, так и в
долгосрочной координации между гиппокампом и новой корой.
Сознание и внимание
401
2.5 . Потеря осознания предсказуемых событий
Хотя обращение внимания на стимул часто приводит различными путями к его
превращению в опыт, имеется основной пример, когда происходит противопо-
ложное: случай обыденных привычек. Как правило, чем более предсказуемым
становится сенсо-моторный навык, тем менее он будет сознательным. Это об-
щепринятое наблюдение над обучением ездить на велосипеде или даже обуче-
нием говорить на новом языке. Уменьшение доступа сознания к привычным
умениям в общем случае называют «автоматизмом», и он сопровождается по-
терей точного сознательного контроля над обычными деталями (см. гл. 2, раз-
дел 5.1).
Мир вокруг нас становится более предсказуемым по мере того, как мы рас-
тем и учимся понимать его. Если бы все было полностью известно и предска-
зуемо, мы, скорее всего, не должны были бы совсем обращать на это внимание.
Наш мозг мог бы все делать автоматически. К счастью, реальный мир полон
изменений и трудных задач, и мы обнаруживаем, что постоянно выбираем наи-
более важные и новые события, на которые нужно обратить внимание. Поток
сознания весь состоит из наших личных заголовков новостей. Мы имеем склон-
ность останавливаться на чем-либо новом или значимом для нас.
Повторяющиеся события имеют тенденцию постепенно исчезать из созна-
ния до тех пор пока не будут иметь особую значимость. Зуд от укуса комара не
может быстро исчезнуть, так как слюна самки комара запускает болевую систе-
му. Боль в своей основе важна, так же как голод или вкус шоколада. Мы не бы-
стро привыкаем к биологически значимым событиям, но нейтральные события
могут исчезнуть из сознания довольно быстро, иногда за секунды. Даже если
гравитация постоянно запускает наше вестибулярное чувство (равновесия), мы
редко обращаем на него внимание, так как оно стало предсказуемым. Тело по
отношению к силе гравитации ставится опять в центр внимания у космонавтов,
парашютистов и детей, которые катаются на колесе обозрения.
Шнайдер (Schneider, 2009) пишет:
Понятия автоматизма, контролируемой обработки информации и сознания тесно
связаны... . Автоматическая обработка информации ... поддерживает параллельный
процесс, требующий слабых усилий, который трудно контролировать. Контролируе-
мая обработка информации происходит из области «общей сферы», которая выпол-
няет последовательную, требующую усилий, обработку в новых или изменяющихся
ситуациях. Однако автоматические процессы в форме автоматического внимания
могут активировать сознательное понимание стимулов, которые были еще не об-
работанны.
Последний пункт очень важен. Мы теряем сознательный доступ к автома-
тическим стандартным операциям, но некоторые из них являются стандартны-
ми операциями с привлечением внимания, т. е. они происходят, чтобы довести
положение вещей до сознания. Хорошим примером являются автоматические
движения глаз. Многие движения глаз являются автоматическими, и в то время
как они представляют двигающийся грузовик в наше поле зрения, изображение
грузовика становится осознанным.
В этих случаях, конечно, автоматизм фактически делает возможным то, что-
бы важная информация стала осознанной. Но в более общем случае практи-
ка приводит к автоматизму и к потере сознательного доступа к умению, часто
402
Глава 8
использующемуся на практике. Большая часть нашей повседневной мысленной
жизни требует автоматических повседневных дел, которые невидимы для нас,
но которые делают возможными наши повседневные дела.
Мозговые исследования практических навыков показали, что фМРТ-
изображение радикально изменяется, когда навык не требует сознания. На
рис. 8 .17 показано сканированное изображение головного мозга одного челове-
ка до и после обучения выполнению прогнозируемой задачи поиска на слух до
уровня автоматизма. В результате практики и обучения наблюдается, что сиг-
нал фМРТ (на BOLD-контрасте, зависящем от уровня кислорода в крови) целой
сети корковых областей опускается ниже порога обнаружения. Такие заметные
снижения в активности коры обычно наблюдаются при работе над часто вы-
полняющимися предсказуемыми задачами.
Однако часть слуховой коры остается активной даже после того, как задачи,
при которых происходит сканирование, становятся автоматическими, а имен-
Рис. 8 .17. Активация на фМРТ до и после практики. Контролируемые и автомати-
ческие варианты одной и той же задачи можно сравнить у одного и того же человека.
Когда задача по слуховому поиску новая, она позволяет осуществлять более точный
сознательный контроль, больший доступ к сознанию и более широко распространен-
ной активации коры. Коул и Шнайдер (Cole, Schneider, 2008) показали, что обработка
информации под контролем сознания включает сеть областей коры, которые показаны
выше. Они, как и следовало ожидать, разделяются на активацию лобной доли (спереди
от центральной борозды) и зависящую от уровня кислорода в крови BOLD-активность
(от англ. blood-oxygenation-level-dependent) в задней части теменной доли. Те же лобные
и теменные зоны активны при сознательной, по сравнению с бессознательной, обработ-
ке стимулов. Эти области называются «общей сферой», потому что они функционируют
в разных задачах, которые используют зрительные или слуховые стимулы, независимо
от того, простыми или сложными являются стимулы или задачи. (Источник: Schneider,
2009.)
Сознание и внимание
403
но — верхний задний гребень задней височной доли. Причина в том, что слу-
ховые входящие сигналы продолжают поступать для анализа. Однако локаль-
ное возбуждение коры, кажется, не распространяется широко, как, вероятно,
при полностью осознаваемых событиях (см. рис. 8 .17).
Изображение рис. 8 .17 вызывает несколько важных вопросов. Один из них
о сети, находящейся под произвольным контролем, которую мы более детально
обсуждаем в гл. 11 и 12. Как можно ожидать, некоторое количество областей
коры, связанных с контролем, группируется вокруг полосы моторной коры, как
раз впереди от центральной борозды. Они включают дорсолатеральную пред-
лобную область (дл-ПлК), часть островка (передний островок), добавочную
премоторную кору (ПМК), дПМК (дорсальную премоторную кору) и нижнее
лобное соединение (НЛС). Как раз над вершиной горы полушария мы видим
активность премоторной коры (дорсальная часть) и переднего пояса (ППК).
Легче думать о всех этих областях как о разных уровнях моторных карт, од-
них — очень специфичных для мышц, а других — на более высших уровнях,
включая просто «побуждение двигаться». Дл-ПлК включена во все произволь-
ные задачи, требующие усилий. Заметьте также, что в «говорящем» полушарии
эти области граничат с зоной Брока, областью коры, воспроизводящей речь, ко-
торая находится близко к моторной коре рта и языка. Эти области лобной доли
имеют сложные названия, но все они подчиняются простому правилу, так как
все они лобные и тесно вовлечены в контроль действий.
Обратите внимание, что эти лобные зоны используют обе поверхности каж-
дого полушария: латеральную (внешнюю) и медиальную (внутреннюю) по-
верхности. По существу они покрывают обе стороны того же холма. Так как
кора в основном является одной огромной поверхностью, ее холмы и долины
не так важны, как продолжающаяся поверхность сама по себе.
Наконец, мы видим BOLD-активность позади этих лобных зон в ЗТК, задней
теменной коре. Вы можете думать об этой области как о центре планирования
действий, начиная с коры более примитивных приматов, до того, как лобная
доля человека не выпятилась так далеко наружу, как сейчас. Так как в мозге со-
держится его собственная история, мы можем думать о человеческом мозге как
об имеющем два центра контроля действий. Оба центра поставляют сигналы в
полосу моторной коры, которая имеет связи с большей частью направлений с
двигательными нейронами, идущими вниз по двигательным путям к спинному
мозгу и черепно-мозговым нейронам для регуляции лица, головы и шеи. Так
как обширное увеличение вперед лобных долей — довольно позднее событие
(на эволюционной временной шкале), мы используем как задний теменной, так
и предлобный центры контроля. Задняя теменная кора вовлечена в планиро-
вание, создание мысленных образов и сенсо-моторную интеграцию. Все эти
функции участвуют в контролируемых (произвольных) процессах, предсказуе-
мая задача становится менее сознательной (Coulthard et al., 2008).
Перестал ли мозг вообще производить обработку поиска на слух в автома-
тически выполняемой задаче (предъявляемой с правой стороны (рис. 8 .17))?
Ясно, что нет. В то время как кора намного менее активна, автоматическая по-
вседневная работа контролируется базальными ганглиями, мозжечком и реф-
лекторными цепями спинного мозга. Они могут все еще использовать чувстви-
тельные входящие сигналы, что обозначено активной слуховой корой, но такие
404
Глава 8
петли с интенсивным движением информации становятся практически полно-
стью подкорковыми. Эти процессы почти бессознательны или намного менее
сознательны с намного менее точным произвольным контролем и меньшей кор-
ковой активностью.
2.6 . Скрытое обучение также требует сознания
Термин скрытое обучение может вводить в заблуждение. Оно не означает об-
учение без участия сознания. Более того, скрытое обучение включает осознан-
ные входящие сигналы, что приводит к скрытым (бессознательным) выводам,
которые выучиваются вместе с сознательными сторонами задачи.
Когда мы практикуемся в сложных сенсомоторных навыках, таких как
спорт, танцы, печатание, чтение или игра на музыкальных инструментах, мы
не осознаем все детали задачи. Скорее мы делаем только часть задачи осознан-
но и позволяем автоматическим сторонам бессознательно присоединяться. То,
что ученые называют «явным», «случайным» или «декларативным» обучени-
ем, включает сознательное обучение, но в данном случае происходит не исклю-
чительно сознательное обучение. Сознательные и бессознательные процессы
часто идут вместе. То, что формально называется «скрытое (бессознательное)
обучение», часто идет как побочный эффект сознательных входящих сигналов.
Сознательные события могут в целом направляться процессами научения, даже
для скрытого обучения.
Например, грамматика нашего первого языка изучается скрытно (бессозна-
тельно). Маленькие дети знакомятся с языком с потрясающей скоростью, но им
нет никакой необходимости знать подробно, какие слова являются существи-
тельными, какие глаголами или наречиями. Дети обращают внимание на новые
слова и предложения и, конечно, начинают их осознавать, когда сами говорят
или слушают. Хорошие воспитатели постоянно обращают внимание детей на
новые слова и новый опыт. Таким образом, сознание постоянно вовлекается в
обучение языка, но закономерности грамматики, вероятно, изучаются скрыто,
как побочный выигрыш обращения внимания и процесса осознания фактов.
2.7. Быстрые ритмы координируют задачи
в состоянии бодрствования
Нативная ЭЭГ бодрствования часто называется «бета-» или «гамма-активно-
стью», но фактически является сложной и переменчивой смесью множества
периодических и непериодических сигналов. Нативную ЭЭГ можно предста-
вить как море с мелкими волнами. В науке возможность анализировать слож-
ные волновые формы с большой скоростью появилась недавно, что связано с
анализом Фурье и сходными подходами на мощных компьютерах. В итоге на-
учные публикации в последнее время редко используют необработанную ЭЭГ.
Это может привести к большой путанице, ведь мы по-прежнему продолжаем
воспринимать ЭЭГ как чисто волновые формы. Если следовать самым послед-
ним тенденциям, то, говоря «бета» или «гамма», мы на самом деле говорим о
диапазоне частот, приблизительно от 13 до 25 Гц для бета, и от 26 до 120 Гц
для гамма. Не существует общепринятых границ между стандартными диапа-
зонами частот, и возможно не следует вводить неизменные числа до того, пока
мы не будем лучше понимать суть явления. В ходе новых исследований выяв-
ляются новые волновые диапазоны, играющие значительную роль в головном
Сознание и внимание
405
мозге. Так, например, недавно обнаружено, что важны высокочастотные (до
1000 Гц) всплески и что волны с очень низкими частотами, до 0,01 Гц, также
присутствуют как при бодрствовании, так и во время сна. Помните, что ритмы
головного мозга — это подвижный объект, так как новые факты появляются
крайне быстро.
В ходе эволюции уже существующие биологические механизмы исполь-
зуются для выполнения новых функций. Древние части мозга, такие как гип-
покамп — часть маленького мозга рептилий, — также обладают гамма- и
тета- (4–7 Гц) волнами. Вполне вероятно, что неокортекс, мощно развившаяся
«новая кора» млекопитающих, позаимствовала указанные древние ритмы, и им
нашлось новое применение. Основные ритмы мозга повсеместно встречаются
в царстве животных. Они отличаются высокой консервативностью среди раз-
ных видов, что позволяет предполагать их важную функцию.
Сейчас высокие уровни гамма-осцилляций показаны для сенсорной обра-
ботки, усиления вниманием сенсорного входа, и для рабочей и долговременной
памяти. В недавней работе продемонстрированы и высокая гамма-активность,
и высокая степень гамма-синхронизации, одновременность в подъемах и спа-
дах ритма в двух разных областях мозга на частотах 20–30 Гц. Синхронизация
естественна и полезна для сигнальных функций в системе с осцилляциями,
такой как мозг. В некоторых случаях полная синхронизация недостижима, и
существует краткий интервал между пиком волны в одном месте (например, в
гиппокампе) и другом месте (например, лобной доле). Для таких случаев лучше
подходит термин когерентность фаз, что несколько похоже на синкопический
музыкальный ритм. Это синхронизация со сдвигом по времени.
У отдельных нейронов время интеграции составляет порядка 10 мс, за этот
период входные сигналы дендритов могут суммировать для увеличения веро-
ятности одиночного аксонального выходного спайка (см. гл . 3). В группе свя-
занных между собой нейронов они могут усиливать уровень возбуждения друг
друга между 30 и 100 Гц, подавая сигналы через синапсы в интервале до 10 мс.
К примеру, если два возбуждающих нейрона передают друг другу сигналы на
50 Гц, то между ними может поддерживаться возбуждающая петля обратной
связи, так как сходящиеся сигналы могут приходить в пределах критическо-
го 10-мс периода. Но уровни возбуждения нейронов ниже 30 Гц могут и не
быть интегрированы нейронами-мишенями, так как отдельные спайки при-
бывают слишком поздно для суммирования эффектов. Поэтому считается, что
группа нейронов, сигнализирующая в бета-гамма диапазонах, сильнее влияет
на нижележащие нейроны, чем сигнализирующие при более низких частотах.
Очевидно, что реальные нейрональные сети головного мозга устроены гораз-
до сложнее и содержат тормозные и возбуждающие компоненты. Тем не менее
указанные исходные положения применимы в целом к нейронам; в поддержку
их получена масса прямых эмпирических фактов.
Передача радиоволн обладает рядом общих черт с синхронностью осцилля-
ций мозга. Обычно на мощных электромагнитных волнах, используемых при
передаче радио сообщений, применяется либо модуляция частот (FM), либо мо-
FM), либо мо-
), либо мо-
дуляция амплитуды (AM). В любом случае базовая частота (число, указанное
на шкале вашего радиоприемника) служит несущей для быстрых изменений
по частоте или амплитуде, отражающих голосовой или музыкальный сигнал.
В радиоприемнике электронная схема настроена на частоту радиостанции, она
406
Глава 8
может отфильтровывать добавленный, модулирующий сигнал, доставляемый
на волне-переносчике (несущей). В итоге вы слышите песню и никогда не слы-
шите частоту носителя.
Описанные сходства не случайны, они отражают базовую физику волн, что
применимо и к ритмам головного мозга, и к электромагнитному излучению.
Наличие FM- и AM-радио позволяет предположить как минимум два различ-
FM- и AM-радио позволяет предположить как минимум два различ-
и AM-радио позволяет предположить как минимум два различ-
AM-радио позволяет предположить как минимум два различ-
радио позволяет предположить как минимум два различ-
ных способа, которыми в мозге идет обработка информации ритмов. Но суще-
ствует значительно больше кодирующих схем. Ритмы мозга могут работать как
часы, в них могут использоваться одиночные толчки или из серии, как в азбуке
Морзе. Разные нейроны могут передавать сигналы различными путями, воз-
можно, используя разные молекулы и синапсы.
Телевидение — это пример пространственно-временного кодирования,
в котором передаваемый сигнал сканирует каждую линию экрана от верха до
самого низа. Мониторы компьютеров используют сходное пространственно-
временное кодирование. Ритмы головного мозга также, по всей вероятности,
координируют зрительно-топические карты, сомато-топические карты и дви-
гательные карты. Как уже упоминалось, в головном мозге много топографиче-
ских карт, представляющих собой сенсорные входные сигнальные пучки или
нейро-мышечные карты на разных уровнях обобщения (см. гл . 5).
В эволюции ритмические свойства нейронов использованы на протяжении
сотен миллионов лет. Поэтому не следует ожидать, что найдется только один
нейрональный код. Сейчас мы знаем, что ритмы головного мозга широко рас-
пространены; знаем также об их связи с установленными функциями.
В конце концов, волны могут взаимодействовать друг с другом. Если поста-
вить радиоприемник рядом с компьютером, то можно слышать шум всякий раз
при нажатии на клавиатуру. Это объясняется тем, что каждая клавиша испуска-
ет электромагнитный сигнал, уходящий в окружающее пространство. Волно-
вая интерференция — это фундаментальное явление в физике распространений
волн. Интерференция может быть очень полезной в мозге, но может и разру-
шать обработку информации нейронами. Мы лишь начинаем разбираться в
роли ритмов головного мозга, но вполне вероятно, что влияние, которое имеет
волновая интерференция, окажется значительным.
2.8 . Гамма-активность играет множество ролей
На рис. 8 .18 приведен классический способ изучения гамма-активности при
зрительном восприятии с применением прямой записи электрической активно-
сти коры головного мозга. Внутричерепная ЭЭГ отличается лучшим соотноше-
нием сигнал/шум по сравнению с поверхностной ЭЭГ, поскольку запись в пер-
вом случае ведется прямо от мозга. Обычно напряжения внутричерепной ЭЭГ
даются в милливольтах; для поверхностной ЭЭГ — в микровольтах. На этом
основании внутричерепная ЭЭГ обладает значительными преимуществами. Но
ее большим недостатком оказывается необходимость в проведении инвазивной
операции. В исследованиях на животных внутричерепные электроды использу-
ются часто, но внутричерепная ЭЭГ на людях строго ограничена пациентами,
которым это необходимо по медицинским показаниям. У больных эпилепсией
внутричерепную ЭЭГ проводят для определения областей мозга, где возника-
ют припадки, и для локализации зон коры, нуждающихся в защите, чтобы не
допустить нарушения важных функций, например речи и зрения. Пациенты во
Сознание и внимание
407
время такой операции находятся в сознании, поэтому могут отвечать на вопро-
сы, описывать свои ощущения. Кора сама по себе не имеет болевых рецепторов,
и местной анестезии в месте надреза оказывается достаточно, чтобы не испы-
тывать дискомфорт. Таким образом, пациент находится в сознании, как обычно,
несмотря на то что происходит непрерывная запись активности его/ее мозга и
ведется стимуляция непосредственно через кору (Crone, 2006; Canolty, 2006).
Рис. 8 .18 . Возникающие гамма-всплески отражают связывание/узнавание зритель-
ных черт. В эксперименте, сделавшем прорыв в своей области, Таллон-Бодри и Бер-
тран (Tallon-Baudry, Bertrand, 1999) показали гамма-активность (желтые подъемы) на
внутричерепной ЭЭГ (при прямой записи мозга) у пациентов-эпилептиков, которых
просили указывать появление видимого треугольника. Выявлялись два скачка гамма-
активности: первый при предъявлении стимула и второй, по которому можно отличать
случаи узнавания треугольника и неузнавания его. Опознавание треугольника происхо-
дило как при предъявлении истинного треугольника, так и мнимого, но не происходило
при предъявлении стимула «нетреугольник». Оба гамма-всплеска, по всей вероятно-
сти, вовлекают как сознательные, так и неосознаваемые аспекты зрительного процесса.
На силуэте головы (в) показано распространение второго гамма-скачка по затылочной
и частично по теменной коре. (Источник: Tallon-Baudry, Bertrand, 1999.)
408
Глава 8
Исследователи Таллон-Бодри и Бертран (Tallon-Baudry, Bertrand, 1999)
просили пациентов, находящихся в сознании, с имплантированными в мозг
электродами для внутричерепной ЭЭГ, рассказывать о видимом треугольнике.
Сначала показывали только истинные треугольники, что вызывало два боль-
ших всплеска активности в гамма диапазоне (выделено желтым на рис. 8.18.
На верхнем графике в части (б) рисунка видно, что два (желтых) гамма подъема
происходят сразу после предъявления стимула (время t = 0). Первый из гамма-
всплесков, между 50 и 150 мс, отражает восприятие зрительного сигнала. Вто-
рой гамма-всплеск наступает через 250–300 мс после стимула, он отмечается
как в случае истинного, так и мнимого треугольника, но не отмечается в случае
контрольного стимула, не похожего на треугольник. Мнимый треугольник Ка-
ниша (рис. 8 .18 (а)) лишен сторон и имеет лишь три белых острых клина на
фоне трех темных дисков. Но испытуемые воспринимают такой треугольник
как истинный. Таким образом, второй гамма всплеск указывает на воспринима-
емый треугольник. Он не развивается в случае предъявления псевдо-треуголь-
ника (нижний график). Момент распознавания треугольника можно считать со-
гласованием между зрительным входным сигналом и памятью. Зрительная кора
при этом восклицает: «Вот и он!» в ответ на видимый треугольник.
Как представлено на рис. 8 .18 (в) вверху, второй гамма-всплеск расположен
над областью первичной зрительной коры испытуемого. Поскольку зрительные
зоны обладают двусторонними связями, вполне вероятно, что зрительные зоны
более высокого уровня также могут осциллировать синхронно с затылочной
корой (область V1). Ниже мы увидим, что одна из предполагаемых функций
гамма-активности — синхронизация зрительных карт.
В серии исследований с применением внутричерепной ЭЭГ, проведенных
в последнее десятилетие, выявлено, что гамма-подъемы (а также подъемы на
других частотах) отмечаются в тесной связи с когнитивными задачами. Они
являются неотъемлемым компонентом механизма передачи сигналов, лежа-
щих в основе обработки информации нейронами. И поскольку именно это об-
суждалось в последнее время, новые факты (вместе с выявленными данными
по ЭЭГ и магнитоэнцефалограммам) имеют важное значение. Заметим, что
измерение быстрых осцилляций с помощью функциональной МЭГ представ-
ляет большую трудность, так как сопровождается пятисекундным повышени-
ем ответа для параметра кислорода крови. При этом метод функциональной
МЭГ обладает своими выдающимися достоинствами и приобрели важность
исследования, объединяющие функциональную МЭГ с электромагнитными
сигналами от мозга.
На рис. 8 .19 показано, как указанные открытия связаны со слухом. В левой
части приведены результаты пассивного слушания, т. е. появление гамма-ак -
тивности (примерно через 200 мс после начала действия стимула). На правой
части рисунка видно, что длительность и частота гамма-активности больше и
шире распространены при «активном слушании», т. е. когда требуется повы-
шенное внимание в процессе прослушивания. К сегодняшнему дню накопле-
но много фактов, демонстрирующих, что активное исполнительное внимание
ведет к увеличению гамма-активности, это описано далее. Также существует
большое количество физиологических данных, свидетельствующих, что актив-
ное слушание улучшает обработку стимула и обучение.
Сознание и внимание
409
Следует отметить, что описанные внутричерепные ЭЭГ-исследования опу-
бликованы в последние десять лет. Их можно рассматривать как значительный
шаг вперед в нашем понимании высокочастотных событий головного мозга.
Если исследования на животных с имплантированными электродами проводи-
лись в течение многих лет, то такие работы на людях послужили подтвержде-
нием полученных данных и их расширением применительно к познавательной
способности человека. Внутричерепная ЭЭГ с имплантированными электро-
дами обладает гораздо лучшей пространственной чувствительностью по срав-
нению с поверхностной ЭЭГ, а также более высокой чувствительностью и от-
ношением сигнал/шум. К тому же можно записать более широкий диапазон
частот. Такие данные, вместе с получаемыми с помощью неинвазивных мето-
дов регистрации мозга, оказывают очень большое влияние на наше представле-
ние о роли головного мозга в человеческих когнитивных способностях.
2.9 . Синхронизация гамма-волн может соединять
визуальные черты в осознаваемые образы
Пока что мы разбирали два рода фактов относительно гамма-ритма в позна-
вательных способностях: амплитуду и синхронность гамма-ритма. Амплиту-
да — это громкость рок-группы, синхронность — это степень того, насколько
Рис. 8 .19 . Гамма-активность в процессе слушания. Слуховое восприятие сопрово-
ждается крупными всплесками гамма-диапазона на внутричерепной ЭЭГ (а), которые
значительно больше при «активном слушании» (б). При таком состоянии, как предпо-
лагается, к сознаваемому слуховому стимулу добавляется исполнительное внимание.
Термином «вызванные гамма» (в) обозначают эндогенную активность мозга, выявля-
ющуюся через 210 мс после начала действия стимула. Она может отражать активность
повторных входов между многочисленными уровнями анализа (такими как акусти-
ка, фонемы речи, слоги, и морфемы (части слов) и слова). (Источник: Tallon-Baudry,
Bertrand, 1999.)
410
Глава 8
каждый попадает в ритм. Вполне вероятно, что обе характеристики взаимос-
вязаны. Однако путем записи с высоким временным разрешением можно от-
делить гамма-синхронность от амплитуды гамма-ритма.
Одна из возможных ролей гамма-синхронности состоит в «связывании» или
интеграции отдельных признаков стимула в значимые объекты (см. рис. 8 .12
и 8.13). Разные зрительные области представляют разные черты, и в некоторой
точке мозг должен их соединять в единый образ. Также эта точка может быть
местом, где зрительная информация становится осознаваемой.
Область распознавания объектов в мозге макаки, как было показано, диффе-
ренцирует между бессознательным и сознательным зрительными входами. Ког-
да мы видим лицо, то многие карты зрительных черт в мозге отвечают гамма-
активностью (рис. 8 .20). Осознаваемые образы сочетают в себе многие черты;
в норме в зрительной сцене мы не видим плывущих точек света или цветовых
точек, или же контрастных граней. Вместо них мы видим зрительные образы
как четкие и осознаваемые.
На рис. 8 .20 непосредственно не видно синхронности, но есть только ампли-
туда в гамма-диапазоне. Один из первых экспериментов, где была продемон-
стрирована гамма-синхронность между зонами головного мозга при сознатель-
ном видении, был опубликован исследователями Энгелом и Зингером (En�el,
Sin�er) (рис. 8.21).
Энгел и Зингер изучали на кошках бинокулярную конкуренцию (см. гл . 6)
как классическую парадигму, которая позволяет нам сравнить обработку созна-
тельного и бессознательного стимулов в зрительных зонах мозга при идентич-
ном физическом входе в оба глаза. Эти ученые регистрировали прямо от глаз-
ных доминантных колонок в зрительной коре, где шло разграничение между
входами через правый и левый глаза. В опыте кошкам проводили операцию, в
результате которой их поля зрения не перекрывались, такое состояние называ-
ется косоглазие (страбизм, гетеротропия) или «амблиопичный глаз», когда оно
развивается естественным образом. Страбизм ведет к одностороннему окуляр-
ному доминированию в зрительной коре, определяя, какой глаз будет домини-
ровать, т. е. какой глаз будет «сознательным». Тогда, при условии что конкури-
рующий стимул становится осознаваемым в правом глазу в сравнении с левым,
доминантное поле коры можно записать отдельно по каждому глазу.
Энгел и Зингер (En�el, Sin�er, 2001) перед каждым глазом кошки демонстри-
En�el, Sin�er, 2001) перед каждым глазом кошки демонстри-
, Sin�er, 2001) перед каждым глазом кошки демонстри-
Sin�er, 2001) перед каждым глазом кошки демонстри-
, 2001) перед каждым глазом кошки демонстри-
ровали решетки, двигающиеся в противоположных направлениях, что давало
разновидность бинокулярной конкуренции, при которой не возможно объеди-
нение в единый сознательный зрительный образ. Результаты эксперимента при-
ведены на рис. 8 .21 . Заметим, что гамма-синхронизация в коре — только по
«сознательному» глазу, а для недоминантного (неосознаваемого) глаза никакой
синхронизации не отмечено.
По совпадению стимулов на реакцию с однозначной двигающейся решет-
кой, кошку как бы спрашивали, что она видит. Можно представить, что экс-
периментатор задавал вопрос: «Видишь эту или ту решетку?». Кошка могла в
такой форме дать «произвольный отчет» по осознаваемому стимулу. Подобные
опыты были всесторонне проведены на макаках, и были получены сравнимые
результаты.
Сознание и внимание
411
Рис. 8 .20 . В восприятии лица задействована гамма-активность на многих зрительных
картах. Лаше с сотрудниками (Lachaux et al., 2005) использовал иЭЭГ для записи во вре-
мя восприятия лиц непосредственно с четырех разных зрительно-топических областей
в коре мозга человека. Стимулом служило упрощенное черно-белое изображение (ввер-
ху), при этом требуется некоторое время, чтобы увидеть в нем лицо человека. В трех из
четырех областей был быстрый интенсивный всплеск гамма-волн, начиная менее, чем
через 100 мс после предъявления стимула (с верхушки, веретенообразной извилины,
области �1, латеральной затылочной извилины [ПБ19] и левой межтеменной борозды).
В четвертой же области, зоне �1, выявлялось снижение в гамма-активности на период
около 700 мс (темная зона), потом следовал высокий ее подъем. По заключению авто-
ров, ответ гамма-диапазона может отличаться по разным зонам мозга. Такие отличия
могут быть обусловлены избирательным подавлением в разных зрительно-топических
картах. Полагают, что ингибирующие нейроны постоянно модулируют общую актив-
ность возбуждающих пирамидных нейронов коры. Поздний гамма-всплеск в �1 может
отражать нисходящую активацию от зоны распознавания лиц (веретенообразная изви-
лина). (Источник: Lachaux et al., 2005.)
412
Глава 8
Следует отметить, что пик активности на рис. 8 .21 происходит с четкой син-
хронностью при времени t = 0 в доминантном комплексе популяций нейронов.
Менее выраженный пик отмечается для t = ± 20 мс и в интервалы, кратные
20 мс. Такие пики уменьшаются по мере того, как зрительно-топические нейро-
нальные карты уходят от полной синхронности. Однако даже самые небольшие
пики происходят с интервалом в 20 мс, что указывает на гамма-синхронизацию
при 50 Гц и составляет один цикл на каждые 20 мс. В отличие от доминантного
комплекса нейронов подавляемый комплекс не демонстрирует никакой синхро-
низации или очень незначительную ее степень. Это также проявлялось по тому,
как кошка не могла указанный стимул соотнести со стимулом–образцом, как бы
отвечая, что «ничего не видно» по недоминантному глазу.
Рис. 8 .21 . Объединение черт в сознании путем гамма-синхронности. Необходимо ли
объединение черт для осознанного восприятия объекта? Многие ученые приводили
аргументы, поддерживающие это положение. Энгел и Зингер (En�el, Sin�er) опублико-
вали классический эксперимент, в котором «доминантный комплекс» нейронов удалось
отдельно связать с левым или правым глазом. Наблюдали непосредственно за левой
или правой зрительной доминантной колонкой в коре кошки, применяя бинокулярную
конкуренцию. Решетки смещались в противоположных направлениях в двух полях зре-
ния, что давало два конкурирующих входа, которые не сливались в единое когерентное
восприятие. Гамма-активность синхронизировалась только в доминантном поле зрения,
для определения которого кошку обучали по однозначному стимулу, на основании чего
шел осознаваемый сигнал. RMA — относительная амплитуда модуляции. (Источник:
En�el, Sin�er, 2001.)
Сознание и внимание
413
Тип экспериментов Энгела и Зингера стал серьезным историческим проры-
вом в нашем понимании сознательного видения у других млекопитающих. Ана-
логичные эксперименты также выполнены и на людях, что обсуждается в гл. 6 .
Сходные результаты по роли ритмов мозга в сознательном видении полу-
чены с помощью совсем иного метода. На рис. 8 .22 приведены данные, недавно
полученные с помощью транскраниальной электрической стимуляции мозга,
а точнее методики с использованием поля переменного тока. График — это тра-
диционная психофизическая кривая, связь физического стимула и субъективно-
го опыта. В данном случае сознательными опытами были «фосфены», зритель-
ные «вспышки», получать которые можно прямой стимуляцией мозга. Впервые
фосфены продемонстрированы на человеке нейрохирургом-новатором Вилде-
ром Пенфилдом (Wilder Penfield) и его сотрудниками около 50 лет назад при
предоперационных исследованиях трудноизлечимых пациентов с эпилепсией
(Penfield and Ro�erts, 1959). Но условия надежного получения фосфенов были
недостаточно хорошо понятны до недавнего времени. Фосфены — это субъек-
тивные опыты (в отличие от иных видов прямой стимуляции зрительной коры),
поэтому они четко говорят нам о зрительном осознании.
В недавно проведенном исследовании Канаи с сотрудниками (Kanai et al.,
2008; рис. 8.22) установлено, что фосфены точно возникают при переменном
токе в бета-диапазоне (около 20 Гц, что иногда называют нижним гамма-диапа-
зоном). Такое открытие тоже носит исторический характер. Психофизическая
функция, описанная Канаи с сотрудниками, выявила, насколько регулярными
теперь должны быть такие явления. Это существенно, так как долго считалось,
Рис. 8 .22 . Транскраниальная стимуляция приводит к возникновению сознательных
«вспышек». Сознаваемые фосфены (визуальные вспышки) возникают при транскра-
ниальной стимуляции переменным током в затылочной доле. Исследователь Канаи с
сотрудниками с помощью транскраниальной стимуляции переменным током описал
спектр частот от 3 до 30 Гц и использовал эти тонкие психофизические кривые, от-
носительно частот транскраниальной стимуляции переменным током, для связи с субъ-
ективными зрительными ощущениями. Пик таких кривых лежит чуть ниже 20 Гц в
бета-диапазоне, поэтому можно предположить, что внутренние ритмы были вызваны
электрическим полем. (Источник: Kanai et al., 2008.)
414
Глава 8
что фосфены сложно получить много раз подряд. Выраженный пик сразу за
20 Гц также привлекает внимание, указывая, что сознательные зрительные
события требуют ритмических событий в мозге, что следует ожидать из выше-
описанных опытов с внутричерепной ЭЭГ-регистрацией. Отметим, что субъек-
тивные уровни пиков возрастают предсказуемым образом по мере увеличения
уровня стимулирующего тока (в микроамперах).
Из приведенных выше фактов следует, что гамма-синхронизация является
одним из компонентов обычных когнитивных задач, но как гамма-синхрони-
зация «выполняет» сложную комплексную задачу? Ведь восприятие слова или
лица требует большой обработки. Она осуществляется очень быстро, для чего
за каждым компонентом задачи должен быстро начинаться следующий компо-
Рис. 8 .23. Множественные синхронные гамма-вспышки при проговаривании слов и их
слушании. Крон и коллеги изучали находящихся в сознании пациентов-эпилептиков с
имплантированными краниально электродами. На основании полученных данных мож-
но заключить, что в мозге возникают синхронные гамма-всплески между разными обла-
стями, задействованными в слушании, восприятии речи, и в образовании речи. Каждая
стадия задания с повторением слов, видимо, вызывает отдельный синхронный гамма-
всплеск в слегка разных районах левого полушария одного пациента. Гамма-синхрон-
ность наблюдали в четырех частях высокой гамма-области, от 57 до 117 Гц (розовые,
голубые и фиолетовые стрелки). Красные горизонтальные стрелки указывают на гамма-
синхронность, отмеченную в особых районах диапазона частот. На графиках красным
цветом обозначена самая высокая амплитуда, желтым и светло-синим — более низкие.
По вертикальным осям графиков — частота, по горизонтальным — интервал до и после
стимула в сотнях миллисекунд. (Источник: Crone et al., 2006.)
Сознание и внимание
415
нент. По независимым фактам можно предположить, к примеру, что исключи-
тельно сенсорный аспект восприятия может происходить до анализа значения,
по крайней мере, при восприятии языка. Нижеописанные эксперименты пока-
зывают, как успешные всплески гамма-синхронизации могут приводить к вы-
полнению разных стадий одной когнитивной задачи.
И снова четкие факты были получены в исследованиях, проведенных на
находящихся в сознании пациентах-эпилептиках, с помощью внутричерепной
ЭЭГ, проводимой для локализации очага эпилепсии и определения тех обла-
стей мозга, повреждение которых при операции приведет к нарушению важных
функций, таких как зрение, слух и речь. В работах Крона и коллег (Crone et al.,
2006) показано, что гамма-синхронизация между локальными областями лево-
го полушария часто временна, что указывает на краткий период координации
между двумя частями мозга. На рис. 8 .23 видна гамма-синхронизация, обуслов-
ленная событиями, у пациента с сетью электродов над левым полушарием, при
запуске ритмов мозга в простом задании называния. Пациенту давали серию
слов, просили их запомнить, и затем повторить по очереди. При этом слуховой
вход и ответ с называнием можно разделить во времени. Фиолетовым цветом
на данном рисунке показаны различные волны в высоком гамма-диапазоне
(67–117 Гц) в разных областях левого полушария. При прослушивании каждого
слова запускались несколько синхронных гамма-всплесков по слуховой коре и
области восприятия речи в коре (зоне Вернике). При произнесении того же сло-
ва отмечалась гамма-синхронизация по полю Брока, как и следовало ожидать.
Обратите внимание, что локальные кортикальные пятна в левом полушарии от-
ражают синхронные всплески в сложных сериях, что показано фиолетовыми,
розовыми и голубыми стрелками.
Пока что мы наблюдали локальную гамма-синхронизацию в ограниченных
областях коры. Как будет описано далее, существует и гамма-синхронизация
больших расстояний, по удаленным зонам коры, которые должны совместно
работать при выполнении определенных задач.
2.10. Тета-ритмы играют множество ролей
Гамма-ритмы не являются единственными важными осцилляциями в головном
мозге; исключительно по историческим причинам в гамма включен широкий
набор осцилляций примерно от 30 до 120 Гц, иногда и выше. Так сложилось в
связи с тем, что быстрые осцилляции было сложнее записать сразу, частично
из-за того, что лучше они видны при имплантированных электродах. Однако
очевидно, что более медленные ритмы играют фундаментальную роль. Тета-
ритм — это разновидность самых медленных осцилляций при нормальном со-
стоянии бодрствования, с циклом от 4 до 7 Гц, непосредственно над дельта-
ритмом, доминирующим при медленноволновом сне. Совсем недавно открыты
медленные осцилляции, которые могут достигать 0,01 Гц (рис. 8 .24).
Видимо, тета «переносят» гамма-осцилляции почти так же, как радиочасто-
ты AM переносят голосовой сигнал. Полагают, что тета играют ключевую роль
в эпизодической памяти и в активностях лобной доли. Тета являются одними из
основных частот–переносчиков в гиппокампе, наблюдаемых как при помеще-
нии событий в эпизодическую память, так и при их извлечении. Считается, что
тета нужны для кодирования и декодирования в неокортексе процессов обуче-
ния из гиппокампа, особенно в лобных долях. На ЭЭГ с поверхности это можно
416
Глава 8
Рис. 8 .24. Уплотнение тета-гамма по 16 разным заданиям. Синхронизация тета-гамма
по 16 разным заданиям, с применением внутричерепной ЭЭГ. (а) Индекс синхрониза-
ции (t = 0 указывает на точную синхронизацию, корреляция уменьшается по мере от-
клонения во времени в каждом направлении). На левых полушариях для испытуемого 2
показаны цветными точками положения электродов соответственно цветной линейке в
центре на (б) и на (в). Отметьте, что пассивное слушание предсказуемых тонов сопро-
вождается тета-гамма-синхронизацией (г). Но, как показано на (в), сложное для рабочей
памяти задание с называнием объектов, отстоящих на два назад от текущего, отлича-
ется большим числом желтого и красного цветов, что указывает на более выраженную
синхронизацию в более трудном задании. (г) Показаны 16 заданий; наиболее высокие
корреляции (красный цвет) для квадратов ближе к диагонали, наиболее сходных друг
с другом. На (д) отражены низкие, но при этом значимые корреляции в синхрониза-
ции между сходными заданиями, по сравнению с непохожими заданиями. (Источник:
Canolty et al., 2006.)
Сознание и внимание
417
наблюдать вдоль средней линии головы, при вызывании событий из эпизодиче-
ской памяти и во время исполнительных заданий, вовлекающих как латераль-
ные, так и медиальные лобные доли.
Существует много способов взаимодействия волн головного мозга: как и
волны на воде, они могут суммироваться, или сглаживать друг друга. Возможна
также модуляция частот волн, вызывающая ускорение или замедление. Более
того, есть указания на модуляцию фазы, так что степень синхронизации между
двумя источниками гамма-волн может измениться с целью передачи опреде-
ленного сигнала. Именно модуляция фаз предложена как механизм кодирова-
ния в пространственной памяти гиппокампа для крыс.
Нет оснований считать, что мозг обладает только одним или двумя типами
кодов. Природа нейронального кодирования — это сейчас один из «горячих»
научных вопросов, который отнюдь не решен.
Мы показали, что тета-осцилляции могут группировать гамма, и в опреде-
ленных условиях альфа также могут группировать гамма-активность. Стериэйд
(Steriade, 2006) выдвинул предположение о том, что медленные осцилляции мо-
Steriade, 2006) выдвинул предположение о том, что медленные осцилляции мо-
, 2006) выдвинул предположение о том, что медленные осцилляции мо-
гут в большинстве случаев работать для группирования более быстрых. Даже
недавно открытые «медленные осцилляции» в пределах от 0,1 до дельта-диа-
пазона могут служить основой для группирования тета, альфа, бета и гамма.
2.11. Альфа-ритмы
В 1929 г. немецкий психиатр Ганс Бергер (Ber�er) впервые получил ЭЭГ че-
Ber�er) впервые получил ЭЭГ че-
) впервые получил ЭЭГ че-
ловека, поместив электрод на голову своего сына в районе затылочной коры.
Ключом к открытию Бергера послужил новый мощный усилитель, способный
усиливать маленькие электрические напряжения с поверхности головы и пе-
реводить их в видимые отклонения следа чернильного пера, оставляемые на
движущейся бумажной ленте. Бергер попросил сына открыть, а затем закрыть
глаза; так он обнаружил, что при закрытых глазах появляется маленькая сину-
соидальная волна. Бергер назвал ее альфа-волной (рис. 8 .25). (Названия альфа,
бета, гамма и прочие отражают лишь исторический ход их открытий.) Тради-
ционно альфа-волну определяют как осцилляции от 8 до 12 Гц. Было обнаруже-
но, что размер альфа-волн увеличивается при внутренних заданиях, например
при устном подсчете и рабочей памяти. Согласно недавно выдвинутой теории,
альфа отражают избирательное подавление не связанных с заданием зон и ак-
тивностей в мозге (Palva, Palva, 2007).
На сегодняшний день ясно, что открытие Бергера явилось историческим для
области изучения мозга и медицины. Но еще несколько десятилетий назад по-
верхностная ЭЭГ с трудом была доступна для анализа. Даже сейчас выполне-
ние ЭЭГ с кожи головы представляет практические затруднения. К примеру,
мы все еще не располагаем надежным способом для определения источников
поверхностных потенциалов. Эту проблему тем не менее можно решить други-
ми методами, включая внутричерепную ЭЭГ, МЭГ, позитронную эмиссионную
томографию, и функциональный ЯМР. Есть много важных практических при-
менений поверхностной ЭЭГ в медицине. Она безоговорочно является самым
простым и доступным по стоимости способом записи деятельности головного
мозга, и продолжается оставаться полезной. Постоянно прилагаются усилия
для повышения ее точности.
418
Глава 8
Головной мозг «полон» пересекающихся электромагнитных волн. Са-
мое примечательное относительно волн Бергера, полученных при закрытых
глазах, — это их удивительная простота. Видимые альфа-волны отчетливо
ритмичны, легко наблюдаемы и связаны с когнитивными событиями. Тради-
ционное объяснение данному факту основано на том, что мозг работает «на
холостом ходу».
Следующее открытие Бергера получило название «альфа-блокирование». У
субъектов, у которых образуются альфа-волны, прерывают процесс открыва-
ния глаз или активный мыслительный процесс или представление чего-либо.
Вместо синусообразных ритмичных волн, на ЭЭГ затылочной зоны коры вы-
являются нерегулярные, быстрые волны с низким напряжением (см. рис. 8 .4).
Такие быстрые волны были названы бета- (13–30 Гц) и гамма-волны (30 Гц и
выше). Сигнал бета-гамма
—
это удобный признак активности сознания при
бодрствовании, стандартно используемый в медицине при постановке диагно-
за. Следует иметь в виду, что данные видимые на ЭЭГ волны — это лишь вол-
ны на поверхности озера осцилляций. В основе нерегулярной ЭЭГ, получаемой
с кожи головы, лежат, как известно, синхронизированные ритмы. Их можно вы-
явить путем математического анализа и посредством записи через электроды,
погруженные глубоко в мозг. Поэтому можно считать, что существует две раз-
новидности сигналов ЭЭГ: альфа-волны, отражающие, как иногда полагают,
«холостой ход мышления», и активный мыслительный процесс, при котором
выявляется сложная, быстрая и нерегулярная активность мозга. Если учесть
сложную организацию головного мозга, то такие ранние наблюдения оказались
удивительно простыми и очень долго сохранялись.
Синусоидальные волны встречаются очень часто; их можно наблюдать в
случае, если любой вращающийся источник испускает сигнал в течение неко-
торого времени. Если прикрепить маленький фонарик на обод велосипедно-
го колеса и потом ночью поехать на этом велосипеде по прямой, то в итоге
луч фонаря прочертит синусную волну. Математик Жозеф Фурье (1768–1830)
Рис. 8 .25. Альфа-волны при открытых и закрытых глазах. Нативная ЭЭГ от затылочной
коры, показывающая нерегулярную активность при бодрствовании с открытыми глаза-
ми (верхняя запись) и спонтанные альфа-волны при закрытых глазах (нижняя запись).
Много исследований проведено с тех пор, как в 1929 г. Бергер открыл это явление, но
по-прежнему альфа-волны остаются в некоторой степени загадкой. Один из основных
вопросов: что происходит с нерегулярной ЭЭГ бодрствования, когда глаза закрыты и
альфа-«праздность» заступает на смену. Альфа-волны затем, по-видимому, активно по-
давляются. В свою очередь, ЭЭГ бодрствования подавляется с появлением альфа-волн
на необработанной, не подвергнутой анализу ЭЭГ. (Источник: Tononi in Laures and
Tononi, 2009.)
Сознание и внимание
419
доказал, что любую сложную волну можно представить как набор синусных
волн в результате процесса, получившего название анализ Фурье. Сейчас ЭЭГ
обычно проводят через высокоскоростной Фурье-анализ, в итоге мы можем
видеть синусообразные волновые компоненты в сложной активности головно-
го мозга. На практике мы часто группируем компоненты волн в диапазоны по
установленным интервалам: от 4 до 7 Гц — для тета, от 8 до 12 Гц для альфа,
и так далее. Однако необходимо учитывать, что природные факты могут не со-
гласовываться с нашими установленными границами. К примеру, разные под-
группы гамма, видимо, выполняют различные задачи.
Согласно недавно проведенным исследованиям, альфа-волны выявляются
при возникновении образов и других формах умственной деятельности (рис.
8.26). Подобно другим медленным ритмам, альфа-волны способны группиро-
вать более быстрые. На внутричерепных записях ЭЭГ видны как альфа-синхро-
низация, так и альфа-десинхронизация. (Десинхронизация напоминает «альфа-
блокирование», которое наблюдал Бергер.)
Мы только начинаем понимать систему нейронного кодирования в головном
мозге. Важно осознавать, что и синхронизация, и ее отсутствие несут в себе
информацию, подобно тому как наличие и отсутствие света представляют со-
бой зрительную информацию — образы тени также необходимы, как и образы
света. В сигнальных кодах использованы и сигналы, и их отсутствие, как в аз-
буке Морзе.
По предположению ученых Палва и Палва (Palva, Palva, 2007), пересекаю-
Palva, Palva, 2007), пересекаю-
, Palva, 2007), пересекаю-
Palva, 2007), пересекаю-
, 2007), пересекаю-
щаяся по частотам синхронизация фаз между альфа-, бета- и гамма-осцилляци-
ями «координирует выбор и поддержание нейрональных представлений объек-
та в рабочей памяти, восприятии и осознании». В некоторых опытных условиях
удается наблюдать и альфа-синхронизацию, и десинхронизацию.
На рис. 8 .26 явно демонстрируется участие альфа-волн при удержании объ-
ектов в рабочей памяти. Как уже указывалось выше, есть вероятность, что
Рис. 8 .26 . Альфа-волны могут группировать гамма-осцилляции в рабочей памяти.
В непосредственном упражнении для памяти выявлен мощный всплеск альфа-актив -
ности (горизонтальные красная и желтая линейки) в течение 3 с, пока испытуемые удер-
живали в памяти некоторое число. Отметим, что наиболее выраженная активность по-
стоянно сконцентрирована около 12 Гц. (Источник: Palva, Palva, 2007.)
420
Глава 8
такое увеличение альфа-активности может также многократно переплетаться с
другими типами волн. Но на основании фактов можно предположить, что альфа
играют функциональную роль в познавательной способности человека и что
они являются не просто «ритмом холостого хода», как считали до недавнего
времени многие исследователи.
3.0 . Внимание улучшает восприятие,
познавательную способность и обучение
С точки зрения здравого смысла, понятия внимание и осознание различаются
между собой. Под вниманием подразумевают способность направить когнитив-
ные ресурсы на определенное событие. К примеру, если вам не удается вспом-
нить термин из этой книги, то вы можете попробовать отбросить все осталь-
ные мысли и «уделить больше внимания» попытке вспомнить нужное слово.
Во внимание вложен «указывающий» или же «направляющий» оттенок смысла.
Осознание же в повседневной речи характеризует опыт — прочтения этой фра-
зы, к примеру, или обдумывания, прочувствования и представления, которые
могут возникнуть после прочтения этих слов. Избирательное внимание дает
возможность выбора между вероятными событиями. Осознание можно пред-
ставлять как переживание/опыт одного события после того, как был сделан
выбор именно его.
Часто мы сами решаем, на что направить наше сознание: продолжить ли
чтение этой книги, или включить телевизор, или прекратить думать о друге
и т. д. Существует и автоматический выбор направления внимания, если сти-
мулы интенсивные, динамичные, или они биологически или личностно важные
(Hahn et al., 2006). Мы непрерывно сканируем мир, даже без явного такого на-
мерения, и часто выбираем значимые события автоматически.
Как большинство млекопитающих, обезьяны могут быть агрессивными дра-
чунами. При столкновении с представляющей угрозу змеей или с соперником
сначала они посылают предупредительные сигналы, используя гримасу угрозы,
крики и соответствующее поведение. На рис. 8 .27 приведена гримаса оскала
макаки. У макак и у людей выявлена повышенная ответная реакция нейронов
зрительной оценки лица на несущие угрозу гримасы, так как они биологиче-
ски крайне важны. По этой причине вполне вероятно, что такие гримасы/вы-
ражения лица угрозы обрабатываются «снизу вверх», как и тревожные звонки.
Такой важный стимул прерывает поток других опытов.
Молодые млекопитающие могут отличить притворную агрессию от истин-
ной. Игровая агрессия — это часть игровых потасовок/драк, что часто нравится
молодым млекопитающим. Лин и соавторы (Lin et al., 2009) выявили, что вни-
мание к выражающим угрозу лицам возникает несознательно, при этом исход
такого процесса обычно сознательный.
3.1. Фланговая задача Познера
Познер с сотрудниками разработали простой тест на зрительное внимание,
получивший название фланговая задача. В нем испытуемого просят обратить
внимание на стимул в обозначенном месте справа или слева от точки фиксации
взгляда (помеченной точкой или плюсом). У людей очень ограничена зона сет-
чатки (желтое пятно), через которое мы фиксируем маленькую часть зритель-
Сознание и внимание
421
ного поля в каждый момент, поэтому можно контролировать точную информа-
цию, доступную для испытуемого. (Мы видим только около 3–6° зрительной
дуги, когда глаза фиксированы на определенной точке. Проверьте сами!)
С помощью данного теста можно проверять как произвольное, так и непро-
извольное внимание, давая испытуемым правильную или ложную информацию
о том фланге, на котором появится стимул (рис. 8 .28), что можно варьировать от
упражнения к упражнению. Путем сочетания верных и ложных указаний перед
стимулом можно получить данные для одного эксперимента по флангам, кон-
тролируемым согласно ожиданию (при правильных указаниях) и неожидаемым
(при ложном указании). Довольно просто примерно за полчаса провести такой
тест через функциональный магнитно-резонансный сканер; по полученным
изображениям мозга можно получить раздельную информацию по упражне-
ниям с совпадением ожидания и результата и упражнениям при неожидаемом
результате. Путем вычитания сканирующих изображений активности мозга
при «неожидаемом внимании» из изображений активности «ожидаемого вни-
мания», Познеру с сотрудниками удалось получить относительно простой кри-
терий для зон мозга, вовлеченных в произвольное зрительное внимание.
Фланговая задача — это самый распространенный метод изучения зритель-
ного внимания, и поэтому мы разберем его детально. Этот тест простой, эффек-
тивный и легко адаптируемый. Например, зрительным стимулом могут служить
эмоциональные лица, что позволит исследовать, как мозг формирует внимание
на эмоциональные события (Fan et al., 2002; Posner and Peterson, 1990).
Рис. 8 .27. Внимание снизу-вверх (восходящее). Гримаса угрозы макаки. Нейроны зри-
тельной оценки лица возбуждаются быстрее, когда макаки видят гримасу угрозы, как на
фотографии, по сравнению с нейтральным выражением. Выражающая угрозу гримаса
является биологически важным сигналом. На графике приведены спайки в секунду ней-
рона, расположенного в зоне зрительной оценки лица, в веретенообразной извилине.
Кривая возбуждения при угрозе выделена красным, нейтральная кривая — черным цве-
том. (Источники: (вверху) Parr, Heintz, 2008; (внизу) �uilleumier, 2005.)
422
Глава 8
Вот как построена фланговая задача.
1. Испытуемые смотрят на точку фиксации взгляда. Они смотрят на цель с из-
вестного расстояния, поэтому угол зрительной дуги, определяемой точкой
фиксации взгляда, всегда постоянен. Ямка (наиболее плотное по фоторецеп-
Рис. 8 .28 . Фланговая задача по оценке внимания. Данный тест используется для иссле-
дования избирательного зрительного внимания и структур мозга, лежащих в его основе.
Испытуемый смотрит только на точку фиксации взгляда. Указательные сигналы, такие
как стрелка, переносят внимание к левому или правому флангу, но движения глаз запре-
щены. Мерой потери и преимущества являются точность и скорость ответа, что зависит
от верности/ложности указательного сигнала. Ответы на стимулы, возникшие на не-
ожидаемой стороне, имеют значения показателей времени и точности ответа хуже, что,
как полагают, отражает способность к отклонению установки внимания, построенной
по указанию. (Источник: Reynolds et al., 2003.)
Сознание и внимание
423
торам место сетчатой оболочки глаза) остается сфокусированной на точке
фиксации; данные от серий, где движения глаз отклонялись от точек фик-
саций, обычно не засчитываются. За пределами ямки, занимающей лишь
2–4° дуги, сетчатка теряет разрешение, она чувствительна только к перепа-
дам света и темноты. (Нормальное для нас восприятие богатого красочного
мира — это результат построения в зрительных зонах мозга, а не просто
запись входных сигналов сетчатки.)
2. При появлении стимулов справа или слева от точки фиксации взгляда за-
метить их можно только невидимым/скрытым вниманием, так как глаза
продолжают фиксироваться на исходной точке. Познер и Петерсон (Posner
and Peterson, 1990) установили, что люди способны избирательно увеличить
эффективность обработки вниманием входных сигналов с сетчатки, распо-
ложенных на несколько градусов от ямки (см. гл . 6).
3. Перед появлением стимула может возникать стрелка, указывающая испыту-
емому, где (слева или справа) следует ожидать появление стимула. Или крат-
кая вспышка левого квадрата информирует испытуемого, что стимул скоро
вспыхнет слева с вероятностью 80%.
4. На приведенном рисунке стимул возникает в ожидаемом квадрате на долю
секунды. Испытуемые реагируют с предельно возможной скоростью. Если
указания верны, то время реакции и точность оптимальны.
5. Но вспышка стимула может возникнуть и на противоположном фланге, т. е.
в неожидаемом месте. Такое неожидаемое событие влечет за собой потерю
в скорости реакции и в ее точности.
Итак, внимание имеет как преимущества, так и издержки. Преимуществом
внимания считается увеличение скорости и точности ответа на ожидаемое рас-
положение, по сравнению с неожидаемым. Потеря в скорости и точности в от-
вете на неожидаемый стимул является издержкой того, что внимание не было
обращено на данное событие. Фланговая задача позволяет дать количественную
оценку соотношения преимуществ-издержек в терминах скорости и точности.
Задание A�T (Fan et al., 2002) является обобщенным вариантом фланговой
задачи, позволяет оценить три разных аспекта внимания: готовность перед
ожидаемым сигналом (после предупредительного указания); ориентирование
к определенному месту в пространстве (применяется стрелка для указания
следующего флангового стимула); и исполнительное внимание, по действию
вопреки ожиданиям, установленным в задании. В таком представлении «испол-
нительное внимание» рассматривается как противоположное к «автоматически
контролируемому вниманию». Хорошим примером исполнительного или про-
извольного внимания служит упражнение Струпа (блок 8.1).
Задание A�T построено так, что каждый компонент внимания реализуется
в свое время, отличное от остальных, позволяя записать изображения мозга по
этим трем отдельным компонентам. В ходе различных условий задания мож-
но избирательно менять готовность, ориентирование и исполнительное вни-
мание. К примеру, если попросить испытуемых запомнить несколько чисел
в ходе выполнения задания, то это снизит исполнительное внимание, так как
задействуется центральная ограниченная способность, но не автоматическое
внимание.
424
Глава 8
Блок 8.1 . Называние цветов по Струпу и исполнительное внимание
Имея секундомер и рис. 8 .29, легко можно повторить исходное упражнение Струпа
(Stroo�, 1935). Сначала нужно прочитать вслух на предельно возможной скорости
список слов в столбцах 1 и 2, не обращая внимания на цвет шрифта. Начинайте,
закрыв все столбцы, кроме 1. Включите секундомер с первым словом и остановите
после последнего. Запишите полученный результат по времени. Теперь закройте все
столбцы, кроме 2-го, и прочтите снова вслух. Для столбцов 3 и 4 задание меняется —
теперь нужно называть цвета шрифта вслух на предельно возможной скорости, не
называя слова и буквы. Это нужно проделать для рядов цветных букв в столбце 3,
затем для рядов цветных слов столбца 4.
Струп отметил три результата. Во-первых, чтение слов происходит быстрее, чем на-
зывание цветов. Это обусловлено тем, что чтение — более привычный навык, по-
этому более автоматический, чем называние цветов (Cattell, 1886; Fraisse, 1969).
Во-вторых, различия были минимальны при прочтении слов в столбцах 1 и 2 (Экс-
перимент 1 Струпа), т. е. неправильный цвет шрифта не влиял на прочтение слов.
В-третьих, переход от букв к словам при назывании цвета шрифта выражался в зна-
чительной разнице по времени между столбцами 3 и 4 (Эксперимент 2 Струпа), т. е .
называние цвета шрифта слов, обозначающих другой цвет, значительно влияло на
результат. Вероятно, большая автоматичность при прочтении слов ведет к тому, что
слова прочитываются, даже когда этого делать не надо, что приводит к противоречию
в ответе на каждый стимул. Поэтому происходит замедление при назывании цветов и
ошибки (прочтение слова вместо называния цвета) возникают более часто.
В современном варианте упражнения Струпа применяются дисплеи, на которые ком-
пьютер выводит по одному цветному слову, вместо большого их числа на карточке,
что дает более тонкий контроль и более точное измерение отдельной реакции и по-
следующих эффектов. Также Струп не использовал совпадающие варианты (КРАС-
НЫЙ напечатанное красным шрифтом; сказать «красный»), а в современных вари-
антах его теста часто это включается. Обе описанные модификации были введены
Далримпль-Элфордом и Будаяром (Dalrim�le-Alford, Budayr, 1966).
Рис. 8 .29 . Упражнение Струпа
Сознание и внимание
425
Можно представить студента в лекционном зале, где одновременно имеются
много разных сенсорных входных сигналов. Студент должен находиться в со-
стоянии готовности, ориентирован на желаемый сенсорный поток, и должен
распределять ресурсы внимания на то, чтобы следить за лекцией, делать записи
и т. д. За эту готовность отвечают районы мозга в правой лобной и теменной
зонах (Witte, Marrocco, 1997). Но для готовности внимания требуется также
состояние бодрствования, что уже зависит от ствола мозга, таламуса и коры.
Зоны мозга, задействованные в ориентировании, расположены в правом височ-
но-теменном соединении и в нижней лобной извилине (соответствующей зоне
Брока в языковом полушарии) (Cor�etta et al., 2000). И наконец, произвольный
контроль внимания реализуется за счет передней поясной коры и латеральной
предлобной коры, ядра подушки таламуса и верхних бугров четверохолмия
(Marrocco, Davidson, 1998) (рис. 8 .30). Для всех указанных зон составлены зри-
Marrocco, Davidson, 1998) (рис. 8 .30). Для всех указанных зон составлены зри-
, Davidson, 1998) (рис. 8 .30). Для всех указанных зон составлены зри-
Davidson, 1998) (рис. 8 .30). Для всех указанных зон составлены зри-
, 1998) (рис. 8 .30). Для всех указанных зон составлены зри-
тельно-топические карты.
Не следует путать избирательное внимание и движения глаз, которые так-
же усиливают качество зрительного входного сигнала. Познер и Петерсон
(Posner, Peterson, 1990) выявили, что переключение внимания возможно тогда,
когда глаза остаются фиксированными на заданную точку. Тем самым мы спо-
собны улучшить обработку на несколько градусов направо или налево от точки
фиксации взгляда, даже без движения глаз. В опытах по зрительному вниманию
регулярно используются такие условия неявного внимания.
Зрительно-топические нейроны отвечают на зрительные стимулы на разных
уровнях анализа (см. гл . 6), что приведено в общих чертах на рис. 8 .31. Каждый
уровень такой графической модели, созданной Итти и Кохом (Itti, Koch), отве-
Itti, Koch), отве-
, Koch), отве-
Koch), отве-
), отве-
Литература
1. Stroop J.R. , 1935. Studies of interference in serial ver�al reactions. J Experimental Psy-
chology, 18, 643–662 . [re�rint. in J Exp Psychol Gen, 121, 15–23 .]
2. Cattel J.M ., 1886. The time it takes to see and name o��ects. Mind, 11 , 63–65.
3. Fraisse P. , 1969. Why is namin� lon�er than readin�? Acta Psychologica, 30, 96–103 .
4. Dalrymple-Alford E.C. , Budayr B. , 1966. Examination of some as�ects of the Stroo�
color-word test. Perceptual and motor skills, 23, 1211–1214.
Рис. 8 .30 . Нейросеть контроля внимания Познера. С помощью теста флангов Познер
с сотрудниками показали активацию нейросети зон мозга при исполнительном вни-
мании. Изображение справа демонстрирует высокую активность таламуса, возможно,
с участием ядер подушки и медиодорсального ядра. На рис. 8.31 приведена модель с ис-
пользованием этих зон. (Источник: Fan et al., 2005.)
426
Глава 8
чает на определенную характеристику стимула: цвет, ориентацию линий, кон-
траст и идентификацию объекта. Это является упрощением зрительной части
мозга, которая несравнимо более гибкая и работает с такими усложнениями,
как постоянные движения глаз и головы, очень узкие границы фовеального зре-
ния, и др. Однако рис. 8 . 31 помогает прояснить данный вопрос.
Рис. 8 .31 . Множественные зрительно-топические карты, отличительные черты и об-
учение. Множественные зрительно-топические карты поддерживают наши сознатель-
ные прочувствования кружек пива и пустынь. Обратите внимание, что человек внизу
рисунка смотрит на холодный напиток жарким днем в Сахаре. Как наш мозг выбирает
холодное пиво? Что мы осознаем? (Источник: Peter Thier et al., 2002; адаптировано из
Itti and Koch, 2001.)
Сознание и внимание
427
Каждая зрительно-топическая карта — это двумерная мозаика нейронов с
довольно узкими рецепторными полями (см. гл . 6). Поэтому можно задать бо-
лее частный вопрос: увеличивает ли внимание, нацеленное на определенное
событие или на определенное местоположение, обработку сигнала в соответ-
ствующей карте и нужном рецепторном поле? Если наблюдатель на рис. 8 .31
испытывает жару и жажду в процессе блуждания по Сахаре, то будет ли его/
ее система внимания усиливать зрительную обработку охлажденных кружек с
пивом, расположенных слева на зрительном поле? Данный вопрос из гораздо
более узкой области, и его можно проверять экспериментально.
3.2 . Модель внимания
Исследователи Итти и Кох (Itti, Koch, 2002) разработали удобную модель вни-
Itti, Koch, 2002) разработали удобную модель вни-
, Koch, 2002) разработали удобную модель вни-
Koch, 2002) разработали удобную модель вни-
, 2002) разработали удобную модель вни-
мания, сочетающую несколько важных признаков. Она показывает упрощенное
послойное представление зрительной системы, со множественными топогра-
фическими зрительными картами. Зрительные карты отображают линейное
ориентирование, интенсивность стимула (контраст), цвет и особенность/отли-
чительную черту. «Отличительная черта (особенность)» определяется в поня-
тиях контраста признака на любой зрительной карте. Для светочувствительных
зон это будет контраст между светлыми и темными пятнами на карте. В чув-
ствительных к движению зонах, таких как MT (средне-височное поле), это мо-
MT (средне-височное поле), это мо-
(средне-височное поле), это мо-
жет быть неподвижный объект на фоне водопада. На карте комбинирования
таких отличительных черт могут быть объединены все выделяющиеся призна-
ки многочисленных зрительных групп в единую общую карту особенностей.
Такая карта будет отражать необычные неожидаемые и выделяющиеся черты
зрительной сцены на любом уровне анализа. Расчет по принципу «Все доста-
ется победителю» выделяет особенное положение на такой комбинированной
карте и подавляет все конкурирующие местоположения. Очевидно, что необхо-
димо знать эту отличительную черту, но она же может ввести в заблуждение:
к примеру, когда вы смотрите музыкальный клип, где много всего привлекает
ваше внимание, у вас появится желание думать о чем-то другом и возникнет по-
требность действовать вопреки тому, что оказывается наиболее ярким по вос-
приятию в данный момент.
Концепция карты отличительной черты отражает значимость, мотиваци-
онную важность и яркость входного сигнала. Многие топографические карты
зрительных зон мозга чувствительны к мотивации и релевантности. Человек
(рис. 8 .31, внизу) представляет себя стоящим в жаркой пустыне, слева с краю
его поля зрения находится холодная кружка пива, точнее, непосредственно за
его прямым полем зрения. Избирательное внимание позволяет значимым сти-
мулам, в данном примере — кружке холодного пива — войти в сознание. Это
можно назвать «нисходящими отклонениями внимания», которые изменяют
карту особенностей на вершине стопки слоев. Предварительное обучение так-
же введено здесь упрощенно в карту особенностей как входной сигнал. Все
топографические карты перекрываются синхронно, решения принимаются по
ним совместно в случаях, когда соответствующие точки на большинстве карт
приводят к одному и тому же общему итогу. Выходом могут являться движения
глаз (что позволит увидеть кружку с холодным пивом) или скрытое смещение
внимания налево, что опять-таки позволит кружке пива войти в зрительное
428
Глава 8
сознание. Сохраняется возможность того, что человек отклонит восприятие
холодного пива в жаркий день и вместо этого сфокусирует свое внимание на
пустыне. Существуют потенциально конкурирующие решения в такого рода
многослойной нейросети.
Важным аспектом модели внимания Итти–Коха является нейросеть «По-
бедителю достается все» (рис. 8 .32). Такие нейросети реализуют «победу»
наиболее активной позиции на объединенных топографических картах, с вход-
ным сигналом от отличительной черты, что представляет такие особенности,
как мотивация, голод и жажда; предварительное знание о значимости, и т. д.
Нейросеть «Победителю достается все» особенно важна, так как она позво-
ляет объяснить сознаваемые опыты двойственных входных сигналов, напри-
мер в случае бинокулярного соперничества. Сознаваемые опыты отличаются
внутренней логичностью, даже когда входные сенсорные сигналы таковыми не
являются. К примеру, большая часть слов английского языка в высокой степе-
ни неоднозначны, но в контексте слова с несколькими оттенками смысла осоз-
нанно воспринимаются только в одном толковании. Таким образом, нейросети
«Победителю достается все» могут участвовать в системе внимания, что отра-
жено на рис. 8 .25, но также они являются важной особенностью сознательного
восприятия. Можно рассматривать сознательное восприятие как результат мно-
гих актов внимания. И в действительности следует допустить, что в головном
мозге между этими двумя механизмами нет различий.
Рис. 8 .32 . Нейросеть «Победителю достается все», в которой самая активируемая точ-
ка на плоской горизонтальной карте подавляет все соседние точки. Вертикальная ось
помечена как «Активность», она может представлять отдельную точку зрительно-то-
пического пространства или суммированную активность многих зрительно-топических
карт. Нейросети «Победителю достается все» очень часто применяются в сетях нейро-
нов по принятию решения, и в мозге при окончательном «решении» по избирательному
вниманию и сознательному восприятию используется механизм «Победителю достает-
ся все». В случае неоднозначного стимула или двойных заданий, как при бинокулярной
конкуренции, мозг переводит одну из двух соперничающих интерпретаций в сознание.
Один из способов для моделирования этого — нейросеть «Победителю достается все».
В случае окулярных доминантных колонн (выше) можно прямо измерить большую об-
ласть коры, которая функционирует именно таким образом, когда зрительная система
получает разные входные сигналы от двух глаз (раздел 2.11). (Источник: Standa�e et al.,
2005.)
Сознание и внимание
429
В обычном смысле слов разница между вниманием и осознанием основана
на том, что есть механизмы контроля внимания, часто определяющие, что ста-
нет осознанным, а что не станет. Имеется много доказательств в пользу такого
утверждения.
Термин внимание используется интуитивно, если существует четкий произ-
вольный или исполнительный аспект. Мы просим человека обратить внимание
на что-нибудь, и это означает, что человек может так поступить или не посту-
пит, в зависимости от исполнительных процессов по принятию решения. По-
этому наиболее простое представление внимания — это произвольный отбор
информации. В экспериментах с участием людей мы постоянно просим участ-
ников обратить внимание на условия стимула в опыте. Произвольное внимание
изучают чаще всего, и, как можно догадаться на основании других глав данной
книги, у человека в него с большой вероятностью вовлечена префронтальная
кора (см. гл . 12).
Как недавно сформулировали Корбетта с соавторами (Cor�etta et al., 2002),
произвольное внимание «участвует в приготовлении и реализации целенаправ-
ленного (нисходящего) отбора на стимулы и ответы». Автоматическое же вни-
мание «не участвует в нисходящем отборе. Вместо этого данная система специ-
ализирована на выделении стимулов, связанных с поведением, особенно если
они особенные или неожидаемые».
При перекрывании двух вышеописанных систем создается полезный пер-
вый захват структур мозга, связанных с избирательным вниманием.
Но если мы вдруг слышим громкий звук, то наше внимание «захватывает-
ся», даже без управления им. Такой захват внимания — важная тема и будет
освещена в следующем разделе по вниманию, определяемому стимулом. Как и
следует ожидать, зрительное внимание может быть больше захвачено лицами
людей, их эмоциональными выражениями и положением тела, по сравнению
с нейтральными стимулами. Интенсивные и неожиданные стимулы, а также
неожидаемые события в целом вызывают в мозге более заметный ответ, чем
контрольные стимулы. Поэтому можно выделить «восходящий» захват избира-
тельного внимания, управляемый стимулом, а также нисходящее целенаправ-
ленное внимание, находящееся под исполнительным контролем.
В реальности произвольное и автоматическое внимание пересекаются. Тре-
нировкой мы привыкаем обращать внимание на звонок телефона. В момент,
когда он зазвонит, мы резко обращаем на него внимание, но происходит это
произвольно или автоматически? Сначала такое действие произвольно, потом
становится более автоматическим. Таким образом автоматическое и произволь-
ное внимание составляют вместе непрерывный ряд. Пожалуй, самое сильное
проявление произвольного внимания происходит тогда, когда мы должны в
течение некоего периода времени напряженно мыслить (см. блок 8.1). Яркий
пример противоположного типа внимания — это случай с громким звуком или
биологически значимым событием (плачущий ребенок), когда сложно не обра-
тить внимание на указанный факт.
Итак, внимание здесь определяется как способность отбирать информацию
в когнитивных целях. На отбор влияют эмоции, мотивация, отличительные чер-
ты, и, по крайней мере, частично, он находится под исполнительным контро-
лем. Следовательно, избирательное внимание функционирует в тесной связи со
430
Глава 8
всеми другими компонентами нашей диаграммы (см. рисунок в начале главы).
Без гибкого произвольного контроля люди не смогли бы справляться с неожи-
данными ситуациями. Мы не могли бы отклониться от правил, даже когда они
уже устарели, или изменить привычкам, связанным с вниманием, с целью из-
влечения преимущества из вновь открывшихся возможностей.
Мы бы медленно отбирали значимые события без внимания, управляемого
стимулом. Человек нуждается и в произвольном, и в автоматическом внима-
нии.
Как уже указывалось, определенные зоны головного мозга отвечают за ре-
ализацию таких функций, как чувственное восприятие, рабочая память, язык
и долговременная память. То же самое верно и для избирательного внимания.
Когда с помощью некоторых методов исследования стали выделять небольшие
зоны мозга, работающие с большой скоростью, мы получили больше фактов о
нейросетях внимания, о том, как в них происходит отбор определенных типов
сенсорной информации. Множество данных сейчас подтверждает основные ис-
ходные факты.
Как показано на рис. 8 .33, внимание рассматривается как избирательная
способность, управляемая либо произвольным контролем, либо стимулом. Ре-
зультат избирательного внимания заключается в улучшении качества отобран-
ной информации, или, как минимум, в попытке этого достичь. Какие имеются
доказательства в пользу усиления внимания?
Рис. 8 .33 . Произвольное внимание: от префронтальной до сенсорной зон коры. Произ-
вольное внимание в восприятии направляется в сенсорную кору лобными и теменными
зонами и, возможно, в другие зоны мозга, для двигательной, языковой и исполнительной
функций. Отметим лобную зону, лобное поле зрения, где осуществляется также про-
извольный контроль над движениями глаз. Теменные зоны, как полагают, участвуют в
пространственно-направляемом внимании. Зрительные области (обозначены красным)
усиливаются механизмами внимания, например гамма-синхронизацией по множествен-
ным зрительно-топическим картам для избирательного пространственного положения
и зрительных признаков. МТ — средне-височное поле. (Источник: Yantis et al., 2008.)
Сознание и внимание
431
Произвольное внимание в головном мозге характеризуется двумя аспекта-
ми (выделено большой желтой стрелкой на рис. 8 .33). Полагают, что лобные
и теменные зоны задействованы в направлении произвольного внимания (обо-
значено синим); в случае зрения целевые области расположены, видимо, в зри-
тельной коре (обозначено красным). Таким образом, в модели внимания надо
прояснить: а) контроль избирательности, б) свойства цели, на которые было
обращено внимание.
На рис. 8. 34 представлено современное предположение о зонах мозга, уча-
ствующих в произвольном внимании, направленном на зрительное местополо-
жение или стимул. Следует отметить, что произвольный контроль внимания
приписывается предлобной (префронтальной) коре. Нисходящая активность
спускается к зрительным картам, связанным с движениями глаз (предлобное,
или префронтальное, поле зрения, теменное поле зрения и верхние бугры чет-
верохолмия), и к зрительно-топическим картам отличительных черт (V1–IT).
Ядро подушки таламуса также содержит зрительно-топическую карту, и, как
предполагается, объединяет знаки/сигналы от отличительных черт, в основном
Рис. 8 .34 . Модель мозга по зрительному вниманию. Шипп проанализировал несколько
моделей мозга по зрительному вниманию. Элементы данной модели сходны с рис. 8 .31,
но при этом компоненты соотносятся с определенными зонами мозга. Заметьте, что,
в дополнение к картам коры, здесь приведены два субкортикальных района с подобием
карты — подушковидное ядро таламуса и верхние бугры четверохолмия. Большая часть
данных зон задействована в контроле за движениями глаз, поэтому не исключено, что в
эволюции зрительно-пространственное внимание развилось на основе избирательного
движения глаз. (Источник: Shi��, 2005.)
432
Глава 8
представляющие контрастные признаки и их местоположения на всех сенсор-
ных картах отличительных черт. Заметим, что данная модель мозга не включает
механизм «Все достается победителю», как постулировано абстрактной моде-
лью на рис. 8 .32.
Нисходящее внимание управляется ожиданиями, и в интервале запаздыва-
ния (рис. 8.35 (а)) испытуемые знают, куда им надо смотреть, но стимул еще
не предъявлен. Однако зрительно-топическая синхронность возникает в двига-
тельно-чувствительных зонах, таких как средне-височное поле (MT), и задняя
межтеменная борозда (зМТБ) в правом полушарии. В течение такого перио-
да внимательного ожидания отмечается значительное сопряжение между MT,
и зМТБ. После указанного интервала запаздывания стимул предъявляется с
одной стороны поля зрения так, что его первый эффект будет в зоне мозга на
Рис. 8 .35. Синхронизирующие множественные карты по усилению внимания. В упраж-
нении по различению двигательного стимула с подсказкой на магнитоэнцефалограмме
видно высокое сопряжение синхронизированных альфа, бета, низких гамма и высоких
гамма в четырех зонах (правого) полушария. Правые полушария «раздуты» математи-
ческими методами, чтобы сделать видимыми кортикальные зоны, скрытые в бороздах.
Возможно, что путем рассылки синхронизированных сигналов в разных сегментах ча-
стотного спектра головной мозг способен одновременно передавать сигналы, не пере-
крывающиеся друг с другом. МТ — средне-височное поле. (Источник: Sie�el et al. 2008.)
Сознание и внимание
433
противоположной (контралатеральной) стороне от самого зрительного стиму-
ла. На рис. 8 .35 время предъявления стимула, альфа и высокие гамма синхро-
низируют области правого полушария.
В период задержки (рис. 8 .35 (б)) возникает преждевременная синхрони-
зация, еще до предъявления стимула, но уже после того, как испытуемому со-
общили о месте будущего расположения ожидаемого подвижного стимула.
В интервале предъявления стимула (рис. 8 .35 (б)) можно наблюдать альфа- и
высокие гамма-волны. Следует подчеркнуть, что лобные поля зрения, участву-
ющие в произвольном контроле внимания и глазных движений, разобщены в
интервале предъявления стимула. Таким образом, существует сложный рису-
нок сопряжения и разобщения между тремя важными зонами зрительного вни-
мания, с синхронизацией и десинхронизацией четырех типов волн в течение
периодов запаздывания и предъявления стимула. Мы можем делать предполо-
жения о типах сигналов между синхронизированными зонами мозга, за счет
чего реализуется гибкий контроль внимания (рис. 8 .37).
3.3 . Опыты по вниманию и опыты сознания
Означают ли термины «внимание» и «осознание» одно и то же? Споры на эту
тему все еще продолжаются. С нашей точки зрения, внимательный отбор ча-
сто приводит к сознательным событиям, а сознательные опыты, в свою очередь,
Рис. 8 .36 . Разделение сознательного восприятия и зрительного внимания. Уарт и Та-
лон-Бодри (Wyart, Tallon-Baudry, 2008) предложили упражнение на внимание (подобное
фланговой задаче, описанной ранее), в котором предъявлялась еле заметная решетка,
близкая к порогу различения. Стимул был примерно в 85% случаев, в половине из
них — выше порога и в половине — ниже порога восприятия. Следовательно, он мог
регистрироваться сознанием примерно в половине случаев, в другой половине — нет,
хотя он также присутствовал. В половине случаев с наличием стимула испытуемым
подсказывали, куда следует смотреть. Поэтому одной независимой переменной было
внимание (определялось по числу серий с подсказками относительно серий без под-
сказок), второй переменной служило осознание видимой решетки. Испытуемых про-
сили определить ориентацию линий в решетке и ее присутствие/отсутствие. Измеря-
ли коэффициент точности и время реакции, регистрировали магнитоэнцефалограмму.
(Источник: Wyart, Tallon-Baudry, 2008.)
434
Глава 8
могут оказывать влияние на внимание. Поэтому предполагается, что все же мож-
но разделять сознательные опыты и отбор, осуществляемый вниманием, но эти
аспекты взаимодействуют. Традиционно используют такое сравнение: луч про-
жектора в темном театре режиссер может попросить направить на конкретного
Рис. 8 .37. Раздельные корреляты мозга по вниманию и осознанию. Уарт и Талон-Бо-
дри (Wyart, Tallon-Baudry, 2008) смогли разделить эффект зрительного осознания и из-
бирательного внимания. (а) Магнитоэнцефалограмма правой затылочной коры в период
от 100 до 500 мс после стимула. (б) Изображение головы. Высокие уровни гамма наблю-
дали в затылочной коре. На графиках внизу (в) показаны времена частот, в цвете приве-
дена шкала F по силе магнитоэнцефалограммы как мера статистической достоверности.
Черный, красный и желтый цвета обозначают высокие уровни гамма, обусловленные
манипуляциями с сознанием (слева) и манипуляциями с вниманием (справа). Зритель-
ное осознание ведет к увеличению силы низких гамма при 250–500 мс (около 60 Гц),
а внимание задействует более высокий гамма-всплеск (около 80 Гц), начинающийся на
350 мс после стимула. Такой результат противоречит интуитивным ожиданиям, так как
можно предположить, что эффект внимания будет предшествовать эффекту осознания.
Остается вопрос — можно ли выявить эффекты внимания ранее в лобно-теменных об-
ластях мозга. Заметьте, что значимые гамма-всплески сопоставляются с тем же локу-
сом в частотно-временном домене на каждом графике в соответствующих сравнениях.
(Источник: Wyart, Tallon-Baudry, 2008.)
Сознание и внимание
435
актера или актрису. Результат такого действия — выделение одного из людей на
сцене. Таким образом, луч прожектора выбирает разных персонажей на сцене,
аналогично тому, как внимание отбирает из массы возможных событий. Осоз-
наваемое событие соответствует появлению того или другого актера в луче про-
жектора. При такой точке зрения содержание сознания и внимания не идентич-
ны друг другу, но они непрерывно взаимодействуют между собой.
Изящный опыт Уарта и Талона-Бодри позволил раздельно управлять созна-
тельным восприятием и избирательным вниманием в рамках одного экспери-
мента (рис. 8 .36 и 8.37) (Wyart, Tallon-Baudry, 2008).
Пока неизвестно, участвуют ли одни и те же области лобно-теменной коры в
сознании и внимании, но, возможно, существует перекрытие. Когда отобранная
сенсорная информация становится сознаваемой, могут активироваться некото-
рые из исходных лобно-теменных локусов, изначально ее выбравшие, таким
образом создается на некоторое время самоподдерживаемая петля обратной
связи.
4.0 . Сон фазы быстрых движений глаз
Не все типичные сновидения происходят в фазу быстрых движений глаз (БДГ),
что определяют физиологическими методами — по быстрым стереотипиче-
ским движениям глаз, по поверхностной ЭЭГ, свойственной бодрствованию,
и по сенсорно-моторному блоку. Довольно большая часть субъективной ум-
ственной деятельности отмечается в медленноволновом сне, хотя и не в ярких
сюжетах, которые мы видим в классических снах. Структура сна (рис. 8 .38)
полезна, но индивидуальные стадии сна могут весьма отличаться. Поэтому мы
будем называть сном фазы быстрых движений глаз (БДГ) те периоды, в которых
присутствуют классические субъективные сны наряду с активностью БДГ.
Аналогично Гобсон и сотрудники (Ho�son et al., 2000) дали такое опреде-
ление сновидениям: «Умственная активность во время сна, которая характери-
зуется живыми сенсомоторными образами, воспринимаемыми как реальность,
несмотря на четкие когнитивные признаки — невозможность или невероят-
ность времени, места, человека и действий; эмоции, особенно страх, восторг
и гнев преобладают над грустью, стыдом и чувством вины, порой достигают
такой силы, что ведут к пробуждению; память даже на очень яркие сны мимо-
летна, быстро угасает после пробуждения, если не предпринять специальные
меры по ее сохранению».
В БДГ-снах заблокированы и сенсорные входы, и моторные выходы, поэто-
му мозг общается только сам с собой. В связи с моторным подавлением могут
возникнуть сны, когда мы как бы парализованы или хотим двинуться во сне,
но чувствуем, что неспособны к этому. Все это может привести к неприятно-
му чувству связанности во сне, если мы осознаем неспособность к движению.
Но мышечное подавление — это абсолютно нормальный компонент сна. Оно
предохраняет нас, прежде всего, чтобы мы не реализовывали в действиях наши
сны.
Память также нарушается в БДГ-снах. Если человека разбудить во время
БДГ-сна, то он может рассказать о ярких зрительных образах, но уже за секун-
ды они забудутся, если не предпринять меры по их повторению. Это подобно
созданию кино во сне, но мы не можем воспроизвести сюжетную линию от
436
Глава 8
одной сцены до другой. Даже люди, отличающиеся хорошим запоминанием
ночных видений, видимо, помнят только незначительную часть своих БДГ-
снов, занимающих от 90 до 120 мин ночью (рис. 8 .38).
БДГ-сны запускаются волнами PGO — активаций варолиева моста-колен-
сны запускаются волнами PGO — активаций варолиева моста-колен-
PGO — активаций варолиева моста-колен-
—
активаций варолиева моста-колен-
чатого тела-затылочной доли, наряду с нейромодуляцией. Кратковременные
подъемы нейронной активности идут от варолиева моста (P), активируют ла-
P), активируют ла-
), активируют ла-
теральное коленчатое ядро (G) таламуса, и затем запускают зрительные образы
через затылочную кору (O). Гобсон и МакКарли (Ho�son, McCarley), выдвинув-
O). Гобсон и МакКарли (Ho�son, McCarley), выдвинув-
). Гобсон и МакКарли (Ho�son, McCarley), выдвинув-
Ho�son, McCarley), выдвинув-
, McCarley), выдвинув-
McCarley), выдвинув-
), выдвинув-
шие раннюю PGO-гипотезу в 1976 г., назвали ее «гипотезой активации-синте-
PGO-гипотезу в 1976 г., назвали ее «гипотезой активации-синте-
гипотезу в 1976 г., назвали ее «гипотезой активации-синте-
за», сделав акцент на активационном всплеске от варолиева моста и его созна-
тельной интерпретации или «синтезе» корой. Обновление гипотезы приведено
позднее (Ho�son et al., 2000) (рис. 8 .39). Пейс-Скотт и Гобсон (Pace-Schott, Ho�-
Pace-Schott, Ho�-
- Schott, Ho�-
Schott, Ho�-
, Ho�-
Ho�-
son, 2002) указали, что БДГ-сны запускаются также изменениями основных мо-
, 2002) указали, что БДГ-сны запускаются также изменениями основных мо-
дуляторов головного мозга, особенно норадреналина, ацетилхолина, серотони-
на (5-HT) и глутамина.
PGO-теория не объясняет содержание снов. Она лишь предполагает, что
кора пытается создать внятную историю из лишенных смысла сигналов ствола
мозга. Но остается загадкой, что же определяет содержание снов. Традицион-
ные идеи, выдвинутые Зигмундом Фрейдом (Si�mund Freud) и Карлом Юнгом
(Karl Jun�), часто воспринимаются скептически; однако сейчас возникли не-
Karl Jun�), часто воспринимаются скептически; однако сейчас возникли не-
Jun�), часто воспринимаются скептически; однако сейчас возникли не-
Jun�), часто воспринимаются скептически; однако сейчас возникли не-
), часто воспринимаются скептически; однако сейчас возникли не-
сколько правдоподобных альтернативных объяснений. Существует общее мне-
ние о большой доле эмоций в содержании снов.
Рис. 8 .38 . Структура сна. Активность таламокортикальной коры во сне меняется очень
значительно. Во время глубокого сна нерегулярные осцилляции ЭЭГ бодрствования
сменяются на высокоамплитудную, медленную и регулярную активность, получившую
название дельта-волны. Четыре стадии способности к пробуждению (Стадии 1–4) от-
личаются по возрастанию дельта-волн, Стадия 4 Не-БДГ почти вся состоит из дель-
та. Примерно каждые 90 мин у спящего человека отмечаются быстрые движения глаз
(БДГ). В это время на ЭЭГ выявляется активность, подобная бодрствованию: быстрая,
нерегулярная и низкая по амплитуде. Люди, разбуженные в БДГ-фазе сна, могут пере-
сказать яркие сновидения, тогда как разбуженные в медленноволновых фазах рассказы-
вают о внутренней речи, сходной с бодрствованием. БДГ-фазы удлиняются во второй
половине ночи. (Источник: Stick�old, Walker, 2007.)
Сознание и внимание
437
Ревонзио (Revonsuo) с коллегами выдвинули теорию моделирования опас-
Revonsuo) с коллегами выдвинули теорию моделирования опас-
) с коллегами выдвинули теорию моделирования опас-
ностей во сне. Данная гипотеза основывается на том, что состояние БДГ-сна
распространено у млекопитающих и птиц, а значит, может иметь важное эво-
люционное значение. Ревонзио (Revonsuo, 2000) предположил, что сны позво-
Revonsuo, 2000) предположил, что сны позво-
, 2000) предположил, что сны позво-
ляют нам повторить в уме ситуации, несущие угрозу. Так люди, получившие
травму, во сне могут вновь переживать события, связанные с ней (в некоторых
случаях даже при бодрствовании). В соответствии с данной теорией учеными
Валли и Ревонзио (�alli, Revonsuo) отмечено, что дети с травмами из зоны во-
�alli, Revonsuo) отмечено, что дети с травмами из зоны во-
, Revonsuo) отмечено, что дети с травмами из зоны во-
Revonsuo) отмечено, что дети с травмами из зоны во-
) отмечено, что дети с травмами из зоны во-
енных действий видят гораздо более пугающие сны, чем дети из других зон.
Другие важные жизненные события, такие как потеря любимого человека, или
даже нормальные стрессовые ситуации, могут также получить отображение в
снах. Спонтанно мысли при бодрствовании возвращаются к повторению теку-
щих задач, и то же самое может происходить со снами. Остается выяснить, бы-
вает ли в снах психофизиологическая адаптация, учитывая, что сны выполняют
функцию, эволюционно связанную с опасностями.
4.1. Сны как состояние сознания
Если людей разбудить в фазу БДГ и попросить описать, что они испытывали за
несколько секунд до пробуждения, то о снах обычно отзываются как о созна-
тельном. ЭЭГ снов фазы БДГ очень напоминает таковую при бодрствовании,
содержит альфа-, бета-, гамма- и тета-волны в сложном сочетании. Поэтому
представляется, что у нас есть два суточных состояния сознания: бодрствование
и БДГ-сны. Электрическая активность бодрствования и БДГ-сна показывает,
что таламо-кортикальная система работает при этом сходным образом. Однако
следует помнить и о важных отличиях БДГ, таких как сенсорное и двигательное
блокирование, высокий уровень эмоциональной активности, и низкий уровень
исполнительного (лобные доли) контроля.
Сюрреалистичная и нерациональная природа снов может быть отражением
низкого уровня исполнительного контроля. Сон — это состояние «гипофрон-
тальное» («гиполобное»), как и дремота, алкогольное опьянение, делириозный
синдром, и прочие состояния низкого самоосознания и самоконтроля (Dietrich,
2002). Более того, лимбические зоны мозга, участвующие в формировании эмо-
ций, обладают во сне более высокой активностью при функциональном магни-
торезонансном изучении по сравнению с бодрствованием (рис. 8.39 и табл. 8 .3).
Некоторые люди после обучения могут во сне понимать, что они сейчас
видят сон, и даже осуществлять некоторый произвольный контроль в течение
сна. Ла Берж и сотрудники (LaBer�e et al., 1981) провели такой опыт «ясного
сна» в лаборатории сна, показав, что некоторые из спящих испытуемых могут
научиться произвольно двигать глазами туда-сюда три раза подряд, по команде,
при звуковом сигнале. Некоторые из находящихся в таком «ясном сне» могли
во сне посчитать до десяти, и затем продемонстрировать экспериментаторам,
что они это проделали, повторив тройное движение глазами как сигнал того,
что прошел интервал в десять секунд. Спящие в «ясном сне» должны были
запомнить и проделать во сне задания, которые они получили при бодрствова-
нии перед этим сном. Для этого им нужно было задействовать исполнительные
функции и использовать семантическую память, а также лингвистическую и
слуховую обработку. Следовательно, видящие «ясный сон» могли контроли-
ровать некоторые из когнитивных функций, которые, как правило, нарушены
в состоянии БДГ-сна.
438
Глава 8
4.2 . Закрепление событий в памяти во время БДГ-сна
Продолжаются споры о роли снов фазы БДГ в закреплении событий в памяти
(в консолидации памяти); публикуются данные за и против такой роли. Види-
мо, эмоциональные эпизоды памяти закрепляются в фазе БДГ в течение послео-
беденного сна (изучать который легче, чем ночной сон) по сравнению с эмоцио-
нально нейтральными событиями (�ishida et al., 2009). Эмоции, как и БДГ-сны,
затрагивают древние системы головного мозга млекопитающих. Существует
трехстороннее перекрывание в зонах мозга, участвующих в эпизодическом обу-
чении (комплекс гиппокампа и неокортекс), в эмоциях (гиппокамп, миндалина,
неокортекс) и в снах (лимбическая активация, таламус и неокортекс). Эмоци-
онально памятные события человечеству необходимо помнить для увеличения
шансов по продолжению рода. Нам следует избегать диких опасных животных
Рис. 8 .39. Функциональные изображения мозга во время фазы сна быстрых движений
глаз (БДГ), активность по функциональному магниторезонансному получению изобра-
жений во время сна. Красные зоны показывают активность с повышением кислорода
крови, к примеру, в зонах эмоций. Синие зоны обозначают сниженную относительно
исходного уровня активность во сне, как в лобных долях. Повышенная активность в
затылочных областях может отражать формирование ярких зрительных образов во
сне. Низкий метаболизм в лобных долях может также объяснять бред, странность и
внезапные разрывы в снах, такие как быстрая смена мест действия, действующих лиц,
и нарушения связности действия. Низкий уровень функционирования лобных долей со-
гласуется с наличием амнезии на сны после пробуждения, а также с низкими уровнями
произвольного контроля. (Источник: Dan�-�u et al, в Squire, 2009.)
Сознание и внимание
439
при следующей встрече с ними; следует помнить, кто из членов клана дал пищу
и кров. Эмоции, связанные с личным и межличностным благополучием: страх,
влечение, безопасность, любовь, голод и сытость, гнев, зависть, гордость; такие
состояния мы обязаны помнить.
В формировании эмоций, как и в закреплении событий в памяти, участвует
гормон стресса кортизол. Его уровень повышается в ночное время, когда воз-
растает и вероятность БДГ-снов (Payne, Lynn �adel, 2004). Более 80% медлен-
Payne, Lynn �adel, 2004). Более 80% медлен-
, Lynn �adel, 2004). Более 80% медлен-
Lynn �adel, 2004). Более 80% медлен-
�adel, 2004). Более 80% медлен-
�adel, 2004). Более 80% медлен-
, 2004). Более 80% медлен-
новолнового сна (МВС) отмечается в первой половине ночи, тогда как количе-
ство снов фазы БДГ удваивается во второй половине ночи.
В ряде исследований выявлено увеличение сопряжения в тета-ритме между
гиппокампом и лобными долями в течение снов фазы БДГ. Поскольку счита-
ется, что для закрепления событий в памяти (консолидации) требуется именно
данный тип тета-координации, эти факты подтверждают роль БДГ в закрепле-
нии событий в памяти (Ho�son, Pace-Schott, 2002). Но самое явное доказатель-
Ho�son, Pace-Schott, 2002). Но самое явное доказатель-
, Pace-Schott, 2002). Но самое явное доказатель-
Pace-Schott, 2002). Но самое явное доказатель-
-Schott, 2002). Но самое явное доказатель-
Schott, 2002). Но самое явное доказатель-
, 2002). Но самое явное доказатель-
ство консолидации памяти основано на расширяющихся знаниях о генной экс-
прессии для синтеза белков, включающихся в состав новых синапсов и других
клеточных структур, что необходимо для улучшения памяти. Пока в основном
такая литература касается других видов млекопитающих. По мере углубления
наших знаний увеличивается число генов, которым приписывается роль в этих
процессах (рис. 8 .40). Мы находимся в центре новой волны фактов и пока мо-
жем привести лишь несколько примеров (см. гл . 16). Однако кажется вероят-
ным, что доказательства консолидации посредством генной экспрессии будут
более четкими, чем все другие факты, доступные на сегодняшний день.
Таблица 8.3
Основные особенности БДГ-снов
Галлюцинации — особенно зрительные и двигательные, но бывают в любом сенсор-
ном проявлении или сразу во всех
Эксцентричность — фантазии (формирование образов странное, необычное или не-
вероятное); прерывистость (образы и сюжет могут меняться, быстро появляться и ис-
чезать); неопределенность (люди, места и события часто причудливо не определены
по меркам бодрствования)
Заблуждение — мы постоянно верим, что мы не спим (если только мы не совершен-
ствуем ясность сознания)
Саморефлексия отсутствует или очень сильно снижена — по сравнению с бодр-
ствованием
Отсутствие стабильности в ориентации — люди, времена и места спутаны, пла-
стичны, не соответствуют друг другу, и прерываются
Линии сюжета сна — объясняют и объединяют все элементы сна вымышленным
образом
Усиленные эмоции — возрастание эмоций и преобладание опасений и возбуждений
Инстинктивные программы — часто встроены (особенно запускаемые адренали-
ном — «бороться или убежать»)
Произвольный контроль — значительно уменьшен
Недостаток памяти — при переходе от сна к бодрствованию, при засыпании и при
переходах от одного сна к другому
Источник: Ho�son J.A. в Squire et al., 2008.
440
Глава 8
На рис. 8 .40 представлен один вид генной экспрессии у крысы, связанный
с консолидацией, как функция состояния мозга. Этот ген называется Zit-268,
его экспрессия выявляется флуоресцентной меткой. Сразу после обучения крыс
умерщвляют, и наблюдают цветную флуоресценцию, выявляющую активность
в различных областях мозга. На данном рисунке приведены шесть левых фрон-
тальных срезов мозга крыс, полученных быстро при бодрствовании, медленно-
волновом сне и БДГ-сне. Видно, что как обогащенная среда, так и контрольное
окружение приводят к уровням экспрессии Zit-268 от среднего до высокого
при бодрствовании (желтый и красный цвет на рисунке). Большинство случаев
Рис. 8 .40 . Генно-контролируемое производство белка для закрепления памяти. Наилуч-
шее подтверждение закрепления в памяти получено в исследованиях на животных по
генной экспрессии для синтеза белков, необходимых при консолидации. Только начи-
нают появляться результаты исследований сотен генов, которые считают участниками
процессов закрепления памяти. На рисунке представлены срезы левых половин коры
мозга крыс, быстро замороженных в каждом из указанных состояний, и помеченных
флуоресцентной меткой на ген Zit-268 . Сравнение проведено для обогащенной среды,
которая, как считается, способствует усиленному обучению, и для контрольных усло-
вий. На нижнем правом рисунке видно, что более высокая генная экспрессия в состо-
янии БДГ наблюдается в группе обогащенной среды по сравнению с контролями. При
бодрствовании выявлена более высокая экспрессия гена Zit-268, чем в состоянии БДГ; в
свою очередь, состояние БДГ превосходит по этому параметру состояние медленновол-
нового сна. (Источник: Kna�ska, Kaczmarek, 2004.)
Сознание и внимание
441
экспрессии происходит при бодрствовании, на втором месте находится БДГ, по-
следнее место занимает медленноволновой сон. Однако следует помнить, что
мы только начинаем исследовать генную экспрессию при консолидации памя-
ти. В обучении задействованы, как считается, сотни разных генов, и это ис-
следование может служить лишь примером того, что появится в ближайшем
будущем (Kna�ska, Kaczmarek, 2004).
5.0 . Глубокий сон: подъемы и спады
Медленноволновой сон (МВС) — наименее осознаваемый период суточного
цикла. В течение МВС может происходить повторное «прокручивание» и за-
крепление в памяти (консолидация) событий, произошедших в течение периода
бодрствования.
Одно из последних открытий касается роли «медленных осцилляций», ко-
торые являются очень широко распространенными кортикальными волнами,
медленными (частота 0,01 Гц), с пиком около 0,75 Гц. Остается дискуссионным
вопрос, являются ли медленные осцилляции (МО) нижней границей традици-
онного дельта-диапазона, легко различимого на необработанной ЭКГ. Считает-
ся, что дельта-волны лежат в пределах от 1 до 3 Гц, но все частоты на ЭКГ не
вполне определены, и пока еще наши знания о них продолжают расширяться.
МО — это серия постепенных пиков и спадов в активности головного мозга,
называемых состояниями подъема и спада. Каждая из таких полуволн может
длиться от 0,5 с до десятков секунд. Как полагают (Destexhe et al., 2007), полу-
волна подъема определяет период активности мозга, напоминающий бодрство-
вание. При состоянии подъема нейроны свободно передают сигналы друг другу,
т. е. происходят тета- и гамма -взаимодействия. Напротив, состояние спада со-
относят со снижением таламокортикальной деятельности до исходного уровня.
Следовательно, состояние подъема МВС можно рассматривать как активное,
сходное с бодрствованием, тогда как состояние спада, видимо, подавляет боль-
шинство межнейронных взаимодействий. Наиболее характерным признаком
МВС могут служить периодические, через каждые несколько секунд, состоя-
ния спада, которые прерывают сходные с бодрствованием состояния подъема на
период, достаточный для того, чтобы предотвратить интегративную обработку.
Представьте, как сильно бы мешала нормальному процессу мышления и разго-
вора необходимость делать паузу через секунду в обычном разговоре.
Полагают, что МО группируют более быстрые волны — тета, альфа, бета и
гамма. Аналогично тому, как медленные приливы и отливы в океане группиру-
ют шестифутовые прибрежные волны, которые расположены на верхушках этих
отливов-приливов. В свою очередь, шестифутовые прибрежные волны группи-
руют мелкие волны, создающие зябь. В море такое группирование медленными
волнами более быстрых выражается в том, что мелкие волны поднимаются на
вершины более медленных и более крупных. Вышеописанная иерархия волн
является заманчивым упрощением, далее будет видно, что есть доказательства
в пользу такого представления.
Несложно представить физиологический механизм группирования более
быстрых ритмов более медленными: состояние подъема МО вызывает мас-
совое возбуждение нейронов, что делает более вероятной деполяризацию
нижележащих нейронов и формирование их аксональных спайков. Поэтому
442
Глава 8
нейроны, участники более быстрых типов волн, получают больше сигналов
по дендритам, что повышает их потенциал, приближая его к порогу деполя-
ризации. Соответственно, медленные состояния подъема можно представлять
как добавление к увеличению потенциала для большого числа нейронов, что
повышает вероятность возбуждения и взаимодействия для таких нейронов. Со-
стояния спадов действуют противоположным образом. Подобный очень общий
эффект медленных волн может реализоваться через объемное проведение, с по-
мощью широко распространенной нейромодуляции, или путем синаптического
межнейронального проведения сигнала (гл. 3).
5.1. Некоторая умственная активность происходит даже
во время медленноволнового сна
При пробуждении в период медленноволнового сна люди примерно в полови-
не случаев рассказывают о продолжающихся умственных процессах. Стериэйд
(Steriade, 2006) писал: «Вопреки утверждению, что сны происходят только в
фазу БДГ, в серии исследований, начавшихся с 1960-х гг., выявлено наличие
процесса мышления, характерного для сна, в течение медленноволнового сна.
Мышление в медленноволновом сне рациональное, с повторами, а в фазу БДГ
внутренние восприятия яркие, мышление становится нелогичным, эмоции бо-
лее выражены». В фазе БДГ сновидения могут занимать до 95% времени; ко-
личество сообщений об умственной активности вне данной фазы снижается до
50%. Стериэйд приходит к выводу, что «головной мозг никогда не «пустует»,
умственная активность имеет место во всех фазах сна» (2006).
Почему нам кажется, что глубокий сон полностью бессознателен? Возмож-
но, потому, что способность к пробуждению снижается по мере того, как в
электрической активности мозга начинают преобладать медленные волны (см.
рис. 8 .18). Сниженная способность к пробуждению означает, что потребуется
более долгое время, чтобы проснуться; более длинный период для полного про-
буждения означает, что наши воспоминания о снах будут хуже. Воспоминание
может померкнуть за секунды. Некоторые люди «просыпаются очень медлен-
но». Это указывает на тот факт, что функции их лобных долей не выходят на
уровень полного бодрствования еще некоторое время после того, как их созна-
ние уже готово к восприятию. В исследовании циркадных ритмов часть людей
называют «жаворонками» (они активны утром), а других — «совами», такие
люди ощущают себя утром вяло, пик их активности наступает позже в течение
дня.
5.2 . Порог пробуждения изменяется на протяжении сна
Глубина сна измеряется порогом пробуждения, т. е. тем, будет ли человек или
животное реагировать на стимул и ориентироваться на него. Порог возбуждения
повышается по мере того, как на ЭКГ растут медленные волны (рис. 8.41). Мед-
ленноволновой сон в основном происходит в первой половине ночи, а основная
доля БДГ — во второй половине. На рис. 8 .41 показано увеличение зон с более
низкой метаболической активностью в головном мозге по мере того, как сон
становится все более глубоким (что определяют по порогу пробуждения). Мета-
болизм зон головного мозга снижается в особом порядке, сначала в медиальных
поясных извилинах, связанных с «самоопределением», и в соответствующих им
таламических ядрах. В медленноволновом сне (3-я и 4-я стадии) метаболизм
Сознание и внимание
443
уменьшается в активных «когнитивных» зонах головного мозга. При этом сен-
сорная кора сохраняет относительную активность, хотя поток сенсорных стиму-
лов и блокирован таламическими переключающими ядрами.
5.3 . Повторное «проигрывание» эпизодов в памяти
и консолидация
Изучение влияния МВС отдельно от БДГ затруднительно, поскольку данные
фазы сна происходят совместно. Но все же медленноволновой сон чаще возни-
кает в первой половине ночи, а БДГ преобладает во второй половине. На этом
основании исследуется консолидация в памяти после «МВС-обогащенных» и
«БДГ-обогащенных» периодов. В целом, «МВС-обогащенные» периоды спо-
собствуют закреплению в памяти явных событий, тогда как «обогащенные бы-
стрым сном» периоды закрепляют процедурные задачи и, возможно, неявные
события. Отмечено увеличение МВС после интенсивного эпизодического об-
учения, и возрастание БДГ после процедурных тренировок.
Другая группа исследований сфокусирована на роли гиппокампа в захвате
эпизодических (осознанных) событий в память (рис. 8.42 и 8.43). Предполага-
ется, что эпизодические воспоминания передаются из гиппокампа в неокор-
текс, при этом они становятся прочными. Гиппокамп играет ключевую роль
при изучении пространства, ориентировании на местности. Удалось точно
определить в гиппокампе животных «клетки пространства», возбуждающиеся
при прохождении конкретной точки лабиринта. Когда крыса пробегает по из-
вестному ей лабиринту, такие клетки возбуждаются последовательно. Позднее
этот же паттерн возбуждения отмечался во время медленноволнового сна, при
этом активировались другие области мозга, такие как таламус, неокортекс и
базальные ганглии (рис. 8.42 и 8.43). Итак, мы можем регистрировать нейро-
Рис. 8 .41 . Снижение регионарного метаболизма с глубиной сна. По мере того как по-
рог пробуждения повышается и все сложнее разбудить спящего, на ЭЭГ наблюдаются
более медленные волны, а метаболическая активность уменьшается в большинстве зон
головного мозга. Однако у глубокого сна есть биологические функции, такие как «про-
игрывание» паттернов возбуждения гиппокампа тех эпизодов, которые произошли за
истекший день. (Источник: Ho�son, J.A., 2009.)
444
Глава 8
ны крыс, последовательно задействованные при изучении этими животными
местоположений; наблюдать, что их мозг вновь повторяет выученную про-
странственную последовательность во время МВС. Тета-ритм представляется
основным ритмом, организующим и передающим между гиппокампом и не-
окортексом, что ведет к консолидации событий в памяти (Jensen, 2005; Buzsaki,
2002).
Ориентирование людей изучали с использованием виртуального города в
видеоигре. После выучивания города у испытуемых наблюдали подобную же
активность гиппокампа, вновь возникающую при МВС. Количество такой ре-
активации коррелировало с обращениями к этим воспоминаниям на следую-
Рис. 8 .42. Медленноволновой сон обеспечивает консолидацию памяти во время «состо-
яний Подъема». Предполагается, что это происходит следующим образом. Три отдела
мозга работают совместно: раннее выучивание эпизодов при бодрствовании кодирует
нестабильное запоминание в гиппокампе и неокортексе. Во время МВС в веретенах
таламуса индуцируются состояния Подъема, запускающие острые спайки гиппокампа,
которые, как полагают, в конечном счете, активируют синхронизированное запомина-
ние в гиппокампе. Так таламическая активность веретена способствует запуску следу-
ющего состояния Подъема, что отображено на ЭЭГ коры. Указанные зоны работают со-
вместно для реактивации недавно помещенных в память событий. Предполагается, что
во время последующего состояния Спада МВС активированные нейроны синтезируют
белки, осуществляющие синаптическую пластичность для более длительного кодиро-
вания в памяти. (Источник: Steriade, 2000.)
Сознание и внимание
445
щий день. Таким образом, полученные данные по человеку и животным приво-
дят к одним и тем же основным выводам (Col�in et al., 2009; �ertes and Kocsis,
1997).
Гиппокамп выполняет множество функций мозга в целом, отражая тот факт,
что у первых млекопитающих он был основным отделом головного мозга.
Крупная система гиппокампа участвует в ориентировании в пространстве и в
выделении эмоционально значимых событий, в целенаправленном контроле по-
ведения, кодировании и извлечении из памяти эпизодических и декларативных
событий, и в консолидации при медленноволновом сне. Различные нейронные
сети (CA1 и CA3) генерируют Острые волны и Пульсацию, объединяющиеся
между собой (как комплекс Узкие волны–Пульсация, показанный справа). Как
видно на предыдущем рисунке, такие ритмы делают возможным восстановле-
ние в памяти событий, выученных при бодрствовании, и их повторение и кон-
солидацию в неокортексе.
Одно из самых лучших доказательств консолидации в памяти медленно-
волнового сна получено при транскраниальной стимуляции мозга для захва-
та медленных осцилляций в коре. Как только такие осцилляции окажутся под
контролем в опыте, возможна проверка гипотезы. На рис. 8 .44 изображено, как
это осуществляется. Испытуемый выучивает задание и через несколько часов
засыпает в специальной лаборатории, в шапочке с электродами для ЭЭГ. Когда
по ЭЭГ наблюдается естественный МВС, то расположенное на голове испыту-
емого электромагнитное кольцо начинает медленно пульсировать (менее 1 Гц),
чтобы увеличить медленные осцилляции. Одновременное записывание отра-
жает медленную волну в коре, наряду с типичными веретеновидными волнами
таламуса и острыми волнами гиппокампа.
В ложном опыте проводится идентичная процедура, но без электромагнит-
ной стимуляции, чтобы контролировать эффект плацебо. Выявлено, что транс-
краниальная стимуляция медленных волн во время МВС улучшает эпизодиче-
скую, но не процедурную память. Видимо, искусственная индукция медленных
волн улучшает условия естественного МВС для консолидации памяти.
Обратите внимание, что известно несколько ритмов головного мозга, играю-
щих существенные роли в консолидации памяти в течение МВС.
Рис. 8 .43 . Острые волны и пульсация в гиппокампе. На схеме гиппокампа показаны
основные группы его нейронов, генерирующие Острые волны и комплексы Пульсации
при активации извлечения из памяти и консолидации в неокортексе в течение медлен-
новолнового сна. (Источник: Beenhaker and Hu�uenard, 2009.)
446
Глава 8
1. Медленные осцилляции (МО) при частоте менее 1 Гц, которые могут пере-
крываться с дельта-диапазоном в 1–3 Гц. Во время состояний подъема воз-
можны пики МО, быстрое возбуждение нейронов и взаимодействия, очень
напоминающие состояние бодрствования. Однако в течение состояний спа-
да МВ кортекс и таламус молчат.
2. Медленные состояния подъема также группируют тета-активность, циклы
которой имеют частоту 4–7 Гц. Известно, что тета участвуют в организации
гиппокампом выученной информации и в передаче такой информации в не-
окортекс.
3. Тета-волны «группируют» активность более быстрых гамма в комплексе
гиппокампа, и ведут кодирование сенсорной информации в гиппокампе
(например, из зрительной коры). Тета также несут сообщения из гиппо-
кампа в лобные доли и другие зоны неокортекса во время консолидации
памяти.
Рис. 8 .44 . Искусственная индукция медленных волн. Отдельные медленные осцилля-
ции (МО) можно усилить индукцией электрического поля на голове. Следует отметить,
что индуцированные МО также вызывают веретена таламуса и острые волны гиппо-
кампа, работающие вместе для осуществления в гиппокампе извлечения из памяти и
консолидации. (Источник: Marshall, Born, 2007.)
Сознание и внимание
447
4. Гамма-активность (30 Гц и выше) входит в тета-активность, служит также
для синхронизации нейронов гиппокампа и неокортекса. Ряд ученых счи-
тает, что именно гамма-активность несет самую детальную информацию,
сгруппированную медленными осцилляциями. Теперь известно, что гамма
имеют очень широкий диапазон (по некоторым оценкам 25–120 Гц, иногда с
активностью даже в 600 Гц и 1200 Гц).
Во всех указанных случаях считается, что более медленные волны повы-
шают вероятность следующей быстрой волны. Состояние подъема медленной
волны увеличивает тета; тета увеличивает гамма; гамма вызывает синхронное
возбуждение отдельных нейронов и их групп в гиппокампе и неокортексе.
6.0. Общее сопоставление
Сознание интересовало человечество с самого начала истории. С улучшением
методов изучения за последние годы достигнут значительный прогресс. В этой
главе рассмотрены некоторые популярные предположения о сознании и внима-
нии.
6.1. Отражает ли сознание функцию глобального
рабочего пространства в головном мозге?
Идея о том, что сознание отличается интегративной функцией, существует
очень давно. Теория глобального рабочего пространства (ТГРП) — это когни-
тивная структура с явной ролью сознания человека. Согласной данной теории,
содержимое сознания включает емкость быстрой интегративной памяти в го-
ловном мозге, где в каждый момент времени может доминировать только один
элемент (Baars, 1988, 2002). Потенциальные события для сознания конкуриру-
Baars, 1988, 2002). Потенциальные события для сознания конкуриру-
, 1988, 2002). Потенциальные события для сознания конкуриру-
ют за доступ в ограниченное рабочее пространство. В ранее описанной модели
по вниманию такое конкурирование осуществляется посредством сети «Все до-
стается победителю» (Itti and Koch, 2000).
По ТГРП доминирующая в данный момент информация широко распро-
страняется по головному мозгу. Так работает нервная система, которую рас-
сматривают как набор широко расположенных сетей, предназначенных для
специальных задач. В системе такого рода координация, контроль и решение
новых задач могут осуществляться путем обмена центральной информацией,
при этом некоторые области, например сенсорная кора, распределяют инфор-
мацию другим отделам. Такая стратегия особенно полезна в случае новых про-
блем, не имеющих известного решения; их можно решить совместной и\или
конкурентной активацией многочисленных специализированных сетей, каждая
из которых может внести вклад в решение. Описанный подход хорошо работает
в компьютерных структурах большого масштаба, которые демонстрируют ти-
пичное поведение ограниченной емкости, когда информация передается путем
глобального рабочего пространства (Baars, 2002).
Теперь есть много фактов, подтверждающих, что сознание — это пер-
вичный фактор для такой функции глобального доступа у человека и других
млекопитающих (Baars, 2002; Dehaene, 2001). Дихаен (Dehaene) и сотрудники
использовали визуальное экранирование с целью сравнения осознанных и не-
осознанных слов, с применением образа/примера, предъявляемого сразу после.
448
Глава 8
Экранированные слова не осознаваемы, но не было препятствия, мешающего
их увидеть. Анализ изображений головного мозга показал, что и осознаваемые
и неосознаваемые слова вызывали в коре активацию зрительной зоны и зоны
распознавания слов, связанных, как известно, с анализом локализации стимула,
формы, цвета и идентификации слова. Однако осознаваемые слова запускали
активность в 12 раз большую в этих зонах, чем идентичные неосознаваемые.
Более того, осознаваемые слова вызывали гораздо большую дополнительную
активность в височной и лобной зонах коры. По-видимому, связанная со стиму-
лом активность в осознанном случае распространяется широко от зрительных
областей в другие области головного мозга.
Вовлечение сознания особенно полезно, когда возникает необходимость в
комбинировании и интеграции новой информации, и в задействовании допол-
нительных ресурсов головного мозга. Вероятно по этой причине эпизодиче-
ская память (память на осознанные события) имеет такое центральное разме-
щение в головном мозге; гиппокамп и неокортекс постоянно взаимодействуют
с целью сохранения и извлечения из памяти эпизодических моментов. Даже
неявное обучение мобилизуется осознаваемыми стимулами, что и описано
в данной главе.
Из ТГРП возникает проверяемая гипотеза относительно осознанных и не-
осознанных аспектов восприятия, обучения, рабочей памяти, произвольного
контроля и личностных систем в головном мозге. Данное положение имеет зна-
чительное сходство с другими биологическими теориями, такими как нейраль-
ный дарвинизм и динамические теории функционирования головного мозга
(Edelman and Tononi, 2000). Дихаэн с сотрудниками разработали эксперимен-
Edelman and Tononi, 2000). Дихаэн с сотрудниками разработали эксперимен-
and Tononi, 2000). Дихаэн с сотрудниками разработали эксперимен-
and Tononi, 2000). Дихаэн с сотрудниками разработали эксперимен-
Tononi, 2000). Дихаэн с сотрудниками разработали эксперимен-
Tononi, 2000). Дихаэн с сотрудниками разработали эксперимен-
, 2000). Дихаэн с сотрудниками разработали эксперимен-
тально проверяемые модели по аспектам данной теории (Dehaene, 2001). Ряд
экспериментов привели, в целом, к положительным результатам.
Дихаэн с сотрудниками предложили нейрональное осуществление архитек-
туры глобального рабочего пространства, что получило название нейрональ-
ное глобальное рабочее пространство (рис. 8.45). Согласно их представлению,
сенсорные стимулы мобилизуют возбуждаемые нейроны с дальнодействую-
щими кортико-кортикальными аксонами, что ведет к рождению паттерна гло-
бальной активности среди нейронов рабочего пространства. Любой из таких
глобальных паттернов может подавлять альтернативные паттерны среди ней-
ронов рабочего пространства, за счет чего блокируется обработка сознанием
альтернативных стимулов. По модели нейронального глобального рабочего
пространства, сознание представляет собой нелинейную функцию особен-
ностей стимула (т. е. постепенное увеличение заметности стимула должно
сопровождаться резким переходом нейронального рабочего пространства в
соответствующий паттерн активности). Такое «воспламенение» рабочего про-
странства соответствует представлению, что сознание работает по принципу
«все-или-ничего», а не в градуальном режиме. Глобальное нейрональное ра-
бочее пространство применялось для моделирования ряда опытных явлений
в сознании. В одном из исследований спонтанная активность рабочего про-
странства блокировала обработку внешних восприятий, аналогично тому как
при слепоте по невниманию, когда нормальные люди, напряженно занятые
умственной работой, не замечают заметного, но не относящегося к текущему
занятию, стимула.
Сознание и внимание
449
Подобный же феномен — это мигание внимания, при котором люди не мо-
гут заметить стимул S2, предъявленный им в коротком временном интервале
(100–500 мс) после первого стимула S1. В данном рассмотрении модели ней-
S1. В данном рассмотрении модели ней-
1. В данном рассмотрении модели ней-
ронального глобального рабочего пространства предъявление стимула S1 вы-
S1 вы-
1 вы-
зывает направленные вверх и вниз сигнальные потоки, приводя к длительной
активности, которая может лежать в основе сознательного ответа на стимул у
человека. Но при предъявлении S2 сразу после S1 происходит активация только
нисходящих потоков, так как глобальная активность сосредоточена на S1. Такая
задержка длительной активности, связанной с S2, может отражать недостаток
доступа в сознание во время эффекта мигания внимания.
К сегодняшнему дню такое представление подтверждают работы разных
исследователей. Наиболее яркой, наверное, можно считать гипотезу трансля-
ции. Согласно этой гипотезе, наполнение сознания мобилизует множественные
источники головного мозга, в таком процессе идет глобальное распределение
Рис. 8 .45. Нейрональное глобальное рабочее пространство, подтверждающее опыты
с сознанием. Дихаэн с соавторами (2001) предложили модель нейронального глобаль-
ного рабочего пространства, относительно длиннодистантного взаимодействия в коре
предлобной зоны. Другие специализированные области коры избирательно включают-
ся в рабочее пространство сознания. Большая часть сетей мозга активируется автома-
тически (бессознательно). Эта модель объясняет основные функции головного мозга,
например перцепционную категоризацию и долговременную память, а также опытные
парадигмы, такие как мигание внимания и зрительное обратное маскирование. (Источ-
ник: Dehaene, с разрешения.)
450
Глава 8
информации (и в мозге, и в исходных моделях искусственного интеллекта, где
использовалось глобальное рабочее пространство). В «модели общества» го-
ловного мозга, сравнимой со всемирной паутиной Интернета, детали обработ-
ки информации происходят в большом числе локальных нейронных сетей. Роль
глобального рабочего пространства, как чата в Интернете, состоит в обеспе-
чении взаимодействия между множественными потоками целенаправленных
процессов, и в распространении доминантных сообщений среди этих потоков,
потенциально конкурирующих между собой. Как и в случае чата в Интернете,
некоторые «голоса» будут доминировать над прочими, наиболее часто домини-
рующие получают наиболее широкое распространение. Одновременно множе-
ственные потоки обработки выходят из чата, или в данном случае — вовне из
глобального рабочего пространства.
Определенное число фактов указывает на то, что осознанное познание четко
связано с широким распространением кортикальной активности, в основном в
направлении лобно-теменной и срединной височной зон (см. гл . 1). Необходи-
мо применять условия сравнения при неосознании в экспериментальных про-
верках любой теории о функциях сознания. По имеющимся сейчас данным, при
таких условиях активируются только локальные зоны, например локальный
зрительный процесс. Эти результаты согласуются с ТГРП, которую разделяют
ныне ряд ученых и философов (Baars, 2002). Баарс и Франклин (Baars, Franklin,
2005) предположили, что осознаваемые моменты крайне важны для мобилиза-
ции таких функций рабочей памяти, как повторение в уме, кодирование, извле-
чение из памяти и преднамеренный отчет.
Альтернативная гипотеза заключается в том, что, поскольку любое осознава-
емое событие подлежит сообщению, предлобно-теменная составляющая может
быть связана с готовностью к такому сообщению, а не с осознанным опытом
как таковым. Такую альтернативу крайне сложно экспериментально проверить
окончательно. Но по мере того, как наше понимание разных функциональных
зон коры становится все лучше, такая возможность может стать реальностью.
Большое число фактов свидетельствует в пользу того, что сознательные со-
бытия отражаются в более широкой кортикальной активности, нежели соответ-
ствующие сравнительные случаи. И главный теоретический вопрос заключает-
ся в выяснении вклада лобно-теменной активности в сознательное восприятие.
Бессознательные состояния отражают другую сторону гипотезы глобальной
трансляции: при таких состояниях отмечают в лобно-теменной области сни-
женный метаболизм в глубоком сне, коме, вегетативных состояниях, потере со-
знания при эпилепсии, и при общей анестезии (Baars et al., 2005; Blumenfeld,
2008). Мы наблюдаем твердые доказательства этого.
На рис. 8.46 из работы Кантеро и соавторов (Cantero et al., 2005) представле-
на длиннодистантная синхронизация гамма-ритма, отражающая быструю пере-
дачу информации, что происходит четко при пробуждении, но не во время МВС
или быстрого сна. Это исследование проведено на эпилептиках перед операци-
ей с имплантированными интракраниально электродными матрицами, т. е. при
максимально возможном на сегодняшний день отношении сигнал/шум. Следу-
ет обратить внимание, что длиннодистантная синхронизация гамма отмечается
в неокортексе, а также между гиппокампом и корой.
Сознание и внимание
451
Как уже упоминалось выше, считается, что гамма-активность переносит-
ся на медленных волнах. Особенно важным является домен в 100 мс, так как
это минимальное время для интеграции в сознании при восприятии к примеру.
Таким образом, два дискретных сенсорных события любой модальности мо-
гут быть интегрированы в единое явление в сознании, только если они отделе-
ны друг от друга не более чем в 100 мс (приблизительно). Интервал от 100 мс
увеличивается при восприятии речи и музыки, танца, когда отдельные явления
интерпретируются в гораздо более протяженной цепи. Но интервал 100 мс хо-
рошо выполняется для таких отдельных явлений, как быстрые щелчки, краткие
звуки, вспышки, сенсорные постукивания и т. д . По другим источникам, также
выявлены 100 мс или серия 10 Гц для микросостояний сознания на основе дан-
ных ЭЭГ и других (Freeman, 2006; Lehmann, 2010).
В этой связи очень интересен факт того, что Доэсбург с коллегами (Does�ur�
et al., 2008) обнаружили при записи зрительных стимулов путем магнитной эн-
цефалографии синхронизацию гамма именно приблизительно в 100 мс на боль-
ших областях коры в ответ на латерализованные зрительные сигналы. Именно
это и следует ожидать при мгновенной «передаче» в сознании мгновенных зри-
тельных входных сигналов (рис. 8 .47). При этом в первые 200 мс после стимула
Рис. 8 .46 . Длиннодистантная гамма-синхронизация при пробуждении, МВС и быстром
сне. В верхнем ряду приведены ЭЭГ неокортекса, в нижнем — ЭЭГ гиппокампа-коры.
При пробуждении гамма-синхронизация стойкая, в среднем около 50%. Гамма счита-
ется в интервале 35–58 Гц. Горизонтальная ось отражает долю от всех возможных со-
четаний электродов, показывающих синхронизацию. (Аналогичные данные получены
для статус-зависимой местной гамма-когерентности.) (Источник: Cantero et al., 2005.)
452
Глава 8
синхронизация отсутствует, но в интервал от 300 до 400 мс происходит взрыв
гамма-синхронизации по всему головному мозгу (синхронизация происходит с
минимально возможным временем задержки, используется здесь как наиболее
общий термин). В крайней справа колонке рис. 8 .47 приведены теменные вос-
принимающие структуры на противоположной стороне коры по отношению к
зрительному сигналу (т. е. контралатерально), где происходит моментальный
взрыв гамма-синхронизации примерно в районе 100 мс, виртуально это и яв-
ляется картиной «глобального транслирования» в головном мозге. Описанные
Рис. 8 .47. Момент активации сознания? Доэсбург (2007) с помощью магнитной энце-
фалографии наблюдал всплеск гамма-синхронизации большого диапазона через 300
мс после начала визуального стимула, латерализованного справа или слева. Значимая
синхронизация на 39 Гц между положениями рецептора показана черными линиями,
десинхронизация показана белыми линиями. Зрительный стимул предъявляли справа
или слева. Через 100 и 200 мс преобладала десинхронизация, отмечены некоторые ос-
цилляции длиннодистантной синхронизации (черные линии). На 300–400 мс преобла-
дает синхронизация в многочисленных местах связывания. В крайней справе колонке
показаны только теменные рецепторы. Черные линии указывают на высокую синхро-
низацию контралательно к зрительному стимулу, что и ожидалось на основании пере-
крестной анатомии коры. Видно, что из теменных источников (P7 слева и Р8 справа)
активность распространяется как к расположенным на той же стороне, так и к кон-
тралатеральным зонам коры. Эти факты согласуются с гипотезой о глобальном транс-
лировании на весь мозг в течение момента осознания, длительностью около 100–300 мс.
Поскольку циклы тета и альфа примерно подходят по этому параметру, предполагается,
что альфа или тета могут переносить синхронизованную гамма-активность кратно в
100 мс всплесках. (Источник: Does�ur� et al., 2007.)
Сознание и внимание
453
данные согласуются и с другими исследованиями, где показана особая роль
тета- и альфа-сигналов, так как эти медленные типы волн осциллируют около
10 Гц, тем самым они могут давать подходящие волны-переносчики для бы-
стрых осцилляций гамма синхронизации.
Из разных источников имеется несколько подтверждений того факта, что
когерентные гамма-всплески связаны с моментами восприятия в сознании и
имеют ритмы в районе тета или альфа. С целью проверки того, применимо ли
это к сознательным явлениям, и что обратное не является необходимым, в ряде
лабораторий использовали бинокулярную конкуренцию, зрительное обратное
экранирование, мигание внимания и другие методы. Доэсбург с коллегами
(Does�ur� et al., 2009) применили бинокулярную конкуренцию для «Выявле-
ния динамики крупно-масштабных осцилляций нейросетей при опосредовании
зрительного восприятия». Их заключение: «Предположим, что отдельные мо-
менты восприятия реализуются временной синхронизаций гамма-волн связан-
ных кортикальных зон; дезинтеграция и повторная интеграция таких ансам-
блей отслеживает время для продолжающихся тета-осцилляций. »
С помощью несколько иных экспериментальных подходов и методов анали-
за Жиллард и его сотрудники (Gaillard et al., 2009) пришли к следующим вы-
водам: «Неосознанная обработка маскированных слов наблюдалась во многих
зонах коры... но не было когерентой длиннодистантной активности нейронов,
поэтому предполагается в этом случае быстро затухающая волна прямой по-
ложительной связи. Напротив, при осознанной обработке немаскированных
(осознаваемых) слов отмечена конвергенция четырех четких нейрофизиоло-
гических признаков: стойкие изменения напряжения, особенно в предлобной
коре, большие увеличения спектральной силы в гамма-полосе, прирост в длин-
нодистантной синхронизации фазы в бета-диапазоне, и возрастание в обуслов-
ленности Грейнджера большого диапазона. Мы считаем, что все это служит
для формирования четких окон в одном и том же рассредоточенном состоянии
сознательной обработки».
На рис. 8 .48 представлено принципиальное различие между условиями со-
знаваемых и неосознаваемых стимулов в экспериментах, проведенных Жил-
лардом и его сотрудниками. Это различие представляется одиночным высоко-
амплитудным скачком синхронизации сразу после 20 Гц, и в районе от 400 до
500 мс после начала действия стимула. На данном графике показана разница
между условиями сознательными и бессознательными (что носит название «не-
маскированный эффект», так как контроль был зрительно обратно маскирован,
поэтому являлся неосознаваемым стимулом).
На сегодняшний день совпадение между данными из разных источников
является удивительным. При тщательном мониторировании головного мозга,
особенно при применении чувствительных методов, таких как внутричерепная
регистрация, наблюдается краткая гамма-синхронизация между удаленными
зонами коры при условиях сознания, но не в точно соответствующим им не-
осознаваемых условиях. Такая гамма-синхронизация, видимо, переносится не-
которыми более медленными формами волн в домене частот тета-альфа. Сход-
ные эффекты отмечены в различных исследованиях, описанных в данной главе.
Похожие типы активности на макроуровне можно получить при поверхност-
ной ЭЭГ или МЭГ, по изменениям состояния сознания во время сна, в других
454
Глава 8
бессознательных условиях и в ряде когнитивных тестов. Учитывая совпадение
результатов из разных исследований, можно уверенно заключить, что формиру-
ющееся сейчас понимание этих актуальных аспектов является верным.
6.2 . Передача сигналов повторным входом и степень
интеграции осознаваемых явлений в головном мозге
Таламокортикальный комплекс обладает интерактивной связностью по срав-
нению с другими структурами, например с мозжечком, и поэтому именно
эта часть головного мозга стоит в центре описания в данной главе. Джеральд
Эдельман (Gerald Edelman) с сотрудниками провели детальные теоретические
исследования таламокортикальной системы в рамках теории нейродарвинизма
(НД). Согласно НД развитие головного мозга и его динамика по природе своей
соответствуют селекционизму, а не инструкционизму, в отличие от компьюте-
ров, с их четкими инструкциями в символах.
Селекционизм обладает четырьмя основными признаками, аналогично ви-
дам живых существ.
1. Набор элементов (нейроны, синапсы, или динамические ансамбли клеток)
обладает разнообразием, как популяции нейрональных групп в головном
мозге.
2. Такие элементы могут размножаться или увеличиваться в числе.
3. Процесс селекции действует на основе разнообразия; например разный
успех при размножении способствует отбору определенных потомков.
Рис. 8 .48. Длиннодистантная гамма-синхронизация в осознаваемом, но небессознатель-
ном видении. Различие между сознаваемым и неосознаваемым зрительным входным
сигналом показано в данном эксперименте кратким, но достоверным статистически
всплеском длиннодистантной гамма-синхронизации несколько выше 20 Гц (стрелка)
Четыре конвергентных метода указывают на различие между этими двумя состояниями,
включая обусловленность Грейнджера, что позволяет статистически вычесть обуслов-
лено связанные активности в комплексе и в параллельных временных сериях, постоян-
но наблюдаемые в головном мозге. (Источник: Gaillard et al., 2009.)
Сознание и внимание
455
4. «Вырождение» или функциональная избыточность, способность разных со-
четаний элементов выполнять одну и ту же функцию, в таких системах явля-
ется врожденной. Функциональная избыточность — стандартный признак
технических систем, а также обычная черта живых организмов.
В головном мозге селекционизм применим и к нейрогенезу, и к сиюминут-
ному функционированию. Отбор в головном мозге ведется по ценности, отра-
жающей особенность данного события, она может быть положительной и от-
рицательной. Ценность аналогична давлению отбора в эволюции. В головном
мозге в это включается удовольствие, боль и эмоциональная окраска. В конеч-
ном счете, согласно НД, пространственно-временная координация в головном
мозге запускается путем повторного входа сигналов, рекурсивного обмена сиг-
налами между зонами нейронов на протяжении реципрокных связей с массо-
вым параллелизмом.
Согласно представлению Джеральда Эдельмана, эволюционно нервные си-
стемы, лежащие в основе сознания, возникали в результате их способности к
параллельной интеграции большого числа сенсорных входов и двигательных
выходов. Это делало возможным различение сенсомоторных сигналов в много-
мерном пространстве, способствуя выбору адаптивного поведения. Первичное
(сенсорное) сознание связано с взаимодействием между перцепционной кате-
горизацией и памятью; при этом первичное сознание работает со ссылкой на
наличие мультимодального фона перцепционных и моторных событий. Оно
обеспечивает животное избирательной селективностью, приспосабливаемо-
стью и способностью планировать.
Эдельман, как и другие теоретики, различал «первичное» сознание и «вы-
сокоорганизованное» сознание, связанное у людей с языком, временной устой-
чивостью, автобиографичной памятью, а также способностью представлять и
планировать будущее.
В гипотезе динамического ядра Эдельман с сотрудниками (2000) предпо-
ложили, что любое конкретное событие сознания сочетает на высоком уров-
не информационно-обогащенное разделение и интеграцию. Нейрональные
механизмы сознательного опыта включают в себя функциональный кластер в
таламокортикальной системе, в пределах которого повторные межнейронные
взаимодействия приводят к серии отдельных состояний. Границы динамиче-
ского ядра со временем могут изменяться: одни нейронные группы будут его
покидать, другие же будут в него включаться.
Согласно заключениям исследователей Тонони и Эдельмана (Tononi, Edel-
Tononi, Edel-
, Edel-
Edel-
man, 2000)), высокие значения параметра, называемого «степень интеграции
нейронов», характеризуют события в сознании. Параметр С отражает степень
того, насколько динамика нервной системы интегрирована и дифференциро-
вана. Модельные исследования позволили считать, что таламокортикальная
система хорошо подходит для формирования динамики высокой степени инте-
грации нейронов.
Тонори предположил, что сознание соответствует способности нервной
системы к интеграции информации, определяемой формально как комбиниро-
вание большого набора состояний, таких как рассматривание объекта наподо-
бие монитора компьютера. Такое информационно-теоретическое предположе-
ние основывается на идее о том, что любое происшествие в сознании тотчас
456
Глава 8
вычеркивает возможности большого числа альтернатив и поэтому является
высокоинформационным выделением. Тонони выдвинул формальный крите-
рий степени сознания, возможного в данной системе, — критерий фи, опреде-
ляемый как количество причинно связанной информации, которая может быть
интегрирована через самое слабое звено данной системы. В рамках этой тео-
рии, сознание, измеряемое по фи, характеризуется как предрасположение или
потенциальная возможность. И степень интеграции нейронов, и фи можно рас-
сматривать как меру пространственного аспекта в балансе между дифференци-
ацией (разделением) и интеграцией в нервной системе.
Сет и его сотрудники (Seth et al., 2006) выдвинули полезные дополнитель-
ные критерии динамики нейронов для оценки других аспектов вышеуказанного
баланса, в том числе степень интеграции во времени (временная степень ин-
теграции) и по разным уровням описания (рекурсивная степень интеграции).
Временная степень интеграции отражает тот факт, что сознание простирает-
ся во времени несколькими способами. По данным многих работ, необходим
интервал в 100 мс для встраивания сенсорного стимула в сценарий сознания,
потенциалы готовности нейронов могут появляться за несколько сотен мил-
лисекунд до осознания намерения действовать. Рекурсивная степень интегра-
ции отражает то, что головной мозг обладает организацией на множественных
уровнях, от молекулярных контактов в пределах отдельных синапсов до по-
вторных взаимодействий между различными областями мозга. Для адекватного
описания нейрональной динамики, касающейся сознания, может понадобиться
сочетание критериев.
Исследователи Эдельман, Тонони и Спорнс предположили, что из четы-
рех возможных таламокортикальных паттернов способов связи (рис. 8 .49) тем
паттерном, который реализует функции сознания, является указанный справа,
названный степенью интеграции. Аппарат связывания, организованный пол-
ностью случайным образом, неспособен поддерживать специфичность содер-
Рис. 8 .49. Четыре возможных таламокортикальных паттерна способа связи. За по-
следние годы произошел прогресс в понимании паттерна соединения в центральных
отделах мозга, таламокортикальной системе. Некоторые предположения о состоянии
сознания и его наполнении включают и утверждения о типе динамического соединения,
которое может быть необходимо для мозга в сознании. К примеру, такие предположения
нацелены на объяснение того, почему мозжечок не считается поддерживающим объем
сознания, в то время как кора больших полушария и таламус в комплексе это осущест-
вляют. (Источник: S�orns and Tononi, 2002.)
Сознание и внимание
457
жимого; система энтропии будет терять информацию. Третий из возможных
паттернов, Интеграция/Объединение, очень полезен в некоторых биологиче-
ских функциях, но не обладает высоким уровнем гибкости, характерным для
функций сознания в головном мозге. Степень интеграции — формальный кри-
терий, учитывающий многомерную различимость между разными входами и
выходами, он также способен интегрировать очень большое количество таких
состояний единым образом. Вышеизложенные предположения можно форма-
лизовать, они были проверены на детальных моделях таламокортикальной си-
стемы, мозжечка и других структур головного мозга (Izhikevich, Edelman, 2009).
6.3 . Требуется ли для сознания личное познание?
Интуитивно большинство из нас считает, что опыт сознания включает и «лич-
ное познание» — свою точку зрения на мир, которая может поддерживаться те-
менными и предлобными зонами коры (Baars et al., 2005; см. гл . 12). Повседнев-
но мы говорим: «Я проснулся», «Я вижу чашку кофе», «Я потерял сознание»,
«Не могу себя контролировать» и т. д. Но существует обширная литература по
философии, которая критически оценивает такую идею. На протяжении многих
лет «наблюдение над собой» высмеивалось как «заблуждения маленького чело-
века». Проблема, по мнению таких критиков, заключается в следующем: чтобы
наблюдение над собой имело смысл, наблюдателю необходимо сесть в голов-
ной мозг, наблюдая за входящим сенсорным потоком. Но чтобы такой малень-
кий наблюдатель внутри мозга имел смысл, он также потребует маленького на-
блюдателя внутри себя, и так до бесконечности. Такое бесконечное обратное
движение ничего не объясняет.
Однако не все варианты наблюдения над собой ведут к бесконечной регрес-
сии (Baars, 1988). Возможна исполнительная способность в лобных и теменных
долях, где заложена интерпретация информации от органов чувств в терминах,
важных для принятия решения, например «Для меня это хорошая новость?
Если нет, какой ущерб это может принести? Следует ли убегать? И буду ли я
испытывать смущение, если убегу?». Существует много доказательств испол-
нительной нейронной сети в головном мозге человека (гл. 12).
Научное представление может сейчас вновь обратиться к идее исполнитель-
ного «Я». Зоны лобной доли головного мозга, показанные на рис. 8 .50, совпа-
дают с поврежденными в классическом случае Финеаса Гейджа и с лобными
зонами других пациентов, страдающих от радикальных изменений личности и
нарушенного самоконтроля. Чувство действия в произвольном контроле также
зависит от медиальных и латеральных лобных зон, а правая теменная зона —
решающая для «перспективы себя» в неврологическом состоянии теменного
игнорирования (Baars 2002; Baars et al., 2003)*
.
Необходима ли «Я система» для опытов сознания? Появляющаяся литерату-
ра может дать нам первую научную зацепку по этим вопросам.
Одной из областей мозга, поддерживающих «перспективу/проекцию себя»
и чувство личного действия в контроле действия, может быть задняя теменная
кора. Эта же область может быть вовлечена в потери сознания во время наркоза
(Alkire, Tononi, 2008).
* Имеется в виду феномен игнорирования (синдром Балинта) при поражении теменной доли правого
полушария. — Прим. ред. перев.
458
Глава 8
Вильям Джеймс указывал, что «Я как наблюдатель» не совпадает со «мной»
как объектом восприятия и мысли. То есть «Я» — это не лицо, которое мы
видим в зеркале, и не воплощенное представление о нас таких, какими могут
нас видеть другие. Скорее «Я» — это точка зрения воспринимающего чело-
века, произвольно действующего и думающего. Вполне возможно, что «Я»
—
умозаключительная сущность, и даже нечто мифически или культурно при-
обретенное. Ведь люди постоянно делают умозаключения о разного рода не-
существующих объектах, и сам по себе факт того, что мы заявляем о суще-
ствовании чего-либо, не доказывает, что это действительно существует. Однако
перспектива «Я» представляется универсальной и фундаментальной в разных
культурах. У маленьких детей с самого раннего возраста есть сенсомоторное
представление, и, вероятно, эмоциональное. Большая часть наших эмоций свя-
зана с впечатлениями от нашего собственного благополучия или опасности при
познании себя. Чувствование боли — это то же самое, что негативная оценка
при познании себя. Считается, что такие эмоции, как стыд, вина и гордость, яв-
ляются «эмоциями самосознания», т. е. чувствами, отражающими, по нашему
представлению, восприятие нас обществом.
Рис. 8 .50. Предлобная медиальная зона может отвечать за осознание «себя как на-
блюдателя». Связанные с собственной личностью зоны коры также являются как бы
транспортными узлами с плотным движением, что выявлено по картирующим иссле-
дованиям белого вещества — основных сигнальных узлов и дорог в коре и таламусе.
Соединения в головном мозге имеют связь с математикой «маленького мира» как струк-
турно, так и функционально, что исчисляется «сигнальными потоками перевозок». (Ис-
точник: Ha�mann et al., 2008.)
Сознание и внимание
459
6.4 . Почему сознаваемые события
подлежат регистрации?
Наши знания о том, что другие обладают сознанием, основываются на том, что
они нам сообщают. Это, к счастью, в большинстве случаев совпадает с нашим
собственным опытом, по крайней мире в перцепционном мире. Любая научная
теория должна, в конечном итоге, объяснить и свои исходные данные. Но до
сегодняшнего дня предприняты лишь несколько попыток объяснения того, по-
чему осознаваемые события подлежат регистрации.
Объяснение, отчасти тривиальное, состоит в том, что вперед направлен-
ное «глобальное транслирование» перцепционного содержимого из сенсорной
коры к лобно-теменным зонам также активирует и зоны речи, в том числе поле
Брока, необходимые для сообщения событий в сознании. Но такое объяснение
не вполне подходит. Люди, страдающие афазией, все равно могут регистриро-
вать события сознания; даже парализованные люди могут движениям глаз вы-
бирать буквы на большом экране-клавиатуре, тем самым «печатать» разговор.
На более глубоком уровне объяснения можно сказать, что на перцепцион-
ные объекты и события можно указать, и это может проделать даже очень ма-
ленький ребенок. Дети со взрослым постоянно играют в игру типа «Посмотри!
Вот игрушка!» и поздней «Посмотри! Вот (и название предмета)». Очевидно,
важной составляющей взросления является узнавание общей социальной и
межличностной реальности объектов, событий и людей. В базовых когнитив-
ных терминах можно считать, что сознание тесно взаимосвязано с тем, что в
лингвистике называют deixis, область объектов, на которые можно «показать».
Определенное количество лингвистических выражений обслуживают процесс
deixis, что может быть фундаментальным аспектом совместной социальной ре-
альности людей. Deixis включает мир восприятий, а также социальный мир, и
даже общее представление о невидимом мире. Дети могут стандартно беседо-
вать о Санта-Клаусе и фее.
С философской точки зрения, сознание может играть роль в объектива-
ции — создании мира объектов, обладающих известными или согласованно
принятыми свойствами, которые сохраняются во времени. Важно, что дорсаль-
ный поток зрительной коры прямо не ведет к сознательному выходу, хотя яв-
ляется существенным как для пространственной локализации себя, так и для
таких действий, как доставание чего-либо. Следовательно, не все зрительные
события могут быть воплощены в объекты, не все они обязательно превратятся
в объекты в сознании. И действительно крайне важно, что дорсальный и вен-
тральный зрительные потоки должны быть разделены друг от друга, чтобы не
было интерференции между четко обозначенными по времени зрительно-дви-
гательными действиями.
Объективация — превращение событий в стабильные объекты восприятия
или мысли — тоже является важным компонентом мышления людей. Люди по-
стоянно совершают логическую ошибку овеществления, превращая абстракт-
ные понятия, например «демократия», в как бы реальные сущности. Такой про-
цесс иногда полезен, но может привести к заблуждениям. Также мы постоянно
замораживаем динамические события, такие как аспекты экономики, биологии
и физики, обращаться с которыми следует более точно с помощью математиче-
ских методов для быстро изменяющихся объектов. Расчеты Ньютона разрабо-
460
Глава 8
таны строго для описания динамических явлений, которые можно маркировать
словами только при очень большем упрощении. Благодаря многочисленным
математическим разработкам стало возможным измерять и описывать динами-
ческие системы (например головной мозг) с высокой точностью. Однако чело-
веческий язык и наше восприятие не успели за темпом математических усовер-
шенствований. Мы по-прежнему говорим об «экономике» и «головном мозге»
по сути в статистических понятиях.
Возможно допустить, что как минимум такая избыточная логическая ошиб-
ка овеществления отражает стиль восприятия и мышления нашего сознания
относительно окружающего мира. Мы биологически приговорены к «мышле-
нию вентральным потоком», хотя наша математика и может иметь дело с более
динамическим миром, чем тот, с которым большинство из нас соприкасается
ежедневно.
Ответы на вышеизложенные вопросы появляются в когнитивной нейронау-
ке. Поэтому становится важным задавать такие вопросы с максимальной точно-
стью и не считать регистрацию сознаваемых событий доказанной, само собой
разумеющейся.
6.5 . Факты по необычным состояниям
Три основных состояния суточного цикла могут представлять собой хорошее
основание для анализа необычных явлений — вызванных приемом препаратов,
психическими расстройствами, и состояний, которых можно достичь после
многолетних тренировок, медитации и йоги. В истории и литературе есть много
описаний необычных состояний рассудка. Но ученые настороженно относятся
к необычным заявлениям. Известно, что такие методы, как неврологическая об-
ратная связь и гипноз, обладают выраженными эффектами, но наше понимание
о связанных с этими методами процессах в головном мозге и результатах еще
недостаточное. Легко ошибиться и занять позицию чрезмерного скептицизма
в данном вопросе или же, напротив, относиться очень легковерно; настоящие
ученые должны быть восприимчивы к новым идеям, но при этом полагаться на
строгие доказательства.
Ряд мистических наблюдений связывают традиционно с эпилепсией, в том
числе известный случай с Ф. Достоевским, который страдал от этого недуга
и описывал сильные мистические «ауры» перед припадками. Первый труд по
эпилепсии приписывают греческому врачу Гиппократу (�–I� в. до н. э.), на-
звание которого — «Об эпилепсии». Обычные больные-эпилептики описывают
измененные состояния сознания, что ассоциировано с гиперсинхронизацией
ЭЭГ, даже без явных припадков (см. рис. 8 .5). Большая часть таких измененных
состояний воспринимаются больными как неприятные. Поскольку гиперсин-
хронизация меняет работу таламокортикальной системы, постольку может су-
ществовать связь между измененными субъективными состояниями и ритмами
головного мозга.
Эффекты психоделических препаратов сравниваются со снами наяву, часто
это яркие зрительные галлюцинации, ложные убеждения, действия как во сне,
мощные эмоциональные столкновения, сложности с осознанием течения вре-
мени, и резкие разрывы от «эпизода» к «эпизоду». Аналогичные наблюдения
известны по разным психотропным соединениям, в том числе ЛСД, марихуане,
и традиционным растительным препаратам. Структурно похожи между собой
Сознание и внимание
461
молекула ЛСД и серотонин, основной нейромодулятор, участвующий, как из-
вестно, в сновидениях быстрой фазы сна. Однако связь между психоделиками
и нормальными снами пока остается гипотетической.
Мании и галлюцинации — характерные симптомы психоза, который может
быть очень опасен. Часто первое проявление шизофренического расстройства
происходит в конце подросткового периода или в молодости, что нередко со-
впадает с крупными изменениями в жизни и стрессом. Как полагают, исполь-
зование марихуаны предрасположенными людьми может повысить у них риск
развития шизофрении. И все же, несмотря на десятилетия исследований в этой
области, шизофрения продолжает оставаться нераскрытой тайной.
Главное различие между психозом и приемом психоделических препаратов
в развлекательных целях заключается в том, являются ли желаемыми возника-
ющие очень необычные ощущения. Препараты для развлечения уже по опре-
делению служат для получения необычных ощущений. При психозах бред и
галлюцинации часто очень расстраивающие, пугающие, и нежелаемые. Они
возникают неожиданно, влияют на обычное течение жизни, могут появляться
болезненные преследующие голоса, горестные галлюцинации пациента о себе;
также они могут быть хроническими. Степень нежелания человеком таких
ощущений — главное различие между психоделиками и расстройством пси-
хики. Можно привести такую аналогию: некоторым людям нравится прыгать
с парашютом с самолета и высоких висячих мостов, они занимаются этим для
удовольствия, выбрав подходящее время и место. Но если бы этим людям при-
шлось проделать то же самое против их воли, так, чтобы действия вышли из-
под их контроля, то те же самые ощущения стали бы шокирующими и травма-
тическими. Психические расстройства — это не вопрос выбора, и страдающие
от них люди не могут по своей воле их остановить.
С другой стороны, на протяжении тысячелетий люди стремились достичь
некоторых экстраординарных состояний. С этой целью были перепробованы
множество методов, в их числе использование растений с психотропными эф-
фектами, длительное голодание, алкогольное опьянение, многолетние физиче-
ские и умственные тренировки, методы визуализации, преднамеренная дезори-
ентация, условия высокогорья, гипоксии, членовредительство (самокалечение),
бурная сексуальная жизнь, ритуалы, прозрачность снов, ритмические танцы,
кружение, монотонное песнопение и музыка, гипноз, разобщающие травмы,
лишение сна и изоляция от общества. Многое из вышеперечисленного остается
настолько же неясным сегодня, как и столетия назад. В буддизме и ведаизме
верят, что люди могут и должны менять свое состояние сознания и ощущение
себя. Отражение такой веры можно встретить в разные эпохи и в разных ме-
стах.
Отчеты людей, испытавших измененное состояние сознания, неубедитель-
ны для скептиков. Простые опросы людей о том, сделало ли их то или иное
занятие более счастливыми или более расслабленными, не могут быть убеди-
тельными доказательствами. К таким же отзывам могут привести и сахарные
пилюли. Почти у каждого занятия есть сторонники, не скупящиеся на похва-
лы. При этом возможны и правдивые отчеты от искренних и проницательных
участников. Но не подлежащие проверке описания частных опытов изменен-
ных состояний сознания не приводят к прогрессу в науке.
462
Глава 8
Для скептиков более убедительны данные по головному мозгу при необыч-
ных состояниях, ведь люди не могут контролировать состояния головного моз-
га. Мы не способны менять свои ЭЭГ и МРТ, чтобы получить доказательства в
пользу приглянувшейся гипотезы. Некоторые психологические задания также
не подвластны личным предубеждениям. Но подавляющее большинство иссле-
дований по медитации и родственным методикам серьезно подвержены вли-
янию предвзято настроенных участников. Такие результаты редко вызывают
доверие ученых-скептиков.
6.5.1 . Гипноз, конверсионные расстройства (при истерии)
и вовлечение мозговых волн
Примерно одна четверть людей сильно подвержена гипнозу, что оценивается по
стандартной процедуре начала гипноза. Гипнотические внушения могут менять
события в головном мозге, например снижается обусловленный событием по-
тенциал в ответ на вспышку света просто при «галлюцинации» ящика, закрыва-
ющего этот свет (S�ie�el, 2003). Мы не знаем, по какой причине гипнотические
внушения так хорошо воздействуют на большинство нормальных людей. Около
5% популяции — это «гипнотически-одаренные» люди, они могут вызвать гал-
люцинацию чувственную, например, жужжание насекомого. Такие люди могут
обладать раздвоением личности (Hil�ard, 1974). С помощью функциональной
МРТ показано, что боль, внушенная гипнозом, активирует в мозге те же зоны,
что и реальная боль; есть и хороший эффект — гипнозом можно уменьшить
боль пациенту, страдающему хроническими болями, но не имеющему других
путей избавления от них (S�ie�el, 2003).
Центром изучения становятся корреляты в головном мозге с гипнозом. Уже
два столетия известно, что люди под воздействием гипноза могут испытывать
легкое чувство потери контроля над произвольными движениями. Распростра-
ненный способ гипноза — говорить испытуемым, чтоб они позволили своим
рукам подниматься свободно вверх. У хорошо поддающихся гипнозу людей это
происходит без чувства дискомфорта. Следовательно, возможно, что гипноз вы-
зывает функциональное разобщение между частью головного мозга, участвую-
щей в произвольном контроле (дорсолатеральная прелобная кора) и способно-
стью контролировать когнитивное противоречие (передняя поясная кора).
Эгнер и его сотрудники (E�ner et al., 2005) по результатам применения функ-
циональной МРТ и ЭЭГ в тесте Струпа при гипнозе заключили: «Гипноз разоб-
щает когнитивный контроль и процессы контроля противоречий лобной доли».
Аналогично Феймонвиль и сотрудники (Faymonville et al., 2006) выявили ши-
рокую нейронную сеть активаций, связанных с гипнозом, но зависимую деак-
тивацию предклинья — зоны головного мозга, ответственной, как считается, за
проекцию себя. Этот результат соответствует разобщению самоконтроля и са-
мовосприятия при гипнозе, особенно у сильно подверженных гипнозу людей.
Сейчас также наблюдается возврат научного интереса к конверсионным рас-
стройствам, что называют истерией в истории болезни. При конверсионных
расстройствах люди испытывают психогенные аналгезию, гипералгезию, во-
ображаемые симптомы медицинских нарушений, паралич кистей и рук, и т. п.;
все это при отсутствии выявляемого нервного повреждения. Симптомы конвер-
сии изменяются и чувствительны к внушению, поэтому врачи в XIX в. пред-
положили, что проблема обусловлена самовнушением пациентов. В настоящее
Сознание и внимание
463
время данная гипотеза остается, однако из этого не следует, что симптомы кон-
версии поддаются лечению внушением.
Легкие симптомы, сходные с конверсией, встречаются крайне часто. Сту-
денты-медики, впервые знакомясь с детальным описанием тяжелых заболева-
ний, часто начинают беспокоиться, поскольку им кажется, что они заметили у
себя «истинные» симптомы болезней. Такой «синдром студента-медика» очень
распространен, и, как правило, со временем он проходит. Возможно, что кон-
версионные расстройства — наиболее длительный и калечащий вариант общей
для всех людей склонности к самовнушению. Эффект плацебо оказывается по-
ложительным проявлением этого же явления.
Диагноз «истерия» подвергался острой критике, отчасти потому, что его ста-
вили только женщинам. Но в XIX веке врачи Жан-Поль Шарко и Пьер Жане про-
демонстрировали, что у мужчин бывают симптомы конверсии, в частности по-
сле тяжелой травмы (например, авария на железной дороге). Тем не менее само
представление о конверсионных расстройствах перестало быть популярным на
десятилетия. В последние годы интерес возник вновь благодаря исследовани-
ям с картинами головного мозга, получаемыми под пристальным контролем.
Действительно существует группа пациентов, страдающая от вышеописанных
проблем, иногда очень болезненных и приводящих к потере дееспособности
(Co�an et al., 2009). Постановка точного диагноза — это шаг вперед, навстречу
дифференциации между конверсионными расстройствами и истинными по-
вреждениями структуры мозга.
6.5.2 . Неврологическая обратная связь
Неврологическая обратная связь определяется как освоение контроля над со-
бытиями в головном мозге путем создания чувственной (сознаваемой) обрат-
ной связи, зависимой от некоего события в мозге. В литературе по исследовани-
ям животных это часто называют «выработкой инструментального условного
рефлекса», что является скорее экспериментальной процедурой, нежели объ-
яснительной конструкцией. Неврологическая обратная связь — разновидность
тренировки биологической обратной связи, что также включает тренировку на
периферические события, например повышение тепла пальца. Просто держа
термометр, отображающий незначительные изменения температуры на пальце,
можно научиться повышать в пальце тепло, что, к примеру, поможет рассла-
биться и снизить артериальное давление.
Исследования биологической обратной связи на животных, с выработкой
инструментального условного рефлекса, на протяжении последних десятиле-
тий демонстрируют положительные результаты. Животные с имплантирован-
ными электродами могут выучиться повышать активность отдельных нейронов
и больших групп нейронов. При наличии медицинских показаний аналогичные
эффекты могут быть достигнуты и у людей. Важно вдуматься в то, насколько
выдающимся является такой итог. В мозге, содержащем миллиарды нейронов,
простая сенсорная обратная связь может помочь выработке произвольного кон-
троля над активацией нейронов, полностью непроизвольной другими способа-
ми, и над другими событиями в головном мозге.
Однако нацеливание неврологической обратной связи на определенные ос-
цилляции в мозге, к примеру, не всегда приводит к увеличению намеченного
464
Глава 8
события. Как ни удивительно, но даже тренировка альфа-неврологической об-
ратной связи не всегда приводит к доказанному повышению активности альфа-
волн. В литературе отсутствуют сведения, акцент сделан в основном на провер-
ке практических результатов. Странная ситуация — аспект «обратной связи» в
неврологической обратной связи иногда утерян.
Парадокс будет не таким разительным, если мы учтем, что ритмы голов-
ного мозга находятся под четким гомеостатическим контролем (John, 2002).
Если неврологическая обратная связь работает, то это не просто сродни пере-
ключению выключателя света в головном мозге; скорее это похоже на подпры-
гивание на большой резиновой лодке, плывущей по озеру: прыжки вызывают
реальные перемены, но мозг по-прежнему плывет, что очень хорошо. Голов-
ной мозг сложен, динамичен и обладает многими устойчивыми состояниями,
благодаря этому организмы выживают при различных изменениях окружаю-
щей среды.
Возникает два вопроса.
• Есть ли результаты поиска обратной связи головного мозга? После
долгих лет исследований ответ, как кажется, должен быть однозначен:
«ДА!». На рис. 8 .51 приведен пример эффектов обратной связи по фМРТ
у пациентов, испытывающих хронические боли.
• Имеет ли обратная связь простой, прямой эффект на мозг, или ее эф-
фекты носят и прямой и косвенный характер? На основании фактов
последнее кажется более вероятным, включая важные положительные
результаты в таких состояниях как синдром дефицита внимания-ги -
перактивность, эпилепсия, депрессия и возможно в других (Gruzelier,
2008) (рис. 8 .52).
Большое число работ по ЭЭГ с неврологической обратной связью показало
значительные результаты, но долговременные исследования часто отсутствуют.
Так как неврологическая обратная связь может работать в тех случаях, когда
другие способы лечения не помогают (как при депрессии и эпилепсии), то дли-
тельные испытания являются жизненно необходимыми.
Рис. 8 .51. Локальная активность головного мозга, контролируемая обратной связью по
фМРТ. Локальная активность головного мозга увеличивается после тренировки обрат-
ной связи по активности фМРТ у пациентов, испытывающих хроническую боль. Цель
тренировок — получить активность в ростральной передней поясной коре, вовлечен-
ной, как известно, в восприятие боли. (Источник: deCharms et al., 2005.)
Сознание и внимание
465
6.5.3 . Сенсомоторный ритм (СМР), обратная связь альфа/тета
и другие наработанные осцилляции
Стерман и Эгнер (Sterman, E�ner, 2006) описали доказательства того, что «невро-
Sterman, E�ner, 2006) описали доказательства того, что «невро-
, E�ner, 2006) описали доказательства того, что «невро-
E�ner, 2006) описали доказательства того, что «невро-
, 2006) описали доказательства того, что «невро-
логическая обратная связь как способ лечения эпилепсии является обоснован-
ной и жизнеспособной альтернативой противосудорожной фармакотерапии».
Это важно как с медицинской, так и с научной точек зрения. Медикаментозное
лечение эпилепсии не всегда эффективно, и наличие другого способа лечения
поможет многим пациентам. С научной точки зрения, эпилепсия (на основании
различных причин) проявляется как медленная синхронизация ЭЭГ, поэтому
способность изменять активность припадков в мозге выявит нам важные факты
о роли ритмов мозга и их произвольном контроле.
Что такое СМР, или сенсомоторный ритм? СМР обнаружен в исследованиях
ЭЭГ на кошках, которых обучали подавлять ранее выученный и усиленный от-
вет. У этих животных выявлен временный, альфа-подобный ЭЭГ-ритм в сен-
сомоторной коре с частотой 12–20 Гц, напоминающий веретена сна на ЭЭГ,
с пиком спектра около 12–14 Гц (Roth et al., 1967). Было решено исследовать
этот ритм непосредственно, путем выработки инструментального условного
Рис. 8.52. Тренировка альфа/тета улучшает качество исполнения музыки и танцев на
высоком уровне. Грузильер (Gruzelier, 2008) показал, что после 20 тренировок невро-
логической обратной связи, направленной на увеличение альфа- и тета-активности,
значительно меняется уровень исполнения у музыкантов консерватории и танцоров.
Такая оценка проводилась независимыми экспертами, не осведомленными об услови-
ях исследования. Выработка неврологической обратной связи дала улучшения, рав-
ные результатам обучения в двух классах консерватории, при других факторах опыта
улучшения не было. Техническое мастерство не совершенствовалось, но повысились
музыкальность, убедительность исполнения, подача себя на сцене и уровень психиче-
ского напряжения. СМР — сенсомоторный ритм, ТМС —транскраниальная магнитная
стимуляция
466
Глава 8
рефлекса для выяснения того, можно ли обучить кошек произвольному вызо-
ву СМР, создавая условия пищевого вознаграждения при ее получении. Кошки
легко обучились такому саморегулированию ЭЭГ, с СМР была ассоциирована
неподвижность тела, всплескам СМР регулярно предшествовало снижение мы-
шечного тонуса. Когда же лаборатории Стермана было поручено изучение эф-
фектов препарата, вызывающего эпилепсию, они открыли, что обученные СМР
кошки менее подвержены химически индуцируемым припадкам. Описанный
эффект распространяется и на людей. Внутричерепные исследования на живот-
ных показали, что СМР возникает в вентробазальных ядрах таламуса и что он
играет ключевую роль в подавлении нежелаемых ответов.
На данный момент показано, что обратная связь с СМР является эффектив-
ным лечением у людей в случаях эпилепсии, синдрома дефицита внимания-ги -
перактивности и нарушениях импульсного контроля (Sterman, 2006). Следова-
Sterman, 2006). Следова-
, 2006). Следова-
тельно, такое лечение возможно. СМР — один из редких примеров (пока) того,
как неврологическая обратная связь привела к всестороннему психобиологи-
ческому пониманию важного медицинского состояния. С улучшением методов
регистрации состояния головного мозга можно ожидать еще больше таких обна-
деживающих результатов. (Сходные результаты выявлены при иной эндогенной
осцилляции мозга, так называемом «Медленном потенциале коры»).
Согласно Стерману (2006), Американская академия детской и подростко-
вой психиатрии приняла во внимание, что неврологическая обратная связь в
случае припадков соответствует критериям «Клинического руководства» для
лечений, основанных на доказательствах. Это сформулировано так: «...указан-
ные результаты получены в очень сложной подгруппе пациентов-эпилептиков,
у которых припадки плохо подлежали контролю, и было доказано отсутствие
ответа на фармакологическое лечение. В среднем для 80% больных, обученных
повышению амплитуд СМР, показаны значительные клинические улучшения»
(Ka�lan, 1975; Seifert an Lu�ar, 1975; (Sterman, McDonald and Stone, 1974).
Следует подчеркнуть, что неврологическая обратная связь по сути все еще
загадочна, так как мы не понимаем роли обратной связи сознания в реализации
произвольного контроля над таламокортикальными формами волн. Это отра-
жает и имеющиеся на настоящее время ограничения. СМР представляет собой
выдающийся пример твердой научной основы, ускорившей разработку методов
лечения серьезных нарушений. Очевидно, что необходимо исследовать СМР
при дефиците внимания и нарушениях импульсного контроля, которые зача-
стую плохо поддаются лечению.
6.5.4 . Ритмическое вовлечение волн головного мозга
Если ритмы мозга — это основной шифр межнейронных сигналов, то крайне
интересно установить, можем ли мы управлять этими ритмами извне. Захват
волн мозга описан при транскраниальной магнитной стимуляции и при транс-
краниальном переменном токе. В нейрохирургии уже полстолетия применяется
прямая электрическая стимуляция коры (см. гл . 1). Не исключено, что можно
использовать также слуховые, зрительные и двигательные ритмы. Но, несмотря
на то что есть много громких заявлений об эффектах слуховых захватов, пока
не продемонстрирована их работа по соответствующим медицинским и науч-
ным стандартам.
Сознание и внимание
467
В идеальном случае внешний сенсорный водитель волн мозга должен быть
нацелен на заданную часть мозга, определенный ритм и должен активироваться
и выключаться в указанное время. Пока остается много невыясненных вопро-
сов об эндогенных ритмах в мозге, об их взаимодействии с внешними входны-
ми сигналами. Как уже говорилось, зрительная детекция сигнала бывает лучше
на вершине альфа-волны, нежели в ее спаде (�anrullen and Koch, 2003). Это
может быть первым примером того, как текущая активность мозга влияет на
обработку входного сигнала. Мозг постоянно активен, и сенсорные входы кор-
ректируют лишь текущие процессы.
Неврологическая обратная связь и такие методы, как акупунктура, исследо-
вались и на животных, что позволило исключить эффект плацебо. Но пока нет
широкого применения методов, казавшихся многообещающими. Сейчас стало
более полным наше знание об осцилляциях мозга, что дает нам большее обо-
снование для обучения неврологической обратной связи. Но мы только начина-
ем понимать детали. Следует помнить, что выработка неврологической обрат-
ной связи всегда включает сознательный сигнал обратной связи (Baars, 2002).
Гипофронтальность — это сниженный метаболизм лобных долей, может
обобщать собой разные измененные состояния мозга. Дитрих (Dietrich, 2002)
показал, что сниженный лобный метаболизм — признак гипнабельности (под-
верженности индивидуума гипнотическому воздействию) и других необычных
состояний рассудка. Сниженный метаболизм лобных долей также отмечается
просто при дремоте и других полусознательных состояниях, при выходе из об-
щей анестезии. Неустойчивость, как при опьянении, — так называют это со-
стояние, в котором мы бодрствуем, но не очень внимательны, исполнительные
функции при этом могут казаться нарушенными. Очевидно, что не все такие
состояния сходны между собой. Низкий метаболизм лобных долей может быть
их единственной общей чертой.
6.5.5 . Медитация и подобные занятия
Медитацию часто описывают как состояние расслабленной бдительности. Под
этим понятием объединены множество умственных техник с весьма разными
эффектами, общий подход — это повторение про себя слова, называемого ман-
тра. Пока не окончен спор о том, может ли быть мантрой любой случайный звук
или же нет. Медитации с мантрами описывают как способ изменить состояние
разума, чему учат такие азиатские течения, как веданта, буддизм, а также тради-
ционная китайская медицина. В Европе и на Ближнем Востоке такие практики
также широко распространены. Поскольку медитации просты (в том смысле,
что не требуют специального оборудования), можно с уверенностью утверж-
дать, что их практиковали в разные времена, в том числе и охотники-собирате-
ли, которые часто исполняли замысловатые ритуалы шаманов.
Другая процедура, называемая «медитация целостности разума», описы-
вается так: «..позволить любым мыслям, чувствам и ощущениям возникать в
процессе сохранения особого состояния внимания: осознание окружения как
бы внимательным и незаинтересованным наблюдателем без анализа и оцени-
вания» (Cahn and Polich, 2006, 2009). Именно эти две техники лучше других
сейчас исследованы, по ним опубликованы сотни статей в научных рецензируе-
мых журналах. Они различаются по степени поощрения свободного, ненаправ-
468
Глава 8
ленного внимания или же нацеленной концентрации. За последние десятилетия
часто указывалось, что при методах медитации увеличивается активность го-
ловного мозга и синхронизация фаз, особенно в тета, альфа и бета до нижнего
гамма-диапазонов ЭЭГ. Также сообщалось о когерентности между полушария-
ми в лобной доле, а также об улучшении функции внимания (Lazar et al., 2000;
Lutz et al., 2004; Tan� et al., 2007).
Удивительным открытием оказалось выраженное снижение метаболизма
при простом повторении мантры, даже у новичков, что отражается в «задержке
дыхания» — спонтанной его остановке, но без последующего компенсаторного
дыхания (рис. 8.53). Это отличается от произвольной задержки дыхания. Так,
например, при плавании мы способны сделать глубокий вдох, затем нырнуть,
а после выныривания возникает потребность в нескольких дополнительных
вдохах. В норме дыхание находится под жестким гомеостатическим контро-
лем, и произвольная задержка дыхания сопровождается последующим компен-
саторным дыханием, чтобы восполнить недостаток кислорода. Поэтому выяв-
ление спонтанной задержки дыхания без его компенсаторного восполнения и
представляет собой такой удивительный факт. Такого явления не наблюдают у
контрольной группы в обычном покое. Отсутствие компенсаторного дыхания
после задержки указывает на то, что энергопотребности действительно сни-
зились больше, чем при обычном состоянии покоя, что отражают изменения
обмена O2 и СO2. (Benson et al., 1975; Hoffmann et al, 1982.)
Герберт Бенсон (Benson) и его сотрудники предположили, что такие резуль-
Benson) и его сотрудники предположили, что такие резуль-
) и его сотрудники предположили, что такие резуль-
таты отражают «релаксационную реакцию», по аналогии с другими физио-
Рис. 8 .53. Спонтанная задержка дыхания при повторении мантры. Повторяющаяся,
спонтанная задержка дыхания при проговаривании про себя мантр совпадает с описы-
ваемыми испытуемыми состояниями «чистого сознания». Задержки дыхания не сопро-
вождаются компенсаторной интенсификацией дыхания, что отличается от произвольно-
го задерживания дыхания, за которым следует его компенсаторное усиление. Задержки
дыхания, как полагают, указывают на моменты «чистого сознания», определяемые как
моменты ясного сознания без определенного его наполнения. Такие состояния и яв-
ляются целью данной практики. Описанное исследование проводилось на практикую-
щих трансцендентную медитацию — технику безмолвных мантр из традиции веданты
в Индии. Однако сходные явления могут происходить и при произвольных мантрах.
(Источник: Benson et al., 1975; Hoffmann et al, 1982.)
Сознание и внимание
469
логическими рефлексами. В расслаблении задействована парасимпатическая
нервная система (конкурируя с симпатической); и кажется вероятным, что су-
ществует путь индукции парасимпатического «включения», для восстановле-
ния физических и умственных сил. Однако много других параметров совпада-
ют с эффектами медитаций с мантрами. Остается загадочным, почему простое
проговаривание в уме слов вызывает такой всеобъемлющий физиологический
ответ и ответ со стороны головного мозга. Наиболее важным, возможно, от-
носительно быстрых физиологических реакций на мантры можно считать сам
факт наличия чего-то удивительного и иного при таком простом действии. Яв-
ное метаболическое снижение позволяет по-новому взглянуть на волны голов-
ного мозга, сопровождающие эти процессы.
Сообщалось, что спонтанные задержки дыхания связаны с периодами «чи-
стого сознания», определяемого как расслабленная бдительность без опреде-
ленного наполнения сознания. Цель мантра-медитации состоит в достижении
«чистого сознания», чтобы такие состояния стали привычными, частью по-
вседневности. Такие эффекты описаны у новичков, у давно практикующих, и
при использовании случайных мантр, как при релаксационном ответе. Согласу-
ющиеся факты получены при измерении метаболизма, тонуса симпатической
нервной системы и, в последнее время, по крупным изменениям в генной экс-
прессии. Но точные функции генов, «связанных с релаксацией», еще не выяс-
нены, и поэтому необходимы дополнительные работы в этой многообещающей
области (Jaco�s et al., 1996; Dusek et al., 2006).
Какие изменения наступают в головном мозге после многолетнего практи-
кования вышеописанных упражнений? На ЭЭГ опытных медитирующих вид-
ны высокие уровни гамма-, альфа- и тета-синхронизации (Lutz et al., 2005).
Исследования такого рода обычно сфокусированы на хорошо подготовленных
людях. К примеру, Лютц (Lutz) и его сотрудники изучали четырех опытных
медитирующих, проведших в упражнениях от 15 000 до 45 000 ч. Но опыт-
ные медитирующие будут значительно различаться во многом. Они напомина-
ют профессиональных спортсменов, танцоров и музыкантов, которые начали
свой путь с детства, имея выдающиеся талант и наклонности. Попытка узнать,
как получен такой результат, равна попытке спросить, почему Тайгер Вудс стал
выдающимся спортсменом: надо было начинать с детства. Необходимо взять
других людей как контрольную группу и провести многолетние исследования,
чтобы в итоге закончить историей одного конкретного случая.
Тем не менее интерес сохраняется, ведь можно наблюдать изменения в го-
ловном мозге, поддающиеся измерению через полчаса медитации, пусть даже
среди исключительных испытуемых. Брока и Вернике изучали лишь очень
небольшое число людей с редкими нарушениями, что позволило им в XIX в.
открыть в коре зоны речи (гл. 1). Интенсивное исследование нескольких па-
циентов привело к значительному прогрессу в понимании роли гиппокампа в
памяти (гл. 1). Есть хорошие прецеденты по изучению небольшого числа выда-
ющихся индивидуумов, когда на получаемые факты не влияют предубеждения
и ожидания.
Однако практически невозможно найти контрольные группы, сравнимые с
медитирующими на высоком уровне. В итоге сложно понять, насколько име-
ющиеся результаты относятся к более широкой популяции. Ответ на такой
470
Глава 8
вопрос заключается, если обратиться к примерам истории, в том, что со вре-
менем применимость выявленных фактов можно проверить другими способа-
ми. Так, поле Брока задействовано в формировании речи, что сейчас мы можем
проверить многими другими подходами и с участием десятков тысяч испытуе-
мых. Но начиналось все с тщательного исследования одного пациента. Случай
с пациентом HM демонстрирует даже лучший методологический подход, ведь
один больной с хирургическим повреждением изучался на протяжении деся-
тилетий во многих лабораториях, т. е. была уверенность, что разные исследо-
ватели работают на одном и том же человеке. Но при работах по измененным
состояниям сознания такого уровня координации еще не удалось достичь. По-
этому в результате мы располагаем многочисленными статьями по опытным
медитирующим, которые могут значительно различаться между собой.
Как объяснение состояний медитации предлагается высокая степень сопря-
жения между зонами в тета-альфа диапазоне (He�ert, 2005). Опытные люди,
практикующие медитацию и сходные техники, описывают состояние полного
спокойствия, сознания без конкретного его наполнения, расширение субъек-
тивного пространства, сознание во сне, и т. п . Исследователи только начинают
уделять серьезное внимание именно субъективным восприятиям опытных ме-
дитирующих, а ведь это и составляет то, ради чего люди занимаются медита-
цией.
В итоге даже спустя четыре десятилетия исследований мы не знаем всего,
что нам хотелось бы, о воздействии медитации. О неврологической обратной
связи в литературе представлено больше сведений, хотя и в этой теме еще очень
много пробелов. Вся имеющаяся научная литература в данной области не впол-
не удовлетворительна; исследований проведено пока недостаточно. Обнадежи-
вает тот факт, что теперь стали более понятны ритмы на ЭЭГ в определенных
зонах мозга, что регистрируется по фМРТ. Раньше ритмы мозга воспринима-
лись просто как корреляты с чем-то интересным, но не было представления
о точной роли, которую они могли бы играть. Знание только о когерентности
альфа-волн можно сравнить со знанием о том, что у людей бьется сердце, без
понимания роли кровообращения и насосной функции сердца. К счастью, на
сегодняшний момент одинокое дерево неинтерпретированных ритмов мозга те-
перь уже стоит в молодом лесу, состоящем из фактов и понимания предполагае-
мых функций. Более глубокое понимание даст нам возможность лучше связать
выявляемые изменения в мозге с медитацией и сходными с ней практиками.
6.5.6 . Опыты выхода из тела можно индуцировать
стимуляцией головного мозга
Наиболее надежные результаты удается получить в исследованиях, посвя-
щенных опытам выхода сознания из тела (ОВТ). Возможность посмотреть на
собственное тело снаружи, безусловно, следует рассматривать как экстраорди-
нарную. В этой сфере достигнут значительный успех. Теперь, к примеру, нам
известно, что прямая стимуляция правой задней париетальной коры может вы-
звать значительные изменения в восприятии тела (рис. 8 .54). Некоторые боль-
ные эпилепсией испытывают большие изменения в восприятии тела, возмож-
но, обусловленные гиперсинхронной активностью, затрагивающей указанную
зону мозга. Известно, что повреждение правой задней теменной коры также
ведет к переменам в восприятии тела. Хорошо описанным нарушением являет-
Сознание и внимание
471
ся теменное игнорирование, ведущее к радикальным изменениям в восприни-
маемом пространстве.
Итак, теменная зона коры задействована в ряде необычных состояний. Те-
перь мы видим растущее количество фактов, касающихся как минимум этого
вида экстраординарных опытов восприятия.
7.0 . Заключение
В этой главе был охвачен большой материал. Хорошая новость в том, что мы
получили лучшее понимание сознания и внимания и умение объединять про-
стые данные, так как кусочки мозаики начали собираться вместе.
Внимание и сознание можно считать взаимодействующими процессами (см.
рис. 8 .55). Сознательное содержание, часто полагают, включает широкое рас-
пределение центрального содержимого. Как только мы видим кролика (осоз-
нанно), мы можем судить, является ли он реальным, является ли он чьим-то до-
машним любимцем, мы можем смеяться или визжать от страха, что он укусит, а
также постараемся вспомнить, не переносят ли кролики бешенство. Суть в том,
что большое многообразие событий в мозге может быть вызвано осознаваемым
объектом, включая большое разнообразие бессознательных процессов. Наобо-
рот, мы можем сконцентрировать ресурсы познания на любом предмете, кото-
рому мы решили посвятить внимание, например на содержании этой книги.
Для того чтобы выучить, мы направляем внимание на предмет опять и опять,
в результате у нас возникает много различных моментов осознания, при этом
в случае необходимости каждый из них способен пробудить дополнительные
познавательные процессы. Изучение трудных тем включает распределение ре-
сурсов внимания во времени. Сознание в каждый данный момент обычно вклю-
чает небольшое количество содержимого, например вид розового куста, кото-
рый может привлечь и мобилизовать много бессознательных функций мозга.
Рис. 8.54. Правая теменная кора и опыты пребывания вне своего тела. (Источник: Ton�,
2003; после Blanke, 2002.)
472
Глава 8
Основные параметры внимания, такие как мотивация, планирование, эмоцио-
нальная необходимость и т. п., становятся направленными на отдельный пред-
мет. Произвольное внимание особенно требует привлечения ресурсов лобной
доли к познанию новой и лучшей информации в интересах некоторой ясной
цели или задачи. Лобно-теменные области коры могут затем увеличить обра-
ботку информации в зрительных областях (например), чтобы обострить, про-
яснить и сделать сознательной некоторую часть зрительного мира. Случайное
и декларативное обучение, кажется, происходит автоматически как функция
содержимого сознания, что, в свою очередь, может управляться произвольным
вниманием. Три основных таламокортикальных мозговых состояния суточного
цикла показывают различные стили действий в диалоге внимание–сознание.
Основная функция медленноволнового сна и сновидений с быстрыми движе-
ниями глаз — закрепление памяти о событиях, которые были первоначально
получены и изучены в бодрствующем состоянии.
Понимание основ сознания и внимания в головном мозге все еще является
огромной трудной задачей. Некоторые подсказки даны в табл. 8 .4, описываю-
щей деятельность мозга в трех основных состояниях. Обратите внимание на
роль нейромодуляции, химических веществ, распространяющихся из неболь-
шого количества нейронов, находящихся в базальных ядрах, которые простира-
ют свои аксоны до вышележащих частей головного мозга. Таламус также игра-
ет важную роль в трех основных состояниях. Однако содержимое сознания,
вероятно, в основном находится под контролем коры.
Рис. 8 .55 . Внимание и сознание. Внимание часто рассматривается по аналогии к пятну
света, двигающемуся к «точке где-то» и выбирающему некоторую часть внешнего мира
для обработки сознанием. Контроль над «точкой внимания» требует работы лобной и
теменной коры, а также подкорковых областей. Недавно было предположено, что созна-
ние, связанное с восприятием, может возбуждать области коры в другом направлении,
начиная с первичной сенсорной коры (сзади) и возбуждая более передние области. Та-
ким образом, мы можем думать о двух потоках взаимодействующей информации, кото-
рые двигаются назад и вперед в коре головного мозга: один выборочный (выборочное
внимание) и другой интегративный и распределительный по своим эффектам (созна-
ние, связанное с восприятием). (Источник: Baars, Fu, 2007.)
Сознание и внимание
473
В табл. 8 .4 представлены основные особенности трех ежедневных состоя-
ний сознания. Заметьте, что активность мозга является наиболее важным, но
не единственным признаком сна или бодрствования. Заметно, что сенсорные
входящие и двигательные исходящие сигналы заблокированы во время сна с
БДГ (чтобы мы бессознательно не выражали наши сны в действиях) и во время
медленноволнового сна. При некоторых нарушениях БДГ-сна торможение дви-
жения в спинном мозге ослаблено, в результате люди могут демонстрировать
галлюцинаторное или бредовое поведение в полубессознательном состоянии.
Некоторые авторы полагают, что таламокортикальный срыв нормального рит-
мического цикла может быть вовлечен в серьезные нарушения мозга, такие как
психоз.
Медленные волны сна можно разлагать на «состояние подъема» (пик мед-
ленной волны) и «состояние спада» (нижняя точка). Состояние подъема позво-
ляет нейронам возбуждаться до уровня их бодрствующего состояния в норме,
а также взаимодействовать друг с другом на секунду или меньше. Состояние
спада блокирует эту активность. Таким образом, ученые предположили, что на
самом деле мы можем не спать на протяжении одной секунды во время МВС.
Когда мы глядим на порог пробуждения на этой секундной шкале, порог про-
буждения может фактически быстро колебаться назад и вперед.
Как волны в океане, распространение сигнала в мозге представлено мно-
жеством многих разных волн. Первыми наблюдаемыми волнами были альфа-
волны, потому что их легко определить: они появляются в необработанной ЭЭГ
над затылочной корой, когда глаза закрыты и в спокойном состоянии. Когда гла-
за открыты, альфа-волны обычно прерываются более быстрой, нерегулярной
низкоамплитудной ЭЭГ, которую отмечают как бета и гамма. Однако сложные
формы волн могут быть разложены на отдельные полосы. Сейчас это расшире-
но в диапазон до 200, иногда до 1000 Гц. Традиционный уровень гамма-волн
все еще возникает от 30 до 120 Гц, но надежнее использовать именно эти зна-
чения. Наши словесные обозначения ритмов мозга произвольны и, вероятно,
изменятся.
Формы волн в мозге взаимодействуют. Ритмы бодрствования могут сумми-
роваться, вычитаться, синхронизироваться, десинхронизироваться, производить
фазовую синхронизацию и иметь сложную структуру. Медленные волны могут
нести более быстрые волны (усложняться). Предположили, что очень медлен-
ные колебания (0,1–,5 Гц), дельта- (0,5–3 Гц), тета- (4–7 Гц), альфа- (8–12), бета-
(13–29) и гамма-волны (30–120) — при некоторых условиях могут усложняться.
Тета-волны обычно несут гамма-активность, состояние подъема дельта-волн во
время медленноволнового сна включает сложную, взаимодействующую высоко-
частотную активность, похожую на бодрствующее состояние.
8.0 . Задания и упражнения к главе 8
1. Какие основные структуры мозга вовлечены в поддержание сознательного
бодрствующего состояние, глубокий сон (медленноволновый сон) и сон с
быстрыми движениями глаз? Как мы переключаемся между состояниями?
2. Какая функция медленноволнового сна наиболее хорошо доказана? Функ-
ция бодрствования? Сна с быстрыми движениями глаз? (Если не установле-
на никакая функция, скажите это.)
474
Глава 8
Т
а
б
л
и
ц
а
8
.
4
А
к
т
и
в
н
о
с
т
ь
м
о
з
г
а
в
о
в
р
е
м
я
б
о
д
р
с
т
в
о
в
а
н
и
я
,
с
н
а
и
с
н
о
в
и
д
е
н
и
й
Т
а
л
а
м
о
к
о
р
т
и
к
а
л
ь
н
ы
й
д
е
й
с
т
в
у
ю
щ
и
й
р
е
ж
и
м
Н
е
о
б
р
а
б
о
т
а
н
н
а
я
Э
Э
Г
(
п
о
-
в
е
р
х
н
о
с
т
н
о
е
э
л
е
к
т
р
о
м
а
г
н
и
т
-
н
о
е
п
о
л
е
)
Н
е
й
р
о
н
ы
и
и
х
в
з
а
и
м
о
-
д
е
й
с
т
в
и
я
Д
р
у
г
и
е
о
с
о
б
е
н
н
о
с
т
и
Б
о
д
р
с
т
в
у
ю
щ
е
е
с
о
з
н
а
н
и
е
Л
ю
д
и
с
о
о
б
щ
а
ю
т
о
н
о
р
м
а
л
ь
-
н
ы
х
с
о
з
н
а
т
е
л
ь
н
ы
х
в
п
е
ч
а
т
-
л
е
н
и
я
х
,
м
о
г
у
т
в
ы
п
о
л
н
я
т
ь
с
т
а
н
д
а
р
т
н
ы
е
з
а
д
а
ч
и
н
а
п
о
-
з
н
а
н
и
е
и
о
с
у
щ
е
с
т
в
л
я
т
ь
п
р
о
-
и
з
в
о
л
ь
н
ы
й
(
и
с
п
о
л
н
и
т
е
л
ь
-
н
ы
й
)
к
о
н
т
р
о
л
ь
н
а
д
с
в
о
и
м
и
с
к
е
л
е
т
н
ы
м
и
м
ы
ш
ц
а
м
и
Б
ы
с
т
р
ы
е
,
с
н
и
з
к
о
й
а
м
п
л
и
т
у
-
д
о
й
,
н
е
р
е
г
у
л
я
р
н
ы
е
ф
о
р
м
ы
в
о
л
н
,
к
о
т
о
р
ы
е
с
о
д
е
р
ж
а
т
м
н
о
г
о
р
а
з
л
и
ч
н
ы
х
п
о
л
о
с
ч
а
с
т
о
т
.
Н
е
-
к
о
т
о
р
ы
е
в
ы
с
о
к
о
с
и
н
х
р
о
н
и
з
и
-
р
о
в
а
н
н
ы
е
в
и
д
ы
д
е
я
т
е
л
ь
н
о
с
т
и
м
о
г
у
т
п
р
о
и
с
х
о
д
и
т
ь
д
а
ж
е
в
т
о
в
р
е
м
я
,
к
о
г
д
а
п
о
в
е
р
х
н
о
с
т
н
а
я
Э
Э
Г
в
ы
г
л
я
д
и
т
р
а
з
н
о
р
о
д
н
о
й
В
ы
с
о
к
о
а
к
т
и
в
н
ы
и
в
з
а
и
м
о
-
д
е
й
с
т
в
у
ю
т
с
д
р
у
г
и
м
и
н
е
й
-
р
о
н
а
м
и
т
а
л
а
м
у
с
а
и
к
о
р
ы
С
в
о
б
о
д
н
о
и
д
у
щ
и
е
с
е
н
с
о
р
н
ы
е
в
х
о
д
я
щ
и
е
и
д
в
и
г
а
т
е
л
ь
н
ы
е
и
с
х
о
д
я
щ
и
е
с
и
г
н
а
л
ы
с
к
о
н
-
к
у
р
е
н
ц
и
е
й
з
а
о
г
р
а
н
и
ч
е
н
н
ы
й
о
б
ъ
е
м
д
л
я
о
с
о
з
н
а
в
а
е
м
ы
х
и
и
с
п
о
л
н
я
е
м
ы
х
с
о
б
ы
т
и
й
.
А
к
т
и
в
н
о
е
р
е
ш
е
н
и
е
п
р
о
б
л
е
м
и
о
б
у
ч
е
н
и
е
.
О
с
о
з
н
а
н
н
ы
е
с
о
б
ы
т
и
я
а
в
т
о
м
а
т
и
ч
е
с
к
и
к
о
-
д
и
р
у
ю
т
с
я
в
п
а
м
я
т
и
г
и
п
п
о
к
а
м
п
а
л
ь
н
ы
м
к
о
м
п
л
е
к
с
о
м
и
н
о
в
о
й
к
о
р
о
й
.
Н
е
к
о
т
о
р
о
е
з
а
-
к
р
е
п
л
е
н
и
е
э
п
и
з
о
д
и
ч
е
с
к
о
й
п
а
м
я
т
и
м
о
ж
е
т
п
р
о
и
с
х
о
д
и
т
ь
в
о
в
р
е
м
я
б
о
д
р
с
т
в
о
в
а
н
и
я
,
н
о
б
о
л
ь
ш
е
й
ч
а
с
т
ь
ю
п
р
о
и
с
х
о
д
и
т
в
о
с
н
е
С
н
о
в
и
д
е
н
и
е
с
б
ы
с
т
р
ы
м
и
д
в
и
ж
е
н
и
я
м
и
г
л
а
з
Н
а
в
о
п
р
о
с
ы
н
а
б
л
ю
д
а
е
м
ы
е
о
т
в
е
ч
а
ю
т
к
а
к
б
у
д
т
о
о
с
о
з
-
н
а
н
н
о
Б
ы
с
т
р
ы
е
с
н
и
з
к
о
й
а
м
п
л
и
т
у
д
о
й
н
е
р
е
г
у
л
я
р
н
ы
е
ф
о
р
м
ы
в
о
л
н
,
к
о
т
о
р
ы
е
с
о
д
е
р
ж
а
т
м
н
о
г
о
р
а
з
-
н
ы
х
п
о
л
о
с
ч
а
с
т
о
т
.
Н
е
к
о
т
о
р
ы
е
в
ы
с
о
к
о
с
и
н
х
р
о
н
и
з
и
р
о
в
а
н
н
ы
е
в
и
д
ы
д
е
я
т
е
л
ь
н
о
с
т
и
м
о
г
у
т
п
р
о
и
с
х
о
д
и
т
ь
д
а
ж
е
в
т
о
в
р
е
м
я
,
к
о
г
д
а
п
о
в
е
р
х
н
о
с
т
н
а
я
Э
Э
Г
в
ы
-
г
л
я
д
и
т
р
а
з
н
о
р
о
д
н
о
й
А
к
т
и
в
н
ы
,
т
е
с
н
о
в
з
а
м
о
д
е
й
-
с
т
в
у
ю
т
с
д
р
у
г
и
м
и
н
е
й
р
о
-
н
а
м
и
т
а
л
а
м
у
с
а
и
к
о
р
ы
З
а
б
л
о
к
и
р
о
в
а
н
н
ы
е
с
е
н
с
о
р
н
ы
е
в
х
о
д
я
щ
и
е
и
д
в
и
г
а
т
е
л
ь
н
ы
е
и
с
х
о
д
я
щ
и
е
с
и
г
н
а
л
ы
.
С
о
д
е
р
-
ж
и
м
о
е
с
о
з
н
а
н
и
я
с
о
з
д
а
е
т
с
я
в
н
у
т
р
и
.
М
о
ж
е
т
п
р
о
и
с
х
о
д
и
т
ь
р
е
ш
е
н
и
е
п
р
о
б
л
е
м
,
н
е
в
ы
р
а
-
ж
е
н
н
о
е
с
л
о
в
а
м
и
.
В
е
р
о
я
т
н
о
,
ч
т
о
б
е
з
у
с
л
о
в
-
н
а
я
и
п
р
о
ц
е
д
у
р
н
а
я
в
и
д
ы
п
а
м
я
т
и
м
о
г
у
т
з
а
-
к
р
е
п
л
я
т
ь
с
я
в
о
в
р
е
м
я
с
о
с
т
о
я
н
и
я
Б
Д
Г
Сознание и внимание
475
Т
а
л
а
м
о
к
о
р
т
и
к
а
л
ь
н
ы
й
д
е
й
с
т
в
у
ю
щ
и
й
р
е
ж
и
м
Н
е
о
б
р
а
б
о
т
а
н
н
а
я
Э
Э
Г
(
п
о
-
в
е
р
х
н
о
с
т
н
о
е
э
л
е
к
т
р
о
м
а
г
н
и
т
-
н
о
е
п
о
л
е
)
Н
е
й
р
о
н
ы
и
и
х
в
з
а
и
м
о
-
д
е
й
с
т
в
и
я
Д
р
у
г
и
е
о
с
о
б
е
н
н
о
с
т
и
С
т
а
д
и
и
с
н
а
1
–
4
С
п
о
с
о
б
н
о
с
т
ь
к
п
р
о
б
у
ж
-
д
е
н
и
ю
с
н
и
ж
а
е
т
с
я
о
т
1
-
й
с
т
а
д
и
и
к
4
-
й
,
с
о
о
б
щ
а
е
т
с
я
о
с
н
и
ж
е
н
и
и
п
р
о
ц
е
с
с
а
м
ы
ш
-
л
е
н
и
я
.
О
д
н
а
к
о
с
о
о
б
щ
а
е
т
с
я
о
н
е
к
о
т
о
р
о
й
м
ы
с
л
и
т
е
л
ь
н
о
й
а
к
т
и
в
н
о
с
т
и
д
а
ж
е
в
о
в
р
е
м
я
г
л
у
б
о
к
о
г
о
с
н
а
В
о
в
р
е
м
я
т
о
г
о
,
к
а
к
с
н
и
ж
а
е
т
с
я
с
п
о
с
о
б
н
о
с
т
ь
к
п
р
о
б
у
ж
д
е
н
и
ю
,
п
о
я
в
л
я
е
т
с
я
у
в
е
л
и
ч
и
в
а
ю
щ
е
е
с
я
к
о
л
и
ч
е
с
т
в
о
м
е
д
л
е
н
н
ы
х
,
в
ы
с
о
-
к
о
а
м
п
л
и
т
у
д
н
ы
х
р
е
г
у
л
я
р
н
ы
х
в
о
л
н
П
р
а
к
т
и
ч
е
с
к
и
с
о
г
л
а
с
о
в
а
н
-
н
а
я
о
б
ш
и
р
н
а
я
а
к
т
и
в
н
о
с
т
ь
п
о
т
и
п
у
«
ш
у
м
–
п
а
у
з
а
»
в
т
а
л
а
м
о
к
о
р
т
и
к
а
л
ь
н
о
й
с
и
-
с
т
е
м
е
.
О
д
н
о
в
р
е
м
е
н
н
а
я
«
б
у
р
н
а
я
»
а
к
т
и
в
н
о
с
т
ь
(
с
о
-
с
т
о
я
н
и
е
п
о
д
ъ
е
м
а
)
с
п
о
с
о
б
-
с
т
в
у
е
т
в
з
а
и
м
о
д
е
й
с
т
в
и
ю
,
п
о
х
о
ж
е
м
у
н
а
б
о
д
р
с
т
в
о
в
а
-
н
и
е
«
П
а
у
з
а
»
и
л
и
с
о
с
т
о
я
-
н
и
е
с
п
а
д
а
п
р
е
п
я
т
с
т
в
у
е
т
о
б
м
е
н
у
с
и
г
н
а
л
а
м
и
м
е
ж
д
у
к
л
е
т
к
а
м
и
З
а
б
л
о
к
и
р
о
в
а
н
н
ы
е
с
е
н
с
о
р
н
ы
е
в
х
о
д
я
щ
и
е
и
д
в
и
г
а
т
е
л
ь
н
ы
е
и
с
х
о
д
я
щ
и
е
с
и
г
н
а
л
ы
.
Н
е
-
д
а
в
н
и
е
э
п
и
з
о
д
и
ч
е
с
к
и
е
в
о
с
п
о
м
и
н
а
н
и
я
п
р
о
-
и
г
р
ы
в
а
ю
т
с
я
в
с
и
с
т
е
м
е
г
и
п
п
о
к
а
м
п
–
н
о
в
а
я
к
о
р
а
в
о
в
р
е
м
я
с
о
с
т
о
я
н
и
й
п
о
д
ъ
е
м
а
м
е
д
л
е
н
-
н
о
в
о
л
н
о
в
о
г
о
р
и
т
м
а
,
и
п
р
о
и
с
х
о
д
и
т
з
а
к
р
е
-
п
л
е
н
и
е
я
в
н
о
й
п
а
м
я
т
и
Э
п
и
л
е
п
т
и
ч
е
с
к
а
я
п
о
т
е
р
я
с
о
з
н
а
н
и
я
М
е
д
л
е
н
н
ы
е
,
в
ы
с
о
к
о
а
м
п
л
и
т
у
д
-
н
ы
е
в
о
л
н
ы
,
и
м
е
ю
щ
и
е
ф
о
р
м
у
п
и
к
о
в
(
с
п
а
й
к
о
в
)
,
р
е
г
у
л
я
р
н
ы
е
.
М
о
г
у
т
н
а
ч
и
н
а
т
ь
с
я
в
г
и
п
п
о
-
к
а
м
п
е
и
л
и
в
и
с
о
ч
н
о
й
д
о
л
е
,
х
о
т
я
с
о
о
б
щ
а
л
о
с
ь
и
о
д
р
у
г
и
х
и
с
т
о
ч
-
н
и
к
а
х
П
р
а
к
т
и
ч
е
с
к
и
с
о
г
л
а
с
о
в
а
н
-
н
а
я
о
б
ш
и
р
н
а
я
а
к
т
и
в
н
о
с
т
ь
п
о
т
и
п
у
«
ш
у
м
–
п
а
у
з
а
»
в
т
а
л
а
м
у
с
е
и
к
о
р
е
.
П
о
л
а
г
а
-
ю
т
,
ч
т
о
г
и
п
е
р
с
и
н
х
р
о
н
н
а
я
а
к
т
и
в
н
о
с
т
ь
в
о
в
л
е
к
а
е
т
в
с
е
р
а
с
ш
и
р
я
ю
щ
и
е
с
я
о
б
л
а
с
т
и
к
о
р
ы
П
о
т
е
р
я
ф
у
н
к
ц
и
й
,
в
к
л
ю
ч
а
я
с
о
з
н
а
н
и
е
,
м
о
ж
е
т
б
ы
т
ь
ч
а
с
т
и
ч
н
о
й
и
л
и
п
о
л
н
о
й
.
М
о
г
у
т
п
р
о
и
с
-
х
о
д
и
т
ь
а
в
т
о
м
а
т
и
ч
е
с
к
и
е
д
е
й
с
т
в
и
я
,
с
х
о
ж
и
е
с
х
о
ж
д
е
н
и
е
м
в
о
с
н
е
476
Глава 8
3. Когда вы научитесь действительно хорошо ездить на велосипеде, что про-
изойдет с вашей кортикальной активностью для сенсомоторного контроля?
4. Как могут быть согласованы две области мозга, которые представляют зри-
тельное поле?
5. Что подразумевается под усложнением мозговых волн? Приведите пример.
6. Каковы основные эффекты выборочного внимания? Какая часть мозга,
предположительно, контролирует целенаправленное внимание?
7. Что подразумевается под «архитектурой сна»? Опишите некоторые ее осо-
бенности и распределение во времени.
8. Каковы некоторые гипотезы о сне с быстрыми движениями глаз? Какой тип
функций он может иметь? Как широко сны с быстрыми движениями глаз
распространены в царстве животных?
9. Каковы основные функциональные и электрофизиологические особенно-
сти сознательного бодрствующего состояния? Какие типы кортикальных
волн встречаются во время сознательного бодрствования?
10. Опишите состояния подъема и спада медленных колебаний.
11. Опишите тренировки с помощью нейронных обратных связей. Для чего
они полезны?
12. Имеются ли убедительные данные, что гипноз отражает истинное состоя-
ние мозга?
13. Какие аргументы «за» и «против» поверхностной (с кожи головы) ЭЭГ по
сравнению с внутричерепной ЭЭГ (интракраниальной иЭЭГ)?
14. Методы медитации были описаны тысячи лет назад. На какие некоторые
их особенности наиболее часто ссылаются? Какие данные мы имеем за и
против таких методов воздействия?
Показано, как память может кодироваться в различных областях коры. Это концепция
нейробиолога Хоакина Фустера (Joaquin Fuster) относительно закодированных в коре
следов долговременной памяти. В задних отделах коры в большей степени располага-
ются перцептивные виды хранимых следов, в то время как в лобной доле — следы ис-
полнительной памяти и памяти, связанной с действиями. Предполагается, что принцип
хранения состоит в том, что память кодируется в неокортексе (видимой обширной части
коры) путем упрочения синаптических связей между многочисленными группами ней-
ронов, отвечающих за высшие когнитивные функции. (Источник: Fuster, 2004.)
Какого качества жизнь (если она вообще возможна в этом случае), какого
качества мировосприятие, какого качества самоощущение могут сохраниться
у человека, утратившего большую часть своей памяти и вместе с ней — свое
прошлое и свою способность ориентироваться во времени?
Оливер Сакс (1995)
Содержание
1.0.Введение.......................................................480
1.1.Общеепредставлениеофункцииобученияипамяти...............483
1.2 . Обучение и память на функциональной схеме работы мозга . . . . . . . . . 485
1.3.Имплицитнаяиэксплицитнаяпамять............................488
2.0.Амнезия........................................................489
2.1.HM:наиболееизученныйпациентсамнезией.....................491
2.2.Краткиесведенияобамнезии...................................491
2.3 . Сохраняющиеся функции при амнезии:
имплицитнаяипроцедурнаяпамять.............................494
2.4.Сохраненноеимплицитноеобучение.............................496
3.0.Какформируютсявоспоминания....................................500
3.1 . Электрически вызванные автобиографические воспоминания. . . . . . . . 501
3.2 . Длительная потенциация и длительная депрессия:
возбуждающиеитормозныеследыпамяти........................504
3.3 . Консолидация: от временного хранения до постоянного. . . . . . . . . . . . . 507
3.4 . Быстрая консолидация: синаптические механизмы,
транскрипциягеновисинтезбелка..............................508
3.5 . Системная консолидация: взаимодействие между медиальными
отделамивисочныхдолейиновойкорой.........................510
4.0.Разновидностипамяти............................................512
4.1 . Эпизодическая и семантическая память:
«припоминания»против«знания»...............................512
4.2 . Эпизодические воспоминания могут со временем
превращатьсявсемантические .................................515
4.3 . Эпизодическая память и семантическая память
частокомбинируются..........................................517
5.0 . Медиальная височная кора в эксплицитном научении и памяти . . . . . . . . . . 518
5.1.Отвлечениевниманиямешаетобучению..........................519
6.0.Префронтальнаякора,сознаниеирабочаяпамять.....................521
6.1 . Работа с памятью: лобная доля целенаправленно работает с памятью . 525
6.2 . Префронтальная кора в эксплицитном (осознанном)
иимплицитном(неосознанном)обученииипамяти................526
6.3.Различныетипырабочейпамяти................................528
6.4 . Префронтальная кора — хранение информации
илиуправлениепроцессом?....................................530
6.5 . Объединение префронтальных и медиальных височных областей
дляобеспеченияпроцессоврабочейпамяти.......................531
7.0.Извлечениеинформацииизпамятииметапознание....................533
7.1.Извлечениеизпамятиложнойинформации.......................534
Глава 9
Обучение и память
480
Глава 9
7.2 . Межполушарная латерализация при извлечении информации . . . . . . 534
7.3 . Тет а -ритмы могут координировать извлечение из памяти . . . . . . . . . . 536
8.0. Другиевидыобучения...........................................537
9.0. Заключение....................................................539
10.0.Заданияиупражнениякглаве9...................................540
1.0 . Введение
Память можно определить как некое стойкое представительство, отраженное в
мыслях, жизненном опыте или поведении. Обучение является приобретением
такого представительства, и в этот процесс вовлекается множество областей
мозга и видов мозговой деятельности. Человеческая память обладает как не-
которыми неожиданными ограничениями, так и некоторыми выдающимися
способностями.
Например, большинство студентов жалуются на то, что хотели бы иметь
«память получше», имея в виду лучшую способность надежно хранить заучи-
ваемую информацию и извлекать ее произвольно, легко и без потерь. Однако
человеческий мозг эволюционировал вовсе не для академических занятий и
экзаменов — это ведь совсем недавние культурные приобретения. Скорее, он
развивался для решения задач, связанных с выживанием. Лучше всего наша
память работает отнюдь не с точными последовательностями символов, что так
хорошо проделывают обычные компьютеры. Наш мозг исключительно хорош,
скорее, для решения сложных, нечетко очерченных и совершенно новых задач,
и именно с такими проблемами нам приходится иметь дело в реальном мире.
Люди исключительно гибко адаптируются к новым условиям. Обучение и
память обеспечивают самый легкий путь к достижению такой гибкости. Имен-
но эта колоссальная способность к обучению позволяет современному челове-
ку использовать свой мозг, исходно приспособленный лишь к неолитической
среде, и применять его с большим успехом в мире компьютеров, науки о мозге
и высшего образования. Должно быть, есть что-то особенное в этой нашей ве-
ликой способности к обучению.
Считается, что хранение информации в памяти сопряжено с обширными
синаптическими изменениями во многих областях коры. Этот процесс, как по-
лагают, включает широкомасштабное хеббовское обучение, происходящее со-
гласно правилу ‘fire and wire’ («нейроны, которые совместно разряжаются, со-
вместно и связаны») (см. гл . 3 и приложение). Таким образом, согласованная
активность нейронов приводит к укреплению связей между ними, как возбуж-
дающих, так и тормозных. Также предполагается, что временные клеточные
ансамбли обеспечивают формирование непосредственной памяти, в то время
как становление долговременной памяти требует более длительных закрепле-
ний синаптических связей, как возбуждающих, так и тормозных. Несмотря на
то что было получено подтверждение этих феноменов, наблюдать измененную
долговременную связь непосредственно во всех тех областях мозга, где , как
считается, она возникает, пока не удается. Таким образом, сам хеббовский след
памяти остается пока чисто логической концепцией.
Некоторые исследователи полагают, что вся кора обладает способностью к
обучению за счет изменения синаптических связей, от задних ее отделов, от-
вечающих за восприятие, до лобной — управляющей и двигательной коры
Обучение и память
481
(Fuster, 2004). Другие больше внимания обращают на височную долю, которая
традиционно ассоциируется в неврологии с функциями памяти. Важнейшие
структуры мозга, которые рассматриваются в этой главе, — это новая кора (ви-
димая внешняя поверхность головного мозга) и медиальные отделы височных
долей, которые содержат два гиппокампа и окружающие их ткани (рис. 9 .1 и
9.2). Медиальная височная доля имеет сложную пространственную структуру
и, чтобы полностью понять ее форму и расположение, необходимо как следует
поупражняться, используя рисунки, приведенные в конце этой главы.
До недавнего времени считалось, что за перевод пережитых событий в па-
мять в основном ответственен гиппокамп, но в настоящее время использование
более качественных методов показало, что к этому причастны и окружающие
его ткани — медиальные отделы височных долей. Эти области перекрывают-
ся с древней обонятельной корой млекопитающих, которая имеет менее шести
клеточных слоев.
Неокортекс, который является «новой» корой современных млекопитаю-
щих, всегда имеет шесть слоев, что служит одним из его самых характерных
признаков. Гигантский неокортекс и медиальная височная кора постоянно вза-
имодействуют друг с другом, поскольку информация непрерывно сохраняется
и извлекается из потока повседневных впечатлений.
Из-за своего древнего эволюционного происхождения медиальные отделы
височных долей обладают множественными функциями. Так, вначале у крыс
Рис. 9 .1 . Медиальные отделы височных долей и гиппокампы. Области, ответственные
за память, обладают сложной пространственной структурой, которую трудно визуали-
зировать. На этом рисунке они представлены с двух точек зрения. Слева располагается
фронтальный поперечный срез мозга, проходящий через медиальные отделы височных
долей (MTL), обведенные красными кружками. Справа изображены гиппокампы обоих
полушарий, на верхушке которых находятся отмеченные красным структуры, миндали-
ны. Следует отметить, что гиппокампы имеют форму петли и располагаются с внутрен-
ней стороны каждой височной доли. Медиальная височная кора включает гиппокампы
и соседние «ринальные» структуры (см. гл . 5). (Источник: Sh��� Fu.)
482
Глава 9
и других млекопитающих было обнаружено, что гиппокамп служит картой
собственного пространственного местоположения. Однако он также кодирует
и обоняние, в связи с чем в медиальной височной коре выделяют части, назы-
ваемые ринальной, энторинальной и периринальной корой (от греч. ринос —
нос).
Медиальные отделы височных долей также взаимодействуют со зритель-
ной областью IT (inferior temporal lobe), нижним отделом височной доли (см.
рис. 9 .2). Читатель, может быть, помнит, что нижневисочная область IT, как
было показано, интегрирует общую информацию о зрительном образе (см.
гл. 6). Разряды нейронов в этой области коррелируют с осознанным зритель-
ным восприятием (She���er�, L���thet�s, 1997). Таким образом, медиальные от-
She���er�, L���thet�s, 1997). Таким образом, медиальные от-
, L���thet�s, 1997). Таким образом, медиальные от-
L���thet�s, 1997). Таким образом, медиальные от-
, 1997). Таким образом, медиальные от-
делы височных долей расположены очень удачно для восприятия обобщенной,
по-видимому, осознанной, зрительной информации.
Рядом с медиальной височной располагается слуховая кора, что указывает
на то, что в медиовисочную может также передаваться и слуховая информация
(см. гл . 7), а возле верхушек гиппокампов располагаются миндалины, основ-
ной центр обработки эмоциональной информации. Таким образом, медиальные
отделы височной коры являются тесно взаимодействующими перекрестками
информационных путей, удобно расположенными для объединения множе-
ства поступающих в мозг сигналов и для координации обучения и извлечения
информации в различных частях неокортекса. Это поистине «центр центров»
каждого полушария большого мозга (рис. 9.3).
Рис. 9.2 . Области, ответственные за память, получают визуальную информацию об
объекте. На этом изображении медиальной поверхности полушария мозга показано,
что медиальные отделы височной доли (MTL) тесно связаны с областью IT, нижней
височной корой. Нижневисочная область IT, по-видимому, обеспечивает сознательное
восприятие зрительного образа (см. гл . 8). Медиальная височная кора включает также
миндалину. Слуховая кора располагается совсем рядом с медиовисочной, с внешней
стороны височной доли. (Источник: Vuilleumier, 2005.)
Обучение и память
483
Большая часть коры головного мозга человека относится к неокортексу,
который за более чем 100 млн лет эволюции млекопитающих «раздулся» из
более древних его областей. Как уже говорилось, считается, что новая кора ко-
дирует долговременную память путем изменения синаптических связей между
миллиардами нейронов. В новой коре и подчиненных ей структурах, особенно
таламусе, имеются буквально триллионы таких синапсов (см. гл . 3). Местопо-
ложение гиппокампа идеально для объединения информации от когнитивных
(неокортекс) и эмоциональных (лимбических) областей и связи этой информа-
ции в след памяти, кодирующий все аспекты событий нашего познавательного
опыта (M�sc�v�tch, 1995).
1.1 . Общее представление о функции обучения и памяти
На рис. 9 .4 показано, что визуальный образ — например, чашки кофе на сто-
ле — вначале формируется в зрительной коре. Затем активность коры, соот-
Рис. 9.3 . Медиальные отделы височной доли (MTL) являются центром, имеющим об-
ширные связи. Эти отделы, включая периринальную кору, имеют весьма обширные свя-
зи со зрительной, слуховой, соматосенсорной, эмоциональной, моторной, управляющей
областями и памятью. Поэтому медиальная височная кора является идеальным местом
для получения, связывания и распределения информации в долговременной памяти.
Обратите внимание, что практически все связи идут в обоих направлениях (двойные
стрелки). (Источник: Murr�y, R�chm���, 2001.)
484
Глава 9
ветствующая воспринятому образу чашки кофе, распространяется на медиаль-
ную височную кору, которая активирует и «связывает» следы памяти, обширно
распределенные как по зрительной коре, так и по другим областям. Некоторые
ученые полагают, что в течение короткого периода времени, составляющего
несколько десятых долей секунды, афферентные зрительные области и другие
«связанные» с ними части новой коры резонируют друг с другом, о чем сви-
детельствует появление согласованной электрической активности в гамма-диа-
пазоне (эти представления часто называют «40-Герцовой гипотезой») (Ll���s,
Pare, 1991; E��el, S���er, 2001; Freeman et al., 2003). Осмысление визуального
стимула, такого как чашка кофе, требует, вероятно, нескольких сотен милли-
секунд. Таким образом, менее чем за секунду зрительная кора распознает объ-
ект, находящийся перед глазами, и активирует медиальную область височной
доли, связывающую различные области неокортекса для запуска формирования
Рис. 9 .4 . Каким образом медиальные височные области помогают хранить и извлекать
из памяти эпизодические воспоминания? Слева: образ чашки кофе на столе активирует
зрительную кору до уровня восприятия объекта (см. гл . 6). В центре: сохранение до-
стигается тогда, когда медиальная височная область координирует следы памяти (в том
числе синаптические изменения), обширно распределенные по многим областям коры.
Они включают семантические ассоциации с чашкой кофе (кофейные зерна на картинке
внизу). Визуальные признаки чашки (ее ручка) также являются частью активирующе-
гося ассоциативного комплекса. Когда на следующий день эпизодическая память (образ
чашки кофе) вновь активируется (например, вопросом: «Тебе понравился кофе, который
я вчера сварила?»), медиальная височная кора снова задействуется в извлечении и орга-
низации обширно распределенных корковых следов памяти. Поэтому зрительной коре
необходимо восстановить образ чашки кофе, который отнюдь не идентичен исходной
чашке, а лишь более или менее точно воссоздает картину соответствующей зрительной
активации, которая перекрывается с исходной картиной. Важно, что зрительная кора
принимает участие в восприятии, обучении и извлечении информации из эпизодиче-
ской памяти
Обучение и память
485
следов памяти. Тем не менее, как будет видно из дальнейшего, постоянная па-
мять требует больше времени для консолидации.
Память предназначена для использования ее в реальном мире, и по этой
причине извлечение информации из памяти так же важно, как и обучение. Ког-
да мы сталкиваемся с напоминанием о специфическом прошлом опыте, каса-
ющемся чашки кофе, следы памяти, связанные с этим событием, вновь «вы-
свечивают» соответствующие области коры. Таким образом воспроизводится
некоторая часть исходного воспоминания, вновь с использованием медиальной
височной области, для объединения следов памяти в когерентный сознатель-
ный опыт. Такой опыт в данном случае — воспоминание о вчерашней чашке
кофе — опять включает использование зрительной коры. Поскольку данный
вопрос является основной темой этой главы, начнем со схематического изобра-
жения, представленного на рис. 9 .4 .
Обучение не ограничивается участием новой коры. Другие типы обучения
используют другие области мозга (см. раздел 8.0). В этой главе, однако, мы
остановимся на обучении и извлечении из памяти ежедневных воспоминаний.
1.2 . Обучение и память на функциональной схеме работы мозга
В соответствии с нашей функциональной схемой, входящий сенсорный сигнал
идет в «рабочее хранилище», являющееся частью рабочей памяти, которая,
в свою очередь, позволяет активно сохранять и управлять информацией (рис.
9.5; см. гл . 2). Рабочая память позволяет нам временно удерживать в памяти
в доступной форме небольшие объемы информации.
Мы обращаемся к этой способности рабочей памяти при решении множе-
ства ежедневных задач, таких как сохранение в уме телефонного номера в те-
чение нескольких секунд, которые требуются для его набора. Рабочая память
также дает нам ощущение непрерывности во времени путем встраивания на-
ших непосредственных сознательных впечатлений в более длительное психо-
логическое настоящее. Точная взаимосвязь между сознательными событиями,
рабочей памятью и избирательным вниманием продолжает обсуждаться. Не-
которые ученые полагают, что рабочая память формирует наши сознательные
впечатления; другие высказывают мнение о том, что рабочая память сама ко-
ординируется когнитивным сознанием (���rs, Fr���l��, 2003). Однако все со-
���rs, Fr���l��, 2003). Однако все со-
, Fr���l��, 2003). Однако все со-
Fr���l��, 2003). Однако все со-
, 2003). Однако все со-
гласны с тем, что все три элемента очень тесно взаимодействуют. Это и есть
самое главное.
Двумя ключевыми свойствами рабочей памяти являются ее относительно
небольшая емкость и ограниченная продолжительность. В своей знаменитой
статье насчет объема непосредственной памяти Джордж Миллер (�e�r�e M�ll-
�e�r�e M�ll-
M�ll-
M�ll-
er, 1956) сделал вывод, что в любой момент времени в непосредственной памя-
, 1956) сделал вывод, что в любой момент времени в непосредственной памя-
ти может храниться только семь (плюс или минус два) элементов. В более позд-
ней работе показано, что истинный объем рабочей памяти может быть даже
меньше — скорее, в отсутствие повторения, рехэрсела*, всего четыре элемента
(C����, 2001; см. гл. 2). Аналогичным образом и время, в течение которого
данный элемент хранится в рабочей памяти, очень короткое — порядка секунд.
Почему система памяти сформировалась с такими ограничениями — неиз-
вестно. Одним из возможных объяснений может быть то, что ограничения раз-
* Рехэрсел (rehe�rs�l) в психологии — намеренное повторение (в уме или вслух) какой-либо информа-
rehe�rs�l) в психологии — намеренное повторение (в уме или вслух) какой-либо информа-
) в психологии — намеренное повторение (в уме или вслух) какой-либо информа-
ции для того, чтобы временно удержать ее в памяти. — Прим. перев.
486
Глава 9
меров рабочей памяти создают преимущество для незначительной по объему,
но наиболее важной для решения текущих задач информации, предохраняя ее
таким образом от интерференции с менее существенной. Соответственно, эле-
менты, хранящиеся в рабочей памяти, должны быстро удаляться, чтобы не бло-
кировать поступление в нее новой информации.
Интерпретация и обращение с материалом, хранящимся в рабочей памяти,
определяются текущими целями и задачами, а также наличными знаниями.
Сенсорная и поступающая изнутри информация осознается при концентра-
ции на ней внимания. Ряд теоретиков утверждает, что, как только информация
становится осознанной, она быстро кодируется в долговременную память (на-
пример, M�sc�v�tch, 1990). Предполагается, что некоторое неосознаваемое об-
M�sc�v�tch, 1990). Предполагается, что некоторое неосознаваемое об-
, 1990). Предполагается, что некоторое неосознаваемое об-
учение также возможно, однако до сих пор был продемонстрирован перевод в
долговременную память лишь реакции бессознательного условнорефлекторно-
го страха (LeD�ux, 1996).
В общем, если предъявление стимула осознано, то оно хорошо коррелирует
с обучением. Хотя детали этой целостной концепции продолжают обсуждаться,
именно с нее удобно начать изложение дальнейшего материала.
На рис. 9 .5 приводятся некоторые свойства обучения и памяти. Как видно,
сознательная когнитивная деятельность приводит к эксплицитному обучению
и извлечению информации из памяти. Очевидный пример — попытка запом-
нить какой-либо термин, относящийся к когнитивной нейронауке. Далеко не
столь очевидно, однако, что, наряду с формированием реакции на осознавае-
мые или эксплицитные стимулы, происходит также и имплицитное обучение
(раздел 2.4).
Таким образом, на рис. 9 .5 демонстрируется как эксплицитное, или осознан-
ное, так и имплицитное, или неосознанное, обучение. Эпизодическая память
(также называемая автобиографической памятью) является хранилищем осоз-
нанных жизненных эпизодов. Семантическая память, обычно рассматриваемая
как память о фактах, также является сознательной в том строгом смысле, что
люди могут точно изложить факты, которым они верят. Это стандартное ра-
бочее определение сознательных мозговых явлений (см. гл . 8). Наконец, воз-
можности перцептивной памяти, такие как способность «научиться слушать»
музыку и воспринимать живопись, также включают в себя сознательные, экс-
плицитные разновидности воспоминаний.
На рис. 9 .5 справа представлено также обучение с помощью имплицитной
памяти. Младенцы, например, слышат речевые последовательности звуков, но
они ведь не изучают эксплицитно грамматические правила и закономерности.
Все это, очевидно, воспринимается неосознанно и будет рассмотрено далее.
В целом, имплицитное обучение часто вызывается эксплицитными, осознан-
ными событиями, однако оно часто выходит далеко за рамки таких событий,
связанных с сознательным опытом (������, �ree���l�, 1995). Привычки, вы-
������, �ree���l�, 1995). Привычки, вы-
, �ree���l�, 1995). Привычки, вы-
�ree���l�, 1995). Привычки, вы-
, 1995). Привычки, вы-
работанные в результате многократного повторения, и двигательные навыки
также в значительной степени являются имплицитными. Как будет видно из
дальнейшего, «прайминг-эффекты» тоже часто являются имплицитными. Кон-
текст-зависимые феномены, связанные с восприятием зрительного простран-
ства, направления падающего света в визуальной картине, абстрактные поло-
жения, высказанные в разговоре и т. д., часто тоже являются имплицитными.
Обучение и память
487
Возникающие эффекты зачастую трудно произносимы, имплицитны и отчасти
неосознанны (���rs, 1988).
Как будет видно из дальнейшего, «центральный регулирующий орган» ра-
бочей памяти играет важную роль в долговременном обучении и извлечении
информации. Например, если вы заучиваете новое слово, то произвольно по-
вторяете его про себя, при этом управляющие системы мозга контролируют по-
вторение слова в уме и блокируют отвлечение внимания. Во время сессии хоро-
шо проконтролировать самого себя «метакогнитивно» — т. е. подумать о своем
собственном мыслительном процессе и определить, является ли уровень усво-
ения материала достаточным, чтобы успешно сдать экзамен. Все приведенные
ситуации являются примерами управляемых процессов (см. гл . 10, 11 и 12).
Рис. 9 .5 . Эксплицитные и имплицитные аспекты памяти. Функциональная диаграмма
обучения и памяти. Рабочая память (вверху) может рассматриваться как входная по от-
ношению к различным типам долговременной памяти, разделенной на эксплицитную и
имплицитную. Эксплицитное обучение и извлечение информации включают осознан-
ное знание как для фактов, так и для автобиографических данных. Память на факты
называется семантической памятью, тогда как автобиографическая память также на-
зывается эпизодической, поскольку в ней отображаются жизненные события. Рабочая
память может управлять эксплицитными воспоминаниями, такими как слова, числа,
семантические факты и автобиографические события. Имплицитное обучение и извле-
чение информации включает так называемые «прайминг-эффекты»*
, а также привычки,
выработанные в результате многократного повторения, и моторные навыки
* Прайминг-эффект (эффект установки, преднастройки, предшествования) — качественное (наличие/
отсутствие) либо количественное (скорость, точность) изменение параметров реакции или действия
человека в ответ на второе появление объекта по сравнению с условием, когда предшествующая
встреча отсутствовала. В отечественной психологии единого аналога термина «pr�m���» до сих пор
не выработано. (М. В. Фаликман. http://v�rtu�lc��l��.cs .msu.su/pr��ects/pr�m���.html) — Прим. перев.
488
Глава 9
1.3 . Имплицитная и эксплицитная память
Имплицитная память не сопровождается осознанием наличия того или иного
воспоминания; к заключению о существовании воспоминания приходят, только
исходя из косвенных эффектов, которые оно оказывает на поведение. Импли-
цитные воспоминания могут извлекаться из памяти непроизвольно. Для иссле-
дования имплицитной памяти широко используются прайминг-эффекты.
Под термином «прайминг» подразумевается эффект стимула по формиро-
ванию готовности к воздействию сходного стимула. Например, демонстрация
картинки лица повышает эффективность обработки изображения следующего
лица, что выражается более быстрой и точной реакцией. Прайминг может быть
как перцептивным, так и концептуальным.
Допустим, вас просят прочесть список слов, а через неделю проверяют, как
вы их запомнили. Можно проверить вашу память непосредственно, попросив
воспроизвести все заученные слова, которые сможете вспомнить, либо пред-
ложить опознать знакомые слова в смешанном списке из старых и новых слов.
Если интервал был достаточно длинным, то вы, вскоре всего, вспомните или
опознаете только немногие слова, или будете путать выученные слова и новые.
Тем не менее если память проверяется косвенным образом и при этом дает-
ся инструкция прочесть знакомые слова из общего списка как можно быстрее,
то испытуемый произносит знакомые слова быстрее, чем новые. Старые сло-
ва при этом являются «праймами». При такой опосредованной проверке никто
не говорит о памяти, и испытуемый обычно даже не осознает, что тестируется
именно память. Однако по тому, насколько быстро испытуемые читают то или
иное слово, можно сказать, оставил ли предыдущий опыт след в их памяти.
Например пациенты с амнезией не могут вспомнить заученные слова вообще.
В случае концептуального или семантического прайминга такие слова, как,
например, «пища», могут повысить эффективность запоминания таких слов,
как «вода», даже если они имеют мало общего перцептивного содержания.
Прайминг можно рассматривать как один из способов подстраиваться под
общее свойство мозга в любой момент включаться в обработку предиктивной
(т. е . связанной с предполагаемыми, будущими событиями.
—
Прим. перев.)
информации.
Перцептивный прайминг основывается на изменениях перцептивного об-
раза в задних отделах новой коры, связанных с обработкой перцептивной ин-
формации. Концептуальный прайминг связан с изменениями концептуальных
систем в префронтальной коре.
1.3.1. Процедурная память
Процедурная память относится к сенсомоторным привычным реакциям и авто-
матизированным навыкам, которые являются, по большей части, бессознатель-
ными. В этих машинальных движениях принимают участие базальные ганглии.
Представьте себе, что едете на велосипеде и заваливаетесь вправо. Как избежать
падения? Многие велосипедисты говорят, что они бы восстановили равновесие,
повернув руль влево, не понимая, что такое действие только ускорит падение.
В действительности во время езды на велосипеде в аналогичной ситуации те же
самые велосипедисты поворачивают руль велосипеда в направлении падения. При-
веденный пример подчеркивает различие между имплицитным и эксплицитным
знанием. Имплицитное обучение относится к способности усваивать разом слож-
Обучение и память
489
ную информацию (например, такие навыки, как езда на велосипеде) в отсутствие
эксплицитного осознавания. Такие случаи, как пример с велосипедом, являются
субъективно неопровержимыми доказательствами существования имплицитных
форм знания, которые отличаются от эксплицитного знания (а возможно, и проти-
воречат ему) ... (Curran, 2001).
Различные классы сенсомоторного обучения опосредуются через нейрон-
ные сети: базальные ганглии — лобная кора (Yin , K���lt��, 2006).
2.0 . Амнезия
В гл. 2 рассматривается случай Клайва Уэринга (Cl�ve We�r���), который жил
с глубокой амнезией с 1985 г., когда в результате вирусной инфекции разру-
шились некоторые области его мозга, ответственные за память. За несколько
дней Уэринг превратился из перспективного молодого музыканта в человека,
который каждое пробуждение воспринимает, как первое, у которого нет почти
никаких воспоминаний о прошлом и который не может ничему научиться.
Мало что изменилось для Уэринга с 1985 г. Хоть он и не может вспомнить
какое-то отдельное определенное событие, некоторые аспекты его памяти со-
храняются. Он может поддерживать нормальный разумный разговор. У него
сохранилась кое-какая кратковременная память, позволяющая удерживать нить
беседы на протяжении нескольких секунд. Он сохранил общее представление
об окружающем мире, обширный словарный запас и невербальное понимание
правил поведения. Уэринг остался искусным музыкантом, способным играть
по нотам сложные фортепьянные пьесы. Хоть он и не помнит никаких опреде-
ленных событий, он тем не менее может воспроизвести некоторые основные
моменты своей жизни. Среди немногочисленных сохранившихся воспомина-
ний — его жена Дебора. Он так радостно приветствует ее всякий раз, как она
входит в его комнату, словно они годами не виделись.
Однако уже через несколько мгновений после ее ухода Уэринг вообще за-
бывает, что она здесь была. В своей недавно опубликованной книге Дебора Уэ-
ринг (2005) рассказывает, в частности, как она вернулась домой после посеще-
ния Клайва в больнице и обнаружила следующие сообщения на автоответчике:
Привет, любимая, это я, Клайв! Сейчас 5 минут пятого, и я не знаю, что здесь про-
исходит. Я только что проснулся и еще ни с кем не разговаривал...
Дорогая? Здравствуй, это я, Клайв. Сейчас 15 минут пятого, и я только что про-
снулся. Все случилось минуту назад, и я хочу тебя видеть...
Дорогая? Это я, Клайв, сейчас 18 минут пятого, и я проснулся. Я открыл глаза
только минуту назад. Я еще ни с кем не разговаривал, я только хочу поговорить с
тобой.
История Уэринга наводит на мысль о том, что амнезия избирательна — опре-
деленные типы памяти могут сохраняться, в то время как другие утрачиваются.
Навыки, такие как способность говорить или играть на фортепьяно, отличают-
ся от наших воспоминаний об отдельных событиях. Таким образом, память не
едина, а состоит из различных типов. История Уэринга также указывает на то,
что различные части мозга могут быть связаны с разными типами памяти.
На рис. 9 .6 показано, насколько специфичной является потеря памяти
у Клайва Уэринга. Органическая амнезия (тип потери памяти, который связан
с повреждением обеих медиальных височных долей) затрагивает эпизодиче-
490
Глава 9
ское обучение и воспроизведение информации. Кроме того, нарушается се-
мантическое обучение, но не извлечение семантической информации. Клайв
Уэринг воспринимает окружающий мир практически таким же, каким он его
воспринимал в 1985 г.; его предыдущее семантическое познание явно остается
незатронутым. Однако новому он не может научиться. Кроме того, он не может
ни познать, ни вспомнить какие-либо отдельные события. Позднее мы увидим,
в чем смысл таких нарушений памяти.
Хотя у всех пациентов с амнезией отмечается потеря памяти, степень ее
различается. Амнезия у Клайва Уэринга напоминает амнезию других паци-
ентов, однако она необычна постоянно возникающим чувством, что он только
что очнулся от некоего бессознательного состояния. У него также наблюдается
«персеверация» в большей степени, чем у большинства пациентов с амнезией,
он повторяет одни и те же мысли и действия вновь и вновь, как в вышеприве-
Рис. 9 .6 . Функциональная диаграмма: типичная потеря памяти при органической амне-
зии. Амнезия, связанная с двусторонним повреждением медиальных височных долей,
высокоспецифична. При ней нарушается восстановление эпизодических (автобиогра-
фических) воспоминаний и блокируется эпизодическое обучение. Кроме того, стано-
вится невозможным изучение новых фактов и концепций (семантическое обучение).
Тем не менее пациенты с амнезией, такие как НМ и Клайв Уэринг, могут вести вполне
вроде бы нормальный разговор, поскольку они все еще в состоянии извлекать ту се-
мантическую информацию, которую усвоили до повреждения мозга. Имплицитное об-
учение и извлечение информации у них также сохраняются. Поскольку эпизодическое
обучение осуществляется при взаимодействии медиальной височной и новой коры,
специфические нарушения у этих больных объясняются, по-видимому, двусторонним
повреждением медиальных височных долей. (Следует отметить, что термины «экспли-
цитный» и «сознательный», по существу, означают одно и то же; оба этих термина при-
меняются при изложении историй болезни (см. гл . 8). Аналогичным образом, термины
«имплицитный» и «бессознательный» являются для наших целей равнозначными.)
Обучение и память
491
денном примере с одними и теми же телефонными сообщениями, которые он
вновь и вновь оставляет своей жене. Эти симптомы могут быть результатом
дополнительного повреждения префронтальных структур, которые в норме по-
зволяют отслеживать наши собственные действия.
2.1. HM: наиболее изученный пациент с амнезией
Мы многое знаем о жизни Клайва Уэринга из многочисленных сообщений в
литературе об этом случае. Однако наиболее изученной жертвой амнезии явля-
ется, безусловно, другой пациент, известный только по инициалам НМ. В слу-
чае НМ мы точно знаем, где располагается повреждение (рис. 9 .7 и 9.8), что
встречается крайне редко. Большинство повреждений мозга являются очень
«размытыми», могут распространяться шире, чем видимые разрушения, и из-
меняться со временем. Вирусная инфекция у Клайва Уэринга явно разрушила
гиппокампальные области с обеих сторон, а также некоторые части лобных до-
лей. У него также могло быть и какое-то другое повреждение мозга, о котором
мы просто ничего не знаем. В случае же с НМ разрушение было нанесено са-
мим хирургом, в результате чего точно известно, что у него с обеих сторон были
удалены лишь медиальные отделы височных долей, причем настолько аккурат-
но, насколько это было возможно в то время.
Объем повреждения мозга и функционального дефицита у НМ был тща-
тельно подтвержден более чем в 100 опубликованных статьях. Это сделало НМ
одним из самых важных пациентов за всю историю науки о мозге (см. блок 9.1).
2.2 . Краткие сведения об амнезии
У НМ отмечаются признаки амнезии в очень «чистой» форме (блок 9.1). В бо-
лее общем смысле амнезия представляет собой любую потерю памяти о лич-
ном опыте и другой информации, несмотря на нормальные протекание других
когнитивных функций. Причина амнезии может быть органической, включая
Рис. 9.7 . Двустороннее повреждение гиппокампа у классического пациента НМ.
В центре: изображении поперечного сканирования мозга, показывающее повреждения
височных долей у НМ. Справа: для сравнения приводится сканированное изображение
нормального мозга, на котором белыми стрелками указаны медиальные отделы височ-
ных долей. Эти области у НМ отсутствуют. Сл ева: приведено изображение мозга с уда-
ленным участком, помогающее понять расположение поперечных срезов. (Источник:
(слева) Aminoff, D�r�ff, 2003; (справа) C�r��� et al., 1997.)
492
Глава 9
инфекцию, инсульт, опухоль, лекарственные средства, кислородное голодание,
эпилепсию и дегенеративные заболевания, такие как болезнь Альцгеймера.
Амнезия также может быть психогенной в результате травмы или внушения
(Nilsson ��� M�r����tsch, 1999).
Как видно из вышеизложенного, органическая амнезия вызывается двусто-
ронним повреждением медиальных отделов височных долей, включающих гип-
покампальную формацию. При этом обычно обнаруживается ряд признаков.
1. Нарушается память, но сохраняется восприятие, узнавание, умственные
способности и поведение. Больные с амнезией нормально выполняют стан-
дартные тесты на интеллект, однако ошибаются в стандартных тестах на
память. Они могут играть в шахматы, решать кроссворды, собирать пазлы,
воспринимать сложные инструкции и рассуждать логически.
2. Нарушается долговременная, но не рабочая память. У людей с амнезией
наблюдаются нормальные результаты теста на запоминание цифр. Однако
результаты ухудшаются, если внимание больных на что-то отвлекается. То
же самое относится к запоминанию слов, историй, зрительных образов, лиц,
мелодий, запахов и осязательных ощущений.
Рис. 9 .8. Медиальные отделы височных долей и повреждения мозга у НМ, вид сни-
з у. ( a) ориентация головы и мозга; (б) изображение нижней поверхности медиальной
височной доли, на котором отмечены основные субрегионы, ответственные за память.
Следует отметить, что название ринальная (обонятельная) кора свидетельствует о древ-
нем происхождении этой области. На всех рисунках сверху указан важный ориентир —
две обонятельные луковицы. (в) Хирургическое повреждение мозга у НМ. В то время
хирурги не знали о важности этой области для памяти. (Источник: (б) �uc�ley, ��ff��,
2006; (в) M�rr�s M�sc�v�tch, личное сообщение.)
Обучение и память
493
Блок 9.1 . Случай НМ
Когнитивная нейронаука о памяти началась, пожалуй, с сообщения Герберта Ско-
вилла и Бренды Милнер (Sc�v�lle, M�l�er, (1957) о нарушении памяти у пациента
НМ после двустороннего удаления у него медиальных отделов височных долей с
целью устранения тяжелых эпилептических приступов. В детстве НМ упал с велоси-
педа и получил травму головы, после чего его стали донимать эпилептические при-
ступы, частота и тяжесть которых нарастала к 30 годам. В качестве крайней меры
Сковилл выполнил операцию по удалению гиппокампа и окружающей ткани с обеих
сторон мозга НМ (см. рис. 9 .8). Хотя операция и снизила частоту приступов у НМ,
она оказала глубокое и непредвиденное влияние на его память. В то время возмож-
ность возникновения такого результата была неизвестна и, если бы Сковилл знал об
этом драматическом последствии, то, несомненно, изменил бы план операции. Это
сообщение Сковилла и Бренды Милнер (1957) о случае НМ было первым, где непо-
средственно продемонстрирована важность гиппокампа и окружающих его структур
для памяти.
В результате НМ не мог вспомнить никакие события своей послеоперационной жиз-
ни — ни с кем он встречался, ни что он делал, ни что происходило в мире вокруг него.
Он не знал, сколько ему теперь лет, и не узнавал свое новое отражение в зеркале.
В дополнение к антероградному (после повреждения) дефициту памяти, НМ так-
же не мог вспомнить события или впечатления за несколько лет, непосредственно
предшествовавших операции — ретроградная амнезия. Несмотря на острую потерю
эпизодической (автобиографической) памяти, другие когнитивные функции, видимо,
оставались незатронутыми. Он мог рассуждать, решать проблемы и поддерживать
нормальный разговор. Его интеллект оставался нормальным и языковые способно-
сти сохранились.
Кратковременная память у НМ сохранилась незатронутой. Результаты его тестов на
рабочую память, например задачи на запоминание ряда цифр, были такими же, как
и у здоровых людей. С юных лет НМ находился под присмотром врачей, и его ум-
ственные способности до операции были подробно описаны. Специфический очаг
поврежденной ткани был четко ограничен и хорошо охарактеризован. У большин-
ства пациентов с амнезией повреждения более обширные и их границы трудно точно
установить. С момента операции пациенту НМ множество раз проводились тестиро-
вания и сканирования, что дало очень полную картину его состояния.
Глядя на рис. 9 .1 и 9.2, можно высказать предположение, что новая кора (наружные
структуры на сканированном изображении мозга) у НМ оставалась явно не затро-
нутой. Как и Клайв Уэринг, НМ мог поддерживать нормальный разговор. Он мог
обсуждать события, непосредственно происходящие в настоящий момент, исполь-
зуя свои общие представления об окружающем мире. Он осознавал происходящее,
осуществлял нормальный волевой контроль над своими действиями и, по-видимому,
был эмоционально хорошо уравновешен. Нарушения возникали только при исследо-
вании его эпизодической памяти, когда выяснялось, что он просто не может вспом-
нить прошлое или приобрести новые знания касательно будущего.
Читателю будет полезно повторить изученный материал о некоторых важных об-
ластях коры (рис. 9 .3, 9.9). В частности, необходимо вспомнить расположение пре-
фронтальных областей лобных долей (кпереди от моторной и премоторной коры).
Вся новая кора играет важную роль в формировании памяти, однако префронтальные
области, возможно, имеют особое значение.
494
Глава 9
3. Нарушается память на недавние, но не на отдаленные события. Особенно
заметна потеря памяти на то, что узнавалось после начала заболевания и
в период, непосредственно предшествовавший ему, но не на информацию,
приобретенную за годы до этого. Таким образом больные страдают в весьма
значительной степени от антероградной амнезии, захватывающей будущее,
но лишь незначительно — от ретроградной амнезии. Длительность и тя-
жесть ретроградной амнезии у больных различаются.
4. Нарушается эксплицитная, но не имплицитная память. Антероградная
(после повреждения) потеря памяти относится только к той информации,
которая может запомниться сознательно, или эксплицитно. Обучение, со-
хранение и извлечение воспоминания без участия сознания или имплицитно
остаются нормальными.
2.3 . Сохраняющиеся функции при амнезии:
имплицитная и процедурная память
Как упоминалось выше, имплицитная память, как правило, оценивается в зада-
чах на прайминг-эффекты (эффекты предшествования). Перцептивное предше-
ствование (перцептивный прайминг) опосредуется сенсорной корой, а концеп-
туальное предшествование (концептуальный прайминг) предполагает, видимо,
активацию как височных, так и префронтальных областей. Пациенты с амне-
зией не утрачивают способности хорошо выполнять прайминг-задачи (на эф-
фекты предшествования), такие как восстановление целого слова по его фраг-
ментам. К примеру, испытуемые просматривают список слов (и в нем, скажем,
встречается слово металл). Затем им предлагают фрагменты слов, которые они
изучали (например, мет_), чтобы проверить, смогут ли они восполнить слово
по его фрагменту. Время изучения списка постепенно уменьшается. Пациенты
с амнезией могут выполнять такие задачи не хуже здоровых испытуемых.
Функциональные нейровизуализационные исследования (фМРТ) подтверж-
дают наличие перцептивного локуса. В отличие от тестов на эксплицитную па-
мять, которые ассоциируются с повышенной активацией во время извлечения
информации тех областей мозга, которые обеспечивают формирование памяти,
таких как медиальная височная и префронтальная кора, перцептивный прай-
минг ассоциируется со сниженной активацией при повторных предъявлени-
ях стимулов тех областей мозга, которые, как считается, опосредуют перцеп-
тивное представительство. Таким образом, повторное предъявление стимула
в виде лиц и слов приводит к снижению активации нижней височной и экс-
трастриарной коры, которые отвечают за восприятие лиц и слов (W���, M�rt��,
1998; Sch�cter et al., 2004).
2.3.1 . Концептуальный прайминг
В случае концептуального прайминга связь между изучением и исследуемыми
предметами основывается на смысловом содержании.
Концептуальные задания бывают на словесную ассоциацию («назовите
первое слово, которое приходит на ум при слове «слон»), на поиск термина
из определенной категории («приведите пример животных») и на общее зна-
ние («какое животное используется в Индии для перевозки тяжелых грузов?»)
(M�sc�v�tch et al., 1993; R�e���er, McDerm�tt, 1993). Концептуальный прайминг
наблюдается в том случае, если изученные слова (например слон) вспомина-
Обучение и память
495
ются чаще, чем неизученные. Поскольку концептуальный прайминг зависит от
смысла, изменение физической формы стимулов оказывает на него незначи-
тельное влияние.
Концептуальный прайминг нарушается у людей с повреждением областей
коры, ответственных за семантику. Таким образом, люди с семантической де-
менцией, у которых дегенеративный процесс затрагивает латеральные и перед-
ние височные доли, неспособны опознавать повторяющиеся объекты, имею-
щие общий смысл, например два телефонных аппарата разного вида. Однако
у них не возникает проблем с опознанием объектов одинаковой формы (�r�h�m
et al., 2000).
Сходным образом у пациентов с болезнью Альцгеймера сохраняется пер-
цептивный прайминг, однако нарушается концептуальный прайминг. В функ-
циональных нейровизуализационных (фМРТ) исследованиях концептуального
прайминга показано вовлечение в процесс областей мозга, в которых осущест-
вляется обработка семантической информации, таких как префронтальная и
латеральная височная области. Как и в тестах на перцептивный прайминг, в те-
стах на концептуальный прайминг отмечается снижение активации этих об-
ластей (�uc��er et al., 1998; Sch�cter et al., 2004). Когда людей, скажем, просят
повторно привести примеры животных, отмечается снижение активации тех
же областей.
В многочисленных исследованиях показано, что у больных с амнезией отме-
чаются нормальные результаты разнообразных тестов на прайминг. Это отно-
сится к большинству тестов на перцептивный и концептуальный прайминг, что
указывает на то, что медиальные височные доли не участвуют в их выполнении.
Рис. 9 .9 . Новая кора: моторная, премоторная и префронтальная области. Вид коры сбо-
ку: моторная область окрашена в фиолетовый цвет. Непосредственно перед ней распо-
лагается премоторная область, в которой кодируется когнитивное намерение выполнить
движение определенными конечностями. Истинная префронтальная кора находится
перед этими моторными областями (светло-зеленого цвета). Голубая область префрон-
тальной доли иногда называется орбитофронтальной корой, поскольку располагается
сразу над обеими глазничными впадинами. Читателю рекомендуется самому найти
другие структуры, которые видны на этом рисунке, в том числе основные доли коры,
сильвиеву борозду и центральную борозду, которая отделяет заднюю половину коры от
передней. (Источник: Sh��� Fu.)
496
Глава 9
2.3.2. Сохранение процедурной памяти при амнезии
Одна из первых демонстраций сохранения памяти при амнезии произошла при
исследовании обучения перцептивным двигательным навыкам, называемым
тестам на процедурную память. Коркин (C�r���, 1965) и Милнер (M�l�er, 1965)
показали, что HM мог обучаться и запоминать задание по преследованию дви-
HM мог обучаться и запоминать задание по преследованию дви-
мог обучаться и запоминать задание по преследованию дви-
жущейся цели, удерживая на ней указатель. Результаты обучения сохранялись у
НМ даже через несколько месяцев, хотя если его внимание отвлекалось, то уже
через несколько минут он не мог вспомнить, что он делал. Такие же результаты
были получены и в ряде других случаев.
Процедурная память зависит от перцептивно-двигательных областей, таких
как базальные ганглии, которые взаимодействуют как с задними, так и с лоб-
ными отделами новой коры (см. гл . 5). У больных с повреждением базальных
ганглиев, связанным с болезнью Паркинсона или Хантингтона, отмечается от-
сутствие улучшения или лишь незначительное улучшение результатов после
обучения сенсомоторным задачам (K�sz����, 1990; ���r�el� et al., 1994).
В серийной задаче на время реакции (SRT) на экране компьютера в одном
из четырех положений возникают точки. При этом участников исследования
инструктируют нажимать кнопку, соответствующую видимому расположению
точек (W�ll���h�m et al., 1989). Одни последовательности появления точек зако-
номерно повторяются, а другие изменяются случайным образом. В ходе тести-
рования реакция на повторяющиеся последовательности ускоряется, тогда как
реакция на случайные последовательности не меняется, даже если участники
не могут осознанно отличить одни последовательности от других (W�ll���h�m,
1998). Пациенты с амнезией хорошо справляются с имплицитным заданием на
время реакции и плохо — с его эксплицитной версией (Nissen, �ullemer, 1987;
Reber, Squ�re, 1998). И вновь пациенты с повреждениями базальных ганглиев,
как при болезни Паркинсона, плохо справляются с обеими задачами на время
реакции (Knopman, Nissen, 1987; V���l et al., 2000).
В функциональных нейровизуализационных исследованиях также показа-
но, что обучение имплицитной задаче на скорость реакции связано с активно-
стью в базальных ганглиях, а не в медиальных височных областях.
2.4 . Сохраненное имплицитное обучение
Академическое обучение обычно эксплицитно: преподаватели объясняют то,
что нужно выучить, а студенты по мере сил стараются это запомнить. Однако
обычное человеческое обучение в основном все же имплицитно. Первобытный
охотник наставлял молодежь, как выследить зверя, как убить его, как содрать с
него шкуру. Таким практическим навыкам гораздо легче обучиться путем мо-
делирования ситуации, чем эксплицитным запоминанием последовательности
действий. Множество тонкостей, связанных с охотой и собирательством, может
даже и названий-то не иметь! Если человек воспринимает какую-то последо-
вательность стимулов, основанную на простых закономерностях, то он эту за-
кономерность выявляет обычно неосознанно, опытным путем. Социальные на-
выки тоже приобретаются, видимо, по большей части имплицитно. Когда дети
учатся говорить, они ведь не опознают слышимые ими слова как существи-
тельные или глаголы. Они, прежде всего, обращают внимание на звуки речи,
а закономерности, лежащие в основе языка, постигаются ими имплицитно.
Обучение и память
497
Мы редко воспринимаем абстрактные вещи — грамматические правила, му-
зыкальную гармонию или утонченную живопись — на уровне сознания. Наши
знания — по большей части не выразить словами, наше обучение — по боль-
шей части имплицитно.
Ноултон с соавторами (K���lt�� et al., 1994, 1996) для исследования им-
плицитного обучения использовал задачу по вероятностному прогнозирова-
нию. Участникам показывали наборы из четырех карт, с помощью которых
с разной степенью вероятности осуществлялся «прогноз погоды» (рис. 9 .10).
После предъявления каждого набора карт участников опрашивали о прогнозе:
«дождь» или «солнечно». Одни наборы предсказывали «дождь» в 20% случаев,
а другие — в 80%. Обучение этой ассоциации у нормальных людей требует
примерно 50 попыток, но чтобы эксплицитно понять, какая последовательность
карт какой тип погоды прогнозирует, требуется значительно больше времени.
То есть предлагаемый принцип прогноза погоды уже постигается имплицитно
до того, как он будет, наконец, понят эксплицитно.
На ранних этапах обучения, когда формируется имплицитная ассоциация,
больные с амнезией выполняют это задание так же, как и контрольные испы-
туемые, но плохо справляются на поздних этапах, когда решение задачи осно-
вывается на эксплицитной, декларативной памяти. А у пациентов с болезнью
Паркинсона (дисфункцией базальных ганглиев), наоборот, отмечаются плохие
результаты в ранних испытаниях, но улучшение на более поздних. Нейрови-
зуализация (фМРТ) показывает активность в базальных ганглиях в течение
всего времени решения задачи, а изменение активности медиальной височной
доли — по ходу выполнения задания (P�l�r�c� et al., 1998).
Рис. 9 .10. Задача на имплицитное обучение «прогнозу погоды» с помощью последо-
вательности из четырех карт, разработанная Ноултоном с соавторами (K���lt�� et al.,
1994). Участники обучались прогнозировать вероятность результата имплицитно еще
до того, как могли сформулировать закономерность эксплицитно. Больные с амнезией
хорошо справлялись с заданием во время имплицитной ранней части процесса обуче-
ния, но так и не могли научиться определять связь между картами и «погодой» экспли-
цитно. (Источник: K���lt�� et al., 2003.)
498
Глава 9
Итак, амнезия, связанная с двусторонним повреждением медиальных височ-
ных долей, связана, по-видимому, главным образом с расстройством эпизоди-
ческой памяти, что является результатом нарушения передачи информации из
рабочей памяти в долговременную.
Поскольку воспоминания, приобретенные задолго до возникновения ам-
незии, остаются относительно сохранными, предполагается, что гиппокамп и
связанные с ним структуры медиальных отделов височной доли необходимы
только для временного удержания информации в памяти до консолидации ее в
других областях мозга, предположительно в новой коре.
Эпизодическая память
Выдающийся психолог Эндель Тулвинг (E��el Tul-
E��el Tul-
Tul-
Tul-
v���, 1985) описал эпизодическую память (способ-
ность запоминать прошедшее событие) как «мыс-
ленное путешествие во времени». Нейрофизиологам
потребовалось двадцать лет, чтобы постичь его идеи,
зато теперь мы понимаем, что эпизодическая память
позволяет нам повторно переживать прошлое, ориен-
тироваться в настоящем и строить планы на будущее
(Sch�cter et al., 2007). Прискорбно, что на память ока-
зывают значительное негативное воздействие много-
численные неврологические (например, болезнь
Альцгеймера, лобно-височная деменция, эпилепсия,
черепно-мозговые травмы и др.) и психиатрические
(например, шизофрения, депрессия, посттравмати-
ческие стрессовые расстройства и др.) заболевания.
Таким образом, понимание функциональной орга-
низации процессов памяти и их нервных субстратов
представляет исключительную важность как для уче-
ных, так и для общества в целом.
До недавнего времени большая часть того, что мы
знали о нервной основе памяти, была получена на
экспериментальных моделях и их сравнении с боль-
ными с амнезией. Таким образом была получена мас-
са информации о различных типах памяти и нейронных системах, которые их обе-
спечивают, однако отсутствовало понимание того, как же эти системы работают в
живом мозге человека. К счастью, когда я еще был студентом, исследователи стали
использовать нейровизуализационные (фМРТ) методы для идентификации нейрон-
ных коррелятов процессов памяти. Эти методы позволили разработать и исследо-
вать модели для объяснения не только самих визуализационных данных, но также и
результатов экспериментальных исследований на системном и молекулярном уров-
нях, и изучения функциональной природы процессов памяти человека.
Конечно, вопрос на 100% состоит в том, узнали ли мы что-либо новое изо всех этих
прелестных картинок? Я, безусловно, думаю, что да. С момента революционной
работы Бренды Милнер (�re��� M�l�er) насчет больного с амнезией по имени Ген-
�re��� M�l�er) насчет больного с амнезией по имени Ген-
M�l�er) насчет больного с амнезией по имени Ген-
M�l�er) насчет больного с амнезией по имени Ген-
) насчет больного с амнезией по имени Ген-
ри Молезон (He�ry M�l��s��, известный как «НМ»), стало ясно, что медиальные
отделы височной доли являются критическими для эпизодической памяти. Визу-
ализационные исследования расширили это представление, продемонстрировав,
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
Рис. 9 .11 . Харан Ранганат
(Ch�r�� R������th, PhD),
Калифорнийский универ-
ситет в Дейвисе, США.
Обучение и память
499
Рис. 9 .12 . Префронтальная область и медиальные отделы височной доли, участвую-
щие в формировании эпизодической памяти. (а) Относительное расположение дор-
солатеральной префронтальной коры (ДЛПФК, голубого цвета), вентролатеральной
префронтальной коры (ВЛПФК, розового цвета), периринальной коры (синего цве-
та), парагиппокампальной коры (зеленого цвета) и гиппокампа (красного цвета) по-
казано на изображении мозга с удаленным участком, чтобы обнажить медиальные
отделы височных долей. (б) Диаграмма, иллюстрирующая текущую модель участия
латеральной префронтальной и медиальной височной областей в формировании
эпизодической памяти. Анатомические связи между областями показаны черными
прямыми, а предполагаемые роли регионов приводятся курсивом. Для простоты на
диаграмме представлены только самые важные анатомические связи между этими
областями и не указаны другие связанные с ними области, которые также могут
играть значительную роль в формировании эпизодической памяти или извлечении
из нее информации. (Источник: R������th, в печати.)
500
Глава 9
что различные области медиальной височной коры обеспечивают формирование
памяти различными путями (рис. 9.12). К примеру, активация периринальной коры
связана с узнаванием знакомых объектов, в то время как активность в гиппокампе и
парагиппокампальной коре связана с припоминанием взаимосвязей между данны-
ми объектами и контекстом, в котором с ними столкнулись (D���� et al., 2007). Эти
результаты весьма схожи с данными, полученными при фокальных повреждениях
медиальных височных долей у крыс и обезьян, подтверждающими удивительный
параллелизм функций этих областей у разных видов (E�che���um et al., 2007). Ви-
зуализационные исследования также показали, что формирование эпизодической
памяти обеспечивается кортикальной сетью, простирающейся далеко за пределы
медиальных височных долей. К примеру, визуализационные данные выявили, что
префронтальная (R������th, �lume�fel�, 2008) и задняя теменная кора (Cabeza et
al., 2008) постоянно вовлекаются в кодирование и извлечение информации из памя-
ти, и роль этих областей только начинает осознаваться (рис. 9.12).
Все вышеприведенное — только намек на те потенциальные возможности, кото-
рые открываются для дальнейших достижений в этой области. Недавний прорыв
в области визуализации высокого разрешения в сочетании с методами анализа
визуализационных данных для обнаружения и классификации пространственных
паттернов приведут к значимым изменениям и усовершенствованиям теорий ней-
рокогнитивных процессов, тех самых, которые обеспечивают нашу поразительную
способность мысленно путешествовать во времени.
Литература
1. C��ez�, R., C��r�mell�, E., Ols��, I. R ., & M�s��v�tch, M. (2008). The p�r�et�l c�rtex
��� ep�s���c mem�ry: A� �tte�t����l �cc�u�t. Nature Review, Neuroscience, 9, 613–
619.
2. D����, R. A., Y��el���s, A. P., & R������th, C. (2007). Im����� rec�llect��� ���
f�m�l��r�ty �� the me���l temp�r�l l��e. A three-c�mp��e�t m��el. Trends in Cognitive
Sciences, 11 , 379–386 .
3. E�che���um, H., Y��el���s, A. P., & R������th, C. (2007). The me���l temp�r�l l��e
��� rec����t��� mem�ry. A��u�l Rev�e� �f Neur�sc�e�ces. 30, 123–152.
4. R������th, C., & �lume�fel�, R. S. (2008). Prefr��t�l c�rtex ��� mem�ry. I� J. H .
�yr�e (E�.), Learning and Memory: A Comprehensive Reference (pp. 261–279).
Oxf�r�: Ac��em�c Press.
5. Sch�cter, D. L., A���s, D. R., & �uc��er, R. L . (2007). Remem�er��� the p�st t�
�m����� the future: The pr�spect�ve �r���. Nature Reviews, Neuroscience, 8, 657–661 .
6. Tulv���, E. (1985). Elements of Episodic Memory. Ne� Y�r�: Oxf�r� U��vers�ty Press.
3.0 . Как формируются воспоминания
Традиционно память считается устойчивой записью события, которая может
быть восстановлена точно в такой же форме, в которой она была впервые за-
фиксирована. В соответствии с таким общепринятым подходом воспоминания
могут быть восстановлены, проверены и воспроизведены как высококачествен-
ная музыкальная запись. Воспоминания также могут быть забыты, что не за-
трагивает другие когнитивные системы.
Имеются, однако, определенные причины в этом усомниться. Одна из них со-
стоит в том, что реальные воспоминания о прошедших событиях редко бывают
точными. С динамической точки зрения, память рассматривается как продукт
динамического процесса: преобразования прошлого под влиянием прошлых и
текущих условий, предвосхищения будущего результата и других когнитивных
Обучение и память
501
процессов. С этой точки зрения память основана на хранении информации, но
не эквивалента ему. Она динамична и изменчива и взаимодействует с другими
процессами. Так, два человека, переживших одинаковое событие, могут иметь
различные воспоминания о нем. Это не значит, что один человек попросту прав,
а другой ошибается. Это значит, что при извлечении воспоминания у каждого
человека может быть свой взгляд, свое знание, своя мотивация и свойства памя-
ти, которые влияют на то, что извлекается.
Память каждого человека со временем изменяется. Большая часть того, что
произошло, забывается в течение нескольких минут или часов, а то, что оста-
ется, обычно реорганизуется и искажается другими знаниями или предубеж-
дениями. Однако мы не хотим, чтобы с компьютерными файлами или книгами
происходило то же самое. Мы не хотим, чтобы файлы со временем разрушались
или перемешивались с соседними файлами. Компьютеры и библиотеки созда-
ны для того, чтобы хранить всю информацию настолько четкой и неизменной,
насколько это возможно. Между тем нормальные воспоминания со временем
действительно тускнеют и часто смешиваются с другими.
Попытайтесь как можно подробнее вспомнить все, что вы делали в позапро-
шлые выходные, точно в том самом порядке, в котором вы все это проделали.
Для этого большинству из нас необходимо сначала вспомнить какие-то ключе-
вые моменты, а затем по ним уже точно восстановить, что происходило. После
этого запускается процесс восстановления последовательности событий. На-
пример, я встретил приятеля до или после разговора с родителями? Ходил ли
я по магазинам, и в каком порядке их посещал? В какой последовательности
в каждом из магазинов я выбирал и покупал товары? Можете проделать то же
самое, скажем, с недавно просмотренным фильмом и затем проверить точность
своей памяти, пересмотрев его.
Чтобы ответить на подобные вопросы, нужно много чего вспомнить, но
все это уж очень отличается от набора имени файла на клавиатуре для полу-
чения к нему доступа или указания шифра книги на библиотечной карточке,
чтобы книгу нашли в хранении. Можно нечаянно спутать то, что происходило
в позапрошлые выходные, с тем, что случилось в другое время. Как видно из
дальнейшего, у некоторых пациентов с мозговыми нарушениями наблюдает-
ся расстройство, называемое конфабуляция, при которой они рассказывают о
ложных воспоминаниях без какого-либо намерения соврать и без какого-либо
осознания того, что их воспоминания неверны.
Одни воспоминания влияют на то, как другие воспоминания организуются
и извлекаются из памяти. Они также формируют наши действия, даже когда мы
этого не осознаем. Наши воспоминания и склонности влияют на наши мысли и
действия и, в свою очередь, подвергаются влиянию с их стороны. Короче гово-
ря, память необходима, чтобы вести повседневную жизнь и строить планы на
будущее. При этом на нее, в свою очередь, оказывают влияние и прошлое (вне
зависимости от того, осознаем мы это или нет) и наши мысли о будущем.
3.1. Электрически вызванные
автобиографические воспоминания
Около 50 лет тому назад хирурги, оперирующие на головном мозге, сообщили,
что бодрствующие пациенты во время стимуляции височной доли описывают
яркие, специфические осознанные воспоминания. Пенфилд и Робертс (Pe��el�,
502
Глава 9
Roberts, 1959) были среди первых, кто использовал точечную электрическую
стимуляцию коры для составления карты функциональных областей. Их целью
было устранить так называемые «эпилептические очаги» в коре, в области руб-
цовой ткани, откуда запускается мощная «электрическая буря» при больших
припадках. Чтобы определить местонахождение эпилептических очагов и из-
бежать повреждения функционально важных областей, кора бодрствующих
пациентов картировалась с использованием низковольтной электростимуляции
специфических областей.
Это возможно только потому, что кора сама по себе не содержит болевых
рецепторов. Поскольку используется лишь местная анестезия для устранения
боли от разреза скальпа, бодрствующие пациенты могут безо всякого вреда для
себя рассказывать о своих ощущениях. Так что хирургия на открытом мозге —
уникальный источник данных для когнитивной нейронауки.
Стимуляция височной доли иногда приводит к внезапному наплыву осоз-
нанных воспоминаний. Например, недавно один больной во время стимуляции
мозга сообщил о следующих впечатлениях (M�r��rty et al., 2001).
1. При четырех расположениях электродов, указанных на рис. 9 .13, — просма-
тривал детские мультики «Флинтстоуны».
2. При других четырех расположениях электродов — прослушивал рок-группу
«Пинк Флойд».
3. Еще при двух расположениях — прослушивал бейсбольный репортаж.
4. Наконец, при четырех последних расположениях — слышал неизвестный
женский голос.
Обратите внимание, что незаштрихованные кружочки на рисунке показыва-
ют расположения электродов, при которых не отмечалось никаких воспомина-
ний вообще. Некоторые из расположений электродов находятся возле слуховой
коры в верхней части височной доли, однако другие — далеко от них. При этом
нет никаких данных о том, чтобы эти слуховые области коры были источником
таких насыщенных впечатлениями воспоминаний о каких-то вещах, которые,
как сообщают больные, они слышали в определенные моменты своей жизни.
Однако этому пациенту было выполнено хирургическое разрушение в меди-
альном отделе височной доли, так что какие-либо обобщения следует делать
с осторожностью.
Отчеты этого больного не могут быть выдумкой, поскольку он не знал, какой
электрод в данный момент стимулирует его височную долю. Ученые скептиче-
ски отнеслись к этим сообщениям, поскольку стимуляция в одном месте может
также активировать и другие области. Кроме того, для эпилептиков характерен
«атипичный» мозг, так как кора обычно изменяется в результате заболевания.
Тем не менее чрезвычайно маловероятно, что за пятьдесят лет истории стиму-
ляции височной доли так много различных больных сообщали бы о случайном
«наплыве» сознательных воспоминаний. После многочисленных эксперимен-
тов на животных, тщательных исследований больных с амнезией и нейровизу-
ализаций при тестировании памяти в настоящее время остается мало сомнений
в том, что височная доля заключает в себе нечто специфическое в отношении
долговременной эпизодической памяти. Одна из приемлемых гипотез состоит в
том, что стимуляция височной доли захватывает ее медиальную часть и каким-
то образом активирует специфические воспоминания.
Обучение и память
503
На рис. 9.14 приводится возможное объяснение этого феномена. То, что мы
наблюдаем у нейрохирургического больного, является стабильным памятным
следом, который вызывается путем непосредственной электрической стиму-
ляции височной доли. Таким образом, поток информации идет из новой коры
(височной) в гиппокампальную систему (медиальную височную кору) и вводит
гиппокампальную систему в резонанс с исходными следами памяти в новой
коре для воссоздания переживания определенного жизненного события или
чего-то очень похожего.
Тем не менее много вопросов все еще остается без ответа, например: какова
связь между электрическим стимулом и нормальным процессом извлечения
эпизодического воспоминания? Почему среди многих миллионов эпизодов,
из которых состоит жизнь обычного человека, в этих обстоятельствах возни-
кают такие весьма специфические воспоминания? Существует ли возможная
Рис. 9 .13 . Автобиографические воспоминания, вызванные стимуляцией височной
доли. Спонтанные сообщения о воспоминании пережитых событий, вызванных элек-
трической стимуляцией мозга у больных с хирургическими разрушениями в медиаль-
ных отделах левой височной доли. Обратите внимание на электродную сетку, которая
была помещена на кору мозга таким образом, как показано на рисунке. Электроды раз-
мещались с интервалом 1 см. Различные электроды всякий раз вызывали воспоминания
разных жизненных эпизодов. Спонтанные отчеты такого рода нередки при стимуляции
височной доли, однако при стимуляциях коры мозга другой локализации таких сообще-
ний обычно не отмечается. (Источник: M�r��rty et al., 2001.)
504
Глава 9
связь между электрическим раздражителем и осцилляторными ЭЭГ феноме-
нами, наблюдаемыми при извлечении информации, например тета-ритмами,
которые, похоже, координируют активность медиальной височной и новой
коры при извлечении информации (см. далее)? Эпизодические воспоминания,
о которых сообщали нейрохирургические больные после электрической сти-
муляции, — поразительные и, видимо, стойкие и хорошо воспроизводимые
феномены. Однако до сих пор нет четкого удовлетворительного объяснения
данного явления.
3.2 . Длительная потенциация и длительная депрессия:
возбуждающие и тормозные следы памяти
Большинство синапсов в коре являются возбуждающими, использующими ней-
ромедиатор глутамат (см. гл. 3). Небольшая их часть использует тормозные
нейромедиаторы, такие как ГАМК (гамма-аминомасляная кислота). Для коди-
ровки следов долговременной памяти путем изменения синаптической прово-
димости эти возбуждающие и тормозные связи должны каким-то образом стать
более стабильными. Эти процессы, как предполагается, происходят за счет так
называемой долговременной потенциации (ДВП) возбуждающих синапсов и
долговременной депрессии (ДВД) тормозных синапсов. Эти явления, которые
наблюдаются в специфических областях, являются просто повышением и сни-
жением вероятности разряда постсинаптического потенциала в ответ на пре-
синаптический спайк.
Явление ДВП было обнаружено в самом гиппокампе с использованием за-
писи одиночной активности одного из нейронных слоев (рис. 9 .15). Запись
активности одиночных клеток широко применяются в исследованиях на жи-
вотных, однако такие записи были сделаны также и у нескольких пациентов
с эпилепсией (Kreiman et al., 2002).
Рис. 9 .14 . Гиппокампальная система (медиальная височная кора) и новая кора при об-
учении и воспроизведении воспоминания. Модель нейронной сети взаимодействия
медиальной височной коры (гиппокампальной системы) с новой корой. Во время ко-
дирования или процесса обучения (а) информация из коры передается в гиппокампаль-
ную систему. Во время извлечения воспоминания из памяти (б) и (в) неокортикальная
импульсация служит активатором перекрывающегося рисунка нейрональной активации
в медиальной височной коре (голубые точки). Гиппокампальная система отвечает акти-
вацией областей новой коры, которые обеспечивают ощущение восстановление некото-
рой части исходного события. (Источник: �luc� et al., 2003.)
Обучение и память
505
Хотя ДВП и ДВД наблюдаются в специфических областях мозга, таких как
гиппокамп, стандартная гипотеза о долговременной памяти подразумевает уча-
стие миллиардов и даже триллионов синапсов в коре и окружающих тканях. До
настоящего времени нет никакого способа учесть все синапсы в этой системе
или хотя бы существенную их часть.
Скорее, имеется ряд экспериментальных данных, демонстрирующих усиле-
ние ДВП и ДВД (рис. 9 .16), дополненных результатами исследований больных
с разрушениями головного мозга и суммарной активности популяций милли-
ардов нейронов, измеренной при помощи ЭЭГ, ВП (вызванные потенциалы),
фМРТ и т. д . Кроме того, имеются подтверждения, полученные в исследова-
Рис. 9 .15. Запись активности одиночных нейронов гиппокампа. Схематическое изо-
бражение записи активности одной клетки при стимуляции изнутри или снаружи гип-
покампальных нейронов. Электрод, помещенный снаружи нейрона, может вызвать
потенциалы электрического поля, которые сходны с ЭЭГ, однако значительно более ло-
кализованы. Потенциалы электрического поля часто отражают активность небольшой
популяции нейронов, в отличие от аксонального или дендритного потенциала одиноч-
ного нейрона. Следует отметить, что очень схожая методика может быть использована
для стимуляции как отдельных клеток, так и небольших групп клеток в гиппокампе.
(Источник: Squ�re et al., 2003.)
506
Глава 9
ниях со стимуляцией — стимуляция височной доли бодрствующих больных во
время нейрохирургических операций и транскраниальной магнитной стимуля-
ции (ТМС) у здоровых испытуемых. Следовательно, то, что известно о памяти,
является, скорее, логическим обобщением, основанным на результатах многих
сотен проведенных исследований. Однако наблюдать непосредственно на уль-
трамикроскопическом уровне множество изменяющихся синаптических соеди-
нений пока не представляется возможным.
Тем не менее в настоящее время достигнуто согласие в отношении того, что
существующие данные согласуются рядом утверждений.
Рис. 9 .16 . Долговременная потенциация в гиппокампе. Следы памяти, как считается,
кодируются в новой и медиальной височной коре путем изменения синаптической про-
водимости между миллиардами нейронов. Эти процессы называются долговременной
потенциацией (ДВП), соответствующей длительному повышению возбуждающей ней-
ДВП), соответствующей длительному повышению возбуждающей ней-
), соответствующей длительному повышению возбуждающей ней-
ропередачи, и долговременной депрессией (ДВД), что соответствует длительному по-
вышению тормозной нейропередачи. Прямое доказательство существования ДВП было
получено при записи активности одиночных клеток гиппокампа, как показано выше. На
нижней части диаграммы приводится три примера изменения ВПСП (возбуждающих
постсинаптических потенциалов) после сильной электрической пресинаптической сти-
муляции (называемой тетанусом). Видно, что через 90 мин ВПСП еще более снижается
(напомним, что более отрицательный потенциал соответствует большей электрической
спайковой активности). Верхний график показывает долгосрочное изменение проводи-
мости синапса, измеренное по углу наклону кривой ВПСП в течение 2 ч после сильного
электрического стимула (тетануса). (Источник: �yr�e in Squ�re et al., 2003.)
Обучение и память
507
1. Эпизодическая афферентная информация первоначально обрабатывается
новой корой.
2. Информация для формирования памяти интегрируется в медиальной височ-
ной коре, содержащей гиппокампы и связанные с ними структуры, а, воз-
можно, также и в таламусе и окружающих областях.
3. Консолидация: затем медиальная височная кора и соседние регионы соеди-
няют и объединяют ряд областей новой коры — процесс, который превра-
щает кратковременные синаптические связи в долговременные следы памя-
ти, хранящиеся как в медиальной височной, так и в новой коре. Считается,
что основным механизмом таких превращений являются ДВП и ДВД.
Последний пункт подробно обсуждается ниже.
3.3 . Консолидация: от временного хранения до постоянного
Дополним картину последними штрихами. В гл. 2 уже излагалась общеприня-
тая гипотеза о взаимосвязи между непосредственной памятью и долговремен-
ной памятью, называемая гипотезой консолидации.
Консолидация обычно определяется как нарастающая стабилизация следов
долговременной памяти, обеспечивающая их относительную устойчивость к
разрушению или распаду. Это тот самый процесс, который отсутствует или
сильно нарушен у пациентов с амнезией, и который объясняет слабую способ-
ность таких пациентов переносить информацию из кратковременной памяти
в долговременную.
На рис. 9 .17 приводится один из вариантов обучения с консолидацией, при
котором входной сигнал вызывает активацию новой коры и гиппокампальных
областей (медиальная височная кора) с образованием новых синаптических
связей между нейронами. Как указывалось выше, непосредственная память ко-
дируется путем усиления синаптической связи между миллиардами нейронов
в новой коре. Нормальный сон, особенно медленноволновая фаза, важен для
перевода этих временных связей в долговечные следы памяти.
Предполагается, однако, что более долговременные следы памяти требуют
синтеза белка, необходимого, например, для роста дендритных шипиков —
крошечных выростов, которые развиваются на верхушке дендритов, порождая
новые синаптические связи между соседними нейронами.
Идея о том, что обучение требует некоторого времени для «закрепления»,
является далеко не новой. Еще в 1904 г. Бёрнхэм (�ur�h�m) писал:
Закрепление впечатлений в памяти зависит от физиологических процессов. Для
того, чтобы данное впечатление достаточно прочно закрепилось, чтобы его мож-
но было воспроизвести даже через длительный интервал, требуется время; а для
того, чтобы данное впечатление закрепилось насовсем, стало частью постоянного
хранилища памяти, может потребоваться очень значительное время. Можно пола-
гать, что эти физиологические процессы — не просто формирование постоянного
запечатления на нервных клетках, но и ассоциация, совместная организация новых
впечатлений со старыми (цит. по: M�sc�v�tch).
На рис. 9.18 приводится принципиальная схема этих двух видов консолида-
ции. Процесс ДВП-ДВД, который обсуждался выше, включает клеточную кон-
солидацию, локальное изменение проводимости в триллионах синапсов. Счи-
тается, однако, что существует также и системная консолидация, при которой
508
Глава 9
может происходить широкомасштабная реорганизация памяти. Имеются весо-
мые подтверждения того, что различные стадии сна могут оказывать различные
эффекты на этот процесс системной консолидации. Обратите внимание, что оба
типа консолидации предполагают активный диалог между медиальной темен-
ной (гиппокампом) и новой корой.
3.4 . Быстрая консолидация: синаптические механизмы,
транскрипция генов и синтез белка
Быстрая или синаптическая консолидация завершается в пределах первых ми-
нут или часов после того, как происходит обучение. Вейлер и коллеги (Weiler
et al., 1995) показали, что она коррелирует с морфологическими изменениями в
самом синапсе. Предъявление стимула запускает каскад нейрохимических со-
бытий на синаптической мембране и внутри клетки, которые повышают си-
наптическую эффективность или проводимость, с помощью которой нейроны,
формирующие след памяти, могут поддерживать связь друг с другом. Первый
из этих процессов включает локальные, кратковременные молекулярные изме-
нения, которые приводят к повышению высвобождения нейромедиатора в за-
тронутом синапсе.
Рис. 9 .17. Этапы обучения, связывания, консолидации и извлечения памяти. На этой
схеме 1-й этап, обучение, состоит из трех элементов (А, В и С). Информация исходно
кодируется в новой коре (например, в зрительной) и затем направляется в медиальную
височную (MTL). На 2-м этапе медиальная височная и новая кора входят в резонанс
друг с другом, благодаря чему начинается формирование следа памяти. На 3-м этапе
прежнего стимула уже нет, и резонанс между медиальной височной и новой корой те-
перь не зависит от внешней поддержки. Этап 4 показывает, каким образом консолида-
ция приводит к долговременным раздельным следам памяти (синаптическим изменени-
ям) в медиальной височной и новой коре, которые теперь сосуществуют отдельно друг
от друга, в то время как обрабатывается другой входной сигнал. На 5-м этапе в качестве
напоминания или сигнала к восстановлению события в памяти показан один из элемен-
тов (элемент В) исходного события (А-В -С). На 6-м этапе следы памяти А-В-С активи-
руются резонансной активностью между медиальной височной и новой корой. На этом
этапе эпизодическое воспоминание извлекается уже в отсутствие исходного стимула.
(Источник: M�rr�s M�sc�v�tch, изменено с разрешения.)
Обучение и память
509
Рис. 9 .18 . Реконсолидация переводит активные нейрональные связи в более длитель-
ные. Считается, что существует два вида консолидации — клеточная и системная. Обе
вызываются активацией медиальной височной и новой коры. Диаграмма подчеркива-
ет участие медиальной височной (также называемой гиппокампальным комплексом) и
новой коры в формировании активных клеточных ансамблей, соответствующих аффе-
рентному обучению, в которых нейроны резонируют друг с другом до тех пор, пока
между ними не установятся более постоянные связи. (Источник: N��er, 2003.)
510
Глава 9
Если стимул достаточно интенсивный и/или повторяющийся, то активиру-
ются дополнительные процессы. Они включают транскрипцию гена и образо-
вание белка, что приводит к долговременным клеточным изменениям, включая
создание новых синапсов, которые обеспечивают формирование и поддержа-
ние долговременной памяти. Эти процессы могут длиться от нескольких часов
до нескольких дней (рис. 9 .19) (Lees et al., 2000; Mc��u�h, 2000; Du���, 2004).
Хотя наука уверенно двигается к пониманию основных клеточных и моле-
кулярных механизмов синаптической консолидации, мы все еще далеки от по-
нимания продолжительной или системной консолидации, которая бурно обсуж-
дается в литературе.
3.5 . Системная консолидация: взаимодействие между
медиальными отделами височных долей и новой корой
Завершение системной консолидации может занять значительно больше вре-
мени и может варьировать от нескольких дней до нескольких лет или десяти-
летий. У больных с повреждениями медиальной височной коры отмечается
ретроградная потеря памяти, которая имеет временную градацию — потеря
памяти на недавнее событие (до амнезии) выражена в большей степени, чем
потеря памяти на более ранние события. Этот временной градиент затрагивает
только эксплицитную память, в то время как имплицитная память остается со-
хранной и стабильной со временем (Sc�v�lle ��� M�l�er, 1957).
Эти наблюдения указывают на то, что медиальная височная кора формирует
кратковременный след памяти, необходимый для эксплицитных воспоминаний
до того момента, как они консолидируются где-то в другой области мозга, пред-
положительно в новой коре (Squ�re, 1992; Squ�re, Alv�rez, 1995). Эта стандарт-
Squ�re, 1992; Squ�re, Alv�rez, 1995). Эта стандарт-
, 1992; Squ�re, Alv�rez, 1995). Эта стандарт-
Squ�re, Alv�rez, 1995). Эта стандарт-
, Alv�rez, 1995). Эта стандарт-
Alv�rez, 1995). Эта стандарт-
, 1995). Эта стандарт-
Рис. 9 .19 . Динамика консолидации. Макго (Mc��u�h, 2000) указывает на то, что име-
Mc��u�h, 2000) указывает на то, что име-
, 2000) указывает на то, что име-
ется три частично перекрывающихся периода действия консолидации. Краткосрочный
относится к «кратковременной памяти» и длится от нескольких секунд до нескольких
часов. Консолидация долговременной памяти совершается на протяжении периода от
нескольких часов до нескольких месяцев. Кроме того, для объяснения некоторых фак-
тов, таких как сохранение давних воспоминаний о ранних жизненных событиях у па-
циентов с амнезией, у которых отсутствует память на события как за на несколько лет
до повреждения мозга, так и после него, необходимо еще ввести понятие долговечной
памяти. (Источник: Mc��u�h, 2000.)
Обучение и память
511
ная модель консолидации не делает никакого различия между отдельными ти-
пами эксплицитной памяти, например, эпизодической и семантической.
Нэйдел и Москович вопреки стандартной модели консолидации пришли к
выводу, что для того, чтобы вызвать в памяти даже старые эпизодические вос-
поминания, необходимо участие медиальной височной коры все то время, пока
воспоминание существует (N��el, M�sc�v�tch, 1997, 1998; M�sc�v�tch, N��el,
1998; N��el et al., 2000). Чтобы вызвать в памяти повторные впечатления, свя-
занные со словами, объектами, людьми и окружающей средой, достаточно уча-
стия только новой коры.
Медиальная височная кора может участвовать в исходном формировании
этих следов в новой коре, однако, раз сформировавшись, они могут существо-
вать сами по себе. Таким образом, уникальные автобиографические воспоми-
нания отличаются от повторных воспоминаний тем, что последние продолжа-
ют нуждаться в участии медиальной височной коры. Повторные воспоминания,
как предполагается, формируют множественные следы, добавляя к ним все но-
вые и новые всякий раз, когда данное событие припоминается.
Результаты нейровизуализационных исследований подтверждают данное
объяснение (блок 9.2). В этих исследованиях обнаружено, что гиппокамп акти-
вируется одинаковым образом во время извлечения недавних и отдаленных ав-
тобиографических воспоминаний (C����y et al., 1999; Ry�� et al., 2000; ��l���
et al., 2004; см. также обзоры: M��u�re, 2000; M�sc�v�tch et al., 2005, 2006). Эти
вопросы продолжают обсуждаться в литературе.
Блок 9.2. Теория множественных следов против традиционной консолидации
Нэйдел и Москович (N��el, M�sc�v�tch, 1997) предложили теорию множественных
следов, согласно которой гиппокампальный комплекс быстро кодирует всю инфор-
мацию, и та становится осознанной. Медиальная височная кора связывает нейроны
новой коры, которые реализуют перевод сознательного опыта в след памяти. Ней-
роны медиальной височной коры действуют как указатель, или индекс, для ансамб-
ля нейронов новой коры, которые образуют представительство этого опыта (Teyler,
D�Sce���, 1986). Поэтому след памяти о каком-либо событии состоит из связанно-
, 1986). Поэтому след памяти о каком-либо событии состоит из связанно-
го ансамбля нейронов и новой, и медиальной височной коры. Формирование таких
следов относительно быстрое — на протяжении секунд или, самое большее, дней
(M�sc�v�tch, 1995).
В этой модели нет места процессу длительной консолидации, который медленно
укрепляет след памяти в новой коре. Вместо этого каждый раз при извлечении из
памяти старого воспоминания создается новый след, опосредованный гиппокампом.
В результате старые воспоминания отображаются большим количеством следов, чем
новые, и поэтому менее подвержены разрушению. Поскольку след памяти распреде-
лен в медиальной височной коре, степень и тяжесть ретроградной амнезии связана с
объемом и локализацией ее повреждения.
В то время как след каждого автобиографического воспоминания уникален, суще-
ствование многих связанных следов облегчает извлечение информации из памяти.
Эпизодические воспоминания объединяются в форме семантических. Таким об-
разом, воспоминания о людях и событиях, связанных с какими-то определенными
эпизодами, становятся отделенными от своих первоисточников. Этот процесс может
создать видимость классической консолидации, однако реальный механизм мозга от-
личается от классической теории.
512
Глава 9
4.0 . Разновидности памяти
Память — не единичное понятие. У Клайва Уэринга (Cl�ve We�r���) память
на отдельные прошлые события его жизни практически полностью разрушена,
однако он может удерживать в памяти информацию о непосредственном про-
шлом в течение примерно 7 с (если его внимание не отвлекается). Ясно, что
одни формы памяти у него были затронуты заболеванием, а другие — нет. Так
обычно и бывает у больных с амнезией, что подтверждает различную нейро-
нальную основу разных типов памяти. Стандартное представление об основ-
ных системах долговременной памяти приводится на рис. 9 .20. Однако точная
взаимосвязь между различными типами памяти все еще остается предметом
дискуссий. Например, перцептивную память относят к недекларативной, но та-
кая классификация скорее произвольна. Другой пример: перцептивная память
проявляется улучшением сенсорного различения на уровне коры мозга, часто
путем реорганизации кортикальных рецептивных полей. Результатом этого
процесса является изменение осознанного восприятия, как, например, при об-
учении игре на гитаре, когда ученик все лучше и лучше распознает отдельные
звуки. Это во многом похоже на эпизодическую память, которая также может
быть в большой степени перцептивной и которую принято считать осознанной.
Тем не менее широко используется классификация памяти, представленная на
рис. 9 .20, и именно такой схеме обучают студентов.
4.1. Эпизодическая и семантическая память:
«припоминание» против «знания»
Как показано на рис. 9 .20, декларативная память может быть разделена на два
типа, эпизодическую и семантическую (Tulv���, 1972). Эпизодическая память
относится к воспоминаниям, которые имеют специфический источник во вре-
мени и пространстве и произошли при определенных жизненных обстоятель-
ствах. Эпизодические воспоминания часто автобиографичны по своей природе,
что позволяет мысленно путешествовать в прошлое, освежая впечатления.
В отличие от этого семантические воспоминания включают факты об окру-
жающем мире, о нас самих и о других общепризнанных вещах. Семантические
воспоминания независимы от пространственных и временных обстоятельств,
в которых они были приобретены. К семантической памяти могут относиться,
скажем, знания о том, что Париж — столица Франции, а Оттава — Канады, или
о том, в какой школе мы учились. В отличие от них к эпизодической памяти мо-
жет относиться какое-то событие, случившееся в Париже, Оттаве или средней
школе (рис. 9.21). Как правило, имеется некий автобиографический источник
воспоминания о специфическом времени, месте и стечении обстоятельств, в ко-
торых это эпизодическое воспоминание было получено. Термины «эпизодиче-
ская» и «автобиографическая» память являются практически синонимами.
Эпизодические воспоминания, как правило:
• имеют отношение к самому себе;
• формируются вокруг определенного периода времени;
• запоминаются осознанно, так, чтобы мы, по-видимому, могли повторно
их пережить;
Обучение и память
513
• подвержены забыванию;
• зависят от обстоятельств времени и места, взаимосвязи с другими вос-
поминаниями и прочего.
Рис. 9 .20 . Классификация типов памяти, предложенная Шактером и Тулвингом (Sch�ct-
Sch�ct-
er, Tulv���). Декларативная память изучена очень подробно и, как считают, является
эксплицитной (сознательной). Недекларативные типы памяти являются бессознатель-
ными или имплицитными, однако это утверждение до сих пор находится под вопросом.
Несмотря на то что, в соответствии с этой диаграммой, недекларативные типы памяти
имеют, несомненно, бессознательные аспекты, до сих пор неясно, может ли такого рода
обучение происходить без осознания афферентного притока. (Источник: Squ�re, 2004.)
Рис. 9.21 . Припоминание: автобиографические эпизоды. Припоминание включает ак-
тивное воссоздание исходного (осознанного) эпизода. Эти сознательные воссоздания
событий, по-видимому, требуют участия гиппокампа. (Источник: M�rr�s M�sc�v�tch,
с разрешения.)
514
Глава 9
Напротив, семантические воспоминания (рис. 9 .22) обычно:
• имеют отношение к общему с другими людьми знанию;
• не относятся к определенному периоду времени;
• дают скорее «ощущение знания», чем полностью сознательного воссоз-
дания исходного события;
• менее подвержены забывания, чем специфические жизненные эпизоды;
• относительно независимы от обстоятельств.
Чтобы изучить типы сознания в тестах на память, Тулвинг (Tulv���, 1985)
ввел процедуру «вспомнить/знать». Она представляет собой следующее: участ-
ников просят провести интроспективный самоанализ при опознании предла-
гаемых им для изучения объектов. Если они полагают, что данный объект им
уже знаком, то они должны решить, помнят они его (т. е . восстановить детали
обстоятельств, при которых они с ним познакомились) или знают (он кажет-
ся хорошо известным) (см. рис. 9.23). Локальная активация гиппокампа, по-
видимому, влияет только на суждение «вспомнить». Память, основанная на
ощущении хорошо известного, обходится без его участия (M�sc�v�tch, McA�-
M�sc�v�tch, McA�-
, McA�-
McA�-
�re�s, 2002; Y��el���s, 2002). Подобным же образом в функциональных ней-
, 2002; Y��el���s, 2002). Подобным же образом в функциональных ней-
Y��el���s, 2002). Подобным же образом в функциональных ней-
, 2002). Подобным же образом в функциональных ней-
ровизуализационных исследованиях активация гиппокампа ассоциируется в
большей степени с припоминанием, чем со знанием (El�r���e et al., 2002; Y��e-
Yone-
linas et al., 2005).
Таким образом, след памяти, опосредованный гиппокампом, заполнен тем
сознанием, которое сопровождало исходный опыт, или тем, что Тулвинг назвал
автоноэтическим сознанием. Это в корне отличается от простого ощущения
знания о чем-то, к которому Тулвинг относит ноэтическое сознание. Ноэтиче-
ское сознание ассоциируется с семантической памятью.
Рис. 9 .22 . Знание: семантическая информация. Семантическая память оценивается по
очень точному ощущению знания чего-либо. Однако она не требует активного воссоз-
дания исходного эпизода и может быть, очевидно, реализована новой корой без помощи
гиппокампального комплекса. (Источник: M�rr�s M�sc�v�tch, с разрешения.)
Обучение и память
515
4.2 . Эпизодические воспоминания могут со временем
превращаться в семантические
В 1958 г. Пенфилд и Милнер (Pe��el� и M�l�er) писали:
Запись потока сознания ... зависит от целостности гиппокампальных структур с
обеих сторон. ... В дальнейшем человек имеет дело уже с тем, что можно назвать
«обобщениями», и он может затребовать их для своих целей. Все жизненные собы-
тия, даже так называемые «незабываемые», постепенно ускользают от возможности
произвольного извлечения, если о них не говорить или не вспоминать их, «прокручи-
вая» рефлексивно. Например, если человек не раз слышал какую-то песню или стих,
он хорошо помнит, о чем там говорится, но быстро забывает обстоятельства, при
которых он это слышал или читал.
Рис. 9.23 . «Потенциалы, связанные с событием» (ПСС) при припоминании по сравне-
нию со знанием. Акт припоминания (извлечения из памяти чего-либо) сопровождается
значительно большей активацией мозга, чем «ощущение знания чего-либо», даже того
же самого. На этих изображениях мозга приводятся так называемые «потенциалы, свя-
занные с событием» (ПСС), записанные, как ЭЭГ, с поверхности кожи головы, и усред-
ненные. Эти результаты согласуются с данными (см. гл . 8), которые указывают на то,
что осознанные стимулы вызывают форвардную активацию, постепенно распространя-
ющуюся за пределы сенсорных областей коры, в то время как неосознанные стимулы
вызывают сугубо локальную активацию. Сознательно извлекаемый материал, видимо,
в большей степени участвует в восстановлении в памяти исходных впечатлений, что
также требует больших умственных усилий, вызывающих высокую активность в пре-
фронтальной коре (см. гл . 10, 11 и 12). (Источник: Duzel et al., 1997.)
516
Глава 9
Более пятидесяти лет назад многие ученые, занимающиеся памятью, согла-
сились бы с этим. Имеется достаточные доказательства того, что семантические
воспоминания могут формироваться из повторяющихся, сходных событий. На-
пример, школьные годы представляют собой, по сути, длинную череду эпи-
зодов. Разумеется, можно припомнить десятки конкретных эпизодов, однако
большая часть школьных лет кажется какой-то «смазанной», оставившей в па-
мяти лишь общее семантическое убеждение, что «я учился в такой-то школе».
На рис. 9 .24 демонстрируется, каким образом эпизодические и семантиче-
ские воспоминания могут быть связаны в мозге. На представленном ниже шу-
точном рисунке приводятся отдельные эпизодические воспоминания: человек,
готовящий на гриле барбекю, дарящий цветы девушке, рисующий картину и
играющий в гольф.
Это отдельные автобиографические воспоминания, запомнившиеся как
осознанные события. На представленной выше небольшой семантической сети
все эти очень специфические и весьма подробные эпизоды объединены в одном
рисунке семантической сети человека, который готовит барбекю, любит, рисует
Блок 9.3 . Ложные ощущения «знания чего-то»: AKP, человек с «дежа вю»
(ощущением «уже пережитого»)
Иногда возникает ложное ощущение, что вы что-то уже знаете, хотя на самом деле с
этим сталкиваетесь впервые. Больной AKP — 80 -летний польский иммигрант в Ан-
AKP — 80 -летний польский иммигрант в Ан-
—
80-летний польский иммигрант в Ан-
глии, бывший дипломированный инженер. Обращение к его семейному врачу было
вызвано проблемами с памятью и жалобами на частые ощущения того, что его жена
описывала как «дежа вю» (ощущение того, что это уже переживалось). Ощущение
«дежа вю» было таким сильным, что мешало повседневной жизни AKP. Он пере-
AKP. Он пере-
. Он пере-
стал читать газеты или смотреть телевизор, поскольку говорил, что уже это видел.
Ощущение «дежа вю» было особенно сильным, когда он ходил гулять, — АКР жа-
ловался, к примеру, что видел всегда одну и ту же птичку на одном и том же дереве,
выводившую одни и те же трели. Он также обращал внимание на номера машин
и рассуждал о том, как строго водители соблюдают расписание, если каждый день
проезжают мимо него точно в одно и то же время. В магазине AKP заявлял, что не
нужно покупать то или это, поскольку он все это уже купил накануне. Были и другие
проявления конфабуляции, в частности он уверял, что был женат три раза на одной и
той же женщине, и у них были три свадьбы в разных странах Европы.
При формальном обследовании было обнаружено, что I� у АКР выше среднего, при-
I� у АКР выше среднего, при-
у АКР выше среднего, при-
чем в некоторых подтестах — выше 90. У него отмечались плохие результаты лишь
в отдельных, но не во всех, тестах на функцию лобной доли. Баллы в стандартных
тестах на память, однако, у него были сниженными, как и баллы в лабораторных
тестах. Он ошибался, например (как и следовало ожидать, исходя из его состояния),
в тесте, где нужно правильно разложить приманки по ловушкам. Как и в реальной
жизни, при тестировании он не только утверждал, что все новое ему уже знакомо, но
и в самом деле верил, что уже с этим сталкивался.
При структурных нейровизуализационных исследованиях (КТ, МРТ) у него выявля-
лась атрофия медиальных отделов височных долей, однако лобные доли выглядели
нормальными. Измерение метаболических показателей также выявило только нару-
шения в медиальных отделах височных долей. Возможно, что в силу этой аномалии
медиальные височные области постоянно генерирует ложные сигналы «памяти». По-
скольку эти сигналы исходят из тех же медиальных височных структур, что и «на-
стоящие», они и воспринимаются мозгом как истинные.
Обучение и память
517
и играет в гольф. Эта семантическая сеть более абстрактная и обобщенная, чем
отдельные эпизоды в жизни человека. Москович (M�sc�v�tch, 2004) полагает,
что нижний ряд эпизодов зависит от медиальной височной доли, а верхний ри-
сунок зависит от элементов новой коры.
Мы можем обобщить эту концепцию, выделив в ней три этапа.
1. Исходно воспоминания являются эпизодическими и зависят от контекста.
2. Со временем эпизодические воспоминания превращаются в семантические.
3. Медиальная височная кора важна для восстановления в памяти эпизодиче-
ских воспоминаний, которые связаны с определенным автобиографическим
контекстом, в котором возникли (блок 9.3).
Согласно этому взгляду, гиппокампальный комплекс (или медиальная ви-
сочная кора) обеспечивает:
• хранение и извлечение подробных, отдаленных автобиографических и
пространственных воспоминаний;
• формирование и усвоение семантической памяти в новой коре.
4.3 . Эпизодическая память и семантическая память
часто комбинируются
Когда вас спрашивают, знакомо ли вам чье-то лицо, скажем, кинозвезды Брэда
Питта, то на чем строится ваш ответ? На семантической, эпизодической памяти
или на той и другой? Вестмакотт и соавторы (Westm�c�tt et al. 2004) показали,
Рис. 9 .24 . Вот каким образом связаны семантические и эпизодические воспоминания:
семантические воспоминания могут быть неокортикальным «остатком» многих эпизо-
дических воспоминаний. Так, к примеру, в памяти содержится много всяких событий,
связанных с каким-то человеком, как это показано на шуточном изображении — как он
готовил барбекю, дарил девушке цветы, рисовал, играл в гольф. Со временем эти эпи-
зоды могут забыться, и остается только семантическое знание, что этот человек все это
делал. Эти следы памяти множественны, они воссоздавались заново всякий раз, когда
мы видели этого человека. Семантическая память требует участия только новой коры
(в особенности височной и лобной долей). Эпизодическая же информация требует уча-
стия как новой коры, так и гиппокампального комплекса. (Источник: M�sc�v�tch, 2004,
с изменениями, с разрешения.)
518
Глава 9
что участвуют оба вида памяти, поскольку результаты семантических тестов
лучше, если у испытуемого имеется также и некоторая эпизодическая память,
связанная с известным именем.
У пациентов с болезнью Альцгеймера наблюдается обширное нарушение
понимания смысла, часто с сохранными перцептивными способностями, на-
зываемое семантической деменцией. Эти больные, у которых дегенерация
передних и латеральных отделов височных долей приводит к семантическим
нарушениям, могут опознавать обычные объекты только в том случае, если эти
объекты как-то с ними связаны. Например, они могут опознать вазу, если она
их собственная, но не могут опознать любую другую вазу (S����e� et al., 1996;
�r�h�m et al., 1999).
Тем не менее удивительно, что если медиальная височная кора повреждается
в раннем детстве, то в большинстве случаев семантическая память развивается
относительно нормально (V�r�h�-Kh��em, M�sh���, 1997). Однако эпизодиче-
V�r�h�-Kh��em, M�sh���, 1997). Однако эпизодиче-
-Kh��em, M�sh���, 1997). Однако эпизодиче-
Kh��em, M�sh���, 1997). Однако эпизодиче-
, M�sh���, 1997). Однако эпизодиче-
M�sh���, 1997). Однако эпизодиче-
, 1997). Однако эпизодиче-
ская память без медиальной височной коры все же остается нарушенной. От-
сюда следует, что в детстве семантическая система может приобретать знания
сама, без помощи эпизодической системы (медиальной височной коры).
5.0 . Медиальная височная кора
в эксплицитном научении и памяти
Медиальная височная кора необходима для сознательного сохранения долго-
временных, эпизодических воспоминаний. У нас нет способа произвольно
«включать» ее. Все, что можно сделать, это сконцентрироваться на запоминае-
мом материале. Как подчеркивалось в гл. 8, в большинстве случаев это значит,
что, когда мы начинаем осознавать запоминаемое, эпизодическая память, види-
мо, срабатывает сама собой.
Это означает, что для сохранения сознательных впечатлений, например,
образа чашки кофе, представленного на рис. 9 .4, по-видимому, необходимо и
достаточно «записать» это сознательное впечатление с помощью медиальной
височной коры при условии, конечно, что мы имеем дело с интактным моз-
гом. След эпизодической памяти состоит из ансамбля медиальных височных и
неокортикальных нейронов, в котором медиальная височная кора играет роль
указателя событий, хранящихся в нейрональных элементах новой коры. Из-
влечение информации происходит, когда сигнал со стороны сознания запускает
медиальную височную кору, которая, в свою очередь, активирует весь ансамбль
нейронов новой коры, ассоциированный с этой информацией. Когда мы восста-
навливаем эпизодические воспоминания, мы восстанавливаем сознательный
опыт (M�sc�v�tch, 1995). Восстановление сознательно пережитых событий во
всех деталях всегда зависит от гиппокампа вне зависимости от того, насколь-
ко давно данное воспоминание было приобретено. Как было показано (M�sc�-
M�sc�-
v�tch, 1992), гиппокампальный комплекс действует как модуль, чьей областью
действия является сознательно усвоенная информация. И похоже, что большая
часть данных, действительно, согласуется с такой точкой зрения.
Однако некоторые недавние исследования подвергли сомнению эту гипо-
тезу. Используя фМРТ, Хенке с соавторами (He��e et al., 2003; De����� et al.,
2005) показали, что гиппокамп может быть активирован путем подпороговой
демонстрации лиц и связанных с ними профессий. Более того, эта активация
коррелирует с результатами последующих тестов на эксплицитную память
Обучение и память
519
в виде подбора пар «лицо-профессия». Сходным образом обнаружено (D�sel��r
et al., 2006), что задне-медиальные отделы височной доли в большей степени
активируются при извлечении информации о старых, хорошо изученных объ-
ектах, даже если испытуемый не знает, что данный объект ему давно знаком.
Наконец, было показано (Sche��� et al., 2003), что гиппокамп активируется при
решении задач на серийное время реакции, если повторные последовательно-
сти имеют ассоциативную связь более высокого порядка.
Имеются также сходные сообщения, полученные при исследовании боль-
ных с амнезией. Остергорд (Oster���r�, 1987) был первым, кто показал, что ре-
Oster���r�, 1987) был первым, кто показал, что ре-
, 1987) был первым, кто показал, что ре-
зультаты некоторых прайминг-тестов связаны со степенью повреждения меди-
альных отделов височной доли.
Позднее Чун и Фелпс (Chu�, Phelps, 1999) показали, что у пациентов с ам-
Chu�, Phelps, 1999) показали, что у пациентов с ам-
, Phelps, 1999) показали, что у пациентов с ам-
Phelps, 1999) показали, что у пациентов с ам-
, 1999) показали, что у пациентов с ам-
незией не обнаруживается неосознаваемого влияния контекста при визуальном
поиске, подтверждая таким образом, что для сохранения контекстуальной ин-
формации, о которой испытуемый не осведомлен, необходима медиальная ви-
сочная кора. Аналогично Райан и Коэн (Ry�� et al., 2000) показали, что у боль-
ных с амнезией не отмечается нормального кругового рисунка движений глаз
при осмотре ранее изученного места, где произведено изменение расположения
объектов, даже несмотря на то, что ни больные, ни здоровые не были осведом-
лены на сознательном уровне о произошедшей перемене.
Это — лишь небольшая часть данных на фоне множества более старых ис-
следований, в которых утверждалось, что гиппокамп связан только лишь с экс-
плицитной памятью. Если результаты недавних исследований будут воспроиз-
ведены и в них не обнаружится ошибок, то это изменит наши представления
о связи гиппокампа с сознанием и памятью.
5.1. Отвлечение внимания мешает обучению
Для успешного обучения необходима концентрация внимания. Невозможно
учиться в окружении отвлекающих факторов. Психологи использовали эту ме-
тодику «рассеивания внимания» или «двойной задачи», чтобы понять вклад
внимания (или сознания) в формирование памяти. В типичном случае испыту-
емых просят обработать целевой материал, такой как слова или картинки, в то
время как их внимание отвлекается какой-то другой задачей, например отсле-
живанием точки на экране или определением трех последовательных нечетных
чисел в бегущей строке с цифрами. В таких условиях, даже если испытуемо-
му дается задача, требующая тщательного, осмысленного анализа материала,
память при рассеянном внимании значительно хуже, чем память при его кон-
центрации. Успешное кодирование требует определенного уровня внимания и,
вероятно, сознания.
Почему именно это так, до конца непонятно. Одно возможное объяснение
состоит в том, что более тщательная обработка информации требует большего
времени для своего завершения, а рассеивание внимания ограничивает время,
выделенное для кодирования. Другое возможное объяснение состоит в том, что
сознание или осознание необходимо для памяти. Если обрабатываемый мате-
риал не полностью осознается, вне зависимости от того, насколько вдумчиво он
обрабатывался, память будет, соответственно, страдать. Третье возможное объ-
яснение состоит в том, что внимание ограничивает обработку или организацию
информации, при этом оба параметра способствуют хорошей памяти.
520
Глава 9
Флетчер с соавторами (Fletcher et al., 1995) в ПЭТ-исследовании (позитрон-
но-эмиссионной томографии) обнаружили, что при рассеивании внимания сни-
жается активация левой нижней префронтальной области. Этот результат был
воспроизведен на молодых и пожилых испытуемых Андерсоном с соавторами
(A��ers�� et al., 2000), показавшими, кроме того, что при рассеивании внима-
ния снижается также активность в левом медиальном отделе височных долей,
важном, как известно, для вербальной памяти.
Как память, так и обучение имеют осознанный и неосознанный аспекты.
Если рассуждать о трех фазах — обучении, удержании информации в памяти
и ее извлечении, — то все возможные варианты можно уложить в матрицу 3×4
(табл. 9 .1). Из этих трех удержание обычно рассматривается как бессознатель-
ное, хотя формируется сознательным опытом. Считается, что обучение часто
требует осознания, и интуитивно понятно, что при обучении определенно ак-
центируется внимание и осознается изучаемый предмет. Это, видимо, и есть
основная стратегия обучения человеческих существ.
Тем не менее имеются некоторые доказательства возможности обучения
без участия сознания, особенно в случае эмоциональных стимулов. Имеются
весомые доказательства для имплицитного обучения, при котором некий логи-
ческий процесс занимает осознаваемый информационный вход и кодирует ре-
зультаты бессознательной обработки осознаваемой информации, поступающей
через этот вход. Тем не менее в задачах на имплицитное обучение испытуемых
всегда просят обратить внимание и осознать набор стимулов (раздел 2.4). При
этом правила и закономерности генерации этих стимулов воспринимаются бес-
сознательно — как в детстве постигается грамматика без эксплицитного знания
ее правил. Однако, чтобы произошло имплицитное обучение, нужно постоянно
слышать произносимые слова и их последовательности.
Термины имплицитная и эксплицитная память используются в контексте
припоминания, т. е . извлечения из памяти хранимой информации.
Таблица 9.1
Эксплицитные и имплицитные типы обучения и памяти
Тип
Обучение
Удержание
в памяти
Извлечение
из памяти
Эпизодическая
память
Сознательное
или эксплицитное
Бессознатель-
ное
Сознательное или
эксплицитное для
процесса обучения
Семантическая
память
Сознательное
или эксплицитное
Бессознатель-
ное
Бессознательное
или имплицитное
для процесса
научения
Имплицитное об-
учение
Сознательные стиму-
лы, однако бессозна-
тельно изучаемые за-
кономерности
Бессознатель-
ное
Бессознательное
извлечение
информации
Подпороговое
обучение (редко
устойчивое и
долговременное)
Бессознательное вос-
приятие целевых сти-
мулов
Бессознатель-
ное
Бессознательное
извлечение
информации
Обучение и память
521
Эксплицитная память — это память с так называемой «осознанной осведом-
ленностью», а именно память о некотором объекте или явлении, о котором дан-
ный человек осведомлен, может продекларировать существование этого объек-
та и прокомментировать его содержание, вербально или невербально (Sch�cter,
1987). Поэтому такая память также известна, как декларативная (Ryle, 1949;
C�he� ��� Squ�re, 1980). Это такая память, к которой обращаются в типичном
повседневном разговоре, когда спрашивают, например: «Ты не забыл позвонить
тетушке и поблагодарить ее за подарок на день рождения?» или «Помнишь, кто
получил премию «Оскар» в номинации лучший актер или актриса?»
6.0. Префронтальная кора, сознание и рабочая память
Префронтальная кора (ПФК) играет главную роль в формировании рабочей па-
мяти. Префронтальная кора как у людей, так и у других приматов находится
перед моторной корой (рис. 9 .25). Большинство исследований рабочей памяти
выполнено в экспериментах на макаках. Очевидно, что для некоторых челове-
Рис. 9 .25. Префронтальная кора (ПФК) у обезьян (сверху) и у людей (снизу). Чаще
всего ПФК делят на верхнюю и нижнюю половины, которые называются дорсолате-
ральной префронтальной корой (ДЛ-ПФК) (фиолетовая область) и вентролатеральной
префронтальной корой (ВЛ-ПФК) (зеленая область). Для ориентировки приводятся пе-
ресекающиеся стрелки, указывающее на дорсальное (В, верхнее), вентральное (Н, ниж-
нее), ростральное (Р, по направлению к носу у людей) и каудальное (К, по направлению
к затылку у людей) направления. (Источник: R������th, 2006.)
522
Глава 9
Рис. 9 .26 . «Отсроченная сверка с образцом» (DMTS, �el�ye� m�tch t� s�mple) у макаки.
В классическом эксперименте макаку обучают выполнять отсроченный ответ на сти-
мул, в данном случае — расположение на экране области красного, белого или синего
цвета. В задаче, называемой «отсроченная сверка с образцом», после периода задержки
макака опознает стимул путем выбора его на экране. По существу, обезьяна сообщает:
«Это то, что я видела». Методика DMTS широко используется на животных, на младен-
DMTS широко используется на животных, на младен-
широко используется на животных, на младен-
цах, еще не умеющих говорить, и других объектах. (Источник: Fuster, 1997.)
Обучение и память
523
ческих способностей, таких как язык, нет прямых параллелей с животными.
Однако в случае исследований рабочей памяти макака стала неизменным важ-
ным источником новых данных.
Впервые такие данные о связи между префронтальной корой и кратковре-
менной памятью появились в 1930-е гг., когда было обнаружено, что обшир-
ные двусторонние повреждения префронтальной коры у животных нарушают
выполнение задачи с отсроченным ответом. В этой задаче животному предъ-
является простой стимул (напр., цвет или местоположение), и его идентич-
ность должна удерживаться в памяти на короткий период задержки так, чтобы
этот стимул мог направить последующий ответ (рис. 9 .26). Комбинируя этот
классический тест с новыми нейрофизиологическими методами, современные
исследователи надежно установили роль префронтальной коры в активном
сохранении информации в рабочей памяти (рис. 9 .27).
Рис. 9 .27. Задача на отсроченный ответ для изучения рабочей памяти у обезьян и лю-
дей. Было предположено, что дорсолатеральная префронтальная кора играет особую
роль в активном хранении информации в рабочей памяти (например, ��l�m��-R���c,
1998). Таким образом, устойчивая активность нейронов в префронтальной области от-
ражает роль этого региона в сохранении специфических образов объектов, которые
должны быть сохранены в мозгу после задержки. Это объяснение подтверждается дан-
ными о том, что индивидуальные нейроны в дорсолатеральной префронтальной коре
селективны для отдельных целевых стимулов. Например, данная клетка может сильно
разряжаться в течение периода задержки, когда мишень находится в верхней левой об-
ласти экрана, но слабо — когда мишень находится в каком-то другом месте на экране.
Такой паттерн указывает на прямое участие внутреннего представительства в целевые
характеристики. На этом рисунке представлен ответ нейронов в префронтальной коре
во время задержки при выполнении «задачи по отсроченной сверке с образцом». У обе-
зьян и у людей получены сходные результаты. (Источник: Curtis ��� D�Esposito, 2003.)
524
Глава 9
Множество исследований на животных было сконцентрировано на специфи-
ческой лобной области, называемой дорсолатеральной префронтальной корой
(ДЛ-ПФК, см. рис. 9.25). (В коре человека дорсальный значит «верхний», а ла-
теральный — «боковой».) Одно из ключевых ранних открытий было сделано в
лаборатории Хоакина Фустера (Fuster, Alex���er, 1971). Фустер и его коллеги об-
Fuster, Alex���er, 1971). Фустер и его коллеги об-
, Alex���er, 1971). Фустер и его коллеги об-
Alex���er, 1971). Фустер и его коллеги об-
, 1971). Фустер и его коллеги об-
учали обезьян выполнять задачу на отсроченный ответ, при которой они должны
были на короткий период задержки запомнить предъявленный цвет, а затем при
тестировании указать на правильный ответ из двух вариантов. Поскольку после
исходного предъявления стимула никакой информации о правильном цвете не
предлагалось, то его идентичность должна была сохраниться в рабочей памяти.
Используя вживленные электроды для записи нейронной активности во время
выполнения задачи (см. гл . 4), авторы обнаружили, что на протяжении периода
задержки отмечалась устойчивая и постоянная активность отдельных нейронов
в дорсолатеральной префронтальной коре обезьяны. Даже после того, как цвет
убирали с экрана дисплея, нейроны в дорсолатеральной префронтальной коре
продолжали разряжаться с повышенной частотой, которая снижалась лишь по-
сле того, как был дан ответ «совпадает/не совпадает» (рис. 9 .26).
С тех пор рисунок устойчивой нейронной активности на период задержки
в дорсолатеральной префронтальной коре воспроизводился множество раз и в
самых разных задачах.
Например, чтобы подтвердить, что вклад префронтальной коры действи-
тельно связан с памятью, а не является просто отражением едва уловимых под-
готовительных жестов, Патриция Голдман-Ракич (P�tr�c�� ��l�m��-R���c) и ее
коллеги разработали версию задачи, при которой обезьяны в течение короткого
времени видят на дисплее мишень, расположенную в одной из нескольких воз-
можных позиций, а затем, после задержки, должны перевести свой взгляд на
другую позицию, чтобы получить награду. Важно, что до окончания периода
задержки обезьянам требуется смотреть прямо вперед, так что нейрональная
активность во время периода задержки не может быть просто побочным про-
дуктом движения глаз, а должна действительно отражать процессы памяти. Та-
кая экспериментальная парадигма также вызывает устойчивую нейрональную
активность в дорсолатеральной префронтальной коре и, что еще важнее, по ко-
личественному выражению величины активности в течение периода задержки
можно предсказать, будут запомнены объекты, или нет; если активность в дор-
солатеральной префронтальной коре в период задержки слабая, то вероятность
забывания выше (Fu��h�sh� et al., 1993).
Спорным остается вопрос относительно того, подразделяется ли пре-
фронтальная кора в соответствии с содержанием информации, которая в ней
хранится, или в соответствии с функцией, которую выполняет каждая ее об-
ласть. Если разделять по содержанию, то дорсолатеральная префронтальная
кора, по-видимому, вовлечена в удержание в памяти информации, в частности,
о пространственном положении, в то время как различные части вентральной
и латеральной префронтальной коры принимают участие в хранении непро-
странственной информации (например, объектов, лиц, слов и т. п.) . С другой
стороны, каждая из этих областей может иметь различные функции, при этом
дорсолатеральная префронтальная кора участвует в манипуляциях с информа-
цией, а вентролатеральная — в ее удержании (вентролатерально, снизу и сбо-
ку). Термин «вентральный» по отношению к коре означает направление вниз;
буквально, ve�trum означает «брюхо» по латыни.
Обучение и память
525
В исследованиях с разрушением мозга обезьян были получены дополни-
тельные сведения о связи префронтальной коры, и, в частности, ее дорсолате-
ральной части, с функцией рабочей памяти. Используя очень точные методи-
ки определения местоположения экспериментально вызванных повреждений,
было показано, что разрушения, ограниченного в точности дорсолатеральной
префронтальной корой, достаточно для нарушения выполнения тестов на ра-
бочую память (Fuster, 1997). Эти данные указывают на причинную роль пре-
Fuster, 1997). Эти данные указывают на причинную роль пре-
, 1997). Эти данные указывают на причинную роль пре-
фронтальной коры в рабочей памяти. Клетки в этой области не только активны
во время задержки, но и их повреждение нарушает рабочую память. Это на-
рушение усугубляется с увеличением продолжительности задержки, подтверж-
дая, что информация быстрее забывается, если не закрепляется с участием
префронтальной коры. Исследования на людях с использованием нейровизу-
ализационных методов подтвердили массу литературных данных, полученных
в экспериментах на животных. В сотнях визуализационных исследований по-
казана активность префрональной коры, когда участники пытаются удержать
в памяти информацию, относящуюся к заданию. В исследованиях с использо-
ванием фМРТ показано, что активность префрональной коры у испытуемых
сохраняется в течение всего периода задержки теста на рабочую память (см.
гл. 4, 10 и 12, а также приложение), что согласуется с результатами работ на
животных.
Нейровизуализационные исследования на людях также внесли изменения в
понятие нагрузки на рабочую память — количество элементов, которые долж-
ны удерживаться в непосредственной памяти (C�he� et al., 1997; Rypm� et al.,
2002). В одном исследовании нагрузка на память варьировала от одного до
восьми элементов, и испытуемым было необходимо удерживать эти объекты
в памяти на протяжении короткой задержки. Было обнаружено, что активация
префронтальной коры положительно коррелирует с количеством элементов в
памяти. Такая «зависимость от нагрузки» в префронтальной коре подтверждает
точку зрения о том, что эта часть мозга принимает участие в хранении инфор-
мации в рабочей памяти (см. гл . 12).
Хотя роль префронтальной коры в рабочей памяти была четко продемон-
стрирована, особый вклад этой области в собственно хранение информации
в рабочей памяти в последнее время был подвергнут сомнению. В некоторых
других корковых и подкорковых областях отмечается сходная, постоянная,
стимул-специфическая активность в ходе коротких задержек. Похоже, что
префронтальная кора — часть более обширной сети мозга, обеспечивающей
формирование рабочей памяти. Другие же данные приводят к предположению
о том, что префронтальная кора сама по себе вообще может не участвовать в
хранении информации, а обеспечивать нисходящую, или исполнительную, под-
держку другим областям, в которых информация действительно хранится.
6.1. Работа с памятью: лобная доля
целенаправленно работает с памятью
В соответствие с этой моделью, как кодирование, так и извлечение сознательно
усвоенной информации посредством гиппокампа и связанных с ним структур
является обязательным и автоматическим. Однако исходя как из обыденного
опыта, так и экспериментальных исследований, известно, что есть все же не-
которая мера контроля того, что кодируется и что извлекается из памяти. Более
526
Глава 9
того, если кодирование обязательное и автоматическое, то информация не мо-
жет быть организована, хотя память, по-видимому, имеет некоторую времен-
ную и тематическую организацию. Каким образом можно «увязать» эту модель
памяти с другими фактами, которые мы знаем о ее работе? Одно возможное
решение состоит в том, что другие структуры, в особенности находящиеся в
лобных долях, контролируют информацию, поступающую в медиальную ви-
сочную систему при кодировании, запускают и направляют ее извлечение, а за-
тем отслеживают и помогают интерпретировать и организовать извлеченную
информацию. Оперируя медиальной височной и диэнцефальной системой,
лобные доли действуют в качестве работающих с памятью структур, которые
контролируют более автоматическую медиальную височную систему и даруют
ей некоторую меру разумности и целенаправленности. Такая дополнительная
система необходима в случае, когда память должна выполнять функции, отлич-
ные от простого сохранения и извлечения прошлых знаний (M�sc�v�tch, 1992).
6.2 . Префронтальная кора в эксплицитном (осознанном)
и имплицитном (неосознанном) обучении и памяти
Как показано на рис. 9 .5, рабочая память помогает получать как эксплицит-
ную (осознанную), так и имплицитную (неосознанную) информацию. Одной
из функций, часто приписываемых сознанию, является интеграция информа-
ции из разных областей. Свет на эту проблему пролило исследование (McI�t�sh
et al., 1998), в котором испытуемые выполняли задачу на выработку следового
условного рефлекса, согласно которой требуется, чтобы испытуемый провел
ассоциацию между цветом и тоном, предъявление которых разделено мертвой
паузой около секунды. Предыдущая работа показала, что такое обусловливание
зависит от гиппокампа. Более того, Кларк и Сквайр (Cl�r�, Squ�re, 1999) пока-
Cl�r�, Squ�re, 1999) пока-
, Squ�re, 1999) пока-
Squ�re, 1999) пока-
, 1999) пока-
зали, что осознанная осведомленность об ассоциации является необходимым
условием этого типа обучения.
Макинтош, используя ПЭТ, показал, что обучение и осознанная осведом-
ленность, которая сопровождает его, были связаны и с лобной активацией, и
с согласованностью в активации многих других областей коры. Макинтош и
соавторы высказали предположение о том, что сознание ассоциируется с ак-
тивацией префронтальной коры, которая, в свою очередь, приводит к согласо-
ванному рисунку активации самых различных областей коры. До сих пор не
установлено, предшествует ли лобная активация осознанной осведомленности
об ассоциации или сопровождает ее. Чтобы это выяснить, необходимо иметь
четкое представление о последовательности активации областей коры, для чего
нужно использовать методы с более высоким временным разрешением, чем у
фМРТ, такие, как вызванные потенциалы (ВП) или магнитоэнцефалография
(МЭГ). До сих пор лишь в очень немногих исследованиях (если таковые во-
обще есть) применяли ВП и МЭГ для изучения динамики сознания и его связи
с памятью.
Если префронтальная кора, по предположению многих, играет ключевую
роль в сознании, то у пациентов с повреждениями лобной доли во всех тестах
на память, зависящей от сознания, должны отмечаться нарушения. Однако су-
ществующие данные указывают на то, что эффекты лобных повреждений зна-
чительно более избирательны и отнюдь не так критичны, как повреждения ме-
диальной височной доли и связанных с ней ядер таламуса.
Обучение и память
527
Префронтальная кора вносит свой вклад в реализацию имплицитного об-
учения и памяти, если при этом требуется провести поиск, упорядочивание,
организацию и сознательное отслеживание. Хорошим примером является им-
плицитное обучение языку (блок 9.4).
Блок 9.4 . Имплицитное изучение языка. В то время, как слова и языковые
последовательности приходится учить, грамматические правила
и закономерности, а, возможно, и значения слов порой усваивают-
ся бессознательно, т. е . с помощью имплицитного обучения
Имплицитное обучение и овладение языком
Последние несколько лет свидетельствуют о возникновении усиливающихся связей
между теорией имплицитного обучения и психолингвистикой. Это, пожалуй, не столь
уж удивительно, ибо овладение языком, как всякое имплицитное обучение, включает
и обучение в случайных условиях. Более того, свободное пользование языком вооб-
ще не требует эксплицитных знаний грамматики. С недавних пор некоторые авторы
стали изучать эту связь эмпирически. К примеру, Саффран и соавторы (S�ffr�� et al.,
1997) показали, что случайного прослушивания искусственного языкоподобного ау-
диоматериала (например, бупадапатубитутибу...), было достаточно, чтобы и дети,
и взрослые испытуемые смогли разделить непрерывную последовательность звуков,
которую они услышали, на искусственные слова (напр., бупада, патуби и т. п.), кото-
рые в ней содержались, что подтвердилось при последующем тестировании на рас-
познание. На основании этих данных Саффран и соавторы выдвинули предположе-
ние о том, что способность разделять слово на части, продемонстрированное этими
испытуемыми, связана с тем, что вероятность повторения одних и тех же слогов вну-
три слов выше, чем между словами. Саффран с коллегами интерпретировали свои
результаты, как пример одной из форм имплицитного обучения. Связь становится
очевидной, если вспомнить, что освоение языка, как имплицитное обучение (�erry,
D�e�es, 1993; Cleerem��s, 1993), вероятно, включает, по крайней мере отчасти, и слу-
, 1993; Cleerem��s, 1993), вероятно, включает, по крайней мере отчасти, и слу-
Cleerem��s, 1993), вероятно, включает, по крайней мере отчасти, и слу-
, 1993), вероятно, включает, по крайней мере отчасти, и слу-
чайное обучение сложной многоуровневой информации. Представление Саффрана с
коллегами о сходстве между освоением языка и имплицитным обучением возникло,
быть может, частично, под влиянием компьютерного моделирования в области ис-
следований памяти. К примеру, коннекционистские модели, такие, как Простая Воз-
вратная Сеть (S�mple Recurre�t Net��r�), были широко и с большим успехом исполь-
зованы в обеих областях — освоении языка и имплицитном обучении (Chr�st���se� et
al., 1998; Re����t��, Ch�ter, 1997). В самом деле, проблемы в обеих областях крайне
схожи: как наилучшим образом выделить определенную структуру из сложной сре-
ды стимула, характеризующейся «глубокими» систематическими закономерностями,
да еще когда обучение происходит скорее случайно, чем намеренно. Ответ в обеих
областях, по-видимому, реализуется с помощью дистрибутивных методов.
Литература
1. S�ffr��, J. R., et al. (1997). I�c��e�t�l l���u��e le�r����, L�ste���� (��� le�r����) �ut �f
the c�r�er �f y�ur e�r. Psychological Science, 8 , 101 –105,
2. �erry, D. C., & D�e�es, Z. (1993). Implicit Learning: Theoretical and Empirical Issues,
L�����: Erl��um.
3. Cleerem��s, A. (1993). Mechanisms of Implicit Learning: Connectionist Models of
Sequence Learning. C�m�r���e: MIT Press.
4. Chr�st���se�, M. H., Alle�, J., Se��e��er�, M. S. (1998). Le�r���� t� se�me�t speech us���
mult�ple cues: � c���ect����st m��el. Language and Cognitive Processes, 3, 221–268.
5. Re����t��, M., ��� Ch�ter, N/ (1997). Pr�����l�st�c ��� ��str��ut����l �ppr��ches t�
l���u��e �cqu�s�t���. Trends in Cognitive Sciences, 1, 273–281 .
528
Глава 9
Несмотря на то что правила грамматики редко применяются осознано и экс-
плицитно, все-таки, чтобы выучить какой-то язык имплицитно, необходимо со-
средоточить внимание, например, на порядке слов в предложении. Вероятно,
бессознательные умозаключения помогают обнаружить правила или законо-
мерности при условии, что «сознательно обращается внимание», например, на
последовательность слов, откуда и выявляются имплицитные закономерности.
6.3 . Различные типы рабочей памяти
Предлагая концепцию рабочей памяти, Бэддли (����eley) и коллеги (2004) при-
����eley) и коллеги (2004) при-
) и коллеги (2004) при-
водят в качестве довода то, что у испытуемых наблюдаются трудности с удер-
жанием элементов в памяти, если их одновременно просят выполнить другие
задания, нарушающие работу специфических компонентов системы рабочей
памяти. В подтверждение своей точки зрения Бэддли и др. показали, что если
испытуемых во время удержания информации в памяти просят громко повто-
рять простой речевой фрагмент (например, определенный артикль «the»), то
объем кратковременной памяти на вербальную информацию резко сокращает-
ся, предположительно из-за того, что эти устные повторы нарушают механизм
повтора, рехэрсела (см. выше) фонологической петли. Существенно важно, что
та же самая задача на речевые повторы оказывает значительно меньший эффект
на рабочую память для зрительно-пространственной информации, поскольку
в этом случае фонологическая петля почти не задействована. В то же время
было обнаружено, что другие задачи (например, визуальное отслеживание дви-
жущегося объекта) нарушают сохранение зрительно-пространственной, но не
вербальной информации.
Мы уже сталкивались с примерами больных, таких как Клайв Уэринг и HM,
у которых, по-видимому, сохранялась кратковременная память, но нарушалась
долговременная. Элизабет Воррингтон и Тим Шоллис (Sh�ll�ce ��� W�rr���t��,
1970) сообщили об одном из первых распознанных случаев пациента KF с про-
KF с про-
с про-
тивоположным примером нарушения — резко нарушенной кратковременной
памятью, но, по всей видимости, интактной долговременной памятью. Напри-
мер, когда KF просили повторить короткий перечень только что произнесенных
чисел (задача на запоминание чисел), он мог достоверно восстановить только
одно или два числа (обычно воспроизводится около семи чисел). Тем не менее
KF сохранял относительно нормальные речевые способности, и мог учиться
и запоминать новую информацию. Описание больного с тяжелым нарушени-
ем кратковременной памяти, который, несмотря на это, все еще был способен
передавать информацию в долговременную память, поставило под сомнение
стандартную гипотезу, утверждавшую, что «воротами» для долговременной па-
мяти служит не что иное, как единая кратковременная память.
Согласно модели рабочей памяти Бэддли при нарушении вербального повторе-
ния для компенсации, может использоваться зрительно-пространственный «аль-
бом» — скечпэд* (система ввода и редактирования информации) (см. рис. 9.5).
*
S�etchp�� — революционная компьютерная программа, созданная Айвеном Сазерлэндом в 1963 г.
S�etchp�� считается предком современных систем автоматизированного проектирования (САПР),
а также крупным прорывом в развитии компьютерной графики в целом. Например, графический
интерфейс пользователя впервые был сделан в S�etchp��. Айвэн Сазерлэнд продемонстрировал, что
компьютерная графика может быть использована для художественного и технического применения,
в дополнение к демонстрации нового (для того времени) способа взаимодействия человека и ком-
пьютера < http://plmpe���.ru/����/S�etchp�� >.
—
Прим. перев.
Обучение и память
529
Действительно, нарушение кратковременной памяти у больного KF и у ряда
сходных пациентов, о которых с тех пор сообщалось, по-видимому, связано с
определенными типами информации. Например, хотя эти пациенты испытыва-
ли значительные затруднения при запоминании вербальных элементов на слух,
их результаты значительно улучшались при зрительном представлении этих же
предметов. Как можно объяснить полученные результаты? Ответ Бэддли со-
стоит в том, что визуально представленные предметы могут кодироваться не-
посредственно в зрительно-пространственный «альбом» — скечпэд, таким об-
разом избегая поврежденной петли вербального повтора — рехэрсела.
Нейровизуализация помогла прояснить особенности различных видов па-
мяти. К ним относится различие между подсистемами сохранения вербального
и зрительно-пространственного материала (например, Sm�th et al., 1996), раз-
деление между хранением и повторением (рехэрселом) при сохранении вер-
бального материала (Paulesu et al., 1993; A� h et al., 1996) и предположение о
наличии центрального исполнительного процессора, который опосредует ра-
боту подчиненных ему подсистем сохранения информации (например, Curtis,
D�Esposito, 2003). В целом, результаты нейровизуализационных исследований
скорее подтверждают основную модель (Sm�th, J����es, 1998; H�rtley, Speer,
2000; He�s��, 2001), нежели опровергают ее.
На рис. 9.28 изображена мозговая сеть, которая по результатам нейрови-
зуализационных исследований непосредственно связана с рабочей памятью
(Curt�s, D�Esp�s�t�, 2003). �ентральный исполнительный орган соответству-
Curt�s, D�Esp�s�t�, 2003). �ентральный исполнительный орган соответству-
, D�Esp�s�t�, 2003). �ентральный исполнительный орган соответству-
D�Esp�s�t�, 2003). �ентральный исполнительный орган соответству-
�Esp�s�t�, 2003). �ентральный исполнительный орган соответству-
Esp�s�t�, 2003). �ентральный исполнительный орган соответству-
, 2003). �ентральный исполнительный орган соответству-
ет дорсолатеральной префронтальной коре (отмечен буквой «D» на рисунке),
подсистема сохранения вербальной информации — левым боковым областям
височно-теменного соединения (отмечена буквой «Р») и вентролатеральной
префронтальной коре, а зрительно-пространственная подсистема сохранения
информации — верхней теменной коре, задним отделам префронтальной коры
Рис. 9 .28 . Области мозга, которые, как полагают, участвуют в вербальной и визуаль-
ной рабочей памяти. Упрощенная модель рабочей памяти (рис. 9 .4 и 9.5). D — дорсо-
латеральная префронтальная область; В — зона Брока, также называемая левой нижней
лобной извилиной; P — фонологическая петля для вербального повторения (рехэрсела),
также называемая надкраевой извилиной; F — лобные глазные поля, которые, как счи-
F — лобные глазные поля, которые, как счи-
—
лобные глазные поля, которые, как счи-
тается, участвуют в зрительно-пространственном скечпэде (наборе изображений, аль-
боме), входящем в модель рабочей памяти Бэддли. (Источник: Curtis, D�Esposito, 2003.)
530
Глава 9
и лобным глазным полям («F»). Однако в качестве прямого подтверждения об-
F»). Однако в качестве прямого подтверждения об-
»). Однако в качестве прямого подтверждения об-
щей схемы эти данные нужно воспринимать с осторожностью, поскольку не-
которые исследователи рассматривают их в качестве подтверждения альтерна-
тивных теорий рабочей памяти (Che�� et al., 2003; R�v�zz� et al., 2005). Кроме
того, порой в нейровизуализационной литературе ставятся под сомнение те
или иные аспекты стандартной модели рабочей памяти. Например, нейрови-
зуализационные данные указывают на то, что различные типы зрительно-про-
странственной информации могут зависеть от различных подсистем хранения
информации. К примеру, сохранение объектной информации и информации о
пространственных расположениях, по-видимому, обеспечивается различными
нервными субстратами — особенность, которая не рассматривалась в тради-
ционной модели рабочей памяти. (Напомним, что такое разделение на «что» и
«где» важно также и при обработке зрительной информации; см. гл . 6 .)
6.4 . Префронтальная кора — хранение информации
или управление процессом?
В начале этого раздела о рабочей памяти мы рассматривали некоторые рабо-
ты, указывающие на то, что префронтальная кора имеет важное значение для
функции рабочей памяти. В соответствие с одним возможным объяснением, эта
область мозга непосредственно участвует в хранении информации. Однако рас-
смотрение данных в контексте многокомпонентной модели Бэддли подтверж-
дает альтернативную точку зрения — а именно что префронтальная кора более
тесно связана с управлением, или с исполнением различных аспектов рабочей
памяти.
Особое свидетельство против представления о функции префронтальной
коры, как «хранителя информации», было также получено в исследованиях на
людях. Выше уже обсуждались больные с повреждением левой височно-темен-
ной области, у которых, по-видимому, было нарушение хранения информации
в рабочей памяти и которые не могли выполнять даже простые задачи на удер-
жание словесно-слуховой информации. Данные, полученные у этих пациентов,
резко отличаются от данных, полученных у больных с повреждением префрон-
тальной коры. При обзоре сообщений о больных с обширными повреждениями
латеральной префронтальной коры (D�Esposito, Postle, 1999) было обнаружено,
что у них не отмечаются вовсе или отмечаются лишь незначительные нару-
шения при решении задач, требующих пассивного сохранения информации на
период задержки (например, вербальные или невербальные задачи на объем
памяти). Однако наблюдались значительные нарушения при решении таких за-
дач, которые требуют мысленных манипуляций или воздействий на информа-
цию в рабочей памяти. Такой характер данных, приведенных в этом обзоре,
указывает на то, что префронтальная кора служит скорее для обеспечения ум-
ственной «работы» над хранимой информацией, чем просто в качестве места
для ее хранения. Лишь у нескольких пациентов, упомянутых в обзоре, имелись
двусторонние повреждения, так что остается открытой возможность хранения
и повторного воспроизведения (рехерсэла) информации за счет двусторонне-
го представительства префронтальной коры, и, таким образом, компенсации
функции из неповрежденного полушария (D�Esposito, Che�, 2006).
Одно возможное объяснение может состоять в том, что различные части
префронтальной коры отвечают за разные вещи. Это предположение основано
Обучение и память
531
главным образом на различиях между дорсальными и вентральными областя-
ми. Сторонники этого взгляда утверждают, что префронтальная кора организо-
вана не по доменам (например, пространственные/непространственные), а по
процессам, при этом вентральные области участвуют в пассивном хранении
информации и удержании объектов в памяти, в то время как более дорсальные
области привлекаются, когда задача требует проведения отбора, контроля, ма-
нипуляции или другой «умственной работы» над этими объектами. Это так на-
зываемое разделение по переработке информации: либо ее «удержание», либо
«манипулирование». Хотя такое представление, казалось бы, может объяснить
широкий диапазон данных, некоторые исследования подвергли сомнению даже
предположение о том, что вентральные области вообще вносят вклад в хране-
ние информации в рабочей памяти (Rush��rth et al., 1997).
В последнее время были приведены доводы против того, что вся префрон-
тальная кора имеет исполнительную функцию в рабочей памяти, скорее различ-
ные ее подотделы выполняют эту функцию на различных уровнях анализа (R��-
Ran-
����th, 2006). Согласно этой точке зрения, основная функция префронтальной
коры состоит в модуляции активности других корковых областей, где хранятся
объекты памяти. В частности, представительства префронтальной коры усили-
вают значимую информацию (или тормозят незначимую), представленную в
других областях коры. Когда информация специфична для отдельных объектов
в памяти, задействуется больше вентральных областей префронтальной коры.
Когда информация касается интеграции (или связи между) множества элемен-
тов, задействуется больше дорсальных областей. Передние же области, распо-
ложенные на лобном полюсе, участвуют в координации и мониторинге актив-
ности в различных областях префронтальной коры для выполнения функций
высшего порядка, таких как планирование. Соответственно, первичная роль
префронтальной коры связана не с рабочей памятью, а с работой с памятью
(M�sc�v�tch, 1992; M�sc�v�tch, W���cur, 1992).
6.5 . Объединение префронтальных и медиальных
височных областей для обеспечения процессов
рабочей памяти
Обычно считается, что рабочая память действует в течение нескольких десят-
ков секунд или нескольких минут. Однако даже в пределах минутного интер-
вала можно найти различия между разными видами памяти. Например, мину-
ту-другую назад вы прочли в этом тексте слово «комбинирование» и пока что,
вероятно, не сможете сознательно вспомнить его. Таким образом, основываясь
на вышеописанной роли медиальной височной коры в эксплицитной памяти,
можно сделать следующий прогноз: если вы сможете вспомнить, что только
что видели слово «комбинирование», то при исследовании с помощью фМРТ
в медиальной височной коре должна появиться активность, когда вас попросят
вспомнить первое слово этого раздела. Наоборот, если у вас нет эксплицитного
воспоминания об этом (даже через короткий период времени), то активность в
медиальной височной коре будет меньше. Однако семантическое воспоминание
о названии этого раздела, основанное на кодировании этого слова в новой коре,
у вас, возможно, все же сохранится.
Таким образом, можно даже описать рабочую память в терминах медиаль-
ной височной, префронтальной, сенсорной коры и т. п. На рис. 9 .29 приводится
532
Глава 9
основанная на большом количестве исследований модель того, как эти области
мозга могут взаимодействовать (R������th, D�Esposito, 2005). Многие детали
данной модели еще обсуждаются и можно ожидать, что появления новых ис-
следований прольет на них свет. Тем не менее похоже, что уже достигнуто со-
гласие относительного того, что полное объяснение функций рабочей памяти
потребует создания мультирегиональной модели подобного рода.
Рис. 9 .29. Связанные области мозга работают сообща для обеспечения процессов визу-
альной рабочей памяти. Согласно одной из точек зрения на визуальную рабочую память,
гиппокамп может кодировать новые объекты рабочей памяти, более широкая система ме-
диальной височной коры может комбинировать их с другими модальностями, а нижняя
височная кора участвует в представительстве зрительного образа высшего порядка. Оба
дорсолатеральных и передних префронтальных отдела коры (фиолетового цвета) связа-
ны с кратковременным поддержанием связей, в то время как вентролатеральные и задние
префронтальные отделы связаны с сохранением в памяти релевантных объектов. Как
указано в тексте, это единственная рабочая гипотеза о функциях данных областей. Од-
нако принято считать, что для полного описания рабочей памяти нужна некоторая функ-
циональная сеть, сходная с вышеописанной. (Источник: R������th, D�Esp�s�t�, 2005.)
Обучение и память
533
7.0 . Извлечение информации из памяти и метапознание
Клайв Уэринг понимает, что что-то явно не так, однако он не имеет понятия
о том, что именно. «Только что я в первый раз проснулся. Я впервые пришел в
сознание» — его единственный способ выразить это. Вот уже более 20 лет он
постоянно выражает сильнейшую неудовлетворенность своим состоянием. По-
этому у Уэринга должно было выработаться некоторое метакогнитивное пред-
ставление о своем собственном когнитивном функционировании, в отличие от
пациента HM, например, который обходится без эмоционального переживания
по поводу того, что он ничего не помнит. Метапознание определяется как спо-
собность иметь представление о наших собственных когнитивных функциях и
быть в состоянии использовать это знание. Многие тяжелые неврологические
больные не имеют метакогнитивного понимания того, что что-то у них идет не
так (M�l�er, Ru��, 1992).
Чтобы извлечение информации было эффективным, извлекаемая информа-
ция должна перекрываться с той информацией, которая была записана или зако-
дирована в памяти. Кроме того, нужно поставить целью вспомнить и сконцен-
трировать внимание на «подсказках», а также ментальном поиске желаемого
воспоминания. При всех этих процессах необходимы мониторинг и проверка,
как и координация различных видов активности. Разные области префронталь-
ной коры участвуют во многих из этих процессов. Медиальная височная об-
ласть в наибольшей степени связана с успешным извлечением эпизодических
воспоминаний. Если встречается «подсказка», то извлечение воспоминания из
памяти часто происходит автоматически (рис. 9 .30). Стратегическое или целе-
направленное извлечение информации, тем не менее, требует внимания и нару-
шается при одновременном решении любой другой требующей усилий задачи.
Рис. 9 .30 . Извлечение семантических воспоминаний с использованием эпизодических
«подсказок» и наоборот. Часто удается извлечь то или иное семантическое воспоми-
нание, типа какого-то факта об окружающем мире, используя «подсказку» из эпизо-
дической памяти; может также возникнуть и иная ассоциация. Так, эпизодическая и
семантическая память продолжают оставаться потенциально связанными, даже если
они существуют относительно независимо друг от друга в гиппокампальном комплексе
(медиальной височной коре) и новой коре. Обратите внимание, что большинство эпизо-
дических воспоминаний для этого семантического блока ассоциаций (вверху) потускне-
ли. (Источник: M�rr�s M�sc�v�tch, личное сообщение.)
534
Глава 9
Метапознание является важным аспектом нормального извлечения инфор-
мации из памяти. След памяти может извлекаться спонтанно или, чаще, с ис-
пользованием «подсказок» или напоминаний. Подсказка может быть простой,
например «вспомните слова, которые вы только что учили», или сложной, на-
пример «детально опишите все, что вы сегодня делали». Такой способ само-
контроля, который мы склонны применить, когда пытаемся вспомнить забытое
слово, является одним из видов метапознания, в формировании которого при-
нимает участие префронтальная кора.
Шактер и коллеги (Sch�cter et al., 1984) показали, что нарушение памяти об
источнике информации у больных с амнезией было связано скорее со степенью
сопутствующей недостаточности лобных долей, чем с тяжестью самой амне-
зии. Распространяя эти данные на более пожилых людей, Глиски (�l�s�y et al.,
1995) показал, что у больных с недостаточностью лобной функции отмечалось
нарушение в тестах на память об источнике информации, однако результаты
тестов на предметную память были нормальными, в то время как у больных
с недостаточностью медиальных отделов височной доли имел место противо-
положный характер нарушений. Сходным образом повреждение префронталь-
ной коры приводит к недостаточности памяти на частоту встречаемости (Sm�th,
M�l�er, 1984) и временной порядок (M�l�er, 1971; Sh�m�mur�, 1984), даже когда
воспоминание о самом объекте сохраняется.
Функциональные нейровизуализационные данные подтверждают клинику.
По сравнению с тестами на предметную память, в тестах на память об источ-
нике сильнее активируются дорсолатеральная префронтальная кора и лобный
полюс (Fletcher, He�s��, 2001; Fletcher et al., 2005), что отмечается также и в те-
стах на временную последовательность (Ny�er� et al., 1996; Cabeza et al., 1997).
Роль метапонимания, формирующегося при участии префронтальной коры,
особенно заметна, когда оно утрачивается. Это как раз тот случай, когда боль-
ные не могут распознать свои собственные ошибки, как мы увидим далее.
7.1. Извлечение из памяти ложной информации
Существуют особенно заметные нарушения извлечения информации из памя-
ти. Исключительно важны случаи конфабуляции — неспособности некоторых
больных с повреждением мозга отчитаться о том, что «воспоминания», которые
они извлекают из памяти, полностью вымышлены. Один такой пример пред-
ставлен в блоке 9.5 .
7.2 . Межполушарная латерализация
при извлечении информации
В свете того, что левое полушарие преимущественно отвечает за речь (гл. 11),
неудивительно, что функции памяти также латерализованы в соответствии
с природой обрабатываемого материала. В рабочей памяти, например, при со-
хранении вербального материала активируются левые области теменной и вен-
тролатеральной префронтальной коры, в то время как сохранение невербаль-
ного материала обычно бывает правосторонним. Области вентролатеральной
префронтальной коры также участвуют и в кодировании информации в долго-
временную память, и при этом, по-видимому, взаимодействуя с медиальной
височной корой. Соответственно, и вентролатеральная префронтальная, и ме-
диальная височная кора при кодировании долговременной памяти демонстри-
руют межполушарную асимметрию (Kelley et al., 1998).
Обучение и память
535
Латерализация может зависеть от использованного материала (вербального,
например), а также от задачи и когнитивного процесса. Эндел Тулвинг с колле-
гами (H���� et al., 2003) выдвинули предположение о том, что обучение, в об-
щем, ассоциируется с более активным участием левой префронтальной коры,
в то время как при извлечении информации из памяти в большей степени при-
нимает участие правая префронтальная кора (рис. 9 .31). Эта модель получила
подтверждение в большом количестве нейровизуализационных исследований.
Некоторые исследователи тем не менее задались вопросом — может ли дан-
ная асимметрия кодирования информации возникать просто в связи с более
высокой зависимостью от вербальных процессов во время обучения по сравне-
нию с извлечением информации. Межполушарная асимметрия, по-видимому,
Блок 9.5 . HW, конфабулирующий больной
HW — 61 -летний больной, который уже более 30 лет живет со своей женой Мартой,
у них четверо детей в возрасте от 22 до 32 лет. Он занимал руководящую должность
на фабрике. У HW аневризма передней соединительной артерии, которая кровос-
HW аневризма передней соединительной артерии, которая кровос-
аневризма передней соединительной артерии, которая кровос-
набжает медиальные отделы лобных долей и базальные отделы переднего мозга.
За хирургическим клипированием аневризмы последовала распространенная дву-
сторонняя ишемия (аноксия) и инфаркт мозговой ткани. Компьютерная аксиальная
томография подтвердила обширное лобное повреждение с недостаточностью меди-
альных и латеральных отделов височных долей.
Такое повреждение мозга привело к проблемам с памятью, отягощенным конфабу-
ляциями и отсутствием осознания этих нарушений. Судя по стандартным тестам,
его умственные способности оставались нормальными. Конфабуляции HW были
спонтанными и, поскольку он верил, что это реальные воспоминания, он пытался
действовать в соответствии с ними, что затрудняло его повседневную жизнь. Напри-
мер, в больнице временами он верил, что он на работе, хотя кругом были кровати,
медсестры и т. п., и он был в больничной одежде. Много раз в 17.00 он собирался
идти домой, и расстраивался и злился, когда ему говорили, что надо остаться.
Ниже приводится отрывок из его беседы с врачом:
Вопрос: Как давно Вы женаты?
Ответ: Около 4 месяцев.
Вопрос: Как зовут Вашу жену?
Ответ: Марта.
Вопрос: Сколько у вас детей?
Ответ: Четверо. (Смеется). Неплохо для четырех месяцев.
Вопрос: Сколько лет Вашим детям?
Ответ: Старшему — 32 . . . младшему — 22 . (Опять смеется).
Вопрос: Как Вы сумели завести четырех детей за четыре месяца?
Ответ: Усыновили.
Вопрос: Кто их усыновил?
Ответ: Мартаия....
Вопрос: Вам не кажется все это немного странным?
Ответ: (Смеется). Да, я согласен, это немного странно.
Вопрос: В Вашей карте записано, что Вы женаты уже более 30 лет. Вам не кажется
это более правильным?
Ответ: Нет.
Вопрос: Вы в самом деле верите, что женаты всего четыре месяца?
Ответ: Да.
536
Глава 9
имеет место в отношении эпизодических воспоминаний, в то время как для
семантической памяти как обучение, так и извлечение более зависимы от меха-
низмов левого полушария.
Результаты недавно проведенного Росси и коллегами исследования (Rossi et
al., 2006) с использованием транскортикальной магнитной стимуляции (ТМС,
см. гл . 4), методики, которая позволяет временно изменить функционирование
коры у здоровых испытуемых, подтверждают предположение Тулвинга. Их
исследование показало, что временное нарушение функции левой префрон-
тальной коры снижает эффективность обучения, определяемую по точности
опознавания. Наоборот, ТМС над правой префронтальной корой снижает эф-
фективность извлечения информации из памяти. Таким образом, по-видимому,
имеется связь между межполушарной латерализацией и различием между
обучением и извлечением информации из памяти.
7.3 . Тета-ритмы могут координировать
извлечение из памяти
Согласно новым литературным данным, во время извлечения информации из
памяти в лобных долях наблюдаются тета-ритмы (5–8 Гц). Глубинные электро-
ды, помещенные в гиппокампы, показывают, что медиальные височные и пре-
фронтальные области могут координироваться при извлечении информации
Рис. 9 .31 . Левое полушарие — для обучения, правое — для извлечения информации
из памяти? Тулвинг с коллегами (H���� et al., 2003) обнаружил, что при эпизодическом
обучении (кодировании) отмечается более высокая активность в левом полушарии, в то
время как при извлечении эпизодического воспоминания наблюдается более высокая
активность в правом полушарии. (Источник: H���� et al, 2003.)
Обучение и память
537
(S��p�s et al., 2005). Эти данные обобщаются на рис. 9 .32, где показано, каким
образом нейрональные сети префронтальных и гиппокампальных структур мо-
гут взаимодействовать для выявления и координации следов памяти при извле-
чении информации.
8.0 . Другие виды обучения
Мы обсудили только некоторые виды памяти. Существуют и другие, опосреду-
емые другими структурами мозга, но о них мы можем только кратко упомянуть
(рис. 9 .33). Например, миндалина опосредует условно-рефлекторную реакцию
страха (см. гл . 14). Мозжечок и базальные ганглии необходимы для формиро-
вания привычных движений и навыков, а также некоторых типов условно-реф-
лекторных реакций. Таламус является одним из важнейших информационных
переключателей мозга, который непрерывно обменивается сигналами с корой.
Поэтому считается, что он принимает участие и в кортикальных механизмах
обучения.
Перцептивное и двигательное обучение включает динамическую организа-
цию и реорганизацию кортикальных карт (гл. 2 и 5). Это называется нервной
пластичностью. Например, потеря пальца изменит представительство пальца
на карте двигательной коры в коре макаки. У людей нейрохирургическое вме-
Рис. 9.32 . Тета-осцилляции могут координировать работу медиальной височной коры
и префронтальной доли во время извлечения информации из памяти. Регулярные ритмы
мозга могут быть предназначены для координирования активности отдельных областей
мозга. Гиппокампальный тета-ритм, как полагают, отражает процессы извлечения из
памяти и координирует работу префронтальной и медиальной височной коры. (Источ-
ник: Je�se�, 2005.)
538
Глава 9
шательство может изменить сенсорные карты тела очень быстро, порой даже
во время операции. Жизнь сама по себе может рассматриваться как процесс по-
стоянного обучения, адаптации и формирования памяти (см. гл . 15). Наконец,
мы теперь знаем, что новые нейроны могут образовываться на протяжении всей
жизни, возникая из стволовых клеток (клеток-предшественников нейронов).
Происходящее замещение некоторых нейронов также включает динамический
процесс обучения и адаптации. У мозга имеется не один, а множество путей
для обучения. И хотя здесь мы сконцентрировали внимание на более стандарт-
ных концепциях обучения и памяти, важно иметь все это в виду.
Рис. 9 .33 . Различные виды памяти опосредуются разными областями мозга. На ри-
сунке представлена обзорная схема множественных систем обучения в мозге. Систе-
ма медиальной височной коры, обсуждавшаяся в этой главе, участвует в обучении и
извлечении из памяти декларативных, эксплицитных воспоминаний; в консолидации
памяти и т. п. Префронтальная кора активируется, по-видимому, в тестах на рабочую
память, при метакогнитивных суждениях о прошлом, включении семантической па-
мяти и концептуальном «прайминге». Перцептивный «прайминг» осуществляется при
участии сенсорной коры, в то время как моторное обучение и некоторые виды классиче-
ского кондиционирования, по-видимому, требуют участия мозжечка. (Источник: M�rr�s
M�sc�v�tch, с разрешения.)
Обучение и память
539
9.0. Заключение
Медиальные отделы височных долей играют ключевую роль в эпизодической
памяти, в которой мы удерживаем информацию об осознанном источнике вос-
поминания. Больные с амнезией вследствие двустороннего повреждения меди-
альных височных долей, соответственно, не могут вспомнить отдельные про-
шлые эпизоды или выучить что-либо новое. Тем не менее у таких пациентов
могут быть сохранены имплицитное обучение и память. У страдающих от се-
мантического дефицита, обычно отмечается повреждение височной доли (осо-
бенно в передней и латеральной частях) и префронтальной коры. Такие паци-
енты с семантической деменцией, например, могут сохранять эпизодическую
память, однако у них отмечаются нарушения при выполнении семантических
задач.
В то время как эксплицитная память оценивается по точности отчетов об
источнике воспоминаний, для оценки имплицитных типов памяти могут потре-
боваться имплицитные же методы, такие как «прайминг» и сенсомоторные за-
дачи. Большая часть нашего обучения происходит имплицитно, например, ов-
ладение языком. Тем не менее важно иметь в виду, что имплицитное научение
требует осознанного и внимательного ориентирования на изучаемые стимулы.
Несознаваемыми при имплицитном научении являются не исходные стимулы,
а логически выведенные закономерности, которые позволяют нам упорядочить
эти стимулы. Сенсомоторные навыки направляются лобной корой при участии
базальных ганглиев и мозжечка. После избыточных упражнений на решение
предсказуемых задач такие приобретенные навыки становятся менее осознан-
ными и, по-видимому, обеспечиваются уже только подкорковыми структурами,
такими, как базальные ганглии.
Рабочая память может быть разделена на зрительную (визуально-простран-
ственный альбом, скечпэд) и вербальную (вербальное повторение, рехэрсэл,
или фонологическая петля). Зачастую делают и дополнительные разделения,
например, на пространственную и зрительную рабочую память.
Полная концепция памяти человека требует участия множества областей
мозга: медиовисочной коры — для эксплицитных эпизодических воспомина-
ний; префронтальной — для метакогнитивной памяти, сохранения и использо-
вания воспоминаний; и сенсорных областей — для перцептивных и сенсорных
воспоминаний. Мозжечок и базальные ганглии во взаимодействии с лобными
долями необходимы для усвоения сенсомоторных навыков. Далее, сенсорные
и моторные половины коры находятся в постоянном взаимодействии друг с
другом, как, например, в случае, когда мы слышим чью-то речь. Наконец, мин-
далина, наряду с ассоциативными зонами лимбической системы, очень тесно
связана с эмоциональным научением.
Разнообразные нарушения памяти могут быть использованы для отделения
различных компонентов памяти друг от друга. Нужно знать некоторые приме-
ры таких нарушений.
540
Глава 9
10.0 . Задания и упражнения к главе 9
1. Заполните недостающие обозначения в функциональной диаграмме, пред-
ставленной на рис. 9 .34. Дайте определение каждому термину.
Рис. 9 .34. Функциональная диаграмма для обучения и памяти
Рис. 9 .35. Соответствующие области
коры мозга
Рис. 9 .36 . Расположение некоторых обла-
стей, ответственных за память
2. Обозначьте на рис. 9 .35 обла-
сти мозга и ассоциированные с
ними типы памяти.
3. Отметьте те области мозга из пред-
ставленных на рис. 9 .36, которые име-
ют отношение к обучению и памяти.
Обучение и память
541
4. Обозначьте на рис. 9 .37 и опишите:
5. Обозначьте сокращения, приведенные на рис. 9 .38. К какой системе они от-
носятся?
Рис. 9 .38 . Области мозга, которые, согласно теории, участвуют в формировании специ-
фических аспектов процесса памяти
Рис. 9 .37. Расположение некоторых областей, ответственных за память
Ɋ
ɂ
ɋ
ɍ
ɇ
Ɉ
Ʉ
1
.
ɉ
ɟ
ɪ
ɜ
ɵ
ɣ
ɩ
ɨ
ɞ
ɯ
ɨ
ɞ
ɤ
ɚ
ɧ
ɚ
ɬ
ɨ
ɦ
ɢ
ɢ
ɦ
ɨ
ɡ
ɝ
ɚ
.
ȼ
ɫ
ɟ
ɝ
ɞ
ɚ
ɧ
ɟ
ɨ
ɛ
ɯ
ɨ
ɞ
ɢ
ɦ
ɨ
ɩ
ɨ
ɦ
ɧ
ɢ
ɬ
ɶ
,
ɤ
ɚ
ɤ
ɪ
ɚ
ɫ
ɩ
ɨ
-
ɥ
ɚ
ɝ
ɚ
ɟ
ɬ
ɫ
ɹ
ɦ
ɨ
ɡ
ɝ
ɜ
ɱ
ɟ
ɪ
ɟ
ɩ
ɟ
.
ɗ
ɬ
ɨ
ɩ
ɟ
ɪ
ɜ
ɵ
ɣ
ɲ
ɚ
ɝ
ɤ
ɩ
ɨ
ɧ
ɢ
ɦ
ɚ
ɧ
ɢ
ɸ
ɩ
ɪ
ɨ
ɫ
ɬ
ɪ
ɚ
ɧ
ɫ
ɬ
ɜ
ɟ
ɧ
ɧ
ɨ
ɝ
ɨ
ɪ
ɚ
ɫ
ɩ
ɨ
ɥ
ɨ
ɠ
ɟ
ɧ
ɢ
ɹ
ɤ
ɨ
ɪ
ɵ
,
ɤ
ɨ
ɬ
ɨ
ɪ
ɚ
ɹ
ɩ
ɨ
ɱ
ɬ
ɢ
ɧ
ɚ
8
5
%
ɫ
ɨ
ɫ
ɬ
ɨ
ɢ
ɬ
ɢ
ɡ
ɛ
ɟ
ɥ
ɨ
ɝ
ɨ
ɜ
ɟ
ɳ
ɟ
ɫ
ɬ
ɜ
ɚ
,
ɡ
ɚ
ɳ
ɢ
ɳ
ɟ
ɧ
ɧ
ɵ
ɯ
«
ɦ
ɚ
ɝ
ɢ
ɫ
ɬ
ɪ
ɚ
ɥ
ɟ
ɣ
»
,
ɫ
ɜ
ɹ
ɡ
ɵ
ɜ
ɚ
ɸ
ɳ
ɢ
ɯ
ɜ
ɫ
ɟ
ɝ
ɥ
ɚ
ɜ
ɧ
ɵ
ɟ
ɨ
ɛ
ɥ
ɚ
ɫ
ɬ
ɢ
ɦ
ɨ
ɡ
ɝ
ɚ
ɞ
ɪ
ɭ
ɝ
ɫ
ɞ
ɪ
ɭ
ɝ
ɨ
ɦ
.
ȼ
ɧ
ɢ
ɡ
ɭ
ɫ
ɩ
ɪ
ɚ
ɜ
ɚ
:
ɤ
ɨ
ɪ
ɚ
ɪ
ɚ
ɫ
ɩ
ɨ
ɥ
ɚ
ɝ
ɚ
ɟ
ɬ
ɫ
ɹ
ɫ
ɜ
ɟ
ɪ
ɯ
ɭ
ɧ
ɚ
ɞ
ɫ
ɬ
ɜ
ɨ
ɥ
ɨ
ɦ
ɦ
ɨ
ɡ
ɝ
ɚ
ɢ
ɩ
ɨ
ɞ
ɤ
ɨ
ɪ
ɤ
ɨ
ɜ
ɵ
ɦ
ɢ
ɡ
ɨ
ɧ
ɚ
ɦ
ɢ
,
ɤ
ɨ
ɬ
ɨ
ɪ
ɵ
ɟ
ɜ
ɧ
ɢ
ɡ
ɭ
ɫ
ɨ
ɟ
ɞ
ɢ
ɧ
ɹ
ɸ
ɬ
ɫ
ɹ
ɫ
ɨ
ɫ
ɩ
ɢ
ɧ
ɧ
ɵ
ɦ
ɦ
ɨ
ɡ
ɝ
ɨ
ɦ
.
ɋ
ɜ
ɹ
ɡ
ɚ
ɧ
ɧ
ɚ
ɹ
ɫ
ɫ
ɨ
ɛ
ɵ
ɬ
ɢ
ɟ
ɦ
ɗ
ɗ
Ƚ
,
ɡ
ɚ
ɪ
ɟ
ɝ
ɢ
ɫ
ɬ
ɪ
ɢ
ɪ
ɨ
ɜ
ɚ
ɧ
ɧ
ɚ
ɹ
ɭ
ɦ
ɨ
ɥ
ɨ
ɞ
ɨ
ɣ
ɞ
ɟ
ɜ
ɭ
ɲ
ɤ
ɢ
(
ɚ
)
,
ɹ
ɜ
ɥ
ɹ
ɟ
ɬ
ɫ
ɹ
ɧ
ɚ
ɩ
ɨ
ɦ
ɢ
ɧ
ɚ
ɧ
ɢ
ɟ
ɦ
ɨ
ɬ
ɨ
ɦ
,
ɱ
ɬ
ɨ
ɫ
ɭ
ɳ
ɟ
ɫ
ɬ
ɜ
ɭ
ɟ
ɬ
ɩ
ɨ
ɫ
ɬ
ɨ
ɹ
ɧ
ɧ
ɵ
ɣ
,
ɦ
ɟ
ɧ
ɹ
ɸ
ɳ
ɢ
ɣ
ɫ
ɹ
ɨ
ɝ
ɪ
ɨ
ɦ
ɧ
ɵ
ɣ
ɩ
ɨ
ɬ
ɨ
ɤ
ɫ
ɢ
ɝ
ɧ
ɚ
ɥ
ɨ
ɜ
ɜ
ɦ
ɨ
ɡ
ɝ
ɟ
,
ɤ
ɨ
ɬ
ɨ
ɪ
ɵ
ɣ
ɦ
ɨ
ɠ
ɧ
ɨ
ɡ
ɚ
ɪ
ɟ
ɝ
ɢ
ɫ
ɬ
ɪ
ɢ
ɪ
ɨ
ɜ
ɚ
ɬ
ɶ
ɜ
ɩ
ɨ
ɜ
ɟ
ɪ
ɯ
ɧ
ɨ
ɫ
ɬ
ɧ
ɨ
ɣ
ɗ
ɗ
Ƚ
Ɋ
ɂ
ɋ
ɍ
ɇ
Ɉ
Ʉ
2
.
ɂ
ɫ
ɩ
ɨ
ɥ
ɶ
ɡ
ɭ
ɣ
ɬ
ɟ
ɢ
ɡ
ɨ
ɛ
ɪ
ɚ
ɠ
ɟ
ɧ
ɢ
ɟ
ɝ
ɨ
ɥ
ɨ
ɜ
ɵ
ɱ
ɟ
ɥ
ɨ
ɜ
ɟ
ɤ
ɚ
(
ɧ
ɚ
ɜ
ɟ
ɪ
ɯ
ɭ
ɫ
ɥ
ɟ
ɜ
ɚ
)
ɞ
ɥ
ɹ
ɩ
ɪ
ɚ
ɜ
ɢ
ɥ
ɶ
ɧ
ɨ
ɣ
ɨ
ɪ
ɢ
ɟ
ɧ
ɬ
ɚ
ɰ
ɢ
ɢ
.
Ɇ
ɵ
ɦ
ɨ
ɠ
ɟ
ɦ
ɜ
ɢ
ɞ
ɟ
ɬ
ɶ
ɦ
ɨ
ɡ
ɨ
ɥ
ɢ
ɫ
ɬ
ɨ
ɟ
ɬ
ɟ
ɥ
ɨ
ɫ
ɥ
ɟ
ɜ
ɨ
ɣ
ɫ
ɬ
ɨ
ɪ
ɨ
ɧ
ɵ
.
Ɉ
ɛ
ɪ
ɚ
ɬ
ɢ
ɬ
ɟ
ɜ
ɧ
ɢ
ɦ
ɚ
ɧ
ɢ
ɟ
ɧ
ɚ
ɬ
ɪ
ɢ
ɨ
ɫ
ɧ
ɨ
ɜ
ɧ
ɵ
ɟ
ɩ
ɥ
ɨ
ɫ
ɤ
ɨ
ɫ
ɬ
ɢ
ɫ
ɟ
ɱ
ɟ
ɧ
ɢ
ɹ
ɦ
ɨ
ɡ
ɝ
ɚ
Ɋ
ɂ
ɋ
ɍ
ɇ
Ɉ
Ʉ
3
.
ɋ
ɩ
ɟ
ɰ
ɢ
ɚ
ɥ
ɢ
ɡ
ɚ
ɰ
ɢ
ɹ
ɨ
ɛ
ɥ
ɚ
ɫ
ɬ
ɟ
ɣ
ɦ
ɨ
ɡ
ɝ
ɚ
,
ɨ
ɛ
ɲ
ɢ
ɪ
ɧ
ɨ
ɟ
ɩ
ɨ
ɤ
ɪ
ɵ
ɬ
ɢ
ɟ
ɦ
ɨ
ɡ
ɝ
ɚ
.
Ʉ
ɨ
ɪ
ɚ
ɩ
ɪ
ɟ
ɞ
ɫ
ɬ
ɚ
ɜ
-
ɥ
ɹ
ɟ
ɬ
ɫ
ɨ
ɛ
ɨ
ɣ
ɩ
ɥ
ɨ
ɫ
ɤ
ɢ
ɣ
ɥ
ɢ
ɫ
ɬ
,
ɦ
ɧ
ɨ
ɝ
ɨ
ɤ
ɪ
ɚ
ɬ
ɧ
ɨ
ɫ
ɥ
ɨ
ɠ
ɟ
ɧ
ɧ
ɵ
ɣ
ɜ
ɜ
ɟ
ɪ
ɯ
ɧ
ɟ
ɣ
ɱ
ɚ
ɫ
ɬ
ɢ
ɱ
ɟ
ɪ
ɟ
ɩ
ɚ
.
Ɉ
ɛ
ɪ
ɚ
ɬ
ɢ
ɬ
ɟ
ɜ
ɧ
ɢ
-
ɦ
ɚ
ɧ
ɢ
ɟ
ɧ
ɚ
ɪ
ɚ
ɫ
ɤ
ɪ
ɚ
ɲ
ɟ
ɧ
ɧ
ɵ
ɟ
ɨ
ɛ
ɥ
ɚ
ɫ
ɬ
ɢ
,
ɨ
ɬ
ɪ
ɚ
ɠ
ɚ
ɸ
ɳ
ɢ
ɟ
ɨ
ɫ
ɧ
ɨ
ɜ
ɧ
ɵ
ɟ
ɮ
ɭ
ɧ
ɤ
ɰ
ɢ
ɢ
ɦ
ɨ
ɡ
ɝ
ɚ
.
ɗ
ɬ
ɢ
ɱ
ɚ
ɫ
ɬ
ɢ
ɤ
ɨ
ɪ
ɵ
ɨ
ɛ
ɟ
ɫ
ɩ
ɟ
ɱ
ɢ
ɜ
ɚ
ɸ
ɬ
ɨ
ɩ
ɪ
ɟ
ɞ
ɟ
ɥ
ɟ
ɧ
ɧ
ɵ
ɟ
ɫ
ɨ
ɫ
ɬ
ɚ
ɜ
ɧ
ɵ
ɟ
ɱ
ɚ
ɫ
ɬ
ɢ
ɫ
ɨ
ɡ
ɧ
ɚ
ɬ
ɟ
ɥ
ɶ
ɧ
ɨ
ɝ
ɨ
ɨ
ɩ
ɵ
ɬ
ɚ
.
Ɂ
ɚ
ɞ
ɧ
ɢ
ɟ
ɱ
ɚ
ɫ
ɬ
ɢ
ɨ
ɬ
ɜ
ɟ
ɱ
ɚ
ɸ
ɬ
ɧ
ɚ
ɫ
ɟ
ɧ
ɫ
ɨ
ɪ
ɧ
ɨ
ɟ
ɜ
ɨ
ɫ
ɩ
ɪ
ɢ
ɹ
ɬ
ɢ
ɟ
,
ɩ
ɟ
ɪ
ɟ
ɞ
ɧ
ɹ
ɹ
ɩ
ɨ
ɥ
ɨ
ɜ
ɢ
ɧ
ɚ
—
ɡ
ɚ
ɦ
ɨ
ɬ
ɨ
ɪ
ɧ
ɭ
ɸ
ɢ
«
ɧ
ɚ
ɩ
ɪ
ɚ
ɜ
ɥ
ɟ
ɧ
ɧ
ɭ
ɸ
ɜ
ɛ
ɭ
ɞ
ɭ
ɳ
ɟ
ɟ
»
ɞ
ɟ
ɹ
ɬ
ɟ
ɥ
ɶ
ɧ
ɨ
ɫ
ɬ
ɶ
:
ɫ
ɨ
ɡ
ɧ
ɚ
ɧ
ɢ
ɟ
,
ɪ
ɚ
ɛ
ɨ
ɱ
ɚ
ɹ
ɩ
ɚ
ɦ
ɹ
ɬ
ɶ
,
ɩ
ɥ
ɚ
ɧ
ɢ
ɪ
ɨ
ɜ
ɚ
ɧ
ɢ
ɟ
,
ɩ
ɪ
ɢ
ɧ
ɹ
ɬ
ɢ
ɟ
ɪ
ɟ
ɲ
ɟ
ɧ
ɢ
ɣ
Ɋ
ɂ
ɋ
ɍ
ɇ
Ɉ
Ʉ
4
.
ɇ
ɟ
ɣ
ɪ
ɨ
ɧ
ɵ
ɧ
ɚ
ɩ
ɨ
ɥ
ɧ
ɹ
ɸ
ɬ
ɫ
ɨ
ɛ
ɨ
ɣ
ɬ
ɟ
ɥ
ɨ
,
ɢ
ɫ
ɩ
ɢ
ɧ
ɧ
ɨ
ɣ
ɦ
ɨ
ɡ
ɝ
ɹ
ɜ
ɥ
ɹ
ɟ
ɬ
ɫ
ɹ
ɝ
ɥ
ɚ
ɜ
ɧ
ɨ
ɣ
ɦ
ɚ
ɝ
ɢ
ɫ
ɬ
ɪ
ɚ
-
ɥ
ɶ
ɸ
,
ɫ
ɜ
ɹ
ɡ
ɵ
ɜ
ɚ
ɸ
ɳ
ɟ
ɣ
ɝ
ɨ
ɥ
ɨ
ɜ
ɧ
ɨ
ɣ
ɦ
ɨ
ɡ
ɝ
ɢ
ɜ
ɫ
ɟ
ɬ
ɟ
ɥ
ɨ
.
ɋ
ɥ
ɟ
ɜ
ɚ
ɧ
ɚ
ɪ
ɢ
ɫ
ɭ
ɧ
ɤ
ɟ
ɦ
ɵ
ɦ
ɨ
ɠ
ɟ
ɦ
ɜ
ɢ
ɞ
ɟ
ɬ
ɶ
ɦ
ɟ
ɞ
ɢ
ɚ
ɥ
ɶ
-
ɧ
ɭ
ɸ
ɩ
ɪ
ɚ
ɜ
ɭ
ɸ
ɩ
ɨ
ɜ
ɟ
ɪ
ɯ
ɧ
ɨ
ɫ
ɬ
ɶ
ɦ
ɨ
ɡ
ɝ
ɚ
.
Ɉ
ɛ
ɪ
ɚ
ɬ
ɢ
ɬ
ɟ
ɜ
ɧ
ɢ
ɦ
ɚ
ɧ
ɢ
ɟ
ɧ
ɚ
ɨ
ɛ
ɳ
ɟ
ɩ
ɪ
ɢ
ɧ
ɹ
ɬ
ɵ
ɟ
ɨ
ɛ
ɨ
ɡ
ɧ
ɚ
ɱ
ɟ
ɧ
ɢ
ɹ
ɧ
ɚ
ɩ
ɪ
ɚ
ɜ
-
ɥ
ɟ
ɧ
ɢ
ɹ
,
ɤ
ɨ
ɬ
ɨ
ɪ
ɵ
ɟ
ɤ
ɚ
ɤ
ɋ
ɟ
ɜ
ɟ
ɪ
,
ɘ
ɝ
,
ȼ
ɨ
ɫ
ɬ
ɨ
ɤ
ɢ
Ɂ
ɚ
ɩ
ɚ
ɞ
ɜ
ɝ
ɟ
ɨ
ɝ
ɪ
ɚ
ɮ
ɢ
ɢ
.
ɇ
ɚ
ɢ
ɛ
ɨ
ɥ
ɟ
ɟ
ɩ
ɪ
ɨ
ɫ
ɬ
ɵ
ɟ
ɬ
ɟ
ɪ
ɦ
ɢ
ɧ
ɵ
ɧ
ɚ
ɩ
ɢ
ɫ
ɚ
ɧ
ɵ
ɫ
ɜ
ɟ
ɪ
ɯ
ɭ
(
ȼ
ɜ
ɟ
ɪ
ɯ
,
ȼ
ɧ
ɢ
ɡ
,
ɢ
ɬ
.
ɞ
.
)
.
ɇ
ɨ
ɚ
ɧ
ɚ
ɬ
ɨ
ɦ
ɵ
ɜ
ɫ
ɟ
ɝ
ɞ
ɚ
ɢ
ɫ
ɩ
ɨ
ɥ
ɶ
ɡ
ɭ
ɸ
ɬ
ɬ
ɟ
ɪ
ɦ
ɢ
ɧ
ɵ
ɥ
ɚ
ɬ
ɢ
ɧ
ɫ
ɤ
ɨ
ɝ
ɨ
ɩ
ɪ
ɨ
ɢ
ɫ
ɯ
ɨ
ɠ
ɞ
ɟ
ɧ
ɢ
ɹ
.
ɂ
ɫ
ɩ
ɨ
ɥ
ɶ
ɡ
ɭ
ɣ
ɬ
ɟ
ɞ
ɚ
ɧ
ɧ
ɵ
ɣ
ɪ
ɢ
ɫ
ɭ
ɧ
ɨ
ɤ
ɞ
ɥ
ɹ
ɫ
ɩ
ɪ
ɚ
ɜ
ɤ
ɢ
,
ɟ
ɫ
ɥ
ɢ
ɜ
ɚ
ɦ
ɩ
ɨ
ɧ
ɚ
ɞ
ɨ
ɛ
ɢ
ɬ
ɫ
ɹ
ɩ
ɨ
ɧ
ɹ
ɬ
ɶ
ɪ
ɚ
ɫ
ɩ
ɨ
ɥ
ɨ
ɠ
ɟ
ɧ
ɢ
ɟ
ɦ
ɨ
ɡ
ɝ
ɚ
ɧ
ɚ
ɪ
ɢ
ɫ
ɭ
ɧ
ɤ
ɚ
ɯ
ɜ
ɷ
ɬ
ɨ
ɣ
ɤ
ɧ
ɢ
ɝ
ɟ
Ɋ
ɂ
ɋ
ɍ
ɇ
Ɉ
Ʉ
5
.
ɋ
ɧ
ɨ
ɜ
ɚ
ɨ
ɛ
ɪ
ɚ
ɬ
ɢ
ɬ
ɟ
ɜ
ɧ
ɢ
ɦ
ɚ
ɧ
ɢ
ɟ
ɧ
ɚ
ɪ
ɚ
ɫ
ɩ
ɨ
ɥ
ɨ
ɠ
ɟ
ɧ
ɢ
ɟ
ɦ
ɨ
ɡ
ɝ
ɚ
ɜ
ɝ
ɨ
ɥ
ɨ
ɜ
ɟ
ɢ
ɩ
ɨ
ɫ
ɦ
ɨ
ɬ
ɪ
ɢ
ɬ
ɟ
,
ɤ
ɚ
ɤ
ɞ
ɜ
ɚ
ɝ
ɢ
ɩ
ɩ
ɨ
ɤ
ɚ
ɦ
ɩ
ɚ
ɜ
ɫ
ɬ
ɪ
ɨ
ɟ
ɧ
ɵ
ɜ
ɧ
ɭ
ɬ
ɪ
ɶ
ɤ
ɚ
ɠ
ɞ
ɨ
ɣ
ɢ
ɡ
ɞ
ɜ
ɭ
ɯ
ɜ
ɢ
ɫ
ɨ
ɱ
ɧ
ɵ
ɯ
ɞ
ɨ
ɥ
ɟ
ɣ
.
ȼ
ɟ
ɪ
ɬ
ɢ
ɤ
ɚ
ɥ
ɶ
ɧ
ɵ
ɣ
ɫ
ɪ
ɟ
ɡ
ɨ
ɬ
ɭ
ɯ
ɚ
ɤ
ɭ
ɯ
ɭ
(
ɤ
ɨ
ɪ
ɨ
ɧ
ɚ
ɪ
ɧ
ɵ
ɣ
)
ɩ
ɨ
ɤ
ɚ
ɡ
ɵ
ɜ
ɚ
ɟ
ɬ
ɝ
ɢ
ɩ
ɩ
ɨ
ɤ
ɚ
ɦ
ɩ
ɬ
ɨ
ɥ
ɶ
ɤ
ɨ
ɤ
ɚ
ɤ
ɦ
ɚ
ɥ
ɟ
ɧ
ɶ
ɤ
ɢ
ɟ
ɨ
ɤ
ɪ
ɭ
ɝ
ɥ
ɵ
ɟ
ɞ
ɢ
ɫ
ɤ
ɢ
(
ɤ
ɪ
ɚ
ɫ
ɧ
ɵ
ɟ
)
.
ȿ
ɫ
ɥ
ɢ
ɜ
ɵ
ɩ
ɪ
ɟ
ɞ
ɫ
ɬ
ɚ
ɜ
ɢ
ɬ
ɟ
ɡ
ɚ
ɩ
ɪ
ɨ
ɤ
ɢ
ɧ
ɭ
ɬ
ɭ
ɸ
ɧ
ɚ
ɡ
ɚ
ɞ
ɝ
ɨ
ɥ
ɨ
ɜ
ɭ
(
ɜ
ɧ
ɢ
ɡ
ɭ
ɪ
ɢ
ɫ
ɭ
ɧ
ɤ
ɚ
)
,
ɜ
ɵ
ɦ
ɨ
ɠ
ɟ
ɬ
ɟ
ɭ
ɜ
ɢ
ɞ
ɟ
ɬ
ɶ
ɦ
ɟ
ɞ
ɢ
ɚ
ɥ
ɶ
ɧ
ɭ
ɸ
ɜ
ɢ
ɫ
ɨ
ɱ
ɧ
ɭ
ɸ
ɞ
ɨ
ɥ
ɸ
(
Ɇ
ȼ
Ⱦ
)
,
ɤ
ɨ
ɬ
ɨ
ɪ
ɚ
ɹ
ɫ
ɨ
ɞ
ɟ
ɪ
ɠ
ɢ
ɬ
ɞ
ɜ
ɚ
ɝ
ɢ
ɩ
ɩ
ɨ
ɤ
ɚ
ɦ
ɩ
ɚ
.
ɗ
ɬ
ɢ
ɫ
ɬ
ɪ
ɭ
ɤ
-
ɬ
ɭ
ɪ
ɵ
ɱ
ɪ
ɟ
ɡ
ɜ
ɵ
ɱ
ɚ
ɣ
ɧ
ɨ
ɜ
ɚ
ɠ
ɧ
ɵ
ɞ
ɥ
ɹ
ɷ
ɦ
ɨ
ɰ
ɢ
ɣ
,
ɡ
ɪ
ɟ
ɧ
ɢ
ɹ
ɢ
ɩ
ɚ
ɦ
ɹ
ɬ
ɢ
Ɋ
ɂ
ɋ
ɍ
ɇ
Ɉ
Ʉ
6
.
ȼ
ɜ
ɟ
ɪ
ɯ
ɭ
ɫ
ɥ
ɟ
ɜ
ɚ
:
ɩ
ɪ
ɨ
ɜ
ɨ
ɞ
ɹ
ɳ
ɢ
ɟ
ɬ
ɪ
ɚ
ɤ
ɬ
ɵ
ɢ
ɡ
ɛ
ɟ
ɥ
ɨ
ɝ
ɨ
ɜ
ɟ
ɳ
ɟ
ɫ
ɬ
ɜ
ɚ
,
ɤ
ɚ
ɤ
«
ɮ
ɨ
ɧ
ɬ
ɚ
ɧ
»
ɩ
ɪ
ɨ
ɜ
ɨ
ɞ
ɹ
-
ɳ
ɢ
ɯ
ɜ
ɨ
ɥ
ɨ
ɤ
ɨ
ɧ
,
ɛ
ɟ
ɪ
ɭ
ɬ
ɫ
ɜ
ɨ
ɟ
ɧ
ɚ
ɱ
ɚ
ɥ
ɨ
ɨ
ɬ
ɬ
ɚ
ɥ
ɚ
ɦ
ɭ
ɫ
ɚ
.
Ⱦ
ɪ
ɭ
ɝ
ɢ
ɟ
ɩ
ɪ
ɨ
ɜ
ɨ
ɞ
ɹ
ɳ
ɢ
ɟ
ɩ
ɭ
ɬ
ɢ
ɢ
ɞ
ɭ
ɬ
ɜ
ɨ
ɜ
ɫ
ɟ
ɯ
ɝ
ɥ
ɚ
ɜ
ɧ
ɵ
ɯ
ɧ
ɚ
ɩ
ɪ
ɚ
ɜ
ɥ
ɟ
ɧ
ɢ
ɹ
ɯ
.
ȼ
ɧ
ɢ
ɡ
ɭ
ɢ
ɫ
ɩ
ɪ
ɚ
ɜ
ɚ
ɩ
ɨ
ɤ
ɚ
ɡ
ɚ
ɧ
ɵ
ɬ
ɪ
ɚ
ɤ
ɬ
ɨ
ɝ
ɪ
ɚ
ɦ
ɦ
ɵ
,
ɫ
ɩ
ɟ
ɰ
ɢ
ɚ
ɥ
ɶ
ɧ
ɵ
ɟ
Ɇ
Ɋ
Ɍ
-
ɢ
ɡ
ɨ
ɛ
ɪ
ɚ
ɠ
ɟ
ɧ
ɢ
ɹ
,
ɤ
ɨ
ɬ
ɨ
ɪ
ɵ
ɟ
ɩ
ɨ
ɤ
ɚ
ɡ
ɵ
ɜ
ɚ
ɸ
ɬ
ɨ
ɫ
ɧ
ɨ
ɜ
ɧ
ɵ
ɟ
ɬ
ɪ
ɚ
ɤ
ɬ
ɵ
ɦ
ɟ
ɠ
ɞ
ɭ
ɝ
ɥ
ɚ
ɜ
ɧ
ɵ
ɦ
ɢ
ɨ
ɛ
ɥ
ɚ
ɫ
ɬ
ɹ
ɦ
ɢ
ɦ
ɨ
ɡ
ɝ
ɚ
.
ɉ
ɨ
ɱ
ɬ
ɢ
ɜ
ɫ
ɟ
ɹ
ɜ
ɥ
ɹ
-
ɸ
ɬ
ɫ
ɹ
ɞ
ɜ
ɭ
ɫ
ɬ
ɨ
ɪ
ɨ
ɧ
ɧ
ɢ
ɦ
ɢ
.
ɂ
ɫ
ɬ
ɨ
ɱ
ɧ
ɢ
ɤ
:
(
ɜ
ɜ
ɟ
ɪ
ɯ
ɭ
ɫ
ɥ
ɟ
ɜ
ɚ
)
I
z
h
i
k
e
v
i
c
h
a
n
d
E
d
e
l
m
a
n
,
2
0
0
8
;
(
ɫ
ɩ
ɪ
ɚ
ɜ
ɚ
)
H
o
f
e
r
a
n
d
F
r
a
h
m
,
2
0
0
6
Ɋ
ɂ
ɋ
ɍ
ɇ
Ɉ
Ʉ
7
.
Ɍ
ɟ
ɥ
ɨ
ɢ
ɦ
ɨ
ɡ
ɝ
ɱ
ɪ
ɟ
ɡ
ɜ
ɵ
ɱ
ɚ
ɣ
ɧ
ɨ
ɬ
ɟ
ɫ
ɧ
ɨ
ɫ
ɜ
ɹ
ɡ
ɚ
ɧ
ɵ
.
ȼ
ɜ
ɟ
ɪ
ɯ
ɭ
ɫ
ɥ
ɟ
ɜ
ɚ
ɩ
ɨ
ɤ
ɚ
ɡ
ɚ
ɧ
ɦ
ɨ
ɡ
ɝ
ɫ
ɦ
ɟ
ɞ
ɢ
-
ɚ
ɥ
ɶ
ɧ
ɨ
ɣ
ɫ
ɬ
ɨ
ɪ
ɨ
ɧ
ɵ
(
ɫ
ɦ
ɨ
ɬ
ɪ
ɢ
ɬ
ɜ
ɩ
ɪ
ɚ
ɜ
ɨ
)
,
ɬ
ɟ
ɫ
ɧ
ɨ
ɜ
ɡ
ɚ
ɢ
ɦ
ɨ
ɫ
ɜ
ɹ
ɡ
ɚ
ɧ
ɧ
ɵ
ɣ
ɫ
ɷ
ɧ
ɞ
ɨ
ɤ
ɪ
ɢ
ɧ
ɧ
ɨ
ɣ
(
ɝ
ɨ
ɪ
ɦ
ɨ
ɧ
ɚ
ɥ
ɶ
ɧ
ɨ
ɣ
)
ɫ
ɢ
ɫ
ɬ
ɟ
ɦ
ɨ
ɣ
,
ɢ
ɦ
ɦ
ɭ
ɧ
ɧ
ɨ
ɣ
ɪ
ɟ
ɝ
ɭ
ɥ
ɹ
ɰ
ɢ
ɟ
ɣ
ɢ
ɚ
ɜ
ɬ
ɨ
ɧ
ɨ
ɦ
ɧ
ɨ
ɣ
ɪ
ɟ
ɝ
ɭ
ɥ
ɹ
ɰ
ɢ
ɟ
ɣ
(
ɫ
ɟ
ɪ
ɞ
ɰ
ɟ
,
ɥ
ɟ
ɝ
ɤ
ɢ
ɟ
,
ɠ
ɟ
ɥ
ɭ
ɞ
ɨ
ɤ
,
ɤ
ɢ
-
ɲ
ɟ
ɱ
ɧ
ɢ
ɤ
ɢ
ɤ
ɪ
ɨ
ɜ
ɟ
ɧ
ɨ
ɫ
ɧ
ɵ
ɟ
ɫ
ɨ
ɫ
ɭ
ɞ
ɵ
)
.
ɍ
ɱ
ɟ
ɥ
ɨ
ɜ
ɟ
ɤ
ɚ
ɧ
ɟ
ɬ
ɩ
ɪ
ɨ
ɢ
ɡ
ɜ
ɨ
ɥ
ɶ
ɧ
ɨ
ɝ
ɨ
ɤ
ɨ
ɧ
ɬ
ɪ
ɨ
ɥ
ɹ
ɧ
ɚ
ɞ
ɷ
ɬ
ɢ
ɦ
ɢ
ɮ
ɭ
ɧ
ɤ
-
ɰ
ɢ
ɹ
ɦ
ɢ
,
ɜ
ɨ
ɬ
ɥ
ɢ
ɱ
ɢ
ɟ
ɨ
ɬ
ɫ
ɤ
ɟ
ɥ
ɟ
ɬ
ɧ
ɨ
ɣ
ɦ
ɭ
ɫ
ɤ
ɭ
ɥ
ɚ
ɬ
ɭ
ɪ
ɵ
ɬ
ɟ
ɥ
ɚ
ɢ
ɝ
ɨ
ɥ
ɨ
ɜ
ɵ
.
ɂ
ɫ
ɬ
ɨ
ɱ
ɧ
ɢ
ɤ
:
L
a
n
e
a
n
d
W
a
g
e
r
,
2
0
0
9
Ɋ
ɂ
ɋ
ɍ
ɇ
Ɉ
Ʉ
8
.
Ȼ
ɨ
ɥ
ɟ
ɟ
ɲ
ɢ
ɪ
ɨ
ɤ
ɢ
ɣ
ɜ
ɢ
ɞ
.
ȼ
ɜ
ɟ
ɪ
ɯ
ɭ
ɫ
ɥ
ɟ
ɜ
ɚ
ɜ
ɵ
ɦ
ɨ
ɠ
ɟ
ɬ
ɟ
ɜ
ɢ
ɞ
ɟ
ɬ
ɶ
ɱ
ɟ
ɥ
ɨ
ɜ
ɟ
ɤ
ɚ
,
ɫ
ɦ
ɨ
ɬ
ɪ
ɹ
ɳ
ɟ
ɝ
ɨ
ɧ
ɚ
ɜ
ɪ
ɚ
ɳ
ɚ
ɸ
ɳ
ɢ
ɣ
ɫ
ɹ
ɡ
ɪ
ɢ
ɬ
ɟ
ɥ
ɶ
ɧ
ɵ
ɣ
ɫ
ɬ
ɢ
ɦ
ɭ
ɥ
,
ɤ
ɨ
ɬ
ɨ
ɪ
ɵ
ɣ
ɜ
ɵ
ɡ
ɵ
ɜ
ɚ
ɟ
ɬ
ɨ
ɛ
ɲ
ɢ
ɪ
ɧ
ɭ
ɸ
ɚ
ɤ
ɬ
ɢ
ɜ
ɚ
ɰ
ɢ
ɸ
ɨ
ɛ
ɥ
ɚ
ɫ
ɬ
ɟ
ɣ
ɦ
ɨ
ɡ
ɝ
ɚ
,
ɧ
ɚ
ɱ
ɢ
ɧ
ɚ
ɹ
ɫ
ɩ
ɨ
ɥ
ɹ
V
1
(
ɬ
ɟ
ɦ
ɧ
ɨ
-
ɫ
ɢ
ɧ
ɢ
ɣ
)
.
ɉ
ɨ
ɥ
ɟ
V
1
,
ɪ
ɚ
ɡ
ɦ
ɟ
ɪ
ɨ
ɦ
ɫ
ɛ
ɚ
ɧ
ɤ
ɨ
ɜ
ɫ
ɤ
ɭ
ɸ
ɤ
ɚ
ɪ
ɬ
ɭ
,
ɫ
ɥ
ɨ
ɠ
ɟ
ɧ
ɨ
ɜ
ɧ
ɭ
ɬ
ɪ
ɶ
ɡ
ɚ
ɬ
ɵ
ɥ
ɨ
ɱ
ɧ
ɨ
ɣ
ɞ
ɨ
ɥ
ɢ
.
ȼ
ɧ
ɢ
ɡ
ɭ
ɞ
ɥ
ɹ
ɫ
ɪ
ɚ
ɜ
ɧ
ɟ
ɧ
ɢ
ɹ
ɩ
ɨ
ɤ
ɚ
ɡ
ɚ
ɧ
ɦ
ɨ
ɡ
ɝ
ɩ
ɥ
ɨ
ɞ
ɨ
ɜ
ɨ
ɣ
ɦ
ɭ
ɲ
ɤ
ɢ
,
ɫ
ɨ
ɞ
ɟ
ɪ
ɠ
ɚ
ɳ
ɢ
ɣ
ɨ
ɤ
ɨ
ɥ
ɨ
1
0
0
0
0
0
ɧ
ɟ
ɣ
ɪ
ɨ
ɧ
ɨ
ɜ
.
Ɉ
ɞ
ɢ
ɧ
ɨ
ɱ
ɧ
ɵ
ɣ
ɧ
ɟ
ɣ
ɪ
ɨ
ɧ
ɩ
ɨ
ɤ
ɚ
ɡ
ɚ
ɧ
ɫ
ɩ
ɪ
ɚ
ɜ
ɚ
,
ɚ
ɪ
ɹ
ɞ
ɨ
ɦ
ɫ
ɧ
ɢ
ɦ
—
ɨ
ɞ
ɢ
ɧ
ɨ
ɱ
ɧ
ɚ
ɹ
ɨ
ɪ
ɝ
ɚ
ɧ
ɢ
ɱ
ɟ
ɫ
ɤ
ɚ
ɹ
ɦ
ɨ
ɥ
ɟ
ɤ
ɭ
ɥ
ɚ
ɚ
ɦ
ɢ
ɧ
ɨ
ɤ
ɢ
ɫ
ɥ
ɨ
ɬ
ɵ
(
ɢ
ɦ
ɟ
ɟ
ɬ
ɚ
ɦ
ɢ
ɧ
ɨ
ɝ
ɪ
ɭ
ɩ
ɩ
ɭ
(
N
H
)
ɢ
ɤ
ɚ
ɪ
ɛ
ɨ
ɤ
ɫ
ɢ
ɥ
ɶ
ɧ
ɭ
ɸ
ɝ
ɪ
ɭ
ɩ
ɩ
ɭ
(
C
O
O
H
)
)
.
ȼ
ɫ
ɟ
ɜ
ɟ
ɪ
ɲ
ɢ
ɧ
ɵ
ɜ
ɮ
ɨ
ɪ
ɦ
ɭ
ɥ
ɟ
ɨ
ɛ
ɨ
ɡ
ɧ
ɚ
ɱ
ɚ
ɸ
ɬ
ɚ
ɬ
ɨ
ɦ
ɵ
ɭ
ɝ
ɥ
ɟ
ɪ
ɨ
ɞ
ɚ
.
R
—
ɛ
ɨ
ɤ
ɨ
ɜ
ɚ
ɹ
ɰ
ɟ
ɩ
ɶ
,
ɤ
ɨ
ɬ
ɨ
-
ɪ
ɚ
ɹ
ɦ
ɨ
ɠ
ɟ
ɬ
ɢ
ɦ
ɟ
ɬ
ɶ
ɪ
ɚ
ɡ
ɧ
ɨ
ɟ
ɫ
ɬ
ɪ
ɨ
ɟ
ɧ
ɢ
ɟ
.
ȼ
ɫ
ɟ
ɭ
ɪ
ɨ
ɜ
ɧ
ɢ
ɚ
ɧ
ɚ
ɥ
ɢ
ɡ
ɚ
ɨ
ɬ
ɦ
ɨ
ɥ
ɟ
ɤ
ɭ
ɥ
ɵ
ɞ
ɨ
ɦ
ɚ
ɤ
ɪ
ɨ
ɫ
ɤ
ɨ
ɩ
ɢ
ɱ
ɟ
ɫ
ɤ
ɨ
ɣ
ɚ
ɧ
ɚ
ɬ
ɨ
ɦ
ɢ
ɢ
ɠ
ɢ
ɡ
ɧ
ɟ
ɧ
ɧ
ɨ
ɜ
ɚ
ɠ
ɧ
ɵ
.
ȼ
ɫ
ɟ
ɛ
ɟ
ɥ
ɤ
ɢ
ɢ
ɦ
ɧ
ɨ
ɝ
ɢ
ɟ
ɧ
ɟ
ɣ
ɪ
ɨ
ɦ
ɟ
ɞ
ɢ
ɚ
ɬ
ɨ
ɪ
ɵ
ɜ
ɤ
ɥ
ɸ
ɱ
ɚ
ɸ
ɬ
ɚ
ɦ
ɢ
ɧ
ɨ
ɤ
ɢ
ɫ
ɥ
ɨ
ɬ
ɵ
Минимальные системные требования определяются соответствующими требованиями про-
граммы Adobe Reader версии не ниже 11-й для операционных систем Windows, Mac OS,
Android, iOS, Windows Phone и BlackBerry, экран 10"
Учебное эл ектронное издание
Серия: «Лучший зарубежный учебник»
МОЗГ, ПОЗНАНИЕ, РАЗУМ: ВВЕДЕНИЕ В КОГНИТИВНЫЕ НЕЙРОНАУКИ
В двух частях
Часть первая
Ведущий редактор канд. биол. наук В. В . Гейдебрехт
Редактор канд. биол. наук О. Б. Мацелепа. Художник Н. А. Новак
Технический редактор Е. В. Денюкова. Компьютерная верстка: В. А. Носенко
Подписано к использованию 25.07.14 .
Издательство «БИНОМ. Лаборатория знаний»
125167, Москва, проезд Аэропорта, д. 3
Телефон: (499) 157-5272, e-mail: binom@Lbz.ru, http://www.Lbz.ru
Книга знакомит читателя:
• с с овременными методами визуализации мозга, позволяющими перейти на
новый уровень наблюдения (в реальном времени) и изучения когнитивных
механизмов и механизмов восприятия;
• с новейшими разработками в области молекулярных основ сознания;
• с результатами новых методов регистрации активности мозга, позволяющими
утверждать, что мозговые ритмы несут информацию как о сознательных, так
и о бессознательных процессах;
• с современными представлениями о строении префронтальной коры, на осно"
вании которых пересмотрена роль лобных долей, считавшихся непостижимыми.
Рассмотрены все возможные нарушения мозговой деятельности: от случаев
повреждения памяти до возникновения слепого зрения, гемиагнозии, прозо"
пагнозии и других расстройств зрительной системы.
Вы узнаете:
• почему люди иногда спонтанно начинают повторять движения за другим чело"
веком и не могут остановиться;
• почему профессиональные музыканты иногда не в состоянии подавить желание
петь во время игры на инструменте;
• как спортсмены и музыканты, используя сенсомоторные области коры голов"
ного мозга, осуществляют мысленные тренировки;
• почему ученые считают, что сознание сегодня является законной и разрешимой
научной проблемой;
• как возникают эмоции и чем обусловлено наше социальное поведение.
Для студентов и специалистов по психологии, биологии, педагогике и медицине.
МОЗГ
ПОЗНАНИЕ
РАЗУМ
COGNITION,
BRAIN,
AND CONSCIOUSNESS
INTRODUCTION TO COGNITIVE
NEUROSCIENCE
Second Edition
Bernard J. Baars
Nicole M. Gage
AMSTERDAM • BOSTON • HEIDELBERG • LONDON • NEW YORK • OXFORD
PARIS • SAN DIEGO • SAN FRANCISCO • SINGAPORE • SYDNEY • TOKYO
Academic Press is an imprint of Elsevier
Москва
БИНОМ. Лаборатория знаний
2014
МОЗГ
ПОЗНАНИЕ
РАЗУМ
ВВЕДЕНИЕ В КОГНИТИВНЫЕ
НЕЙРОНАУКИ
Перевод 2
го английского издания
под общей редакцией
профессора, д
ра биол. наук
В. В . Шульговского
Б. Баарс, Н. Гейдж
ЛУЧШИЙ ЗАРУБЕЖНЫЙ УЧЕБНИК
ЛУЧШИЙ ЗАРУБЕЖНЫЙ УЧЕБНИК
В двух томах
2
ЭЛЕКТРОННОЕ ИЗДАНИЕ
УДК 612
ББК 28.707
М74
Серия основана в2006 г.
Переводчики:
В. Н. Егорова (гл. 6, 8), М. А. Каменская (гл. 1, 2),
В. М. Ковальзон (гл. 5, 7, 9, 13), А. В. Любителев (гл. 3, 16),
О. Б. Мацелепа (гл. 4, 10–12, 14, 15)
М74
Мозг, познание, разум: введение в когнитивные нейронауки
[Электронный ресурс] : в 2 ч. Ч. 2 / под ред. Б. Баарса, Н. Гейдж ;
пер. с англ. под ред. проф. В . В . Шульговского. — Эл. изд. — Элек-
трон. текстовые дан. (1 файл pdf : 467 с.) . — М . : БИНОМ. Лабо-
ратория знаний, 2014. — (Лучший зарубежный учебник). — Систем.
требования: Adobe Reader XI ; экран 10".
ISBN 978-5-9963-2353-1 (Ч. 2)
ISBN 978-5-9963-1452-2
В книге изложены ре зультаты новейших исследований связи строения
мозга и способно сти человека к познанию. В наглядной и до ступной фор-
ме представлена взаимо связь строения мозга, психологических функций,
эффектов во сприятия и умственных спо собно стей человека. Выводы о сно-
ваны на данных, полученных в психологии, биологии, медицине, биохимии
и физике. Показано, что когнитивные механизмы и механизмы во сприя-
тия, которые изучались на уровне поведения, в настоящее время могут
наблюдаться непо средственно на уровне работы с амого мозга благодаря
использованию новейших методов визуализации мозговых процессов.
Для студентов и преподавателей биологических, психологиче ских и ме-
дицинских вузов, педагогов, всех интере сующихся когнитивной нейронаукой
и работающих в этой области.
УДК 612
ББК 28.707
Деривативное электронное издание на основе печатного аналога: Мозг,
познание, разум: введение в когнитивные нейронауки : в 2 ч. Ч . 2 / под ред.
Б. Баарса, Н. Гейдж ; пер. с англ. под ред. проф. В . В . Шульговского. —
М. : БИНОМ. Лаборатория знаний, 2014. — 4 64 с. : ил . — (Лучший зарубеж-
ный учебник).
В соответствии со ст. 1299 и 1301 ГК РФ при устранении ограничений, уст ановленных
техническими средствами защиты авторских прав, правообладатель вправе требовать
от нарушителя возмещения убытков или выплаты компенсации
ISBN 978-5-9963-2353-1 (Ч. 2)
ISBN 978-5-9963-1452-2
c
○ 2010 Elsevier Ltd. All rights reserved.
This edition of COGNITION, BRAIN, AND
CONSCIOUSNESS, INTRODUCTION TO COGNITIVE
NEUROSCIENCE by Bernard Baars and Nicole Gage
is published by arrangement with ELSEVIER LIMITED
of The Boulevard, Langford Lane, Kidlington, Oxford,
OX5 1GB, UK. ISBN 978-0 -12-375070-9 .
Книга «Мозг, познание, разум: введение
в когнитивные нейронауки» под ред. Б. Баарса,
Н. Гейдж опубликована с разрешения ELSEVIER
LIMITED, The Boulevard, Langford Lane, Kidlington,
Oxford, OX5 1GB, UK
c
○ Перевод и оформление. БИНОМ. Лаборатория
знаний, 2014
4
Глава10.Мышлениеиспособностьрешатьзадачи.............11
1.0.Рабочаяпамять.................................................15
1.1 . Рабочая память перекрывается с вниманием, осознанными
событиямииэпизодическимивоспоминаниями.....................15
2.0.Эксплицитноерешениезадач.....................................19
2.1.Исполнительныйконтрольприрешениизадач......................22
3.0.Умственнаянагрузкаикорковаяактивность.........................27
4.0.Использованиесуществующихзнаний.............................30
4.1 . Практика и тренировка могут изменить
сетевоевзаимодействиевмозге...................................31
4.2.Семантическаяпамять..........................................32
4.3.Абстрактныепонятия,прототипыинейронныесети.................34
4.4.Знанияформируютсяввиденейроннойсети........................35
4.5.Умозрительныенедостатки......................................37
4.6.Определениеколичестваичисла..................................39
5.0.Имплицитноемышление.........................................40
5.1.Чувствооналичиизнания.......................................41
6.0.Заключение....................................................47
7.0.Заданияиупражнениякглаве10..................................48
Глава11.Речь..............................................51
1.0.Введение......................................................51
2.0.Природаязыка..................................................53
2.1.Биологическиеаспекты.........................................56
2.2.Истокиязыка..................................................62
2.3.Речьпротивязыка..............................................67
3.0.Звукиустнойречи...............................................67
4.0.Планированиеиформированиеречи...............................71
5.0. Эволюционные аспекты производства и восприятия речи . . . . . . . . . . . . .74
6.0.Словаиихзначения.............................................76
6.1.Культурнаясокровищницасловиидей.............................78
6.2.Узнаваниесинонимов...........................................79
6.3 . Современные данные о словах и их значении фрагментарны . . . . . . . . . . 80
7.0.Синтаксис,включениеисинтагматизация...........................81
Оглавление
6
Оглавление
8.0.Просодияимелодикаречи......................................82
9.0.Значимыеутверждения.........................................84
10.0.Объединенныеобразыречи.....................................84
11.0.Заключение..................................................88
12.0.Заданияиупражнениякглаве11.................................88
Глава 12.Цели, управляющий контроль и действие . . . . . . . . . . . .91
1.0.Введение......................................................91
1.1.Множествосложныхфункцийлобныхдолей.......................92
1.2.Отмолчащихдолейкоргануцивилизации.........................93
2.0.Филогенезионтогенез...........................................93
2.1.Филогенез.....................................................94
2.2.Онтогенез.....................................................95
3.0.Обзорфункций.................................................95
3.1.«Памятьбудущего»...............................................96
3.2.Самовосприятиеиуправляющиефункции...........................96
4.0.Болеедетальноерассмотрениелобныхдолей........................97
4.1.Нейроанатомияинейрофизиологиялобныхдолей...................97
4.2.Какустановитьграницыпрефронтальнойкоры.....................97
4.3.Обширныепроводящиепутилобныхдолей.........................97
5.0. Более детальное рассмотрение функций лобных долей . . . . . . . . . . . . . . .100
5.1.Функциирегуляции............................................100
5.2.Социальноевзрослениеинравственноеразвитие...................102
6.0.Нейровизуализацияуправляющегомозга..........................104
6.1.Вниманиеивосприятие........................................104
6.2.Рабочаяпамять................................................107
6.3.Управляющиефункцииимоторныйконтроль.....................110
6.4.Принятиерешения.............................................112
6.5.Усвоениеправил..............................................114
7.0.Нарушениялобныхдолей.......................................115
7.1.Хрупкиелобныедоли..........................................115
7.2.Синдромыповреждениялобныхдолей............................116
7.3.Повреждение лобных долей и асоциальное поведение. . . . . . . . . . . . . . . 119
7.4 . Другие клинические повреждения, связанные с повреждением
лобныхдолей.................................................120
8.0. Современный взгляд на организационные принципы лобных долей . . . .122
9.0. К единой теории управляющего контроля: заключение . . . . . . . . . . . . . . .123
10.0.Заданияиупражнениякглаве12 ...............................124
Глава13.Эмоции..........................................127
1.0.Введение.....................................................127
1.1.Триединыймозг...............................................128
1.2.Базовыеэмоцииирольрефлексивногосознания....................130
2.0.ЭмоциональныесистемымозгапоПанксеппу......................130
2.1.Переживаниеэмоций..........................................134
Оглавление
7
3.0.Системастраха................................................134
3.1 . Сознательное и бессознательное восприятие страха:
«нижний»и«верхний»путипоЛеду.............................138
3.2.Неосознаваемыйстрах.........................................139
3.3.Аффективноеслепозрение......................................142
3.4 . Взаимодействие познавательных и эмоциональных функций
приреакцияхстраха...........................................143
3.5.Имплицитноеэмоциональноеобучениеипамять...................144
3.6.Эмоциональнаямодуляцияэксплицитнойпамяти...................144
3.7.Влияниеэмоцийнавосприятиеивнимание........................145
3.8.Эмоцииисоциальноеповедение.................................147
3.9.Торможениеирегуляцияэмоций.................................147
4.0. Система положительного подкрепления: любить, хотеть, учиться . . . . . .151
4.1 . Новое понимание «подкрепления»: от подкрепления
к предсказанию подкрепления и далее —
кошибкампредсказанияподкрепления...........................152
4.2.Подкрепление—этобольшечемобучение........................154
4.3.«Путьподкрепления»инаркотики...............................157
4.4.Сигналыподкреплениявлияютнавнимание.......................158
5.0.Заключение...................................................159
6.0.Заданияиупражнениякглаве13.................................159
6.1.Контрольныезадания..........................................159
6.2.Графическиеупражнения.......................................160
Глава 14. Социальная когнитивная деятельность:
восприятие ментального состояния других людей . . . . . . . . . . . . 163
1.0.Введение.....................................................163
1.1 . Термины, используемые в отношении
социальнойкогнитивнойдеятельности...........................164
1.2.Важностьперспективы:первое,второеитретьелица...............165
1.3 . Подходы к восприятию ментального состояния других людей . . . . . . . . 165
2.0. Организационная основа для социальной когнитивной
деятельности..................................................168
2.1.Намерение...................................................168
2.2.Распознаваниеглаз............................................169
2.3.Разделенноевнимание.........................................170
2.4.Теорияразуманаивысшегопорядка..............................170
3.0.Зеркальныенейроныивосприятиенамерений......................171
3.1.Отдействийкнамерениям .....................................171
3.2.Распознавание глаз и восприятие направлениявзора . . . .. . . . . .. . . . . .178
3.3.Разделенноевнимание.........................................180
3.4.СпособностивысокогопорядкаизТР.............................182
3.5. Восприятие других как похожих и непохожих: Я-Оно в мозге . . . . . . . . 187
3.6.Восприятиелиц...............................................190
4.0.Заключение...................................................192
5.0.Заданияиупражнениякглаве14.................................193
8
Оглавление
5.1.Контрольныезадания..........................................193
5.2.Графическиеупражнения ......................................193
Глава15.Развитие.........................................195
1.0Введение.....................................................195
1.1 . Новые технологии для исследования развивающегося мозга. . . . . . . . . . 196
1.2.Тайнаразвитиямозга:старыеиновыевопросы....................197
2.0. Пренатальное развитие: от бластоцисты до младенца . . . . . . . . . . . . . . . .197
2.1.Эпигенез.....................................................198
2.2.Анатомияразвивающегосямозга................................199
2.3.Миграциянейронов............................................204
2.4.Пересмотрроливрожденногоиприобретенного ...................207
2.5 . Пренатальный слуховой опыт: восприятие голо са и музыки
дорождения..................................................210
3.0.Развивающийсямозг:изменениядлиноювжизнь...................211
3.1.Начало изавершение постнатальногоразвития мозга . . .. . . . . .. . . . . .211
3.2.Региональныеособенностиразвитиямозга........................213
4.0.Развитиемозгаисознания.......................................215
4.1.Первыйгоджизни:взрывнойростиразвитие......................221
4.2 . Детство и подростковый возраст:
динамичноеипоэтапноеразвитие...............................234
5.0.Ранние повреждения мозга и пластичность развития . . . . . . . . . . . . . . . .252
6.0.Заключение...................................................256
7.0.Заданияиупражнениякглаве15.................................256
7.1.Контрольныезадания..........................................256
Глава16.Геныимолекулымышления.......................259
1.0.Введение.....................................................260
1.1 . Основные малые молекулы клетки также используются
длянейрональнойсигнализации.................................261
1.2.Живыеорганизмыиспользуютферменты.........................263
1.3.Однаитажемолекуламожетнестиразныесигналы................264
1.4 . Гематоэнцефалический барьер защищает внутреннюю
средумозга...................................................264
1.5.Модельныеорганизмы.........................................266
2.0.Генывэволюции,онтогенезеиповседневнойжизни.................268
2.1 . Разные события с ДНК происходят в различные
промежуткивремени...........................................270
2.2.Развитиемозга:нейрогенез,затемсинаптогенез....................271
2.3.Какчеловекполучилтакойбольшоймозг?........................274
3.0.Экспрессияирегуляциягенов....................................275
3.1.Обработкаинформациивклетке.................................275
3.2.ДНКвысшихуровнейрегулируетостальнуюДНК..................277
3.3.Ошибкиприразвитиимозга....................................279
3.4Гены,контролирующиеразвитие.................................279
3.5 . Программирование генов: за пределами центральной догмы . . . . . . . . . 281
Оглавление
9
3.6.Средаможетперестраиватьхроматинвядреклетки ................282
3.7.Обучениекакэпигенетическийпроцесс...........................283
4.0. Нейроны и глиальные клетки как обработчики сигналов . . . . . . . . . . . . .283
4.1.Нейроныисинапсывкачествепереключателей....................284
4.2.Химическаясаморегуляция.....................................285
4.3.Мембраны,ионныеканалыиаксоновыеспайки....................286
5.0. Синаптическая передача: от производства до расчистки . . . . . . . . . . . . . .290
5.1.«Большаядвойка»:глутаматиГАМК.............................290
5.2.ГАМК:основнойтормозныймедиатор............................292
5.3.Нейроглиятакже можетобрабатыватьинформацию. . . . .. . . . . .. . . . . .294
5.4 . Производство, высвобождение и удаление нейромедиаторов . . . . . . . . . 295
5.5.Высвобождениевеществавсинапсе..............................295
5.6.Синапсыирецепторывкачествеконтрольныхточек................298
5.7.Рецепторыраспознаютсигнальныемолекулы......................298
5.8.Трансмиттерзависимыеионныеканалы...........................302
5.9 . Очистка синапса: дезактивация, диффузия и утилизация
медиатора....................................................306
6.0.Нейромодуляторы..............................................308
7.0.Обучение.....................................................310
7.1.Гиппокампальныйкомплекс.....................................312
7.2 . Глутамат, ГАМК и долговременная депрессия и потенциация . . . . . . . . . 314
7.3 . Глутаматные синапсы в качестве модели
синаптическогообучения.......................................315
7.4.Эпигенетическиемеханизмыобучения............................322
7.5.Нейротропныефакторыприобучении............................323
8.0.Заключение...................................................324
9.0.Заданияиупражнениякглаве16.................................327
Приложение. Методы для изучения живого мозга . . . . . . . . . . . . .331
1.0.Историческиепредпосылки.....................................331
1.1.Корреляциямозгаиразума......................................331
1.2.Регистрацияактивностимозга...................................333
2.0.Методы......................................................335
2.1.Электроэнцефалография.......................................335
2.2.Магнитоэнцефалография.......................................344
2.3.Позитронно-эмиссионнаятомография............................350
2.4.Магнитно-резонанснаятомография..............................353
2.5.МРТ —методбудущего........................................370
2.6.Оптическоекартирование.......................................372
3.0.Мультимодальныеметодывизуализациимозга.....................374
3.1.Одновременнаявизуализацияразнымиметодами....................374
3.2.Визуализациягенетики.........................................376
4.0.Заключение...................................................379
Литература...............................................381
Глоссарий................................................433
Высший уровень решения задач. Микронезийская навигация — пример чрезвычайно
сложного для успешной реализации практического опыта. Если бы не этот навык, то
жители островов Тихого океана не могли бы осуществлять навигацию на маленьких
суднах, путешествовать от острова к острову, проплывая сотни миль, как они делают
в течение нескольких тысяч лет. Звездный компас (вверху) является изобретением
океанийских мореплавателей, которые придерживались определенного направления
плавания, измеряя угол восходящих и заходящих звезд в ночном небе относительно
Полярной звезды. Высокий уровень знаний по навигации разрабатывался, сохранялся
и передавался. Каноэ с балансиром — очень эффективный способ передвижения на
длинные дистанции, но вряд ли безопасный.
Привлечение произвольного внимания обычно ассоциируется с субъектив-
ным ощущением умственного усилия.
Л. Наккаш, С. Деан, Л. Коэн, М.-О. Абер,
Э. Гишар-Гомес, Д. Галано и Ж. - К. Виллер (2004)
Содержание
1.0.Рабочаяпамять...................................................15
1.1 . Рабочая память перекрывается с вниманием, осознанными событиями
иэпизодическимивоспоминаниями...............................15
2.0.Эксплицитноерешениезадач.......................................19
2.1.Исполнительныйконтрольприрешениизадач......................22
3.0.Умственнаянагрузкаикорковаяактивность............................27
4.0.Использованиесуществующихзнаний................................30
4.1 . Практика и тренировка могут изменить
сетевоевзаимодействиевмозге..................................31
4.2.Семантическаяпамять..........................................32
4.3.Абстрактныепонятия,прототипыинейронныесети.................34
4.4.Знанияформируютсяввиденейроннойсети.......................35
4.5.Умозрительныенедостатки......................................37
4.6.Определениеколичестваичисла.................................39
5.0.Имплицитноемышление............................................40
5.1.Чувствооналичиизнания.......................................41
6.0.Заключение......................................................47
7.0.Заданияиупражнениякглаве10....................................48
У человека решение задач осуществляется двумя способами, эксплицитным
и имплицитным. Эти два варианта значительно различаются. Эксплицитное
мышление содержит четкие, осознаваемые конечные и промежуточные цели и
точно определенные шаги для того, чтобы прийти от начальной точки к реше-
нию. В качестве примера из повседневной жизни можно привести умственный
арифметический счет. Эксплицитное мышление требует более значительного
контроля со стороны высших отделов мозга, повышенную умственную нагруз-
ку, более частое привлечение сознания, и более широкое вовлечение корковых
областей мозга.
В отличие от этого имплицитное принятие решения может быть более при-
вычным, так как мы обучаемся и практикуем большое число навыков с начала
жизни. Такие навыки по поиску и принятию решения с практикой становят-
ся более искусными, имплицитными (не осознаваемыми) и автоматическими.
Примером может служить понимание этого написанного предложения или ло-
гическое завершение предсказуемого предложения, так что мы можем сказать,
какие слова должны быть в конце. Имплицитное принятие решения требует
меньшего контроля со стороны исполнительных отделов по сравнению с экс-
плицитным вариантом, меньшего привлечения сознания, меньше умственной
Глава 10
Мышление и способность
решать задачи
12
Глава 10
нагрузки и вовлечения корковых областей. С другой стороны, данный вид ре-
шения задач в большей степени зависит от долговременной памяти и высоко-
тренированных навыков. Очень часто неявными целями обучения является
перевод эксплицитного принятия решения в имплицитное.
В области мышления продолжается исторический спор между локальными
и распределенными функциями мозга. С помощью методов визуализации мы
обнаруживаем гораздо больше локальных областей и процессов, чем раньше.
В связи с этим нам может показаться, что теория локализационизма функций
мозга выигрывает этот спор, но этого (еще) не случилось. Несмотря на то что
более маленькие и специфичные участки мозга могут быть исследованы, дока-
зательства широкого распределения процессов до сих пор очевидны (см. гл . 9).
В связи с этим некоторые теоретики разрабатывают модели, которые включают
в себя локализационный и распределенный подходы к проектированию ней-
ронных сетей.
Произвольное внимание и непосредственная память
Любая задача, которую мы пытаемся решить в нашей по-
вседневной жизни, создает спрос на произвольное вни-
мание — о чем мы даем себе отчет, — а также на объем
непосредственной памяти. Также от нас требуется кон-
тролировать, на чем сфокусировать внимание. Нельсон
Кован посвятил свою успешную карьеру изучению этих
«ограниченных емкостных свойств» человеческого мозга
вместе с их постоянными взаимодействиями с огромным
хранилищем долговременной памяти (рис. 10 .1 и 10.2).
По словам Кована, «то, что служит главным емкостным
ограничением рабочей памяти... очень тесно связано с
содержанием сознательной психической деятельности»
(2008).
Произвольное внимание, согласно исследованиям Ко-
вана, еще более ограничено, чем ранее считалось пси-
хологами. При тщательном проведении экспериментов
с исключением возможности мысленного повторения и
группировки предметов объем произвольного внимания сокращается до четырех
и менее объектов. Согласно же традиционным взглядам непосредственная память
может сохранять «семь плюс или минус два» различных объекта, но это справед-
ливо только при их мысленном повторении человеком. Если испытуемых просят
произносить последовательность звуков «the... the... the» для предотвращения
перечисления запоминаемых предметов, объем памяти сразу падает до четырех.
Ограничение памяти четырьмя объектами ассоциируют с участием задней темен-
ной коры в этом процессе, однако до сих пор продолжается спор о мозговой основе
ограниченной емкости памяти.
«Магическое число четыре» является достаточно стабильным для объектов разного
типа — чисел, коротких слов, простых звуков, зрительных объектов и т. д . Кован
считает, что это применимо как для абстрактных понятий, так и для перцептив-
ных событий, образов и внутренней речи. Для того чтобы мысленно сложить два
числа, мы должны на несколько секунд, пока мы производим вычисление суммы,
задержать их в памяти. Для понимания смысла длинного предложения человеку
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
Рис. 10.1 . Нельсон
Кован (Nelson Cowan),
кандидат наук, факуль-
тет психологических
наук,
Университет
Миссури, Колумбия,
США
Мышление и способность решать задачи
13
необходимо сохранить его первую часть в непосредственной памяти для того, что-
бы постичь смысл второй части. Предшествующее предложение служит отличным
примером этого явления. Однако существуют данные о кодировании каждого зна-
чимого куска длинного предложения в семантическую единицу, например фраза
«Для понимания смысла длинного предложения»... Люди очень искусны в семан-
тическом кодировании информации из-за постоянной тренировки при попытках
понять сказанное при общении. Мы стараемся преобразовать слова в смысл так
быстро, как мы это можем.
Проблема заключается в том, как наилучшим образом использовать ограниченные
ресурсы, с которыми мы родились. Одним из подходов служит мысленное повто-
рение последовательности чисел. С тренировкой предел нашей рабочей памяти по-
вышается до традиционного «семь плюс или минус два объекта». Но в этом случае
мы уже используем как произвольное внимание, так и емкость непосредственной
памяти для внутренней речи и мысленного повтора.
Мы также можем снизить заполнение непосредственной памяти, написав заметку
или выделяя предложение в книге. Но письменность является относительно недав-
ним изобретением человека, не считая такие возможности, как аудио- или видео-
записи. До возникновения письменности человечество все равно было способно
к решению сложных задач, в чем мы убеждаемся на примерах культур, которые не
используют письмо. Они опираются на свою долговременную память (ДВП), мне-
моническую схему и тщательно выработанные знания об устройстве мира. Многие
культуры используют такие речевые схемы, как стихотворные ритмы или аллите-
рации, мелодичный напев или стихотворный шаг. Именно поэтому маленькие дети
легко выучивают алфавит посредством песни. Это является неплохим способом ос-
воить и анатомию мозга.
Рис. 10 .2 . Когнитивный психолог Нельсон Кован предполагает, что непосредствен-
ная память может быть представлена как активированная часть долговременной.
Например, мысленное повторение слов активирует существующий словарный запас
в длительно хранимом знании языка. Любопытно, что для сохранения в долговре-
менной памяти новых ассоциаций между уже известными элементами необходима
одновременная фокусировка на них внимания. Очень важно воспринимать непосред-
ственную память и концентрирование внимания, внутреннее повторение и другие
компоненты «рабочей памяти» (РП) как постоянно существующие аспекты нормаль-
ной когнитивной деятельности. Таким образом, элементы РП могут быть частично
заимствованы из ДВП, и внутренняя речь может запросто использовать мозговые
возможности «внешней речи». (Источник: Nelson Cowan, из личной переписки.)
14
Глава 10
Человек может решать задачи при помощи произвольного внимания и непосред-
ственной памяти при условии использования их совместно с огромными библио-
теками долговременной памяти. Каждое прочитанное вами слово задействует на
короткое время немного ресурсов ограниченного произвольного внимания, но тот
факт, что вы знаете около 100 000 слов и их значения, позволяет обрабатывать боль-
шие «блоки слов», а не просто бессмысленные всплески и бульканье звуков.
Часть ДВП, задействованной в текущих процессах, Кован называет активной памя-
тью. Во время чтения данного параграфа ваша активная память содержит информа-
цию о том, что вы освоили из этой главы, а также все аналогичные знания из других
источников. Как у авторов, все наше стремление направлено на попытки связать
научные факты со знанием, которое студенты могут напрямую использовать.
Кован указывает на существование как временного, так и емкостного предела. Из
рис. 10.2 видно, что фокус внимания, в котором находятся осознаваемые в настоя-
щий момент события, имеет емкостной предел. В противоположность этому, «ак-
тивная память» на рис. 10 .2 может иметь временной предел, т. е . исчезать со вре-
менем. Например, при сосредоточении внимания на чтении этого предложения вы
можете заметить постепенное забывание его через несколько секунд.
Согласно предположениям Кована, данная система с ограниченными возможностя-
ми может, на самом деле иметь оба из этих двух пределов, в зависимости от фокуса
внимания. Если мы настраиваем узкий фокус внимания, то только один объект мо-
жет быть сознательно воспринят, но при широком фокусе могут быть восприняты
до четырех различных объектов. Как известно, такая схема работает для зрения.
Зрительное внимание концентрируется или нет на различных деталях, и мы можем
наблюдать, как в мозге активируются перекрывающиеся области коры при взаимо-
действии процессов контроля над зрительным вниманием и движением глаз (гл. 8).
Таким образом, зрительное внимание и движения глаз могли совместно развиваться
в процессе эволюции позвоночных животных. Схожий скрытый «потенциал увели-
чения внимания», возможно, эволюционировал вместе с изменением фокусировки
в зависимости от интенсивности света. Эта гипотеза, по крайней мере, может быть
проверена при помощи современных методов визуализации работы мозга (Cowan,
2008).
На некоторые вопросы до сих пор нет однозначного ответа. Например, Оберауэр и
Биалкова (Oberauer, Bialkova, 2009) считают, что существует сознательный фокус
внимания, нацеленный только на один объект, наряду с «внешней границей», в ко-
торой содержатся от трех до пяти объектов. Но данное несогласование в предель-
ном числе воспринимаемых объектов не является существенным.
Узкое горлышко в потоке мысленной информации имеет сильный эффект на про-
цессы обучения и решения задач. Люди различаются между собой способностями
направлять и удерживать фокус внимания, игнорировать отвлекающие события и
продолжать выполнять сложное задание. Эти различия коррелируют с индивиду-
альными особенностями когнитивной деятельности, как общий уровень интеллек-
та, упорство и время подготовки реакции. Кроме того, они развиваются предсказу-
емым образом в течение жизни.
Халфорд, Кован и Эндрюс (Halford, Cowan and Andrews, 2007) выдвинули пред-
положение о существовании особой связи между рабочей памятью и логическим
мышлением, как в задаче «Рыцари и мошенники» (2007; http://en.wikipedia.org/wiki/
Knights_and_Knaves). Задача про рыцарей и мошенников может быть решена раз-
мещением элементов (А и Б) в группы (рыцари или мошенники).
Емкостной предел для рабочей памяти и мышления может быть соотнесен с числом
связей в ячейках координатной сетки или взаимосвязей. Рабочая память ограничена
приблизительно четырьмя активными объектами, тогда как образы при логическом
мышлении лимитированы четырьмя взаимосвязанными переменными.
Мышление и способность решать задачи
15
Литература
1. Cowan, N. (2008). What are the differences between long-term, short-term, and
working memory? Progress in Brain Research, 169, 323–338 .
2. Halford, G.S., Cowan, N. and Andrews, G. (2007). Separating cognitive capacity from
knowledge: A new hypothesis. Trends in Cognitive Sciences, 11 (6), 236–242 .
3. Oberauer, K. and Bialkova, S. (2009). Accessing information in working memory: Can
the focus of attention grasp two elements at the same time? Journal of Experimental
Psychology General, 138 (1), 64–87 .
1.0 . Рабочая память
В широком смысле, термин «рабочая память» (РП) является частью принятия
решения, речи и мышления (гл. 2). Рабочая память — это «набор умственных
процессов по удержанию лимитированного объема информации во временно
доступном состоянии для обеспечения когнитивной деятельности» (Cowan
et al., 2005). Рабочая память нам необходима при выполнении арифметических
операций, для поддержания разговора и для решения задачи по поиску пути —
каким образом отсюда я могу дойти до дома? Вы не можете понять смысла
предложения, которое вы сейчас читаете, без существования набора слов, идей
и синтаксиса в оперативной памяти.
Когда мы думаем над какой-нибудь задачей, мы постоянно пользуемся вну-
тренней речью и зрительно-пространственными образами, тем самым направ-
ляя внимание на то, что в данный момент имеет первостепенное значение. Как
было написано в гл. 8, внимание обеспечивает восприятие нами сенсорной ин-
формации, внутренней речи и формирования программы действия. И наконец,
то, о чем мы думаем в настоящий момент, взаимодействует с тем, что мы уже
знаем — с воспоминаниями, знаниями и навыками в долговременной памяти.
Каждое движение глаз, которое вы совершаете, основано на длительно практи-
куемом навыке (см. гл . 9). Таким образом, все составные части схемы на рис.
10.3 вовлекаются в процессы мышления и решения задач.
Удивительным является то, что большая часть данной главы вовсе не о цвет-
ных участках схемы, а о ряде серых прямоугольников внизу рисунка. Цветные
блоки отражают активные процессы, те, что требуют нейрональных залпов и
интеграции ответов. Но они очень часто имеют сходные пути, что и предыду-
щие процессы, которые к этому моменту уже сформировали устойчивые связи
нейронных сетей. Исследования с визуализацией мозговой активности могут
не показывать такие устойчивые образования, так как они закодированы в силе
взаимодействия между нейронами. Следовательно, методы, подобные фМРТ,
могут представлять в меньшем количестве объем долговременно хранимых
знаний.
1.1 . Рабочая память перекрывается с вниманием,
осознанными событиями и эпизодическими
воспоминаниями
Эксперименты в основном нацелены на разделение рабочей памяти и схожих
процессов. Например, в некоторых экспериментах на рабочую память на кошках
или обезьянах регистрируют активность одиночных нейронов в префронталь-
ной коре в задачах с отсроченным выбором по образцу. В этих экспериментах
16
Глава 10
животному предъявляют стимул, затем после исчезновения стимула следует пе-
риод ожидания, по окончании которого оно должно указать запомненный сти-
мул среди ряда других. Распространенные данные показывают, что нейроны
продолжают разряжаться во время периода задержки, что интерпретируется ис-
следователями как рабочая память, с помощью которой животное держит в уме
предъявленный стимул, пока ждет возможности совершить требуемое действие
(Goldman-Rakic, 1995, см. гл . 9). Селективное внимание выявляется на людях в
случае наличия двух конкурирующих сенсорных входов. Сознательная когни-
тивная деятельность сравнивает сознательные (о которых испытуемые дают от-
чет) и бессознательные (без отчета) стимулы при помощи экспериментальных
методик с несовпадением изображения на сетчатке (см. гл . 6 и 8). Изучение
процессов извлечения эпизодической памяти может включать опрос испыту-
емых об их вчерашнем обеде. Используя условия для точного сравнения, мы
обнаруживаем, что «загораются» разные участки мозга. Новые и достоверные
знания часто получают при помощи описанных методик.
В то же время на рис. 10.4 и 10.5 показана обширная активация фронтальных
и теменных областей коры головного мозга во время выполнения четырех за-
дач на выявление процессов, которые обычно отделяют друг от друга: рабочая
память, внимание, извлечение эпизодической памяти и осознанное восприятие.
Пересечение областей огромное. Это не значит, что данные процессы имеют
одинаковое теоретическое обоснование. Они могут быть разделены экспери-
ментальными методами. Однако не следует забывать, что мозг продолжает рабо-
тать независимо от того, какие эксперименты мы разрабатываем. В данной главе
Рис. 10 .3 . Функциональная схема процесса решения задачи. Рабочая память (РП) по-
стоянно вовлечена в решение задач. Однако РП также помогает использовать сохранен-
ную ранее информацию, такую как набор слов родного языка и их смысл, набор привы-
чек и моторных навыков, а также значительного количества вариантов памяти, которые
показаны в серых блоках внизу рисунка
Мышление и способность решать задачи
17
Рис. 10 .4 . Предположительные области, отвечающие за РП. Области мозга, обычно во-
влеченные в процессы образования РП. (Источник: Schneider and Chein, 2003.)
Рис. 10 .5 . Перекрывающиеся
области мозга, ответственные за
РП, селективное внимание, из-
влечение автобиографических
воспоминаний и сознательное
восприятие. Этот рисунок схе-
матически показывает обшир-
ную активацию фронтальных
и теменных областей мозга во
время четырех разных актив-
ностей мозга, которые обычно
отделяют друг от друга: РП,
внимание, извлечение эпизо-
дической памяти и осознанное
восприятие. Существует значи-
тельное перекрывание актив-
ных областей и поэтому совер-
шенно не очевидно, что данные
процессы могут быть разде-
лены. Внимание = выделение
стимула, РП = отсроченный вы-
бор, сознательное восприятие
= сравнение видимого объекта
с неосознаваемым, извлечение
эпизодической памяти = осоз -
нание автобиографического со-
бытия, извлеченного из памяти.
В то время как эти четыре ак-
тивности мозга отделяют друг
от друга в экспериментах, они
имеют множество перекрыва-
ющихся свойств. (Источник:
Naghavi and Nyberg, 2005.)
18
Глава 10
мы будем использовать термин «рабочая память» при рассмотрении сознатель-
ной и бессознательной когнитивной деятельности, восприятия и долговремен-
ной памяти. Большинство активных сторон рабочей памяти требуют сознания и
являются произвольными. Таким образом, было высказано предположение, что
функции пассивной рабочей памяти могут находиться под контролем сознатель-
ных/произвольных процессов, как перцептивный вход, извлечение, повторение,
планирования действия и ответные реакции (Baars и Franklin, 2003).
Пределы рабочей памяти создавали для человечества немало трудностей до
изобретения и распространения письменности. Сегодня мы можем набросать
заметки на листе бумаги или посмотреть информацию по Интернету. Но в пери-
од неолита никто не мог написать список продуктов перед началом охоты или
сбором пищи. Вероятно, именно поэтому используемые языки содержат много
приспособлений и приемов для упаковывания большого объема информации в
небольшие блоки: предложения, слова, фразы, поговорки, имена нарицатель-
ные, имена собственные и местоимения, все из которых можно рассматривать
как указатели и напоминания (рис. 10.6). Мы также концентрируем большой
объем знаний в виде абстракций и классификационных схем (блок 10.1). Боль-
Рис. 10.6 . Некоторые участки лобной коры, задействованные в высших мыслительных
процессах. Лобные доли также необходимы для продвинутых навыков, простирающих-
ся от возможности понимать намерения других людей до рассуждений, воображения и
самопознания. Эти лобные процессы постоянно находятся во взаимодействии с задни-
ми и подкорковыми областями мозга. (Источник: Wicker et al., 2003.)
Мышление и способность решать задачи
19
шая часть нашего словарного запаса состоит из подобных ярлыков. Словарный
запас естественных языков — сокровищница блоков значимой информации, со-
бранных под влиянием культуры (см. гл . 11).
Каждый блок может указывать на достаточно большой объем знания. В дан-
ной книге слово «мозг» может служить связью между всем тем, что вам из-
вестно в когнитивной нейронауке. Обращаясь к знанию посредством коротких
ярлыков, мы можем оптимизировать наши когнитивные ограничения.
2.0 . Эксплицитное решение задач
Были описаны (Unterrainer, Owen, 2006) основные условия эксплицитного ре-
шения задач.
В начале объекту требуется создать мысленное представление о текущей
ситуации и цели. Далее эти представления должны быть связаны путем опре-
деления, какие требуется совершить действия для изменения текущего поло-
жения в целевое. Таким образом, задача имеет три основные характеристики:
1) начальная стадия, или стадия, на которой объект определяет начальные ус-
ловия; 2) стадия цели, или стадия решения, которую объект пытается достичь;
3) шаги, которые объект предпринимает для трансформации начального состо-
яния в требуемое, которое изначально может быть неочевидным (Sternberg and
Ben-Zeev, 2001).
Решение задачи можно представить как поиск пути через лабиринт точек
выбора между возможными промежуточными целями к конечной цели. В стан-
дартной головоломке под названием «ханойская башня» предъявляют задачу,
как показано на рис. 10.7 . Целью головоломки является перемещение трех ко-
лец в крайнее правое положение и расположение их в той же последовательно-
сти, как было вначале. Эти действия требуют составления промежуточных це-
лей. Схему возможных вариантов называют пространством задачи (рис. 10.9).
Некоторая последовательность действий ведет к правильному решению задачи,
Рис. 10 .7 . Ханойская башня. Шеллис (Shallice, 1982) использовал головоломку под на-
званием «башни Лондона» (или «ханойская башня») для диагностирования поврежде-
ний фронтальных отделов головного мозга. В задаче требуется перемещать кольца по-
следовательно по одному до совпадения с финальной схемой. Уровень сложности этой
загадки может изменяться, как показано в правой половине рисунка. Задачи с башнями
стандартизировали для клинических испытаний, и в результате было проведено боль-
шое количество исследований. Наличие инструкции составлять план действий улучша-
ет выполнение у людей с неповрежденными лобными областями коры головного мозга.
(Источник: Miller and Wallis, из Squire et al., 2003.)
20
Глава 10
Блок 10.1 . Каким образом профессиональные шахматисты используют из-
вестные игровые позиции
Специальная память для шахматных позиций
Большая часть того, что известно о профессиональной компетенции, относится к ра-
ботам Де Грота (De Groot, 1946), Чейза и Саймона (Chase, Simon, 1973). Выявление
существования четких различий в выполнении задачи на память, включающей кра-
тковременное предъявление шахматных позиций, взятых из турнирных игр, у игро-
ков разного уровня стало одним из неоценимых вкладов, который сделал Де Грот.
Обычно игроки уровня мастера спорта и выше могут воспроизвести правильно все
расположение фигур, тогда как более слабые шахматисты справляются гораздо хуже
(рис. 10.8).
Однако Чейз и Саймон не обнаружили различий в эффективности воспроизведения
случайных позиций у трех групп испытуемых: мастеров спорта, у игроков класса
А и новичков при использовании модели CHREST (Chunk Hierarchy and REtrieval
STructures — иерархия блоков и структуры, ответственные за извлечение). Такое
одинаково плохое выполнение задачи со случайным расположением фигур и вели-
колепное воспроизведение позиций мастерами спорта и гроссмейстерами нагляд-
но проиллюстрировали принцип, что знание является ключом к профессиональной
компетенции. Данный пример, став классическим открытием, широко цитируется в
учебниках по когнитивной психологии и статьях на тему профессионализма.
Однако ранняя версия CHREST, полученная путем доработки и дополнения MAPP
(Memory-Aided Pattern Recognizer — средство распознавания паттерна посредством
памяти; Simon and Gilmartin, 1973), делала противоположные предположения от-
носительно воспроизведения случайного расположения фигур. При моделировании
Рис. 10 .8 . (a) Варианты расположения
шахматных фигур, которые обычно ис-
пользуют в исследованиях памяти шах-
матистов. Изображенное слева распо-
ложение фигур было взято из игровой
партии мастеров, а справа — случайное
расположение фигур, полученное путем
их перемешивания. (б) Среднее число (ус-
реднено по 13 опытам) фигур, размещен-
ных правильно в соответствии с функцией
координат (игра или случайность), и уро-
вень мастерства. В среднем каждый раз
предъявлялось изображение из 25 фигур,
а время предъявления составляло ≤ 10 с.
Планки погрешностей («усы») отобража-
ют стандартные ошибки среднего. (Ис-
точник: Gomet and Simon, 1966.)
Мышление и способность решать задачи
21
в то время как другая нет. На рис. 10.10 показаны области мозга, в частности
дорсолатеральная префронтальная кора (ДЛ-ПФК), активные во время реше-
ния головоломки о ханойской башне.
игры в шахматы CHREST поисходит обучение на базе партий мастеров спорта, иден-
тифицируя контуры фигур в разных положениях. Как и было предположено, воз-
можность модели запоминать игровые позиции улучшалась с увеличением числа и
размера блоков. Однако у модели было зарегистрировано небольшое, но четкое уве-
личение числа правильного воспроизведения случайных позиций фигур. Различия в
воспроизведении являются результатом легко объяснимого механизма: чисто случай-
но обширная дифференцированная сеть с большей вероятностью будет содержать
тот же паттерн локализации фигур, что и в случае непроизвольного их расположения.
Методичный обзор экспериментов, в которых шахматистов просили воспроизвести
расположение случайно расставленных фигур (Gobet, Simon, 1966), содержал 12 ис-
следований, в которых мастера спорта показали превосходство, и только одну работу
(Chase, Simon, 1973), в которой профессионалы показали результаты хуже, чем но-
вички. Несмотря на то что в большинстве исследований различия в точности воспро-
изведения не были достоверными из-за недостатка статистических данных, этот эф-
фект отчетливо проявлялся при суммировании результатов всех работ (рис. 10 .8 (б)).
Тот факт, что создание блоков прецептивной информации создает преимущество для
профессиональных шахматистов даже при воспроизведении случайного расположе-
ния фигур, доказывает правомерность блоковых теорий над выдающимися знаниями
или схемами (De Groot, 1946). Более подробно см. другой обзор (Gobet, 1998).
Литература
1. De Groot, A.D. (1946). Het Denken van den Schaker. Noord Hollandsche.
2. Chase W.G. and Simon, H.A . (1973) Skill in chess. American Scientist, 61, 393–403 .
3. Simon, H.A. and Gilmartin, K.J. (1973). A stimulation of memory for chess positions.
Cognition Psychology, 5, 29–46 .
4. Gobet, F. and Simon, H.A . (1966). Recall of rapidly presented random chess positions is a
function of skill. Psychonomic Bulletin and Review, 3, 159–163.
5. Gobet, F. (1998). Expert memory: a comparison of four theories. Cognition, 66, 115–152.
Рис. 10 .9. Пространство задачи. Пространство задачи для ханойской башни показыва-
ет все возможные расклады, точки выбора и пути решения
22
Глава 10
Решение задачи с башнями скорее можно отнести к головоломкам, чем к
реальным жизненным проблемам. Действительными задачами для людей мо-
гут служить, например, поиск пути из одного местоположения в другое через
трудную и опасную территорию, поиск пищи и убежища, стремление защитить
детей и соплеменников от опасностей, желание сохранить взаимоотношения
или стремление преуспеть в конкурентной борьбе. Жизненные трудности очень
редко можно четко описать, и ставки могут быть очень высоки. Несмотря на
это, даже решение простых задач может продемонстрировать важные особен-
ности реальных целей. Такие понятия, как промежуточные и конечные цели,
точки выбора в пространстве задачи, издержки и выгоды, выявляются во мно-
гих случаях. В то время как составление полного пространства задачи в реаль-
ной жизни очень редко удается, одной из удачных стратегий ее решения может
быть выделение в большой задаче промежуточных целей, которые можно опи-
сать точно и полно.
2.1. Исполнительный контроль при решении задач
Клише о том, что «генералы склонны вести прошлую войну», можно прило-
жить к решению многих задач. В науке, например, существует тенденция, даже
среди выдающихся ученых, концентрироваться только на одном основном под-
ходе, который, однако, не всегда работает. Знаменитым примером этого служит
Альберт Эйнштейн, у которого сформировалось стойкое неприятие квантовой
теории, несмотря на то что она смогла объяснить непонятые ранее феномены.
Своими философскими воззрениями Эйнштейн отрицал идею того, что физика
может быть более статистической, нежели детерминистической, и более 30 лет
пытался найти альтернативу квантовой теории. В истории науки можно найти
сотни подобных примеров. По всей видимости, «зацикленность» является до-
статочно распространенной трудностью в решении задач у человека.
Рис. 10 .10 . Дорсолатеральная префронтальная кора при решении задачи с башнями.
Такие задачи вызывают увеличение мозговой активности в центре ДЛ-ПФК, являющей-
ся ключевой областью для высших контрольных функций. (Источник: Unterrainer and
Owen, 2006.)
Мышление и способность решать задачи
23
Очень просто вызвать функциональную зацикленность при решении задач
(Duncker, 1945). Для этого достаточно только поощрять людей использовать
один путь для достижения цели, даже в том случае, когда он больше не рабо-
тает. Функциональную зацикленность можно наблюдать в целом ряде экспе-
риментов. В языке существуют «предложения заблуждения» (англ. garden-path
sentences), которые сбивают с пути и вызывают непонимание читателей отно-
сительно синтаксиса предложения. Очень распространенным примером этого
служит предложение «The horse raced past the barn fell». Это грамматически
правильное предложение может заставить читателя задуматься на несколько
минут. Проблема состоит в том, что в предложении существует внешняя часть,
«The horse ... fell» (Лошадь ... упала), и внутренняя часть, «raced past the barn»
(мчалась мимо амбара). Его можно перефразировать таким образом:«The horse
which raced past the barn fell» (Лошадь, которая мчалась мимо амбара, упала).
Однако в английском языке маркер «which» является необязательным элемен-
том для подчиненных предложений.
Таким образом, предложение ведет читателя по неправильному пути непо-
нимания, и только в конце предложения мы обнаруживаем это, когда остается
лишний глагол, «fell» (упала). Такие «сбивающие» предложения намного более
распространены, чем может показаться на первый взгляд, так как обычно мы
интерпретируем фразы в контексте всего сказанного. Например, мы можем на-
блюдать за лошадьми, бегающими по племенной ферме, и можем видеть, как
одна лошадь промчалась мимо амбара, тогда как другая проскакала мимо фер-
мерского дома.
Зацикленность и заблуждения в психологических установках очень рас-
пространены. На рис. 10.11 показан висконсинский тест сортировки карточек
(ВТСК), который был разработан для введения дезориентировки в обычный
тест по сортировке карточек. Испытуемых просят угадать, какая из карточек
Рис. 10 .11 . Висконсинский тест сортировки карточек (ВТСК) вынуждает испытуемого
принять определенное правило выбора, например «желтый цвет указывает на правиль-
ность карточки». В тот же момент в игру вводится другое правило, например число или
форма изображенного на карточке предмета имеет значение. Это становится испыта-
нием для нашей способности выдвигать альтернативные первому правилу гипотезы,
и люди с повреждениями лобных долей обычно плохо справляются с ситуацией смены
правил. (Источник: Grant, 1993.)
24
Глава 10
правильная, на основании ее цвета, числа или формы. Они получают ответ-
ную реакцию после каждого выбора. На начальной стадии испытуемые полу-
чают вознаграждение за одно условие, например за правильный выбор желтого
объекта. В тот же момент правило выбора меняется без ведома испытуемого.
Время и число проб, потребовавшихся испытуемому для изменения тактики
выбора карточек, используются в качестве показателей способности проверять
различные гипотезы.
Пациенты с повреждениями передних отделов лобной коры часто демон-
стрируют отсутствие гибкости мышления и показывают низкие результаты в
ВТСК. Персеверация неэффективной стратегии является маркером таких за-
болеваний, как болезнь Альцгеймера (Ridderinkhof et al., 2002). ВТСК особенно
часто используется, когда повреждения лобных долей слишком малы, чтобы
быть замеченными на стандартных мозговых сканах.
Изменение правил очень тяжело переносится, если человек умственно утом-
лен или сонный, или недееспособен. Даже смена одной деятельности на дру-
гую требует затраты дополнительных умственных усилий сверх того, что обыч-
но вовлечено в выполнение рутинных и автоматизированных действий (рис.
10.12). Таким образом, сонным водителям гораздо труднее принимать быстрые
решения в непредсказуемой ситуации на дороге, предположительно из-за сни-
жения активности лобных долей. Показано, что люди, работающие в ночные
смены, совершают больше ошибок по сравнению с работающими в дневную
смену, и за эти отличия тоже, вероятно, ответственны лобные отделы коры го-
ловного мозга.
На рис. 10.13 показана модель исполнительных функций (Schneider, Chein,
2003). Мы будем использовать данную схему для подведения итогов наших те-
кущих знаний об эксплицитном решении задач. Область ДЛ-ПФК (дорсолате-
ральная префронтальная кора) ответственна за определение цели. Следователь-
но, при решении головоломки по ханойскую башню определение конечных и
промежуточных целей будет отражено в этой области. ПЦК (передняя цингу-
лярная кора) регистрирует ошибки и конфликты между целями (см. гл . 8, 11 и
12). ЗТК (задняя теменная кора) вовлечена в контроль внимания (см. гл . 8), в то
время как в среднюю височную кору (СВК) поступает эпизодическая память,
автобиографическое хранилище прошлого опыта (гл. 9). И наконец, таламус
(отмечен как ТАЛ) считается «входным и отчетным реле». Последнее обозна-
чение может быть спорным и в реальности обозначает таламокортикальную
воротную систему (см. гл . 5 и 8).
Мы уже наблюдали активацию ДЛ-ПФК при решении головоломки про
ханойскую башню и при смене задач (выше). На рис. 10.14 представлена во-
Рис. 10 .12 . Смена деятельности. Об-
ласти мозга с высокой активностью во
время смены задач перекрываются с
областями, задействованными в других
исполнительных процессах. Сравните
с рис. 10.19 (с. 29). (Источник: Braver
et al., 2003.)
Мышление и способность решать задачи
25
влеченность ДЛ-ПФК и ПЦК в пяти различных заданиях, требующих испол-
нительных функций, включая конфликт ответа (как в тесте Струпа), новизну,
загрузку текущей памяти и даже трудности восприятия.
На рис. 10.15 объединена активация ПЦК во время различных заданий,
включая эмоционально окрашенные. Как мы можем ожидать, области испол-
нительных функций могут быть вовлечены в работу при большом числе усло-
вий, которые требуют гибкого принятия решений. В противоположность этому,
Рис. 10 .13 . Возможная модель. На
внешней стороне каждого полушария
пик активности во время решения
задач возникает в дорсолатеральной
префронтальной коре (ДЛ-ПФК). Во
время конфликтной ситуации при
решении задачи или в случае оши-
бок активность возникает в передней
части цингулярной коры (поясной
извилины) (ПЦК). Эти две области
активны, когда решаемая задача труд-
ная. (Источник: Schneider and Chein,
2003.)
Рис. 10 .14 . Умственное усилие активирует исполнительные области. Слева: Дункан
и Оуэн (2000) показали, что пять различных заданий, вовлекающих все мыслительные
усилия, активируют латеральные и средние области лобной доли. Пять заданий вклю-
чают конфликт ответов (зеленые точки), новизну задания (розовые), число элементов в
рабочей памяти (желтые), необходимую задержку в рабочей памяти (красные) и труд-
ности восприятия, как неясность зрительного стимула (синие). МТ — мозолистое тело;
НЛБ — нижняя лобная борозда; СБ — сильвиева борозда. Справа: другой ракурс на
ПЦК (переднюю цингулярную кору) и ДЛ-ПФК (дорсолатеральную префронтальную
кору) (MacLeod and MacDonald, 2003). (Источник: (слева) Duncan and Owen, 2000;
(справа) MacLeod and MacDonald, 2003.)
26
Глава 10
как мы узнаем далее, бессознательные памятные следы и навыки не требуют
вовлечения этих исполнительных областей. Вместо этого считается, что они
отражаются в активации обширных очагов в височной коре и других областях
мозга (Moscovitch et al., 2006).
Эта модель, таким образом, демонстрирует основополагающие функции.
1. Поддержание стремления к цели и определение промежуточных целей.
2. Сохранение фокуса внимание на решении задачи.
3. Тормозное влияние на отвлекающие мысли и эмоции.
4. Метакогнитивный мониторинг требуемого качества сенсорных, моторных,
речевых функций и текущей памяти.
Далее мы увидим, как исполнительные процессы постоянно используют
сохраненную информацию, включая блоки данных, которые нужны для того,
чтобы обойти пределы способностей.
Рис. 10.15. Опознавание ошибок и разрешение. На данном рисунке, как и на рис. 10 .12,
показана активация передней цингулярной коры (ПЦК) несколькими различными за-
дачами. Предыдущие исследования показали, что опознавание несоответствия (как в
Эффекте Струпа) вызывает повышение активности в ПЦК. Однако такие различные
проблемы, как двусмысленные эмоциональные картинки, вызванная тревога, ожидае-
мая, неожиданная и испытанная боль, вызванные эмоции и ожидаемые эмоциональные
картинки, показывают достоверную активацию ПЦК. Другие исследования локализуют
эмоциональные аспекты активации ПЦК в ее передней части, а когнитивную актива-
цию, как опознавание ошибок, в верхней части ПЦК. (Источник: Botviinck et al., 2004.)
Мышление и способность решать задачи
27
3.0 . Умственная нагрузка и корковая активность
Задачи, требующие большего усилия, вызывают более обширную активацию
головного мозга, которая простирается за пределы исполнительных областей
лобных отделов. Например, в классической работе (Smith, Jonides, 1998) обна-
ружена чрезвычайно обширная активация коры головного мозга с увеличени-
ем загрузки памяти. Авторы использовали методику «n раз назад» (рис. 10.16).
Испытуемым предъявляли серию фигур для запоминания. В случае условия
1-назад им необходимо было вспомнить предыдущий предмет; в случае 2-на-
зад — предмет, показанный перед предыдущим; в случае 3-назад — предмет,
показанный три слайда назад. Задачу типа n раз назад можно считать челове-
n раз назад можно считать челове-
раз назад можно считать челове-
ческим эквивалентом «беличьего» колеса. Испытуемый должен постоянно ра-
ботать для того, чтобы справляться с потоком информации. Уровень сложности
нарастает очень быстро с увеличением числа предметов, которые нужно дер-
жать в памяти. Результаты этого исследования представлены на рис. 10.17.
Однако силу связей между клетками мозга мы не можем наблюдать, даже
несмотря на современные методы регистрации. Изображение на рис. 10.18 сви-
детельствует о том, что умственная нагрузка изменяет силу взаимодействия
между нейронами, т. е . интенсивность нейронной сигнализации между корко-
выми областями.
Судя по активности мозга, когнитивные процессы являются одними из ос-
новных факторов в его функционировании. Это может показаться странным.
В конце концов, задания, которые требуют вовлечения когнитивных процес-
сов, не всегда являются самыми трудными или сложными. Предложение, кото-
рое вы сейчас читаете, гораздо сложнее, чем задача n раз назад. Почему такой
сложный процесс, как обработка речи, не требует активации огромного числа
областей мозга? На рис. 10.19 предложен ответ: уровень активации коры (как
Рис. 10 .16 . Загрузка рабочей памяти: тест «n раз назад». В случае 0-назад испытуемый
должен сообщать только то, что он видит. Со сменой n от 0 до 3 испытуемому требуется
держать в голове большее количество предметов, сохранять порядок их предъявления и
одновременно замечать новый слайд для того, чтобы добавить его в память. Трудность
задания возрастает очень сильно с увеличением числа предметов, которые необходимо
держать в рабочей памяти. (Источник: Smith and Jonides, 1998.)
28
Глава 10
Рис. 10 .17. Задания, требующие значительного умственного усилия, вызывают актива-
цию обширных областей мозга. Обратите внимание, что увеличение загрузки рабочей
памяти в степени прогрессии увеличивает вовлечение в работу регионов мозга, включая
фронтальные и затылочные области мозга и мозжечок. Таламус и базальные ганглии не
показаны на рисунке, но считается, что и они вовлекаются в данную активацию. (Ис-
точник: Smith and Jonides, 1997, рис. 10 .)
Мышление и способность решать задачи
29
Рис. 10 .18 . При увеличении сложности задания изменяется сила взаимодействия ней-
ронных сетей. Цветные отрезки на обеих схемах мозга указывают силу связи между
исполнительными и другими областями мозга. Паттерн связей изменяется при увели-
чении загрузки рабочей памяти. ЗТК — задняя теменная кора; СЛИ — средняя лобная
извилина; НЛИ — нижняя лобная извилина. (Источник: Honey et al., 2002.)
Рис. 10 .19. Исполнительная активность падает с тренировкой. После тренировки, при
выполнении ожидаемого навыка, корковая активность значительно снижается. Этот
обобщающий рисунок показывает снижение активности в исполнительных областях
(по сравнению с рис. 10 .12). Однако в других корковых регионах также наблюдается де-
кремент активности с увеличением тренировки. Считается, что контроль за рутинными
навыками осуществляется подкорковыми структурами, как базальные ядра и мозжечок.
В то же время периодические корковые связи могут сохраниться для решения задач с не-
предсказуемыми промежуточными вариантами. (Источник: Schneider and Chein, 2003.)
30
Глава 10
минимум) падает с тренировкой и увеличением автоматизма. В то время как
анализ речи является чрезвычайно сложным процессом, он в течение длинного
периода времени практикуется у всех людей. Именно поэтому нейронные раз-
ряды или метаболические реакции мозга не могут являться непосредственным
маркером некоторых умственных процессов. Напротив, данные параметры от-
ражают вовлечение мозговых ресурсов, которые необходимы для решения но-
вых или неожиданных задач. Как только навык, даже самый сложный, освоен,
для его выполнения требуется все меньше корковой активности. Навыки, до-
веденные до автоматизма, также имеют меньший сознательный доступ и про-
извольный контроль, как и было показано в поведенческих опытах (Schneider,
1995; Baars, 2002).
4.0 . Использование существующих знаний
Одной из проблем, которая рассматривается в этой главе, является постоянное
использование человеческим мозгом стратегии группировки информации (чан-
кинг) для решения задач, превышающих лимитированные возможности непо-
средственной памяти. Каким образом мозгу удается справиться с этим?
Для того чтобы понять механизм чанкинга, необходимо пересмотреть обыч-
ную концепцию рабочей памяти и усилить степень, в которой мыслительные
операции используют существующие знания, закодированные в сетях приобре-
тенных синаптических связей (см. гл . 3). Рабочая память в решающей степени
зависит от сохранности информации в долговременной памяти. Ваше понима-
ние этого предложения зависит от памяти, в которой сохранены слова, струк-
тура предложения и смысл. Следовательно, как предложили Кован с коллегами
(Cowan et al., 2001), мы можем поменять ролями рабочую и долговременную
память и считать, что рабочая память (РП) основывается на хранимой много
лет информации (см. рис. 10.2).
Современные методы визуализации мозга могут выявить активную нейрон-
ную сигнализацию, но не синаптические связи, которые кодируют длительное
обучение. Поэтому результаты регистрации активности мозга выглядят так же,
как снимок потока движения машин в городе из космоса. Мы легко можем каж-
дый день видеть свет от автомобильных фар, движущийся в разные стороны.
Но с нашими текущими приборами трудно наблюдать за медленными процес-
сами строительства новых дорог, автомагистралей и автостоянок, которые де-
лают возможным движение машин. Тем не менее вся эта система зависит от
физического расположения улиц и дорог, которое можно считать своего рода
постоянной памятью города. Внешний наблюдатель может многое узнать ис-
ходя из схемы движения машин в городе — один район может быть финансо-
вым центром, в другом районе может находиться аэропорт. Но по выходным
движение транспортных потоков может изменяться. В плохую погоду больше
людей могут остаться дома. В зависимости от неизвестных условий движение
машин может происходить по другим маршрутам, однако все равно будут ис-
пользоваться постоянно существующие улицы и дороги. Просто наблюдение за
прохождением сигнала обнаруживает лишь часть тех процессов, которые про-
текают в мозге.
Таким образом, знание схемы соединения и силы взаимодействия — улицы
и магистрали — является неотъемлемой частью головоломки, которую все еще
Мышление и способность решать задачи
31
трудно понять. Но мы уверены, что долговременная память имеет решающее
значение (см. блок 10.1). Мы рассмотрим доказательства важности изменений
долговременной памяти в следующем разделе.
4.1. Практика и тренировка могут изменить
сетевое взаимодействие в мозге
Как обсуждалось ранее, считается, что долговременная память кодируется
в связях между нейронами. Если воспоминания закодированы в синаптических
связях, они не могут непосредственно появляться при измерении мозговой ак-
тивности, например при помощи фМРТ. Связи и активные процессы в мозге
не всегда коррелируют между собой, а на самом деле, как мы узнали в пре-
дыдущем разделе, более тренированное и экспертное решение задачи может
вызывать меньшую активацию коры. Таким образом, долгосрочное хранение
информации не отражается в фМРТ, магнитоэнцефалографии (МЭГ), электро-
энцефалографии (ЭЭГ), а также при использовании других стандартных изме-
рений.
В настоящее время разрабатываются другие способы оценки эффективно-
сти синаптических связей в нейронных сетях головного мозга. Один из возмож-
ных вариантов показан на рис. 10.20, взятом из классического исследования
(Maguire et al., 2000), проведенного на лондонских таксистах, которые вынуж-
дены сдавать экзамены на знание сложной карты улиц Лондона. Какие измене-
ния можно наблюдать в активности головного мозга таксистов после многолет-
ней работы в условиях реальной пространственной задачи?
Рис. 10 .20 . Долговременные измене-
ния в строении мозга у профессионалов.
Магуайр с соавторами обнаружили, что
у лондонских таксистов размер задней
части правого гиппокампа больше по
сравнению с размерами у контрольной
группы, что соотносится с представле-
ниями о роли гиппокампа в процессах
пространственного
ориентирования.
Однако данная методика не в состоянии
подтвердить то, что число синаптиче-
ских связей увеличилось в результате
профессионализма. Различия в размере
можно объяснить и другими причина-
ми, такими как миелинизация, измене-
ние числа глиальных клеток и размеров
нейронов. В то же время абсолютный
размер соответствующих областей моз-
га взаимосвязан с профессионализмом
и по результатам других работ. ЛП —
левое полушарие; ПП — правое полу-
шарие. (Источник: Maguire et al., 2000.)
32
Глава 10
4.2 . Семантическая память
Мы уже обратили внимание на тесную взаимосвязь между сенсорной и мотор-
ной системами в мозге и их вклад в реализацию эндогенных или «внутренних»
когнитивных функций. Известно, что зрительные образы обрабатываются в
зрительной коре, а внутренняя речь — в слуховой коре. Это эмпирическое пра-
вило дополняет рис. 10.21 тем, что семантические функции, вероятно, связаны
с активацией височной доли (в том числе медиальной височной и гиппокам-
пальной областей, невидимых на рисунке). Более общие понятия, как полага-
ют, кодируются в левой латеральной височной доле за областью, отвечающей
за кодирование уникальных концепций. Центр, отвечающий за биологическое
движение — довольно сложные движения животных, — расположен недалеко
от области ЗМ (зрительно-моторной области), в то время как действия с исполь-
зованием самодельных устройств и инструментов активируют у людей другую
часть затылочно-височной области. Существует доказательство того, что ис-
пользование инструментов может также активировать центры, прилегающие к
соматосенсорной и моторной коре. При помощи фМРТ (Hauk et al., 2004) до-
казано, что «слова, обозначающие движения лица, руки или ноги (например,
лизать, подбирать, или ударять) ... дифференциально активировали зоны коры,
лежащие вдоль границы моторной области, которые либо непосредственно
прилегали, либо перекрывались с регионами мозга, активирующимися реаль-
ными движениями языка, пальцев или ног». Это подтверждает основное по-
ложение, что для сложных и биологически важных семантических процессов
необходимо участие областей мозга, ранее адаптированных к взаимодействию
с сенсомоторным миром.
В одном из недавних обзоров (Martin, Chao, 2001) написано:
Распределенные сети разрозненных областей коры головного мозга активны в те-
чение обработки информации об объекте. Распределение этих областей меняется
в зависимости от семантических категорий. Те же зоны активны, по крайней мере
частично, когда стимулы из одной категории узнаются, называются, воображаются
испытуемым, а также при чтении и ответах на вопросы о них...
При анализе одновременно всех этих данных видно, что вентральную затылоч-
но-височную кору лучше рассматривать не как мозаику разрозненных областей,
отвечающих за конкретные категории, но, скорее, как некое пространство свойств,
отражающее хранящуюся информацию обо всех особенностях формы объектов,
которую разделяют члены одной из категорий...
Ориентированная на признаках модель может объяснить то наблюдение, что про-
извольная категория объектов, таких как стулья, вызывает паттерн нейронной
активности, отличный от других объектов категории (т. е. лиц и домов). Очевид-
но, было бы трудно, а также неразумно утверждать, что есть «представительство
стула» в головном мозге. Существует слишком много категорий и слишком мало
нейронного пространства для размещения дискретных, специфичных для данной
категории модулей. В реальности не существует пределов числа категорий разных
объектов. Ориентированные на признаках модели могут обеспечить необходимую
гибкость для представления бесконечного множества объектов.
Иными словами, височно-префронтальная система, показанная на рис. 10.21,
не может являться словарем всех семантических категорий, которые мы знаем.
Их слишком много. Скорее, она может представлять только важные характери-
стики основных категорий, которые могут указывать на их определенное чис-
Мышление и способность решать задачи
33
ло категорий, представленных в обширных корковых сетях. Мы можем найти
зоны в этой огромной области, которые преимущественно отвечают активацией
на предъявление домов, чем лиц, или больший ответ вызывают инструменты,
чем дома или автомобили. Область представительства «инструментов» может
кодировать широкий спектр объектов, которые используются в качестве орудий
труда — не только молоток и зубила, но, возможно, и компьютеры и телефон-
ные компании. Было бы очень интересно посмотреть, может ли одна из кате-
горий (например, человеческого лица) продемонстрировать «использование в
качестве орудий», как если бы этот объект определили как инструмент. Напри-
мер, маскарадные маски с формой человеческого лица могут быть «использо-
ваны в качестве инструмента», чтобы напугать людей в канун Дня всех святых
(Хеллоуин). С помощью таких экспериментов можно проверить гипотезу о том,
что области представительства «инструментов» могут на самом деле кодиро-
вать более абстрактные положения, чем положение «может быть использован
для достижения конкретной цели».
Неврологи предполагают, что «представление об объекте формируется с по-
мощью сенсорных и моторных признаков и свойств, приобретенных опытным
путем». Это является мощным организационным принципом для общих поня-
тий в головном мозге.
Рис. 10 .21 . Области мозга, вовлеченные в семантическую память. Показано обобщение
локализации областей мозга, ответственных за семантическую память, в левом полуша-
рии (б) и на изображении нижней поверхности височной доли (а). Может потребоваться
некоторое время для понимания пространственного расположения нижней поверхно-
сти височной доли. Обратите внимание на то, что существует семантический градиент
между областями, отвечающими за специфические зрительные объекты и абстрактные
зрительные формы. Семантическая рабочая память вызывает постоянно циклирующую
активность между височной и лобной областями, в которую вовлекаются и подкорковые
связи между этими областями. (Источник: Martin and Chao, 2001.)
34
Глава 10
Возможные противоречивые факты получены в исследованиях на больных
эпилепсией при регистрации одиночных нейронов от электродов, импланти-
рованных в височную долю. Некоторые из этих исследований показали очень
конкретные ответы на определенную категорию, например на предъявление раз-
ных фотографий президента Б. Клинтона (Kreiman et al., 2002). Видимо, такие
высокоспецифичные нейронные ответы представляют один из узлов в большой
сети или набор перекрывающихся сетей, непосредственно связанных с восприя-
тием политических деятелей или известных людей. Вполне возможно, что с раз-
витием возможностей более детально исследовать височную долю, можно будет
локализовать сети, содержащие высокоспецифичную информацию.
Как показано на рис. 10.21, префронтальные области, расположенные в не-
посредственной близости к зоне Брока, могут обеспечивать «систему семанти-
ческой рабочей памяти», что свидетельствует об отличии ее от фонологическо-
го и зрительно-пространственного компонентов рабочей памяти.
4.3 . Абстрактные понятия, прототипы и нейронные сети
Интуитивно мы стремимся иметь некоторые сбивающие с пути суждения о че-
ловеческой когнитивной деятельности. Одним из них является представление
о том, что мы держим в наших головах картины, которые представляют собой
окружающий нас перцептивный мир. Однако данные свидетельствуют о том,
что вместо этого мы склонны использовать визуальные образы, которые явля-
ются прототипными напоминаниями категорий окружающего мира, а не точны-
ми образами из этих категорий, таких как стулья и кинозвезды. По всей види-
мости, у нас имеется сеть перцептивных, когнитивных и двигательных знаний
о стульях и их использовании. Такие сети могут быть доступны при использо-
вании картинок с изображением прототипов стульев (рис. 10.22). Люди предпо-
читают такие прототипы образов, но они не являются точными изображениями
всех стульев, когда-либо нам известных (Rosch, 1975; Barsalou, 1999). Скорее,
они являются особыми членами целых категорий — стулья или кинозвезды.
Деревянный стул на рис. 10.22 может не являться обычным стулом, который вы
обычно видите и используете. Но в такой схеме тестирования этот визуальный
образ придет на ум легче, чем пластиковый или металлический стулья, которые
мы склонны видеть чаще.
Барсалоу (Barsalou, 1999, 2005) выдвинул предположение, что у людей су-
1999, 2005) выдвинул предположение, что у людей су-
ществует сильная перцептивная склонность, даже когда они имеют дело с аб-
страктными категориями. Одной из причин является то, что человек изначально
не наделен пониманием абстракций. В ранний период происходят сенсорное,
двигательное и эмоциональное исследования, которые, с эволюционной точки
зрения, служат для выживания. Развивающийся мозг может выбрать ранние по-
пуляции нейронов и связей, прежде чем мы постигнем взрослые представления
о мире.
Такие же данные получены в работах Барсалоу и других исследователей при
изучении «перцептивных символических систем». Разве мы представляем ав-
томобиль c рис. 10.23 таким же образом, как инженер, т. е. имеющим набор ха-
c рис. 10.23 таким же образом, как инженер, т. е. имеющим набор ха-
рис. 10.23 таким же образом, как инженер, т. е. имеющим набор ха-
рактеристик двигателя и колес? Вместо этого существуют доказательства того,
что абстрактное понятие автомобиля в большей степени зависит от восприятия
перцептивных особенностей внешнего вида автомобиля и когнитивного вос-
произведения его движения (Barsalou et al., 2003).
Мышление и способность решать задачи
35
4.4 . Знания формируются в виде нейронной сети
Другим общепринятым представлением является то, что обычные слова и по-
нятия кажутся простыми, поскольку они часто имеют простые названия, та-
кие как «автомобиль», «мозг» и «человек». Это положение также весьма об-
манчиво. Ментальные образы, включая слова, зрительные образы и понятия,
следует рассматривать с точки зрения тщательно разработанных сетей знаний.
Когнитивные доказательства существования такой сети были многочисленны
еще до появления методов визуализации мозга. Действительно, такие свиде-
тельства могут быть найдены в научных работах середины ХХ в. (Kuhn, 1962).
Рис. 10 .22 . Набор зрительных про-
тотипов, которые обозначают более
абстрактные категории (Источник:
Laeng et al., 2003.)
36
Глава 10
Очень наивно, но мы склонны рассматривать основные научные идеи, такие
как «гравитация» и «молекула», в качестве отдельных понятий. Вследствие
этого некоторые философы предполагают, что между ньютоновской теорией
силы и физическими наблюдения, которые способствовали ее развитию, долж-
ны быть взаимнооднозначные соответствия. Однако другие философы указали
на отсутствие наблюдаемых корреляций ньютоновской силы — можно только
измерить массу и ускорение. «Сила» — логически выведенное понятие в физи-
ке (см. гл . 1). Теория Ньютона является не только списком положений, а сетью
тщательно установленных идей, подтвержденных стандартными эксперимен-
тами и предсказуемыми наблюдениями, с собственными математическими вы-
кладками. Таким образом, научные теории представляют собой семантические
сети, а не просто коллекцию классифицированных наблюдений. То же самое
относится и к словам в естественном языке.
Эти положения имеют фундаментальное значение для понимания того, как
мозг формирует знания. При визуализации деятельности мозга мы не видим
абстрактных классов объектов. Скорее, мы можем наблюдать перцептивные
объекты в сенсорных областях мозга, которые постепенно преобразуются в
более абстрактные формы изображений (см. рис. 10.23). Идеи оказываются
представленными в коре головного мозга в виде сложных сетей возникшего
взаимодействия, а не в виде локализованной файловой системы с аккуратно
расположенными понятийными категориями. Мозг — очень практичный орган,
всегда близко взаимодействующий с сенсомоторной и мотивационной окружа-
ющей средой, а не является абстрактной логической машиной. (Мозг, конечно,
может решать логические задачи, но большую часть времени он ориентирован
на более приземленные процессы.)
Рис. 10 .23 . Немодальная и перцептивная символьные системы. Барсалоу предпола-
гает, что люди думают с помощью «перцептивной символической системы» (а), а не
абстрактными категориями со списком особенностей. (б) Такое предположение согла-
суется с основной тенденцией мозга развиваться на базе данной информации, особенно
информации ранних лет жизни. (Источник: Barsalou, 2003.)
Мышление и способность решать задачи
37
Важно понимать необходимость существования семантической сети, а не
простых понятийных связей, при рассмотрении нейронной основы когнитив-
ных единиц, таких как слова, идеи и образы. Например, на рис. 10.24 показано,
как одно личное имя может быть представлено в виде нейронной сети.
Поскольку смежные понятия часто имеют общие признаки, та же линия
рассуждения свидетельствует, что нейроны, чувствительные к определенным
свойствам, могут участвовать в более чем одной сети понятий. На рис. 10.25
проиллюстрирована эта гипотеза для понятия «тигр» и «слон».
Чрезвычайно трудно выявить специфическую корковую локализацию кон-
кретных понятий. Некоторые исключения показаны на рис. 10.26.
4.5 . Умозрительные недостатки
В одном из исследований (Martin, Chao, 2001) сообщается:
У пациентов с повреждением левой префронтальной коры (ЛПК) часто наблю-
даются трудности извлечения слов в ответ на конкретные просьбы (например,
произнести слова, начинающиеся с конкретной буквы; назвать предметы, при-
надлежащие к конкретной семантической категории)... Это свидетельствует о
том, что ЛПК играет главную и очень важную роль в извлечении лексической и
семантической информации. Пациенты с повреждением височных долей часто
сталкиваются с трудностями именования объектов и извлечения информации о
характеристике объекта... Это предполагает, что информация, характеризующая
объект, может быть сохранена, по крайней мере частично, в височных долях.
Рис. 10 .24. Предположительная сеть для одного слова. Каждое слово в нашем лекси-
коне соотносится с сетью взаимосвязанных перцептивных, семантических, моторных и
лексических узлов. (Источник: Lokendra Shastri, с разрешения.)
38
Глава 10
Однако некоторые языковые проблемы могут быть довольно четкими и
ограниченными. Например, в нижней части височной доли соседние области,
вероятно, соотносятся с разными понятиями, такими как стулья, лица и дома.
Инструменты, животные и транспортные средства вызывают увеличение ак-
тивности в разных областях. Это вовсе не означает, что эти конкретные зоны
являются единственными, участвующими в образовании таких понятий, как
«молоток» или «велосипед». Скорее, мы будем наблюдать пики активностей в
разных частях височной доли, и уже зарегистрированы случаи локальных по-
вреждений этих областей (блоки 10.2 и 10.3).
В одной из работ проведено тщательное исследование больной, известной
как ЭВ, у которой были трудности в именовании животных (Caramazza and Ma-
Ma-
hon, 2003; блок 10.2). Предъявляя испытуемой фотографии с определенной ча-
, 2003; блок 10.2). Предъявляя испытуемой фотографии с определенной ча-
стотой и степенью сходности, можно тестировать в контролируемых условиях
знания ЭВ нарисованных категорий. У нее наблюдалась большая трудность в
назывании животных по сравнению с другими категориями. ЭВ также было
трудно просто узнать изображение с животным, но не картинки других катего-
рий. Ее трудности не ограничивались изображениями, а также распространя-
лись на произнесение названия животных. Однако пациентка нормально справ-
лялась с выполнением сложной зрительной обработки изображений, таких как
узнавание лица и сравнение картинок.
И наконец, у ЭВ возникли проблемы с основными характеристиками жи-
вотных, но не признаками других объектов. На такие вопросы, как «Киты ле-
тают?» и «У коровы есть грива?», она отвечала неправильно примерно в одной
трети случаев. При использовании других категорий она выполнила задания
с лучшим результатом. Хотя предположение о локальном повреждении более
правдоподобно, существует и альтернативное толкование фактов (Tyler and
Moss, 2001).
Рис. 10 .25. Перекрывающиеся се-
мантические сети для двух поня-
тий. В мозге могут существовать
нейроны, чувствительные к опреде-
ленным свойствам и принимающие
участие в работе более чем одной
семантической сети. (Источник:
Hodges and Patterson, 1997.)
Мышление и способность решать задачи
39
4.6 . Определение количества и числа
К удивлению многих исследователей, в последние годы появились отличные
подтверждающие данные о локализации в теменной коре области, отвечающей
за определение и оценку чисел. Полагают (Dehaene et al., 2004), что области
мозга, показанные на рис 10.27, преимущественно отвечают за процесс оценки
количества, хотя могут иметь и другие функции. Однако когда мы производим
арифметические вычисления в уме, как и следовало ожидать, активизируются
области внутренней речи, что, вероятно, связано с процессами мысленного со-
провождения речью этапов вычитания или умножения (рис. 10.28). Было бы
интересно исследовать людей, которые обладают опытом в использовании ви-
зуальных образов в арифметике, например использующие счеты при вычисле-
ниях. Мы не ожидаем, что у таких людей во время арифметического счета будет
наблюдаться активация речевых областей, напротив, активироваться должны
будут области в затылочной и теменной коре.
Рис. 10.26 . Области повышенной активности в ответ на понятия «животные», «автомо-
били», «инструменты» и «овощи». Понятийные категории трудно локализовать в мозге.
Исключения составляют отдаленные категории, такие как показаны на рисунке. Так же
как и на других изображениях мозга, мы можем наблюдать только различия в актива-
ции областей в ответ на предъявления, например, животных и инструментов. Области,
имеющие разную активность в зависимости от предъявленного объекта, расположены
в средней височной области. Некоторые ученые считают, что специфичные категории
могут служить главными указателями для извлечения более точных сетей, имеющих
отношение к слонам и яблокам, нежели демонстрировать особый специфичный ней-
ронный код для каждого объекта категории. (Источник: Caramazza and Mahon, 2003.)
40
Глава 10
5.0 . Имплицитное мышление
Рассказывают, что, будучи ребенком, Вольфганг Амадей Моцарт подшучивал
над отцом Леопольдом (который одновременно являлся и его учителем музыки)
таким образом, что, играя на клавесине какой-то отрывок, нарочно не заканчи-
Рис. 10.27. Сеть для определения чисел? Обнаружено, что эта сеть в теменной обла-
сти, особенно во внутритеменной борозде, вовлечена в определение числа и количества.
(Источник: Dehaene et al., 2004.)
Рис. 10 .28 . Мысленный арифметический счет использует фонологическую петлю вну-
тренней речи. Обратите внимание на перекрывание фонологических процессов (таких
как мысленное повторение списка слов) и задачи на вычитание. Предположительно,
в данном случае, внутренняя речь вовлечена в процесс вычитания. (Источник: Wynn
в Ramachandran, 2002.)
Мышление и способность решать задачи
41
вал его. В западной музыке почти всегда отрывок произведения завершается
определенным набором нот или аккордов, дабы закончить его на том же тони-
ческом уровне, что и вначале. Во времена Моцарта это правило нельзя было
игнорировать. Леопольду Моцарту приходилось вставать с постели, спускаться
вниз и доигрывать последовательность нот, крутящуюся у него в голове, пре-
жде чем возвращаться снова спать. Ему было просто невыносимо слышать не-
законченную мелодию. Вы можете достигнуть этого эффекта, если при прослу-
шивании любимой песни выключите ее, когда она почти закончилась.
Завершение ожидаемой последовательности является одним из примеров
имплицитного решения задачи. Мы не говорим себе постоянно, что надо дослу-
шивать музыку до конца. Но как только мы начинаем, возникает ощущение, что
это необходимо сделать. Эта цель имплицитная или неосознанная, так же как
и правила гармонии и мелодичности. Не все люди могут объяснить эти основ-
ные правила, но они оказывают значительные эффекты независимо от этого.
Если сравнить прослушивание песни с решением задачи про ханойскую баш-
ню (выше), то восприятие музыка настолько неосознанный процесс, насколько
осознанным является решение задачи про ханойскую башню.
Для большинства людей процесс решения задачи состоит из эксплицитных
и имплицитных компонентов. Существует тенденция недооценивать слож-
ность имплицитной когнитивной деятельности, потому что она является не-
осознаваемой. Более того, в наших профессиональных, высокотренированных
навыках это может являться наиболее эффективным механизмом при решении
проблем. Эксплицитная сторона в решении задачи более эффективно исполь-
зуется во временных исполнительных функциях в навыках, которые в другой
ситуации являются привычными. Например, вождение автомобиля может быть
«автоматизированным» действием значительную часть времени, так как явля-
ется рутинным и предсказуемым занятием. Но как только поток машин стано-
вится более плотным и непредсказуемым, когда начинает спускать колесо или
когда кто-то начинает отвлекать разговорами, исполнительный контроль над
вождением становится необходим. Таким образом, существует гибкая смена
более контролируемой и более автоматизированной деятельности друг другом
в процессе вождения автомобиля. Эти принципы могут быть также соотнесены
с другими процессами решения задач.
5.1. Чувство о наличии знания
Чувства знания и «расплывчатые» или «неясные» сознательные события встре-
чаются очень часто (James, 1890). Они включают хорошо изученные чувства
«на кончике языка» и «ощущение знания». Очень просто вызвать ощущение
«на кончике языка». Необходимо дать человеку определение достаточно ред-
кого, но известного слова и спросить, возникло ли у него ощущение, что он
почти знает ответ, но не совсем. Эффективными вопросами могут стать «Как
называется динозавр-вегетарианец?» или «Какими двумя словами обозначают
технологию создания искусственных конечностей?».
Считается, что такие субъективно неясные, но осознанные события, направ-
ляют интуитивное решение задач, включая вербальные и зрительные проблемы
(Bowers et al., 1990), поддерживают непрерывность при извлечении воспомина-
ния во время состояния, когда необходимая информация крутится «на кончике
42
Глава 10
Блок 10.2 . Специфические умозрительные недостатки пациентки ЭВ
Показательный случай специфичного для данной категории семантическо-
го нарушения: пациентка ЭВ
Для того чтобы оценить удивительные свойства специфичных для данной категории
семантических нарушений, рассмотрим случай пациентки ЭВ (Caramazza, Shelton,
1998). У этой пациентки наблюдалось непропорциональное нарушение восприятия
семантической категории ‘животные’ по сравнению с другими категориями. Далее
мы рассмотрим эмпирические характеристики нарушения пациентки ЭВ.
Называние изображений
На основе предъявления набора изображений (Snodgrass and Vanderwart, 1992), по-
добранных относительно степени известности и частоты, а также зрительной слож-
ности и степени известности показано, что у ЭВ наблюдаются непропорциональ-
ные нарушения при назывании животных по сравнению с называнием не животных
(табл. 10.1). Это указывает на то, что нарушение специфической категории при на-
зывании картинок нельзя объяснить неконтролируемыми переменными стимулов
(Steward et al., 1992; Funnel and Sheridan, 1992).
Таблиц а 10.1
Выполнение задания по называнию картинок,
сопоставленных по набору объектов, пациенткой ЭВ
Сопоставление по частоте встре
чае мости и степени знакомства
Сопоставление по зрительной
сложности и степени знакомства
Животные
Не животные
Животные
Не животные
ЭВ
12/22 (55%) 18/22 (82%)
7/17 (41%)
16/17 (94%)
Контроль 11/11 (100%) 10,8/11 (98%)
16,6/17 (98%) 16/17 (94%)
Размах
11
10–11
16–17
16–17
Выполнение задания по называнию картинок у пациентки ЭВ выявило не только ко-
личественные различия в числе правильных ответов при предъявлении животных и
не животных, но и качественные отличия. При предъявлении изображения живот-
ного ЭВ либо называла животное неправильно, либо вовсе не узнавала его, тогда как
при демонстрации рисунков с не животными ЭВ узнавала их, но не могла воспроиз-
вести названия (рис. 10 .29(а)).
Рис. 10 .29 . (а) Примеры сделанных ЭВ ошибок при назывании животных. (б) При-
меры стимулов из теста на определение реальности изображенного объекта. (в) При-
меры изображений из «Теста на определение частей» или «Теста на определение
головы».
Мышление и способность решать задачи
43
Проблема с называнием объектов у пациентки ЭВ распространялась только на ка-
тегорию «животные», и не наблюдалась в других, таких как «овощи/фрукты», при
назывании которых результативность была максимальна.
Идентификация звуков
У пациентки ЭВ наблюдались также нарушения при назывании животных по срав-
нению с не животными на основе их звуковой характеристики (8/32; 25% правильно
против 20/32; 63% правильно: z 3,06, р < 0,05), что указывает на то, что нарушение в
назывании связано не только со зрительным восприятием.
Определение объектов
ЭВ дали задание определить («да» или «нет»), является ли предъявленный объект
реальным (см. рис. 10 .29(б) для примеров стимулов). Степень результативности вы-
полнения данного задания интерпретируется как показатель целостности системы
зрительно-структурного описания (т. е . система с модально-специфическим входом,
отвечающая за хранение образов относительно формы или очертания объектов, и
обеспечивающая доступ к информации о понятиях). Результаты выполнения задания
по выявлению реальных и вымышленных животных у ЭВ были значительно ниже
нормы (36/60: 60% правильно; среднее по контролю: 54/60: 84% правильно). Но ре-
зультаты оказались в пределах нормы при разделении реальных и вымышленных
объектов, не являющихся животными (55/60: 92% правильно; среднее по контролю:
50,5/60: 84% правильно).
Определение частей
Пациентку ЭВ просили определить, какая из двух предъявленных голов (или частей)
принадлежит безголовому телу (или объекту с отсутствующей частью) (см. рис.
10.29(в) для примеров стимулов). У ЭВ наблюдались значительные затруднения в вы-
полнении этого задания с животными (60% правильно; среднее нормы = 100%), но
она прекрасно справлялась с набором обычных стимулов (97% правильно; среднее
нормы = 97%).
Зрительное восприятие
ЭВ справлялась в рамках нормы реакции с набором заданий на сложное зрительное
восприятие, как зрительное сличение и узнавание лиц. Эти данные указывают на то,
что у ЭВ отсутствует общее нарушение восприятия зрительных сложных стимулов,
и позволяют выдвинуть предположение о том, что повреждение процесса принятия
правильного решения о реальности животного зависит от категории объекта.
Суждения о центральном свойстве
ЭВ просили определить, насколько правильно данное свойство приписывают опре-
деленному объекту (см. табл. 10 .2 для примеров стимулов). Очень значительные на-
рушения у ЭВ обнаружены относительно свойств животных (65% правильно; раз-
брос контроля 85–100%), но выполнение задания было в пределах нормы в задачах с
не животными (95% правильно; разброс контроля 86–100%). Нарушения одинаково
проявлялись в случаях зрительно/перцептивного и функционально/ассоциативного
знания о живых существах (65% правильно для обоих вариантов знания), но ока-
зались в пределах нормы для обоих типов знания для категории не животных (зри-
тельно/перцептивное: 93,5% правильно; разброс нормы 86–100%; функционально/
ассоциативное: 98% правильно; разброс нормы 92–100%). Эффективность ответов
на вопросы о центральных свойствах объектов указывает на то, что нарушение паци-
ентки не ограничено ответной деятельностью.
44
Глава 10
Блок 10.3 . Ассоциативная зрительная агнозия
Исследование ситуации ассоциативной зрительной агнозии
Пациентом являлся мужчина 47 лет, страдающий острым снижением кровяного дав-
ления, что в результате привело к повреждению мозга. Его психическое состояние
и речевые функции были в норме, зрительная острота составляла 20/30 с правосто-
ронней гомонимной гемианопсией (слепотой в правой половине зрительного поля).
Единственным сильным нарушением у него была неспособность узнать большин-
ство зрительных стимулов. Первые три недели в больнице пациент был не в состо-
янии идентифицировать обычные предметы, предъявленные зрительно, а также не
знал, что находится у него в тарелке, пока не попробует еду на вкус. Он мог иденти-
фицировать объекты моментально при тактильном прикосновении.
Когда ему был показан стетоскоп, он описал его как «длинный шнур с круглой шту-
кой на конце» и поинтересовался, является ли эта вещь часами. Он идентифицировал
открывалку как ключ. На просьбу дать название зажигалке, он ответил: «Я не знаю»,
но смог определить ее после того, как обследующий врач зажег ее. Он сказал, что «не
уверен», когда ему была показана зубная щетка. Он никогда не мог правильно опи-
сать или продемонстрировать в действии предмет, который не мог назвать. Если он
неправильно идентифицировал объект, его демонстрация этого предмета в действии
соотносилась с ошибочным названием. Идентификация незначительно улучшилась,
когда подсказывали категорию объекта (например, что-то съедобное) или когда па-
циента просили показать названный предмет. Когда ему называли правильное имя
объекта, он обычно быстро кивал и часто говорил: «Да, теперь я это вижу». Затем,
он, как правило, мог выделить различные части ранее неопознанного предмета так
же быстро, как и нормальный субъект (например, мундштук и чаша трубки и шнур-
ки, подошва и каблук у туфли). Похожая нерешительность никогда не возникала с
орально или тактильно определенными объектами.
После проведения трех недель в больничной палате его способность называть пред-
меты улучшилась настолько, что он мог идентифицировать большое число обычных
объектов, но эта способность варьировала; он мог один раз назвать предмет пра-
вильно, а в другой раз ошибиться. Эффективность выполнения снижалась в том слу-
чае, когда экспериментатор закрывал какую-то часть объекта. Пациент мог сравнить
Таблиц а 10.2
Примеры вопросов на центральное свойство
Зрительно/перцептивные
Функционально/ассоциативные
У коровы есть грива?
У кита есть большой хвостовой
плавник?
У кита есть восемь ног?
Кит летает?
Орлы откладывают яйца?
Корова является сельскохозяйственным
животным?
Литература
1. Caramazza, A. and Shelton J.R. (1998). Domain-specific knowledge systems in the brain:
the animate-inanimate distinction. Journal of Cognitive Neuroscience, 10, 1–34 .
2. Snodgrass, J. and Vanderwart, M. (1992). A standardized set of 260 pictures: norms for
name agreement, familiarity, and visual complexity. Journal of Experimental Psychology.
[Human Learning] 6, 174–215.
3. Stewart, F. et al. (1992). Naming impairments following recovery from herpes simple[
encephalitis. The Quarterly Journal of Experimental Psychobiology A, 44, 261–284 .
4. Funnel, E. and Sheridan, J.S. (1992). Categories of knowledge? Unfamiliar aspects of living
and nonliving things. Cognitive Neuropsychology, 9, 135–153.
Мышление и способность решать задачи
45
языка» (Metcalfe, 1986) и влияет на оценочные задания и задачи на принятие ре-
шения (Yzerbyt et al., 1998). На эту тему существует небольшое число работ, но
было показано (Maril et al., 2001) , что в состоянии «на кончике языка», которое
можно легко вызвать, высокая активность наблюдается в префронтальных от-
делах, т. е. в тех же областях, которые связывают с сохранением непрерывности
процесса при решении задачи (Duncan and Owen, 2000) (рис. 10.31).
идентичные предметы, но не мог сгруппировать их по категориям (одежда, пища).
Он мог нарисовать очертания предметов (ключ, ложка и т. д.), которые не мог опре-
делить.
Пациент не узнавал членов своей семьи, больничный персонал и даже свое лицо в
зеркале. Иногда у него были трудности в отделении очертания морды животного от
лица человека, но пациент всегда точно идентифицировал их как лицо. Возможность
узнавать рисунки предметов пострадала очень значительно, и даже после длительно-
го тестирования он мог назвать только 1 или 2 предмета на картинках. Он всегда мог
правильно назвать геометрические фигуры (круг, квадрат, треугольник, куб). Удиви-
тельно то, что он мог великолепно срисовать предмет с изображения, но все равно
не мог назвать его (рис. 10 .30). Он легко мог сопоставить рисунки объектов, которые
не мог идентифицировать, и у него отсутствовала трудность в отделении сложных
абстрактных паттернов, отличающихся друг от друга очень незначительно. Время
от времени он не справлялся с таким разделением, так как включал в изображение
недостатки бумаги или типографских чернил. Он никогда не мог сгруппировать ри-
сунки объектов по классам, если только не называл их до этого правильно. Чтение,
как вслух, так и просто для понимания, было ограничено значительно. Пациент мог
читать, запинаясь, большинство печатных букв, но часто путал буквы «К» c «Р» и
«Л» с «Т» и наоборот. Он мог читать слова медленно, произнося их вслух по буквам.
(Источник: Hillary R. Rodman, Luiz Pesson, and Leslie G. Ungerleider. Цит. из Rubens
and Benson, 1971.)
Рис. 10 .30 . Зрительная агнозия.
Пациент, страдающий ассоци-
ативной зрительной агнозией,
способен копировать обыкно-
венные рисунки, показанные
выше, но не может назвать их
(Rubens and Benson, 1971). (Ис-
точник: Squire et al., 2003.)
46
Глава 10
Существуют хорошие свидетельства того, что спонтанные мысли ориенти-
рованы на проблему, даже если цель мыслительного процесса не эксплицитна
(Greenwald et al., 2002). Содержание таких спонтанные мыслей часто описыва-
ют как чьи-то «текущие заботы». В социальных психологических эксперимен-
тах обнаружили, что можно при помощи опытных условий выделять одну из
текущих забот, например трудности в общении, показывая для этого фильмы с
изображением этих забот. Таким образом, студенты, которые определили свои
отношения с родителями как конфликтные, при демонстрации провокационно-
го фильма сообщили о преобладании у них мыслей относительно конфликтных
ситуаций.
Однако схожие паттерны решения задач начали встречаться довольно часто,
когда стали более внимательно проводить исследования на рутинный «поток
сознания» (Singer, 1994). «Всплески» в потоке мыслей являются обыденными
событиями и могут быть вызваны экспериментальным путем в опытах с ощу-
щением «на кончике языка» (рис. 10.32) и в хорошо известном тесте отдален-
ных ассоциаций (Kihlstrom, 1996).
Рис. 10 .31. Требующее усилия размышление при поиске слов. На данном рисунке пред-
ставлены локализация повышенной активности в задаче с ощущением «на кончике язы-
ка» (НКЯ) и временная динамика изменений активности (полученная при помощи по-
тенциалов, связанных с событием). Обратите внимание на активизацию классических
исполнительных областей. В то время как обычно эти области не рассматривают отно-
сительно сознательного опыта, тот факт, что состояние «на кончике языка» может быть
зарегистрировано поддающимся проверке способом, свидетельствует о «смутном» со-
знании. НИ — неизвестный, И — известный. (Источник: Maril et al., 2001.)
Мышление и способность решать задачи
47
6.0. Заключение
Рабочая память является частью процесса принятия решения. Полностью экс-
плицитное решение задач, по-видимому, очень редкое явление в реальном мире.
Однако выполнение эксплицитных головоломок, например «ханойская башня»,
очень чувствительно к повреждениям лобных долей, а также различным влия-
ниям, как дремота, наркотические эффекты, скука, когнитивная перегрузка или
отвлекающие события, которые приводят к нарушениям процессов решения
задач. Таким образом, задания на эксплицитное решение проблем служат по-
лезными показателями функционирования мозга. Кроме того, такие задачи, как
Висконсинский тест сортировки карточек, могут быть использованы для регу-
лирования «фиксированных» представлений о структуре задачи для того, что-
бы проверять когнитивную гибкость под влиянием неожиданных изменений в
условиях задачи. Испытуемые с повреждениями лобных долей чувствительны
и к такой смене условий постановки задачи.
Из-за ограниченных возможностей рабочей памяти, внимания, сознательных
процессов и произвольного контроля, основной стратегией при решении задач
является чанкинг и другие компоненты долговременной памяти для смещения
рутинных аспектов решения задач в сторону более объемных систем памяти.
Профессиональные шахматисты, например, знают много прогнозируемых шах-
матных позиций, что освобождает их рабочую память для того, чтобы обраба-
тывать новые и неожиданные позиции фигур. Долговременная семантическая
память еще до конца не изучена, но мы знаем, что во время этих процессов
Рис. 10 .32 . Неожиданное
«озарение» при решении за-
дачи. Мощность альфа- и гам-
ма-активностей в разложенной
ЭЭГ в момент «озарения» при
выполнении
ассоциативной
задачи на слова. (а) Обратите
внимание, что непосредствен-
но перед правильным ответом
альфа-активность уменьшает-
ся так же, как увеличивается
мощность гамма-активности.
(б) Увеличение гамма-ритма
связывают с активными и син-
хронизированными процесса-
ми в сети нейронов связанных
областей, вовлеченных в про-
цесс решения задачи. (в) Непо-
средственно перед решением
(отмечено минусовыми значе-
ниями в секундах) наблюда-
ется сдвиг активности ЭЭГ в
левое полушарие. (Источник:
Jung-Beeman et al., 2004.)
48
Глава 10
активизируются височные и префронтальные области, а также зоны средней
височной извилины, ответственные за эпизодические возможности обучения.
В то время как имплицитное мышление является высокоэффективным, оно
чувствительно и к инертности и недостаткам гибкого контроля. Оптимальная
стратегия решения задач включает в себя и эксплицитные, и имплицитный под-
ходы. Процессы на грани сознания обычно вовлекаются в задачах на «чувство
знания» и «на кончике языка». Эти задания могут выявить метакогнитивные
знания непрерывного процесса имплицитного решения задач. В настоящий мо-
мент считается, что данные процесс активируют исполнительные области пре-
фронтальной коры.
7.0 . Задания и упражнения к главе 10
1. Заполните ячейки на рис. 10.33. Обратите внимание, что ячейки парные.
Одна ячейка соответствует определенной области мозга, тогда как другая —
ее основной функции, которая была обсуждена в этой главе.
2. Аналогичное задание для рис. 10.34. Что связывает эти два рисунка? На-
зовите области мозга и укажите, в обеспечение каких процессов памяти они
вовлечены.
3. Приведите пример группировки информации в задаче на профессионализм.
Почему это необходимо для нормального функционирования мозга чело-
века?
Рис. 10 .33
Мышление и способность решать задачи
49
Рис. 10 .34
Активность мозга в ответ на одно произнесен-
ное слово (начиная с верхнего изображения).
Обратите внимание, насколько быстро актив-
ность распространяется на области, вне слу-
ховой коры. К третьему сверху изображению
активность распространилась как на левую
нижнюю лобную извилину (Л-НЛИ), так и на
область центральной борозды, теменную кору
и даже в затылочную долю. Предъявленные
МЭГ-изображения демонстрируют негатив-
ность рассогласования (НР), большую волну
электромагнитной активности, возникающей
в ответ на неожиданный стимул. (Источник:
Pulvermüller et al., 2006.)
Г-н Брока в своем докладе представил строение мозга пятидесятиоднолет-
него мужчины с отсутствием речи, который, находясь под его наблюдением,
умер в больнице Бисетр... Когда пациент в возрасте 21 года поступил в Бисетр,
у него уже некоторое время отсутствовала речевая функция; он был неспосо-
бен произнести больше одного слога, который обычно повторял несколько раз
подряд; независимо от того, какой вопрос ему был задан, он всегда отвечал
тан, тан в сочетании с довольно разнообразными выразительными жестами.
По этой причине, по всей больнице он был известен только по имени Тан.
Пьер Поль Брока (1861) «Потеря речи, хроническое ухудшение
и частичное разрушение левой передней доли мозга»,
впервые опубликовано в Bulletin de la Societe
Anthropologique, 2, 235–238.
Перевод с франц. Christopher D. Green.
http://psychclassics.yorku.ca/Broca/perte-e.htm
Содержание
1.0.Введение........................................................51
2.0.Природаязыка...................................................53
2.1.Биологическиеаспекты........................................56
2.2.Истокиязыка.................................................62
2.3.Речьпротивязыка.............................................67
3.0.Звукиустнойречи................................................67
4.0.Планированиеиформированиеречи................................71
5.0.Эволюционные аспекты производства и восприятияречи. . . . . . . . . . . . . . . . 74
6.0.Словаиихзначения..............................................76
6.1.Культурнаясокровищницасловиидей...........................78
6.2.Узнаваниесинонимов..........................................79
6.3. Современные данные о словах и их значении фрагментарны . . . . . . . . . 80
7.0.Синтаксис,включениеисинтагматизация.............................81
8.0.Просодияимелодикаречи.........................................82
9.0.Значимыеутверждения............................................84
10.0.Объединенныеобразыречи........................................84
11.0.Заключение.....................................................88
12.0.Заданияиупражнениякглаве11....................................88
1.0 . Введение
Язык является, прежде всего, инструментом человеческой мысли и культуры.
Это одна из основных вех развития детей, не имеющая близких аналогов у дру-
гих видов. К своему четвертому дню рождения мы уже справляемся с пробле-
мой понимания нашей первой фонологии, нашего первого основного лексикона
и синтаксиса. В течение этих лет мы узнаем новые слова весьма быстрыми тем-
пами. Хотя наше понимание синтаксиса еще не полностью сформировано, дети
обучаются ему без видимых трудностей. Кроме того, маленькие дети знают,
как использовать свои новые речевые навыки для достижения важных целей
(рис. 11 .1). Даже прежде, чем они освоят речь, у них уже имеется отличное
понимание устройства окружающего их мира. Это является достижением в раз-
витии мозга, которое обеспечивается хорошими воспитателями, при помощи
познавательной и эмоциональной стимуляции, опыта, и, конечно, родного язы-
ка, как подарка культуры.
Для иллюстрации научных вопросов, связанных с речью, можно попросить
ко го-нибудь повторить только что услышанное предложение. Вы, возможно,
Глава 11
Речь
52
Глава 11
удивитесь: люди редко могут дословно воспроизвести предложение уже через
пару секунд после прослушивания. Это поисходит не потому, что у людей пло-
хая память, а из-за того, что мы стремимся сохранить смысл, а не услышанные
нами слова (Sachs, 1967). Поэтому большинство людей выдают пересказ того,
что они слышали — другое предложение с аналогичным значением — а не ис-
ходное предложение. Как только речь понята, мы склонны забывать ее «внеш-
нюю форму». Она выполнила свою цель.
Рис. 11.1 . Уровни речи — анализ и генерация. Схема уровней анализа и генерации
языка. Каждый уровень очень сложен, но обрабатывается опытным говорящим субъек-
том бессознательно и в течение нескольких секунд. (Источник: Baars, адаптировано из
Miller, 1991.)
Речь
53
Таким образом, в течение нескольких секунд звуковой сигнал, вероятно,
проходит следующие этапы анализа.
• Акустический анализ — перевод звуков в лингвистические элементы та-
кие, как фонемы; кодирование и преобразование фонем для построения
слогов и морфем (смысловых единиц).
• Лексическая идентификация — присвоение слов входному сигналу, вы-
бранных из словаря с десятком тысяч слов.
• Синтаксический анализ — определение существительных, глаголов и дру-
гих грамматических категории и строительство синтаксических основ.
• Семантика — создание семантической сети из лексической и синтакси-
ческой структур.
• Соотнесение с дискурсом и разговором — как выявленный смысл связан
с предыдущими концепциями в разговоре или споре?
• Прагматичные и социальные выводы — какова цель говорившего, и как
это влияет на мою цель?
2.0 .
Природа языка
По мнению Баррет и соавторов (Barrett et al., 2003), низшие обезьяны думают:
«А что теперь?», тогда как антропоидные обезьяны задаются вопросом: «А что
если?». Таким образом, приматы с большей площадью фронтальной коры мо-
гут думать о воображаемых событиях не только в понятиях «здесь и сейчас»,
но также и в терминах «там и тогда». Вроде бы это не большая разница, но язык
и множество культурных достижений, основанных на речи, превратились в по-
леты на луну и зубные щетки, и, вероятно, в некоторые события, которые не
столь приятны. Эволюционная история человеческого вида, во многих отноше-
ниях, — это история развертывания языка в погоне за личными и культурными
целями (блок 11.1).
Блок 11.1 . Неоднозначность существует на всех речевых уровнях
Если посмотреть на рис. 11.1, то создается впечатление, что существует точное со-
отношения между звуковым входом и фонетическим анализом, или между словами
и их значениями. Такое впечатление складывается обычно у людей, свободно вла-
деющих речью, и это совершенно неверно. Язык изобилует неясностями на каждом
уровне анализа, как на входе, так и на выходе, т. е . существуют точки выбора при об-
работке речи (см. гл . 10). Например, значительное число часто встречающихся слов
имеет несколько значений, так что соответствия между лексикой и концептуальными
представлениями, представленные в верхней части рис. 11 .1, всегда имеют альтерна-
тиву. Так же часто существуют неясности в акустическом анализе, они присутствуют
и в известных синтаксических неоднозначностях, как, например, показано на древо-
видной диаграмме двойственной структуры на рис. 11 .2 (Chomsky, 1957).
Недавняя оценка лексической неоднозначности в английском языке выявила в сред-
нем по два наиболее часто используемых толкования для каждого слова (Miller,
1991). Более привычные слова, как правило, имеют большее число значений, так что
слово «set» в английском языке имеет более двух десятков значений согласно Ок-
сфордскому словарю английского языка (Simpson and Weiner, 1989).
Точки выбора существуют и при производстве речи и языка. Мы начинаем это чув-
ствовать, когда пишем сочинение, когда сталкиваемся с выбором между двумя сло-
вами, имеющими немного различающийся смысл. Синонимы и парафразы создают
54
Глава 11
точки выбора при производстве речи, а неясности на различных уровнях анализа
создают точки выбора при восприятии речи.
Точки выбора в потоке обработки требуют определенного разрешения, затрагиваю-
щего часто различные уровни анализа. Они превращают простые переходы из одного
языкового уровня на другой в лабиринты, которые, как правило, приводят к «комби-
наторному взрыву». Таким образом, для n двойных точек выбора число возможных
путей через лабиринт экспоненциально возрастает как 2
n
. Это число растет настоль-
ко быстро, что толкование автоматизированного языка и речи считается достаточно
трудным процессом.
Главное решение такого затруднения называется «нисходящим» или «прогностиче-
ским» процессом обработки: когда мы сталкиваемся с неоднозначным словом, таким
как «set» в предложении «Please set the table» (Пожалуйста, накройте на стол), мы мо-
жем сказать, основываясь на смысловом контексте, что в данном действии участвуют
Рис. 11 .2 . Синтаксическая неоднозначность. Существует два способа понять предло-
жение «They are flying planes», или «Пилоты водят самолеты», или «Самолеты лета-
ют». Назначение субъекта (they, англ. они) неоднозначно, и в результате неоднознач-
ной становится вся структура предложения. Местоимения как «they» в английском
языке являются богатым источником неопределенности, так как принимают условия
как должное. Носители языка, как правило, не замечают такие неоднозначности, но
новички в данной языковой группе имеют склонность недопонимать их. Синтакси-
ческая неоднозначность является примером принятия решения в точках выбора при
анализе языка
Речь
55
тарелки. Фраза «Please, set an example» («Пожалуйста, приведите пример») требует
чего-то совсем иного. В этих случаях для разрешения неоднозначности необходи-
мы семантические знания. Таким образом, более высокие уровни анализа необходи-
мы для определения направления в точках выбора на нижних уровнях. «Иерархия»
языка, так же как и иерархия процессов обработки зрительной информации (гл. 3),
требует потока информации во всех направлениях. Предполагаемая схема потока ин-
формации представлена на рис. 11 .3 .
Рис. 11.3 . Информация не переходит строго с одного уровня на другой в системе
иерархии языковой обработки. Так же как иерархия процессов обработки зрительной
информации, рассмотренная в гл. 3, физические сигналы речи в мозге присутству-
ют в виде множества различных точек выбора, в системе как ввода, так и вывода
информации. При обработке входящих речевых сигналов существуют значительное
число неоднозначностей в звуках, значениях слов и синтаксисе, которые решаются
при помощи информации об ожидаемом значении из других иерархических уровней
анализа. Не существует четкого потока речевой информации снизу вверх, так же как
нет простого соответствия зрительных карт. То же самое относится и к выведению
речевой информации. (Источник: Baars.)
56
Глава 11
2.1.
Биологические аспекты
Практически все люди обучаются говорить в течение первых нескольких лет
жизни, тогда как ничего подобного не отмечено у других видов. Существуют
интересные исключения, например шимпанзе и гориллы могут освоить язык
глухонемых при помощи подражательного обучения и тренировки в раннем
возрасте. Но, как у вида, у других ныне живущих приматов в отличие от людей
отсутствует способность осваивать речь. Это отсутствие не значит, что мож-
но свести к минимуму значение специальных возможностей Бонобо и других
приматов, или других видов, таких как африканские серые попугаи Алекс и
Артур (Pepperberg, 2002); просто оно указывает на то, что речь является чисто
человеческой особенностью, для которой мы биологически и культурно подго-
товлены. Именно поэтому мы великолепно справляемся с этой особенностью.
Дети обучаются языку на определенных этапах развития и даже могут обра-
зовывать свои собственные «креолы», управляемые определенными правилами
языки, когда они растут в смешанной языковой среде без доминирующего язы-
ка (см. блок 11.2). При взрослении становится все труднее приобрести навыки
носителя языка. Устная речь является сложной биологической надстройкой на
ранее существовавшую вокальную и слуховую физиологию. Образование речи,
как правило, происходит в доминирующем полушарии (обычно с левой сто-
роны), хотя раннее повреждение головного мозга может привести к переходу
этой функции на недоминирующую сторону (гл. 1). Ген FOXP2, по-видимому,
должен точно экспрессироваться для нормального развития человеческой
речи, хотя этот ген широко встречается среди всех видов позвоночных (Vargha-
Khadem et al., 2005).
Повторение предложения задействует конкретные области мозга при ус-
ловии его нормального развития. Наиболее известными примерами являются
зоны Брока и Вернике (гл. 1), но речь и язык вовлекают в процесс гораздо более
широкий набор корковых и подкорковых областей. «Речевая кора» становится
специализированной для данной функции только в рамках эволюции гоминид,
т. е. в течение менее чем 3 млн лет из 200 млн лет эволюции млекопитающих.
До гоминид зона Брока могла быть вовлечена вместе с голосовым трактом в
контроль других функций: она, в конце концов, непосредственно примыкает
к области гомункулуса, отвечающей за двигательный контроль рта и языка.
В общем, как писал Мезалун (Mesalum, 1990):
Когнитивные проблемы будут решены не путем последовательного и иерархиче-
ского продвижения к предопределенным целями, а вместо этого путем совмест-
ного и взаимодействующего рассмотрения многочисленных возможностей и труд-
ностей до достижения удовлетворительного результата. Конечная организация
психической деятельности характеризуется почти бесконечным богатством и гиб-
костью. Согласно этой модели, сложное поведение переходит на уровень мульти-
фокусной нервной системы, а не конкретных анатомических структур, порождая
такие взаимодействия между мозгом и поведением, которые одновременно и лока-
лизованы и распределены.
Нисходящие или прогнозируемые процессы обработки представляются универсаль-
ным свойством когнитивного мозга.
Речь
57
Многие составные части разговорного языка такие, как контроль голоса и слух,
должны были возникнуть очень рано в эволюции. Но есть предположение, что
полный арсенал языковых навыков, возможно, стал доступен гораздо позднее в
эволюции, а именно 30–100 тыс. лет назад, а не 3 млн лет назад как считалось
ранее. Конечно, рабочая лексика живого языка, по-видимому, значительно рас-
ширилась за последнее время, в то же время синтаксис, возможно, стал проще.
Все это позволяет предположить, что речь и язык обусловлены биологиче-
ской готовностью, во многих отношения схожей с развитием ловкости рук, зре-
ния, умственным развитием и социальными отношениями. Как и другие гене-
тически обусловленные свойства, речь зависит от большого числа переменных.
Какой язык мы приобретем в детстве, насколько хорошо мы изучим его, какой
словарный запас (и, следовательно, концептуальную систему) приобретем, и
многое другое зависит от нашего опыта. Тем не менее биологический субстрат
языка является важным источником понимания, как мы увидим далее. Он не
сводит людей до уровня некоторых более простых видов, а вместо этого под-
черкивает огромную сложность и адаптивное значение языка. Наши культур-
ные и личные достижения в значительной степени зависят от биологической
предрасположенности нашего вида.
В гл. 1 была затронута тема открытия в XIX в. Пьером-Полем Брока и
Карлом Вернике зон мозга, участвующих в производстве и восприятии речи,
соответственно. Их результаты, полученные на небольшом числе пациентов
с повреждениями головного мозга, вызвали более чем вековую дискуссию по
вопросу локализации речевой функции в мозге. Эта дискуссия продолжается и
сегодня, зачастую с привлечением, например, данных нейросетевого моделиро-
вания речевой функции и сложных методик регистрации мозговой активности
(гл.3и4).
На рис. 11.4 дано классическое представление о локализации зоны Брока,
отвечающей за производство речи, и зоны Вернике, участвующей в восприятии
и понимании речи. Кроме того, в XIX веке врачами был выявлен целый ряд раз-
личных афазий (нарушений речи). Наиболее известной из них является прово-
дниковая афазия, обусловленная повреждением дугообразного пучка, «малень-
Рис. 11 .4. Классические речевые зоны мозга. Традиционное расположение зон Брока и
Вернике в левом полушарии на основе данных, полученных на неврологических паци-
ентах с нарушениями понимания и воспроизведения речи. Правое полушарие обладает
функциями, воспринимающими речь, которые не отображены на рисунке. ВВИ— верх-
няя височная извилина; ЦБ — центральная борозда. (Источник: Standring, 2005.)
58
Глава 11
кого аркообразного пучка», соединяющего зоны Брока и Вернике (Catani and
Ffytche, 2005). Одним из наиболее важных современных достижений является
возможность изучения больших трактов белого вещества, которые соединяют
области коры между собой и заполняют значительный объем полушарий голов-
ного мозга. В ближайшие несколько лет трактография (гл. 4) и другие методы
изучения связей в живом мозге позволят узнать много нового как о корковой
специализации, так и о взаимодействиях между корковыми зонами.
Зона Вернике граничит со слуховой корой в области латеральной борозды
и верхней височной извилины (ВВИ на рис 11.4). Зона Брока непосредственно
прилегает к областям моторной коры, отвечающим за ротовую полость и фор-
мирование звуков. Наше современное понимание речевой локализации гораздо
шире, однако полезно запомнить, что расположение этих областей с определен-
ным соседством имеет функциональный смысл.
В 1861 г., когда Брока обнаружил у пациентов повреждения левой нижней
лобной извилины (Л-НЛИ), которая теперь называется зоной Брока, вспыхнула
большая дискуссии среди неврологов, которые испытали трудности в поиске
пациентов с точно таким же повреждением и такими же симптомами. Эта дис-
куссия продолжалась и в следующем столетии. Мы могли бы ожидать, что этот
спор разрешится благодаря использованию сложных методов визуализации
мозга или при исследовании сотен пациентов при проведении прямой стиму-
ляции мозга во время хирургических операций на открытом головном мозге
(Penfield and Roberts, 1959; Ojemann, 2003). Удивительно, но этого не произо-
шло. Дискуссии о локализации функций продолжается и сегодня.
Однако нельзя сказать, что никаких новых данных не было получено за бо-
лее чем вековую историю исследования. Большинство ученых считают, что
зона Брока является необходимой для нормального производства речи. Но го-
раздо больше процессов происходит в этой части головного мозга, чем предпо-
лагалось ранее, к тому же известны случаи отсутствия речевых нарушений при
повреждении зоны Брока. С постоянным улучшением методов нейровизуали-
зации появляется возможность изучать все меньшие и меньшие области коры
головного мозга, что позволяет исследователям выделять новые подразделения
левой нижней лобной извилины (Л-НЛИ).
Мозг не хранит воспоминания в отдельных, специально выделенных обла-
стях. Однако существует общее представление, что долговременная память мо-
жет участвовать в активации областей, связанных сенсорными, моторными, мо-
тивационными и языковыми функциями. В коре существуют многочисленные
локальные соединения в рамках одной вертикальной колонки и между группа-
ми колонок; также есть более отдаленные связи между соседними регионами
бóльших размеров; и существует огромная протяженная система магистралей
от одного конца головного мозг до другого, соединяющая задние и передние об-
ласти, правое и левое полушария, и имеющие восходящие и нисходящие ветви
в сенсорные и моторные представительства черепно-мозговых нервов и спин-
ного мозга (см. гл . 5). Такая высоко взаимосвязанная система больше похожа на
Всемирную паутину, чем на организацию студенческого городка.
Прежде чем мы перейдем к выводам о том, что язык и мышление локализо-
ваны в легко обнаруживаемых областях мозга, было бы полезно ознакомиться
с обширным обзором литературы (Vigneau et al., 2006). На рис. 11 .5 продемон-
Речь
59
стрированы результаты крупного мета-анализа более чем 125 экспериментов
сканирования мозга, в результате которого было выявлено более 700 областей
повышенной активности в левом полушарии. Различными цветами обозначены
области, отвечающие за фонологию, семантику (деятельность, связанная с кон-
цепцией), а также обработку предложений или текстов. Степень перекрытия
этих областей очень высока. Это не такая модель, с которой мы сталкиваемся
при изучении сенсорных областей, например зрительных, где четко локализо-
ванные зрительные карты, такие как поле V1, могут быть соотнесены топогра-
V1, могут быть соотнесены топогра-
1, могут быть соотнесены топогра-
фически с картой сетчатки.
Эти факты имеют сложную теоретическую интерпретацию. Они свидетель-
ствуют в пользу того, что в большинстве случаев работа мозга осуществляет-
ся благодаря распределенной сети речевой функций, аналогичной Всемирной
паутине. Интернет-чаты работают и в том случае, когда участники переписки
находятся в совершенно разных частях мира. Идея о распределении мозговых
функций, по-видимому, поддерживает взгляд на работу мозга с точки зрения его
взаимосвязей, как описано в гл. 3 (см. также «Дарвинистский подход», гл. 3).
Выполнив статистический кластерный анализ сотен наборов данных, иссле-
дователи смогли выделить более конкретный локус, вовлеченный в функцио-
нирование классической петли рабочей памяти (рис. 11 .6). Анализ позволяет
выделить три самостоятельных петли рабочей памяти для процессов обработки
фонологической, семантической и контекстной информации. Но все равно су-
ществует загадка, которая занимает ученых, по крайней мере, начиная с вре-
мен Пьера-Поля Брока: какие данные указывают нам на реально происходящие
события? Вполне возможно, что статистический анализ выявит «истинную»
природу областей мозга, показанных на рис. 11 .6 . Данное предположение было
Рис. 11 .5 . Широкое распределение речевых зон в головном мозге может согласоваться
с региональной специализацией. Обобщенные данные более чем 100 экспериментов по
нейровизуализации мозга при помощи фМРТ и ПЭТ указывают на активацию около 730
зон (Vigneau et al., 2006). Точки синего цвета указывают области, активирующиеся при
обработке фонологии, — речевых звуков. Регионы, выделенные красным, наибольшую
активность проявляют при семантических процессах, при выявлении значения слов и
фраз, а области, отмеченные зелеными точками, активируются при обработке информа-
ции целых предложений и текстов. Перекрытие областей и широкое распределение трех
функциональных областей поражает. (Источник: Vigneau et al., 2006.)
60
Глава 11
бы логичным, если бы существовал случайный разброс данных, обусловлен-
ный методологическими и биологическими причинами. Но каждая отмечен-
ная функциональная точка на рис. 11 .5 является результатом работы многих
коллективов и указывает на то, что не существует такого случайного разброса.
Таким образом, перед нами стоит выбор между принятием на основе данных
кластерного анализа точки зрения о том, что функции в мозге строго локализо-
ваны, или, опираясь на рис. 11 .6, склониться в сторону распределенности моз-
говых функций. Совершенно не очевидно, какая из точек зрения верна.
Тем не менее по некоторым основным моментам существует определенное
соответствие. Например, обе иллюстрации демонстрируют постоянное взаимо-
действие между сенсорной (задней) и моторной (передней) частями коры го-
ловного мозга. Несмотря на то, что левое полушарие изучают в связи с левосто-
ронней ассиметрией у большинства людей, важно понимать, что локализация
речевых функции не ограничивается левым полушарием. Существуют убеди-
тельные свидетельства двустороннего функционирования при обработке вход-
ных сигналов речи и языка — восприятия речи и понимания. Однако по причи-
нам, которые не очень хорошо понятны, воспроизведение речи, планирование и
контроль, локализуются в левом полушарии у более чем у 90% населения. Как
мы увидим далее, правое полушарие головного мозга может даже иметь свой
собственный способ понимания сложных средств выражения мысли, таких как
анекдоты, метафоры и ирония, а левая сторона воспринимает сказанное бук-
вально (Zaidel et al., 2000).
Рис. 11.6 . Петля рабочей памяти для фонологии, семантики и предложений. Широкий
разброс пиков активности на рис. 11 .5 может быть упорядочен при помощи кластер-
ного анализа, в результате которого в левом полушарии можно выявить центры, ответ-
ственные за речевые процессы. Однако этот анализ должен затем объяснить первичную
обширную активацию областей при обработке фонологии, семантики и предложений.
Ответы в этом случае не так очевидны. ВВБз — верхняя височная борозда, задняя;
УИ — угловая извилина; ВКИ — верхняя краевая извилина; Л2з — средняя лобная из-
вилина, задняя; НЛ3п — покрышка нижней лобной извилины; Л3н — нижняя лобная
извилина. Термин «покрышка» употребляется для обозначения участка коры, который
прикрывает скрытые области островковой зоны и сильвиевой борозды. Лобные изви-
лины пронумерованы сверху вниз (Л1 — верхняя, Л2 — средняя, Л3 — нижняя. (Ис-
точник: Vigneau et al., 2006.)
Речь
61
Мы вернемся к этим идеям еще раз, когда будем изучать, как мозг обеспе-
чивает человеческую речь и мышление. Разные исследователи предлагают не-
сколько различающиеся трактовки большого объема имеющейся по данной те-
матике литературы. Мы тщательно объединили данные работ по исследованию
поврежденного мозга, регистрации единичных нейронов, позитронно-эмисси-
онной томографии (ПЭТ), функциональной магнитно-резонансной томографии
(фМРТ), магнитоэнцефалографии (МЭГ), электроэнцефалографии (ЭЭГ), вы-
званных потенциалов и т. д. Не все полученные результаты можно легко соот-
нести между собой, однако такая организация информации очень полезна.
Например, предложено рассматривать классическую область Брока более ши-
роко (Hagoort, 2005). Автор выдвигает идею разделения функций, которая проде-
монстрирована на рис. 11 .7 . Границу зоны Брока в настоящее время отодвигают
вперед, включая в нее серую овальную область, которая предположительно вы-
полняет функцию объединения речевых звуков, значений и синтаксических взаи-
мосвязей. Эта зона включает также два участка, расположенных в височной доли
и ответственных за память, в том числе и МВД (медиальную височную долю,
скрытую при рассмотрении с левой стороны). Наконец, контролирующие функ-
ции принадлежат дорсолатеральной префронтальной коре (ДЛ-ПФК).
Как мы знаем из трагических случаев проведения хирургических операций
на левом полушарии у маленьких детей, левосторонняя ассиметрия языковой
функции может сместиться в другое полушарие, если возникают значительные
повреждения этой области в раннем детстве (см. гл . 2). Латерализация функций
также наблюдается у некоторых других видов, но причины ее все еще недоста-
точно изучены. Необходимо уяснить, работая с этой темой, что широкое рас-
пространение активности речевой функции не ограничивается только левым
полушарием. Оно расширяется как некий огромный мегаполис и в полушарие,
не имеющее центров речи. Анатомически большим мостом между полушария-
ми является мозолистое тело, которое расходится в обе стороны и двусторонне
Рис. 11 .7 . Л -НЛИ: расширенное представление зоны Брока. Левая нижняя лобная из-
вилина (Л-НЛИ) является более точным обозначением обширной области, вовлеченной
в планирование и производство речи. Выдвинуто также предположение, что Л-НЛИ
служит «зоной конвергенции» для речи, т. е . это область, в которой различные речевые
параметры объединяются вместе в единую схему перед тем, как быть отправленными в
моторные зоны коры. (Источник: Hagoot, 2005.)
62
Глава 11
соединяет полушария, так что нейроны во фронтальной коре с одной стороны
посылают аксоны, которые достигают соответствующих точек фронтальной
коры на другой стороне (см. гл . 5). Для осуществления многих функций работа
обоих полушарий головного мозга интегрирована.
В то же время повреждение зоны Брока ведет к нарушению воспроизведе-
ния речи, а разрушение зоны Вернике и соседних областей вызывает ухудше-
ние восприятия и понимания речи (рис. 11 .8). Исторически Брока и Вернике не
ошиблись в своих выводах, они просто не обладали полным набором фактов.
На рис. 11 .11 представлена модель устной речи в коре (Hickok, Poeppel,
2007). Обратите внимание, что верхние районы височной доли, ближе к задней
части сильвиевой борозды, содержат слуховые области, которые представляют
фонемы или, возможно, «слоги», упомянутые выше, являются звуковыми пред-
ставителями речи (рис. 11 .11). Это вероятная модель, но как уже говорилось,
мы еще не имеем необходимой пространственной разрешающей способности
методов, чтобы знать, есть ли более микроскопическое разделение функций у
чувствительных нейронов, аналогичное найденному в зрительной коре.
2.2 .
Истоки языка
Люди тысячелетиями задавались вопросом об истоках речи. Если ребенок рас-
тет в безъязыковой среде, сможет ли он сам создать язык? Между прочим, при-
рода обладает очень похожими на этот примерами. В некоторых изолированных
сообществах, например на маленьких островах, взрослые люди, владеющие
разными языками, могут поселяться в одном месте. Взрослые, говорящие сво-
бодно на новом языке, очень редко воспринимают «акцент» (фонологию) или
грамматику языка, если его выучили после пубертатного периода. Вместо это-
го они имеют склонность говорить на «ломаной» версии новоприобретенно-
го языка, достаточной для общения с взрослыми людьми по всему свету, но,
как правило, без живой беглости и богатой выразительности их родного языка.
Рис. 11.8 . Модель Вернике–Гешвинда. Наиболее известная модель зон мозга, основан-
ная на неврологических данных, была предложена Гешвиндом (1979). В то время как
ученые продолжают интенсивно изучать модель Вернике–Гешвинда, возникновение
методов нейровизуализации привело к появлению новых данных. (Источник: Weems
and Reggia, 2006.)
Речь
63
Дети в таких двойных сообществах поступают очень интересно: они создают
свои собственные языки, которые называют «креолами», так как они впервые
были исследованы в креольских общинах на Карибский островах (блок 11.2;
Bickerton, 1984, 1990).
Блок 11.2 . Креольские языки создаются детьми,
выращенными в многоязыковой среде
Креолы — это языки с простой синтаксической структурой, которые возникают тог-
да, когда нескольким людям с разной языковой принадлежностью приходиться об-
щаться друг с другом. Креолы возникают при иммиграциях и оккупации или в тех
случаях, когда люди с разной культурой вынуждены работать вместе. Взрослые люди
в таких случаях вырабатывают примитивный гибридный язык (пиджин, гибридный
язык с искажением морфологического и фонетического облика слов), скудную си-
стему коммуникации с ограниченным набором существительных, глаголов и про-
изводных, сопровождаемую интенсивной жестикуляцией. Дети людей с гибридным
языком усваивают родной язык родителей, но не их пиджин; вместо этого они в до-
полнение к родному языку создают для общения с себе подобными креолы основан-
ные на гибридном языке. Все креольские языки обладают набором свойств, которые
могут быть чрезвычайно схожими с признаками полностью сформированных язы-
ков. Таким образом, дети в смешанных сообществах формируют более полноценный
язык, чем их родители.
1. Грамматика основывается на порядке слов; в противоположность этому, большин-
ство человеческих языков чаще опираются на интонацию (утверждение и отрица-
ние — см. обсуждение выше).
2. Существуют семь частей речи: существительное, местоимение, прилагательное,
глагол, наречие, артикли и предлоги; многие человеческие языки упускают одну
или несколько перечисленных категорий или добавляют новые.
3. Существительные могут быть в единственном, множественном или неопределен-
ном числе, во многих языках отсутствует последний вариант.
4. Три частицы используются в качестве вспомогательных глаголов для обозначения
результативных, нерезультативных и повторяющихся действий; данное разделе-
ние отсутствует во многих языках.
5. Существует система спряжения единичных глаголов; в большинстве языков име-
ется основное правило для спряжения подавляющей части глаголов, а также осо-
бые исключения для часто употребляемых (неправильных) глаголов.
6. Вопросительные предложения строятся на основе интонации, а не порядке слов;
во многих языках используются оба варианта.
Бикертон (Bickerton) указывает на то, что во многих случаях перечисленные свой-
ства креолов отсутствуют в речи родителей детей, создавших креолы. Таким обра-
зом, вполне вероятно, что креольские языки могут быть связанными с врожденными
специфическими особенностями языка.
Однако даже креолы не могут напрямую проиллюстрировать эти свойства. Если бы
все только что перечисленные свойства креольских языков были врожденными, было
бы невозможно объяснить, каким образом в языках возникают другие особенности.
Исходные свойства языка должны быть более абстрактными, нежели обнаруженные
в креолах, и должны позволять развитие всех существующих на сегодняшний день
видов языков. Вместо того чтобы рассматривать креолы как непосредственную ил-
люстрацию универсальных особенностей языка, лучше воспринимать их как воз-
можность узнать, какие из особенностей языка легче развиваются. Возможно, имен-
но этими чертами обладали языки человека на ранних этапах его развития.
(Источник: Squire et al., 2003.)
64
Глава 11
Как произнесенные предложения обрабатываются в мозге
Мы общаемся преимущественно при помощи уст-
ной речи. Произносимые предложения содержат не
только семантическую и синтаксическую информа-
цию, выражающую значения слов и их грамматиче-
ские связи (кто кому что делает), но также и просо-
дическую информацию — мелодию и ритм языка.
Просодия обеспечивает нас информацией о грани-
цах синтаксических фраз, а также сообщает об эмо-
циональном тоне, модулирующем значение. Иссле-
дования с нейровизуализацией продемонстрировали
обработку семантической и синтаксической инфор-
мации нейронными сетями левого полушария (рис.
11.10), включающими нижнюю лобную область
(зону Брока и области кпереди от нее) и верхнюю
и среднюю височные области (преимущественно
зону Вернике) (Friederici, 2002; Hickok and Poeppel,
2007). Функциональная сеть для синтаксиса состоит
из зоны Брока (Stormswold et al., 1996) и наиболее
задней части нижней лобной извилины и борозды (Bornkessel et al., 2004); семантика
задействует области нижней лобной извилины кпереди от зоны Брока (Thompson-
Schill et al., 1997) и часть средней и верхней височных извилин (Demonet et al., 1994).
При обработке синтаксически сложных предложений мы видим активацию сетей,
связывающих зону Брока и заднюю часть верхней височных извилин (Friederici
et al., 2006a). Недавние работы показали, что в пределах лобно-височной сети зона
Брока выполняет обработку синтаксических иерархий независимо от значения по-
следовательности слов (Friederici et al., 2006b; Makuuchi et al., 2009). Задние части
височной доли участвуют при обработке синтаксически сложных значимых предло-
жений (Bornkessel et al., 2004; Friederici et al., 2009). Последние данные указывают
на то, что задняя часть височной коры помогает объединять значение и синтаксис
для понимания смысла предложения.
Просодическая информация — мелодия языка — обрабатывается в правом полу-
шарии, в особенности в верхних височных и нижних лобных областях (Meyer et al.,
2002). Связанные с событиями потенциалы (ССП) указывают, что интонационная
кайма фразы может помочь или наоборот запутать при синтаксическом разборе
строения предложения при его прослушивании (Steinhauer et al., 1999). Например,
два предложения: (а) Так как Джон постоянно бегает две мили, для него это кажется
короткой дистанцией и (б) Так как Джон постоянно бегает, две мили для него кажут-
ся короткой дистанцией — синтаксически явно различаются. Если прочитать эти
предложения внимательно, то понятно, что они оба являются осмысленными нор-
мальными предложениями — просто у них различная интонационная кайма. В пред-
ложении (а) мы заканчиваем часть фразы на слове «мили»; в предложении (б) изме-
няем интонацию после слова «бегает». Интонационный акцент после слова «бегает»
в предложении (а) привел бы к неправильному синтаксическому разбору фразы.
Это предполагает наличие сильных взаимодействий между левополушарной об-
работкой синтаксиса и правополушарной обработкой просодической информации.
Такое межполушарное взаимодействие обусловливается тяжами белых волокон,
соединяющими два полушария, т. е . мозолистым телом. Данные по регистрации
ССП у пациентов с повреждениями задней трети мозолистого тела указывают на
необходимость этой структуры для взаимосвязи синтаксической и просодической
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
Рис. 11 .9 . Анжела Д. Фри-
деричи (Angela D. Friederici,
PhD), Институт человече-
ской когнитологии и науки о
мозге общества Макса План-
ка, Лейпциг, Германия
Речь
65
информации (Freiderici et al., 2007). Это позволяет двум типам информации влиять
друг на друга на ранних этапах обработки, что помогает ускорить наше понимание
произнесенного предложения (Eckstein et al., 2006).
Литература
1. Bornkessel, I., Fiebach, C.J., Friederici, A.D., and Schlesewsky, M. (2004). «Capacity»
reconsidered: Interindividual differences in language comprehension and individual
alpha frequency. Experimental Psychology, 51, 279–289.
2. Demonet, J.F., Price, C., Wise, R., and Frackowiak, R.S.J . (1994). Differential activation
of right and left posterior sylvian regions by semantic and phonological tasks —
A positron-emission tomography study in normal human-subjects. Neuroscience
Letters, 182, 25–28 .
3. Eckstein, K., and Friederici, A.D. (2006), It’s early: event-related potential evidence
for initial interaction of syntax and prosody in speech comprehension. Journal of
Cognitive Neuroscience, 18, 1696–1711 .
4. Friederici, A.D., (2002). Towards a neural basis of auditory sentence processing.
Trends in Cognitive Sciences, 6, 78–84 .
5. Friederici, A.D ., Fiebach, C.J., Schlesewsky, M., Bornkessel, I., and von Cramon, D.Y.
(2006a). Processing linguistic complexity and grammaticality in the left frontal cortex.
Cerebral Cortex, 16, 1709–1717 .
6. Friederici, A.D., Bahlmann, J., Heim, S., Schubotz, R.I., and Anwander, A. (2006b).
The brain differentiates human and non-human grammars: Functional localization and
structural connectivity. Proceedings of the National Academy of Sciences of the USA,
103, 2458–2463 .
7. Friederici, A.D., von Cramon, D.Y., and Kotz, S.A . (2007). Role of the corpus callosum
in speech comprehension. NeuroReport, 20, 563–568.
8. Hickok, G., and Poeppel, D. (2007). Opinion — The cortical organization of speech
processing. Nature Reviews Neuroscience, 8, 39–402 .
Рис. 11 .10 . Схематическое изображение речевых областей в левом полушарии
с указанием основных значимых извилин и борозд. Цифрами отмечены имеющие
отношение к речевой функции поля Бродмана (ПБ), выделенные на основании их
микроскопических клеточных характеристик (Brodmann, 1909). Классическими
речевыми областями являются зона Брока (ПБ 44 и 45) и зона Вернике (ПБ 22).
Термины «верхняя» или «нижняя» обозначают расположение извилины в опреде-
ленной доле мозга (например, верхняя височная извилина) или в пределах поля
Бродмана (например, верхняя часть ПБ 44). Термины «передняя» или «задняя» ука-
зывают на положение в пределах извилины (например, задняя часть верхней височ-
ной извилины). (Источник: Friederici.)
66
Глава 11
9. Makuuchi, M., Bahlmann, J., Anwander, A., and Friederici, A.D . (2009). Segregating
the core computational faculty of human language from working memory. Proceedings
of the National Academy of Sciences of the USA, 106, 8362–8367 .
10. Meyer, M., Alter, K., Friederici, A.D ., Lohmann, G., and von Cramon, D.Y. (2002).
FMRI reveals brain regions mediating slow prosodic modulations in spoken sentences.
Human Brain Mapping, 17, 73–88 .
11. Steinhauer, K., Alter, K., and Friederici, A.D . (1999). Brain potentials indicate
immediate use of prosodic cues in natural speech processing. Nature Neuroscience, 2,
191–196.
12. Stormswold, K., Caplan, D., Alpert, N., and Rauch, S. (1996). Localization of syntactic
comprehension by positron emission tomography. Brain and Language, 52, 452–473 .
13. Thompson-Schill, S.L., D’Esposito, M., Aguirre, G.K., and Farah, M.J . (1997). Role
of the left inferior prefrontal cortex in retrieval of semantic knowledge: A reevaluation.
Proceedings of the National Academy of Sciences of the USA, 94, 14792–14797.
Рис. 11 .11 . Модель устной речи Хикока–Пёпла. На верхней половине рисунка изо-
бражена модель обработки устной речи (Hickok, Poeppel, 2004). На нижней половине
рисунка показаны области мозга, которые, вероятно, отражают стадии этой модели (со-
кращения см. на рис. 11.10). Обратите внимание, что системы первичного восприятия
речи для перевода акустической и фонетической информации звуков в значение лока-
лизуются билатерально в левом и правом полушариях головного мозга, тогда как бо-
лее поздние процессы опосредуются зонами левого полушария. (Источник: Hickok and
Poeppel, 2007, с разрешения.)
Речь
67
2.3 .
Речь против языка
Филологи очень часто отмечают различия между речью и языком частично из-
за поражающей пластичности человеческого мозга при освоении разных вход-
ных и выходных модальностей. В данный момент вы используете свои глаза
и, возможно, руки для того, чтобы делать заметки. Вы обучились читать, писать
или печатать с поражающей легкостью. Вы также можете освоить язык жестов,
азбуку Брайля и паралингвистические системы символов, такие как математи-
ка, логика и компьютерное программирование. Таким образом, чисто устная
и слуховая природа речи является не столь необходимой.
Однако точное и быстрое разделение речи и языка не определено. Челове-
ческий мозг подготовлен для восприятия устной словесной речи. Подавляющее
большинство детей обучаются говорить в первые годы жизни, но письменность
и чтение осваиваются ими позже и имеют меньший процент успеха. Тот факт,
что мы можем тренировать удивительную речевую пластичность, обусловлен-
ную потребностями и возможностями, не отрицает биологическую значимость
разговорного языка.
3.0 .
Звуки устной речи
В гл. 7 обсуждалась звуковая природа речи, состоящая из быстро распростра-
няющихся трехмерных воздушных пузырей высокого и низкого давления, кото-
рые вызывают вибрационные колебания чувствительных мембран внутреннего
уха. Мы также можем рассмотреть речь с точки зрения формирования голосо-
вого выхода.
Человеческий голосовой тракт является в своей основе трубкой с двумя
гибкими клапанами, расположенными непосредственно над легкими и диа-
фрагмальными мышцами, которые вместе создают давление воздуха, когда мы
выдыхаем (рис. 11.12). Таким образом, голосовой тракт похож на мембран-
ный гудок, например саксофон, с пластинкой вибрирующей на верхнем конце
трубы. Голосовые связки могут вибрировать быстрее или медленнее, производя
более высокие или низкие звуки. Мы произносим согласные путем перекрыва-
ния голосовой трубки в одном из нескольких возможных мест. В английском
языке для произношения используют сжимание губ (/b/ /p/ /m/), прижимание
языка к средней или задней части неба (/g/ /k/ /ng/ /r/) и стискивание зубов
(/th/ /the/ /s/ /z/ /v/ /w/). Однако полное препятствование прохождению потока
воздуха вызывает отсутствие звука вообще, поэтому именно быстрый переход
от закрытия к открытию воздушного потока вызывает формирование так назы-
ваемых взрывных согласных звуков (/b/ /p/ /t/ /d/ /k/ /g/). В других языках поток
воздуха формируется немного отличающимися способами, однако физические
принципы произношения звуков одинаковые.
Голосовые вибрации как эхо распространяются через голову и тело, вызывая
вибрации во всех полостях головы и туловища. Значит, например, зажимая нос
можно менять качество голоса. Эти вибрирующие воздушные полости также
позволяют нам выделять различия в голосах. Голоса детей звучат выше, чем
взрослых, только потому, что у детей вибрирующие полости меньше.
Таким же образом как перекрывание голосовой щели вызывает образование
согласных, придание ротовой полости разных форм ведет к произнесению раз-
ных гласных звуков. Пение очень близко к речи; оно основано на растягивании
68
Глава 11
длины гласных звуков и настраивании их на определенную высоту при помощи
голосовых связок. Но даже обычная речь имеет определенную мелодичность,
называемую интонационным контуром (мелодическим рисунком). В англий-
ском языке вопросительные интонации заключаются в повышении высоты не-
скольких последних слогов фразы. Также при помощи интонационной мелодии
языка отражается эмоциональная окраска речи.
Даже ритмичность в музыке имеет близкую аналогию с голосовыми стрес-
совыми паттернами, какие, например, используются в поэзии и рэповой му-
зыке. Таким образом, пение и голосовые интонации, ритм и вокальные жесты
можно рассматривать как процессы, использующие одни и те же голосовые воз-
можности в немного измененной форме.
Рис. 11 .12 . Голосовой тракт человека использует уже существовавшие механизмы кон-
троля дыхания, движений ротовой полости, языка, голосовой щели и гортани, унас-
ледованные от предковых видов. Голосовой тракт представлен трубкой с источником
настраиваемых вибраций на голосовых связках, двух полосках ткани в гортани. Каче-
ство голосовых звуков зависит от резонанса вибрации подвижных тканей и воздушных
полостей в голове и туловище. В то время как согласные получаются при ограничении
сжимания голосовой трубки, гласные образуются в большинстве случаев при движени-
ях языка и губ для придания определенной формы ротовой полости. Это имеет такой же
эффект, как изменение частотного резонанса или формантов в голосовом тракте. Таким
образом, гласные звуки и пары согласные-гласные звуки являются минимальными фи-
зическими единицами при производстве речи (если принять шипящие звуки как /h/ /th/
/tha/ /s/ и т. д. как сходные с гласными). (Источник: Standring, 2005.)
Речь
69
В заключение можно сказать, что люди способны изменять форму голосово-
го тракта всевозможными способами, что является редкой (но не уникальной)
адаптацией среди видов животных.
До изобретения печатания устная речь была наиболее быстрым и точным
навыком, который развивали люди. Как было указано в гл. 7, различие между
произнесением слогов /pa/ и /ba/ заключается только в нескольких миллисе-
pa/ и /ba/ заключается только в нескольких миллисе-
/ и /ba/ заключается только в нескольких миллисе-
ba/ заключается только в нескольких миллисе-
/ заключается только в нескольких миллисе-
кундах между началом голосовых вибраций и открытием рта. В случае про-
изнесения слога /pa/ губы открываются чуть раньше, чем начинается произ-
pa/ губы открываются чуть раньше, чем начинается произ-
/ губы открываются чуть раньше, чем начинается произ-
водство звука, в то время как для слога /ba/ произнесение звука незначительно
опережает размыкание губ. Являясь хорошо натренированными читателями и
писателями, мы склоняемся к представлению о том, что язык строится из по-
следовательности букв, но это заблуждение. Устная речь включает набор арти-
куляционных движений, которые формируют частотные вибрации в голосовом
тракте. В то время как мы способны зрительно различать отдельные буквы (на
этой странице), не существует такого понятия, как отдельная изолированная
устная согласная. Мы не можем произнести /k/ без воспроизведения соответ-
k/ без воспроизведения соответ-
/ без воспроизведения соответ-
ствующего звука, даже если это просто длинное /hhhh/, только потому, что го-
hhhh/, только потому, что го-
/, только потому, что го-
лосовая щель должна закрыться и открыться для произнесения звука. По этой
причине физика звука свидетельствует о том, что простые слоги (как /ba/ и /pa/)
и отдельные гласные могут являться элементарными единицами речи. Неко-
торые теоретики предполагают, что люди используют слоговую азбуку, т. е. не
только алфавит фонем, а более широкий набор разговорных знаков из комбина-
ции гласных и согласно-гласных звуков (Levelt and Wheeldon, 1994). Слоговая
азбука должна быть больше, чем просто набор фонем, а также отражать способ
производства звуков речи.
Из этих положений следует, что фонемы человеческого языка являются аб-
страктными понятиями, так же как зрительные объекты. Наши глаза не видят
деревья, вместо этого они получают проекции светлого и темного на сетчатку,
которые интерпретируются корой головного мозга как трехмерные деревья, на-
ходящиеся на определенном расстоянии от глаз. Аналогично, фонемы — аб-
страктные категории звуков, которые очень сильно зависят от соседствующих
речевых сигналов. Когда человек говорит /ba/ и /bee/, акустическая информация
от открывания рта, приводящего к возникновению согласного звука /b/, значи-
b/, значи-
/, значи-
тельно отличается в обоих случаях. В случае произнесения /bee/ происходит
быстро нарастающий переход для выделения согласной, в то время как для
/ba/ — быстро снижающийся переход (см. гл . 7). В то же время мы восприни-
маем согласную /b/ как одну и ту же в обоих случаях. Это очень типично для
человеческих языков, которые, как правило, имеют около сорока абстрактных
фонем, обозначающих, в свою очередь, значительно более широкий набор фи-
зических звуков, слышимых нашим ухом. Восприятие абстрактных фонем яв-
ляется видоспецифическим признаком.
Отражается ли восприятие фонем активацией четких групп нейронов коры
со специфическими свойствами? Мы не знаем ответа на этот вопрос. В случае
зрения, клетки со специфическими свойствами, отвечающие на цвет, ориента-
цию линий и т. д., были впервые обнаружены у обезьян, макак-резусов, при по-
мощи регистрации единичных нейронов. Такие эксперименты невозможно про-
водить на людях, а макаки биологически не адаптированы для восприятия речи.
70
Глава 11
По этим причинам для ответов требуется возникновение улучшенных методов
визуализации деятельности мозга (но см. рис. 11 .13 и 11.14 для ознакомления
с современными исследованиями мозговых процессов восприятия речи и рече-
подобных звуков). Сообщения о первых высокоточных фМРТ-исследованиях
слуховой коры макак были опубликованы перед выходом этой книги (Petkov
et al., 2006). В следующие несколько лет мы можем узнать, существуют ли
нейроны со специфическими свойствами в слуховой коре. Из-за того что мозг
имеет тенденцию выполнять схожие функции одинаковым путем, это предпо-
ложение кажется разумным. Но точных доказательств пока нет.
Рис. 11.13 . Показаны различные уровни активности слуховой коры в ответ на разные
речеподобные звуки. Современные исследования показывают более детальное разби-
ение слуховых и воспринимающих речь областей. С усовершенствованием методов
визуализации станет возможным локализовать группы чувствительных речевых нейро-
нов, что уже сделано для зрительной коры. (Источник: Langers et al., 2003.)
Рис. 11 .14 . Высовывание языка без произнесения звуков. Даже в случае, когда не про-
износятся звуки, простое выдвижение языка вперед вызывает активизацию «речевых»
областей мозга в обоих полушариях. Из этого следует, что язык и ротовая полость
имеют широкие представительства в моторной коре в противоположность, например,
представительству спины. В моторной коре наиболее хорошо представлены важные
функции двигательного контроля и в то же время мало представлены менее важные.
(Источник: Dogil et al., 2002.)
Речь
71
4.0 .
Планирование и формирование речи
Формирование выходной информации в речевых зонах головного мозга по
крайней мере также сложно и адаптивно, как восприятие и понимание речи.
На рис. 11 .15 представлены связи между восприятием и формированием речи.
На рис. 11 .16 продемонстрована современная модель выхода информации на-
чиная с концептуального уровня. Считается, что грамматическое кодирование
предложений включает леммы, как формулы для перевода семантики в мор-
фемы и фонемы, которые управляют голосовыми движениями. Очень важно
понимать, что все эти лингвистические единицы абстрактны, таким образом,
любая лингвистическая фонема, к примеру, может быть произнесена несколь-
кими способами в зависимости от соседних фонем. Голосовой тракт является
физической системой, и требуется время для перемещения языка, например, от
зубов (формируя согласную /th/) до мягкого неба в задней части ротовой поло-
th/) до мягкого неба в задней части ротовой поло-
/) до мягкого неба в задней части ротовой поло-
сти для произнесения звука /g/. В результате существует некая небрежность в
создаваемом акустическом сигнале из-за смазывания голосовой жестикуляции.
В английском языке, например, в слове /tan/ предопределен последний звук /n/,
в то время как произносится звук /ae/. Следовательно, звук /ae/ произносится в
нос или «сочленяется». Таким образом, реальные звуки речи изменяются, даже
несмотря на то, что абстрактные фонемы изображаются одинаково в случае
звука /ae/ в словах «act» и «tan». Если вы внимательно прислушаетесь, то смо-
ae/ в словах «act» и «tan». Если вы внимательно прислушаетесь, то смо-
/ в словах «act» и «tan». Если вы внимательно прислушаетесь, то смо-
act» и «tan». Если вы внимательно прислушаетесь, то смо-
» и «tan». Если вы внимательно прислушаетесь, то смо-
tan». Если вы внимательно прислушаетесь, то смо-
». Если вы внимательно прислушаетесь, то смо-
жете услышать разницу в произнесении.
Обратите внимание, что эта ситуация имеет аналогии в зрительной системе.
В зрительной коре поле V1 отражает информацию, получаемую от сетчатки.
Изображение получается очень детальным из-за маленьких рецептивных полей
Рис. 11.15. Пути между областями, ответственными за производство и восприятие
речи. Представлены результаты трактографического исследования связей между об-
ластями воспроизведения и восприятия речи. Изображение, полученное при помощи
трактографии, демонстрирует дугообразный пучок, соединяющий зоны Брока и Верни-
ке, на нем также можно отметить наличие дополнительных путей. Левая часть рисунка
отражает математически определенное весовое соотношение связей в левом и правом
полушариях соответственно. (ЗБ — Зона Брока; ЗВ — Зона Вернике; КП — крючковид-
ный пучок; ВВИ — верхняя височная извилина; СВИ — средняя височная извилина).
(Источник: Parker et al., 2005.)
72
Глава 11
и высокой оптической разрешающей способностью. И даже несмотря на то, что
последующие зрительные области тоже имеют зрительнотопические карты,
они менее детализированы и более абстрактно отображают признаки зритель-
ного стимула, такие как цвет, принадлежность к объекту и другие. Представле-
ние о том, что в зрительной системе происходит абстрагирование информации
по мере перехода на более высокие уровни подтверждено значительным чис-
лом фактов. То же мы можем ожидать и в случае речи, однако доказательства
этого малочисленны.
Следует заметить, что не только кора головного мозга контролирует рече-
вую функцию. Действия, изначально инициируемые произвольно в лобной
коре, также контролируются базальными ядрами и мозжечком. Традиционно
считается, что базальные ядра вовлечены в контроль автоматизированных ком-
понентов движений, таких как особенное произношение звука /r/ по сравнению
с /l/, что, например, является очень сложным для носителей японского языка.
У англоязычных людей схожие трудности возникают при произнесении зву-
ков французского языка /u/ и /oo/. Такое высоко тренированное произнесение
сложных звуков не полностью контролируются корой. В дополнении к этому
мозжечок, по сложившимся представлениям, требуется для контроля точных
тонких движений.
Однако современные данные нейровизуализации свидетельствуют об уча-
стии и мозжечка, и базальных ядер в чисто когнитивных аспектах мозговой
активности, включая рабочую память и некоторые типы ассоциативного обуче-
ния (рис. 11 .17).
Рис. 11 .16 . Производство речи: от значения до артикуляции. Модель производства речи.
Сравните с рис. 11 .1 . (Источник: Dogil et al., 2002.)
Речь
73
Несмотря на то что речевые области мозга были известны с XIX в., точ-
ные функции зон Брока и Вернике до сих пор до конца неясны (см. гл. 1). Не-
правильно думать, что Брока и Вернике ошиблись в своих предположениях о
производстве и понимании речи. Напротив, с увеличением точности наших на-
блюдений, мы обнаруживаем больше функций для этих классических областей
Рис. 11 .17. (а) Нейронная сеть для производства речи: дополнительная моторная об-
ласть (1) и цингулярная моторная область (2) связаны с основной моторной корой (3).
Подкорковая активация происходит в таламусе (4), базальных ядрах (не показаны),
красном ядре (6) и мозжечке (5). Дополнительно задняя часть верхней височной изви-
лины в обоих полушариях активируется при производстве речи (7). В мозговом стволе
такие области, как ядро подъязычного нерва (8), также иннервируют процессы произ-
водства речи. Корковые зоны совместно с базальными ядрами, таламусом и мозжечком
участвуют в речевом контроле. (б) Подкорковые дополнительные области, такие как
базальные ядра, вовлечены в последовательные двигательные акты речи. Мозжечок и
таламические ядра (не показаны) также играют важную роль. (Источник: (а) Soros et
al., 2006; (б) Angerine, 2002.)
74
Глава 11
речи в левом полушарии, а также функции, связанные с речью, вне традици-
онных границ этих областей. Наши знания постоянно пересматриваются. Мы,
можно сказать, исследуем новую планету, и с приближением нашего космиче-
ского зонда к ней необходимо картировать более детально удивительную гео-
графию нового мира.
5.0 .
Эволюционные аспекты производства
и восприятия речи
Наш высокоспециализированный голосовой аппарат настроен на производство
устной речи. Он эволюционировал из неречевых голосовых органов предковых
видов. Они, в свою очередь, были сформированы на основе дыхательного аппа-
рата, который одинаков у нас и других видов наземных животных, так же как и
нейромышечный механизм жевания и глотания. Например, молодые древесные
гиббоны в дикой природе издают звуки очень похожие на звуки, издаваемые
человеческим ребенком во время игр. Однако вскоре после рождения челове-
ческие малыши начинают лепетать, экспериментируя при этом со слогами, ко-
торые значительно отличаются от неречевых звуков (Schirmer and Kotz, 2006).
Анатомически вокализация требует двойного контроля со стороны дыха-
тельной системы и системы оценки эмоциональных выражений лиц (гл. 2).
В социально провокационных ситуациях голосовые звуки, от воркования ре-
бенка до печального стона, крика злости или возгласа горя, могут быть изда-
ны с минимальным исполнительным контролем латеральной префронтальной
коры. На рис. 11 .18 схематически изображен двойной контроль вокализации.
Как вы сами видите, это непростая система, эволюционировавшая многие мил-
лионы лет.
В данной главе нам важно рассмотреть правую половину этой диаграммы,
а именно корковый контроль речевой функции, начинающийся от префрон-
тальной коры и зоны Брока (в широком понимании), в то время как в случае
входящей речевой информации мы интересуемся восприятием и пониманием
речи.
Корковые речевые области очень близко связаны со слухом (в случае сен-
сорного входа) и с корковыми представительствами ротовой полости и голосо-
вого тракта (для выхода). Классические зоны Брока и Вернике прилегают к кор-
ковым регионам производства и понимания речи соответственно (рис. 11 .19).
На рис. 11 .20 представлено расположение рта и голосового аппарата на гомун-
кулусе моторной коры, которая прилегает к зоне Брока. Прямая электрическая
стимуляция точек карты моторной коры приводит к сокращению определенных
мышц, однако стимуляция премоторных областей, таких как ПБ 6 (поле Брод-
мана 6), вызывает «желание подвигать» соответствующей частью тела. Счита-
ется, что результат стимуляции моторной коры находится под контролем врача,
а не пациента. Зону Брока можно рассматривать как премоторную область коры
для речи, т. е. корковую зону, отвечающую за «намерение заговорить». Стиму-
ляция зоны Брока у пациента в состоянии сознания, вызывает блокировку на-
мерения говорить (Quinones-Hinojosa et al., 2003).
Очень многое стало известно о корковых представительствах речи и языка,
области производства и восприятия речи расширились, а также разделились на
более маленькие специализированные зоны. Дополнительные функции были
Речь
75
открыты для классических зон Брока и Вернике, как и в случае «зеркальных
нейронов», которые рассматриваются в гл. 14 . Восприятие речи (но не произ-
водство) также задействует «неговорящее» полушарие головного мозга (правое
у большинства людей), несмотря на то, что планирование и производство речи
обычно лимитируется левой стороной.
Рис. 11 .18 . Речь могла возникнуть от социально обусловленного производства звуков.
Вокализация имеет двойной контроль, так же как дыхание, лицевая мимика, движения
глаз и другие двигательные системы. Левую ветвь диаграммы часто называют эмоци-
ональным двигательным путем. Начинаясь в префронтальной коре, она следует клас-
сическим путем, отвечающим за вокализацию у млекопитающих, который включает
лимбическую систему и околоводопроводное серое вещество мозга (ОСВ). ОСВ играет
огромную роль в вокализации горя, например, когда детенышей крысы отлучают от ма-
тери. Как и эмоциональный двигательный путь, правая ветвь диаграммы берет начало
в префронтальной коре, а затем следует по более изученным ступеням от зоны Брока
до моторной коры и оттуда к черепно-мозговым нервам, участвующим в голосовом кон-
троле. Оба пути получают входы от базальных ядер и мозжечка. Правый путь находится
под бóльшим произвольным контролем. (Источник: Holstege et al., 2004.)
76
Глава 11
6.0.
Слова и их значения
Английский язык принадлежит индоевропейской языковой группе. Эта группа
включает в себя персидский язык и некоторое число языков населения Индий-
ского полуострова. Другие языковые группы иначе формируют такие осново-
полагающие единицы речи как слова. Финский и турецкий языки называют
«агглютинирующими», потому что в них морфемы соединяются в длинное
высказывание, как невероятно длинное английское слово «antidisestablishmen-
antidisestablishmen-
tarianism» (англ., против отделения церкви от государства). Такая длинная цепь
морфем является нормальным явлением для агглютинирующих языков. Такие
тональные языки, как китайский и тибетский, применяют противоположный
Рис. 11 .19. Производство и пони-
мание речи очень сильно перекры-
ваются. Области, ответственные за
производство и понимание речи, по-
стоянно обмениваются информацией
как напрямую через внутрикорковые
пути, так и не напрямую, в то время
как мы слышим собственный голос.
Как было сказано в гл. 2, существует
достаточно доказательств того, что
люди в течение дня практически по-
стоянно беседуют с собой мысленно.
К тому же исследования во время
сна продемонстрировали наличие
внутренней речи во время дремоты
и сна. S1 — стимул 1, S2 — стимул
2, W — зона Вернике, ДПК — дор-
сальная префронтальная кора. (Ис-
точник: Dogil et al., 2002.)
Рис. 11 .20 . В гомункулусе моторной
коры (ПБ 5) мышечный контроль ро-
товой полости, челюсти, языка, голо-
совых связок, а также действия жева-
ния и глотания располагаются около
зоны Брока для контроля речевой
функции (ПБ 6, 44 и 45). «Моторный
гомункулус» впервые был открыт
Вальдером Пенфильдом при помощи
электрической стимуляции моторной
коры у бодрствующих пациентов во
время диагностических операций
(Penfield and Roberts, 1959). Обрати-
те внимание, что процессы жевания,
вокализации и глотания отмечены
около ротовой области гомункулуса.
(Источник: Standring, 2005.)
Речь
77
подход, формируя предложения из довольно коротких слов, модулируемых при
помощи богатого мелодического видоизменения слогов, так что слово «Chang»
может иметь довольно разное значение в зависимости от тонального контура.
Однако во всех языках существуют лексические единицы некоего рода — фраг-
менты из одного или более слогов, которые обозначают категории значений или
взаимоотношения между категориями.
На рис. 11 .21 показана современная модель того, как мозг переводит одни
лингвистические коды в другие. Как было продемонстрировано на рис. 11 .1,
язык можно рассматривать как систему с двойной иерархией, с одной входной
стороны, от звука к смыслу, с другой — выходной — от смысла к голосовым
движениям. Но мы можем повторять бессмысленный набор слогов, таких как
«fronk», даже несмотря на то, что они не имеют определенного смысла, и из
рис. 11 .15 следует, что такое перекодирование от звука к речи может происхо-
дить в нижней теменной коре. Традиционно считается, что зона Брока отвечает
за перевод мысли в речевую артикуляцию, и в настоящее время более точный
термин для нее — левая нижняя лобная извилина (Л-НЛИ), потому что эти про-
цессы затрагивают активацию более обширных областей. Более ранний термин
«зона Вернике» для понимания речи сейчас чаще заменяют ВТЗ-объединением
(височно-теменно-затылочное). Такая локализация функции логична, так как
височная доля необходима для понятий, основанных на памяти, включая те,
которые получены путем абстрагирования от зрительных объектов, как разли-
чия между животными и неживотными, рабочими инструментами и частями
тела. Как было рассмотрено в гл. 9 и 10, существует возможность, что эти очень
специфичные семантические области служат маркерами для обширных клас-
сов слов и понятий, нежели просто локализованными семантическими полями.
Рис. 11 .21 . Активация мозга в ответ на слова, имеющие значение, и бессмысленные
слова. Слова являются не просто последовательностью фонем. Несмотря на то что моз-
говой базис значений слов остается неизученным (вероятно, из-за того, что значения
слов являются частью долговременной памяти, см. гл . 9), слова со значением вызывают
активацию определенных областей мозга по сравнению с бессмысленными словами.
В данном исследовании при сравнении были выявлены три области: височно-теменная
переходная область (втПО), задняя часть средней височной извилины (зСВИ) и огибаю-
щая извилина (ОИ). (Источник: Friederici and Kotz, 2003.)
78
Глава 11
6.1.
Культурная сокровищница слов и идей
Язык используется как средство для обмена смысловым контекстом. В то время
как фонемы определяют как минимальные речевые единицы, которые обуслав-
ливают различия в значении, слова, в отличие от фонем, относятся к предметам.
Слова — основные строительные блоки смысла. Однако предложения и в осо-
бенности утверждения, построенные на основе предложений, могут быть взяты
за основную «смысловую формулу», аналогичную математическим формулам.
Мы думаем и общаемся при помощи предложений. Многие высказывания, раз-
умеется, сжаты, т. е . они обозначают целую мысль всего лишь несколькими
словами. Тем не менее они выражают семантическое утверждение некого рода.
Как было предложено в гл. 10, слова естественного языка — это безмерное
наследство полезных блоков смысловых знаний, полученных за века. Действи-
тельно во многих случаях можно проследить происхождение слова за более
чем тысячелетний период времени. Одним из удобных примеров служит ан-
глийское слово «quality» (качество), которое произошло от латинского перевода
(сделанного Цицероном) древнегреческого высказывания «po io tes» или «како-
po io tes» или «како-
io tes» или «како-
io tes» или «како-
tes» или «како-
tes» или «како-
» или «како-
во состояние». Цицерон, вероятно, встречался с этим выражением в диалогах
Сократа. До того момента, как изобретенное им слово qualitas стало популярно,
в европейских языках не было способа для определения «состояния» звука, вку-
са или других событий. Мы всегда можем использовать более длинные фразы,
выражая идею, для которой не существует одного слова, но слова позволяют
нам работать с понятиями как с целыми блоками в нашей очень ограниченной
рабочей памяти. Таким образом, изобретение новых и полезных слов является
реальным вкладом в нашу способность понимать мир.
Науку сложно представить без такого слова, как «качество». Однако потре-
бовалось некое осознание в определенной точке интеллектуальной истории, бо-
лее двадцати столетий назад, для того, чтобы сформировать понятие «качество»
в том смысле, в котором мы использует его как абстрактное существительное.
Вероятно, сегодня все современнее языки имеют некий термин для обозначе-
ния «качества», потому что мы уже не можем без него думать. Так как совре-
менные языки состоят из десятков тысяч слов и ежедневно образуются новые
слова, мы должны умножить изобретение Цицерона в десятки тысяч раз для
того, чтобы прочувствовать всю совокупность знаний, предаваемую каждому
новому носителю языка.
Тезаурус Роже (идеографический словарь), впервые составленный Питером
Марком Роже (1779–1869), — одна из исторических попыток классифициро-
вать слова естественного языка по семантическим категориям для того, чтобы
можно было найти слова согласно значению. В тезариусе слова с одинаковым
значением сгруппированы вместе, создавая тем самым семантическую сокро-
вищницу идей; действительно слово «тезаурус» означает «сокровищница».
Современные попытки в этом направлении осуществляются с применением
вычислительных методов, но, характеризуя наше знание основных понятий,
можно сказать, что это фантастически смелое предприятие. Система WordNet
Джорджа А. Миллера из Принстонского университета — еще одна попытка,
сделанная в данном направлении (http://wordnet.princeton.edu/), он-лайн систе-
http://wordnet.princeton.edu/), он-лайн систе-
://wordnet.princeton.edu/), он-лайн систе-
wordnet.princeton.edu/), он-лайн систе-
. princeton.edu/), он-лайн систе-
princeton.edu/), он-лайн систе-
. edu/), он-лайн систе-
edu/), он-лайн систе-
/), он-лайн систе-
ма лексических ссылок. За несколько лет эта система классифицировала бо-
лее 200 000 пар слово-значение для английского языка. Аналогичные попыт-
Речь
79
ки предпринимаются и для других языков. По подсчету WordNet в английском
языке существуют 128 000 слов с единственным значением и около 80 000 слов
с множеством значений. Однако в данный подсчет не были включены слова с
несколькими синтаксическими ролями, как слово «book», которое может быть
и существительным (англ., книга), и глаголом (регистрировать арестованного
человека). Слово «книга» в английском языке может также употребляться в ка-
честве прилагательного (например, книжные знания, книжный переплет, кни-
голюб и т. д.).
Таким образом, число слов и значений, которые мы как искушенные носи-
тели языка знаем, поистине огромно. Оксфордский английский словарь (ОАС)
является еще одной значительной попыткой проследить историю становления
обычных английских слов. Очевидно, однако, что действительные истоки слов
потеряны во времени, так как изначально язык был только разговорным сред-
ством, которое приобрело стандартизированный печатный вид значительно
позже изобретения письменности, два или три тысячелетия назад. Тем не ме-
нее ОАС дает нам некое восприятие полного объема человеческого словарного
лексикона и понятий.
6.2 .
Узнавание синонимов
Гительман с соавторами (Gitelman et al., 2005) совершили ультрасовременную
попытку определить области специфичные для речи, используя идентичные
когнитивные тесты для одной группы испытуемых, смотрящих на орфогра-
фию (правила написания), семантику и фонологию в одном опыте. (Синтак-
сис включает последовательность слов, и поэтому рассмотрим его позже.) Ис-
пытуемым демонстрировали пары слов, как на рис. 11.22, и просили отвечать
только «одинаковые» они или «разные». В случае предъявления слов «rain» и
«reign» испытуемые должны были сказать, что данные слова являются омофо-
reign» испытуемые должны были сказать, что данные слова являются омофо-
» испытуемые должны были сказать, что данные слова являются омофо-
нами (ОМО), т. е . словами, которые независимо от разного написания имеют
одно звучание. В случае «boat» и «ship» испытуемые должны были определить
их как синонимы (СИН), разные слова с одним значением. А при предъявле-
нии «aunt» и «tuna» испытуемые должны были назвать их анаграммой (пере-
aunt» и «tuna» испытуемые должны были назвать их анаграммой (пере-
» и «tuna» испытуемые должны были назвать их анаграммой (пере-
tuna» испытуемые должны были назвать их анаграммой (пере-
» испытуемые должны были назвать их анаграммой (пере-
мешанными буквами, АНА); в качестве контроля для определения схожести
им показывали наборы из четырех согласных, которые или совпадали между
Рис. 11 .22 . Примеры стимулов (Gitelman et al., 2005). Условия эксперимента представ-
лены на рисунке и включают фонологические (омофон, ОМО), семантические (сино-
ним, СИН) и орфографические (анаграмма, АНА) контрасты при сходстве и различии
пар слов. (Источник: Gitelman et al., 2005.)
80
Глава 11
собой, или нет. Контрольный эксперимент позволяет исключить процессы,
задействованные при чтении, зрительном восприятии и т. д., активность при
контроле вычитают из активности в других условиях. Было проведено фМРТ-
исследование при предъявлении каждой пары одинаковых-разных слов. Срав-
нения были проведены для каждого испытуемого отдельно с целью исключе-
ния межиндивидуальных различий. Слова были тщательно отобраны для того,
чтобы обеспечить равное число существительных и глаголов, в то время как
другие параметры, например частота предъявления слов, контролировалось
статистически.
На рис. 11.23 представлены результаты, полученные для пар ОМО и СИН.
6.3 .
Современные данные о словах
и их значении фрагментарны
Как было указано в гл. 10, нам известна только пара областей мозга, которая
по-разному отвечает на определенные смысловые значения, такие как инстру-
менты, животные и не животные и другие. Функция этих областей до сих пор
неясна. Они могут выступать в роли индексов в обширной сети смысловых вза-
имосвязей неокортекса (Moscovitch, 1992). Существует возможность того, что с
изобретением более совершенных методов регистрации активности мозга, бу-
дут выявлены группы нейронов подобно тезариусу, отвечающие на определен-
Рис. 11 .23. Активация областей левого по-
лушария головного мозга в ответ на разные
значения слов. Вверху: фМРТ-изображение
при сравнивании омофонов — слов с разным
написанием, но одинаковым произношением,
например англ. «rain» и «reigh». Внизу: фМРТ-
изображение при сравнивании синонимов,
таких как англ. «boat» и «ship». Активность
в ответ на предъявление контрольного набо-
ра стимулов была вычтена из каждого фМРТ-
изображения для того, чтобы исключить актив-
ность мозга, связанную с чтением и другими
схожими процессами. Активация, связанная со
смыслом слов (СИН, внизу), более широко рас-
пределена, чем активация мозга при сравнении
омофонов. Активация происходит в верхней
височной извилине и борозде (ВВИ и ВВБ) и
на конце средней височной извилины (СВИ).
Активируется нижняя лобная извилина (НЛИ),
как и латеральная орбитофронтальная извили-
на (ЛОИ). В верхней части левого полушария
мозга при условии СИН отмечена небольшая
активация в дополнительном моторном поле
(ДМП), области, которая ассоциируется с на-
мерением совершить движение. При выпол-
нении задачи по сравнению омофонов (ОМО)
также наблюдается активация в задней цен-
тральной борозде (ЗЦБ). (Источник: Gitelman
et al., 2005.)
Речь
81
ные семантические категории (см. гл . 10). Но с такой же вероятностью смыс-
ловые значения слов широко распределены по мозгу и являются по существу
сетью, а значит, должны вызывать активацию обширных областей мозга.
Однако не следует считать, что семантика не затрагивает определенных об-
ластей коры головного мозга. Некоторое число исследований выявило регионы
мозга, высокочувствительные к различиям между значениями слов. Однако на
данный момент эти методы демонстрируют очень широкие изменения в коре (и
других областях).
7.0 .
Синтаксис, вк лючение и синтагматизация
Синтаксис является наиболее характерной чертой языка. На рис. 11 .24 пред-
ставлена древовидная схема синтаксической структуры предложения с прида-
точным предложением. Древовидные структуры являются пошаговыми, т. е.
позволяют вставлять предложения внутри других предложений. И хотя теорети-
чески могут существовать множество уровней синтаксической рекурсии, мозг
человека лимитирован до восприятия одного или двух вставочных предложе-
ний. Это в основном обусловлено пропускной способностью рабочей памяти.
Когда мы тщательно разрабатываем план действия, как в задачах о ханойской
башне, при игре в шахматы или других сложных задачах, мы обычно исполь-
зуем другие механизмы памяти. Таким образом, схема предложения, представ-
ленного на рис. 11 .24, является типичной для одного предложения. Обратите
внимание на рис. 11 .25, на котором отмечены области, вероятно, участвующие
в обработке синтаксической информации.
Древовидная схема синтаксической структуры предложения напоминает
пространство решений задачи, что было рассмотрено в гл. 10 (см. блок 11.3).
Это сходство не является случайностью. Во многих случаях синтаксис дает нам
основу для формулирования предложения, также как подцели и цели позволяют
Рис. 11 .24 . Древовидная схема синтаксической структуры предложения позволяет
включать в них утверждения. Основная синтаксическая древовидная схема, содержа-
щая вложенное предложение. (Источник: Gitelman et al., 2005.)
82
Глава 11
планировать серию действий, например осуществить навигацию от одной точ-
ки в другую или сделать ход в шахматах. Синтаксис, таким образом, можно ис-
пользовать в качестве когнитивного инструмента для планирования, к которому
люди хорошо приспособлены. При разговоре на родном языке планирование
речи является практически имплицитным процессом (см. гл . 10).
8.0 .
Просодия и мелодика речи
Речь и пение являются схожими действиями; с физической точки зрения, пе-
ние — просто удлинение и настройка по высоте гласных звуков. Если растя-
гивать гласные звуки любого слова, например «когнитивная нейронаука», то
получиться монотонное пение. А если изменять высоту каждого слога, то полу-
читься небольшая песенка. Как и речь, музыка и танец являются видоспеци-
фичными для людей процессами. (Биологический смысл этих процессов труд-
но определить: функция языка для выживания очевидна, а музыки?)
Определенный вариант музыкальной шкалы используется повсеместно в
мире. Все музыкальные звуки делятся на октавы. В западной музыке мажор-
ная гамма воспринимается как более радостная, чем минорная. Музыкальные
ноты, так же как и гласные звуки языка, основаны на физическом резонан-
се труб, например бамбуковые флейты или натяжения тетивы. Когда вибри-
рующий столб воздуха наполовину короче трубы, звук становится на октаву
выше. В случае если струна гитары натянута вдвое сильнее, то звук также
получается на октаву выше. Маленькие животные производят высокие звуки,
а большие — низкие из-за различной акустики резонансных полостей. Значит,
существует физическая основа для восприятия звуков, имеющая биологиче-
ское применение. Это объясняет необходимость убегать при реве льва, а не при
стрекотании сверчка.
Рис. 11 .25. Синтаксис вызывает активацию различных областей мозга. Гродзински и
Фридеричи (2006) предположили, что разные корковые области отвечают за разные син-
таксические функции, что и продемонстрировано на рисунке. (Источник: Grodzinsky
and Friederici, 2006.)
Речь
83
Блок 11.3 . Некоторые основы синтаксиса
Структурные блоки синтаксического знания
Действие
Описание
Примеры
Основные взаимосвязи между словами и фразами
ЛЕКС
Лексические связи, которые име-
ют синтаксическую значимость.
Например, аргументная структу-
ра глагола – тип и число аргумен-
тов, которые требуются согласно
структуре естественного языка
1. Аргумент: Он бежал/спал/умер;
2. Аргументы: Он увидел/ударил/преследовал
Марию;
3. Аргументы: Он подарил/передал/послал Сью
подарок
СЛИЯНИЕ Группа принудительных струк-
туроформирующих
операций,
которые разбивают предложения
на иерархические ступени. Дан-
ный пример демонстрирует как
правила синтаксического СЛИ-
ЯНИЯ создают предложения из
набора лексических категорий
(«исчислений»). СЛИЯНИЕ фор-
мирует фразовые узлы (ИЧ =
именная часть, ГГ = глагольная
группа, ПО = предложный обо-
рот) из объединенных категорий
(ОПРеделительные Существи-
тельное, Глагол, Предлог), и в
итоге все сливается в «основу»,
узел предложения
Категории: (ОПР = a, the (в англ. языке); C =
мужчина, женщина, дерево; Г = увидел; ПР =
около
Результат повторного слияния: The man saw
the woman near the tree (англ., Мужчина увидел
женщину около дерева)
Отношения зависимостей в предложении
ДЕЙСТВИЕ Главное действие в древовидной
диаграмме (созданной при помо-
щи СЛИЯНИЯ). Оно связывает
слышимую фразу (= ИЧ, ГГ,
ПО) с одной или более непроиз-
носимой, однако синтаксически
активной, позицией «■» в изме-
ненном варианте предложения
Sam knows that he saw the ballet dancer on Monday
(англ., Сэм уверен, что видел балетного танцора
в понедельник) => Which dancer does Sam know
that he saw ■ on Monday? (Какого танцора Сэм
знает, что увидел ■ в понедельник?
ДЕЙСТВИЕГ Соединяет различные позиции,
которые может занимать гла-
гол. Только в одной позиции
находится глагол, другие («♦»)
–
пустые. Такие отношения по-
казаны для вопросов типа да/
нет в английском языке и для не-
мецкого языка, в котором глагол
«sah» (видел) и его производная
«gesehen» (видел) занимают раз-
ные позиции
В английском: John is tall => Is John ♦ tall?
В немецком: Hans hat Maria gesehen => Hans sah
Maria ♦ => Gestern Sah Hans ♦ Maria ♦
СВЯЗКА
Отношения, которые определя-
ют взаимодействие местоимений
и возвратных местоимений с ИЧ,
на которую они в этом же пред-
ложении опираются
John looked at himself (англ., Джон посмотрел
на себя)
Mary asked John to help her (англ., Мария попро-
сила Джона помочь ей
84
Глава 11
В человеческой речи эмоции выражаются посредством интонации и каче-
ства звука. У людей в грустном состоянии имеется тенденция к снижению ин-
тонационного контура, что, вероятно, отражает понижение давления в нижних
отделах голосовой трубки. Радость обычно выражается приподнятыми звука-
ми. Эмоции выражаются в речи музыкальными интервалами. Деление на ок-
тавы универсально, несмотря на то, что в некоторых культурах это делают по-
другому. Эмоциональные выражения и сигнализация могла предшествовать,
а затем совместно формироваться с денотативным языком. Разные музыкаль-
ные интервалы, т. е. двухтональные последовательности, несут разную эмоци-
ональную окраску.
Даже ритмичность является свойством нормальной речи, что можно на-
блюдать при расстановке ударений в фразе, например: «The rain / in Spain /
falls mainly / in the plain /» (англ., в Испании дождь выпадает, в основном, на
равнинах). Вы интуитивно выделяете ударные слоги в предложениях. Даже в
языках с минимальным количеством ударных слогов, как во французском, все
равно присутствует мелодическая форма у фраз. Малыши лепечут нараспев, и
взрослые непроизвольно начинают при общении с маленькими детьми (и жи-
вотными) использовать преувеличенные интонации. Несмотря на то что у раз-
ных культур наблюдаются разные музыкальные формы, вероятно, существует
некая универсальная (возможно биологическая) взаимосвязь между языком и
музыкой.
9.0.
Значимые утверждения
Мы говорим при помощи не единичных слов, а целых высказываний, т. е. се -
мантически значимых утверждений о мире. Назначение синтаксиса — создание
таких утверждений, но даже в тех случаях, когда синтаксис нарушен, при не-
которых видах афазий у людей все равно сохраняется возможность мыслить и
выражать свои мысли в виде определенных высказываний. Например, больные
с афазией могут указывать на желаемую пищу, выражать чувства дискомфорта
или удовольствия или произносить несколько «паралингвистических» утверж-
дений о них самих и окружающем мире. Предложена (Shastri, 2002) модель ко-
Shastri, 2002) модель ко-
2002) модель ко-
дирования смысловых утверждений в виде нейронной сети гиппокампальных
и кортикальных нейронов. На рис. 11 .26 проиллюстрирована идея, как разные
нейроны могут отражать разные части предложения. Координируя разряды
нейронов (например, при помощи синхронизации в частоте гамма-ритма), мож-
но представить, как целое высказывание отражено в головном мозге.
10.0 . Объединенные образы речи
В XIX в. психолог Вильгельм Вундт выдвинул предположение, что производ-
ство языка начинается с Gesamtvorstelung, объединенного мысленного пред-
ставления предложения, которое человек собирается произнести (Blumenthal,
1979). Эти объединенные представления нуждаются во многих уровнях описа-
ния: семантическом, синтаксическом, фонетическом и, возможно, голосовом,
прагматическом (т. е. включающим цели говорящего) и других. Как только та-
кая многоуровневая структура активации мозга готова, она начинает посылать
команды для перевода параллельной структуры объединенных представлений
в последовательность голосовых движений. На рис. 11 .27 продемонстрирова-
Речь
85
но современное понимание этой идеи. Обратите внимание на существование
параллельных и перекрывающихся уровней описания и контроля, каждый из
которых, вероятно, выражается в чем-то вроде нейронной сети. В данном слу-
чае каждый уровень соотнесен с характерными вызванными потенциалами
(Friederici, 2002). Например, известно, что семантическая информация вызыва-
ет генерацию большой негативной волны с латентным периодом 400 мс после
возникновения стимула (называется N400).
Если это кажется чрезвычайно сложным, значит, можно считать эту схему
верной. Также всегда необходимо понимать, что современный язык является
определенно человеческим достижением. Человеческая ветвь эволюции отде-
лилась от других гоминид около трех миллионов лет назад, и считается, что та-
кие основополагающие элементы человеческого интеллекта, как язык, настой-
Рис. 11 .26 . Создание утверждений затрагивает активацию обширной нейронной сети
головного мозга. На уровне нейронных сетей предложение «Мамы футболистов, ве-
роятно, владеют мини-вэнами», может быть представлено набором нейронных групп,
разряжающихся в ответ на основные компоненты высказывания: мини-вэны, мамы,
футболисты, взаимоотношения типа «владеют чем-то» и подобные. Такие сети для ут-
верждений не были обнаружены в мозге. Данная модель всего лишь предлагает нейрон-
ные сети в том виде, в каком мы их понимаем. (Источник: Shastri, 2002.)
86
Глава 11
чивость при решении задач, совместное и конкурирующее поведение, возникли
порядка 100 000 лет назад. В то время как наш словарный запас — результат ве-
кового культурного развития, биологическая подготовленность нашего мозга и
тела для такой деятельности имеет множество эволюционных слоев, некоторые
из которых общие с млекопитающими или другими сухопутными позвоночны-
ми. Такая сложная система, как изображенная на рис. 11 .27, эволюционировала
многие поколения.
В гл. 3 и 6 продемонстрировано, что у зрительной системы как минимум
один уровень интеграции, «где все сходиться», который располагается в нижне-
височной коре (область НВ) (Sheinberg and Logothetis, 1997). В данной области
нейроны отвечают не на единичные сигналы от сетчатки или отдельные харак-
теристики, такие как цвет или светлые границы, а на весь зрительный стимул.
Рис. 11 .27. Объединение всего. Нейрокогнитивная модель обработки устного предло-
жения. В прямоугольниках отражены функциональные процессы, в эллипсах — соот-
ветствующие мозговые области, выявленные при помощи фМРТ, ПЭТ или ССП. Ней-
роанатомические уточнения (указанные в скобках) получены благодаря фМРТ или ПЭТ.
Компоненты ССП височных отделов соотнесены с временными структурами (с левой
стороны) на основе их функциональной корреляции, нежели локализации источника.
ПБ — поля Бродмана; РЛПН — ранняя левая передняя негативность; ССП — связанные
с событием потенциалы; НЛИ — нижняя лобная извилина; СВИ — средняя височная
извилина; ВВИ — верхняя височная извилина. (Источник: Friederici, 2002.)
Речь
87
Существует вероятность, что у речевой функции имеется такая же система ин-
теграции (Hagoort, 2005; рис. 11 .28).
Идея существования для некоторых функций областей, где «все соединяется
воедино», является очень привлекательной и может быть смоделирована как
в виде нейронных сетей (например, McClelland, 1986; Shanahan, 2005), так и
в символьных моделях когнитивной деятельности (Baars and Franklin, 2003).
Эта идея очень древняя и восходит к Аристотелю, который предполагал, что
человеческое сознание должно обладать возможностью соединять воедино
информацию от всех органов чувств, например зрения и слуха, для создания
мультимодальных объектов мира — гитары, которую можно видеть, трогать и
слышать (Baars, 1988). Аристотель полагал, что должен существовать «здравый
смысл» — некое главное сенсорное чувство, которое объединяет все специаль-
ные чувства в одно целое. Правдоподобным кажется размещение такой функ-
ции в теменных и лобных отделах головного мозга. Однако многие области
мозга имеют центральные нервные связи от ретикулярной формации мозгового
ствола к миндалине, энторинальной коре, префронтальным долям и таламу-
су. Некоторые теоретики считают, что всю кору или таламо-кортикальную си-
стему следует рассматривать как огромную сеть для интеграции, разделения и
распределения сигналов (например, Edelman, 1989; Edelman and Tononi, 2000;
Freeman, 2004). Кто-то может считать, что мозг состоит из узлов, связанных
с некоторыми понятиями сознания, что обсуждалось в гл. 8. Идея Хагоортa
о том, что левая НЛИ является анатомическим центром для интеграции языка и
понятий, — одна из версий этой базовой идеи. Однако, как было сказано выше,
привлекательность и возможность идей нельзя рассматривать как доказатель-
ство, и данные гипотезы нуждаются в дополнительных фактах.
Рис. 11 .28 . Область интеграции. Хагоорт (2005) указал, что зона Брока имеет функции,
выходящие за пределы только контроля речи. Однако он полагает, что расширенное по-
нимание зоны Брока может рассматриваться как область объединения речи и семанти-
ческой информации. Для того чтобы избежать путаницы с классической зоной Брока,
эту часть коры головного мозга называют согласно ее расположению левой нижней лоб-
ной извилиной (Л-НЛИ). (Источник: Hagoort, 2005.)
88
Глава 11
11.0 . Заключение
Речь является исключительно человеческой способностью, позволяющей пере-
давать культурные знания через пространство и время. Мозговые механизмы
речи до сих пор выясняются, но отправными точками все равно являются от-
крытия XIX в. Зона Брока для производства речи и зона Вернике для понимания
речи — только часть областей мозга, задействованных в этом процессе. Совре-
менные открытия расширили и поделили классические речевые зоны, так что
левая нижняя лобная извилина (Л-НЛИ) является более уместным названием
для зоны Брока, а задние слуховые и речевые зоны теменной и височной коры
больше подходят под функции зоны Вернике. Однако существует постоянная
взаимосвязь между передними и задними речевыми областями, и такое жесткое
разделение их в некой мере искусственно. В дополнении можно сказать, что
есть факты, указывающие на существенную роль правого полушария в воспри-
ятии речи.
Каждый уровень лингвистического восприятия продемонстрирован с помо-
щью методов нейровизуализации (см. рис. 11 .1). Тем не менее у нас недоста-
точно детализированных знаний относительно речевых зон коры, аналогичных
тому, что мы знаем о зрительных областях. Одна из основных причин этому —
отсутствие подходящей модели на животных, как макаки-резусы для изучения
зрения. Однако современные технологии, которые можно применять на челове-
ке, начинают достигать необходимого уровня детализации, и мы скоро сможем
обнаружить специфичные области ввода и вывода речевой информации.
Очень многие процессы речи и языка зависят от долговременной памяти,
однако многие ученые считают, что мы в конечном итоге должны искать ши-
роко распространенные кортикальные сети, которые отвечали бы за словарный
запас, синтаксис и семантику языка. Вероятно, эти сети зависят от синаптиче-
ских контактов большого числа нейронов. Современные методы только сейчас
начинают достигать такого уровня, которые позволят получить доступ к сете-
вым процессам нейронов.
12.0 . Задания и упражнения к главе 11
1. Заполните ячейки иерархической обработки речи на рис. 11 .29.
2. Приведите примеры «нисходящей» или «обусловленной ожиданием» обра-
ботки входящей речевой информации. Что происходит при выводе инфор-
мации?
3. Какие существуют доказательства биологической приспособленности чело-
века к языковым процессам?
4. В чем сходство человеческого голосового аппарата с музыкальными инстру-
ментами?
5. На рис. 11.30 отметьте следующее:
а) корковую область, вероятнее всего, связанную с восприятием речи;
б) область для планирования речи;
в) область двигательного контроля голосового тракта, например языка;
г) область для координации восприятия и производства речи (возможно, ее
придется нарисовать).
6. Какова роль, если таковая имеется, в речевой функции у правого полушария?
Речь
89
7. Назовите примерный объем лексикона (словарного запаса естественного
языка). Что известно об организации такого числа слов и понятий?
8. Объясните, что такое синтаксическая неоднозначность. Что дает неодно-
значность для понимания процессов обработки речи?
Рис. 11 .29 . Иерархия процессов обработки лингвистической информации
Рис. 11 .30
Реконструкция травмы, полученной Финеасом Гейджем. Реконструкция
несчастного случая на железной дороге, произошедшего в 1848 г. с двадцати
пятилетним Финеасом Гейджем. Обратите внимание на возможное повреждение
орбитофронтальных областей и медиальной поверхности лобных долей.
Подобные ранения приводят к повреждению мозга от отека, кровотечения,
повышения температуры, инфекции, воспаления и физической деформации
тканей, которые распространяются далеко за пределы области непосредственной
травмы. Таким образом, на самом деле мы не знаем объем повреждений мозга
у этого классического неврологического пациента. (Источник: Squire et al., 2003.)
Он порывист, непочтителен, временами с грубой профанацией предается
удовольствиям (чего ранее не наблюдалось у него), не проявляет уважения к
товарищам, не выносит отказов или советов, если они противоречат его же-
ланиям... Будучи ребенком по интеллектуальным способностям и действиям,
он обладал неудержимыми животными инстинктами, свойственными силь-
ному мужчине... Его разум так радикально изменился, что друзья и знакомые
стали утверждать, что он «больше не Гейдж».
Харлоу Дж. М . Восстановление после проникновения
железного прута через голову.
Publ Mass Med Soc 1868; 2: 327–347. Процитировано в:
BMJ 1998; 317: 1673–1674 (19 декабря)
«Больше не Гейдж»: как железный прут прошел через голову.
Содержание
1.0.Введение........................................................91
1.1.Множествосложныхфункцийлобныхдолей.......................92
1.2.Отмолчащихдолейкоргануцивилизации.........................93
2.0.Филогенезионтогенез.............................................93
2.1.Филогенез....................................................94
2.2.Онтогенез....................................................95
3.0.Обзорфункций....................................................95
3.1.«Памятьбудущего»................................................96
3.2.Самовосприятиеиуправляющиефункции............................96
4.0.Болеедетальноерассмотрениелобныхдолей.........................97
4.1.Нейроанатомияинейрофизиологиялобныхдолей..................97
4.2.Какустановитьграницыпрефронтальнойкоры.....................97
4.3.Обширныепроводящиепутилобныхдолей........................97
5.0.Болеедетальноерассмотрениефункцийлобныхдолей.................100
5.1.Функциирегуляции...........................................100
5.2.Социальноевзрослениеинравственноеразвитие..................102
6.0.Нейровизуализацияуправляющегомозга.............................104
6.1.Вниманиеивосприятие........................................104
6.2.Рабочаяпамять...............................................107
6.3.Управляющиефункцииимоторныйконтроль.....................110
6.4.Принятиерешения............................................112
6.5.Усвоениеправил..............................................114
7.0.Нарушениялобныхдолей...........................................115
7.1.Хрупкиелобныедоли.........................................115
7.2.Синдромыповреждениялобныхдолей...........................116
7.3. Повреждение лобных долей и асоциальное поведение . . . . . . . . . . . . . . 119
7.4 . Другие клинические повреждения, связанные с повреждением
лобныхдолей................................................120
8.0 . Современный взгляд на организационные принципы лобных долей . . . . . . . 122
9.0.Кединойтеорииуправляющегоконтроля:заключение.................. 123
10.0.Заданияиупражнениякглаве12..................................124
1.0 . Введение
Что объединяет данные поведенческие проявления? Заучивание телефонного
номера друга с листка бумаги. Принятие решения о том, что сначала надо поза-
ниматься, а потом только идти играть в баскетбол. Разработка нового маршрута,
ко гд а дорога закрыта на ремонт. Концентрирование внимания во время чтения
Глава 12
Цели, управляющий контроль
и действие
92
Глава 12
данного предложения. Изменение решения по поводу поднятия ставок во вре
мя игры в покер. Все эти виды поведения — и многие другие — определяются
лобными долями.
1.1 .
Множество сложных функций лобных долей
Лобные доли называют центром контроля таких функций, как концентриро
вание внимания на одном определенном предмете, планирование и пересмотр
планов, когда это требуется, мониторинг событий в окружающем мире — эти
сложные задачи составляют часть нашей повседневной жизни. Лобные доли
также описывают как «доли действия», ответственные за планирование физи
ческих движений и инициирования активности в моторной системе. Они слу
жат «домом» для нашей личности и социальных принципов поведения. Каким
образом одна область мозга может обеспечить нормальное выполнение таких
сильно различающихся функций?
Ответ на эти вопросы до сих пор окончательно не найден, однако многое
мы знаем о функциях лобных долей из большого числа исследований. В экс
периментальных работах на животных роль лобных долей изучается при помо
щи регистрации единичных нейронов. У людей определение функций лобных
долей происходит за счет исследования мозговых повреждений и заболеваний.
Ключевой фигурой в наших знаниях о лобных долях мозга является Финеас
Гейдж, железнодорожный рабочий с обширным повреждением фронтальных
отделов головного мозга вследствие несчастного случая на работе. Неожидан
ное изменение личности у Финеаса после травмы пролило свет на роль лобных
долей в формировании индивидуальности и социальных навыков. Изучение
людей с повреждением лобных долей изза болезни, мозговой травмы или раз
рушения способствовало разработке функциональных категорий для лобных
долей в зависимости о того, где происходили повреждения или разрушения и
какие нарушения поведения вследствие этого наблюдались. Недавние иссле
дования с помощью метода нейровизуализации на здоровых испытуемых обе
спечили ученых новой информацией о локализации паттернов активности в
лобных долях во время выполнения заданий, требующих участия различных
функций лобной коры, таких как произвольное или управляющее внимание и
принятие решений.
Лобные доли содержат мозаику различных типов клеток и корковых обла
стей, различающихся по типу клеточной структуры, анатомическим особенно
стям и имеющимся связям. В отличие от сенсорных областей коры у лобных
долей нет единой функции — они не специализированы для обработки речевых
звуков или узнавания лиц. Скорее, лобные доли вовлечены во все аспекты чело
веческой когнитивной деятельности.
В данной главе мы представим результаты нейровизуализационных иссле
дований, которые обеспечили новой информацией о том, где, какие и каким
образом определенные области лобных долей активируются при выполнении
когнитивных задач. Далее мы рассмотрим нарушения работы лобных долей,
которые наблюдаются у пациентов с повреждением этих и связанных с ними
областей. В конце мы увидим, как соотносятся результаты экспериментов с по
мощью методов нейровизуализации и исследований пациентов и что они дают
нам для понимания роли лобных долей в поведении человека.
Цели, управляющий контроль и действие
93
1.2.
От молчащих долей к органу цивилизации
Ученым потребовалось много времени для того, чтобы понять важность лоб
ных долей для когнитивной деятельности. Но когда это произошло, сформиро
валась сложная и четкая картина функций лобных долей. Основное внимание в
данной главе будет уделено префронтальной коре (ПФК), наиболее фронтально
расположенному отделу лобных долей, кпереди от моторных зон. Выделяют
много подобластей в ПФК, но обычно рассматривают четыре области с чет
ко определенной функциональной ролью в когнитивной деятельности: дорсо-
латеральная ПФК (ДЛПФК), вентролатеральная ПФК (ВЛПФК), передняя
ПФК (ППФК) и медиальная ПФК (МПФК) (рис. 12 .1).
ПФК расположена кпереди от первичной моторной коры. Раньше считалось,
что лобные доли являются «молчащими», потому что их трудно соотнести с
одиночными, легко выделяемыми функциями. Однако за последнее десятиле
тие с развитием технологий визуализации появилась возможность выделить
более дробные области в пределах ПФК для таких функций, как управляющий
контроль, поиск конфликтных событий, эмоций и рабочей памяти. Тем не ме
нее существует немалый спор по этому вопросу.
2.0 .
Филогенез и онтогенез
Если вам кажется, что только у человека лобные доли головного мозга играют
важную роль в когнитивной деятельности, то вы совершенно правы! Сильное
увеличение лобных долей во время эволюции и их дальнейшее созревание в
течение жизни уникальны для человека по сравнению с другими живыми ор
ганизмами.
Рис. 12 .1 . Основные деления префронтальной коры. Префронтальная кора может быть
разделена на латеральные (сбоку), медиальные (по средней линии), вентральные (сни
зу) и фронтальные области. Латеральные области, в свою очередь, делятся на верхние
(дорсальные) и нижние (вентральные) половины, разделенные большой горизонталь
ной складкой, нижней латеральной бороздой. (Источник: Simons and Spiers, 2003.)
94
Глава 12
2.1.
Филогенез
В эволюции лобные доли значительно увеличились в размере только у чело
векообразных обезьян. Эти области претерпели сильное развитие только на
поздних стадиях эволюции. Согласно Бродману (1909), ПФК или ее аналоги
занимают 29% от поверхности коры у человека, 17% у шимпанзе, 11,5% у гиб
бона и макак, 8,5% у лемура, 7% у собак и только 3,5% у кошки (рис. 12 .2).
В то время как у китов и дельфинов объем мозга тоже достаточно большой, у
морских млекопитающих получила наибольшее развитие теменная, а не пре
фронтальная кора.
Рис. 12 .2 . Префронтальная кора расширяется в ходе эволюции млекопитающих и при
матов. Значительно увеличенная ПФК является определенно особенностью человека
и приматов. У других животных с большим мозгом, как у китов и дельфинов, большее
развитие получила теменная кора, а не префронтальная. На правом нижнем рисунке
изображен мозг человека, а слева — мозг шимпанзе. (Источник: Squire et al., 2003.)
Цели, управляющий контроль и действие
95
2.2 .
Онтогенез
Как и определение целей, прогнозирование и планирование, так и развитые
лобные доли являются однозначно чисто человеческой особенностью. В 1928 г.
невролог Тилней предложил всю эволюцию человека назвать «эрой лобных до
лей», однако научный интерес к ПФК возник гораздо позже. Только понемногу
она стала приоткрывать свои секреты таким великим ученым и клиницистам,
как Хьюлингс Джексон (Hughlings Jackson, 1884) и Александр Лурия (Alexander
Luria, 1966), а за последнее десятилетие таким исследователям, как Антонио
Дамасио (An�onio �amasio, 1993), Хоакин Фустер (Joaquin �us�er, 1997), Па
An�onio �amasio, 1993), Хоакин Фустер (Joaquin �us�er, 1997), Па
�amasio, 1993), Хоакин Фустер (Joaquin �us�er, 1997), Па
�amasio, 1993), Хоакин Фустер (Joaquin �us�er, 1997), Па
1993), Хоакин Фустер (Joaquin �us�er, 1997), Па
Joaquin �us�er, 1997), Па
�us�er, 1997), Па
�us�er, 1997), Па
1997), Па
трисия ГолдманРакич (Pa�ricia GoldmanRakic, 1987), Доналд Стасс и Фрэнк
Бенсон (�onald S�uss, �rank Benson, 1986) и другим.
3.0 .
Обзор функций
Если лобные доли являются «органом цивилизации», какова же их функция?
Каков их «цивилизующий» эффект? Функции лобных долей не поддаются про
стому описанию. Они не вовлечены ни в какие простые, легко определимые
процессы. Пациенты с поражением лобных долей, как правило, сохраняют спо
собность к передвижению, использованию речи, опознанию объектов и даже к
запоминанию информации. Однако ПФК играет центральную роль в формули
ровании целей и задач, затем в разработке плана действий, необходимых для
достижения этих целей. Они выделяют когнитивные процессы, необходимые
для следования плану, координирования этих действий и выполнения их в опре
деленной последовательности. И наконец, ПФК отвечает за способность оце
нивать наши действия как успех или неудачу по отношению к нашим стремле
ниям (табл. 12 .1).
Таблица 12.1
Некоторые обычные функции префронтальной коры
1. Планирование, постановка целей и инициация действия
2. Контроль результатов и адаптация к ошибкам
3. Умственное усилие для достижения трудных целей
4. Взаимодействие с другими областями для достижения целей (базальными ган
глиями, ядрами таламуса, мозжечком, моторной корой)
5. Создание мотивации, желания совершать действия
6. Инициация речи и зрительных образов
7. Признание целей других людей, участие в общественном сотрудничестве и со
ревновании
8. Регуляция эмоциональных импульсов
9. Переживание эмоций
10. Сохранение и обновление рабочей памяти
11. Активное мышление
12. Облегчает осознание произошедшего опыта
13. Непрерывное внимание при наличии постоянных дистракторов
14. Принятие решения, перемещение внимания и изменение стратегии
15. Планирование и составление последовательности выполнения действий
16. Объединение звуков, синтаксиса и значений в речь
17. Разрешение противоречий между целями
96
Глава 12
3.1.
«Память будущего»
Д. Ингвар (�. �ng�ar, 1985) ввел новый термин: «память будущего». Он обра
�ng�ar, 1985) ввел новый термин: «память будущего». Он обра
�ng�ar, 1985) ввел новый термин: «память будущего». Он обра
1985) ввел новый термин: «память будущего». Он обра
Он обра
обра
щался к одной из самых важных функций сложных организмов: составлению
планов и использованию их для направления поведения. В отличие от прими
тивных организмов, человек — активное существо, а не реактивное. Переход от
реактивного типа поведения к наиболее активному является центральной лини
ей эволюции нервной системы. Человек способен формировать цели, представ
ления о будущем. Далее мы действуем согласно нашим планам. Но для того,
чтобы направлять наше поведение по намеченному пути, эти ментальные обра
зы будущего должны стать частью нашей памяти; таким образом формируется
«память будущего».
Человеческий разум дальновиден, он, скорее, действует с опережением со
бытий, нежели противодействует им. Он руководствуется целями, планами,
надеждами, амбициями и мечтами, всем тем, что относится к будущему, а не
прошлому. Эти когнитивные процессы опираются на функционирование лоб
ных долей и развиваются совместно с ними. Лобные доли обеспечивают орга
низм способностью создавать нейронные модели как предпосылки для дости
жения успехов в действиях, модели того, чего еще на самом деле не существует,
но что вы хотите воплотить в жизнь.
Для того чтобы создавать планы на будущее, мозг должен быть способным к
переформированию предыдущего опыта так, что он не копирует прошлый или
настоящий опыт в действительности. Для достижения этого организм должен
выйти за рамки простой возможности создавать внутренние образы, модели
внешнего мира. Он должен приобрести способность манипулировать этими
моделями и изменять их. Мы можем утверждать, что изготовление инстру
ментов, одна из основных отличительных особенностей когнитивной деятель
ности приматов, зависит от обсуждаемых способностей, так как инструменты
не существуют в готовом виде в природе и должны быть придуманы, прежде
чем их можно было бы смастерить. Нейронные механизмы для формирования
и хранения «образов будущего» — необходимая предпосылка для изготовления
инструментов, а значит, и для начала человеческой цивилизации.
Можно утверждать, что способность языка формулировать новые идеи так
же опирается на эти особенности. Возможность манипулировать и изменять
внутренние образы зависит от функционирования ПФК, что, вероятно, и сдела
ло ее критической для развития речи.
3.2 .
Самовосприятие и управляющие функции
Создание цели предполагает вопрос «что мне надо?», a не «что есть». Таким
образом, способность постановки цели должна быть жестко связана с возник
новением внутренних образов самого себя. Не сюрприз, что самовосприятие
также сложным образом зависит от работы лобных долей.
Все эти функции можно назвать метакогнитивными, а не просто когнитив
ными, так как их нельзя отнести ни к какому конкретному умственному про
цессу, и они обеспечивают всеобъемлющую организацию всей когнитивной
деятельности. По этой причине некоторые авторы объединяют все функции
лобных долей под названием управляющих функций, по аналогии с государ
ственным или корпоративным устройством.
Цели, управляющий контроль и действие
97
4.0 .
Более детальное рассмотрение лобных долей
Как уже было рассмотрено выше, нейроанатомия и нейрофизиология лобных
долей с множеством сильно различающихся по клеточной и анатомической
структуре областей отражает их разнообразные и сложные функции. Еще один
уровень сложности добавляет высоко разветвленная сеть связей со многими
областями мозга. В данном разделе мы рассмотрим основные подразделения
лобных долей и выявим удивительные нейронные пути, соединяющие эти по
добласти с остальным мозгом.
4.1.
Нейроанатомия и нейрофизиология лобных долей
Границы лобных долей обычно проводят по крупным анатомическим струк
турам, а зоны Бродмана используют для разделения лобной коры на функцио
нальные области. Следующие границы на поверхности коры головного мозга
используют для выделения лобных долей: на дорсолатеральной поверхности
мозга лобная кора начинается кпереди от латеральной (или сильвиевой) бороз-
ды и центральной борозды, медиально граница проходит ориентировочно по
продолжению центральной борозды на медиодорсальную поверхность мозга, и
по воображаемой линии проекции латеральной борозды на медиовентральную
поверхность мозга (Carpen�er and Paren�, 1995).
4.2 .
Как установить границы префронтальной коры
Более четко границы лобной коры можно определить, используя карту полей
Бродмана (Brodmann, 1909). Поля Бродмана выделены на основе типов нейро
Brodmann, 1909). Поля Бродмана выделены на основе типов нейро
, 1909). Поля Бродмана выделены на основе типов нейро
нов и их связей. Согласно этому, ПФК содержит поля Бродмана с номерами 8,
9, 10, 11, 12, 13, 44, 45, 46 и 47 (�us�er, 1997) (рис. 12.3). Эти поля характеризу
�us�er, 1997) (рис. 12.3). Эти поля характеризу
, 1997) (рис. 12.3). Эти поля характеризу
ются преобладанием так называемых гранулярных клеток в основном в �V слое
(Campbell, 1905, из �us�er, 1997).
Другой метод выделения ПФК основан на подкорковых проекциях. Дор
сомедиальное ядро таламуса является, по сути, точкой конвергенции, «вер
шиной» интеграции информации, внутри специфических таламических ядер
(рис. 12 .4). Значит, ПФК определяется областью проекций от дорсомедиально
го ядра таламуса в кору головного мозга.
И наконец, ПФК иногда определяют по биохимическим проекциям. Соглас
но такому варианту, ПФК — это область, получающая проекции из мезокор
тикальной дофаминергической системы. Эти различные методы очерчивания
границ ПФК выделяют приблизительно одинаковую область.
4.3 .
Обширные проводящие пути лобных долей
ПФК соединена напрямую с каждой отдельной функциональной единицей
мозга (�au�a, 1972). Она связана как с наивысшими уровнями интеграции ин
�au�a, 1972). Она связана как с наивысшими уровнями интеграции ин
, 1972). Она связана как с наивысшими уровнями интеграции ин
формации в мозге, так и с премоторной корой, базальными ядрами и мозжеч-
ком, всеми структурами, участвующими в моторном контроле движений. ПФК
также имеет проекции с дорсомедиальным ядром таламуса, которое считают
высшим уровнем интеграции информации внутри таламуса; с гиппокампом и
медиальной височной корой, областями, критическими для памяти; с цингуляр-
ной корой, ответственной за эмоции и разрешение неоднозначностей. Кроме
того, ПФК связана с миндалиной, регулирующей большинство эмоций и соци
альное поведение, и гипоталамусом, ответственным за поддержание жизненно
98
Глава 12
важных гомеостатических функций организма. Также ПФК имеет широкие свя
зи с ядрами ствола мозга, вовлеченными в процессы бодрствования, реакции
пробуждения, общей бдительности и регуляции циклов медленного и БДГсна.
Схематическое (но далеко не исчерпывающее) изображение этой сложной си
стемы связей показано на рис. 12 .5 .
Этот уникальный набор связей ПФК с другими областями мозга делает ее
наиболее приспособленной частью мозга для координирования и интеграции
Рис. 12 .3 . Поля Бродмана (ПБ) в лобных долях. Поля, лежащие впереди моторной
коры, относят к префронтальной коре. (ПБ 4 и 6 называются моторными и премотор
ными областями.) Однако границы не являются четкими. Иногда более правильно гово
рить о постепенном переходе от «когнитивных» областей к первичной моторной коре
(ПБ 4), которая напрямую контролирует произвольную мускулатуру. (Источник: адап
тировано Бернардом Дж. Баарсом из M. �ubin, с разрешения; рисунки Shawn �u.)
Цели, управляющий контроль и действие
99
работы многих мозговых структур (рис. 12 .6). Такая обширность связей под
вергает лобную кору повышенному риску различных повреждений. Некото
рые ученые согласны с предположением, возникшим в конце X�X в. (Hughlings
Jackson, 1884), что ПФК содержит карту всей коры. Согласно этой гипотезе,
префронтальные области необходимы для нормальной когнитивной деятель
ности. Вследствие того что любой аспект нашей умственной работы может
в принципе находиться в фокусе нашего сознания, должна существовать об
ласть конвергенции всех существующих нервных структур. Это ведет к прово
кационному предположению, что эволюция сознания, наивысшего проявления
развитого мозга, шла параллельно эволюции ПФК.
Рис. 12 .4 . Границы ПФК можно определить по основным проекциям от таламических
ядер. Желтым цветом обозначены области мозга, которые относят к ПФК, так как они
получают проекции от медиодорсального ядра таламуса. Многие ученые считают, что
связь между корой и таламусом настолько тесная и близкая, что таламус можно назвать
дополнительным слоем коры. Таким образом, надо говорить о единой таламокортикаль
ной системе. (Источник: S�andring, 2005.)
100
Глава 12
5.0 .
Более детальное рассмотрение
функций лобных долей
5.1.
Функции регуляции
Как было рассмотрено ранее, в настоящее время считается, что управляющие
функции тесно связаны с работой лобной коры. Фундаментальная работа по
определению системы управляющих функций была проведена Александром
Лурией в 1966 г. (Luria, 1966). Тогда Лурия предположил существование си
Luria, 1966). Тогда Лурия предположил существование си
, 1966). Тогда Лурия предположил существование си
стемы, ответственной за контролирование стремлений, формулирование целей,
планирование действий для достижения этих целей, определение соответству
Рис. 12 .5 . Связи префронтальной коры со структурами, вовлеченными в моторную де
ятельность. Три области на латеральной поверхности коры больших полушарий (пре
фронтальная, премоторная и моторная кора) взаимосвязаны с таламусом, базальными
ядрами и мозжечком при помощи повторяющихся аксональных путей, что чрезвычайно
важно для контроля моторной активности. ЦМ — центральное медианное ядро; МД —
медиодорсальное ядро; ВП — вентральное переднее ядро; ВЛ — вентролатеральное
ядро. (Источник: �us�er, 2008.)
Цели, управляющий контроль и действие
101
ющих целям когнитивных процессов, построение последовательности актива
ции этих процессов, постепенный переход между ними и оценку результата
действия (рис. 12.7).
Следовательно, два основных типа когнитивных процессов, вовлеченных в
управляющий контроль, наиболее заметно отражены в научной литературе.
1. Способность организма направлять свое поведение при помощи внутрен
них образов (GoldmanRakic, 1987) — формулирование планов и далее по
GoldmanRakic, 1987) — формулирование планов и далее по
Rakic, 1987) — формулирование планов и далее по
Rakic, 1987) — формулирование планов и далее по
, 1987) — формулирование планов и далее по
строение поведения на их основе.
2. Способность организма не только управлять своим поведением согласно
внутренним образам, но и «переключаться», когда происходят непредвиден
ные события (Milner, 1982).
Для того чтобы эффективно справляться с такими переключениями, необхо
дима определенная особенность — гибкость мышления, способность быстро
реагировать на неожиданные события окружающей среды. Иногда это можно
назвать сменой когнитивной установки. К тому же управляющая система необ
ходима для процессов планирования и генерации новых идей (Goldberg, 2001).
Рис. 12.6 . (а) Проводящие волокна, изображенные зеленым цветом на среднесаггиталь
ном срезе мозга, направляются к префронтальной коре, а волокна синего цвета — к пре
моторной и моторной коре. (б) Продольный вид этих проводящих путей. (Источник:
Hofer and �rahm, 2006.)
102
Глава 12
Фустер (�us�er, 1997) расширил исходную теорию Лурии предположением
о том, что так называемая управляющая система может быть функционально
однородной, так как она отвечает как за внешние, так и внутренние (например,
логическое обоснование) действия. В общем функции управляющего контроля
не являются исключительно человеческим свойством. Однако особенностью
человеческих управляющих функций является предел, до которого они могут
объединять такие факторы, как время, новизна информации, сложность и воз
можная неоднозначность.
5.2 .
Социальное взросление и нравственное развитие
В то время как множество исследований по нейровизуализации и поведенче
ских работ были направлены на изучение внимания, рабочей памяти и процес
сов управляющего контроля в ПФК, стало известно, что лобные доли также
необходимы для развития социального восприятия (ключ к пониманию роли
лобных долей как «органа цивилизации»). Способность к сознательному кон
тролю над своими поступками не является врожденной, а постепенно развива
ется в течение жизни и очень важна для социального взросления.
5.2.1 . Ранний жизненный опыт и развитие орбитофронтальной коры
Аллан Шор (Allan Schore, 1999) считает, что раннее общение между матерью
и ребенком необходимо для нормального развития орбитофронтальной коры
Рис. 12 .7 . Префронтальные области коры координируют планирование и действия.
Объединенная модель формирования целей в префронтальной коре (в дорсолатераль
ной ее части) и разрешения конфликтных ситуаций в передней цингулярной коре. Па
мять участвует в воспроизведении сохраненных ранее планов действий и контекстной
информации (височная доля), а задняя теменная кора контролирует процессы простран
ственного внимания, необходимого для действий. Таламус является узлом коммутации
этих множественных функций. (Источник: Schneider and Chein, 2003.)
Цели, управляющий контроль и действие
103
в первые месяцы жизни. Например, стрессовый период в раннем возрасте ве
дет к перманентному повреждению орбитофронтальной коры, что усиливает
предрасположенность к психиатрическим заболеваниям. Это подразумевает,
что ранние социальные контакты помогают развитию мозга. Ученые уже дав
но знают, что ранняя сенсорная стимуляция активирует развитие зрительной
коры в затылочных областях, в то время как сенсорная депривация приводит к
задержке развития. Значит, возможно, что социальное общение активирует раз
витие лобной коры, таким же образом, как зрительная стимуляция ведет к раз
витию затылочных долей.
5.2.2. Развитие моральных качеств и лобная кора
Существует ли возможность того, что в развитие моральных качеств вносит
вклад лобная кора, так же как развитие зрения зависит от затылочной коры,
а речи — от височной? ПФК — ассоциативная кора лобных долей, «долей
действия». Задняя ассоциативная кора кодирует основную информацию об
окружающем мире. Она содержит таксономическую классификацию всех су
ществующих вещей и помогает относить отдельные объекты к определенным
категориям. По такой аналогии, ПФК может содержать классификацию всех
одобренных, моральных действий. Может ли так произойти, что повреждение
или недоразвитие ПФК приведет к моральной агнозии, так же как повреждение
и плохое развитие задней ассоциативной коры ведет к возникновению пред-
метной агнозии?
Сообщение Дамасио (�amasio) и его коллег дает некоторые доказательства
для данной гипотезы (Anderson et al., 1999). Дамасио исследовал молодых муж
чину и женщину, получивших повреждение лобных долей в раннем возрасте.
У обоих наблюдалось антисоциальное поведение: ложь, мелкая кража, прогу
лы. По утверждению Дамасио, эти пациенты не только не могли действовать в
рамках общественного порядка, но и не были способны оценить свои поступки
как морально недопустимые.
Орбитофронтальная кора не является единственной областью лобных до
лей, ответственной за социальное зрелое поведение. Передняя поясная кора за
нимает переднее медиальное положение в мозге и имеет тесные связи с ПФК.
Переднюю поясную кору традиционно связывают с эмоциями. Согласно Май
клу Познеру, она также оказывает влияние на социальное развитие при помощи
регуляции негативных переживаний (Posner and Ro�hbar�, 1998).
5.2.3. Возраст взросления и развитие лобных долей
Определение социальной зрелости менялось в течение всей истории обще
ства, также как и возраст «совершеннолетия». В современном западном обще
стве возраст 18 лет записан в законе как возраст социальной зрелости. С этого
времени разрешается голосовать, и человека считают ответственным за свои
поступки. В возрасте 18 лет также практически завершается развитие лобной
коры. Созревание разных структур мозга может быть измерено несколькими
способами. Одним из самых распространенных способов является измерение
миелинизации проводящих путей (Yakole� and Lecours, 1967). Лобные доли не
могут полностью выполнять свою роль до того, как все проводящие волокна к
отдаленным областям мозга будут полностью миелинизированы.
104
Глава 12
Одинаковый возраст социальной зрелости и практически полного созрева
ния лобных долей является больше чем просто совпадением. Без явных науч
ных данных, но благодаря накоплению опыта общество понимает, что только
к определенному возрасту индивидуум становится способным к адекватному
контролю над своими импульсами, стремлениями и желаниями. До этого вре
мени человек не может полностью отвечать за свои поступки ни по закону, ни с
моральной стороны. Эти способности напрямую зависят от зрелости и функци
ональной целостности лобной коры.
6.0.
Нейровизуализация управляющего мозга
В настоящее время интенсивно исследуются при помощи методик нейровизу
ализации (ПЭТ и фМРТ) два основных типа когнитивных процессов, близко
связанных с системой управляющего контроля лобных долей. Мы подвели итог
нескольких главных работ, разделив результаты на три секции: внимание и вос-
приятие, рабочая память, управляющие функции и моторный контроль. Но
сначала несколько слов предостережения об использовании методов функцио
нальной нейровизуализации для исследования сложных процессов: как мы уже
говорили, ПФК участвует во множестве когнитивных и управляющих процес
сов, таких как фокусировка внимания, сохранение на короткое время информа
ции в памяти, смена фокуса внимания при необходимости и принятие решения.
Таким образом, любая задача, использующаяся в исследованиях, обязательно
задействует эти сложные и перекрывающиеся процессы в лобных долях и дру
гих областях мозга. К тому же простое участие в экспериментах задействует
испытуемого — и его лобные доли — сложным образом (�us�er, 2008).
Для того чтобы разобраться во множестве процессов, задействованных в ре
шение любой задачи, ученым, исследующим функции лобной коры, надо очень
аккуратно разрабатывать экспериментальные модели для точного определения
вовлеченных процессов, например разделение являющихся частью практиче
ски всех заданий внимания и рабочей памяти. Далее в этом разделе мы приво
дим краткий обзор большого числа работ по нейровизуализации процессов в
префронтальной коре. Как мы уже упоминали в гл. 4, существование сходных,
взаимодополняющих сведений в разных лабораториях является ключевым под
ходом к пониманию сложных мозговых функций, так как применение различ
ных методик и групп испытуемых обеспечивает доказательство центрального
понятия или гипотезы о том, какие области мозга и сети обеспечивают управ
ляющие процессы.
6.1.
Внимание и восприятие
Представьте, что вы являетесь испытуемым в фМРТисследовании. Ваша зада
ча — смотреть на экран и, если на нем появляется изображение мужского лица,
вам необходимо нажать на одну кнопку, а при появлении женского лица — дру
гую кнопку. Легко, правда? Какие области мозга будут задействованы при вы
полнении данной задачи? Основываясь на знаниях о сенсорной системе мозга,
вы можете предположить, что будет задействована зрительная кора. Также вы
скажите, что активируется область лиц фузиформной коры (см. гл . 6). И вы
будете правы. Но существуют и другие аспекты этого задания, которые тоже,
Цели, управляющий контроль и действие
105
в свою очередь, приведут к активации мозга: фокусирование внимания на экра
не; принятие решения о том, является ли лицо мужским или женским; подго
товка к нажатию на кнопку; и собственно нажатие на кнопку (все это зависит от
функционирования лобной коры). К тому же обнаружение активации во многих
областях мозга за пределами сенсорной коры является центральным открытием
работ по нейровизуализации зрительного и слухового восприятия.
Но сначала несколько слов о внимании. В этой главе о функциях лобной
коры мы обсудим произвольное внимание, т. е. внимание, при котором мы мо
жем контролировать, на какой объект направить фокус внимания. Например,
когда вы читаете это предложение, вы фокусируете внимание на словах и их
значениях, а не на формате и размере шрифта или тиканье часов около вас. Про
извольное внимание также называют управляющим вниманием или нисходящим
вниманием ( �opdown) для того, чтобы подчеркнуть, что этот процесс находится
под нашим контролем. Другой тип внимания является непроизвольным, т. е.
автоматическим, часто стимулзависимым процессом, при котором наше вни
мание может быть «захвачено» какимлибо событием в окружающей среде.
Например, телефонный звонок, неожиданный выход солнца изза туч, аромат
кофе — все это может временно привлечь ваше внимание без сознательных
усилий с вашей стороны. Автоматические процессы внимания могут мешать
произвольным, но оба этих типа внимания являются критическими для способ
ности планировать и совершать действия независимо от того, дописываете ли
вы сочинение к утреннему уроку или отпрыгиваете в сторону при звуках сире
ны автомобиля. Взаимодействие процессов произвольного и непроизвольного
внимания является ключевой функцией лобных долей.
В ранних работах для локализации паттернов устойчивого произвольного
внимания и получения важной информации об участии в процессах произволь
ного внимания префронтальных областей использовали ПЭТ (Pardo, �ox and
Raichle, 1991). Приблизительно в то же время Майклом Познером и Стивом
Питерсоном была сделана основополагающая работа по изучению сети произ
вольного внимания, вовлеченного в решение перцептивных задач (Posner and
Pe�erson, 1990). Они выделили переднюю систему внимания, имеющую три от
, 1990). Они выделили переднюю систему внимания, имеющую три от
дельные нейронные сети для обеспечения оповещающих, ориентирующих и
управляющих (разрешающих конфликтные ситуации) функций произвольного
внимания, которые очень важны для многих процессов. За многие годы Познер
с коллегами разработали тест на сети внимания, что позволило им отдельно из
мерять и регистрировать активность областей мозга, ответственных за функци
онирование этих сетей произвольного внимания. Передняя система внимания
состоит из трех ключевых сетей: оповещающей, для поддержания состояния
напряженности, например, быть готовым рассмотреть рисунки на этой страни
це; ориентирующей, для подготовки к восприятию изображения нового лица
на экране; управляющей, для принятия решения о половой принадлежности
предъявленного лица.
Эти сети отражены на изображениях мозга (рис. 12 .8 –12 .10), при этом на
рис. 12 .8 детально отражены участки фронтопариетальной системы мозга, яв
ляющейся частью оповещающей сети. На рис. 12 .9 показана активация в те
менных областях коры, необходимых для ориентировочной сети, а на рис. 12 .10
показаны области лобной коры и других структур мозга, активирующихся для
участия в управляющей или разрешающей конфликтные ситуации сети.
106
Глава 12
Второе центральное положение, вытекающее из этих и других исследова
ний передней системы внимания, состоит в том, что уровень активации ПФК
соотносится с уровнем внимания, необходимым для осуществления задания;
при этом для задач, требующих более высокий уровень внимания, наблюдается
более сильная активация (Posner et al., 1988; Pardo et al., 1990). Передняя по
ясная извилина (ППИ), относящаяся к передней цингулярной коре, — часть
префронтальной области, обычно активирующейся при выполнении заданий,
требующих концентрации внимания (Posner et al., 1988; Raichle, 1994).
Рис. 12 .8. Результаты фМРТисследования оповещающей сети внимания. На срезах от
ражена активация таламуса оповещающей системой внимания. (Источник: �an et al.,
2005.)
Рис. 12 .9 . Результаты фМРТисследования ориентационной сети внимания. На срезах
отражена активация теменных областей мозга. (Источник: �an et al., 2005.)
Рис. 12.10 . Результаты фМРТисследования управляющей сети внимания. На срезах
отражена активация ППК. (Источник: �an et al., 2005.)
Цели, управляющий контроль и действие
107
6.2 .
Рабочая память
Способность сохранять в памяти на предельный момент времени информа
цию является центральной функцией сознания. Эта способность — рабочая
память — близко связана с системой произвольного внимания. К тому же опи
сать рабочую память можно и как систему направленного внутрь произволь
ного внимания, необходимую для контроля внутренних представлений (�us�er,
2008). Центральная проблема при определении областей мозга, ответственных
за рабочую память, — как изучать рабочую память и отделить ее от других
управляющих процессов, таких как внимание и принятие решения? Для того
чтобы решить этот вопрос, была разработана двойная парадигма, включающая
две задачи, каждая из которых в отдельности не требовала участия рабочей па
мяти (�’Esposi�o et al., 1995). Однако при решении обеих задач вместе про
исходило вовлечение рабочей памяти, таким образом, исследователям удалось
выделить процессы специфичные рабочей памяти. Они идентифицировали
области ПФК, активирующиеся вовлечением рабочей памяти, что обеспечило
ученых новой информацией о различных аспектах центральных управляющих
функций ПФК.
В своей книге «Префронтальная кора» Фустер (�us�er, 2008) представил ме
�us�er, 2008) представил ме
2008) представил ме
таанализ нескольких исследований по нейровизуализации рабочей памяти для
построения схематической модели областей мозга, активирующихся зритель
ной или вербальной рабочей памятью. Эти результаты продемонстрированы на
рис. 12 .11 –12 .13.
На рис. 12 .11 схематически изображены области мозга как видимые с лате
ральной поверхности мозга, так и спрятанные внутри, активирующиеся в экс
периментах по выявлению процессов рабочей памяти. Как видно на рис. 12.12,
Рис. 12.11. Изображение левого полушария мозга, использующееся в следующих рисун
ках для отображения зон, активированных рабочей памяти. Зоны, расположенные на по
верхности коры, отмечены белыми подписями, а расположенные на медиальной поверх
ности, — серыми. (ДМП — дополнительное моторное поле.) Внизу рисунка представлена
временная схема типичного опыта на зрительную рабочую память (РП), отсроченный вы
бор по образцу с лицами. Первая прямоугольная метка на оси времени отражает время
предъявления лица, являющегося образцом для сравнения; вторая метка — время предъ
явления двух лиц для выбора одного, подобного предыдущему. Период отсрочки между
предъявлениями двух групп стимулов длится 20 с. (Источник: �us�er, 2008.)
108
Глава 12
паттерны активации при выполнении задания на зрительную память наблю
даются в затылочных областях (так как используются зрительные стимулы) и
лобной коре (изза свойства задачи). На рис. 12 .13 представлен сходный пат
терн активации, однако в данном случае он возникает в задаче на вербальную
память, и активация происходит в височной коре при предъявлении звуковых
стимулов, а также в лобной коре.
Рис. 12 .12 . Относительная (выше исходного уровня) корковая активация в течение ше
сти временных отрезков (отмеченных желтым треугольником) выполнения задания на
зрительную рабочую память по схеме с рис. 12.11. Активация областей на латераль
ной поверхности мозга показана красным цветом, на медиальной — розовым. (1) При
предъявлении образца области активации ограничиваются зрительной и задней инфе
ротемпоральной корой; (2) в начале период задержки активация распространяется на
латеральную префронтальную кору, переднюю поясную, переднюю инферотемпораль
ную и фузиформную кору; (3) в середине задержки она сохраняется в префронталь
ной, инферотемпоральной и фузиформной коре; (4) в завершении периода задержки
активация смещается к премоторной коре, но сохраняется в инферотемпоральной и
фузиформной коре; (5) при ответе (выборе лица аналогичного образцу) активация на
блюдается в зрительной, инферотемпоральной и фузиформной коре и распространяется
на моторные области (включая фронтальное глазодвигательное поле), дополнительное
моторное поле (ДМП) и орбитофронтальную кору; (6) после завершения этой пробы,
активация задерживается в передней поясной и орбитофронтальной коре. (Источник:
�us�er, 2008.)
Цели, управляющий контроль и действие
109
Одна из гипотез участия ПФК в процессах рабочей памяти заключается в
том, что функция ПФК состоит в выборе материала или информации для реше
ния задачи, тогда как в сенсорных областях происходит активное сохранение
этих материалов или информации во время их использования при выполнении
задания (Cur�is and �’Esposi�o, 2003; Wager and Smi�h, 2003). Одновременная
активация ПФК и сенсорных областей мозга в когнитивных процессах была
Рис. 12 .13 . Корковая активация в разные моменты времени (желтые треугольники)
выполнения задачи на вербальную рабочую память. Стимулом является предъявление
слова через наушники. Активация областей на латеральной поверхности мозга показа
на оранжевым цветом, в медиальной поверхности или областей внутри борозд и изви
лин — желтым. (1) При предъявлении стимулаобразца область активации ограничива
ется слуховой корой, верхней височной извилиной и нижней лобной корой; (2) в начале
периода задержки активация распространяется на латеральную префронтальную, пе
реднюю поясную, верхнюю височную и теменную ассоциативную кору; (3) в середине
задержки она сохраняется в префронтальной и теменновисочной коре; (4) в заверше
нии периода задержки активация остается в префронтальной и теменновисочной коре
и появляется в премоторных областях; (5) при ответе (сообщение о предъявлении слова
образца на экране) активация затрагивает зрительную и теменновисочную кору, рас
пространяется на фронтальное глазодвигательное поле, дополнительное моторное поле
(ДМП), нижнюю лобную и орбитофронтальную кору; (6) после завершения этой пробы
активация задерживается в передней поясной, орбитофронтальной коре и речевых об
ластях. (Источник: �us�er, 2008.)
110
Глава 12
описана как «реверберирующий ввод» с вовлечением сетей в цикл восприятия-
действия (рис. 12 .14) (�us�er, 2008).
6.3 .
Управляющие функции и моторный контроль
Мы описали основные функции ПФК, как возможность планировать наши дей
ствия (либо мыслительные, либо физические) и придерживаться этого плана.
Мысленное планирование моторных действий от начальных абстрактных об
разов до настоящего кодирования моторных актов происходит в лобной коре.
Современный взгляд на организацию этих процессов на нейронном уровне сле
дующий: абстрактные образы/планирование действий возникают в передней
части ПФК, активность смещается назад (т. е . ближе к моторным областям) с
уменьшением абстрактности образов и началом формирования моторных ко
дов движений (�us�er, 2008). Уровень мозговой активности для планирования
и управления сложным поведением соотносится с уровнем сложности дей
ствия. Головоломку «башни Лондона» очень часто используют для исследова
ния управляющих функций лобной коры (рис. 12 .15) (Shallice, 1982). Данная
задача представляет собой головоломку с множеством шагов до успешного ее
завершения. Для того чтобы решить ее, испытуемому необходимо составить
план действий. Ученые обнаружили активацию в ДЛПФК левого полушария
при решении головоломки с более высоким уровнем ее у тех испытуемых, для
которых данная задача была сложной (Morris et al., 1993).
Однако обучение и тренировка совершенно меняет этот эффект, так как хо
рошо усвоенным, но в то же время сложным поведенческим актам соответ
ствует низкий уровень мозговой активности (рис. 12 .16) (Poldrack et al., 2005;
Posner and Raichle, 1994). Высокоавтоматизированное поведение, например
Рис. 12 .14 . Схематическое изображение мозга обезьяны, отражающее реципрокные
взаимодействия корковых областей в процессах рабочей памяти на вершине цикла вос
приятиедействие. Префронтальная кора взаимодействует при помощи ревербераций с
разными корковыми областями в зависимости от модальности предъявленного стимула
(образец или подсказка) в четырех задачах на рабочую память (указаны под соответ
ствующими картинками). (Источник: �us�er, 2008.)
Цели, управляющий контроль и действие
111
Рис. 12 .15. Головоломка башни Лондона. (Источник: �us�er, 2008.)
Рис. 12 .16 . Активация областей мозга на ПЭТизображениях при выполнении задания
по образованию глаголов. Испытуемому предъявляется серия существительных, со
ставленная экспериментатором, к каждому слову необходимо придумать подходящий
глагол. Контролем к данному заданию служит простое чтение слов с экрана монитора.
На рисунке представлены результаты вычитания изображений мозга при разных зада
чах в трех вариантах (слева направо): выполнение задания нетренированными испыту
емыми, испытуемые натренированы на одном списке слов, им предъявлена новая серия
существительных. (Источник: �us�er, 2008 из Posner and Raichle, с разрешения.)
112
Глава 12
завязывание шнурков или запирание двери, вызывает очень небольшую актива
цию ПФК. Таким образом, чувство того, что эти виды действий являются про
стыми, требующими мало усилий для их осуществления, коррелирует с низким
уровнем активности в ПФК.
Мы кратко затронули функции передней поясной коры (ППК) (поле Бродма
на 24 на рис. 12 .3). Какова роль ППК в управляющих функциях? Несмотря на
то что точная роль ППК до сих пор исследуется, существует гипотеза об ее тор
мозном влиянии на процессы лобной коры. Если это действительно так, то ППК
станет частью орбитомедиальной ПФК, которая помогает снизить эффекты от
влекающих событий на управляющие функции мозга (рис. 12 .17) (Bush et al.,
2000). Фустер (2008) высказал предположение, что данный тормозный эффект,
который помогает противостоять отвлекающим моментам, может являться об
ратной стороной процесса управляющего внимания. Таким образом, успешное
концентрирование на задание зависит как от способности фокусировать на нем
внимание, так и от возможности игнорировать отвлекающие моменты.
6.4 .
Принятие решения
В повседневной жизни мы встречаемся с двумя типами ситуаций, которые тре
буют двух разных стратегий поведения. Такие ситуации, как планирование рас
ходов и запоминания номера телефона или имени, являются детерминирован-
ными. Каждая такая проблема имеет только одно решение, соответствующее
данной ситуации, а все остальные варианты являются ложными. Осуществляя
поиск правильного решения, мы вовлечены в процесс истинного принятия ре-
шения. Ситуации выбора, что одеть, какой фильм посмотреть или по какому
карьерному пути пойти, являются неопределенными. Они не имеют, по сути,
правильного решения. Делая выбор в такой ситуации, мы осуществляем адап-
тивное (зависящее от субъекта) принятие решение.
Рис. 12.17. Разделение когнитивных и эмоциональных областей. Обобщенные резуль
таты большого числа исследований передней поясной коры, показывающие разделение
когнитивных и эмоциональных областей. (Источник: Bush et al., 2000.)
Цели, управляющий контроль и действие
113
В школе, когда нам задают задачу, мы должны найти правильное решение.
И существует, как правило, только один правильный ответ. Но, кроме выпуск
ных экзаменов, проверочных работ и викторин, основанных на фактах, боль
шинство принимаемых нами каждый день решений не имеют определенного
правильного ответа. Выбор, который мы делаем, не вытекает напрямую из
текущей ситуации. Он основан на взаимодействиях между свойствами самой
ситуации и нашими особенностями, нашими стремлениями, сомнениями и
предыдущим опытом. Префронтальная кора является центральной структурой,
обеспечивающей данной процесс. Поиск решения в детерминированных ситу
ациях часто осуществляется по алгоритму. Это все чаще поручают различным
приборам: калькуляторам, компьютерам, справочникам. Но принятие решения
в отсутствии конкретного правильного ответа, по крайней мере, до сих пор,
является исключительно человеческой прерогативой. В некотором смысле сво
бода выбора возможна только при наличие неопределенности.
Разрешение неопределенности часто требует сначала формулировки вопро
са, т. е. сведения ситуации к одному вопросу, который имеет один правильный
ответ. Как в данный момент мы разрешим существующую неоднозначность,
зависит от наших приоритетов, которые могут меняться в зависимости от ситу
ации. Невозможность разрешить неопределенность ведет к нерешительности,
колебаниям и непоследовательному поведению.
В то же время у индивидуума должна присутствовать определенная гибкость
для усвоения разных перспектив одной ситуации, но в разное время. Организм
должен быть способен разрешить неопределенности множеством разных спо
собов в зависимости от текущей ситуации и легко переключаться между ними.
Борьба с врожденной неопределенностью является одной из основных функ
ций лобных долей. Исследования пациентов с повреждениями лобных долей
показали, что к решению по определению двойственных ситуаций такие люди
подходят иначе, чем здоровые люди. Потеря способности принимать решения
является одним из самых распространенных признаков ранней деменции. По
ражение других областей мозга не влияет на рассматриваемый процесс.
Где конкретно в лобной коре происходят процессы принятия решения? Мно
жество данных указывают на участие орбитомедиальной части ПФК, особен
но когда присутствуют эмоциональные факторы в принятии решения. Пред
ставьте, что вы играете в покер. Вы должны постоянно принимать решения о
следующем ходе, имея недостаточно информации — сбросить ли карты, под
нять ли ставку, какой ход предпримут соперники? Для того чтобы преуспеть в
этой игре, необходимо постоянно оценивать факторы риска и вознаграждения.
Такой процесс был исследован при помощи игровой задачи �owa gambling �ask
(Bechara, �amasio, �ranel and Anderson, 1994). Эти и многие другие ученые по
Bechara, �amasio, �ranel and Anderson, 1994). Эти и многие другие ученые по
, �amasio, �ranel and Anderson, 1994). Эти и многие другие ученые по
�amasio, �ranel and Anderson, 1994). Эти и многие другие ученые по
, �ranel and Anderson, 1994). Эти и многие другие ученые по
�ranel and Anderson, 1994). Эти и многие другие ученые по
and Anderson, 1994). Эти и многие другие ученые по
and Anderson, 1994). Эти и многие другие ученые по
Anderson, 1994). Эти и многие другие ученые по
Anderson, 1994). Эти и многие другие ученые по
, 1994). Эти и многие другие ученые по
казали активацию орбитальной или медиальной части ПФК при выполнении
оценки рисков и вознаграждения.
Вам приходилось находиться в состоянии внутреннего конфликта при при
нятии какоголибо решения? С настоящим чувством того, что «часть вас» хочет
прислушаться к сердцу, а другая «часть вас» — к голове? Данный вид внутренних
конфликтов между эмоциями и рациональным мышлением отражает взаимодей
ствие между орбитомедиальной ПФК с ее связями и подкорковыми структурами
эмоций и латеральной ПФК, связанной с управляющими областями коры.
114
Глава 12
6.5 .
Усвоение правил
Для того чтобы направлять ежедневно наши действия в сложной жизни, необхо
димо выработать метод сокращения всех процессов планирования и действий.
Люди великолепно усваивают новые правила. Мы постоянно разрабатываем и
обучаемся стратегиям для упрощения нашей сложной жизни. Так же как и стра
тегический план или схема, правила помогают нам повысить эффективность
нашей деятельности. Очень хорошим примером умственной гибкости человека
в усвоении правил и, что очень важно, в их смене, является висконсинский тест
сортировки карточек (рис. 12 .18).
Исследования по нейровизуализации усвоения правил показали, что, как и
в процессах внимания и рабочей памяти, нейронная активность в лобной коре
увеличивается с увеличением сложности свода правил, который необходимо
усвоить и выполнить (рис. 12 .19) (Bunge, 2004).
Нейровизуализация позволила пролить свет на множество различных про
цессов, выполняемых или управляемых ПФК, от фокусировки внимания на
стимуле в окружающем пространстве до наблюдения за его изменениями, со
хранением информации в памяти и принятия сложных решений. Множество
этих процессов сильно перекрываются во времени и в областях мозга, поэтому
ученые до сих пор выясняют, какие именно части лобной коры ответственны за
эти процессы. Несмотря на то что мы находимся на ранних стадиях понимания
того, как и где в ПФК выполняются управляющие процессы, методы нейрови
зуализации начинают вырисовывать более четкую картину функционирования
ПФК.
Рис. 12 .18 . Висконсинский тест сортировки карточек (ВТСК). Карточки можно разде
лить на совпадающие по цвету изображений, по форме изображенных предметов, по их
числу и даже на группу с несовпадающими свойствами. В начале игры экспериментатор
определяет правило, по которому необходимо сортировать карточки (например, соот
носить карточки по цвету), но не сообщает его игроку. Игрок должен сам вычислить
это правило в ходе игры. Во время эксперимента ученый несколько раз меняет правила
игры, и игрок должен усваивать новые правила методом проб и ошибок. Это тест на
умственную гибкость — возможность усваивать новые правила и приспосабливаться
к новым условиям при необходимости. (Источник: �us�er, 2008.)
Цели, управляющий контроль и действие
115
7.0 .
Нарушения лобных долей
Мы обсудили многие функции лобных долей, включая произвольное внимание,
рабочую память, принятие решения и даже личностные качества. Что произой
дет при повреждении этой критической области? Или при нарушении ее разви
тия? Ответы на эти вопросы так же сложны, как и сама лобная кора.
7.1.
Хрупкие лобные доли
Дисфункции лобных долей очень часто являются результатом не только повреж
дения непосредственно их самих (Goldberg, 1992). Лобные доли как бутылоч
Goldberg, 1992). Лобные доли как бутылоч
, 1992). Лобные доли как бутылоч
Рис. 12 .19 . В данном исследовании ученые стремились выявить области мозга, ак
тивирующиеся при выполнении испытуемыми простых или сложных абстрактных
«правил». За ключевым периодом предъявления правила следовал длинный период за
держки (7–15 с). Области мозга, активирующиеся в большей степени при следовании
сложным правилам, нежели простым, показаны на верхнем рисунке. Активность обла
стей мозга в период задержки (когда испытуемый держит правило в голове) показана на
среднем (простые правила) и нижнем (сложные правила) рисунках. (Источник: Bunge,
2004, адаптировано �us�er, 2008.)
116
Глава 12
ное горлышко, как точка схождения эффектов практически любого поврежде
ния мозга. Существуют реципрокные взаимоотношения между повреждениями
лобной коры и других структур мозга. Повреждения лобной коры вызывают
широкую неравномерную волну эффектов на весь мозг. В то же время повреж
дения других структур мозга оказывают сильный эффект на функционирование
лобных долей. Причиной этого является уникальное свойство лобных долей
работать как «узел коммутации» в необыкновенно обширной системе мозговых
связей. Это делает дисфункции лобных долей наиболее распространенными, но
и наименее специфичными среди других неврологических, психиатрических
заболеваний и нарушений развития (Goldberg, 1992).
Чрезвычайно низкий порог функциональных нарушений лобных долей со
гласуется с предположением об «эволюции и деградации» (Hughlings Jackson,
1884). Согласно этой гипотезе, филогенетически (эволюционно) более моло
дые структуры мозга первыми подвергаются нарушениям. Уникальная уязви
мость лобных долей является обратной стороной богатства их связей с другими
структурами. Дисфункции лобных долей не всегда оказываются результатом
прямого их разрушения. Вместо этого, причиной могут являться рассеянные,
распространенные или отдаленные повреждения мозга.
7.2 .
Синдромы повреждения лобных долей
Всю важность управляющих функций можно понять, анализируя их наруше
ния вследствие мозговой травмы. Пациенты с повреждением лобных долей со
храняют, по крайней мере в определенной степени, способность развивать по
отдельности большинство когнитивных навыков (Luria, 1966). Основные спо
Luria, 1966). Основные спо
, 1966). Основные спо
собности, такие как чтение, письмо, простые вычисления, словесное общение
и двигательная активность, остаются, как правило, не затронутыми. Такой па
Рис. 12 .20 . Орбитофронтальная область прилегает сверху к орбитам глаз. Орбитофрон
тальная кора (зеленые полосы) может быть выделена на вентральной стороне лобных
долей. Она участвует в процессах выявления будущих наград, изменения вероятности
вознаграждения и выбора цели. Повреждение орбитофронтальной области может при
вести к потере контроля над поведением. (Источник: �a�idson and �rwin, 1999.)
Цели, управляющий контроль и действие
117
циент будет хорошо справляться с психологическими заданиями в отдельности
на каждый процесс. Однако любая комплексная активность, требующая коор
динации множества когнитивных навыков в последовательный, направленный
на достижение цели процесс, будет чрезвычайно сильно нарушена.
Повреждения разных участков лобной коры ведут к отдельным клиниче
ским синдромам. Наиболее распространены дорсолатеральный и орбитофрон
тальный префронтальные синдромы (рис. 12 .20) (Goldberg and Cos�a, 1985).
7.2.1 . Дорсолатеральный префронтальный синдром
Наиболее часто встречающиеся симптомы дорсолатерального префронтально
го синдрома — изменение личности, поведение, зависящее от окружения, и
персеверация поведения.
Изменение личности
По клиническим проявлениям пациент с дорсолатеральным лобным синдро
мом характеризуется сниженной способностью инициировать поведенческие
акты. У пациента происходит «уплощение эмоций», т. е. наблюдается моно
тонная речь и чувство равнодушия. Такой человек не грустит и не радуется,
как будто у него нет какоголибо настроения. Такое состояние равнодушия со
храняется независимо от происходящих хороших или плохих событий. Однако
наиболее заметный симптом у пациента с дорсолатеральным синдромом — су
щественное нарушение способности инициировать действия. Один раз начав
любую деятельность, такой пациент так же неспособен закончить или изменить
ее самостоятельно. Комбинация «инертности инициации и завершения» наблю
дается при различных нарушениях, затрагивающих лобную кору, в том числе
при хронической шизофрении.
Вследствие даже относительно легкой черепномозговой травмы пациенты
часто становятся равнодушными и лишаются инициативы и стремлений. Дан
ные нарушения могут быть незаметными, и членам семьи или врачам не всегда
ясно, что эти изменения имеют неврологическую природу и являются резуль
татом легкой формы синдрома лобных долей. Эти симптомы часто объединя
ют термином изменение личности, но само понятие личность имеет широкий
нейробиологический контекст. Лобная кора оказывает максимальное влияние,
по сравнению с любыми другими структурами мозга, на нашу личность, и по
вреждение лобных долей может вызвать абсолютное изменение личности.
Поведение, зависящее от окружения
Другим распространенным симптомом дорсолатерального префронтального
синдрома является нахождение пациента под властью разных отвлекающих со
бытий, а значит, неспособного следовать внутренним планам. Крайняя форма
может принимать вид поведения, напрямую зависящего от окружения. Такой
пациент будет пить из пустой чашки, наденет чужую куртку или будет рисо
вать на столе карандашом только потому, что чашка, куртка или карандаш на
ходились в его поле зрения независимо от того, что данные действия не имеют
смысла. Данный феномен интенсивно исследовался французским неврологом
Франсуа Лермиттом, который и придумал новый термин — поведение исполь-
зования (Lhermi��e, 1983). Наиболее крайнее выражение поведения, зависящего
от окружения, принимает форму прямого подражания по названием эхолалия
(имитация речи) или эхопраксия (имитация действий).
118
Глава 12
Такие пациенты покажут плохие результаты (S�uss et al., 2001) в тесте
Струпа (S�roop, 1935). Чем интересен тест Струпа? Он требует от нас противо
S�roop, 1935). Чем интересен тест Струпа? Он требует от нас противо
, 1935). Чем интересен тест Струпа? Он требует от нас противо
стояния немедленным импульсам. Стремление прочитать слова является есте
ственным свойством любого образованного человека при виде написанного
материала. Но задача теста заключается в назывании цветов. Для того чтобы
справиться с заданием, нам надо следовать внутреннему плану, идущему враз-
рез с естественным, устоявшимся стремлением (см. также гл. 2 и 8).
Когда неврологические заболевания затрагивают лобные доли, способность
следовать определенным курсом теряется и пациенты оказываются во власти
неожиданных стимулов окружающей среды и отклоняющих внутренних об
разов. Способность легко отвлекаться — свойство многих неврологических и
психиатрических заболеваний и, как правило, результат повреждения в лобной
коре. Например, дефицит внимания/гиперактивность, характеризующийся вы
соким уровнем рассеянности, связывают с нарушениями работы лобных долей
(Barkle�, 1997). Роль лобной коры в направлении поведения на основе внутрен
Barkle�, 1997). Роль лобной коры в направлении поведения на основе внутрен
, 1997). Роль лобной коры в направлении поведения на основе внутрен
них образов при выполнении теста Струпа, при эмоционально значащем зада
нии и у пациентов с дефицитом внимания/гиперактивностью была выявлена в
многочисленных работах (см. рис. 12 .17).
Умственная ригидность и персеверации
Наша способность поддерживать умственную стабильность должна находить
ся в равновесии с умственной подвижностью. Независимо от того, как сильно
мы сфокусированы на действии или мышлении, наступает такой момент, ког
да ситуация требует радикально сменить деятельность. Возможность сменять
мысленную установку также важна, как и сохранение прежнего образа мысли.
Свойство переключаться с одной активности или идеи на другую настолько
естественно и автоматично, что мы принимаем его как должное. В действи
тельности же оно требует сложного нервного аппарата, основанного на работе
лобных долей. Умственная подвижность или способность видеть вещи в новом
свете, креативность и оригинальность мысли — все это зависит от функциони
рования лобной коры. Более глубокие формы умственной ригидности приводят
к обсессивно-компулятивному расстройству (ОКР), при котором дисфункция
хвостатого ядра влияет на связанную с ним лобную кору (Rauch et al., 1994).
Повреждения лобных долей очень часто приводят к крайней степени ум
ственной ригидности, которая значительно расшатывает когнитивную дея
тельность пациента. Часто более детальное рассмотрение выполнения таким
пациентом нескольких заданий показывает, что полное переключение с одной
задачи на другую невозможно, и фрагменты выполнения предыдущего задания
накладываются на новое. Этот феномен носит название персеверации.
Существует простой нейропсихологический тест, чувствительный даже
к незначительному нарушению умственной подвижности. Это Висконсинский
тест сортировки карточек (Gran� and Berg, 1948), для выполнения которого ис
Gran� and Berg, 1948), для выполнения которого ис
and Berg, 1948), для выполнения которого ис
and Berg, 1948), для выполнения которого ис
Berg, 1948), для выполнения которого ис
Berg, 1948), для выполнения которого ис
, 1948), для выполнения которого ис
пытуемому необходимо рассортировать карточки с изображением простых
геометрических фигур по трем разным категориям согласно одному простому
правилу (см. рис. 12.18). Принцип классификации изначально неизвестен ис
пытуемому, и его необходимо вычислить методом проб и ошибок. Как только
испытуемый овладевает правилом, оно внезапно, без предупреждения меняет
ся снова и снова. Это задание требует создания плана, направления действий
Цели, управляющий контроль и действие
119
с помощью внутренних образов, умственной подвижности и рабочей памяти,
т. е . всех аспектов функционирования лобной доли.
7.2.2. Орбитофронтальный префронтальный синдром и самоконтроль
Орбитофронтальный префронтальный синдром по многим показателям про
тивоположен дорсолатеральному синдрому. Такие пациенты поведенчески и
эмоционально расторможены. Их поведение редко спокойное, настроение ме
няется от эйфории до ярости, а импульсный контроль над действиями слаб или
несущественен. Их способность подавлять порыв к получению немедленного
вознаграждения сильно повреждена. Они делают, что хотят, когда хотят, без ка
койлибо заботы о соблюдении социальных норм и законодательных актов. Они
неспособны предвидеть последствия своих поступков. Пациенты, страдающие
орбитофронтальным синдромом (вследствие мозговой травмы, цереброваску
лярного заболевания или деменции), могут осуществлять кражи, вести себя
сексуально агрессивно, безрассудно водить машину и осуществлять другое
антисоциальное поведение. Таких пациентов характеризуют как эгоистичных,
хвастливых, инфантильных, вульгарных и сексуально возбудимых. Юмор у
таких людей непристойный, их шутки, называемые Witzelsucht (неадекватная
веселость), больше характерны пьяным подросткам (Oppenheim, 1889). И если
пациенты с дорсолатеральным синдромом характеризуются отсутствием лич
ности, то орбитофронтальные пациенты выделяются «незрелыми» личностны
ми качествами.
7.2.3 . Синдром ретикулофронтального рассогласования
В том случае, когда лобные доли сами по себе не повреждены, но у пациента
присутствует лобная симптоматика, проблема может быть в проводящих путях,
соединяющих лобную кору с другими структурами мозга. Повреждение этих
связей может привести к развитию синдрома ретикулофронтального рассогла-
сования (Goldberg et al., 1989).
В стволе мозга расположены ядра, ответственные за бодрствование и ак
тивацию остального мозга. Существует сложная система взаимосвязей между
лобной корой и ретикулярными ядрами ствола мозга, отвечающими за актива
цию и бодрствование. Эту систему можно описать как петлю. С одной стороны,
активация лобной коры зависит от восходящих проекций. С другой стороны,
существуют проводящие пути из лобной коры к ретикулярным ядрам вентраль
ного ствола мозга. Через эти проекции лобные доли контролируют различные
структуры мозга, модулируя уровень их активации. Если лобные доли являются
устройством принятия решения, то мозговой ствол можно назвать усилителем,
помогающим провести эту информацию к остальным структурам мозга. Нис
ходящие проекции — пути, по которым инструкции от лобных долей попадают
к вентральным ядрам ствола мозга.
Легко предположить как повреждение связей лобной коры с мозговым ство
лом может привести к нарушению управляющих функций без непосредствен
ного повреждения лобных долей.
7.3 .
Повреждение лобных долей и асоциальное поведение
Взаимосвязь между повреждением лобной коры и асоциальным поведением
сложна и вызывает немалый интерес. Было высказано предположение, осно
120
Глава 12
ванное на нескольких работах, что процент мозговых травм выше среди пре
ступников, чем в целом по популяции, а также при сравнении агрессивных и не
агрессивных преступников (Vola�ka et al., 1995; Raine et al., 1997). По причине
мозговой анатомии и строения черепа, закрытая мозговая травма с большей ве
роятностью ведет к повреждению лобных долей, особенно орбитофронтальной
коры. Более того, повреждения верхней части мозгового ствола также очень
часто встречаются при закрытых мозговых травмах, даже в легких случаях, и с
большой вероятностью они могут вызвать дисфункцию лобных долей в отсут
ствии прямого их повреждения, вызывая «синдром ретикулофронтального рас
согласования» (Goldberg et al., 1989).
Адриан Райне с коллегами (Raine et al., 1997) исследовали мозг осужден
ных убийц при помощи ПЭТсканирования и обнаружили аномалии в ПФК.
В 2000 г. также изучали мозг мужчин, страдающих антисоциальным расстрой
2000 г. также изучали мозг мужчин, страдающих антисоциальным расстрой
ством личности и обнаружили 11%е снижение доли серого вещества в лобной
коре этих пациентов. Причина снижения количества серого вещества не ясна,
но Райне считает, что оно является, по крайней мере, частично врожденным,
и вызванным факторами окружающей среди, например плохим обращением и
воспитанием. Связь между дисфункцией лобной коры и асоциальным поведе
нием поднимает важные социальные, моральные и законодательные вопросы,
лежащие далеко за пределами этой главы.
7.4 .
Другие клинические повреждения,
связанные с повреждением лобных долей
ПФК вовлечена в широкий круг нарушений (Goldberg, 1992; Goldberg and Bou
Goldberg, 1992; Goldberg and Bou
, 1992; Goldberg and Bou
Goldberg and Bou
and Bou
and Bou
Bou
Bou
gako�, 2000) и, к тому же, необязательно наличие прямого повреждения лоб
, 2000) и, к тому же, необязательно наличие прямого повреждения лоб
ных долей для возникновения лобной симптоматики. Лобная кора чрезвычайно
чувствительна к большому числу других нарушений. Такие заболевания как
шизофрения (�ng�ar and �ran�en, 1974; �ran�en and �ng�ar, 1975), травматиче
�ng�ar and �ran�en, 1974; �ran�en and �ng�ar, 1975), травматиче
and �ran�en, 1974; �ran�en and �ng�ar, 1975), травматиче
and �ran�en, 1974; �ran�en and �ng�ar, 1975), травматиче
�ran�en, 1974; �ran�en and �ng�ar, 1975), травматиче
�ran�en, 1974; �ran�en and �ng�ar, 1975), травматиче
, 1974; �ran�en and �ng�ar, 1975), травматиче
�ran�en and �ng�ar, 1975), травматиче
and �ng�ar, 1975), травматиче
and �ng�ar, 1975), травматиче
�ng�ar, 1975), травматиче
�ng�ar, 1975), травматиче
, 1975), травматиче
ское повреждение мозга (ТПМ) (�eu�sch and Eisenberg, 1987), синдром Туретта
(�oure��e, 1885; Shapiro and Shapiro, 1974; Sacks, 1992) и дефицит внимания/
гиперактивность (Barkle�, 1997) включают в себя нарушения функционирова
Barkle�, 1997) включают в себя нарушения функционирова
, 1997) включают в себя нарушения функционирова
ния лобной коры. Например, при ТПМ дисфункция лобных долей возникает
либо вследствие прямого их повреждения, либо разрушения корковостволо
вых связей (Goldberg et al., 1989). Управляющие функции также страдают при
деменции и депрессии.
Для наличия лобной симптоматики совершенно не обязательно наличие
прямого повреждения лобных долей. Существует много условий, при которых
(основываясь на данных функциональной нейровизуализации и нейропсихо
логических исследованиях) присутствуют дисфункции лобной коры, но отсут
ствует информация об их структурных повреждениях.
Дефицит внимания и гиперактивность
Префронтальная кора и ее связи с вентральным стволом мозга играют очень
важную роль в механизмах внимания. Когда речь идет о дефиците внимания
и гиперактивности, мы тоже обычно затрагиваем эти системы. Истинные при
чины их повреждения различны. Они могут быть наследственными или воз
никать в раннем возрасте, могут быть биохимическими или структурными.
Как мы уже знаем, нарушение функционирования лобных долей наблюдается
Цели, управляющий контроль и действие
121
в большом числе заболеваний, изза высокой вероятности возникновения по
вреждений в них. Диагноз дефицит внимания и гиперактивность обычно ста
вят в случае легкой дисфункции лобных долей или их связей при отсутствии
других, сравнительно серьезных нарушений. Вследствие высокой вероятности
дисфункции лобных долей по разным причинам, предрасположенность к дефи
циту внимания и гиперактивности тоже должна быть высока.
Для того чтобы лучше понять вероятный механизм дефицита внимания/
гиперактивности, нам необходимо лучше понять механизм еще одного вида
управляющих функций — селективного внимания. Цель действия должна быть
определена, и она должна эффективно направлять поведение в течение опреде
ленного времени. Мы уже знаем, что формирование целей и следование им обе
спечивается префронтальной корой. Префронтальная кора оказывает влияние
на другие области коры, вовлеченные в обработку поступающей информации.
В зависимости от имеющейся цели различные части задней коры должны на
ходиться в состоянии оптимальной активности. Выбор этих областей осущест
вляется префронтальной корой. Префронтальная кора оказывает это влияние
посредством ядер вентрального ствола мозга, которые селективно активируют
обширную область коры через восходящие проекции. Префронтальная кора
контролирует это активирующее влияние на заднюю кору через свои нисхо
дящие проекции к вентральной части ствола мозга. Наконец, префронтальная
кора модифицирует свои контролирующие импульсы на ядра ствола в зависи
мости от полученного ответа из задней коры.
В итоге внимание необходимо рассматривать как циклический процесс,
включающий множество переключений между префронтальной корой, вен-
тральным мозговым стволом (и, вероятно, неспецифическими ядрами средней
линии таламуса) и задней частью коры больших полушарий (рис. 12 .21). Раз
рыв цепи в любой точке цикла нарушает внимание, таким образом создавая
одну из форм синдрома дефицита внимания. Следовательно, любое повреж
дение префронтальной коры или ее проекций может привести к нарушениям
внимания.
Рис. 12 .21 . Цикл внимания, объединяющий лобную кору, мозговой ствол и заднюю
кору. Голдберг (2001) высказал предположение, что функции внимания управляются
не только лобными долями, но и через причинноследственную петлю, соединяющую
лобную кору с ретикулярной формацией ствола мозга и задними областями коры. Эта
гипотеза согласуется с данными об активирующей системе ствола мозга. (Источник:
Elkhonon Goldberg, с разрешения.)
122
Глава 12
8.0 .
Современный взгляд на организационные принципы
лобных долей
Выявлены некоторые принципы организации функций лобной коры после де
сятилетий исследований на приматах и людях с использованием множества экс
периментальных моделей и методов. Лидером в области исследований функ
ций лобных долей является Фустер, предложивший модель функционирования
лобной коры. Вот ее положения.
1) Вся корковая поверхность лобных долей направлена на представление и
производство действий любого уровня биологической сложности.
2) Нейронный субстрат для формирования любого действия идентичен суб
страту для его представления.
3) Этот субстрат имеет иерархическую организацию с наиболее простыми
действиями на нижних ступенях иерархии, в орбитофронтальной и моторной
коре, и наиболее сложными и абстрактными — в латеральной префронтальной
коре.
4) Функции лобных долей также имеют иерархическую организацию, в ко
торой простые функции способствуют выполнению более значимых. (�us�er,
2008).
Положение об иерархической организации функций лобных долей, от
простых к сложным, от абстрактных к конкретным, было разработано благо
даря исследованиям с нейровизуализацией процессов в лобной коре у чело
века. В одном из обзоров (Badre, 2008) приведено обобщение современных ре
Badre, 2008) приведено обобщение современных ре
2008) приведено обобщение современных ре
зультатов, включая модели, разработанные Кёхлином, Д’Эспосито (Koechlin,
�’Esposi�o)и другими, для подтверждения иерархической организации лобных
долей в переднезаднем направлении, основанной на уровне абстрагирования
(рис. 12 .22).
Выводы нейровизуализации
Несмотря на то что для исчерпывающего объяснения функций лобных долей
многие детали необходимо более тщательно изучить, современные технологии
и методы нейровизуализации обогатили наше знание о сложной роли лобных
долей, и особенно ПФК, в когнитивной деятельности человека. Но, несмо
тря на то, что исследования по нейровизуализации направлены к выявлению
функций в отдельных областях мозга, например произвольного внимания или
рабочей памяти, они дают совершенно иную картину управляющих функций
человека, когда работы проводят на пациентах с повреждениями лобных долей.
Как результаты исследований по нейровизуализации соотносятся с данными,
полученными на пациентах с лобным синдромом? Эти две группы данных не
так просто свести воедино. Работы по нейровизуализации обеспечили ученых
большим объемом информации об отдельных областях мозга и локализации
регионов, активирующихся при разных аспектах функционирования лобных
долей, однако это результаты не соотносятся с данными о поведении людей,
страдающих повреждением лобных долей. Это вызов для когнитивных ней
робиологов продолжить поиск единых данных в разных исследовательских
группах и при использовании множества методов для выявления основопола
гающих организационных принципов сложной и исключительно человеческой
ПФК, «органа цивилизации».
Цели, управляющий контроль и действие
123
9.0.
К единой теории управляющего контроля:
заключение
В заключение надо сказать, что после многолетнего пренебрежения к этой
теме, управляющие функции стали центром все увеличивающегося количества
исследовательских работ.
В результате ранних нейровизуализационных работ по исследованию функ
ций лобных долей образовалась сложная сеть областей мозга, специализиру
ющихся на сенсорных модальностях, речевых и неречевых процессах, объ
екториентированных и пространствоориентированных, а также «где» и «что»
путей. Этот подход, призванный пролить свет на скрытые элементы, для ПФК
выдал большое число данных, но в итоге не создал четкой картины организаци
онных принципов ПФК, к тому же полученные результаты плохо соотносятся с
большим объемом информации о синдромах повреждения лобных долей.
Рис. 12 .22 . Результаты экспериментов по реагированию, определению свойств, про
странственных и контекстных экспериментов и сравнение их с моделью Кёхлина
(Koechlin et al., 2003). Иерархия относительно абстрактности вносит маленький вклад в
рострокаудальный градиент в модели Кёхлина с соавторами (2003) и Бадре, Д’Эспосито
(2007). Сферы диаметром в 8 мм (в пределах фокуса каждого эксперимента) были цен
трированы относительно максимального ответа на манипуляции Бадре, Д’Эспосито
(2007) над реакциями (i), свойствами (iii), пространством (�) и контекстом (�i) (красные)
и сенсорные (ii), контекстные (i�) и эпизодические (�i) манипуляции Кёхлина с коллега
ми (2003) (синие). Эти сферы размещены на объемной поверхности �alairach. Обратите
внимание, что сферы точно отображают только взаимное расположение, а не реальные
размеры областей активации в данных экспериментах. Пунктирные линии разделяют
манипуляции, проводимые на одном уровне абстракции в репрезентативной иерархии.
Эквивалентные эпизодические и контекстные контрольные манипуляции двух экспери
ментов также отображены на схеме. (Источник: Badre, 2008, адаптировано из Badre and
�’Esposi�o, 2007, с разрешения.)
124
Глава 12
Совершенно другой подход берет начало от работ русского нейропсихолога
Александра Лурия (1966). Продолжение и расширение исходной теории Лурия
можно проследить в современной тенденции к опровержению точки зрения о
четкой локализации функций мозга и склонению к предположению о распре
деленном принципе функциональной организации коры (Goldberg et al., 1989;
Goldberg, 1992; �us�er, 2003). Согласно градиентной теории Голдберга, функци
, 1992; �us�er, 2003). Согласно градиентной теории Голдберга, функци
�us�er, 2003). Согласно градиентной теории Голдберга, функци
, 2003). Согласно градиентной теории Голдберга, функци
ональная организация гетеромодальной ассоциативной коры (например, пре
фронтальной коры) является динамичной и последовательной. Полимодальная
ассоциативная кора развивается в непрерывной последовательности. При такой
последовательности (градиенте) функционально близкие аспекты когнитивной
деятельности расположены в анатомически близких областях ассоциативного
неокортекса.
Существует еще одна теория, выдвинутая Фустером (2003) как продолжение
теории функциональных систем Лурии. Фустер соглашается с тем, что когни
тивные функции не имеют отдельного коркового представительства. В своей
теории он вводит единицу когнит (cogni�), которая по определению представ
cogni�), которая по определению представ
), которая по определению представ
ляет любое представительство знаний в коре больших полушарий. Когниты
являют собой динамические структуры, которые на нейронном уровне прибли
зительно совпадают с нейронными группами и их связями. Согласно Фустеру,
когнитивные функции являются результатом обмена информацией как внутри,
так и между когнитами, и разные когнитивные функции получаются из мно
жества перекрывающихся когнитов. Ключевое положение теории Фустера со
стоит в том, что разные когниты (нейронные сети) имеют идентифицируемое
корковое распределение, но когнитивные функции, которые основаны на этих
сетях, не имеют точной локализации, так как разные функции могут опираться
на одни и те же нейронные сети.
Согласно этим теориям, корковые представительства управляющего контро
ля имеют как распределенную, так и локализационную природу. Можно ска
зать, что управляющий контроль отвечает за действия как внутренние, так и
внешние и служит для объединения таких факторов, как время, новизна, слож
ность и вероятная неоднозначность.
10.0 . Задания и упражнения к главе 12
1. Назовите несколько функций, приписываемых лобным долям.
2. Обсудите современный взгляд на специфичность префронтальных функций.
3. На схематическом изображении мозга на рис. 12 .23:
а) раскрасьте основные доли мозга;
б) отметьте главные ориентиры, такие как центральная борозда и Сильвиева
борозда;
в) назовите основные извилины (зеленые линии) и борозды (пунктирные
красные линии).
Цели, управляющий контроль и действие
125
Рис. 12 .23
Для Чарльза Дарвина было очевидно безо всяких доказательств, что жи-
вотные наделены разумом: «... низшие животные, как и человек, — писал он
в 1871 г. — демонстрируют чувства удовольствия и боли, радости и страда-
ния». «Тот факт, что низшие животные испытывают те же эмоции, что и
мы, настолько очевиден, что не стоит утомлять читателя перечнем этих
деталей». (Однако) ...веря, что для движения вперед прежде всего необходима
четкая формулировка проблем и открытие путей для их решения, я для начала
хотела бы задать вот какой вопрос: отчего эта «способность переживать
эмоции», о которой с такой легкостью, обсуждая умственные способности
животных, говорит Дарвин, все еще остается фундаментальной проблемой
биологии?
Мариан Стемп Докинс (2006)
Основа «эмоционального мозга» у всех млекопитающих (а, возможно, и не только у
них) устроена весьма консервативно. Вверху слева: мозг человека, каким он видится
от средней линии (медиальный срез); показано, как информация распространяется от
зрительной коры (поле V1) к миндалине, эмоциональному «узлу связи» нейронального
потока, поступающего к ней от самых разных источников. Например, змеи вызывают
чувство безотчетного страха у человека и других млекопитающих. Наоборот, птенцы
и детеныши млекопитающих вызывают ощущение мира и покоя (��������, 1���). Оче�
��������, 1���). Оче�
, 1���). Оче�
видно, что гигантский неокортекс, характерный для человека, оказывает мощное влия�
ние на работу эмоциональных систем, собранных в лимбической области (сердцевине
мозга), и свойственных всем млекопитающим.
Содержание
1.0.Введение.......................................................127
1.1.Триединыймозг..............................................128
1.2.Базовыеэмоцииирольрефлексивногосознания...................130
2.0.ЭмоциональныесистемымозгапоПанксеппу.........................130
2.1.Переживаниеэмоций..........................................134
3.0.Системастраха..................................................134
3.1 . Сознательное и бессознательное восприятие страха:
«нижний»и«верхний»путипоЛеду.............................138
3.2.Неосознаваемыйстрах.........................................139
3.3.Аффективноеслепозрение.....................................142
3.4 . Взаимодействие познавательных и эмоциональных функций
приреакцияхстраха...........................................143
3.5.Имплицитноеэмоциональноеобучениеипамять..................144
3.6.Эмоциональнаямодуляцияэксплицитнойпамяти.................. 144
3.7.Влияниеэмоцийнавосприятиеивнимание.......................145
3.8.Эмоцииисоциальноеповедение................................147
3.9.Торможениеирегуляцияэмоций................................147
4.0 . Система положительного подкрепления: любить, хотеть, учиться . . . . . . . . . 151
4.1 . Новое понимание «подкрепления»: от подкрепления к предсказанию
подкрепления и далее — к ошибкам предсказания подкрепления . . . . . 152
4.2.Подкрепление—этобольшечемобучение.......................154
4.3.«Путьподкрепления»инаркотики...............................157
4.4.Сигналыподкреплениявлияютнавнимание......................158
5.0.Заключение.....................................................159
6.0.Заданияиупражнениякглаве13...................................159
6.1.Контрольныезадания..........................................159
6.2.Графическиеупражнения......................................160
1.0 . Введение
Помните, гд е вы были утром 11 сентября 2001 г.? Скорее всего, вы помните этот
день, и в то же время вам будет трудно вспомнить, скажем, 9 сентября того же
года. Почему так? События 11 сентября глубоко врезались в память из-за силь-
ных эмоций, с которыми они были связаны. В предыдущих главах мы обсудили
такие высшие когнитивные функции мозга, как память, управляющие функции и
речь. Наши эмоции прочно сцеплены с системами мозга, которые обеспечивают
эти когнитивные функции. В этой главе мы рассмотрим эмоциональные системы
мозга и их взаимодействие с когнитивными процессами. В конце 1990-х гг. для
описания новой области, изучающей нейробиологию эмоций, был введен новый
Глава 13
Эмоции
128
Глава 13
термин «аффективная нейронаука». Хотя к настоящему времени об управлении
эмоций мозгом известно уже достаточно много, мы сфокусируемся на том, как
системы эмоций формируют когнитивные процессы и влияют на них.
1.1 . Триединый мозг
Пол Маклин в 1�60�х гг. ввел концепцию «триединого мозга», чтобы описать
три функционально различных слоя («этажа») мозга млекопитающих. Она
легла в основу сложившегося позже восприятия общей функциональной орга�
низации мозга. И хотя некоторые аспекты теории триединого мозга довольно
противоречивы, все равно полезно мысленно делить мозг млекопитающих на
три «слоя». С точки зрения Маклина, в ходе эволюции позвоночных мозг раз�
вился в трехслойный орган, в котором слои, несмотря на тесную взаимосвязь,
сохраняют некоторую разделенность в силу различий своего эволюционного
происхождения. Хорошо видно влияние каждого из слоев на поведение челове�
ка, особенно когда речь идет о познании и эмоциях.
Самый старый слой мозга называют рептильным мозгом. Он образован
стволом мозга (продолговатый мозг, мост, мозжечок, средний мозг, бледный
шар и обонятельные луковицы) — структурами, преобладающими в мозге змей
и ящериц. Этот слой мозга слабо задействован в обучении на индивидуальном
опыте и, скорее, вновь и вновь способствует воспроизведению фиксированных
инстинктивных поведенческих программ. У человека эта часть мозга отвечает
за жизненно важные функции, такие как дыхание, регуляция сердечного ритма,
поддержание гомеостатического равновесия. В этой главе мы не будем уделять
особого внимания рептильному мозгу.
Мозг млекопитающих расположен поверх рептильного мозга (рис. 13.1).
В основном он состоит из частей, формирующих лимбическую систему. Сло�
во «лимбический» происходит от латинского limbus и означает «граница, край,
кромка». Термин относится к области внутри больших полушарий мозга,
Рис. 13 .1 . Триединый мозг: оранжевым обозначен неокортекс, зеленым — мозг млеко�
питающих, желтым — рептильный мозг
Эмоции
129
примыкающей к стенке боковых желудочков (заполненных жидкостью полостей).
Лимбическая система была впервые описана в конце XIX в., но понимание ее
роли в эмоциях пришло только после работы нейроанатома Пейпеца, опублико�
ванной в 1�37 г. Лимбическую систему раньше называли также кругом Пейпеца.
Перечень составных частей лимбической системы зависит от того, к какому
автору мы обратимся. До сих пор не существует общепринятого мнения о том,
что же входит в состав лимбической системы! Некоторые нейробиологи счита�
ют, что вообще больше не следует говорить о лимбической системе. Но все же
полезно использовать этот термин, чтобы охватить систему связанных между
собой подкорковых структур, поддерживающих наши эмоции. Обычно к лим�
бической системе относят миндалину, гиппокамп, парагиппокампальную кору,
поясную извилину, гипоталамус и вентральный стриатум/прилежащее ядро.
Лимбическая система играет ключевую роль в формировании эмоций у че�
ловека. Влияние лимбической системы на наш сознательный опыт проявляется
в виде некоторой дополнительной значимости (положительной или отрица�
тельной оценке или ощущении) и заметности («выделяемости») определенных
мыслей и образов. Эта часть мозга есть у всех млекопитающих. Типичные для
млекопитающих эмоциональные реакции легко распознаются у домашних со�
бак и кошек.
Что же касается адаптивной роли лимбической системы, то можно отметить,
что она содержит в себе несколько подсистем, сформировавшихся под давлени�
ем таких факторов эволюции млекопитающих, как реагирование на опасность,
репродуктивные потребности, забота о потомстве и пище. Например, минда�
лина и гипоталамус взаимодействуют между собой, образуя систему раннего
предупреждения об опасности. Эта система автоматически запускает направ�
ленное на выживание поведение при столкновении со стимулами, похожими
на те, с которыми животное уже сталкивалось в опасных ситуациях. Однако
обработка стимулов миндалиной имеет куда меньшую точность к деталям по
сравнению с сенсорными зонами коры мозга. Узнавание стимулов лимбической
системой работает по принципу «похожести», и в результате, скажем, человек
порой вначале шарахается от каких�то неясных темных витков в траве и только
потом соображает, что это поливальный шланг, а не змея. Следуя ассоциациям,
возникшим в ходе эволюционного отбора, лимбическая система соответствует
правилу: «лучше лишний раз перестраховаться, чем потом жалеть о случив�
шемся». В ходе своего познавательного опыта именно под влиянием миндали�
ны человек совершает непроизвольные поступки. И только несколько мгнове�
ний спустя наше рациональное «я» испытывает смущение от совершения явно
глупого поступка. Можно легко найти и другие примеры взаимодействия лим�
бической и корковой систем в нашем повседневном поведении. Далее мы еще
коснемся этого подробнее.
Лимбическая система тесно взаимосвязана с корой, в особенности через
орбитальные извилины вентромедиальной префронтальной коры (ВМ�ПФК) и
островную кору. Из�за этих тесных связей ВМ�ПФК еще называют чувству�
ющей частью думающего мозга. ВМ �ПФК
—
это ворота между лимбической
системой и неокортексом.
Неокортекс, или мозг приматов, — эволюционно последний слой мозга. Он
состоит из складчатого плаща, покрывающего большие полушария, и ряда под�
130
Глава 13
корковых ядер, например базальных ганглиев. У человека и других приматов
мозговой плащ чрезвычайно разросся по сравнению с другими млекопитаю�
щими. Человеческий неокортекс — вместилище сложнейших когнитивных,
лингвистических, моторных, сенсорных и социальных способностей. Неокор�
текс дает нам значительную гибкость и креативность при адаптации к изме�
няющимся условиям окружающей среды. Кора служит и для контроля, и для
социализации порожденных лимбической системой эмоций. Оценка ситуаций
корой необходима также и для более тонкого проявления эмоций, чем то, на что
способна лимбическая система в одиночку.
1.2.
Базовые эмоции и роль рефлексивного сознания
Эмоциональные ответные реакции управляются разными путями.
1. Классически обусловленные (рефлекторные) реакции на стимулы, ранее до�
ставлявшие удовольствие или причинявшие боль, — опосредуются подкорко�
выми системами.
2. Когнитивные оценки контекстных стимулов — неокортексом.
Нейробиологи традиционно уделяли основное внимание классическим ис�
следованиям условных рефлексов на животных моделях. На возможность су�
ществования в мозге сразу нескольких эмоциональных систем практически не
обращали внимания. В настоящее время считается, что возникновение чуства
страха связано с дофаминергической «системой подкрепления» миндалины.
Однако же объяснению нейрональной базы огромного разнообразия испыты�
ваемых нами эмоций явно не уделяется достаточно внимания. С развитием все
более точных методов визуализации мозга эта ситуация может измениться.
Хотя формирование условного рефлекса сохраняет свою значимость для
объяснения обучения путем эмоциональных ассоциаций, аффективная нейро�
биология предлагает и другую концепцию. Предполагается, что эмоции млеко�
питающих порождаются несколькими отдельными генетически детерминиро-
ванными сетями мозга, каждая из которых служит определенной адаптивной
функции, порождает свою уникальную моторную деятельность при активации,
обладает уникальными «условиями вызова» или запускающими стимулами и
каждая является нейрональным субстратом определенных осознаваемых эмо�
циональных состояний. Психо�этологический подход позволяет увидеть работу
эмоций в контексте адаптации.
2.0 .
Эмоциональные системы мозга по Панксеппу
Яак Панксепп (J��� ��������, 1���) предложил свое функциональное определе�
ние эмоциональной системы в мозге (рис. 13.2).
1. Лежащие в основе контуры генетически предрасположены безусловно от�
вечать на стимулы, эволюционно важные для данного вида.
2. Эти контуры запускают моторные программы, а также изменения в вегета�
тивном и гормональном статусе в ответ как на закономерные воздействия
окружающей среды, так и на случайные факторы.
3. Они подстраивают чувствительность сенсорных систем так, чтобы они от�
вечали на стимулы, существенные для вызванной эмоции.
4. Положительная обратная связь нейрональной активности означает, что эмо�
циональное возбуждение переживет вызвавшие его обстоятельства.
Эмоции
131
5. Эмоциональные контуры могут находиться под сознательным контролем.
6. Эмоциональные контуры реципрокно влияют на выше лежащие системы
принятия решений и анализа, и на сознание.
7. Контур способен порождать субъективно четко различимые ощущения
(не показано на рис. 13.2).
Панксепп (1���) описал небольшой набор «вшитых» в мозг млекопитаю�
щих эмоциональных систем. (Мы последуем за Панксеппом и будем обозначать
системы прописными буквами, которые будут напоминать нам, что мы говорим
об эмоциональных системах, а не просто о сознательных ощущениях, связан�
ных с ними или с одиночными мозговыми структурами.) Первые четыре эмоци�
ональные системы появляются у всех млекопитающих вскоре после рождения.
• ПОИСК — аппетитивная* система, которая заставляет млекопитающих
интересоваться окружающим миром и стимулирует целенаправленное
поведение, направленное на разнообразные объекты, например на пищу,
укрытие, полового партнера.
• СТРАХ — это система, которая отвечает на боль и угрозу существования
и приводит к хорошо известным реакциям: бегству, борьбе или замира�
нию.
• ЯРОСТЬ — эта система управляет гневом и запускается неудовлетворен�
ностью, телесным раздражением, ограничением свободы передвижения.
• ПАНИКА — система, которая отвечает запуском плача и крика у детены�
шей в ответ на их отделение от тех, кто о них заботится.
Эти четыре эмоциональные системы и их взаимные тормозящие и активиру�
ющие связи показаны на рис. 13.3 .
* Не путать с клеточными слоями коры! — Прим. перев.
Рис. 13 .2 . Функции эмоциональных систем: (1) безусловный сенсорный вход, (2) коор�
динированные физиологические и поведенческие реакции, (3) пропускание входящих
сигналов, (4) положительная обратная связь, (5) когнитивная деятельность, активиру�
ющая эмоции, (6) эмоциональный контроль когнитивной деятельности. (Источник:
��������, 2006.)
132
Глава 13
Вдобавок к четырем фундаментальным эмоциональным системам, на опре�
деленных стадиях развития млекопитающего включаются еще три специализи�
рованных системы.
• СТРАСТЬ — система, координирующая сексуальное поведение и чув�
ства.
• ЗАБОТА — система, которая у взрослых является аналогом системы ПА�
НИКИ детенышей; ЗАБОТА работает и у матерей, и у отцов, способствуя
социальным связям и заботе о потомстве.
• ИГРА — система, которая организует драку в форме игры, спонтанно
вспыхивающую у детенышей млекопитающих; эта система поддержива�
ет смех и может быть нейрональным субстратом радости.
Поскольку каждая из этих эмоциональных систем имеет свою собственную
«принципиальную схему», понимание организации эмоций в мозге становится
весьма затруднительным!
Рис. 13 .3. Четыре основных эмоциональных системы млекопитающих показаны вме�
сте с базовыми формами поведения. ДА — дофамин. (Источник: ��������, 1���.)
Эмоции
133
Большая часть того, что мы знаем об эмоциональных системах млекопита�
ющих (табл. 13.1), получена в исследованиях на животных. Изучение эмоций
на людях ограничено этическими запретами на использование инвазивных ме�
тодик и на типы и интенсивность эмоциональных стимулов, которые можно
применять для вызывания реакций. Лабораторные исследования эмоций че�
ловека основываются на наблюдении за поведением и на нейровизуализации
после относительно мягких заданий, вызывающих эмоции, таких, как прямая
сенсорная стимуляция и условные рефлексы, просмотр изображений эмоци�
онально насыщенных событий и генерация эмоций путем обращения к эмо�
ционально нагруженным воспоминаниям. Дополнительные данные удается
получить при обследовании неврологических больных с повреждениями раз�
личных зон мозга.
Таблица 13.1
Основные эмоциональные системы мозга и связанные с ними зоны
(Источник: Panksepp, 2006)
Основная
эмоциональная
система
Соответствующие зоны мозга мле-
копитающих
Соответствующие
эмоциональные
ощущения
СТРАХ/тревога
От центральной и латеральной минда�
лины — к медиальному гипоталамусу
и дорсальному ОВСВ
Страх, тревога
ПОИСК/
предвкушение
Мезолимбические эфференты ВОП в
прилежащее ядро перегородки; мезо�
кортикальные эфференты ВОП в орби�
тофронтальную кору; от латерального
гипоталамуса к ОВСВ
Интерес, любопытство
ГНЕВ/ярость
От медиальной миндалины к опорному
ядру концевой полоски (ОЯКП); от ме�
диального и перифронтального гипо�
таламуса к ОВСВ
Ярость, презрение
ПАНИКА/
отделение от мате�
ри, дистресс
Передняя поясная кора, ОЯКП, пре�
оптическая область; дорсомедиальный
таламус, ОВСВ
Грусть, робость,
вина/стыд
СТРАСТЬ/
сексуальность
Кортикомедиальная
миндалина,
ОЯКП; преоптическая область, ВОП,
ОВСВ
Эротические
ощущения,
ревность
ЗАБОТА/опека
Передняя поясная кора, ОЯКП; преоп�
тическая область, ВОП, ОВСВ
Любовь
ИГРА/радость
Дорсомедиальный гипоталамус; пара�
фасцикулярная область, ОВСВ
Радость, счастье
ОВСВ — околоводопроводное серое вещество, расположено в глубине среднего мозга,
окружая сильвиев водопровод, простирается в ростро�каудальнои направлении от зад�
ней комиссуры до голубого пятна. ОЯКП — опорное ядро концевой полоски, скопление
подкорковых ядер, расположенное медиальнее базальных ганглиев и выше гипоталаму�
са. ВОП — вентральная область покрышки, расположена в среднем мозге.
134
Глава 13
2.1.
Переживание эмоций
Переживание эмоций нельзя изучать на животных, хотя, наблюдая за их по�
веденческими реакциями, можно догадаться, что они чувствуют. В настоящее
время идут напряженные дебаты о том, позволяют ли проведенные исследова�
ния говорить о «чувствах» у других млекопитающих, кроме человека. Похоже,
что сейчас общее мнение специалистов склоняется в пользу принятия этого по�
ложения — на основании общности нейроанатомии эмоций и систем, лежащих
в основе сознательного опыта.
Дамасио и соавторы (2000) изучали нейрональный субстрат эмоциональ�
ных ощущений человека. Они просили испытуемых извлечь из памяти и за�
ново пережить эмоционально насыщенные личные воспоминания, и при этом
наблюдали, какие области головного мозга активируются при различных эмо�
циональных состояниях. Используя позитронно�эмисионную томографию
(ПЭТ), исследователи обнаружили весьма различные нейрональные картины
активности для страха, счастья, грусти и гнева (рис. 13.4). Они отметили раз�
личные картины активации и деактивации в коре и подкорковых зонах, связан�
ных с представительством и регуляцией эмоций, а также с гомеостазом всего
организма, включая островковую кору, вторичные соматосенсорные зоны, зону
поясной извилины и ядра ствола мозга и гипоталамуса.
Поскольку эти эмоциональные ощущения были ответом на внутренние пе�
реживания и не требовали ни восприятия внешних стимулов, ни мимических
или двигательных выражений, Дамасио и его коллеги полагали, что итоговые
нейрональные карты представляют собой корреляты различных эмоциональ�
ных переживаний и пришли к следующему выводу:
«Нейрональные картины, возникающие во всех этих структурах, образуют много�
мерные карты внутреннего состояния организма и формируют основу для столь важ�
ного аспекта психических состояний, известного как переживания».
3.0 .
Система страха
Мы более подробно рассмотрим описанные Панксеппом системы СТРАХА и ПО�
ИСКА — две достаточно подробно изученные эмоциональные системы, влияние
которых на когнитивные функции уже хорошо известно. Затем мы расширим ох�
ват и взглянем на систему страха у людей и на систему подкрепления, включая
эмоциональные аспекты подкрепления: предпочтения, желания, обучение.
Нейрональная система страха направлена прежде всего на избегание боли
и повреждающих организм воздействий. Ее основой являются центральное и
латеральное ядра миндалины, связанные с медиальным гипоталамусом и дор�
сальным околоводопроводным серым веществом среднего мозга.
Рис. 13.4 . Результаты ПЭТ�визуализации при самогенерации у испытуемых эмоций ра�
дости и печали (верхний ряд), страха и гнева (нижний ряд). Красным и желтым обозна�
чены зоны повышенной метаболической активности, фиолетовым и синим — понижен�
ной. mb — средний мозг, ob — орбитофронтальная кора, i� — островок, bf — основание
переднего мозга, �c — передняя поясная кора, � — главная борозда, hy� — гипоталамус,
�c — задняя поясная кора, �u — вторичная сенсомоторная кора, стимул 2. (Источник:
D�m��io et al., 2000.)
Эмоции
135
136
Глава 13
Эта система отвечает как на безусловные стимулы (громкие звуки, неясные
очертания и резкие движения объекта, болевые стимулы, страшные лица), так
и на условные стимулы (классические условные сигналы опасности, воспоми�
нания, образы), поступающие от таламуса, а также сенсорной и ассоциативной
коры (рис. 13.5). Реципрокные эфферентные пути возвращают сигналы обрат�
ной связи в те же области таламуса и коры и подстраивают обработку сенсор�
ной информации в соответствии с эмоциональным фоном (рис. 13.6). Ясно, что
эфферентные (выходные) пути из миндалины в кору столь же сложны и на�
сыщены, как и афферентные (входные) пути из коры в миндалину. Афферент�
ные сигналы поступают в миндалину по четырем путям. Обонятельная инфор-
мация, играющая важную роль для млекопитающих, поступает в миндалину
напрямую из обонятельной коры без дополнительной обработки в таламусе.
Возможно, с этим связана присущая запахам способность вызывать эмоцио�
нальные воспоминания. Висцеральная информация достигает миндалины через
гипоталамус и зону перегородки через пограничную полоску (stria terminalis).
Эмоционально-значимая информация о внутреннем состоянии организма при�
ходит от гипоталамуса, таламуса, ствола мозга, а также орбитальной коры и
передней поясной коры через вентральный путь. И наконец, сенсорная инфор�
мация приходит напрямую от структур височной доли, таких как первичная
слуховая кора и гиппокамп.
Сама миндалина является совокупностью ядер и внутренних путей, выпол�
няющих различные функции в формировании эмоций. Выделяют базолате-
ральный комплекс, центромедиальное ядро и кортикальное ядро. Базолатераль�
ный комплекс можно далее разделить на латеральные, базальные и добавочные
базальные ядра.
Латеральная миндалина, которая является афферентом для всего остального
базолатерального комплекса и центромедиального ядра, получает информацию
от сенсорных систем. Центромедиальное ядро — основной выход базолате�
рального комплекса, и у млекопитающих связано с эмоциональным возбужде�
нием. Корковое ядро связано с восприятием запахов и феромонов, оно получает
информацию от обонятельной луковицы (рис. 13.7).
Эфферентные пути от миндалины отражают афферентные пути и возвраща�
ют сигналы в подкорковые области и ствол мозга. Для рассмотрения взаимосвя�
зи когнитивных функций и эмоций важно, что существуют прямые эфферент�
ные пути из миндалины в энторинальную кору, кору нижней области височной
доли и, наконец, в зрительную кору, включая так называемую фузиформную
лицевую область, F FA (англ. fu�iform f�c� �r��)* (рис. 13.7). Миндалина и кора
работают совместно по принципам сверху вниз и снизу вверх, настраивая мозг
для адаптивных ответов на значимые угрозы со стороны окружающей среды.
И хотя большая часть исследований нейропсихологических функций и ра�
боты миндалины была проведена на крысах и других модельных млекопитаю�
щих, наиболее важные результаты постепенно накапливаются в ходе изучения
человека. Считается, что подкорковые эмоциональные системы мозга являются
консервативными в ряду млекопитающих; одни и те же структуры мозга одина�
* Область распознавания лиц, расположенная на вентральной поверхности веретенообразной из�
вилины височной доли. Относится к парагиппокампальной области. Открыта Джюстиной Сер�
джент (J. S�rg��t) с соавторами в 1��2 г. с помощью ПЭТ и детально исследована Нэнси Кэнвишер
(N. K��wi�h�r) с соавторами в 1��7 г. с помощью фМРТ. — Прим. перев.
Эмоции
137
Рис. 13 .5. Афферентные пути миндалины
Рис. 13 .6. Эфферентные пути миндалины
138
Глава 13
ково работают у различных групп животных. Большинство исследователей по�
лагает, что полученные на крысах или других млекопитающих данные можно
будет применить и к нейропсихологии человека. Однако, чтобы подтвердить эту
гипотезу, необходимо провести еще множество исследований. В одной из ран�
них работ, подтвердивших роль миндалины в связанном со страхом обучении у
человека, Лабар и соавторы (L�B�r et al., 1���) с помощью фМРТ показали, что
формирование условнорефлекторной реакции страха у испытуемых вызывало
усиленный BOLD�сигнал (от англ. blood oxygen level dependent — зависимый
от уровня кислорода в крови) от миндалины. По величине этой BOLD�реакции
можно было предсказать силу условного ответа.
3.1.
Сознательное и бессознательное восприятие страха:
«нижний» и «верхний» пути по Леду
Леду (L� Doux, 1��6) обозначил два сенсорных пути, проводящие информа�
цию о пугающих стимулах к миндалине, как «нижний» и «верхний». Нижний
путь — быстрый, проходящий от рецептора через сенсорный таламус к минда�
лине, минуя кору больших полушарий (рис. 13.�). Как уже говорилось в начале
этой главы, обработка сигнала на прямом таламо�амигдалярном пути (нижнем
пути) обладает низким пространственным разрешением и потому может отве�
чать только на простые стимулы или грубые характеристики сложных стиму�
лов. Эта «быстрая и грубая» обработка обеспечивает автоматические бессозна�
Рис. 13 .7 . Ядра миндалины и их эфферентные связи с сенсорной корой. (Источник:
Vuill�umi�r, 2005.)
Эмоции
139
тельные реакции на общие черты потенциально опасных стимулов еще до того,
как мы успеваем об этом задуматься.
Таламо�кортико�амигдалярный путь (верхний) длиннее, зато позволяет про�
водить сложный анализ стимулов, учитывающий их контекст, и реагировать со�
знательно и обдуманно. Верхний путь больше подвержен влиянию социальных
норм и личностных особенностей принятия решений, и, значит, может отра�
жать культурно�специфические эмоциональные реакции.
3.2 .
Неосознаваемый страх
Ряд исследований поддерживает гипотезу о том, что эмоциональные стимулы
могут обрабатываться путем чередования сознательного и бессознательного
путей. Рассмотрим одно из них, проведенное Вюйемьером и его коллегами
(Vuill�umi�r et al., 2002). Они искали различные нейрональные ответы на пуга�
ющие или нейтральные стимулы, которые поступали при наличии или отсут�
ствии их осознания у больного с правотеменным неглектом (сенсорным невни�
манием) и зрительным угасанием из�за повреждения правой нижней области
теменной коры (рис. 13.�).
Теменной неглект — это нарушение восприятия тела и зрительно�простран�
ственной информации у людей с повреждениями теменной коры.
Больные с повреждениями в правой теменной доле могут испытывать труд�
ности с восприятием стимулов, которые им показывают в контралатеральной
Рис. 13 .8. Два пути системы страха: нижний и верхний
140
Глава 13
(с противоположной стороны) повреждению части поля зрения — слева. В ла�
боратории при демонстрации одиночных стимулов в правой или левой полови�
не поля зрения пациенты с неглектом не испытывают затруднений с восприяти�
ем. Однако, когда им одновременно показывают два разных стимула в правой
и левой половинах поля зрения, пациенты с правосторонним теменным неглек�
том часто не замечают стимула, показанного слева. Этот феномен называют
выпадением. Он позволяет исследователям изучать обработку стимула при на�
личии или отсутствии его сознательного восприятия.
Вюйемьер и соавторы (2002) показывали испытуемому картинки с выраже�
нием страха на лицах, нейтральными лицами или домиком, все вместе или по
отдельности, как показано на рис. 13.10 и 13.11 . Были получены важные резуль�
таты: оказалось, что существует резервная обработка лиц в правом полушарии,
сохраняющаяся, несмотря на теменное повреждение и заявленную больным
неспособность видеть стимулы.
Находки Вюйемьера и его коллег.
1. Изображения лиц с выражением страха (но не нейтральных!) активировали
левую миндалину, экстрастриатную (зрительную) кору, орбитофронтальную
кору с обеих сторон, правую и левую веретенообразную извилину — и когда
испытуемый их видел, и при их выпадении.
2. Обработка миндалиной изображений испуганных лиц не менялась от того,
видел ли испытуемый эти лица, или они выпадали. Изображения лиц с вы�
ражением страха могут обрабатываться бессознательно и независимо от по�
вреждений в теменной области.
3. Сознательное восприятие повышало активность левой веретенообразной
извилины, теменной и префронтальной коры левого полушария по сравне�
нию с выпадением. Сознание изменяет кортикальную обработку.
Полученные в другой серии результаты на испытуемых без повреждений
мозга подтверждают, что внимание к стимулам (имеющим или не имеющим
Рис. 13 .9. Поврежденная нижняя область теменной коры правого полушария у испыту�
емого. (Источник: Vuill�um�ir et al., 2002.)
Эмоции
141
отношение к заданию) и эмоциональная значимость стимулов (пугающие или
нейтральные) осуществляют независимый вклад в визуальное восприятие
(Vuill�um�ir et al., 2001).
Активность веретенообразной извилины вентральной височной доли выше,
когда на стимул обращают внимание; ее активация сильнее для восприятия изо�
бражений испуганных лиц, независимо от уровня внимания (рис. 13.12).
Рис. 13 .10 . Односторонние стимулы, использованные Вюйемьером с соавторами (2002).
Когда испытуемый фиксировал взор на светлом квадрате, изображения проецировались
либо для контралатерального правого, либо для левого полушария. Испытуемый видел
их в течение 100% времени предъявления. (Источник: Vuill�umi�r et al.,2002.)
Рис. 13 .11 . Двусторонние (билатеральные) стимулы. Когда испытуемый фиксировал
взор на светлом квадрате, он отчитывался о невозможности увидеть лицо (проецирую�
щиеся в его поврежденную кору правого полушария) приблизительно в 65% времени
предъявления. (Источник: Vuill�umi�r et al., 2002.)
142
Глава 13
Все эти результаты подтверждают, что существуют независимые сознатель�
ные и бессознательные пути к эмоциональной системе страха в миндалине.
3.3 .
Аффективное слепозрение
Некоторое время назад было обнаружено (��i��r��t�, 1��6), что больные с по�
��i��r��t�, 1��6), что больные с по�
, 1��6), что больные с по�
вреждениями зрительной коры сохраняют способность реагировать на визуаль�
ные детали объектов, предъявляемых в соответствующей повреждению слепой
зоне поля зрения. И хотя такие пациенты не осознают эти стимулы и не могут
описать их, они, тем не менее, могут отвечать адекватным поведением на появ�
ление определенных объектов, например, совершать правильные хватательные
движения руками или «угадывать» с вероятностью выше случайной направ�
ление движения, ориентацию и цвет объектов. Такую способность называют
слепозрением. В последнее время исследователи смогли сымитировать слепоз�
рение в лаборатории, производя транскраниальную магнитную стимуляцию
(ТМС), чтобы на время прервать переработку информации в зрительной коре
(Ro et al., 2004). Эта работа, похоже, подтверждает устоявшееся мнение о том,
что подобная бессознательная обработка визуальной информации зависит от
эволюционно древнего верхнего четверохолмия.
Рис. 13 .12 . Активность фузиформной лицевой области височной доли: роль внимания
и эмоций. Сл ева вверху: коронарный срез, показывающий активность фузиформной об�
ласти. Справа вверху: горизонтальный срез, показывающий активность фузиформной
области. Внизу: эффекты внимания и эмоций на уровень активации фузиформной об�
ласти. (Источник: Vuill�umi�r et al., 2001.)
Эмоции
143
Для описания способности пациентов с повреждениями зрительной коры
реагировать на эмоциональные свойства стимула, предъявляемого им в слепой
зоне, был предложен термин аффективное слепозрение (d� G�ld�r et al., 2000;
H�ywood, K��tridg�, 2000).
Де Гелдер и его соавторы (d� G�ld�r et al., 2005) смогли исследовать взаи�
модействие сознательной и бессознательной обработки эмоциональной инфор�
мации у больного с обширными повреждениями левой зрительной коры, одно�
временно показывая картинки с различными выражениями лиц в интактной и
слепой зонах поля зрения. Изучая отчет пациента об осознанно воспринятых
стимулах, ученые могли выявить влияние неосознаваемых стимулов на созна�
тельный отчет. Когда осознаваемый и неосознаваемый стимулы страха совпа�
дали (испуганные видимое и невидимое лица), то доля правильных определе�
ний выражения лица была высокой.
Однако для неконгруэнтных (несовпадающих) лиц (счастливое видимое
лицо, спаренное с испуганным невидимое лицом, или наоборот), вероятность
правильного определения падала до уровня случайной. В испытаниях с конгру�
энтными (совпадающими) лицами повышалась активность левой миндалины,
что навело исследователей на мысль о влиянии бессознательно воспринятых
невидимых испуганных лиц на сознательное восприятие, и что этот эффект
опосредуется миндалиной и верхним четверохолмием. Дополнительный экс�
перимент также был посвящен влиянию видимых и невидимых лиц на вос�
приятие, на этот раз, слухового сигнала — эмоционального голоса (радостного
или испуганного). Когда совпадали испуганное лицо и испуганный голос, го�
лос воспринимался намного точнее, чем когда они не совпадали, независимо
от того, воспринималось ли лицо сознательно или нет. Невидимые счастливые
лица и эмоциональные рисунки без лиц не вызывали подобного улучшения.
Исследователи пришли к выводу, что распознавание пугающих стимулов про�
исходит инстинктивно и не зависит от их осознания. Существуют, видимо, от�
дельные корковые и подкорковые пути восприятия стимулов, связанных с чув�
ством страха.
3.4 .
Взаимодействие познавательных
и эмоциональных функций при реакциях страха
Теперь, когда мы уяснили принципиальную схему входов и выходов миндали�
ны, мы можем перейти к описанию взаимодействия когнитивных функций и
эмоций, зависящих и от миндалины, и от связанных с ней зон.
В недавнем обзоре роли миндалины в обработке эмоциональных процессов
(�h�l�� ��d L�Doux, 2005) выделено пять областей, где исследования и на жи�
вотных, и на людях выделили сходное взаимодействие когнитивных функций и
эмоций, проходящее с участием миндалины:
1) имплицитное эмоциональное обучение и память;
2) эмоциональная модуляция памяти;
3) влияние эмоций на восприятие и внимание;
4) эмоции и социальное поведение;
5) торможение и регулирование эмоций.
Рассмотрим принципиально важные исследования, проведенные на людях.
144
Глава 13
3.5 .
Имплицитное эмоциональное обучение и память
Имплицитная память уже обсуждалась в одной из предыдущих глав. Эта такой
вид обучения, который проявляется в поведенческих показателях, но не под�
дается произвольному извлечению из памяти или сознательному отчету. К ней
относят различные процедурные навыки, языковую грамматику и классически
обусловленные ассоциации. Имплицитная эмоциональная память включает
хранение классических условнорефлекторных эмоциональных отношений, к
которым невозможно сознательно обратиться или отчитаться о них. Информа�
ция об этом типе памяти была получена в том числе и от больных с различными
неврологическими нарушениями.
Пациенты с поврежденными миндалинами, но сохранными гиппокампами,
не демонстрируют физиологических реакций, характерных для условнореф�
лекторного страха (повышения частоты сердечных сокращений, электрокож�
ные реакции и др.), но могут вспомнить и дать отчет об эпизодических воспо�
минаниях, касающихся обстоятельств, сопровождавших условнорефлекторное
обучение. И наоборот, пациенты с повреждениями гиппокампа и уцелевшими
миндалинами неспособны дать сознательный отчет о событиях, происходящих
при формировании условного рефлекса страха, но демонстрируют нормаль�
ные для этой реакции физиологические изменения (B�ch�r� et al, 1��5). Это
расхождение между зависимым от миндалины физиологическим выражением
условного рефлекса страха и зависимым от гиппокампа эпизодическим созна�
тельным отчетом о происходивших событиях подтверждает, что в хранение
эмоциональных воспоминаний вовлечено множество систем.
Клинические психологи, работающие с жертвами травмирующих событий
(катастрофы, надругательства, войны и др.), также интересуются имплицитной
эмоциональной памятью.
3.6 .
Эмоциональная модуляция эксплицитной памяти
Несколько лет назад были получены психологические доказательства того, что
умеренное эмоциональное возбуждение (обычно — страх) во время какого�то
события приводит к лучшему сохранению эксплицитных воспоминаний о нем.
Например, фильмы ужасов запоминаются лучше, чем нейтральные, причем
наблюдается линейная зависимость между степенью эмоционального напряже�
ния в ходе просмотра (измеренного по самоотчету и ПЭТ�активности миндали�
ны) и уровнем запоминания (C�hill et al., 1��6). Если же речь идет о предсказа�
нии взаимодействия эмоций и консолидации памяти, то возникает классическая
колоколообразная кривая. Слишком сильная активация миндалины приводит к
потере эксплицитных воспоминаний об эмоциональных событиях (C�hill ��d
McG�ugh, 1���).
Похоже, что пути консолидации эксплицитной памяти зависят от восприя�
тия эмоциональных стимулов миндалиной, за которым следует активация гипо�
таламуса и гипофиза, приводящая в итоге к выбросу вначале гормона мозгового
вещества надпочечников адреналина, а затем — гормона коры надпочечников
кортизола. И адреналин, и кортизол (часто называемые гормонами стресса), ви�
димо, влияют на зависимое от гиппокампа формирование эксплицитной памяти
(рис. 13.13). У пациентов с двусторонним повреждением миндалин не наблю�
дается улучшенного запоминания эмоциональных стимулов (C�hill et al., 1��5).
Эффект возбуждения из�за выброса гормонов стресса также можно предотвра�
Эмоции
145
тить введением антагонистов β�адренорецепторов («β�блокаторов», например,
пропанолола) (C�hill et al., 1��4), блокирующих рецепторы адреналина. Пере�
возбудимость у больных с посттравматическим стрессовым расстройством
(ПТСР) можно смягчить, применив пропанолол при экстренной помощи вскоре
после травмирующих событий (�itm�� et al., 2002).
3.7.
Влияние эмоций на восприятие и внимание
Одной из определяющих черт эмоциональных систем по критериям Панксеппа
(о которых рассказывалось в начале этой главы) является их способность вли�
ять на более высокоуровневые процессы. Свидетельства такого влияния были
получены на животных: условнорефлекторная реакция страха меняла предста�
вительство условного стимула в слуховой коре. Работая с морскими свинками,
Бакин и соавторы (B��i� et al., 1��6) обнаружили, что выработка классического
условного рефлекса на тоны сдвигает частоту настройки отдельных нейронов
коры в сторону условного стимула. Исследователи отметили, что эта пластич�
ность рецептивного поля ассоциативна, высоко специфична, быстро развивает�
ся и сохраняется неопределенно долго.
Сходные, но более ограниченные результаты были получены и на человеке.
Как мы уже знаем, миндалина имеет прямые и непрямые пути, связывающие ее
с сенсорной корой, которые могут служить для положительной обратной связи
с процессами восприятия.
Рис. 13 .13. Гормоны стресса и консолидация эксплицитной памяти. Адреналиновый
путь отмечен зеленым цветом. Кортизоловый путь — красным. Обратите внимание, что
оба пути начинаются в миндалине, доходят до надпочечников и возвращаются к минда�
лине и гиппокампу, пройдя через весь организм
146
Глава 13
Исследования по нейровизуализации с использованием методов зрительной
маскировки и подсознательного условнорефлекторного страха (Morri� et al.,
2001) показали, что испытуемые, которым в качестве подсознательных услов�
ных стимулов предъявляли изображения лиц, а в качестве безусловного стиму�
ла — неприятный громкий шум, в ходе испытаний демонстрировали повыше�
ние реактивности миндалины и зрительной коры в ответ на условный стимул.
Исследователи пришли к выводу, что «параллельные связанные с обучением
реакции, наблюдаемые в вентральной миндалине и зрительной коре, согласу�
ются с предположением, что таким образом эфференты «обратной связи» от
базальных ядер миндалины опосредуют зависимую от эмоций модуляцию зри�
тельной обработки».
Видимо, миндалина играет роль в определении того, как прошедшие мимо
внимания, но важные стимулы получают доступ к сознанию с помощью
врéменной обратной связи с получающими сенсорную информацию областями
коры. Активность миндалины может мгновенно сделать кортикальные области
более восприимчивыми к определенным адаптивно важным стимулам.
Пример такой повышенной восприимчивости к значимым стимулам можно
увидеть в тесте Струпа (блок 13.1), когда персонально эмоционально�нагру�
женные слова сравниваются с другими положительными, отрицательными и
нейтральными словами.
В тестировании участвовали ветераны войны во Вьетнаме (McN�lly et al.,
1��0) и гражданские лица, пережившие психическую травму (T�ylor et al.,
2006). Слова, имевшие персональную значимость для испытуемых (похорон�
ный мешок, Вьетнам, медицинская эвакуация — для ветеранов, или револь�
вер, инцест, 11 сентября, пожар — для гражданских жертв терактов и наси�
лий), вновь и вновь быстрее достигали сознания испытуемых, чем нейтральные
слова, и соответственно, испытуемые с большей задержкой сообщали о цвете,
которым было напечатано данное слово. Однако больные с двусторонним по�
вреждением миндалин не демонстрируют ожидаемого заострения внимания на
эмоциональных словах, характерного для нормальных испытуемых. Хотя паци�
енты с повреждениями миндалины понимают значение слов, они не проявляют
избирательного обострения внимания по отношению к вербальным стимулам
с негативным значением (A�d�r�o� ��d �h�l��, 2001).
Блок 13.1 . Эмоциональный тест Струпа
Версия эмоционального теста Струпа, вызывающая реакцию у ветеранов
войны во Вьетнаме (Источник: McNally et al., 1990)
Контроль
Связанные с войной
Негативные Нейтральные
Позитивные
OOOOOOO перестрелка
микробы
смесь
преданный
OOOOOOO медицинская эвакуация брань
миллионер
приятный
OOOOOOO Вьетнам
грязный
бето н
счастливый
OOOOOOO похоронный мешок
моча
вход
дружба
Эмоции
147
3.8 .
Эмоции и социальное поведение
Двустороннее повреждение миндалин у обезьян вызывает резкие изменения в
поведении, включая исчезновение страха перед опасными стимулами, поедание
несъедобных объектов и нетипичное сексуальное поведение (синдром Клюве�
ра–Бьюси). Однако подобных изменений поведения у людей с повреждениями
миндалин не было отмечено, вероятно, потому, что человек обладает высоко
развитыми неокортикальными системами социального контроля и торможения,
отсутствующими у других приматов. Имеются лишь некоторые данные, что та�
ким пациентам свойственны более тонкие нарушения, например, при оценке
эмоциональных выражений лица.
Как мы уже отмечали в начале главы, миндалина реагирует на простые со�
ставляющие сложных стимулов, например на мелкие частотно�пространствен�
ные детали изображений лиц (Vuill�um�ir et al., 2003). В соответствии с этим
общим правилом полученные данные показывают, что миндалина реагирует на
широко открытые глаза на испуганных или удивленных лицах. В ходе уточняю�
щего исследования, проведенного Уэйленом и коллегами (�h�l�� et al., 2004),
выяснилось, что миндалина избирательно реагирует на предъявленные ниже
порога сознательного восприятия белки испуганных глаз по сравнению с бел�
ками радостных глаз (рис. 13.14).
И наконец, Адольфс и соавторы (Adol�h� et al., 2005) обнаружили, что
больные с двусторонним повреждением миндалин при оценке выражения на�
рисованного лица не смотрят на глаза. Этот недостаток может быть причиной
характерных для таких пациентов сложностей с определением чужих эмоций.
Больные с амигдалярными нарушениями недооценивают выраженность эмо�
ций и переоценивают надежность и контактность других людей по сравнению
со здоровыми испытуемыми.
3.9 .
Торможение и регуляция эмоций
Обучение с участием страха — это стабильное и долговременное обучение,
крайне устойчивое к изменениям и волевому контролю. Очень трудно предот�
вратить реакцию испуга. И хотя такая реакция адаптивна в том смысле, что
Рис. 13 .14. Миндалина реагирует на белки испуганных глаз сильнее, чем на белки
счастливых глаз. Справа — зависимый от уровня кислорода в крови (BOLD) сигнал
от левой вентральной миндалины на белки испуганных глаз. (Источник: �h�l�� et al.,
2004, из �h�l�� ��d L�Doux, 2005.)
148
Глава 13
позволяет избегать предсказуемых опасностей окружающей среды, однако если
она неуместна или чрезмерна, то может заблокировать всякое реагирование.
В связи с этим исключительно важно исследовать возможности модификации
или устранения эмоциональных ассоциаций с помощью их угашения или ин�
версии, реконсолидации и эмоциональной регуляции.
Угашение — один из Павловских методов работы с условными рефлексами,
при котором условный стимул (УС), ранее связанный с негативным безуслов�
ным стимулом (БС), в ряде испытаний предъявляется в одиночку. Испытуемый
усваивает, что УС в этой ситуации больше не связан с неприятным событи�
ем. Однако по прошествии времени, в другой обстановке или при повторном
предъявлении БС — УС все равно будет вызывать реакцию страха. Это показы�
вает, что приобретенная реакция страха хранится в памяти, и угашение работа�
ет путем торможения этой реакции.
Нейробиология угашения сейчас уже хорошо изучена. Угашение крити�
чески зависит от работы NMDA�рецепторов* в миндалине. При блокировке
NMDA�рецепторов у крыс угашение условнорефлекторного обучения наруша�
ется. При повышении активности NMDA�рецепторов угашение усиливается.
Не так давно агонисты NMDA были впервые использованы для стимуляции
угашения у людей — больных с агорафобией (R���l�r et al., 2004). Исследова�
тели обнаружили, что две дозы D�циклосерина в сочетании с экспозиционной
терапией** значимо уменьшали агорафобию по сравнению с терапией подвер�
гания в сочетании с плацебо при оценке по самоотчету и по кожногальваниче�
ской реакции на БС. Улучшения сохранялись по прошествии трех месяцев по�
сле лечения. Эти результаты позволяют надеяться на развитие методов лечения
отрицательных эмоций, основанных на понимании нейробиологии восприятия
страха миндалиной.
Инверсное (реверсивное) обуславливание (условнорефлекторное переключе-
ние) схоже с угашением тем, что оно меняет сопряженность между условными
стимулами и реакциями. Согласно этой парадигме, испытуемые сначала обуча�
ются бояться одного условного стимула (УС+ ), за которым неизменно следу�
ет вызывающий страх безусловный стимул (БС), например громкий шум, и не
бояться другого условного стимула (УС–), за которым никогда не следует БС.
В следующей фазе зависимости обращаются (инвертируются) так, что ней�
тральный ранее стимул оказывается спарен с БС.
Моррис и Долан (Morri� ��d Dol��, 2004) использовали такое условнореф�
лекторное переключение, чтобы исследовать роль миндалины и орбитофрон�
тальной коры в условных реакциях страха. Они использовали предъявление
нейтрального выражения лица как УС+ и громкий шум — как БС. Было об�
наружено, что вначале в ходе выработки условного рефлекса в миндалине раз�
вивается сильная двусторонняя активация в ответ на УС+. После переключения
новое лицо, предъявляемое в качестве УС, вызывает усиленные ответы в орби�
тофронтальной коре, в то время как прежний УС продолжает провоцировать
* NMDA�рецептор — ионотропный рецептор глутамата, селективно связывающий N�метил �D�
аспартат (NMDA).
** Экспозиционная терапия, или терапия подвергания, — метод поведенческой терапии, включающий
экспозицию или подвергание больного устрашающему его объекту или воздействию в обстановке,
исключающей всякую опасность, и при успокаивании больного психотерапевтом с целью снижения
уровня тревоги и страха (прим. пер.).
Эмоции
149
повышенную активность в правой вентральной миндалине. Орбитофронталь�
ная кора способна быстро перестраивать реакции страха, а миндалина демон�
стрирует устойчивые, неинвертируемые ответы на предыдущие связанные со
страхом стимулы. Видимо, кора и лимбическая система работают по разным
принципам: кора быстро реагирует на новые связи, а лимбическая система бо�
лее консервативна и хранит информацию о старых запускающих стимулах.
Эти сведения говорят о нейробиологической основе «состояния раздвоения
сознания», когда одна часть личности как бы знает, что опасности больше нет,
но другая все равно реагирует на старые сигналы. С такой непроизвольной ре�
акцией, независимой от изменившихся обстоятельств, хорошо знакомы страда�
ющие посттравматическим стрессовым расстройством (ПТСР).
Память, включая и память о страхе, становится постоянной в процессе кон�
солидации, связанной с синтезом белка. При извлечении воспоминания вновь
дестабилизируются и становятся подвержены дальнейшим манипуляциям и
изменениям, предшествующим реконсолидации. Есть данные о том, что ре�
консолидация памяти о страхе у крыс сопряжена с дополнительным синтезом
белка в миндалине (N�d�r et al., 2000). Инфузия антибиотика, прерывающего
синтез белка, устраняет условную реакцию при ее тестировании на следующий
день. У человека до настоящего времени прямых исследований синтеза белка и
реконсолидации памяти опубликовано не было. Но это может быть многообе�
щающим направлением будущих исследований с возможностью найти способ
смягчения симптомов ПТСР.
Наконец, эмоциональные реакции в какой�то степени могут регулироваться
корковыми нисходящими влияниями на миндалину. Переоценка и пересмотр
событий — основные стратегии, с помощью которых человек справляется с
эмоциональным стрессом. Можно переоценить обстоятельства и попытаться
увидеть их в другом свете, как бы имеющими другое значение и влияние на
нашу жизнь. Например, посчитать не в том месте сделанный поворот на шоссе
не досадной оплошностью и потерей времени, а возможностью увидеть что�то
новое. Таким образом, можно изменить собственный эмоциональный настрой:
раздражение сменится любопытством. Переоценка может приводить и к осла�
блению, и к усилению аффекта. Она меняет наше субъективное состояние и
физиологические реакции. В целом, стратегии переоценки активируют одно�
временно как префронтальную и переднюю поясную кору, вовлеченные в ког�
нитивный контроль, так и миндалину, усиливая эмоциональные реакции.
Окснер и его коллеги (Och���r et al., 2004) исследовали фМРТ�корреляты
переоценки предъявленных изображений негативных событий.
Они искали области активации мозга в то время, как испытуемые произ�
вольно увеличивали или уменьшали отрицательные эмоции, связанные с де�
монстрацией изображений неприятных сцен, например, больного в палате. Ис�
пытуемых разделили на две группы по типу предлагаемой стратегии оценки
события. В группе самофокусировки их просили сосредоточиться на внутрен�
нем содержании негативного события и либо усилить отрицательное впечат�
ление, представив себя самого или кого�то из близких в качестве персонажа на
картинке, либо ослабить его, увеличив межличностную дистанцию от собы�
тия, изображенного на рисунке, как бы взглянув на него с отстраненной, объек�
тивной точки зрения. В группе фокусировки на ситуации испытуемых просили
150
Глава 13
оценить аспекты ситуации, либо представив, что дела у больного пойдут на
поправку (ослабление негативного аффекта), либо что все будет только хуже
(усиление негативного аффекта).
И ослабление, и усиление негативных эмоций вовлекало префронтальную и
переднюю поясную области коры, которым часто приписывают роль когнитив�
ного контроля. Активация миндалины увеличивалась или уменьшалась в соот�
ветствии с примененной стратегией оценки. Повышение активности вовлекало
области исключительно ростромедиальной префронтальной коры левого полу�
шария, участвующей в извлечении эмоциональных знаний, тогда как ослабле�
ние аффекта активировало исключительно области латеральной и орбитальной
префронтальной коры правого полушария, участвующей в поведенческом тор�
можении (рис. 13.15).
Кроме того, результаты показали, что «самофокусировка» вовлекает меди�
альные префронтальные области (ПБ 32), участвующие во внутренней обра�
ботке информации и «самоотносимой» оценке. Регуляция при «фокусировке на
ситуации» вовлекала латеральные префронтальные области (ПБ 46), участвую�
щие в поддержании и манипуляции информацией об окружающем мире с по�
мощью рабочей памяти (рис. 13.16).
Эти данные указывают на то, что различные формы переоценки событий
вовлекают как общие, так и уникальные для них нейрональные системы. Раз�
Рис. 13 .15. Усиление и подавление эмоциональных процессов. Активация неокортек�
са при применении стратегии оценки картинок по сравнению с просмотром картинок
без оценки. (а) Активация неокортекса, связанная с усилением отрицательных эмоций.
Слева и по центру — латеральные изображения, справа — медиальный срез. Видна
активация левой дорсолатеральной, медиальной префронтальной и передней поясной
коры. (б) Подавление аффекта. Видна двусторонняя активация латеральной и медиаль�
ной префронтальной коры, включая многие левосторонние регионы — те же, что уча�
ствуют в усилении аффекта. (Источник: Och���r et al., 2004.)
Эмоции
151
личные области префронтальной коры в зависимости от цели и использованной
стратегии по�разному модулируют активность миндалины.
Мы видим, что система страха, порождающая негативные эмоции, имеет
комплексные связи с когнитивными системами в коре больших полушарий.
Теперь обратимся к другой хорошо изученной эмоциональной системе моз�
га — системе ПОИСК по Панксеппу. В исследованиях на человеке эти аспекты
эмоциональной работы мозга обычно называют системами или путями поло�
жительного подкрепления, соответствующими человеческим эмоциям любви
и желания. Взглянем на некоторые примеры взаимодействия когнитивного и
эмоционального аспектов с участием этой системы.
4.0 .
Система положительного подкрепления:
любить, хотеть, учиться
Панксепп (��������, 1���) описал систему ПОИСК как аппетитивную систе�
му внутреннего побуждения, заставляющую млекопитающих интересоваться
окружающим миром и стимулирующую целенаправленное поведение по отно�
шению к целому ряду объектов, таких как пища, укрытие, сексуальный партнер
и т. д . При таком подходе система ПОИСК включает классические пути подкре�
пления в стволе мозга, а также ряд других подкорковых структур. В отличие от
системы СТРАХ, запускающей затаивание, замирание или бегство, обеспечива�
ющие самосохранение при их активации, ПОИСК — несущая положительный
заряд, энергетизирующая система, которая побуждает животных выходить на
взаимодействие с окружающей средой, активно искать корм и исследовать.
В 1�54 г. Олдс (Old�) и Милнер (Mil��r) обнаружили, что крысы обучаются
работать ради электрического раздражения подкорковых зон, включая область
вокруг латерального гипоталамуса. Последующие исследования показали,
что электрическое самораздражение этих зон является мощным подкреплени�
ем. Крысы стимулировали зону латерального гипоталамуса до 2000 раз в час
Рис. 13 .16 . Регуляция при «фокусировке на ситуации». Указаны зоны активации, уни�
кальные для снижения аффекта при стратегиях оценки. Слева: «фокус на ситуации», об�
ратите внимание на ПБ 46 (обведено). Справа: «самофокусировка», обратите внимание
на ПБ 32 (обведено). (Источник: Och���r et al., 2004.)
152
Глава 13
и предпочитали самораздражение пище, вплоть до истощения. Развитие ми�
кроэлектродной техники в 1�70�х гг. позволило все более и более точно карти�
ровать области мозга, отвечающие за подкрепление.
Первичные пути подкрепления у млекопитающих.
1. Мезолимбический дофаминовый путь: дофаминергические нейроны вен�
тральной области покрышки (VTA) ствола мозга проецируются в прилежа�
щее ядро (n. accumbens) переднего мозга.
2. Мезокортикальный дофаминовый путь: дофаминергические нейроны VTA
дают проекции в орбитофронтальную кору (рис. 13.17). VTA располагается
в мосте среднего мозга в окружении других ядер и проводящих путей. При�
лежащее ядро находится рядом с миндалиной, кпереди от нее и ниже стриа�
тума (базальных ганглиев) в каждом полушарии.
4.1.
Новое понимание «подкрепления»:
от подкрепления к предсказанию подкрепления
и далее — к ошибкам предсказания подкрепления
Понимание феномена подкрепления и роли дофаминергических структур сред�
него мозга развивалось со временем. В течение долгого времени считалось, что
дофаминовая система реагирует на получение подкрепляющего стимула, напри�
мер электросамораздражения, внутримозгового введения наркотиков, пищи,
сладкой воды или условных стимулов, предшествующих положительному под�
креплению.
Более поздние исследования показали, однако, что нейроны этой системы
сильнее реагируют на ожидание подкрепления, чем на его получение (Schult�,
1���, 2002). В подтверждение этой гипотезы говорят факты.
Рис. 13.17. Дофаминергические пути системы положительного подкрепления — мезо�
лимбический и мезокортикальный
Эмоции
153
1. Дофаминергические нейроны избирательно разряжаются в ответ на неожи�
данное подкрепление и новые, привлекающие внимание события; при по�
вторном предъявлении ожидаемого подкрепления ответ быстро снижается.
2. Дофаминергические нейроны перестают отвечать на ожидаемое подкрепле�
ние, постепенно начиная реагировать на условные стимулы, предсказываю-
щие его.
3. Дофаминергические нейроны тормозятся пропуском ожидаемого подкре�
пления.
Система подкрепления начинается с безусловных ответов на сенсорные
аспекты подкрепляющих объектов — безусловных стимулов, таких как сладкий
вкус, мускусные запахи и др. Поскольку первичным подкрепляющим событием
на самом деле является поступление дофамина в прилежащее ядро, безуслов�
ные стимулы (БС) сами по себе подкреплением не являются. Они представляют
собой лишь первичные подсказки о предстоящих внутренних процессах. С по�
явлением опыта все более отдаленные, поступающие из окружающей среды
сигналы (условные стимулы, УС) начинают предсказывать появление вкусных
и приятно пахнущих объектов. И эти новые подсказки теперь тоже запускают
дофаминергические реакции в среднем мозге. Со временем оказывается, что
наибольшее удовольствие доставляет не само подкрепление, а стимулы, сооб�
щающие о его будущем получении. Так, в жизни больше радует перспектива
повышения зарплаты или свидания, чем само их «получение» (�i��, 2002).
К сожалению, по крайней мере с точки зрения физиологии дофаминергической
системы, желанный объект после его обретения действительно становится уже
не таким желанным...
Постепенно, в свете все новых данных, концепция предсказания подкрепле�
ния сменилась на точку зрения, что дофаминергические нейроны реагируют на
ошибки в предсказании подкрепления. Например, со временем исчезает акти�
вация в ответ на такие УС, за которыми наверняка следует подкрепление. Но
это исчезновение не связано с общей потерей реакции на само подкрепление,
поскольку дофаминергическая система все так же реагирует на подкрепления
вне тестовой ситуации. Кроме того, ответ дофаминергических нейронов пода�
вляется, когда УС указывает на то, что подкрепление должно появиться, а оно
отсутствует или запаздывает. Поскольку во время ожидания подкрепления ни�
каких новых стимулов не появляется, этот подавленный ответ нельзя считать
реакцией на другой стимул. Подавление дофаминергического ответа, когда
подкрепления пропускаются, рассматривается как реакция на отсутствие пред�
сказанного события.
Экспериментальные данные подтверждают, что реактивность дофаминер�
гических нейронов смещается в сторону таких ситуаций, в которых подкрепле�
ния совершенно непредсказуемы. Доходит до того, что к легко предсказуемому
подкреплению уже не удается «привязать» УС. Давний эмпирический тест для
проверки этой гипотезы — так называемая «парадигма блокирования». В ее
рамках можно показать, что новый стимул не будет ассоциироваться с под�
креплением, если его предъявлять в таких условиях, когда подкрепление и так
возникает в 100% случаев. На обезьянах с помощью микроэлектродной записи
ответов индивидуальных нейронов было получено прямое эмпирическое до�
154
Глава 13
казательство того, что дофаминергическая система действительно вовлекается
в опосредование этого поведенческого обучения.
Исследование Тоблера и его соавторов (Tobl�r et al., 2003), обобщенное Ан�
глессом (��gl���, 2004), показало, что дофаминергические нейроны активиру�
��gl���, 2004), показало, что дофаминергические нейроны активиру�
, 2004), показало, что дофаминергические нейроны активиру�
ются не всеми заметными стимулами. В этой работе обучали обезьян, давая им
награду каждый раз, когда зажигался свет. Затем одновременно со светом стали
включать тон и не давать награду (рис 13.1�), т. е. тон предсказывал отсутствие
подкрепления. При предъявлении только тона дофаминергический нейрон тор�
мозился. Англесс на основании этих данных предположил, что дофаминергиче�
ские нейроны не активируются неспецифически в ответ на заметные стимулы;
активность нейрона модулируется «предсказательной способностью» стиму�
лов, а не просто наличием подкрепления.
В настоящее время теория ошибки предсказания подкрепления является
наиболее широко принятым объяснением механизма работы дофаминергиче�
ской системы положительного подкрепления.
4.2 .
Подкрепление — это больше чем обучение
Теория ошибок предсказания подкрепления делает основной упор на аспектах
подкрепления, связанных с обучением. Шульц (Schult�, 2002), цитируя ранние
взгляды Торндайка, подчеркивал роль системы положительного подкрепления
во «впечатывании» ассоциаций между условными стимулами и их последстви�
ями. В ответ на это ряд других исследователей (B�rridg�, Robi��o�, 1���, 2003;
Рис. 13 .18 . Пример торможения одиноч�
ного дофаминергического нейрона стиму�
лом, предваряющим отсутствие подкре�
пления. В верхней части каждого рисунка
находится гистограмма общего количества
потенциалов действия с течением времени
(каждый столбец — 10 мс) при повторном
предъявлении стимулов; штрихи под каж�
дой гистограммой обозначают отдельные
потенциалы действия в ходе каждой се�
рии. Стимул А сочетается с подкреплени�
ем (А+ ) и возбуждает дофаминовый ней�
рон. При одновременном предъявлении А
и Х подкрепления не поступает (АХ–) и,
таким образом, Х предваряет отсутствие
подкрепления. При предъявлении только
Х дофаминергический нейрон тормозится
(Х–). Контрольные стимулы также предъ�
являли, но они на рисунке не показаны.
(Источник: ��gl���, 2004; из Tobl�r et al.,
2003.)
Эмоции
155
��������, 2005) отметил, что теория предсказания подкрепления обходит вни�
манием важные гедонические и мотивационные компоненты подкрепления, ко�
торые не должны быть проигнорированы.
Показано, что гедоническое ощущение «нравится» разные виды млекопита�
ющих демонстрируют сходным лицевым выражением, не зависящим от дофами�
новой системы. Реакции предпочтения/симпатии модулируются сетью мозговых
структур, включающей оболочку (скорлупу) прилежащего ядра, вентральную
часть бледного шара и парабрахиальное ядро ствола (B�rridg�, Robi��o�, 2003;
рис. 13.1�). Реакции предпочтения сладкой воды у крыс не подвержены влиянию
активации или подавления со стороны мезолимбической дофаминовой системы,
но усиливаются при микроинъекции опиоидных или ГАМК агонистов. Разруше�
ния, снижающие уровень дофамина в прилежащем ядре и приводящие к выра�
женной афагии (нежеланию есть), не нарушают вкусовых предпочтений (�yv�ll
��d B�rridg�, 2000). Антагонисты дофаминовых рецепторов зачастую не могут
подавить субъективное удовольствие от амфетаминов или сигарет у человека.
И наконец, активация дофаминергической системы амфетаминами у человека
лучше коррелирует с субъективным уровнем желания наркотика или пищи, чем
с субъективным уровнем удовольствия (L�yto� et al ., 2002). С субъективной точ�
ки зрения, дофаминовая система делает пищу и наркотики более желанными и
соблазнительными, но не делает их приятнее, аппетитнее или вкуснее, когда они
уже получены. Привлекательность явно зависит от отдельной системы, включа�
ющей оболочку (скорлупу) прилежащего ядра.
Рис. 13 .19 . Упрощенная схема подкорковых путей предпочтения и желания в мозге
крысы. Пути «предпочтения» показаны зелеными стрелками, пути «желания» — жел�
тыми, когнитивная обработка признаков отмечена голубым. (Источник: B�rridg� ��d
Robi��o�, 2003.)
156
Глава 13
В свете этих данных будет логичным разделять обучающие аспекты подкре�
пления, т. е. способность подкрепляемого поведения закрепляться, и гедониче�
ские аспекты предпочтения. Гомология в поведенческих реакциях на различ�
ные вкусовые стимулы позволяет оценить гедоническое влияние этих стимулов
(B�rridg�, 2000). Среди сходных признаков «склонности и предпочтения» у раз�
личных видов млекопитающих — высовывание языка при опробовании на вкус
сладкой воды. Среди характерных реакций отвращения — наоборот, широко
открытый рот в ответ на горький вкус хинина (рис. 13.20).
Третьим компонентом системы подкрепления является элемент желания
или побуждения (B�rrig� ��d Robi��o�, 1���). Желание отделимо от склонности/
предпочтения и обучения как субъективно, так и неврологически (рис. 13.21).
Мы можем хотеть чего�то, например, наркотиков или сигарет, даже если они
нам не нравятся. Обучение может мотивироваться желанием, но может прохо�
дить и без него. Соответственно, обучение может проходить и без субъективно�
го ощущения «нравится», как в случае обученного получения внутримозговых
введений наркотика за нажатие на рычаг без сопровождающего чувства под�
крепления.
Желание «превращает простую сенсорную информацию о подкреплении и
его признаках (образах, звуках и запахах) в привлекательные, желанные и за�
хватывающие побуждения» (B�rridg� ��d Robi��o�, 2003). В системе поощре�
��d Robi��o�, 2003). В системе поощре�
Robi��o�, 2003). В системе поощре�
ния компонента желания, в отличие от компоненты предпочтения (у человека,
страсти) сильно зависит от мезолимбического дофаминергического пути.
Рис. 13 .20 . Реакции предпочтения: объективые показатели гедонических аспектов эмо�
ции. Гомологичные аффективные лицевые выражения у младенца, детеныша орангу�
тана и взрослой крысы в ответ на «понравившуюся» сахарозу (вверху) и «не понравив�
шийся» хинин (внизу). (Источник: B�rridg� ��d Robi��o�, 2003.)
Эмоции
157
4.3 .
«Путь подкрепления» и наркотики
Все наркотики влияют на систему «желания» через дофаминергические пути
и их связи в среднем мозге (рис. 13.22). Животные обучаются нажимать рычаг
ради получения внутримозговых микроинъекций наркотиков и вырабатывают
условное пространственное предпочтение тех мест в клетке, где они их полу�
чали. Поведение таких животных в подобных экспериментах позволило опре�
делить, какие области мозга отвечают за подкрепляющие свойства кокаина,
амфетамина, никотина, алкоголя и опиатов (S����g�l ��d ��i��, 1���; I��moto
��d �i��, 2004). У человека исследовали влияние различных агонистов и анта�
�i��, 2004). У человека исследовали влияние различных агонистов и анта�
�i��, 2004). У человека исследовали влияние различных агонистов и анта�
, 2004). У человека исследовали влияние различных агонистов и анта�
гонистов наркотиков на лиц, страдающих наркотической зависимостью.
Естественные виды подкрепления и наркотики по�разному действуют на
мезолимбическую дофаминергическую систему. Еда, а также опиоиды косвен�
но влияют на систему положительного подкрепления, снижая активность тор�
мозных интернейронов, в норме ингибирующих дофаминергические нейроны
вентральной области покрышки; в результате активность этих нейронов воз�
растает.
Кокаин и амфетамин, напротив, напрямую действуют на прилежащее ядро,
поддерживая высокий уровень дофамина в синапсах соответствующих ней�
ронов; кокаин ингибирует обратный захват дофамина, а амфетамин заметно
повышает его выброс. Тонкости механизмов действия различных наркотиков
Рис. 13 .21 . Схема психологических компонентов желания, обучения и предпочтения и
измеряемых видов поведения, используемых для их исследования. (Источник: B�rridg�,
200�.)
158
Глава 13
все еще исследуют. Однако уже ясно, что указанные в табл. 13.2 вещества вли�
яют на поведение и субъективные ощущения через воздействие на мезолим�
бическую дофаминергическую систему (S����g�l, ��i��, 1���; I��moto, �i��,
2004).
4.4 .
Сигналы подкрепления влияют на внимание
В соответствии с теорией ошибок предсказания подкрепления, сигналы, указы�
вающие на доступность наркотика, быстро становятся условными стимулами
и вызывают у наркомана сильную тягу. Такие сигналы не только активируют
дофаминергическую систему и создают субъективное ощущение потребности,
но и влияют на когнитивные функции. Потребление наркотиков сдвигает вни�
мание к связанным с ними сигналам в ущерб всем остальным стимулам. Мно�
гочисленные исследования с использованием вышерассмотренной парадигмы
Струпа показали, что связанные с наркотиками слова или изображения отвле�
кают внимание от задания по называнию цветов у потребляющих наркотики
испытуемых, в отличие от не принимающих (H��t�r et al. 2006).
Соответствующие исследования с использованием визуализации мозга по�
казали, что у курящих испытуемых связанные с курением изображения вы�
зывают значительно более сильную реакцию в прилежащем ядре и соседнем
вентральном стриатуме, чем у некурящих, хотя в реакциях на нейтральные изо�
бражения разницы между группами не отмечается (D�vid et al., 2005).
Рис. 13 .22 . Препараты и их влияние на систему «желания». Прямая и косвенная ак�
тивация мезолимбической системы — дофаминергических нейронов вентральной по�
крышки (VTA) — естественным положительным подкреплением и препаратами. До�
фаминергические нейроны группы A10 находятся в VTA и дают проекции в оболочку
прилежащего ядра (NAC). ГАМК�ергические интернейроны в VTA и длинная петля об�
ратной связи с участием ГАМК�ергических нейронов NAC осуществляют тормозный
контроль (минусы на рисунке) нейронов А10. Различные дофаминовые рецепторы NAC
(D1 и D2) опосредуют эффекты подкрепления. (Источник: S����g�l ��d ��i��, 1���.)
Эмоции
159
Таблица 13.2
Вещества и их влияние на поведение: области мозга,
вовлеченные в подкрепляющее действие
естественного положительного подкрепления и наркотиков
Агент
Область воздействия
Пища, питье,
секс
Вызывают высвобождение дофамина в вентральной области по�
крышки (V TA ), ингибируя ГАМК�ергические интернейроны, ко�
торые обычно сами ингибируют дофаминергические нейроны
Мю� и дель�
та�опиоиды
Опиоиды воздействуют косвенно посредством торможения ГАМК�
ергических нейронов в V TA и NAC (прилежащее ядро n. accum-
bens), которые обычно ингибируют дофаминергические нейроны
Амфетамин
Напрямую повышает выброс дофамина в прилежащем ядре
Кокаин
Напрямую блокирует систему транспорта/обратного захвата до�
фамина в оболочке (скорлупе) прилежащего ядра
Фенциклидин
(�C�)
Напрямую блокирует NMDA�рецепторы в оболочке (скорлупе)
прилежащего ядра
Каннабис
Действует в задней V TA и оболочке (скорлупе) прилежащего ядра
Этанол
Действует косвенно через заднюю V TA , тормозя ГАМК�
ергические интернейроны
Никотин
Прилежащее ядро/вентральный стриатум
5.0 .
Заключение
В мозге млекопитающих существуют несколько эмоциональных систем. У каж�
дой из них есть своя врожденная схема ответа на связанные с выживанием
стимулы, поступающие из характерной для данного вида окружающей среды.
У таких, как система страха или подкрепления, есть условные и безусловные
реакции на значимые «условия вызова», поддерживаемые обособленными ней�
ронными сетями: страх обеспечивает миндалина и связанные с ней структуры,
положительное подкрепление зависит главным образом от мезолимбических и
мезокортикальных путей к V TA. Каждая из этих систем в той или иной степени
поддается когнитивному контролю и, в свою очередь, сама влияет на высшие
функции принятия решений, оценки событий и сознания. Каждая из систем
формирует субъективно четко различимые эмоции.
6.0.
Задания и упражнения к главе 13
6.1.
Контрольные задания
1. В чем заключаются ключевые различия между мозгом пресмыкающихся и
млекопитающих?
160
Глава 13
2. Приведите приметы «прошитых» эмоций, описанных Панксеппом.
3. Почему система страха изучена лучше других?
4. Что означают термины «верхний путь» и «нижний путь» при описании моз�
говых систем страха?
5. Каким образом исследование аффективного слепозрения помогает понять
обработку эмоциональной информации мозгом?
6. Какие факторы разделяют «подкрепление» и «обучение» в исследовании
ПОИСК?
6.2 . Графические упражнения
1. Обозначьте на рис. 13.23 области триединого мозга.
2. Обозначьте на рис. 13.24 афферентные связи миндалины.
3. Обозначьте на рис. 13.25 эфферентные связи миндалины.
Рис. 13.23
Эмоции
161
Рис. 13 .25
Рис. 13 .24
Человек — животное социальное.
Приписывают Аристотелю
Если вам кажется, что эту же картину можно увидеть и у людей, то это потому, что наш
социальный мозг имеет много сходств и совпадений с мозгом других приматов. Два
больших полушария мозга на рисунке ниже свидетельствуют о том, что социальная
когнитивная деятельность часто активирует верхнюю височную борозду (ВВБ) в мозге
человека. (Источник: de Waal, 2004 (верхний рисунок); Allison et al., 2000 (нижний ри-
сунок).)
Содержание
1.0.Введение.......................................................163
1.1 . Термины, используемые в отношении
социальнойкогнитивнойдеятельности...........................164
1.2.Важностьперспективы:первое,второеитретьелица............... 165
1.3 . Подходы к восприятию ментального состояния других людей. . . . . . . . 165
2.0 . Организационная основа для социальной когнитивной деятельности. . . . . . 168
2.1.Намерение...................................................168
2.2.Распознаваниеглаз............................................169
2.3.Разделенноевнимание.........................................170
2.4.Теорияразуманаивысшегопорядка..............................170
3.0.Зеркальныенейроныивосприятиенамерений........................171
3.1.Отдействийкнамерениям.....................................171
3.2.Распознаваниеглазивосприятиенаправлениявзора ............... 178
3.3.Разделенноевнимание.........................................180
3.4.СпособностивысокогопорядкаизТР............................182
3.5. Восприятие других как похожих и непохожих: Я-Оно в мозге . . . . . . . . 187
3.6.Восприятиелиц..............................................190
4.0.Заключение.....................................................192
5.0.Заданияиупражнениякглаве14...................................193
5.1.Контрольныезадания..........................................193
5.2.Графическиеупражнения......................................193
1.0 . Введение
В этой главе мы познакомимся с одним из аспектов когнитивной деятельности
человека, который отличает нас от других млекопитающих — способностью
понимать друг друга как сознательных существ с собственным внутренним
миром. У других представителей млекопитающих есть частичная способность
подстраиваться под психологические состояния членов своего вида (табл. 14 .1).
Они воспринимают разновидности поз, вокализации и лицевой экспрессии как
сигналы и координируют свои действия на их основе. У некоторых приматов
обнаружены «зеркальные нейроны», которые позволяют им определять общ-
ность своих действий и действий других обезьян. Однако ни для каких видов
животных не показана способность понимать, использовать или зависеть от
субъективных действий других представителей вида. Это является исключи-
тельно человеческой особенностью.
Глава 14
Социальная когнитивная
деятельность: восприятие
ментального состояния
других людей
164
Глава 14
«Вопрос чужих умов» не нов для философского круга. Однако психологи-
ческие и нейробиологические исследования нашего понимания мысленных
состояний достаточно современны. Пока еще психологи и нейробиологи не
пришли к согласию по поводу использующихся общих определений терминов
для описания и объяснения социального познания. Поэтому далее, прежде чем
начать изучение социальной когнитивной деятельности, познакомимся с неко-
торыми терминами.
Таблица 14.1
Восприятие в социальной когнитивной деятельности и в науке
Восприятие
от первого лица
Восприятие
от второго лица
Восприятие
от третьего
лица
Тип данных
Феноменологиче-
ские данные
Сопереживающие осоз-
нание
Обычные эмпи-
рические данные;
чувственные
данные
Метод сбора
данных
Контролируемая
интроспекция
внутренних состоя-
ний; традиционная
психофизика
Отражение при помощи
эмпатической сонастро-
енности; внутреннее
сканирование эмоцио-
нальных и когнитивных
ответов при наличии
другого субъекта
Объективное
наблюдение
за внешними
объектами
Лингвистические
маркеры
Я чувствую, Я знаю Ты думаешь, Ты выгля-
дишь
Оно есть,
Он/она есть
1.1 .
Термины, используемые в отношении
социальной когнитивной деятельности
В исследовательской литературе термины, относящиеся к аспектам социальной
когнитивной деятельности, очень часто взаимозаменяемы и используются раз-
ными способами разными учеными.
Под эмпатией понимают чувство восприятия чувств других. В латинском
языке это слово означает «чувствовать внутри» или «чувствовать с». В то же
время теория разума (ТР, см. раздел 1.3) часто используется для того, чтобы
подчеркнуть идею, что мы обычно имеем комплексное метакогнитивное пони-
мание как нашей мыслительной деятельности, так и мышления других, вклю-
чая когнитивные и чувственные аспекты. Фрис и Фрис (Frith and Frith, 1999)
представили схожий термин «ментализация» для обозначения идеи, что если у
нас есть развитая теория разума, то мы воспринимаем себя и других не только
как сенсорных объектов, но и как субъективных индивидов с психическим со-
стоянием. Мы можем предвидеть и использовать для направления нашего по-
ведения психическое состояние окружающих нас субъектов. Чтение мыслей,
так же как и ментализация, определяет нашу способность подстраивать наше
поведение к состоянию и предвиденным действиям других.
Философы используют термин интенциональность, когда говорят о том, что
наш разум и ментальные состояния всегда «о чем-то другом», а физические
объекты, например части тела, нет. Наши мысли всегда имеют какой-то объ-
Социальная когнитивная деятельность: восприятие состояния других людей
165
ект. Например, мы можем думать «о» стуле, книге или идее так, как не может
наш желудок, рука или зуб. Разум всегда имеет умственное состояние; разум
представляет объекты и события вне себя. Неясно, понимают ли другие виды
умышленную природу разума представителей своего вида. У человека суще-
ствует врожденное понимание содержания умственной деятельности других.
Совершенно другой смысл имеет термин «преднамеренность», т. е. спо-
собность формировать образ конечной цели и строить поведение для ее до-
стижения. Будьте осторожны, не перепутайте эти два очень похожих термина!
Способности теории разума позволяют нам понимать намерения других и раз-
делить с ними внимание к общему фокусу.
И наконец, термин интерсубъективизм подчеркивает нашу способность ко-
ординировать взаимное общение в свете нашего представления о субъективиз-
ме и интенциональности других. Когда эта способность отсутствует, мы легко
видим нарушения социального обмена у других. Примеры можно найти в за-
болеваниях аутистического спектра, иногда в трудности распознавания эмоций
при шизофрении и в отсутствии сострадания у психопатов и при пограничном
личностном расстройстве.
1.2.
Важность перспективы: первое, второе и третье лица
Научный эксперимент обычно строится с перспективы третьего лица. Это зна-
чит, что исследователи принимают объективную точку зрения, рассматривают
все доказательства, будучи физическими объектами. Даже люди воспринима-
ются в качестве объектов, чувственных поверхностей. Внутренний умственный
мир также рассматривается при помощи объективных методов как наблюдение
за поведением или нейровизуализация. Недавно ученые, интересующиеся со-
знанием, стали обсуждать дополнительный способ научного познания, который
позволяет собирать информацию с перспективы первого лица, т. е. использо-
вать феноменологические данные от интроспекции или самоотчета. До опреде-
ленного предела, самоотчет под контролируемыми обстоятельствами является
достаточно устоявшимся научным методом, например в психофизике и при из-
учении восприятия.
А что происходит с перспективой второго лица? В данном случае другой
человек воспринимается как субъект, имеющий определенные умственные со-
стояния, а не просто объект. Эта перспектива мало используется в психологии
и нейронауке, но хорошо известна в философии и клинический психологии.
В настоящее время исследование социальной когнитивной деятельности подхо-
дит все ближе к рассмотрению проблем с перспективы второго лица. Но более
точно можно сказать, что мы воспринимаем перспективу третьего лица (объ-
ективный взгляд) на других людей в то время, как они выполняют деятельность
второго лица.
1.3.
Подходы к восприятию
ментального состояния других людей
Как только нам исполняется четыре года (независимо от того, нормально мы
развиваемся или имеем отставание в развитии), мы подаем признаки пони-
мания мыслей других людей — этот процесс Крис Фрис (Chris Frith) назвал
«ментализацией». Мы становимся способными узнавать и реагировать в ответ
на невидимые, внутренние субъективные закономерности, которые стоят за
166
Глава 14
поведением других людей. Мы будем называть полностью развившиеся спо-
собности понимать и предсказывать наши собственные и чужие мысли тео-
рией разума (ТР). ТР может быть рассмотрена с точки зрения трех принципов:
модульного принципа, теории теорий и имитационной теории.
Например, согласно модульной теории, принадлежащей Саймону Барон-Коэну
(�. �aron-Cohen), модуль теории разума (МТР) у людей формируется отдель-
�. �aron-Cohen), модуль теории разума (МТР) у людей формируется отдель-
�aron-Cohen), модуль теории разума (МТР) у людей формируется отдель-
�aron-Cohen), модуль теории разума (МТР) у людей формируется отдель-
-Cohen), модуль теории разума (МТР) у людей формируется отдель-
Cohen), модуль теории разума (МТР) у людей формируется отдель-
, модуль теории разума (МТР) у людей формируется отдель-
но, но основывается на мыслительных способностях, общих для людей, нече-
ловекообразных обезьян и других млекопитающих. Только у человека может
полностью развиться МТР. Эта теория очень хорошо согласуется с данными,
полученными при изучении аутизма.
Социальный мозг
Традиционно областью исследования когнитивной
нейронауки являлись люди и их мозг в отдельности,
но за последние несколько лет наблюдается чрезвы-
чайное увеличение интереса к происходящему при
взаимодействии людей друг с другом. В связи с этим
появились два модных слова: теория разума, или спо-
собность читать мысли другого человека, и зеркаль-
ные нейроны, нейроны, которые отвечают разрядом
на наши действия и активируются, когда мы видим,
что это действие выполняет кто-то другой. Для меня
самым поразительным результатом всех этих иссле-
дований является понимание, насколько сильно мы
погружены в наш общественный мир. Мы можем чув-
ствовать себя независимыми представителями, но на
самом деле нас постоянно координирует так или иначе
поток социальных сигналов, на которые мы отвечаем
независимо от нашего желания. В большинстве случа-
ев мы даже не осознаем влияние этих сигналов.
Эта идея подтверждена результатами многих экспе-
риментов, но у меня есть две любимые работы для
иллюстрации таких автоматизированных ответов.
В первой работе (�lakemore et al., 2005) использова-
ли фМРТ для того, чтобы посмотреть, что происходит,
когда мы видим, как прикасаются к другим людям. Добровольцев сканировали, в
то время как они наблюдали, как прикасаются к лицам других испытуемых, а так-
же при прикосновениях к их лицам в тех же точках. Полученные результаты (рис.
14.2) четко демонстрируют, что одна и та же область соматосенсорной (тактильная
информация от тела) коры активировалась как при прикосновениях к испытуемым,
так и в случае, когда они видели дотрагивания до других. Локализация активности
зависела от места прикосновения: лицо или шея, правая сторона или левая. Дан-
ный эффект отсутствовал при наблюдении за прикосновениями к неодушевленным
предметам, например при дотрагавании к электрическому фену в форме головы.
В большинстве случаев люди не отдают себе отчет о том, что они воспринимают
прикосновения к другим. Только в очень редких случаях синестезии эта активность
пробивается к сознанию, и пациенты говорят: «когда я вижу, как к чьему-то лицу
прикасаются, я чувствую это прикосновение у себя на лице».
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
Рис. 14 .1 . Кристофер
Фрис (Christopher Frith,
PhD), Wellcome Trust
Centre for Neuroimaging,
Университетский
кол-
ледж Лондона, Велико-
британия
Социальная когнитивная деятельность: восприятие состояния других людей
167
Во втором эксперименте (Liepelt et al., 2009) использовали классическую психоло-
гическую методику измерения времени реакции. В данном исследовании испытуе-
мые должны были нажимать на кнопки первым (указательным) или вторым (сред-
ним) пальцами как можно быстрее. Сигнал к началу выполнения предъявлялся на
экране в виде изображения руки (рис. 14 .3). Цифры 1 и 2, появлявшиеся между пер-
вым и вторым пальцами, указывали, на какую кнопку надо нажимать. Уникальной
особенностью данной работы было то, что в некоторых случаях первый и второй
пальцы на предъявляемых картинках придерживались металлическими скобками.
В этих случаях время реакции испытуемых было значительно больше, несмотря на
то, что их пальцы были абсолютно свободны. Этот эффект был очень специфичным.
Только реакция тех пальцев, которые были фиксированы на рисунке, замедлялась.
Мы не сознаем этого, но лишь один вид того, что происходит с окружающими нас
людьми, изменяет наши чувства и поведение. Вероятно, мозг — это исходно со-
циальный орган.
Рис. 14 .2 . Данные области мозга акти-
вируются, когда вы чувствуете прикос-
новения к своему лицу или когда видите,
как дотрагиваются до лица другого че-
ловека. �I — первичная сенсомоторная
кора. (Источник: �lakemore et al., 2005/)
Рис. 14 .3 . Экспериментальная модель и результаты для задачи на реакцию паль-
цев. (Источник: Liepelt et al., 2009.)
168
Глава 14
Литература
1. �lakemore, �.J ., �ristow, D., �ird, G., Frith, C. and Ward, J. (2005). �omatosensory
activations during the observation of touch and a case of vision-touch synaesthesia.
Brain, 128 (7), 1571–1583.
2. Liepelt, R., Ullsperger, M., Obst, K., �pengler, �., von Cramon, D.Y. And �rass, M.
(2009). Contextual movement constraints of others modulate motor preparation in the
observer. Neuropsychologia, 47 (1), 268–275.
Теория теорий предполагает развитие ТР как примитивных, внутренних
принципов в цикле развития, почти как в теории диалога Пиаже. Эта теория
предсказывает внезапные изменения в поведении при получении нового зна-
ния, что и наблюдается у детей в возрасте от трех до четырех лет.
Согласно имитационной теории, мы понимаем мысли других людей при
помощи внутренней имитации или «проигрывания» возможных мысленных
состояний других людей в разных ситуациях. Наличие ответной реакции зер-
кальных нейронов как на собственные, так и на чужие действия можно считать
подтверждением теории имитации.
Вполне вероятно, что все три теории необходимы для объяснения челове-
ческой способности к «ментализации». Как мы узнаем далее, существуют от-
дельные умения, развивающиеся у млекопитающих и человека, и работающие
отдельные модули. Мы можем потерять один модуль, но у нас все равно оста-
нутся другие. Система, позволяющая нам подражать другим, функционирует,
вероятно, через внутреннюю имитацию действий других. И наконец, взрослые
люди имеют утонченные способности к социальному восприятию, которые по-
зволяют им делать умозаключения о внутренних состояниях окружающих лю-
дей. Мы действуем так, будто имеем сложный набор правил о собственных и
чужих умственных состояниях, который можно назвать имплицитной теорией.
2.0 .
Организационная основа
для социальной когнитивной деятельности
Симон Барон-Коэн выдвинул предположение, что полностью развитая теория
разума состоит из четырех типов навыков, которые развиваются независимо
(�imon �aron-Cohen, 1995). Этими навыками являются детекция чужих наме-
рений, распознавание направления взора, разделенное внимание и сложная си-
стема внутренних знаний об окружающих, называемая модулем теории разума.
Некоторые из этих навыков присутствуют как у млекопитающих и нечеловеко-
образных обезьян, так и у людей. Однако только у нормально развивающихся
людей появляется модуль теории разума. Сначала мы рассмотрим модель Ба-
рона-Коэна (рис. 14.4), а затем будем использовать ее как основу для система-
тизации большого объема данных исследований по социальной когнитивной
деятельности.
2.1.
Намерение
Первым компонентом ТР является детектор намерений (ДН). Это способность
воспринимать намерения или целевые действия во многих формах биологиче-
ских и небиологических движений. Например, когда мы смотрим, как кружат-
ся листья на автомобильной стоянке, у нас появляется ощущение, что листья
Социальная когнитивная деятельность: восприятие состояния других людей
169
«хотят собраться вместе». Мы приписываем определенную цель куче листьев.
Или когда смотрим, как передвигают пластилиновых персонажей по нарисо-
ванной местности в пластилиновых мультфильмах, мы с готовностью припи-
сываем кусочкам пластилина определенные намерения и другие умственные
состояния. Аналогично, когда мы видим людей или животных, вовлеченных в
какую-либо деятельность, мы понимаем их цели и желаемый результат их дей-
ствий. Мы интерпретируем действие как намерение.
2.2 .
Распознавание глаз
Вторым компонентом модели является детектор направления взора (ДНВ), т. е.
способность выделять глаза и стимулы, похожие на них, и определять направ-
ление взора. У многих млекопитающих есть способность собирать и использо-
вать информацию о направлении взора. Например, кошки используют взгляд
как часть доминирующего поведения; подчиненные кошки должны отводить
взгляд от доминантной кошки. Люди с первых часов жизни ищут и фокуси-
руются взглядом на глазах своих воспитателей. У нас есть также тенденция
Рис. 14 .4 . Схематическая диаграмма модели теории разума, созданной Бароном-Коэ-
ном, содержит детектор направления взора в верхнем правом углу рисунка, направля-
ющий информацию к детектору намерений в верхней левой части и к механизму раз-
деленного внимания в центре. МРВ также получает входы от ДН и взаимодействует
с модулем теории разума. (Источник: �aron-Cohen, 1995.)
170
Глава 14
находить стимулы, похожие на глаза, в неживых объектах, например лицо на
поверхности луны и у машины, на сучковатых стволах деревьев и неровностях
гор. «Язык глаз», похоже, является фундаментальным способом обмена мыс-
ленными состояниями среди людей.
Вместе детектор намерения и детектор направления взора вовлечены в диа-
дические (двусторонние) взаимодействия. В этом случае один человек вос-
принимает, а другой является объектом восприятия. Но на этой стадии еще
не обязательно наличие обмена мысленными состояниями. И ДНВ, и ДН об-
наруживаются у животных как часть их способностей социального общения.
Третий модуль ТР является исключительно частью человеческой социальной
когнитивной деятельности.
2.3 .
Разделенное внимание
Механизм разделенного внимания (МРВ) — приобретаемая к концу первого
года жизни способность понимать, что если кто-то меняет направление взгляда,
то он «смотрит» на какой-то объект. Мы узнаем, что «смотреть» значит «ви-
деть», и это является дополнением к простой способности воспринимать на-
правление взора. Мы понимаем, что можем посмотреть туда же и увидеть тот
же объект. Изменение направления взгляда и показывание пальцем являются
способами привлечения внимания собеседника.
Младенцы до года, а также большинство приматов и других млекопитаю-
щих не способны к разделенному вниманию. Вы можете наблюдать это у ва-
ших любимых питомцев. В то время как домашняя собака способна поймать
мяч и принести его обратно, она не будет следовать взглядом за вашим взо-
ром, когда вы посмотрите в направлении мяча, лежащего в траве. Также она не
последует в направлении, показанном пальцем, когда вы попытаетесь указать
местоположение мяча. Собаки обладают значительным интеллектом, но у них
нет разделенного внимания. Аналогично ребенок в возрасте 6 месяцев не бу-
дет следовать глазами за взглядом родителей, в то время как годовалый малыш
будет. Способность к разделенному вниманию — отличительная особенность
человека.
2.4 .
Теория разума наивысшего порядка
Финальным компонентом полностью развившейся теории разума, по мнению
Барона-Коэна, является модуль теории разума (МТР) — комплексная база зна-
ний, содержащая правила социального общения, которые развиваются ко вре-
мени достижения нами четырехлетнего возраста.
• Видимость и реальность не всегда совпадают, например камень может
иметь форму яйца, но яйцом он не является. Я могу притворяться соба-
кой, но ею не являюсь.
• Человек, сидящий на стуле, может быть чем-то занят, т. е. думает, пред-
ставляет или вспоминает что-то (маленькие дети не понимают этого).
• У других людей есть мысленные состояния, так же как и физические со-
стояния.
• Я могу знать то, чего не знают окружающие; Я могу одурачить или обма-
нуть их; Я понимаю смысл игры в прятки.
• Мое умственное состояние в прошлом отличается от такого в настоящем.
Социальная когнитивная деятельность: восприятие состояния других людей
171
• Выражения лиц являются отражением мысленных состояний, так же
как и физического состояния; Я могу отличить выражение удивления
от зевания.
ТР не является аналогом интеллекта или уровня I�. Дети, отстающие в раз-
I�. Дети, отстающие в раз-
. Дети, отстающие в раз-
витии, и взрослые демонстрируют полноценные способности ТР, несмотря на
низкий уровень интеллекта, в то время как люди, страдающие аутистическими
нарушениями, могут иметь высокий уровень I�, но значительно поврежденные
способности ТР.
3.0 .
Зеркальные нейроны и восприятие намерений
Где в мозге происходит социальная когнитивная деятельность? Некоторые со-
временные исследования системы зеркальных нейронов на макаках пролили
свет на локализацию социальных процессов в мозге. Первые исследования
при помощи методов нейровизуализации, таких как фМРТ, наличия подобной
системы у человека дали удивительные результаты, подтверждающие возмож-
ность ее существования у людей. Однако более новые работы бросают тень на
эти ранние результаты. Существование зеркальных нейронов у человека до сих
пор является спорным и горячо обсуждаемым фактом в области социальной
когнитивной нейронауки. Мы начнем с рассмотрения работ, сделанных на ма-
каках, а затем познакомимся с данными существования зеркальных нейронов у
человека.
3.1.
От действий к намерениям
Система зеркальных нейронов (СЗН) — группа корковых нейронов, которые,
как считается, позволяют понимать намерения других, просто наблюдая за со-
вершаемыми ими действиями. Зеркальные нейроны были впервые обнаружены
в лобной коре макак, и вскоре после этого они были найдены и в теменной
коре. Эти нейроны обладают уникальным свойством: отдельный зеркальный
нейрон отвечает разрядом не только при восприятии определенного действия,
но и при выполнении этого же действия наблюдателем. Исследователи сразу
же ухватились за возможность того, что зеркальные нейроны служат средством
сравнения собственных действий с действиями окружающих. В дополнение к
тому зеркальные нейроны рассматривают как основу для прогнозирования це-
лей и намерений других посредством внутреннего сравнения образов действий
окружающих с собственным репертуаром активности.
Исходная работа о зеркальных нейронах была сделана Риззолатти (Ri��o-
Ri��o-
latti) с коллегами в университете города Парма в Италии в начале 1990-х гг.
Они обнаружили зеркальные нейроны во фронтальной моторной области коры
обезьяны в поле F5. При помощи регистрации одиночных нейронов коры мозга
макак они зафиксировали, что такие нейроны отвечали разрядом, когда экспе-
риментатор брал орехи руками с подноса, а также когда сама обезьяна брала эти
орехи; нейрон не отвечал, если орех просто лежал на подносе или его брали при
помощи пинцета. Очевидно, что этот нейрон «настроен» отвечать разрядом на
акт хватания пищи руками. На рис. 14 .5 показана обезьяна с устройством для
микроэлектродной регистрации на голове; она сидит в аппарате. На рис. 14 .6
изображена экспериментальная схема, использовавшаяся Риззолатти с коллега-
ми для регистрации ответов одиночных нейронов при разных условиях.
172
Глава 14
Для того чтобы убедиться, что ответы нейронов были не артефактом от вза-
имодействия обезьяны и экспериментатора или ожидания пищи, исследователи
также зарегистрировали нейронные ответы обезьяны, когда она смотрела на
процессы хватания пищи, происходящими между экспериментатором и второй
обезьяной. Ответы зеркальных нейронов были такими же, т. е . они отвечали
на акт хватания второй обезьяной пищи, и молчали при передвижении пищи
и опять разряжались, когда вторая обезьяна брала еду (рис. 14 .7).
Исследования на макаках были сделаны методом регистрации одиноч-
ных нейронов в префронтальной коре. Существование зеркальных нейронов
у обезьян считается доказанным. Аналогичных исследований с регистрацией
одиночных нейронов на людях проведено не было; данные о системе зеркаль-
ных нейронов (СЗН) человека получены при помощи нейровизуализации ак-
тивности множества нейронов. Таким образом, индивидуальные зеркальные
нейроны у человека не исследовались. Тем не менее существует значительное
сходство между системой зеркальных нейронов, исследованной при помощи
Рис. 14 .5 . Обезьяна с микроэлектродом, присоединенным к ее голове, выполняет за-
дания внутри аппарата для фМРТ. (а) микроэлектрод; (б) катушки фМРТ; (в) колыбель.
(Источник: Tammer et al., 2006.)
Социальная когнитивная деятельность: восприятие состояния других людей
173
Рис. 14 .6 . Наблюдаемые действия и оди-
ночные ответы зеркальных нейронов.
(а) Экспериментатор размещает кусочки
пищи на доске, пододвигает их к обезья-
не, которая хватает еду. Изображение на
верху картинки демонстрирует данное
поведение. Индивидуальные нейронные
ответы в течение этого времени отобра-
жены посередине. А внизу этого рисунка
находится гистограмма с общим числом
нейронных разрядов в каждом временном
отрезке по 20 мс. Обратите внимание на
многочисленность ответов, когда экспе-
риментатор берет в руки еду, отсутствие
разрядов во время передвижения доски,
и второй всплеск активности нейронов
когда обезьяна сама берет пищу. (б) Экс-
периментатор берет пищу при помощи
пинцета, кладет на доску, пододвигает ее
к обезьяне, и она руками берет еду. Об-
ратите внимание, что разряды нейронов
наблюдаются только, когда обезьяна ру-
ками берет пищу. (в) Обезьяна хватает
кусочек пищи в темноте. Множественные
разряды нейронов наблюдаются даже при
совершении действия в полной темноте.
(Источник: Ri��olatti et al., 1996.)
Рис. 14 .7 . Наблюдение за действиями
другой обезьяны может вызвать разряды
зеркальных нейронов. Зеркальные нейро-
ны отвечают пачкой разрядов в ответ на
увиденное действие другой обезьяны (а),
на действие экспериментатора (б) и на
действие самой регистрируемой обезьяны
(в). (Источник: Ri��olatti et al., 1996.)
174
Глава 14
регистрации одиночных клеток у обезьян, и системой зеркальных нейронов че-
ловека, определенной методом нейровизуализации групповых нейронных от-
ветов. Риззолатти с коллегами (2002) обобщили гомологии человека и обезьяны
на рис. 14.8.
Область F5 коры макаки (фиолетовая на рис. 14.8) и ПБ 44 у человека (ро-
F5 коры макаки (фиолетовая на рис. 14.8) и ПБ 44 у человека (ро-
5 коры макаки (фиолетовая на рис. 14.8) и ПБ 44 у человека (ро-
зовая) кодируют движения руки, аналогичные исследованным в исходной ра-
боте по зеркальным нейронам. Поле F4 макаки и вентральная премоторная
кора (вПМ или нижняя часть ПБ 6) у человека (красная на рис. 14 .8) настроены
на движения руки или кисти в сторону определенного местоположения. Это
Рис. 14 .8 . Гомология локализации и функционирования зеркальных нейронов у людей
и обезьян. (а) Латеральная поверхность коры правого полушария мозга макак. (б) Лате-
ральная поверхность коры правого полушария мозга человека. PE, PEc — область коры
мозга макак, верхняя теменная область; FEF — фронтальное глазодвигательное поле;
PG — задняя нижняя теменная область; PF, PFG — передняя нижняя теменная область;
�1 — стимул 1; IP — внутритеменная борозда; C — центральная борозда; Al — нижняя
дугообразная борозда; A� — верхняя дугообразная борозда; P — главная борозда. (Ис-
точник: Ri��olatti et al., 2002.)
Социальная когнитивная деятельность: восприятие состояния других людей
175
значит, что зеркальные нейроны в этих областях не просто отражают основные
движения руки и запястья, но, скорее, отвечают за движения к определенной
цели, например дотянуться до какой-то точки пространства.
Примечание: как вы помните, поле Бродмана 44 (ПБ 44) в левом полушарии
называют зоной Брока. Ее традиционно рассматривали как корковую область,
ответственную за речь. Современные исследования имитации у людей пока-
зали, что ПБ 44 имеет свойства зеркальных нейронов, включая отображение
действий рукой или запястьем (Iacoboni, 2005) (рис. 14 .9). Таким образом, зона
Брока имеет разные функции.
Очень долгое время сложностью при исследовании зеркальных нейронов яв-
лялось разделение простого восприятия движения (это хватание, достижение
или держание) и детекции намерения (когда у лица есть цель, которая дости-
гается действием: пить чай или убирать после чая). Финальное доказательство,
объединяющее зеркальные нейроны с восприятием намерения, было получе-
но в исследованиях поведения людей в разных условиях. Центральный вопрос
этих работ: СЗН отвечает на действие независимо от контекста или отвечает
на действие в определенном контексте? Положительный ответ на первый во-
прос означает, что зеркальные нейроны ответственны за восприятие движений.
Вторая альтернатива позволяет расширить их функции: СЗН необходима для
восприятия намерения.
Марко Якобони с коллегами собрали доказательства, используя метод фМРТ
во время наблюдения за участниками действий в контексте и без, для подтверж-
Рис. 14.9 . Упрощенное изображение левого полушария мозга с выделением гипоте-
тического расположения фронтальной системы зеркальных нейронов. (Примечание:
Нижняя теменная кора = огибающая извилина = ПБ 40; вентральная премоторная
кора = нижняя часть ПБ 6; задняя нижняя лобная извилина = покрышечная часть
НЛИ = ПБ 44).
176
Глава 14
дения того, что СЗН отвечает, скорее, на намерения, чем на конкретные дви-
жения. Они обнаружили, что действия, включенные в контекст (Намерение на
рис. 14 .10), вызвали значительное увеличение фМРТ-сигнала в задней части
нижней лобной извилины и в прилегающей области вентральной премоторной
коры, где представлены движения руки, по сравнению с другими двумя услови-
ями (Контекст или Действие). Для того чтобы продемонстрировать, что наме-
рение может быть воспринято отдельно от действия, исследователи сравнили
фМРТ-изображения двух условий. Эти сравнения представлены на рис. 14 .11 .
Последующие работы (Dinstein et al., 2007, 2008) и (Carama��a et al., 2009),
однако, не подтвердили эти ранние открытия. Динштейн с коллегами обнаружи-
ли, что в то время как области теменной доли (передняя межтеменная борозда)
Рис. 14 .10 . Два разных типа контекста, действия и намерения использовали в своей
работе Якобони с коллегами (2005): пить чай и убирать после чаепития. Левый набор
картинок относится к условиям контекста, центральные изображения — условия дей-
ствия, а правые — условия намерений. (Источник: Iacoboni et al., 2005.)
Рис. 14.11. фМРТ-изображения экспериментов Якобони с коллегами (2005). В верхней
части рисунка показан ответ в условиях Намерение минус Действие; на нижней — в ус-
ловиях Намерение минус Контекст. (Источник: Iacoboni et al., 2005.)
Социальная когнитивная деятельность: восприятие состояния других людей
177
активировались при восприятии и выполнении движений, паттерны этой ак-
тивности сильно различались для случаев выполнения движения и простого
наблюдения за ним (см. рис. 14 .12 для иллюстрации модели эксперимента).
Исследователи предполагают, что эти результаты указывают на существование
различных субпопуляций нейронов, отвечающих на восприятие и совершение
движений (рис. 14 .13).
Похожее фМРТ-исследование провели Карамаза с коллегами (2009) и по-
казали результаты о приспособлении нейронов из области предположительно
содержащей зеркальные нейроны. Они обнаружили, что эти области мозга из-
меняют активность, когда движения кисти руки было увидено, но не когда то же
движение было выполнено испытуемым.
В целом эти и другие современные работы по изучению СЗН человека дают
доказательства того, что процессы восприятия и совершения двигательного
акта у человека и обезьян не имеют такого большого сходства, как считалось
ранее. Исследование СЗН является хорошим примером того, как научные тео-
рии разрабатываются и меняются в процессе накопления экспериментальной
базы, новые и исправленные гипотезы проверяются по мере переработки и со-
вершенствования экспериментов и теорий.
Рис. 14 .12 . Экспериментальная парадигма Динштейна с коллегами (2008). Испытуе-
мые играли в «камень, ножницы, бумага» в сканнере. Когда испытуемый видел сло-
ва «НАЧАЛИ» в начале пробы, он выполняет игровые движения (складывание руки в
жест «камня», «бумаги» или «ножниц»). После непродолжительного перерыва (3–6 с)
испытуемому предъявляют вариант хода оппонента. Эти пробы повторяют 36 раз.
(Источник: Dinstein et al., 2008.)
178
Глава 14
3.2 .
Распознавание глаз и восприятие направления взора
Распознавание глаз у особей своего вида и координация своего поведения на
основе этого восприятия наблюдается во многих классах животных от репти-
лий до человека. Четкое понимание роли глаз в социальном общении требует от
нас разобраться в исследованиях восприятия глаз и взора. В обзоре, включаю-
щем работу с множеством классов животных (Emery, 2000), был продемонстри-
Emery, 2000), был продемонстри-
2000), был продемонстри-
рован эволюционный тренд в сторону усложнения восприятия взора. В табл.
14.2 представлено обобщение этих данных.
В то время как рыбы, рептилии, птицы и другие млекопитающие имеют
способность воспринимать стимулы, похожие на глаза (пару горизонтально
расположенных темных кругов), и определять направление взора, только у че-
ловекообразных обезьян (гориллы, шимпанзе, бонобо и орангутанг) и челове-
ка существует разделенное внимание, и только они могут использовать «язык
глаз» для понимания мысленных состояний других особей.
Несколько типов процессов восприятия глаз и взора показаны на рис. 14.14 .
Где в мозге происходят эти события? Верхняя височная борозда (ВВБ) реги-
стрирует глаза и стимулы, им подобные. Более сложные процессы обработки
взора (направление взора и определение смещения взора) затрагивают связи
между ВВБ и областями теменной коры, а именно межтеменной бороздой
(МТБ; Haxby et al., 2000) (рис. 14 .15). В дополнении связи ВВБ и МТБ с такими
подкорковыми структурами, как миндалина, позволяют выявлять социальную
и эмоциональную значимости взгляда, включая угрозу.
Рис. 14.13 . Мозговая активность при выполнении и наблюдении движений. Результаты
работы Динштейна с коллегами (2008) «камень, ножницы, бумага» показали активацию
разных областей мозга выполнением и наблюдением схожих движений рукой. Эти дан-
ные противоречат теории о существовании зеркальных нейронов у человека. ПНТБ —
передняя часть нижне-теменной борозды.
Социальная когнитивная деятельность: восприятие состояния других людей
179
Таблица 14.2
Этологический обзор восприятия глаз/взора среди классов животных
Группа
субъектов
Опре-
деление
глаз
Пристальное
смотрение
Следование
за взором
Объединен
ное внима-
ние
Соотнесе-
ние мыслей
Рыбы
?
?
?
?
Рептилии
?
?
?
Птицы
?
(?)
?
Грызуны
?
?
?
?
Собаки
(домашние)
?
?
?
Полуобезьяны ?
?
?
Обезьяны
Высшие
приматы
(?)
Человек
–
6 месяцев
–
9 месяцев
–
12 месяцев
–
18 месяцев
–
24 месяца
–
48 месяцев
–
Аутизм
(?)
?
–
синдром
Дауна
–
Шизофрения
?
?
?
–
Поврежде-
ния минда-
лины
(?)
(?)
?
(?)
— подтвержденные данные; — нет данных; ? — не тестировали или противоречи-
вые данные (Источник: Emery, 2000)
Социально важную информацию от глаз и направления взора относят к
изменяющимся аспектам человеческого лица. Мы также используем зритель-
ную информацию для определения постоянных аспектов отдельных лиц как
отличительные черты. Эти типы восприятия лиц обрабатываются в отдельной
области височной коры, о чем мы расскажем в следующем разделе данной
главы.
180
Глава 14
Наконец, присвоение мысленных состояний требует вовлечения связей ВВБ
с СЗН и медиальных областей лобной коры (Iacoboni, 2005). Мы рассмотрим в
деталях присвоение мысленных образов.
3.3 .
Разделенное внимание
Разделенное внимание — социальный навык, присущий только человеку и че-
ловекообразным приматам. Запомните, что разделенное внимание — это боль-
ше, чем просто смотреть на один и тоже объект, на который смотрит другой ин-
дивид. Разделенное внимание обладает дополнительным свойством, т. е . двое
наблюдателей не только смотрят на один объект, но также знают, что каждый из
них на него смотрит. Это является триадической (трехсторонней) активностью.
Разделенное внимание позволяет нам косвенно понять, что: «Я знаю, что ты
смотришь на это». Приматы и люди понимают, что если особь своего вида смо-
трит на что-то, то у нее формируется внутренний образ этого объекта. Смотреть
Рис. 14 .14 . Процессы, связанные с глазами и взором. (1) При взаимном взгляде внима-
ние индивидуумов А и Б направлено друг на друга. Отведенный взгляд подразумевает,
что индивидуум А смотрит на Б, но у Б фокус внимания смещен в другую сторону.
(2) Следование за взором происходит, когда индивидуум А понимает, что взор Б на-
правлен не на него, и следует направлению взора Б к другой точке пространства. (3)
Объединенное внимание является почти тем же, что и следование за взором, но отлича-
ется наличием фокуса внимания (объекта), на который и смотрят индивидуумы А и Б.
(4) Разделенное внимание является комбинацией взаимного и объединенного внимания,
когда фокус внимания А и Б направлен как на объект объединенного внимания, так
и на друг друга («Я знаю, что ты смотришь на Х, и ты знаешь, что я смотрю на Х»).
(5) Теория разума опирается как на варианты 1–4, так и на социальные навыки высокого
порядка, которые позволяют индивидуумам предвидеть, что намереваются сделать дру-
гие с объектом, потому что они намерены что-то сделать с объектом сами. (Источник:
Emery, 2000.)
Социальная когнитивная деятельность: восприятие состояния других людей
181
значит видеть. Если я хочу увидеть то же, что и ты, я могу проследить за твоим
взором (см. блок 14.1).
Как эти разделенные процессы формируются у людей? Показано (�rooks,
Melt�off, 2005), что младенцы начинают отслеживать направления поворота го-
2005), что младенцы начинают отслеживать направления поворота го-
ловы взрослых начиная с 9 месяцев; однако в этом возрасте дети следят за по-
воротом головы независимо от того, открыты или закрыты глаза. К 12 месяцами
дети более часто отслеживают направления поворота головы, если у взрослого
открыты глаза. Значит, младенцы начинают отслеживать именно направление
взора, а не просто повороты головы. Разделенное внимание сформировано.
У ребенка сформировалось внутреннее знание о том, что необходимо смотреть,
чтобы видеть (рис. 14.16).
Какие области мозга обеспечивают функционирование разделенного вни-
мания? Мы знаем уже, что ВВБ обеспечивает выделение глаз. Для того чтобы
перейти от простого восприятия направления взора к разделенному вниманию,
необходимо задействовать префронтальную кору. Уильямс с коллегами (Wil-
Wil-
liams, 2005) изучали взрослых, когда они испытывали разделенное внимание, и
сравнивали параметры этого состояния с контрольным (рис. 14 .17).
Рис. 14 .15. Верхняя височная борозда и межтеменная борозда — области детекции глаз
и взора
Блок 14.1 . Эксперимент по слежению за взором
Попробуйте этот эксперимент на разделенное внимание: когда вы находитесь в об-
ществе друзей или одноклассников, переместите ваш взгляд в угол помещения, не
говоря при этом ничего. Посмотрите, переместит ли кто-нибудь свой взор вслед за
вами. Спросите, почему они сделали это.
182
Глава 14
Результаты фМРТ-анализа показали, что разделенное внимание задействует
лобные области, включая вентромедиальную префронтальную кору (ВМ-ПФК),
левую верхнюю лобную извилину (ПБ 10), поясную извилину и хвостатое ядро.
ВМ-ПФК ассоциируют с процессами регистрации мысленных состояний дру-
гих особей, что мы и обсудим далее. Уильямс с коллегами утверждают, что ПБ
10 ответственно за сличение восприятия и действия (рис. 14 .18).
3.4 .
Способности высокого порядка из ТР
3.4.1 . Приписывание мысленных состояний себе и окружающим
В предыдущих разделах мы узнали, что парацингулярная извилина вовлечена
в процессы подражательного обучения, а вентромедиальная префронтальная
кора (ВМ-ПФК) — в разделенное внимание. В этом разделе мы рассмотрим
роль медиальной поверхности ПФК в приписывании мысленных состояний
другим, или в «ментализации», как говорит Крис Фрис.
Рис. 14 .16 . Ребенок 18 ме-
сяцев показывает пальцем.
Показывание пальцем яв-
ляется признаком триади-
ческих взаимодействий
Рис. 14 .17. Стимулы, использующиеся для создания объединенного внимания. Когда
мы смотрим на красный кружок на левой половине рисунка, у нас возникает ощущение,
что мужчина смотрит туда же; рассматривание красного кружка на правой половине не
приводит к появлению чувства разделенного внимания. (Источник: Williams et al., 2005.)
Социальная когнитивная деятельность: восприятие состояния других людей
183
Медиальная поверхность префронтальной коры может быть разделена на
три части сверху вниз: дорсомедиальная префронтальная кора (ДМ-ПФК), ме-
диальная префронтальная кора (М-ПФК) и вентромедиальная префронтальная
кора (ВМ-ПФК). (Данное разделение основано на соглашении и было выпол-
нено по координатам системы Талайраха в трехмерной проекции, но об этом
мы не будем говорить здесь.) ДМ-ПФК включает область коры в верхней части
медиальной поверхности префронтальной коры. М -ПФК занимает среднее по-
ложение на медиальной поверхности ПФК. ВМ -ПФК расположена на нижней
поверхности лобной доли и внизу медиальной стороны префронтальной коры
(рис. 14 .19).
Рис. 14 .18. Нейронная
сеть для разделенного
внимания
Рис. 14 .19 . Деление префронтальной коры: ДМ-ПФК, М-ПФК и ВМ-ПФК. Верхняя
лобная извилина, поясная (ПБ 24) и парацингулярная (ПБ 32) извилины являются важ-
ными областями для мыслительной деятельности
184
Глава 14
Поясная и парацингулярная извилины, которые иногда рассматривают вме-
сте как переднюю поясную кору (ППК), образуют пояс (от лат. cingulum) вокруг
мозолистого тела. К тому же в зависимости от того, как сформирован индиви-
дуальный мозг, парацингулярная извилина может образовывать борозду. С по-
мощью рис. 14 .20 можно разобраться в анатомии поясной и парацингулярной
извилин. Области мозга, активирующиеся при процессах принятия перспекти-
вы и позиции намерения, показаны на рис. 14 .21 .
3.4.2. Перспектива принятия и намеренная позиция
Перспектива принятия — социальный навык, лежащий в основе человеческого
сочувствия. Он позволяет нам понимать, что думает и чувствует другой чело-
век в болезненных ситуациях. Например, когда испытуемых просят предста-
вить боль, которую они или другой человек почувствовали бы (по сравнению
с ощущениями искусственной руки), если бы их пальцы прищемило дверью
Рис. 14 .20. Анатомия поясной и парацингулярной извилин: на верхнем левом рисунке
изображено левое полушарие, на верхнем правом — правое полушарие мозга. Парацин-
гулярная борозда покрашена синим цветом, поясная борозда — розовым, а мозолистая
борозда — фиолетовым. На нижнем рисунке эти области мозга, включая ростральную
часть поясной области, показаны более подробно. (Источник: Heckers et al., 2004.)
Социальная когнитивная деятельность: восприятие состояния других людей
185
машины, активируется медиальная префронтальная кора, а также другие об-
ласти коры, отвечающие за восприятие боли (Jackson et al., 2006). В частности,
парацингулярная кора (ПБ 32) М-ПФК значительно активируется в обоих слу-
чаях; она регистрирует воображаемую собственную боль и чужую боль. К тому
же, когда испытуемые представляли свою боль, активация наблюдалась в по-
ясной коре (ПБ 24). Эти различия позволяют нам не только сочувствовать боли
окружающих людей, но и отделять свое чувство боли от чужих.
Или представьте, что вы играете с другим человеком: игроки и спортсмены
рассказывают о «психологическом воздействии» на противников. Большин-
ство, о чем они говорят, сводится к чтению мыслей другого человека. Для того
чтобы это сделать, необходимо поставить себя на место этого индивида, или,
если сказать иначе, проявить эмпатию. Когда мы играем в «камень, ножницы,
бумага» против невидимого человека (по сравнению с игрой против невиди-
мого компьютера), мы принимаем намеренную позицию, т. е . мы действуем
так, как если бы противник являлся субъективным существом. И мы можем
наблюдать изменение метаболической активности в мозге, которая совпадает
с принятием нами намеренной позиции. На рис. 14.22 представлена активация
передней парацингулярной коры (ПБ 32), т. е . та часть медиальной префрон-
тальной коры, которая задействована в восприятии своих и чужих ментальных
состояний (Gallagher et al., 2002).
Нас иногда просят принимать решения за других, учитывая, что они сами бы
хотели. В исследовании студентов-медиков просили сформировать представле-
ние о пациенте для того, чтобы сделать предполагаемые решения относительно
их здоровья. Результаты ПЭТ-сканирования опять выявили уникальную актив-
ность в медиальной лобной коре, в этом случае в медиальной верхней лобной
извилине (рис. 14 .23) (Ruby and Decety, 2003).
ПЭТ-активация возникает в дорсомедиальной префронтальной коре (ДМ-
ПФК) и медиальной теменной (предклинье), когда испытуемые смотрят фильм,
в котором актеры вовлечены в социальные взаимодействия (рис. 12 .24) (Iaco-
Iaco-
Рис. 14 .21 . Перспектива
принятия и намеренная по-
зиция. (а) Значительная ме-
диальная префронтальная
активация на собственную
боль по сравнению с искус-
ственной. (б) Значительная
активация М-ПФК в ответ
на боль других людей по
сравнению с искусствен-
ной болью. (в) Изменение
местного кровотока на ПЭТ-
диаграмме в М-ПФК; соб-
ственная — синяя линия;
чужая — желтая; искус-
ственная рука — красная.
(Источник: Jackson et al.,
2006.)
186
Глава 14
Рис. 14 .22. Активация пе-
редней части парацингу-
лярной коры в тот момент,
когда мы думаем о нашем
сопернике.
(Источник:
Gallagher et al., 2002.)
Рис. 14 .23 . Формирова-
ние восприятия другого
человека для принятия за
него или нее медицинско-
го решения. Активация
медиальной ПФК показа-
на градиентом от желтого
к красному цвету. (Ис-
точник: Ruby and Decety,
2003.)
Рис. 14 .24. Размышление
о социальных взаимоот-
ношениях. Дорсомедиаль-
ная префронтальная кора
и медиальная теменная
кора формируют систему
для мышления о социаль-
ных связях. (Источник:
Iacoboni et al., 2004.)
Социальная когнитивная деятельность: восприятие состояния других людей
187
boni et al., 2004). Эти области не активируются, если испытуемые видят, как
один человек вовлечен в социальную деятельность. Красным цветом обозна-
чены области, которые активируются во время социальных взаимоотношений,
но не во время индивидуальных действий. Синие области показывают наличие
активности во время просмотра эпизодов с социальной активностью по сравне-
нию с состояние покоя.
Якобони и его коллеги (2004) сделали следующее предположение:
Система для размышлений о социальных взаимоотношениях в дорсомедиальной
префронтальной коре и медиальной теменной коре является частью мозгового со-
стояния по умолчанию; она может постоянно, часто без усилия или намерения, оце-
нивать и анализировать прошлые, настоящие или возможные будущие социальные
отношения всякий раз, когда выполняемые действия не требуют полного внимания.
3.5 .
Восприятие других как похожих и непохожих:
Я-Оно в мозге
Для социальной психологии важен вопрос о том, как вовлекается мозг, когда
мы воспринимаем окружающих либо похожих на нас самих, либо отличающих-
ся от нас. Когда других людей не воспринимают как принадлежащих к вашей
социальной группе, вы можете начать себя оправдывать в обращении с ними
иначе. К тому же мы можем отрицать возможность наличия у них высших пси-
хических состояний и чувств. С непохожими «другими» с большей вероятно-
стью будут обходиться как с объектом, по Буберу, согласно понятию Я-Оно (см.
блок 14.2).
Исследование Митчелла с коллегами (Mitchell, 2006) выявляет разность в
активации областей медиальной префронтальной коры у испытуемых, которых
Блок 14.2 . В 1923 г. Философ Мартин Бубер написал в своей ставшей
классической книге «Я и Ты» о перспективе второго человека
Бубер идентифицировал два фундаментально различных способа взаимодействия с
другими людьми и объектами, Я-Оно и Я-Ты. Я-Оно предполагает восприятие дру-
гих в качестве объектов; Я-Ты предусматривает эмпатическое восприятие других
как субъектов. Обычно мы легко переключаемся между
этими двумя способами восприятия других людей. Как
только мы принимаем одну или другую позицию, наше
внутреннее состояние меняется. За последние десять лет
эти внутренние состояния были изучены с помощью ме-
тодов нейровизуализации. Бубер писал:
Основные слова исходят от существа человека.
Когда говорится Ты, говорится и Я сочетания Я-Ты.
Когда говорится Оно, говорится и Я сочетания Я-Оно....
Нет Я самого по себе, есть только Я основного слова
Я-Ты и Я основного слова Я-Оно....
Тот , кто говорит Ты, не обладает никаким Нечто как
объектом. Ибо там, где есть Нечто, есть и другое Не-
что; каждое Оно граничит с другими Оно; Оно суще-
ствует лишь в силу того, что граничит с другими. Но
когда говорится Ты, нет никакого Нечто. Ты безгранично.
Рис. 14 .25. Мартин
Бубер (Martin �uber,
1878–1965)
188
Глава 14
просили сделать выводы о людях, имеющих похожие либо не похожие полити-
ческие взгляды. Они выяснили, что области вентромедиальной префронталь-
ной коры (ВМ-ПФК) активируются во время суждений о похожих людях, тогда
как области в дорсомедиальной ПФК были активны во время суждений о не
похожих индивидуумах (рис. 14 .26).
Митчелл с коллегами, основываясь на результатах, заключили, что мы мо-
с коллегами, основываясь на результатах, заключили, что мы мо-
жем активно развивать другую систему стратегий социальной когнитивной де-
ятельности при восприятии непохожих на нас людей. Меньшее привлечение
ВМ-ПФК к восприятию непохожих индивидуумов подразумевает меньшее
приписывание им чувств и эмоций. В следующем разделе мы рассмотрим раз-
ные стратегии восприятия других людей.
3.5.1 . Когнитивная или чувственная эмпатия?
Психологи иногда различают сочувствие эмоциональное и разумное. Они ука-
зывают на различия между способностями теории разума, призванными по-
мочь понять убеждения других, и способностью к сочувствию, которая помо-
гает нам понять, что чувствует другой человек.
Рис. 14 .26 . Восприятие дру-
гих как похожих и не похожих.
(а) Вентромедиальная ПФК
активируется при решении
о схожести других. (б) Дор-
сомедиальная ПФК активиру-
ется в результате решения о
различии других. (Источник:
Mitchell et al., 2006.)
Социальная когнитивная деятельность: восприятие состояния других людей
189
Шамэй-Тсори с коллегами (�hamay-Tsoory, 2005) обнаружили различия,
отражающие это положение, когда тестировали взрослых, страдающих пора-
жениями вентромедиальной ПФК, при выполнении социальных логических
задач. Исследователи просили испытуемых отвечать на вопросы после трех
прочитанных рассказов, содержащих (1) ошибочные утверждения второго по-
рядка, (2) социальную бестактность (faux pas) и (3) иронию. В блоке 14.3 при-
ведены примеры историй и вопросов к ним. Ученые считают, что понимание
Блок 14.3 . История для определения трех типов общественного суждения
Ошибочные утверждения второго порядка
Хана и Бенни сидят в кабинете и обсуждают их встречу с начальником. Бенни ставит
открытую чернильницу себе на стол. Во время этого действия несколько капель чер-
нил проливаются, поэтому он покидает кабинет в поисках тряпки, чтобы вытереть
пролитые чернила. Пока Бенни нет в кабинете Хана убирает чернильницу в ящик
стола. В то время как Бенни отсутствует в кабинете, он подсматривает в замочную
скважину и видит, как Хана убирает чернильницу. Бенни возвращается в кабинет.
После рассказа истории, задают четыре вопроса:
Вопрос на утверждения: Как думает Хана, знает ли Бенни, где находится черниль-
ница?
Вопрос о реальности (определение степени понимания): Где на самом деле находит-
ся чернильница?
Вопрос на память: Где оставил чернильницу Бенни?
Вопрос на логику: Где будет находиться пятно от чернил?
Ирония
Саркастический вариант: Джо пришел на работу, и вместо того, чтобы начать тру-
диться, он сел отдыхать. Его начальник заметил это поведение и сказал: «Джо, не
работай слишком интенсивно!»
Нейтральный вариант: Джо пришел на работу и немедленно приступил к делу. Его
начальник заметил это поведение и сказал: «Джо, не работай слишком интенсивно!»
После каждого варианта рассказа задают два вопроса:
Вопрос о фактах (определение степени понимания): Джо много работал?
Вопрос отношения (определение понимания истинного отношения говорившего):
Считал ли начальник, что Джо много трудится?
Узнавание социальной бестактности (faux pas)
Девятилетний мальчик Майк поступил в новую школу. Он находился в одной из ка-
бинок школьного туалета, когда туда вошли Джо и Питер, другие школьники, и стали
разговаривать, стоя у раковин.
Джо сказал: «Ты знаешь нового мальчика из нашего класса? Его зовут Майк. Правда,
что он очень странный? Он такой маленький!» Майк вышел из кабинки, и Джо с
Питером его увидели. Питер сказал: «О, привет, Майк! Ты идешь сейчас играть в
футбол?»
Испытуемому задают следующие вопросы:
Вопросы на узнавание faux pas:
Кто-нибудь сказал то, чего ему не следовало говорить?
Кто сказал то, чего ему не следовало говорить?
Почему это нельзя было говорить?
Почему они так сказали?
Контрольные вопросы (определение степени понимания):
Где находился Майк во время разговора Джо с Питером?
(Источник: �hamay-Tsoory et al., 2005).
190
Глава 14
социальной бестактности и иронии требует большего вовлечения эмоциональ-
ных процессов, в то время как определение ошибочных утверждений опирается
в большей степени на когнитивные процессы.
Было обнаружено, что пациенты с повреждениями вентромедиальной пре-
обнаружено, что пациенты с повреждениями вентромедиальной пре-
фронтальной коры справлялись с заданием на ошибочные утверждения как
здоровые испытуемые и пациенты с повреждениями задних областей коры
выполнили задание великолепно. Однако пациенты с повреждением ВМ-ПФК
показали плохие результаты в тестах на иронию и faux pas по сравнению с дру-
гими испытуемыми. Согласно интерпретации исследователей, ВМ-ПФК явля-
ется необходимой для чувственной эмпатии, но не для когнитивных навыков
сочувствия.
Похоже, что навыки сочувствия следуют главному плану других когнитив-
ных способностей: факты об окружающем мире, убеждения, сенсорные обра-
зы обрабатываются дорсальной и латеральной корой. Чувства, эмоциональные
ценности и социальная значимость зависят от работы вентральной и медиаль-
ной коры, которые тесно взаимосвязаны с подкорковой лимбической системой.
3.6 .
Восприятие лиц
Восприятие постоянных черт человеческих лиц происходит в веретеновидной
области лиц (ВОЛ), которая является частью нижней височной доли (рис. 14 .27).
Пример того, как выглядит ВОЛ на изображения ПЭТ/МРТ, можно взять из
некоторых работ (Caldara et al., 2006) (рис. 14 .28). Эти исследователи сравнили
корковую активацию, когда испытуемые рассматривали рисунки объектов или
Рис. 14 .27. Вид на полушария
мозга снизу с обозначением
веретеновидной области лиц
(ВОЛ)
Социальная когнитивная деятельность: восприятие состояния других людей
191
лиц. На изображении мы смотрим на нижнюю поверхность мозга. Височные
доли занимают практически всю внешнюю часть на этой картинке. Правая и
левая ВОЛ, области восприятия лиц, отчетливо отмечены красным цветом. Ря-
дом с ВОЛ расположены еще области нижней височной доли, правая и левая па-
рагиппокампальные извилины (ПГИ), отвечающие на такие неодушевленные
предметы, как дома или обувь.
Хэксби с коллегами (Haxby et al., 2002) собрали результаты этих исследо-
ваний воедино и создали модель областей восприятия лиц в мозге (рис. 14 .29).
Они сформировали иерархическую систему взаимосвязанных областей мозга,
отвечающую за восприятие изменяющихся и постоянных аспектов лиц.
В этой модели первичный зрительный анализ свойств лиц происходит в зри-
тельной коре, нижней затылочной извилине (НЗИ). НЗИ посылает информа-
цию в верхнюю височную борозду (ВВБ), где обрабатываются изменяющиеся
черты лица, например глаза; оттуда информация о глазах объединяется с про-
странственной информацией от межтеменной борозды (МТБ) для формирова-
ния информации о направлении взора. Данные от ВВБ также направляются в
миндалину, где к ним присоединяются социальные и чувственные значения, и
в слуховую кору, где обрабатываются движения губ. Постоянные аспекты лица,
индивидуальные черты, обрабатываются латеральной веретеновидной извили-
ной (ВОЛ), которая взаимосвязана с височной долей, где воспринимается спец-
ифичная информация об имени и биографических данных.
Рис. 14.28 . Вид на нижнюю
поверхность полушарий мозга
с обозначением красным цве-
том веретеновидных областей
лиц (ВОЛ), а синим — пара-
гиппокампальных
извилин
(ПГИ). (Источник: Caldara
et al., 2006.)
192
Глава 14
4.0 .
Заключение
Способность к социальному общению у людей является сложной и многогран-
ной. Она обеспечивается множеством систем взаимосвязанных корковых и под-
корковых областей. В процессе эволюции простая оценочная и поведенческая
система лимбического мозга была надстроена растущими полушариями моз-
га, которые сделали возможным сложную когнитивную деятельность. Вполне
вероятно, что наши большие и развитые полушария мозга эволюционировали
частично в связи с необходимостью справляться с давлением увеличивающей-
Рис. 14 .29 . Модель для восприятия лиц. В левой части схемы показаны первичные зри-
тельные области для восприятия лиц, в центре — области меняющихся и постоянных
черт лица, в правой части схемы показана дальнейшая обработка информации о свой-
ствах лица. (Источник: Haxby et al., 2002,)
Социальная когнитивная деятельность: восприятие состояния других людей
193
ся сложности человеческого общества и требованиями социальной когнитив-
ной деятельности. Увеличивающаяся социальная и когнитивная сложность в
окружающей среде идет в ногу с увеличением сложности в соответствующих
структурах мозга.
5.0 .
Задания и упражнения к главе 14
5.1.
Контрольные задания
1. Кратко объясните, что понимают под термином «теория разума».
2. Что по Фрису значит ментализация?
3. Почему механизмы разделенного внимания так важны для развития челове-
ка? Как они развиваются?
4. Как отличается система зеркальных нейронов от других нейронных систем?
Какова их роль в социальной когнитивной деятельности?
5. Какую роль играет контекст для понимания намерений?
6 Почему перспектива принятия является ключевым социальным навыком?
5.2 .
Графические упражнения
1. Отметьте ключевые области мозга (показаны оттенками красного и розового
на рис. 14 .30), предположительно относящиеся к фронтальной системе зер-
кальных нейронов.
Рис. 14 .30
Жизнь – это пламя, которое горит лишь раз, но оно загорается снова,
когда ребенок рождается.
Джордж Бернард Шоу
Зрелый мозг содержит полностью сформированные нервные проводящие пути, соеди-
няющие все области мозга между собой. До недавнего времени очень мало было извест-
но о развитии проводящих волокон у младенцев. Используя технологию визуализации
распределения тензора, созревание проводящих путей можно исследовать на детях в
возрасте 1–4 месяцев. Степень созревания проводящих волокон закодирована в цвете
(от синего к красному). (Источник: Debois et al., 2009.)
Содержание
1.0.Введение.......................................................195
1.1 . Новые технологии для исследования развивающегося мозга . . . . . . . . . 196
1.2.Тайнаразвитиямозга:старыеиновыевопросы....................197
2.0.Пренатальноеразвитие:отбластоцистыдомладенца..................197
2.1.Эпигенез....................................................198
2.2.Анатомияразвивающегосямозга................................199
2.3.Миграциянейронов...........................................204
2.4.Пересмотрроливрожденногоиприобретенного...................207
2.5 . Пренатальный слуховой опыт:
восприятиеголосаимузыкидорождения.........................210
3.0.Развивающийсямозг:изменениядлиноювжизнь.......................211
3.1.Начало и завершение постнатального развития мозга. . . . . .. . . . . .. . . 211
3.2.Региональныеособенностиразвитиямозга.......................213
4.0.Развитиемозгаисознания.........................................215
4.1.Первыйгоджизни:взрывнойростиразвитие.....................221
4.2 . Детство и подростковый возраст: динамичное и поэтапное развитие . . 234
5.0.Ранниеповреждениямозгаипластичностьразвития...................252
6.0.Заключение.....................................................256
7.0.Заданияиупражнениякглаве15...................................256
7.1.Контрольныезадания..........................................256
1.0 . Введение
В этой главе мы дадим обзор того, как происходит рост и развитие человека в
течение его жизни: от пренатальных стадий до младенчества, от детского воз-
раста до взросления. Более детально мы рассмотрим ранние этапы развития
мозга и когнитивных процессов, так как в первые годы жизни у человека про-
исходит резкий взрыв изменений, связанных с развитием нервной системы, по
мере того, как ребенок познает мир. Мы исследуем роль врожденного и приоб-
ретенного в развитии мозга и мышления, раскрывая взаимовлияния генетиче-
ски запрограммированных свойств и опыта.
Область нейронауки о развитии когнитивных процессов, т. е . исследование
созревания мозга и соответствие этапов развития мозга человека с мыслитель-
ной деятельно стью, является достаточно молодой. Без преувеличения скажем,
изобретение новых неинвазивных методов для исследования мозговых функ-
ций у младенцев и детей привело к революционным изменениям в знаниях о
Глава 15
Развитие
196
Глава 15
том, что малыши и маленькие дети знают об окружающем их мире. Централь-
ная область исследования развития мозга и его влияния на поведение опирается
на комбинацию множества технологий и экспериментальных подходов для вы-
явления сложной взаимосвязи мозга и разума.
В этой главе мы кратко рассмотрим технологии, позволяющие наблюдать
развитие младенцев и детей. Также мы познакомимся с анатомическим разви-
тием мозга от пренатальных до постнатальных стадий жизни. Далее проследим
развитие мозга и разума в детстве и подростковом возрасте. А в конце главы
рассмотрим долговременные эффекты раннего перинатального повреждения
мозга и пластичность мозга в детстве. На всем протяжении главы мы познако-
мимся с современными экспериментальными исследованиями развития мозга
и когнитивных функций. Однако необходима осторожность в суждениях, так
как данная область исследований чрезвычайно молода, и мы только начинаем
узнавать связи между мозгом и поведением.
1.1 .
Новые технологии для исследования
развивающегося мозга
Появление новых методов исследования человеческого мозга подробно рас-
смотрено в гл. 4 . Электроэнцефалография/ связанные с событиями потенциалы
(ЭЭГ/ССП) и фМРТ — два метода, использующиеся в исследованиях на ново-
Рис. 15.1 . Методы для исследования мозговых процессов у новорожденных и малень-
ких детей: (вверху) многоэлектродное измерение ЭЭГ; (слева внизу) установка МРТ,
подготовленная для обследования новорожденных и детей; (справа) специальное крес-
ло для младенцев для проведения МРТ. (Источник: Ghislaine Dehaene-Lambertz, с раз-
решения.)
Развитие
197
рожденных и маленьких детях (рис. 15 .1). В то время как эти методы внесли
революционные изменения в область молодой когнитивной нейронауки, нигде
этот эффект так сильно не чувствовался, как в исследованиях развития мозга
и соответствующих поведенческих функций. Изучение поведения и мозговых
функций взрослых людей дало нам представление о распределении по областям
мозга типичных мозговых функций, таких как речь, эмоции и память. Эти ис-
следования дают нам информацию о результатах повреждения или заболевани-
ях мозга. Однако картина нарушений, обнаруженных при мозговых поврежде-
ниях у взрослых людей, значительно отличается от таковых при возникновении
травм на раннем этапе развития. Таким образом, изобретение новых методов
нейровизуализации позволяет нам понять, как устроены различные области
мозга и когнитивные функции на разных этапах развития.
Новые и сложные методы исследования анатомических изменений в про-
цессе развития также способствовали обогащению наших знаний о формиро-
вании мозга (рис. 15 .2). Эти методы позволяют прослеживать развитие серого
вещества во всем мозге, а также оценивать формирование проводящих путей
между и внутри полушарий мозга.
1.2.
Тайна развития мозга: старые и новые вопросы
В этой главе мы обратимся к некоторым вопросам, которые задавали ученые на
протяжении многих лет. Главным вопросом в развитии человека является опре-
деление роли врожденного и приобретенного. Схожая проблема — выяснение
границ пластичности мозга в результате новых событий в окружающей среде и
восстановления после повреждения. Можно сформулировать и новые вопросы,
которые невозможно было поставить в связи с недостаточным развитием экспе-
риментальных подходов и методов, например, что ребенок уже знает к моменту
рождения? Каковы отставленные эффекты повреждения мозга в раннем воз-
расте? Как различается скорость формирования мозговых процессов в разных
областях и полушариях мозга? Мы рассмотрим дополнительные данные о пу-
тях развития трех основных областей когнитивной науки: речи, произвольного
поведения и социальных навыков.
2.0 .
Пренатальное развитие: от бластоцисты до младенца
Большая часть этой главы будет направлена на рассмотрение развития мозга
и соответствующих когнитивных процессов во время младенчества и детства.
Прежде чем начать такое обсуждение, мы рассмотрим основные процессы,
протекающие до рождения, во время пренатального развития. Очень мало из-
вестно о сенсорных, перцептивных или когнитивных процессах у плода в утро-
бе (in utero), поэтому современные работы концентрируются на исследовании
того, что ребенок познает до рождения. Этот предшествующий рождению опыт
может быть критичным для дальнейшего развития. И независимо от того, явля-
ется опыт положительным — слышать голос матери или ее сердцебиение, или
негативным — испытывать злоупотребление матерью алкоголя — он может
оказывать отставленные эффекты на мышление и социальное развитие. Давай-
те начнем наш раздел о пренатальном развитии с обсуждения экспрессии генов
и роли окружающей среды.
198
Глава 15
2.1.
Эпигенез
Центральной проблемой в области знаний о развитии человека является вопрос
о влиянии врожденных особенностей и приобретенных свойств. Предопреде-
ляет ли набор генетической информации то, какими мы станем? Или наш опыт
делает нас такими, какие мы есть? На самом деле и опыт, и гены влияют на
развитие человека. Разворачивается ли сначала экспрессия генов, за которой
следует формирование структур и функций, которые потом испытывают влия-
ние со стороны окружающей среды? Или опыт — локальная среда, в пределах
одной клетки, системы или мозга в ц елом — оказывает влияние на экспрессию
генов? Взаимодействие между генами и средой чрезвычайно сложное и возни-
кает задолго до рождения. Мы начинаем раздел когнитивной нейронауки раз-
вития человека с обсуждения природы эпигенеза.
Рис. 15.2 . Система обработки МРТ-изображений. Верхний ряд: типичная последова-
тельность обработки изображения начинается с трансформации нативной записи маг-
нитно-резонансного (МР) образа в стереотаксические рамки; в результате этого про-
цесса формируется «зарегистрированное» изображение на шаблоне мозга. Следующий
этап включает поточечное распределение мозговой ткани на три основных класса: се-
рое вещество (в красном цвете), белое вещество (в белом) и спинномозговая жидкость
(зеленая). Каждое бинарное изображение (0 — отсутствие ткани; 1 — наличие ткани)
фильтруется (или сглаживается) для получения «плотностных» изображений; изобра-
жение плотности серого вещества (СВ), показанное на этом рисунке, свидетельствует
о том, что в каждой точке локальная концентрация СВ имеет значение от 0 до 1 (более
«теплый» цвет, от синего к красному, отображает более высокую плотность СВ). Ниж-
ний ряд: нелинейная регистрация нативного изображения по отношению к шаблону
мозгового строения позволяет выявлять локальные особенности строения мозга; поле
искажения количественно определяет различия между шаблоном и образцом во всем
мозге. Комбинация нелинейной регистрации и определения тканей позволяет автома-
тически выделить различные мозговые структуры, как лобная кора или миндалина.
Другие методы формируют изображения карт корковой толщины или идентифицируют
борозды коры больших полушарий. (Источник: Paus, 2005.)
Развитие
199
Эпигенез, или разворачивание генетической информации в зависимости от
специфического окружения, является ключом к современным вопросам о раз-
витии. Разные взгляды на эпигенез лежат в основе разных концепций развития
в когнитивной нейронауке. Готтлиб и Халперн (Gottlieb, Halpern, 2002) удачно
разделили два понятия «предопределенный эпигенез» и «вероятностный эпи-
генез». Предопределенный эпигенез предполагает существование однонаправ-
ленного пути от генов к анатомии мозга и изменениям в строении мозга и его
когнитивных функций. Как гипотетический пример можно привести синтез
некоего нейрохимического соединений в результате экспрессии гена. Высокие
уровни концентрации данного соединения могут активировать определенный
нервный процесс в мозге, и эта дополнительная активность в мозге может при-
вести к улучшению выполнения сложных задач по сравнению с предыдущими
возможностями. Эти увеличенные когнитивные способности можно наблю-
дать, когда ребенок начинает справляться с задачами, которые ранее были ему
не под силу. В отличие от этого вероятностный эпигенез предполагает нали-
чие двусторонних взаимодействий между генами, структурными изменения-
ми в мозге и функциями (рис. 15 .3). Наличие двусторонних взаимодействий
подразумевает, что не только гены могут вызывать изменения в поведении,
но и сенсорная информация, полученная ребенком, может влиять на паттер-
ны экспрессии генов. Например, как далее будет рассказано, новорожденные
дети имеют простые нейронные сети, которые смещают взгляд ребенка на лица
других людей (�ohnson, 1991). Это рано возникшее внимание к лицам в резуль-
�ohnson, 1991). Это рано возникшее внимание к лицам в резуль-
, 1991). Это рано возникшее внимание к лицам в резуль-
тате вызывает активацию определенных нервных областей в зрительной коре
ребенка и формирует их для восприятия лиц. Нейроанатомические изменения,
лежащие в основе этих процессов, являются результатом дифференциальной
экспрессии генов.
2.2 .
Анатомия развивающегося мозга
Большая часть развития мозга приходится на первые недели после оплодотво-
рения, и мы рассмотрим этот процесс в данном разделе. Вскоре после зачатия,
оплодотворенная клетка претерпевает ряд клеточных делений, в результате чего
появляется группа пролиферирующих клеток (называющаяся бластоцистой),
которая по внешнему виду напоминает гроздь винограда (рис. 15 .4). Через не-
сколько дней бластоциста дифференцируется в трехслойную форму (зародыше-
вый диск). Каждый из трех слоев впоследствии дифференцируется в основные
системы органов, при этом эндодерма (внутренний слой) образует внутренние
органы (пищеварительную, дыхательную и другие системы), мезодерма (сред-
Рис. 15.3 . Изображение системы психобиологического развития. (Источник: Gottlieb
and Halpern, 2002.)
200
Глава 15
Рис. 15.4 . Образование бластоцисты. Ранние процессы развития животных происхо-
дят по консервативному пути; после оплодотворения следует дробление, которое пре-
вращает яйцеклетку в многоклеточную бластулу. Анимальный и вегетативный полюсы
являются результатом исходной асимметрии ооцита, а вторая ось, дорсовентральная в
данном примере, образуется после оплодотворения. В процессе гаструляции некоторые
клетки уходят с поверхности эмбриона внутрь, что приводит к формированию трех-
слойной структуры, свойственной большинству многоклеточных животных. (Источ-
ник: Sanes et al., 2006.)
Рис. 15.5 . Нервная трубка. Конечная организация
нервной трубки начинает формироваться вско-
ре после ее смыкания. Самая вентральная часть
нервной трубки уплощается в хорошо различи-
мую вентральную пластинку. Дорсальная часть
нервной трубки формируется в ткань дорсальной
пластинки. Отчетливая борозда, sulcus limitans,
образуется между дорсальной и вентральной ча-
стями нервной трубки на всем ее протяжении.
(Источник: Sanes et al., 2006.)
Развитие
201
ний слой) станет скелетом и мышечной системой, а эктодерма (внешний слой)
разовьется в кожные покровы и нервную систему (включая органы восприятия).
Формирование нервной системы начинается с процесса нейруляции. Часть
эктодермы начинает складываться, образуя полый цилиндр, называющийся
нервной трубкой (рис. 15 .5).
Нервная трубка претерпевает изменения в трех измерениях: продольная дли-
на, длина окружности и радиус. По длине нервная трубка дифференцируется в
отделы центральной нервной системы, с формированием переднего и среднего
мозга на одном конце и спинного мозга на другом (рис. 15 .6). Часть нервной
трубки, которая станет спинным мозгом, разделяется на множество одинаковых
сегментов, в то время как на другом конце образуется и формируется последо-
вательность выпуклостей и изгибов. Через пять недель после оплодотворения
эти выпуклости становятся протоформами отделов мозга. Одна выпуклость
дает основу для формирования полушарий, вторая развивается в таламус и ги-
поталамус, третья становится средним мозгом, другие формируют мозжечок и
продолговатый мозг.
Рис. 15.6 . Образование головного и спинного мозга из нервной трубки у позвоноч-
ных животных. На рисунке показан вид с латеральной (верхняя часть) и дорсальной
(нижняя часть) поверхности на человеческий эмбрион на разных стадиях эмбриональ-
ного развития (а, б, в). Основные три части мозга (а) образуются в виде трех мозго-
вых пузырей или вздутий, и называются передний мозг (prosencephalon), средний мозг
(mesenceephalon) и ромбовидный мозг (rhombencephalon). На следующей стадии раз-
вития мозга (б) происходит дальнейшее его подразделение, при этом передний пузырь
делится на парные конечные пузыри и промежуточный мозг, ромбовидный мозг — на
задний мозг и продолговатый мозг. Эти основные отделы мозга могут быть соотнесе-
ны с конечной анатомической организацией зрелого мозга (в). (Источник: Sanes et al.,
2006.)
202
Глава 15
Формирование сенсорной и моторной систем происходит по оси, перпен-
дикулярной поверхности нервной трубки, при этом дорсальная (верхняя) часть
образует в основном сенсорную кору, а вентральная (нижняя сторона) развива-
ется в моторную кору. Различные ассоциативные области коры, а также высшие
отделы сенсорной и моторной коры преимущественно исходят из ткани между
этими сторонами.
В радиальном измерении нервная трубка образует определенные слои,
свойственные взрослому мозгу. Выпуклости увеличиваются в размере в ради-
альном направлении и становятся более характерными. В этих выпуклостях
клетки пролиферируют (рождаются), мигрируют (перемещаются) и диффе-
ренцируются в разные типы. Большинство клеток в мозге образуются в специ-
альных пролиферативных зонах (рис. 15 .7). Эти области лежат на внутренней
поверхности трубки, около полого центра (который образует позже желудочки
мозга). Одной из таких областей пролиферации является вентрикулярная зона,
которая, возможно, является филогенетически древней (Nowakowski, 1987).
Другая, субвентрикулярная зона, формирует филогенетически более молодые
структуры мозга, как неокортекс (т. е. «новая» кора, так как появляется только
у млекопитающих). Эти две зоны содержат различные типы глиальных (опор-
ные и питающие клетки) и нервных клеток, и в них происходят разные формы
клеточной миграции.
Рис. 15.7 . Судьба и миграция
клеток нервного гребня. В одну
клетку-предшественник
был
введен радиоизотопный маркер,
и ее потомки были прослежены
во время их миграции из нерв-
ной трубки. Некоторые стали
сенсорными нейронами, другие
дифференцировались в Шван-
новские клетки или нейроны
автономной нервной системы.
Среда, через которую проходи-
ли эти клетки при миграции,
повлияла на вид их дифферен-
цировки. (Источник: согласно
Bronner-Fraser and Fraser, 1991;
Sanes et al., 2006.)
Развитие
203
Нейроны и глиальные клетки образуются путем деления клеток в зоне про-
лиферации, где образуются клоны (клон представляет собой группу клеток,
образовавшуюся в результате деления одной предшествующей клетки — та -
кие клетки-предшественницы формируют линии клеток) (рис. 15 .8). Нейроны
образуются из нейробластов, каждый из которых дает начало ограниченному
числу определенных нейронов. В некоторых случаях нейробласты формируют
конкретные типы нейронов. Например, меньше дюжины нейробластов обра-
зуют все клетки Пуркинье коры мозжечка, при этом каждый нейробласт дает
начало примерно 10 000 клеток (Nowakowski, 1987).
Рис. 15.8 . Базовая родословная клеток центральной нервной системы позвоночных
животных. В результате работ с различными клеточными культурами и исследований
in vivo были установлены связи между различными классами клеток в нервной систе-
ме. Ранние клетки нервной трубки имеют способность порождать огромное количество
клеток-потомков, в результате чего их иногда называют клетками-основательницами
или стволовыми клетками, которые претерпевают симметричное клеточное деление
для образования, как дополнительных стволовых клеток, так и клеток-предшественниц.
(Термин «стволовые клетки» также используется для описания сохраняющихся клеток-
предшественниц во взрослом организме.) Считается, что ранние стволовые клетки мо-
гут также образовывать клетки-предшественницы, которые имеют способность к огра-
ниченному числу клеточных делений. По этой причине клоны клеток-предшественниц,
меченные на более поздних стадиях эмбриогенеза, имеют меньшее число потомков. Тем
не менее поздние клетки-предшественницы способны формировать как нейроны, так
и макроглию, олигодендроциты и астроциты. И хотя в исследованиях in vitro отдель-
ных частей нервной системы, а именно зрительного нерва, показано, что астроциты
и олигодендроциты имеют общего предшественника — глиального предшественника
О2А, в спинном мозге мотонейроны и олигодендроциты имеют общее происхожде-
ние. Следовательно, родословная клеток может различаться для разных областей ЦНС.
(Источник: Sanes et al., 2006.)
204
Глава 15
2.3 .
Миграция нейронов
После образования новые нейроны должны мигрировать из зоны пролифе-
рации в определенные области, где они будут функционировать во взрослом
мозге. Наиболее распространенным способом миграции является пассивное
замещение клеток. В этом случае молодые клетки просто выталкиваются от
зоны пролиферации вновь образовавшимися клетками. В результате этого фор-
мируется градиент, в котором наиболее старые клетки оказываются на внешней
поверхности мозга, в то время как наиболее молодые на внутренней. Такой тип
миграции наблюдается при формировании таких структур как таламус и мно-
гих других областей ствола мозга.
Второй более активный вид миграции заключается в том, что молодые клет-
ки перемещаются за образовавшиеся в предыдущем цикле деления клетки,
создавая при этом градиент изнутри-наружу (рис. 15 .9). Такой вид миграции
происходит в коре больших полушарий и подкорковых структурах, имеющих
ламинарное строение (разделенное на параллельные слои) (рис. 15 .10).
Наиболее подробно активная миграция была изучена Паско Ракичем (Pasko
Rakic, 1988) в пренатальной развивающейся коре, а также была предложена
модель радиальной единицы. Как было сказано ранее, большинство корковых
Рис. 15.9 . Гистогенез в коре головного мозга происходит в три стадии. На первой ста-
дии гистогенеза вся толщина коры головного мозга состоит из клеток-предшественниц,
которые образуют вентрикулярную зону (ВЗ). На следующей стадии развития первые
нейроны вступают в клеточный цикл (красные) и собираются в области предпластин-
ки, прилегающей к поверхности мягкой мозговой оболочки. Нейроны предпластинки
могут быть разделены на клетки субпластинки и находящиеся на поверхности клетки
Кахаля–Ретциуса. На третей стадии коркового гистогенеза вновь образовавшиеся ней-
роны (красные) мигрируют вдоль отростков радиальных глиальных клеток и формиру-
ют слой между клетками Кахаля–Ретциуса и субпластинкой. Этот слой называется кор-
ковой пластинкой, и большинство нейронов коры головного мозга будут расположены
в нем. (Источник: Sanes et al., 2006.)
Развитие
205
нейронов у человека образуются вне самой коры, в области, расположенной
под формирующейся корой, в «пролиферативной зоне». Обратите внимание,
что у человека кора больших полушарий наиболее развита по сравнению с дру-
гими видами. Значит, все эти клетки должны мигрировать, чтобы занять свое
конечное положение в коре. Выдвинутая Ракичем «модель радиальной едини-
цы» для описания процессов дифференцировки неокортекса дает объяснения,
как образуются и региональные структуры, и слоистые структуры коры боль-
ших полушарий головного мозга млекопитающих (Rakic, 1988). Согласно этой
модели, ламинарное строение коры головного мозга определяется тем, что каж-
Рис. 15.10 . Гистогенез пирамидных нейронов глубоких слоев, V и VI. (а) После обра-
зования и миграции клеток Кахаля–Ретциуса и клеток субпластинки следующими об-
разующимися клетками коры головного мозга являются пирамидные нейроны глубоких
слоев, V и VI, аксоны которых идут к подкорковым структурам. (б) Следующие образу-
ющиеся клетки — интернейроны в IV слое коры. (в) И наконец, образуются пирамид-
ные клетки в верхних слоях, II и III. Они посылают проекции к другим областям коры
головного мозга. ПЗ — промежуточная зона, КП — корковая пластинка. (Источник:
Sanes et al., 2006.)
206
Глава 15
дая значимая единица деления дает начало примерно ста нейронам. Потомство
этих пролиферативных единиц мигрируют вверх по одному и тому же отростку
радиальной глиальной клетки. При этом нейрон, образовавшийся последним,
мигрирует дальше своих более старых собратьев. Отросток радиальной гли-
альной клетки очень длинный, берет начало от глиальной клетки и простира-
ется снизу вверх через всю поверхность коры. По сути, отростки радиальной
глиальной клетки являются канатом, который обеспечивает образование одной
радиальной колонки внутри коры из потомков одной пролиферативной едини-
цы (рис. 15 .11).
Существует интересное следствие из модели радиальной единицы, кото-
рое объясняет видовые различия коры головного мозга. Как утверждает Ра-
кич (1988), увеличение числа клеточных делений на стадии пролиферативной
единицы приведет к увеличению числа корковых колонок, а значит, и общей
площади поверхности коры головного мозга. Напротив, один дополнительный
Рис. 15.11 . Модель радиальной единицы П. Ракича (1988). Отростки радиальных гли-
альных клеток простираются от вентрикулярной зоны (ВЗ) до корковой пластинки (КП)
через несколько областей: промежуточную зону (ПЗ) и зону субпластинки (СП). РГ —
отросток радиальной глиальной клетки. Каждый мигрирующий нейрон (МН) проходит
через ПЗ и СП, которые содержат ожидающие терминальные окончания таламической
радиации (ТР) и кортико-корковых афферентов (КК). Как было описано в тексте, после
достижения корковой пластинки нейроны мигрируют мимо своих предшественников к
краевой зоне (КЗ). (Источник: Rakic et al., 2009.)
Развитие
207
цикл делений на более поздних стадиях, вне пролиферативной зоны, приве-
дет только к увеличению размера колонки на одну клетку (примерно на 1%).
Это хорошо соотносится с фактом отсутствия сильных различий между видами
млекопитающих в общей структуре слоев коры, в то время как площадь по-
верхности коры может различаться между видами в 100 или более раз. Таким
образом, видовые различия возникают (по крайней мере, частично) из-за числа
«циклов» клеточного деления, которые происходят в пролиферативной зоне.
Так, эмбрион в течение ранних недель созревания проходит ряд сложных
процессов, формирующих основы центральной нервной системы. Очень важно
учитывать, что пренатальное развитие мозга не является пассивным процессом,
основанным на разворачивании генетического материала. Напротив, с ранних
стадий взаимодействия между клетками необходимы, включая передачу элек-
трических сигналов между нейронами. Вот один пример: паттерны спонтан-
ных разрядов клеток в зрительной системе (перед открытием глаз в процессе
развития) дают сигналы, которые, вероятно, определяют слоистую структуру
зрительного таламуса, латеральных коленчатых тел (ЛКТ) (Shatz, 2002; O’Leary
and Naka�awa, 2002). Таким образом, импульсы разрядов, присущие развиваю-
Naka�awa, 2002). Таким образом, импульсы разрядов, присущие развиваю-
Naka�awa, 2002). Таким образом, импульсы разрядов, присущие развиваю-
, 2002). Таким образом, импульсы разрядов, присущие развиваю-
щемуся организму, могут играть важную роль для определения особенностей
строения мозга задолго до того, что сенсорные входы из внешнего мира произ-
ведут какой-либо эффект.
2.4 .
Пересмотр роли врожденного и приобретенного
Как мы уже упоминали в начале этого раздела, влияние среды в пренатальный
период на развертывание генетической информации и развитие мозга могут
иметь долгосрочные последствия, как положительные, так и отрицательные.
Развивающийся младенец очень восприимчив к событиям, происходящим в его
окружении. Одним из таких события является подверженность воздействию
тератогенов. Тератоген — это агент окружающей среды, который вызывает
нарушения в пренатальном периоде. Примерами тератогенов являются лекар-
ства отпускаемые по рецептам, и также без них, кофеин из кофе и прохлади-
тельных напитков, наркотики (например кокаин и героин), продукты табака и
марихуаны, алкоголь. Эффекты тератогена(ов) могут быть комплексными в за-
висимости от дозы, времени появления в пренатальном развитии и генетиче-
ских данных матери, так как некоторые организмы более восприимчивы к их
действию, чем другие. Пренатальная консультация для женщин, готовящихся
стать матерями, обеспечивает их знаниями о потенциальных источниках тера-
тогенов, и таким образом снижает вероятность появления повреждений мозга
детей от тератогенов.
Мозг в пренатальном периоде чрезвычайно восприимчив к действию алко-
голя. Злоупотребление алкоголем матери во время беременности имеет долго-
срочные эффекты на когнитивную деятельность, речь и социальное развитие,
и называется плодным алкогольным синдромом (ПАС) (�ones and Smith, 1973;
�ones, 1975). Современные исследования детей, подростков и молодых взрос-
, 1975). Современные исследования детей, подростков и молодых взрос-
лых с тяжелым ПАС выявили отличия в плотности серого вещества у паци-
ентов с ПАС по сравнению с контрольной группой людей соответствующего
пола и возраста. Характерными результатами были уменьшение плотности се-
рого вещества во фронтальных и теменных областях и увеличение в височных
208
Глава 15
областях и нижней части теменной доли (рис. 15 .12). Как мы узнаем дальше,
эти области мозга созревают в течение всего детства до позднего подросткового
периода; а значит, такие различия в плотности серого вещества у людей с ПАС
указывают на то, что алкоголь оказывает повторяющиеся и длительные эффек-
ты на развитие мозга и когнитивной деятельности на всем протяжении жизни.
Длительные повторные обследования, оценивающие долговременные эффекты
курения табака или марихуаны во время беременности, выявили новые данные
об их влиянии на развитие когнитивной деятельности у ребенка. Фрид с колле-
гами (Fried et al., 2003) проследили в Канаде группу детей от рождения до взрос-
ления. Используя батарею нейрофизиологических тестов для выявления уров-
ня когнитивных функций, как грамотность речи, зрительно-пространственное
восприятие, речевые способности и внимание, Фрид с коллегами обнаружили
рано возникающие (к трем годам) и продолжительные когнитивные наруше-
ния, вызванные курением табака или марихуаны матерью во время беремен-
ности. Специфичные эффекты пренатального влияния табачного дыма очень
сильно отличаются от эффектов марихуаны, хотя они оба вредны. Воздействие
сигаретного дыма вызывало снижение общего интеллекта у ребенка совмест-
но с нарушениями слуховых процессов и вербальной рабочей памяти, кото-
рые продолжались с раннего детства (3 лет) до подросткового периода (16 лет)
(Fried et al., 2003). Влияние курения марихуаны не влияло на общий интеллект,
однако высшие психические функции, такие как внимание и рабочая память, в
особенности зрительная обработка, например зрительная интеграция, анализ
и объяснение, у таких детей были повреждены. Данные нарушения также на-
блюдались в раннем возрасте (3 года) и продолжались в подростковом возрасте.
Когнитивные нарушения, выявленные нейрофизиологическими тестами,
определили для дальнейшего исследования пренатального влияния табака и
марихуаны специфичные области мозга. Процессы зрительно-пространствен-
Рис. 15.12. Различия в плотности серого вещества у детей с плодным алкогольным
синдромом (ПАС) и нормально развивающихся детей из контрольной группы. Теплы-
ми (красным, желтым) цветами отмечены положительные изменения, указывающие на
увеличение плотности серого вещества (а значит, уменьшение при нормальном форми-
ровании) в этих областях по сравнению с контролем. Обратите внимание, что у детей с
ПАС обнаружено увеличение плотности серого вещества в области височной доли, пре-
имущественно в левом полушарии. (Источник: Sowell et al., 2002 из To�a et al., 2006.)
Развитие
209
ной интеграции используют для изучения лобных долей у взрослых (см. гл . 6).
Фрид с коллегами (Smith et al., 2006) продолжили исследование с помощью
фМРТ долговременных эффектов пренатального воздействия марихуаны у мо-
лодых людей (18–22 года) из Канадской группы. Результаты работы выявили
различия в паттернах нервной активности в лобных отделах, задействованных
при выполнении зрительно-пространственных задач. Эти характерные осо-
бенности заключались в уменьшении активности в областях правого полуша-
рия (рис. 15.13, верхняя часть) и увеличении активности левого полушария
(рис. 15 .13, нижняя часть). Авторская интерпретация результатов заключалась
в том, что нейронные сети правого полушария, необходимые для зрительно-
пространственной кратковременной памяти, менее активны у детей, подвергав-
шихся пренатальному воздействию марихуаны. Несмотря на то что на данном
этапе исследований сложно объяснить, почему увеличивается активность в ле-
вом полушарии, можно предположить, что области левого полушария задей-
ствованы в компенсации сниженной активности в правом полушарии. Опре-
деленно требуется большее число исследований для того, чтобы полностью
понять эффекты пренатального воздействия курения табака и марихуаны. Од-
нако работа Фрида с коллегами обеспечила нас данными о том, что эти наруше-
ния возникают в раннем возрасте и сохраняются длительное время. Четким вы-
водом из данного исследования является то, что ранние инсульты, вызванные
тератогенами, такими как табак или марихуана, могут привести к когнитивным
нарушениям на всю жизнь и соотносятся в различными паттернами коркового
развития.
Рис. 15.13. Эффекты пренатального воздействия марихуаны у молодых взрослых
(18–22 лет), измеренные при помощи фМРТ. Участки лобных долей, задействованные
при решении задач на зрительно-пространственную рабочую память, в правом полуша-
рии менее активны (верхняя часть рисунка), в то время как в левом полушарии (нижняя
часть) наблюдается повышенная активность по сравнению с контролем соответствую-
щего возраста. (Источник: Smith et al., 2006.)
210
Глава 15
2.5 .
Пренатальный слуховой опыт:
восприятие голоса и музыки до рождения
Что уже знают дети к моменту рождения? Важно ли разговаривать будущей
матери со своим еще не рожденным ребенком? Читать ребенку? Петь своему
ребенку? Есть ли влияние в последующем на речевые, музыкальные и когни-
тивные функции? Другими словами, существует ли способность к восприятию
у нерожденного ребенка и как она влияет на последующее когнитивное разви-
тие? Этот вопрос не давал покоя физиологам развития более ста лет (Kisilevsky
and Low, 1998), однако систематические исследования восприятия у плода не
проводились до 1980-х гг. Как можно измерить ответную реакцию плода на
звуки? Обычно исследователи измеряют частоту сердечных сокращений и ино-
гда движения тела в ответ на различные звуки. Эти первые работы выявили, что
примерно к 30-й неделе созревания плод слышит и отвечает на простые звуки.
К 37–42-й неделе плод начинает различать типы речевых звуков (как гласные,
слоги из согласных и гласных) (Lecanuet et al., 1987; Groome et al., 1999).
Открытие того, что плод слышит и различает звуки до рождения, привело
к исследованиям, что же плод знает об определенных звуках, а именно голо-
се матери. ДеКаспер с коллегами исследовали слуховые предпочтения у ново-
рожденных в ряде работ в 1980-х гг. Они выяснили, что новорожденные дети
предпочитают слышать голос своей матери, а не голос незнакомой женщины
(DeCasper and Fifer, 1980). Эти данные вызвали спор об основах восприятия
речи: отражает ли предпочтение голоса матери новорожденным генетически
обусловленную предрасположенность к человеческой речи? Или предпочтения
новорожденных детей отражают их пренатальный опыт? Современная рабо-
та, выполненная Кисилевски с коллегами (Kisilevsky et al., 2003), дополнила
результаты ДеКаспера более детальными данными о том, что плод на поздних
стадиях созревания отличает голос своей матери от голоса посторонней жен-
щины. Они отмечали, что частота сердечных сокращений у плода увеличива-
лась, когда он слышал голос матери, и уменьшалась, когда плод слышал го-
лос незнакомого человека. Несмотря на то что результаты работ Кисилевски
с коллегами (Kisilevsky et al., 2003) не разрешают спор между врожденным и
приобретенным о том, является ли генетической или приобретается с опытом
способность к четкому восприятию речи у ребенка, они дают нам уникальные
факты, что плод не только может различать знакомые и незнакомые голоса, но
у него также существует уже до рождения особое предпочтение голоса матери.
В настоящее время наблюдается всплеск интереса к эффектам пренаталь-
ного воздействия музыки. Такое повышение внимания обусловлено утверж-
дениями о том, что пренатальное восприятие музыки улучшает интеллекту-
альные или когнитивные функции ребенка при последующем развитии. Эти
утверждения основаны на отдельных примерах, без должной строгой научной
основы. Давайте познакомимся с данными, которые мы действительно имеем
о восприятии музыки сразу после рождения: существует много работ по ис-
следованию способности к восприятию музыки у младенцев в первые месяцы
жизни. Есть сообщения о том, что уже в возрасте двух месяцев ребенок вос-
принимает ритмический паттерн в музыке (Trehub and Hannon, 2005). Авторы
утверждают, что существует много параллелей между ранними музыкальными
знаниями у ребенка и особенностями у взрослого человека, которые подтверж-
Развитие
211
дают мнение о наличии врожденной или биологической основы для восприя-
тия музыки.
Очень мало известно о восприятии музыки до рождения. Тем не менее Ки-
силевски с коллегами (Kisilevsky et al., 2004) сообщают, что плод на поздних
сроках созревания (33–37-я неделя развития) может различать изменения рит-
ма в музыкальных фрагментах. Важно, что на более ранних этапах созревания
(28 недель) у плода не отмечен данный эффект, что привело Кисилевски к вы-
воду о развитии способности к восприятию музыки in utero, а именно возмож-
ности более зрелого плода выделять особенности музыкальных фрагментов
по сравнению с плодом на более ранних сроках созревания. В целом работы
Кисилевски с коллегами доказали, что слуховое восприятие более высокого по-
рядка материнского голоса и музыки не только происходит до рождения, но и
претерпевает изменения, созревает.
Исключает ли восприятие плодом человеческого голоса и музыка до рожде-
ния идею о генетической предрасположенности к речи и музыки? Не совсем.
Толкование сложных взаимодействий экспрессии генов и опыта, которые лежат
в основе человеческой речи и восприятия музыки, продолжает исследоваться в
области когнитивной нейронауки развития. Разрабатываются новые техноло-
гии и экспериментальные парадигмы для того, чтобы ответить на этот очень
важный вопрос о развитии человека.
3.0 .
Развивающийся мозг: изменения д линою в жизнь
3.1.
Начало и завершение постнатального развития мозга
В то время как общее строение мозга новорожденного ребенка в значительной
степени соотносится с таковым у взрослых, и большинство нейронов достигли
своего конечного местоположения, во время постнатального развития проис-
ходят значительные дополняющие изменения в мозге. В частности, объем моз-
га увеличивается в четыре раза с момента рождения до взросления в связи со
многими причинами, но в основном не из-за увеличения числа нейронов. Об -
разование и миграция нейронов происходит у человека практически полностью
в период пренатального развития. И хотя в гиппокампе и других областях может
появляться некоторое число нейронов, преобладающее большинство нейронов
уже присутствует к седьмому месяцу внутриутробного развития (Rakic, 1995).
Вероятно, наиболее заметным изменением в период постнатального раз-
вития является увеличение в размере и сложности строения дендритического
древа большинства нейронов. Пример грандиозного увеличения дендритиче-
ских отростков у человека в постнатальном периоде показан на рис. 15 .14 . По
мере того как дендритические отростки клетки разрастаются, они очень часто
приобретают специализацию и характерные свойства.
В дополнение к более распространенным процессам, протекающим по ис-
ходящим и приходящим волокнам клетки, происходит стабильное увеличение
плотности синапсов в большинстве областей коры головного мозга у человека
(Huttenlocher et al., 1982; Huttenlocher, 1990, 1994). Процесс формирования но-
Huttenlocher, 1990, 1994). Процесс формирования но-
, 1990, 1994). Процесс формирования но-
вых синапсов, синаптогенез, начинается примерно в одинаковое время в мо-
мент рождения во всех исследованных на данный момент областях коры, но
наибольшая интенсивность этого процесс и окончательное увеличение плотно-
212
Глава 15
сти контактов происходит в разное время в разных областях. Например, в зри-
тельной коре интенсивный синаптогенез происходит с 3 до 4 месяцев, и макси-
мальная плотность, равная примерно 150% от зарегистрированной у взрослых
людей, наблюдается между 4 и 12 месяцами. В отличие от этого, в префрон-
тальной коре плотность контактов увеличивается намного медленнее и до-
стигает пика только после первого года жизни, хотя синаптогенез начинается
в тоже время, что и в зрительной коре.
Миелинизация является еще одним дополняющим процессом. Миелиниза-
ция — это увеличение жировой оболочки, которая окружает нейронные отрост-
ки и окончания и увеличивает эффективность электрической передачи. Из-за
того что миелинизация в корковых областях продолжается многие годы после
рождения, существуют различные предположения о ее роли в поведенческом
развитии (Yakovlev and Lecours, 1967; Parmelee and Si�man, 1983; Volpe, 1987).
По причине увеличенного содержания жировой ткани в мозге вследствие ми-
елинизации волокон, структурная фМРТ может четко контрастировать белое
и серое вещество, что позволяет производить количественное определение их
объема в процессе развития (Sampaio and Truwit, 2001). Несмотря на то что су-
Sampaio and Truwit, 2001). Несмотря на то что су-
and Truwit, 2001). Несмотря на то что су-
and Truwit, 2001). Несмотря на то что су-
Truwit, 2001). Несмотря на то что су-
Truwit, 2001). Несмотря на то что су-
, 2001). Несмотря на то что су-
ществуют противоречия при интерпретации снимков, сделанных детям младше
6 месяцев, существует соглашение, что к возрасту двух лет облик мозговых
структур становится схожим с таковым у взрослых, и все основные проводящие
тракты могут наблюдаться к трехлетнему возрасту (Huttenlocher and Dabholkar,
1997; Bour�eois, 2001). Изменения в протяженности белого вещества вызывает
интерес в связи с тем, что предположительно отражает межрегиональные вза-
имодействия в развивающемся мозге. Хотя увеличение протяженности бело-
го вещества наблюдается еще и в подростковом возрасте, преимущественно во
фронтальных отделах мозга (Huttenlocher et al., 1982), наиболее быстро данные
изменения происходят в первые 2 года. Миелинизация начинается с рождения
в мосту и мозжечковых ножках и к трем месяцам охватывает оптическую ради-
ацию и валик мозолистого тела. В возрасте 8–12 месяцев становится различи-
мым белое вещество фронтальных, теменных и затылочных областей.
Поразительно, но в постнатальном развитии мозга у человека происходят и
значительные регрессивные события. Одним из количественно определяемых
Рис. 15.14 . Древовидное ветвление дендритов. Рисунки Конела (1939–1967) клеточной
структуры нейрона из зрительной коры человека по препаратам, окрашенным по Голь-
джи. (Источник: Quartz, 1999, из Conel, 1953.)
Развитие
213
нейроанатомических показателей регрессивных событий является плотность
синапсов, указывающая на существование периода утраты или сокращения си-
напсов (Huttenlocher, 1990, 1994). Как и продолжительность синаптогенеза и
последующие пики в плотности контактов, время сокращения синаптической
плотности варьирует в областях коры головного мозга, по крайней мере, у лю-
дей. Например, синаптическая плотность в зрительной коре начинает возвра-
щаться на уровень, свойственный взрослым людям, после двух лет, в то время
как аналогичные значения достигаются в префронтальной коре только к под-
ростковому периоду. Huttenlocher (1990, 1994) предполагает, что это играет
важную роль в очевидной пластичности молодого мозга.
Таким образом, начало и завершение последовательности развития мож-
но наблюдать с помощью ряда различных измерений нейроанатомического и
физиологического развития в коре человека. Однако необходимо постоянно
помнить, что не все показатели могут отражать этот процесс (например, миели-
низация) и что измерения плотности синаптических контактов являются ста-
тическими снимками динамического процесса, в котором одновременно проис-
ходят и дополняющие, и регрессивные события.
В то время как большая часть данного раздела была посвящена коре головно-
го мозга, такие структуры мозга, как гиппокамп и мозжечок, также подвергают-
ся постнатальному развитию. Действительно, существование постнатального
развития у некоторых подкорковых структур (таких как гиппокамп, мозжечок
и таламус) вызывает некий парадокс; с одной стороны, наличие большого ко-
личества поведенческих и нейрофизиологических данных указывает на функ-
ционировании этих структуру при рождении, с другой — есть данные о пост-
натальном развитии и/или функциональной реорганизации. Одно возможное
решение данной проблемы заключается в том, что кора больших полушарий
развивается в постнатальном периоде, поэтому ее взаимодействия с подкорко-
выми структурами также претерпевают определенные изменения. Таким обра-
зом, в то время как некоторые подкорковые структуры способны функциониро-
вать относительно независимо от коры на раннем этапе жизни, созревание коры
требует от них некоторых структурных и функциональных приспособлений.
3.2 .
Региональные особенности развития мозга
По сравнению с другими животными людям требуется значительно более дли-
тельное время для становления самостоятельными взрослыми существами. На-
пример, постнатальное развитие коры у человека происходит почти в четыре
раза дольше, чем у нечеловекообразных обезьян. Отрицательной стороной та-
кого медленного развития является то, что в течение многих лет ребенок чрез-
вычайно зависим от опеки членов семьи. Положительный момент затянувше-
гося развития заключается в том, что мозг человека обладает гораздо большими
возможностями изменяться и формироваться под воздействием опыта и обще-
ния с другими людьми.
Как было предложено выше, начало и завершение дополняющих и регрес-
сивных процессов развития возникают в разных областях и долях мозга в раз-
ное время. Эти события чрезвычайно сильно зависят от опыта и отражают фор-
мирование синапсов и ветвление дендритов, необходимых для когнитивного и
сенсорного развития, обучения и интегративных процессов, которые возникают
на всем протяжении младенчества и детства и продолжаются в подростковом
214
Глава 15
периоде. Динамика развития мозга от момента зачатия до позднего подрост-
кового возраста представлена на рис. 15 .15 (Casey et al., 2005). Пренатальные
изменения показаны в левой части рисунка и отражают преимущественно про-
цессы нейруляции, клеточной пролиферации и миграции. Постнатальные изме-
нения включают такие процессы развития, как синаптогенез и ветвление ден-
дритов. Например, сенсорные области для восприятия зрительной и слуховой
информации формируются раньше префронтальных областей, ответственных
за произвольные функции (Casey et al., 2005).
Как объективно измерить развитие мозга в течение жизни? Один из спосо-
бов для прослеживания изменений развивающегося мозга состоит в измерении
плотности серого вещества в областях коры головного мозга. Большое охваты-
вающее разных испытуемых исследование 176 человек от 7 до 87 лет выявило
различные паттерны уменьшений плотности серого вещества в течение всей
Рис. 15.15. Ход развития человеческого мозга. Мозг человека претерпевает значитель-
ные изменения как в структурной, так и функциональной организации, что отражает
динамические взаимодействия синхронно возникающих прогрессивных и регрессив-
ных событий. Несмотря на то что в шестилетнем возрасте вес мозга составляет около
90% от веса мозга взрослого человека, в мозге продолжают происходить динамические
изменения в подростковом периоде и на ранних этапах взросления. На рис. 15.15 проил-
люстрированы некоторые изменения, связанные с развитием, включая пролиферацию
и миграцию клеток, преимущественно на стадии созревания плода, и региональные
изменения синаптической плотности в постнатальном периоде и затянувшееся разви-
тие во взрослом возрасте. Современные неинвазивные методы нейровизуализации не
имеют разрешающей способности, достаточной для разделения процессов, лежащих в
основе изменений компонентов серого и белого вещества в процессе развития. (Источ-
ник: Casey et al., 2005 из Thompson and Nelson, 2001.)
Развитие
215
жизни (To�a et al., 2006). В верхней лобной борозде плотность серого вещества
быстро уменьшается начиная с подросткового периода. Однако в верхней ви-
сочной борозде уменьшение плотности серого вещества происходит постепен-
но в течение всей жизни (рис. 15 .16).
В то время как мы начинаем прослеживать паттерны развития отдельных об-
ластей мозга в течение жизни, центральный вопрос заключается в индивидуаль-
ных различиях созревания мозга и соотношении этого с когнитивным развитием.
Другими словами, какова связь между мозгом и поведением? Отражают ли раз-
ные паттерны развития мозга разные уровни интеллектуальных способностей?
Эти вопросы были исследованы в современной работе с нейровизуализацией
плотности серого вещества в большой группе из более 300 детей и подростков
(Shaw et al., 2006). Данные исходно были разделены на три группы в зависи-
мости от уровня коэффициента IQ. «Превосходная» группа имела значения IQ
от 121 до 149; «высокая» — от 109 до 120; и «средняя» группа имела IQ от 83
до 108. Результаты исследования утверждают о наличии различий в паттернах
развития мозга в зависимости от общего уровня интеллекта (рис. 15 .17). Важ-
ное открытие заключается в том, что высокие уровни коэффициента интеллекта
связаны с наличием более тонкой коры, особенно в лобных и височных долях,
в раннем детстве. В позднем периоде детства был отмечен противоположный
эффект — наличие более толстой коры у детей с высоким уровнем IQ.
Важное открытие этой работы — разные паттерны развития лобных обла-
стей мозга у детей из «превосходной» группы по сравнению с другими груп-
пами. В частности, существуют различия в темпе утолщения и утончения коры
в период от раннего детства до подросткового возраста и взросления. Авторы
сделали заключение, что различия в плотности серого вещества само по себе
не приводило к появлению превосходных интеллектуальных способностей.
Напротив, они предположили, что динамические свойства развития коры со-
относятся с уровнем интеллекта, влияя на способность выделять больше ин-
формации из окружающей среды. Один из вопросов, поднятых этой работой,
заключается в том, обусловлены ли генетической предрасположенностью дина-
мические изменения в паттернах развития мозга у детей «превосходной» груп-
пы? Или они отражают различия в окружающей среде? Или зависят от комби-
нации генетических влияний и опыта?
Результаты современных работ по исследованию развития мозга на протя-
жении жизни помогают создавать новые модели когнитивного развития чело-
века и соотношения с познанием. Эти работы еще находятся на раннем этапе
в ряде продолжающихся исследований сложных процессов созревания мозга.
Но, несмотря на ранние стадии изучения, на настоящий момент данные указы-
вают, что мозг динамически меняется на всем протяжении детства и подрост-
кового возраста.
4.0 .
Развитие мозга и сознания
Мы знаем, что мозг претерпевает значительные изменения в ходе развития в
детстве. Важным вопросом в области когнитивной нейронауки развития явля-
ется вопрос о том, как изменения в мозге отражают развитие когнитивных про-
цессов. Новые технологии позволяют нам отмечать развитие мозга и соотносить
с сознанием. Возникновение долговременных междисциплинарных исследова-
216
Глава 15
Развитие
217
Рис. 15.16 . Картирование мозговых изменений во времени. Изменения мозга во вре-
мя развития могут быть выделены при помощи соотнесения статистических моделей с
временной зависимостью и данных, полученных по сечению (т. е . у всех членов группы
в определенное время), продольно (т. е . прослеживая индивидуальных испытуемых по
мере взросления) или обоими способами одновременно. Измерения, такие как корковая
толщина, наносят на изображение коры, используя цветовой код. (а, б) Траектория по-
тери серого вещества в течение жизни у человека, основанная на исследовании группы
из 176 человек в возрасте от 7 до 87 лет (Sowell et al., 2003). Точки, перенесенные на
изображение мозга (б), отражают уменьшение плотности серого вещества в определен-
ных областях; (а) яркие участки указывают на области, в которых плотность серого ве-
щества быстро уменьшается во время подросткового периода (верхняя лобная борозда)
или имеет более пологую динамику уменьшения в течение жизни (верхняя височная
борозда) (в, г). (Источник: To�a et al., 2006.)
Рис. 15.17. Корреляция между уровнем интеллекта IQ и корковой плотностью. (а) Кор-
реляции Пирсона для всех 307 испытуемых были преимущественно положительными и
значимыми (p > 0,05), со значениями r от 0 до 0,10 (зеленый/желтый), кроме передней
височной коры (которая показала отрицательную корреляцию с r в пределах от 0 до –1;
синий/фиолетовый). (б) В группах разного возраста отрицательные корреляции в самой
младшей группе указывали на то, что более высокий уровень IQ соотносится с более
тонкой корой, преимущественно во фронтальных и височных областях. Это отношение
меняет знак в более позднем возрасте, когда плотность большей части коры положи-
тельно коррелирует с уровнем IQ. (Источник: Shaw et al., 2006.)
218
Глава 15
Как учатся младенцы
Речь – самые сложные звуки, которые люди расшиф-
ровывают. Центральный вопрос для ученых, иссле-
дующих развитие человека: как младенцы осваива-
ют процесс расшифровки этого сложного сигнала?
Вы, скорее всего, в детстве или подростковом воз-
расте изучали иностранный язык в школе. В данном
процессе вам помогали некоторые факторы, кото-
рые отсутствуют у маленьких детей, осваивающих
их первый язык: во-первых, вы осознавали, что учи-
те новый язык; во-вторых, вы могли сопоставлять
новую информацию с уже полученными знаниями
вашего первого языка (например, вы можете вы-
учить, что aqua – это слово для обозначения воды);
в-третьих, вы обладаете большим объемом суще-
ствующих знаний о строении своего языка (напри-
мер, вы знаете, что отдельные звуки/фонемы вместе
образуют слоги и слова).
У младенцев нет таких металингвистических знаний. Они просто находятся в высо-
ко мотивированном состоянии для общения с членами семьи и опекунами. Поэтому
некоторые ученые задаются вопросом, как младенцы обучаются расшифровывать
речь? Как они начинают понимать, что практически непрерывный речевой поток
содержит отдельные слова и фразы? Другими словами, как младенцы выделяют
информацию о границах слова в речи?
У доктора Дженни Саффран из Университета Висконсина есть некоторые удиви-
тельные объяснения того, как дети справляются с этим. Доктор Саффран считает,
что важным аспектом при первом обучении языку является то, что младенцы не-
осознанно вычленяют статистические закономерности в речевом потоке (Saffran
et al., 1996; Aslin et al., 1998). Вот как, по ее мнению, это происходит: в любом
языке существуют звуковые паттерны, которые возникают с определенной вероят-
ностью. Несмотря на то, что все языки имеют множество индивидуальных фонем,
специфичных для этого языка звуков, они формируют ограниченное множество.
И варианты формирования из них слов — их фонология — также конечны. Как
считает доктор Саффран, младенцы выделяют условную вероятность (УВ) звуко-
вых паттернов в речи (рис. 15.19). Значит вероятность, с которой определенный
звуковой паттерн будет следовать за другим предъявленным звуковым паттерном,
обусловливается вероятностью первого звукового паттерна.
Например, в английском языке за слогом «ba» очень часто следует слог «by» для
создания слова «baby» (от англ. малыш), и данная комбинация возникает намного
чаще, чем комбинация слогов «ba» и «sin» для формирования слова «basin» (от англ.
миска). Тогда УВ у таких комбинаций слогов соответствует вероятности «by», обу-
словленной вероятностью «ba», и вероятности «sin», обусловленной вероятностью
ПЕРСПЕКТИВЫ КОГНИТИВНОЙ НАУКИ
Рис. 15.18 . Дженни Саф-
фран (�enny Safran, PhD),
Центр Вейсмана, Универси-
тет Висконсина, Мэдисон,
штат Висконсин, США
Рис. 15.19 . Схема искусственной грамматики, которую использует Саффран. (Ис-
точник: Saffran et al., 1996/)
Развитие
219
«ba». И, несмотря на то что вероятность появления в речевом потоке слога «ba»
одинакова, УВ значительно различается при комбинировании с «by» или «sin».
Согласно представлениям доктора Саффран, младенцы выделяют такие вероятно-
сти и научаются формировать границы слов, используя статистические обучающие
механизмы. Определение УВ в речи позволяет младенцам определять границы слов
и в конечном счете усваивать язык.
Работа Дж. Саффран с коллегами в буквальном смысле перевернула образ рас-
суждений ученых из области развития и речи о процессе обучения у младенцев.
Это привело к появлению новой волны исследований, включая новые работы под
руководством доктора Саффран в Лаборатории детского обучения. Вот некоторые
основные направления текущего исследования, проводимого в ее лаборатории.
1. Естественная речь: в ранних работах доктор Саффран использовала упрощенный
набор стимулов и грамматики для выявления формирования восприятия речи у
младенцев. В текущих работах, она исследует процессы обучения у маленьких
детей с помощью естественных языковых стимулов. Результаты на настоящий
момент показывают, что младенцы так же хорошо и даже лучше обучаются есте-
ственной речи (Pelucchi et al., 2009).
2. Обучение в реальном времени: в лаборатории Саффран наблюдают в реальном
времени, как младенцы обучаются, исследуя эффекты статистических способ-
ностей по мере того, как они возникают. Понимание языка у взрослых также об-
легчается статистической информацией — знанием того, что последует дальше.
Доктор Саффран в настоящий момент проверяет гипотезу о том, что дети также
используют статистическую информацию во время понимания речи, что измеря-
ется прослеживанием за взором.
3. Объединение клинических данных: в лаборатории Саффран исследовали, как
обучаются языку нормально развивающиеся младенцы и дети. В настоящий мо-
мент они расширяют свою работу для того, чтобы посмотреть обучаются ли та-
ким же образом дети с речевыми нарушениями или потерей слуха (Evans et al.,
2009).
Несмотря на то что доктор Саффран не изучает мозговые основы этих рано воз-
никающих механизмов обучения у младенцев, ее работа поднимает интересные
вопросы: где в мозге младенцев происходят эти процессы? Отражают ли они про-
явление основных механизмов обучения, которые вовлечены в освоение языка, но
также используются и для остального когнитивного развития? Или они представля-
ют множественные — возможно параллельные — системы обучения, которые объ-
единяются для формирования основы умозрительных представлений младенцев и
детей? Эти вопросы в области когнитивной нейронауки детского развития оста-
ются пока без ответа, и нам придется подождать того, что новые работы выяснят
мозговые механизмы, лежащие в основе обучения у младенцев!
Литература
1. Saffran, �.R., Aslin, R.N . and Newport, E.L. (1996). Statistical learnin� by 8-month-
old infants. Science, 274 (5294), 1926–1928.
2. Aslin, R.N ., Saffran, �.R ., and Newport, E.L . (1998). Computation of conditional
probability statistics by 8-montj-old infants. Psychological Science, 9 (4), 321–324 .
3. Pelucchi, B., Hay, �. and Saffran, �.R . (2009). Statistical learnin� in a natural lan�ua�e
by 8-month-old infants. Child Development, 80 (3), 674–685.
4. Evans, �.L., Saffran, �.R . and Robe-Torres, K. (2009). Statistical learnin� in children
with specific lan�ua�e impairment. Journal of Speech, Language, and Hearing
Research, 52, 321–335.
220
Глава 15
ний в комбинации с работами по нейровизуализации корковых изменений и по-
веденческие исследования выполнения когнитивных задач дадут нам хорошие
возможности выявить взаимосвязь между развитием мозга и сознанием. В дан-
ном разделе мы обсудим современные данные о связи между созреванием моз-
га и процессами развития ребенка. Во время того, как мы будем освещать не-
которые современные работы, необходимо помнить, что область когнитивной
нейронауки развития чрезвычайно молода. Первые фМРТ-исследования детей
были опубликованы в 1995 г. (Casey et al., 1995). Таким образом, несмотря на
то, что представленные далее результаты очень содержательны, ответ на вопрос
о нервной основе приобретения речи, процессов когнитивного контроля и со-
циальной когнитивной деятельности до сих пор изучаются в лабораториях по
всему миру.
Мы обсудили функциональные основы для понимания когнитивных процес-
сов. Ученые из области когнитивной нейронауки развития пытаются понять,
как системы и процессы, наблюдаемые у здоровых взрослых людей (рис. 15 .20),
развиваются и формируются в период младенчества, детства и подросткового
возраста. Снова в данном разделе будет рассмотрен вопрос о роли врожденного
и приобретенного. Предрасположен ли мозг человека к речи? Восприятию лиц?
Или эти процессы основываются на жизненном опыте? Так как подобные во-
просы находятся в центре интереса при исследовании человеческого развития,
мы остановимся на работах в трех основных областях когнитивной нейронау-
ки развития, которые могут пролить свет на эти вопросы: возникновение речи;
развитие механизмов произвольного поведения; и развитие социальной когни-
тивной деятельности с уклоном в восприятии лиц. В данном разделе мы дадим
краткий обзор современных работ, выполненных на детях первого года жизни.
Далее мы предоставим результаты таких же исследований детей старшего воз-
раста и подростков. И наконец, проанализируем эффекты ранних (пренаталь-
ных) повреждений этих систем.
Рис. 15.20 . Функциональная структура развития, которая является адаптированным ва-
риантом общей функциональной структуры, представленной в гл. 2
Развитие
221
4.1.
Первый год жизни: взрывной рост и развитие
Мозг человека увеличивается в четыре раза в период от рождения до взросле-
ния. Многие изменения, возникающие при развитии, происходят во время пер-
вого года жизни. Во время этого 12-месячного отрезка младенец развивается
из крохотного существа с очень маленьким набором произвольных движений в
активного сорванца, который тянется к привлекательным объектам, произносит
множество речевых звуков, ползает и даже ходит, исследуя мир вокруг.
4.1.1. Формирование речевого мозга: речевые способности младенца
Помните исследования, демонстрирующие, что ребенок может слышать и вы-
делять голос матери до рождения? Такие работы обеспечили веские доказатель-
ства тому, что новорожденный ребенок уже имел опыт восприятия человече-
ской речи. Мы уже обсуждали роль врожденного и приобретенного, а именно
генетическую предрасположенность к речи или влияние опыта. Этот спор явля-
ется важным в области когнитивной нейробиологии развития, и много идет ра-
бот для определения, что знает о языке младенец. Большинство исследований
маленьких детей (младше 12 месяцев) сфокусировано на классах и категориях
речевых звуков — фонологии. Работы со старшими детьми также включают
изучение их знаний о семантике (смысловой основе) и синтаксисе (граммати-
ческой основе).
Является ли язык «биологически специализированным»? Один из подходов
решить этот вопрос — посмотреть, маленькие дети обладают особой чувстви-
тельностью к человеческой речи. Если существуют определенные нервные кор-
реляты обработки речи, наблюдаемые на очень раннем этапе жизни, это может
указывать на наличие нервных процессов, связанных с речью, до появления
значимого опыта.
Один из примеров такого подхода имеет отношение к возможности дискри-
минировать значимые для речи звуки, фонемы (см. гл . 7). В каждом челове-
ческом языке есть набор звуков, разделяемых на отдельные фонемы, которые
обычно при написании заключают между косыми чертами: /п/, например, для
отображения звуков (фонов), которые накладываются на фонему /п/. Вспомни-
те проблему «нехватки постоянства», которую обсуждали в гл. 7: различающи-
еся фонемы, которые произносятся до и после произнесения /п/, меняют его
акустические свойства. Таким образом, не существует единого инвариантного
физического свойства, которое однозначно определит /п/. Вместо этого пред-
ставление фонемы /п/ должно основываться на некоем абстрактном (не только
физическом) свойстве. Этот аспект человеческой речи интенсивно использо-
вался в исследованиях восприятия речи, в которых фонемы, различающиеся од-
ним признаком, создаются синтезирующим речевые звуки программным обе-
спечением так, что они различаются на градуированный шаг, как например, /б/
и /п/, различающиеся только в исходном произношении (вибрации голосовых
связок).
Англоязычные взрослые, при прослушивании градуированного фонети-
ческого перехода от речевых звуков «ba» к «pa» , будут воспринимать звуки
промежуточных форм между /ba/ и /pa/, которые будет являться либо одним,
либо другим. Другими словами, мы имеет категориальную границу между
ними двумя. Поведенческие эксперименты выявили, что маленькие дети также
222
Глава 15
обладают способностью к улучшенной (категориальной) дискриминации при-
меняющихся в речи фонетических границ, как /ba/ и /pa/. То есть градуирован-
ba/ и /pa/. То есть градуирован-
/ и /pa/. То есть градуирован-
pa/. То есть градуирован-
/. То есть градуирован-
ный фонетический переход от /ba/ к /pa/ также воспринимается младенцами
как неожиданный категориальный сдвиг. Эти наблюдения изначально вызвали
воодушевление как доказательство существования у человека механизма, спец-
ифичного для восприятия человеческой речи. Однако более современные ис-
следования показали, что другие виды, такие как шиншиллы, обладают сход-
ными способностями акустической дискриминации. Сходство указывает на то,
что эта способность может отчасти отражать основные характеристики систе-
мы обработки слуховых сигналов у млекопитающих, а не исходный механизм,
специфичный для устной речи (Werker and Vouloumanos, 2001).
Интригующе, но в отличие от взрослых, человеческие младенцы могут ис-
ходно выделять очень широкий диапазон фонетических конструкций, включая
и такие, которые отсутствуют в их родном языке. Например, японские младен-
цы, но не взрослые, могут различать звуки «р» и «л» . Однако примерно к 10
месяцам данная способность ограничивается фонетическими конструкциями
родного языка. Эти результаты могут отражать ранние речевые перцептивные
процессы в младенчестве, которые основываются на физических или акустиче-
ских свойствах всех речевых звуков и развиваются позже в механизмы, меньше
опирающиеся на физические признаки и более на абстрактные образы фонем
их родного языка. Следовательно, роль опыта имеет сильное влияние на фор-
мирование знания языка у младенца.
Если бы можно было выявить мозговые корреляты этих процессов, стало
бы возможным изучать механизмы, лежащие в основе такого специфичного
к речи выборочного снижения чувствительности. Для решения этого вопро-
са использовали связанные с событием потенциалы (ССП). Когда компонен-
ты ССП различаются по латентности (следования за событием) и простран-
ственным распространением, можно утверждать об активации в мозге разных
нервных сетей. В одном из экспериментов (Dehaene-Lambertz, Dehaene, 1994)
младенцам предъявляли серию стимулов, в которой последовательность четы-
рех идентичных слогов (стандартных) завершалась пятым, который был либо
идентичным, либо фонетически отличным (девиантным) от них. Исследовате-
ли синхронизовали ССП по времени начала предъявления слога и получили два
пика разной локализации на черепе. Первый пик возникал около 220 мс после
предъявления стимула и не уменьшался в длинном ряду стимулов (исключение
составляет первое предъявление) или увеличивался в ответ на новый стимул.
Таким образом, области генерации этого компонента, вероятно, первичные и
вторичные слуховые поля в височной доле, не проявляют чувствительности к
незначительным акустическим различиям, которые определяют фонетическую
информацию.
Второй компонент достигал максимума около 390 мс после предъявления
стимула и снова не проявлял привыкания в повторению одного слога, кроме
случая первого его предъявления. Однако когда в последовательность был вве-
ден девиантный слог, амплитуда компонента возвратилась к первому исходно-
му значению. Значит, нервные генераторы этого компонента также располо-
жены в височной области, но в ограниченной и расположенной в задней ее
части, чувствительны к фонетической информации. Необходимы последующие
Развитие
223
исследования для выяснения, происходит ли восстановление амплитуды второ-
го компонента вследствие категориального восприятия или причина в измене-
нии звука.
В дальнейших работах для изучения ранних коррелятов восприятия речи
были применены два метода визуализации с лучшей пространственной разре-
шающей способностью (Dehaene-Lambertz et al., 2006). Эти авторы измеряли
активацию мозга у трех месячных младенцев в ответ на произнесение речи в
прямом и обратном порядке. Они выяснили, что прямая и обратная речь активи-
ровали различные, хотя и близко расположенные, области мозга в речевой зоне
левой височной доли (рис. 15 .21). Было также обнаружено, что прямая речь
активирует правую префронтальную кору, в то время как обратная речь нет. Эти
результаты доказали, что начиная с третьего месяца жизни у младенцев наблю-
дается левосторонняя латерализация функции речи, которая также активирует
фронтальные области мозга.
Общие выводы выше изложенных работ подкреплены сходящимися резуль-
татами новых методологий — ближней инфракрасной спектроскопии (БИКС).
Рис. 15.21 . Уже к возрасту трех месяцев у малышей наблюдается левосторонняя лате-
рализация процессов человеческой речи. Изображения мозга слева направо отражают:
(I) активацию мозга в височной доле в ответ на обратную речь (оранжевый); (II) моз-
говой ответ в более высоком мозговом срезе, демонстрирующий активацию в ответ на
обратную речь и прямую речь (синий); (III) активацию мозга в более высоком срезе
на прямую речь; (IV) различие в активации мозга при предъявлении прямой и обрат-
ной речи, состоящее в активации правой префронтальной коры. (Источник: Dehaene-
Lambertz et al., 2006.)
224
Глава 15
В одной из работ Мелер (Mehler) с коллегами (Pena et al., 2003) проигрывали
нормальную обращенную к младенцу речь или такие фрагменты в обратном
порядке во время измерения колебаний концентрации общего гемоглобина в
областях в левом и правом полушариях мозга. Они обнаружили, что в левой
височной области происходит более сильная активация при предъявлении нор-
мальной речи по сравнению с обратной речью или тишиной, что привело их к
выводу: новорожденные уже рождаются с предрасположенностью левого полу-
шария к обработке речи.
Понимание языка и восприятие речи являются наиболее активно развива-
ющимися областями когнитивной нейронауки развития. Использование пере-
крывающихся методик и частое сравнение обычной и нетипичной линий разви-
тия делают данную область основным претендентом на грандиозный прорыв в
следующем десятилетии (обсуждение слуховых механизмов, лежащих в основе
развития речи у детей с аутизмом, см. Ga�e et al., 2003a, b).
4.1.2. Развитие управляющего мозга: что знают младенцы?
Критическим аспектом в когнитивном развитии младенца в первый год жиз-
ни является способность узнавать новое об их окружении. Новые объекты при
смотрении на них будут приковывать внимание маленького ребенка более дли-
тельное время, чем объекты, привычные для взора. В то время как для младен-
ца чрезвычайно важно смотреть на новые предметы, также важно ориентиро-
ваться относительно других объектов в окружающей среде. Смена действий по
смотрению на новый предмет и перевод внимания на другие элементы окружа-
ющей среды обеспечивают младенцам возможность учиться выявлять свойства
новых объектов и дают большой диапазон такого опыта, изменяя фокус их вни-
мания, что является критичным для когнитивного развития.
Произвольный контроль направления внимания к новым стимулам, ориен-
тация, поддержание целей и контроль над хватательными движениями требуют
участия наиболее фронтально расположенных областей мозга, префронталь-
ной коры (ПФК) (см. гл. 12). Как было рассмотрено ранее, лобная кора у чело-
века требует наиболее длительного периода постнатального развития, с нейро-
анатомическими изменениями, происходящими даже в подростковом возрасте,
по сравнению с любой другой областью мозга (Huttenlocher, 1990; Giedd et al.,
1999). По этой причине данную область мозга часто ассоциируют с развитием
когнитивных способностей в детстве.
Одна из наиболее исчерпывающих попыток связать когнитивные изменения
с мозговыми изменениями, лежащими в их основе, касается возникновения у
малыша чувства постоянства объекта. Неисчезаемость объекта — способность
сохранять образ объекта в уме после его исчезновения за другим объектом
или покрывалом (рис. 15 .22). В частности, Пиаже (Pia�et) наблюдал, что дети
младше 7 месяцев не способны правильно находить после короткого периода
задержки спрятанный объект, если объект помещали в другое место по срав-
нению с предыдущими и успешными попытками. В особенности младенцы в
данном возрасте делают характерные повторяющиеся ошибки, когда неодно-
кратно тянутся к укрытию, где предмет был найден в непосредственно предше-
ствующий пробе. Этот характерный паттерн ошибок, названный «А не Б», был
рассмотрен в работе Пиаже (1954) как доказательство неспособности младен-
цев понять, что объекты не исчезают или постоянны, когда скрываются из виду.
Развитие
225
С возраста 7 месяцев малыши начинают лучше справляться с этой задачей при
увеличении периода задержки от 1 до 5 с (Diamond, 1985, 2001).
Было обнаружено (Diamond, Goldman-Rakic, 1989), что младенцы обезьян
также совершают ошибки в адаптированной версии теста Пиаже о постоян-
стве объектов. Похожие ошибки наблюдали у взрослых обезьян с повреждени-
ем дорсолатеральной префронтальной коры (ДЛ-ПФК). Повреждения других
Рис. 15.22 . Поведенческие тесты у малышей. (а) Задание по извлечению объекта, с ко-
торым ребенок не справляется до возраста 9 месяцев. На глазах у малыша эксперимен-
татор прячет предмет в первое место, и ребенок успешно его находит. После нескольких
таких подходов экспериментатор прячет предмет во второе место, но ребенок продолжа-
ет искать в предыдущем месте (Pia�et, 1954). (б) Метод зрительного привыкания может
быть использован для демонстрации того, что уже с четырехмесячного возраста малы-
ши воспринимают нарисованный слева объект как непрерывную палочку, двигающую-
ся за блоком. У детей происходит снятие эффектов привыкания (обнаружение новизны)
при предъявлении двух коротких палочек; это указывает на то, что они перцептивно
«заполняли» пространство за нарисованным блоком в пробах с привыканием. Малы-
ши младше 4 месяцев только частично справлялись с таким заданием, что зависело от
сложности изображений на экране. (в) Младенец наблюдает за двумя последовательно-
стями событий, возможных и невозможных, в которых заслонка вращается по направ-
лению к трехмерному кубу. При «возможном» варианте событий заслонка останавлива-
ется, когда достигает твердого объекта. При невозможном событии заслонка проходит
через объект. Уже в четыре месяца дети начинают удивляться (смотреть дольше), когда
им демонстрируют невозможные действия, что указывает на то, что они воспринимают
объект как трехмерный и (как правило) несжимаемый. (Источник: �ohnson, 2001.)
226
Глава 15
областей коры не вызывали таких эффектов, что указывает на определенную
роль ДЛ-ПФК для выполнения этого задания.
Результаты, связывающие данные изменения в поведении с развитием моз-
га, также исходят из ЭЭГ-исследований малышей человека (Fox and Bell, 1990;
Bell, 1992a, b; Bell and Fox, 1992). В данных работах увеличение ЭЭГ-ответов
во фронтальных отделах мозга коррелирует со способностью правильно отве-
чать после более длительных периодов задержки задания с отсроченным выбо-
ром. Совсем недавно работы по оптической визуализации выявили у младенцев
корреляцию между успешным выполнением АБ-задачи и уровнем насыщения
крови кислородом в префронтальной коре (Baird et al., 2002).
Схожие данные о необходимости развития лобной коры получены из об-
следований детей с нарушением нейрохимического баланса в префронтальной
коре вследствие фенилкетонурии (ФКН) (Welsh et al., 1990; Diamond, 2001).
Даже при лечении это метаболическое расстройство может привести к опреде-
ленным последствиям в виде уменьшения уровня нейромедиатора дофамина в
дорсолатеральной префронтальной коре. В результате данное снижение уровня
дофамина в дорсолатеральной префронтальной коре у младенцев и детей вызы-
вает нарушение выполнения задач, вовлекающих активацию префронтальной
коры, как задач на постоянство объектов, и не влияет на выполнение заданий,
зависящих от других областей коры, как тест отставленного не сопоставления
по образцу (Welsh et al., 1990; Diamond, 2001).
Таким образом, сходные данные из нескольких источников подтверждают
точку зрения, что развитие ДЛ-ПФК позволяет младенцам успешно выполнять
задания на постоянство объектов. Согласно Диамонду (Diamond, 1991), крити-
Diamond, 1991), крити-
1991), крити-
ческие для выполнения заданий процессы, вовлекающие ДЛ-ПФК, включают
способность сохранять информацию во время периода задержки и подавлять
доминирующие (подкрепленные прежде) ответы. Однако недавно полученные
факты указывают на то, что гипотеза развития ДЛ-ПФК не полная и требует
некоторых модификаций или совершенствований. Один из фактов обусловлен
другим заданием, для выполнения которого также требовалась активация ДЛ-
ПФК: отставленный глазодвигательный ответ (рис. 15 .23). Было обнаружено
(Gilmore, �ohnson, 1995), что младенцы начинают успешно выполнять это за-
Gilmore, �ohnson, 1995), что младенцы начинают успешно выполнять это за-
�ohnson, 1995), что младенцы начинают успешно выполнять это за-
�ohnson, 1995), что младенцы начинают успешно выполнять это за-
1995), что младенцы начинают успешно выполнять это за-
дание в более раннем возрасте, чем указывается в заданиях по извлечению объ-
ектов, несмотря на то что оно также требует сохранять пространственную ин-
формацию в период отсрочки и подавлять предустановленные ответы.
Одним вариантом объяснений такого несходства между выполнением те-
ста Пиаже на постоянство объекта и результатами последней работы (Gilmore,
�ohnson, 1995) является то, что задания, требующие в качестве ответов движе-
1995) является то, что задания, требующие в качестве ответов движе-
ния глазами (смотрение), более простые, так как планирование движений глаза-
ми развивается более быстро по сравнению с другими формами двигательных
актов, например хватания. Это предположение согласуется с несколькими ра-
ботами, показавшими, что младенцы могут успешно выполнять аналог теста
на постоянство объекта уже в возрасте 4 или 5 месяцев, когда в качестве ответа
определяют направление взора, а не движение руки (Lecuyer et al., 1992).
Для того чтобы объяснить различия результатов выполнения заданий
с движением глаз и хватательных движений конечности, предположили, что
у младенцев отсутствуют навыки планирования действий для координации
Развитие
227
последовательности двигательных актов при извлечении спрятанного предмета
(Diamond, 1991; Baillar�eon, 1993). Для проверки этой идеи обучали семиме-
Diamond, 1991; Baillar�eon, 1993). Для проверки этой идеи обучали семиме-
, 1991; Baillar�eon, 1993). Для проверки этой идеи обучали семиме-
Baillar�eon, 1993). Для проверки этой идеи обучали семиме-
, 1993). Для проверки этой идеи обучали семиме-
сячных младенцев получать предметы, расположенные в удалении, при помо-
щи нажатия на кнопку или потянув за тряпочку (Munakata et al., 1994). Малыши
извлекали предметы в случае помещения прозрачной перегородки между ними,
но не в случае установки достаточно непроницаемой ширмы, делающей объ-
ект невидимым. Так как выполнение заданий с прозрачной и непроницаемой
перегородками требовало одинакового планирования действий, было сделано
заключение, что оно никак не влияет на различия между задачами на смотрение
и хватание. Мунаката выдвинул альтернативную теорию различий, выполнен-
ную в виде модели нервной сети. Эта модель иллюстрирует, что слабые вну-
тренние образы стимула могут успешно вызывать простую ответную реакцию,
как движение глазами, но не способны инициировать более сложный моторный
выход, как хватание.
Альтернативный подход к пониманию роли префронтальной коры в когни-
тивном развитии был усовершенствован несколькими учеными, которые пред-
положили, что эта область играет важную роль в приобретении новой инфор-
мации и навыков. С этой точки зрения, вызов, который получает мозг малыша,
например, при обучении доставать предмет, эквивалентен, в некотором роде,
таковому у взрослого человека при его столкновении со сложными моторными
навыками, такими как обучение вождению автомобиля. Из этого можно сде-
лать три прогноза: (1) корковые области, необходимые для выполнения опреде-
ленного навыка, будут меняться в процессе обучения; (2) префронтальная кора
играет роль в организации или размещении информации в других областях
Рис. 15.23 . Задача на отставленный глазодвигательный ответ, разработанная для при-
менения на младенцах. Малыш смотрит на три монитора, на которых появляются яркие
цветные стимулы. В начале каждой пробы на среднем экране предъявляли фиксаци-
онный стимул. Как только ребенок направлял на него свой взор, стимул-подсказка на
короткий период времени загорался на одном из боковых мониторов. После кратковре-
менной подсказки, центральный стимул продолжал светиться еще в течение 1–5 с, перед
предъявлением двух целевых стимулов на боковых экранах. Измерение отставленных
смещений взора на цель в направлении предшествующей подсказки выявило, что мла-
денцы способны сохранять информацию о положении подсказки в течение нескольких
секунд. (Источник: Gilmore and �ohnson, 1995.)
228
Глава 15
коры; и (3) развитие включает установление иерархических структур управле-
ния с локализацией высшего уровня контроля в лобной коре. Современные дан-
ные, свидетельствующие об активации ПФК в младенчестве, также подтверди-
ли это положение. Ограниченное число фМРТ и ПЭТ исследований, которые
проводили на младенцах, часто выявляли активацию в ПФК, даже когда этого
эффекта не наблюдалось у взрослых людей. Например, в фМРТ-исследовании
восприятия речи у трехмесячных детей (Dehaene-Lambertz et al., 2006) авторы
наблюдали правостороннюю активацию ДЛ-ПФК при дискриминации (пря-
мой) речи у бодрствующих, но не спящих малышей (см. рис. 15 .21). Схожий
паттерн активации в ДЛ-ПФК был найден в ответ на предъявление лиц в этом
же возрасте (Tzourio-Mazoyer et al., 2002). Это является доказательством ак-
тивации в первые месяцы, по крайней мере, некоторых областей ПФК, однако
возможно, что данная активация является пассивной, так как не играет роли
в управлении поведением младенца.
Исследования с регистрацией ССП процессов развития часто выявляли
изменение активности в лобных отведениях у малышей, и некоторые авторы
предполагают, что данная активность имеет важные последствия для поведе-
ния. Эти эксперименты основаны на измерении паттернов активации, предше-
ствующей началу саккады. Например, обнаружено (Csibra et al., 1998, 2001),
что саккадические пре-потенциалы, которые обычно регистрируют у взрослых
людей в задних отведениях, у шестимесячных детей выявляются в лобных ка-
налах. Так как эти потенциалы связаны по времени с возникновением движе-
ния, то разумно предположить, что они являются результатом процессов, не-
обходимых для планирования или выполнения действий.
Дальнейшие доказательства необходимости ПФК для развития с самого ран-
него детства получены из работ по изучению длительных и широко распростра-
ненных эффектов перинатального повреждения ПФК. Избирательное перина-
тальное повреждение значимых областей, которые активируются у взрослых
людей, оказывает, в худшем случае, умеренный эффект на малышей, который
впоследствии практически полностью исчезает. В отличие от этого перинаталь-
ное повреждение лобной коры и ПФК часто вызывает как немедленные, так
и отсроченные последствия. Эти обобщения ряда работ свидетельствуют, что
ПФК может играть важную структурирующую или облегчающую роль с ран-
них этапов постнатального развития.
В начале данного раздела поднят вопрос относительно того, как согласовать
данные о продолжающемся до подросткового периода нейроанатомическом
развитии лобной коры, с одной стороны, и данные о функционировании этой
области с первых месяцев жизни, с другой стороны. Одним из возможных вари-
антов объяснения этой проблемы служит то, что представления, возникающие
в этих областях исходно слабы и способны только контролировать некоторые
типы реакций, такие как саккады, но не другие, например хватание (Munakata
et al., 1994). Другие решения этого вопроса основываются на предположении
(Diamond, 1991), что разные области лобной коры по-разному отсрочены в раз-
Diamond, 1991), что разные области лобной коры по-разному отсрочены в раз-
1991), что разные области лобной коры по-разному отсрочены в раз-
витии, и высказывании (Thatcher, 1992), что префронтальные области играют
непрерывную роль в циклической реорганизации остальных отделов коры.
Подтверждаются данные гипотезы или нет, но существует важная причина,
почему определенный уровень функционирования ПФК жизненно необходим
Развитие
229
с первых недель постнатального периода и даже раньше (Fulford et al., 2003).
Способность формировать и поддерживать цели, хоть и на короткий период
времени, является основополагающим свойством для генерации усилий при
выполнении действий по хватанию объекта. Первые и часто исходно безрезуль-
татные попытки выполнить двигательный акт обеспечивают базовый опыт, не-
обходимый для последующего развития.
4.1.3. Развитие социального мозга: лица и места
Как было описано в гл. 14, социальная природа является основной характери-
стикой человеческого мозга. Множество корковых областей, включая верхнюю
височную борозду (ВВБ), веретеновидную область лиц (ВОЛ) и орбитофрон-
тальную кору, вовлечены в «социальный мозг». Один из основных вопросов
когнитивной нейронауки касается истоков «социального мозга» у человека и
изобилия теоретических доводов о степени их приобретения вследствие опыта.
Одним из аспектов функционирования социального мозга, который был
предметом исследования, является восприятие и обработка лиц (блок 15.1).
Долгая история исследований развития восприятия лиц у маленьких детей
Блок 15.1 . Предпочтения лиц у новорожденных
За последнее десятилетие было опубликовано более дюжины работ о зрительном
предпочтении лиц новорожденными детьми. Большинство авторов заключили, что
новорожденные приспособлены к восприятию стимулов, обладающих определенны-
ми характеристиками, но были высказаны также две альтернативные точки зрения.
Первое мнение («сенсорная гипотеза») состоит в том, что все зрительные предпо-
чтения новорожденных могут быть очень просто объяснены в терминах относитель-
ной видимости стимулов. Зрительная система новорожденных ограничена нижней
частью набора пространственных частот, свойственного взрослому человеку. Было
предположено, что новорожденные дети предпочитают смотреть на лица всего лишь
потому, что амплитуда на разных частотах у этих стимулов наилучшим образом со-
впадает с чувствительностью зрительной системы ребенка. Сенсорная гипотеза впа-
ла в немилость, так как даже при контроле амплитуды фазовая информация (конфигу-
рация) все равно смещала предпочтения младенцев к лицам. Попытки симулировать
предпочтения новорожденных в моделях нервных сетей, основанных на сенсорной
гипотезе, вряд ли смогут объяснить другие эксперименты с реалистичными лицами
внутри сложных зрительных образов, окружающих новорожденных.
Вторая альтернативная точка зрения указывает на наличие у человека от рождения
комплексных способностей к восприятию лиц. Результаты, использовавшиеся для
подтверждения этого мнения, включают: предпочтение изображений привлекатель-
ных лиц; данные, указывающие, что новорожденные чувствительны к наличию на
лице глаз; и доказательства того, что они предпочитают смотреть на лица, поддержи-
вающие с ними зрительный контакт. В дополнение к недоразвитию коры к моменту
рождения, все эти результаты могут объясняться низкочастотным пространственным
детектором конфигурации лиц. Например, малыши более старшего возраста пред-
почитают привлекательные лица, так как они наиболее приближены к среднему или
прототипному лицу. Пересмотр реалистичных изображений лиц через простран-
ственные частотные фильтры, присущие новорожденными, демонстрирует, что ме-
ханизм, чувствительный к конфигурациям лица, может быть активирован предпо-
чтительно (1) с помощью большинства предъявляемых прототипных конфигураций
лиц, (2) присутствием (или отсутствием) открытых глаз, и (3) направленным или от-
веденным взором (см. рис. 15.23).
230
Глава 15
относит нас к работам Фантца, сделанным более 40 лет назад (Fantz, 1964).
За последнее десятилетие многочисленные исследования рассматривали корко-
вую основу обработки лиц у взрослых, включая выделение областей, которые
могут быть специально предназначены для этих целей (см. гл . 6). Несмотря на
этот большой объем данных, удивительно мало известно о развитии когнитив-
ных процессов восприятии лиц (блок 15.2).
В обзоре всех доступных в 1980-х гг. литературных источников (�ohnson,
Morton, 1991) выявлено два явно противоречащих массива данных: в то время
как господствующая точка зрения и большая часть доказательств подтвержда-
ют ту идею, что младенцы постепенно в течение первых месяцев жизни узнают
о взаиморасположении признаков, составляющих лицо, результаты как мини-
мум одного исследования указывают на то, что новорожденный ребенок 10 мин
от роду будет дольше прослеживать взглядом за стимулом, подобным лицу, чем
за различными разрозненными паттернами лица (Goren et al., 1975). Данные о
том, что новорожденные продемонстрировали избирательный ответ на лица,
использовались некоторыми учеными для усиления нативистской точки зрения
на сознание младенца. В отличие от этого данные о постепенном развитии в
течение нескольких месяцев способностей воспринимать лица часто цитиру-
ются теоретиками, которые считают, что данный навык осваивается в процессе
обучения и является результатом познания мира.
Блок 15.2 . Почему в веретеновидной извилине формируется область лиц?
Центральный спор в когнитивной нейробиологии касается истоков и специфики
веретеновидной области лиц (ВОЛ) и других областей коры, чувствительных к ли-
цам. Согласно одной точке зрения, ВОЛ селективно активируется лицами благода-
ря генетически обусловленным и специализированным вычислительным свойствам
этой области. Другие ученые предполагают, что эта область вовлечена в обработку
зрительных стимулов в сфере перцептивной компетенции, и большинство людей
становятся специалистами по обработке лиц. С точки зрения развития, появляется
альтернативный «средний» вариант формирования ВОЛ. Согласно ему, части вере-
теновидной коры специализируются на обработке лиц в результате нескольких вы-
нуждающих факторов. Во-первых, наличие подкорковых связей, рассмотренных
в этой главе, гарантирует, что новорожденный предпочтительно ориентирует взор
на лица, и, следовательно, фокусирует изображение их на фовеа, обеспечивая тем
самым сенсорные входы к зрительным корковым путям. Во-вторых, проекционные
паттерны коры к подкорковым путям могут увеличить активацию определенных об-
ластей, включая веретеновидную кору, когда лицо возникает в поле зрения ребенка.
В-третьих, части веретеновидной коры, ставшие чувствительными к лицам, полу-
чают фовеальный корковый зрительный вход и переходят на «объектный уровень»
обработки зрительных стимулов в вентральном пути. Таким образом, информация от
обоих путей конвергирует в ВОЛ. Это и другие возможные условия, такие как муль-
тимодальные входы и основное смещение уровня экспрессии генов между правым
и левым полушариями мозга, объединяются для обеспечения специализации опре-
деленного развивающегося коркового участка к восприятию связанных с лицами
стимулов. Отсюда следует, что этот процесс специализации веретеновидной коры на
лица неизбежен и стоек к некоторым нарушениям в нормальных движущих факто-
рах. Однако такой неизбежный результат достигается без генетически обусловлен-
ных специализированных паттернов связей в ВОЛ.
Развитие
231
И хотя остаются некоторые вопросы, открытие того, что новорожденные
дети предпочтительно отвечают на стимулы, соответствующие лицам, счита-
ется доказанным (�ohnson, 2005b) (рис. 15 .24). Современные данные из многих
источников указывают на возможное участие подкорковых структур, включая
верхние бугры четверохолмия, подушку и миндалину, в обработке лиц (�ohnson,
2005b). Несмотря на то что данная сеть выявляется только у взрослых людей
с помощью функционального оптического картирования или после приобре-
тенных мозговых травм, она может оказывать основное влияние на поведение
новорожденного ребенка, у которого корковые сети обработки зрительной ин-
формации относительно недоразвиты.
Как мы можем исследовать восприятие лиц у младенцев? В некоторых ла-
бораториях наблюдали изменения связанных с событием потенциалов (ССП)
при рассматривании лиц взрослыми испытуемыми. В частности, интерес был
вызван компонентом N170 (негативная волна, возникающая после примерно
170 мс), который показал сильную связь с обработкой информации о лицах
в ряде исследований на людях. В особенности временные параметры и ам-
плитуда этого компонента изменялись в зависимости от того, находилось или
нет в зрительном поле добровольного испытуемого изображение лица. Важ-
ным свойством компонента N170 является то, что у взрослых людей этот от-
N170 является то, что у взрослых людей этот от-
170 является то, что у взрослых людей этот от-
вет высокоспецифичен. Например, компонент N170 проявляет разный ответ
на предъявление человеческих лиц в нормальной ориентации по сравнению с
очень близкими стимулами (перевернутые человеческие лица или лица обезьян
в нормальном положении). В то время как точные мозговые генераторы ком-
понента N170 не определены, специфичный ответ данного компонента может
быть использован как индекс степени специализации корковой обработки для
Рис. 15.24 . Восприятие лиц у новорожденных. Данные демонстрируют степень поворо-
та головы и глаз при прослеживании изображений схематичного лица, зашифрованного
лица и пустого (не разрисованного) шаблона. Младенец следует взором за изображени-
ем лица гораздо дольше, чем за другими стимулами. (Источник: �ohnson et al., 1991.)
232
Глава 15
восприятия прямых человеческих лиц. По этой причине была проведена (de
Haan, �ohnson et al., 2002) серия исследований по изучению формирования ком-
понента N170 в первые недели и месяцы постнатального периода.
Первый вопрос, изучавшийся в этих работах с регистрацией ССП, стоял в
определении времени возникновения сенситивного к лицам компонента N170.
В ряде работ был идентифицирован компонент вызванных потенциалов у мла-
денцев, который обладал многими свойствами компонента N170 у взрослых,
но имеющего более длительный латентный период (240–290 мс). Исследова-
ние изменений данного ответа у детей в возрасте 3, 6 и 12 месяцев выявило,
что: (1) компонент регистрируется, начиная с трехмесячного возраста (хотя
его развитие продолжается до середины детства), и (2) компонент в процессе
взросления становится более специфически настроенным на восприятие пря-
мых человеческих лиц. Для того чтобы расширить второе утверждение, было
показано, что в то время, как различные ССП ответы на прямые и перевер-
нутые лица были зарегистрированы у двенадцатимесячных детей и взрослых,
у трех- и шестимесячных детей различий обнаружено не было. Таким образом,
исследование данного сенситивного к лицам компонента ССП согласуется с
предположением об увеличении специализации корковых процессов с возрас-
том, что также согласуется с современными данными функциональной нейро-
визуализации.
В своем исследовании Шерф с коллегами (Sherf et al., 2007) проигрывали
короткие отрывки фильмов с лицами, объектами или зданиями детям (5–8 лет),
подросткам (11–14 лет) и взрослым. Они показали, что у детей отсутствовала
активация специфичных для восприятия лиц областей (рис. 15 .25). Эта рабо-
та доказывает, что, хотя обработка лиц может обладать исходным смещением
в нервных процессах, опыт играет ключевую роль в развитии селективной для
восприятия лиц коры (Kadosh and �ohnson, 2007).
Сходные результаты об увеличении в процессе развития специализации об-
ластей для обработки лиц получены также в поведенческом исследовании, в ко-
тором проверяли удивительную теорию о том, что по мере того, как обработка
«сужается» до человеческих лиц, младенцы теряют способность различать не-
человеческие лица. В то время как шестимесячные младенцы могут различать
индивидуальные лица как людей, так и обезьян, дети в 9 месяцев и взрослые
различают только человеческие лица (Pascalis et al., 2002). Эти результаты наи-
более веские, так как они демонстрируют предсказанную способность у малень-
ких детей, которая отсутствует у взрослых.
Необходимо выйти за рамки относительно простого восприятия лиц, и по-
говорить о более сложном свойстве зрелого социального мозга — обработке
информации о глазах. Существует два важных момента в обработке информа-
ции о глазах. Во-первых, возможность определять направление взгляда другого
индивида для перемещения своего собственного внимания на тот же объект
или точку в пространстве. Восприятие отведенного взгляда может вызвать ав-
томатическое переключение внимания в этом же направлении, что позволяет
установить «обоюдное внимание». Общее внимание к объектам является кри-
тичным для ряда аспектов когнитивного и социального развития, включая вы-
учивание слов. Вторым важным моментом восприятия направления взгляда
является определение направленного взгляда, что позволяет образовать обо-
Развитие
233
юдный взгляд со смотрящим. Обоюдный взгляд (зрительный контакт) является
основой для установления коммуникативной связи между людьми и считается
чрезвычайно важным для нормального социального развития. Общепринято,
что восприятие направления взора очень важно для взаимоотношений матери и
ребенка и обеспечивает существенную основу для социального развития.
В ряде экспериментов с четырехмесячными младенцами, использующими
простую парадигму подсказок взглядом, было показано (Farroni et al., 2000),
что только после периода зрительного контакта наблюдаются эффекты под-
сказок. Другими словами, обоюдный взгляд с прямым лицом может включать
механизмы внимания, благодаря которым смотрящий младенец воспользуется
подсказкой для последующего движения. В общих словах, критическими свой-
ствами для подсказок с помощью взора у младенцев являются: (1) боковые дви-
гающиеся элементы и (2) предшествующий краткий период зрительного кон-
такта с прямым лицом.
Вследствие удивительного наблюдения того, что необходим период направ-
ленного взгляда перед эффективными подсказками у младенцев, могут быть
исследованы ранние источники развития определения зрительного контакта.
Как уже известно, у новорожденных детей существует предрасположенность
к ориентации взгляда в направлении стимулов, похожих на лица (см. выше),
предпочтение к лицам с открытыми глазами и стремление к подражанию не-
Рис. 15.25. Активация мозга в ответ на предъявление лиц (показана красным), зданий
(показана зеленым) и объектов (показана синим) детям 5–8 лет (слева), подросткам
11–14 лет (в ц ентре) и взрослым (справа). Результаты показали, что у детей в возрасте
5–8 лет не наблюдается избирательность к лицам, как у подростков и взрослых. (Ис-
точник: Kadosh and �ohnson, 2007.)
234
Глава 15
которых лицевых выражений. Предпочтительное внимание к лицам с направ-
ленным взором убедительно доказало бы, что новорожденные подготовлены к
выделению социально значимой информации. По этой причине мы исследова-
ли распознавание направления зрительного взора у детей с самого рождения.
Фаррони с коллегами тестировали здоровых новорожденных детей, предъявляя
им пару стимулов, одно лицо со взглядом, направленным прямо на младенца,
и второе лицо с отведенным взглядом. Результаты показали, что время фикса-
ции было значительно больше для лица с направленным взором. Далее число
ориентаций было больше с прямым взором, чем с отведенным.
Во втором эксперименте схожие результаты по разной обработке направлен-
ного взора были получены при регистрации связанных с событием потенциалов
(ССП) с черепа младенцев, смотрящих на лица. Фаррони с коллегами изучали
четырехмесячных детей, предъявляя им те же стимулы, что и в экспериментах с
новорожденными детьми, и обнаружили различия между двумя направлениями
взоров во времени и пространственном расположении ранее выделенного чув-
ствительного к лицам компонента ССП, который обсуждался ранее. Выводом
из этих работ является то, что прямой зрительный контакт улучшает обработку
лиц у детей, начиная с возраста четырех месяцев.
Помимо восприятия лиц и направления взора, существует значительное
число более сложных аспектов социального мозга, таких как согласованное
восприятие человеческих действий и подходящее приписывание окружающим
намерений и целей. Исследование когнитивной основы этих способностей
у младенцев и детей является прологом для работ следующего десятилетия.
Один из способов, который возможен для ответа на эти вопросы, — изучение
генетических расстройств развития, при которых социальные аспекты когни-
тивной деятельности либо селективно нарушены (аутизм), либо селективно не
повреждены, несмотря на другие когнитивные нарушения (синдром Вильсона).
В первый год жизни происходят быстрые изменения как в мозге, так и по-
ведении. Несмотря на то что мы в отдельности рассмотрели появление речи,
произвольного контроля и социального сознания в этом оживленном периоде
первого года жизни, эти три области когнитивной деятельности человека слож-
ным образом взаимодействуют друг с другом в процессе развития. Например,
повышенный интерес младенцев к лицам поможет им освоить речь. Способ-
ность фокусироваться на новом объекте помогает малышам осваивать окружа-
ющий мир. Несмотря на то что мы еще недостаточно полно понимаем процес-
сы взрывного роста мозга и их связи с поведением малышей, такие работы, как
рассмотренные в данном разделе, помогают выявлять сложные взаимосвязи
между мозгом и разумом.
4.2 .
Детство и подростковый возраст:
динамичное и поэтапное развитие
Хотя первый год жизни является уникальным этапом быстрого развития че-
ловека, требуются годы для созревания многих структур мозга и когнитивных
процессов. Как уже упоминалось ранее, область когнитивной нейронауки раз-
вития очень молода. В данном разделе мы рассмотрим результаты некоторых
работ по изучению развития областей мозга, обеспечивающих речь, произволь-
ные действия и социальную когнитивную деятельность у детей и подростков и
их связь с поведением. Несмотря на то что эти исследования очень информатив-
Развитие
235
ны, необходимо постоянно помнить, что процесс соотнесения активации мозга
с выполнением когнитивных задач очень сложен. Например, может случиться
так, что мозговая активность, которую мы наблюдаем у взрослых людей (когда
когнитивные процессы сформированы), может задействовать разные области
мозга при развитии. Таким образом, простое сравнение активации интересу-
ющих областей мозга в экспериментах по нейровизуализации у групп детей и
взрослых не может обеспечить необходимый уровень точности для формулиро-
вания предположений о мозге и поведении. Аналогично, разные когнитивные
стратегии или механизмы психологической адаптации у детей и взрослых так
же могут влиять на сеть мозговых структур, задействованных в выполнении
определенных заданий.
С учетом этих предостережений познакомимся с обзором результатов от-
носительно развития нервных систем речи, произвольного контроля и социаль-
ного познания.
4.2.1 . Языковой мозг: приобретение речи
Речь не является единичным процессом: для того чтобы словесно выражать
свои мысли, нам надо пройти через стадии формулирования понятий, обла-
чения их в слова нашего мысленного лексикона, обращения к мнемонической
грамматике для формирования предложений и наложения этой информации
на звуковые образы артикуляции. Следовательно, система речи имеет множе-
ство стадий вычисления для многих категорий или аспектов языка. Интуитив-
но предполагается, что в раннем периоде жизни младенцы формируют свое
знание о языке преимущественно на основе устной речи, которую они слышат
в своем окружении. Поэтому неудивительно, что работы на маленьких детях
(младше 12 месяцев) обычно фокусируются на фонологии человеческой речи.
Исследования младенцев более старшего возраста, детей и подростков обычно
затрагивают другие аспекты языка: лексическо-семантический (основанный на
значениях) и синтаксический (основанный на грамматике).
Одинаковым ли образом развиваются разные аспекты речи и лежат ли в их
основе схожие процессы развития мозга? Этот вопрос задают ученые из об-
ласти когнитивной нейронауки развития при изучении нервного субстрата че-
ловеческой речи. Вот краткий обзор известных на настоящий момент фактов.
Мы уже обсуждали использование связанных с событием потенциалов
(ССП) для измерения динамики мозговых ответов на предъявления стимулов.
Некоторые компоненты ССП, которые мы охарактеризуем далее, используются
для изучения процессов формирования речи у младенцев и маленьких детей.
• Негативность рассогласования (НР) — негативный компонент, возни-
кающий через 100–250 мс после начала стимула, отражает способность
мозга различать акустические/фонетические признаки звуков
• N400 — негативная волна, возникающая около 400 мс, имеет генераторы
в центрально-теменных областях. Считается, что N400 отражает лекси-
N400 отражает лекси-
400 отражает лекси-
ческо-семантические процессы при восприятии слов и предложений
• Левая передняя негативность (ранняя, Р/ЛПН) возникает около 150–
350 мс и отражает текущие синтаксические процессы
• Р600 — позитивная волна, возникающая примерно через 600 мс после
стимула и имеющая центрально-теменное распределение. Р600 отражает
процессы, вовлеченные в синтаксическую проверку и повторный анализ.
236
Глава 15
Фридеричи (2005) описывает динамику развития речи у младенцев и детей
в современном обзоре литературы о детской речи (рис. 15 .26). Согласно этому
обзору, фонологические и интонационные процессы развиваются относитель-
но рано (с рождения до 9 месяцев), в то время как лексические, семантические
и синтаксические процессы развиваются позже (к 1–3 годам), по мере того как
младенцы и маленькие дети получают информацию на уровне слов и предло-
жений.
В течение первых двух лет жизни происходит интенсивный рост речевых
знаний и навыков во время того, как ребенок начинает лепетать, произносить
простые звуки («мама»), слова («собака») и словосочетания из двух или трех
слов («хочу сока»). К двухлетнему возрасту ребенок, как правило, может про-
износить большое число слов. Но что двухлетний ребенок понимает о значении
этих слов? Они просто повторяют услышанные слова/звуки? Или они обладают
знанием, выходящим за рамки простого повторения звуков? Ученые разрабо-
тали уникальные экспериментальные парадигмы для исследования семантиче-
ского знания у младенцев и маленьких детей. Один из методов заключается
Рис. 15.26 . Схематический обзор стадий развития восприятия устной речи и их ССП
коррелятов, которые обеспечивают возможность изучения фонологических, семанти-
ческих и синтаксических процессов. Стадии развития могут рассматриваться как взаи-
мосвязанные ступени, на которых новая информация извлекается и обрабатывается на
основе полученных ранее знаний. Как только базовые фонологические процессы сфор-
мированы, фонемные знания используются для определения и представления первых
лексических форм и создания большой базы лексических и семантических знаний, ко-
торая потом используется для выявления значения в контексте предложения. Приведен-
ная динамика разных стадий развития приблизительна и основана на опубликованных
работах по ССП. Данная схема также отражает взаимосвязь между возрастом и ком-
понентами ССП, зарегистрированными в различных рассмотренных в тексте работах.
(Источник: Friederici, 2005.)
Развитие
237
в предъявлении картинки с объектом, знакомым для ребенка, например нари-
сованной уткой, вслед за которой следует произнесение слова «утка» (условие
совпадения) или слова, которое не подходит к картинке («кошка», условие не
совпадения). Эти точные процедуры тестирования позволили ученым обсле-
довать очень маленьких детей с целью определения уровня их семантических
знаний. Результаты одного такого исследования представлены на рис. 15.27, где
показано, что уже к возрасту 14 месяцев у детей формируется семантическое
контекстное знание, что отражено в N400 в ответ на совпадающие и не совпа-
N400 в ответ на совпадающие и не совпа-
400 в ответ на совпадающие и не совпа-
дающие события (Friederici, 2005).
Что знают двухлетние дети о правилах грамматики своего родного языка?
Уровень обработки предложений исследовали у двухлетних детей и взрослых,
предъявляя им контрастирующие предложения правильного грамматического
строения («The lion roars» или «Лев рычит») и неправильного строения («The
lion in the roars» или «Лев в рычании»). Двухлетние дети способны различать
правильно сформулированное предложения от ошибочного, что отражено в
компонентах Р/ЛПН и Р600 (рис. 15 .28).
Что эти работы с регистрацией ССП говорят нам о процессах формирования
речи в детстве? Несмотря на то что такие исследования находятся на начальном
этапе, результаты отражают развитие методов регистрации ССП и экспери-
ментальных парадигм, которые предоставляют эффективные чувствительные
показатели формирования речевых навыков. Несмотря на то что необходимо
Рис. 15.27. Семантическая обработка совпадающих и несовпадающих слов. Мозговые
ответы детей в возрасте (а) 14 месяцев, (б) 19 месяцев и (в) взрослых. Изображение объ-
екта демонстрировали на экране в течение 4 с, через 900 мс после появления картинки
акустически предъявляли неопределенный артикль, вслед за которым через 1 с произ-
носили слово, либо совпадающее с картинкой (например, изображение утки и произне-
сение слова «утка»), либо нет (изображение утки и слово «кошка»). Жирные линии на
графике отображают ССП в ответ на произнесение совпадающего слова, а пунктирная
линия — несовпадающего слова. ССП показаны для одного регистрирующего электро-
да (PZ-отведение). Негативная волна (N400), возникающая около 400 мс, наблюдалась
во всех возрастных группах. ((а) Адаптировано с разрешения из Friedrich and Friederici,
2005. (б, в) адаптировано из Friedrich and Friederici, 2005. Источник: Friederici, 2005.)
238
Глава 15
Рис. 15.28 . Синтаксические процессы у двухлетних детей. (а) Мозговые ответы у
двухлетних детей и взрослых на нарушения структуры фразы в простых предложени-
ях. ССП показаны от двух отведений (F7, PZ) для последнего слова в синтаксически
правильной ситуации (жирная линия) и синтаксически неправильной ситуации (пун-
ктирная линия). Ранняя левая передняя негативность (РЛПН) и поздняя позитивность
(Р600), участвующие у взрослых в ответах на ошибки в структуре фразы, также зареги-
стрированы у двухлетних детей. (б) Топографические карты различий в паттернах ак-
тивации мозга при вычитании активности мозговых ответов на правильные условия из
ответов на неправильные условия. Показано топографическое распределение эффектов
для двухлетних детей (слева) и взрослых (справа). (Адаптировано из Oberecker et al.,
2005. Источник: Friederici, 2005.)
Рис. 15.29 . Связанные с возрастом изменения белого вещества в двух областях мозга.
Увеличение плотности белого вещества показано во внутренней капсуле (левый рису-
нок) и дугообразном пучке в левом полушарии (правый рисунок) для группы из 111
детей в возрасте от 4 до 17 лет. (С разрешения из Paus et al., 1999. Источник: Paus et al.,
2005.)
Развитие
239
выполнить еще много исследований для того, чтобы полностью понять всю
сложность приобретения человеческой речи, существующие работы обеспечи-
вают ученых новыми важными данными о языковых знаниях у очень малень-
ких детей.
Корковые области коры продолжают развиваться в период детства и под-
росткового возраста, как уже было сказано в этой главе. Как развиваются и со-
зревают в детстве области мозга, ответственные за восприятие и производство
речи? К классическим речевым зонам относят область лобной доли (функци-
онально выделена как зона Брока) и область височной доли (функционально
определена как зона Вернике), которые обеспечивают процессы производства и
восприятия речи. В течение всего периода детства между этими областями об-
разуются связи вследствие опыта по восприятию и производству устной речи.
В своих исследованиях Паус с коллегами (Paus et al., 1999) использовали фМРТ
для изучения образования связи между зонами Брока и Вернике. Они прово-
дили измерения плотности белого вещества в большой группе (N = 111) детей
в возрасте от 4 до 11 лет. Их результаты показали зависимые от возраста изме-
нения во внутренней капсуле (рис. 15 .29, слева) и в левом дугообразном пуч-
ке, проводящем пути, соединяющие зоны Брока и Вернике в левом полушарии
(рис. 15 .29, справа).
Обратите внимание, что измерения корковой плотности используют для вы-
явлений паттерна развития коры. Существуют ли определенные изменения в
анатомическом строении областей мозга, которые обеспечивают речевую функ-
цию? Этот вопрос изучали в длительном обследовании группы из 45 нормально
развивающихся детей в возрасте от 5 до 11 лет (To�a et al., 2005). Увеличение
толщины коры примерно на 0,10–0,15 мм/год обнаружено в областях соответ-
ствующих зонам Брока и Вернике (рис. 15 .30).
Рис. 15.30 . Общегодовая степень изменений в толщине коры. Средняя доля в измене-
ниях корковой плотности показана в миллиметрах согласно цветовой схеме на рисунке
справа (максимальная потеря серого вещества показана в оттенках красного цвета и
максимальное увеличение серого вещества в оттенках синего). 45 детей в возрасте от 5
до 11 лет были дважды обследованы (с двухлетним перерывом). (Источник: To�a et al.,
2005, с разрешения из Sowell et al., 2004.)
240
Глава 15
Отражают ли результаты утолщения корковых зон, обеспечивающих речь,
происходящие динамические изменения речи у ребенка? Вероятно, что да, но
необходимы дополнительные исследования связи анатомических изменений
с речевыми навыками для того, чтобы мы полностью понимали взаимодействие
мозга и разума в процессах развития речи.
В этом разделе мы осветили данные о том, что очень маленькие дети уже
имеют сложные знания о семантике и синтаксисе родного языка. Мы также
продемонстрировали, что речевые области мозга продолжают развиваться в пе-
риод детства и подросткового возраста. Однако существует множество вопро-
сов о приобретении языка, которые остаются без ответа. Центральный вопрос,
который остается неразрешенным, касается взаимовлияния врожденного (гене-
тической предрасположенности к речи) и приобретенного (роли опыта). Другие
аспекты приобретения языка, которые в настоящий момент активно исследуют-
ся, заключаются в развитии речевой системы у двуязычных и мультиязычных
детей.
4.2.2. Произвольный мозг: получение когнитивного контроля
Даже маленькие младенцы должны освоить, какая информация в окружающем
их мире является важной, а какая не важна или не существенна. Эти механиз-
мы обучения входят в общею категорию «когнитивного контроля» и находят-
ся в фокусе значительного числа исследований развития младенцев и детей.
Вспомните, что задача «А не Б» применялась для изучения способности мла-
денцев игнорировать или подавлять иррелевантную информацию и тормозить
доминантную реакцию (Pia�et, 1937, 1954; Diamond, 1985). Эти способности
становятся все более важными в течение детства по мере того, как окружаю-
щая среда становится сложнее. Представьте шестилетнего ребенка на уроке в
первом классе: он должен внимательно слушать учителя или сосредоточиться
на задании, несмотря на большое число отвлекающих событий вокруг него, на-
пример, разговоры между детьми, падающие учебники, скрип стульев. Направ-
ление ресурсов внимания к значимым аспектам окружения и прочь от менее
важных является жизненно необходимым элементом в развитии.
У взрослых считается, что ДЛ-ПФК вовлечена в функции когнитивного кон-
троля. Мы знаем из гистологических и нейроанатомических работ на развиваю-
щихся детях, что ПФК имеет длительный период развития и достигает зрелой,
взрослой стадии только к середине или концу подросткового возраста. Резуль-
таты поведенческих исследований функций произвольного контроля у детей и
подростков показали схожую динамику развития способностей к когнитивному
контролю. Открытым вопросом в области когнитивной нейронауки развития
является вопрос о существовании взаимосвязи между поздно созревающими
областями мозга и поздно развивающимися навыками произвольного контроля.
Работы по изучению нейронных основ произвольного контроля только не-
давно были выполнены на детях. Кейси с коллегами (Casey et al., 2005) про-
вели основополагающие исследования произвольного поведения, используя
фМРТ и поведенческие методы для того, чтобы измерить паттерны нервной
активации в мозге во время выполнения детьми заданий, вовлекающих обла-
сти ПФК. В одном эксперименте они регистрировали связанные с событием
фМРТ-изображения, в то время как дети и взрослые были вовлечены в пара-
Развитие
241
дигму «�o-no �o» (Durston et al., 2002). При выполнении данной парадигмы ис-
пытуемые должны подавлять ответную реакцию при предъявлении определен-
ного зрительного стимула в последовательности других стимулов (например,
один герой Покемон в последовательности других героев Покемон). Сложность
задания увеличивалась с увеличением числа «�o» стимулов, предшествующих
предъявлению «no �o» стимула. Успешное подавление реакции ассоциировали
с более сильной активацией префронтальных областей у детей по сравнению с
взрослыми. В то время как у взрослых активация некоторых префронтальных
областей увеличивалась с увеличением числа предшествовавших «�o» проб
(соответствует повышению необходимости затормозить), у детей эти области
продемонстрировали максимальную активацию во время всех проб. Наряду с
низким уровнем поведенческого выполнения у детей в этой и других задачах
на подавление реакции эти данные свидетельствуют, что функциональное раз-
витие некоторых областей ПФК необходимо для сформировавшихся навыков
подавлять существующие намерения.
Основным результатом работы Кейси с коллегами является обнаружение,
что у маленьких детей наблюдается более широкая, распространенная актива-
ция мозга при выполнении заданий на когнитивный контроль по сравнению с
взрослыми. Во время развития эти области созревают и мозговая активность,
коррелирующая с успешностью выполнения заданий (например время реакции
и точность) становится более локальной и точно настроенной. На рис. 15.31
представлена иллюстрация из новой статьи (Casey et al., 2005), являющейся
обзором литературных данных о развитии когнитивного контроля. Основное
положение о том, что мозговая активация становится более локальной и огра-
ниченной с увеличением возраста ребенка, показано с упоминанием работ, про-
демонстрировавших увеличение и уменьшение активации с возрастом.
Другой подход для выявления роли фронтальных отделов в заданиях, вовле-
кающих функции произвольного контроля, был продемонстрирован в совре-
менном фМРТ-исследовании на детях в возрасте 8–12 лет (Konrad et al., 2005).
Авторы использовали популярную модель для исследования процессов внима-
ния у взрослых (Posner and Petersen, 1990) и соединили ее с задачей на сети
внимания (ЗСВ), для которой хорошо известно, как она активирует процессы
внимания (Fan et al., 2002) (рис. 15 .32).
Модель внимания, предложенная Познером с коллегами, описывает различ-
ные (отдельные, но сильно перекрывающиеся) нейронные сети для:
1) настораживания при появлении новой и значимой информации,
2) ориентации и переориентации к значимой информации или стимулу,
3) произвольного контроля над процессами внимания (Posner and Petersen,
1990).
Например, представьте шестилетнего ребенка на уроке в первом классе. Для
того чтобы справляться со сложной окружающей средой, содержащей много
сенсорной информации, конкурирующей за его внимание, он должен быть со-
бран и находиться в состоянии готовности. Далее ученик должен быть спосо-
бен легко приспосабливать фокус своего внимания, например, для восприятия
новых сенсорных входов или переориентировать его на значимую информа-
цию, когда необходимо. И наконец, у него должен быть способ контролировать
эти ресурсы внимания.
242
Глава 15
Эти процессы находились в центре внимания исследований на взрослых
людях. У взрослых области мозга, активирующиеся при настораживании, рас-
положены во фронтальных и теменных областях правого полушария (Witte and
Marrocco, 1997). Области мозга, участвующие в ориентировании, расположены
Рис. 15.31. Развитие функций коры у человека, измеренное при помощи современных
методов визуализации, отражает точную настройку распространенных сетей нейроана-
томических областей мозга. В совокупности, работы по нейровизуализации развития
процессов когнитивного контроля указывают на существование общего паттерна уве-
личения степени вовлечения в эти процессы медленно созревающей префронтальной
коры (ссылки указаны красным цветом), в особенности латеральной префронтальной
коры и вентральной префронтальной коры, и уменьшения степени участия сенсорных
областей низшего уровня (синие ссылки), включая области экстрастриарной и верете-
новидной коры и задних областей теменной коры. Важно отметить, что специфичная
активация изменяется в зависимости от требований задания так, что задачи на рабо-
чую память и тест Струпа активируют другие области, нежели задания на подавление
реакции. Такой паттерн активации, который был зарегистрирован при выполнении
большого числа различных парадигм, предполагает, что высшие когнитивные функции,
обеспечиваемые ассоциативной корой, становятся более локальными или точно настро-
енными с развитием, в то время как активность других областей, не связанных с опреде-
ленными когнитивными функциями, затухает. П — передний, З — задний. (Источник:
Casey et al., 2005.)
Развитие
243
в правом полушарии на границе теменной и височных областей и в нижней лоб-
ной извилине (Corbetta et al., 2000). Области, ответственные за произвольный
контроль, включают переднюю поясную и латеральную префронтальную кору
(Marrocco and Davidson, 1998). Конрад с коллегами (Konrad et al., 2005) исполь-
зовали данные зоны мозга, которые продемонстрировали активность у взрос-
лых во время выполнения ЗСВ, как области интереса для фМРТ-исследования
16 мальчиков в возрасте 8–12 лет и 16 взрослых мужчин.
Результаты исследования процессов настораживания представлены на
рис. 15 .33 . Дети не продемонстрировали активационные паттерны, как у взрос-
лых, в заранее выделенных областях. Ключевые активационные различия меж-
ду детьми и взрослыми наблюдались в правой передней поясной извилине (рис.
15.33 (а)) и в среднем мозге (рис. 15 .33 (б)). Авторы предполагают, что данные
различия в паттернах активации происходят вследствие модуляции процессов
выполнения задания взрослыми при помощи нисходящих механизмов внима-
ния, которые развиты не полностью у детей в возрасте 8–12 лет.
Результаты исследования процессов переориентации показали типичный
паттерн правополушарной активации у взрослых, в то время как у детей зареги-
стрировано более диффузное и широкое распространение активации во время
переориентации. В итоге, большее увеличение активации происходит в правом
полушарии на границе височной и теменной коры у взрослых по сравнению
Рис. 15.32 . Экспериментальная парадигма: модифицированная версия задачи на сети
внимания (Fan et al., 2002). Данный рисунок иллюстрирует динамику предъявления че-
тырех разных подсказок и двух целевых стимулов. (Источник: Konrad et al., 2005.)
244
Глава 15
с детьми (рис. 15 .34(а)); более сильная активация наблюдается в скорлупе и
островке у детей по сравнению с взрослыми (рис. 15 .34(б)); и увеличение ак-
тивации зарегистрировано в верхней лобной извилине у детей по сравнению с
взрослыми (рис. 15.34(в)). Эти данные могут указывать на вовлечение этих об-
ластей мозга в процессы выполнения задания вследствие незрелых механизмов
когнитивного контроля у маленьких детей.
Результаты исследования процессов произвольного внимания показали
увеличение активации в верхней теменной коре и нижней лобной извилине
у взрослых по сравнению с детьми (рис. 15 .35(а, б)) и увеличение активации
верхней височной извилины и верхней лобной извилины у детей по сравнению
с взрослыми (рис. 15.35(в, г)).
В целом результаты работы Конрада с коллегами показали поразительные
различия паттерна мозговой активации у детей и взрослых при выполнении за-
даний на внимание. И хотя к данным результатам необходимо относится с осто-
рожностью, так как все еще требуется проведение большого числа исследова-
ний с похожими парадигмами и измерениями, результаты Конрада с коллегами
свидетельствуют о незрелости лобно-теменных сетей при выполнении заданий
на когнитивный контроль, что соотносится с другими данными (рассмотрено
в Casey et al., 2005).
Рис. 15.33 . Результаты на-
стораживания. Разная кар-
тина активации отмечена
у взрослых и детей при
проведении сравнения при
помощи двухвыборочного
критерия (ограниченная
psvc
< 0,05, порог больше
5 объемных
элементов,
показано Т1-изображение,
усредненное по группе).
(а) Увеличенная активация
в правой поясной извилине
у взрослых по сравнению
с детьми. (б) Увеличенная
активность ствола мозга
у взрослых по сравнению
с детьми. На графиках
представлено
изменение
BOLD-сигнала в процен-
тах, отдельно для каждой
группы, в зависимости от
типа пробы (объединенные
для совпадающих и не-
совпадающих стимулов),
от относительного уровня
максимальной активации.
(Источник: Konrad et al.,
2005.)
Развитие
245
Рис. 15.34 . Разная картина активации отмечена у взрослых и детей при проведении
сравнения условий переориентации при помощи двухвыборочного критерия (ограни-
ченный psvc
< 0,05 или p < 0,1, исправленный для множественных сравнений анализа
целого мозга, порог больше 5 объемных элементов, показано Т1-изображение, усред-
ненное по группе). (а) Бóльшая активация области на границе височной и теменной
коры в правом полушарии у взрослых по сравнению с детьми. (б) Более сильная актива-
ция скорлупы и островка у детей по сравнению с взрослыми. (в) Увеличение активации
верхней лобной извилины у детей по сравнению с взрослыми. На графиках представле-
но изменение BOLD-сигнала в процентах, отдельно для каждой группы, в зависимости
от типа пробы (объединенные для совпадающих и несовпадающих стимулов), от отно-
сительного уровня максимальной активации. (Источник: Konrad et al., 2005.)
246
Глава 15
Общий вывод из этих фМРТ-исследований когнитивного контроля у детей
обеспечивает доказательство того, что системы внимания у них функциониру-
ют, но еще незрелы по сравнению с взрослыми. Дети демонстрируют паттер-
ны активации, которые более диффузны и затрагивают более широкие области
мозга, чем паттерны активации у взрослых. Эти данные могут отражать незре-
лую природу систем в детстве, которые в процессе развития, с опытом, стано-
вятся более локальными и специализированными.
Дальнейшие работы по исследованию когнитивного контроля и развития
ПФК у детей должны будут учесть многие другие аспекты длительного пути
развития лобной коры и соответствие с поведением. Некоторые исследования
могут сосредоточиться на изучении половых различий в функциях произволь-
ного контроля и связанных с ними паттернах активации мозга. Как вы могли
заметить, большинство работ, приведенных в данном разделе, основаны на
Рис. 15.35. Разная картина активации отмечена у взрослых и детей при проведении
сравнения условий произвольного внимания при помощи двухвыборочного критерия
(ограниченный psvc
< 0,05 или p < 0,1, исправленный для множественных сравнений
анализа целого мозга, порог больше 5 объемных элементов, показано Т1-изображение,
усредненное по группе). (а) Бóльшая активация верхней теменной коры и (б) нижней
лобной извилины у взрослых по сравнению с детьми. (в) Увеличение активации верхней
височной извилины и (г) верхней лобной извилины у детей по сравнению с взрослыми.
На графиках представлено изменение BOLD-сигнала в процентах, отдельно для каждой
группы, в зависимости от типа пробы (объединенные для совпадающих и несовпадаю-
щих стимулов), от относительного уровня максимальной активации. (Источник: Konrad
et al., 2005.)
Развитие
247
зрительном предъявлении стимулов. Требуется провести значительное число
работ по выявлению сетей лобных долей, обеспечивающих контроль слухового
внимания, а также сетей, которые обеспечивают выполнение заданий, для ко-
торых необходима интеграция мультимодальной информации (например, зри-
тельной и слуховой).
4.2.3. Социальный мозг: восприятие лиц в детстве
Вопросы восприятия лиц у человека находились в центре внимания многих по-
веденческих работ и работ с нейровизуализацией на взрослых людях, младен-
цах, детях и подростках. Почему данная область исследований так важна для
когнитивной нейронауки? Выявление областей мозга, которые специализиру-
ются на восприятии лиц представителей своего вида, может пролить свет на
спор о роли врожденного и приобретенного в развитии. Имеем ли мы врожден-
ную предрасположенность к выделению, фокусировке на лицах и толкованию
выражений лиц? Или наш обширный опыт с восприятием лиц обеспечивает
способность к обработке информации, которая не специфична для лиц, но ис-
пользует сети восприятия зрительных объектов? Исследование процессов вос-
приятия лиц в ходе развития может помочь ученым выявить, как генетическая
предрасположенность взаимодействует с опытом.
Вспомните, что в исследованиях восприятия лиц на младенцах использо-
вали регистрацию ССП и, в частности, проводили определение степени изме-
нения компонента N170 в ответ на предъявление лиц по сравнению с предъ-
N170 в ответ на предъявление лиц по сравнению с предъ-
170 в ответ на предъявление лиц по сравнению с предъ-
явлением других зрительных объектов. Результаты этих работ, а также работ,
выполненных с помощью методов нейровизуализации (МЭГ, фМРТ), доказы-
вают, что процесс восприятия лиц менее специализирован у маленьких детей
(5–10 лет), с более диффузной и менее четко выделенной активацией мозга на
лица, по сравнению со старшими детьми (11–14 лет) (Gathers et al., 2004; Ayl-
Ayl-
ward et al., 2005; Kyllainen et al., 2006). У старших детей наблюдается также
билатеральная активация веретеновидной области лиц на предъявление лиц по
сравнению с предъявлением изображений домов, чем у младших детей, и этот
паттерн активации коррелирует с возрастом (Aylward et al., 2005). Тем не менее
эти исследования указывают, что в то время как старшие дети демонстрируют
более локальную активацию, чем младшие, паттерн активности у них все же не
совпадает с таковым у взрослых, что свидетельствует о медленно протекающем
процессе созревания.
Во многих работах ученые используют в качестве стимулов изображения
лиц с нейтральными или радостными выражениями; однако они оставляют от-
крытым вопрос о том, как дети воспринимают эмоциональную информацию в
лицах. Понимание эмоциональных подсказок в выражениях лиц является жиз-
ненно необходимым аспектом социального познания у человека. Исследование
обработки эмоциональных признаков лица является очень сложным процессом,
и, к тому же этот вопрос недостаточно изучен у взрослых, не говоря о детях.
Далее мы осветим две современные работы, проведенные на детях, по исследо-
ванию эмоциональных признаков лиц: одна использует фМРТ для выделения
областей мозга, вовлеченных в расшифровку сигналов эмоционального лица,
а во второй работе регистрируют ЭЭГ с выделением компонента N170 для из-
N170 для из-
170 для из-
учения динамики обработки эмоциональных сигналов. Необходимо отметить,
что исследования развития процессов восприятия эмоциональных признаков
248
Глава 15
лиц находятся только в самой начальной стадии: несмотря на то что эти работы
предоставляют полезную информацию для понимания развития и созревания
этих процессов, требуется значительное число исследований, чтобы полностью
понять все хитрости восприятия лиц у человека.
В одной из работ (Batty, Taylor, 2006) изучалось появление компонента N170
в ответ на предъявление изображений лиц с шестью разными эмоциональными
выражениями (радость, удивление, грусть, страх, злость и отвращение), а так-
же лиц с нейтральным выражением в большой (82) группе детей в возрасте
от 4 до 15 лет. Авторы сообщили, что только у старших детей (14–15 лет) пат-
терн ответа компонента N170 совпадал с таковым у взрослых. Но даже в этой
группе детей ответы компонента N170 не демонстрировали картину взрослых
при дискриминации эмоциональных стимулов. Авторы предположили, что ин-
терпретация сложной эмоциональной информации человеческих лиц является
медленно созревающим процессом, который еще не завершается к середине
подросткового периода.
Похожее исследование было выполнено при помощи фМРТ и предъявления
таких же восьми эмоциональных категорий (Lobau�h et al., 2006). В данной
работе принимали участие дети в возрасте 10–12 лет. Ставилась задача по опре-
Ставилась задача по опре-
задача по опре-
делению половой принадлежности: детей просили определить, является ли
предъявляемое лицо мужским или женским. Таким способом авторы пытались
понять нервную основу имплицитного узнавания эмоциональных сигналов во
время восприятия лиц. Сначала авторы выявили области мозга, которые по-
авторы выявили области мозга, которые по-
выявили области мозга, которые по-
казали разную картину активации в ответ на предъявление лиц по сравнению
с простым смотрением. Результаты показаны на рис. 15 .36 . Этими областями
активации являлись веретеновидная/парагиппокампальная извилина правого
полушария, которая продемонстрировала более сильную активацию при предъ-
явлении лиц, и верхняя височная и задняя височная извилины обоих полуша-
рий, в которых отмечено снижение активности при демонстрации лиц.
Второе важное открытие данной работы состоит в том, что активация мозга
в ответ на негативные эмоции (отвращение, страх) была существенно сильнее,
чем при предъявлении лиц с положительными (радость) или нейтральными вы-
ражениями (рис. 15.37). Наиболее сильный ответ наблюдался при предъявле-
нии лиц с выражениями страха. По мнению авторов, эти результаты свидетель-
ствуют о становлении функциональных эмоциональных сетей уже к возрасту
10 лет.
Что же говорят нам результаты этих работ по поводу того, как дети воспри-
нимают сложные эмоциональные сигналы лиц? Очень важно помнить, что
измерения, проводимые в этих двух работах (регистрация ССП с определе-
нием латентного периода и амплитуда N170 и фМРТ исследование гемодина-
N170 и фМРТ исследование гемодина-
170 и фМРТ исследование гемодина-
мических изменений в мозге), очень сильно различаются. К тому же, в одном
исследовании (Batty and Taylor, 2006) участвовала большая (82) группа детей
с сильным разбросом по возрасту (4–15 лет), в то время как в другой работе
принимала участие небольшая (10 человек) группа детей близкого возраста
(10–12 лет). Различающиеся задания и методы, применявшиеся в данных ис-
следованиях, делают интерпретацию результатов очень проблематичной, не-
смотря на то что в этих работах изучали очень похожие процессы, эмоцио-
нальные сигналы в лицах.
Развитие
249
Результаты, представленные здесь, свидетельствуют о разных паттернах
мозгового ответа у детей при восприятии эмоциональных сигналов на лицах.
Требуется провести еще очень большое число работ, прежде чем мы сможем
полностью понять участие мозговых областей и путь развития обработки эмо-
циональных признаков при восприятии лиц. Некоторыми аспектами будущих
исследований могут стать изучение восприятия эмоциональных стимулов
у мальчиков и девочек и определение, обращают ли дети больше внимания на
некоторые части лица (например, глаза) по сравнению с взрослыми.
Что мы узнали о развитии речи, произвольных функций и социального со-
знания в детстве и подростковом периоде? Центральное открытие из множества
информационных источников (гистология, патология, нейровизуализация) —
корковые области, обеспечивающие процессы высших психических функций.
Они имеют длительный путь развития, простирающийся до середины или кон-
ца подросткового возраста. Мы обладаем значительным объемом поведенче-
ских данных, указывающих на схожие паттерны активности в задачах, затра-
гивающих более сложные аспекты этих когнитивных функций, с достижением
эффективности при выполнении задач уровня, свойственного взрослым, толь-
ко к завершению подросткового периода. Объединение этой информации для
соотнесения развития мозга с когнитивными функциями до сих пор остается
сложной проблемой для ученых.
Рис. 15.36 . Результаты ответов на предъявления лиц и при простом смотрении. (а) Об-
ласти мозга, окрашенные в синий цвет, проявили увеличение активации при предъявле-
нии лиц по сравнению с простым смотрением. Обратную картину наблюдали в областях
желтого/красного цвета. Стрелками (б–д) указаны пики значений в областях. Среднее
процентное изменение (± SEM) указано для выбранных областей. (б) Большая область
веретеновидной/парагиппокампальной извилины правого полушария (20/40/16) была
более активна при предъявлении лиц. (в, г) Верхняя височная извилина обоих полу-
шарий (в: 48/32/+ 4, г: + 44/32/+ 8). (д) Задняя височная извилина (44/52/4) продемон-
стрировала уменьшение активации при предъявлении лиц. С — смотрение, Л — лицо.
(Источник: Lobau�h et al., 2006.)
250
Глава 15
В то время как мы сделали существенный шаг вперед, множество сложно-
стей в каждой области сознания остается невыясненным. Несмотря на то что
мы понимаем процессы возникновения речи у маленьких детей, выявление
корковых областей для сложной системы речи у взрослых еще продолжается,
и совсем мало известно о свойствах возникающего языка. Аналогично, про-
должаются работы по исследованию того, как ПФК обеспечивает выполнение
Развитие
251
заданий, требующих вовлечения систем внимания, планирования и рабочей
памяти. Определение взаимосвязи между этими структурами и навыками или
требованиями для выполнения определенных задач остается еще нерешенным
вопросом у взрослых и продолжает исследоваться у детей. Восприятие лиц
и интеграция эмоциональных, лингвистических и прагматических признаков
лиц в социальное знание также продолжает исследоваться у взрослых, причем
очень мало известно о возникающих паттернах и взаимосвязях между этими
факторами для понимания социального поведения человека.
Важным направлением в исследованиях когнитивной нейронауки развития
является объединение методик — например, фМРТ и ЭЭГ — с измерениями
поведенческих ответов, что позволит получить полные результаты относи-
тельно аспектов когнитивных функций высшего порядка. Объединение фМРТ
с ЭЭГ, например, может обеспечить большое пространственное разрешение
относительно выявления областей активации мозга совместно с большой вре-
менной разрешающей способностью для выявления динамики. Другое важное
направление — проведение долговременных исследований, которые помогут
проследить развитие отдельных детей. В данном случае, сначала получают «ис-
ходные» показатели, а затем проводят их повторное измерение через опреде-
ленные промежутки времени. И наконец, новые экспериментальные парадигмы
показывают, что малыши знают и понимают намного больше об окружающем
их мире, чем считалось ранее. Новые подходы к измерению сознания младен-
цев и выявлению значимой активности мозга обеспечат важными представле-
ниями об изменениях, происходящих в первые годы жизни.
Рис. 15.37. Результаты предъявления эмоциональных лиц. (а) Различия заданий по пер-
вой латентной переменной. Первый паттерн наиболее четко отделил отвращение (синий,
отрицательный показатель) от страха (красный, положительный показатель) и менее
четко позволяет различить удивление (голубой) и радость (серый). (б) Области мозга,
активность которых отличалась в ответ на страх и отвращение. Желтый/красный цвет:
области, где ответ на страх был выше, чем на отвращение; синий цвет: области, в кото-
рых ответ на отвращение был больше, чем на страх. (в–е) Локализация и доверительный
интервал (ДИ = 7) в ответах объемных элементов в блоках, указанных для выбранных
областей (среднее 8 проб после начала предъявления). (в, г) Миндалина обоих полуша-
рий (в: + 20/4/8; г: 24/4/8). (д) Правая верхняя височная извилина (+ 48/40/+ 8). (е) Левая
верхняя височная извилина/нижняя огибающая извилина (48/40/+ 20). (ж) Различия
заданий по второй латентной переменной. Этот паттерн наиболее четко разделил от-
вращение и страх (синий и красный, положительные значения) от грусти (зеленый, от-
рицательные значения). Удивление (голубой) и радость (серый) также отличались от
отвращения и страха. (з) Области мозга, активность которых наиболее сильно разли-
чалась в случаях предъявления лиц с выражениями отвращения и страх по сравнению
с выражениями грусти. Желтый/красный цвет: области мозга, где ответ на отвращение
и страх были больше, чем на грусть; синий цвет: области, в которых ответ на грустные
лица был сильнее, чем на лица с выражениями отвращения и страха. (и–л) Среднее
процентное число изменений в ответах объемных элементов (ДИ = 7) для выбранных
областей. (и) Левая верхняя височная извилина (44/28/+ 8). (к, л) Передняя поясная кора
(к: + 12/+ 36/+ 24, ПБ 32; л: + 8/+ 8/+ 28, ПБ 24). Контрольная цветная полоса отражает
параметры стабильности объемных элементов. Левая сторона мозга расположена на ри-
сунке справа. (Источник: Lobau�h et al., 2006.)
252
Глава 15
5.0 .
Ранние повреж дения мозга
и пластичность развития
Мы уже говорили о важности долгосрочных исследований для выявления ин-
дивидуального развития и его конечного результата. Этот тип исследования
особенно важен, когда оценивают долговременное влияние ранних (перина-
тальных) травм мозга. Мы уже знаем из предыдущих глав, что у взрослых по-
вреждения мозга вследствие инсульта, заболевания или травмы обычно при-
водят к нарушениям когнитивной деятельности, которые являются достаточно
серьезными, и полного восстановления функций, как правило, не наблюдается.
Что происходит, если повреждения происходят у новорожденных? Этот вопрос
важен для решения проблемы о роли врожденного и приобретенного. Рассмо-
трите гипотезу о том, что некоторые системы мозга, например языка, имеют
сильное генетическое предрасположение к развитию в определенной области
мозга. Если в этой предопределенной области произойдет инсульт на раннем
этапе развития, сформируется ли нормальным образом речь у ребенка? Или
речь разовьется с нарушениями вследствие ранних и не восстанавливающихся
повреждений этих областей? С альтернативной точки зрения, если опыт играет
доминантную роль в развитии определенной области мозга, которая настраива-
ется для выполнения речевой функции, сможет ли ребенок сформировать нор-
мальную речь, несмотря на ранние мозговые повреждения?
Эффекты перинатальных мозговых повреждений интенсивно изучаются
в работах в области нейробиологии на животных. Эффекты ранних поврежде-
ний мозга и их влияние на последующее когнитивное развитие менее изучены
на людях. Одной из причин этого является то, что одиночные, унилатеральные
(только в одном полушарии) пре- или перинатальные мозговые инсульты отно-
сительно редко встречаются. Обычно последствия ранних мозговых инсультов
более широкие и объединяются с другими неврологическими проявлениями
(рис. 15 .38, слева). В таких случаях обширного повреждения в купе с другими
травматическими событиями очень трудно сравнивать когнитивное развитие с
развитием ребенка без ранних травм и повреждений. В случае если перинаталь-
ное повреждение ограничено определенной областью (рис. 15.38, справа и в
центре), долговременные эффекты обычно менее выражены. В данном разделе
мы остановимся на следующих вариантах повреждений: ранних, локальных,
унилатеральных травмах.
Несмотря на то что мы только начинаем понимать всю сложность послед-
ствий ранних травм на последующее развитие у человека, серия длительных
исследований (Stiles et al., 2005) пролила свет на долгосрочные эффекты пери-
2005) пролила свет на долгосрочные эффекты пери-
натальных травм, и мы рассмотрим их некоторые результаты. Долговременный
проект Сан Диего (Stiles et al., 2002) является самым большим исследованием
в США долгосрочных эффектов перинатальных повреждений мозга. Стайлс с
коллегами проследили, начиная с 1989 г., когнитивное развитие нескольких со-
тен детей с перинатальными повреждениями мозга.
Основное внимание в этих исследованиях было уделено речевому развитию
у детей с перинатальными мозговыми травмами. Одно ключевое открытие со-
стояло в том, что в то время как у взрослых локальное мозговое повреждение
(обычно вследствие разрушений речевых центров приводит к длительным на-
рушениям, у младенцев, перенесших перинатальную травму мозга, наблюдает-
Развитие
253
ся иная картина нарушений. Несмотря на то что у таких детей происходит от-
ставание в развитии речи (например, способность понимать слова развивается
только к 9–12 месяцам, а способность произносить слова к 12–15 месяцам), они
в основном догоняют в речевом развитии своих нормальных сверстников к воз-
расту 5 лет. Очень важный момент, однако, состоит в том, что при тщательном
тестировании выявляются некоторые нарушения даже в пятилетнем возрасте,
особенно при сложных структурах предложений (Reilly et al., 2004). Эти дети с
ранними мозговыми травмами, в конце концов, достигают языковой компетен-
ции, но данные, полученные в длительной работе Стайлса с коллегами (Stiles et
al., 2005), указывают, что их языковые способности находятся на уровне ниже
среднего. Таким образом, несмотря на то что дети осваивают множество навы-
ков и умений в отношении речи, в течение детства, подросткового и, возможно,
взрослого возраста остаются некоторые ключевые нарушения вследствие очень
ранних повреждений важных для понимания и обработки речи областей мозга.
Другое направление изучения когнитивной деятельности в долговременном
проекте Сан Диего касалось пространственного восприятия. Пространственное
распознавание и эффекты, наблюдаемые у взрослых, от полученных повреж-
дений были целью многих нейропсихологических исследований. Центральное
открытие заключается в том, что существует полушарная асимметрия в распоз-
навании зрительных стимулов, с левым полушарием, ответственным за выде-
ление информации о признаках (частной), и правым полушарием за выделение
информации (общей) о конфигурации стимулов (рис. 15 .39).
Функциональные МРТ-исследования нормально развивающихся подрост-
ков демонстрирует подобную асимметрию полушарий, с преобладанием ак-
тивации в затылочно-теменных областях правого полушария при обработке
общих признаков и затылочно-теменных областях левого полушария при вос-
приятии частных свойств (рис. 15 .40). В отличие от этого пятнадцатилетний
Рис. 15.38 . Крупномасштабные и незначительные перинатальные повреждения мозга.
Структурные МРТ-изображения мозга в осевом сечении, полученные у трех детей с пе-
ринатальным повреждением мозга. Слева: большое одностороннее разрушение, затра-
гивающее большую часть одного полушария мозга. В центре: небольшие разрушения в
одной доле мозга. Справа: глубокие разрушения, затрагивающие подкорковые области.
(Источник: Stiles et al., 2005.)
254
Глава 15
подросток, которые пережил перинатальное повреждение правого полушария
мозга, демонстрирует повышенную активацию левого (не поврежденного) по-
лушария при обработке информации как о частных, так и общих свойствах,
а у тринадцатилетнего подростка с перинатальным повреждением левого полу-
шария более сильная активация наблюдается при восприятии общих и частных
свойств в правом (не поврежденном) полушарии мозга (рис. 15.40). Таким об-
разом, фМРТ-данные обследования двух подростков с перинатальной травмой
указывают на долговременные эффекты, влияющие на механизмы простран-
ственного восприятия. Однако они также свидетельствуют об удивительной
пластичности мозга, что проявляется в вовлечение в процессы пространствен-
ного восприятия областей неповрежденного полушария.
Что же мы узнали из длительных исследований Стайлса с коллегами долго-
временных эффектов ранних повреждений мозга? И какую информацию они
предоставляют о сложной и высоко перекрывающейся роли врожденного и
приобретенного в развитии человека? Несмотря на то что работы в этом на-
правлении находятся на ранних этапах, результаты на сегодняшний день свиде-
тельствуют о том, что ранние повреждения мозга приводят к длительным, хотя
и практически неуловимым, нарушениям. Второе важное открытие заключает-
ся в том, что, несмотря на ранние травмы и отставание в развитии вследствие
этих поражений мозга, дети развиваются и приобретают высшие когнитив-
ные функции, хотя иногда и на более низком уровне. В целом, эти результаты
Рис. 15.39. Общее и частное: примеры зрительно-пространственных нарушений. При-
меры воспроизведения по памяти иерархических стимулов у взрослых пациентов с
инсультом в правом или левом полушариях. (С разрешения из Delis et al., 1986.) Слева:
пример стимулов, которые необходимо воспроизвести. В центре: пациент с правосто-
ронним повреждением мозга обычно воспроизводит частные признаки стимула (дета-
ли), но упускает общие (совокупные) аспекты стимула, т. е ., в данном примере, «М»
или треугольник. Справа: пациенты с левополушарными повреждениями обычно вос-
производят общий вид стимула, но упускают частные детали. (Источник: Stiles et al.,
2005.)
Развитие
255
доказывают, что повреждение некоторых областей приводит к длительным на-
рушениям, даже в случае возникновения травмы при рождении или в раннем
детстве. Это в определенной степени доказывает, что некоторые системы име-
ют генетическую предрасположенность в функционировании и претерпевают
длительные нарушения при повреждениях. Более радужный вывод, который
Рис. 15.40 . Общие и частные процессы в мозге. Данные активации, полученные при
помощи фМРТ, двух подростков с пренатальной локальной мозговой травмой при вы-
полнении задания на восприятие иерархических изображений и сравнение с нормаль-
ными подростками. Каждый ребенок участвовал в отдельных циклах нейровизуализа-
ции, в которых просили обращать внимание либо на общие, либо на частные признаки
стимулов. В отличие от контрольной группы, которая продемонстрировала различный
паттерн латерализации общих и частных процессов, у этих двух детей с разрушениями
активация в основном наблюдалась в неповрежденном полушарии. На рисунке пред-
ставлены изображения активации мозга у двух детей с перинатальной мозговой трав-
мой. (Источник: с разрешения из Stiles et al., 2005,)
256
Глава 15
следует из этих исследований, говорит о значительной мозговой пластичности:
когнитивные функции, нарушенные при ранних повреждениях, могут развить-
ся по альтернативному пути.
6.0.
Заключение
В этой главе мы проследили стадии развития человека от эмбриона до младен-
ца и подростка. Несмотря на то что область когнитивной нейронауки развития
чрезвычайно молода, изложенные результаты отвечают на важные вопросы о
развитии человеческого мозга и соответствии этого процесса с когнитивной
деятельностью. Актуальная тема в большинстве споров в области развития че-
ловека касается роли врожденного и приобретенного. Из представленных здесь
данных вы можете видеть, что на каждую стадию человеческого развития ока-
зывают влияние генетические эффекты и биологические предпосылки. Анало-
гично, каждая стадия испытывает влияние окружающей среды, независимо от
уровня (клетка, система или целый мозг).
Появление новых неинвазивных методов изучения развития человека обе-
спечивает возможность исследовать новые вопросы о когнитивном развитии,
например: что знает малыш до рождения? Понимает ли младенец грамматику
языка? Как развивается у младенца и ребенка чувство о собственном «я»? Эти
и другие вопросы будут заданы в будущих исследовательских работах, направ-
ленных на открытие сложных паттернов развития человеческого мозга и его
связи с сознанием.
7.0 .
Задания и упражнения к главе 15
7.1.
Контрольные задания
1. Как изменили технологии нейровизуализации исследования развития у мла-
денцев и детей?
2. К чему в развитии человека относят термин «двусторонние влияния»? По-
чему они важны?
3. Приведите пример влияния врожденного и приобретенного на пренатальное
развитие.
4. Какие эффекты оказывают употребление матерью алкоголя, табака и мари-
хуаны на не рожденного еще ребенка?
5. Какие области мозга созревают и формируются в раннем детстве? Какие об-
ласти созревают позже?
6. Развитие представлений у ребенка о неисчезаемости объекта является фоку-
сом многих работ. Опишите эксперимент для изучения формирования вос-
приятия о постоянстве объекта и возможные его результаты для: (а) шести-
месячных детей; (б) двенадцатимесячных детей.
Человеческий мозг был способен к творчеству уже тогда, когда формирование
современного человека едва завершилось. Наскальная живопись пещеры Ляско,
возраст которой оценивается в 16 000 лет. (Источник: http://en/.wikipedia.org/wiki/
Lasceau.)
Содержание
1.0.Введение.......................................................260
1.1 . Основные малые молекулы клетки также используются
длянейрональнойсигнализации.................................261
1.2.Живыеорганизмыиспользуютферменты.........................263
1.3.Однаитажемолекуламожетнестиразныесигналы................264
1.4 . Гематоэнцефалический барьер защищает внутреннюю среду мозга . . . 264
1.5.Модельныеорганизмы.........................................266
2.0.Генывэволюции,онтогенезеиповседневнойжизни....................268
2.1 . Разные события с ДНК происходят в различные промежутки времени. 270
2.2.Развитиемозга:нейрогенез,затемсинаптогенез...................271
2.3.Какчеловекполучилтакойбольшоймозг?........................274
3.0.Экспрессияирегуляциягенов......................................275
3.1.Обработкаинформациивклетке................................275
3.2.ДНКвысшихуровнейрегулируетостальнуюДНК.................277
3.3.Ошибкиприразвитиимозга....................................279
3.4.Гены,контролирующиеразвитие................................279
3.5 . Программирование генов: за пределами центральной догмы . . . . . . . . . 281
3.6.Средаможетперестраиватьхроматинвядреклетки................ 282
3.7.Обучениекакэпигенетическийпроцесс..........................283
4.0.Нейроныиглиальныеклеткикакобработчикисигналов.................283
4.1.Нейроныисинапсывкачествепереключателей....................284
4.2.Химическаясаморегуляция.....................................285
4.3.Мембраны,ионныеканалыиаксоновыеспайки...................286
5.0.Синаптическая передача: от производства до расчистки . . . . . . . . . . . . . . . . 290
5.1.«Большаядвойка»:глутаматиГАМК............................290
5.2.ГАМК:основнойтормозныймедиатор...........................292
5.3.Нейроглиятакжеможетобрабатыватьинформацию................ 294
5.4 . Производство, высвобождение и удаление нейромедиаторов . . . . . . . . . 295
5.5.Высвобождениевеществавсинапсе.............................295
5.6.Синапсыирецепторывкачествеконтрольныхточек...............298
5.7.Рецепторыраспознаютсигнальныемолекулы.....................298
5.8.Трансмиттерзависимыеионныеканалы..........................302
5.9 . Очистка синапса: дезактивация, диффузия и утилизация медиатора . . . 306
6.0.Нейромодуляторы................................................308
7.0.Обучение.......................................................310
7.1.Гиппокампальныйкомплекс....................................312
7.2 . Глутамат, ГАМК и долговременные депрессия и потенциация. . . . . . . . 314
Глава 16
Гены и молекулы мышления
260
Глава 16
7.3 . Глутаматные синапсы в качестве модели синаптического обучения . . . 315
7.4.Эпигенетическиемеханизмыобучения...........................322
7.5.Нейротропныефакторыприобучении...........................323
8.0.Заключение.....................................................324
9.0.Заданияиупражнениякглаве16...................................327
1.0 . Введение
Что, по-вашему, может быть общего у шоколадной конфеты и укуса пчелы?
И то, и другое воспринимается через воздействие сигнальных молекул на клет-
ки мозга. Вкус шоколада и зуд от укуса возникают в результате ответа мозга на
соответствующее раздражение. В повседневной жизни мы встречаемся с огром-
ным разнообразием молекул, содержащихся в кофе, аспирине, витаминных
препаратах, пиве, маслах и жирах, медикаментах, сигаретном дыме и бензино-
вых испарениях. Яды насекомых и змей содержат нейротоксины — вещества,
влияющие на нейрональную передачу сигнала. В яйцах содержится холин, из
которого в организме синтезируется нейромедиатор ацетилхолин — молекула,
необходимая для поддержания бодрствующего состояния. Молоко содержит
аминокислоту триптофан, которая является биохимическим предшественником
серотонина — именно поэтому стакан теплого молока на ночь способствует
хорошему сну. Кофе, чай и шоколад содержат некоторое количество кофеина,
который конкурентно ингибирует с нейромедиатором аденозином (рис. 16.1).
Нейротрансмиттеры, как вы можете видеть, представлены в повседневной жиз-
ни самым широким образом.
Самые обычные продукты питания содержат большие количества
нейроактивных молекул, среди которых можно отметить сахара (энергетические
потребности мозга удовлетворяются главным образом ими), холин и целые
полипептиды, содержащиеся в молоке, мясе, яйцах и орехах. Кальций используется
для осуществления разнообразных функций клетки, включая и нейрональную
передачу сигнала. Все вышеперечисленное мы получаем из пищи, которую едим, и
воздуха, которым дышим.
Рис. 16 .1 . Чаю, кофе? А может, аденозина? Трехмерная структура молекулы определя-
ет большую часть ее биологических свойств. Кофеин и теофиллин по своей структуре
близки к нейромедиатору аденозину, и, связываясь с аденозииновыми рецепторами, они
повышают внимание. Аденозин также является частью основной биологической макро-
эргической молекулы АТ Ф (аденозинтрифосфата); нейромедиаторы часто синтезиру-
ются из существующих предшественников
Гены и молекулы мышления
261
Все живые существа имеют в своей основе углеродные цепочки, из которых
пу тем различных модификаций и получается бесконечное разнообразие веществ
в живой клетке. При таких модификациях в состав молекулы включаются атомы
водорода, азота, кислорода, кальция, магния и некоторых других элементов. Белки
представляют собой длинные полимерные цепочки, состоящие из аминокислот —
молекул, имеющих NH2
- и COOH-группы. Все аминокислоты могут быть получены
из белковой пищи, которая содержит 12 незаменимых аминокислот.
1.1 . Основные малые молекулы клетки так же используются
для нейрональной сигнализации
Лексикон ученых изобилует различными специальными названиями и терми-
нами, и количество названий и типов молекул, имеющих отношение к нервной
деятельности, поначалу может ошеломить. Слово «каннабиноиды», происхо-
дит от латинского названия растения, содержащего психоактивные вещества —
конопли Cannabis sativa; при этом окончание «-оиды» говорит нам о том, что
это целый класс молекул, первый представитель которого был обнаружен в
означенном растении. Эндоканнабиноиды, как мы можем видеть из названия,
являются эндогенными каннабиноидами, синтезируемыми в организме.
Вещества часто называют по первой из их функций, которую удалось устано-
вить. Так, серотонин был впервые описан как вещество, повышающее кровяное
давление (от лат. tonus — давление и serum — сыворотка). Окончание «-ин»
говорит нам о белковой природе вещества (такое же окончание имеет более рас-
пространенное в западных странах слово «протеин», используемое вместо слова
«белок»). Хотя в кровотоке это вещество действительно служит для регуляции
давления, в мозге оно выполняет совсем другие функции — регулирует цикл сна
и бодрствования и влияет на настроение и сон. Однако первоначальное название
серотонина сохранилось, что может облегчить его запоминание. Как вы можете
видеть, химические вещества часто имеют собственную историю.
Для запоминания названий тех или иных соединений могут быть полезны
мнемонические приемы. Чтобы запомнить то или иное слово, постарайтесь най-
ти похожие на него слова среди тех, что вы уже знаете (так, слово «серотонин»
может звучать для вас как «серая туника», что и можно использовать; особенно
хорошо такая хитрость работает, если вы можете представить это визуально).
При запоминании визуальной информации, примерами которой могут высту-
пать функциональная карта мозга или схема строения синапса, замечательно
работает рисование этих структур. Обучение — это активный и интерактивный
процесс, и чем больше вы можете сделать с материалом этой главы, тем лучше
вы его запомните. Если вы сможете своими словами рассказать основные мо-
менты, то это будет означать, что у вас отныне имеется прочный фундамент для
дальнейшего понимания.
Сам мозг также предоставляет нам определенные шаблоны для запомина-
ния. Один из них состоит в том, что многие молекулы используются для выпол-
нения более чем одной задачи. Большинство веществ, которые обсуждаются в
настоящей главе, выполняют общие функции в организмах самых разных жи-
вотных — начиная с плодовой мушки дрозофилы и моллюска аплизии и закан-
чивая человеком. Соединения, вовлеченные в энергетический метаболизм и пе-
редачу наследственной информации, также используются и для нейрональной
сигнализации. Все клетки поддерживают определенную разность потенциалов
262
Глава 16
на своей мембране, но только нейроны могут использовать ее для генерации по-
тенциала действия вдоль аксона. Все клетки используют АТФ в качестве пере-
носчика энергии, и в то же время из него может синтезироваться нейромедиа-
тор. Глутаминовая кислота — одна из основных аминокислот в клетке, а также
основной возбуждающий медиатор в мозге. И подобных примеров — десятки.
И тот факт, что основные вещества клетки используются для нейрональной
сигнализации, облегчает их запоминание.
Низшие животные часто используются в качестве экспериментальных мо-
делей для изучения тех или иных биологических процессов. Синапсы имеют
очень малые размеры, сложно устроены и требуют для своего изучения инва-
зивных вмешательств, проведение которых на человеке не очень приемлемо с
этической точки зрения. По этой причине синапсы изначально изучались на
модельных животных и культурах клеток. И эти методики все еще продолжают
приносить плоды.
Животные имеют определенный набор высококонсервативных генов, ве-
ществ и нервных клеток. Говоря «высококонсервативные», мы подразумеваем,
что такие структуры обнаруживаются у животных, принадлежащих к разным
видам, классам, а порой и типам. Так, человек и насекомые используют сход-
ный набор нейромедиаторов. Конечно, не подразумевается, что человек устро-
ен так же, как дрозофила, но дрозофила становится модельным организмом,
приемлемым для изучения свойств некоторых человеческих нейромедиаторов.
Электрическая активность мозга движущейся плодовой мушки практически в
точности повторяет электроэнцефалограмму просыпающегося мозга человека.
Такие сходства в биологии организмов, десятки миллионов лет эволюциониро-
вавших независимо, составляют одну из интереснейших сторон изучения био-
логии существ, имеющих мозг.
Обратив внимание на более близкие к нам с эволюционной точки зрения
организмы, можно обнаружить, что млекопитающие имеют примерно те же
структуры в головном мозге, что и мы. Новая кора, и особенно ее фронтальные
доли, как раз относятся к таким структурам. Мозг, таким образом, содержит
свою эволюционную историю.
Молекулы работают на наиболее фундаментальном уровне биологического
контроля. Гены представляют собой участки молекулы ДНК, почти два метра
которой содержится в каждой клетке и транскрибируемой в РНК, которая затем
транслируется в белки (рис. 16.2).
Мозг не может работать без постоянного притока кислорода и глюкозы, ко-
торая составляет примерно четверть от всей потребляемой телом энергии. Он
также нуждается в белках, витаминах, минеральных веществах, маслах и жи-
рах — последние нужны для построения мембран клетки и миелиновых обо-
лочек аксонов. Некоторые жиры способны поднимать настроение (это было
продемонстрировано для омега-3 -жиров). Такие вещества, как дофамин и аце-
тилхолин, необходимы для получения удовольствия, для бодрствования и фор-
мирования кратковременной памяти.
Молекулярные механизмы работы мозга поначалу вполне могут парализо-
вать своей сложностью. В задачи главы не входит их полное описание — вме-
сто этого она познакомит вас с основными положениями. Несмотря на очевид-
ную сложность молекулярных процессов, возможно представить механизмы,
Гены и молекулы мышления
263
при помощи которых эффект того или иного вещества увеличивается или
уменьшается. Агонистом дофамина называют любое вещество, усиливающее
его действия. Так, знаменитое лекарство L-ДОФА представляет собой один из
его метаболических предшественников и при введении в организм усиливает
выработку дофамина, а следовательно, и его эффект. SSRI-антидепрессанты яв-
ляются селективными ингибиторами реабсорбции серотонина — они уменьша-
ют скорость поглощения и распада серотонина. Если же то или иное вещество
не распадается, то происходит его накопление, и, как следствие, — усиление
действия. Поэтому SSRI-антидепрессанты также являются агонистами серото-
нина.
В противоположность агонистам антагонисты уменьшают действие того
или иного медиатора. Примером антагониста может служить препарат налок-
сон, применяемый при лечении передозировки опиатов. Налоксон препятству-
ет связыванию опиатов с рецептором, блокируя доступ к местам его связывания
и уменьшая таким образом действие опиатов. Как и большинство психоактив-
ных веществ, опиаты имеют важное значение при терапии, но при этом их ис-
пользование сопряжено с определенным риском.
В конечном итоге, как бы ни был сложен конкретный механизм дейстия того
или иного вещества, психоактивные соединения увеличивают или снижают ин-
тенсивность этого действия.
Эта глава познакомит вас с молекулами, важными для нервной деятельно-
сти. Конечной же целью изучения этих молекул является понимание того, как
именно молекулы участвуют в передаче и обработке сигналов, лежащей в осно-
ве функционирования человеческого мозга.
1.2 . Живые организмы используют ферменты
Все известные на данный момент живые организмы спообны существовать
в весьма узком диапазоне температуры, давления и кислотности. Люди научи-
лись готовить пищу 10 тыс. лет назад, а выплавлять сталь — около двух тысяч;
Рис. 16 .2 . ДНК и ферментативные реакции. В широком смысле все вещества в живой
клетке подвержены влиянию ферментов, в том числе и сама ДНК. Ферменты ускоряют
химические реакции, связывая молекулы субстрата. Они способны расщеплять, транс-
портировать и высвобождать другие молекулы, подобно наномашинам. Ферментатив-
ные реакции протекают гораздо быстрее обычных. ДНК выступает одновременно и как
хранилище наследственной информации, и как источник матриц для синтеза большин-
ства молекул клетки. (Источник: National Institute of General Medical Sciences, 2009.)
264
Глава 16
именно тогда были заложены основы современной цивилизации. Однако тело
человека не может переносить экстремальных значений температуры, давления
и кислотности — они попросту убьют его. Это связано во многом с тем, что все
организмы используют для своего существования ферментативный катализ, ко-
торый осуществляется, как несложно догадаться, ферментами. Все молекулы
мозга действуют через то или иное воздействие на ферменты.
Самой простой аналогией, которая позволила бы представить себе механизм
работы фермента, является, пожалуй, сравнение фермента и субстрата с замком
и ключом; при этом именно пространственное строение этих молекул позволя-
ет осуществлять связывание одного другим, перенос, изменение конформации
или передачу сигналов. Многие витамины являются необходимыми составны-
ми частями ферментов, без которых жизнедеятельность клетки невозможна.
Витамин фолиевая кислота, также известный как витамин B12, задействован
в осуществлении важных функций в мозге. Синаптическая передача сигнала
представляет собой фермент-опосредованную сигнализацию.
1.3 . Одна и та же молекула может нести разные сигналы
Конопля производит тетрагидроканнабинол вовсе не для того, чтобы человек
получал при его употреблении некие ощущения. Кофейное дерево синтезирует
кофеин не для обеспечения человека напитком к завтраку. Означенные веще-
ства вовлечены в соответствующих организмах совсем в другие процессы, не-
жели те, в которых они участвуют у нас в мозге.
Но в чем же тогда причина такого сильного влияния растительных веществ
на мозг? Ведь очень много различных лекарственных препаратов было впервые
выделено именно из растений. Аспирин (ацетилсалициловая кислота), к при-
меру, был впервые получен из коры ивы. Кураре входит в состав яда для стрел,
которым пользуются аборигены Гайаны; именно это вещество не так давно
сыграло ключевую роль в открытии ацетилхолина. Тысячи нейроактивных ве-
ществ были изначально выделены из растений и животных. Как вы можете ви-
деть, химизм жизни высококонсервативен — одно и то же вещество может быть
и инсектицидом, и обладать при этом психоактивностью.
Одно и то же соединение может иметь различные функции даже в одном и
том же организме. Адреналин стимулирует многие ткани тела, переводя их в
режим повышенной готовности, при котором учащается сердцебиение и уси-
ливается кровоснабжение легких и мышц. Адреналин секретируется надпочеч-
никами (лат. ad-
—
«над», renes — «почки», -in — окончание, указывающее
на белковую природу). В мозге это вещество называется эпинефрином (греч.
эпи- — « над», нефрос — «почка», -ин
—
окончание, указывающее на белковую
природу).
В центральной нервной системе норэпинефрин является основным нейро-
медиатором, обусловливающим бодрствование, что отличается от его гормо-
нальных функций в остальном теле. Одно и то же вещество, таким образом,
может выполнять различные функции в разных частях тела.
1.4 . Гематоэнцефалический барьер защищает
внутреннюю среду мозга
Способность адреналина выполнять разные функции в теле и мозге обусловле-
на его неспособностью проникать из тела в мозг и наоборот; это явление носит
Гены и молекулы мышления
265
название компартментализации и позволяет вашему мозгу сохранять нормаль-
ный уровень эпинефрина, когда вы чем-либо возбуждены. Гематоэнцефаличе-
ский барьер был открыт в начале прошлого века, когда было показано, что си-
няя краска, инъецированная в мозг, не распространяется по остальному телу, и
наоборот. На рис. 16.3 приведены основные свойства гематоэнцефалического
барьера.
Образно гематоэнцефалический барьер можно представить как раздели-
тельную стену между телом и мозгом. Он позволяет мозгу регулировать хими-
ческий состав своей внутренней среды вне зависимости от состава крови. Точ-
Рис. 16 .3 . Мозг окружен несколькими последовательными слоями защиты. Гематоэн-
цефалический барьер подстилает кровеносные сосуды мозга и полости ликвора. Это
позволяет мозгу поддерживать свою собственную внутреннюю среду, которая отделена
от других частей тела и окружающей среды. Большие молекулы, такие как белки, не-
способны преодолеть гематоэнцефалический барьер в случае, если они не предназна-
чены для транспортировки в мозг. Похожие образования защищают мозг плода и череп
взрослого от проникновения нежелательных веществ. (Источник: Saunders et al., 2008.)
266
Глава 16
но так же организм в целом отделен от внешней среды кожей, препятствующей
свободному проникновению различных соединений извне в организм. Каждая
клетка мозга имеет несколько барьеров, первым из которых является ее мем-
брана; за ней следуют глиальные клетки и гематоэнцефалический барьер, кото-
рые, в свою очередь, окружены периферическими тканями и, наконец, кожей.
В самой клетке содержится еще один барьер — оболочка ядра, внутри которой
заключена одна из важнейших структур клетки — генетический аппарат. Весь
наш организм с этой точки зрения представляет собой биоконтейнеры внутри
биоконтейнеров.
Головной и спинной мозг имеют свою собственную систему обращения
жидкости, отдельную от кровотока. Она заполнена так называемой церебро-
спинальной жидкостью, или ликвором. Ликвор служит для транспротировки
различных веществ и клеток. Постоянный, но при этом контролируемый при-
ток различных соединений в мозг обеспечивается кровью и ликвором.
Гормонами называют сигнальные молекулы, доставляемые к органам-ми -
шеням кровотоком, тогда как нейрональные мессенджеры не распространяют
своего действия за пределы нервной системы. Однако мозг пронизан большим
количеством кровеносных сосудов самого разного размера. Как мы уже видели,
МРТ позволяет отслеживать изменения в головном мозге, обусловленные уров-
нем кислорода в крови; они связаны в первую очередь с усилением притока
крови для удовлетворения возросших потребностей участка мозга в кислороде.
Селективность гематоэнцефалического барьера при этом позволяет кислороду
и глюкозе свободно диффундировать сквозь него и вместе с тем препятствует
свободной диффузии молекул большего размера, таких как адреналин.
1.5 . Модельные организмы
Горох, насекомые, черви и крысы занимают почетное место в науке, посколь-
ку они долгое время служили в качестве объектов для экспериментов, кото-
рые нельзя было проводить на людях (рис. 16.4). Каждый из перечисленных
организмов занял свое место в результатах исследований, которые были бы
трудноосуществимы на других моделях. В наши дни на передний план вышел
круглый червь C. elegans, главным образом потому, что имеется возможность
идентификации каждого нейрона в его теле. В силу высокой консервативности
биологических процессов данные, получаемые при его изучении, а равно и при
изучении мышей и дрозофил, позволяют судить о процессах, происходящих в
мозге человека. Синаптическое обучение изначально было исследовано на ги-
гантском аксоне морского зайца Aplisia, поскольку он был достаточно велик
для изучения под световым микроскопом. И лишь несколько десятков лет спу-
стя были получены прямые доказательства наличия синаптического обучения у
человека. Можно сказать, что биологи в своих экспериметах изначально всегда
смотрят не туда. Поэтому важно не иметь предубеждений относительно недо-
пустимости тех или иных явлений или процессов.
Изучение дрозофил и гороха началось еще в конце XIX в. После биогене-
тической революции последних десятилетий было проведено большое коли-
чество экспериментов, включавших нокаутные и нокиновые организмы. При
использовании ретровирусов возможно ввести новые гены в организм или от-
ключить уже имеющиеся. И хотя на настоящий момент имеется масса экспе-
Гены и молекулы мышления
267
риментальных данных, адекватной замены непредвзятому обдумыванию про-
блемы альтернатив до сих пор нет. Нейробиология — отнюдь не устоявшаяся
наука, и в процессе изучения ее проблем сюрпризы более чем возможны.
Первым из открытых человеком нейромедиаторов стал ацетилхолин. Он
был открыт при изучении нервно-мышечного синапса подопытных животных,
находящегося за пределами ЦНС. Открытие этого нейромедиатора оказало
огромное влияние на нейробиологию. Так, последующие исследования позво-
лили установить, что существуют два разных холинергических рецептора, об-
условливающих различные ответы. Первый из этих рецепторов носит название
никотинового, поскольку отвечает на никотин — вещество, содержащееся в та-
баке. Второй рецептор называется мускариновым, так как, по аналогии с пер-
вым, активируется мускарином — токсином мухомора Muscarina amanita.
Сам по себе ацетилхолин имеет две разных функции: за пределами ЦНС он
выступает в качестве нейромышечного возбуждающего медиатора, тогда как в
мозге он является основным нейромодулятором, функции которого сильно от-
личаются от выполняемых им на периферии. Этот факт в корне изменил в свое
время понимание функций ацетилхолина. Холинергическая система мозга слу-
жит для перевода организма из состояния бодрствования в состояние быстрого
сна. Вот и еще один пример выполнения одним и тем же веществом разных
функций.
Изучение ацетилхолинового синтеза также позволило обнаружить фермент,
деактивирующий ацетилхолин. Ацетилхолинэстераза разрушает ацетилхолин
после его высвобождения в синаптическую щель. Очистка синапса от медиа-
Рис. 16 .4 . Кора головного мозга мыши: измерение экспрессии гена. Экспрессия проис-
ходит во всем мозге. На этом рисунке показан мозг мыши, обработанный репортерным
флуоресцентным красителем. Краситель реагирует на экспрессию генов под контролем
промотора АRC, которые задействованы в обучении и нейрональной пластичности. От-
крытия, сделанные на других видах, позволяют пролить свет на механизмы контроля
активности генов в мозге человека. (Источник: Eguchi and Yamaguch, 2009.)
268
Глава 16
тора является одним из ключевых моментов в его функционировании. Если бы
подобной очистки не происходило, постсинаптическая клетка находилась бы
под действием нейромедиатора даже после того, как сигнал, вызвавший его вы-
брос, перестал поступать. Это — одна из многочисленных причин, по которым
содержание биоактивных молекул в организме строго регулируется. (Оконча-
ние -аза в названии фермента указывает на то, что он разрушает молекулу, к
которой специфичен; в данном случае — молекулу ацетилхолина).
2.0 . Гены в эволюции, онтогенезе и повседневной жизни
Положение генетики долгие годы было спорным, поскольку она говорит о био-
логичности и наследуемости межрасовых различий. Важно подходить к гене-
тике со всей возможной аккуратностью и ответственностью, поскольку на всех
нас оказывают влияние те гены, которые мы несем. Различия между людьми
теперь рассматриваются в основном как результат происхождения на разных
континентах и от различных предков. Однако такие различия существуют от-
носительно недавно. Наши предки-гоминиды эволюционировали в Африке в
течение нескольких миллионов лет (рис. 16.5). И лишь примерно 30–70 тыс. лет
назад Homo sapiens sapiens начал расселяться по всему миру; также не редки
были миграции и внутри самой Африки. Геном человека принял свой совре-
менный вид, по-видимому, в результате снижения численности популяции при-
мерно до 5000 особей около 100 000 лет назад. Такие события в эволюционной
истории вида снижают генетическую вариабельность, из-за чего геномы совре-
менных людей различаются примерно на 1–2%.
Большая часть психологических и мозговых различий наследуема примерно
на 50%. Главной задачей на сегодняшний день является определение конкрет-
ных функций того или иного гена — например, его принадлежность к развитию
генетических заболеваний. Каждый человек индивидуален с биохимической
точки зрения, чем, к примеру, обусловлены различные ответы разных людей на
одно и то же лекарство. И сейчас мы стоим лишь у истоков понимания таких
различий.
Наши предки почти наверняка были темнокожими, что вызвано высокой ин-
тенсивностью солнечного света в экваториальных областях. Солнечный свет
вызывает распад фолиевой кислоты — витамина, необходимого для размноже-
ния. Продолжительное облучение может также вызывать рак кожи, но вместе
с тем солнечный свет необходим для синтеза витамина D. В областях с повы-
шенной интенсивностью солнечной радиации люди вырабатывают большие ко-
личества пигмента меланина для защиты от ее избытка. Именно этот пигмент
обусловливает окраску кожи; имеются небольшие различия в его структуре
у разных популяций людей.
Существует предположение, что первые современные с поведенческой точ-
ки зрения люди — люди, способные абстрактно мыслить, использующие ору-
дия, огонь и имеющие речь — примерно соответствовали людям современного
южноафриканского племени Сан. Интересно отметить, что строение и биохи-
мия мозга проявляют очень высокую консервативность. Внешние же различия
между людьми отражают адаптации их предков к различным температурам,
переносчикам болезней, социальным условиям и пище в местах, где они неког-
Гены и молекулы мышления
269
да обитали. Так, молочные продукты требуют наличия ферментов для усвоения
лактозы; при этом немало людей таких ферментов во взрослом состоянии не
имеют. Некоторые группы людей выработали ферменты для усвоения молока,
которое отличается от материнского по составу, как, например, молоко коров и
коз. Другие приспособления были вызваны низкими температурами, токсинами
и инфекционными заболеваниями.
Несмотря на эти изменения, мы мало отличаемся от наших предков, живших
сотни тысяч лет назад. В начале этой главы был приведен образец наскальной
живописи из пещеры, найденной в 1940 г. во Франции, неподалеку от дерев-
ни Ласко. Возраст рисунков оценивается примерно в 16 000 лет. Когда Пабло
Пикассо увидел эти рисунки вскоре после их обнаружения, он разочарованно
сказал: «Мы не придумали ничего нового в искусстве». И хотя мы не знали,
как думали первобытные люди, их рисунки, тем не менее, близки и понятны
нам.
Рис. 16 .5 . Четыре миллиона лет эволюционной истории гоминид. Человекообразные
существа обитали на деревьях уже 4 млн лет назад, т. е. до того времени, когда обитал
предок человека, названный Люси (см. выше). Эта схема отражает современные пред-
ставления о местоположении Homo sapiens среди его предков и родственных видов.
На сегодняшний день бонобо и шимпанзе являются нашими наиболее близкими род-
ственниками среди гоминид (нижний правый угол). Однако несколько тысяч лет назад
сосуществовало несколько видов гоминид (верхний правый угол). (Источник: Kaas and
Preuss, из Squire et al. 2003 .)
270
Глава 16
Таблица 16.1
Исследование риска заболеваний нервной системы методом близнецов
Заболевание
Совпадение у одно-
яйцевых близнецов
Совпадение у дву-
яйцевых близнецов
λs
Рассеянный склероз
25% [54]
5% [54]
20
Аутизм
75% [60]
16% [60]
45 [61]
Мигрень
50% [62]
21% [62]
4 [63]
Болезнь Альцгеймера 59% [64]
32% [64]
5
Шизофрения
50% [65]
4% [65]
10 [66]
Полиомиелит
36%
6%
Рахит
91%
23%
Туберкулез
32%
14%
Изучение близнецов помогает установить паттерны наследования генов. Для однояйце-
вых близнецов величина совпадения может быть весьма высокой, тогда как у двуяйце-
вых она по тем или иным причинам бывает ниже.
2.1. Разные события с ДНК происходят
в различные промежутки времени
Различные события происходят с ДНК в очень разные по продолжительности
промежутки времени. Гены могут работать в течение геологических периодов
времени — сотен миллионов лет. Также они работают в поколениях, продолжи-
тельность которого у человека составляет 15–30 лет. Однако даже в пределах
24 ч они способны менять свою активность. На настоящий момент известно,
что они контролируют цикл сна и бодрствования, обучение и нейрональную
пластичность, иммунный ответ, старение, развитие заболеваний и выздоров-
ление. Ваш мозг в настоящий момент подвергается генетическим процессам,
позволяя вам запоминать то, что вы читаете.
Запомните, что гены выполняют три основные функции: они направляют
эволюцию мозга, к примеру развитие лобных долей, отвечающих за речь, со-
циальные взаимоотношения и способность распознавать выражения лица. Они
контролируют развитие мозга на протяжении всей жизни человека, начиная с
внутриутробного развития и заканчивая смертью. В нервных клетках генами
контролируются мгновенные события, такие как обучение и погружение в сон.
По всему миру люди давно отмечали сходство между родственниками, и в
этом нет ничего нового. Но влияние генов на поведение человека долгое время
оспаривалось в научной среде. Большая часть доводов против такого влияния
была опровергнута в последние несколько десятков лет, когда началось под-
робное изучение близнецов, братьев и сестер, а также многодетных и просто
больших семей. Исследователи обнаружили сходства, выраженность которых
зависела от степени родства. Наибольшее сходство было отмечено среди одно-
яйцевых близнецов, несколько меньшим оно было для близнецов разнояйце-
вых; еще меньшим — для братьев и сестер и прочих родственников (табл. 16.1).
В некоторых семьях частота встречаемости определенного заболевания оказа-
лась заметно выше средней. Такие семьи впоследствии изучались с целью уста-
новления причин и механизмов развития таких заболеваний.
Гены и молекулы мышления
271
Одним из наиболее известных примеров является семья КЕ из Великобрита-
нии. В этой семье на протяжении трех поколений наблюдались тяжелые нару-
шения речи; при этом подвержено им было около половины ее членов. Во всех
трех поколениях существовала повышенная вероятность потери участка ДНК с
геном FOXP2, которая затем наследовалась аутосомно и доминантно. Изучение
генома семьи выявило делецию в 7-й хромосоме, вызванную мутацией.
Очень быстро распространилось предположение о том, что открыт ген речи.
Это имело определенные последствия. В предыдущих главах описывалось поле
Брока (латеральный участок нижней фронтальной извилины, чаще всего в ле-
вом полушарии), которое отвечает за речь. Магнитно-резонансная томография
членов семьи КЕ показала сниженную активность поля Брока при произне-
сении фраз и выявила дефекты базальных ганглиев и их контроля голосового
тракта. Однако мутации гена FOXP2 затрагивают не только произношение, но
и понимание речи.
Тот факт, что человек обучается речи очень рано, в то время как его ближай-
шие эволюционные родственники — шимпанзе и бонобо — к ней вообще не
способны, свидетельствует в пользу наличия у человека некого особого свой-
ства, определяющего способность говорить. Манипулирование словами и сим-
волами сформировалось в эволюции человека сравнительно недавно — при-
близительно 50–100 тыс. лет назад. Это позволяет выдвинуть FOXP2 в качестве
одного из основных кандидатов на роль «гена речи».
Интерес к этому предположению, тем не менее, несколько ослаб после того,
как было установлено, что этот же ген отвечает за голосовую сигнализацию у
певчих птиц, эхолокацию у летучих мышей, а также за звуковые сигналы у мы-
шей, обезьян и даже рептилий. Некоторые птицы, как, например, африканский
серый попугай, способны воспроизводить человеческую речь, однако функции
такой способности отличаются от таковых у человека. Попугаи — превосход-
ные звукоподражатели и отлично приспособленные к окружающей среде со-
циальные животные. Поэтому FOXP2 может определять функцию, одинаковую
у человека и попугаев — а также у аллигаторов. Наиболее очевидной из таких
функций является контроль развития и функционирования голосового тракта.
Сейчас предполагают, что FOXP2 контролирует работу других генов, участву-
ющих в развитии мозга, легких и кишечника.
Все данные указывают на то, что наличие языка у человека не сводится к
единственному свойству. Язык определяется целым набором свойств, некото-
рые из которых имеются и у других видов — к примеру, особенности строения
легких и дыхательной мускулатуры, контроль дыхания и вокализации, способ-
ность к оценке окружения и действиям согласно этому окружению и способ-
ность обмениваться информацией с другими представителями вида. В насто-
ящий момент ведутся интенсивные работы по поиску генов, принимающих
участие в развитии речи. Эти исследования помогут найти лекарства для людей
с дефектами речи, а возможно, даже учить новые слова и языки.
2.2 . Развитие мозга: нейрогенез, затем синаптогенез
Разные гены и молекулы играют различные роли на разных стадиях развития.
Полезно будет понять этот простой факт в терминах первого и второго репер-
туаров нейрональной вариации и селекции Джеральда Эдельмана. В первом
репертуаре во время роста плода десятки миллиардов нейронов растут, образуя
272
Глава 16
головной и спинной мозг, а также периферические нервы. В это же время проис-
ходит дифференцировка нейронов на различные классы. На момент рождения,
когда эти процессы в большей степени завершаются, человек имеет наиболь-
шее число нейронов за весь период жизни (рис. 16.6). Это вызвано тем, что ней-
роны постоянно отмирают, если образуют нестабильные или непродуктивные
соединения. В мозге новорожденного отмирает большое количество нейронов
в процессе дифференцировки их соединений. Такой механизм размножения и
отмирания в большой степени подчиняется дарвиновским представлениям об
эволюции — наиболее загруженные работой нейроны и циклы выживают, тогда
как остальные уничтожаются. Таким образом, функциональные пути в мозге
появляются на достаточно ранних стадиях развития, и по мере приближения
момента рождения плод проводит большую часть времени в состоянии, похо-
жем на состояние быстрого сна, показывая характерную энцефалограмму с бы-
стыми движениями глаз.
Когда в начале третьего триместра плод начинает использовать свои органы
чувств и учиться, активируется второй репертуар. Приток новых нейронов по-
Рис. 16 .6. Окружающая среда и гены на протяжении жизни. Среда и гены взаимодей-
ствуют на протяжении всей жизни. (а) Пролиферация: незадолго после оплодотворе-
ния нейроны начинают делиться и расти — этот процесс носит название нейрогенеза.
(б) Дифференциация и миграция: клетки дифференцируются по форме и функциям,
мигрируют и образуют отростки. (в) Направление роста аксона играет важную роль в
этом слабо исследованном процессе. (г) Аксоны и дендриты образуют синапсы с дру-
гими нейронами; успешные синапсы сохраняются, бесполезные — исчезают. (д) Сети
в мозге делают возможной обработку информации. На момент начала развития мозга
синапсов в нем нет, тогда как в полностью сформировавшемся мозге их порядка тысячи
триллионов. (Источник: Yehezkel, 2008.)
Гены и молекулы мышления
273
степенно сходит на нет, тогда как новые синапсы начинают возникать в огром-
ных количествах — порядка триллионов. За таким интенсивным синаптогене-
зом следует отмирание синапсов. На этой стадии отбору подвергаются уже не
нейроны, а синапсы (см. гл . 3 и 10). Синапсы, образующие работающие со-
единения, сохраняются, тогда как не служащие для выполнения тех или иных
функций исчезают. Синаптические соединения мозга определяют его функци-
онирование, и поэтому второй репертуар работает на протяжении всей жизни
человека — образуются новые синапсы, а нефункционирующие деградируют.
Во время функционирования первого репертуара нейрогенеза образуются
и развиваются миллионы нейронов, гены и молекулы управляют ростом мозга
и направляют нервные окончания к их органам-мишеням. После установления
основных структур мозга те же гены и молекулы переходят на регуляцию актив-
ности мозга. К примеру, ГАМК, в норме являющаяся тормозным медиатором,
во время развития мозга действует как медиатор возбуждающий и принимает
участие в направлении роста аксонов.
Результатом такой адаптивной стратегии является огромная адаптивная ва-
риабельность (рис. 16.7). Стоит помнить, что человек, как и множество других
видов животных, вынужден постоянно иметь дело с угрозой голода, обезвожи-
вания, новых болезней, мутаций, появлением новых социальных или половых
взаимоотношений, а также многими другими обстоятельствами. Когда человек
Рис. 16.7. Нейроны образуются на протяжении жизни. Гиппокампальное обучение
крысы ориентированию в лабиринте: означает ли оранжевый цвет стенки то, что слева
от нее находится пища? В областях СА1 и СА3 гиппокампа имеются так называемые
клетки места, ответственные за обучение поведению относительно ориентиров и от-
слеживание скорости и положения тела при прохождении лабиринта. Хотя масштабный
нейрогенез завершается к моменту рождения, некоторые части мозга сохраняют актив-
ные нейрональные стволовые клетки на протяжении всей жизни. Такими областями яв-
ляются зубчатая извилина гиппокампа и субвентрикулярная зона, расположенная ниже
латеральных желудочков. Новые клетки мозга обусловливают повышенную пластич-
ность. (Источник: Kempermann, 2008.)
274
Глава 16
одомашнил коров и коз и начал использовать их молоко, люди с неработающим
генетическим аппаратом усвоения молока столкнулись с реальной угрозой сво-
ему здоровью. Все при этом зависело от их биохимических свойств.
Гены и молекулы отвечают как за наше здоровье, так и за болезни. Мы об-
судим когнитивные, социальные и нейродегенеративные расстройства мозга.
Однако большинство генов в нашем теле активно лишь в некоторые периоды
времени — когда это требуется для роста и поддержания мозга и тела человека.
2.3 . Как человек получил такой большой мозг?
Мозг человека за последние несколько сот тысяч лет быстро увеличивался в
размерах, значительно опережая тело. Данный факт свидетельствует о нали-
чии в геноме определенных изменений, ответственных за такое опережение,
но что это за изменения — неизвестно. Во всей живой природе не существует
аналогов передних полей фронтальных долей головного мозга человека. Тем не
менее строение мозга приматов сходно со строением мозга других млекопита-
ющих, как видно из рис. 16.8, где одинаковые зоны показаны одинаковыми цве-
тами. Строение коры в классе млекопитающих высококонсервативно, однако
неокортекс и фронтальные доли у наиболее продвинутых его представителей
имеют тенденцию к увеличению.
Рис. 16 .8 . Приматы и мыши. Хотя мозг этих двух групп сильно различается, основные
зоны в нем очень консервативны (окрашенные зоны на рисунках). Гиппокампальная
область очень сходна по строению у всех млекопитающих, и даже у млекопитающих
и птиц, и имеет гомологии с гиппокампальными областями рептилий и рыб. Внешняя
часть мозга человека представлена в основном церебральной корой и мозжечком (вни-
зу). У грызунов неокортекс выглядит как плоская поверхность над обонятельной частью
мозга. Неокортекс фактически представляет собой шестислойную плоскую структуру,
но у человека и китов он настолько обширен, что его поверхность образует складки —
извилины. (Источник: Eichenbaum, 2003.)
Гены и молекулы мышления
275
Homo sapiens выделяется интенсивным расширением церебральной коры
префронтальных долей, которую иногда называют «органом цивилизации» (см.
гл. 12). Это выделяет человека среди прочих животных. Неокортекс человека
примерно в 1000 раз больше неокортекса ранних млекопитающих. Другие мле-
копитающие с крупным мозгом, такие как слоны и дельфины, также имеют раз-
витую кору, однако неокортекс занимает у них меньшую площадь относительно
всей коры. Наиболее важными свойствами новой коры являются контроль со-
циального поведения и исполнительный контроль, а также участие в языковой
коммуникации. Исполнительный контроль позволяет человеку строить пла-
ны и работать в группах. Язык и символизм служат отличительными чертами
человеческой культуры. Социальное мышление является ключом к созданию
эффективного общества, но оно также обусловливает агрессию по отношению
к чужакам, которая контролируется серотониновой и моноаминоксидазной
системами. Агрессия может быть полезна охотникам и собирателям, которые
должны быть готовы защититься от угрозы или убить жертву или соперника,
но в высокоорганизованных социальных группах эта функция в значительной
мере утрачивает свою полезность.
3.0 . Экспрессия и регуляция генов
Каждая клетка человеческого тела несет свою копию генома нашего вида в фор-
ме длинной двухцепочечной молекулы ДНК, закрученной в двойную спираль.
Генетическая информация экспрессируется в белки, определяющие фенотип,
развитие организма и регуляцию его функций. В каждой клетке ядерная ДНК
транскрибируется в матричную РНК (мРНК), которая затем транслируется
с образванием белков.
Формальное определение гена гласит, что он является «участком цепи ДНК,
ответственным за единицу наследования, который ассоциирован с регулятор-
ными последовательностями, транскрибируемыми участками и/или другими
функциональными последовательностями» (Pearson, 2006). Последние иследо-
вания показали, что в геноме имеется гораздо больше регуляторных последо-
вательностей, чем предполагали ранее. Простая экспрессия белка с гена встре-
чается очень редко. Геном — сложная функциональная сеть, а не словарь для
перевода последовательности нуклеотидов в последовательность аминокислот.
В некоторых случаях, как в случае наследуемых по Менделю признаков
дрозофилы, можно наблюдать, как с ДНК идет экспрессия белков, определяю-
щих хорошо различимый признак — например, окраску глаза. Однако большая
часть генов работает гораздо шире, а один и тот же признак могут контролиро-
вать несколько генов.
ДНК прямо или непрямо экспрессируется в протеом — видоспецифичный
набор белков, состоящих из аминокислот, имеющих аминогруппу NH3 и кар-
боксильную группу СООН. Белки, наряду с жирами и малыми молекулами, яв-
ляются основными веществами в клетке.
3.1. Обработка информации в клетке
Центральная догма молекулярной биологии говорит о наличии в клетке трех
типов информационных молекул и трех основных путях взаимодействия между
ними (рис. 16.9).
276
Глава 16
1. Репликация ДНК
Первый путь передачи информации заключается в репродукции ДНК, которая
позволяет передавать геном родителей потомкам. Этот путь носит название
репликации и традиционно рассматривается как практически неизменный для
всех живых организмов. Большая часть мутаций, затрагивающих его, по всей
видимости, летальны и не сохраняются. Это же свойство лежит в основе того,
что некоторые виды, такие как акулы, не меняются вот уже сотни миллионов
лет. Поскольку, согласно второму закону термодинамики, все системы со вре-
менем утрачивают структуру, такая стабильность генетической информации не
может не удивлять.
Считается, что во время репликации информация копируется абсолютно
точно, однако иногда все же происходят мутации и дрейф генов, позволяющие
виду приспосабливаться к новым условиям, затрагивающим его выживание
или размножение.
2. Транскрипция: ДНК-мРНК
ДНК, которую можно образно назвать чертежом организма, копируется в РНК
в процессе транскрипции. В первом приближении на транскрипцию можно
смотреть как на простое копирование. Однако не стоит забывать, что факторы
транскрипции оказывают сильное влияние на этот этап экспрессии.
3. Трансляция: мРНК-белок
Все живые организмы имеют свои белки — полимеры аминокислот. Факто-
ры, контролирующие активность ДНК и РНК, ферменты, факторы транскрип-
Рис. 16 .9 . Центральная догма молеку-
лярной биологии. Конвейер жизни. Цен-
тральная догма молекулярной биологии
гласит о том, что информация в клетке
идет от ДНК через РНК к белку, и более
никак. Множество исследований опро-
вергли постулат о том, что других путей
передачи информации не существует, но
о них будет сказано позже. (Источник:
Baars.)
Гены и молекулы мышления
277
ции — вс это белки (рис. 16.10). Они обеспечивают основные структурные и
функциональные особенности тела. Трансляция мРНК в белок завершает цепь
экспрессии ДНК.
3.2 . ДНК высших уровней регулирует остальную ДНК
Большая часть геномной ДНК представлена некодирующими участками и не-
посредственно в белки не экспрессируется. Около 3% некодирующей ДНК осу-
ществляет регуляторные функции, контролируя другие участки ДНК. В каче-
стве примера можно привести гены контроля развития тела, контроля развития
мозга и гены, отвечающие за циркадные циклы. Эти гены можно назвать гена-
ми пространственного и временного планирования.
Гены гомеобоксов, называемые Нох, высококонсервативны среди позвоноч-
ных. Именно поэтому человек, мышь и саламандра имеют вытянутое тело с
парой передних и парой задних конечностей, длинный пищеварительный тракт,
а также спинной и головной мозг. Гены пространственного планирования тела
контролируют другие гены, обусловливающие развитие органов и кончностей.
Гены Нох полезно представить как группу генов высшего порядка, контролиру-
ющих и взаимодействующих с генами низшего. На рис. 16.11 показано, каким
образом гены Нох определяют строение передней части тела, шеи и головы у
самых разных животных. На рис. 16.12 показано, как некоторые участки мозга
регулируются генами этого же семейства.
Рис. 16 .10. ДНК: простые единицы дают множество комбинаций. Учитывая неверо-
ятную сложность живых организмов, основные элементы генетического кода оказыва-
ются удивительно просты. Четыре азотистых основания образуют пары, являющиеся
буквами генетического кода (Г — гуанин, Т — тимин, Ц — цитозин и А — аденин).
На рисунке показана хромосома характерной Х-образной формы. Хромосома состоит
из ДНК, окруженной различными белковыми молекулами. (Источник: Министерство
энергетики США.)
278
Глава 16
Рис. 16.11. Гены Нох определяют раз-
витие органов относительно краниокау-
дальной оси. Натуралисты с очень ран-
них времен отмечали сходства между
организмами самых разных групп. Од-
ним из таких свойств является билате-
ральная симметрия всех позвоночных.
На сегодняшний день известно, что
такой план строения тела обусловлива-
ется высококонсервативными генами
Нох, крайне широко распространенны-
ми в царстве животных. Существуют,
однако, и другие планы строения тела,
например у ракообразных и моллю-
сков. Mb — средний мозг; Fb — пе-
редний мозг. (Источник: Chambers and
McGonnell, 2002.)
Рис. 16.12. Гены Нох и анатомия мозга. Гены Нох отвечают не только за общий план
строения тела, но и за развитие некоторых участков мозга. Обратите внимание на трех-
мерный атлас мозга (а) и дифференциально окрашеный атлас участков головного мозга
мыши (б). Линейка внизу (в) отражает уровень экспрессии Нох и некоторых других ге-
нов в мозге мыши. OB — обонятельная луковица, Ctx — кора, Bnst — ствол мозга, Hi —
гиппокамп, Hy — гипоталамус, IC — нижний холмик четверохолмия, Cb — мозжечок,
Pons — часть мозга, Med — продолговатый мозг, Str — стриатум, Sp Crd — спинной
мозг. (Источник: Zapala et al., 2005. Из Zapala, Вarlow and Hovattaa, 2009.)
Гены и молекулы мышления
279
Активность генов Нох проявляется через контроль факторов транскрип-
ции — белков, регулирующих переписывание информации ДНК в РНК-матрицы.
Это один из способов регуляции экспрессии гена. Подобная регуляция может
осуществляться на многих стадиях — от ДНК до белка.
Чтобы сформировались характерные для человека полушария переднего
мозга, необходимо, чтобы в геноме была информация о том, где и на какой ста-
дии развития требуется повышенное количество нейронов. Так водопроводчик
должен знать план всего дома чтобы решить, где именно разместить тот или
иной вентиль. Рост коры головного мозга также следует рассматривать в кон-
тексте роста всего тела. В этом случае можно сказать, что план тела по отноше-
нию к плану мозга является планом более высокого порядка.
Ошибки в регуляторных генах могут давать самые серьезные последствия.
Некоторые болезни мозга возникают по причине нарушения развития, ошибок
в контроле временной генетической регуляции роста нейронов. В то же вре-
мя гены, контролирующие развитие, могут быть мишенями для лекарственных
препаратов. Ниже мы увидим примеры как первого, так и второго.
3.3 . Ошибки при развитии мозга
Для понимания развития неокортекса можно изучать не только те гены, которые
контролируют этот процесс, но и обусловливающие ошибки в нем. На сегод-
няшний день известно около шести генов, влияющих на размер неокортекса,
но, по-видимому, этот перечень неполный. Ген под названием microcephalin
был открыт у семей, подверженных микроцефалии — болезни, при которой ра-
стущий мозг замещается ликвором. До некоторой степени сходные врожденные
заболевания носят названия анэнцефалии и гидроцефалии. Наличие выживших
после даже столь тяжелых нарушений развития людей говорит о крайне высо-
кой адаптивной способности мозга.
Наиболее ярким примером вышесказанного может служить состояние, на-
зываемое гидроцефалией. Младенцы с таким расстройством совсем не имеют
неокортекса (рис. 16.13). На сегодняшний день считается, что при этом забо-
левании неокортекс частично развивается in utero, но затем по непонятным
причинам еще до рождения рассасывается. Молодая кора, возможно, способ-
на передавать часть своих свойств окружающим тканям перед исчезновением.
И хотя большая часть младенцев с этим заболеванием погибает в первый год
жизни, некоторые из них живут дольше и демонстрируют любопытные пове-
денческие свойства (рис. 16.14).
Дети-гидроцефалы могут выглядеть абсолютно нормальными в первые не-
сколько месяцев жизни, привязывая к себе родных. Позже, когда симптомы ста-
новятся явными, некоторым семьям бывает трудно свыкнуться с мыслью, что
ребенок для них потерян. Родители таких детей формируют группы поддержки,
часто размещают информацию в Интернете.
3.4 . Гены, контролирующие развитие
Один из способов быстрого эволюционного ответа на изменившиеся условия
среды является сдвиг стадий развития. Так, антропологи установили, что по
сравнению со скелетом неандертальца скелет современного человека более
грацилен — кости в нем более тонкие, а сам он дает меньшую защиту вну-
тренним органам. Одним из способов превращения мощного тела в грацильное
280
Глава 16
служит продление детского и подросткового периодов. Такая «ювенилизация»
происходит довольно часто. К примеру, кошки и собаки были одомашнены не в
последнюю очередь при помощи селективной ювенилизации, поскольку дете-
ныши млекопитающих легче привыкают к животным, заменяющим мать, чем
взрослые. С точки зрения наших домашних животных, мы сами приходимся им
матерями и вожаками стай.
Ювенилизация — еще один пример регуляторных изменений в ДНК — вы -
сокоуровневого плана, модифицирующего активность тех или иных участков
ДНК. Регуляторные гены могут быть отличными мишенями для лекарств, по-
скольку имеют столь обширное влияние на организм. Сдвиг стадий развития
позволяет единомоментно изменить огромный набор характеристик. Изучение
взаимодействия эволюции и развития на сегодняшний день является интенсив-
но разрабатываемой областью, иногда называемой «evo-devo».
Одним из генов развития является упомянутый уже ген FOXP2. У ряда ви-
дов он контролирует развитие легких и голосовой мускулатуры. ДНК, извлечен-
ная из ископаемых останков неандертальцев, несла тот же самый FOXP2, что и
ДНК современных людей. Тем не менее неандертальцы были менее способны к
Рис. 16.13. Отсутствие коры у ребенка-гидроцефала. Результаты магнитно-резонанс-
ного сканирования ребенка с гидроцефалией. Темная верхняя половина черепа на сним-
ках заполнена жидкостью. Мозговой ствол и мозжечок, по всей видимости, остаются
неповрежденными. Эти части мозга ответственны за основные реакции засыпания и
подъема, и дети с заболеванием могут иметь практически нормальные циркадные рит-
мы некоторое время после рождения. Неожиданным является тот факт, что симптомы
болезни могут проявляться спустя несколько месяцев после рождения. Причины забо-
левания на настоящий момент не установлены. (Источник: Merker, 2007.)
Гены и молекулы мышления
281
абстрактному мышлению и языку, чем современные люди. Так что история гена
FOXP2 все еще далека от завершения.
3.5 . Программирование генов:
за пределами центральной догмы
Если вы, готовясь к экзамену, не поспите одну ночь, то тем самым вы окажете
некоторое не слишком большое влияние на генетическую программу, управля-
ющую вашим циклом сна и бодрствования. Если сделать так несколько раз, то
влияние будет сильнее и долговременнее. Экспрессия генов в клетках челове-
ка может меняться в некоторых пределах путем включения или выключения
участков ДНК (рис. 16.15 и 16.16). Такой эпигенетический контроь влияет на
поток информации, описанный в центральной догме.
Открытие эпигенетических программ показало, что причинно-следственные
сети в мозге могут быть куда сложнее, чем ученые привыкли думать. К при-
меру, на внутреннюю среду плода во время беременности оказывает влияние
здоровье матери, прием ею алкоголя и лекарств, а также уровень стресса. Даже
качество материнской заботы может оказывать влияние на эпигенетическую
регуляцию. Существует эпигенетическое нарушение, называемое алкогольным
синдромом плода, при котором происходит отставание и нарушения в развитии
мозга плода; вызвано оно приемом матерью алкоголя в период беременности.
Разумеется, не все нарушения такого рода вызываются чрезмерным присут-
ствием того или иного вещества. Токсины также оказывают эпигенетическое
влияние на человека в течение его жизни. Некоторые ученые полагают, что ау-
тизм может быть эпигенетическим заболеванием, а его причиной являются не
гены или окружающая среда сами по себе, а неизвестное взаимодействие меж-
ду ними.
Таким образом, молекулярная генетика теперь рассматривает экспрессию
генов не как цепь, а как сеть, в которую вовлечены гены, их продукты и факто-
ры внешней среды.
Рис. 16.14. Некоторые дети могут выглядеть нормальными и без коры головного моз-
га. Дети-гидроцефалы (девочка справа) во многом похожи по поведению на своих нор-
мальных сверстников, чем привязывают к себе родных. (Источник: Merker, 2007.)
282
Глава 16
3.6 . Среда может перестраивать хроматин в ядре клетки
Хромосомы состоят из длинных цепей ДНК, скрученных в плотные клубки
в ядре клетки и окруженных огромным количеством различных молекул. Че-
ловек имеет 23 пары хромосом и приблизительно 30 000 генов. ДНК закручена
вокруг гистоновых октамеров — белков, благодаря которым хромосома имеет
компактную структуру; при этом образуется так называемый хроматин. (На са-
мом деле белков, участвующих в компактизации ДНК, значительно больше. —
Прим. перев.)
Кокаин и многие другие вещества способны изменять конфигурации этих
белков. В зависимости от типа изменения хроматин может разрежаться и уплот-
няться, активируя или заглушая активность генов намотанной на него ДНК.
Реструктурирование хроматина, по всей видимости, лежит в основе таких адап-
тивных процессов, как обучение и память, а также в основе заболеваний, таких
Рис. 16 .15. Эпигенетика: изменение экс-
прессии генов под влиянием окружаю-
щей среды. Открытие альтернативных
описанным в центральной догме путей
передачи информации иногда называ-
ют третьей биологической революцией.
Многие внешние факторы могут вы-
зывать сайленсинг (молчание) участков
ДНК или активировать их, либо изме-
нять соотношение между ДНК, РНК и
белком. Транскрипционная регуляция
происходит на уровне транскрипции
ДНК в РНК. (Источник: Baars.)
Рис. 16 .16 . Эпигенетические факто-
ры могут включать и выключать гены.
Участки ДНК могут включаться и вы-
ключаться эпигенетически. Октамеры
гистонов (зеленые) служат катушками,
на которые наматывается нить ДНК, тем
самым компактизируясь из нити длиной
почти 2 м в хромосомы, длина которых
не превышает сотен микрометров. Для
активации или деактивации участков
хромосомы ДНК наматывается или отде-
ляется от гистонов. (Источник: Nacional
Institute of Medical Sciences, 2009.)
Гены и молекулы мышления
283
как рак, судороги, шизофрения и депрессии. Некоторые медикаменты — напри-
мер, антидепрессанты — также способны, по всей видимости, изменять конфи-
гурацию хроматина наряду с другими эффектами.
У грызунов прикосновение и вылизывание матерью детенышей понижают
конденсированность ДНК в нейронах, позволяя РНК-полимеразе инициировать
транскрипцию с деконденсированных областей. Пониженное внимание матери
ведет к молчанию ДНК, поскольку она остается плотно намотанной на гистоны.
Вещества, обусловливающие такое влияние, — трихостатин А и метионин, —
доноры метильных групп. На рис. 16.17 зеленые шестиугольники с буквой А
обозначают ацетилированные N-концы гистонов, наличие которых вызывает
деконденсацию ДНК. Справа на рисунке надписью «РНКп» обозначена РНК-
полимераза, транскрибирующая ДНК в пре-мРНК.
3.7. Обучение как эпигенетический процесс
Как мы увидим, эпигенетика в настоящий момент становится все более актуаль-
ной при изучении проблем обучения. Нейромедиаторы изменяют экспрессию
генов в клетках, из которых происходят, равно как и в клетках, на которые воз-
действуют. Поскольку ДНК работает как программа для производства молекул,
включая и нейромедиаторы, смысл в этом есть как для пресинаптической клет-
ки, так и для клетки постсинаптической, и для окружающих глиальных клеток.
Эти явления подробнее рассмотрены в разделе «Обучение» настоящей главы.
4.0 . Нейроны и глиальные клетки
как обработчики сигналов
Все живые клетки имеют некоторые общие фундаментальные свойства и струк-
туры: ядро, содержащее генетическую информацию, окисление глюкозы и из-
бавление от токсинов, двухслойную липидную мембрану. Все клетки обмени-
ваются информацией при помощи сигнальных молекул, проходящих сквозь
мембрану посредством различных механизмов транспорта. В этих отношениях
Рис. 16 .17. Материнская забота может модифицировать экспрессию ДНК. Вылизыва-
ние детенышей грызунов их матерями может активировать или подавлять экспрессию
генов. (Источник: Champagne and Curley, 2009.)
284
Глава 16
нейроны подобны любой другой животной клетке, однако они также специали-
зированы в электрохимическом проведении и обработке сигнала.
Для выполнения задач проведения и обработки нейроны образуют множество
отростков — самое большое среди всех клеток животных — и функционируют
одновременно и как передатчики, и как переключатели, а возможно, что даже как
небольшие вычислительные устройства. Нейроны отличаются возможностью
быстро проводить электрохимический сигнал по своему аксону, преобразуя
мембранный потенциал в потенциал действия (ПД) (см. гл . 1 и 3). Также важно
помнить, что нейроны соединены между собой как при помощи классических
синапсов, описанных десятки лет назад, так и неклассическими способами.
Не так давно было установлено, что глиальные клетки, ранее считавшиеся
поддерживающими элементами нервной ткани, играют важную роль в проведении
сигналов. Глиальные клетки составляют примерно половину массы мозга, что
свидетельствует о важности выполняемых ими функций. Традиционно считалось,
что при проведении сигнала глиальным клеткам отводились «эпизодические роли»,
однако в некоторых случаях они способны сыграть и главную. Глия проводит свои
сигналы путем распространения волн кальция, движущихся медленнее, чем ПД по
аксонам нейронов. При этом нейроглиальные клетки взаимодействуют с нейронами
при помощи нейромедиаторов и являются единственным местом в мозге, где
производятся ГАМК и глутамат.
4.1. Нейроны и синапсы в качестве переключателей
Нейрон можно рассматривать как переключатель — биологическую версию
электрического переключателя. Потенциал действия по аксону либо идет, либо
нет. Точно так же действуют и синапсы, передавая химический сигнал или не
передавая его. Входные ответвления нейрона — дендриты — можно таким об-
разом сравнить с интегратором, суммирующим импульсы десятков тысяч вхо-
дящих сигналов за очень короткий период времени. Этот механизм не может не
наводить на мысли об аналоговом компьютере. Таким образом, поток информа-
ции в каждом нейроне и на каждом синапсе может быть остановлен, модифи-
цирован или усилен. И хотя мозг отнюдь не идентичен компьютеру, он похож на
него в плане анализа, фильтрации, хранения, и контроля информации многими
его элементами. Поскольку мозг содержит примерно сто миллиардов нейронов
и около триллиона синапсов, правильнее рассматривать его в качестве большой
сети переработчиков информации, до некоторой степени напоминающей Ин-
тернет.
Полезно рассмотреть мозг и в качестве огромного адаптивного органа. В не-
котором смысле он подобен биологическим видам, так как в нем постоянно
происходит появление, отбор и вымирание синапсов. Но процессы, происходя-
щие в мозге, длятся секунды, часы или дни, а не сотни тысяч лет, как в случае
видов.
Обратите внимание, что все процессы на рис. 16.18 — саморегулируемые,
что позволяет поддерживать концентрацию биомолекул в требуемых пределах.
Ауторецепторы контролируют синтез и высвобождение медиатора, контроли-
руя таким образом частоту прохождения импульсов по нейрону. Транспортные
молекулы возвращают молекулы медиатора в клетку, выбросившую их, для
повторного использования. Везикулярные транспортеры переносят молекулы
медиатора в везикулы — небольшие мембранные пузырьки, из которых они
Гены и молекулы мышления
285
высвобождаются в синаптическую щель. Некоторые ферменты расщепляют
медиаторы, инактивируя их, а в клетку транспортируются их части. Обрати-
те внимание, что мембранные рецепторы выглядят как тонкие нити, много раз
пронизывающие мембрану насквозь. Это позволяет не забывать, что мембран-
ные рецепторы имеют белковую природу и могут быть классифицированы по
количеству пересечений мембраны полипептидной цепью.
Основные вопросы синаптической передачи — как медиаторы синтезиру-
ются, доставляются к синапсу, высвобождаются и вызывают ответ постсинап-
тической клетки. Затем постсинаптическая клетка должна превратить молеку-
лярный сигнал в потенциал действия (рис. 16.19).
4.2 . Химическая саморегуляция
Химические реакции в мозге жестко регулируются. Если синапс выделяет
слишком мало медиатора, сигнал не будет передан. Избыток медиатора токси-
чен и убивает нейроны. По этой причине во всех метаболических путях име-
ются петли отрицательной обратной связи, которые не дают количеству тех или
Рис. 16 .18 . Генерализованный синапс. Два нейрона разделены синаптической щелью,
в которую один из них высвобождает нейромедиатор, диффундирующий к мембране
постсинаптической клетки. В соответствии с этим один из нейронов называют преси-
наптическим, а другой — постсинаптическим. Глиальная клетка выполняет второсте-
пенные функции, однако на настоящий момент известно, что она также может пере-
рабатывать информацию. Поэтому при рассмотрении синапсов следует принимать во
внимание три клетки — пресинаптическую, постсинаптическую и глиальную. (Источ-
ник: Byrne and Roberts, 2004.)
286
Глава 16
иных молекул подняться выше определенного предела. Непосредственно в си-
напсе работает несколько механизмов для удаления лишних молекул медиато-
ра — это и снижение скорости его высвобождения, и их пассивная диффузия за
пределы синаптической щели, и транспорт в клетку происхождения, и химиче-
ская их деактивация, и переработка, и удаление глиальными клетками. Очистка
синапса жизненно важна. Синапсы со слишком большим количеством медиа-
тора могут перестать работать, а нейроны, образовавшие их, — погибнуть. Хо-
рошим примером служит оксид азота, который работает в качестве медиатора
во многих синапсах (в частности, именно это вещество опосредует активность
препарата «Виагра»), но при этом становится ядовитым, если не удаляется с
достаточно высокой скоростью. Это вещество диффундирует за пределы си-
наптической щели в течение нескольких миллисекунд, если вброшенное его
количество было не слишком большим.
Примером химической саморегуляции в мозге являются ауторецепторы.
Нейронам необходимо контролировать их секрецию — поскольку при ее отсут-
ствии они могут отравить сами себя, а также потому, что при слишком большом
количестве медиатора сигнал, идущий через синапс, никогда не прекратится.
Ели вы слышите одну и ту же фразу сотни раз, она теряет информативность. То
же самое справедливо и для мозга. Сигнал информативен только тогда, когда он
посылается через разумные промежутки времени. По этим причинам каждый
нейрон должен регулировать секрецию нейромедиаторов.
4.3 . Мембраны, ионные каналы и аксоновые спайки
В настоящем разделе мы вернемся к вопросам, рассмотренным нами в гл. 1 и 3.
Все живые клетки работают как крошечные батарейки, поддерживая на
своих мембранах разность потенциалов от –70 до –100 мВ (рис. 16.20). Это
возможно благодаря активному трансмембранному транспорту ионов против
Рис. 16 .19 . Нексусы, или щелевидные контакты, образуют прямые соединения между
клетками. Они делают возможным прямое электрическое сообщение между клетками.
Большая часть ГАМК-ергических контактов в коре — щелевидные, что повышает син-
хронность ответа многочисленных кортикальных нейронов. (Источник: Cunningham
and Le Beau, из Squire, 2009.)
Гены и молекулы мышления
287
градиентов их концентраций, закачивающих отрицательные ионы внутрь клет-
ки и выкачивающих положительно заряженные наружу*
. Для поддержания ра-
боты этих насосов, а также для большинства других функций клетки требуется
постоянный приток макроэргических метаболитов — из-за этого нейроны по-
стоянно потребляют энергию. Именно поэтому на поддержание работы мозга
уходит столько ресурсов.
В отличие от большинства других клеток нейроны способны превращать
свой трансмембранный потенциал в электрохимический сигнал, как показано
на рис. 16.21. Дендриты нейрона принимают сигнал от соседних клеток, кото-
рый меняет потенциал мембраны. Когда разность потенциалов в области ак-
сонного бугорка изменяется примерно на 70 мВ, аксон дает спайк — быструю
волну деполяризации мембраны. Ионы по разные стороны мембраны устрем-
ляются сквозь нее, поскольку она на короткое время становится для них про-
* Потенциал покоя складывается как из переноса зарядов в результате пассивного движения ионов по
их концентрационным градиентам, так и из переноса зарядов в результате активного транспорта.
Электрически асимметричный активный транспорт поддерживает внутриклеточные концентрации
Na+ и K+ и переносит эти ионы через мембрану в соотношении 3/2. Во многих клетках существуют
транспортные системы для ионов Cl– против его концентрационного градиента. — Прим. перев.
Рис. 16 .20 . Основы нейро-
нальной сигнализации. Все
клетки поддерживают транс-
мембранную разность потен-
циалов, однако только нейро-
ны способны с ее помощью
давать аксональные спайки.
Из рисунка видно, как дисба-
ланс между положительными
ионами вне клетки и отрица-
тельными ионами в клетке
дает мембранный потенци-
ал, который поддерживается
постоянным перекачиванием
ионов. При прохождении вол-
ны деполяризации мембран-
ный потенциал обращается,
но немедленно после этого
он восстанавливается. Для
его восстановления нейрон
должен непрерывно потре-
блять энергию. (Источник:
Shepherd, 2003 в Squire, по
Spencer and Kandel, 196.)
288
Глава 16
ницаема. В конце концов волна электрохимической активности достигает си-
напса. Мембранный потенциал аксона при этом локально повышается с –70 мВ
до + 30 мВ, однако он возвращается к исходному значению за несколько милли-
секунд. Результатом такой активности является волна деполяризации, идущая
от тела клетки к синапсу.
Эта система способна включаться и выключаться, работая подобно электри-
ческому триггеру, а возможно, и как более сложный элемент. Для того чтобы
возбудить нейрон, требуется деполяризовать его мембрану. Это можно сделать,
приложив внешний ток к клетке для обращения полярности ее мембраны, —
примерно то же самое происходит при переключении полярности батарейки.
Также можно ввести нейромедиатор в непосредственной близости от синапса,
чтобы заставить постсинаптический нейрон деполяризоваться — эффект до-
стигается абсолютно такой же. Деполяризация мембраны индуцирует возник-
новение аксонного спайка, который, в свою очередь, опосредует высвобожде-
ние нейроном медиатора в синаптическую щель.
Выше описана классическая модель возбуждающего нейрона. Есть возмож-
ность ингибировать постсинаптический нейрон, гиперполяризуя его мембрану.
При этом разность потенциалов на мембране опускается ниже –70 мВ. Гипер-
поляризованной мембране требуется больше энергии, чтобы деполяризоваться.
В результате возможность прохождения спайка по аксону резко снижается.
Рис. 16 .21 . Ток ионов через мем-
брану клетки. Мембрана представ-
ляет собой бислой липидов, которые
упакованы подобно двум слоям спи-
чек с направленными наружу голов-
ками. Эти головки, обозначающие
головки липидов, гидрофильны,
а хвосты — гидрофобны. Из-за та-
кого распределения гидрофильных
и гидрофобных участков молекулы
фосфолипидов спонтанно образуют
липидные бислои, в которых гидро-
фильные головки обращены в вод-
ную среду, а гидрофобные хвосты —
друг к другу. Мембрана клетки в
первом приближении представляет
собой пузырь, образованный таким
бислоем, который отделяет внутрен-
нее пространство клетки от внешней
среды слоем толщиной в две моле-
кулы. (Источник: McCormick, по
Byrne and Roberts, 2009.)
Гены и молекулы мышления
289
Проще описать возбуждение и торможение в терминах спонтанного возбуж-
дения нейрона. В нормальных условиях нейроны дают спайки с некоторой по-
стоянной частотой, даже если их не стимулировать. Гипотетической частотой
можно выбрать один спайк в секунду. При стимуляции нейрона возбуждающим
медиатором частота возрастает. Однако при торможении медиатором тормоз-
ным эта частота может снижаться практически до нуля. Как уже отмечалось
ранее, самым распространенным тормозным медиатором является ГАМК, а са-
мым распространенным возбуждающим — глутамат.
Липидный бислой образуется спонтанно при суспендировании липи-
да в воде. Один из концов молекулы фосфолипида имеет высокое сродство к
воде — гидрофилен, тогда как другой — низкое (гидрофобен). Из-за этого мо-
лекулы фосфолипидов выставляют свои гидрофильные концы воду, а гидро-
фобные — прячут, сближая их друг с другом. Этим и обусловлено спонтанное
образование липидных бислоев в воде и принятие ими шарообразной формы.
Липидный бислой одновременно достаточно подвижен и весьма прочен и име-
ется во всех без исключения живых клетках. В бислой встроены многочислен-
ные белки, такие как ионные насосы, рецепторы и ферменты, а также многие
другие структуры, обеспечивающие сотни различных процессов.
Все мембранные каналы, упоминавшиеся при работе нейрона, относятся
к классу потенциалзависимых. Как мы уже упоминали, на мембране сохраняет-
ся постоянная разность потенциалов. Для ее поддержания требуется непрерыв-
но расходовать энергию — в частности, на работу трансмембранных ионных
насосов. Разность потенциалов на мембране снимается всякий раз, когда нейрон
стимулируют электрически или когда происходит деполяризация. Эти события
вызывают локальный обмен ионами на мембране в ту или другую сторону.
На рис. 16.22 подробно представлен один из потенциалзависимых ионных
каналов. Каждый тип канала имеет характерное для него число пересечений
мембраны полипептидной цепочкой. Такие пересечения носят название транс-
мембранных сегментов и формируют субъединицы. Это сравнительно простое
описание позволяет достаточно точно классифицировать очень большую часть
трансмембранных белков, среди которых — ионные каналы и рецепторы.
Рис. 16.22 . Потенциалзависимый ионный канал. Здесь показана схема укладки бел-
ковой цепи потенциалзависимого натриевого канала, проходящей через мембрану на-
сквозь и образующей при этом шесть трансмембранных сегментов, каждый из которых
повторен четыре раза (DI-DIV). В его состав, помимо большой α-субъединицы, входит
также небольшая β-субъединица. (Источник: Squire, 2009 [Encyclopedia of NS].)
290
Глава 16
5.0 . Синаптическая передача:
от производства до расчистки
Информационные молекулы нервной системы можно разделить на два клас-
са — медиаторы и модуляторы. При этом медиаторы служат для локальной
передачи сигнала в синапсах, хотя таких синапсов может насчитываться де-
сятки миллиардов. Нейромодуляторы секретируются нейронами коры и рас-
пространяются в широкой области. Клетки, секретирующие модуляторы, раз-
ветвляются на весьма обширные участки мозга. На концах их ответвлений и
секретируются эти молекулы. Таким образом, модуляторы влияют на работу
обширных участков мозга, тогда как медиаторы действуют на уровне синапсов
(см. разд. 6 .0).
5.1. «Большая двойка»: глутамат и ГАМК
Глутамат и ГАМК являются двумя наиболее распространенными нейромедиа-
торами в мозге. Девяносто процентов нейронов коры используют глутамат —
основной возбуждающий медиатор, увеличивающий вероятность развития
аксонного ПД на постсинаптическом нейроне при высвобождении в синаптиче-
скую щель. В человеческом мозге глутамат чаще всего используется крупными
пирамидными нейронами коры и более глубоких структур мозга. Также этот
медиатор нередко используется в модифицируемых синапсах, обусловливая
обучение.
ГАМК, в противоположность глутамату, является основным тормозным ме-
диатором коры головного мозга. Ингибирующие синапсы снижают вероятность
прохождения ПД по аксону постсинаптического нейрона. ГАМК распростране-
на во вставочных нейронах, окружающих пирамидальные клетки, — полагают,
что в этом случае она служит для регуляции непрерывной возбуждающей ак-
тивности коры.
Для функционирования мозга не требуется постоянной активности всех воз-
буждающих синапсов. В этом случае в мозге образовались бы петли положи-
тельной обратной связи, усиливающиеся с каждым циклом. Кора будет пере-
гружена, как в случае эпилептических припадков. Избыток глутамата токсичен
и ведет к явлению, называемому эксайтотоксичностью. Большая часть ущерба
от припадков происходит не непосредственно от них, а от избыточного высо-
бождения глутамата. Это похоже на взрыв бака с горючим в горящем автомоби-
ле: взрыв наносит гораздо больше ущерба, чем пламя, его вызвавшее. Нейроме-
диаторы полезны только в строго определенных количествах.
Глутамат (Glu) также отлично подходит для рассмотрения процесса образо-
вания нейрмедиаторов из существовавших молекул. Глутамин является одной
из аминокислот, которые организм животных получает с пищей. Мозг, в свою
очередь, использует глутамин для передачи возбуждающего сигнала.
Мы можем чувствовать вкус глутамата в пище, как было установлено япон-
скими учеными в 1907 г. при исследовании соевого соуса. Вкус глутамата —
пятый базовый вкус, помимо четырех основных, к которому у нас имеются
отдельные рецепторы; он носит название умами. Вкус глутамата помогает
определить съедобность и свежесть пищи — особенность, крайне необходимая
для охотников и собирателей первобытного мира.
Мы можем рассмотреть показанную на рис. 16.23 систему из трех клеток
как линию производства определенных количеств нейромедиатора глутамата,
Гены и молекулы мышления
291
транспортировки их в синапс с использованием везикулярного транспорта и
выброса их в синаптическую щель. Небольшая овальная органелла в верху
клетки — митохондрия, производящая большую часть клеточного АТФ. Вся эта
система питается глюкозой и кислородом, диффундирующими через мембраны
из капилляра справа. Глюкоза используется для получения энергии, а также для
синтеза нейромедиатора глутамата.
Заметьте, что постсинаптическая клетка имеет два типа рецепторов глутама-
та, которые мы обсудим позже. Метаботропные рецепторы используют для от-
вета метаболических путей клетки. Ионотропные рецепторы активируют ион-
ные каналы, о которых мы уже говорили: натриевые, калиевые и кальциевые.
Астроцит в середине схемы также важен для работы всей системы. Он за-
хватывает глюкозу, разлагает ее и превращает АДФ в АТФ в своих митохондри-
ях, направляет глутамин в пресинаптическую клетку, где из него синтезируется
глутамат, и захватывает избыточный глутамат, диффундирующий из синап-
тической щели. Последнее очень важно, поскольку глутамат, при длительном
нахождении вне клетки, токсичен. Токсичность глутамата, как полагают, ста-
новится причиной серъезных заблеваний мозга. (Такие расстройства также на-
зывают эксайтотоксичностью, поскольку глутамат является основным возбуж-
дающим медиатором мозга.)
Рис. 16.23 . Глутаматергическая сигнализация требует участия трех клеток. Три клетки
работают вместе для обеспечения работы глутаматергической сигнализации. Обратите
внимание на кровеносный капилляр, обеспечивающий астроцит и нейроны глюкозой и
кислородом. Глюкоза также является одним из промежуточных метаболитов при синте-
зе глутамата. V
m
—
мембранный потенциал верхнего нейрона, на котором показаны не-
сколько спайков, обусловливающих высвобождение медиатора в синаптическую щель,
PGK — фосфоглицераткиназа. (Источник: Magistretti, из Square et al., 2008.)
292
Глава 16
И хотя для доставки глутамата к месту его действия требуется немало ухищ-
рений, сигнальная система с его применением является одной из наиболее рас-
пространенных и древних. Первые млекопитающие, жившие 200 млн лет на-
зад, имели такой же механизм. Он же функционирует даже у круглого червя
С. elegans. Глутаматергическая сигнализация крайне точна по времени, ее ней-
ромедиатор может быстро удаляться из внеклеточного пространства; она так-
же не оставляет токсичных соединений во внеклеточной среде. В то же время
практически все биохимические процессы, особенно окислительные, произво-
дят некоторое количество токсичных веществ и могут быть очень вредными
при длительном функционировании.
5.2 . ГАМК: основной тормозный медиатор
Возбуждающие нейроны необходимы для работы мозга, однако если бы в нерв-
ной системе были только они, то в мозге быстро произошла бы перегрузка, по-
скольку каждый глутаматергический нейрон возбуждал бы остальные. Тормоз-
ные нейроны балансируют возбуждающие для поддержания требуемого уровня
активности. Возбуждающие и тормозные нейроны взаимодействуют, создавая
регулярные ритмы — наиболее важную часть межрегиональной сигнализации
в мозге.
Глутаматный метаболический путь производит также основной тормозный
нейромедиатор — ГАМК (гамма-аминомасляную кислоту) (рис. 16.24). Если
Рис. 16 .24 . ГАМК-ергический синапс. Обратите внимание, что для производства ГАМК
используется глутамин и его производные. Транспортер — это особая белковая молеку-
ла, транспортирующая молекулы медиатора обратно в клетку для повторного исполь-
зования. Ауторецепторы принимают участие в саморегуляции синапса. GAT— пере-
носчик ГАМК, GAD — глутаматдекарбоксилаза, VIAAT — везикулярный транспортер
тормозных медиаторов, Gln — глутамин, Glu — глутаминовая кислота. (Источник:
Deutch and Roth, из Squire et al., 2008.)
Гены и молекулы мышления
293
в синаптическую щель высвобождается ГАМК, то вероятность прохождения
ПД по аксону постсинаптического нейрона снижается. Эффект ГАМК заклю-
чается в гиперполяризации, а не деполяризации мембраны, и снижении веро-
ятности прохождения ПД даже в случае одновременного получения клеткой
возбуждающего сигнала. Таким образом достигается баланс между возбужда-
ющими и тормозными сигналами, приходящими к клетке с интервалами при-
мерно в 10 мс, который и обусловливает ее текущую активность (см. гл . 3).
Пирамидные нейроны коры обладают по большей части возбуждающей ак-
тивностью, тогда как олигодендроциты — тормозной (см. рис. 16.25). Таким
образом, волны возбуждения, непрерывно проходящие по всему мозгу, локаль-
но ингибируются ГАМК-секретирующими клетками. Такие клетки называют
ГАМК-ергическими, тогда как возбуждающие носят название глутаматергиче-
ских.
Ингибирование необходимо для регуляции возбуждающих нейронов и соз-
дания барьеров на пути прохождения волны возбуждения. Для предотвращения
перегрузки мозга возбуждением существует множество ГАМК-ергичеких си-
напсов, регулирующих уровни возбуждения.
ГАМК используется в подавляющем большинстве быстрых тормозных си-
напсов практически в любой части мозга. Большинство транквилизаторов уси-
ливают ГАМК-ергическую активность мозга. Расслабляющий эффект спиртно-
го также основан на его влиянии на ГАМК-опосредованные процессы. Однако
ГАМК имеет, помимо торможения, еще и другие функции.
Рис. 16 .25. Все контуры в коре включают как возбуждающие, так и тормозные нейроны.
Пирамидные клетки на этой схеме выделены голубым, а тормозные интернейроны — крас-
ным. Зеленым помечен вход из-за пределов системы (например, из зрительного тракта), воз-
буждающий кортикальные нейроны. «Гладкие» нейроны, обозначенные на схеме S1, также
являются возбуждающими. Микроконтуры такого типа распространены в коре и определяют
различия между одинаковыми с анатомической точки зрения областями мозга. (Источник:
Douglas and Martin, 2009.)
294
Глава 16
С некоторым упрощением мы можем свести описанное к нескольким основ-
ным принципам. Гены подвергаются различным процессам как на протяжении
миллионов лет, так и на протяжении человеческой жизни, дней, часов и даже
минут. Биохимические пути клеток используют достаточно ограниченный на-
бор веществ, отобранный и сохраняемый эволюцией. Человек по многим био-
химическим характеристикам сходен с широким рядом животных, но имеет
тем не менее определенные отличия.
Как мы увидим, эти различия обусловлены в большинстве своем активно-
стью высокоуровневой регуляторной ДНК, а не ДНК, непосредственно экс-
прессирующейся в белки. Мы не отличаемся от большинства животных по
химическому составу. Отличия заключаются главным образом в более высоко-
уровневой организации генома, опосредующей рост и функционирование об-
ширного неокортекса, и особенно переднего мозга. Человек — новая мелодия,
сложенная из нот жизни.
5.3 . Нейроглия также может обрабатывать информацию
Нейроны — основные клетки, проводящие и обрабатывающие сигналы в го-
ловном мозге, но не единственные. Мы уже упоминали тесную взаимосвязь
нейронов с глиальными клетками. Существует много разных типов нейронов
и глии, но мы для простоты будем рассматривать их как два единых класса об-
рабатывающих сигналы клеток.
В настоящее время все еще ведутся дискуссии относительно роли глиальных
клеток в обработке информации. Нейромедиаторы, высвобождаемые нейрона-
Рис. 16 .26 . Глиальные клетки используют кальциевые волны для передачи сигна-
ла. Глиальные клетки могут передавать кальциевый сигнал путем пассивной диффу-
зии. Эти клетки не требуют синапсов для передачи возбуждения. (Источник: Haas and
Kettenmann, из Squire, 2009.)
Гены и молекулы мышления
295
ми, вызывают в глиальных клетах — в особенности астроцитах — так назы-
ваемые кальциевые волны. Астроциты в пределах определенных рецепторных
полей способны также отвечать на световое раздражение, подобно клеткам
сетчатки. Они селективно восприимчивы к направленности и частоте стимула.
Таким образом, глиальные клетки способны вести себя подобно нейронам. Од-
нако они не идентичны нейронам, что вызывает споры относительно их роли
в основных когнитивных задачах, таких, как сенсорное восприятие и память.
Поскольку для передачи сигнала от одной глиальной клетки к другой не
требуется синапсов, полагают, что сигнал, передаваемый ими, распространя-
ется очень быстро. Это происходит благодаря наличию щелевых контактов. На
рис. 16.26 показано, как стимул (красная молния) вызывает высвобождение
АТФ глиальной клеткой. АТФ вызывает прохождение кальциевых волн, кото-
рые диффундируют через мембраны соседних клеток, распространяясь таким
образом дальше. Концентрации АТФ и глутамата во внеклеточном простран-
стве также могут увеличиваться, как показано на рисунке.
5.4 . Производство, высвобождение
и удаление нейромедиаторов
Каждая клетка — в некотором смысле микроскопическое химическое произ-
водство. Как и всякое производство, оно начинается с сырья, примером кото-
рого может служить триптофан в стакане молока. Триптофан превращается в
серотонин за несколько этапов, каждый из которых контролируется ДНК ядра
пресинаптического нейрона. После окончания синтеза вещества его транспор-
тируют к месту действия — синапсу — мембранные везикулы. Везикулы затем
сливаются с мембраной клетки, высвобождая свое содержимое в синаптиче-
скую щель.
Производство того или иного вещества идет в несколько определенных ста-
дий, называемых метаболической цепью или каскадом, каждый шаг которого
точно контролируется ферментами. На рис. 16.27 показан катехоламиновый ме-
таболический путь, который производит такие важные вещества, как дофамин,
норадреналин и адреналин. Регуляция количеств производимого вещества в та-
ком пути возможна путем регуляции активности соответствующих ферментов
или поступления исходных веществ, таких как L-ДОФА. Не стоит также за-
бывать, что каждый шаг каскада контролируется, поэтому добавление избыточ-
ных количеств исходных веществ не приведет к производству продуктов выше
определенной нормы. Так, если на автозавод поставить лишние десять тонн
стали, то эта сталь не будет использована. Ее необходимо хранить или утили-
зировать. Поскольку каждый шаг процесса контролируется, вмешательством
извне можно повлиять на него только в сильно ограниченных рамках. Однако
если создается недостаток исходных веществ, как в случае дефицита фолиевой
кислоты у беременных, такая ситуация может очень существенно повлиять на
количества производимого вещества.
5.5 . Высвобождение вещества в синапсе
На рис. 16.28 показано несколько последовательных стадий высвобождения
нейромедиатора в синаптическую щель. Молекулы медиатора свободно диф-
фундируют от пресинаптического нейрона и определяются рецепторами пост-
синаптического. В мозге имеются триллионы таких синапсов. Заметьте, что
296
Глава 16
величина перепада потенциала зависит от недавней активности. Если до этого
по клетке проходила волна возбуждения, некоторое количество медиатора было
выброшено в синаптическую щель, а его количество внутри клетки, таким об-
разом, понизилось — в результате в ответ на следующую волну возбуждения
будет выброшено только оставшееся количество медиатора. Таким образом,
производство и высвобождение веществ нейроном напоминают людей, пере-
дающих по цепочке ведра с водой: скорость передачи ведра следующему чело-
веку будет зависеть от того, насколько быстро он передаст дальше свое ведро.
В классической модели синапса предполагается, что везикулы (желтые)
транспортируют медиатор по аксону к синапсам, где сливаются с плазмалем-
мой и высвобождают свое содержимое в синаптическую щель. Некоторые ней-
роны, тем не менее, высвобождают нейромедиатор с использованием других
механизмов. Каждая везикула может содержать медиатор в количестве несколь-
ких тысяч молекул, что позволяет точно регулировать концентрацию высво-
бождаемого вещества как в пресинаптической клетке, так и непосредственно
в синаптической щели.
Рис. 16 .27. Катехоламиновый метаболический путь, служащий для производства мно-
гих нейромедиаторов. Катехоламиновый путь включает основные вещества, связанные
с нервной деятельностью, такие как дофамин, норадреналин и адреналин. В результа-
те один метаболический путь способен синтезировать несколько сигнальных веществ.
Одно из задействованных в нем веществ — L-ДОФА (дофамин) — используется в каче-
стве лекарства от болезни Паркинсона и увеличивает количество дофамина в мозге. Эти
вещества имеют сходное строение. (Источник: Baars, 2009.)
Гены и молекулы мышления
297
Везикулы несут кальцийсвязывающие белки, которые отрываются от по-
верхности при касании везикулой плазмалеммы синапса. Эти белки работают в
качестве сенсоров, отслеживающих момент слияния мембраны везикулы и пре-
синаптической мембраны. В силу этого кальций играет важную роль в точно
отрегулированном по времени и количеству процессе высвобождения нейроме-
диатора в синаптическую щель (рис. 16.29).
Рис. 16 .28 . Везикулы выбрасывают
свое содержимое в синаптическую
щель. Упрощенное изображение вы-
свобождения нейромедиатора из ве-
зикул, транспортирующих медиатор
от места синтеза, расположенного по-
близости от ядра. Везикулы движутся
по аксону, направляемые микротрубоч-
ками. (а) Высвобождение медиатора
привело к выбросу двух одинаковых
по размеру порций. (б) Первый вы-
брос привел к некоторому исчерпанию
запаса везикул в синапсе, поэтому во
второй раз порция выброшенного ве-
щества меньше. (Источник: Schwarz,
из Squire et al., 2003.)
Рис. 16 .29 . Кальций опосредует высвобождение нейромедиатора. Кальций имеет много
сигнальных функций. В мозге (и некоторых других органах, включая сердце), концен-
трация кальция регулируется ритмами потенциалов. В нейронах входящий ток кальция
может опосредовать высвобождение нейромедиатора на синапсах. (Источник: Snutch,
из Squire, 2009.)
298
Глава 16
5.6 . Синапсы и рецепторы в качестве контрольных точек
Психоактивные вещества чаще всего действуют на тот или иной, определенный
шаг синаптической сигнализации. Так, L-ДОФА, упоминавшийся нами ранее,
действует как исходное вещество на стадии синтеза нейромодулятора дофами-
на у больных с болезнью Паркинсона. При болезни Паркинсона происходит
отмирание дофаминергических нейронов черного вещества, L-ДОФА позво-
ляет отсрочить наступление наиболее тяжелых последствий этого процесса.
(Для устранения самой возможности появления нарушений отмершие клетки
следует заменять, и множество исследований сейчас направлено на разработку
метода трансплантации дофаминергических клеток в черное вещество.)
В других случаях действие препарата может быть направлено на продление
времени нахождения медиатора в синаптической щели. Одним из примеров та-
ких препаратов могут служить селективные ингибиторы обратного захвата се-
ротонина (SSRI, selective serotonin reuptake inhibitors) — психоактивные препа-
раты, эффективные при депрессиях и некоторых других расстройствах. Захват в
данном случае — не совсем корректный термин для процесса транспортировки
серотонина в клетку происхождения и его последующей реутилизации — если
этот процесс ингибирован, то серотонин может в течение более длительного
времени стимулировать постсинаптическую клетку.
ГАМК-ергические препараты, такие как бензодиазепины, увеличивают
количество ГАМК в тормозных синапсах головного мозга. Не следует также
недооценивать влияние здоровой диеты, поставляющей все основные предше-
ственники — вещества, из которых в клетках синтезируются белки и другие
необходимые соединения.
Одной из основных задач медицинских исследований является выявление
новых контрольных точек тела и мозга. Блокаторы каналов делают невозмож-
ным ток ионов, таких как натрий, калий и магний, через мембрану (широко
используются в кардиологии и при лечении расстройств мозга). Эти места кон-
троля следует держать в памяти. И хотя каждая из таких контрольных точек
крайне сложна, принципы, лежащие в основе, довольно просты.
Существует множество типов нейронов и нейроноподобных возбудимых
клеток. Нейроглия ранее считалась тканью, поддерживающей нейроны, но не-
давно было установлено, что ее клетки также способны проводить и обраба-
тывать сигнал. Клеток нейроглии в мозге в десять раз больше, чем нейронов.
(Греческое слово «глия» означает «клей» — глия ранее считалась своеобразным
клеем, склеивающим нейроны.)
5.7. Рецепторы распознают сигнальные молекулы
С известным упрощением молекулы рецепторов можно рассматривать как
замки, которые пытаются отпереть ключами — молекулами нейромедиаторов
(рис. 16.30). Метафору с ключом и замком любят использовать при описании
фермент-субстратных взаимодействий. На самом деле нейрональные рецеп-
торы являются только одним из классов рецепторов, которые имеются даже у
одноклеточных организмов, с полным отсутствием нервной системы. Клетка-
мишень нейромедиатора называется постсинаптической, а та, которая его вы-
бросила, — пресинаптической; это родственные термины. В результате синапс
рассмтривается как черный ящик, на входе которого находится пресинапти-
Гены и молекулы мышления
299
ческая часть, а на выходе — постсинаптическая. Если же избрать объектом вни-
мания нейрон, то входы и выходы нашего ящика заметно поменяются. Важно
помнить об этом различии, дабы избежать неправильного понимания информа-
ции относительно микроструктуры мозга.
Глутаматный рецептор был и остается одним из главных опытных объектов
при изучении передачи сигнала в нейронах. На рис. 16.31 показаны основные
принципы глутаматергической (возбуждающей) сигнализации. Обратите вни-
мание на тот факт, что глутамат имеет рецепторы двух типов, один из кото-
рых — ионотропный, а другой — метаботропный, благодаря которому глутамат
может оказывать влияние на метаболические пути клетки-мишени.
В первом случае мы можем рассматривать ионотропный рецептор как набор
каналов, перекачивающих ионы с одной стороны мембраны на другую, как и в
любом другом месте мембраны нейрона. Однако в нашем случае нас интересу-
ют только каналы на постсинаптической мембране. Ионные каналы — молеку-
лярные ворота, способные открываться и закрываться в ответ на присутствие
определенного вещества или на изменение мембранного потенциала.
Мы можем спуститься еще на один уровень ниже, и тогда мы увидим, что
ионотропный эффект глутамата обусловлен работой двух различных молеку-
лярных машин, одна из которых задействует N-метил-D -аспартат (NMDA),
а другая — α -амино-3 -гидрокси-5 -метил-4 -изоксазолпропионовой кислоты
(АМРА). Оба этих вещества необходимы для процесса обучения, и мы еще
встретим их в этой главе. Баланс между NMDA и АМРА определяет тип обу-
чения, который будет происходить на постсинаптическом глутаматергическом
элементе. NMDA отвечает за долговременное потенцирование (LTP, long-time
potentiation), делая синапс более возбудимым. АМРА отвечает за долговре-
менное подавление (LTD, long-time depression), делающее синапс менее воз-
будимым. Эти термины до некоторой степени аналогичны возбуждению и по-
давлению, но, в отличие от них, изменяют не мгновенные, а долговременные
свойства синапса. В терминах Хэбба это описывается как два типа логических
изменений в силе синапса после обучения.
Рис. 16 .30 . Рецепторы и длинные белковые молекулы, заякоренные в мембране. Длин-
ная полипептидная цепь несколько раз пронизывает мембрану клетки, образуя рецептор
или канал. (Источник: Squire et al.)
300
Глава 16
Метаботропные рецепторы могут быть очень сложными, поскольку в эво-
люционном плане они должны были стать частью уже существовавшей на мо-
мент их возникновения метаболической сети клетки. Однако основная идея и
в этом случае весьма проста: нейромедиатор глутамат связывается с рецептор-
ным комплексом и активирует G-белок (назван так потому, что использует для
работы гуанозинтрифосфат (ГТФ) и гуанозиндифосфат (ГДФ) — в английском
варианте GDP и GTP). G -белки плотно изучались как часть базовой биологии
клетки, в которой они контролируют метаболичекие пути и активность генов.
Рецепторы, связанные с G-белками, также называют вторичными мессенджера-
ми, поскольку они активируются первичным — молекулой медиатора — и пере-
дают сигнал дальше. Вторичные мессенджеры могут опосредовать активацию
третичных, которые глубже интегрированы в аппарат регулируемой функции.
Рис. 16 .31 . Ионотропные и метаботропные глутаматные рецепторы. Глутамат активи-
рует два типа рецепторов — ионотропные и метаботропные. Ионотропные рецепторы
открывают и закрывают ионные каналы — точно так же, как изменение потенциала от-
крывает и закрывает натриевые и калиевые каналы, контролирующие трансмембранный
ток ионов. Метаботропные рецепторы, изображенные справа, регулируют активность
метаболических путей клетки, что эволюционно предшествовало развитию специали-
зированной нервной системы. Рецепторы, связанные с G-белками, также называют вто-
ричными мессенджерами. (Источник: Forsythe and Barnes-Davies, 1997.)
Гены и молекулы мышления
301
Рецепторы, связанные с G-белками, имеют немаловажное практическое зна-
чение. Очень большое количество лекарств использует в качестве цели именно
этот уровень нейрональной передачи сигнала и регуляции метаболической ак-
тивности.
Метаботропные рецепторы еще более разнообразны, чем ионотропные, —
для глутамата их существует даже не два, а по меньшей мере восемь. Разуме-
ется, описывать каждый из этих типов подробно мы не будем. Однако полезно
помнить причины существования такого разнообразия. Одной из таких причин
является то, что глутамат задействован в 90% возбуждающих синапсов коры.
Но эти синапсы имеют самые разнообразные функции — от визуального вос-
приятия до обеспечения эмоциональных реакций. Ситуация, возникающая при
этом, до некоторой степени аналогична той, что имеет место в сети Интернет.
Каждый компьютер в сети используют одни и те же универсальные правила
кодировки — ASCII. Но вместе с тем все компьютеры имеют уникальные IP-
адреса, позволяющие отправлять данные с одной машины на другую и не оши-
баться при этом. Так же и разные типы рецепторов могут служить для иденти-
фикации различных типов синапсов, которые, возможно, характерны только для
своей области мозга, либо осуществляют определенную функцию. Поскольку
мозг использует распространенные вещества, должны существовать различия
в синапсах, использующих эти вещества. (На самом деле и то, и другое имеет
место. Но глутамат и ГАМК все же используются в разных рецепторных сис-
темах.)
После того как активируется достаточно постсинаптических рецепторов,
происходит деполяризация постсинаптической клетки, обусловленная либо то-
ком ионов через мембрану, либо более сложными механизмами. Если на клетку
подается достаточно деполяризующего сигнала в течение небольшого периода
времени, по ее аксону проходит волна потенциала действия. Также это может
оказать влияние на потенциал мембраны дендритов. Этот потенциал, как сви-
детельствуют последние исследования, тоже может иметь определенную роль
при обработке информации. Однако здесь мы сосредоточимся на аксональном
ПД как на основной сигнальной функции нейрона.
Глутаматный рецептор имеет длинную эволюционную историю, как можно
предположить из его тесной связи с рецепторами и основными биологически-
ми молекулами, такими как глюкоза и аминокислота глутамин. Японские, ки-
тайские и французские кулинары характеризуют вкус глутамата как «свежий»,
«мясной» или «бульонный» и считают, что он обязательно должен присутство-
вать у качественной белковой пищи. Сейчас этот вкус называют «умами»; этот
вкус входит в число тех пяти, к которым человек имеет вкусовые рецепторы.
С биологической точки зрения, этот вкус соответствует свежему мясу. Посколь-
ку млекопитающие должны быть способны отличать свежее мясо от испорчен-
ного, глутаматный вкусовой рецептор с этой точки зрения дал им значительное
преимущество. Одним из рецепторов умами является рецептор mGlu номер 4
(mGlu4), показанный на рис. 16.31. На рис. 16.32 представлено его детальное
строение в качестве мембранного рецептора на языке. Это — еще один пример
высокой консервативности многих биологических молекул.
Связанные с глутаматом нарушения лежат в основе многих расстройств
мозга, включая депрессию, гипервозбудимость (особенно из-за его сходств с
302
Глава 16
ГАМК) и повреждения после инсульта (эксайтотоксичность). Глутаматные си-
напсы по разным оценкам потребляют до 90% энергии, идущей на поддержа-
ние работы мозга. Они очень важны при обучении и, поскольку глутамат явля-
ется основным возбуждающим медиатором коры, множестве других функций.
5.8 . Трансмиттерзависимые ионные каналы
Лигандом называется молекула, связывающаяся с рецептором (от лат. ligare —
«связывать»). Глутамат — один из нейромедиаторов, способных открывать
ионные каналы в мембране, что позволяет ионам свободно двигаться через
мембрану, приводя таким образом к деполяризации. Как показано на рис. 16.33,
существует множество других молекул, способных открывать мембранные ка-
налы — среди них ацетилхолин, ГАМК, серотонин (5НТ) и гликоген. На этом
рисунке изображено несколько разрезов через мембрану разных трансмиттер-
зависимых каналов, а также их вид со стороны мембраны. Зеленым показан
специфический рецептор серотонина (ГАМК) и ацетилхолина (R-рецептор). На
другом рисунке показан ионотропный глутаматный рецептор (оранжевый), рас-
познающий также NMDA, АМРА и каинат (открыт позже). АТФ — основное
макроэргическое соединение клетки — также может выступать в качестве ли-
ганда, открывающего ионные каналы.
Рис. 16 .32 . Умами — вкус глутамата. Японские, китайские и французские кулинары
издавна считают умами вкусом свежести. Впервые он был выделен в 1907 г. в Японии
как вкус соевого соуса, и является важным вкусовым компонентом бульонов и соусов.
За восприятие этого вкуса отвечает глутаматный рецептор mGlu4 (один из метаботроп-
ных рецепторов, встречающийся, помимо языка, также в мозге), наряду с некоторыми
другими. Обратите внимание, что рецептор активирует G-белок, обусловливающий по-
вышение концентрации цАМФ — производного АТФ. (Источник: Squore et al., 2008.)
Гены и молекулы мышления
303
Сигнальные пути, включающие G-белки, чрезвычайно важны. Около поло-
вины имеющихся на сегодняшний день лекарств влияют именно на них. Ре-
цепторы, связанные с G-белками, чувствительны к свету, запахам, гормонам и
нейромедиаторам (рис. 16.34).
C учетом того факта, что ежесекундно в мозге происходят триллионы раз-
личных событий, может показаться странным, что в коре имеются лишь два
основных медиатора — глутамат и ГАМК. На самом же деле нейромедиаторов
на сегодняшний момент известно более ста, и это, без сомнения, не все. Однако
возможности кодировки сигнала не ограничиваются только разнообразием ме-
диаторов, ведь для каждого из них может существовать более одного рецептора
(на рис. 16.35 показаны результаты позитронно-томографического исследова-
ния дофаминовых рецепторов в мозге человека и обезьяны). В результате один
медиатор может давать различные эффекты в разных типах синапсов в зависи-
мости от типа рецепторов.
Многозадачность — широко распространенное в биологии явление. К при-
меру, все сухопутные позвоночные имеют легкие, при помощи которых у них
осуществляется газообмен. Однако рептилии, помимо газообмена, используют
легкие для того, чтобы издавать рычащие звуки. Млекопитающие пошли еще
Рис. 16 .33 . Трансмиттерзависимые ионные каналы. TRP-канал (от англ. transient
receptor potential) (Источник: Gopala-krishnan и Briggs, 2007.)
304
Глава 16
дальше и способны издавать при помощи легких большой набор звуков, ре-
гулируя поток воздуха при помощи голосовых связок; человек, при помощи
развитой мускулатуры губ, способен еще сильнее менять звучание, придавая
ему вид гласных и согласных. Таким образом, звуковая коммуникация целиком
базируется на легких, изначально возникших совсем для других целей. Исполь-
зование первичных адаптаций для других целей встретится нам еще не еди-
ножды.
Нейроактивные вещества, которые мы обсуждаем в этой главе, в той или
иной степени используют для работы эволюционно более древние механизмы
мозга. Глутамат синтезируется из аминокислоты глутамина, которая, в свою
очередь, синтезируется из глюкозы. Метаботропные рецепторы регулируют ра-
боту основных метаболических путей, имеющихся в каждой клетке. Этот мо-
мент важен, поскольку позволяет лучше понять и запомнить то невероятное
число веществ и реакций, которые имеют место в мозге. Одной из причин тако-
го упрощения понимания как раз и является использование основных молекул
организма для специфических целей мозга.
Рис. 16.34 . Метаботропные и ионотропные рецепторы. Сверху: молекулы медиатора
связываются с ионотропным рецептором (а) и метаботропным рецептором, связанным
с G-белком (б). G -белки являются частью энергетического метаболизма любой клетки,
связанного с цАМФ. В мозге же древние биохимические процессы, такие, как цАМФ-
зависимый путь, используются для передачи сигнала. Снизу: схемы полипептидной
цепи рецепторов обоих типов. Обратите внимание на различие в количестве и организа-
ции трансмембранных участков. Показанные на рисунке группы NH3 и COOH имеются
на концах любой полипептидной цепи в силу строения ее мономеров — аминокислот.
(Источник: Squire et al., 2003.)
Гены и молекулы мышления
305
На рис. 16.36 показан еще один пример такого использования — на этот раз
для АТФ, который является основным макроэргическим веществом в клетке.
АТФ, как показано на рисунке, также используется в качестве нейромедиато-
ра — иногда совместно с глутаматом или ГАМК и всегда при посредстве гли-
альной клетки.
Рис. 16 .35. Распределение дофаминовых рецепторов D2/D3 в мозге. При использова-
нии радиоактивной молекулы, связывающейся с рецептором (в данном случае — с ре-
цепторами D2 и D3), возможно произвести сканирование всего мозга с использованием
протонно-эмиссионной томографии. Слева показано распределение рецепторов D2/D3
в мозге человека, справа — в мозге макака-резуса. Обратите внимание на их высокое
содержание в таламусе (Th) у человека и остальных приматов. DVR — отношение плот-
ности распределения рецепторов к объему, Pit — гипофиз, HCd — головка хвостатого
ядра, Pu — скорлупа, TCd — хвост хвостатого ядра. (Источник: Christian et al., 2009.)
Рис. 16 .36 . Энергетическая молекула АТФ также используется в качестве нейромедиа-
тора. (Источник: Verkhratsky and Krishtal из Squire, 2009.)
306
Глава 16
5.9 . Очистка синапса: дезактивация, диффузия
и утилизация медиатора
Регуляция концентраций различных веществ в организме крайне важна. Наша
жизнь зависит от наличия правильных количеств требуемых веществ, а при по-
вышенном или пониженном их содержании может развиваться заболевание.
Все реакции в клетке имеют в качестве побочных продуктов радикалы — ча -
стицы с неспаренным электроном на внешнем уровне. Чаще всего ими явля-
ются активные формы кислорода — например, пероксид водорода или гидрок-
сид-радикал, хотя ими могут быть и части белковых молекул, как, например,
аминная фракция аминокислот. Из-за особенностей строения электронной обо-
лочки радикалы крайне реакционноспособны и реагируют практически с лю-
бой заряженной молекулой. Их действие иногда называют кросс-линкингом
—
в результате созданных радикалами случайных химических связей происхо-
дит сморщивание кожи. Такое действие радикалов является одним из главных
механизмов старения и связанных с ним болезней, таких как атеросклероз. Для
защиты от радикалов в организме имеется целый набор веществ, называемых
антиоксидантами, способных реагировать с ними с образованием безопасных
соединений, — среди таких антиоксидантов можно назвать витамины и неко-
торые жиры.
Воспаление — нормальный физиологический ответ на повреждение, необ-
ходимый для его устранения. Тем не менее хроническое воспаление является
одним из основных факторов дегенеративных заболеваний, таких как нейроде-
генеративные болезни Паркинсона и Альцгеймера. Таким образом, регуляция
продолжительности воспаления также важна для поддержания нормального
функционирования нервной системы.
Мозг — наиболее интенсивно потребляющий энергию орган человеческого
тела, особенно при современном образе жизни, при котором человек далеко
не весь день проводит в движении. Как следствие, мозг производит большое
количество свободных радикалов и повреждений, способных вызвать воспале-
ние. Синапс и внеклеточное пространство — два наиболее уязвимых при не-
правильном контроле места, поскольку они задействованы в механизмах само-
регуляции клетки (что со временем вызовет еще большие нарушения).
Очистка синапса — один из хороших примеров необходимости регуляции
количества потенциально токсических веществ. Мы уже отмечали, что глута-
мат — наиболее распространенный возбуждающий медиатор — может быть
токсичен, если его не удалять из синапса. Эксайтотоксичность является наи-
более распространенной причиной повреждений мозга после инсульта. При
инсульте происходит разрыв кровеносных сосудов головного мозга. Однако
влияние вытекшей крови и повреждения других тканей не столь вредоносно,
как последующее повреждение свободными радикалами, воспалением и экс-
айтотоксичностью. В силу этого лечение при инсульте в последние годы на-
правлено именно на устранение этих эффектов, развивающихся сразу же после
разрыва сосуда.
В нервной системе имеется множество механизмов поддержания концен-
трации потенциально опасных веществ в норме. Перечислим три основных
типа очистки синапса.
Гены и молекулы мышления
307
1. Диффузия молекул медиатора, при помощи которой производится удаление
оксида азота NO. Роль NO в качестве нейромедиатора была неизвестна еще
около десяти лет назад, однако за последние годы установлено, что он игра-
ет важную роль во многих синапсах — наиболее известным примером его
влияния является препарат «Виагра». Помимо роли в синаптической переда-
че, NO выполняет в нервной системе много других функций. Поскольку при
нормальных условиях он представляет собой газ, секретируется он в очень
малых количествах, что позволяет ему быстро и безопасно диффундировать
за пределы синаптической щели.
2. Деградация. Ацетилхолин расщепляется ферментом ацетилхолинэстеразой.
В процессе ферментативной деградации происходит распад молекулы меди-
атора на несколько неактивных частей, образующиеся в результате продук-
ты захватываются и утилизируются пресинаптическими нейронами.
3. Транспортировка в пресинаптический нейрон. Дофамин — наиболее из-
вестный пример такой транспортировки. На рис. 16.37 показан результат
протонно-эмиссионного исследования мозга с использованием радиоактив-
но меченого вещества, способного связываться с молекулой переносчика
дофамина. Слева показан посмертный разрез мозга; в центре — протонно-
эмиссионная томограмма мозга здорового человека, на которой видна вы-
сокая концентрация переносчика дофамина, особенно в области базальных
ганглиев и таламуса (хвостатое и чечевицеобразное ядра); справа приведен
результат исследования пациента с болезнью Паркинсона, при которой про-
исходит нарушение синтеза дофамина. На последнем изображении виден
сниженный уровень дофаминового переносчика, что, возможно, отражает
пониженное содержание дофамина в мозге пациента. Хотя не исключе-
но, что при болезни Паркинсона происходит и более прямое повреждение
транспортера дофамина.
Рис. 16 .37. Транспортер возвращает дофамин в клетку происхождения. Протонно-резо-
нансные томограммы меченых молекул переносчика дофамина. Отмечены хвостатое и
чечевицеобразное ядра. Слева: анатомический фронтальный срез мозга на том уровне,
на котором производилось исследование. В центре: томограмма нормального мозга с
большим содержанием метки в хвостатом и чечевицеобразном ядрах. Справа: резуль-
тат исследования мозга пациента с болезнью Паркинсона, вызванной отмиранием до-
фаминергических нейронов черного вещества, аксоны которых простираются далеко в
передний мозг. Базальные ганглии, контролируемые нейронами черного вещества, уча-
ствующими в контроле моторной активности, которая нарушается при развитии болез-
ни Паркинсона. (Источник: Newberg and Alavi, 2009.)
308
Глава 16
6.0. Нейромодуляторы
Нейромодуляция отличается от опосредованной медатором передачи сигнала
тем, что вещество, осуществляющее ее, широко распространено во многих ча-
стях мозга. При этом одно и то же вещество может выполнять различные роли
в зависимости от того, где оно было секретировано. Ацетилхолин и глутамат
могут быть как медиаторами, так и модуляторами. В качестве нейромодулято-
ров эти вещества могут менять активность целых областей мозга, влияя на мил-
лионы нейронов и синапсов. Так, нейромодуляторы могут изменять потенциал
покоя целых популяций клеток, изменяя в них таким образом синаптическую
сигнализацию. Нейромодуляторы регулируют баланс между восприятием сен-
сорной информации и внутренними процессами, переключая мозг между фа-
зами бодрствования и быстрого и медленного сна. При бодрствовании сенсор-
ная информация обрабатывается в максимальном объеме, однако эта обработка
сильно снижена или полностью заблокирована во время быстрого и медленно-
го сна. Эти различия как раз и контролируются нейромодуляторами.
Медиатор-опосредованная передача сигнала гораздо быстрее, чем нейромо-
дуляция, и в большей степени ионотропна, тогда как нейромодуляция скорее
метаботропна. При этом в одно и то же время может действовать несколько
модуляторов. Так, мозг находится в бодрствующем состоянии тогда, когда в
определенных областях коры и таламуса высока концентрация ацетилхолина,
серотонина, норэпинефрина и дофамина (см. гл . 8). Из данных табл. 16.2 вид-
но, для чего мозгу в бодрствующем состоянии требуется поддерживать мно-
жество обусловленных разными нейромодуляторами функций одновременно:
это и внимание, и мотивация, и точная работа сенсорных систем (высокое от-
ношение сигнал-шум), а также точное планирование и двигательный контроль.
В состоянии бодрствования человек должен быть способен использовать при
необходимости все эти функции одновременно.
Клетки, осуществляющие нейромодуляцию, локализованы в базальной об-
ласти переднего мозга и мозговом стволе, откуда они выбрасывают нейромоду-
лятор в обширные зоны переднего мозга (рис. 16.39). Состояние бодрствования
опосредовано нейромодуляторами ацетилхолином, норэпинефрином и дофами-
ном. Вместе они делают возможным выполнение различных когнитивных задач.
Каждая из таких задач требует сочетания локального действия нейромедиато-
ров, таких как глутамат и ГАМК, с более обширным влиянием нейромодулято-
ров, контролирующих общее состояние мозга, обусловливающее возможность
обработки сенсорной информации или интеграции долговременной памяти.
Черное вещество (лат. substantia nigra) представляет собой небольшой клин
нервных клеток, расположенный над стволом мозга, аксоны нейронов которо-
го простираются далеко вверх и модулируют активность миллиардов других
нейронов (рис. 16.38). Все черное вещество содержит примерно 10 000 дофми-
нергических нейронов — крайне мало по сравнению с десятками миллиардов
нейронов коры. Однако их роль не стоит недооценивать — при их потере, как,
например, в случае болезни Паркинсона, ущерб может быть огромен. Дофами-
новая модуляция обусловливает основные реакции, такие как удовольствие или
желание награды, сон и бодрствование, никотиновая зависимость и рабочая
память, произвольный контроль движения и целенаправленное поведение. Как
видите, набор функций действительно огромен. (Как и большинство других
Гены и молекулы мышления
309
Таблица 16.2
Основные функции нейромодуляторов
Нейромодулятор
Некоторые основные функции
Норэпинефрин
Поддерживае т высокое отношение сигнал-шум в сенсорных
сис темах
Серотонин
Защищает от перегрузки информацией и уменьшае т взаимо-
действие между разными сенсорными каналами
Витамины и ацетилхолин:
Дофамин
Поддерживает психомоторную и мотивационную фокуси-
ровку и внимание (сознательное возбуждение)
Ацетилхолин
Регулирует степень внимания и сознательного возбужде-
ния во всех сенсорных системах
Бета-эндорфин
Противодействует нарушениям гомеостаза; обуславливает
удовольствие
Кортиколиберин
Обусловливает развитие стресса и негативных эмоцио-
нальных реакций
Нейропептиды
Вазопрессин/окситоцин Вазопрессин отвечает за мужскую настойчивость; оксито-
цин — за женскую заботливость и мягкость
Холецистокинин
Регулирует эмоциональные реакции, чувства, половые ре-
акции, исследование и боль
Источник: Northoff and Panksepp, 2008
Рис. 16 .38 . Нейромодуляция: базальные ядра влияют на множество протекающих
в мозге процессов. Нейромодулирующая активность базальных ядер регулирует ак-
тивность коры и базальных ганглиев. (Источник: National institute of general medical
sciences, 2009.)
310
Глава 16
структур мозга, все нейромодулирующие ядра парные. Каждое ядро черного
вещества состоит из двух частей — компактной и сетчатой.)
Большинство синапсов комбинирует локальную передачу сигнала и нейро-
модуляцию. К примеру, нейрон зрительной коры может вести себя по-разному в
зависимости от общего состояния мозга, таких как сон и бодрствование, которые,
в свою очередь, обусловлены активностью нейронов ядер ствола мозга, аксоны
которых достигают коры и доставляют туда нейромодуляторы.
7.0 . Обучение
В гл. 9 описаны процессы, происходящие при обучении на уровне мозга. Здесь
же мы рассмотрим молекулярные аспекты этих процессов. Достижения в этой
области имеют и будут иметь в дальнейшем огромное влияние не только на
терапию соответствующих заболеваний, но и на улучшение обучаемости в те-
чение жизни.
Рис. 16 .39. Пять основных нейромодуляторов. Обратите внимание на то, что нейроны,
производящие все пять, находятся в небольших ядрах под корой и распределяют моду-
ляторы на большие области мозга. При этом всего несколько тысяч нейронов способны
регулировать активность многих миллиардов клеток, контролируя сон и бодрствование,
удовольствие и боль, внимание и рабочую память. Если модулирующие клетки гиб-
нут, как в случае дофаминергических клеток при болезни Паркинсона, результат может
быть крайне разрушительным и даже привести к смерти. (Источник: Gu, рис. 1, с из-
менениями 2002.)
Гены и молекулы мышления
311
Все области мозга так или иначе задействованы в обучении, запоминании и
пластичности, которые могут рассматриваться как методы построения долго-
временной адаптивной изменчивости. Наиболее типичным примером такой
адаптации является обучение по Хэббу, которое можно определить как образо-
вание синапсов только координированного прохождения ПД по сети нейронов.
Таким образом, нейроны соединяются так, чтобы образовывать работающую
сеть. Эта модель достаточно неплохо отражает нейрональный уровень обуче-
ния. Основным положением этой модели является то, что обучение такого типа
должно происходить практически в любой части мозга, в которой имеются та-
кие синапсы. На рис. 16.40 показан широкий набор процессов обучения, а также
области мозга, в которых они имеют место. Как можно видеть, не существует
отдельной области, ответственной за обучение. Любая функция мозга адапти-
руется к изменяющимся условиям. Одним из самых знаменитых примеров та-
кого процесса является адаптация сенсорной коры при раннем визуальном об-
учении у котят (Hubel и Wiesel, 1968). В повседневной жизни мы сталкиваемся
с подобного рода обучением, когда внимательнее вслушиваемся в уже знакомое
Рис. 16 .40. За обучение ответственны все зоны в мозге. За память отвечают те же об-
ласти, под чьим контролем находятся активные задачи. Таким образом, сенсорная па-
мять задействует селективное усиление и настройку синапсов соответствующих полей
сенсорной коры. Миндалина, гиппокамп и медиальная префронтальная доля вовлечены
в разные аспекты эмоционального обучения. Декларативная память контролируется
гиппокампальным комплексом, однако после консолидации она хранится также и в но-
вой коре (см. гл . 12). Таким образом, обучение того или иного типа происходит во всех
частях мозга. (Источник: Eichenbaum, из Squire, 2003.)
312
Глава 16
музыкальное произведение, что позволяет нам получить дополнительный опыт
из уже встречавшегося физического явления.
Фронтальная область коры отвечает за обучение исполнительным функциям
(см. гл. 12), рабочей памяти, речи и всему, что связано с языком, а также, ра-
зумеется, за моторный контроль. Мерзених с соавторами (1984) показали, что
утрата пальца вызывает соответствующую адаптивную реорганизацию мотор-
ных и сенсорных карт ладони и пальцев в коре. До ампутации все пять пальцев
были представлены одинаково (это было продемонстрировано регистрацией
ответа соответствующих областей коры при стимуляции пальцев); после ам-
путации ответ на сигналы от утраченного пальца снизился в пользу усиления
ответа на сигналы от соседних. Это позволяет оптимизировать работу коры под
оставшиеся пальцы. Этот принцип применим ко всем моторным и сенсорным
областям коры, а также, возможно, к ее префронтальным областям (Merzenich
et al., 1984).
Сходная редукция наблюдается при частом использовании одного и того же
пальца. В случае, если одно и то же движение пальца раз за разом повторяется,
соответствующая область коры будет уменьшаться, поскольку кора будет рас-
сматривать это как чрезмерное событие. Тем не менее субкортикальная иннер-
вация пальца при этом не исчезает, поскольку мозг по-прежнему контролирует
и отслеживает движения пальца.
7.1. Гиппокампальный комплекс
Декларативная память осознанных событий, по всей видимости, обусловли-
вается медиальной частью височных долей, которую мы назвали гиппокам-
пальным комплексом (он шире, чем гиппокамп отдельно). На рис. 8 .16 (гл.
8) показан динамический контур временного хранения осознанных эпизодов,
Рис. 16 .41 . Культура среза гиппокампа. Срез гиппокампа способен сохранять жизне-
способность в культуре, что позволяет экспериментаторам обрабатывать его различ-
ными нейромедиаторами и регистрировать ответ множества нейронов одновременно.
И хотя чаще всего используют срез гиппокампа крысы, основные принципы его работы
применимы и для человека. На рисунке показаны гиппокампальные нейроны трех ти-
пов — СА1, СА3 и шафферовы коллатерали. (Источник: Stevens and Sullivan, 1998.)
Гены и молекулы мышления
313
накопленных в бодрствующем состоянии, и включающий сенсорную кору, та-
ламус и гиппокампальный комплекс. Следует помнить, что при эпизодическом
обучении также задействуется большое количество неявных циклов, в резуль-
тате чего сознательные стимулы дают бессознательные последствия, которые
также оказывают влияние на мозг. Хорошо изученным примером подобного
служит роль обучения последовательности слов у детей и неявное обучение
грамматическим правилам в результате явного обучения. Трехлетний ребенок
может сказать «Мама, самолет!», но неявное выражение несет большую нагруз-
ку, чем эти слова. Оно означает «Мама, посмотри на меня и посмотри туда, куда
я показываю, — там летит самолет». Неявное значение также воспринимается
как матерью, так и ребенком. По всей видимости, не существует такого явле-
ния, как изолированное явное обучение — все явные сознательные процессы
также имеют неявную часть.
Считается, что эпизодическое обучение происходит как минимум в две
стадии, первая из которых происходит в течение секунд, минут и часов и за-
действует гиппокампальную область (MВД), которая принимает зрительный и
другие сигналы (на рис. 16.41 показан препарат гиппокампа). В случае зрения,
к примеру, в височной доле происходит интеграция множества потоков в изо-
бражения конкретных объектов и событий. Именно эти осознанные объекты
и события и записываются в немедленную память. На рис. 16.42 показан срез
гиппокампа, из которого очевидна огромная сложность и разнообразие собы-
тий молекулярного кодирования, происходящих при гиппокампальном обуче-
нии. Немедленная память лабильна и потому уязвима к разного рода влияниям.
Электрический шок, примененный непосредственно после обучения, нарушит
Рис. 16 .42 . Активность эндоканна-
биноидов в гиппокампе. Эндогенные
каннабиноидные вещества играют
важную роль в обучении и пластич-
ности. (а) На этой фотографии пока-
зан трансверсальный срез гиппокам-
па, окрашенный при помощи метода
гибридизации in situ для выделения
разных тканей. Коричневым окра-
шены клетки, содержащие мРНК ре-
цептора СВ1. Ядра клеток окрашены
синим. ГАМК-ергические нейроны
отмечены черной стрелкой, глутама-
тергические пирамидальные — бе-
лой. На снимке (б) показана совмест-
ная экспрессия мРНК рецептора СВ1
(красная) и маркера ГАМК (GAD).
(в) Совместная экспрессия мРНК
рецептора СВ1 (красная) и марке-
ра транспортера глутамата типа 1
(VGLUT1). Обратите внимание на
большую сложность и разнообразие
веществ, участвующих в гиппокам-
пальном обучении. (Источник: Lutz
and Marsicano, из Squire et al., 2009.)
314
Глава 16
вновь записанную в память информацию, однако старая информация к такому
воздействию устойчива.
Одним из частых вопросов, возникающих при изучении памяти, является
вопрос о молекулярных и клеточных процессах, ее обусловливающих. Пока-
зано, что даже для немедленной памяти, контур которой показан на рис. 8 .11,
имеют место молекулярные и эпигенетические изменения. Этот аспект мы рас-
смотрим позже (Wang, 2003)
7.2 . Глутамат, ГАМК и долговременные депрессия
и потенциация
Если предположить, что при обучении имеет место образование новых связей
между нейронами, то получится, что может иметь место либо увеличение, либо
уменьшение вероятности образования связи. Традиционно эти явления называ-
ют долговременной потенциацией и договременной депрессией соответствен-
но. До некоторой степени они сходны с возбуждением и торможением, хотя
электрохимические механизмы, лежащие в их основе, не идентичны. За годы
попыток установить истинный механизм долговременных депрессии и потен-
циации стало ясно, что такой механизм не один, поскольку существует большое
количество разнообразных нейронов, синапсов, нейроактивных веществ и мно-
гих прочих свойств и компонентов. Для выполнения одной и той же фундамен-
тальной функции в природе существует, как правило, несколько механизмов.
Одной из причин огромного разнообразия путей и механизмов на всех уровнях
организации является повышенная устойчивость системы с широким набором
Рис. 16 .43. Измерение долговременных потенциации и депрессии. Обратите внима-
ние на то, что на рисунке показаны только вход (тетанический шок) и выход (мВ/с)
препарата. В случае препарата среза гиппокампа можно регистрировать активность как
одиночного синапса, так и целой области клеток, такой как поля СА1 и СА3. (Источник:
Byrne, из Squire et al., 2003.)
Гены и молекулы мышления
315
путей в случае изменения внешних условий (Edelman и Gally, 1992). Инженеры
при создании различных устройств также снабжают их резервными системами
для повышения надежности в экстремальных условиях.
На рис. 16.43 показано, каким образом долговременные депрессии и потен-
циации могут быть исследованы с применением срезов гиппокампа и нейро-
нальных препаратов на его основе. Стимул, обозаченный как «тетанус», пред-
ставляет собой небольшой электрический удар. По вертикальной оси отложены
милливольты за миллисекунду, что отображает состояние мембранного потен-
циала нейрона во времени. Это значение становится приблизительно в два раза
больше после тетанического шока и длится более 90 мин. Такое поведение сви-
детельствует о том, что в повышении мембранного потенциала участвуют ме-
ханизмы, отличные от возбуждающей или тормозной синаптической передачи.
7.3 . Глутаматные синапсы в качестве модели
синаптического обучения
Первой детально изученной системой долговременной потенциации был глута-
матергический синапс — наиболее распространенный возбудительный синапс
в мозге.
На рис. 16.44 показано условно-рефлекторное обучение выбору направле-
ния по Павлову для одного из наиболее изученных организмов — плодовой
мушки Drosophila melanogaster. Мозг этого животного содержит примерно
100 000 нейронов. Мушка отлично летает, выбирает направление, распознает
образы, обучается и имеет социальное поведение, включающее схватки самцов
за самку или пищу и отбор подходящего партнера для размножения. Одним из
важнейших чувств для этого насекомого является обоняние. Запах фруктовых
сахаров помогает мушке находить пищу, прочие запахи могут предупреждать
о присутствии токсинов или служить феромонами для привлечения особей
противоположного пола. Мушку можно научить ассоциировать тот или иной
запах с неприятным событием, таким как удар током, поскольку электриче-
ский тетанус влияет на сигнализацию во всей нервной ткани. Передача сигнала
между нейронами осуществляется электрохимически, вследстие чего она чув-
ствительна к электрическому воздействию. Также нельзя не принимать во вни-
мание, что шок болезнен и неприятен и, следовательно, запускает мехнизмы
отслеживания и избегания угроз.
На рис. 16.44 (а) показана экспериментальная установка, представляющая
собой небольшой Т-образный лабиринт, на концах которого размещены две ка-
меры с двумя разными запахами. Запах 1 служит в качестве условного стиму-
ла для нежелательного воздействия, в качестве которого здесь выступает удар
электрическим током. Если запах 1 сочетается с ударом током по лапкам мушки,
то она быстро учится следовать в направлении безопасного запаха — запаха 2,
двигаясь от точки выбора по направлению от запаха 1. Поток воздуха помогает
мухе двигаться и препятствует ее возвращению в пройденные части лабиринта.
Вторая часть рис. 16.44 демонстрирует принятую ныне модель обучения
на глутаматергических синапсах Drosophila. Теоретический вопрос здесь про-
стой и следует непосредственно из хэббовской теории обучения: если два ней-
рона деполяризуются одновременно, будут ли они образовывать связи? Будет
ли синапс отмечать происходящие одновременно события, такие как запах 1 и
электрический ток? И может ли синапс быть усилен и будет ли он с большей
316
Глава 16
вероятностью передавать сигнал в следующий раз, когда животное встретит за-
пах 1? Эти вопросы могут быть простыми, однако молекулярные механизмы,
необходимые для выполнения поставленных задач, нельзя назвать простыми
(на рис. 16.45, 16.46 и 16.47 приведены схемы этих механизмов). Стоит отме-
тить, что поддерживающие у D. melanogaster обучение глутаматергические си-
напсы высококонсервативны, а следовательно, данные, полученные на дрозо-
филе, применимы и для человека.
Рис. 16 .44 . Условно-рефлекторное обучение (по Павлову) плодовой мушки. Класси-
ческая выработка условного рефлекса у дрозофилы делает возможным изучение ассо-
циативного обучения в относительно небольшой нервной системе, используя запах в
качестве сигнала о нежелательном воздействии — ударе электрическим током. Ионо-
тропные рецепторы глутамата носят название NMDA и АМРА. Они непосредственно
открывают ионные каналы на мембране, среди которых — кальциевые и магниевые
каналы. Обратите внимание на частоту прохождения ПД для двух нейронов (обозначена
набором вертикальных линий). Долговременная потенциация требует дополнительных
и более глубоких событий в клетке для развития структурных изменений в синапсах.
(Источник: Glanzman, 2005.)
Гены и молекулы мышления
317
Одним из интересных способов усилить синаптические связи между двумя
клетками является их соединение друг с другом при помощи молекул клеточ-
ной адгезии. На рис. 16.48 показано, каким образом, в общих чертах, работает
этот механизм.
Обучение требует структурных изменений в нейронах — роста дендритов,
образования новых синапсов и настройки метаболических путей для удов-
летворения возросших потребностей клетки. Большую роль в обучении, по-
видимому, играет образование связей между нейронами по принципу Хэбба,
которое подчиняется правилу «одновременно деполяризующиеся нейроны об-
разуют связи друг с другом» (см. гл . 3 и 10). Поскольку образование связей тре-
бует образования больших по размерам и более разветвленных контактов, эпи-
генетический контроль экспрессии непременно вовлекается в этот процесс. По
определению обучение представляет собой «обусловленные опытом изменения
в головном мозге». Опыт, накапливающийся за время нашего бодрствования,
Рис. 16 .45. Долговременная потенциация с участием рецепторов NMDA и АМРА. Де-
тальное рассмотрение зависимости рецептора NMDA от частоты прохождения ПД при
долговременной потенциации. Слева: в отсутствие глутамата в синаптической щели
канал рецептора NMDA блокирован ионом магния (черная точка). В центре: показан
тот же синапс при базовой скорости высвобождения медиатора из пресинаптической
клетки, при которой с рецептором NMDA также не происходит никаких событий. Глу-
тамат (красный) присутствует в синаптической щели, однако канал рецептора NMDA
по-прежнему блокирован ионом магния. Справа: высокая частота сигналов от преси-
наптической клетки активирует рецептор NMDA. Таким образом, синапс работает как
переключатель с высокой полосой пропускания, который активируется только при по-
даче сигнала высокой частоты. (Источник: Doherty, Fitzjohn and Collingridge, из Squire
et al., 2009.)
318
Глава 16
составляет входные данные от окружающей среды, а изменения в мозге, проис-
ходящие в результате таких воздействий, требуют новых структурных элемен-
тов и связей между ними, дабы активные нейрональные сети могли адаптиро-
ваться к полученным данным.
Подобно автомобильной фабрике, каждая клетка отвечает на адаптив-
ные требования производством требуемых количеств необходимого продукта
(рис. 6 .49).
Мы описали обучение в терминах Хэбба в виде усиления синаптической свя-
зи между нейронами. Такое описание в некоторой степени отражает действи-
тельность, однако это — только часть адаптивного аппарата мозга. Считается,
что могут существовать и отличные от хэббовских механизмы обучения. Ведь
адаптация к окружающей среде, к нам самим и к окружающим — это то, что
Рис. 16 .46 . Долговременная потенциация: детальное рассмотрение рецептора NMDA.
Обучение происходит не только на уровне отдельных нейронов. Чаще оно проис-
ходит как сетевое событие и задействует множество возбуждающих и тормозных си-
напсов. Вверху слева: глутаматергическая клетка, иннервирующая тормозный ГАМК-
ергический нейрон, который, в свою очередь, снижает вероятность прохождения ПД по
клетке внизу рисунка. В результате клетка-мишень получает как возбуждающие, так и
тормозные импульсы. Справа: мембранные потенциалы, обусловленные ГАМК и глу-
таматом, причем рецептор АМРА дает выброс в положительную область, а рецептор
NMDA перекрыт ионом магния (черная точка). Представлены два рецептора ГАМК
—
ГАМКА и ГАМКB
, влияющие на клетку по-разному. Возбудительный постсинапти-
ческий потенциал является нижней границей активации системы, за которой следует
прохождение ПД с периодом рефрактерности. Контуры, подобные этому, могут быть
сложными и способны к адаптации. (Источник: Doherty, Fitzjohn and Collingridge, из
Squire et al., 2009.)
Гены и молекулы мышления
319
делает мозг. За сотни миллионов лет эволюции мозг выработал множество ме-
ханизмов адаптации, каждый из которых включает обучение в широком смысле
этого слова. Термины «обучение», «память» и «синаптическая пластичность» в
настоящий момент стали практически синонимами. Исторически термин «об-
учение» был принят под влиянием ассоциативного обучения, особенно класси-
ческих (Павловских) и инструментальных условных рефлексов. Пластичность
была впервые изучена на примере сенсорного обучения в работе Хубеля (Hubel)
и Визеля (Wiesel) по адаптации доминирования глаза у котят, лишенных воз-
можности видеть одним из глаз. Пластичность изучалась с учетом импринтинга
в критические периоды — быстрого обучения, имеющего место у детенышей
млекопитающих, и, возможно, даже у нерожденного плода. Импринтинг важен
для развития зрительной коры, зрительно-моторного обучения, обучения по-
ниманию речи и многих других функций.
Тем не менее сейчас термин «пластичность» имеет более широкое значе-
ние. Известно, что взрослые особи обладают кортикальной пластичностью на
протяжении всей жизни, хотя она и меньше ранней пластичности зрительного
мозга. Серьезные изменения сенсорной или моторной части, такие как поте-
ря пальца, запускают механизмы пластичности в сенсорно-моторной области
коры. Однако даже повторение одного и того же действия — к примеру, набор
на клавиатуре того или иного слова — также ведет к изменениям кортикальной
Рис. 16.47. Активация транскрипции и трансляции при долговременной потенциации
и депрессии. Долговременная депрессия и потенциация требуют синтеза белка для уве-
личения количества синапсов и увеличения количества нейроактивных веществ (чаще
всего белков) во всех частях системы. При сильной стимуляции синапсов активируется
как трансляционный (мРНК), так и транскрипционный (ДНК-мРНК) механизм
320
Глава 16
карты того пальца, которым мы это делаем, а также соседних пальцев. Поэтому
пластичность нельзя отделять от других адаптивных функций нервной систе-
мы. Иногда бывает полезно подчеркнуть существующие между этими явлени-
ями различия, однако мы будем считать их частями одного и того же явления.
Термин «память» также имеет свою собственную историю, в течение ко-
торой он находился под влиянием природы хранимой информации. Однако в
случае мозга это влияние не столь выражено. Одной из причин этого является
то, что разделить разные типы информации в мозге практически невозможно.
Рис. 16.48 . Молекулы клеточной адгезии стабилизируют синапс. Молекулы кадгерина
(красные) удерживают мембраны синапса поблизости друг от друга, позволяя осущест-
влять синаптическую передачу. Молекулы клеточной адгезии имеют множество функ-
ций и, по всей видимости, необходимы для обучения и синаптической пластичности.
NMDAR — NMDA-рецептор, AMPAR — AMPA-рецептор. (Источник: Tai, Kim and
Schuman, 2008.)
Гены и молекулы мышления
321
Существа, имеющие мозг, начинают учиться сразу же после помещения в
новую среду, которая может быть любой. Новые стимулы вызывают массиро-
ванный ответ, включающий масштабную активацию мозга. Когда животное
почувствует себя в безопасности в новой среде, оно начнет собирать инфор-
мацию, осматривая и обнюхивая окружение. Новизна сама по себе является
достаточным стимулом для обострения внимания и начала обучения.
Грызуны в лабораторных экспериментах чувствительны к запахам экспе-
риментатора, к примеру к запаху его кошки. Для лабораторной крысы запах
хищника означает прямую угрозу, и некоторые исследователи отмечают тен-
денцию у грызунов к замиранию от страха. В таком состоянии обучение сильно
затруднено либо животное научится не тому, что предполагал экспериментатор.
Обучение в экспериментальных задачах требует, чтобы животное чувствовало
себя достаточно безопасно для исследования.
В настоящей лаборатории, как и в реальной жизни, происходит сразу не-
сколько типов обучения. При рассмотрении событий, происходящих при этом
в мозге, весь набор адаптивных событий становится очевиден.
Рис. 16 .49 . Некоторые способы усиления синаптической сигнализации. Существует
много путей усиления синапса. Для серотонина (5-НТ) на этом рисунке приведено че-
тыре различных способа. Один из них заключается во внесении изменений в структур-
ные элементы клетки — цитоскелет, в результате чего происходит увеличение числа
синапсов. Серотонин также может обусловливать образование синаптичеких отростков,
управляя экспрессией генов для увеличения стабильности синапсов. (Синапсы, как и
любой другой элемент клетки, непрерывно обновляются.) Нейромедиатор также мо-
жет запустить структурные изменения в уже существующих синапсах, увеличивая их
или усиливая выброс медиатора. Кроме того, серотонин способен действовать в каче-
стве вторичного мессенджера, усиливая трансляцию (с участием мРНК) или запуская
транскрипцию молчавшего ранее гена (обусловливая эпигенетические изменения в экс-
прессии ДНК), что, в свою очередь, ведет к усилению выброса медиатора. (Источник:
Sossin, 2008.)
322
Глава 16
7.4 . Эпигенетические механизмы обучения
Как уже указывалось ранее, эпигенетические факторы в настоящее время слу-
жат объектом пристального интереса при изучении поцессов обучения. При об-
учении имеет место генетический контроль нейронов и глии, задействованных
в этой задаче. Нейромедиаторы влияют на экпрессию генов как в клетке-мише-
ни, так и в клетке, из которой они происходят. Поскольку в ДНК закодирован
синтетический аппарат различных веществ, это имеет место и для пресинап-
тичекой клетки. Однако это справедливо и для постсинаптического нейрона,
и для окружающей нейроны глии.
На рис. 16.50 показана зависимость экспрессии генов в постсинаптической
клетке от активности. Периодическая стимуляция нейрона вызывает экспрес-
сию специфических генов, делающих обучение возможным. Тета-ритмы, как
известно на настоящий момент, возникают в гиппокампе во время обучения и
существуют еще некоторое время спустя, выступая в качестве платформы для
кортикально-гиппокампальной консолидации. Таким образом, тета-ритмы, по-
видимому, оказывают влияние на нейроны, в которых они образуются. Такая
ритмическая стимуляция запускает эпигенетические процессы в популяциях
клеток.
Этот механизм использует важный кальцийзависимый фермент, называе-
мый CamK (кальмодулинзависимая киназа), который помогает индуцировать
пластичность, изменяя экспрессию генов. Как показано на рис. 16.50, ионы
кальция входят в клетку, когда рецептор NMDA открывает соответствующий
канал (а также при открытии потенциалзависимых кальциевых каналов). Ионы
кальция связываются с белком кальмодулином (СаМ), который активирует бел-
ки, регулирующие экспрессию генов.
Молекулярные механизмы мозга являются основными мишенями лекарств,
предназначенных для лечения нервных расстройств, таких как нарушения на-
строения, помешательства, инсульт, травмы головного мозга, половые девиа-
ции, кома, нарушения внимания и памяти, бессонница, сонливость и многих
других. Добавьте к этому широкий набор восстановительных медикаментов и
Рис. 16 .50. Зависимость экспрессии гена от активности. NMDAR — NMDA-рецептор.
(Источник: Nelson and Monteggia, из Squire, 2009.)
Гены и молекулы мышления
323
наркотических веществ, влияние которых на мозг зачастую не изучено, и вы
поймете, насколько велико разнообразие психо- и нейроактивных веществ.
Относительно новый класс препаратов, называемых ноотропными (от греч.
ноос — разум и тропос — движение), создан для усиления способности чело-
века к обучению и обострения его внимания. Так что недалек тот день, когда
студенты, вместо того, чтобы употреблять перед экзаменом невероятное коли-
чество кофе, будут пользоваться эффективными и безопасными ноотропными
препаратами.
7.5 . Нейротропные факторы при обучении
При обучении игре на гитаре мотонейроны спинного мозга должны структур-
но усиливаться. Для построения нейрона больших размеров и с более развет-
вленными дендритами необходимо задействовать механизмы роста клетки,
называемые нейротропными факторами (рис. 16.51 и 16.52). Один из таких
факторов носит название BDNF (brain-derived neurotrophic factor). Недавние ис-
следования показали, что BDNF, инъецированный в везикулы мозга грызунов,
Рис. 16 .51. Факторы роста при обучении. Тело поддерживается в работоспособном со-
стоянии веществами, называемыми факторами роста, такими как человеческий гормон
роста. Для обучения важны два фактора роста — нейротропин и BDNF (brain-derived
neurothropic factor). Регуляция высвобождения нейротропина осуществляется скоро-
стью прохождения ПД по нейронам и током нейромедиатора через синапсы. (Источник:
Berardi et al., 2003.)
324
Глава 16
усиливает обучение и продлевает фазы медленного сна. Это соотносится с
представлениями о роли медленного сна в консолидации памяти — считается,
что во время его течения происходят структурные изменения, обусловливаю-
щие долговременную память.
8.0 . Заключение
В этой главе мы рассмотрели способы, при помощи которых основные био-
молекулы — такие как глюкоза, холин или ионы кальция — используются для
нужд нейрональной сигнализации в мозге. Эти «молекулы мышления» участву-
ют в передаче сигналов, которые позволяют нам справляться с обыденными
задачами. Живые организмы используют ферментативный катализ, ускоряю-
щий течение химических реакций. Все нейроактивные вещества действуют
посредством влияния на ферменты. Основные биологические соединения вы-
сококонсервативны — их можно обнаружить у большинства видов животных,
даже очень далеких друг от друга филогенетически. Важно отметить, что эти
вещества способны выполнять несколько различных функций в зависимости от
локализации (табл. 16.3).
Рис. 16.52. Физические упражнения стимулируют образование факторов роста мозга.
Существует баланс между стрессовыми факторами и постоянным обновлением мозга
и тела. Упражнения усиливают образование факторов роста, которые помогают под-
держивать когнитивные функции, синаптическую пластичность, нейрогенез и функ-
ционирование сосудов. Также упражнения снижают повышенное кровяное давление
и инсулинорезистентность. Не слишком интенсивные упражнения помогают снизить
хроническое воспаление. Было показано, что они сокращают риск нейродегенератив-
ных заболеваний, таких как болезни Паркинсона и Альцгеймера. (Источник: Cotman,
Berchtold and Christie, 2007.)
Гены и молекулы мышления
325
Все клетки человеческого тела несут полную копию генома нашего вида в
своих ядрах, представляющую собой двухцепочечную молекулу ДНК, скру-
ченную в хорошо известную каждому двойную спираль. Информация, закоди-
рованная в геноме при помощи генетического кода, экспрессируется в белки,
которые и выполняют большинство функций в клетке. В каждом ядре ДНК
транскрибируется в матричную РНК, которая затем транслируется в белки.
Центральная догма молекулярной биологии гласит, что поток информации в
клетке движется по направлению от ДНК к мРНК и затем к белкам, и никог-
да — назад. (Однако существует явление обратной транскрипции, при котором
по РНК-матрице синтезируется комплементарная цепь ДНК. — Прим. перев.)
Гены и молекулы играют разные роли на разных стадиях развития: они на-
правляют эволюцию мозга — например, переднего — включая язык, социаль-
ные взаимоотношения и способность распознавать выражения лиц. Они кон-
тролируют развитие мозга в течение всей жизни — от роста мозга плода и до
его старения.
Гены нервных клеток также контролируют такие события, как обучение и
засыпание.
Окружающая среда и генетический аппарат взаимодействуют на всех ста-
диях развития мозга, включая пролиферацию, дифференциацию и миграцию,
направление роста аксона и образование синапсов. Экспрессия генов в клетках
мозга человека может меняться в определенных пределах селективным вклю-
чением или выключением участков ДНК. Такой эпигенетический контроль осу-
ществляется в некотором отдалении от описанного в центральной догме потока
информации. Эпигенетический контроль в настоящее время стал объектом при-
стального внимания при изучении процессов обучения.
Мессенджерные молекулы мозга делятся на два типа — медиаторы и мо-
дуляторы. Нейромедиаторы используются для локальной передачи сигнала в
синапсах, хотя таких синапсов могут быть многие миллиарды. На сегодняшний
день известно более 100 нейромедиаторов. Эти вещества находятся под стро-
гим регуляторным контролем: если медиатора в синапсе недостаточно, сигнал
на нем прервется. В то же время избыток медиатора токсичен и способен уби-
вать нервные клетки. Для предотвращения подобного в любом метаболическом
пути существует множество петель отрицательной обратной связи, которые не
дают превысить допустимое количество того или иного вещества.
Глутамат и ГАМК являются двумя наиболее распространенными нейроме-
диаторами. Девяносто процентов нейронов коры головного мозга используют в
качестве медиатора глутамат — возбуждающий медиатор, увеличивающий ве-
ротность прохождения ПД по аксону постсинаптического нейрона при выбро-
се в синаптическую щель. При избытке глутамата развиваются нежелательные
явления, называемые эксайтотоксичностью. Эксайтотоксичность, обусловлен-
ная избытком глутамата, является одним из основных негативных явлений при
повреждениях и заболеваниях мозга. Именно поэтому для нормальной работы
мозга требуется поддерживать уровень глутамата на минимально необходимом
уровне для передачи сигнала.
Глутамат способен запускать два рецепторных каскада, один из которых
задействует NMDA, второй — АМРА-рецепторы. Оба этих рецептора играют
важные роли в процессе обучения. Соотношение NMDA и АМРА определяет
326
Глава 16
тип обучения, который будет происходить в постсинаптическом глутаматер-
гическом нейроне. NMDA ассоциирован с долговременной потенциацией, де-
лающей синапс более возбудимым. АМРА же опосредует долговременную де-
прессию, при которой синапс становится менее возбудимым. Впервые система
долговременной потенциации была детально описана для глутаматергического
синапса, который является самым распространенным возбуждающим синап-
сом мозга. Долговременная потенциация и депрессия требуют синтеза белка
для увеличения числа синапсов и увеличения притока нейроактивных веществ
(чаще всего белковой природы) ко всем частям системы.
Мозг потребляет больше энергии, чем любой другой орган нашего тела. Как
следствие, он производит в процессе работы большое количество свободных
радикалов и создает множество потенциальных очагов воспаления. Хрониче-
ское воспаление является основным фактором дегенеративных заболеваний,
включая и нейродегенеративные, такие как болезни Паркинсона и Альцгейме-
ра. Подавление очагов воспаления совершенно необходимо для поддержания
работы нервной системы. Синапс и внеклеточное пространство наиболее под-
вержены потере контроля за химическим составом, поскольку они не включе-
ны в системы саморегуляции клетки. Очистка синапса является хорошим при-
мером необходимости контроля содержания потенциально токсичных веществ.
В нервной системе существует несколько путей поддержания концентрации
потенциально вредных молекул в синапсе в пределах нормы. Тремя основными
способами являются диффузия, деградация и реутилизация нейромедиатора.
Выше мы отмечали, что действие медиатора носит локальный характер и не
распространяется за пределы отдельного синапса. Нейромодуляторы же влияют
на активность больших областей мозга, хотя и синтезируются в относительно
компактных скоплениях клеток. Эти клетки — обычно их количество находит-
ся в районе нескольких тысяч — простирают свои отростки на очень обширные
зоны мозга. На концах этих отрстков происходит высвобождение модулятора.
Таким образом, нейромодуляторы регулируют активность больших областей
мозга, тогда как медиаторы действуют на уровне отдельных синапсов. Нейро-
модуляция, обусловленная небольшими ядрами базальной части мозга, регули-
рует множество процессов в коре и базальных ганглиях.
Стоит иметь в виду, что одно и то же соединение может выполнять раз-
личные функции в зависимости от места секреции. Ацетилхолин и глутамат
могут быть как нейромедиаторами, так и нейромодуляторами. В качестве ней-
ромодуляторов они могут регулировать активность обширных областей моз-
га, влияя на синаптическую сигнализацию в миллиардах нейронов и синапсов.
К примеру, нейромодуляторы способны изменить потенциал покоя популяции
клеток, модифицировав таким образом в них синаптическую сигнализацию.
Переключением нейромодуляторов между бодрствованием и фазами медлен-
ного и быстрого сна достигается баланс между обработкой сенсорного входа и
внутренними процессами. В бодрствующем состоянии сенсорная информация
обрабатывается в полном объеме, тогда как во время сна ее приток снижен или
отсутствует. Это различие контролируется нейромодуляцией.
Тело поддерживается в работоспособном состоянии при участии веществ,
называемых факторами роста, такими как человеческий гормон роста. Два
таких фактора играют важную роль при обучении — нейротропин и BDNF.
Гены и молекулы мышления
327
Содержание нейротропина контролируется количеством проходящих волн
ПД и высвобождением медиатора на синапсах. Также выработка факторов ро-
ста происходит при физических упражнениях. Постоянное обновление тела и
мозга действует как противовес стрессу и прочим деструктивным факторам.
Упражнения повышают содержание факторов роста, помогая поддерживать
мыслительные функции, синаптическую пластичность, нейрогенез и функци-
онирование сосудов. Также упражнения снижают повышенное кровяное дав-
ление и инсулинорезистентность. Не слишком интенсивные упражнения также
помогают снять хроническое воспаление.
В этой главе мы рассмотрели вещества, делающие возможным проведение
и обработку сигналов, что служит основой человеческого разума. И хотя эта
область сложна, ее понимание поможет по-новому взглянуть на фундаменталь-
ные процессы, лежащие в основе функционирования нашего разумного мозга.
9.0. Задания и упражнения к главе 16
1. Назовите пять высококонсервативных молекул. Каковы их общие функции?
Какую роль они играют в мозге?
2. События с ДНК происходят на протяжении разных по масштабу промежут-
ков времени. Каких именно?
3. Каким образом гены Нох влияют на мозг? Как гены влияют на развитие моз-
га? Какова роль FOXP2?
4. Что называется центральной догмой молекулярной биологии? Какие изме-
нения в соответствии с новыми данными в нее были внесены? Дайте опре-
деление эпигенетических процессов и приведите пример.
5. Информационные вещества в мозге делятся на два класса. Что это за клас-
сы? Как различается их действие?
6. Какое нейроактивное вещество наиболее распространено в коре головного
мозга? А какое занимает второе место по распространенности? Какие функ-
ции они выполняют?
7. Каким образом одна и та же сигнальная молекула может иметь несколько
разных рецепторов? С каким хорошо известным примером этого вы позна-
комились?
8. Каким образом нормальная передача сигналов может привести к наруше-
нию функций мозга?
9. Какие два типа «событий обучения» известны для синаптического уровня?
Какие уровни молекулярных механизмов клетки считаются вовлеченными в
каждый из них?
10. Какие два типа клеток вовлечены в обработку информации в мозге?
328
Глава 16
Т
а
б
л
и
ц
а
1
6
.
3
П
р
о
ч
и
е
н
е
й
р
о
м
е
д
и
а
т
о
р
ы
К
л
а
с
с
в
е
щ
е
с
т
в
а
П
р
и
м
е
р
К
л
е
т
к
и
п
р
о
и
с
-
х
о
ж
д
е
н
и
я
М
е
с
т
о
х
р
а
н
е
н
и
я
В
ы
с
в
о
б
о
ж
д
е
н
и
е
и
п
р
о
ч
и
е
м
е
х
а
н
и
з
м
ы
с
и
г
н
а
л
и
з
а
ц
и
и
И
н
а
к
т
и
в
а
ц
и
я
Р
е
ц
е
п
т
о
р
ы
А
ц
е
т
и
л
х
о
л
и
н
А
ц
е
т
и
л
х
о
-
л
и
н
Н
е
й
р
о
н
ы
В
е
з
и
к
у
л
ы
Д
е
п
о
л
я
р
и
з
а
ц
и
я
Ф
е
р
м
е
н
т
а
т
и
в
н
а
я
(
а
ц
е
т
и
л
х
о
л
и
н
э
с
т
е
-
р
а
з
о
й
)
И
о
н
о
т
р
о
п
н
ы
е
(
н
и
к
о
т
и
н
о
в
ы
е
)
и
с
в
я
з
а
н
н
ы
е
с
G
-
б
е
л
к
а
м
и
(
м
у
-
с
к
а
р
и
н
о
в
ы
е
)
А
м
и
н
о
к
и
с
л
о
-
т
ы
Г
л
у
т
а
м
а
т
Н
е
й
р
о
н
ы
В
е
з
и
к
у
л
ы
Д
е
п
о
л
я
р
и
з
а
ц
и
я
Б
е
л
к
и
-
п
е
р
е
н
о
с
ч
и
к
и
И
о
н
о
т
р
о
п
н
ы
е
и
с
в
я
з
а
н
н
ы
е
с
G
-
б
е
л
к
а
м
и
М
о
н
о
а
м
и
н
ы
Д
о
ф
а
м
и
н
Н
е
й
р
о
н
ы
В
е
з
и
к
у
л
ы
Д
е
п
о
л
я
р
и
з
а
ц
и
я
Б
е
л
к
и
-
п
е
р
е
н
о
с
ч
и
к
и
С
в
я
з
а
н
н
ы
е
с
G
-
б
е
л
к
а
м
и
П
е
п
т
и
д
ы
Э
н
к
е
ф
а
-
л
и
н
Н
е
й
р
о
н
ы
В
е
з
и
к
у
л
ы
,
ч
а
с
т
о
о
т
-
д
е
л
ь
н
о
о
т
н
е
б
е
л
к
о
в
ы
х
н
е
й
р
о
м
е
д
и
а
т
о
р
о
в
П
е
р
и
о
д
и
ч
е
с
к
а
я
д
е
п
о
л
я
р
и
-
з
а
ц
и
я
Ф
е
р
м
е
н
т
а
т
и
в
н
а
я
(
д
л
я
м
н
о
г
и
х
п
е
п
т
и
-
д
о
в
с
о
о
т
в
е
т
с
т
в
у
ю
-
щ
и
е
ф
е
р
м
е
н
т
ы
и
л
и
д
р
у
г
и
е
м
е
х
а
н
и
з
м
ы
н
е
и
з
в
е
с
т
н
ы
)
С
в
я
з
а
н
н
ы
е
с
G
-
б
е
л
к
а
м
и
р
е
ц
е
п
т
о
р
ы
μ
и
δ
,
о
п
и
а
т
н
ы
е
р
е
ц
е
п
т
о
р
ы
D
-
а
м
и
н
о
-
к
и
с
л
о
т
ы
D
-
с
е
р
и
н
А
с
т
р
о
ц
и
-
т
ы
(
г
л
и
я
)
Н
е
и
з
в
е
с
т
н
о
А
к
т
и
в
а
ц
и
я
с
е
р
и
н
о
в
о
й
р
а
ц
е
м
а
з
ы
п
о
с
л
е
в
ы
с
в
о
-
б
о
ж
д
е
н
и
я
D
-
с
е
р
и
н
а
п
р
и
п
о
м
о
щ
и
н
е
и
з
в
е
с
т
н
ы
х
м
е
-
х
а
н
и
з
м
о
в
Б
е
л
к
и
-
п
е
р
е
н
о
с
ч
и
к
и
М
о
д
у
л
я
т
о
р
н
ы
й
(
г
л
и
ц
и
н
о
в
ы
й
)
с
а
й
т
н
а
р
е
ц
е
п
-
т
о
р
е
N
M
D
A
Гены и молекулы мышления
329
К
л
а
с
с
в
е
щ
е
с
т
в
а
П
р
и
м
е
р
К
л
е
т
к
и
п
р
о
и
с
-
х
о
ж
д
е
н
и
я
М
е
с
т
о
х
р
а
н
е
н
и
я
В
ы
с
в
о
б
о
ж
д
е
н
и
е
и
п
р
о
ч
и
е
м
е
х
а
н
и
з
м
ы
с
и
г
н
а
л
и
з
а
ц
и
и
И
н
а
к
т
и
в
а
ц
и
я
Р
е
ц
е
п
т
о
р
ы
П
у
р
и
н
ы
А
д
е
н
о
з
и
н
Н
е
й
р
о
н
ы
,
в
о
з
м
о
ж
н
о
,
д
р
у
г
и
е
к
л
е
т
к
и
А
Т
Ф
х
р
а
н
и
т
с
я
в
в
е
з
и
-
к
у
л
а
х
и
м
о
ж
е
т
б
ы
т
ь
г
и
д
р
о
л
и
з
о
в
а
н
а
д
о
а
д
е
-
н
о
з
и
н
а
п
о
с
л
е
в
ы
с
в
о
-
б
о
ж
д
е
н
и
я
;
в
ы
с
о
к
и
й
у
р
о
в
е
н
ь
в
н
е
к
л
е
т
к
и
с
о
-
о
т
в
е
т
с
т
в
у
е
т
н
е
с
о
х
р
а
-
н
я
е
м
о
м
у
а
д
е
н
о
з
и
н
у
(
1
)
В
ы
с
в
о
б
о
ж
д
е
н
и
е
и
з
в
н
е
-
к
л
е
т
о
ч
н
ы
х
п
у
р
и
н
о
в
э
к
з
о
-
ф
е
р
м
е
н
т
а
м
и
(
2
)
К
о
н
т
р
о
л
и
р
у
е
м
а
я
д
и
ф
-
ф
у
з
и
я
и
з
к
л
е
т
к
и
Б
е
л
к
и
-
п
е
р
е
н
о
с
ч
и
к
и
С
в
я
з
а
н
н
ы
е
с
G
-
б
е
л
к
а
м
и
Г
а
з
ы
N
O
Н
е
й
р
о
н
ы
Н
е
х
р
а
н
я
т
с
я
К
а
л
ь
ц
и
й
з
а
в
и
с
и
м
а
я
р
е
г
у
л
я
-
ц
и
я
с
и
н
т
а
з
ы
о
к
с
и
д
а
а
з
о
т
а
,
в
р
е
з
у
л
ь
т
а
т
е
ч
е
г
о
п
р
о
и
с
-
х
о
д
и
т
с
и
н
т
е
з
о
к
с
и
д
а
а
з
о
т
а
,
к
о
т
о
р
ы
й
н
е
м
е
д
л
е
н
н
о
в
ы
-
с
в
о
б
о
ж
д
а
е
т
с
я
Д
и
ф
ф
у
з
и
я
.
В
о
з
м
о
ж
-
н
о
,
ф
е
р
м
е
н
т
а
т
и
в
н
а
я
д
е
г
р
а
д
а
ц
и
я
М
н
о
г
и
е
ф
е
р
-
м
е
н
т
ы
,
с
о
д
е
р
-
ж
а
щ
и
е
п
е
р
е
х
о
д
-
н
ы
е
м
е
т
а
л
л
ы
,
т
а
к
и
е
к
а
к
г
у
а
-
н
и
л
а
т
ц
и
к
л
а
з
а
Л
и
п
и
д
ы
А
н
а
д
а
м
и
н
Н
е
й
р
о
н
ы
Н
е
и
з
в
е
с
т
н
о
;
п
о
в
с
е
й
в
и
д
и
м
о
с
т
и
,
н
е
х
р
а
н
я
т
-
с
я
В
о
з
м
о
ж
н
о
,
в
ы
с
в
о
б
о
ж
д
а
-
ю
т
с
я
в
с
и
н
а
п
т
и
ч
е
с
к
у
ю
щ
е
л
ь
,
о
д
н
а
к
о
м
е
х
а
н
и
з
м
д
и
ф
ф
у
з
и
и
в
в
о
д
н
о
й
с
р
е
д
е
н
е
я
с
е
н
Т
р
а
н
с
п
о
р
т
с
п
о
с
л
е
-
д
у
ю
щ
и
м
г
и
д
р
о
л
и
-
з
о
м
С
в
я
з
а
н
н
ы
е
с
G
-
б
е
л
к
а
м
и
(
к
а
н
-
н
а
б
и
н
о
и
д
н
ы
е
)
(
1
)
Д
а
н
н
ы
е
п
р
и
м
е
р
ы
н
е
п
о
л
н
о
с
т
ь
ю
м
о
г
у
т
б
ы
т
ь
п
р
и
м
е
н
е
н
ы
к
с
о
о
т
в
е
т
с
т
в
у
ю
щ
е
м
у
к
л
а
с
с
у
.
П
у
р
и
н
о
в
о
е
с
о
е
д
и
н
е
н
и
е
а
д
е
н
о
з
и
н
т
р
и
ф
о
с
ф
а
т
(
А
Т
Ф
)
,
н
е
с
м
о
т
р
я
н
а
п
о
в
с
е
м
е
с
т
н
о
с
т
ь
и
с
п
о
л
ь
з
о
в
а
н
и
я
е
г
о
к
а
к
п
е
р
е
н
о
с
я
щ
у
ю
э
н
е
р
г
и
ю
м
о
л
е
к
у
л
у
,
м
о
ж
е
т
п
е
р
е
д
а
в
а
т
ь
и
и
н
ф
о
р
м
а
ц
и
ю
,
н
а
-
х
о
д
и
т
с
я
в
в
е
з
и
к
у
л
а
х
с
о
в
м
е
с
т
н
о
с
д
р
у
г
и
м
и
н
е
й
р
о
м
е
д
и
а
т
о
р
а
м
и
(
а
ц
е
т
и
л
х
о
л
и
н
о
м
и
л
и
д
о
ф
а
м
и
н
о
м
)
и
в
ы
с
в
о
б
о
ж
д
а
е
т
с
я
п
р
и
д
е
п
о
л
я
р
и
з
а
ц
и
и
,
т
о
г
д
а
к
а
к
п
р
и
н
а
д
л
е
ж
а
щ
и
й
к
п
у
р
и
н
а
м
а
д
е
н
о
з
и
н
в
ы
с
в
о
б
о
ж
д
а
е
т
с
я
о
б
л
е
г
ч
е
н
н
о
й
д
и
ф
ф
у
з
и
е
й
.
А
Т
Ф
в
з
а
и
м
о
д
е
й
с
т
в
у
е
т
к
а
к
с
и
о
н
о
т
р
о
п
н
ы
м
и
(
Р
2
Х
-
р
е
ц
е
п
т
о
р
а
м
и
)
,
т
а
к
и
р
е
ц
е
п
т
о
р
а
м
и
,
с
в
я
з
а
н
н
ы
м
и
с
G
-
б
е
л
к
а
м
и
(
P
2
Y
-
р
е
ц
е
п
т
о
р
а
м
и
)
,
т
о
г
д
а
к
а
к
а
д
е
н
о
з
и
н
в
о
з
д
е
й
с
т
в
у
е
т
т
о
л
ь
к
о
н
а
с
в
я
з
а
н
-
н
ы
е
с
G
-
б
е
л
к
а
м
и
р
е
ц
е
п
т
о
р
а
м
и
.
(
2
)
К
а
к
у
п
о
м
я
н
у
т
о
в
т
е
к
с
т
е
,
г
л
и
я
м
о
ж
е
т
,
в
н
е
к
о
т
о
р
ы
х
с
л
у
ч
а
я
х
,
б
ы
т
ь
н
а
п
р
я
м
у
ю
в
о
в
л
е
ч
е
н
а
в
н
е
й
р
о
п
е
р
е
д
а
ч
у
:
н
а
п
р
и
м
е
р
и
н
а
к
т
и
в
и
р
у
я
н
е
й
р
о
м
е
д
и
а
т
о
р
,
к
а
к
г
л
у
т
а
м
а
т
,
и
л
и
в
ы
п
у
с
к
а
я
А
Т
Ф
и
л
и
d
-
с
е
р
и
н
.
Содержание
1.0 . Исторические предпосылки ........................................ 331
1.1 . Корреляция мозга и разума ..................................... 331
1.2 . Регистрация активности мозга .................................. 333
2.0 . Методы ......................................................... 335
2.1 . Электроэнцефалография ....................................... 335
2.2.Магнитоэнцефалография......................................344
2.3 . Позитронно-эмиссионная томография ........................... 350
2.4 . Магнитно-резонансная томография ............................. 353
2.5 . МРТ – метод будущего ........................................ 370
2.6 . Оптическое картирование ...................................... 372
3.0 . Мультимодальные методы визуализации мозга ........................ 374
3.1 . Одновременная визуализация разными методами ................. 374
3.2 . Визуализация генетики ........................................ 376
4.0 . Заключение ..................................................... 379
1.0 .
Исторические предпосылки
1.1 .
Корреляция мозга и разума
Понимание сложных функций мозга, включая сенсорное восприятие, двига-
тельные функции, сознание и мышление, всегда вызывало наше любопытство.
Мозг активируется, когда мы вовлечены в процессы решения задачи, например
размышление, разговор или целенаправленные действия. Активация сопрово-
ждается локальными изменениями мозгового кровообращения в активирован-
ной, т. е. задействованной, области мозга, и это лежит в основе работ по функ-
циональному картированию мозга. В данном разделе мы дадим краткий обзор
некоторых важных направлений в этой области.
Давно было замечено, что разрушение большого полушария мозга приводит
к контралатеральному параличу, потери способности двигать частью тела. Если
разрушение было локализовано в левом полушарии, могли возникнуть пробле-
мы с речью. В XIX и начале XX вв. функциональная организация мозга изуча-
лась на базе клинико-патологических случаев. Подробное описание симптомов
впоследствии (после смерти пациента) соотносилось с результатами вскрытия,
что позволяло определить локализацию определенных функций в мозге. Основ-
ным ограничением этого метода является то, что повреждения мозга возникали
в результате несчастных случаев, а значит, неспецифичные по локализации, так
как повреждения диффузные и, как правило, затрагивают области, незначи-
Приложение
Методы для изучения
живого мозга
Томас Рамсой, Даниэла Балслев и Олаф Паульсон
332
Приложение
мые для исследуемого неврологического заболевания. Более того, адаптивные
способности мозга могут изменять с течением времени неврологические рас-
стройства. При помощи клинико-патологических корреляций мы узнаем о на-
рушениях, которые возникают при повреждении определенной области мозга,
т. е. какую функцию выполняет данная область мозга. Например, изучение раз-
рушений выявило участие зон Брока и Вернике в нормальной речевой функции.
Эти ограничения обеспечили стимул для параллельного развития других
методов, которые позволяют проводить систематические исследования взаи-
мосвязи между мозгом и поведением у человека. Новые методы нейровизуали-
зации, предложенные в конце ХХ в., измеряют изменения мозговой активности,
которые коррелируют с изменениями в поведении. Современное оборудование
для нейровизуализации позволяет регистрировать изменения активности в объ-
емах мозга менее 1 мм3, происходящие за десятки миллисекунд (рис. А.1). По-
пулярность новых методов визуализации характеризуется постоянным увели-
чением их доли в объеме всех исследовательских работ. Например, в 2005 г.
каждое пятое опубликованное исследование было зарегистрировано в PubMed
под ключевыми словами ПЭТ и МРТ (рис. А .2).
Клинико-патологическая корреляция и новые методы визуализации мозга
дают информацию о функциональной организации мозга, но фундаменталь-
ные различия разделяют эти методы. Так, клинико-патологическая корреляция
Рис. А.1 . За и против различных методов визуализации мозга. Различные методы ре-
гистрации активности мозга имеют разную разрешающую способность. В то время
как одни методы обладают высокой пространственной разрешающей способностью,
но низкой временной, другие имеют противоположные свойства. (Источник: Thomas
Ramsøy, Daniela Balslev and Olaf Paulson, с разрешения.)
Методы для изучения живого мозга
333
позволяет выяснить, какие нарушения возникают, если определенная область
мозга травмирована, т. е . для выполнения каких функций данная область пред-
назначена. В противоположность этому, ПЭТ и фМРТ (функциональная МРТ)
демонстрируют активацию областей мозга, задействованных в выполнении
задания, по сравнению с контрольными условиями. Таким образом, из клини-
ко-патологических корреляций мы знаем, что зоны Брока и Вернике являются
необходимыми для нормальной функции речи, однако сенсорные и моторные
области, вовлеченные в процессы артикуляции, также будут активированы.
По этим причинам усложненные конструкции экспериментальных парадигм
становятся неотъемлемой частью современных работ по картированию мозга.
Состояние выполнения задания и контрольное состояние должны, в принци-
пе, быть идентичными, кроме наличия единичной психологической функции,
локализацию которой и изучают в данном эксперименте. На практике очень
сложно, если не невозможно, полностью контролировать все поведенческие
параметры. Так, например, в парадигме, разработанной для выявления нейрон-
ных коррелятов пространственного внимания и основанной на предъявлении
зрительных стимулов либо в подсказанной точке экрана либо нет, изменения
активности мозга могут отражать непроизвольные движения глазами или уси-
лие испытуемого для предотвращения саккад в направлении стимула.
1.2.
Регистрация активности мозга
Мозг активируется, когда мы вовлечены в решение задачи, размышляем, гово-
рим или совершаем целенаправленное движение и т. д. Такая активация сопро-
вождается изменением в гемодинамике в активированной области мозга. Это
создает основу для некоторых исследований по функциональному картирова-
нию мозга, включая доминирующие в настоящее время методы фМРТ. Далее в
этом разделе будет дан краткий обзор нескольких основополагающих методов
в этой области.
Изменения в гемодинамики во время сенсорных, моторных и когнитив-
ных процессов впервые наблюдал Массо около 130 лет назад в Италии (Masso,
1881). У него был пациент с дефектом черепа, позволяющим проводить наруж-
ные наблюдения за «пульсацией мозга». Он наблюдал изменения в пульсации
мозга не только в ответ на звон церковных колоколов, но и когда пациент читал
молитву, без произнесения ее вслух. Примерно через 10 лет было выдвинуто
Рис. А.2 . Впечатляющее увеличение применения методов нейровизуализации в иссле-
дованиях мозга. В 2005 г. каждая пятая опубликованная работа была зарегистрирована
в PubMed под ключевыми словами ПЭТ или МРТ. (Источник: Thomas Ramsøy, Daniela
Balslev and Olaf Paulson, с разрешения.)
334
Приложение
предположение о существовании связи между функциями мозга и его кровос-
набжением (циркуляцией крови) (Roy, Sherrington, 1890). Эта теория получила
доказательство и сформировала основу для интенсивных исследований.
В конце 1920-х гг. Фултон (�ulton, 1928) наблюдал пациента с внутричереп-
�ulton, 1928) наблюдал пациента с внутричереп-
1928) наблюдал пациента с внутричереп-
ным артериовенозным пороком развития — состоянием, при котором кровь на-
прямую из артериальной кровеносной системы переходит в венозную систему
без прохождения через систему капилляров. Пациент жаловался на странный
звук (шум) в голове, который становился громче во время чтения. Шум и его
усиление можно было зарегистрировать. Это свидетельствовало о том, что ге-
модинамические изменения некого рода возникали при чтении (по крайней
мере, у этого пациента). В конце 1950-х гг. было показано, что стимуляция
ствола мозга вызывает появление паттернов активации в ЭЭГ (электроэнцефа-
лограмме), а также увеличение кровоснабжения мозга (Ingvar and Söderberg,
1956, 1958).
В середине 1960-х гг. Купер (Cooper et al., 1966) наблюдал локальное уве-
личение парциального давления кислорода в мозге при активации у пациентов
во время хирургических операций. Это исследование еще сильнее подтвердило
гипотезу, что гемодинамика изменяется вследствие мозговой активации. Более
того, это была первая работа, проиллюстрировавшая изменения во взаимосвязи
кровотока и метаболизма во время активации.
Прорыв в исследованиях локализации мозговых функций был совершен в
начале 1960-х гг., когда был предложен метод внутриартериальных инъекций
инертных газов с радиоактивной меткой. В одной из работ с поверхности от-
крытого мозга у экспериментальных животных измеряли разрушение изотопа
криптона85 (�assen and Ingvar, 1961). Позже этот метод был адаптирован для ис-
�assen and Ingvar, 1961). Позже этот метод был адаптирован для ис-
and Ingvar, 1961). Позже этот метод был адаптирован для ис-
and Ingvar, 1961). Позже этот метод был адаптирован для ис-
Ingvar, 1961). Позже этот метод был адаптирован для ис-
Ingvar, 1961). Позже этот метод был адаптирован для ис-
, 1961). Позже этот метод был адаптирован для ис-
следований на людях с применением гамма-излучающих изотопов ксенона133,
который мог быть зарегистрирован через череп. Используя многочисленные
внешние детекторы импульсов (сенсоры, измеряющие ионизированную ради-
ацию), стало возможно измерять кровоток в более чем в 250 областях. Этот
метод интенсивно применялся в 1970-х гг. для функциональной локализации
областей мозга и был, на самом деле, первым методом для функционального
картирования мозга (�assen et al., 1978). Одним недостатком метода являлась
необходимость введения метки непосредственно во внутреннюю сонную арте-
рию, что ограничивало исследование кругом пациентов, проходящих ангиогра-
фическое обследование в диагностических целях.
В 1970-х гг. (Sokoloff et al., 1977) был представлен метод С14-дезоксиглюкозы
для измерения метаболизма глюкозы в интересующих областях мозга у экс-
периментальных животных. Позже данный метод был переделан для ПЭТ-
исследований на людях с применением фторированной дезоксиглюкозы (ФДГ),
меченной позитрониспускающим �18
. Хорошо известно, что изменения в кро-
вотоке и в метаболизме глюкозы сцеплены. Но, несмотря на то что метод изме-
рения локального метаболизма глюкозы в мозге является значительным дости-
жением, он не применяется широко при изучении функциональной мозговой
активации по причине ограниченного временного разрешения (15–45 мин) по
сравнению с ПЭТ и фМРТ, которые будут рассмотрены ниже.
Введение в 1980-х гг. ПЭТ и в 1990-х гг. фМРТ позволило проводить неин-
вазивные исследования локальных изменений в гемодинамике мозга как прояв-
Методы для изучения живого мозга
335
ления изменений в локальной активности мозга. Нетравматичная природа этих
методов позволяет проводить работы не только на пациентах, но и на нормаль-
ных здоровых испытуемых, что позволяет расширить исследовательские рабо-
ты по изучению функциональной активации мозга.
2.0 .
Методы
ПЭТ и фМРТ являются доминирующими методами исследования функцио-
нальной мозговой активации (рис. А.3). Однофотонная эмиссионная компью-
терная томография (ОЭКТ) имеет сходные с ПЭТ свойства, но обладает более
низкой разрешающей способностью, в связи с чем менее пригодна для из-
учения функциональной активации. Другие методы основаны на регистрации
электрических и магнитных сигналов работающего мозга, образуемые актив-
ностью нейронов. К этим методам относят электроэнцефалографию (ЭЭГ) и
магнитоэнцефалографию (МЭГ), которые обладают превосходной временной
разрешающей способностью, равной около 10 мс, но ограниченной способно-
стью для пространственной локализации. Исследования с помощью ЭЭГ стали
особенно интересными, когда их стали совмещать с фМРТ, потому что объеди-
няются преимущества: хорошая пространственная разрешающая способность
фМРТ и хорошее временное разрешение ЭЭГ. МЭГ относится к интенсивно
расширяющейся области исследований.
Все эти методы нацелены на изучение поведения и измеряют изменения
мозговой активности. Дополнительная категория методов, включая работы по
разрушению или транскраниальной магнитной стимуляции (ТМС), позволяет
проводить манипуляции с мозгом для выявления их эффектов на поведение.
ТМС создает электрический ток в мозге, который, в зависимости от параме-
тров, либо возбуждает, либо тормозит активность нейронов. Используя кон-
фигурацию «виртуального разрушения», этим методом можно снижать возбу-
димость определенных областей мозга, что приводит к имитации разрушений
мозга. ТМС — многообещающее средство для подтверждения взаимосвязи
между мозгом и поведением, выявленной с помощью функциональной нейро-
визуализации. Если активированная область мозга является основополагающей
для выполнения задания, то ее инактивация изменит поведение субъекта при
выполнении этого задания.
2.1.
Электроэнцефалография
Электроэнцефалография — нейрофизиологическое измерение электрической
активности в мозге. Это измерение проводят, располагая регистрирующие
электроды либо на поверхности головы, либо непосредственно на поверхно-
сти коры мозга. Записываемая таким образом активность была впервые назва-
на электроэнцефалограммой (ЭЭГ) немецким психиатром Хансом Бергером
(Berger and Gloor, 1969). До него работы по ЭЭГ технологиям были выполнены
Ричардом Катоном (�aton, 1875) и Владимиром Владимировичем Правдич-Не-
�aton, 1875) и Владимиром Владимировичем Правдич-Не-
, 1875) и Владимиром Владимировичем Правдич-Не-
минским (Pravdich-Neminsky, 1913). Сегодня ЭЭГ применяют в клинических
обследованиях для выявления мозговых повреждений, эпилепсии и других
мозговых расстройств. Во многих административно-территориальных едини-
цах по всему миру ЭЭГ используют для констатации смерти мозга. ЭЭГ также
используют в когнитивных исследованиях из-за его высокой временной разре-
336
Приложение
шающей способности, и результаты таких работ часто сравнивают с результа-
тами, полученными другими методами.
Амплитуда ЭЭГ бодрствующих и здоровых испытуемых имеет разброс от
нескольких мкВ до примерно 75 мкВ. ЭЭГ сигнал, по существу, формируется
градуированными постсинаптическими потенциалами, возникающими в телах
нейронов и больших дендритах. Пирамидные нейроны корковых слоев III–V
вносят основной вклад в сигнал, и активность этих нейронов синхронизируется
ритмическими разрядами таламических ядер. От степени синхронизации кор-
ковой активности зависит амплитуда ЭЭГ. Неудивительно, что большая часть
сигнала, зарегистрированного в ЭЭГ, исходит из внешних слоев коры, располо-
женных рядом с электродом, а также складчатое строение коры вносит больший
вклад в электрическую суммацию сигнала, нежели во взаимопогашение.
Рис. А.3 . BO�D-сигнал в ответ на стиму-
ляцию пальца. Когда палец правой руки
стимулируют слабым электрическим то-
ком, изменения кровотока можно зареги-
стрировать в левом полушарии в сомато-
сенсорном корковом представительстве
тела. Это зависящая от уровня кислорода
в крови активность или BO�D-сигнал в
фМРТ. (а) Усредненные изображения моз-
га. (б) Изменения активности, зависящей
от уровня кислорода в крови (гемодинами-
ческий ответ), после нервной активации.
(Источник: Mc�onigle в �rackowiak et al.,
2004.)
Методы для изучения живого мозга
337
ЭЭГ, зарегистрированная с поверхности черепа, представляет собой пассив-
ное проведение токов, вызванных суммацией активности большого скопления
нейронов. Локальная десинхронизация в ЭЭГ является результатом увеличения
взаимодействия в подгруппе нейронов, вовлеченных в «совместную работу», и
выражается в понижении амплитуды сигнала. Таким образом, по нативной за-
писи ЭЭГ, даже нетренированным глазом, можно определить уровень синхро-
низации. Профессиональный взгляд также способен выделить патологические
паттерны, присущие таким состояниям, как эпилепсия. Как вы можете сами
увидеть на рис. А .4, в нативной записи ЭЭГ можно выделить основные разли-
чия в нормальной и патологической активности мозга.
Рис. А.4. Нативная запись ЭЭГ во время разных состояний. (а) Запись ЭЭГ здорового
и бодрствующего испытуемого, на которой можно выявить разные низкоамплитудные и
высокочастотные ритмы активации, отражающие стремительные взаимодействия меж-
ду группами нейронов. (б) При засыпании мозг может находиться на разных уровнях
сна, от сна с быстрыми движениями глаз (БДГ-сон) до стадии глубокого сна. Стадия
глубокого сна характеризуется высокоамплитудными и низкочастотными колебаниями,
которые свидетельствуют о снижении взаимодействия между областями мозга. (в) И на-
конец, эпилептические припадки на ЭЭГ отображаются как высокоамплитудными, так
и относительно высокочастотными паттернами, во всех регистрируемых отведениях.
Это указывает на то, что большая часть мозга оказывается вовлеченной в припадок. Та-
кой паттерн активности служит указанием областей, вовлеченных во взаимную петлю
активации, которая, в конечном итоге, приводит к хаосу и неинформативным процессам
в мозге. (Источник: Thomas Ramsøy, Daniela Balslev and Olaf Paulson, с разрешения.)
338
Приложение
В общем, мы можем рассматривать записи ЭЭГ двумя способами. Во-
первых, спонтанная активность — активность, которая протекает постоянно у
живых индивидуумов и регистрируется с поверхности черепа или непосред-
ственно с коры. Измерения таких сигналов называют энцефалограммой, кото-
рая является результатом измерения электрических сигналов в определенном
временном окне, но без учета дополнительных временных факторов. Амплиту-
да ЭЭГ составляет около 100 мкВ, если регистрируется от поверхности черепа,
и равна около 1–2 мВ при измерении от поверхности коры (внутричерепная
регистрация). Диапазон частот сигнала составляет от значений меньше 1 Гц
примерно до 50 Гц. На рис. А.4 представлены разные виды спонтанной ЭЭГ-ак -
тивности. Сегодня ЭЭГ интенсивно применяют в клинике, особенно для выяв-
ления эпилепсии, но с недавнего времени интерес к этой методике возрастает в
связи с комбинацией ЭЭГ с другими методами визуализации, имеющими более
высокую пространственную разрешающую способность.
Вторым способом обработки ЭЭГ записи является выделение вызванных
потенциалов (ВП) и связанных с событием потенциалов (ССП), т. е . компо-
нентов ЭЭГ, которые возникают в ответ на стимул (например, слуховой, сома-
тосенсорный или зрительный вход). Эти ответы, как правило, находятся ниже
уровня шума и, их обычно невозможно выделить в нативной записи ЭЭГ. Для
того чтобы увидеть эффекты от стимулов, необходимо предъявить один сти-
мул несколько раз, а затем усреднить ответную активность по этим эпохам.
Таким образом, сигналы, зарегистрированные с момента предъявления стиму-
лов, группируются в одну категорию. Усреднение электрических сигналов по
времени нивелирует спонтанные флуктуации в ЭЭГ, и становится возможным
выявить и отобразить ответный сигнал во временном окне, свойственном всем
предъявлениям стимула. Так, для зрительных стимулов мы можем выделить
несколько волн увеличения и уменьшений интенсивности сигнала (рис. А .5).
Каждый компонент на сегодняшний день идентифицирован как значимый кор-
релят когнитивных процессов. Например, при регистрации ЭЭГ у читающего
слова испытуемого можно выявить ранние компоненты, отражающие ранние
процессы обработки зрительной информации и процессы привлечения внима-
ния, и поздний пик после 400 мс, который отражает обработку семантической
информации. Этот компонент называется Р400, где Р — позитивная волна сиг-
нала.
Примером хорошо известного компонента ССП является негативность рас-
согласования (НР), которую также иногда называют полем рассогласования.
Представьте, что вы слушаете поток одинаковых звуковых стимулов, и неожи-
данно звук меняется. Мозг отвечает на этот девиантный звук, отличающийся
по высоте, длительности или громкости, независимо от того, сосредоточен ли
испытуемый на последовательности стимулов.
На записи ССП этот компонент возникает с латентным периодом в 150–250 мс
после девиантного стимула и генерируется в первичной и не первичной слу-
ховой коре. Несмотря на то что точная природа компонента НР еще не опре-
делена, считается, что он отражает автоматический и ранний ответ на нару-
шение автоматически сформированной кратковременной нервной модели или
памятного следа о физических или абстрактных свойствах окружающей сре-
ды (Näätänen and Winkler, 1999; Näätänen, Paavilainen, Rinne, and Alho, 2007).
Методы для изучения живого мозга
339
Рис. А.5. Нативная ЭЭГ и два способа ее обработки. (а) Расположение регистрирующих
электродов на определенных точках черепа согласно специальной системе. (б) Запись с
каждого электрода, или отведения, может быть представлена в виде отдельных кривых
линий во времени, которые и формируют нативную запись ЭЭГ. События, как предъ-
явление слуховых стимулов, могут быть наложены на временную ось, что дает пред-
ставление о динамике ЭЭГ, а также служит для усреднения данных. События на рисунке
показаны только для иллюстрации. (в) Анализ ССП выполнен при помощи усреднения
нативных записей ЭЭГ относительно момента предъявления стимулов. При усреднении
спонтанная активность взаимно уничтожается, в то время как изменения сигнала в от-
вет на предъявление стимулов будут видимы. Таким образом, можно выделить несколь-
ко компонентов. Они включают ранние позитивные и негативные волны, которые отра-
жают процессы активации подкорковых структур (например, ствола мозга), и поздние
волны, которые вовлечены в более сложные, когнитивные процессы. Примером такого
компонента служит Р400 (Р400 — позитивный пик с латентным периодом около 400 мс
от предъявления стимула, не показан на рисунке), который считается индикатором се-
мантической обработки. И наконец, современные разработки позволяют одновременно
анализировать данные о частотных характеристиках ЭЭГ и пространственной локали-
зации таких компонентов. (г) Топография распределения спектра мощности ЭЭГ при
спокойном бодрствовании. Можно видеть, что дельта-активность локализуется в пре-
фронтальной части мозга с максимальным фокусом в A�8 отведении. Тета-активность
регистрируется по средней линии в лобных отделах с максимальной мощностью в ��z
отведении. Альфа-активность распределена по средней линии в теменно-затылочной
области с максимумом в точке PO8. Альфа-активность локализуется в затылочной об-
ласти с максимумом, зарегистрированным в отведении PO4. Бета-активность широко
распространена по поверхности головы, но можно выделить максимум в отведении
��z. Гамма-активность локализуется билатерально в височных областях с максимумом
в точке ��T7h. (Источник: (а) и (г) Andrew et al., 2006; (б) и (в) авторы главы, с разре-
шения.)
340
Приложение
Блок А.1 . Электромагнетизм при медитации
Была проведена очень интересная работа по исследованию мозговой активности у
медитирующих буддистов (�utz et al., 2004), с использованием регистрации ЭЭГ.
В данной работе ЭЭГ регистрировали у группы профессионально медитирующих
людей и незнакомых с медитацией членов контрольной группы во время спокойно-
го бодрствования и разных стадий медитации. Использовали три вида состояний:
стандартное бодрствование, состояние медитации и стадию паузы между разными
стадиями медитации. Авторы работы выявили, что во время медитации в группе про-
фессионально медитирующих испытуемых чрезвычайно сильно повышен уровень
гамма-активности. Также исследователи отметили фазовую синхронизацию между
отдаленными лобными и теменными областями мозга. Из этих открытий сделан вы-
вод, что медитация улучшает взаимосвязь отдаленных областей мозга. Интересно,
но результаты также показали, что активность мозга в состоянии покоя до медита-
ции различалась у профессионалов и наивных испытуемых. Это свидетельствует в
пользу того, что солидный опыт медитации может изменять работу мозга, хотя на
данный момент мы можем только делать предположения на счет исходной причины
и следствия (рис. А .6).
Рис. А.6 . Измерения актив-
ности мозга при медитации:
показано, что абсолютная
мощность гамма-активности
при мысленном напряжении
значительно больше у про-
фессионально
медитирую-
щих испытуемых (справа)
по сравнению с немедити-
рующими
контрольными
субъектами (слева). Цветовая
шкала отражает процент ис-
пытуемых в каждой группе,
у которых отмечено увеличе-
ние гамма-активности в со-
стоянии мыслительной актив-
ности. (Источник: �utz et al.,
2004.)
Методы для изучения живого мозга
341
В качестве альтернативы, также возможно, что НР отражает изменения в ней-
ронных сетях, т. е . ответ новых вовлеченных сетей, по сравнению с сетями,
которые обрабатывали повторяющийся стимул, оказывается сильнее (Näätänen,
1992; Jääskeläinen et al., 2004).
Метод ССП демонстрирует, как можно идентифицировать определенные
паттерны активации в мозге на основании изменений во времени интенсив-
ности сигнала. Несмотря на то что метод ЭЭГ обладает высокой временной
разрешающей способностью, но очень низким пространственным разрешени-
ем, непрекращающееся совершенствование технологий значительно улучши-
ло пространственную разрешающую способность этого метода. Сегодня ЭЭГ
регистрируют, размещая сотни электродов на поверхности черепа, и обычно
используют для этого специальную шапочку с электродами. Таким образом,
метод исследования с регистрацией ВП/ССП может предоставить значимую
информацию как о временных параметрах, так и, в настоящее время, о точной
локализации активности, вызванной мыслительными процессами (блок А.1).
2.1.1. Внутричерепная регистрация ЭЭГ и глубинные электроды
Не всегда регистрацию ЭЭГ проводят с поверхности черепа. В некоторых слу-
чаях возможно также регистрировать электрические параметры напрямую от
мозга. Существуют в основном два способа для этого: размещение внутриче-
репной сетки электродов на часть поверхности мозга или погружение глубин-
ных электродов в мозг. Оба способа инвазивны и проводятся только на людях
как часть клинического обследования, обычно для поиска эпилептического оча-
га при планировании операции. Для проведения операции хирургам необходи-
мо знать не только источник эпилептического припадка, но также локализацию
важных когнитивных функций. Если возможно, то хирурги избегают удаления
мозговой ткани, которая вовлечена в выполнение когнитивных функций как
память или речь. Следовательно, пациент должен находиться в бодрствующем
состоянии при проведении исследований для того, чтобы выполнять разные
когнитивные задания и сообщать о каких-либо изменениях, происходящих во
время обследования. Во время имплантации электродов ученым иногда позво-
ляют в течение ограниченного времени провести несколько тестов с такими
пациентами.
В исследовании с глубинными электродами на людях (Quiroga et al., 2005)
зарегистрирован ответ единичных нейронов в медиальной височной коре при
предъявлении пациентам изображений лиц и объектов. Поразительно, но под-
группа нейронов селективно отвечала на предъявление сильно отличающихся
изображений определенных знаменитых людей, объектов природы и других
объектов. Иными словами, исследователи нашли один нейрон, который специ-
фически отвечал на фотографии актрисы Дженнифер Энистон, другой нейрон
отвечал значительно активнее на предъявление изображения актрисы Холли
Берри, а еще один нейрон — на изображение Сиднейского оперного театра
(рис. А.7). Это открытие демонстрирует существование инвариантного, разбро-
санного и детального нейронного кода, который может быть важен для перево-
да зрительных образов в память.
В то время как глубинные электроды вживлены в мозг, можно перена-
править электрический поток и стимулировать некоторые области мозга. Та-
ким образом, можно грубо активировать высокоспецифичную область мозга.
342
Приложение
1,0
Рис. А.7 . Обнаружение нейрона в области задней части левого гиппокампа, селективно
реагирующего на различные изображения актрисы Дженнифер Энистон. Ответы на 30
из 87 предъявленных фотографий, показаны на рисунке (вкладка: ответы на изображе-
ния Дженнифер Энистон). Числа указывают на номер изображения; каждый столбик
отображает количество зарегистрированных нейронных спайков. Аналогичные резуль-
таты наблюдали для других нейронов при предъявлении стимулов, включая изображе-
ния актрисы Холли Берри и Сиднейского оперного театра. (Источник: Quiroga et al.,
2005.)
Методы для изучения живого мозга
343
С использованием этого метода (Barbeau et al., 2005) были обнаружены разли-
чия в субъективном отчете испытуемых при стимуляции гиппокампа или при-
легающей к нему области периринальной коры, областей медиальной височной
коры. При стимуляции гиппокампа пациенты сообщали развернутые эпизоди-
ческие воспоминания с подробными деталями со сценическим и эпизодиче-
ским содержанием. При стимуляции же периринальной коры (рис. А.8) паци-
енты сообщали воспоминания об определенном объекте, а после рассказывали
развернутые эпизодические воспоминания. В то время как данное исследова-
ние ограничено только двумя областями экспериментальной стимуляции, и эти
результаты должны быть интерпретированы с большой осторожностью, данное
открытие указывает на то, что разные области в пределах медиальной височной
коры играют разную роль в процессах запоминания и воспроизведения памя-
ти. Вероятно, гиппокамп вовлечен в процессы воспроизведения целых эпизо-
дических воспоминаний, тогда как периринальная кора более ответственна за
память об определенных объектах.
Внутричерепную регистрацию также проводят и на нечеловекообразных
обезьянах — макаках -резус. Животные «модели» когнитивной деятельности
человека дают ученым возможность для дальнейшего исследования нейронных
коррелятов когнитивных функций. Одним из важных открытий при использо-
Рис. А.8 . Стимуляция периринальной коры приводит к воспоминаниям об объектах.
(а) Реконструкция изображения мозга пациента с локализацией точек имплантации
каждого электрода. Зеленая линия показывает точку имплантации электрода, проходя-
щего через периринальную область. Красные и синие точки указывают на другие обла-
сти имплантации электродов в мозге. (б) Изображение КТ показывает место контактов
электродов (белые пунктирные линии). (в) Аксиальный срез мозга на МРТ-изображении,
демонстрирующий тонкие следы, оставленные проходящим электродом (белая линия).
(г) Коронарный срез мозга на МРТ-изображении с указанием точки стимуляции (зеле-
ные точки) в глубине затылочно-височной борозды в правой височной доле, в области
соответствующей периринальной коре (см. текст). (Источник: Barbeau et al., 2005.)
344
Приложение
вании этого подхода является обнаружение зеркальных нейронов (Gallese et al.,
1996).
В общем, ЭЭГ представляет целый набор различных способов регистрации
(и стимуляции), каждый из которых вносит значительный вклад в изучение вза-
имосвязи мозга и разума. С высоким временным разрешением ЭЭГ дает важ-
ную информацию, которую можно совместить с результатами других методов
визуализации, которые обладают высокой пространственной разрешающей
способностью. Мы вернемся к обсуждению комбинации таких данных методов
в конце этого приложения.
2.2 .
Магнитоэнцефалография
Магнитоэнцефалография (МЭГ) — это измерение магнитных полей, созданных
электрической активностью в мозге, проводимое, как правило, вне черепа. Это
очень интересный метод для исследования функциональной активации и связей
внутри мозга. Пространственное разрешение метода при использовании наибо-
лее современных приборов достигает всего нескольких миллиметров, а времен-
ное разрешение, как и у ЭЭГ, составляет миллисекунды. Это позволяет реги-
стрировать распространение активности из одной области в другую (блок А.2).
2.2.1 . Принцип МЭГ
Физический принцип МЭГ основан на наблюдении Г. Х . Эрстеда в 1820 г. того,
что электрический ток в проводе будет создавать вокруг него окружающее ци-
клическое магнитное поле. Так как импульсы, распространяющиеся в мозге,
имеют электрическую природу, образуется большое количество маленьких ло-
кальных магнитных полей. Магнитное поле от одного нейрона находится ниже
порога регистрации; однако совмещение полей от одной области от примерно
50 000 активных нейронов может дать измеримое магнитное поле.
Давайте для простоты представим единичный электрический провод, кото-
рый окружен циклическим магнитным полем ортогональным к направлению
электрического тока. Если это соотнести с электрическим током в мозге, элек-
трическим диполем, параллельным поверхности черепа, то магнитное поле бу-
дет выходить из головы с одной стороны от электрического потока и входить
обратно с другой стороны. Изменения в электрическом токе в мозге и маг-
нитном поле вызовут электрические токи в круглых контактах, расположенных
параллельно поверхности черепа. Индуцированный ток изменит направление
при перемещении электрода с одной стороны на другую сторону тока в мозге
(рис. А.9). Если, напротив, электрический ток в мозге будет располагаться пер-
пендикулярно поверхности черепа, то магнитное поле не будет образовывать
градиента снаружи от черепа, и внешний электрод будет находиться в покое.
Электромагнитные сигналы в мозге — конечный результат ионных токов в
дендритах нейронов и внеклеточном пространстве как обратных токов, возни-
кающих во время проведения импульса и синаптической передачи. Потенциал
действия не вызывает значимых полей, так как токи в потенциалах действия
текут в обратном направлении, взаимно уничтожая магнитные поля.
Из вышеизложенного обсуждения о направлении электрических диполей и
образованных магнитных полей следует, что нейроны, расположенные в стен-
ках борозд коры, с ориентацией параллельной поверхности головы, формируют
измеримые магнитные поля, которые можно зарегистрировать снаружи черепа.
Методы для изучения живого мозга
345
Значит, нейроны, расположенные на поверхности и в основании борозд, име-
ют ориентацию, которая вызывает магнитные поля с минимальным градиентом
вне черепа и, следовательно, нерегистрируемые. Но отклонение радиального
направления выпуклых источников на 10–20° может быть достаточным для по-
лучения различимых сигналов. Поэтому вполне вероятно, что выпуклые ис-
точники вблизи черепа, а значит и около регистрирующего аппарата, вносят
основной вклад в МЭГ-сигнал.
Рис. А.9 . Магнитоэнцефалография и ее анализ. Испытуемых помещают в сканер, кото-
рый имеет большой набор экранированных датчиков. Сам сигнал возникает от сетевого
эффекта в дендритах нейронов во время синаптической передачи и во внеклеточном
пространстве как обратные токи (внизу). Потенциалы действия не вызывают воспри-
нимаемые поля, так как токи, вызываемые потенциалами действия, текут в обратном
направлении и магнитные поля взаимно уничтожаются. Левая вст авка: магнитные поля
вследствие болезненной (эпидермальной) стимуляции, где (а) зарегистрированные дан-
ные, (б) и (в) остаточные магнитные поля, полученные вследствие фильтрации сигналов
соматосенсорных процессов из зарегистрированных данных. Две нижние линии пока-
зывают временное развитие силы источника во время болезненной стимуляции. Правая
вставка: информация о локализации МЭГ-данных, наложенная на изображения МРТ.
(Источник: 4D Neuroimaging, Сан-Диего, с разрешения.)
346
Приложение
2.2.2. Регистрация МЭГ
Магнитный сигнал, испускаемый работающим мозгом, чрезвычайно мал, не-
сколько фемтотесла (17 фТл = 10–15
Тл *). Следовательно, должны использоваться
чрезвычайно чувствительные и дорогие приборы, такие как сверхпроводящее
квантовое интерференционное устройство СКВИД (SQUID, Superconducting
Quantum Interference Device). СКВИД является ультрачувствительным при-
Interference Device). СКВИД является ультрачувствительным при-
Interference Device). СКВИД является ультрачувствительным при-
Device). СКВИД является ультрачувствительным при-
Device). СКВИД является ультрачувствительным при-
). СКВИД является ультрачувствительным при-
бором для определения магнитного потока. Он действует как преобразователь
ток–напряжение и обеспечивает систему достаточной точностью для опреде-
ления нейромагнитных сигналов. Для того чтобы регистрировать такие слабые
магнитные поля, необходимо экранировать все внешние магнитные сигналы,
включая магнитное поле Земли. Подходящая магнито-экранированная комна-
та может быть создана из мю-металла (никелевого сплава), который обладает
способностью отражать высокочастотный шум, в то время как алгоритмы по-
давления помех уменьшают низкочастотные синфазные помехи. С достаточ-
ным экранированием СКВИД становится преобразователем ток–напряжение
с низким уровнем помех и высоким коэффициентом усиления, обладающим
достаточной чувствительностью для регистрации нейромагнитных сигналов,
достигающих всего лишь нескольких фемтотесла.
Первые регистрации магнитных ритмов мозга были проведены около
40 лет назад с применением магнитометра и индукционной катушкой в экра-
нированной комнате (�ohen, 1968). Современные системы содержат до 300 ка-
�ohen, 1968). Современные системы содержат до 300 ка-
, 1968). Современные системы содержат до 300 ка-
налов СКВИД, расположенных вокруг головы и имеющих уровень шума около
5–7 фТл. Эти параметры можно сравнить с общим магнитным полем мозга,
достигающим от 100 до 1000 фТл.
2.2.3. Анализ данных
Основная техническая трудность МЭГ заключается в том, что проблема опре-
деления изменений в мозге, исходя из измерений магнитного поля вне головы
(«обратная проблема»), не имеет, по существу, единого решения (рис. А .10).
Проблема поиска наилучшего решения является объектом интенсивных ис-
следований в настоящее время. Адекватное решение может быть получено с
использованием моделей, основанных на первоначальных знаниях активности
мозга и характеристик головы, а также с использованием алгоритмов по ло-
кализации. Некоторые исследователи считают, что более сложные, но реали-
стичные модели источников и мозга увеличивают качество решения. Однако
они также увеличивают возможность локальных минимумов и, потенциаль-
но, ухудшают числовые свойства системы, тем самым увеличивая эффекты от
ошибок модели. Многие эксперименты используют простые модели, уменьшая
возможные ошибки в источниках и снижая время вычисления при поиске ре-
шений. Алгоритмы по локализации используют данные модели об источнике
и строении мозга для поиска вероятного расположения генератора локального
поля. Альтернативная методика заключается в выполнении сначала независи-
мого компонентного анализа для того, чтобы выделить отдельные источники, а
* Тесла — количество всего магнитного потока (магнитной «силы») на площадь поверхности. Следо-
вательно, уменьшение площади воздействия, как правило, приведет к увеличению плотности маг-
нитного потока. Тесла — это единица измерения для описания интенсивности (плотности) магнитно-
го поля. Магнитное поле Земли на широте 50o равно 58 мкТл (5,8 × 10–5 Тл), а на экваторе на широте
0o составляет 31 мкТл (3,1 × 10–5 Тл).
Методы для изучения живого мозга
347
затем производится локализация каждого источника по отдельности. Показано,
что этот метод уменьшает соотношение сигнал-шум в данных, правильно раз-
деляя не нейронные источники шума от нейронных источников, и дает обеща-
ющие результаты по изолированию очаговых нейронных источников.
В основном алгоритмы локализации действуют по принципу последова-
тельного уточнения. Система инициируется первым предположением. Далее
запускается цепь, в которой используется модель для генерации магнитного
поля, которое должно возникнуть из предположения, и это предположение из-
меняется для уменьшения различий между данным вычисленным полем и ре-
ально зарегистрированным полем. Этот процесс повторяют до тех пор, пока не
достигнуто схождение вычисленного и измеренного полей.
Другой подход основан на отрицании обратной проблемы и использует оце-
ночный алгоритм для локализации источников. Одним вариантом такого подхода
служат технологии второго порядка, названные магнитометрия синтезирован-
ной апертуры (SAM, synthetic aperture magnitometry), которая использует линей-
SAM, synthetic aperture magnitometry), которая использует линей-
, synthetic aperture magnitometry), которая использует линей-
synthetic aperture magnitometry), которая использует линей-
aperture magnitometry), которая использует линей-
aperture magnitometry), которая использует линей-
magnitometry), которая использует линей-
magnitometry), которая использует линей-
), которая использует линей-
ное взвешивание каналов сенсора для фокусировки луча на данном положении
источника. Этот подход, еще известный как «beam forming» (лучеобразующий),
имеет некоторые преимущества над более традиционными технологиями лока-
лизации источников, так как большинство источников в мозге распределены и
не могут быть описаны точечным положением, таким как диполь тока.
Решение затем накладывается на МРТ-изображения для получения изобра-
жений магнитных источников (ИМИ). Два набора данных соединяются при
помощи определения контрольных точек, отмеченных при МРТ липидными
маркерами и отмеченных во время МЭГ электрическими катушками, которые
испускают магнитное поле. Локализация этих контрольных точек затем исполь-
зуется для создания координатной сетки, чтобы наложение («одновременная
регистрация») функциональной МЭГ на результаты структурной МРТ стало
возможным.
Рис. А.10 . Магнитный гради-
ент нейронов на поверхности
и в основании борозды не име-
ет такой ориентации, которая
увеличивает их вклад в запись
МЭГ. Изменения магнитного
поля, возникающие в нейро-
нах, расположенных в стенках
борозды, лучше регистриру-
ются приборами МЭГ. (Источ-
ник: Adams et al., 2004.)
348
Приложение
Блок А.2 . Чувство фантомной конечности и пластичность мозга
Хорошим примером использования МЭГ в исследованиях психологических или
перцептивных вопросов является работа, проведенная Янгом с коллегами (Yang
et al., 1994) (см. также Ramachandran and Hirstein, 1998; Ramachandran and Rogers-
Ramachandran, 2000). Эти ученые изучали пластические изменения, которые воз-
никают в мозге после ампутации конечности. Фантомные ощущения конечностей
возникают практически у всех людей, перенесших ампутацию. Это значит, что люди,
потерявшие руку, продолжают испытывать ощущения от руки, несмотря на то что ее
нет! Такой феномен хорошо известен в истории — например, лорд Нельсон, фантом-
ная рука которого привела его и других к мысли о существовании нематериальной
души без физического тела. Сегодня мы можем выявлять изменения, происходящие
в мозге у этих несчастных людей.
Существует возможность создать фантомные ощущения у таких пациентов. Напри-
мер, у человека с ампутированной рукой можно вызвать фантомное чувство руки,
щекоча подбородок или челюсть на стороне ампутированной руки. Делая это, Янг с
коллегами обнаружили, что области, которые в норме активируются стимулирова-
нием неповрежденной руки, становятся активными вследствие стимулирования как
лица, так и ампутированной конечности. Эти результаты свидетельствуют о том, что
ампутация ведет к перекартированию первичной соматосенсорной коры и что даже у
взрослых отдельные области мозга могут менять паттерны функциональных связей.
Сегодня считается, что ампутация приводит к трем стадиям изменений в соматосен-
сорной коре. Ампутация ведет к потере входной информации от исходной сенсорной
области. Это приводит к функциональным изменениям в уже существующих связях
в пределах соматосенсорной коры. И наконец, продолжение этих функциональных
изменений приведет к изменениям в физических связях между нейронами.
Рис. А.11 . В норме прилегающие области, отражающие разные части тела, напри-
мер области лица и рук, тесно взаимосвязаны. Это облегчает создание строго специ-
фичных образов, иначе говоря, «это вход от руки» и «этот вход не от лица». Когда
рука ампутирована (или лишена трудоспособности на длительный срок), происхо-
дит значительное перераспределение в соматосенсорной коре. В МЭГ-исследовании
(Yang et al., 1994) четко показано, что стимуляция лица приводила к фантомным ощу-
щениям в потерянной (ампутированной) руке, что коррелировало с совместной акти-
вацией областей представительств лица и рук в соматосенсорной коре. Четко видно,
что у пациента произошло перераспределение в коре. (Источник: Yang et al., 1994.)
Методы для изучения живого мозга
349
Критика использования этого метода в клинической практике заключа-
ется в том, что цветные области с четкой границей накладываются на МРТ-
изображения: неспециалист может не понять, что цвета отражают не определен-
ную физиологическую структуру из-за относительно низкой пространственной
разрешающей способности МЭГ, а, в значительной степени, облако вероят-
ностного расположения на основе статистических вычислений. Но, когда изо-
бражения магнитных источников подтверждают другие результаты, они могут
иметь клиническое применение.
2.2.4. Взаимосвязь с другими методами регистрации
МЭГ развивается с начала 1970-х гг., но только в настоящее время этот метод
получил значительное развитие благодаря современным достижениям в вы-
числительных алгоритмах и техническом обеспечении и имеет хорошее про-
странственное разрешение и невероятно высокое временное разрешение. Так
как МЭГ измеряет напрямую активность нейронов, ее временное разрешение
сопоставимо с регистрацией нейронов внутричерепными электродами. Пре-
имущества МЭГ дополняют таковые других методов измерения активности
мозга, как ЭЭГ, ПЭТ и фМРТ, чьи положительные стороны, в свою очередь,
дополняют МЭГ. Необходимо отметить важные преимущества МЭГ, которые
заключаются в том, что биосигналы, которые она измеряет, не распределены по
черепу, как в ЭЭГ (кроме случаев наличия металлических имплантов), кроме
того, этот метод совершенно неинвазивен по сравнению с ПЭТ.
В исследовательских работах МЭГ в основном применяют для определения
динамики активности, так как она не может быть измерена при помощи фМРТ.
Вследствие различных технологических и методологических трудностей при
локализации источников с помощью МЭГ, она играет вторую роль при созда-
нии функциональных карт коры мозга человека, так как подтверждение пред-
ложенных карт требовало бы применения других методов, прежде чем карты
будут признаны научным сообществом.
В клинике применение МЭГ до настоящего времени ограничивалось осо-
быми случаями по выявлению и локализации эпилептических источников у па-
циентов с эпилепсией и по локализации корковой патологии при планировании
операций у пациентов с мозговыми опухолями или инкурабельной эпилепсией.
Обычно прилегающие области, отражающие разные части тела, например области
лица и рук, имеют тесные взаимосвязи. Это облегчает создание специфичных об-
разов, иначе говоря, «это вход от руки» и «этот вход не от лица». Когда рука ам-
путирована (или лишена трудоспособности на длительный срок), происходит зна-
чительное перераспределение в соматосенсорной коре. Это было четко показано в
МЭГ-исследовании Янга и его коллег (рис. А .11).
В данной работе стимуляция лица приводила к фантомным ощущениям и коррелиро-
вала с взаимной активацией представительств лица и руки в соматосенсорной коре.
Важной особенностью данного исследования было то, что незатронутое полушарие
служило контролем для сравнения. Значит, в дополнении к сравнению испытуемых
с неампутированными конечностями, мы можем сравнить активацию между полу-
шариями. Работа Янга и его коллег считается одним из первых исследований, про-
демонстрировавших пластические изменения в человеческом мозге.
350
Приложение
Физиологическая основа нейровизуализации сигнала заключается в сопря-
жении между локальной корковой активацией и кровотоком, и, далее разобще-
нии между током и потреблением кислорода во время активации. Если говорить
простыми словами, то увеличение активации мозговой ткани ведет к увеличе-
нию интенсивности кровообращения, которое может быть измерено ПЭТ. По-
вышение интенсивности кровотока значительно превышает увеличение потре-
бления кислорода, так что концентрация дезоксигемоглобина уменьшается. Это
локальное снижение концентрации дезоксигемоглобина и лежит в основе сиг-
нала фМРТ.
Эпилептический припадок можно рассматривать как экстремальную нефи-
зиологичную активацию, при которой все нейроны разряжаются с максималь-
ной частотой. При этих условиях кровоток значительно возрастает, потребле-
ние кислорода также увеличивается, но не в таком объеме, в результате чего
увеличивается содержание кислорода в венозной крови в мозге (Brodersen et
al., 1973). Это соотносится с наблюдениями Купера с коллегами, которые ранее
описали увеличение давления кислорода в мозговой ткани. В интересном ПЭТ-
исследовании показано, что во время нормальной физиологической активации
мозга кровоток, измеренный при помощи 15О
2
, и фосфорилирование глюкозы,
измеренное при помощи 18�-ФДГ, увеличиваются параллельно, в то время как
потребление кислорода повышается незначительно (�ox and Raichle, 1986; �ox
et al., 1988).
Теоретические вычисления показывают, что значительная часть метаболи-
ческой энергии тратится на проведение потенциалов действия по аксонным
коллатералям (Attwell and �aughlin, 2001). Однако эмпирические исследования
(�auritzen, 2001, �ogothetis et al., 2001) с использованием совместной регистра-
ции электрофизиологических и гемодинамических параметров показали, что
увеличение кровотока и BO�D (зависимого от уровня кислорода в крови) сиг-
BO�D (зависимого от уровня кислорода в крови) сиг-
(зависимого от уровня кислорода в крови) сиг-
нала отражают повышение синаптической активности и локальных градиентов
потенциалов на дендритах, нежели увеличение частоты разрядов в постсинап-
тических нейронах. Таким образом, выделение возбуждающих, а также тор-
мозных нейромедиаторов приводит к увеличению метаболических процессов,
которые повышают интенсивность кровотока и BO�D-сигнал.
2.3 .
Позитронно-эмиссионная томография
Современный ПЭТ-сканер содержит более 10 000 импульсных детекторов, ко-
торые регистрируют распад изотопов, испускающих позитроны. Когда изото-
пы распадаются, испускаемый позитрон сразу же сталкивается с нормальным
электроном, который его поглощает и формирует два гамма-луча с энергией
511 кэВ* с углом между ними, равным 180°. Когда два детектора сканера ре-
гистрируют фотон с энергией 511 кэВ одновременно, тогда можно точно ска-
зать, что произошел распад изотопа на линии, соединяющей эти два детектора.
При нормальной интенсивности излучения вероятность того, что два детектора
одновременно зарегистрируют два фотона от распада двух разных позитронов,
очень мала. Таким образом, становится возможным трехмерная реконструкция
изображения распределения изотопа в поле зрения ПЭТ-камеры.
* Электрон-вольт (эВ) — количество кинетической энергии, приобретаемой одним свободным элек-
троном при прохождении через разность электростатических потенциалов в 1 вольт в вакууме.
Килоэлектрон-вольт (кэВ) равен 1000 эВ.
Методы для изучения живого мозга
351
Наиболее часто используется в ПЭТ-исследованиях функциональной акти-
вации мозга изотоп H2
15
O. Изотоп 15О является испускающим позитрон изото-
пом со временем полужизни около 2 мин. По этой причине данный изотоп дол-
жен производиться в циклотроне в непосредственной близости от ПЭТ-камеры.
H2
15
O вводят внутривенно, и он достигает мозга пропорционально кровос-
вводят внутривенно, и он достигает мозга пропорционально кровос-
набжению. Несмотря на то что вода не полностью диффундирует, уровень ее
диффузии через гематоэнцефалический барьер в мозге велик, и большая часть
воды, поступающая по сосудистому руслу в мозг, диффундирует в ткани. Позже
произойдет вымывание меченой воды из мозга также в пропорции к перфузии.
Таким образом, становится возможным вычислить мозговой кровоток. Суще-
ствует возможность провести числовые вычисления; однако в многочисленных
исследованиях функциональной активации отмечены только относительные
изменения. Вследствие быстрого распада 15O, доза радиации, получаемая ис-
, доза радиации, получаемая ис-
пытуемым при обследовании, относительно мала, особенно при использовании
ПЭТ-сканера в 3D-конфигурации (трехмерная регистрация с высокой счетной
способностью). Можно провести около 24 исследовательских теста на актива-
цию до получения дозы радиоактивного облучения, меньше чем в два раза пре-
вышающей годовую фоновую радиацию. Так как в основе ПЭТ лежат радио-
активные вещества, существуют этические ограничения высоты дозы, которую
испытуемый может получить за определенный период времени. В результате
это оказывает сильное влияние на выбор методики исследования.
Другим испускающим позитрон изотопом, который используют в исследо-
ваниях функциональной активации, является 10С в форме диоксида углерода
10СО
2
(�aw et al., 2001). Период полураспада 10С составляет только 19 с, что
оказывает очень незначительное радиоактивное воздействие и поэтому позво-
ляет провести до 64 ПЭТ-сканирований с таким же радиационным облучением,
Рис. А .12 . Связывание с рецептором 5-HT2A, измеренное при помощи ПЭТ и совме-
щенное со структурным МРТ-изображением. (Источник: Adams et al., 2004.)
352
Приложение
что и при получении 12 изображений при использовании H2
15O. Короткое вре-
. Короткое вре-
мя жизни изотопа 10С позволяет проводить исследования через более короткие
промежутки времени или прослеживать активацию при повторных введениях
10С-меток. 10СО
2
получил ограниченное распространение потому, что более вы-
сокую разрешающую способность можно получить при помощи фМРТ без по-
лучения радиоактивного облучения. И все же метод 10СО
2
может представлять
интерес в областях, где применение фМРТ ограничивается чувствительными
артефактами, например в областях, расположенных около назальных синусов,
как орбитофронтальная кора.
Недавно прогресс был достигнут в области функциональной визуализации
нервной передачи; волнующие данные были получены при наблюдении in vivo
сигнальной системы мозга относительно выделения нейромедиаторов. Ярким
примером этого может служить ПЭТ-демонстрация конкуренции между эндо-
генным дофамином и антагонистом дофаминовых рецепторов D2 [11С]raclo-
raclo-
pride с радиоактивной меткой или ОЭКТ-индикатором 123I-IBZM (рис. А .13).
Связывание рецепторов в стриатуме с радиоактивными лигандами уменьша-
ется с увеличением уровня дофамина, вызванного успешным прохождением
уровня в компьютерной игре (Koepp et al., 1998). До сих пор этот подход в ос-
новном использовался для визуализации работы D2-рецепторов. Несмотря на
попытки выявить ПЭТ-маркеры, чувствительные, например, к высвобождению
серотонина (Pinborg et al., 2004), таких подходящих соединений еще не выяви-
ли (блок А.3).
Рис. А .13 . Дофаминергический синапс в исходном состоянии (слева) и после фармако-
логической стимуляции амфетамином. Внизу: уровень связывания [11С]raclopride (в ста-
ционарном подходе) при увеличении уровня дофамина. (Источник: �aruelle, 2000.)
Методы для изучения живого мозга
353
2.3.1 . Анализ изображений
Технический прием «областей интереса» является наиболее часто применяю-
щимся подходом обработки изображений при сравнении групп. Он заключает-
ся в выведении на экран изображений каждого субъекта и очерчивании с помо-
щью шарового манипулятора или ручки области интереса. Средняя активность
в области интереса каждого испытуемого затем сравнивается с контролем.
Основным недостатком данного метода служит отсутствие учета структурных
различий в мозге, которые могут существовать между субъектами. В связи с
тем что выделение областей интереса происходит человеком, необходимо пред-
полагать, что оператор знает, где именно надо нарисовать границы. На функ-
циональном изображении заключение о том, что структурные границы могут
быть определены, является центральным. Существуют два возможных реше-
ния данной проблемы. Первое заключается в том, чтобы регистрировать изо-
бражения со структурными данными для каждого испытуемого наряду с функ-
циональными изображениями. Затем технологии для обработки данных могут
применяться для соотнесения полученных результатов воксел к вокселу, так
что получают полностью совмещенную карту для каждого испытуемого. ПЭТ-
изображение на рис. А.12 показывает усредненное 3D-изображение совмещен-
D-изображение совмещен-
изображение совмещен-
ных результатов ПЭТ и МРТ.
2.4 .
Магнитно-резонансная томография
МРТ является методом визуализации, который преимущественно используется
в медицинских обследованиях для получения четких изображений внутреннего
строения человеческого тела. Он основан на принципах ядерного магнитного
резонанса, что является спектроскопическим методом, который ученые ис-
пользуют для получения информации о химических и физических свойствах
молекул. По существу, МРТ начиналось как томографический метод визуали-
зации — метод для получения изображений внутреннего строения тела. Сегод-
ня МРТ значительно расширило свои границы и сейчас включает ряд разных
подходов, которые выявляют структуру, функцию, взаимосвязи и химический
состав любой части тела.
МРТ основано на поглощении и испускании энергии в радиочастотном диа-
пазоне электромагнитного спектра. Человеческое тело в основном состоит из
жировой ткани и воды — ткани тела содержат большое число атомов водоро-
да. Эти водородные ядра и составляют основу сигнала МРТ. Воксел — объем-
ный элемент, который представляет значение в трехмерном пространстве. Он
аналогичен пикселю, который служит для представления данных в двумерных
изображениях. Воксел часто используют для представления и анализа меди-
цинских и научных результатов. При визуализации мозг разделяют на опреде-
ленное число вокселей, и в пределах каждого воксела можно проводить разные
статистические тесты. Каждый воксел (рис. А.15) в человеческом теле состоит
из более чем одного вида ткани. Приближение к вокселу выявляет клетки и
в каждой клетке содержатся молекулы воды. Каждая молекула воды состоит
из одного атома кислорода и двух атомов водорода. Если мы приблизимся при
рассмотрении одного атома водорода за пределы электронного облака, то мы
увидим ядро, состоящее из одного протона.
354
Приложение
Блок А.3 . Изменения нейрохимии при психиатрических расстройствах
В работе Адамса с коллегами (Adams et al., 2005) пациенты, страдающие обсессивно-
компульсивным расстройством (ОКР), были обследованы на связывание серотонина с
рецепторами в мозге (при помощи связывания [18H]altanserin с 5-HT2A-рецепторами).
ОКР является психиатрическим заболеванием, которое затрагивает 2–3% населения
во всем мире и классифицируется как тревожное расстройство. Симптомы включают
повторяющиеся, нежелательные мысли (мании) и сознательные, ритуализированные
действия (компульсивные побуждения), обычно направленные мрачными предчув-
ствиями, вызванными манией. ОКР обычно лечат при помощи селективных инги-
биторов обратного захвата серотонина (СИОЗС), химических соединений, которые
позволяют серотонину дольше находиться в синаптической щели.
При сравнении группы ОКР-пациентов с контрольной группой аналогичного пола
и возраста, Адамс с коллегами обнаружили увеличение связывание [18H]altanserin с
рецепторами в хвостатом ядре. Это указывает на то, что у пациентов с ОКР имеется
большее число активных 5-HT2A-рецепторов по сравнению со здоровыми контроль-
ными испытуемыми (рис. А.14). Считается, что такое увеличение серотониновых
рецепторов является компенсаторным механизмом, вследствие низкого уровня серо-
тонина у пациентов с ОКР. Система серотонина, которая затронута, включает орби-
тофронтальную префронтальную кору, хвостатое ядро и бледный шар.
Ученые провели повторное сканирование пациентов с ОКР после введения им в
течение 12 недель СИОЗС, препарата, который увеличивал количество доступного
серотонина у этих пациентов. Эти результаты не выявили различий в связывании
серотонина между контрольной группой и пациентами. Другими словами, у них не
было после лечения большего числа активных 5-HT2A-рецепторов по сравнению
с контрольной группой.
ПЭТ-исследования, аналогичные этому, выявляют важную информацию о нейрохи-
мии психиатрических заболеваний, и они расширяют наше понимание, предоставляя
новые данные о нейрохимическом влиянии психиатрических препаратов как СИОЗС.
Рис. А.14 . Уровень активации 5-HT2A рецепторов у больных с ОКР без лечения
и здоровых испытуемых
Методы для изучения живого мозга
355
В магнитном поле, как в МРТ-сканере, магнитная ориентация каждого ато-
ма водорода выстраивается по направлению линий магнитного поля, и протон
вращается в данной ориентации (рис. А.16(б) и (в)). Приложение короткого
электромагнитного (радиочастотного) импульса временно искажает осевое на-
правление атома в магнитном поле (рис. А.16(г)). Когда радиочастотный им-
пульс заканчивается, атомы начинают возвращаться к направлению по линии
магнитного поля, этот процесс называется релаксация (рис. А.16(д)). Во время
этой стадии атом теряет свою энергию, испуская ее, что обеспечивает полу-
чение информации о его окружении. Релаксация зависит от двух параметров:
Время 1 и Время 2. Возвращение атома в исходное положение относительно
магнитного поля называется продольной релаксацией, а время, равное пример-
но 1 мс, требуемое для возвращения в это состояние определенного количе-
ства ткани, называют Временем 1 или Т1. Это является основой для получе-
ния Т1-взвешенных изображений, которые дают основную часть структурных
МРТ-изображений. Т2-взвешенные изображения основаны на локальном сме-
щении по фазе спинов, вследствие приложения поперечного энергетического
импульса; это время поперечной релаксации называют «Время 2» или Т2.
Рис. А.15. Воксел как минимальный объемный элемент мозга. Вокселы мозга. Воксел
представляет единицу объема в трехмерном пространстве. В мозге разрешение сканера
определяет, насколько маленькими будут размеры воксела. Параметры, как повышение
силы поля в сканере, увеличивают пространственное разрешение, а значит, улучшают
возможность выделения отдельных структур мозга. Воксел мозга выделяет сигнал из
одной части мозга, где локальное молекулярное окружение влияет на магнитный ответ.
В иллюстративных целях воксел, показанный на рисунке, намного больше, чем реально
используемые при МРТ-сканированиях. (Источник: Jones et al., 2002.)
356
Приложение
Параметры Т1 и Т2 обеспечивают основу для большинства способов меди-
цинской визуализации. В разных частях тела, например мозге, различные тка-
ни имеют разную скорость возникновения Т1 и Т2 релаксации. Три основные
ткани мозга — серое вещество (СВ), белое вещество (БВ) и цереброспинальная
жидкость (ЦСЖ). Влияние этих тканей обусловливает разную интенсивность
сигнала — контраст — которая позволяет легко разделять их. Варьируя разные
параметры при сканировании, например частоту и амплитуду радиочастотного
импульса или время от возбуждения до регистрации, становится возможным
выделять разные признаки тканей и их различия.
Это можно четко увидеть на снимках, показывающих различия между Т1- и
Т2-взвешенными изображениями (рис. А .17). На рисунке показано, что опухоль
мозга, которая практически не видна на Т1-взвешенном изображении, отчетли-
во выделяется на Т2-взвешенном изображении. Причиной этого служит то, что
опухоль содержит значительно больше воды и поэтому дает больший сигнал
при Т2-взвешенной визуализации в связи с более медленным временем про-
дольной релаксации. Так как продольная релаксация происходит в воде очень
Рис. А.16 . Сигнал, лежащий в основе метода фМРТ. (а) Вне сканера атомы в мозге ори-
ентированы в произвольном направлении. (б) Когда испытуемого помещают в сканер,
атомы выстраиваются в линию по направлению вектора магнитного поля (В0). Однако
упорядочивание не совершенно, так как соседние атомы влияют друг на друга. (в) При
таком начальном уровне атомы вращаются вокруг оси y, т. е. вектора В0 поля. (г) Когда
прикладывают радиочастотный (РЧ) импульс, он оказывает влияние на спины атомов,
направляя их «вниз». Процесс называется стадией неравновесного состояния, и при
возвращении в равновесное состояние относительно В0 поля атом испускает энергию,
которую он получает от РЧ-импульса. (д) Локальное окружение атома, т. е. серое ве-
щество, белое вещество, кость или цереброспинальная жидкость, определяет скорость
релаксации. Эта основа различий на МРТ-изображении делает возможным определение
разных тканей в теле
Методы для изучения живого мозга
357
быстро, дополнительное количество воды в опухоли не оказывает влияния на
сигнал в Т1-взвешенном изображении.
Применение технологий структурного сканирования является очевидным
выбором при исследовании изменений в мозге в связи со старением, мозговы-
ми травмами или дегенеративными заболеваниями, такими как болезнь Альц-
геймера. На основе визуализации морфологии (строения) мозга становится
возможным постановка вопросов относительно того, как изменяется структура
мозга в результате развития, старения или дегенеративных заболеваний. Напри-
мер, были продемонстрированы возрастные изменения в разных частях коры и
мозжечка (Jernigan et al., 2001). Используя структурные МРТ изображения и
вырисовывая анатомические области, исследователи смогли сравнить относи-
тельные размеры этих структур в разном возрасте. На рис. А .18 показано, что
в мозге в процессе старения происходят значительные изменения. Более того,
существует нелинейная взаимосвязь между возрастом и объемом мозга, что
опровергло ранние предположения о старении мозга. И наконец, значительным
открытием в этой области стало заключение о том, что существуют региональ-
ные различия в старении мозга.
2.4.1 . Функциональная МРТ
В то время как Т1- и Т2-взвешенные изображения являются главными при визу-
ализации структуры мозга, МРТ также дает возможность выявлять различные
функции мозга. В основном существуют два подхода: BO�D фМРТ (с опреде-
BO�D фМРТ (с опреде-
фМРТ (с опреде-
лением уровня кислорода в крови) и перфузионная МРТ. В то время как BO�D
МРТ основана на серии сложных процессов, которые связывают активацию
мозга с изменениями в сосудистой системе и относительном уровне кислорода
в локальном кровотоке, перфузионная МРТ измеряет мозговой кровоток (МК)
или мозговой объем крови (МОК).
BO�D фМРТ является наиболее применимым и широко известным спосо-
фМРТ является наиболее применимым и широко известным спосо-
бом определять мозговую активацию при помощи МРТ. Мозговая активация ме-
няет относительные концентрации оксигенированного и дезоксигенированного
гемоглобина — крови с кислородом и без него, соответственно — в локальном
Рис. А.17. Т1- и Т2-взвешенные изображения. Обратите внимание, что опухоль практи-
чески не видна на Т1-взвешенном изображении(T1W), в то время как на Т2-взвешенном
изображении ее можно легко локализовать — круглое темное пятно в левом полуша-
рии мозга на правом рисунке. TRA — трактография, I�1 — независимый компонент 1.
(Источник: Nakai et al., 2004.)
358
Приложение
Рис. А .18 . Старение мозга происходит неравномерно в разных его областях. Вверху:
пример структурных границ мозга, нарисованных как области интереса. Обратите вни-
мание, что срезы состоят как из серого вещества, так и из белого; белое вещество отмече-
но более светлым цветом. Все вокселы белого вещества, которые отражают увеличение
сигнала в группе старых людей по сравнению с более молодыми взрослыми, отмечены
желтым цветом. Внизу: возрастные изменения в двух областях мозга, таламусе и при-
лежащем ядре (отмечены стрелкой), в возрасте от 10 до 100 лет. Можно отметить два
основных эффекта. Во-первых, в обеих структурах наблюдаются три зависящих от вре-
мени фазы: (1) стремительное уменьшение объема обеих структур в подростковом воз-
расте и раннем периоде взрослой жизни; (2) плоская и относительно стабильная фаза
до позднего периода взрослой жизни; (3) потеря объема в позднем периоде взрослой
жизни. Во-вторых, у этих структур наблюдается разная траектория потери в объеме.
(Источник: (вверху) Jernigan et al., 2001; (внизу) Jernigan, с разрешения.)
Методы для изучения живого мозга
359
кровотоке. В то время как оксигенированная кровь является диамагнетиком
и не меняет МРТ-сигнал, дезоксигенированная кровь — парамагнетик и ведет
к падению МРТ-сигнала. Если в области больше дезоксигенированной крови,
то BO�D-сигнал падает, если больше оксигенированной крови в данной обла-
BO�D-сигнал падает, если больше оксигенированной крови в данной обла-
сигнал падает, если больше оксигенированной крови в данной обла-
сти, то это приводит к увеличению сигнала. Таким образом, изменение BO�D-
сигнала происходит в четыре стадии (рис. А .19).
1. Начальное снижение сигнала, вызванное комбинацией негативного мета-
болического и неметаболического BO�D-эффекта. Другими словами, если
группа нейронов разряжается, она поглощает больше кислорода. Когда это
происходит, локальный уровень оксигенированной крови падает и повы-
шается относительный уровень дезоксигенированной крови в этой области.
В дополнение к этому происходит расширение кровеносных сосудов, кото-
рое приводит к дальнейшему снижению BO�D-сигнала.
2. Длительное увеличение сигнала или положительный BO�D-эффект вслед-
BO�D-эффект вслед-
- эффект вслед-
ствие повышения кровотока и соответствующего сдвига соотношения де-
зокси/оксигемоглобина. Когда нейроны возвращаются к исходному уровню
активации, их увеличенное потребление кислорода заканчивается. В то же
Рис. А.19 . Сигнал, зависящий от уровня кислорода в крови, упрощенно представлен в
виде четырех стадий. Стадия 1: увеличение нейронной активации ведет к повышению
потребления кислорода из крови, что приводит к снижению количества оксигенирован-
ной крови и повышению дезоксигенированной, что, в свою очередь, приводит к паде-
нию BO�D-сигнала. Стадия 2: сосудистый ответ на увеличение потребления кислорода
приводит к значительному повышению количества новой оксигенированной крови, в
то же время потребление кислорода падает с уменьшением активности нейронов. Ста-
дия 3: нормализация кровотока и уровня дезокси/оксигемоглобина (не показано). Ста-
дия 4: постстимульный провал, вызванный медленным восстановлением объема крови
360
Приложение
время приток новой оксигенированной крови увеличен вследствие предыду-
щих требований. По мере увеличения оксигенации крови, сигнал продолжа-
ет увеличиваться. Снижение потребления кислорода совместно с отсрочкой
притока оксигенированной крови приводит к значительному выбросу от-
носительного уровня оксигенированной крови. Это превышение вызывает
главный эффект BO�D-сигнала.
3. Длительное снижение сигнала, который вызван возвращением к нормально-
му кровоснабжению и нормальному соотношению дезокси/оксигемоглобина.
4. Постстимульный спад, вызванный медленным восстановлением объема
крови в мозге.
Измерение начального падения наиболее близко отражает нейронную акти-
вацию, так как оно отражает первое снижение интенсивности сигнала вслед-
ствие потребления оксигенированной крови. Однако изменения сигнала на дан-
ной стадии настолько малы, что могут быть измерены только очень мощными
МРТ-сканерами, которые очень дороги и к тому же не одобрены для использо-
вания на людях. Таким образом, вторая фаза — длительное увеличение сигна-
ла — используется в большинстве BO�D фМРТ-исследованиях. Этот сигнал
служит непрямым измерением нервной активации, так как он является резуль-
татом отсроченного сосудистого выброса оксигенированной крови как ответа
на потребность в оксигенированной крови в этой области. Несмотря на то что
BO�D-ответ является непрямым и отсроченным измерением нервной активно-
-ответ является непрямым и отсроченным измерением нервной активно-
сти, он обладает временной разрешающей способностью порядка нескольких
миллисекунд. К тому же этот метод обладает очень высокой пространственной
разрешающей способностью, которая составляет 1 мм. С современным техни-
ческим обеспечением разрешение стало еще меньше, что привело МРТ к суб-
миллиметровой шкале.
Существуют разные способы проведения фМРТ-исследований. В целом,
можно выделить две категории экспериментальных парадигм: блочная схема и
связанная с событием схема. Так же как и при ПЭТ-исследованиях, блочная схе-
ма была наиболее распространена в начальном периоде использования фМРТ.
Как правило, блочная схема состоит в предъявлении стимулов блоками, каж-
дый из которых содержит набор из одинаковых стимулов. Например, блочная
схема может быть использована при изучении непрерывного внимания, когда
испытуемого инструктируют нажимать на кнопку каждый раз при предъявле-
нии буквы «Х» на экране в ряду других букв. Обычно блоки разделены одина-
ковыми по времени периодами покоя, хотя можно составить блочную схему
эксперимента и без периодов для отдыха. Блочная схема может использоваться,
если необходимо увидеть разницу между процессами обработки информации о
личности или расположении объекта (рис. А .20).
При использовании связанной с событием схемы стимулы предъявляют-
ся в случайном или псевдослучайном порядке. При этом отдельные стимулы
разных условий предъявляют в произвольном порядке, с вероятностным по-
явлением стимула. Этот подход позволяет выявлять различные изменения в за-
висимости от пробы, например правильные или неправильные реакции. Свя-
занная с событием парадигма лежит ближе к традиционным психологическим
экспериментам и позволяет проводить запоздалый анализ с сортировкой проб
(точность, выполнение, время реакции и т. д.). Эта схема наиболее эффектив-
Методы для изучения живого мозга
361
на, так как исходная произвольность предъявления стимулов гарантирует, что
подготовительные или опережающие эффекты (которые свойственны блочной
схеме) не повлияют на связанные с событием ответы. Пример связанной с со-
бытием парадигмы представлен на рис. А .21 .
Когда надо применять блочную или связанную с событием схему в экспе-
рименте? В основном если необходимо исследовать связанные с ситуацией со-
бытия (например, «процесс кодирования» или «процесс воспроизведения»), то
блочная схема будет наилучшим выбором. Однако если вы хотите исследовать
процессы, связанные с предметами (как правильные или ошибочные реакции),
оптимальным является применение связанной с событием парадигмы. В опре-
деленных экспериментальных ситуациях можно совместить блочную и связан-
ную с событием схемы, что позволяет как прослеживать общие различия между
стадиями, так и выявлять различия между частными случаямиь (рис. А .23).
При рассмотрении экспериментальной парадигмы необходимо учитывать
еще один важный параметр сравнения двух состояний. Существуют несколько
разных параметров, из которых можно выбрать. В некоторых случаях, особен-
но в старых работах по нейровизуализации, сравнения проводили между со-
стояниями выполнения задания и покоя (см. рис. А .20). Сегодня более распро-
странено сравнение двух или более активных состояний. В целом, мы можем
выделить сравнения внутри субъекта и между субъектами. При использовании
внутри-субъектной схемы, сравнение проводят между двумя различными со-
Рис. А.20 . Блочная схема. Интервалы отдельных проб (X и Y) разделены периодами
отдыха. Обратите внимание, порядок предъявления проб X и Y произвольный. В такой
схеме эксперимента во время X-проб требуется «читать буквы», а во время Y-проб —
«читать цифры»
Рис. А.21 . Связанная с событием схема. В отличие от блочной схемы в связанной с
событием парадигме внимание уделяется отдельным предъявлениям. Значит, порядок
предъявления стимулов более случаен, и это хорошо для использования в психологиче-
ских исследованиях, когда нам необходимо избежать предсказывания испытуемым по-
рядка следования стимулов. Примером этой парадигмы может служить исследование,
в котором стимул предъявляется на очень короткое время, и испытуемый должен дать
отчет, видел ли он стимул. Ответы, а также эндогенные процессы (например, намере-
ние) будут различаться в зависимости от длительности стимула. Испытуемые в такой
ситуации не способны предсказать свой следующий ответ (или восприятие стимула)
362
Приложение
Блок А.4 . BOLD-природа сознательного восприятия
Какова нейронная судьба стимула, который сознательно воспринят? Как отличается
этот процесс от случая, когда стимул обработан, но сознательно не воспринят? Су-
ществуют ли такие ситуации, когда мы получаем смутное ощущение того, что стимул
присутствовал, но не можем его описать? Если так, то как мозг работает при таком
восприятии?
Нейронные корреляты сознания исследуются уже более десяти лет с использовани-
ем методов нейровизуализации. Эти данные показывают, что сознательное воспри-
ятие – в отличие от бессознательного восприятия – вовлекает в активацию сеть об-
ластей мозга (Baars, 2002b). Эта сеть включает префронтальную и теменную кору, и
очень часто височную кору. Также считается, что распространение информации по
сети происходит через кортикоталамокортикальные связи, т. е . сигналы, возникаю-
щие в коре, распространяются к обширным областям через таламические ядра.
Недавно проведенные поведенческие работы выявили, что необходимо пересмот-
реть устоявшиеся различия между сознательным и бессознательным восприятием
(Ramsøy and Overgaard, 2004). Если испытуемым позволено использовать более под-
робные отчеты об их опыте, они часто отмечают, что некоторый их опыт можно оце-
нить как смутный. Это состояние можно охарактеризовать как восприятие наличия
чего-то, без способности его идентифицировать. Возникает один вопрос: как этот
опыт соотносится с сознательным и бессознательным восприятием? Также очень
важно понять, существуют ли различия в нервном субстрате для такого опыта.
Этот вопрос был изучен при предъявлении обратно замаскированных зритель-
ных стимулов различной длительности при сканировании испытуемых в фМРТ
(�hristensen et al., 2006). Данная парадигма ведет к появлению разных уровней
восприятия стимула, и по результатам прошлых поведенческих работ (Ramsøy and
Overgaard, 2004) испытуемым было позволено отойти от дихотомической модели
ответов и давать отчет о стимулах как о неувиденных, смутно или четко воспри-
нятых. Простые зрительные стимулы предъявляли на короткое время, равное при-
мерно 100 мс. Для каждого стимула испытуемый должен был давать отчет о том,
насколько четко он воспринял стимул. Это позволило сравнивать уникальные пат-
терны активации нервных процессов, лежащих в основе четкого и смутного вос-
приятия.
Во-первых, сравнения между отчетами о четком восприятии и отсутствии его, т. е .
традиционные различия между сознательным и бессознательным восприятием, по-
казали ранее отмеченную обширную сеть областей мозга. Она включала теменные
и префронтальные области коры, височную долю (веретеновидную извилину), кото-
рые хорошо известны своей ролью в процессах восприятия объектов (рис. А .22(а)).
Показано, что при сознательном восприятии также активируется большая часть та-
ламуса в обоих полушариях.
Эти результаты подтверждают «глобалистическую» точку зрения на сознание в моз-
ге, которая заключается в том, что сознательное восприятие требует активации об-
ширных областей мозга.
Что тогда происходит при смутном восприятии? При сравнении отчетов о смутном
восприятии и отсутствии его, исследователи обнаружили активацию сходной сети,
но не настольно обширной, как при сознательном восприятии (рис. А.22(б)). Сравне-
ние состояний смутного и четкого восприятия дало возможность идентифицировать
области, отвечающие за переход от обнаружения к идентификации (рис. А.22(в)). Из
этого следует, что, когда мы обнаруживаем что-то или испытываем смутное ощу-
щение видения чего-то, у нас активирована обширная сеть областей мозга, с осо-
бой активацией в префронтальной коре. Если смутное восприятие стимула стано-
вится полностью осознанным, то активность сети значительно увеличивается как по
Методы для изучения живого мозга
363
амплитуде, так и по распространению. В целом, исследование Кристенсена с кол-
легами дало новую и важную информацию для нашего понимания сознательного и
бессознательного восприятия.
Рис. А.22 . фМРТ-исследование (�hristensen et al., 2006) показывает различие в ак-
тивации нейронных сетей в ответ на четко воспринятые зрительные стимулы по
сравнению со смутным восприятием. Верхние два ряда (а) демонстрируют значи-
тельную активацию при четком восприятии (С) по сравнению с отсутствием вос-
приятия стимула (N). Следующие два ряда (б) показывают активность при смутном
или мимолетном восприятии стимула (V) по сравнению с N. (Источник: �hristensen
et al., 2006.)
364
Приложение
стояниями обработки, между разными условиями во время одного состояния
или в зависимости от времени. Межсубъектная схема позволяет исследовать
групповые эффекты несколькими способами, включая сравнения между па-
циентами и здоровыми испытуемыми, между разными группами больных или
между группами испытуемых, перенесших разные воздействия (например,
обучение). Список некоторых возможных схем экспериментов представлен в
табл. А.1 . Таким образом, нужно обязательно обращать внимание, какие срав-
нения производят в данном конкретном эксперименте.
Рис. А.23 . Совмещение блочной и связанной с событием схем экспериментов возмож-
но при определенных условиях. При этом анализ можно фокусировать на уровне бло-
ков, например, сравнение активации при кодировании и воспроизведении информации,
или используя поведенческие данные из фазы воспроизведения, выявлять нейронную
активацию внутри блоков кодирования для рассмотрения только успешных процессов
кодирования (т. е . игнорируя данные о стимулах, которые впоследствии забываются).
(Источник: Thomas Ramsøy, Daniela Balslev and Olaf Paulson, с разрешения.)
Таблица А.1
Разные схемы проведения исследований с нейровизуализацией
Внутрисубъектные схемы Примеры
Разные стадии обработки
Чтение вслух слов против самостоятельного выдумы-
вания слов
Сравнение эффектов содер-
жимого в одном состоянии
Обработка изображений лиц против домов
Длительное исследование
Повторное тестирование одних и тех же когнитивных
функций:
• Эффекты обучения
• Эффекты старения
• Эффекты заболеваний (только для заболеваний,
которые меняются со временем)
Межсубъектные схемы
Примеры
Эффекты манипуляций
Процессы внимания в группе принимающих экстази
против не принимающих
Эффекты обучения
Эффект обучения жонглированию на активацию мотор-
ной коры
Пациенты и контрольная
группа
Функции рабочей памяти у пациентов с шизофренией
против контрольной группы
Разные группы пациентов
Процессы обработки у пациентов с болезнью Альцгей-
мера против пациентов с семантической деменцией
Методы для изучения живого мозга
365
При изучении эффектов одного условия всегда необходимо сравнивать его
с другими условиями для получения значимой информации. Давайте проил-
люстрируем это на примере изучения восприятии эмоциональных выражений
лиц. В стандартном эмоциональном задании (Del Ben et al., 2005) испытуе-
мых просят смотреть на изображения человеческих лиц. Половина предъяв-
ляемых лиц женские, и в задание испытуемого входит сообщать, является ли
лицо женским или мужским. В то же время лица различаются относительно
эмоций, которые они выражают. Лица могут быть либо нейтральными, либо
вызывающими отвращение (например, грустными или испуганными). Во вре-
мя этого эксперимента наше внимание направлено на различия нейтральных и
отвратительных лиц. Во время обработки данных мы сначала определяем, ког-
да были показаны отвратительные и нейтральные лица. Затем мы определяем
интенсивности сигналов для нейтральных и вызывающих отвращение лиц, и
получаем среднюю интенсивность сигнала для нейтральных и отвратительных
лиц, соответственно. Средний уровень сигнала и его распределение для каждо-
го условия показаны на рис. А.24 . Если мы заинтересованы в идентификации
областей, которые вовлечены в обработку эффектов вызывающих отвращение
лиц, мы берем среднюю активность при предъявлении отвращающего стимула
и вычитаем из нее активность в ответ на нейтральные стимулы. Таким образом,
вся активация, которая является общей для двух условий, включая первичную
Рис. А.24 . фМРТ при воспри-
ятии вызывающих отвраще-
ние эмоциональных лиц. При
смотрении на вызывающие
отвращение лица активиру-
ются области эмоций в мозге.
В последовательности предъ-
являемых лиц некоторые лица
вызывали чувство отвращения
(т. е. были испуганы, грустны
или показывали отвращение),
другие были нейтральными.
Если мы вычтем активность,
связанную с восприятием ней-
тральных лиц, из активности
вызывающих отвращение лиц,
мы получим исходный сиг-
нал, который специфичен для
восприятия вызывающих от-
вращение лиц. Здесь показана
активация миндалины в обоих
полушариях и орбитофрон-
тальной коры у отдельного ис-
пытуемого при использовании
этого метода. (Источник: Del
Ben et al., 2005.)
366
Приложение
зрительную обработку и элементарное восприятие лиц, исключается. В конце
мы получаем данные о том, в каких областях мозга наблюдается повышение
(или снижение) активности при восприятии вызывающих отвращение лиц.
BO�D фМРТ сегодня является, вероятно, наиболее используемым методом
измерения активности мозга. Он доказал свою значимость в каждой области
когнитивной нейронауке, и он постоянно совершенствуется как инструмент для
исследований. С увеличением технического обеспечения этот метод приведет
к лучшему пониманию временных и пространственных процессов в мозге и
мышлении (блок А.4).
2.4.2. Мечение артериального спина (МАС)
Изменяя магнитные свойства крови, текущей в мозге, измерения при помощи
МРТ можно сделать чувствительными к перфузии крови в мозге. Метод, на-
зываемый мечением артериальных спинов, использует идею «мечения» крови в
сонной артерии, с приложением коротких радиочастотных импульсов. Это из-
меняет магнитные свойства у небольшого объема крови. По мере того как кровь
течет через мозг, можно измерять магнитную восприимчивость определенной
области или всего мозга. Перфузионно-взвешенное изображение может быть
получено при вычитании исходного изображения мозга без намагниченной
крови из изображения мозга с намагниченной кровью. Таким образом, можно
измерять перфузию крови через определенную область мозга, а также сравни-
вать перфузионные изображения между группами (блок А.5).
2.4.3. Другие специальные варианты МРТ
Диффузионно-тензорная визуализация (ДТВ) и трактография
МРТ можно использовать для измерения движения молекул воды во времени.
В свободной и неограниченной (изотропной) среде вода будет диффундировать
равномерно во всех направлениях, что также известно под названием броунов-
ского движения. Если измерять диффузию в такой среде, то результирующее
изображение будет иметь форму сферы. Если вода находится в ограниченной
(анизотропной) среде, она не может свободно диффундировать, а может толь-
ко двигаться вдоль структур, которые образуют для нее физические границы.
Например, в стакане с водой молекулы воды в центре могут свободно переме-
щаться во всех направлениях. Потенциал движения для одной молекулы одина-
ковый во всех направлениях. Однако если в стакан с водой поместить трубочку,
то потенциал движения молекул в трубочке будет значительно ограничен. Если
измерять диффузию воды в этой трубочке, она больше не будет сферической
или эквипотенциальной, а образует продолговатое облако (рис. А .26).
В биологической среде, например в мозге, диффузия воды сильно ограни-
чена. Однако существует систематическая разница между белым и серым ве-
ществом. В то время как серое вещество обладает незначительной внутренней
структурой, белое вещество состоит из волокон, которые ограничивают диффу-
зию молекул в одном направлении больше, чем в другом. Так же как помещен-
ные в стакан с водой несколько трубочек препятствуют движению воды через
их стенки, так и нервные волокна запрещают диффузию молекул в поперечном
направлении, но не в продольном (см. рис. А.26(б)). Связи внутри мозга со-
стоят из пучков волокон (аксонов). Используя диффузионно-тензорную визу-
ализацию, возможно измерять относительное направление и связанность этих
Методы для изучения живого мозга
367
трактов белого вещества. Таким образом, ДТВ может использоваться для выяв-
ления изменений белого вещества при неврологических заболеваниях, а также
можно узнать, как определенная область мозга связана с другими, прослеживая
волокна из этой области (метод называется прослеживанием волокон). Исполь-
зование ДТВ у здоровых испытуемых рассмотрено в блоке А.6 (рис. А.27).
МР-спектроскопия
Магнитно-резонансная спектроскопия (МРС) очень близко связана с магнитно-
резонансной томографией. Оба этих метода используют магнитные свойства
атомных ядер для получения информации о биологическом образце. Однако
Блок А.5 . МАС при болезни Альцгеймера
и умеренных когнитивных нарушениях
Хотя значительные когнитивные нарушения могут быть выявлены на ранних стадиях
болезни Альцгеймера (БА), включая нарушения памяти, в мозге возникает значи-
тельное число физиологических изменений. Исследуя перфузию крови у таких паци-
ентов, можно увидеть области, которые имеют распространенное снижение метабо-
лизма или кровотока. В подобной работе (Johnson et al., 2005) авторы использовали
мечение артериального спина для сравнения уровня перфузии в группе с болезнью
Альцгеймера с группой здоровых пожилых испытуемых и с группой пациентов,
страдающих умеренными когнитивными нарушениями, которые являются предиаг-
ностическим синдромом, при котором часть пациентов имеет высокую предрасполо-
женность к БА.
Исследователи обнаружили значительное снижение уровня перфузии в группе БА
по сравнению с контрольной группой в некоторых областях теменной коры, задней
части поясной коры и лобной коры. На более низком статистическом уровне группа
с УКН продемонстрировала снижение перфузии в правой нижней теменной доле,
там же, где наблюдается наибольший эффект в группе БА. Эта работа демонстрирует,
что сканирование перфузии при помощи МАС может использоваться для выявления
различий между группами с разной стадией болезни Альцгеймера (рис. А.25).
Рис. А.25. МАС при болезни Альцгеймера. (Источник: Johnson et al., 2005.)
368
Приложение
существует ключевое различие: в то время как МРТ измеряет распределение
магнитного поля, МРС определяет количество сигнала от каждой химической
среды (химического или молекулярного распределения). Другими словами,
МРТ позволяет исследовать определенную область внутри организма или об-
разца, но дает относительно мало информации о химической или физической
природе этой области. МРС, которая является методом ЯМР, обеспечивает
огромной объем химических данных об этой области. Частотное распределе-
ние, называемое спектром, является аналогом оптическому спектру веществ,
которые ответственны за их видимые цвета. Таким образом, МРС может опре-
делять химический состав данной области мозга. МРТ и МРС можно объеди-
нить для получения пространственного распределения химических веществ
Рис. А.26 . Диффузия воды зависит от локального окружения. (а) В свободной, без огра-
ничений среде (например, в стакане воды) вода может свободно диффундировать. Диф-
фузия является изотропной; она имеет возможность двигаться в любом направлении.
Если молекула воды физически ограничена, то она больше не может свободно пере-
мещаться в любом направлении. Это анизотропная диффузия; молекула не может дви-
гаться в любом направлении. В среде волокон, таких как в белом веществе мозга (б),
молекулы воды жестко ограничены аксональными волокнами. Таким образом, можно
визуализировать проводящие пути мозга и, более того, установить целостность (гомо-
генность) белого вещества в данной области. (в) Такая визуализация формирует типич-
ные окрашенные ДТВ-изображения мозга, которые иллюстрируют траектории в белом
веществе
Методы для изучения живого мозга
369
Блок А.6 . Способность к чтению и связи мозга
Чтение является сложным когнитивным навыком и результатом координации между
множеством разных областей мозга. Для того чтобы такая взаимосвязь между об-
ластями возникла, необходимо наличие физических контактов между этими обла-
стями. Это могут быть как прямые связи между двумя и более областями, так и
непрямые связи через одну или более дополнительные структуры. И хотя многие
исследования с использованием функциональной визуализации обращаются к во-
просу о взаимодействии между такими областями, они не дают четких результатов
о структурных связях, которые присутствуют в мозге для обеспечения этого взаимо-
действия.
В одной из работ (Beaulieu et al., 2005) авторы обратились к этой проблеме, исследуя
связи мозга у детей с различными способностями к чтению. Сравнивая связи бело-
го вещества и изменения целостности со способностью детей читать, они выявили
пять областей белого вещества, которые служили хорошими маркерами способности
детей читать.
Исследователи продолжили искать различные взаимосвязи между этими регионами.
Используя каждую область белого вещества как «отправную точку», ученые смог-
ли выявить и визуализировать связи каждого кластера. Их можно разделить на три
основные группы: одни связи идут между передней и задней частями мозга; другие
слева направо; и третьи идут от верхней части мозга вниз.
Таким образом, удалось определить, какие области белого вещества важны для раз-
вития способности к чтению у детей, и в то же время показать, какими проводящими
путями они представлены. Вслед за этим открытием появилась возможность фор-
мулировать более четкие гипотезы о нервном субстрате формирования способности
чтения, о том, как эти области могут быть функционально связаны, и исследовать их
при помощи других методов, например фМРТ.
Рис. А .27. Паттерны связей в мозге, выявленные с использованием ДТВ. (а) Пока-
заны области мозга (фиолетовые), которые были выбраны для исследования. Четыре
области находились в левом полушарии и одна в правом. (б) Показаны тракты белого
вещества (желтые) для выбранных областей, которые имеют передне-заднюю (спе-
реди мозга к затылку) ориентацию. (в) Показаны тракты белого вещества (красные)
для выбранных областей, которые имеют ориентацию слева направо. (г) Показаны
тракты белого вещества (синие) для выбранных областей, которые имеют дорсо-
вентральную (сверху вниз в мозге) ориентации. (Источник: Beaulieu et al., 2005.)
370
Приложение
в мозге, что и показано на рис. А .28. Он иллюстрирует спектр из определенной
области мозга и соответствующее химическое распределение в этом регионе.
Если сравнить распределение в здоровой области с областью пораженного бо-
лезнью мозга, то можно увидеть значительные различия. Так, в здоровом по-
лушарии мозга сигнал от N-ацетиласпартата (NAA) наивысший, среди других
химических соединений. NAA является маркером созревания аксонального
белого вещества в живом мозге (Bjartmar et al., 2002). В отличие от этого тот
же сигнал значительно снижается, если мы производим измерения в той же об-
ласти в поврежденном полушарии мозга. Даже без таких серьезных эффектов
МРС может использоваться при изучении дегенерации тканей мозга при таких
болезнях, как множественный склероз (De Stefano et al., 2005), инсульт (Saun-
Saun-
ders, 2000) и болезнь Альцгеймера (�alini et al., 2005), а также эффекты злоу-
потребления некоторыми веществами (Magalhaes, 2005; Reneman et al., 2006).
2.5 .
МРТ — метод будущего
Магнитно-резонансный сканер с его высокой пространственной разрешающей
способностью и широким применением является незаменимым инструментом
в когнитивной нейронауке. Усовершенствования в МРТ происходят во всем,
но, в целом, можно выделить две категории развития: технические и анали-
тические устройства. Технические усовершенствования включают создание
сканеров, создающих более сильное поле. Увеличение силы магнитного поля
Рис. А.28 . Спектроскопическое МРТ определяет химические различия при па-
тологии мозга. Слева: (зеленый) здоровая часть мозга имеет нормальные уровни
N-ацетиласпартата (NAA) (зеленый пик), в то время как на правой стороне NAA-
уровень падает, а уровень лактата значительно изменяет (красный) падение. (Источ-
ник: �ars Hanson, Датский исследовательский центр магнитного резонанса, Госпиталь
Университета Копенгагена, Дания.)
Методы для изучения живого мозга
371
улучшает способность сканера регистрировать сигналы. Так, заменяя сканер
с силой поля 1,5 Тесла (1,5 Т) сканером с силой поля 3 Т, мы получаем более
высокое пространственное разрешение как для структурных, так и функцио-
нальных изображений (рис. А.29). Несмотря на то что 3 Т и 4 Т являются сегод-
ня стандартами используемой силы поля, появляются сканеры с еще большей
силой поля, такие как 7 Т и 11 Т. Сканирование коры полем в 7 Т и выше дает
возможность выделять разные слои в коре (�atterpekar et al., 2002; Zhao et al.,
2006). Это очень важно не только потому, что мы получаем лучшее разрешение
для уже созданных исследований, но и из-за того, что у ученых появятся новые
идеи по изучению мозга и пониманию его работы.
Совершенствования силы поля дополняются другими техническими успеха-
ми, такими как улучшение аппарата, генерирующего магнитные импульсы, или
регистрирующего устройства, которое воспринимает сигналы. Эти происходя-
щие изменения, очень вероятно, внесут значительный вклад в возможность ска-
нировать мозг с чрезвычайно высокой разрешающей способностью и иногда в
формирование новых подходов получения данных. Несмотря на то что данные
усовершенствования обещают усиление разрешения, они также связаны с не-
которыми специфическими трудностями. Например, повышение силы поля,
что улучшает соотношение сигнал–шум в большинстве областей мозга, ведет к
значительной потере сигнала в некоторых областях. Так как BO�D-сигнал (см.
ранее) зависит от относительного объема оксигенированной крови в регионе,
области, которые имеют кислород, не являющийся частью крови, влияют на
сигнал. Медиальная височная доля и вентральная префронтальная кора лежат
рядом с носовыми воздушными пазухами. В результате BO�D-сигнал в этих
областях испорчен сигналом из не мозговых областей, заполненных кислоро-
дом. При переходе от силы поля в 1,5 Т к 3 Т, это стало проблемой, которую
нельзя игнорировать. фМРТ-данные по активации гиппокампа, миндалины и
орбитофронтальной коры были искажены потерей сигнала при 3 Т. Для того
чтобы избежать этого, сейчас в исследованиях используют специализирован-
Рис. А.29 . Увеличение силы поля ведет
к повышению разрешающей способно-
сти. При сравнении изображений одно-
го испытуемого, сделанных на сканерах
с силой поля 1,5 Т и 3 Т, можно увидеть
увеличение разрешения изображений.
Здесь патологические изменения мозга,
возникающие при склерозе, лучше вид-
ны при использовании 3 Т (б и г), тогда
как некоторые повреждения не видны
дажена1,5Т(аив,см.стрелкунаг).
Все изображения были получены при
введении контрастного агента гадоли-
ния. (Источник: Nielsen et al., 2006.)
372
Приложение
ные последовательности, которые уменьшают артефакты в этих областях. Од-
нако с переходом от стандартных 3 Т или 4 Т сканеров к сканерам с силой поля
7 Т и выше эти проблемы возникают снова. Только усовершенствования в мето-
дах погашения шума могут устранить эти недостатки.
Другой областью прогресса является усовершенствование программно-
го обеспечения. За несколько лет в области нейровизуализации произошел
значительный прорыв в числе способов обработки данных. Множество этих
улучшений привязано не только к МРТ, а также значимы для большинства или
всех подходов по нейровизуализации. В то время как в ранние годы фМРТ-
визуализация анализировала блоки проб, позже, новая связанная с событием
парадигма привела к образованию новых аналитических методов. Одним из
них является анализ паттернов нервной активации с помощи динамического
причинно-следственного моделирования (ДПСМ). В общем, ДПСМ является
способом определить, как активация в мозге вызвана или возникает в опреде-
ленное время. Существует два способа проанализировать эти данные. Анализ
функциональной взаимосвязи фокусируется на корреляции между областями
мозга. Например, кто-то может спросить, является ли случайной одновремен-
ная активация дорсолатеральной префронтальной коры (ДЛ-ПФК) и гиппокам-
па во время задачи на кодирование. Так, можно вычислить, насколько сильно
коррелируют две (или более) области мозга — иными словами, силу относи-
тельной корреляции двух или более областей. Второй способ анализа заклю-
чается в поиске эффективной взаимосвязи между областями. При этом ученые
пытаются выйти за пределы простой корреляции и вычислить причинно-след-
ственную связь и эффекты взаимодействия между областями. В нашем при-
мере с гиппокампом и ДЛ-ПФК, ученые не только задают вопрос о том, как
коррелируют эти области, но и как одна область вызывает активацию в другой.
Сегодня оба этих подхода постоянно развиваются и интенсивно используются
для анализа результатов нейровизуализации. Однако так как они опираются на
специфическую экспериментальную парадигму и регистрацию данных, могут
быть использованы только специально созданные схемы экспериментов.
2.6 .
Оптическое картирование
Оптическое картирование — современное дополнение к методам визуализации
мозга. Лазерный луч из света ближней инфракрасной области размещают над
черепом. Пучок оптических волокон используют как детекторы и размещают
на несколько сантиметров вдали от источника света. Детекторы регистрируют,
как луч света из лазерного источника изменяется, вследствие либо поглоще-
ния, либо отражения, при прохождении через мозговую ткань. Информация ис-
пользуется двумя способами. Во-первых, можно измерять концентрацию хими-
ческих веществ в мозге, определяя степень поглощения света в этой области.
Во-вторых, этот метод может дать информацию о физиологических свойствах
мозга, которые связаны с уровнем разрядов нейронов. Эти сведения получа-
ют, измеряя рассеяние света, что является показателем набухания глиальных
клеток и нейронов. Таким образом, оптическое картирование обеспечивает
мгновенное измерение как источника, так и динамики нейронной активации
в пределах определенной области мозга.
В работе с использованием оптического картирования (Sato et al., 2005) из-
учалась активность соматосенсорной коры на стадии планирования хирурги-
Методы для изучения живого мозга
373
ческой операции у больных с опухолями или эпилепсией. Было обнаружино,
что организация соматосенсорной коры имеет прилегающие области ответов,
например, для пальцев, и области имеют определенное функциональное пере-
крытие, что соответствует предыдущим литературным данным об исследова-
ниях на человеке и животных (рис. А.30). Также заслуживает внимания необ-
ходимость рассмотрения взаимосвязи между этими результатами и данными
исследования пластичности мозга при фантомных ощущениях конечностей,
рассмотренной в блоке А.2 . Эта работа подтверждает возможность использо-
вания оптического картирования при подготовке к хирургическим операциям.
Рис. А.30. Оптическое картирование: видно, как кора отвечает на стимуляцию. Внутрен-
ний оптический ответ левой соматосенсорной коры 47-летнего пациента на стимуляцию
пальцев. Зарегистрированные оптические ответы для иллюстрации окрашены и нало-
жены на изображение сосудов. (Источник: адаптировано из Sato et al., 2005.)
374
Приложение
3.0 .
Мультимодальные методы визуализации мозга
Сегодня нейронаука опирается на определенное число различных методов (мо-
дальностей) визуализации, и мы рассмотрели только самые важные. Каждый
подход продолжает давать новые результаты, которые вносят вклад в наше по-
нимание мозга и когнитивных процессов. Но, как показано на рис. А .1, каждый
метод визуализации имеет свои преимущества и недостатки, особенно относи-
тельно их пространственного и временного разрешения. Очевидное решение
этой проблемы — сравнить результаты работ, использовавших разные техно-
логии визуализации. Однако, так как каждый метод отличается от остальных,
мы никогда не сможем получить идентичных данных. Вместо этого мы полу-
чаем дополнительную информацию об области нашего интереса. Например,
если наше исследование нацелено на изучение процессов чтения вслух, мы мо-
жем сравнить результаты ЭЭГ и фМРТ. Таким образом, мы получаем хорошее
представление о локализации областей, которые ответственны за чтение вслух
(фМРТ) и в то же время получаем результаты о миллисекундных изменениях
уровней активации при выполнении этого задания (ЭЭГ).
Модальности визуализации можно сравнить разными способами. В дополне-
ние к этому, для сравнения результатов разных исследований очень часто при-
влекают технологии визуализации. ПЭТ, как правило, регистрируется совмест-
но со стандартной МРТ или КТ структур мозга. То же самое обычно делают для
фМРТ: изображения активации снимаются совместно со структурными изобра-
жениями. Это дает нам возможность лучше определить, где в мозге возникают
выявленные изменения. Активационные изображения сами по себе несут очень
мало информации о локализации активации, чтобы быть значимыми.
3.1.
Одновременная визуализация разными методами
В то время как большую часть сравнений производят между данными, зареги-
стрированными отдельно, например ПЭТ-изображения и изображения струк-
турной МРТ, существует возможность одновременной регистрации данных.
Например, ЭЭГ и ПЭТ были совмещены для изучения вклада областей мозга
в разные стадии сна (Dang-Vu et al., 2005). Регистрация ЭЭГ и ПЭТ произво-
дилась одновременно, пока испытуемый спал. Затем исследователи идентифи-
цировали отдельные стадии сна по записям ЭЭГ, сфокусировались на дельта-
активности во время не-БДГ (НБДГ) сна и сравнили с БДГ сном и нормальным
бодрствованием (см. рис. А .31). Ученые обнаружили отрицательную корреля-
цию между мощностью дельта-активности и локальным мозговым кровотоком
в вентомедиальной префронтальной коре, основании переднего мозга, стриату-
ме, передней части островка, и предклинье. Эти результаты дают представле-
ние об областях, которые различаются по уровню дельта-активности в зависи-
мости от уровня сознания.
Аналогично, Лауфс с коллегами (�aufs et al., 2003) совместили ЭЭГ и МРТ
при обследовании субъектов в состоянии покоя. Работы по изучению «состо-
яния покоя» должны рассматриваться как спорные, так как ни разум, ни мозг
никогда не пребывают в состоянии «покоя». Лауфс с коллегами выявили раз-
ные паттерны активности, возникающие на разных фазах покоя у испытуемого,
и обнаружили, что состояние покоя имеет на ЭЭГ «переплетенные и тем не ме-
нее раздельные динамические мозговые процессы». Другими словами, работы
Методы для изучения живого мозга
375
по нейровизуализации указывают, что мозг у испытуемых в состоянии покоя за-
нят разными процессами в разные периоды времени. Более того, ученые смогли
выделить отдельные нервные сети, лежащие в основе ритмов ЭЭГ, таких как
альфа- и бета-активности.
Необходимо отметить, что существует значительная разница в совместной
регистрации ЭЭГ с ПЭТ или фМРТ. В то время как ПЭТ-сканеры не вызывают
артефактов, которые непреодолимы для ЭЭГ, МР-сканер образует артефакты в
ЭЭГ такие значительные, что только совсем недавно стало возможно успеш-
но использовать их комбинацию. Один подход заключается в регистрации ЭЭГ
между запусками фМРТ, и таким образом получать данные о состоянии со-
знания, т. е . бодрствует ли испытуемый или спит. Использование надлежащей
фильтрации шума позволило фильтровать артефакты от сканера и получать
необходимые ЭЭГ-измерения непосредственно во время регистрации фМРТ.
Таким образом, стало возможно совмещать высокое временное разрешение
ЭЭГ и высокое пространственное разрешение МРТ. И все же фильтрование ар-
тефактов является проблемой — мы можем наблюдать значительную потерю
исходного сигнала, а некоторые остаточные артефакты остаются (даже после
фильтрации). Необходимо с осторожностью интерпретировать результаты со-
вмещенных измерений, таких как ЭЭГ/фМРТ.
Рис. А.31 . Мозговой субстрат для дельта-ритма во время не-БДГ сна. Иллюстрации
отображают изменения в мозговом кровотоке (МК) как функции от мощности дель-
та-ритма во время глубокого сна. Активация показана на среднем саггитальном срезе
(сверху) и разных аксиальных срезах (снизу). (Источник: Dang-Vu, 2005.)
376
Приложение
3.2 .
Визуализация генетики
В области мультимодальных методов визуализации важные шаги были сделаны
для дополнения нашего понимания того, как гены формируют разум. Гены дей-
ствуют на молекулярном уровне тела, т. е . действуют как строительные блоки
для нейронов. Таким образом, индивидуальные различия в генетической ин-
формации могут повлиять на работу мозга значительным образом.
Естественная вариация в промоторной зоне гена (названного 5-HTT�PR) се-
HTT�PR) се-
) се-
ротонинового транспортера может быть связана с различиями в образовании
серотонина, а также его обратного захвата. У индивидуумов с двумя копия-
ми длинной (l) модификации промоторного участка отмечена более высокая
концентрация мРНК серотонинового транспортера, а следовательно, и повы-
шенный уровень захвата серотонина, по сравнению с индивидуумами с дву-
мя короткими (s) версиями участка. У таких людей будет относительно низкая
концентрация транспортера и, в результате, относительно высокий уровень
серотонина в синаптической щели. Значит, генетический состав, которым вы
обладаете в этой области, влияет на количество доступного серотонина в моз-
ге. Просто имея генетическую информацию относительно гена серотониново-
го транспортера, сейчас возможно предсказать уровень активного серотонина
у любого человека.
Совмещение нейровизуализации и определения генотипа (Hariri et al., 2002)
показало, что набор генетической информации оказывает влияние на ответы
миндалины на предъявления лиц с разными выражениями (рис. А .32). Испы-
туемые с короткой генетической версией продемонстрировали повышенный
уровень активации миндалины, вероятно, вызванный более высокими концен-
трациями у них серотонина. Эта генетически обусловленная разница в возбу-
Рис. А.32 . Гены, гнев и миндалина. (а) Самоотчет об уровне гнева (на шкале ГНЕВ)
выше у мужчин с высоким уровнем ТТ и СТ-пар последовательностей гена DARPP-32,
по сравнению с СС-последовательностью этого отрезка ДНК у тех же испытуемых.
(б) На МРТ показано, что у этих испытуемых объем серого вещества (тел нервных
клеток) к левой миндалине значительно ниже, чем в контрольной группе. (Источник:
Reuter et al., 2009.)
Методы для изучения живого мозга
377
димости миндалины может вносить вклад в увеличение тревожности и страха,
которые обычно ассоциируются с коротким генетическим участком. По суще-
ству, данная работа является мощной демонстрацией того, что может вызывать
индивидуальные различия в работе мозга (рис. А .33). Сегодня многие ученые
склоняются к мысли, что область «визуализации генетики» обещает дать значи-
тельные новые данные о работе мозга и разуме.
Транскраниальная магнитная стимуляция
Основываясь на результатах разных методов визуализации, можно получить
улучшенное понимание взаимосвязи между локальной мозговой активацией и
сознанием. Однако эти работы очень страдают от корреляций: они часто про-
сто подразумевают взаимосвязь между активацией и сознанием. Представьте,
что при выполнении определенного задания испытуемым вы обнаруживаете,
что три области мозга, похоже, играют важную роль для этого процесса. Но
для того, чтобы выйти за рамки этих результатов, вам необходимо исследовать
эффекты разрушения этих областей. Повреждения мозга у пациентов — цен-
ный источник для проведения таких работ, но разрушения очень редко следуют
анатомическим границам или рамкам функциональной активации. Поэтому ис-
пользование методов, которые обратимы и влияют на важные процессы в ма-
ленькой области мозга, очень привлекательно.
Транскраниальная магнитная стимуляция (ТМС) является одним из та-
ких методов. В ТМС быстроменяющиеся магнитные поля (электромагнитная
индукция) используются для создания слабых электрических токов в мозге
Рис. А.33 . Многоуровневое понимание мозга и разума. Четыре уровня понимания ра-
зума и мозга. Визуализация генетики позволяет определять генетические эффекты на
уровне обработки информации в мозге, что дает более четкие биологические связи с
генами, а также обязательное промежуточное звено поведения. (Источник: адаптиро-
вано из Hariri et al., 2006.)
378
Приложение
(рис. А .34). Хотя точная природа процессов ТМС полностью еще не понятна,
ученые выделяют два вида ТМС: единично/парно импульсную ТМС (е/пТМС)
и повторяющуюся ТМС (пТМС).
В е/пТМС импульсы вызывают деполяризацию и генерацию потенциалов
действия в популяциях нейронов неокортекса. Если ее применить к первичной
зрительной коре, возбуждения этой области заставят человека видеть малень-
кие вспышки света (фосфены). Если ее приложить к моторным представитель-
ствам руки в коре, то можно вызвать движение руки или изменять силу напря-
жения мышц в руке.
Аналогично, е/пТМС можно использовать для грубого торможения актива-
ции в определенной области неокортекса. Например, благодаря прикладыванию
ТМС-импульсов билатерально к височной доле были (Overgaard et al., 2004)
продемонстрированы изменения в осознании восприятии зрительных стиму-
лов, но не в способности правильно угадывать принадлежности стимулов.
Таким образом, ТМС может быть использована для того, чтобы вызывать
временные и обратимые «выключения» областей у здоровых испытуемых. Этот
метод можно использовать для временного нарушения функций определенной
области. Если это повлияет на выполнение задания, можно предположить, что
данная часть мозга была вовлечена в выполнение задания в тот конкретный
момент.
Рис. А.34 . (а) ТМС. Когда исследователи двигают ТМС-катушку около головы испы-
туемого, мощное (• 2 Т) и кратковременное (• 300 мс) изменяющееся магнитное поле
безболезненно проходит через кожу и кость. Из-за того что сила магнитного поля падает
очень быстро с увеличением расстояния от ТМС-катушки, оно может распространиться
только на несколько сантиметров, т. е . только поверхностные области коры можно сти-
мулировать эффективно. Индуцированное электрическое поле вызывает электрические
токи в прилежащих нейронах, следовательно, стимулирует выбранные области коры.
(б) тСПТ (транскраниальная стимуляция переменным током). Этот метод основан на
прикладывании к электродам переменного тока. Электрический ток подается непрерыв-
но с низкой интенсивностью (1 мА) в течение периода времени в несколько секунд для
достижения изменений в корковой активности. Форма стимулирующего сигнала пред-
ставляет собой синусоиду, и можно использовать разную частоту во время стимуляции
(до 250 Гц). (Источник: Thut and Miniussi, 2009.)
Методы для изучения живого мозга
379
Второй ТМС-метод, повторяющаяся ТМС, вызывает эффекты, которые
длятся дольше времени стимуляции. Прикладывая повторяющуюся последова-
тельность ТМС-импульсов, можно значительно увеличить или уменьшить воз-
будимость кортикоспинальных или кортикокорковых путей. Хотя механизмы,
вовлеченные в пТМС, еще до конца не поняты, было выявлено, что уровень
возбудимости зависит от интенсивности стимуляции, ориентации катушки и
частоты стимуляции. Сделано предположение, что пТМС работает, меняя си-
наптическую эффективность, как долговременная потенциация (ДВП) и долго-
временная депрессия (ДВД) (�itzgerald et al., 2006).
В другом исследовании (Babiloni et al., 2007) показано, что пТМС вентраль-
ной теменной коры влияла как на процессы внимания, так и на осознание ис-
пытуемым наличия стимулов.
В последние годы исследования также выявили, что приложение пТМС
к определенным областям может привести к нейрофизиологическим изменени-
ям. Например, работы показали, что пТМС префронтальной коры вызывает из-
менения уровня дофамина в стриатуме и хвостатом ядре (Straffella et al., 2001;
2003), вероятно, действуя через кортикостриарные пути.
Несмотря на различное применение, ТМС относится к методам, которые
еще набирают популярность среди ученых. К тому же этот метод продемон-
стрировал некие перспективы для лечения депрессии и других эмоциональных
расстройств, а также болезни Паркинсона.
4.0 .
Заключение
Нейровизуализация развилась и стала одним из самых главных подходов к наше-
му растущему пониманию человеческого мозга и разума. Усовершенствования
каждого метода — будь то оптическое картирование, ЭЭГ или фМРТ — быстро
меняют не только способы обработки информации, но и способы постановки
вопросов. Продолжающиеся инновации, как в технике, так и в программном
обеспечении, делают невозможным предсказать будущее нейровизуализации.
Однако некоторые тренды, скорее всего, будут продолжены, включая увеличе-
ние разрешающей способности во всех методах и новые способы совмещения
данных, полученных разными методами.
Мы должны всегда помнить, что в действительности мы изучаем. Иссле-
дование человеческого разума и его мозга основано на биологических строи-
тельных блоках нашего тела: наших генах. Наши гены определяют, как и когда
наш мозг развивается, и, следовательно, они являются рецептом для нашего
разума. Вот что мы должны пытаться понять. Для того чтобы это сделать, нам
надо понять уровни, на которых мы работаем в поисках понимания самих себя.
Давайте разберемся в четырех уровнях. Во-первых, то, как мы понимаем гены,
формирует знание о составе клеточных компонентов, таких как белки и нейро-
медиаторы. Во-вторых, мы должны понять, как работает единичный нейрон и
как нейроны взаимодействуют в сети. В -третьих, мы можем изучить мозг и вы-
явить морфологические и функциональные факторы, влияющие на мышление.
И в-четвертых, мы можем изучать эффекты любого из предыдущих уровней на
наше мышление, поведение и даже личность.
380
Приложение
Литература
1. Babiloni, C., Vecchio, �., Rossi, S., De Capua, A., Bartalini, S., Ulivelli, M., and
Rossini, P.M . (2007). Human ventral parietal cortex plays a functional role on vi-
, P.M . (2007). Human ventral parietal cortex plays a functional role on vi-
P.M . (2007). Human ventral parietal cortex plays a functional role on vi-
suospatial attention and primary consciousness. A repetitive transcranial magnetic
stimulation study. Cereb Cortex, 17(6), 1486–1492.
2. �itzgerald, P.B., �ountain, S., and Daskalakis, Z.J. (2006). A comprehensive
review of the effects of rTMS on motor cortical excitability and inhibition. Clin
Neurophysiol, 11 7 (12), 2584–2596.
3. Jääskelänen, I.P., Ahveninen, J., Bonmassar, G., Dale, A.M ., Ilmoniemi, R.J.,
�evanen, S., et al. (2004). Human posterior auditory cortex gates novel sounds
to consciousness. Proceedings of the National Academy of Sciences, USA, 101,
6809–6814.
4. Näätänen, R. (1992). Attention and brain function. Hillsdale, NJ: Erlbaum.
5. Näätänen, R., Paavilainen, P., Rinne, T., and Alho, K. (2007). The mismatch
negativity (MMN) in basic research of central auditory processing: A review.
Clinical Neurophysiology, 118 , 2544–2590.
6. Näätänen, R., and Winkler, I. (1999). The concept of auditory stimulus
representation in cognitive neuroscience. Psychological Bulletin, 125, 826–859.
7. Overgaard, M., Nielsen, J.�., and �uglsang-�rederiksen, A. (2004). A TMS study
of the ventral projections from V1 with implications for the finding of neural
correlates of consciousness. Brain Cogn, 54(1), 58–64.
8. Strafella, A.P., Paus, T., Barrett, J., and Dagher, A. (2001). Repetitive transcranial
magnetic stimulation of the human prefrontal cortex induces dopamine release in
the caudate nucleus. J Neurosci, 21(15), R�157.
9. Strafella, A.P., Paus, T., �raraccio, M., and Dagher, A. (2003). Striatal dopamine
release induced by repetitive transcranial magnetic stimulation of the human
motor cortex. Brain, 126(Pt 12), 2609–2615.
Abraham, W. C ., & Robins, A. (2005). Memory retention — the synaptic stability
versus plasticity dilemma. Trends in Neurosciences, 28(2), 73–78.
Achard, S., Salvador, R., Whitcher, В., Suckling, J., & Bullmore, E. (2007).
A resilient, low-frequency, small-world human brain functional network
with highly connected association cortical hubs. January 4, 2006. Journal of
Neuroscience, 26(1), 63–72 .
Adams, К. H ., Hansen, E. S., Pinborg, L. H ., et al. (2005). Patients with obsessive-
compulsive disorder have increased 5-HT2a receptor binding in the caudate
nuclei. International Journal of neuropsychopharmacology, 8(3), 391–401.
Adams, К. H ., Pinborg, L. H . Svarer, C., et al. (2004). A database of [(18)f]-
altanserin binding to 5-ht(2a) receptors in normal volunteers: Normative data
and relationship to physiological and demographic variables. Neuroimage, 21(3),
1105–1113.
Adelman, G., & Smith, В. H . (2004). Encyclopedia of neuroscience (3rd ed.).
Amsterdam: Elsevier.
Adolphs, R., Gosselin, F., Buchanan, T. W., Tranel, D., Schyns, P., & Damasio, A. R.
(2005). A mechanism for impaired fear recognition after amygdala damage.
Nature, 433(7021), 68–72 .
Ahissar, M., & Hochstein, S. (2004). The reverse hierarchy theory of visual perceptual
learning. Trends in Cognitive Sciences, 8(10), 457–464.
Albright, T. D . (1984). Direction and orientation selectivity of neurons in visual area
MT of the macaque. Journal of Neurophysiology, 52(6), 1106–1130.
Albright, T. D . (1992). Form-cue invariant motion processing in primate visual
cortex. Science, 255(5048), 1141–1143.
Aleksander, I. (2005). The world in my mind, my mind in the world.
Aleksander, I., Browne, C., Dunmall, В., & Wright, T. (1998). Towards visual
awareness in a neural system. In S. I. Kasabov (Ed.), Brain-like computing and
intelligent information systems (pp. 513–533). Berlin: Springer.
Alitto, H. J., & Ursey, W. M. (2003). Corticothalamic feedback and sensory
processing. Current Opinion in Neurobiology, 13(4), 440–445 .
Alkire, M. Т., Hudetz, A. G., & Tononi, G. (2008). Consciousness and anesthesia.
Science, 322(5903), 876–880 .
Alle, H., & Geiger, J. R. (2006). Combined analog and action potential coding in
hippocampal mossy fibers. Science, 311(5765), 1290–1293.
Allison, Т., Puce, A., & McCarthy, G. (2000). Social perception from visual cues:
Role of the STS region. Trends in Cognitive Sciences, 4(7), 267–278.
Литература
382
Литература
Aminoff, M., & Daroff, R. (Eds.), (2003). Encyclopedia of the neurological sciences.
San Diego: Academic Press.
Anderson, A. K ., & Phelps, E. A . (2001). Lesions of the human amygdala impair
enhanced perception of emotionally salient events. Nature, 421(6835), 305–309 .
Anderson, N. D., Iidaka, Т., Cabeza, R., Kapur, S., McIntosh, A. R., & Craik, F. I.
(2000). The effects of divided attention on encoding- and retrieval-related
brain activity: A PET study of younger and older adults. Journal of Cognitive
Neuroscience, 12(5), 775–792.
Anderson, S. W., Bechara, A., Damasio, H., Tranel, D., & Damasio, A. R. (1999).
Impairment of social and moral behavior related to early damage in human
prefrontal cortex. Nature Neuroscience, 2(11), 1032–1037.
Andoh, J., Artiges, E., Pallier, C., et al. (2006). Modulation of language areas with
functional MR image-guided magnetic stimulation. Neuroimage, 29(2), 619–627.
Andrew, C. N., Chen, F. - J., Wang, L., & Arendt-Nielsen, L. (2006). Mode and site
of acupuncture modulation in the human brain: 3d (124-ch) EEG power spectrum
mapping and source imaging. Neuroimage, 29(4), 1080–1091.
Angecine, J. В ., Jr., in Ramachandran (2002) Encyclopedia of the human brain. San
Diego: Elsevier.
Arthurs, O. J., & Boniface, S. (2002). How well do we understand the neural origins
of the fMRI bold signal? Trends in Neurosciences, 25(1), 27–31.
Atkinson, R. C., & Shiffrin, R. M . (1968). Human memory: a proposed system and
its control processes. The psychology of learning and motivation: II. Oxford:
Academic Press.
Attwell, D., & Laughlin, S. B . (2001). An energy budget for signaling in the grey
matter of the brain. Journal of Cerebral Blood Flow and Metabolism, 21(10),
1133–1145.
Awh, E., Armstrong, К. M ., & Moore, T. (2006). Visual and oculomotor selection:
Links, causes and implications for spatial attention. Trends in Cognitive Sciences,
10(3), 124–130.
Awh, E., Jonides, J., Smith, E. E., Schumacher, E. H., Koeppe, R. A., & Katz, S.
(1996). Dissociation of storage and rehearsal in verbal working memory: Evidence
from positron emission tomography. Psychological Science, 7(1), 25–31.
Aylward, E. H . Park, J. E., Field, К. M ., et al. (2005). Brain activation during
face perception: Evidence of a developmental change. Journal of Cognitive
Neuroscience, 17(2), 308–319.
Baars, B. J. (1983). Conscious concepts provide the nervous system with coherent,
global information. In R. J. Davidson (Ed.), Consciousness and self-regulation.
New York: Plenum Press.
Baars, B. J. (1986). What is a theory of consciousness a theory of? The search for
criterial constraints on theory. Imagination Cognition Personality, 6(1), 3.
Baars, B. J. (1988). A cognitive theory of consciousness. New York: Cambridge
University Press.
Baars, B. J. (2002). The conscious access hypothesis: Origins and recent evidence.
Trends in Cognitive Sciences, 6(1), 47–52 .
Baars, B. J., Banks, W. P., & Newman, J. B. (2003a). Essential sources in the scientific
study of consciousness. Cambridge: MIT Press.
Литература
383
Baars, B. J., & Franklin, S. (2003). How conscious experience and working memory
interact. Trends in Cognitive Sciences, 7(4), 166–172 .
Baars, B. J., Ramsoy, T. Z., & Laureys, S. (2003b). Brain, conscious experience and
the observing self. Trends in Neurosciences, 26(12), 671–675.
Bacon, E. R., Chatterjee, S., & Williams, M. (2007). Sleep. In Triggle, D. J., & Taylor,
J. В., Comprehensive Medicinal Chemistry II. Vol. 6,139–167.
Baddeley, A. (2000). The episodic buffer: a new component of working memory?
Trends in Cognitive Sciences, 4(11), 417–423.
Baddeley, A. (2003). Working memory: Looking back and looking forward. Nature
Reviews Neuroscience, 4(10), 829–839 .
Baddeley, A. D. (2002). Is working memory still working? European Psychologist,
7(2), 85.
Baddeley, A. D., & Hitch, G. J. (1974). Working memory. In G. A. Bower (Ed.),
Recent advances in learning and motivation: Vol. 8 (pp. 47 –90). New York:
Academic Press.
Baddeley, Т. C., Davidson, I. G ., Glidewell, C., Low, J. N ., Skakle, J. M . & Wardell,
J. L . (2004). Supramolecular structures of substituted alpha, alpha’-trehalose
derivatives. Acta Crystallographica. Section B, 60(Pt 4), 461–471 .
Baillargeon, R. (1993). The object concept revisited: New directions in the
investigation of infant’s physical knowledge. In С. E . Granrud (Ed.), Visual
perception and cognition in infancy (pp. 265–315). Hillsdale: Lawrence Erlbaum
Associates.
Baird, A. A., Kagan, J., Gaudette, Т., Walz, K. A., Hershlag, N., & Boas, D. A. (2002).
Frontal lobe activation during object permanence: Data from near-infrared
spectroscopy. Neuroimage, 16(4), 1120–1125 .
Bakin, J. S., South, D. A ., & Weinberger, N. M. (1996). Induction of receptive field
plasticity in the auditory cortex of the guinea pig during instrumental avoidance
conditioning. Behavioral Neuroscience, 110(5), 905–913.
Banaji, M. R ., & Greenwald, A. G. (1995). Implicit gender stereo-typing in
judgments of fame. Journal of Personality and Social Psychology, 68(2),
181–198.
Bar, M., Tootell, R. В., Schacter, D. L ., et al. (2001). Cortical mechanisms specific to
explicit visual object recognition. Neuron, 29(2), 529–535.
Barbeau, E., Wendling, F., Regis, J., et al. (2005). Recollection of vivid memories
after perirhinal region stimulations: Synchronization in the theta range of spatially
distributed brain areas. Neuropsychologia, 43(9), 1329–1337.
Bargh, J. A . (2006). What have we been priming all these years? On the development,
mechanisms, and ecology of nonconscious social behavior. European Journal of
Social Psychology, 1.
Bargh, J. A ., & Chartrand, T. L . (1999). The unbearable automaticity of being. The
American Psychologist, 54(7), 462.
Bargh, J. A., & Williams, E. L . (2006). The automaticity of social life. Current
Directions Psychological Science, 15,1–4 .
Barkley, R. A. (1997). ADHD and the nature of self-control. New York: The Guilford
Press.
Barlow, H. В., Blakemore, C., & Pettigrew, J. D. (1967). The neural mechanism of
binocular depth discrimination. Journal of Physiology, 193(2), 327–342.
384
Литература
Baron-Cohen, S. (1995). Mindblindness: An essay on autism and theory of mind.
Boston: MIT Press/Bradford Books.
Barrett, N. A., Large, M. M ., Smith, G. L., et al. (2003). Human brain regions required
for the dividing and switching of attention between two features of a single object.
Brain Research. Cognitive Brain Research, 17(1), 1–13.
Barsalou, L. W. (1999). Perceptual symbol systems discussion 610–660 . The
Behavioral and Brain Sciences, 22(4), 577–609.
Barsalou, L. W. (2005). Continuity of the conceptual system across species. Trends in
Cognitive Sciences, 9(7), 309–311.
Barsalou, L. W., Kyle Simmons, W., Barbey, A. K., & Wilson, C. D. (2003).
Grounding conceptual knowledge in modality-specific systems. Trends in
Cognitive Sciences, 7(2), 84–91.
Bartels, A., & Zeki, S. (2005). Brain dynamics during natural viewing conditions —
a new guide for mapping connectivity in vivo. Neuroimage, 24(2), 339–349.
Batty, M., & Taylor, M. J. (2006). The development of emotional face processing
during childhood. Developmental Science, 9(2), 207–220.
Beauchamp, M. S. (2005). See me, hear me, touch me: Multisensory integration in
lateral occipital-temporal cortex. Current Opinion in Neurobiology, 15(2), 145–
153.
Beaulieu, C., Plewes, C., Paulson, L. A., et al. (2005). Imaging brain connectivity in
children with diverse reading ability. Neuroimage, 25(4), 1266–1271 .
Bechara, A., Tranel, D., Damasio, H., Adolphs, R., Rockland, C., & Damasio, A. R.
(1995). Double dissociation of conditioning and declarative knowledge relative
to the amygdala and hippocampus in humans. Science, 269(5227), 1115–1118.
Beck, D. M., Rees, G., Frith, C. D ., & Lavie, N. (2001). Neural correlates of change
detection and change blindness. Nature Neuroscience, 4(6), 645–650.
Bell, M. A. (1992a). Electrophysiological correlates of object search performance
during infancy. Proceedings of the Vllth International Conference on Infant
Studies. Miami Beach.
Bell, M. A . (1992b). A not В task performance is related to frontal EEG asymmetry
regardless of locomotor experience. Proceedings of the VIIIth International
Conference on Infant Studies. Miami Beach.
Bell, M. A., & Fox, N. A . (1992). The relations between frontal brain electrical
activity and cognitive development during infancy. Child Development, 63(5),
1142–1163.
Ben-Ari, Y. (2008). Neuro-archaeology: Pre-symptomatic architecture and signature
of neurological disorders. Trends in Neurosciences, 31(12), 626–636.
Bennett, M. R. (1999). The early history of the synapse: From Plato to Sherrington.
Brain Research Bulletin, 50(2), 95–118.
Berardi, N., Pizzorusso, Т., Ratto, G. M ., & Maffei, L. (2003). Molecular basis of
plasticity in the visual cortex. Trends in Neurosciences, 26(7), 369–378.
Berger, H., & Gloor, P. (1969). Hans Berger on the electroencephalogram of man:
The fourteen original reports on the human electroencephalogram. Amsterdam:
Elsevier.
Berridge, К. C. (2000). Measuring hedonic impact in animals and infants:
Microstructure of affective taste reactivity patterns. Neuroscience and
Biobehavioral Reviews, 24(2), 173–198.
Литература
385
Berridge, К. C., & Robinson, Т. E . (1998). What is the role of dopamine in reward:
Hedonic impact, reward learning, or incentive salience? Brain Research. Brain
Research Reviews, 28(3), 309–369 .
Berridge, К. C., & Robinson, Т. E . (2003). Parsing reward. Trends in Neurosciences,
26(9), 507–513.
Berry, D. C ., & Dienes, Z. (1993). Implicit learning: Theoretical and empirical
issues. Erlbaum.
Bickerton, D. (1984). The language bioprogram hypothesis. The Behavioral and
Brain Sciences, 7(2), 212–218.
Bickerton, D. (1990). Language and species. Chicago: University of Chicago Press.
Binder, J. (1997). Functional magnetic resonance imaging. Language mapping.
Neurosurgery Clinics of North America, 8(3), 383–392.
Binder, J. R., Frost, J. A., Hammeke, T. A., et al. (2000). Human temporal lobe
activation by speech and non-speech sounds. Cerebral Cortex, 10(5), 512–528.
Bjartmar, C., Battistuta, J., Terada, N., Dupree, E., & Trapp, B. D. (2002).
N-acetylaspartate is an axon-specific marker of mature white matter in vivo:
A biochemical and immuno-histochemical study on the rat optic nerve. Annals of
Neurology, 51(1), 51–58.
Blake, R., & Logothetis, N. K. (2002). Visual competition. Nature Reviews.
Neuroscience, 3(1), 13–21 .
Blanke, O., Ortigue, S. Landis, Т., & Seeck, M. (2002). Stimulating illusory own-
body perceptions. Nature, 419(6904), 269–270.
Blemenfled, H. (2005). Consciousness and epilepsy: Why are patients with absent
seizures absent? In S. E . Laureys (Ed.), Progress in Brain Research, Vol. 150,
chapter 20.
Blumenthal, J. L ., & Elman, J. L . (1979). Wilhelm Wundt, the founding father we
never knew. Contemporary Psychology, 24(7), 547–550.
Blumstein, S. E . (1997). A perspective on the neurobiology of language. Brain and
Language, 60(3), 335–346.
Boatman, D. F. (2006). Cortical auditory systems: Speech and other complex sounds.
Epilepsy & Behavior, 8(3), 494–503.
Bodamer, J. L. (1947). Die prosopagnosie. Eur Arch Psychiatr Clinical Neuroscience,
179(1), 6–53.
Borgstein, J., & Grootendorst, C. (2002). Half a brain. Lancet, 359(9305), 473–475 .
Botvinick, M. M ., Cohen, J. D ., & Carter, C. S . (2004). Conflict monitoring and
anterior cingulate cortex: An update. Trends in Cognitive Sciences, 8(12), 539–
546.
Bourgeois, J. P. (2001). Synaptogenesis in the neocortex of the newborn: The
ultimate frontier for individuation? The handbook of developmental cognitive
neuroscience. Cambridge: MIT Press (pp. 23–34).
Bouvier, S. E., & Engel, S. A . (2006). Behavioral deficits and cortical damage loci in
cerebral achromatopsia. Cerebral Cortex, 16(2), 183–191.
Bowers, K. S ., Regehr, G., Balthazard, C., & Parker, K. (1990). Intuition in the
context of discovery. Cognitive Psychology, 22(1), 72–110.
Braver, T. S., Reynolds, J. R., & Donaldson, D. I . (2003). Neural mechanisms of
transient and sustained cognitive control during task switching. Neuron, 39(4),
713–726.
386
Литература
Bregman, A. S . (1990). Auditory scene analysis: The perceptual organization of
sound. Cambridge: MIT Press.
Broadbent, D. E . (1957). A mechanical model for human attention and immediate
memory. Psychological Review, 64(3), 205–215 .
Broadbent, D. E . (1982). Task combination and selective intake of information. Acta
Psychologica (amst), 50(3), 253–290.
Broca, P. (1861a). Pert de la parole, ramollissement chronique et destruction partielle
du lobe anterieur gauche en cerveau. Bulletin de la Societe Anthropology, 2, 235–
238.
Broca, P. (1861b). Remarques sur le siege de la faculte du langage articule, suives
d’une observation d’aphemie (pert de la parole). Bulletin de la Societe Anatomica,
6(330–357), 398–407.
Brodersen, P., Paulson, О. В., Bolwig, T. G., Rogon, Z. E ., Rafaelsen, O. J., & Lassen,
N. A . (1973). Cerebral hyperemia in electrically induced epileptic seizures.
Archives of Neurology, 28(5), 334–338.
Brodman, K. (1909). Vergleichende lakalisationslehre der grosshirnrinde: In ihren
prinzipien dargestellt anfgrund des zellenbaues. Leipzig: Verlag von Johann
Ambrosisus Barth.
Brooks, R., & Meltzoff, A. (2002). The importance of eyes: how infants interpret
adult looking behavior. Developmental Psychology, 38(6), 958–966 .
Brooks, R., Meltzoff, A. (2003). Gaze following at 9 and 12 months: A developmental
shift from global head direction to gaze. Poster presented at SRCD conference.
Society for Research in Child Development, Tampa.
Brown, M. C . (2003). Audition. In L. R. Squire, F. E. Bloom, S. K . McConnell, J. L .
Roberts, N. C. Spitzer, & M. J. Zigmond (Eds.), Fundamental neuroscience (2nd
ed.) . San Diego: Elsevier.
Brown, R. E ., & Milner, P. M . (2003). The legacy of Donald O. Hebb: More than the
Hebb synapse. Nature Reviews. Neuroscience, 4(12), 1013–1019.
Brown, R., & McNeill, D. (1966). The ‘tip of the tongue’ phenomenon. Journal of
Verbal Learning and Verbal Behavior, 5(4), 325.
Brugge, J. F., & Howard, M. A . (2003). Hearing. In L. R. Squire, F. E . Bloom, S. K.
McConnell, J. L . Roberts, N. C. Spitzer, & M. J. Zigmond (Eds.), Fundamental
neuroscience (2nd ed.) . San Diego: Elsevier.
Brugge, J. F., & Merzenich, M. M. (1973). Responses of neurons in auditory
cortex of the macaque monkey to monaural and binaural stimulation. Journal of
Neurophysiology, 36(6), 1138–1158.
Buccino, G., Vogt, S.
;
Ritzi, A., Fink, G., & Zilles, K. (2004). Neural circuits
underlying imitation learning of hand actions: An event-related fMRI study.
Neuron, 42, 323–334.
Buckley, M. J., & Gaffan, D. (2006). Perirhinal cortical contributions to object
perception. Trends in Cognitive Sciences, 10(3), 100–107.
Buckner, R. L ., Goodman, J., Burock, M., et al. (1998). Functional-anatomic correlates
of object priming in humans revealed by rapid presentation event-related fMRI.
Neuron, 20(2), 285–296.
Bullier, J. (2001). Feedback connections and conscious vision. Trends in Cognitive
Sciences, 5(9), 369–370.
Литература
387
Bunzeck, N., Wuestenberg, Т., Lutz, K., Heinze, H. J., & Jancke, L. (2005). Scanning
silence: Mental imagery of complex sounds. Neuroimage, 26(4), 1119–1127 .
Burgess, N., & Hitch, G. (2005). Computational models of working memory: Putting
long-term memory into context. Trends in Cognitive Sciences, 9(11), 535–541 .
Burgess, N., & Hitch, G. J. (1999). Memory for serial order: A network model of the
phonological loop and its timing. Psychological Review, 106(3), 551–581.
Bush, G., Luu, P., & Posner, M. I . (2000). Cognitive and emotional influences in
anterior cingulate cortex. Trends in Cognitive Sciences, 4(6), 215–222 .
Byrne, J. H ., & Roberts, J. L . (2004). From molecules to networks: An introduction to
cellular and molecular neuroscience. Elsevier Academic Press.
Cabeza, R., Mangels, J., Nyberg, L., et al. (1997). Brain regions differentially
involved in remembering what and when: A PET study. Neuron, 19(4), 863–870.
Cahill, L., Babinsky, R., Markowitsch, H. J., & McGaugh, J. L . (1995). The amygdala
and emotional memory. Nature, 377(6547), 295–296.
Cahill, L., Haier, R. J., Fallon, J., et al. (1996). Amygdala activity at encoding
correlated with long-term, free recall of emotional information. Proceedings of
the National Academy of Sciences of the United States of America, 93(15), 8016–
8021.
Cahill, L., & McGaugh, J. L . (1998). Mechanisms of emotional arousal and lasting
declarative memory. Trends in Neurosciences, 21(7), 294–299.
Cahill, L., Prins, В., Weber, M., & McGaugh, J. L . (1994). Beta-adrenergic activation
and memory for emotional events. Nature, 371(6499), 702–704.
Caldara, R., Segher, M., Rossion, В., Lazeyras, F., Michel, C., & Hauert, C. (2006).
The fusiform face area is tuned for curvilinear patterns with more high-contrasted
elements in the upper part. Neuroimage, 31, 313–319.
Canolty, R. Т., Edwards, E., Dalai, S. S., Soltani, M., Nagarajan, S. S., Kirsch, H. E.,
Berger, M. S., Barbara, N. M., & Knight, R. T. (2006). High gamma power is
phase-locked to theta oscillations in human neocortex. Science, 313, 1626–1628.
Caplan, D. N ., & Gould, J. L. (2003). Language and communication. In L. R. Squire,
F. E . Bloom, S. K . McConnell, J. L . Roberts, N. C. Spitzer, & M. J. Zigmond
(Eds.), Fundamental neuroscience (ed.). San Diego: Elsevier.
Caramazza, A., & Mahon, B. Z . (2003). The organization of conceptual knowledge:
The evidence from category-specific semantic deficits. Trends in Cognitive
Sciences, 7(8), 354–361.
Caramazza, A., & Shelton, J. R. (1998). Domain-specific knowledge systems in
the brain the animate-inanimate distinction. Journal of Cognitive Neuroscience,
20(1), 1–34.
Carpenter, G. A ., & Grossberg, S. (1991). Pattern recognition by self-organizing
neural networks. Cambridge: MIT Press.
Carpenter, M. В., & Parent, A. (1995). Carpenter’s human neuroanatomy (9th ed.).
Baltimore: Lippincot, Williams & Wilkins.
Carroll, D. N. (1999). Psychology of language. Pacific Cove, CA: Brooks/Cole
Publishing Co.
Casey, B. J., Cohen, J. D., Jezzard, P., et al. (1995). Activation of prefrontal cortex
in children during a nonspatial working memory task with functional MRI.
Neuroimage, 2(3), 221–229.
388
Литература
Casey, B. J., Tottenham, N., Liston, C., & Durston, S. (2005). Imaging the developing
brain: What have we learned about cognitive development? Trends in Cognitive
Sciences, 9(3), 104–110.
Catani, M., & ffytche, D. H. (2005). The rises and falls of disconnection syndromes.
Brain, 128(Pt 10), 2224–2239.
Caton, R. (1875). The electric currents of the brain. British Medical Journal, 2(1),
278.
Cattell, J. M . (1886). The time it takes to see and name objects. Mind, 11, 63–65.
Chambers, D., & McGonnell, I. M . (2002). Neural crest: Facing the facts of head
development. Trends in Genetics, 18(8), 381.
Champagne, F. A ., & Curley, J. P. (2009). Epigenetic mechanisms mediating the long-
term effects of maternal care on development. Neuroscience and Biobehavioral
Reviews, 33, 593–600 .
Chartrand, T. L., & Bargh, J. A. (1999). The chameleon effect: The perception-behavior
link and social interaction. Journal of Personality and Social Psychology, 76(6),
893–910.
Cheeseman, J., & Merikle, P. M . (1984). Priming with and without awareness.
Perception & Psychophysics, 36(4), 387–395.
Cheeseman, J., & Merikle, P. M . (1986). Distinguishing conscious from unconscious
perceptual processes. Canadian Journal of Psychology, 40(4), 343–367.
Chein, J. M . (2004). Evaluating models of working memory: fMRI and behavioral
evidence on the effects of concurrent irrelevant information. Dissertation. USA:
University of Pittsburgh.
Chein, J. M ., Ravizza, S. M ., & Fiez, J. A. (2003). Using neuro-imaging to evaluate
models of working memory and their implications for language processing.
Journal of Neurolinguistics, 16(4), 315–339 .
Chein, J. M ., & Schneider, W. (2005). Neuroimaging studies of practice-related
change: fMRI and meta-analytic evidence of a domain-general control network
for learning. Brain Research. Cognitive Brain Research, 25(3), 607–623.
Cherry, E. C. (1953). Some experiments on the recognition of speech, with one and
with two ears. The Journal of the Acoustical Society of America, 25, 975.
Chomsky, N. (1957). Syntactic structures. The Hague: Mouton.
Christensen, M. S., Ramsøy, T. Z ., Lund, Т. E., Madsen, К. H., & Rowe, J. B. (2006).
An fMRI study of the neural correlates of graded visual perception. Neuroimage,
32(4), 1711–1725 .
Christian, В. Т., Vandehey, N. Т., Fox, A. S., et al. (2009). The distribution of D2/
D3 receptor binding in the adolescent rhesus monkey using small animal PET
imaging. Neuroimage, 44,1334–1344.
Christiansen, M. H., Allen, J., & Seidenberg, M. S . (1998). Learning to segment
speech using multiple cues: A connectionist model. Language and Cognitive
Processes, 13(2/3), 221–268.
Chun, M. M ., & Phelps, E. A . (1999). Memory deficits for implicit contextual
information in amnesic subjects with hippocampal damage. Nature Neuroscience,
2(9), 844–847.
Clark, R. E ., & Squire, L. R. (1999). Human eyeblink classical conditioning: Effects
of manipulating awareness of the stimulus contingencies. Psychological Science,
10(1), 14–18.
Литература
389
Clarke, S., Bellmann Thiran, A., Maeder, P., et al. (2002). What and where in human
audition: Selective deficits following focal hemispheric lesions. Experimental
Brain Research, 147(1), 8–15.
Cleeremans, A. (1993). Mechanisms of implicit learning: Connectionist models of
sequence learning. MIT Press.
Cleeremans, A., Destrebecqz, A., & Boyer, M. (1998). Implicit learning: News from
the front. Trends in Cognitive Sciences, 2(10), 406–415 .
Cohen, D. (1968). Magnetoencephalography: Evidence of magnetic fields produced
by alpha-rhythm currents. Science, 162(843), 784–786.
Cohen, J. D ., et al. (1990). On the control of automatic processes: A parallel distributed
processing account of the Stroop effect. Psychological Revieiv, 97, 332–361.
Cohen, J., Perlstein, W., Braver, Т., et al. (1997). Temporal dynamics of brain
activation during a working memory task. Nature, 386(6625), 604–608 .
Cohen, N. J., & Squire, L. R. (1980). Preserved learning and retention of pattern-
analyzing skill in amnesia: Dissociation of knowing how and knowing that.
Science, 110(4466), 207–210.
Cole, M. W, & Schneider, W. (2007). The cognitive control network: Integrated
cortical regions with dissociable functions. Neuroimage, 37, 343–360 .
Colom, R., Jung, R. E., & Haier, R. J. (2006). Distributed brain sites for the g-factor
of intelligence. Neuroimage, 31(3), 1359–1365.
Computerised Wisconsin card sort task. (2003). Version (4 ed.), Vol. (WCST):
Psychological Assessment Resources.
Conway, A. R., Cowan, N., & Bunting, M. F. (2001). The cocktail party phenomenon
revisited: The importance of working memory capacity. Psychonomic Bulletin &
Revieiv, 8(2), 331–335.
Conway, M. A., Turk, D. J., Miller, S. L ., et al. (1999). A positron emission tomography
(PET) study of autobiographical memory retrieval. Memory, 7(5–6), 679–702.
Cooper, R., Crow, H. J., Walter, W. G ., & Winter, A. L. (1966). Regional control
of cerebral vascular reactivity and oxygen supply in man. Brain Research, 3(2),
174–191.
Corbetta, M., Akbudak, E., Conturo, Т. E ., et al. (1998). A common network of
functional areas for attention and eye movements. Neuron, 21(4), 761–773.
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, M. P., & Shulman, G. L.
(2000). Voluntary orienting is dissociated from target detection in human posterior
parietal cortex. Nature Neuroscience, 3(3), 292–297.
Corbetta, M., Kincade, J. M ., & Shulman, G. L. (2002). Neural systems for visual
orienting and their relationships to spatial working memory. Journal of Cognitive
Neuroscience, 14(3), 508–523.
Corbetta, M., & Shulman, G. L . (2002). Control of goal-directed and stimulus-driven
attention in the brain. Nature Reviews. Neuroscience, 3(3), 201–215 .
Corkin, S. (1965). Tactually-guided maze learning in man: effects of unilateral cortical
excisions and bilateral hippocampal lesions. Neuropsychologia, 3, 339–351.
Corkin, S., Amaral, D. G., Gonzalez, R. G ., Johnson, K. A ., & Hyman, В. T. (1997).
H. M .’s medial temporal lobe lesion: Findings from magnetic resonance imaging.
The Journal of Neuroscience, 17(10), 3964–3979.
Cosmelli, D. David, O., Lachaux, J. P., et al. (2004). Waves of consciousness:
Ongoing cortical patterns during binocular rivalry. Neuroimage, 23(1), 128–140.
390
Литература
Coulthard, E. J., Nachev, P., & Husain, M. (2008). Control over conflict during
movement preparation: Role of posterior parietal cortex. Neuron, 58(1), 144–157 .
Cowan, N. (2001). The magical number 4 in short-term memory: A reconsideration
of mental storage capacity. Discussion 114–185. The Behavioral and Brain
Sciences, 24(1), 87–114.
Cowan, N., Izawa, C., & Ohta, N. (2005). Working-memory capacity limits in
a theoretical context. Human learning and memory: Advances in theory and
application. The 4th Tsukuba International Conference on Memory. Mahwah:
Lawrence Erlbaum Associates, Publishers, p. 155 .
Cowey, A., & Stoerig, P. (1995). Blindsight in monkeys. Nature, 373(6511), 247–249.
Cowey, A., & Walsh, V. (2000). Magnetically induced phosphenes in sighted, blind
and blindsighted observers. Neuroreport, 11(14), 3269–3273.
Cowey, A., & Walsh, V. (2001). Tickling the brain: Studying visual sensation,
perception and cognition by transcranial magnetic stimulation. Progress in Brain
Research, 134, 411–425 .
Craig, A. D . (2005). Forebrain emotional asymmetry: A neuro-anatomical basis?
Trends in Cognitive Sciences, 9(12), 566–571.
Crick, F., & Koch, C. (1995). Are we aware of neural activity in primary visual
cortex? Nature, 375(6527), 121–123.
Crick, F., & Koch, C. (2003). A framework for consciousness. Nature Neuroscience,
6(2), 119–126.
Crone, N. E ., Sinai, A., & Korzeniewska, A. (2006). High-frequency gamma
oscillations and human brain mapping with electrocorticography in Neuper, C., &
Klimesch, W. (Eds.), Progress in Brain Research, 159, 276–295.
Csibra, G., Tucker, L. A., & Johnson, M. H . (1998). Neural correlates of saccade
planning in infants: A high-density ERP study. International Journal of
Psychophysiology, 29(2), 201–215 .
Csibra, G., Tucker, L. A., & Johnson, M. H. (2001). Differential frontal cortex
activation before anticipatory and reactive saccades in infants. Infancy, 2(2), 159–
174.
Cumming, B. G. (2002). An unexpected specialization for horizontal disparity in
primate primary visual cortex. Nature, 418(6898), 633–636.
Curran, T. (2001). Implicit learning revealed by the method of opposition. Trends in
Cognitive Sciences, 5(12), 503–504.
Curtis, С. E., & D’Esposito, M. (2003). Persistent activity in the prefrontal cortex
during working memory. Trends in Cognitive Sciences, 7(9), 415–423.
Cusack, R. (2005). The intraparietal sulcus and perceptual organization. Journal of
Cognitive Neuroscience, 17(4), 641–651.
D’Esposito, M., & Chen, A. J. (2006). Neural mechanisms of prefrontal cortical
function: Implications for cognitive rehabilitation. Progress in Brain Research,
157, 123–139.
D’Esposito, M., & Postle, B. R. (1999). The dependence of span and delayed-response
performance on prefrontal cortex. Neuropsychologia, 37(11), 1303–1315.
D’Haenen, H., Boer, J. A ., & Willner, P. (2002). Biological psychiatry. John Wiley
& Sons.
Dalrymple-Alford, E. C, & Budayr, B. (1966). Examination of some aspects of the
Stroop color-word test. Perceptual and Motor Skills, 23,1211–1214 .
Литература
391
Damasio, A. R. (1989). Time-locked multiregional retroactivation: A systems level
proposal for the neural substrates of recall and recognition. Cognition, 33, 25–32.
Damasio, A. R., Grabowski, T. J., Bechara, A., et al. (2000). Subcortical and cortical
brain activity during the feeling of self-generated emotions. Nature Neuroscience,
3(10), 1049–1056.
Damasio, H., Grabowski, Т., Frank, R., Galaburda, A. M ., & Damasio, A. R. (1994).
The return of Phineas Gage: Clues about the brain from the skull of a famous
patient. Science, 264(5162), 1102–1105.
Dang-Vu, Т. Т., Desseilles, M., Laureys, S., et al. (2005). Cerebral correlates of delta
waves during non-REM sleep revisited. Neuroimage, 28(1), 14–21 .
Dapretto, M., Davies, M. S., Pfeifer, J. H., et al. (2006). Understanding emotions in
others: Mirror neuron dysfunction in children with autism spectrum disorders.
Nature Neuroscience, 9(1), 28–30 .
Darwin, C. (1872). The expression of the emotions in man and animals. London: John
Murray.
Daselaar, S. M ., Fleck, M. S ., Prince, S. E ., & Cabeza, R. (2006). The medial temporal
lobe distinguishes old from new independently of consciousness. The Journal of
Neuroscience, 26(21), 5835–5839.
Davatzikos, C., Ruparel, K., Fan, Y., et al. (2005). Classifying spatial patterns of
brain activity with machine learning methods: Application to lie detection.
Neuroimage, 28(3), 663–668 .
David, S. P., Munafo, M. R., Johansen-Berg, H., et al. (2005). Ventral striatum/
nucleus accumbens activation to smoking-related pictorial cues in smokers
and nonsmokers: A functional magnetic resonance imaging study. Biological
Psychiatry, 58(6), 488–494.
Davidson, R. J., & Irwin, W. (1999). The functional neuroanatomy of emotion and
affective style. Trends in Cognitive Sciences, 3(1), 11–21 .
de Gelder, В., Morris, J. S., & Dolan, R. J. (2005). Unconscious fear influences
emotional awareness of faces and voices. Proceedings of the National Academy
of Sciences of the United States of America, 102(51), 18682–18687.
de Gelder, В., Vroomen, J., Pourtois, G., & Weiskrantz, L. (2000). Affective blindsight:
Are we blindly led by emotions? Response to Heywood and Kentridge. Trends in
Cognitive Sciences, 4(4), 126–127 .
De Groot, A. D. (1946). Het denken van den schaker. Amsterdam: Noord Hollandsche.
de Haan, M., Pascalis, O., & Johnson, M. H . (2002). Specialization of neural
mechanisms underlying face recognition in human infants. Journal of Cognitive
Neuroscience, 14(2), 199–209.
De Stefano, N., Bartolozzi, M. L ., Guidi, L., Stromillo, M. L ., & Federico, A. (2005).
Magnetic resonance spectroscopy as a measure of brain damage in multiple
sclerosis. Journal of the Neurological Sciences, 233(1–2), 203–208.
De Waal, F. B. (2004). Peace lessons from an unlikely source. PLoS Biology, 2(4),
E106.
DeCasper, A. J., & Fifer, W. P. (1980). Of human bonding: new-borns prefer their
mothers’ voices. Science, 208(4448), 1174–1176.
Decety, J., Chaminade, Т., Gre’zes, J., & Meltzoff, A. (2002). A PET exploration of
the neural mechanisms involved in reciprocal imitation. Neuroimage, 15, 265–272.
392
Литература
DeFelipe, J. (2002). Sesquicentenary of the birthday of Santiago Ramon у Cajal, the
father of modern neuroscience. Trends in Neurosciences, 25(9), 481–484.
Degonda, N., Mondadori, C. R., Bosshardt, S., et al. (2005). Implici associative
learning engages the hippocampus and interacts with explicit associative learning.
Neuron, 46(3), 505–520.
Dehaene, S. (2001). Calculating creative connections. Trends in Cognitive Sciences,
5(8), 364.
Dehaene, S., Molko, N., Cohen, L., & Wilson, A. J. (2004). Arithmetic and the brain.
Current Opinion in Neurobiology, 14(2), 218–224 .
Dehaene, S., & Naccache, L. (2001). Towards a cognitive permanence of
consciousness: Basic evidence and a workspace framework. Cognition, 79,1–37.
Dehaene, S., Naccache, L., Cohen, L., et al. (2001). Cerebral mechanisms of word
masking and unconscious repetition priming. Nature Neuroscience, 4(7), 752–
758.
Dehaene-Lambertz, G., & Dehaene, S. (1994). Speed and cerebral correlates of
syllable discrimination in infants. Nature, 370(6487), 292–295.
Dehaene-Lambertz, G., Dehaene, S., & Hertz-Pannier, L. (2002). Functional
neuroimaging of speech perception in infants. Science, 298(5600), 2013–2015.
Dehaene-Lambertz, G., Hertz-Pannier, L., & Dubois, J. (2006). Nature and nurture in
language acquisition: Anatomical and functional brain-imaging studies in infants.
Trends in Neurosciences, 29(7), 367–373.
Delaney, P. F., Ericsson, K. A ., & Knowles, M. E . (2004). Immediate and sustained
effects of planning in a problem-solving task. Journal of Experimental
Psychology. Learning, Memory, and Cognition, 30(6), 1219–1234.
Del-Ben, С. M ., Deakin, J. F., McKie, S., et al. (2005). The effect of citalopram
pretreatment on neuronal responses to neuropsychological tasks in normal
volunteers: an fMRI study. Neuropsychopharmacology, 30(9), 1724–1734.
Dell, G. S., & Sullivan, J. M. (2004). Speech errors and language production:
Neuropsychological and connectionist perspectives. In В. H . Ross (Ed.), The
psychology of learning and motivation (pp. 63 –108). San Diego: Elsevier.
Dennett, D. (2001). Are we explaining consciousness yet? Cognition, 79(1–2), 221–
237.
Desimone, R., Albright, T. D ., Gross, C. G., & Bruce, C. (1984). Stimulus-
selective properties of inferior temporal neurons in the macaque. The Journal of
Neuroscience: The Official Journal of the Society for Neuroscience, 4(8), 2051–
2062.
Deutsch, G., & Eisenberg, H. M. (1987). Frontal blood flow changes in recovery
from coma. Journal of Cerebral Blood Flow and Metabolism, 7(1), 29–34.
Diamond, A. (1985). Development of the ability to use recall to guide action, as
indicated by infants’ performance on AB. Child Development, 56(4), 868–883 .
Diamond, A. (1991). Neuropsychological insights into the meaning of object concept
development. In S. G. R. Carey (Ed.), The epigenesis of mind: Essays on biology
and cognition (pp. 67–110). Hillsdale: Lawrence Erlbaum Associates.
Diamond, A. (2001). A model system for studying the role of dopamine in the
prefrontal cortex during early development in humans: Early and continuously
treated phenylketonuria. In C. A. L. Nelson (Ed.), Handbook of developmental
cognitive neuroscience (pp. 433–472). Cambridge: MIT Press.
Литература
393
Diamond, A., & Goldman-Rakic, P. S . (1989). Comparison of human infants and
rhesus monkeys on Piaget’s AB task: Evidence for dependence on dorsolateral
prefrontal cortex. Experimental Brain Research, 74(1), 24–40.
Dietl, Т., Trautner, P., Staedtgen, M., et al. (2005). Processing of famous faces
and medial temporal lobe event-related potentials: A depth electrode study.
Neuroimage, 25(2), 401–407.
Dietrich, V., Nieschalk, M., Stoll, W., Rajan, R., & Pantev, C. (2001). Cortical
reorganization in patients with high frequency cochlear hearing loss. Hearing
Research, 158(1–2), 95–101.
Dixon, N. F. (1971). Subliminal perception: The nature of a controversy. McGraw-Hill.
Doesburg, S. M ., Roggeveen, А. В ., Kitajo, K., & Ward, L. M . (2007). Large-Scale
Gamma-Band Phase Synchronization and Selective Attention Cerebral Cortex
doi:10.1093/ cercor/bhm073.
Dogil, G., Ackermann, W., Grodd, W., et al. (2002). The speaking brain: A tutorial
introduction to fMRI experiments in the production of speech, prosody, and
syntax. Journal of Neurolinguistics, 15(1), 59–90.
Dolan, R. J., Fink, G. R., Rolls, E., et al. (1997). How the brain learns to see objects
and faces in an impoverished context. Nature, 389(6651), 596–599.
Doniger, G. M., Foxe, J. J., Schroeder, С. E., Murray, M. M., Higgins, B. A., &
Javitt, D. C. (2001). Visual perceptual learning in human object recognition areas:
A repetition priming study using high-density electrical mapping. Neuroimage,
13(2), 305–313.
Downing, P. E ., Bray, D., Rogers, J., & Childs, C. (2004). Bodies capture attention
when nothing is expected. Cognition, 93(1), B27–B38.
Draine, S. C., & Greenwald, A. G . (1998). Replicable unconscious semantic priming.
Journal of Experimental Psychology. General, 127(3), 286–303.
Drake, R., Vogl, W., & Mitchell, A. (Eds.), (2005). Gray’s anatomy for students.
Edinburgh: Churchill Livingston.
Driver, J., & Mattingley, J. B. (1998). Parietal neglect and visual awareness. Nature
Neuroscience, 1(1), 17–22 .
Driver, J., & Vuilleumier, P. (2001). Perceptual awareness and its loss in unilateral
neglect and extinction. Cognition, 79(1–2), 39–88 .
Dronkers, N. F., & Ogar, J. (2003). Aphasia. In M. Aminoff & R. Daroff (Eds.),
Encyclopedia of the neurological sciences. San Diego: Academic Press.
Dubois, J., et al. (2008). Asynchrony of the early maturation of white matter bundles
in healthy infants: Quantitative landmarks revealed noninvasively by diffusion
tensor imaging. Human Brain Mapping, 29, 14–27 .
Dudai, Y. (2004). The neurobiology of consolidations, or, how stable is the engram?
Annual Review of Psychology, 55, 51–86.
Dudley, H. (1939). The automatic synthesis of speech. Proceedings of the National
Academy of Sciences of the United States of America, 25(7), 377–383 .
Duncan, J., & Owen, A. M . (2000). Common regions of the human frontal lobe
recruited by diverse cognitive demands. Trends Neuroscience, 23(10), 475–483.
Duncker, K. (1945). On problem-solving. Psychological Monographs, 58(5).
Durston, S., Thomas, К. M., Worden, M. S., Yang, Y., & Casey, B. J. (2002). The effect
of preceding context on inhibition: An event-related fMRI study. Neuroimage,
16(2), 449–453.
394
Литература
Duzel, E., Yonelinas, A. P., Mangun, G. R., Heinze, H. J., & Tulving, E. (1997).
Event-related brain potential correlates of two states of conscious awareness in
memory. Proceedings of the National Academy of Sciences of the United States of
America, 94(11), 5973–5978.
Eagleman, D. M . (2001). Visual illusions and neurobiology. Nature Reviews
Neuroscience, 2(12), 920–926.
Edelman, G. M . (1989). The remembered present: A biological theory of
consciousness. New York: Basic Books Inc.
Edelman, G. M, & Gaily, J. A . (1992). Nitric oxide: Linking space and time in the
brain. PNAS, 89(24), 11651–11652.
Edelman, G. M ., & Mountcastle, V. B . (1978). The mindful brain: Cortical
organization and the group-selective theory of higher brain function. Oxford:
Massachusetts Institute of Technology Press.
Edelman, G. M ., & Tononi, G. (2000). A universe of consciousness: How matter
becomes imagination. New York: Basic Books Inc.
Eguchi, M., & Yamaguchi, S. (2009). In vivo and in vitro visualization of gene
expression dynamics over extensive areas of the brain. Neuroimage, 44, 1274–
1283.
Eichenbaum, H. B . (2003). Learning and memory: Brain systems. In L. R. Squire,
J. L . Roberts, N. C. Spitzer, M. J . Zigmond, S. K. McConnell, & F. E. Bloom
(Eds.), Fundamental neuroscience (2nd ed.) (pp. 1297–1328). Amsterdam:
Academic Press.
Eisenberger, N. I ., & Lieberman, M. D. (2004). Why rejection hurts: A common
neural alarm system for physical and social pain. Trends in Cognitive Sciences,
8(7), 294–300 .
Ekman, P. (2003). Emotions inside out. 130 years after Darwin’s ‘the expression of
the emotions in man and animal’. Annals of the New York Academy of Sciences,
1000, 1–6.
Eldridge, L. L ., Sarfatti, S., & Knowlton, B. J. (2002). The effect of testing procedure
on remember-know judgments. Psychonomic Bulletin & Review, 9(1), 139–145 .
Emery, N. J. (2000). The eyes have it: The neuroethology, function and evolution of
social gaze. Neuroscience and Biobehavioral Reviews, 24(6), 581–604.
Engel, A. K ., & Singer, W. (2001). Temporal binding and the neural correlates of
sensory awareness. Trends in Cognitive Sciences, 5(1), 16–25 .
Engel, A. K, Fries, P., Koenig, P., & Singer, W. (2001). Dynamic predictions:
Oscillations and synchrony in top- down processing. Nature Reviews Neuroscience,
2, 704–716.
Epstein, R., & Kanwisher, N. (1998). A cortical representation of the local visual
environment. Nature, 392(6676), 598–601.
Falini, A., Bozzali, M., Magnani, G., et al. (2005). A whole brain MR spectroscopy
study from patients with Alzheimer’s disease and mild cognitive impairment.
Neuroimage, 26(4), 1159–1163.
Fan, J., McCandliss, B. D ., Fossella, J., Flombaum, J. I ., & Posner, M. I. (2005). The
activation of attentional networks. Neuroimage, 26(2), 471–179.
Fan, J., McCandliss, B. D ., Sommer, Т., Raz, A., & Posner, M. I. (2002). Testing
the efficiency and independence of attentional networks. Journal of Cognitive
Neuroscience, 14(3), 340–347.
Литература
395
Fantz, R. L . (1964). Visual experience in infants: Decreased attention to familiar
patterns relative to novel ones. Science, 146, 668–670.
Farah, M. J., Tanaka, J. W., & Drain, H. M . (1995). What causes the face inversion
effect? Journal of Experimental Psychology. Human Perception and Performance,
21(3), 628–634.
Farroni, Т., Johnson, M. H., Brockbank, M., & Simion, F. (2000). Infants’ use of gaze
direction to cue attention: The importance of perceived motion. Visual Cognition,
7(6), 705–718.
Fatterpekar, G. M ., Naidich, T. P., Delman, B. N ., et al. (2002). Cytoarchitecture of
the human cerebral cortex: MR microscopy of excised specimens at 9.4 tesla.
American Journal of Neuroradiology, 23(8), 1313–1321.
Faubert, J., Diaconu, V., Pitto, M., & Pitto, A. (2002). Residual vision in the blind
field of hemidecorticated humans predicted by a diffusion scatter model and
selective spectral absorption of the human eye. Vision Research, 39(10), 149–157 .
Feldman, J. (2003). What is a visual object? Trends in Cognitive Sciences, 7(6), 252–
256.
Felleman, D. J., & Van Essen, D. C. (1991). Distributed hierarchical processing in the
primate cerebral cortex. Cerebral Cortex, 2(1), 1–17 .
Filley, С. M . (2002). Neuroanatomy. In V. S . Ramachandran (Ed.), Encyclopedia of
the human brain. San Diego: Academic Press.
Finkelstein, G. (2000). Why Darwin was English. Endeavour, 24(2), 76–78.
Fishman, Y. I., Reser, D. H., Arezzo, J. C ., & Steinschneider, M. (2001). Neural
correlates of auditory stream segregation in primary auditory cortex of the awake
monkey. Hearing Research, 152(1–2), 167–187.
Fletcher, P. C., Frith, C. D ., Grasby, P. M., Shallice, Т., Frackowiak, R. S ., & Dolan,
R. J. (1995). Brain systems for encoding and retrieval of auditory-verbal memory.
An in vivo study in humans. Brain, 128(Pt 2), 401–416.
Fletcher, P. C., & Henson, R. N . (2001). Frontal lobes and human memory: Insights
from functional neuroimaging. Brain, 224(Pt 5), 849–881.
Fletcher, P. C., Zafiris, O., Frith, C. D., et al. (2005). On the benefits of not trying:
Brain activity and connectivity reflecting the interactions of explicit and implicit
sequence learning. Cerebral Cortex, 15(7), 1002–1015.
Fogassi, L., Ferrari, P. F., Gesierich, В., Rozzi, S., Chersi, F., & Rizzolatti, G. (2005).
Parietal lobe: From action organization to intention understanding. Science,
308(5722), 662–667.
Fourier, J. (1822). The analytical theory of heat (English transl Freeman, 1878).
Republished 1955. New York: Dover.
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen, D. C., & Raichle, M.
E. (2005). The human brain is intrinsically organized into dynamic, anticorrelated
functional networks. Proceedings of the National Academy of Sciences of the
United States of America, 102(27), 9673–9678.
Fox, N. A ., & Bell, M. A. (1990). Electrophysiological indices of frontal lobe
development. In A. Diamond (Ed.), The development and neural bases of higher
cognitive functions: Vol. 608 (pp. 677–698). New York: New York Academy of
Sciences.
Fox, P. Т., & Raichle, M. E . (1986). Focal physiological uncoupling of cerebral
blood flow and oxidative metabolism during somatosensory stimulation in human
396
Литература
subjects. Proceedings of the National Academy of Sciences of the United States of
America, 83(4), 1140–1144 .
Fox, P. Т., Raichle, M. E., Mintun, M. A ., & Dence, C. (1988). Nonoxidative glucose
consumption during focal physiologic neural activity. Science, 242(4864), 462–
164.
Frackowiak, R. S . J. (Ed.). (2004). Human brain function (2nd ed.): Vol. xvi. Boston:
Elsevier Academic Press.
Fraisse, P. (1969). Why is naming longer than reading? Acta Psychologica, 30, 96–
103.
Franzen, G., & Ingvar, D. H. (1975). Abnormal distribution of cerebral activity in
chronic schizophrenia. Journal of Psychiatric Research, 22(3), 199–214 .
Freeman, W. J . (1991). The physiology of perception. Scientific American, 284, 78–85.
Freeman, W. J. (2004). Origin, structure, and role of background EEG activity. Part 1
Analytic amplitude. Clinical Neurophysiology, 115(9), 2077–2088.
Freeman, W. J., Burke, В. C ., & Holmes, M. D. (2003). Aperiodic phase resetting
in scalp EEG of beta-gamma oscillations by state transitions at alpha-theta rates.
Human Brain Mapping, 19(4), 248–272 .
Freeman, W„ Holmes, M., West, G., & Vanhatalo, S. (2006). Fine spatiotemporal
structure of phase in human intracranial EEG. Clinical Neurophysiology, 117(6),
1228–1243.
Fried, P. A ., Watkinson, В., & Gray, R. (2003). Differential effects on cognitive
functioning in 13- to 16-year- olds prenatally exposed to cigarettes and marihuana.
Neurotoxicology and Teratology, 25(4), 427–436.
Friederici, A. D . (2002). Towards a neural basis of auditory sentence processing.
Trends in Cognitive Sciences, 6(2), 78–84.
Friederici, A. D . (2005). Neurophysiological markers of early language acquisition:
From syllables to sentences. Trends in Cognitive Sciences, 9(10), 481–488.
Friederici, A. D., & Kotz, S. A. (2003). The brain basis of syntactic processes:
Functional imaging and lesion studies. Neuroimage, 20(Suppl 1), S8–S17.
Fries, P. (2005). A mechanism for cognitive dynamics: Neuronal communication
through neuronal coherence. Trends in Cognitive Sciences, 9(10), 474–480.
Fries, P., Fernandez, G., & Jensen, O. (2003). When neurons form memories. Trends
in Neurosciences, 26(3), 123–124 .
Fries, P., Schroder, J. H., Roelfsema, P. R., Singer, W., & Engel, A. K. (2002).
Oscillatory neuronal synchronization in primary visual cortex as a correlate of
stimulus selection. The Journal of Neuroscience, 22(9), 3739–3754.
Friston, K. (2003). Learning and inference in the brain. Neural Networks, 16(9),
1325–1352.
Frith, C. D ., & Frith, U. (1999). Interacting minds — a biological basis. Science,
286(5445), 1692–1695.
Fulford, J., Vadeyar, S. H ., Dodampahala, S. H ., et al. (2003). Fetal brain activity in
response to a visual stimulus. Human Brain Mapping, 20(4), 239–245 .
Fulton, J. F. (1928). Observations upon the vascularity of the human occipital lobe
during visual activity. Brain, 51, 310–320.
Funahashi, S., Bruce, C. J., & Goldman-Rakic, P. S. (1989). Mnemonic coding
of visual space in the monkey’s dorsolateral prefrontal cortex. Journal of
Neurophysiology, 61(2), 331–349.
Литература
397
Funahashi, S., Bruce, C. J., & Goldman-Rakic, P. S. (1993). Dorsolateral prefrontal
lesions and oculomotor delayed- response performance: Evidence for mnemonic
‘scotomas’. The Journal of Neuroscience, 13(4), 1479–1497.
Funnel, E., & Sheridan, J. S. (1992). Categories of knowledge? Unfamiliar aspects of
living and nonliving things. Cognitive Neuropsychology, 9, 135–153.
Fuster, J. M . (1985). Temporal organization of behavior. Human Neurobiology, 4(2),
57–60 .
Fuster, J. M . (1997). Network memory. Trends in Neurosciences, 20(10), 451–459.
Fuster, J. M . (2003). Cortex and mind: Unifying cognition. New York: Oxford
University Press.
Fuster, J. M . (2004). Upper processing stages of the perceptionaction cycle. Trends in
Cognitive Sciences, 8(4), 143–145 .
Fuster, J. M ., & Alexander, G. E . (1971). Neuron activity related to short-term
memory. Science, 173(997), 652–6654.
Gabrieli, J. D., Keane, M. M., Stanger, B. Z., Kjelgaard, M. M., Corkin, S., &
Growdon, J. H. (1994). Dissociations among structural-perceptual, lexical-
semantic, and event-fact memory systems in Alzheimer, amnesic, and normal
subjects. Cortex, 30(1), 75–103.
Gage, N. M ., & Roberts, T. P. (2000). Temporal integration: Reflections in the ml00
of the auditory evoked field. Neuroreport, 11(12), 2723–2726.
Gage, N. M ., Roberts, T. P., & Hickok, G. (2002). Hemispheric asymmetries in
auditory evoked neuromagnetic fields in response to place of articulation contrasts.
Brain Research. Cognitive Brain Research, 14(2), 303–306 .
Gage, N. M ., Siegel, В., Callen, M., & Roberts, T. P. (2003a). Cortical sound
processing in children with autism disorder: An MEG investigation. Neuroreport,
14(16), 2047–2051.
Gage, N. M ., Siegel, В., & Roberts, T. P. (2003b). Cortical auditory system maturational
abnormalities in children with autism disorder: An MEG investigation. Brain
Research. Developmental Brain Research, 144(2), 201–209.
Gage, N., Poeppel, D., Roberts, T. P., & Hickok, G. (1998). Auditory evoked ml00
reflects onset acoustics of speech sounds. Brain Research, 814(1–2), 236–239.
Gage, N., Roberts, T. P., & Hickok, G. (2006). Temporal resolution properties of
human auditory cortex: Reflections in the neuromagnetic auditory evoked ml00
component. Brain Research, 1069(1), 166–171 .
Galaburda, A. M ., & Pandya, D. N . (1983). The intrinsic architectonic and connectional
organization of the superior temporal region of the rhesus monkey. The Journal of
Comparative Neurology, 221(2), 169–184.
Gallagher, H., Jack, A., Roepstorff, A., & Frith, C. (2002). Imaging the intentional
stance in a competitive game. Neuroimage, 16, 814–821.
Gallese, V., Fadiga, L., Fogassi, L., & Rizzolatti, G. (1996). Action recognition in the
premotor cortex. Brain, 119(Pt 2), 593–609 .
Ganis, G., Thompson, W. L ., & Kosslyn, S. M . (2004). Brain areas underlying visual
mental imagery and visual perception: An fMRI study. Brain Research. Cognitive
Brain Research, 20(2), 226–241 .
Gastaut, D. (1958). The role of the reticular system in establishing conditioned
reactions. In H. M . Jasper, L. D. Proctor, R. S. Knighton, W. C. Noshay, & R. T.
Costello (Eds.), The reticular formation of the brain. Boston: Little, Brown.
398
Литература
Gathers, A. D., Bhatt, R., Corbly, C. R., Farley, А. В., & Joseph, J. E. (2004).
Developmental shifts in cortical loci for face and object recognition. Neuroreport,
25(10), 1549–1553.
Gauthier, I., Skudlarski, P., Gore, J. C., & Anderson, A. W. (2000). Expertise for cars
and birds recruits brain areas involved in face recognition. Nature Neuroscience,
3(2), 191–197.
Geschwind, N. (1979a). Anatomical and functional specialization of the cerebral
hemispheres in the human. Bulletin Et Memoires De L’Academie Royale De
Medecine De Belgique, 134(6), 286–297.
Geschwind, N. (1979b). Specializations of the human brain. Scientific American,
241(3), 180–199.
Geschwind, N., & Galaburda, A. M. (1985a). Cerebral lateralization. Biological
mechanisms, associations, and pathology. I. A hypothesis and a program for
research. Archives of Neurology, 42(6), 428–459.
Geschwind, N., & Galaburda, A. M . (1985b). Cerebral lateralization. Biological
mechanisms, associations, and pathology. II . A hypothesis and a program for
research. Archives of Neurology, 42(6), 521–552 .
Geschwind, N., & Galaburda, A. M. (1985c). Cerebral lateralization. Biological
mechanisms, associations, and pathology. III . A hypothesis and a program for
research. Archives of Neurology, 42(6), 634–654.
Geschwind, N., & Levitsky, W. (1968). Human brain: Left-right asymmetries in
temporal speech region. Science, 262(837), 186–187.
Gevins, A., Leong, H., Smith, M. E ., Le, J., & Du, R. (1995). Mapping cognitive
brain function with modern high-resolution electroencephalography. Trends in
Neurosciences, 18(10), 429–436.
Gibbs, S. E ., & D’Esposito, M. (2006). A functional magnetic resonance imaging
study of the effects of pergolide, a dopamine receptor agonist, on component
processes of working memory. Neuroscience, 239(1), 359–371.
Giedd, J. N ., Blumenthal, J., Jeffries, N. O ., et al. (1999). Brain development during
childhood and adolescence: A longitudinal MRI study. Nature Neuroscience,
2(10), 861–863.
Gilboa, A., Winocur, G., Grady, C. L ., Hevenor, S. J., & Mscovitch, M. (2004).
Remembering our past: Functional neuroanatomy of recent and very remote
personal events. Cerebral Cortex, 14(11), 1214–1225.
Gilmore, R. O ., & Johnson, M. H . (1995). Working memory in infancy: Six-month-
olds’ performance on two versions of the oculomotor delayed response task.
Journal of Experimental Child Psychology, 59(3), 397–418.
Giocomo, L. M ., & Hasselmo, M. E . (2007). Neuromodulation by glutamate and
acetylcholine can change circuit dynamics by regulating the relative influence of
afferent input and excitatory feedback. Molecular Neurobiology, 36, 184–200.
Girard, P., Salin, P. A., & Bullier, J. (1991). Visual activity in areas V3a and V3
during reversible inactivation of area VI in the macaque monkey. Journal of
Neurophysiology, 66(5), 1493–1503.
Gitelman, D. R., Nobre, A. C., Sonty, S., Parrish, Т. В., & Mesulam, M. M . (2005).
Language network specializations: An analysis with parallel task designs and
functional magnetic resonance imaging. Neuroimage, 26(4), 975–985.
Литература
399
Glisky, E. L ., Polster, M. R., & Routhieaux, В. C. (1995). Double dissociation
between item and source memory. Neuropsychology, 9(2), 229–235.
Gluck, M. A., Meeter, M., & Myers, С. E. (2003). Computational models of the
hippocampal region: Linking incremental learning and episodic memory. Trends
in Cognitive Sciences, 7(6), 269–276.
Gobet, F. (1998). Expert memory: A comparison of four theories. Cognition, 66,
15–152 .
Gobet, F., & Simon, H. A. (1996). Recall of rapidly presented random chess positions.
Psychonomic Bulletin & Review, 3, 159–163.
Goebel, R., Muckli, L., Zanella, F. E ., Singer, W., & Stoerig, P. (2001). Sustained
extrastriate cortical activation without visual awareness revealed by fMRI studies
of hemianopic patients. Vision Research, 41(10–11), 1459–1474 .
Gold, J. M ., Berman, K. F., Randolph, C., Goldberg, T. E ., & Weinberger, D. R.
(1996). PET validation of a novel prefrontal task: Delayed response alternation.
Neuropsychology, 20(1), 3–10.
Goldberg, E. (1992). Introduction: The frontal lobes in neurological and psychiatric
conditions. Neuropsychol Neuropsychiatr and Behavioral Neurology, 5(4), 231–
232.
Goldberg, E. (2001a). The executive brain. New York: Oxford University Press.
Goldberg, E. (2001b). The executive brain: Frontfl lobes and the civilized mind (Vol.
xix). New York: Oxford University Press.
Goldberg, E., Bilder, R. M ., Hughes, J. E., Antin, S. P., & Mattis, S. (1989). A reticulo-
frontal disconnection syndrome. Cortex, 25(4), 687–695.
Goldberg, E., Bilder, R. M ., Hughes, J. E., Antin, S. P., & Mattis, S. (1989). A reticulo-
frintal disconnection syndrome. Cortex, 25(4), 687–695.
Goldberg, E., & Bougakov, D. (2000). Novel approaches to the diagnosis and treatment
of frontal lobe dysfunction. International Handbook Of Neuropsychological
Rehabilitation. Critical Issues in Neuropsychology, 93–112 .
Goldberg, E., & Costa, L. D . (1981). Hemisphere differences in the acquisition and
use of descriptive systems. Brain and Language, 24(1), 144–173.
Goldberg, E., & Costa, L. D. (1985). Qualitative indices in neuropsychological
assessment: An extension of Luria’s approach to executive deficit following
prefrontal lesion. In I. Grant & К. M . Adams (Eds.), Neuropsychological assess-
ment of neuropsychiatric disorders (pp. 48–64). New York: Oxford University
Press.
Goldberg, E., Harner, R., Lovell, M., Podell, K., & Riggio, S. (1994). Cognitive
bias, functional cortical geometry, and the frontal lobes: Laterality, sex, and
handedness. Journal of Cognitive Neuroscience, 6(3), 276–293.
Goldberg, E., Kluger, A., Griesing, Т., Malta, L., Shapiro, M., Ferris, S. (1997,
August 17–22). Early diagnosis offrontal-lobe dementias. Jerusalem, Israel: Paper
presented at the Eighth Congress of International Psychogeriatric Association.
Goldman-Rakic, P. S. (1987). Circuitry of primate prefrontal cortex and regulation
of behavior by representational memory. Handbook Physiologica, 5, 373–417 .
Goldman-Rakic, P. S. (1995). Cellular basis of working memory. Neuron, 24(3),
477–485.
Goldman-Rakic, P. S. (1998). The prefrontal landscape: Implications of functional
architecture for understanding human mentation and the central executive.
400
Литература
In A. C. Roberts, T. W. Robbins, & L. Weiskrantz (Eds.), The prefrontal cortex:
Executive and cognitive functions (pp. 87–102). Oxford: Oxford University Press.
Goodale, M. A ., & Humphrey, G. K . (1998). The objects of action and perception.
Cognition, 67(1–2), 181–207.
Goodale, M. A ., & Milner, A. D. (1992). Separate visual pathways for perception and
action. Trends in Neurosciences, 25(1), 20–25 .
Goodale, M. A., Milner, A. D ., Jakobson, L. S., & Carey, D. P. (1991). A neurological
dissociation between perceiving objects and grasping them. Nature, 349(6305),
154–156.
Gopalakrishnan, M., & Briggs, C. A . (2007). Ion channels: Ligand gated. In D. J.
Triggle, & J. B. Taylor (Eds.), Comprehensive Medicinal Chemistry II, Vol. 2,
877–918.
Gore, A. C., Roberts, J. L., Squire, L. R., Bloom, F. E., McConnell, S. K., Roberts,
J. L ., Spitzer, N. C ., & Zigmond, M. J. (Eds.) . (2003). Fundamental neuroscience
(2nd ed.) . San Diego: Elsevier Academic Press.
Goren, C. C ., Sarty, M., & Wu, P. Y. (1975). Visual following and pattern discrimination
of face-like stimuli by newborn infants. Pediatrics, 56(4), 544–549.
Gottesmanri, С. (1999). Neurophysiological support of consciousness during waking
and sleep. Progress in Neurobiology, 59(5), 469–508.
Gottlieb, G., & Halpern, С. T. (2002). A relational view of causality in normal and
abnormal development. Development and Psychopathology, 14(3), 421–435.
Graham, K. S., Murre, J. M ., & Hodges, J. R . (1999). Episodic memory in semantic
dementia: A computational approach based on tracelink. Progress in Brain
Research, 121, 47–65.
Graham, N. L., Patterson, K., & Hodges, J. R. (2000). The impact of semantic memory
impairment on spelling: Evidence from semantic dementia. Neuropsychologia,
38(2), 143–163.
Grant, D. A ., & Berg, E. A. (1948). A behavioral analysis of degree of reinforcement
and ease of shifting to new responses in a Weigl-type card-sorting problem.
Journal of Experimental Psychology, 38, 404–411 .
Grant, D. A., & Berg, E. A . (1993). Wisconsin card sorting test. Lutz, FL: Psychological
Assessment Resources.
Greenwald, A. G., Banaji, M. R., Rudman, L. A., Farnham, S. D., Nosek, B. A.,
& Mellott, D. S . (2002). A unified theory of implicit attitudes, stereotypes, self-
esteem, and self- concept. Psychological Review, 109(1), 3–25 .
Griffiths, T. D„ & Warren, J. D . (2002). The plenum temporale as a computational
hub. Trends in Neurosciences, 25(7), 348–353.
Grill-Spector, K., Kushnir, Т., Edelman, S., Avidan, G., Itzchak, Y., & Malach, R.
(1999). Differential processing of objects under various viewing conditions in the
human lateral occipital complex. Neuron, 24(1), 187–203.
Grill-Spector, K., Kushnir, Т., Hendler, Т., & Malach, R. (2000). The dynamics of
object-selective activation correlate with recognition performance in humans.
Nature Neuroscience, 3(8), 837–843.
Grodzinsky, Y., & Friedrici, A. D . (2006). Neuroimaging of syntax and syntactic
processing. Current Opinion in Neurobiology, 16(2), 240–246.
Литература
401
Groome, L. J., Mooney, D. M., Holland, S. В., Smith, L. A., Atterbury, J. L., &
Dykman, R. A . (1999). Behavioral state affects heart rate response to low-intensity
sound in human fetuses. Early Hum Dev, 54(1), 39–54 .
Gross, C. G . (1992). Representation of visual stimuli in inferior temporal cortex.
Philosophical Transactions of the Royal Society of London. Series B, Biological
Sciences, 335(1273), 3–10.
Gu, Q. (2002). Neuromodulatory transmitter systems in the cortex and their role in
cortical plasticity. Neuroscience, 111(4), 815–835.
Habib, R., Nyberg, L., & Tulving, E. (2003). Hemispheric asymmetries of memory:
The hera model revisited. Trends in Cognitive Sciences, 7(6), 241–245 .
Hagoort, P. (2005). On broca, brain, and binding: A new framework. Trends in
Cognitive Sciences, 9(9), 416–423.
Hahn, S., Carlson, C., Singer, S., & Gronlund, S. D. (2006). Aging and visual search:
Automatic and controlled attentional bias to threat faces. Acta Psychologica
(Amst).
Haier, R. J., Siegel, В. V., Jr., MacLachlan, A., Soderling, E., Lottenberg, S., &
Buchsbaum, M. S. (1992). Regional glucose metabolic changes after learning a
complex visuospatial/motor task: A positron emission tomographic study. Brain
Research, 570(1–2), 134–143.
Handy, Т. C., Gazzaniga, M. S„ & Ivry, R. B. (2003). Cortical and subcortical
contributions to the representation of temporal information. Neuropsychologia,
41(11), 1461–1473.
Hanon, E. E., & Trehub, S. E. (2005). Tuning in to musical rhythms: Infants learn
more readily than adults. Proceedings of the National Academy of Sciences of the
United States of America, 102(35), 12639–12643.
Harenski, C. L ., & Hamann, S. (2006). Neural correlates of regulating negative
emotions related to moral violations. Neuroimage, 30(1), 313–324.
Hariri, A. R., Brown, S. M., Williamson, D. E., Flory, J. D., de Wit, H., & Manuck,
S. B. (2006). Preference for immediate over delayed rewards is associated with
magnitude of ventral striatal activity. The Journal of Neuroscience, 26(51),
13213–13217.
Hariri, A. R., Mattay, V. S., Tessitore, A., et al. (2002). Serotonin transporter genetic
variation and the response of the human amygdala. Science, 297(5580), 400–403.
Hartley, A. A., & Speer, N. K. (2000). Locating and fractionating working memory
using functional neuroimaging: Storage, maintenance, and executive functions.
Microscopy Research and Technique, 52(1), 45–53.
Hauk, O., Johnsrude, I., & Pulvermuller, F. (2004). Somatotopic representation of
action words in human motor and premotor cortex. Neuron, 41(2), 301–307.
Haxby, J. V., Gobbini, M. I., Furey, M. L., Ishai, A., Schouten, J. L., & Pietrini, P.
(2001). Distributed and overlapping representations of faces and objects in ventral
temporal cortex. Science, 293(5539), 2425–2430.
Haxby, J., Hoffman, E. A., & Gobbini, M. I . (2000). The distributed human neural
system for face perception. Trends in Cognitive Sciences, 4(6), 223–233.
Haxby, J., Hoffman, E., & Gobbini, M. (2002). Human neural systems for face
recognition and social communication. Biological Psychiatry, 51, 59–67.
Hayhoe, M., & Ballard, D. (2005). Eye movements in natural behavior. Trends in
Cognitive Sciences, 9(4), 188–194.
402
Литература
Haynes, J. D., Deichmann, R., & Rees, G. (2005). Eye-specific effects of binocular
rivalry in the human lateral geniculate nucleus. Nature, 438(7067), 496–499.
Hebb, D. O . (1949). The organization of behavior: A neuropsychological theory.
Oxford: Wiley.
Heckers, S., Weis, A., Deckersbach, Т., Goff, D., Mrecraft, R., & Bush, G. (2004).
Anterior cingulate cortex activation during cognitive interference in schizophrenia.
The American Journal of Psychiatry, 161, 707–715 .
Heimer, L„ & Van Hoesen, G. W. (2006). The limbic lobe and its output channels:
Implications for emotional functions and adaptive behavior. Neuroscience and
Biobehavioral Reviews, 30(2), 126–147 .
Hendry, S. H., Squire, L. R., Bloom, F. E., McConnell, S. K., Roberts, J. L., Spitzer,
N. C ., Zigmond, M. J. (Eds.), et al. (2003). Fundamental neuroscience (2nd ed.).
San Diego: Elsevier Academic Press.
Henke, K., Mondadori, C. R., Treyer, V., Nitsch, R. M., Buck, A., & Hock, C. (2003).
Nonconscious formation and reactivation of semantic associations by way of the
medial temporal lobe. Neuropsychologia, 41(8), 863–876.
Henson, R. N . (2001). Repetition effects for words and non-words as indexed by
event-related fMRI: A preliminary study. Scandinavian Journal of Psychology,
42(3), 179–186.
Hermann, L. (1870). Eine erscheinung simultanen contrastes. Pfliigers Archiv :
European Journal of Physiology, 3,13–15 .
Hesslow, G. (2002). Conscious thought as simulation of behaviour and perception.
Trends in Cognitive Sciences, 6(6), 242–247 .
Hester, R., Dixon, V., & Garavan, H. (2006). A consistent attentional bias for drug-
related material in active cocaine users across word and picture versions of the
emotional stroop task. Drug and Alcohol Dependence, 81(3), 251–257 .
Heywood, C. A., & Kentridge, R. W. (2000). Affective blindsight? Trends in Cognitive
Sciences, 4(4), 125–126.
Hickok, G., & Poeppel, D. (2004). Dorsal and ventral streams: A framework for
understanding aspects of the functional anatomy of language. Cognition, 92, 67–
99.
Hillyard, S. A., Hink, R. F., Schwent, V. L ., & Picton, T. W. (1973). Electrical signs of
selective attention in the human brain. Science, 182(108), 177–180.
Hobson, J. A ., & Stickgold, R. (1995). Sleep. Sleep the beloved teacher? Current
biology, 5(1), 35–36 .
Hodges, J. R., & Graham, K. S . (2001). Episodic memory: Insights from semantic
dementia. Philosophical Transactions of the Royal Society of London. Series B,
Biological Sciences, 356(1413), 1423–1434.
Hodges, J. R., & Patterson, K. (1997). Semantic memory disorders. Trends in
Cognitive Sciences, 1(2), 68–72.
Hodgkin, A. L., & Huxley, A. F. (1952). Currents carried by sodium and potassium
ions through the membrane of the giant axon of loligo. Journal of Physiology,
11 6 (4), 449–472 .
Hofer, S., & Frahm, J. (2006). Topography of the human corpus callosum revisited —
Comprehensive fiber tractography using magnetic resonance diffusion tensor
imaging. Neurolmage, 32, 989–994.
Литература
403
Hofer, S., Merboldt, K. D., Tammer, R., & Frahm, J. (2008). Rhesus monkey and
human share a similar topography of the corpus callosum as revealed by diffusion
tensor MRI in vivo. Cerebral Cortex, 18,1079–1084.
Holmes, G. (1918). Disturbances of vision by cerebral lesions. The British Journal of
Ophthalmology, 2, 353–384.
Holstege, G. J., Mouton, L., & Gerrits, N. M . (2004). Chapter 36. In G. Paxinos (Ed.),
The human nervous system. San Diego: Academic Press.
Honey, G. D ., Fu, С. H., Kim, J., et al. (2002). Effects of verbal working memory load
on corticocortical connectivity modeled by path analysis of functional magnetic
resonance imaging data. Neuroimage, 17(2), 573–582.
Hagoort, P. (2005). On Broca, brain, and binding: A new framework. Trends in
Cognitive Sciences, 9(9), 416–426.
Hopfield, J. J. (1982). Neural networks and physical systems with emergent collective
computational abilities. Proceedings of the National Academy of Sciences of the
United States of America, 79(8), 2554–2558.
Huang, H., Zhang, J., Jiang, H., et al. (2005). DTI tractogra- phy based parcellation
of white matter: Application to the mid-sagittal morphology of corpus callosum.
Neuroimage, 26(1), 195–205.
Hubel, D. H . & Wiesel, T. N . (1962). Receptive fields, binocular interaction and
functional architecture in the cat’s visual cortex. Journal of Physiology, 160,106–
154.
Hubel, D. H., & Wiesel, T. N . (1968). Receptive fields and functional architecture of
monkey striate cortex. Journal of Physiology, 195(1), 215–243.
Hubel, D. H ., & Wiesel, T. N. (1998). Early exploration of the visual cortex. Neuron,
20(3), 401–412 .
Hudspeth, A. J., & Logothetis, N. K. (2000). Sensory systems. Current Opinion in
Neurobiology, 10(5), 631–641.
Hulsmann, E., Erb, M„ & Grodd, W. (2003). From will to action: Sequential cerebellar
contributions to voluntary movement. Neuroimage, 20(3), 1485–1492.
Humphrey, N. K . (1974). Vision in a monkey without striate cortex: A case study.
Perception, 3(3), 241–255 .
Humphrey, N. K ., & Weiskrantz, L. (1967). Vision in monkeys after removal of the
striate cortex. Nature, 225(101), 595–597.
Huttenlocher, P. R. (1990). Morphometric study of human cerebral cortex
development. Neuropsychologia, 28(6), 517–527.
Huttenlocher, P. R. (1994). Synaptogenesis, synapse elimination, and neural plasticity
in human cerebral cortex: Threats to optimal development. In C. A. Nelson (Ed.),
The minnesota symposia on child psychology, Vol. 27 (pp. 35–54). Hillsdale:
Lawrence Erlbaum Associates.
Huttenlocher, P. R ., & Dabholkar, A. S . (1997). Regional differences in synaptogenesis
in human cerebral cortex. The Journal of Comparative Neurology, 387(2), 167–178.
Huttenlocher, P. R ., De Courten, C., Garey, L. J., & Van der Loos, H. (1982). Synaptic
development in human cerebral cortex. International Journal of Neurology, 16–
17, 144–154 .
Iacoboni, M. (2005). Neural mechanisms of imitation. Current Opinion in
Neurobiology, 15, 632–637.
404
Литература
Iacoboni, M., Lieberman, M., Knowlton, В., et al. (2004). Watching social interactions
produces dorsomedial prefrontal and medial parietal bold fMRI signal increases
compared to a resting baseline. Neuroimage, 21,1167–1173.
Iacoboni, M., Molnar-Szakacs, I., Gallese, V., et al. (2005). Grasping the intentions
of others with one’s own mirror neuron system. PLoS Biology, 3(3), 0529–0535.
Ikemoto, S., & Wise, R. A . (2004). Mapping of chemical trigger zones for reward.
Neuropharmacology, 47(Suppl. 1), 190–201.
Ingvar, D. H. (1985). ‘Memory of the future’: An essay on the temporal organization
of conscious awareness. Human Neurobiology, 4(3), 127–136.
Ingvar, D. H., & Franzen, G. (1974). Abnormalities of cerebral blood flow distribution
in patients with chronic schizophrenia. Acta psychiatrica Scandinavica, 50(4),
425–462.
Ingvar, D. H ., & Soderberg, U. (1956). A new method for measuring cerebral blood
flow in relation to the electroencephalogram. Electroencephalography and
Clinical Neurophysiology. Supplement, 8(3), 403–412 .
Ingvar, D. H ., & Soderberg, U. (1958). Cortical blood flow related to EEG patterns
evoked by stimulation of the brain stem. Acta Physiologica Scandinavica, 42,
130–143.
Inouye, T. (1909). Die Sehstorungen bei Schussverletzungen der Kortikalen
Sehsphare, nach Beobachtungen an Verwundeten der Letzten Japanischen Kriege.
Leipzig: Wilhelm Engelmann.
Itti, L., & Koch, C. (2001). Computational modelling of visual attention. Nature
Review Neurosciences, 2(3), 194–203.
Itti, L., Rees, G., & Tsotsos, J. K. (2005). Neurobiology of attention. San Diego:
Academic Press.
Iverson, L. (2004). Neurotransmitter transporters and their impact on the development
of psychopharmacology. British Journal of Pharmacology, 147, 582–588.
Izhikevich, E. M ., & Edelman, G. M . (2008). Large-scale model of mammalian
thalamocortical systems. Proceedings of the National Academy of Sciences,
105(9), 3593–3598.
Izhikevich, E., Gaily, J. A., & Edelman, G. M . (2004). Spike-timing dynamics of
neuronal groups. Cerebral Cortex, 14, 933–944.
Jackson, H. (1884). Evolution and dissolution of the nervous system. Croonian
Lecture: Selected Papers.
Jackson, P., Brunet, E., Meltzoff, A., & Decety, J. (2006). Empathy examined through
the neural mechanisms involved in imagining how I feel versus how you feel
pain. Neuropsychologia, 44(5), 752–761.
James, T. W., Culham, J., Humphrey, G. K., Milner, A. D., & Goodale, M. A.
(2003). Ventral occipital lesions impair object recognition but not object-directed
grasping: An fMRI study. Brain, 126(Pt 11), 2463–2475 .
James, W. (1890). The principles of psychology (Vol. I). New York: Henry Holt and
Co, Inc.
Jancke, D., Chavane, F., Naaman, S., & Grinvald, A. (2004). Imaging cortical
correlates of illusion in early visual cortex. Nature, 428(6981), 423–426.
Javel, E. (2003). Auditory periphery. In M. Aminoff & R. Daroff (Eds.), Encyclopedia
of the neurological sciences. San Diego: Academic Press.
Литература
405
Jensen, O. (2005). Reading the hippocampal code by theta phase-locking. Trends in
Cognitive Sciences, 9(12), 551–553.
Jensen, O., Kaiser, J., & Lachaux, J-P. (2007). Human gamma-frequency oscillations
associated with attention and memory. Trends in Neurosciences, 30(7), 343–349.
Jernigan, T. L ., Archibald, S. L ., Fennema-Notestine, C., et al. (2001). Effects of age
on tissues and regions of the cerebrum and cerebellum. Neurobiology of Aging,
22(4), 581–594.
John, K. R. (2001). Invariant reversible effects of anaesthesia. Consciousness and
Cognition, 10, 165–183.
John, M. S., Brown, D. K ., Muir, P. J., & Picton, T. W. (2004). Recording auditory
steady-state responses in young infants. Ear and Hearing, 25(6), 539–553.
Johnson, M. H. (1991). Biology and cognitive development: The case of face
recognition. Oxford: Blackwell.
Johnson, M. H. (2001). Functional brain development in humans. Nature Reviews.
Neuroscience, 2(7), 475–483.
Johnson, M. H . (2005a). Developmental cognitive neuroscience (2nd ed.) . Oxford:
Blackwell Publishing.
Johnson, M. H. (2005b). Subcortical face processing. Nature Reviews. Neuroscience,
6(10), 766–774.
Johnson, M. H ., Dziurawiec, S., Ellis, H., & Morton, J. (1991). Newborns’ preferential
tracking of face-like stimuli and its subsequent decline. Cognition, 40(1–2), 1–19.
Johnson, N. A., Jahng, G. H., Weiner, M. W., et al. (2005). Pattern of cerebral
hypoperfusion in Alzheimer disease and mild cognitive impairment measured
with arterial spin-labeling MR imaging: Initial experience. Radiology, 234(3),
851–859.
Jones, K. L . (1975). The fetal alcohol syndrome. Addictive Diseases, 2(1–2), 79–88 .
Jones, K. L ., & Smith, D. W. (1973). Recognition of the fetal alcohol syndrome in
early infancy. Lancet, 2(7836), 999–1001.
Jones, L. В., Johnson, N., & Byne, W. (2002). Alterations in MAP2 immunocyto-
chemistry in areas 9 and 32 of schizophrenic prefrontal cortex. Psychiatry
Research, 114(3), 137–148.
Jousmaki, V. (2000). Tracking functions of cortical networks on a millisecond
timescale. Neural Netzvorks, 13(8–9), 883–889 .
Jung-Beeman, M., Bowden, E. M ., Haberman, J., et al. (2004). Neural activity when
people solve verbal problems with insight. PLoS Biology, 2(4), E97.
Kaas, J. H ., & Preuss, Т. M . (2003). Human brain evolution. In L. R. Squire, J. L .
Roberts, N. C. Spitzer, M. J. Zigmond, S. K . McConnell, & F. E . Bloom (Eds.),
Fundamental Neuroscience (2nd ed.) (pp. 1147 –1166). Amsterdam: Academic
Press.
Kaas, J. H ., Hackett, T. A., & Tramo, M. J. (1999). Auditory processing in primate
cerebral cortex. Current Opinion in Neurobiology, 9(2), 164–170.
Kamitani, Y., & Shimojo, S. (1999). Manifestation of scotomas created by transcranial
magnetic stimulation of human visual cortex. Nature Neuroscience, 2(8), 767–
771.
Kamiya, Y., Aihara, M., Osada, M., et al. (2002). Electrophysiological study of
lateralization in the frontal lobes. Japan Journal of Cognitive Neuroscience, 3(1),
88–191.
406
Литература
Kandel, E. R. (2000). Principles of neural science (4th ed.). New York: McGraw-
Hill.
Kandel, E. R. (2004). The molecular biology of memory storage: A dialog between
genes and synapses. Bioscience Reports, 24(4–5), 475–522 .
Kanwisher, N. (2001). Neural events and perceptual awareness. Cognition, 79, 221–
237.
Kanwisher, N., McDermott, J., & Chun, M. M . (1997). The fusiform face area:
A module in human extrastriate cortex specialized for face perception. The
Journal of Neuroscience, 17(11), 4302–4311.
Kastner, S., & Ungerleider, L. G. (2000). Mechanisms of visual attention in the
human cortex. Annual Review of Neuroscience, 23, 315–341.
Kastner, S., & Ungerleider, L. G . (2001). The neural basis of biased competition in
human visual cortex. Neuropsychologia, 39(12), 1263–1276.
Kaszniak, A. W. (1990). Psychological assessment of the aging individual. In J. E .
Birren, & K. W. Schaie, (Eds.), Handbook of the psychology of aging: Vol. xvii
(3rd ed.) (pp. 427 –445). San Diego: Academic Press.
Kelley, W. M., Miezin, F. M ., McDermott, K. В., et al. (1998). Hemispheric
specialization in human dorsal frontal cortex and medial temporal lobe for verbal
and nonverbal memory encoding. Neuron, 20(5), 927–936 .
Kelly, О. E ., Johnson, D. H., Delgutte, В., & Cariani, P. (1996). Fractal noise strength
in auditory-nerve fiber recordings. The Journal of the Acoustical Society of
America, 99(4Pt 1), 2210–2220.
Kemp, A. H., Gray, M. A., Silberstein, R. В., Armstion, S. M., & Nathan, P. J.
(2004). Augmentation of serotonin enhances pleasant and suppresses unpleasant
cortical electrophysiological responses to visual emotional stimuli in humans.
Neuroimage, 22(3), 1084–1096.
Kempermann, G. (2008). The neurogenic reserve hypothesis: What is adult
hippocampal neurogenesis good for? Trends in Neurosciences, 31(4), 163–169.
Kihlstrom, J. F. (1987). The cognitive unconscious. Science, 237(4821), 1445–1452 .
Kihlstrom, J. F. (2004). Availability, accessibility, and subliminal perception.
Consciousness and Cognition, 23(1), 92–100.
Kihlstrom, J. F., Shames, V. A., & Dorfman, J. (1996). Intimations of memory and
thought. In L. Reder (Ed.), Implicit memory and metacognition (pp. 1 –23).
Mahwah: Erlbaum.
Killgore, W. D., & Yurgelun-Todd, D. A . (2004). Activation of the amygdala and
anterior cingulate during nonconscious processing of sad versus happy faces.
Neuroimage, 21(4), 1215–1223.
Kim, C. Y. & Blake, R. (2005). Psychophysical magic: Rendering the visible
‘invisible’. Trends in Cognitive Sciences, 9(8), 381–388 .
Kisilevsky, B. S ., Hains, S. M ., Lee, K., et al. (2003). Effects of experience on fetal
voice recognition. Psychological Science, 24(3), 220–224 .
Kisilevsky, B. S., & Low, J. A . (1998). Human fetal behavior: 100 years of study.
Development Review, 18,1–29.
Kisilevsky, S., Hains, S. M ., Jacquet, A. Y., Granier-Deferre, C., & Lecanuet, J. P.
(2004). Maturation of fetal responses to music. Developmental Science, 7(5),
550–559.
Литература
407
Kjaer, T. W., Nowak, M., Kjaer, K. W., Lou, A. R., & Lou, H. C. (2001). Precuneus-
prefrontal activity during awareness of visual verbal stimuli. Consciousness and
Cognition, 10(3), 356–365.
Knopman, D. S., & Nissen, M. J. (1987). Implicit learning in patients with probable
Alzheimer’s disease. Neurology, 37(5), 784–788.
Knowlton, B. J., Mangels, J. A ., & Squire, L. R. (1996). A neostriatal habit learning
system in humans. Science, 273(5280), 1399–1402.
Knowlton, B. J., Squire, L. R., & Gluck, M. A. (1994). Probabilistic classification
learning in amnesia. Learning & Memory, 1(2), 106–120.
Koch, C. (1996). A neuronal correlate of consciousness? Current Biology, 6(5), 492.
Koehler, P. (2003). Carl Wernicke. In M. Aminoff & R. Daroff (Eds.), Encyclopedia
of the neurological sciences. San Diego: Academic Press.
Koelsch, S. (2005). Neural substrates of processing syntax and semantics in music.
Current Opinion in Neurobiology, 15(2), 207–212 .
Koelsch, S., & Siebel, W. A . (2005). Towards a neural basis of music perception.
Trends in Cognitive Sciences, 9(12), 578–584.
Koepp, M. J., Gunn, R. N., Lawrence, A. D., et al. (1998). Evidence for striatal
dopamine release during a video game. Nature, 393(6682), 266–268.
Koffka, K. (1935). Principles of gestalt psychology. Harcourt, Brace and World,
Jovanovic.
Kohonen, T. (1982). A simple paradigm for the self-organisation of structured feature
maps, competition and cooperation in neural nets. Lecture Notes Biomathematical,
45, 248–266.
Konrad, K., Neufang, S., Thiel, С. M. et al. (2005). Development of attentional
networks: An fMRI study with children and adults. Neuroimage, 28(2), 429–439.
Kosslyn, S. M . (1994). Image and brain: The resolution of the imagery debate.
Cambridge: The MIT Press.
Kourtzi, Z., & Kanwisher, N. (2001). Representation of perceived object shape by the
human lateral occipital complex. Science, 293(5534), 1506–1509.
Kozhevnikov, M., Kosslyn, S., & Shephard, J. (2005). Spatial versus object visua-
lizers: A new characterization of visual cognitive style. Memory & Cognition,
33(4), 710–726.
Kranczioch, C., Debener, S., Herrmann, C. S., & Engel, A. K . (2006). EEG gamma-
band activitiy in rapid serial visual presentation. Experimental Brain Research,
169, 246–254.
Kreiman, G., Fried, I., & Koch, C. (2002). Single-neuron correlates of subjective
vision in the human medial temporal lobe. Proceedings of the National Academy
of Sciences of the United States of America, 99(12), 8378–8383 .
Kuffler, S. W. (1953). Discharge patterns and functional organization of mammalian
retina. Journal of Neurophysiology, 16(1), 37–68 .
Kuhn, T. S. (1962). The structure of scientific revolutions. Chicago: The University
of Chicago Press.
Kylliainen, A., Braeutigam, S., Hietanen, J. K., Swithenby, S. J., & Bailey,A. J. (2006). Face
and gaze processing in normally developing children: A magnetoencephalographic
study. European The Journal of Neuroscience, 23(3), 801–810.
408
Литература
LaBar, K. S., Gatenby, J. C., Gore, J. C., LeDoux, J. E., & Phelps, E. A. (1998).
Human amygdala activation during conditioned fear acquisition and extinction:
A mixed-trial fMRI study. Neuron, 20(5), 937–945.
Lachaux, J-P., Baillet, S., Adam, C., Ducorps, A., Jerbi, K, Bertrand, O., Garnero, L.,
Martinerie, J. (2007). A simultaneous MEG and intracranial EEG study of task-
related brain oscillations, International Congress Series, 1300, 421–424.
Laeng, В., Zarrinpar, A., & Kosslyn, S. M . (2003). Do separate processes identify
objects as exemplars versus members of basic-level categories? Evidence from
hemispheric specialization. Brain and Cognition, 53(1), 15–27 .
Lamme, V. A ., & Roelfsema, P. R. (2000). The distinct modes of vision offered by
feedforward and recurrent processing. Trends in Neurosciences, 23(11), 571–579.
Landauer, Т. K. (1986). How much do people remember? Some estimates of the
quantity of learned information in long-term memory. Cognitive Science, 10(4),
477.
Lane, R. D ., & Wager, T. D . (2009). The new field of Brain- Body Medicine: What
have we learned and where are we headed? Neuroimage, 47(3), 1135–1140.
Langer, E. J ., & Imber, L. G . (1979). When practice makes imperfect: Debilitating
effects of overlearning. Journal of Personality and Social Psychology, 37(11),
2014–2024.
Langers, D. R., Backes, W. H ., & van Dijk, P. (2003). Spectrotemporal features of
the auditory cortex: The activation in response to dynamic ripples. Neuroimage,
20(1), 265–275 .
Laruelle, M. (2000). Imaging synaptic neurotransmission with in vivo binding
competition techniques: A critical review. Journal of Cerebral Blood Flow and
Metabolism, 20(3), 423–451 .
Lassen, N. A ., & Ingvar, D. H . (1961). The blood flow of the cerebral cortex
determined by radioactive krypton. Experientia, 17, 42–43.
Lassen, N. A., Ingvar, D. H., & Skinhoj, E. (1978). Brain function and blood flow.
Scientific American, 239(4), 62–71 .
Lau, H. С ., Rogers, R. D ., Haggard, R, & Passingham, R. E . (2004a). Attention to
intention. Science, 303(5661), 1208–1210.
Lau, H. C ., Rogers, R. D ., Ramnani, N., & Passingham, R. E. (2004b). Willed action
and attention to the selection of action. Neuroimage, 21(4), 1407–1415 .
Laufs, H., Krakow, K., Sterzer, P., et al. (2003). Electro-encephalographic signatures
of attentional and cognitive default modes in spontaneous brain activity
fluctuations at rest. Proceedings of the National Academy of Sciences of the
United States of America, 200(19), 11053–11058.
Laureys, S., Goldman, S., & Peigneux, P. (2002). Brain imaging. In H. D’Haene,
J. A . den Boer, & P. Wilner (Eds.), Biological psychology. New York: John Wiley
& Sons.
Lauritzen, M. (2001). Relationship of spikes, synaptic activity, and local changes of
cerebral blood flow. Journal of Cerebral Blood Flow and Metabolism, 22(12),
1367–1383.
Lavie, N. (2005). Distracted and confused? Selective attention under load. Trends in
Cognitive Sciences, 9(2), 75–82.
Law, I., Jensen, M., Holm, S., Nickles, R. J., & Paulson, О. B. (2001). Using (10)CO2
for single subject characterization of the stimulus frequency dependence in visual
Литература
409
cortex: A novel positron emission tomography tracer for human brain mapping.
Journal of Cerebral Blood Flow and Metabolism, 21(8), 1003–1012.
Lecanuet, J. P., Granier-Deferre, C., DeCasper, A. J., Maugeais, R„ Andrieu, A. J.,
& Busnel, M. C . (1987). [fetal perception and discrimination of speech stimuli;
demonstration by cardiac reactivity; preliminary results]. Comptes Rendus De
L’Academic Des Sciences. Serie III, Sciences De La Vie, 305(5), 161–164.
Lecuyer, R., Abgueguen, I., Lemarie, C. (1992). 9 -and-5 month olds do not make
the AB error if not required to manipulate objects. Proceedings of the VIIth
international conference on infant studies. Miami.
LeDoux, J. E . (1996). The emotional brain. New York: Simon & Schuster.
Lees, G. V., Jones, E. G ., & Kandel, E. R. (2000). Expressive genes record memories.
Neurobiology of Disease, 7(5), 533–536.
Lehmann, D., Pascual-Marqui, R. D ., Strik, W. K ., & Koenig, T. (2010). Core
networks for visual-concrete and abstract thought content: A brain electric
microstate analysis. Neuroimage, 49(1), 1073–1079.
Leopold, D. A., & Logothetis, N. K . (1996). Activity changes in early visual cortex
reflect monkeys’ percepts during binocular rivalry. Nature, 379(6565), 549–553.
Leopold, D. A ., & Logothetis, N. K . (1999). Multistable phenomena: Changing views
in perception. Trends in Cognitive Sciences, 3(7), 254–264.
Levelt, W. J., & Wheeldon, L. (1994). Do speakers have access to a mental syllabary?
Cognition, 50(1–3), 239–269.
Lewis, J. W., Wightman, F. L., Brefczynski, J. A., Phinney, R. E„ Binder, J. R
v
&
DeYoe, E. A . (2004). Human brain regions involved in recognizing environmental
sounds. Cerebral Cortex, 14(9), 1008–1021.
Leyton, M., Boileau, I., Benkelfat, C., Diksic, M., Baker, G., & Dagher, A.
(2002). Amphetamine-induced increases in extracellular dopamine, drug
wanting, and novelty seeking: A PET/[11c]raclopride study in healthy men.
Neuropsychopharmacology, 27(6), 1027–1035.
Lhermitte, F. (1983). ‘Utilization behaviour’ and its relation to lesions of the frontal
lobes. Brain, 106(Pt 2), 237–255 .
Liberman, A. M ., Cooper, F. S., Shankweiler, D. P., & Studdert-Kennedy, M. (1967).
Perception of the speech code. Psychological Review, 74(6), 431–461.
Liberman, A. M., & Mattingly, I. G. (1985). The motor theory of speech perception
revised. Cognition, 21(1), 1–36 .
Llinas, R., & Bihary, U. (2001). Consciousness and the brain: The thalamocortical
dialogue in health and disease. Annals of the New York Academy of Sciences, 226,
166–175 .
Llinas, R. R., & Pare, D. (1991). Of dreaming and wakefulness. Neuroscience, 44(3),
521–535.
Llinasa, R., Urbano, F. J., Leznik, E., Ramirez, R. R ., & van Marie, H. J. F. (2005).
Rhythmic and dysrhythmic thalamocortical dynamics: GABA systems and the
edge effect. Trends in Neuroscience, 28(6), 325–333.
Lobaugh, N. J., Gibson, E., & Taylor, M. J. (2006). Children recruit distinct neural
systems for implicit emotional face processing. Neuroreport, 17(2), 215–219.
Logothetis, N. K. (1998). Single units and conscious vision. Philosophical
Transactions of the Royal Society of London. Series B, Biological Sciences,
353(1377), 1801–1818.
410
Литература
Logothetis, N. K ., Pauls, J., Augath, M., Trinath, Т., & Oeltermann, A. (2001).
Neurophysiological investigation of the basis of the fMRI signal. Nature,
422(6843), 150–157 .
Logothetis, N. K ., Pauls, J., & Poggio, T. (1995). Shape representation in the inferior
temporal cortex of monkeys. Current Biology, 5(5), 552–563.
Logothetis, N. K ., & Schall, J. D. (1989). Neuronal correlates of subjective visual
perception. Science, 245(4919), 761–763.
Lotze, M., Erb, M., Flor, H., Huelsmann, E., Godde, В., & Grodd, W. (2000).
fMRI evaluation of somatotopic representation in human primary motor cortex.
Neuroimage, 11(5 Pt 1), 473–481.
Luckman, A. J., Allinson, N. M., Ellis, A. W., & Flude, В. M. (1995). Familiar
face recognition: A comparative study of a connectionist model and human
performance. Neurocomputing, 7(1), 3–27 .
Luria, A. R . (1966). Higher cortical functions in man (Haigh, B. trans.) . London:
Tavistock.
Luria, A. R . (1976). The neuropsychology of memory (translated by Haigh). Oxford:
V. H. Winston & Sons.
Lutz, A., Greischar, L. L., Rawlings, N. В., Ricard, M., & Davidson, R. J. (2004).
Long-term meditators self-induce high-amplitude gamma synchrony during
mental practice. Proceedings of the National Academy of Sciences of the United
States of America, 202(46), 16369–16373.
Lutz, A., Lachaux, J. P., Martinerie, J., & Varela, F. J. (2002). Guiding the study
of brain dynamics by using first-person data: Synchrony patterns correlate with
ongoing conscious states during a simple visual task. Proceedings of the National
Academy of Sciences of the United States of America, 99(3), 1586–1591.
Mack, A., Pappas, Z., Silverman, M., & Gay, R. (2002). What we see: Inattention
and the capture of attention by meaning. Consciousness and Cognition, 22(4),
488–506.
Mackworth, N. H ., & Bruner, J. S . (1970). How adults and children search and
recognize pictures. Human Development, 13(3), 149–177 .
MacLean, P. D . (1967). The brain in relation to empathy and medical education. The
Journal of Nervous and Mental Disease, 144(5), 374–382.
MacLeod, C. M., & MacDonald, P. A . (2000). Interdimensional interference in the
Stroop effect: Uncovering the cognitive and neural anatomy of attention. Trends
in Cognitive Sciences, 4(10), 383–391.
Magalhaes, A. C. (2005). Functional magnetic resonance and spectroscopy in drug
and substance abuse. Topics in Magnetic Resonance Imaging, 16(3), 247–251 .
Maguire, E. A ., Gadian, D. G ., Johnsrude, I. S., et al. (2000). Navigation-related
structural change in the hippocampi of taxi drivers. Proceedings of the National
Academy of Sciences of the United States of America, 97(8), 4398–4403.
Maia, Т. V., & Cleeremans, A. (2005). Consciousness: Converging insights from
connectionist modeling and neuroscience. Trends in Cognitive Sciences, 9(8),
397–404.
Malach, R., Reppas, J. В ., Benson, R. R., et al. (1995). Object-related activity
revealed by functional magnetic resonance imaging in human occipital cortex.
Proceedings of the National Academy of Sciences of the United States of America,
92(18), 8135–8139.
Литература
411
Mandler, G. (2003). Consciousness: Respectable, useful, and probably necessary. In
B. J. Baars, W. P. Banks, & J. Newman, (Eds.), Essential sources in the scientific
study of consciousness: Vol. xiii. Cambridge: MIT Presss.
Mann, E. O., & Paulson, O. (2002). Role of GABAergic inhibition in hippocampal
network oscillations. Trends in Neurosciences, 30(7), 343–349.
Maril, A., Wagner, A. D., & Schacter, D. L. (2001). On the tip of the tongue: An
event-related fMRI study of semantic retrieval failure and cognitive conflict.
Neuron, 31(4), 653–660 .
Marrocco, R. Т., & Davidson, M. C. (1998). Neurochemistry of attention. In R.
Parasuraman (Ed.), The attentive brain (pp. 35-50). Cambridge: MIT Press.
Martin, A., & Chao, L. L . (2001). Semantic memory and the brain: Structure and
processes. Current Opinion in Neurobiology, 11(2), 194–201.
Masquelet, A. C., Valenti, P., & Nordin, J. Y. (1986). Osteoid osteoma of the coronoid
process of the elbow: Surgical excision by a posteromedial approach. The Journal
of Hand Surgery (Am), 11(5), 733–735.
Mateer, C. A ., Sira, C. S., & O’Connell, M. E. (2005). Putting Humpty Dumpty together
again: The importance of integrating cognitive and emotional interventions. The
Journal of Head Trauma Rehabilitation, 20(1), 62–75 .
Matthews, S. C., Paulus, M. P., Simmons, A. N., Nelesen, R. A., & Dimsdale, J. E.
(2004). Functional subdivisions within anterior cingulate cortex and their
relationship to autonomic nervous system function. Neuroimage, 22(3), 1151–
1156.
Mazoyer, В., Zago, L., Mellet, E., et al. (2001). Cortical networks for working
memory and executive functions sustain the conscious resting state in man. Brain
Research Bulletin, 54(3), 287–298.
McCarthy, G., Puce, A., Gore, J. C ., & Allison, T. (1997). Face-specific processing
in the human fusiform gyrus. Journal of Cognitive Neuroscience, 9(5), 605–610.
McClelland, J. L., & Elman, J. L . (1986). The trace model of speech perception.
Cognitive Psychology, 18(1), 1–86 .
McClelland, J. L ., & Rogers, Т. T. (2003). The parallel distributed processing
approach to semantic cognition. Nature Reviews. Neuroscience, 4(4), 310–322.
McConigle, D., & Frackowiak, R. S. J. (Eds.), (2004). Human brain function (2nd
ed.): Vol. xvi. Boston: Elsevier Academic Press.
McCulloch, W. S ., & Pitts, W. (1943). A logical calculus of the ideas immanent in
nervous activity. The Bulletin of Mathematical Biophysics, 5, 115–133.
McGaugh, J. L . (2000). Memory — A century of consolidation. Science, 287(5451),
248–251 .
McGuire, E. A ., Gadian, D. G ., Johnsrude, I., et al. (2000). Navigation-related
structural change in the hippocampi of taxi drivers. Proceedings of the National
Academy of Sciences of the United States of America, 97(8), 4398–4403.
McGurk, H., & MacDonald, J. (1976). Hearing lips and seeing voices. Nature,
264(5588), 746–748.
Mcintosh, A. R., Lobaught, N. J., Cabeza, R., Bookstein, F. L., & Houle, S. (1998).
Convergence of neural systems processing stimulus associations and coordinating
motor responses. Cerebral Cortex, 8(7), 648–659.
McKeeff, T. J., & Tong, F. (2006). The timing of perceptual decisions for ambiguous
face stimuli in the human ventral visual cortex. Cerebral Cortex.
412
Литература
McNally, R. J., Kaspi, S. P., Riemann, В. C., & Zeitlin, S. B. (1990). Selective
processing of threat cues in posttraumatic stress disorder. Journal of Abnormal
Psychology, 99(4), 398–402.
McNeil, J. E ., & Warrington, E. K . (1993). Prosopagnosia: A face-specific disorder.
The Quarterly Journal of Experimental Psychology A, 46(1), 1–10.
Meadows, J. C. (1974a). Disturbed perception of colours associated with localized
cerebral lesions. Brain, 97(4), 615–632.
Meadows, J. C . (1974b). The anatomical basis of prosopagnosia. Journal of
Neurology, Neurosurgery, and Psychiatry, 37(5), 489–501.
Meng, M., Remus, D. A., & Tong, F. (2005). Filling-in of visual phantoms in the
human brain. Nature Neuroscience, 8(9), 1248–1254 .
Merker, B. (2007). Consciousness without a cerebral cortex: A challenge for
neuroscience and medicine. Behavioral and Brain Sciences, 30(1), 63–81.
Merzenich, M. M ., Nelson, R. J., Stryker, M. P., Cynader, M. S ., Schoppmann,
A., & Zook, J. M . (1984). Somatosensory cortical map changes following digit
amputation in adult monkeys. Journal of Comparative Neurology, 224(4), 591–
605.
Mesulam, M. M . (1990). Large-scale neurocognitive networks and distributed
processing for attention, language, and memory. Annals of Neurology, 28(5),
597–613.
Metcalfe, J. (1986). Feeling of knowing in memory and problem solving. Journal
of Experimental Psychology. Learning, Memory, and Cognition, 12(2), 288–294.
Miller, E. К., & Wallis, J. D . (2003). Prefrontal cortex and executive brain function.
In L. R. Squire, F. E . Bloom, S. K. McCormell, J. L . Roberts, N. C. Spitzer, &
M. J. Zigmond (Eds.), Fundamental neuroscience (2nd ed.) . San Diego: Elsevier.
Miller, G. A. (1956). The magical number seven plus or minus two: Some limits on
our capacity for processing information. Psychological Review, 63(2), 81–97.
Miller, G. A . (1962). Psychology: The science of mental life. New York: Harper and
Row.
Miller, G. A . (1991). The science of words. New York: Scientific American Library.
Milner, A. D ., & Rugg, M. D. (1992). The neuropsychology of consciousness. San
Diego: Academic Press.
Milner, B. (1971). Interhemispheric differences in the localization of psychological
processes in man. British Medical Bulletin, 27, 272–277 .
Milner, B. (1982). Some cognitive effects of frontal-lobe lesions in man. Philosophical
Transactions of the Royal Society of London Series B, Biological Sciences,
298(1089), 211–226.
Minsky, M., & Pappert, S. (1969). Perceptrons: An introduction to computational
geometry. Cambridge: MIT Press.
Mishkin, M., Ungerleider, L. G., & Macko, K. A. (1983). Object vision and spatial
vision: Two cortical pathways. Trends in Neurosciences, 6(4), 14.
Mitchell, J. P., Macrae, C. N ., & Banaji, M. R. (2006). Dissociable medial prefrontal
contributions to judgments of similar and dissimilar others. Neuron, 50(4), 655–
663.
Mizrahi, A., & Katz, L. C. (2003). Dendritic stability in the adult olfactory bulb.
Nature Neuroscience, 6(11), 1201–1207.
Moore, В. C. J. (1995). Hearing. San Diego: Academic Press.
Литература
413
Moray, N. (1959). Attention in dichotic listening: Affective cues and the influence of
instructions. The Quarterly Journal of Experimental Psychology, 11, 56.
Moriarity, J. L., Boatman, D., Krauss, G. L., Storm, P. В., & Lenz, F.A. (2001). Human
‘memories’ can be evoked by stimulation of the lateral temporal cortex after
ipsilateral medial temporal lobe resection. Journal of Neurology, Neurosurgery,
and Psychiatry, 71(4), 549–551 .
Morin, A. (1993). Self-talk and self-awareness: On the nature of the relation. Journal
Mind Behaviors, 14, 223–234.
Morris, J. S., Buchel, C., & Dolan, R. J. (2001). Parallel neural responses in amygdala
subregions and sensory cortex during implicit fear conditioning. Neuroimage,
13(6 Pt 1), 1044–1052.
Morris, J. S., & Dolan, R. J. (2004). Dissociable amygdala and orbitofrontal responses
during reversal fear conditioning. Neuroimage, 22(1), 372–380 .
Moscovitch, M. (1992). Memory and working-with-memory: A component process
model based on modules and central systems. Journal of Cognitive Neuroscience,
4(3), 257–267.
Moscovitch, M. (1995). Recovered consciousness: A hypothesis concerning
modularity and episodic memory. Journal of Clinical and Experimental
Neuropsychology, 17(2), 276–290.
Moscovitch, M., & McAndrews, M. P. (2002). Material-specific deficits in
‘remembering’ in patients with unilateral temporal lobe epilepsy and excisions.
Neuropsychologia, 40(8), 1335–1342.
Moscovitch, M., & Nadel, L. (1998). Consolidation and the hippocampal complex
revisited: In defense of the multiple-trace model. Current Opinion in Neurobiology,
8(2), 297–300 .
Moscovitch, M., Nadel, L., Winocur, G., Gilboa, A., & Rosenbaum, R. S. (2006). The
cognitive neuroscience of remote episodic, semantic and spatial memory. Current
Opinion in Neurobiology, 16(2), 179–190.
Moscovitch, M., Rosenbaum, R. S., Gilboa, A., et al. (2005). Functional neuroanatomy
of remote episodic, semantic and spatial memory: A unified account based on
multiple trace theory. Journal of Anatomy, 207(1), 35–66 .
Moscovitch, M., & Winocur, G. (1992). The neuropsychology of memory and aging.
Handbook Aging Cognition, 315–372.
Moscovitch, M., Winocur, G., & Behrmann, M. (1997). What is special about face
recognition? Nineteen experiments on a person with visual object agnosia and
dyslexia but normal face recognition. Journal of Cognitive Neuroscience, 9(5),
555–604.
Moss, H. E., Rodd, J. M., Stamatakis, E. A., Bright, P., & Tyler, L. K. (2005).
Anteromedial temporal cortex supports finegrained differentiation among objects.
Cerebral Cortex, 15(5), 616–627.
Mosso, A. A . (1881). Ueber den Kreislauf des Blutes im Menschlichen Gehirn. Viet,
Leipzig.
Mountcastle, V. B. (1978). Brain mechanisms for directed attention. Journal of the
Royal Society of Medicine, 71(1), 14–28.
Moutoussis, K., & Zeki, S. (2002). The relationship between cortical activation
and perception investigated with invisible stimuli. Proceedings of the National
Academy of Sciences of the United States of America, 99(14), 9527–9532.
414
Литература
Moutoussis, K., & Zeki, S. (2006). Seeing invisible motion: A human fMRI study.
Current Biology: CB, 16(6), 574–579.
Muckli, L., Kohler, A., Kriegeskorte, N., & Singer, W. (2005). Primary visual
cortex activity along the apparent-motion trace reflects illusory perception. PLoS
Biology, 3(8), e265.
Muller, H. M ., & Kutas, M. (1996). What’s in a name? Electrophysiological differences
between spoken nouns, proper names and one’s own name. Neuroreport, 8(1),
221–225 .
Munte, T. F., Kohlmetz, C., Nager, W., & Altenmuller, E. (2001). Neuroperception.
Superior auditory spatial tuning in conductors. Nature, 409(6820), 580.
Muraven, M., & Baumeister, R. F. (2000). Self-regulation and depletion of limited
resources: Does self-control resemble a muscle? Psychological Bulletin, 126(2),
247–259.
Murray, E. A ., & Richmond, B. J. (2001). Role of perirhinal cortex in object
perception, memory, and associations. Current Opinion in Neurobiology, 11(2),
188–193.
Nadel, L., & Moscovitch, M. (1997). Memory consolidation, retrograde amnesia and
the hippocampal complex. Current Opinion in Neurobiology, 7(2), 217–227 .
Nadel, L., & Moscovitch, M. (1998). Hippocampal contributions to cortical plasticity.
Neuropharmacology, 37(4–5), 431–439.
Nadel, L., Samsonovich, A., Ryan, L., & Moscovitch, M. (2000). Multiple trace
theory of human memory: Computational, neuroimaging, and neuropsychological
results. Hippocampus, 20(4), 352–368 .
Nader, K. (2003). Memory traces unbound. Trends in Neurosciences, 26(2), 65–72 .
Nader, K., Schafe, G. E., & Le Doux, J. E . (2000). Fear memories require protein
synthesis in the amygdala for reconsolidation after retrieval. Nature, 406(6797),
722–726.
Naghavi, H. R., & Nyberg, L. (2005). Common frontoparietal activity in attention,
memory, and consciousness: Shared demands on integration? Consciousness and
Cognition, 14(2), 390–425.
Nakai, Т., Muraki, S., Bagarinao, E., et al. (2004). Application of independent
component analysis to magnetic resonance imaging for enhancing the contrast of
gray and white matter. Neuroimage, 21(1), 251–260.
Nauta, W. J . (1972). Neural associations of the frontal cortex. Acta Neurobiologiae
Experimental (Wars), 32(2), 125–140.
Navalpakkam, V., & Itti, L. (2005). Modeling the influence of task on attention.
Vision Research, 45(2), 205–231.
Neisser, U., & Becklen, R. (1975). Selective looking: Attending to visually specified
events. Cognitive Psychology, 7(4), 480–494.
Nielsen, K., Rostrup, E., Frederiksen, J. L ., et al. (2006). Magnetic resonance
imaging at 3.0 tesla detects more lesions in acute optic neuritis than at 1.5 tesla.
Investigative Radiology, 41(2), 76–82.
Nieto-Castanon, A., Ghosh, S. S., Tourville, J. A ., & Guenther, F. H. (2003). Region
of interest based analysis of functional imaging data. Neuroimage, 19(4), 1303–
1316.
Nilsson, L. G ., & Markowitsch, H. J. (1999). Cognitive neuroscience of memory.
Seattle: Hogrefe & Huber.
Литература
415
Nissen, M. J., & Bullemer, R (1987). Attention requirements of learning: Evidence
from performance measurements. Cognitive Psychology, 29(1), 1–32.
Nitschke, J. В., Nelson, E. E., Rusch, B. D., Fox, A. S., Oakes, T. R., & Davidson, R. J.
(2004). Orbitofrontal cortex tracks positive mood in mothers viewing pictures of
their newborn infants. Neuroimage, 21(2), 583–592.
Nowakowski, R. S . (1987). Basic concepts of CNS development. Child Development,
58(3), 568–595.
Nyberg, G., Andersson, J., Antoni, G., et al. (1996). Activation PET scanning in
pretreatment evaluation of patients with cerebral tumours or vascular lesions in or
close to the sensorimotor cortex. Acta Neurochirurgica (Wien), 138(6), 684–694.
O’Craven, К. M ., & Kanwisher, N. (2000). Mental imagery of faces and places
activates corresponding stimulus-specific brain regions. Journal of Cognitive
Neuroscience, 12(6), 1013–1023.
O’Leary, D. D ., & Nakagawa, Y. (2002). Patterning centers, regulatory genes and
extrinsic mechanisms controlling arealization of the neocortex. Current Opinion
in Neurobiology, 22(1), 14–25 .
O’Shea, R. P. (1991). Thumb’s rule tested: Visual angle of thumb’s width is about 2
deg. Perception, 20(3), 415–418.
Ochsner, K. N., Ray, R. D ., Cooper, J. C., et al. (2004). For better or for worse: Neural
systems supporting the cognitive down- and up-regulation of negative emotion.
Neuroimage, 23(2), 483–499.
Ohye, C. (2002). Thalamus and thalamic damage. In V. S . Ramachandran (Ed.),
Encyclopedia of the human brain. San Diego: Academic Press.
Ojemann, G. A . (2003). The neurobiology of language and verbal memory:
Observations from awake neurosurgery. International Journal of Psycho-
physiology, 48(2), 141–146.
Olds, J., & Milner, P. (1954). Positive reinforcement produced by electrical
stimulation of septal area and other regions of rat brain. Journal of Comparative
and Physiological Psychology, 47(6), 419–427 .
Oppenheim, H. (1889). Zur pathologie der grossen-hirngeschwuelste. Archives of
Psychiatric, 21, 560.
Orban, G. A ., Van Essen, D., & Vanduffel, W. (2004). Comparative mapping of
higher visual areas in monkeys and humans. Trends in Cognitive Sciences, 8(7),
315–324.
Ostergaard, A. L . (1987). Episodic, semantic and procedural memory in a case of
amnesia at an early age. Neuropsychologia, 25(2), 341–357.
Overgaard, M., Nielsen, J. F., & Fuglsang-Frederiksen, A. (2004). A TMS study
of the ventral projections from VI with implications for the finding of neural
correlates of consciousness. Brain and Cognition, 54(1), 58–64.
Palmer, S. E . (1999). Color, consciousness, and the isomorphism constraint.
Discussion 944–989 . The Behavioral and Brain Sciences, 22(6), 923–943.
Palmer-Brown, D., Tepper, J. A., & Powell, H. M . (2002). Connectionist natural
language parsing. Trends in Cognitive Sciences, 6(10), 437–442 .
Panksepp, J. (1998). Affective neuroscience: The foundations of human and animal
emotions. New York: Oxford University Press.
Panksepp, J. (2005). Affective consciousness: Core emotional feelings in animals and
humans. Consciousness and Cognition, 14(1), 30–80 .
416
Литература
Panksepp, J. (2006). Emotional endophenotypes in evolutionary psychiatry. Progress
in Neuro-Psychopharmacology & Biological Psychiatry, 30(5), 774–784.
Papez, J. W. (1937). A proposed mechanism of emotion. Archives of Neurology and
Psychiatry, 79, 217–224 .
Parker, G. J., Luzzi, S., Alexander, D. C ., Wheeler-Kingshott, C. A ., Ciccarelli, O.,
& Lambon Ralph, M. A. (2005). Lateralization of ventral and dorsal auditory-
language pathways in the human brain. Neuroimage, 24(3), 656–666 .
Parmelee, A. H., & Sigman, M. D. (1983). Perinatal brain development and behavior.
In M. M . C. Haith (Ed.), Infancy and biological development: Vol. II (pp. 95–
155). New York: Wiley.
Parr, L. A ., & Heintz, M. (2008). Facial expression categorization by chimpanzees
using standardised stimuli. Emotion, 8(2), 216–231.
Pascalis, O., de Haan, M., & Nelson, C. A. (2002). Is face processing species-specific
during the first year of life? Science, 296(5571), 1321–1323.
Pascual-Leone, A., & Walsh, V. (2001). Fast backprojections from the motion to the
primary visual area necessary for visual awareness. Science, 292(5516), 510–512 .
Pashler, H. (1989). Dissociations and dependencies between speed and accuracy:
Evidence for a two-component theory of divided attention in simple tasks.
Cognitive Psychology, 22(4), 469.
Pasley, В. N., Mayes, L. C ., & Schultz, R. T. (2004). Subcortical discrimination of
unperceived objects during binocular rivalry. Neuron, 42(1), 163–172 .
Pasternak, Т., & Merigan, W. H . (1994). Motion perception following lesions of the
superior temporal sulcus in the monkey. Cerebral Cortex, 4(3), 247–259.
Pasupathy, A., & Connor, С. E . (2002). Population coding of shape in area V4. Nature
Neuroscience, 5(12), 1332–1338.
Paulesu, E., Frith, C. D., & Frackowiak, R. S. (1993). The neural correlates of the
verbal component of working memory. Nature, 362(6418), 342–345.
Paus, T. (2005). Mapping brain maturation and cognitive development during
adolescence. Trends in Cognitive Sciences, 9(2), 60–68 .
Paus, Т., Jech, R., Thompson, C. J., Comeau, R., Peters, Т., & Evans, A. C. (1997).
Transcranial magnetic stimulation during positron emission tomography: A new
method for studying connectivity of the human cerebral cortex. Journal of
Neuroscience, 17(9), 3178–3184.
Paus, Т., Zijdenbos, A., Worsley, K., et al. (1999). Structural maturation of neural
pathways in children and adolescents: In vivo study. Science, 283(5409), 1908–
1911.
Paxinos, G., & Mai, J. (Eds.), (2004). The human nervous system (2nd ed.). San
Diego: Academic Press.
Pearson, H. (2006). Genetics: What is a gene? Nature, 441, 398–401.
Репа, M., Maki, A., Kovacic, D., et al. (2003). Sounds and silence: An optical
topography study of language recognition at birth. Proceedings of the National
Academy of Sciences of the United States of America, 100(20), 11702–11705.
Penfield, W., & Milner, B. (1958). Memory deficit produced by bilateral lesions in the
hippocampal zone. AMA Archives of Neurology and Psychiatry, 79(5), 475–497.
Penfield, W., & Roberts, L. (1959). Speech and brain mechanisms. Princeton:
Princeton University Press (URL:http://pup.princeton.edu).
Литература
417
Pepperberg, I. M . (2002). In search of King Solomon’s Ring: Cognitive and
communicative studies of grey parrots (Psittacus erithacus). Brain, Behavior and
Evolution, 59(1–2), 54–67.
Peretz, I., & Zatorre, R. J. (2005). Brain organization for music processing. Annual
Review of Psychology, 56, 89–114 .
Perky, C. W. (1910). An experimental study of imagination. The American Journal of
Psychology, 21(3), 422.
Pessoa, L., Kastner, S., & Ungerleider, L. G. (2003). Neuroimaging studies of
attention: From modulation of sensory processing to top-down control. The
Journal of Neuroscience, 23(10), 3990–3998 .
Petkov, С. I., Kayser, C., Augath, M., & Logothetis, N. K. (2006). Functional
imaging reveals numerous fields in the monkey auditory cortex. PLoS Biology,
4(7), E215.
Petrides, M. (1994). Frontal lobes and working memory: Evidence from
investigations of the effects of cortical excisions in nonhuman primates. Handbook
Neuropsychology, 9, 59–82.
Petrides, M. (1995). Impairments on nonspatial self-ordered and externally ordered
working memory tasks after lesions of the mid-dorsal part of the lateral frontal
cortex in the monkey. The Journal of Neuroscience, 15(1 Pt 1), 359–375.
Petrides, M., & Milner, B. (1982). Deficits on subject-ordered tasks after frontal- and
temporal-lobe lesions in man. Neuropsychologia, 20(3), 249–262.
Phelps, E. A ., & LeDoux, J. E . (2005). Contributions of the amygdala to emotion
processing: From animal models to human behavior. Neuron, 48(2), 175–187.
Piaget, J. (1937/1964). The construction of reality in the child. New York: Basic
Books.
Piaget, J. (1954). The construction of reality in the child (Cook, M. trans.) . New York:
Basic Books.
Piazza, M., Izard, V., Pinel, P., Le Bihan, D., & Dehaene, S. (2004). Tuning curves
for approximate numerosity in the human intraparietal sulcus. Neuron, 44(3),
547–555 .
Pinborg, L. H ., Adams, К. H ., Yndgaard, S., et al. (2004). [18f]altanserin binding
to human 5HT2a receptors is unaltered after citalopram and pindolol challenge.
Journal of Cerebral Blood Flow and Metabolism, 24(9), 1037–1045.
Pitman, R. K., Sanders, К. M., Zusman, R. M ., et al. (2002). Pilot study of secondary
prevention of posttraumatic stress disorder with propranolol. Biological
Psychiatry, 51(2), 189–192.
Plant, G. Т., Laxer, K. D., Barbara, N. M., Schiffman, J. S., & Nakayama, K. (1993).
Impaired visual motion perception in the contralateral hemifield following
unilateral posterior cerebral lesions in humans. Brain, 116(Pt 6), 1303–1335.
Plourde, G., Belin, P., Chartrand, D., et al. (2006). Cortical processing of complex
auditory stimuli during alterations of consciousness with the general anesthetic
propofol. Anesthesiology, 104(3), 448–457 .
Poeppel, D. (2001). Pure word deafness and the bilateral processing of speech code.
Cognitive Science, 25, 679–693 .
Poldrack, R. A ., Desmond, J. E ., Glover, G. H., & Gabrieli, J. D. (1998). The neural
basis of visual skill learning: An fMRI study of mirror reading. Cerebral Cortex,
8(1), 1–10.
418
Литература
Pollen, D. A . (1999). On the neural correlates of visual perception. Cerebral Cortex,
9(1), 4–19.
Polonsky, A., Blake, R., Braun, J., & Heeger, D. J. (2000). Neuronal activity in human
primary visual cortex correlates with perception during binocular rivalry. Nature
Neuroscience, 3(11), 1153–1159.
Ponton, C. W., Don, M., Eggermont, J. J., Waring, M. D., Kwong, В., & Masuda,
A. (1996a). Auditory system plasticity in children after long periods of complete
deafness. Neuroreport, 8(1), 61–65.
Ponton, C. W., Don, M., Eggermont, J. J., Waring, M. D., & Masuda, A. (1996b).
Maturation of human cortical auditory function: Differences between normal-
hearing children and children with cochlear implants. Ear and Hearing, 17(5),
430–137.
Portas, С. M., Krakow, K., Allen, P., Josephs, O., Armony, J. L., & Frith, C. D. (2000).
Auditory processing across the sleep-wake cycle: Simultaneous EEG and fMRI
monitoring in humans. Neuron, 28(3), 991–999 .
Posner, M. I. (1980). Orienting of attention. The Quarterly Journal of Experimental
Psychology, 32(1), 3–25 .
Posner, M. I . (2003). Attentional mechanisms. In M. Aminoff & R. Daroff (Eds.),
Encyclopedia of the neurological sciences. San Diego: Academic Press.
Posner, M. I., Cohen, Y., & Rafal, R. D . (1982). Neural systems control of spatial
orienting. Philosophical Transactions of the Royal Society of London. Series B,
Biological Sciences, 298(1089), 187–198.
Posner, M. I., & Desimone, R. (1998). Beyond images. Current Opinion in
Neurobiology, 8(2), 175–177 .
Posner, M. I., & Petersen, S. E . (1990). The attention system of the human brain.
Annual Review of Neuroscience, 13, 25–42 .
Posner, M. I., & Raichle, M. E . (1997). Images of mind. New York: Scientific
American Library/Scientific American Books.
Posner, M. I ., & Rothbart, M. K. (1998). Attention, self- regulation and consciousness.
Philosophical Transactions of the Royal Society of London. Series B, Biological
Sciences, 353(1377), 1915–1927.
Posner, M. I., & Rothbart, M. K . (2005). Influencing brain networks: Implications for
education. Trends in Cognitive Sciences, 9(3), 99–103.
Posner, M. I ., Walker, J. A., Friedrich, F. J ., & Rafal, R. D. (1984). Effects of parietal
injury on covert orienting of attention. The journal of Neuroscience, 4(7), 1863–1874.
Potter, R., Kopp, G., & Green, H. (1947). Visible speech. New York: Van Nostrand
Reinhold.
Pravdich-Neminsky, V. V. (1913). Ein versuch der Registrierung der Elektrischen
Gehirnerscheinungen. Zentral-blatt Physiology, 27, 951–960 .
Pulvermuller, F., Shtyrov, Y., Ilmoniemi, R. J., & Marslen-Wilson, W. D. (2006).
Tracking speech comprehension in space and time. Neuroimage, 31(3), 1297–
1305.
Quartz, S. R. (1999). The constructivist brain. Trends in Cognitive Sciences, 3(2),
48–57 .
Quinones-Hinojosa, A., Ojemann, S. G ., Sanai, N., Dillon, W. P., & Berger, M. S.
(2003). Preoperative correlation of intraoperative cortical mapping with magnetic
Литература
419
resonance imaging landmarks to predict localization of the Broca area. Journal of
Neurosurgery, 99(2), 311–318.
Quiroga, R. Q., Reddy, L., Kreiman, G., Koch, C., & Fried, I. (2005). Invariant visual
representation by single neurons in the human brain. Nature, 435(7045), 1102–
1107.
Raaijmakers, J. G ., & Shiffrin, R. M. (1992). Models for recall and recognition.
Annual Review of Psychology, 43, 205–234.
Raichle, M. E ., Fiez, J. A., Videen, Т. O ., et al. (1994). Practice-related changes in
human brain functional anatomy during nonmotor learning. Cerebral Cortex,
4(1), 8–26.
Raine, A., Buchsbaum, M., & LaCasse, L. (1997). Brain abnormalities in murderers
indicated by positron emission tomography. Biological Psychiatry, 42(6), 495–
508.
Raine, A., Lencz, Т., Bihrle, S., LaCasse, L., & Colletti, P. (2000). Reduced prefrontal
gray matter volume and reduced autonomic activity in antisocial personality
disorder discussion 128–129. Archives of General Psychiatry, 57(2), 119–127 .
Rajan, R., Irvine, D. R., Wise, L. Z ., & Heil, P. (1993). Effect of unilateral partial
cochlear lesions in adult cats on the representation of lesioned and unlesioned
cochleas in primary auditory cortex. The Journal of Comparative Neurology,
338(1), 17–49.
Rakic, P. (1988). Specification of cerebral cortical areas. Science, 241(4862), 170–
176.
Rakic, P. (1995). Corticogenesis in human and nonhuman primates. In M. S.
Gazzaniga (Ed.), The cognitive neurosciences. Cambridge: MIT Press.
Rakic, P. (2005). Less is more: Progenitor death and cortical size. Nature Neuroscience,
8(8), 981–982.
Ramachandran, V. S . (2002). Encyclopedia of the human brain. San Diego: Academic
Press.
Ramachandran, V. S., & Hirstein, W. (1998). The perception of phantom limbs. The
D. O . Hebb lecture. Brain, 121(Pt 9), 1603–1630.
Ramachandran, V. S., & Rogers-Ramachandran, D. (2000). Phantom limbs and
neural plasticity. Archives of Neurology, 57(3), 317–320.
Ramsey, T. Z ., & Overgaard, M. (2004). Introspection and subliminal perception.
Phenomenological Cognitive Science, 3(1), 1–23.
Ranganath, C. (in press). Binding items and contexts: The cognitive neuroscience of
episodic memory. Current Directions in Psychological Science.
Ranganath, C. (2006). Working memory for visual objects: Complementary roles of
inferior temporal, medial temporal, and prefrontal cortex. Neuroscience, 139(1),
277–289.
Ranganath, C., & D’Esposito, M. (2005). Directing the mind’s eye: Prefrontal,
inferior and medial temporal mechanisms for visual working memory. Current
Opinion in Neurobiology, 15(2), 175–182.
Rauch, S. L., Jenike, M. A., Alpert, N. M., et al. (1994). Regional cerebral blood flow
measured during symptom provocation in obsessive-compulsive disorder using
oxygen 15-labeled carbon dioxide and positron emission tomography. Archives of
General Psychiatry, 51(1), 62–70.
420
Литература
Ravizza, S. M ., Behrmann, M., & Fiez, J. A . (2005). Right parietal contributions
to verbal working memory: Spatial or executive? Neuropsychologica, 43(14),
2057–2067.
Reber, P. J., & Squire, L. R. (1998). Encapsulation of implicit and explicit memory in
sequence learning. Journal of Cognitive Neuroscience, 10(2), 248–263.
Redington, M., & Chater, N. (1997). Probabilistic and distributional approaches to
language acquisition. Trends in Cognitive Sciences, 1(7), 273–281.
Rees, G. (2001). Seeing is not perceiving. Nature Neuroscience, 4, 678–680 .
Rees, G., Kreiman, G., & Koch, C. (2002). Neural correlates of consciousness in
humans. Nature Reviews. Neuroscience, 3(4), 261–270.
Rees, G., & Lavie, N. (2001). What can functional imaging reveal about the role of
attention in visual awareness? Neuropsychologia, 39(12), 1343–1353.
Rees, G., Russell, C., Frith, C. D., & Driver, J. (1999). Inattentional blindness versus
inattentional amnesia for fixated but ignored words. Science, 286(5449), 2504–2507.
Reid, R. C. (2003). Vision. In L. R . Squire, F. E . Bloom, S. K . McConnell, J. L .
Roberts, N. C. Spitzer, & M. J . Zigmond (Eds.), Fundamental neuroscience (2nd
ed.) . San Diego: Elsevier.
Reilly, J., Losh, M., Bellugi, U., & Wulfeck, B. (2004). ‘Frog, where are you?’
narratives in children with specific language impairment, early focal brain injury,
and Williams syndrome. Brain and Language, 88(2), 229–247 .
Reneman, L., de Win, M. M., van den Brink, W., Booij, J., & den Heeten, G. J. (2006).
Neuroimaging findings with MDMA/ ecstasy: Technical aspects, conceptual
issues and future prospects. Journal of Psychopharmacology, 20(2), 164–175 .
Ress, D., & Heeger, D. J. (2003). Neuronal correlates of perception in early visual
cortex. Nature Neuroscience, 6(4), 414–420.
Ressler, K. J., Rothbaum, В. O., Tannenbaum, L., et al. (2004). Cognitive enhancers
as adjuncts to psychotherapy: Use of d-cycloserine in phobic individuals to
facilitate extinction of fear. Archives of General Psychiatry, 61(11), 1136–1144.
Reynolds, J. H., & Desimone, R. (2003). Interacting roles of attention and visual
salience in V4. Neuron, 37(5), 853–863.
Reynolds, J. H ., Gottlieb, J. P., & Kastner, S. (2003). Attention. In L. R. Squire,
F. E . Bloom, S. K. McConnell, J. L . Roberts, N. C. Spitzer, & M. J. Zigmond
(Eds.), Fundamental neuroscience (2nd ed.) . San Diego: Elsevier.
Ribeiro, S., Gervasoni, D., Soares., et al. (2004). Long-lasting novelty-induced
neuronal reverberation during slow- wave sleep in multiple forebrain areas. PLoS
Biology, 2(1).
Ridderinkhof, K. R., Span, M. M ., & van der Molen, M. W. (2002). Perseverative
behavior and adaptive control in older adults: Performance monitoring, rule
induction, and set shifting. Brain and Cognition, 49(3), 382–401.
Rizzolatti, G., Fadiga, L., Gallese, V., & Fogassi, L. (1996). Premotor cortex and the
recognition of motor actions. Cognitive Brain Research, 3, 131–141 .
Rizzolatti, G., Fogassi, L., & Gallese, V. (2002). Motor and cognitive functions of the
ventral premotor cortex. Current Opinion in Neurobiology, 12, 149–154 .
Ro, Т., Shelton, D., Lee, O. L., & Chang, E. (2004). Extrageniculate mediation of
unconscious vision in transcranial magnetic stimulation-induced blindsight.
Proceedings of the National Academy of Sciences of the United States of America,
101(26), 9933–9935.
Литература
421
Robinson, R. (2004). fMRI beyond the clinic: Will it ever be ready for prime time?
PLoS Biology, 2(6), el50.
Rodman, H. R., Gross, C. G., & Albright, T. D. (1989). Afferent basis of visual
response properties in area MT of the macaque. I. Effects of striate cortex removal.
The Journal of Neuroscience, 9(6), 2033–2050.
Rodman, H. R., Pessoa, L., & Ungerleider, L. (2003). Visual perception of objects. In
L. R. Squire, F. E . Bloom, S. K . McConnell, J. L . Roberts, N. C . Spitzer, & M. J.
Zigmond (Eds.), Fundamental neuroscience (2nd ed.) . San Diego: Elsevier.
Roediger, H. L., & McDermott, К. B . (1993). Implicit memory in normal human
subjects. Handbook Neuropsychology, 8, 63–131.
Rosch, E. (1975). Cognitive representations of semantic categories. Journal of
Experimental Psychology. General, 104(3), 192–233.
Rosen, S. D ., Paulesu, E., Nihoyannopoulos, P., et al. (1996). Silent ischemia as
a central problem: Regional brain activation compared in silent and painful
myocardial ischemia. Annals of Internal Medicine, 124(11), 939–949.
Rosenblatt, F. (1958). The perception: A probabilistic model for information storage
and organization in the brain. Psychological Review, 65(6), 386–408.
Rosenblatt, F. (1962). Principles of neurodynamics: Perceptrons and the theory of
brain mechanisms. Washington, DC: Spartan.
Rossi, S., Pasqualetti, P., Zito, G., et al. (2006). Prefrontal and parietal cortex in
human episodic memory: An interference study by repetitive transcranial magnetic
stimulation. The European Journal of Neuroscience, 23(3), 793–800 .
Roy, C. S., & Sherrington, C. S. (1890). On the regulation of the blood supply of the
brain. Journal of Physiology, 11, 85–108.
Rozin, P. (1976). The evolution of intelligence and access to the cognitive unconscious.
Progress in Psychobiology and Physiological Psychology, 6, 245–280.
Rubens, А. В., & Benson, D. F. (1971). Associative visual agnosia. Archives of
Neurology, 24(4), 305–316.
Ruby, P., & Decety, J. (2003). What you believe versus what you think they believe:
A neuroimaging study of conceptual perspective-taking. The European Journal of
Neuroscience, 17, 2475.
Rumelhart, D. E ., Hinton, G. E ., & Williams, R. J. (1986). Learning internal
representations by error propagation. In D. E . Rumelhart & J. L . McClelland
(Eds.), Parallel distributed processing: Explorations in the microstructure of
cognition, vol. 1: Foundations. Cambridge: MIT Press.
Rumelhart, D. E ., & McClelland, J. L . (Eds.), (1986a). Parallel distributed processing:
Explorations in the microstructure of cognition (Vol. 2). Cambridge: MIT Press.
Rumelhart, D. E., & McClelland, J. L. (Eds.), (1986b). Parallel distributed processing:
Foundations (Vol. 1: Foundations). Cambridge: MIT Press.
Rushworth, M. F., Nixon, P. D., Eacott, M. J., & Passingham, R. E . (1997).
Ventral prefrontal cortex is not essential for working memory. The Journal of
Neuroscience, 17(12), 4829–4838.
Rutkowski, R. G., & Weinberger, N. M. (2005). Encoding of learned importance
of sound by magnitude of representational area in primary auditory cortex.
Proceedings of the National Academy of Sciences of the United States of America,
102(38), 13664–13669.
422
Литература
Ryan, J. D., Althoff, R. R., Whitlow, S., & Cohen, N. J. (2000). Amnesia is a deficit
in relational memory. Psychological Science, 11(6), 454–461.
Ryle, G. (1949). The concept of mind. London: Hutchinson.
Rypma, В., Berger, J. S ., & D’Esposito, M. (2002). The influence of working-
memory demand and subject performance on prefrontal cortical activity. Journal
of Cognitive Science, 14(5), 721–731.
Sachs, J. D . S. (1967). Recognition memory for syntactic and semantic aspects of
connected discourse. Perception Psychophys, 2(9), 437–442 .
Sacks, O. (1984). A leg to stand on. New York: Simon & Schuster.
Sacks, O. (1985). The man who mistook his wife for a hat: And other clinical tales.
New York: Simon & Schuster.
Sacks, O. (1992). Tourette’s syndrome and creativity. British Medical Journal,
305(6868), 1151–1515 .
Saffran, J. R., Newport, E. L ., Aslin, R. N ., Tunick, R. A., & Barrueco, S. (1997).
Incidental language learning: Listening (and learning) out of the corner of your
ear. Psychological Science, 8(2), 101–105.
Salzman, C. D ., Britten, K. H ., & Newsome, W. T. (1990). Cortical microstimulation
influences perceptual judgements of motion direction. Nature, 346(6280), 174–177.
Sampaio, R. C., & Truwit, C. L . (2001). Myelination in the developing brain. In C.
A. L . Nelson (Ed.), The handbook of developmental cognitive neuroscience (pp.
35–15). Cambridge: MIT Press.
Sanes, D. H ., Reh, T. A ., & Harris, W. A . (2006). Development of the nervous system
(2nd ed.) . Amsterdam, Boston: Elsevier Academic Press.
Sasaki, Y., & Watanabe, T. (2004). The primary visual cortex fills in color. Proceedings
of the National Academy of Sciences of the United States of America, 101(52),
18251–18256.
Sato, K., Nariai, Т., Tanaka, Y., et al. (2005). Functional representation of the finger
and face in the human somatosensory cortex: Intraoperative intrinsic optical
imaging. Neuroimage, 25(4), 1292–1301.
Saunders, D. E . (2000). MR spectroscopy in stroke. British Medical Bulletin, 56(2),
334–345.
Saunders, N. R., Ek, C. J., Habgood, M. D., & Dziegielewska, К. M . (2008). Barriers
in the brain: A renaissance? Trends in Neurosciences, 31(6), 279–286.
Schacter, D. L . (1987). Implicit expressions of memory in organic amnesia: Learning
of new facts and associations. Human Neurobiology, 6(2), 107–118.
Schacter, D. L ., Dobbins, I. G ., & Schnyer, D. M. (2004). Specificity of priming: A
cognitive neuroscience perspective. Nature Reviews. Neuroscience, 5(11), 853–862.
Schacter, D. L ., Harbluk, J. L ., & McLachlan, D. R. (1984). Retrieval without
recollection: An experimental analysis of source amnesia. Journal of Verbal
Learning and Verbal Behavior, 23, 593–611.
Schacter, D. L ., & Tulving, E. (1994). Memory systems. Cambridge: MIT Press.
Scheibel, M. E ., & Scheibel, A. B. (1965). Activity cycles in neurons of the reticular
formation. Recent Advances in Biological Psychiatry, 8, 283–293.
Schirmer, A., & Kotz, S. A. (2006). Beyond the right hemisphere: Brain mechanisms
mediating vocal emotional processing. Trends Cign Sciences, 10(1), 24–30 .
Литература
423
Schneider, P., Scherg, M., Dosch, H. G ., Specht, H. J., Gutschalk, A., & Rupp, A.
(2002). Morphology of Heschl’s gyrus reflects enhanced activation in the auditory
cortex of musicians. Nature Neuroscience, 5(7), 688–694.
Schneider, W. (2009). Automaticity and consciousness. Elsevier Encyclopedia of
Consciousness. In W. Banks (Ed.), Encyclopedia of consciousness (1st ed) (pp.
83–92). Amsterdam: Academic Press.
Schneider, W. X . (1995). Vam: A neuro-cognitive model for visual attention control of
segmentation, object recognition and space-based motor action. Visual Cognition,
2, 331.
Schneider, W., & Chein, J. M . (2003). Controlled and automatic processing: Behavior,
theory, and biological mechanisms. Cognitive Sciences, 27, 525–559.
Scholz, J., Klein, M. C., Behrens, Т. E . J., & Johansen-Berg, H. (2009). Training
induces changes in white-matter architecture. Nature Neuroscience, 12, 1370–
1371.
Schore, A. (1999). Affect regulation and the origin of the self: The neurobiology of
emotional development. Hillsdale: Lawrence Erlbaum Associates.
Schultz, W. (1998). Predictive reward signal of dopamine neurons. Journal of
Neurophysiology, 80(1), 1–27 .
Schultz, W. (2002). Getting formal with dopamine and reward. Neuron, 36(2), 241–
263.
Schwarzlose, R. F., Baker, С. I ., & Kanwisher, N. (2005). Separate face and body
selectivity on the fusiform gyrus. Journal of Neuroscience, 25(47), 11055–11059.
Scott, S. K . (2005). Auditory processing - speech, space and auditory objects. Current
Opinion in Neurobiology, 15(2), 197–201.
Scott, S. K ., & Wise, R. J. (2004). The functional neuroanatomy of prelexical
processing in speech perception. Cognition, 92(1–2), 13–45 .
Scoville, W. В., & Milner, B. (1957). Loss of recent memory after bilateral
hippocampal lesions. Journal of Neurology, Neurosurgery, and Psychiatry, 20(1),
11–21 .
Seitz, A., & Watanabe, T. (2005). A unified model for perceptual learning. Trends in
Cognitive Science, 9(7), 329–334.
Serences, J. Т., & Yantis, S. (2006). Selective visual attention and perceptual
coherence. Trends in Cognitive Sciences, 10(1), 38–45 .
Seth, A. K ., Izhikevich, E„ Reeke, G. N., & Edelman, G. M . (2006). Theories and
measures of consciousness: An extended framework. Proceedings of the National
Academy of Sciences, 103(27), 10799–10804.
Shallice, T. (1982). Specific impairments of planning. Philosophical Transactions of
the Royal Society of London. Series B, Biological Sciences, 298(1089), 199–209.
Shallice, Т., & Warrington, E. K. (1970). Independent functioning of verbal memory
stores: A neuropsychological study. The Quarterly Journal of Experimental
Psychology, 22(2), 261–273.
Shamay-Tsoory, S., Tomer, R., Berger, В., Goldsher, D., & Aharon-Perestz, J.
(2005). Impaired ‘affective theory of mind’ is associated with right ventromedial
prefrontal damage. Cognitive and Behavioral Neurology, 18, 55–67.
Shanahan, M. P. (2005). Consciousness, emotion, and imagination: A brain-inspired
architecture for cognitive robotics. Proceedings AISB 2005 symposium on next
generation approaches to machine consciousness, 26–35.
424
Литература
Shannon, R. V., Zeng, F. G ., & Wygonski, J. (1998). Speech recognition with altered
spectral distribution of envelope cues. The Journal of the Acoustical Society of
America, 104(4), 2467–2476.
Shapiro, A. K., & Shapiro, E. (1974). Gilles de la Tourette’s syndrome. American
Family Physician, 9(6), 94–96 .
Sharma, R., & Sharma, A. (2004). Physiological basis and image processing in
functional magnetic resonance imaging: Neuronal and motor activity in brain.
Biomedical Engineering Online, 3(1), 13.
Shastri, L. (2002). Episodic memory and cortico-hippocampal interactions. Trends in
Cognitive Sciences, 6(4), 162–168.
Shatz, C. J. (2002). Emergence of order in visual system development. In M. H .
Johnson, Y. Munakata, & R. O . Gilmore (Eds.), Brain development and cognition:
A reader (2nd ed.) (pp. 231–244). Oxford: Blackwell, Oxford.
Shaw, P., Greenstein, D., Lerch, J., et al. (2006). Intellectual ability and cortical
development in children and adolescents. Nature, 440(7084), 676–679.
Sheinberg, D. L ., & Logothetis, N. K . (1997). The role of temporal cortical areas in
perceptual organization. Proceedings of the National Academy of Sciences of the
United States of America, 94(7), 3408–3413.
Shepard, R. C., & Cooper, L. (1982). Mental images and their transformations.
Cambridge: MIT Press.
Shepherd, G. M . (2004). In Paxinos, G. (Ed.), The human nervous system (2nd ed.).
San Diego: Academic Press.
Sherman, S. M ., & Guillery, R. W. (1998). On the actions that one nerve cell can
have on another: Distinguishing ‘drivers’ from ‘modulators’. Proceedings of the
National Academy of Sciences USA, 95(12), 7121–7126.
Sherrington, С. S . (1947). The integrative action of the nervous si/stem. Cambridge:
Cambridge University Press Quoted by Joseph Bogen http://www.its.caltech.
edu/~ jbogen/ text/mentali.htm.
Shevrin, H., & Dickman, S. (1980). The psychological unconscious: A necessary
assumption for all psychological theory. The American Psychologist, 35(5),
421–434.
Shiffrin, R. M ., Huber, D. E ., & Marinelli, K. (1995). Effects of category length
and strength on familiarity in recognition. Journal of Experimental Psychology.
Learning, Memory, and Cognition, 21(2), 267–287.
Shiffrin, R. M ., & Schneider, W. (1977). Controlled and automatic human information
processing: II. Perceptual learning, automatic attending and a general theory.
Psychological Review, 84(2), 127.
Shimamura, A. P., & Squire, L. R . (1984). Paired-associate learning and priming
effects in amnesia: A neuropsychological study. Journal of Experimental
Psychology. General, 113(4), 556–570.
Shipp, S. (2005). The importance of being agranular: A comparative account of visual
and motor cortex. Philosophical Transactions of the Royal Society of London,
Series B, Biological Sciences, 360(1456), 797–814.
Siapas, A. G ., Lubenov, E. V., & Wilson, M. A. (2005). Prefrontal phase locking to
hippocampal theta oscillations. Neuron, 46(1), 141–151 .
Silver, M. A., Ress, D., & Heeger, D. J. (2005). Topographic maps of visual spatial
attention in human parietal cortex. Journal of Neurophysiology, 94(2), 1358–1371.
Литература
425
Simon, H. A ., & Chase, W. G. (1973). Skill in chess. American Scientist, 61, 393–403.
Simon, H. A ., & Gilmartin, K. J. (1973). A simulation of memory for chess positions.
Cognitive Psychology, 5, 29–16.
Simons, D. J. (2000). Attentional capture and inattentional blindness. Trends in
Cognitive Sciences, 4(4), 147–155 .
Simons, J. S., & Spiers, H. J. (2003). Prefrontal and medial temporal lobe interactions
in long-term memory, Nature Reviews. Neuroscience, 4(8), 637–648.
Simpson, J. A ., & Weiner, E. S. C. (1989). The Oxford English Dictionary (2nd ed).
Oxford, New York: Oxford University Press/Clarendon Press.
Singer, J. L. (1993). Experimental studies of ongoing conscious experience discussion
116–122 . Ciba Foundation Symposium, 174, 100–116.
Singer, W. (1994). A new job for the thalamus. Nature, 369, 444–445 .
Singh, K. D ., Smith, А. Т., & Greenless, M. W. (2000). Spatiotemporal frequency
and direction of sensitivities of human visual areas measured using fMRI.
Neuroimage, 12(5), 550–564.
Smith, A. M., Fried, P. A., Hogan, M. J., & Cameron, I. (2006). Effects of prenatal
marijuana on visuospatial working memory: An fMRI study in young adults.
Neurotoxicology and Teratology, 28(2), 286–295.
Smith, E. E ., & Jonides, J. (1997). Working memory: A view from neuroimaging.
Cognitive Psychology, 33(1), 5–12 .
Smith, E. E., & Jonides, J. (1998). Neuroimaging analysis of human working memory.
Proceedings of the National Academy of Sciences of the United States of America,
95(20), 12061–12068.
Smith, E. E., Jonides, J., & Koeppe, R. A. (1996). Dissociating verbal and spatial
working memory using PET. Cerebral Cortex, 6(1), 11–20.
Smith, M. L ., & Milner, B. (1984). Differential effects of frontal-lobe lesions on
cognitive estimation and spatial memory. Neuropsychologia, 22(6), 697–705.
Snodgrass, J. G ., & Vanderwart, M. (1980). A standardized set of 260 pictures:
Norms for name agreement, image agreement, familiarity, and visual complexity.
Journal of Experimental Psychology. [Human Learning], 6(2), 174–215 .
Snowden, J. S., Griffiths, H. L ., & Neary, D. (1996). Semantic-episodic memory
interactions in semantic dementia: Implications for retrograde memory function.
Cognitive Neuropsychology, 13(8), 1101–1139.
Sokoloff, L., Reivich, M., Kennedy, C., et al. (1977). The [14C]deoxyglucose method
for the measurement of local cerebral glucose utilization: Theory, procedure,
and normal values in the conscious and anesthetized albino rat. Journal of
Neurochemistry, 28(5), 897–916.
Soros, P., Sokoloff, L. G., Bose, A., Mcintosh, A. R., Graham, S. J., & Stuss, D. T.
(2006). Clustered functional MRI of overt speech production. Neuroimage, 32(1),
376–387.
Sowell, E. R., Peterson, B. S., Thompson, P. M., Welcome, S. E., Henkenius, A. L., &
Toga, A. W. (2003). Mapping cortical change across the human life span. Nature
Neuroscience, 6(3), 309–315.
Sowell, E. R., Thompson, P. M ., Peterson, B. S., et al. (2002). Mapping cortical gray
matter asymmetry patterns in adolescents with heavy prenatal alcohol exposure.
Neuroimage, 17(4), 1807–1819.
426
Литература
Spanagel, R., & Weiss, F. (1999). The dopamine hypothesis of reward: Past and
current status. Trends in Neurosciences, 22(11), 521–527 .
Sporns, O., & Tononi, G. (2002). Classes of network connectivity and dynamics.
Complexity, 7, 28–38 .
Squire, L. R . (1992). Declarative and nondeclarative memory: Multiple brain systems
supporting learning and memory. Journal of Cognitive Neuroscience, 4(3), 232–243.
Squire, L. R. (2004). Memory systems of the brain: A brief history and current
perspective. Neurobiology of Learning and Memory, 82, 171–177 .
Squire, L. R., & Alvarez, P. (1995). Retrograde amnesia and memory consolidation:
A neurobiological perspective. Current Opinion in Neurobiology, 5(2), 169–177 .
Squire, L. R., Berg, D., Bloom, F. E., du Lac, S., & Ghosh, A. (Eds.), (2008).
Fundamental neuroscience (3rd ed.) . San Diego: Elsevier.
Srinivasan, R., Russell, D. P., Edelman, G. M ., & Tononi, G. (1999). Increased
synchronization of neuromagnetic responses during conscious perception. The
Journal of Neuroscience, 19(13), 5435–5448.
Stahnisch, F. W., & Nitsch, R. (2002). Santiago Ramon у Cajal’s concept of neuronal
plasticity: The ambiguity lives on. Trends in Neurosciences, 25(11), 589–591.
Standage, D. I ., Trappenberg, T. P., & Klein, R. M . (2005). Modelling divided visual
attention with a winner-take-all network. Neural Networks, 18(5–6), 620–627.
Standring, S. (Ed.). (2005). Gray’s anatomy: The anatomical basis of clinical practice
(39th ed.) . Edinburgh: Churchill Livingstone.
Stark, С. E . L., & Okado, Y. (2003). Making memories without trying: Medial
temporal lobe activity associated with incidental memory formation during
recognition. The Journal of Neuroscience, 23(17), 6748–6753.
Stephan, К. M ., Thaut, M. H„ Wunderlich, G., et al. (2002). Conscious and
subconscious sensorimotor synchronization — prefrontal cortex and the influence
of awareness. Neuroimage, 15(2), 345–352.
Steriade, M. (2006). Grouping of brain rhythms in corticothalamic systems.
Neuroscience, 137(4), 1087–1106.
Sternberg, R. J., & Ben-Zeev, T. (2001). Complex cognition: The psychology of
human thought. Oxford: Oxford University Press.
Stewart, F., Parkin, A. J., & Hunkin, N. M . (1992). Naming impairments following
recovery from herpes simplex encephalitis: Category-specific? The Quarterly
Journal of Experimental Psychology. A, 44(2), 261–284.
Stickgold, R„ Hobson, J. A ., Fosse, R., & Fosse, M. (2001). Sleep, learning, and
dreams: Off-line memory reprocessing. Science, 294(5544), 1052–1057.
Stiles, J., Bates, E. A., Thai, D., Trauner, D. A., & Reilly, J. (2002). Linguistic and
spatial cognitive development in children with pre- and perinatal focal, brain
injury: A ten-year overview from the San Diego Longitudinal Project. In M. H.
Johnson, Y. Munakata, & R. O . Gilmore (Eds.) . Brain development and cognition:
A reader (2nd ed.) (pp. 272 –291). Oxford: Blackwell Publishing.
Stiles, J., Reilly, J., Paul, В., & Moses, P. (2005). Cognitive development following
early brain injury: Evidence for neural adaptation. Trends in Cognitive Sciences,
9(3), 136–143.
Stoerig, P., Zontanou, A., & Cowey, A. (2002). Aware or unaware: Assessment of
cortical blindness in four men and a monkey. Cerebral Cortex, 12(6), 565–574 .
Литература
427
Strauss, G. I., Hogh, P., Moller, K., Knudsen, G. M., Hansen, B. A., & Larsen, F. S.
(1999). Regional cerebral blood flow during mechanical hyperventilation in
patients with fulminant hepatic failure. Hepatology, 30(6), 1368–1373.
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. Journal of
Experimental Psychology, 18, 643–662.
Stuss, D. Т., Floden, D., Alexander, M. P., Levine, В., & Katz, D. (2001). Stroop
performance in focal lesion patients: Dissociation of processes and frontal lobe
lesion location. Neuropsychologia, 39(8), 771–786.
Sussman, E. S . (2005). Integration and segregation in auditory scene analysis. The
Journal of the Acoustical Society of America, 117(3 Pt 1), 1285–1298.
Sutherland, G. R., & McNaughton, B. (2000). Memory trace reactivation in
hippocampal and neocortical neuronal ensembles. Current Opinion in
Neurobiology, 10(2), 180–186.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic atlas of the human brain:
3-dimensional proportional system: An approach to cerebral imaging. Stuttgart:
Thieme Medical Publishers.
Tallal, P. (2004). Improving language and literacy is a matter of time. Nature Reviews.
Neuroscience, 5(9), 721–728.
Tammer, R., Ehrenreich, L., Boretius, S., Watanabe, Т., Frahm, J., & Michaelis, T.
(2006). Compatibility of glass- guided recording microelectrodes in the brain
stem of squirrel monkeys with high-resolution 3D MRI. Journal of Neuroscience
Methods, 153(2), 221–229.
Tanaka, K. (1996). Inferotemporal cortex and object vision. Annual Review of
Neuroscience, 19, 109–139.
Tansey, E. M . (2006). Pavlov at home and abroad: His role in international physiology.
Autonomic Neuroscience: Basic and Clinical, 125, 1–11 .
Taylor, F. В., Lowe, K., Thompson, C., et al. (2006). Daytime prazosin reduces
psychological distress to trauma specific cues in civilian trauma posttraumatic
stress disorder. Biological Psychiatry, 59(7), 577–581.
Teyler, T. J., & DiScenna, P. (1986). The hippocampal memory indexing theory.
Behavioral Neuroscience, 100(2), 147–154 .
Thatcher, R. W. (1992). Cyclic cortical reorganization during early childhood. Brain
and Cognition, 20(1), 24–50.
Thier, P., Haarmeier, Т., & Ignashchenkova, A. (2002). The functional architecture of
attention. Current Biology, 12(5), R158–R162.
Thottakara, P., Lazar, M., Johnson, S. C., & Alexander, A. L. (2006). Application
of Brodmann’s area templates for ROI selection in white matter tractography
studies. Neuroimage, 29(3), 868–878.
Tiitinen, H., Salminen, N. H., Palomaki, K. J., Makinen, V. Т., Alku, P., & May, P.
J. (2006). Neuromagnetic recordings reveal the temporal dynamics of auditory
spatial processing in the human cortex. Neuroscience Letters, 396(1), 17–22 .
Toga, A. W., Thompson, P. M., & Sowell, E. R. (2006). Mapping brain maturation.
Trends in Neurosciences, 29(3), 148–159.
Tong, F. (2003). Primary visual cortex and visual awareness. Nature Reviews
Neuroscience, 4(3), 219–229.
Tong, F., & Engel, S. A . (2001). Interocular rivalry revealed in the human cortical
blind-spot representation. Nature, 412(6834), 195–199.
428
Литература
Tong, F., Nakayama, K., Moscovitch, M., Weinrib, O., & Kanwisher, N. (2000).
Response properties of the human fusiform face area. Cognitive Neuropsychology,
17, 257–279.
Tong, F., Nakayama, K., Vaughan, J. Т., & Kanwisher, N. (1998). Binocular rivalry
and visual awareness in human extrastriate cortex. Neuron, 21(4), 753–759.
Tononi, G. (2004). An information integration theory of consciousness. BMC
Neuroscience, 5, 42.
Tononi, G., Srinivasan, R., Russell, D. P., & Edelman, G. M . (1998). Investigating
neural correlates of conscious perception by frequency-tagged neuromagnetic
responses. Proceedings of the National Academy of Sciences of the United States
of America, 95(6), 3198–3203.
Tootell, R. В., Dale, A. M., Sereno, M. I., & Malach, R. (1996). New images from
human visual cortex. Trends in Neurosciences, 19(11), 481–189.
Tourette, G. (1885). Etude sur une affection nerveuse caracterisee par de
l’incorrdination motrice accompagne d’ echolalie et de copralalie. Archives of
Neurology, 9, 158–200.
Treisman, A. M ., & Gelade, G. (1980). A feature-integration theory of attention.
Cognitive Psychology, 12(1), 97–136.
Tsao, D. Y., Freiwald, W. A ., Tootell, R. В., & Livingstone, M. S. (2006). A cortical
region consisting entirely of face-selective cells. Science, 311(5761), 670–674.
Tulving, E. (1972). Episodic and semantic memory. In E. Tulving, W. Donaldson, &
G. H. Bower (Eds.), Organization of memory (pp. 381–403). New York: Academic
Press.
Tulving, E. (1985). How many memory systems are there. The American Psychologist,
40(4), 385–398 .
Tulving, E. (2002). Episodic memory: From mind to brain. Annual Review of
Psychology, 53, 1–25 .
Tyler, L. K., & Moss, H. E . (2001). Towards a distributed account of conceptual
knowledge. Trends in Cognitive Sciences, 5(6), 244–252.
Tzourio, N., Massioui, F. E ., Crivello, F., Joliot, M., Renault, В., & Mazoyer, B.
(1997). Functional anatomy of human auditory attention studied with pet.
Neuroimage, 5(1), 63–77.
Tzourio-Mazoyer, N., De Schonen, S., Crivello, F., Reutter, В., Aujard, Y., &
Mazoyer, B. (2002). Neural correlates of woman face processing by 2-month-old
infants. Neuroimage, 15(2), 454–461.
Uncapher, M. R., & Rugg, M. D. (2009). Selecting for memory? The influence of
selective attention on the mnemonic binding of contextual information. Journal
of Neuroscience, 29(25), 8270–8279.
Ungerleider, L. G ., & Mishkin, M. (1982). Two cortical visual systems. In M. A.
Ingle, M. A . Goodale, & J. W. Mansfield (Eds.), Analysis of visual behavior.
Cambridge: The MIT Press.
Ungless, M. A . (2004). Dopamine: The salient issue. Trends in Neurosciences,
27(12), 702–706.
Unterrainer, J. M ., & Owen, A. M. (2006). Planning and problem solving: From
neuropsychology to functional neuroimaging. Journal of Physiology Paris, 99(4–
6), 308–317.
Литература
429
Vakil, E., Kahan, S., Huberman, M., & Osman, A. (2000). Motor and non-motor
sequence learning in patients with basal ganglia lesions: The case of serial reaction
time (SRT). Neuropsychologia, 38(1), 1–10.
Valet, M., Sprenger, Т., Boeck, H., et al. (2004). Distraction modulates connectivity
of the cingulo-frontal cortex and the midbrain during pain — an fMRI analysis.
Pain, 109, 399–408.
Van Essen, D. C . (2005). A population-average, landmark- and surface-based (PALS)
atlas of human cerebral cortex. Neuroimage, 28(3), 635–662.
VanRullen, R., & Koch, C. (2003). Is perception discrete or continuous? Trends in
Cognitive Sciences, 7(5), 207–213.
Varela, F., et al. (2001). The brainwave: Phase synchronization and large-scale
integration. Nature Neuroscience, 2, 229–238.
Vargha-Khadem, F., Gadian, D. G ., Copp, A., & Mishkin, M. (2005). Foxp2 and
the neuroanatomy of speech and language. Nature Reviews. Neuroscience, 6(2),
131–138.
Vargha-Khadem, F., & Mishkin, M. (1997). Speech and language outcome after
hemispherectomy in childhood. Paediatrica Epilepsy Syn Surgical Treatment,
774–784.
Vigneau, M., Beaucousin, V., Herve, P. Y., et al. (2006). Metaanalyzing left hemisphere
language areas: Phonology, semantics, and sentence processing. Neuroimage,
30(4), 1414–1432.
Vogeley, K., Kurthen, M., Falkai, P., & Maier, W. (1999). Essential functions of the
human self model are implemented in the prefrontal cortex. Consciousness and
Cognition, 8(3), 343–363 .
Volavka, J., Mohammad, Y., Vitrai, J., Connolly, M., Stefanovic, M., & Ford, M.
(1995). Characteristics of state hospital patients arrested offenses committed
during hospitalization. Psychiatric Services, 46(8), 796–800 .
Volpe, J. J. (1987). Brain death determination in the newborn. Pediatrics, 80(2),
293–297.
Vuilleumier, P. (2005). How brains beware: Neural mechanisms of emotional
attention. Trends in Cognitive Sciences, 9(12), 585–594.
Vuilleumier, P., Armony, J. L ., Clarke, K., Husain, M., Driver, J., & Dolan, R. J.
(2002). Neural response to emotional faces with and without awareness: Event-
related fMRI in a parietal patient with visual extinction and spatial neglect.
Neuropsychologia, 40(12), 2156–2166.
Vuilleumier, P., Armony, J. L ., Driver, J., & Dolan, R. J. (2001). Effects of attention
and emotion on face processing in the human brain: An event-related fMRI study.
Neuron, 30(3), 829–841.
Vuilleumier, P., Armony, J. L ., Driver, J., & Dolan, R. J . (2003). Distinct spatial
frequency sensitivities for processing faces and emotional expressions. Nature
Neuroscience, 6(6), 624–631.
Walsh, V., & Cowey, A. (2000). Transcranial magnetic stimulation and cognitive
neuroscience. Nature Reviews. Neuroscience, 1(1), 73–79.
Wang, X-J. (2001). Synaptic reverberation underlying mnemonic persistent activity.
Trends in Neurosciences, 24(8), 455–463.
Ward, L. M. (2003). Synchronous neural oscillations and cognitive processes. Trends
in Cognitive Sciences, 7(12), 553–559.
430
Литература
Wearing, D. (2005). Forever today: A true story of lost memory and never-ending
love. London: Corgi Books.
Weems, S. A ., & Reggia, J. A . (2006). Simulating single word processing in the
classic aphasia syndromes based on the Wemicke-Lichtheim-Geschwind theory.
Brain and Language, 98(3), 291–309 .
Weiler, I. J., Hawrylak, N., & Greenough, W. T. (1995). Morphogenesis in memory
formation: Synaptic and cellular mechanisms. Behavioural Brain Research,
66(1–2), 1–6 .
Weiskrantz, L. (1986). Blindsight: A case study and implications. Oxford: Oxford
University Press.
Weiskrantz, L., Warrington, E. K ., Sanders, M. D., & Marshall, J. (1974). Visual
capacity in the hemianopic field following a restricted occipital ablation. Brain,
97(4), 709–728.
Welsh, M. C., Pennington, B. F., Ozonoff, S., Rouse, В., & McCabe, E. R. (1990).
Neuropsychology of early-treated phenylketonuria: Specific executive function
deficits. Child Development, 61(6), 1697–1713.
Werbos, P. (1974). Beyond regression: New tools for prediction and analysis in the
behavioral sciences. Cambridge: Harvard University.
Werker, J. F., & Vouloumanos, A. (2001). Speech and language processing in infancy:
A neurocognitive approach. In C. A. L. Nelson (Ed.), Handbook of developmental
cognitive neuroscience (2nd ed) (pp. 269–280). Cambridge: MIT Press.
Wernicke, C. (1874). Der Aphasische Symptomencomplex: Eine Psychologische
Studie auf Anatomischer Basis. Breslau: Cohn and Weigert.
Wertheimer, M. (1912). Experimentelle Studien iiber Sehen von Bewegung. Zeits
Psychology, 61, 161–265.
Wessinger, С. M ., VanMeter, J., Tian, В., Van Lare, J., Pekar, J., & Rauschecker,
J. P. (2001). Hierarchical organization of the human auditory cortex revealed
by functional magnetic resonance imaging. Journal of Cognitive Neuroscience,
13(1), 1–7 .
Westmacott, R., Black, S. E ., Freedman, M., & Moscovitch, M. The contribution of
autobiographical significance to semantic memory: Evidence from Alzheimer’s
disease, semantic dementia, and amnesia. Neuropsychologia, 42(1), 25–48.
Wexler, M. (2001). Two new face illusions. Trends in Cognitive Sciences, 5(4), 137.
Whalen, P. J., Kagan, J., Cook, R. G., et al. (2004). Human amygdala responsivity to
masked fearful eye whites. Science, 306(5704), 2061.
Wheeler, M. E ., & Buckner, R. L . (2004). Functional-anatomic correlates of
remembering and knowing. Neuroimage, 21, 1337–1349.
White, J. A. (2002). Action potentials. In V. S. Ramachandran (Ed.), Encyclopedia of
the human brain. San Diego: Academic Press.
Wicker, В., Keysers, C., Plailly, J., Royet, J., Gallese, V., & Rizzolatti, G. (2003a).
Both of us disgusted in my insula, the common neural basis of seeing and feeling
disgust. Neuron, 40(3), 655–664.
Wicker, В., Ruby, P., Royet, J. P., & Fonlupt, P. (2003b). A relation between rest and
the self in the brain? Brain Research. Brain Research Reviews, 43(2), 224–230.
Wigg, C. L ., & Martin, A. (1998). Properties and mechanisms of perceptual priming.
Current Opinion in Neurobiology, 8, 227–233.
Литература
431
Williams, J., Waiter, G., Perra, O., Perrett, D., & Whiten, A. An fMRI study of joint
attention experience. Neuroimage, 25, 133–140.
Willingham, D. B. (1998). A neuropsychological theory of motor skill learning.
Psychological Review, 105(3), 558–584.
Willingham, D. В., Nissen, M. J., & Bullemer, P. (1989). On the development of
procedural knowledge. Journal of Experimental Psychology. Learning, Memory,
and Cognition, 15(6), 1047–1060.
Wilson, B. A ., Baddeley, A. D ., & Kapur, N. (1995). Dense amnesia in a professional
musician following herpes simplex virus encephalitis. Journal of Clinical and
Experimental Neuropsychology, 17(5), 668–681.
Wise, R. A . (2002). Brain reward circuitry: Insights from unsensed incentives.
Neuron, 36(2), 229–240.
Witte, E. A., & Marrocco, R. T. (1997). Alteration of brain noradrenergic
activity in rhesus monkeys affects the alerting component of covert orienting.
Psychopharmacology (Berl), 132(4), 315–323.
Woldorff, M. G ., Tempelmann, C., Fell, J., et al. (1999). Lateralized auditory
spatial perception and the contralaterality of cortical processing as studied with
functional magnetic resonance imaging and magnetoencephalography. Human
Brain Mapping, 7(1), 49–66 .
Wolfe, J. M . (2003). Moving towards solutions to some enduring controversies in
visual search. Trends in Cognitive Sciences, 7(2), 70–76.
Wunderlich, K., Schneider, K. A ., & Kastner, S. (2005). Neural correlates of
binocular rivalry in the human lateral geniculate nucleus. Nature Neuroscience,
8(11), 1595–1602.
Wyvell, C. L ., & Berridge, К. C. (2000). Intra-accumbens amphetamine increases
the conditioned incentive salience of sucrose reward: Enhancement of reward
‘wanting’ without enhanced ‘liking’ or response reinforcement. The Journal of
Neuroscience, 20(21), 8122–8130.
Yakovlev, P. I., & Lecours, A. R. (1967). The myelogenetic cycles of regional
maturation of the brain. In A. Minokowski (Ed.), Regional development of the
brain in early life (pp. 3 –70). Philadelphia: Davis.
Yang, Т. Т., Gallen, С. C., Ramachandran, V. S., Cobb, S., Schwartz, B. J., &
Bloom, F. E . (1994). Noninvasive detection of cerebral plasticity in adult human
somatosensory cortex. Neuroreport, 5(6), 701–704.
Yantis, S. (2008). The Neural basis of selective attention. Current Directions in
Psychological Science, 17(2), 86–90 .
Yarbus, A. F. (1967). Eye movements and vision. New York: Plenum Press.
Yin, H. H ., & Knowlton, B. J. (2006). The role of the basal ganglia in habit formation.
Nature Reviews. Neuroscience, 7(6), 464–476.
Yonelinas, A. P. (2002). The nature of recollection and familiarity: A review of 30
years of research. Journal of Memory and Language, 46(3), 441–517 .
Yonelinas, A. P., Otten, L. J., Shaw, K. N ., & Rugg, M. D . Separating the brain regions
involved in recollection and familiarity in recognition memory. The Journal of
Neuroscience, 25(11), 3002–3008 .
Yzerbyt, V. Y., Lories, G., & Dardenne, B. (Eds.), (1998). Metacognition: Cognitive
and social dimensions. Thousand Oaks: Sage Publications.
432
Литература
Zaidel, E., Kasher, A., Soroker, N., Batori, G., Giora, R., & Graves, D. (2000).
Hemispheric contributions to pragmatics. Brain and Cognition, 43(1–3), 438–
443.
Zaidel, E., Zaidel, D. W., & Bogen, J. E . (2002). The split brain. In V. S . Ramachandran
(Ed.), Encyclopedia of the human brain. San Diego: Academic Press.
Zapala, M. A., Hovatta, I., Ellison, J. A., et al. (2005). Adult mouse brain gene
expression patterns bear an embryologic imprint. Reprinted in Zapala, M. A.,
Barlow, C., Hovattaa, I. (2009). Molecular Anatomy of the Mammalian brain.
Elsevier Encyclopedia of Neuroscience, 2009. Proceedings of the National
Academy of Sciences of the United States of America, 19, 10357–10362.
Zatorre, R. J. (2002). Auditory cortex. In V. S. Ramachandran (Ed.), Encyclopedia of
the human brain. San Diego: Academic Press.
Zatorre, R. J., & Halpern, A. R. (2005). Mental concerts: Musical imagery and
auditory cortex. Neuron, 47(1), 9–12 .
Zeki, S. (2003). Improbable areas in the visual brain. Trends in Neurosciences, 26(1),
23–26.
Zeki, S. M . (1974). Functional organization of a visual area in the posterior bank of
the superior temporal sulcus of the rhesus monkey. Journal of Physiology, 236(3),
549–573.
Zeki, S. M . (1977). Colour coding in the superior temporal sulcus of rhesus monkey
visual cortex. Proceedings of the Royal Society of London. Series B, Biological
Sciences, 197(1127), 195–223.
Zhao, F., Wang, P., Hendrich, K., Ugurbil, K., & Kim, S. G . .Cortical layer-dependent
BOLD and CBV responses measured by spin-echo and gradient-echo fMRI:
Insights into hemodynamic regulation. Neuroimage, 30(4), 1149–1160.
Zheng, X., & Rajapakse, J. C. (2006). Learning functional structure from FMR
images. Neuroimage, 31(4), 1601–1613.
L-ДОФА (L — левовращающая молекула; ДОФА — дигидроксифенилаланин) —
первый использовавшийся в медицине агонист дофаминовых рецепторов для лече-
ния пациентов с болезнью Паркинсона, направленный на стимуляцию синтеза до-
фамина в черной субстанции. L-ДОФА является естественным предшественником
в синтезе дофамина
автоматизм процессов — высокотренированные навыки или привычки, которые
осуществляются с минимальным произвольным усилием и вовлечением сознания.
См. гл. 3, «Рабочие гипотезы»
автономная нервная система (АНС) — подразделение периферической нервной
системы, которая ответственна за поддержание гомеостаза и регуляции отдыха и
активности. Физиологические процессы, контролируемые АНС, например кровя-
ное давление и потоотделение, в основном непроизвольные и неосознанные. См.
гл. 5, «От вопроса «где?» до вопроса «что?»: функциональная роль областей мозга»
агонист (от лат. соперник) — молекула, которая связывается с рецептором мембра-
ны нейрона, повышая тем самым вероятность разряда постсинаптического нейро-
на. См. гл . 16, «Запоминание слов»
аденозин — тормозный нейромедиатор, который обеспечивает сон и подавляет
пробуждение и бодрствование. См. гл. 16
адреналин (от лат. ad-
—
сверху; renes — почка; -in — белок) — или эпинефрин,
является периферическим гормоном и мозговым нейромедиатором. Как циркуля-
торный гормон, он выделяется надпочечниками, расположенными на верхней по-
верхности почек, и готовит организм к реакции борьбы или бегства. См. гл . 16
аксон — длинный, тонкий отросток нервной клетки (нейрона), который проводит
электрические импульсы от тела клетки. См. гл . 1, «Некоторые исходные положе-
ния», и гл. 3, «Введение»
активность, зависящая от уровня кислорода в крови (BOLD) — физический
сигнал магнитной природы, который отражает поступление кислорода к опреде-
ленным областям мозга. Активность BOLD является физической основой для
функциональной магнитно-резонансной томографии. См. гл . 4
акустический анализ (от греч. akoustikos — для слушания) — процесс интерпре-
тации физической звуковой энергии, как лингвистической и музыкальной, так и
звуков окружающей среды (например, хлопанье дверью или звуки заведенной ма-
шины). См. гл . 11, «Введение»
альфа-волны — регулярные электромагнитные волны, регистрируемые в мозге
или на поверхности головы и отражающие активность большой популяции нейро-
Глоссарий
434
Глоссарий
нов. Альфа-волны имеют частоту от 7,5 до 13 Гц и возникают преимущественно в
затылочной области при расслабленном бодрствовании с закрытыми глазами. На-
против, при открывании глаз, а также в состояниях дремоты и сна, альфа-волны
уменьшаются. См. гл. 4, «Ряд полезных инструментов»
аминокислота — составляющая часть молекулы белка. Аминокислоты жизненно
необходимы и выполняют множество метаболических функций. Все аминокислоты
содержат как NH3 (амино), так и COOH (карбоксильную) группы. См. гл . 16
амнезия (от греч. a -mnēsia — не память) — потеря памяти. Есть два типа амнезии:
антероградная (потеря памяти о событиях после травмы) и ретроградная (потеря
памяти о событиях до травмы). См. гл . 9, «Амнезия»
анализ слуховых образов — процесс, благодаря которому слуховая система рас-
щепляет и организует звуки окружающей среды. См. гл . 7, «Введение»
анализ Фурье — назван в честь французского математика и физика Жозефа Фурье,
который показал, что любую сложную функцию можно разложить в конечную по-
следовательность синусоидальных функций. В музыке, например, это значит, что
любой сложный звук может быть разложен в ряд простых тонов (синусоидальных
волн). Анализ Фурье часто применяют для разложения ЭЭГ-записей и других сиг-
налов мозга по частотным диапазонам. См. гл . 4, «Ряд полезных инструментов»
антагонист (от лат. соперник) — молекула, которая связывается с рецептором
и блокирует или подавляет его активность. См. агонист
антероградная амнезия (от нового лат. antero-
—
вперед; и греч. a -mnēsia – не па-
мять) — тип амнезии, при которой события, происходящие после мозговой травмы,
не кодируются в долговременную память, хотя сохраняются воспоминания о собы-
тиях до травмы. См. гл . 9, «Амнезия». См. ретроградная амнезия
ассоциативный процесс (от лат. ad- + sociare — соединять) — процесс, в котором
один или более сенсорных и/или ответных событий связываются вместе в мозге.
См. гл . 5, «От вопоса «где?» до вопроса «что?»: функциональная роль областей
мозга»
аутистические расстройства — ряд состояний, характеризующихся нарушением
в социальном восприятии и общении, циклическими действиями и другими сим-
птомами
афазия (от греч. а — без; от лат. phasia — речь) — потеря речевой функции вслед-
ствие мозговой травмы, например повреждение зоны Брока, ответственной за про-
изводство речи, или зоны Вернике, ответственной за понимание речи. См. гл. 1,
«Немного истории и продолжающаяся дискуссия», и гл. 7, «Восприятие речи»
афазия Вернике (от греч. a — без; от лат. phasia — речь) (см. Вернике, Карл. См.
гл. 1, «Немного истории и продолжающаяся дискуссия»)
аффективное слепозрение (от лат. affectus — эмоция) — неспособность различать
эмоциональные выражения лиц. См. гл . 13, «Система страха»
ацетилхолин — нейромедиатор в центральной и периферической нервной систе-
ме. См. гл. 16
ацетилхолинэстераза — фермент, который разрушает ацетилхолин для освобож-
дения синапса
Глоссарий
435
базальные ядра (от лат. basis — основа; от лат. набухание или разрастание) —
большое скопление подкорковых структур рядом с таламусом, ответственное за
двигательный контроль, автоматизм, сознание, эмоции и обучение. См. гл . 5, «Раз-
витие мозга от основания до вершины»
Барон-Коэн, Саймон (род. 1958) — исследователь аутизма, предположил, что у
детей формируется теория разума, состоящая из четырех навыков: определение на-
мерений других, определение направления взора, разделенное внимание с другими
и имплицитное знание о других людях. См. гл. 14, «Заключение»
белое вещество — в мозге белое вещество состоит из компактных пучков миели-
низированных аксонов, которые соединяют множество областей серого вещества
друг с другом. Белое вещество названо так за свой вид или большое скопление ми-
елинизированных аксонов нейронов, которые образуют видимую оболочку мозго-
вых структур. См. гл . 1, «Немного истории и продолжающаяся дискуссия»
бессознательное восприятие — обработка сенсорного стимула без осознания на-
личия этого стимула. См. гл. 6, «Манипуляции со зрительным сознанием»
бета-волны — диапазон иррегулярных электромагнитных волн, регистрируемых
от мозга или поверхности скальпа, отражающий активность большой популяции
нейронов. Бета-волны имеют частоту выше 12 Гц, и связан с состоянием сознатель-
ного нормального бодрствования. Низкоамплитудные β-волны с множественной и
изменяющейся частотой нередко связывают с процессами активного, интенсивного
мышления и активной концентрации. См. гл. 4, «Ряд полезных инструментов»
бинокулярная диспаратность (от лат. bi- — два; oculus — глаз; disparare — разде-
лять) — различие в восприятии левым и правым глазами местоположения объекта,
вследствие горизонтального расположения глаз. Мозг использует бинокулярную
диспаратность для получения информации об объеме от изображений с сетчатки
обоих глаз. См. гл. 6, «Функциональная организация зрительной системы»
бинокулярное соревнование (от лат. bi- — два; oculus — глаз) — переменное вос-
приятие, которое возникает, когда разные стимулы предъявляются в каждый глаз,
и мозг не может соединить их в один согласованный образ. Вместо этого образ
от каждого глаза возникает и исчезает из сознания за несколько секунд. См. гл. 6,
«Связь деятельности мозга и зрительного опыта», гл. 8 . См. неоднозначное вос-
приятие
бихевиоризм — философское направление в физиологии, которое предполагает,
что любые действия организма, включая движения, мышление и ощущение, не-
обходимо рассматривать как поведение. См. гл. 1, «Наука возвращается к представ-
лениям о сознании»
большой мозг (см. кора больших полушарий)
Бродбент, Дональд (1926–1993) — британский когнитивный психолог, развивший
важные понятия селективного внимания и рабочей памяти. См. гл. 2, «Ограничен-
ные и обширные функции»
Брока, Пьер-Поль (1824–1880) — французский хирург, изучавший повреждения
мозга у пациентов с афазией, неспособностью говорить, в то время как они пони-
мали речь. После смерти пациентов Брока проводил посмертное определение по-
врежденной области в левой нижней лобной извилине, которую сейчас называют
зоной Брока. См. гл . 1, «Немного истории и продолжающаяся дискуссия»
436
Глоссарий
Бубер, Мартин (1878–1965) — родившийся в Австрии еврейский философ, широ-
ко известный благодаря философскому диалогу «Я и Оно», который подчеркивает
взаимосвязи между осознанными личностями, вместо того чтобы сводить их к объ-
ектам. См. гл . 14, «Заключение»
Везалий, Андреас (1514–1564) — бельгийский физиолог, который создал первый
точный атлас анатомии человеческого тела в 1543 г., названный «О строении чело-
веческого тела». Книга Везилия стала основополагающей вехой в развитии науки
и медицины в эпоху Ренессанса. См. гл . 1, «Немного истории и продолжающаяся
дискуссия»
везикула (от лат. vesicular — маленький пузырь) — маленькие пузырьки, напол-
ненные молекулами нейромедиаторов, которые путешествуют по аксону к синап-
тическим терминалям, где они сливаются с синаптической мембраной для высво-
бождения нейромедиаторов в синаптическую щель при возникновении потенциала
действия. Нейромедиаторы диффундируют в щели и запускают деполяризацию на
постсинаптической мембране, в конечном счете вызывая другой спайк. Везикулы
необходимы для передачи сигналов между нейронами и постоянно формируются в
клетке. См. гл. 3 и 16
вентральный (от лат. venter — живот) — нижняя часть мозговых структур
вентромедиальная префронтальная кора (от лат. venter — живот; medialis – в
центре) — нижняя часть медиальной поверхности лобных долей, преимуществен-
но у людей и приматов. Это область, простирающаяся назад от носа, включена в
процессы эмоций, связь матери и ребенка, процессы страха и риска при принятии
решений. См. гл . 12
вербальная диспраксия (от греч. dys — поврежденный; praxis — действие) — со -
стояние, при котором нарушена способность говорить. См. гл . 16
вербальное повторение — мысленное повторение слов при помощи внутренней
речи (компонента рабочей памяти) для их запоминания. Внутренняя речь включает
спонтанные комментарии относительно текущих забот, целей и эмоций. См. гл . 2,
«Классическое понятие оперативной памяти»
веретенообразная область лиц (от лат. fusus — веретено, из-за ее формы) — спе-
циализированная область на медиальной поверхности височной доли, которая отве-
чает селективно на предъявления зрительных образов лиц по сравнению с другими
объектами. См. гл. 6, «Функциональная организация зрительной системы»
Вернике, Карл (1848–1905) — немецкий физиолог и первооткрыватель специаль-
ной области коры, ответственной за понимание речи. Эта область сейчас называ-
ется зоной Вернике и ассоциирована с повреждениями, известными как афазия
Вернике или рецептивная афазия. Пациенты с такими нарушениями не способны
понимать речь, включая свою собственную, но могут производить беглую (но не
всегда значащую) речь. См. гл. 1, «Немного истории и продолжающаяся дискуссия»
верхний — сверху. В анатомии мозга является синонимом слова дорсальный
височная доля — височные доли являются частью коры больших полушарий, уча-
ствующей в зрительном восприятии, восприятии звуков и речи, кодировании и из-
влечении памяти. Они находятся на боковой поверхности мозга под латеральной
бороздой. В профиль мозг человека напоминает перчатку для бокса, у которой боль-
шой палец – височная доля. Височная доля окружает гиппокамп и миндалину, по-
Глоссарий
437
этому участвует в функциях эмоции и памяти. Медиальная поверхность височной
доли (лучше видимая при рассмотрении мозга снизу) является древним палеокортек-
сом, включая обонятельную кору. См. гл. 1 и 5
височная пластинка (planum temporale; от лат. planum — плоская поверхность;
temporalis — у виска) — часть слуховой коры, участвующая в анализе звуков и
восприятии речи. См. гл . 7, «Функциональное картирование обработки слуховой
информации»
внимание, привлеченное стимулом — привлечение внимания новым резким сти-
мулом. См. гл . 8
внимание — выделение некоторых сенсорных, когнитивных или двигательных со-
бытий и вытеснение других. Внимание часто описывают как фокус на некоторых
сознательных событиях. См. гл . 8, «Селективное внимание»
возвратные связи — большая часть мозговых связей является двусторонней, т. е .
активность в точке А запускает активность в точке Б и наоборот. См. гл. 3, «Рабочие
положения»
вокодер (от англ. voice — голос и coder — кодирующее устройство) — техно-
логия, созданная компанией Bell Telephone Laboratories для передачи высоко-
отфильтрованной речи с минимальными потерями разборчивости. Эта технология
лежит в основе ушных имплантов. См. гл . 7, «Восприятие речи» и Блок 7.1
волевой акт — произвольный контроль за действиями, противопоставлен автома-
тическому контролю, как происходит при высокотренированных автоматизирован-
ных действиях. Многие нарушения мозга вызывают потерю произвольного контро-
ля. См. гл. 12
временная огибающая — возрастание и падение интенсивности речевых сигна-
лов во времени. См. гл . 7, «Восприятие речи»
вызванный потенциал (от лат. potentia — сила) — также называют связанные с
событием потенциалы (ССП). Довольно стереотипные паттерны электрической
разности потенциалов, регистрируемых от мозга, после усреднения потенциалов
относительно времени или события. Традиционно ВП получают, усредняя несколь-
ко ЭЭГ-записей относительно момента предъявления стимула. Несмотря на то что
точные источники генерации ВП неизвестны, ВП являются крайне чувствитель-
ными к когнитивным и эмоциональным переменным. См. гл . 4, «Ряд полезных ин-
струментов»
выходные функции — мозговые процессы, контролируемые лобными долями и
включающие высшие произвольные функции, планирование действий и моторный
выход. См. гл . 2, «Классическое понятие оперативной памяти»
гамма-волны — полоса быстрой низкоамплитудной электромагнитной активно-
сти, ассоциированной с бодрствованием и активным мышлением, регистрируемая
от мозга или поверхности скальпа, вероятно, отражающая активность большой по-
пуляции нейронов. Считается, что пик гамма-активности приходится на 40 Гц, а
диапазон имеет границы от 25 до 60 Гц. Однако более высокочастотная активность
также часто относится к гамма-активности. Считается, что гамма отражает связи
областей для обеспечения определенных функций. См. гл . 4, «Ряд полезных ин-
струментов»
438
Глоссарий
гамма-аминомасляная кислота (ГАМК) — основной тормозный нейромедиатор
в центральной нервной системе. Играет важную роль в регулировании нейронной и
поведенческой возбудимости, включая сон. ГАМК необходима для мозговой актив-
ности и регулирования мышечного тонуса. См. гл . 16
ганглий (от лат. вздутие) — большое скопление нейронов. Основные подкорко-
вые структуры можно рассматривать как ганглии, например базальные ядра. Как
правило, это большие структуры с множеством функций. Обычно они состоят из
подразделений, которые, в свою очередь, имеют слоистое или складчатое строение.
См. гл. 5, «От вопроса «где?» до вопроса «что?»: функциональная роль областей
мозга»
Гейдж, Финеас (1823–1860) — уникальный пациент с повреждением мозга в ре-
зультате несчастного случая на железной дороге, который продемонстрировал
удивительные резервные когнитивные возможности, несмотря на значительное по-
вреждение лобных долей. Гейдж был железнодорожным рабочим, которому шпа-
лоподбойка, длиной в два фута, прошла через верхнюю часть орбиты левого глаза
и вышла в середине черепа, когда произошел случайный взрыв нестабильного за-
ряда динамита. Несмотря на то что у Гейджа не отмечалась утрата восприятия, мо-
торного контроля или речи, в его личности произошли изменения, которые теперь
определяют как результат повреждения лобных долей, особенно значительна была
потеря способности контролировать импульсы и долговременные мотивации. См.
гл.2и12
Гельмгольц, Герман фон (1821–1894) — немецкий врач, физик и исследователь
сенсорной системы. Его работы по зрению и слуховой системе до сих пор изуча-
ют, но он широко известен благодаря демонстрации физического закона сохране-
ния энергии с использованием электрической стимуляции препарированной лапки
лягушки. Гельмгольц первым предположил, что зрительная система создает «бес-
сознательное умозаключение», которое является нечто большим, чем просто вход
световой информации в глаза. Эта идея в то время была спорной, но сейчас широко
принята. См. гл. 1, «Немного истории и продолжающаяся дискуссия»
гематоэнцефалический барьер — барьер между кровеносной системой и тканью
мозга, который создают клетки, окружающие кровеносные сосуды в мозге. Он про-
пускает маленькие молекулы, например кислород и глюкозу, препятствует прохож-
дению больших молекул. См. гл. 16
гемодинамика (от hema — кровь; dynamos — сила) — изучение изменения крово-
тока, в особенности в мозге, например показателя локальной активности нейронов.
См.гл.1и4
геном (от греч. genea — поколение, раса) — полная генетическая копия, определя-
ющая биологический вид. См. гл . 16
генотип —геном, часто сопоставляемый с фенотипом определенной особи, кото-
рый является физиологическим отображением генетического кода. См. гл . 16
гештальт (от нем. форма) — 1) перцептивный образ, который нельзя разбить на
простые субкомпоненты; 2) направление в психологии, основанное на немецком
представлении о Gestalt, которое характеризуют следующим высказыванием: «Це-
лое является чем-то большим, чем просто сумма составляющих его частей». Геш-
тальтпсихология оказала значительное влияние на исследование процессов воспри-
ятия. См. гл . 6, «Введение»
Глоссарий
439
гидроанэнцефалия (от греч. hydr- — вода; encephalon — голова) — внутриутроб-
ное нарушение, заключающееся в тяжелом повреждении мозга, которое включает
отсутствие нормального развития коры; отсутствующая ткань замещается водяни-
стой цереброспинальной жидкостью. Гидроанэнцефалия является более тяжелым
нарушением, чем гидроцефалия, при которой развитие мозга более полное, несмо-
тря на то, что происходит накопление цереброспинальной жидкости в желудочках.
См. гл. 16
гиперколонка (от греч. hyper — сверху; лат. columna — вершина) — группа коло-
нок коры, которые часто содержат близко связанные нейроны. См. гл . 5, «Введение»
гипоталамус (от лат. hypo — под; греч. thalamos — палата) — основной нейроэндо-
кринный орган в каждой половине мозга, имеющий витальные роли в регулирова-
нии питательных веществ в крови, контроль мотиваций, голода и других основных
жизненных функций. Гипоталамус расположен под каждым таламусом, сразу над
стволом мозга. Он близко связан с гипофизом и эпифизом. См. гл . 5, «Развитие
мозга от основания до вершины»
гипотеза консолидации — процесс, благодаря которому новые памятные собы-
тия переводятся в памятные следы долговременной памяти. Воспоминания могут
храниться в тех же областях мозга, которые обеспечивают активные моментальные
функции как восприятие и контроль речи. Консолидация может основываться на
синаптических изменениях в определенных областях мозга, что делает активные
нейронные связи более эффективными. См. гл . 2, «Консолидация кратковременных
событий в долговременную память»
гипофиз — главная железа организма. Это эндокринная железа размером с горо-
шину как будто свисает с гипоталамуса, расположенного в основании мозга. Гипо-
физ совместно с гипоталамусом регулируют стадии развития и гомеостаз. См. гл . 5,
«Развитие мозга от основания до вершины»
гиппокамп (греч. морской конек, от hippos — лошадь, kampos — морское чудови-
ще) — в мозге человека парная дугообразная структура в медиальных височных
долях. Гиппокамп является частью лимбической системы и играет основную роль
в кодировании и извлечении эпизодической и семантической памяти и в про-
странственной навигации. См. гл . 2, «Классическое понятие оперативной памяти»
гистогенез (от греч. histos — стрела, очертания, луч, сплетение; греч. gignesthai —
рождаться) — формирование специфичных клеток из менее дифференцированных
клеток-предшественниц. См. гл . 15, «Пренатальное развитие: от бластоцисты до
младенца».
глия (от греч. glia — клей) — не нервные клетки в мозге, которые выполняют под-
держивающие функции, обеспечивают нейрохимический гомеостаз, образуют за-
щитную миелиновую оболочку вокруг нейронов и обрабатывают информацию.
См. гл . 15, «Пренатальное развитие: от бластоцисты до младенца»
глутамат (от лат. gluten — клей) — молекула, производная глутаминовой кислоты,
которая является самым распространенным возбуждающим нейромедиатором в
мозге. См. гамма-аминомасляная кислота (ГАМК). См. гл. 3 и 16.
гомеобокс (Hox) (от греч. homos — одинаковый) — ДНК-последовательность, ко-
торая регулирует строение тела у животных и растений. Гены ��� обычно регули-
��� обычно регули-
обычно регули-
руют эффекты других генов
440
Глоссарий
гомункулус (лат. — ма ленький человек) — искаженная карта человеческого тела
в первичной соматосенсорной коре (сенсорный гомункулус) и в первичной мо-
торной коре (моторный гомункулус). Губы, руки, ступни и половые органы имеют
большее число сенсорных нейронов, чем другие части тела, и гомункулус имеет
соответственно искаженные большие губы, руки, стопы и гениталии. Каждое полу-
шарие имеет сенсорный и моторный гомункулус противоположной части тела. Эти
карты тела были открыты нейрохирургом Уайлдером Пенфилдом в Университете
Монреаля в 1950-х и 1960-х гг. См. гл . 5, «От вопроса «где?» до вопроса «что?»:
функциональная роль областей мозга»
Дарвин, Чарльз (1809–1882) — английский натуралист, который впервые в совре-
менном мире высказал убедительную мысль о том, что все живущие виды возникли
от одного общего предка в течение длительного периода времени, так как «выжи-
вает наиболее приспособленный» и наиболее успешно размножающийся. Дарвин
также опубликовал труд о важных наблюдениях эмоциональных выражений у лю-
дей и животных. См. гл . 1, «Немного истории и продолжающаяся дискуссия»
двигательная слепота (акинетопсия) — симптом, вызванный повреждением об-
ластей мозга, которые отвечают на восприятие действия, например, зона �� зри-
�� зри-
зри-
тельной коры, и приводящий к неспособности воспринимать движение. См. гл . 6,
«Области мозга, необходимые для визуального восприятия: исследование повреж-
дений»
дежа вю (déjà vu, от франц. «уже видел») — чувство, что уже переживал текущий
момент прежде. См. гл . 9, «Вариации памяти» и блок 9.3
Декарт, Рене (1596–1650) — французский философ, математик, ученый и писа-
тель, который провел большую часть жизни в Дании. Декарт, названный «Отцом
современной философии», был очень внимательным исследователем мозга. Часто
его считают основателем современной философии разума/тела. См. гл. 1, «Немного
истории и продолжающаяся дискуссия»
декларативная память (от лат. declarare — сделать видимым) — способность
помнить факты и мнения. Тип эксплицитной памяти. См. гл . 2, «Классическое
понятие оперативной памяти»
дельта-волны — диапазон медленных, высокоамплитудных электромагнитных
волн, характерных для глубокого сна и регистрируемых от поверхности мозга или
скальпа и отражающих активность большой популяции нейронов. Частота дельта-
активности ниже 2,5 Гц. Дельта-волны присутствуют в ЭЭГ и в состоянии бодр-
ствования, но появляются в нативной (не обработанной) записи ЭЭГ только когда
начинают доминировать в состоянии сна и дремоты. См. гл . 4, «Ряд полезных ин-
струментов»
дендрит (от греч. dendron — дерево) — одна из многочисленных тонких, ветвящих-
ся микроскопических ветвей, простирающихся от тела нейрона. Как правило, ден-
дриты получают синаптический вход от других нейронов, и тем самым являются
входными воротами для нейронов. См. гл . 3, «Введение»
Джеймс, Уильям (1842–1910) — американский психолог и философ. Джеймс поды-
тожил исследования XIX в. о человеке и мозге. Его принципы психологии (1893)
использовались широко в психологии как вступительное слово вплоть до 1920-х гг.
Джеймс сильно повлиял на работы западного течения в литературе о разуме, пси-
Глоссарий
441
хологии обучения и религиозных и мистических переживаниях. В философии он
считается основным приверженцем прагматизма, и его считают родоначальником
европейской феноменологии. Он был братом писателя-романиста Генри Джеймса и
Алисы Джеймс. См. гл . 1, «Немного истории и продолжающаяся дискуссия»
динамическое причинно-следственное моделирование (от греч. dynamikos —
мощный; лат. causa — причина; modulus — маленькая мера) — метод интерпре-
тации мозговых данных, например в функциональной магнитно-резонансной то-
мографии (фМРТ), который помогает установить причинно-следственную связь
между мозговой активностью во время выполнения определенного задания. См. гл .
4, «Корреляция и причинная связь»
дискурс — обсуждение обществе, часто отражающее определенные политические
взгляды. См. гл . 11, «Введение»
диффузионная трактография (от лат. diffusus — распределение; trahere — тянуть;
греч. graphein — писать) — метод визуализации мозга, основанный на диффузии
молекул воды, направленный на выявление основных нервных путей в мозге. См.
гл. 4 и приложение
долговременная депрессия (ДВД) (от лат. deprimere — подавлять) — длительное
снижение активности синапса. Наряду с долговременной потенциацией (ДВП),
ДВД считается синаптической основой обучения и долговременной памяти. См.
гл. 3, «Адаптация и обучение массивов нейронов»
долговременная потенциация (ДВП) (от лат. potentia — сила) — длительное
усиление синаптического контакта. Наряду с ДВД, ДВП считается синаптической
основой обучения и долговременной памяти. См. гл. 3, «Адаптация и обучение
массивов нейронов»
дорсальный (от dorsum — спина) — верхняя часть мозговых структур, также на-
зывается верхней
дорсолатеральная префронтальная кора (от лат. dorsum — спина; latus — сторо-
на; frons — лоб; греч. cortex — кора) — префронтальная область, ответственная за
планирование моторных действий, исполнительный контроль, самоконтроль, эмо-
ции и рабочую память. См. гл. 2, «Центральное управление»
дорсолатеральный синдром (от лат. dorsum — спина; latus — сторона; frons —
лоб) — нарушения, возникающие вследствие повреждения дорсолатеральной
префронтальной коры, такие как эмоциональная уплощенность (отсутствие эмо-
ций) и нарушение способности переключать и инициировать новые действия. См.
гл. 12, «Нарушения лобных долей»
дофамин — один из главных нейромедиаторов и нейромодуляторов, который об-
разуется в нескольких частях мозга, включая черную субстанцию и вентральную
тегментальную зону. Дофамин имеет ряд функций, включающих контроль произ-
вольных движений, подкрепление и подавление лактации (образования молока) у
женщин, сон, настроение, внимание и обучение
дугообразный пучок — пучок аксональных волокон, преимущественно тех, ко-
торые соединяют зоны Брока и Вернике в левом полушарии. См. гл . 1, «Немного
истории и продолжающаяся дискуссия»
442
Глоссарий
желудочки — четыре маленькие полости в мозге, содержащие циркулирующую
спинномозговую жидкость. В стенках желудочков были найдены области со ство-
ловыми клетками. См. гл . 5, «Развитие мозга от основания до вершины»
задачи на сети внимания (AN�, attenti�n network task) — обобщение фланкерно-
го теста, метод исследования зрительного внимания. ANT позволяет изучить три
аспекта внимания: приведение в состояние готовности перед появлением ожидае-
мого сигнала, ориентация в направлении определенного местоположения, где ожи-
дается появление стимула, и произвольное внимание для того, чтобы действовать
против ожидаемых событий, запрограммированных этим тестом. См. гл . 8
задний — в анатомии мозга задний является синонимом каудальному
затухание — процесс, при котором выученное поведение перестает подкрепляться
и вследствие этого уменьшается частота его выполнения. При теменном игнориро-
вании, после повреждения правой теменной доли, появляется возможность стимула,
предъявленного в левой части зрительного поля, подавлять стимулы, предъявленные
в правой части поля. См. гл . 13, «Система страха»
затылочная доля — затылочные доли мозга, которые содержат первичную область
зрительной коры, являются самыми маленькими из четырех долей коры больших
полушарий человека. См. гл . 1 и 5
зеркальный нейрон — нейрон, который разряжается как при совершении живот-
ным определенного действия, так и при наблюдении за другим животным, совер-
шающим это же действие. Большая группа зеркальных нейронов была обнаружена
у приматов, и считается, что такие клетки имеются и у других видов, включая птиц.
У человека мозговая активность, подобная зеркальным нейронам, была выявлена в
премоторной коре и нижней теменной коре. Однако существуют многочисленные
споры относительно этих открытий. См. гл . 14
зона MT (V5, средняя височная) — часть зрительной коры, которая отвечает за
зрительное восприятие движения. См. гл. 6, «Функциональная организация зри-
тельной системы»
зона Брока — левая нижняя лобная извилина, или ее задняя часть, согласно со-
общению Пьера-Поля Брока в 1861 г., ответственна за нарушение способности
говорить у пациента при сохранении у него способности понимать речь. Другие
функции также были приписаны зоне Брока. См. гл. 1, «Немного истории и про-
должающаяся дискуссия», и гл. 7, «Восприятие речи»
зона Вернике — область, расположенная в верхней задней части височной доли и
необходимая для понимания речи. См. гл . 1, «Немного истории и продолжающаяся
дискуссия»
зрительная агнозия (от греч. agnosia — отсутствие знания) — состояние, при ко-
тором человек испытывает трудности опознавания объектов, что вызвано повреж-
дением областей коры, ответственных за узнавание объектов, например, нижняя
височная кора. См. гл . 6, «Области мозга, необходимые для визуального восприя-
тия: исследование повреждений»
зрительная обратная маскировка — осознанный зрительный образ может быть
стерт последующим зрительным объектом, как решетчатый экран, даже несмотря
на то, что этот образ физически достигает сетчатки. См. гл . 8
Глоссарий
443
зрительно-пространственный скетчпад — способность удерживать зрительную
и пространственную информацию одновременно в рабочей памяти. См. гл. 2,
«Классическое понятие оперативной памяти»
зрительные фантомы — форма перцептивного заполнения, при которой зритель-
ные образы возникают перед окружающими формами и цветами, что создает иллю-
зию. См. гл . 6, «Связь деятельности мозга и зрительного опыта»
иконическая память (также называют кратковременной памятью) — способ-
ность воспроизводить что-то в течение 10–30 с без заучивания. Рабочую память и
сенсорный след можно рассматривать как особый вид иконической памяти. См. гл .
2, «Классическое понятие оперативной памяти»
иллюзия движения — вид перцептивного восприятия, при котором мозг создает
иллюзию движения, когда ряд неподвижных изображений молниеносно следуют
друг за другом. См. гл. 6, «Связь деятельности мозга и зрительного опыта»
имплицитная память (от лат. implicitus — неявный) — бессознательная память, ко -
торая может появиться вследствие сознательных или бессознательных событий. См.
гл. 2, «Классическое понятие оперативной памяти», гл. 9, «Введение»
интенциональность — направление психических событий, их способность отра-
жать проявления внешнего мира. Отличают от намерения, мысленной цели. См. гл .
14, «Заключение»
интерауральная разница уровня звука (от лат. inter-
—
между; auris — ухо) —
метод определения локализации источника звука, при котором мозг выявляет ма-
ленькую разницу в громкости звука, возникающего между двух ушей, распростра-
няющегося к голове под углом. См. гл . 7, «Функциональное картирование слуховых
процессов».
интерауральный интервал (от лат. inter-
—
между; auris — ухо) — Метод лока-
лизации звука, когда мозг выявляет задержку на доли секунды прихода звука от
отдаленного источника к дальнему уху по сравнению с ближним ухом. См. гл. 7,
«Функциональное картирование обработки слуховой информации»
интерсубъективность — способность разделять субъективный опыт, как, напри-
мер, способность младенцев указывать и называть игрушку и получать одобрение
от взрослых. Критический аспект социального общения и обучения речи. См. гл .
14, «Заключение»
интонационный контур — «мелодия» нормальной речи. В английском и других
языках вопросы обычно произносят с особой интонацией, которая отлична от ин-
тонации утвердительных фраз. См. гл . 11, «Звуки устной речи»
ионотропный рецептор (от греч. ion — идти; tropos — направление) — группа
мембранных рецепторных каналов для заряженных частиц, таких как Ca+ 2
,Cl
–
и
Na+
. Ионотропные рецепторы могут открываться или закрываться нейромедиато-
рами. Иногда называются рецепторами, связанными с каналом
ипсилатеральный (от лат. ipse — сам; latus — сторона) — на той же стороне тела.
См. контралатеральный. См. гл. 2, «Действие»
искусственная нервная сеть (известна также как нейронные модели) — искус-
ственная нервная сеть является имитационной, упрощенной моделью функцио-
нирования мозга. Большинство имеют относительно не большой размер. Однако
444
Глоссарий
они чрезвычайно важны для понимания принципов нервных процессов. См. гл . 3,
«Введение»
исполнительное внимание (известно также как произвольное, целевое, или нис-
ходящее внимание) — акт волевого фокусирования внимания на потоке осознава-
емых событий, в то время как другие события игнорируются. См. также селектив-
ное внимание, внимание, направленное на стимул. См. гл. 8
исполнительные функции (также называют исполнительным контролем или
функциями лобных долей) — способности, такие как планирование, когнитивная
пластичность, произвольные действия, абстрактное мышление, усвоение правил,
инициирование правильных действий и подавление неправильных, контроль им-
пульсных актов и регуляция эмоций. См. гл . 12, «Введение»
классический условный рефлекс (также называют Павловским условным реф-
лексом) — форма ассоциативного обучения, при которой люди или животные об-
учаются тому, что произвольный стимул (как звонок) свидетельствует о появле-
нии биологического стимула (например, еды). И. П. Павлов назвал эти стимулы
условным стимулом (УС) и безусловным стимулом (БС). Если УС неоднократно
предъявляется перед БС, у животного начинает формироваться реакция в ответ на
УС. Предположение Павлова о том, что условные рефлексы являются основой для
человеческого обучения, уже несправедливо. Однако классический условный реф-
лекс широко применяется в исследованиях, и считается, что он необходим для вы-
ученных тревожных расстройств и пищевого предпочтения. См. гл . 13, «Введение»
клеточные ассамблеи (также называемые нейроном Хебба) — активные сети свя-
занных нейронов отражают некий сенсорный вход или схожее событие. Согласно
гипотезе 1949 г. Дональда О. Хебба, «нейроны, которые разряжаются вместе, об-
разуют связь», так что спонтанные разряды вызывают усиление синаптических
связей в клеточных группах. См. учение Хебба. См. гл . 3, «Рабочие гипотезы»
когнитивная нейронаука (от лат. cognoscere — знать) — возникающее слияние
двух отдельных областей науки, когнитивной психологии и нейронауки. Большин-
ство исследований в области когнитивной нейронауки используют комбинацию
психологических методов с одновременной регистрацией мозговой активности.
См. гл. 1, «Введение»
когнитивная нейронаука развития — исследования нормального развития мозга
и мыслительных способностей. См. гл . 15, «Введение»
когнитивная установка — состояние умственной подготовки к некому событию
или реакции. См. гл . 12, «Детальное рассмотрение функций лобных долей»
компьютерная томография (от лат. computare — учитывать; греч. tomos — срез;
graphein — писать) — физиологические записи, в которых трехмерное изображе-
ние структуры тела (например, мозга) строится компьютером на базе серии изобра-
жений срезов. См. гл. 4, «Введение»
коннекционизм (от лат. com- + nectere — соединять вместе) — исследования ис-
кусственных или биологических нейронных сетей. См. гл. 3, «Адаптация и обуче-
ние массивов нейронов»
контралатеральный (от лат. «противоположно стороне») — противоположная
сторона тела или мозга. См. гл . 2, «Действие». См. ипсилатеральный
Глоссарий
445
конфабуляции (от лат. fibula — рассказ) — неврологический симптом, при кото-
ром рассказывают ложные воспоминания или ощущения без намерения соврать.
См. гл. 9, «Воспоминания состоят из этого»
координаты Талайраха — точная трехмерная координатная система мозга чело-
века, позволяющая локализовать любую точку мозга с точностью до миллиметра.
См. гл. 5, «Введение»
кора больших полушарий — внешняя поверхность большого мозга, самая боль-
шая часть человеческого мозга, разделенная на два симметричных больших полу-
шария мозга. В коре в основном разделяют шесть разных клеточных слоев, содер-
жащих тела нейронов, которые выглядят серыми. Далеко расположенные друг от
друга нейроны посылают аксоны в другие части коры, в таламус и другие области
мозга, которые покрываются белыми поддерживающими клетками (миелин). В ре-
зультате, вертикальный срез коры выглядит для невооруженного глаза как белая
масса и тонкой серой оболочкой сверху, которые соответственно называют белым
и серым веществом. Кора больших полушарий играет важную роль в сенсорном
восприятии, пространственной локализации, производстве и восприятии речи, па-
мяти, внимании, эмоциях, мотивациях, планировании действий, произвольном
контроле, мышлении, высших исполнительных функциях и сознании. См. гл .
1, «Некоторые исходные положения», и гл. 5, «Развитие мозга от основания до вер-
шины»
кора (см. кора больших полушарий)
корковая колонка (от греч. cort — кожа, кожура; лат. columna — вершина) — ци-
линдрические срезы шести слоев коры, которые часто содержат связанные нейро-
ны. Колонки имеют диаметр около 0,5 мм и высоту 2,5 мм. Они могут образовывать
гиперколонки, которые в свою очередь могут являться частью более больших об-
разований. См. гл . 5, «Введение»
корковая цветовая слепота — неспособность различать цвета, вызванная по-
вреждением воспринимающих цвет областей зрительной коры. См. гл . 6, «Области
мозга, необходимые для зрительного восприятия: изучение разрешений»
коронарный (от лат. corona — корона) — вертикальный срез мозга, который делит
его на переднюю и заднюю половины. См. гл . 1, «Некоторые начальные положе-
ния»
креол — истинный язык, который дети спонтанно развивают в многоязыковой сре-
де. Очень часто креолы возникают в островных популяциях, где популяции с раз-
ным языком перекрываются. «Креолы» противопоставляют «пиджинам», которые
являются диалектами, обычно использующимися взрослыми людьми как второй
язык. Креолы уникальны, так как они имеют полноценную грамматику в отличие от
гибридных языков. Спонтанность возникновения креолов свидетельствует в пользу
того, что младенцы и дети имеют универсальную биологическую предрасположен-
ность к речи. См. гл . 11, «Природа языка» и блок 11.2
латеральная борозда (от лат. latus — сторона) (также называют сильвиевой бороз-
дой) — эта хорошо выделяемая борозда отделяет теменные доли мозга от лобных
и височных долей. См. гл. 1, «Некоторые исходные положения»
латеральное коленчатое тело — ядро в таламусе, имеющее коленопободный слой
клеток. Это основной центр переключения информации между сетчаткой глаза и
446
Глоссарий
первичной зрительной корой (областью �1). См. гл . 6, «Функциональная органи-
�1). См. гл . 6, «Функциональная органи-
1). См. гл . 6, «Функциональная органи-
зация зрительной системы»
латеральное торможение (от лат. latus — сторона) — способность нейронов тор-
мозить активность соседних клеток в том же слое нейронов. См. гл . 3, «Рабочие
гипотезы»
латеральный (от лат. latus — сторона) — на боковой поверхности мозга. См. гл . 1,
«Некоторые исходные положения»
латеральный затылочный комплекс — область мозга на латеральной поверхно-
сти затылочной доли мозга, которой отводят основную роль в зрительном распоз-
навании объектов. См. гл . 6 «Функциональная организация зрительной системы»
лексикон (от греч. lexis — слово) — словарный запас языка. См. гл . 11, «Введение»
лексическое отождествление (от греч. lexis — слово) — процесс отождествления
слов со звуками речи. См. гл . 11, «Введение»
лимбическая система (от лат. limbus — граница) — древняя группа структур мозга,
вовлеченных в процессы эмоций, памяти, обоняния и контроля действий, состоя-
щая из гиппокампа, миндалины, таламуса, гипоталамуса и поясной извилины.
Лимбическая система имеет тесные связи с эндокринной системой и автономной
нервной системой. См. гл. 13, «Введение»
лобные доли — большая область коры, расположенная в передней части каждо-
го полушария мозга, граничит сзади с теменными долями и примыкает к верх-
ней передней границе височных долей. Исполнительные функции лобных долей
включают способность предвидеть последовательность действий, планировать и
принимать решения, говорить, преодолевать неуместные импульсы и разрешать
конфликты целей, понимать мысленные состояния других, держать информацию в
рабочей памяти. См. гл . 1, «Немного истории и продолжающаяся дискуссия», и гл.
5, «Введение»
локализация звука — определение местоположения звука часто основывается на
бинауральном различии между ушами в громкости и времени прихода звука. См.
гл. 7, «Функциональное картирование обработки слуховой информации»
магнитно-резонансная томография (МРТ, от лат. resonare — звучать; imago —
имитация) — основанная на спиновом резонансе ядер атомов, МРТ является ме-
тодом визуализации внутренней структуры тела, включая мозг. Функциональная
МРТ (фМРТ) регистрирует мозговую активность, и очень часто данные фМРТ со-
вмещают со снимком мозга, полученным при помощи МРТ. См. гл. 4, «Введение»
магнитоэнцефалография (МЭГ, от греч. en - + kephale — в голове; graphein — пи -
сать) — метод визуализации, основанный на регистрации магнитных полей работа-
ющего мозга. МЭГ является удобным неинвазивным методом с хорошей временной
и пространственной разрешающей способностью. См. гл. 4, «Ряд полезных инстру-
ментов» и приложение
медиальная височная доля (МВД) — нижняя часть височных долей, которая
расположена симметрично середины мозга и содержит эволюционно более древ-
ние обонятельные структуры, а также функции кодирования и извлечения памяти
и эмоции. См. гл. 5, «От вопорса «где?» до вопроса «что?»: функциональная роль
областей мозга»
Глоссарий
447
медиальный — к средней линии тела. Среднесаггитальный. См. гл . 1, «Некото-
рые исходные положения»
ментализация (от лат. mens — разум) — способность понимать себя и других, не
только как сенсорных объектов, но и как субъективных существ со своим менталь-
ным состоянием. См. гл . 14, «Заключение»
метаботропный рецептор (от греч. metabole — изменение; tropos — направле-
ние) — нейронные рецепторы, которые используют существующую систему мета-
болизма клетки для активации постсинаптического ответа. Их противопоставляют
ионотропным рецепторам, использующим ионные каналы на мембране клетки
метакогнитивный (от греч. meta — над; лат. cognere — знать) — знание о разуме.
Собственное знание о памяти, восприятии или произвольных действиях. См. гл . 8
миграция нейронов — перемещение нервных клеток из области деления в зоны
их конечной локализации во взрослом мозге. См. гл . 15, «Пренатальное развитие:
от бластоцисты к младенцу»
миелин (от лат. myel — мякоть) — оболочка из глиальных клеток, окружающая
аксоны многих нейронов. Миелинизированные аксоны выглядят белыми, поэтому
нервные волокна относят к белому веществу мозга. См. гл . 1 и 3
миндалина (от amygdale — миндаль) — парное небольшое миндалевидное ско-
пление нейронов, расположенное внутри височных долей. Они являются частью
лимбической системы и играют важную роль как в эмоциях, например страх и до-
верие, так и в процессах обучения. См. гл. 1, «Немного истории и продолжающаяся
дискуссия»
митохондрия (от греч. mitos — нить; chondrion — зерно) — органелла, в которой
производится энергия; присутствует в клетках большинства животных. Считается,
что митохондрии произошли от сине-зеленых водорослей, которые мигрировали в
клетки животных и установили с ними взаимовыгодное сожительство или симби-
оз. Митохондрии являются основным источником энергетической молекулы АТФ
(аденозинтрифосфат), и необходимы для жизни животных. Они имеют ядро с соб-
ственной молекулой ДНК, но большая часть их ДНК была перенесена в ядро клет-
ки-хозяина. Так как производство энергии является фундаментальным процессом
жизни, метаболическая система митохондрий также вовлекается к функции пере-
дачи нейронами информации. См. гл . 16
модель нейронных сетей (так же называют искусственные нейронные сети (ИНС))
—
имитация, упрощенные модели для обеспечения определенных функций мозга.
Большинство из них относительно невелики и не отражают высокую сложность
мозга. Однако они чрезвычайно важны для улучшения понимания процессов, про-
исходящих в мозге. См. гл . 3
модель радиальной единицы — модель нейронной миграции, предложенная уче-
ным Паско Ракичем, которая постулирует, что в развивающейся коре головного
мозга клетки образуются в основании колонки, и каждая новая клетка мигрирует
дальше своей предшественницы. См. гл . 15, «Пренатальное развитие: от бластоци-
сты к младенцу»
модульная теория (от лат. modulus — маленькое измерение; греч. theoria — пред-
положение) — теория, которая постулирует, что различные функции мозга могут
быть локализованы в разных областях или сетях мозга. См. гл . 14, «Заключение»
448
Глоссарий
мозжечок (от лат. cerebrum — мозг; �e�e�e���� значит «маленький мозг») — боль-
�e�e�e���� значит «маленький мозг») — боль-
значит «маленький мозг») — боль-
шая область мозга, расположенная ниже и сзади от затылочных долей больших
полушарий. Мозжечок играет важную роль в интеграции сенсорного восприятия,
точного моторного контроля и сенсомоторной координации. Недавние исследова-
ния указывают на его участие в когнитивной деятельности. См. гл . 1, «Некоторые
исходные положения», и гл. 5, «Введение»
мозолистое тело (���p�s �a���s��, от лат. corpus — тело; лат. callosum — жесткий)
—
массивный мост из волокон между правым и левым полушариями, состоящий
из более чем 100 млн нейронных аксонов. На срезе он окрашен в белый цвет, так
как аксоны покрыты белыми миелинизирующими клетками. См. гл. 1, «Некоторые
исходные положения»
морфема (от греч. morphe — форма) — наименьшая лингвистическая единица, ко-
торая может нести значимую информацию самостоятельно. В английском языке,
префиксы и суффиксы относят к морфемам (например, ‘pre-’ и ‘post-’). См. гл . 11,
«Введение»
мост — выступающая передняя часть ствола мозга. Мост передает сенсорную
информацию между мозжечком и передним мозгом и спинным мозгом, помогает
контролировать сон и бодрствование, отвечает за регуляцию дыхания. Также гене-
рирует сигналы во время БДГ-сна, которые интерпретируются как зрительно вос-
принимаемые повествовательные сны. См. гл . 5, «Развитие мозга от основания до
вершины»
мультистабильное восприятие – изменяющиеся зрительные образы при воспри-
ятии двойственных стимулов. См. гл . 6, «Связь деяельности мозга и зрительного
опыта»
мысленная гибкость — способность смещать когнитивную установку. Способ-
ность быстро отвечать на непредвиденные изменения в окружающей среде. См. гл .
12, «Детальное рассмотрение функций лобных долей»
мысленная ригидность (от лат. rigidus — негибкий) — неспособность быстро от-
вечать на непредвиденные изменения окружающей среды или переключать ког-
нитивную установку. Серьезные формы мысленной ригидности вызывают обсес-
сивно-компульсивное расстройство. Повреждения лобных долей часто вызывают
сильную мысленную ригидность. См. гл. 12, «Повреждения лобных долей»
надвисочная пластина — плоская область коры в Сильвиевой борозде, где распо-
лагается первичная и вторичная слуховая кора и часть зоны Вернике. См. гл . 5, «От
вопроса «где?» до вопроса «что?»: функциональная роль областей мозга»
неглект — вид повреждения правой височной доли мозга, при котором пациент не-
способен сознательно воспринимать левую половину поля зрения. См. гл . 6, «Обла-
сти мозга, необходимые для визуального восприятия: исследование повреждений»
неисчезаемость объектов — знание того, что воспринимаемые объекты продол-
жают существовать, даже если их нельзя увидеть или потрогать. Данный навык
развивается у детей с 7 месяцев. См. гл . 15, «Развитие мозга и сознания»
нейробласт (от греч. neuron — нерв; blastos — почка, побег) — делящаяся клетка в
раннем эмбриогенезе, которая дифференцируется в нейроны или глиальные клетки.
См. гл. 15, «Пренатальное развитие: от бластоцисты к младенцу»
Глоссарий
449
нейрогенез (от греч. neuron — нервная клетка; genesis — начало) — возникновение
и дифференциация нейронов из клеток-предшественниц. См. гл. 15, «Пренатальное
развитие: от бластоцисты к младенцу»
нейромедиатор (нейротрансмиттер; от греч. neuron — нерв; лат. trans — двигаться
сквозь) — химическое соединение, которое передает сигнал от одного нейрона к
другому через синаптическую щель. Некоторые нейромедиаторы упакованы в ве-
зикулы, которые собираются у мембраны пресинаптического нейрона около си-
напса и выделяют содержимое в синаптическую щель, где они связываются с ре-
цепторами на мембране постсинаптического нейрона. Выделение нейромедиатора
обеспечивается потенциалом действия в пресинаптическом нейроне. Существует
незначительное выделение медиатора даже в отсутствии потенциала действия. См.
гл.1,3и16
нейромодулятор (от греч. neuron — нерв; модулировать обозначает «влиять» или
«регулировать») — определенное нейрохимическое соединение, которое имеет ши-
рокое влияние в обширных областях мозга. Нейромодуляторы отличают от ней-
ромедиаторов, молекул, оказывающих локальный синаптический эффект. См. гл .
1и16
нейрон (от греч. neuron — нерв) — нервная клетка, которая передает информацию
посредством электрических сигналов. Нейроны являются центральным компонен-
том человеческого головного мозга, спинного мозга и периферических нервов. Су-
ществует множество типов нейронов, от сенсорных рецепторов и моторных единиц
и нейроэндокринных клеток до пирамидных нейронов, которые имеют длинные
аксоны, интернейронов с короткими древовидными отростками и большого коли-
чества специализированных клеток. См. гл . 3
нейронная доктрина (от греч. neuron — нерв; doctor — учитель) — теория, ав-
торство которой приписывают испанскому гистологу Сантьяго Рамон-и-Кахалу,
постулирующая, что «нервная система состоит из большого числа анатомически и
генетически самостоятельных нервных единиц (нейронов)». Это одно из основопо-
лагающих положений в науке о мозге в прошлом столетии. Однако открытие боль-
шого числа электрических синапсов (щелевых контактов) может поставить под со-
мнение некоторые аспекты нейронной доктрины. См. гл . 1, «Немного истории и
продолжающаяся дискуссия»
нейронный дарвинизм — теория, предложенная ученым Джеральдом Эдельма-
ном, которая постулирует, что нейроны образуют и создают связи между собой,
основываясь на принципах Дарвина. Во время биологической эволюции виды адап-
тируются при помощи воспроизведения, мутаций, которые приводят к возникнове-
нию различных форм, и селекции из возникающего спектра частично различных
организмов. Нейронный дарвинизм предполагает, что мозг развивается сходным
образом, как при воспроизведении, изменении и селекции развивающихся нейро-
нов, так и на поздних стадиях селекции синаптических связей. Мозг селектирует,
нежели просто следует инструкции, в отличие от программ вычислительного ком-
пьютера. См. гл . 3, «Адаптация и обучение массивов нейронов»
нейропептид (от греч. neuron — нерв; лат. peptidia — маленькие усваиваемые ча-
стицы) — пептид, который является короткой цепочкой аминокислот. Нейропепти-
450
Глоссарий
ды действуют как локальные нейромедиаторы или нейрогормоны, влияя на регуля-
цию аппетита, гормон роста и восприятие боли. См. гл . 1 и 3
нейропередача — электрохимическая сигнализация между нервными клетками.
См.гл.3и16
нейротоксин (от греч. neuron — нервная клетка; toxicum — яд) — химическое со-
единение, которое нарушает функционирование нервной системы, часто блокируя
передачу нервных импульсов. См. гл . 16
нейруляция — раннее развитие центральной нервной системы у зародыша по-
звоночных животных. См. гл . 15, «Пренатальное развитие: от бластоцисты к мла-
денцу»
неоднозначное восприятие — сенсорное событие, которое изменяется в зависи-
мости от двух перцептивных интерпретаций. См. гл . 6, «Связь деятельности мозга
и зрительного опыта»
неокортекс (от лат. neo — новая; греч. cort — кора) — самая большая и наиболее
видимая часть коры больших полушарий человека. Эта новая, с эволюционной точ-
ки зрения, кора противопоставляется древней коре медиальной височной доли,
гиппокампа и обонятельного мозга. См. гл . 1, «Немного истории и продолжающа-
яся дискуссия»
нервная трубка — цилиндрическая структура в развивающихся эмбрионах позво-
ночных, которая становится центральной нервной системой. См. гл. 15, «Прена-
тальное развитие: от бластоцисты к младенцу»
нижний (от лат. inferus — нижний) — внизу
норэпинефрин (от греч. epi — сверху; nephros — почка; -in — белок) (также на-
зывают норадреналином) — химическое вещество мозга, имеющее двойную роль в
качестве циркуляторного гормона и химического нейромедиатора. См. адреналин,
норадреналин. См. гл . 16
область интереса — область мозга, выбранная для изучения в исследованиях по
нейровизуализации, для того чтобы можно было сделать статистически достовер-
ные предположения об активности мозга в этой зоне. См. гл . 4
обратная связь — 1) в целенаправленных системах сигнал из окружающей среды,
указывающий на степень ошибок при достижении цели; 2) в нейронауке и психоло-
гии, сигнал окружающей среды, отражающий некое нейронное событие. Этот тип
нервной обратной связи часто позволяет людям обучаться контролировать спон-
танную нейронную активность; 3) в нервных сетях, поток информации, возвраща-
ющий выходной сигнал к входному слою сети. Некоторые теоретики четко разде-
ляют обратную связь и повторный вход сигнала в таламокортикальной системе
мозга. См. нейронный дарвинизм; см. гл . 3 и 6, «Теории зрительного осознания:
где оно происходит?»
обучение Хебба — согласно Дональду О. Хеббу, «нейроны, которые разряжаются
вместе, образуют между собой связь», т. е . два нейрона укрепляют свою синапти-
ческию связь, если они активизируются в один и тот же момент. Этот процесс фор-
мирует нейронные сети. Он был предложен Дональдом Хеббом в 1949 г., и носит
еще название правила Хебба. См. гл . 3, «Адаптация и обучение массивов нейронов»
Глоссарий
451
онтогенез (от греч. ontos — быть; geneia — начало) — развитие или ход развития
индивидуального организма. См. гл . 12, «Филогенез и онтогенез»
орбитофронтальная кора (от лат. orbis — круг, орбита, относится к части мозга,
расположенной непосредственно над глазницами или орбитами глаз; греч. cortex —
кора) — часть лобной доли. См. гл . 12, «Нарушения лобных долей»
орбитофронтальный синдром (от лат. orbis — круг, орбита, относится орбитоф-
ронтальной коре, расположенной непосредственно над глазницами или орбитами
глаз; греч. cortex — кора) — специфический синдром повреждения лобных долей,
при котором пациенты характеризуются наличием эйфории, гиперактивности и по-
терю импульсного контроля. См. гл . 12, «Нарушения лобных долей»
островок (от лат. «островок») — область, спрятанная в и под латеральной бо-
роздой, покрытая височными и теменными долями, выглядит как остров, если
осторожно оттянуть закрывающую его ткань. Островок отвечает за «интуитивные
чувства», такие как чувство тошноты и отвращения, и, вероятно, эмоциональные
чувства и влечения. См. гл . 5, «От вопроса «где?» до вопроса «что?»: функциональ-
ная роль областей мозга»
оттенение — экспериментальная парадигма для исследования селективного вни-
мания, при выполнении которой испытуемые должны немедленно повторять услы-
шанную устную речь. Тренировка позволяет испытуемым повторять сказанное с
периодом задержки меньше одной секунды. Метод оттенения требует очень много
ресурсов, так что другие потоки речи не могут быть восприняты в то же самое
время. См. гл . 8
Павлов, Иван Петрович (1849–1936) — русский физиолог. Получил Нобелевскую
премию по физиологии и медицине в 1904 г. за исследования пищеварительной
системы. Широко известен за описание классических условных рефлексов, когда
произвольный стимул, например звон колокольчика, становится сигналом биоло-
гического стимула, например пищи, и вызывает слюноотделение. Предположение
Павлова о том, что условные рефлексы являются основой обучения у человека,
в настоящее время отвергнуто. Однако павловские условные рефлексы широко
применяются в исследованиях и необходимы в клинических работах по изучению
предпочтения и отвращения пищи и, возможно, тревожных расстройств. См. гл . 1,
«Немного истории и продолжающаяся дискуссия»
палеокортекс (от греч. paleo — старый, древний; cortex — кора) — эволюционно
древняя область коры головного мозга, включающая медиальные височные доли,
обонятельную кору и гиппокамп. См. гл . 2, «Классическое понятие оперативной
памяти»
память места — память об определенном периоде времени, месте или условия,
когда произошло событие. См. гл. 9, «Виды памяти»
память — долговременные образы в мозге, которые отображаются в мышлении,
опыте или поведении. См. гл. 9
Панксепп, Джаак (р. 1943) — основатель аффективной нейронауки, исследова-
ния мозговой основы эмоций. Панксепп известен благодаря работам по изучению
смеха других млекопитающих (при щекотании), а также высокочастотного крика
бедствия, который возникает при разделении крыс-матерей и их детенышей. См. гл .
13, «Эмоциональная система мозга Панксеппа»
452
Глоссарий
парагиппокампальная область мест (ПГОМ, от греч. para — перед; см. гиппо-
камп) — область около гиппокампа, которая отвечает сильнее при предъявлении
пейзажей или зрительных сцен, чем при предъявлении отдельных изображений до-
мов или лиц. См. гл . 6, «Функциональная организация зрительной системы»
паралингвистический (от греч. para — около; лат. lingua — язык) — невербаль-
ный аспект лингвистического общения, учитывающий интонацию голоса, жесты,
социальную дистанцию и зрительный контакт. См. гл . 11, «Значимые утверждения»
париетальный неглект — состояние, при котором повреждение правой теменной
доли вызывает отсутствие сознательного восприятия левой половины эгоцентрич-
ного пространства. См. гл . 13, «Система страха»
патология — исследование фундаментальных причин заболеваний и, в особенно-
сти, структурных и функциональных изменений, вызванных этими заболеваниями.
См. гл. 12, «Нарушения лобных долей»
паттерн разряда — частота и картина потенциалов действия, которые могут пере-
давать важную информацию в мозге. См. гл. 3, «Массивы и карты»
первичная зрительная кора (или �1) — первая корковая зона зрительной систе-
�1) — первая корковая зона зрительной систе-
1) — первая корковая зона зрительной систе-
мы, расположенная в затылочной доле. См. гл. 6, «Функциональная организация
зрительной системы»
первичная моторная кора — область мозга, которая напрямую контролирует ске-
летную (произвольную) мускулатуру. Она соответствует моторному гомункулусу
и работает совместно с другими сенсорными и моторными областями, как премо-
торная кора
первичная соматосенсорная кора — сенсорный гомункулус (карта тела), рас-
положенный на постцентральной извилине коры больших полушарий, является
первичной корковой областью обработки чувствительности тела
передний (anterior, от ante — спереди от) — расположенный спереди от чего-либо.
См. гл. 1 и цветн. вклейку в начале книги
передняя комиссура — большой пучок нервных волокон, соединяющий два боль-
ших полушария мозга. См. гл . 5, «Введение»
передняя поясная кора (от лат. ante — перед, спереди от; cingulum — кушак, пояс;
cortex — кора) — передняя часть поясной коры. Передняя поясная кора вовлечена в
управление произвольными процессами. См. гл . 2,
периакведуктальное серое вещество (от греч. peri — около; лат. aqueductus —
водопровод) — серое вещество, окружающее проток (водопровод) в среднем мозге
и стволе мозга, по которому цереброспинальная жидкость циркулирует между же-
лудочками и спинным мозгом. См. гл . 13, «Система страха»
периферическая нервная система (ПНС) — обширная сеть нейронов за предела-
ми головного и спинного мозга. ПНС включает сенсомоторные нейроны, располо-
женные ниже шеи и нейроны автономной нервной системы, которые иннервируют
гладкую мускулатуру пищеварительного тракта, сердце и кровеносную систему.
См. гл. 5, «Введение»
персеверация (от лат. perseverare — продолжать) — симптом, включающий не-
уместное и бесконтрольное повторение определенных слов, фраз и жестов. См. гл .
10, «Эксплицитное решение задач»
Глоссарий
453
перспектива второго лица — межличностная точка зрения, что подразумевает
встречу двух субъективных взглядов, своего и другого лица. См. гл. 14, «Заключе-
ние»
перспектива первого лица — сознательная точка зрения на себя или «Я». См. гл .
14, «Заключение»
перспектива третьего лица — общественный взгляд на событие. Общественный
взгляд на определенную гипотезу необходим в науке. См. гл . 14, «Заключение»
перцептивная память —длительные изменения способности воспринимать окру-
жающий мир, например способность воспринимать звуки речи или способность
узнавать зрительные объекты при изменении ориентации и уровня освещенности.
См. гл. 2, «Классическая рабочая память»
перцептивное заполнение — основное свойство сенсорного восприятия, при ко-
тором мозг достраивает отсутствующие части зрительных объектов или сцен, часто
находящиеся за пределами прямого сенсорного входа. См. гл . 6, «Связь деятельно-
сти мозга и зрительного опыта»
пластичность (от греч. plastikos, от plassein — шаблон, форма) — способность
мозга адаптироваться и реорганизовать процессы при изменении требований окру-
жающей среды или потребностей, а также вследствие мозговой травмы. См. гл . 7,
«Восприятие музыки»
плодный алкогольный синдром (ПАС) — повреждение мозга у плода вследствие
потребления алкоголя матерью, сильная угроза для здоровья. См. гл . 15, «Прена-
тальное развитие: от бластоцисты к младенцу»
поведение, зависящее от окружения — нарушение лобных долей, при котором
пациенты имитируют действия осматривающего их врача или находят невозмож-
ным отказаться от использования расположенного перед ними предмета. См. гл. 12,
«Нарушения лобных долей»
повторный вход — в нейронном дарвинизме, резонансная петля между двумя ней-
ронами или группами нейронов, так что нейрон А активирует нейрон Б и наобо-
рот. Повторный вход может также наблюдаться и между популяциями нейронов.
Считается, что это основной механизм передачи сигналов между областями мозга,
который близко связан с мозговыми ритмами
подавление вспышкой — вариант теста на бинокулярное соревнование, в котором
изображение, предъявляемое в один глаз, подавляется вспышками, предъявляемы-
ми в другой глаз. См. гл. 8
позитронная эмиссионная томография (ПЭТ) (от лат. emitterre — испускать;
греч. tomos — срез; graphein — писать) — позитроны — положительно заряженные
субъядерные частицы, производимые ускорителем частиц. ПЭТ является низкора-
диоактивным методом визуализации, который позволяет получать вычисленные
карты строения мозга или тела, из которых можно создать трехмерное изображе-
ние. См. гл. 4
полушарная латерализация (от лат. lateralis — сторона) — степень, с которой
некоторые мозговые функции выполняются преимущественно одним полушарием
головного мозга, наиболее яркий пример, производство речи в левом полушарии у
большинства людей. См. гл . 5, 9 и 11
454
Глоссарий
поля Бродмана — около 100 корковых полей, выделенных и определенных немец-
ким неврологом Корбинианом Бродманом, исходно основывались на микроско-
пической анатомии нейронов в разных областях коры. Они до сих пор интенсивно
используются и в основном имеют различные функции. См. гл . 5, «Введение»
последовательная группировка — один из способов организации слуховой си-
стемы человека звуков в персептивно значимые элементы. Если свойства звуков
повторяются в той же последовательности, они могут быть сгруппированы вместе.
См. гл. 7, «Функциональное картирование обработки слуховой информации»
постцентральная извилина — выступающая складка в теменной доле мозга
человека, расположенная непосредственно за центральной бороздой. Она содер-
жит первичную соматосенсорную кору, первую область коры, отвечающую на
чувствительность тела, или соматосенсорный гомункулус, который отображает
противоположную или контралатеральную половину тела
потенциал действия — в нейронах электрохимический сигнал возникает около
сомы и распространяется по аксону к синаптической терминали. Также его называ-
ют «спайком» или «нейронным разрядом». См. гл . 3, «Введение»
поясная (цингулярная) кора (от. лат. cingulum — пояс; cortex — кора) — часть
коры на медиальной (внутренней) поверхности каждого полушария. Она участвует
в исполнительных функциях, разрешении конфликтных целей и эмоциях
префронтальная кора (от лат. prae — спереди; frons — лоб) — большая, передняя
часть лобных долей, не включающая моторную кору. Префронтальная кора отве-
чает за произвольные функции, и содержит зону Брока, «орган цивилизации». См.
гл. 2, «Классическое понятие оперативной памяти»
продольная борозда — глубокая щель, разделяющая правое и левое полушария
мозга позвоночных. См. гл . 1, «Некоторые исходные положения»
промежуточный мозг (диэнцефалон, от греч. dia — через; enkephalos — мозг) —
часть мозга, содержащая таламус, гипоталамус и заднюю часть гипофиза
проприоцепция (от лат. proprius — свой; perception — восприятие) — чувство о
себе, включающее восприятие своего местоположения в пространстве, основанное
на внутренних и внешних сенсорных входах
пространство задачи — схема возможных решений, часто строится в форме дере-
ва. См. гл . 10, «Эксплицитное решение задач»
протеом (по аналогии с геномом) — совокупность всех белков, экспрессируемых
геномом
процедурная память (от лат. procedure — способ делать что-то) — форма импли-
цитной памяти эквивалентная памяти навыков или знанию того, как выполнить
задачу. По всей видимости, является преимущественно неосознанной. Эта память
часто очень долговременная
прямая связь — 1) переход сигнала от простых к более сложным стадиям обра-
ботки; 2) в сенсомоторной системе подготовка внутренней траектории реакции для
получения более точной обратной связи, когда действие выполнено. Эта стратегия
используется в контроле быстрых движений у птиц и людей, а также в машинах,
например в аэродинамически нестабильных реактивных самолетах; 3) в нервных
сетях переход информации от ранних к более высоким слоям сети
Глоссарий
455
Пэнфилд, Уайлдер (1891–1976) — нейрохирург и ученый из Монреаля, кото-
рый выполнил пионерскую работу по хирургическому лечению эпилепсии. Перед
операциями он проводил исследовательскую стимуляцию мозга у бодрствующих
пациентов (которые были избавлены от боли с помощью локальной анестезии
хирургической поверхности). Таким образом, пациенты могли давать отчет об ощу-
щениях от электрической стимуляции мозга. Пэнфилд с коллегами смогли опреде-
лить функции областей мозга, которые ранее изучали только при помощи посмерт-
ных исследований или при повреждениях. См. гл . 4, «Ряд полезных инструментов»
рабочая память — когнитивная способность хранить и использовать в течении
10–30 с новую информацию. Рабочая память включает центральный исполни-
тель, рабочее хранение, вербальное повторение и зрительно-пространствен-
ный скетчпад. См. гл . 2, «Классическое понятие оперативной памяти»
разум — аспекты интеллекта и сознания, которые обнаруживаются в мышлении,
восприятии, памяти, эмоциях, воле и воображении, включая все сознательные и
бессознательные когнитивные процессы мозга. См. гл . 1, «Немного истории и про-
должающаяся дискуссия»
Ракич, Паско (р. 1933) — нейроанатом, который показал, что нейронная миграция
осуществляется радиально и рострально, как расходящиеся спицы движущегося
колеса. См. гл . 15, «Пренатальное развитие: от бластоцисты к младенцу»
Рамон-и-Кахал, Сантьяго (1852–1934) — испанский исследователь-первооткры-
ватель в области микроскопического изучения мозга. Его часто считают родона-
чальником науки о мозге. Существует большое число его детальных рисунков сре-
зов мозговой ткани. См. гл . 1, «Немного истории и продолжающаяся дискуссия»
распространение неоновых цветов — иллюзия восприятия, при которой белая
область вблизи цветных или черных объектов кажется раскрашенной. См. гл . 6,
«Связь деятельности мозга и зрительного опыта»
репликация — 1) копирование генома при наследовании; 2) копирование экспе-
римента с использованием точно таких же методов для верификации полученных
результатов на новой группе испытуемых
ретикулофронтальный синдром разобщения (от лат. reticulum — сеть; frons —
лоб) — нарушения произвольных функций, вызванные повреждением путей, со-
единяющих лобную кору с сетчатыми (ретикулярными) структурами ствола мозга
и среднего мозга. См. гл . 12, «Нарушения лобных долей»
ретикулярная формация (от лат. reticulum — сеть) — часть мозгового ствола, ко-
торая вовлечена в регуляцию цикла сон–бодрствование и многих других функций.
Она получает коллатеральные входы от всех сенсорных и моторных систем, а также
от высших отделов мозга. Эволюционно это одна из самых древних частей мозга.
См. гл . 5, «От вопроса «где?» до вопроса «что?»: Функциональная роль областей
мозга»
ретровирус (от лат. re tro — назад; virus — яд, отравлять) — вирус, который функ-
ционирует в обратном направлении от обычного пути ДНК–РНК–белок. Ретрови-
русы являются РНК-паразитами, которые могут встраиваться в ДНК клетки-хозяи-
на. Вирус ВИЧ является первым детально изученным ретровирусом. Ретровирусы
используются для того, чтобы вставлять или удалять определенные генные после-
довательности в клеточную ДНК
456
Глоссарий
ретроградная амнезия (от лат. retrogrades — идти назад; греч. a -mnesia — без
памяти) — вид потери памяти о событиях, произошедших до мозговой травмы.
Противопоставляют с антероградной амнезией. См. гл . 9, «Амнезия»
рефлекс переключения — метод, основанный на использовании классических ус-
ловных рефлексов, когда животное сначала обучают бояться одного стимула и не
испытывать страх при предъявлении другого. На следующей стадии эти рефлексы
меняют местами, и животное должно приспособиться к быстрому переключению
стимулов, определяющих болевое раздражение. См. гл . 13, «Система страха»
рефлекторная цепь — или рефлекторная дуга, является относительно простым пу-
тем, обеспечивающим рефлекторное поведение. Наиболее известный пример — ко-
ленный (или пателлярный) спинальный рефлекс, который возникает даже в случае
отделения спинного мозга от головного. Однако спинальные рефлексы достаточны
быстры, сложны и координированны и могут вовлекать ствол мозга и вестибуляр-
ную (балансирующую) систему, как в случае переворота тела кошки при падении.
Обычно рефлексы функционируют в близкой связи с произвольной деятельностью,
осуществляемой через лобные доли, мозжечок и базальные ядра. Мозговые рефлек-
сы, такие как зрачковый рефлекс, находятся под взаимным контролем автономной,
зрительной и эмоциональной областей мозга. См. гл. 3, «Рабочие гипотезы»
рецептивное поле (от лат. recipere — брать) — рецептивным полем в зрительной
системе, например, является область зрительного поля, которая может активиро-
вать или подавить разряды клетки. Рецептивное поле рецептора сетчатки, таким
образом, отличается от рецептивных полей нейронов более высоких уровней, уча-
ствующих в обработке движения или идентификации зрительных объектов. Анало-
гичные рецептивные поля были обнаружены для зрительного внимания в теменной
коре. Рецептивные поля также существуют и для других сенсорных систем, таких
как слуховая и соматосенсорная. См. гл . 3 и для более детального рассмотрения
рецептивных полей в зрительной системе см. гл . 6
саггитальный (от лат. sagittal — стрела) — любое сечение мозга, проходящее па-
раллельно медиальному или срединному срезу. См. гл . 1, «Некоторые исходные
положения»
селективное внимание — способность концентрировать внимание на одном
аспекте окружающей среды, в то время как остальные игнорируются. Оно возни-
кает произвольно, как, например, принятие решение о чтении интересной книги в
шумном автобусе, или когда сенсорное восприятие направлено на биологически
или персонально значимый стимул. См. гл . 2 и 8
селекционизм — теория мозга, построенная на основе нейронного дарвинизма. Во
время биологической эволюции виды адаптируются при помощи размножения, му-
таций, которые приводят к появлению новых форм, и селекции между полученными
вариантами немного различающихся особей. Нейронный дарвинизм предполагает
существование похожего метода селекции растущих нейронов и их синаптических
связей. См. гл . 3, «Адаптация и обучение массивов нейронов»
семантика — область науки о значениях в языке. См. гл . 11, «Введение»
семантическая память — вид декларативной памяти, который включает значе-
ния, фактические верования, категории и другое общее знание, выходящее за преде-
лы собственного опыта. См. гл. 2, «Классическое понятие оперативной памяти»
Глоссарий
457
семантический дефицит — форма афазии, заключающаяся в потере определенной
категории значений, например названий животных. См. гл . 7, «Восприятие речи»
сенсорная система — часть нервной системы, ответственной на обработку сен-
сорной информации. Сенсорная система состоит из чувствительных рецепторов,
проводящих путей и, преимущественно, задней коры, вовлеченной в восприятие
сенсорных стимулов. Классические виды чувствительности имеют и подвиды:
боль и щекочущее чувство, запуск световыми рецепторами мелатонина как гормо-
на, регулирующего сон, чувство равновесия и другие. Не все сенсорные системы
находятся под сознательным контролем; кровяное давление, например, очень редко
бывает осознано. Классические системы чувств начинаются с рецепторной поверх-
ности, содержащей многие миллионы рецепторов, например сетчатка или базиляр-
ная мембрана. См. гл . 2, «Введение»
серое вещество — внешние слои коры больших полушарий для невооруженного
взгляда. Серое вещество состоит из тел десятков миллионов нейронов, которые
посылают белые аксоны во всех направлениях вниз. См. белое вещество. См. гл .
5, «Введение»
серотонин (от лат. serum — кровь, tonus — давление) — нейромедиатор, впервые
открытый как гормон, регулирующий кровяное давление. У серотонина множество
функций в головном и спинном мозге, а нарушения системы серотонина считаются
ответственными за некоторые заболевания
сетчатка — слой световых рецепторов, расположенных на внутренней поверхно-
сти глаза. Светочувствительные рецепторы сетчатки (палочки и колбочки) запуска-
ют химическую реакцию, которая приводит к изменению электрического потен-
циала на мембране клетки. Это может запустить активность ганглиозных клеток
сетчатки, аксоны которых образуют зрительный нерв, оканчивающийся в зритель-
ных релейных ядрах таламуса. См. гл . 6, «Функциональная организация зритель-
ной системы»
сеть — 1) двумерная решетка сенсорных рецепторов, например сетчатка; 2) двух-
мерная сетка элементов более высокого уровня в зрительной иерархии. Многие
«слоистые» структуры мозга можно представить в виде таких сетей. См. гл. 3,
«Массивы и карты»
Сильвиева борозда или латеральная борозда — хорошо различимая борозда,
отделяющая лобную и теменную доли от расположенной снизу височной доли.
См. гл. 5
симультанная группировка — если два звука имеют схожее начало и конец, они
могут быть сгруппированы. Это один из способов организации слуховой системой
человека звуков в значимые элементы. См. гл . 7, «Функциональное картирование
обработки слуховой информации»
синапсы — это межнейрональные контакты с крошечными зазорами с мембраной
следующего нейрона, благодаря которым нейроны обмениваются информацией
при помощи химических медиаторов. Синапсы являются основными вычисли-
тельными элементами мозга как точки контроля за движением для потоков инфор-
мации. В мозге насчитывают десятки миллиардов нейронов, но более нескольких
триллионов синапсов
458
Глоссарий
синаптическая щель — зазор между двумя нейронами, которые передают друг
другу информацию посредством нейромедиаторов. См. гл . 3
синаптический прунинг — селективная потеря синапсов в мозге, когда некоторые
возможные связи не используются. См. обучение Хебба, нейронный дарвинизм.
См. гл. 15, «Развивающийся мозг: изменения длиною в жизнь»
синаптогенез — формирование синапсов в мозге. См. гл . 15, «Развивающийся
мозг: изменения длиною в жизнь»
синдром Балинта — мозговое нарушение, которое характеризуется симультанной
агнозией, неспособностью воспринимать два или более объектов за один раз. Мо-
жет также включать неспособность указывать на объект и произвольно смещать
взор. См. гл . 6, «Области мозга, необходимые для визуального восприятия: иссле-
дование повреждений»
синдром ломкой хромосомы Х — генетическое расстройство, сопровождающее-
ся рядом физических, когнитивных, эмоциональных и поведенческих нарушений.
Наиболее частая причина умственной отсталости. См. гл . 16
синдром нарушения внимания (гиперактивность) (СНВ, СНВГ) — устойчивый
характер импульсивности и сложности поддерживать длительное произвольное вни-
мание, с компонентом гиперактивности или без него. Существует спор, является ли
это истинным заболеванием или нормальной адаптацией, которая может быть очень
сложна на сегодняшний день. Он затрагивает от 3 до 5% детей во всем мире и сохра-
няется во взрослом возрасте у 30–50% этих людей. См. гл . 12, «Дефицит внимания и
гиперактивность»
синтаксис — правила и закономерности предложений в естественном языке. См.
гл. 11, «Введение» и «Синтаксис, включение и синтагматизация»
синтаксический анализ — идентификация грамматической структуры на основе
слов, фонем и морфем. См. гл . 11, «Введение»
система зеркальных нейронов — предполагаемая система, образованная зер-
кальными нейронами в мозге человека, которая, как считают, необходима для со-
циальной когнитивной деятельности. См. гл . 14
скотома — поврежденная или утраченная часть сетчатки или нейронной сети зри-
тельного поля высшего уровня. Скотомы обычно замещаются информацией от со-
седних с поврежденной областью нейронов. См. гл . 6
слепозрение — разновидность мозговой травмы, при которой пациент дает отчет о
некоторых зрительных стимулах без возникновения у него субъективного ощуще-
ния о том, что он их видел, вследствие повреждения первичной корковой области
зрительной системы, поля �1. См. гл . 6, «Области мозга, необходимые для визуаль-
�1. См. гл . 6, «Области мозга, необходимые для визуаль-
1. См. гл . 6, «Области мозга, необходимые для визуаль-
ного восприятия: исследование повреждений»
слепота невнимания — заслуживающий доверия экспериментальный феномен,
при котором человек не может видеть вещи, которые в обычной жизни четко раз-
личимы. См. гл . 8
словесная глухота — неспособность понимать смысл устной речи, с сохранением
способности слышать звуки, говорить, читать и писать. Связано с двусторонним
повреждением верхней задней части височных долей или их подкорковых связей.
См. гл. 7, «Восприятие речи»
Глоссарий
459
слуховая кора — часть коры больших полушарий, вовлеченная в процесс обра-
ботки звуков, например зона Вернике и извилина Гешля. См. гл. 7, «Центральная
слуховая система»
сознание (��ns�i��sness, от лат. con — вместе; scientia — знание) — психическая
ориентированность, бодрствование. Сознание подразумевает нахождение в воспри-
имчивом и легко реагирующем состоянии по отношению к окружающей среде, в
отличие от состояний сна или комы. Синонимы включают эксплицитное мышление
и фокальное внимание. См. гл . 1, «Немного истории и продолжающаяся дискуссия»
и «Возвращение сознания в науку»; и гл. 8
спектрограф (от лат. spectrum — появление; греч. graphon — писать) — прибор,
сконструированный во время Второй мировой войны в Bell Telephone Laboratories
для анализа звуковых сигналов и построения спектрограммы, которая показыва-
ет частоту звука во времени. См. гл . 7, «Функциональное картирование обработки
слуховой информации»
спектроскопия ближнего инфракрасного света (от лат. infra — снизу, внутри;
греч. erythros — красный) — метод измерения световых волн, расположенных ниже
красного цвета в спектре длин волн (около 800 нм до 2500 нм). Его используют
для измерения количества гемоглобина в крови как показателя местной активности
мозга. См. гл. 15, «Развитие мозга и сознания»
специфичность доменов — функциональная специфичность областей мозга или
механизмов. Идея о том, что каждая когнитивная функция может иметь свою об-
ласть или нейронную сеть в мозге, в противоположность представлениям об общих
механизмах для множества когнитивных функций. См. гл . 12, «Детальное рассмо-
трение функций лобных долей»
специфичность процессов — утверждение, что когнитивные процессы могут за-
трагивать разное число различных областей мозга. См. гл . 12, «Детальное рассмо-
трение функций лобных долей»
среднесаггитальный (от лат. sagitta — стрела) — срез по средней линии, идущий
от носа к затылку. См. гл . 1, «Некоторые исходные положения»
ствол мозга — нижняя часть мозга, соединенная со спинным мозгом. Все основ-
ные двигательные и сенсорные пути проходят через него, включая зрительный и
слуховой нервы. Ствол мозга также регулирует сердечно-сосудистую и дыхатель-
ную системы, поддерживает состояние бодрствования, медленноволновый сон
(МВС) и БДГ-сон. См. гл . 5, «Развитие мозга от основания до вершины»
таламокортикальная система (от греч. thalamus — палата; cort — кора) — цен-
тральный переключатель мозга, включающий кору и таламус и позволяющий по-
току сигналов свободно перемещаться в обоих направлениях. См. гл . 5, «Введение»
таламус (от греч. thalamus — комната, палата) — парная симметричная структура
с формой яйца, являющаяся основным переключателем информации для коры и ре-
гулятором потока информации в таламокортикальной системе. См. гл. 1, «немного
истории и продолжающаяся дискуссия»
теменная доля — большая область мозга, расположенная над затылочной долей
и сзади от лобных долей. Теменная область интегрирует сенсорную информацию
разной модальности и содержит постоянно обновляющиеся карты расположения
тела и окружающих объектов. См. гл . 1 и 5
460
Глоссарий
теория разума — способность принимать мысленное состояние (убеждения, по-
требности, намерения) других людей. См. гл. 14, «Заключение»
теория симуляции — предположение, что люди иногда мысленно симулируют или
имитируют то, что, по их мнению, испытывают другие, для лучшего понимания.
См. гл. 14, «Заключение»
теория — представления о развитии детьми имплицитных теорий о других людях,
а также развитие учеными теорий после многолетних исследований и развития. См.
гл. 14, «Заключение»
тератоген — химический или иной фактор, вызывающий нарушения развития. См.
гл. 15, «Пренатальное развитие: от бластоцисты к младенцу»
терминальный — дистальный конец аксона. См. гл . 3, «Введение»
тест Струпа — тест, названный в честь американского психолога Джона Ридли
Струпа, который впервые описал данный феномен на английском языке в 1935 г.
Когда название цвета, например синий, зеленый или красный, написано цветом,
отличным от значения написанного цвета (например, слово «красный» написано
синими чернилами), испытуемый испытывает сложности называть цвет слова и де-
лает это медленнее и более склонен ошибаться, чем в случае совпадения цвета и
смысла написанного слова. Этот феномен еще известен как эффект Струпа. Эффект
Струпа используется для активации областей конфликтов в мозге и объясняет схо-
жие парадигмы, такие как «эмоциональный тест Струпа». См. гл . 8
тета-волны — регулярные электромагнитные волны с частотой 3,5–7,5 Гц. Одна
из ролей тета является координация гиппокампального кодирования памяти с кор-
ковыми сайтами синаптического хранения памяти. Тета также участвуют в обрат-
ном процессе извлечения эпизодической памяти. Считается, что тета-ритм вовле-
кает синхронные разряды многих нейронов, осуществляемые холинергической
модуляцией
тормозный контроль (от лат. in- —
не; habēre — иметь, полагать) — нервные им-
пульсы, которые снижают или прекращают определенную активность или ответ.
См. гл. 12
транскраниальная магнитная стимуляция (ТМС) — относительно неинва-
зивный метод, использующий сильные электромагнитные поля около головы для
стимуляции или подавления корковых нейронов. ТМС имеет хорошее временное и
пространственное разрешение. См. гл. 4, «Ряд полезных инструментов»
транскрипция (от. лат. trans — поперек; scribere — писать) — копирование ДНК
в матричную РНК. См. гл. 16
трансляция — перевод матричной РНК в активные белки
триединый мозг (от лат. tri — три) — модель, предложенная Полем МакЛином для
того, чтобы охарактеризовать мозг человека с точки зрения трех основных эволю-
ционных стадий развития, включает комплекс пресмыкающихся, лимбическую си-
стему (млекопитающих) и неокортекс (приматов и других млекопитающих с боль-
шим мозгом, с сильным развитием лобных долей у людей). См. гл . 13
умозаключение — приведение выводов, основанных на знаниях, а не на прямом
наблюдении. См. гл . 1, «Некоторые исходные положения»
Глоссарий
461
Уэринг, Клайв (р. 1938) — выдающийся британский классический музыкант, пе-
ренесший в сорок лет вирусное поражение мозга, которое разрушило оба гиппо-
кампа и некоторые области лобной коры. Случай Уэринга стал широко известен
благодаря стараниям его жены, Деборы Уэринг, поднять общественный интерес к
таким медицинским проблемам. Уэринг живет в сиюминутном мире, без способно-
сти хранить информацию для последующего извлечения. Несмотря на проблемы с
памятью, он способен играть на пианино и дирижировать исполнением музыкаль-
ных произведений, которые он знал до травмы См. антероградная амнезия. См.
гл. 2, «Классическая понятие оперативной памяти»
фенотип (от греч. phainein — показывать; typos — тип) — любой физиологический
признак организма, который является оттиском ДНК или генотипа. См. гл . 16
Фехнер, Густав (1801–1887) — немецкий физик, математик и родоначальник пси-
хофизики. Фехнер утверждал, что решил спор тела с разумом, представив общую
логарифмическую взаимосвязь между субъективной сенсорной интенсивностью и
физической интенсивностью стимула для многих сенсорных модальностей. См. гл .
1, «Немного истории и продолжающаяся дискуссия»
филогенез (от греч. phylon — раса; geneia — исток) — эволюционная история раз-
вития отдельного вида организмов или генетически близкой группы организмов от
одного предшественника. См. гл . 12, «Филогенез и онтогенез»
фланкерный тест — метод изучения зрительного внимания, при котором испы-
туемого просят реагировать как можно быстрее на предъявление стимула в цен-
тральной точке фиксации или в точке, расположенной от центра на определенном
расстоянии. Целевой стимул сопровождается предъявлением отвлекающих сти-
мулов, например букв или стрелок. Фланкерный тест проводит количественную
оценку скорости испытуемого и точности выполнения перемещения внимания в
ожидаемую и неожидаемую точку появления стимула. Перемещение внимания в
неожиданную точку требует исполнительного внимания, так как необходимо от-
вергнуть подготовленное, ожидаемое перемещение. Фланкерный тест обычно тре-
бует от испытуемого подавлять произвольные движения глаз, так что изменение в
точности и скорости ответа на стимул является исключительно результатом импли-
цитного смещения внимания. См. гл. 8
фонема (от греч. phone — звук) — в речи человека наименьшая лексически само-
стоятельная категория звуков, например гласные и согласные звуки. См. гл . 7, «Вос-
приятие речи»
фонологический дефицит — вид афазии, при которой звуки речи не могут быть
идентифицированы как фонемные категории. См. гл . 7, «Восприятие речи»
фосфен (от греч. phos — свет; phainein — показывать) — световые точки в зри-
тельном поле, которые возникают при механической, электрической или другой
стимуляции сетчатки или зрительной коры. Отчет о фосфенах также дают и меди-
тирующие люди при сенсорной изоляции и под влиянием наркотиков, например,
галлюциногенов. См. гл . 6
Фрейд, Зигмунд (1856–1939) — австрийский врач-невролог, основавший школу
психоанализа в психологии, оказавший также значительное влияние на культуру.
Фрейд широко известен благодаря своей теории о бессознательном и созданию
клинической практики по психоанализу. Этот исследователь нервной системы
462
Глоссарий
разработал раннюю модель нервной сети и открыл новый химический краситель
(хлорид золота), позволявший отчетливо выделять под микроскопом определенные
нейроны ствола мозга. Несмотря на то что многие идеи Фрейда перестали поль-
зоваться популярностью, некоторые мозговые феномены, например роль лобных
долей в регулировании эмоциональных импульсов и конфликтов между целями,
хорошо согласуются с его взглядами. См. гл . 1, «Немного истории и продолжающа-
яся дискуссия»
функциональная избыточность —встроенные дублирующие функции в системе
для предотвращения полного сбоя функционирования в критических ситуациях.
Например, у млекопитающих есть два легких: если одно выйдет из строя, для ды-
хания останется еще одно. Мозг имеет множество избыточных способностей. См.
гл. 3, «Функциональная избыточность»
функциональная магнитно-резонансная томография (фМРТ) — особый тип
магнитно-резонансной томографии (МРТ), который измеряет локальное снабже-
ние кислородом ткани, связанное с нервной активностью в определенных областях
мозга. Благодаря фМРТ существует когнитивная нейронаука. См. гл. 4, «Введение»,
«фМРТ и ПЭТ: Непрямое измерение нейронной активности»
функциональная фиксированность — когнитивная установка, которая блокирует
у человека новые варианты действий, восприятия или решения проблем. См. гл . 10,
«Эксплицитное решение задач»
функция передачи звука — метод выявления местоположения звука в звуковом
пространстве. См. гл. 7, «Функциональное картирование обработки слуховой ин-
формации»
хроматин (от греч. chroma — цвет) — вещество в хромосомах каждой клетки,
включающее ДНК, немного РНК, белки и регуляторные молекулы, которые изме-
няют экспрессию ДНК
центральная борозда — эта складка коры больших полушарий, разделяющая
теменные доли от лобных долей, а также первичную моторную кору от пер-
вичной соматосенсорной коры. Центральная борозда четко разделяет входные и
выходные области мозга. См. гл . 1, «Некоторые исходные положения»
центральная догма молекулярной биологии — гипотеза, согласно которой ге-
нетическая информация реализуется только в одном направлении, от ДНК к ма-
тричной РНК, а затем к активным белкам. Центральная догма была впервые сфор-
мулирована Френсисом Криком в 1958 г. и была модифицирована более поздними
открытиями, свидетельствующими о наличии многочисленных цепей, которые дей-
ствуют в обратную сторону. Однако ДНК является хорошо защищенной молекулой,
которая очень редко изменяется, хотя ее экспрессия постоянно регулируется — за-
пускается или блокируется эпигенетическими событиями. См. гл . 15 и 16
центральная нервная система (ЦНС) — головной и спинной мозг. Все нейроны
за пределами ЦНС относят к периферической нервной системе (ПНС). См. гл . 5,
«Введение»
центральный исполнитель — мозговые процессы планирования, принятия реше-
ния, абстрактного мышления, понимания правил, инициации и торможения дей-
ствий, решения конфликтов целей и гибкого контроля над вниманием. Эти функции
Глоссарий
463
относятся к рабочей памяти и вовлекают лобные доли. См. гл . 2, «Классическое
понятие оперативной памяти» и «Центральный исполнитель»
цереброспинальная жидкость (ЦСЖ) — внутренняя циркуляция в спинном и го-
ловном мозге. ЦСЖ обусловливает защищенный обмен молекулами и клетками, не
связанный с кровеносной системой
чанкинг — эффективный способ использовать лимитированную кратковременную
память, с превращением больших объемов знаний в маленькие символические еди-
ницы, правила или закономерности, или «кусок», «чанк». В естественном языке
существительные можно рассматривать как чанки, так как они позволяют нам об-
ращаться к большому объему информации при помощи единичных слов. См. гл . 2,
«Ограниченость функциональных способностей»
Черчланд, Патриция (род. 1943) — канадско-американский философ, которая вы-
играла премию Макартура в 1991 г. «Нейрофилософия» Черчланд заключается в
том, что популярные понятия о разуме необходимо будет пересматривать по мере
того, как мы будем узнавать о лежащем в их основе функционировании мозга. См.
гл. 1, блок 1.2
чтение мыслей — попытка соотнести свои действия с мыслями других. В боль-
шинстве случаев много людей неточно понимают мысли других людей. См. гл. 14,
«Заключение»
щелевой контакт (или электрический синапс) — прямое соединение двух мембран
нейронов или глиальных клеток. Ранее считали, что такие контакты очень редки, но
сейчас полагают, что они существуют во всем мозге, особенно в ГАМК-ергических
синапсах в верхнем слое коры
Эдельман, Джеральд М. (род. 1929) — американский иммунолог и нейробиолог,
который получил Нобелевскую премию за работу о структуре молекул антител.
Эдельман разработал теоретическую основу для нейронного дарвинизма, которая
накладывает принципы селектирования Дарвина на мозг, в отличие от принципов
вычислительных компьютеров. См. гл. 3 и 16
эксайтотоксичность (от лат. excitare – поднимать; toxicum — яд) — повреждение
нейронов, вызванное поступлением глутамата, обычного возбудительного нейро-
медиатора. Эксайтотоксичность является основной причиной мозговой дегенера-
ции и после травматических повреждений. См. гл . 16
эксплицитная память (от лат. explicitus — ясный) — тип памяти, включающий
сознательное, намеренное воспроизведение памятных следов и опыта. См. импли-
цитная память, имплицитное обучение. См. гл . 2, «Классическое понятие опера-
тивной памяти», и гл. 9, «Введение»
электроэнцефалография (ЭЭГ) (от греч. ēlectron — солнечный свет; en- + kephalē
—
в голове; graphein — писать) — электрическая активность, которая, как правило,
регистрируется с поверхности скальпа и иногда с поверхности коры, отражающая
электромагнитные поля огромного числа активных нейронов. См. гл . 4, «Ряд полез-
ных инструментов», и приложение
эмпатия (от греч. empatheia — страсть) — способность разделять чувства другого
и понимать его. См. гл. 14, «Заключение»
энзим (фермент, от греч. en - + zyme — дрожжи) — молекулы, которые ускоряют
химические реакции
464
Глоссарий
эпигенез (от греч. epi — после; genesis — рождение, исток) — факторы, отличные
от ДНК, которые влияют на развитие клеток во время внутриутробного развития
(беременности) и после рождения. В отличие от центральной догмы молекулярной
биологии, по которой ДНК кодирует транспортную РНК, приводя к образованию
белков для строительства и функционирования всех клеток, эпигенез подразумевает
существование обратного влияния. Например, многочисленные физиологические
факторы и факторы окружающей среды могут приводить к экспрессии некоторых
генов (ДНК) или ее подавлению. ДНК является основной молекулой, кодирующей
фенотип, дающей план развития вида от одного поколения к другому. Но внешние
факторы могут влиять на активацию и торможение ДНК, как выключатели. См. гл .
15и16
эпигенетика (от греч. epi — после; genesis — рождение, исток) — изменение экс-
прессии генов, вызванное другими механизмами, нежели прямой экспрессией ДНК
через матричную ДНК и соответствующие белки. См. гл . 15 и 16
эпизодическая память (от греч. episeidos — вставка) — память на осознанные
события, особенно те, которые могут быть явно воспроизведены, как время, место,
события, ассоциативные эмоции и другая контекстная информация. Формирование
новой эпизодической памяти требует участия средней височной доли, особенно
области гиппокампа и связи его с корой больших полушарий. См. гл . 2 и 9
эхолалия (от греч. ēchō — повторять; laliá — лепетание) — постоянная и не кон-
тролируемая имитация речи других. Эхолалия может наблюдаться при аутизме,
синдроме Туретта, афазии, пороках развития, шизофрении, синдроме Аспергера,
болезни Альцгеймера и других. См. гл. 12, «Нарушения лобных долей»
эхопраксия (от греч. ēchō — повторять; praxis — действие) — неврологический
симптом, включающий повторяющуюся, непроизвольную имитацию движений
других людей. См. гл . 12, «Нарушения лобных долей»
Минимальные системные требования определяются соответствующими требованиями про-
граммы Adobe Reader версии не ниже 11-й для операционных систем Windows, Mac OS,
Android, iOS, Windows Phone и BlackBerry, экран 10"
Учебное эл ектронное издание
Серия: «Лучший зарубежный учебник»
МОЗГ, ПОЗНАНИЕ, РАЗУМ: ВВЕДЕНИЕ В КОГНИТИВНЫЕ НЕЙРОНАУКИ
В двух частях
Часть вторая
Ведущий редактор канд. биол. наук В. В . Гейдебрехт
Редактор канд. биол. наук О. Б. Мацелепа. Художник Н. А. Новак
Технический редактор Е. В. Денюкова. Компьютерная верстка: В. А. Носенко
Подписано к использованию 25.07.14 .
Издательство «БИНОМ. Лаборатория знаний»
125167, Москва, проезд Аэропорта, д. 3
Телефон: (499) 157-5272, e-mail: binom@Lbz.ru, http://www.Lbz.ru
Книга знакомит читателя:
• с с овременными методами визуализации мозга, позволяющими перейти на
новый уровень наблюдения (в реальном времени) и изучения когнитивных
механизмов и механизмов восприятия;
• с новейшими разработками в области молекулярных основ сознания;
• с результатами новых методов регистрации активности мозга, позволяющими
утверждать, что мозговые ритмы несут информацию как о сознательных, так
и о бессознательных процессах;
• с современными представлениями о строении префронтальной коры, на осно"
вании которых пересмотрена роль лобных долей, считавшихся непостижимыми.
Рассмотрены все возможные нарушения мозговой деятельности: от случаев
повреждения памяти до возникновения слепого зрения, гемиагнозии, прозо"
пагнозии и других расстройств зрительной системы.
Вы узнаете:
• почему люди иногда спонтанно начинают повторять движения за другим чело"
веком и не могут остановиться;
• почему профессиональные музыканты иногда не в состоянии подавить желание
петь во время игры на инструменте;
• как спортсмены и музыканты, используя сенсомоторные области коры голов"
ного мозга, осуществляют мысленные тренировки;
• почему ученые считают, что сознание сегодня является законной и разрешимой
научной проблемой;
• как возникают эмоции и чем обусловлено наше социальное поведение.
Для студентов и специалистов по психологии, биологии, педагогике и медицине.