










































































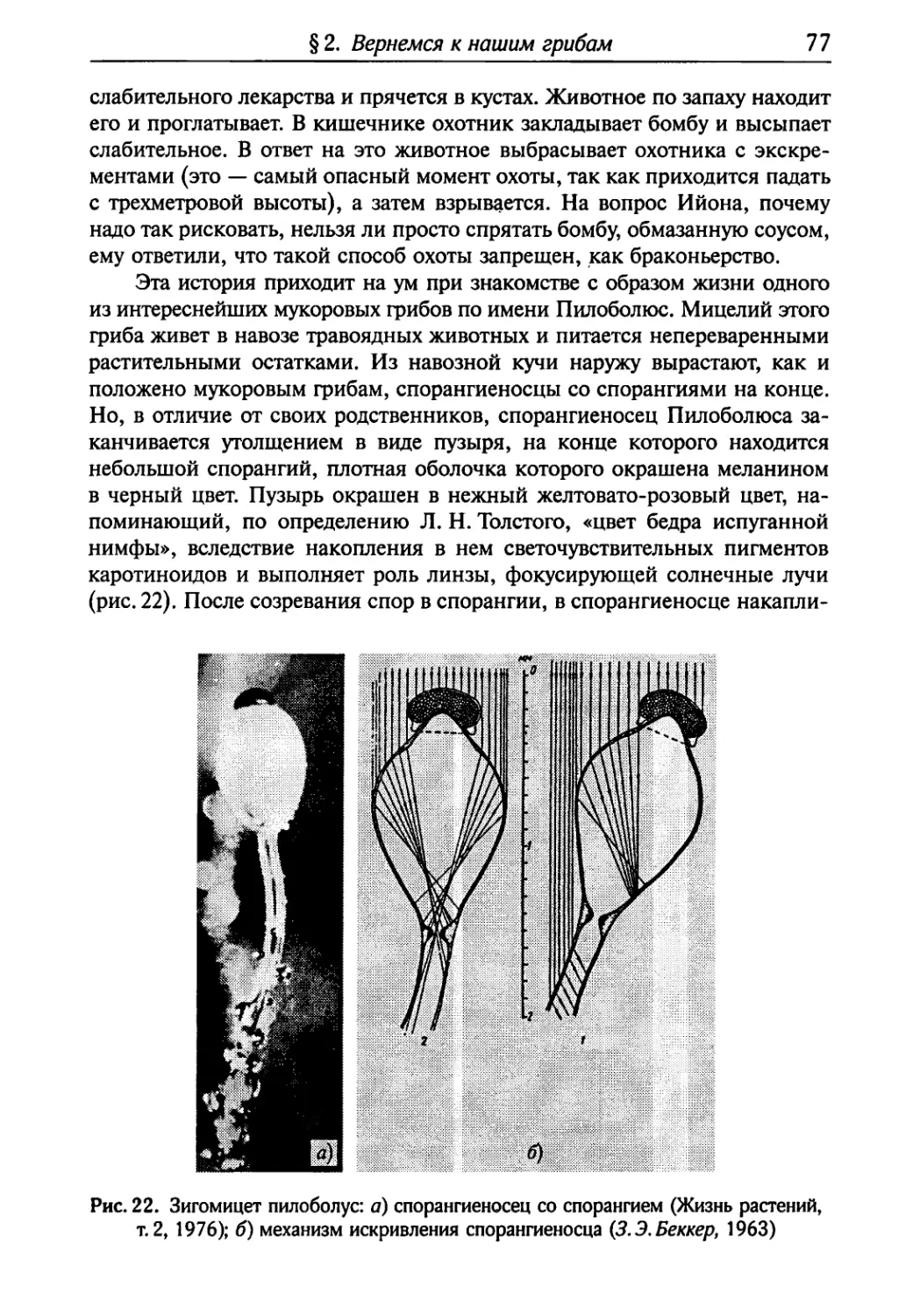

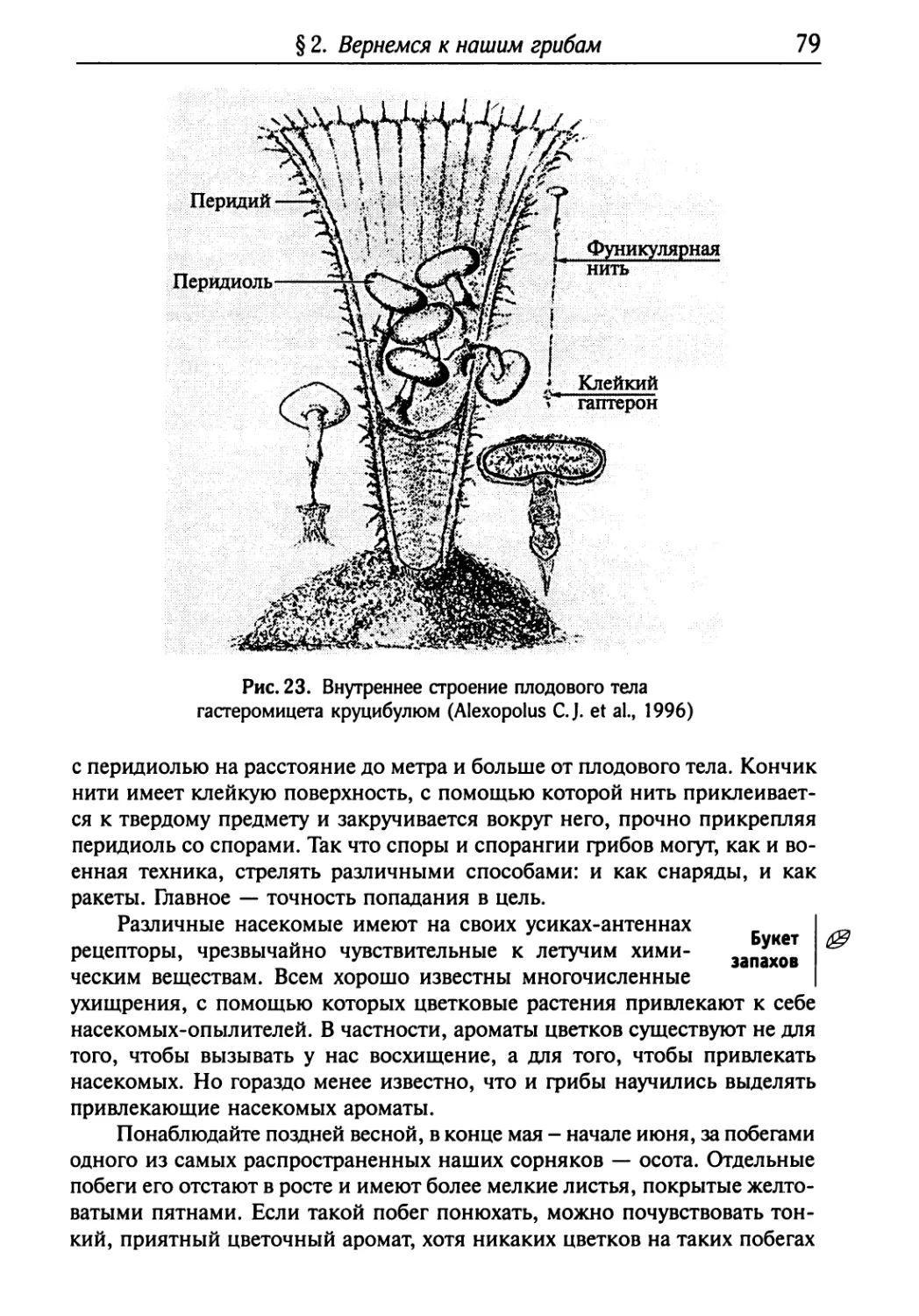









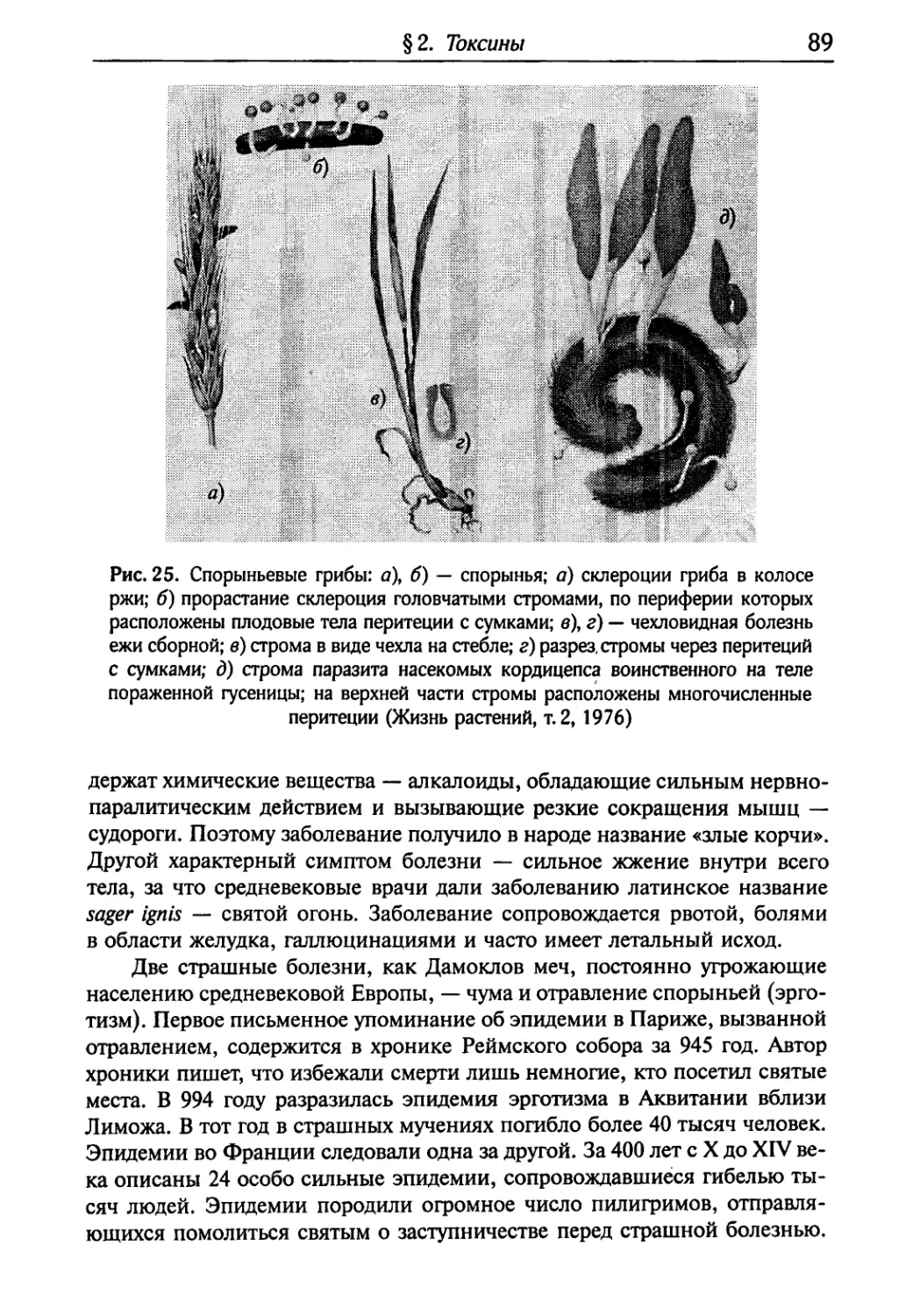
























































































































































Текст
ББК 28о 28.073 28.5 41
Дьяков Юрий Таричанович
Занимательная микология.— М.: Книжный дом «ЛИБРОКОМ»,
2013. — 240 с. (НАУКУ — ВСЕМ! Шедевры научно-популярной
литературы (биология).)
В настоящей книге в популярной форме описаны особенности строения, фи¬
зиологии и образа жизни большой группы организмов — грибов, их биотические
связи с растениями и животными. Приведены сведения о вреде, который наносят
грибы как возбудители болезней полезных растений, домашних животных и чело¬
века и как разрушители продуктов человеческой деятельности, а также рассказы¬
вается о том, как люди используют грибы, получая из них продукты питания,
лекарства и другие полезные вещества.
Книга предназначена для студентов биологических, сельскохозяйственных
и медицинских вузов, школьных учителей биологии, а также широкого круга любо¬
знательных читателей.
Издательство «Книжный дом “ЛИБРОКОМ”».
117335, Москва, Нахимовский пр-т, 56.
Формат 60x90/16. Печ. л. 15. Зак. № ВП-89.
Отпечатано в ООО «ЛЕНАНД».
117312, Москва, пр-т Шестидесятилетия Октября, 11А, стр. II.
ISBN 978-5-397-03209-4 © Книжный дом «ЛИБРОКОМ», 2012
Все права защищены. Никакая часть настоящей книги не может быть воспроизведена или
передана в какой бы то ни было форме и какими бы то ни было средствами, будь то элек¬
тронные или механические, включая фотокопирование и запись на магнитный носитель,
а также размещение в Интернете, если на то нет письменного разрешения владельца.
НАУЧНАЯ И УЧЕБНАЯ ЛИТЕРАТУРА
URSS
E-mail: URSS@URSS.ru
Каталог изданий в Интернете:
http://URSS.ru
Телефакс (многоканальный):
I + 7 (499) 724 25 45
Содержание
Предисловие 5
Введение 7
Гиава 1. Кто такие грибы 10
§ 1. Определение 10
§ 2. Система грибов 16
§ 3. Как и где возникли грибы 21
§ 4. Сколько видов грибов живет на Земле 23
Пшва 2. Мицелиальный образ жизни 26
§ 1. Грибной мицелий и его роль 26
§ 2. Строение и рост 28
§3. Питание 32
§4. Что можно сделать из разветвленных ниток 33
§5. Дрожжи 36
§6. Грибы-экстремалы 40
§ 7. Грибы-поденки и грибы-секвойи:
продолжительность жизни разных грибов 42
§ 8. Что такое грибная особь 44
§ 9. Есть ли у грибов иммунитет? 47
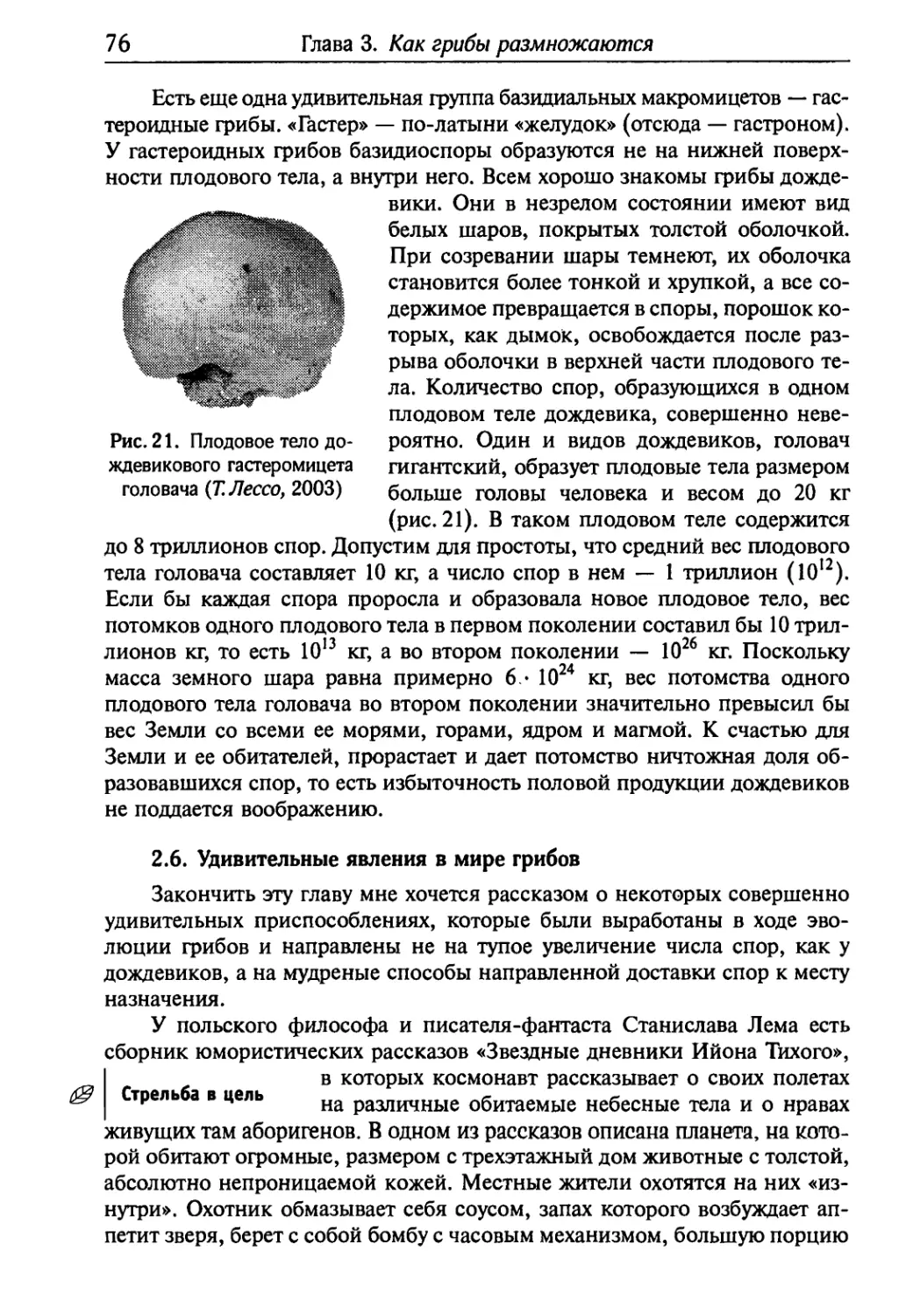
Гпава 3. Как грибы размножаются §2
§1.0 том, как размножаемся мы и наши меньшие братья .... 52
§ 2. Вернемся к нашим грибам 59
Бгава 4. Вторичные метаболиты грибов 82
§ 1. Первичный и вторичный метаболизм 82
§2. Токсины 83
§3. Фитогормоны 102
§4. Пигменты 105
4
Содержание
Нгава 5. 1}шбы и растения 108
§ 1. Грибы-симбионты. Микоризные грибы 109
§2. Грибы-паразиты 114
§3. Грибы-сапротрофы 137
Нгава 6. Лишайники: два в одном 143
§ 1. Что такое лишайник 143
§ 2. Обитатели лишайниковой «коммуналки» 145
§ 3. Как выглядят лишайники 145
§4. А что у них внутри? 148
§5. Как они размножаются 149
§6. Как они питаются и где живут 150
§ 7. Какую роль играют лишайники
в биосфере и в биоценозах 152
§8. Возможные пути использования лишайников 154
Нгава 7. 1}шбы и животные 157
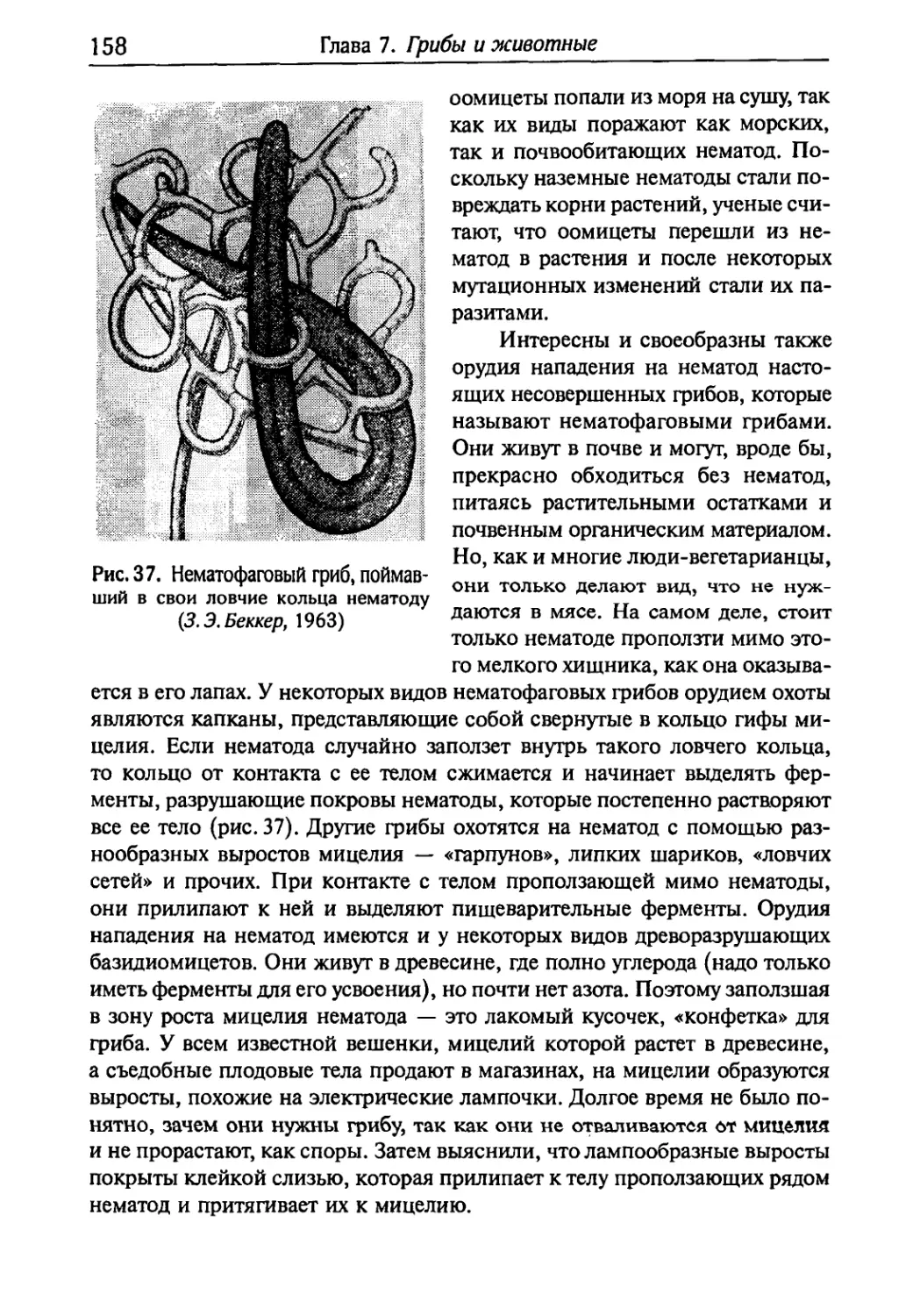
§ 1. Грибы и нематоды 157
§2. Грибы и насекомые 159
§ 3. Грибы и жвачные животные 167
Глава 8. Эти удивительные создания миксомицеты 170
§1. Сетчатые слизевики, или лабиринтуломицеты 171
§2. Паразитические слизевики, или фитомиксины 171
§3. Настоящие миксомицеты, или миксогастриды 172
§4. Клеточные миксомицеты: диктиостелиды и акразии 176
Глава 9. 1}шбы и человек 182
§ 1. Как грибы портят нам кровь 182
§ 2. Как люди используют грибы 196
Нгава 10. 1}шбы — модели в научных исследованиях 216
Словарь основных терминов 224
Источники иллюстраций 236
Предисловие
Очень давно, в школьные годы я прочитал две кни-
0 п u п Занимательно
ги: «Занимательную математику» Я. И. Перельмана и необычном
«Занимательную минералогию» А. Е. Ферсмана. Книги,
помнится, понравились, но сейчас, к сожалению, от них не осталось в па¬
мяти не только содержания, но и впечатления. Лет пятнадцать назад, уже
старым человеком, я встретился с книгой М. Л. Гаспарова «Занимательная
Греция», которую прочитал взахлеб, потом еще раз — спокойнее и более
внимательно, так что многое из того, чего не знал ранее, даже запомнил.
И у меня возникло желание написать занимательную, по возможности,
книгу о жизни организмов, которыми я всю жизнь занимаюсь профессио¬
нально, — о грибах, но все не хватало времени. Наконец, освободившись
от административных обязанностей, я приступил к этой книге, работу над
которой облегчало многолетнее чтение лекций студентам и написание ря¬
да статей и книг в области микологии.
Конечно, грибы — не древние греки, а я как беллетрист — далеко
не Гаспаров. К тому же огромное количество грибов, фу — противно вы¬
говорить — образуют плесени или паразитируют на растениях, животных
и даже человеке. Однако и жизнь паразитов не только достойна внимания,
но и полна интереснейших тайн. По этому случаю вспомню одну историю.
Когда я нахожусь на кухне, я слушаю радио. Однажды слушал беседу с из¬
вестной грузинской пианисткой Элисо Вирсаладзе. Она рассказывала, что
ее бабушка, профессор грузинской консерватории, давала дома частные
уроки игры на фортепьяно детям. И выговаривая нерадивым ученикам,
она говорила им: «Паразитолог увлекается паразитами, а ты не можешь
увлечься музыкой!». То есть, по ее мнению на низшей точке шкалы, раз¬
битой по степени увлечений, находятся паразиты, а на высшей — музыка.
Думаю, это не совсем так. Есть профессии, и есть внепрофессиональные
интересы. Конечно, предложенная бабушкой Вирсаладзе шкала в какой-
то мере отражает последние — немузыкантов, увлекающихся музыкой,
гораздо больше, чем непаразитологов, увлекающихся паразитами, но для
профессионала, если он занимается чем-то не только ради денег, его про¬
фессиональные занятия — самые интересные занятия на свете. Тем более,
что изучение биологии паразитов открывает такие хитросплетения их при¬
способлений, что захватывает дух. А об интересном надо писать интересно,
6
Предисловие
ибо, по мнению великого немецкого естествоиспытателя Александра Гум¬
больдта, «вдохновение поэта, украшение речи и сладкие звуки задушевной
тоски не только нигде не вредят точности изображения физических яв¬
лений, но, напротив, как это обыкновенно случается там, где искусство
черпает из ясного источника, они еще более возвышают живительное впе¬
чатление, производимое величием и истиной картин природы».
Еще одно обстоятельство усложняет написание популярной книги
о жизни грибов, как, впрочем, и других организмов, — необходимость
привлечения хотя бы и в самом поверхностном изложении данных совре¬
менной экспериментальной биологии, сложных для понимания неподго¬
товленного читателя и набитых чуждыми для русского языка терминами.
А чтобы читатель не потонул во время плавания, столкнувшись с терми¬
нологическими рифами, в конце книги помещен словарик общебиологи¬
ческих и микологических терминов, встречающихся в книжке, к которому
автор предлагает обращаться в процессе чтения.
Желаю любознательному читателю успешно проплыть между Сцил-
лой и Харибдой этих трудностей и познакомиться с новой для себя ин¬
формацией о разнообразии объектов живой природы, а молодым людям
пусть эта книжка поможет в выборе профессии, так как изучение инте¬
реснейшего мира грибов может дать им много положительных эмоций.
Введение
Микология — наука о грибах. Чем же она занимательна, и почему о
ней надо писать не только статьи и книги, предназначенные для специали¬
стов, занимающихся биологией грибов, или справочники для грибников-
любителей, занимающихся «тихой охотой», но и популярные книги просто
для любознательных читателей? Думаю, для этого есть несколько причин.
1. Все живые организмы, окружающие человека и имеющие доста¬
точные размеры тела, чтобы быть доступными для наблюдений за ними,
всегда привлекали внимание. Сколько народного творчества посвящено
животному миру — от античных басен Эзопа и памятника Средневековой
Европы лиса Ренара, воспетого Гете в «Рейнеке-Лисе», до народных сказок
разных стран. Животные в народном творчестве наделены определенными
чертами характера, которые используются и для характеристики людей.
Хитрый, как лиса, упрямый, как осел, глупый, как ворона (хотя интел¬
лект у вороны будет повыше, чем у осла и лисицы); царь зверей — лев,
царь птиц — орел: рассказывать об этом можно до бесконечности. То же
самое можно сказать и о растениях. Легенды о цветении папоротника
в ночь на Ивана Купалу (хотя папоротник не образует цветков по опре¬
делению), таинственные корешки, которые собирал гауфский маленький
Мук... Издавна человек использовал животных и растения не только для
сугубо потребительских целей (для достижения полезного), но и для удо¬
вольствия, достижения приятного, причем не только для чисто интел¬
лектуального удовольствия, какое получаешь, обучив разговорной речи
попугая, скворца или ворона, но и для достижения иллюзорных сверхо¬
щущений, впадения в транс или религиозный экстаз при потреблении
растений, обладающих наркотическими свойствами.
Мой школьный товарищ все сочинения по лите¬
ратуре начинал фразой: «Пушкин (Гоголь, Тургенев, Г^и ы и МИФЫ
Фадеев...) занимает особое место в русской литературе». И был прав. Гри¬
бы, как и писатели, тоже занимают особое положение. Необычность их
форм, внезапное массовое появление и такое же внезапное исчезновение,
отменные вкусовые качества и частые случаи отравления грибами, — все
это вызывало очень серьезное, даже мистическое отношение к грибам
у населения. В сказке Кэрролла «Алиса в стране чудес» Гусеница расска¬
зывает Алисе, что, если съесть кусочек мухомора с одной стороны, то
8
Введение
вырастешь очень большой, а если съесть с другой стороны, то станешь
очень маленькой. По-видимому, этот эпизод навеян рассказами об ощуще¬
ниях людей, поевших мухоморы. Г&плюциногенные свойства некоторых
грибов также укрепляли веру в их сверхъестественные свойства. Грибы
явились не только героями многих сказок, мифов, легенд разных наро¬
дов, но и вошли в практику религиозных культов. Галлюциногенные грибы
применялись и применяются до сих пор в религиозных культах разных
стран — от Мексики до Чукотки. Необычная форма плодовых тел многих
грибов, имеющая сходство с гениталиями человека, также явилась источ¬
ником разнообразных мифов. В одном из якутских сказаний описывается
время, когда на деревьях росли фаллосы, потом они упали, и некоторые
приросли к мужчинам, а другие, упав на землю, превратились в грибы.
Широко распространены также мифы о небесном происхождении гри¬
бов, которые попадают на землю после вспышки молнии и удара грома.
А среди аборигенов Сибири бытовали рассказы о мухоморолюдях, которые
объединены в особое племя; они могут схватить человека и показывать
ему «потусторонний мир» (по-видимому, эти легенды возникли вследствие
галлюцинаций, вызванных поеданием мухоморов). Есть также упомина¬
ния о девушках-мухоморах, заманивающих своими чарами охотников в та¬
ежную глушь (прямо, как древнегреческие морские ведьмы-сирёны). Ми¬
фотворчество, связанное с грибами, продолжается и в наше время. В годы
перестройки по телевидению был показан фильм о том, что группа людей,
называвших себя большевиками, ела слишком много грибов и под вли¬
янием этого устроила Октябрьский переворот, а их вождь Ленин вообще
является грибом. Авторы фильма откровенно ерничали, но если бы они
знали, что эти их странные высказывания не так уж далеки от истины...
2. Отношение к грибам как к непонятным, даже мистическим ор¬
ганизмам, не укладывающимся в нормальные системы, разделяющее все
живые существа на растения и животные, присуще не только фольклору,
но и серьезным ученым — от римлянина Плиния Старшего, считавшего,
что появление грибов связано с грозой, до великого систематика Лин¬
нея, которой в своей системе растительного мира отнес грибы к группе,
названной им «Хаос», потому что ее нельзя было связать с какой-либо
другой группой. А ботаник XVIII века Вейан назвал грибы изобретениями
дьявола, которые созданы для того, чтобы портить нервы систематикам.
Естественная (филогенетическая) система грибов, отражающая их поло¬
жение среди других групп организмов, их связи с растениями, животными
и друг с другом, начала создаваться немного более полусотни лет назад бла¬
годаря серьезным методологическим достижениям в области цитологии,
биохимии, физиологии и молекулярной биологии. Ее создание продол¬
жается и сейчас.
3. Бытующее представление о грибах как об организмах, состоящих
из шляпки и ножки, неверно вдвойне. Во-первых, так устроены не грибы,
Введение
9
а лишь органы их размножения, а само тело этих «грибов» находится в поч¬
ве и совершенно на них не похоже; во-вторых, такие органы размножения
имеют далеко не все грибы, а лишь не самая большая их часть. Огромное
число грибов имеет микроскопические органы размножения, они всю¬
ду окружают нас, но мы обращаем на них внимание толь в тех случаях,
когда заведется «грибок» на пальцах ног, вызывая нестерпимый зуд, или
когда на даче все плоды земляники покрылись серым пушистым нале¬
том и стали непригодными для употребления. Но если в любой комнате
Вашей квартиры поставить открытую микробиологическую чашку Петри,
заправленную питательной средой, то на второй-третий день поверхность
среды покроется колониями грибов. И если материал, склеивающий плит¬
ки на стенах Вашей ванной комнаты, покроется черным налетом, то это
тоже грибы. И если вдруг половые доски в домике на даче под давлением
Ваших шагов, ломаются, и пол проваливается, — это тоже грибы.
4. Наконец, мир грибов удивителен еще и своим образом жизни.
Их тело, называемое талломом, ни на что не похоже, уникально, мно¬
гие способы размножения и способы обмена генетической информаци¬
ей присущи только им. Грибы, как инопланетяне, — особый, ни с чем
не сравнимый мир, который начал открывать свои тайны совсем недавно,
после того, как были разработаны методы, позволившие вскрывать самые
интимные процессы, в жизни их клеток и клеточных конгломератов.
Кажется, этих аргументов достаточно, чтобы взять в руки книжку
с предложенным названием, а уж какой она получилась, судить не мне.
Глава 1
Кто такие грибы
§1. Определение
Чем меньше человек знает, тем легче ему жить. Как
сказал три тысячи лет назад Екклезиаст, «...во многой
мудрости много печали, и кто умножает познания, умно¬
жает скорбь». Это глубочайшее высказывание примени¬
мо к любой области человеческого знания, применимо
оно и к микологии.
Определить свойства той или иной группы организ¬
мов означает очертить границы, отделяющие эту группу
от других групп на огромном панно, называемом био¬
той. Чем меньше знаний, тем проще это сделать. В од¬
ном детском рассказе две девочки сидели около ручейка
и спорили, живой он или неживой? В качестве аргумен¬
тов, подтверждающих, что ручеек живой, были такие:
«ручеек бежит» и «когда на него дуешь, то он морщит¬
ся». А вот дихотомическая классификация природы, ко¬
торую изложил в книге «Диалектика» знаменитый хри¬
стианский богослов Иоанн Дамаскин, живший в VII в.:
все существа он разделил на телесные и бестелесные;
телесные, в свою очередь, — на одушевленные и неоду¬
шевленные; одушевленные — на наделенных чувства¬
ми животных и бесчувственные растения; животных —
на словесных и бессловесных; словесных — на смертных
и бессмертных (ангелов); смертных — на людей, волов,
коней, собак и т. п. (уже здесь — логическая ошибка,
так как бессловесные животные объединены со словес¬
ными людьми); а людей — на отдельные лица, каковы
Петр, Павел, Иоанн.
Все эти рассуждения касаются и отношения к гри¬
бам. Мало сведущий человек оперирует лишь неболь¬
шой выборкой организмов, привлекших его внимание
крупными размерами, яркой окраской, частой встре¬
чаемостью или практическим использованием. Любой
грибник-любитель, не задумываясь, разделит встречен-
§1. Определение
11
ные в лесу организмы на грибы и
не грибы. Даже впервые увиденные
редкие виды, поразившие нашед¬
шего оригинальными формами, та¬
кие как весёлка или земляная звез¬
дочка (рис. 1), любознательные со¬
биратели несут не к ботаникам или
зоологам, а к микологам. Однако
люди, профессионально занимаю¬
щиеся биологическим разнообра¬
зием, значительно менее категорич¬
ны в разделении таксонов высокого
ранга.
Во-первых, некоторые организ¬
мы, относимые к разным группам,
имеют перекрывающиеся признаки,
вследствие чего границы между груп¬
пами становятся размытыми. На¬
пример, любой небиолог скажет,
чем растения отличаются от живот¬
ных. Растения ведут прикреплен¬
ный образ жизни, тогда как живот¬
ные подвижны. Вот как, например,
описал различия между растениями и животными фило¬
соф Освальд Шпенглер в своей знаменитой книге «Закат
Европы»: «Растение существует не само по себе. Оно об¬
разует часть ландшафта, в котором случай заставил его пустить корни...
А вот животное способно выбирать. Оно освобождено от связанности
всего прочего мира. Всякий рой мошкары, одинокая птица, пролетающая
в сумерках, лисица, подстерегающая птичий выводок — самостоятельные
малые миры в другом, большом мире. Инфузория, которая влачит в капле
воды... невидимое для человеческого глаза существование, ...свободна и
независима перед лицом целого мироздания. Дуб-великан, с одного из ли¬
стьев которого свешивается эта капля, — нет! Связанность и свобода — так
можно выразить глубочайшее и коренное различие растительного и жи¬
вотного существования».
Однако любой биолог знает, что есть множество растений, тела кото¬
рых не прикреплены к субстратам. Ряска плавает на поверхности водое¬
мов; перекати-поле разносится по сухой степи с помощью ветра. А уж сре¬
ди низших растений-водорослей свободно живущих организмов сколько
угодно, причем многие из них имеют светочувствительный аппарат и жгу¬
тики, позволяющие выбирать вектор движения — активно плыть в на¬
правлении света. С другой стороны есть много видов животных, прикреп¬
ленных к субстратам, например, колонии кораллов или морские «цветы»
Рис. 1. Удивительные грибы — гастеро-
мицеты: а) весёлка (фаллус); б) диктио-
фора (дама с вуалью); в) решеточник;
г) земляная звездочка. (Жизнь растений,
т. 2, 1976)
Растения или
животные?
12
Глава 1. Кто такие грибы
актинии. Далее. Растения в массе зеленые, по¬
тому что имеют в клетках хлоропласты с зе¬
леным пигментом хлорофиллом. С помощью
хлорофилла растения могут, используя энер¬
гию солнечного света, из простейших неорга¬
нических веществ углекислого газа и воды со¬
здавать органические соединения, необходи¬
мые для построения своего тела и выполне¬
ния им различных физиологических функций.
Многие любители растений выращивают до¬
ма пальмы или фикусы. Через несколько лет
эти растения могут вырасти до потолка ком¬
наты, однако количество земли в горшке при
этом почти не убавляется, так как растения
питаются в прямом смысле воздухом, вернее
находящимися в нем парами воды и углекис¬
лым газом, черпая из почвы только минераль-
Рис. 2. Побеги паразитиче- ные элементы — азот, фосфор и др. А жи-
ского растения заразихи на вотныв хлорофилла не имеют, нм необщцц-
корнях подсолнечника. (Бе- мы готовые органические вещества, запасен-
ляев И. М., Горленко М. Вные в растениях и в других животных, по-
Дьяков Ю.Т. и др., 1970) этому они имеют аппарат поглощения пищи
(ротовое отверстие) и пищеварительную си¬
стему, где эта пища разрушается с помощью разнообразных ферментов
и всасывается внутрь тела. Однако известно много растений-паразитов,
которые присасываются к стеблям или корням других растений и всасыва¬
ют из них готовые органические со¬
единения (рис. 2). Хлорофилл они
теряют за ненадобностью. Есть и
хищные растения, имеющие при¬
способления для ловли насекомых,
тела которых они переваривают сво¬
ими ферментами. И животные, ока¬
зывается, могут фотосинтезировать.
Морской моллюск зеленая элизия
имеет листообразное плоское тело
и вытянутый вперед ротовой аппа¬
рат. Элизия ползает по дну и пи¬
тается нитчатыми водорослями, но
хлоропласты съеденных водорослей
не перевариваются и могут в тече¬
ние нескольких месяцев функцио¬
нировать в организме элизии, снаб¬
жая его органическими вещества¬
Рис. 3. Морской моллюск зеленая
элизия (Rumpho М. Е. et al., 2000)
§1. Определение
13
ми — продуктами фотосинтеза. В геноме (ядре) элизии были обнаружены
гены, необходимые для такого длительного функционирования хлоро-
пластов пне своих клеток. Эти гены были когда-то перенесены ИЗ ядра
водоросли в ядро питающегося ею моллюска и закрепились там. Вот и
получился «ползающий лист», животное по своему строению и растение
по способу питания (рис. 3). А уж в процессе эволюции такие переходы
происходили неоднократно. Предок паразитического простейшего живот¬
ного трипаносомы когда-то поглотил одноклеточную зеленую водоросль,
превратившуюся в хлоропласт, и стал растением (водорослью) — эвгле¬
ной, а малярийный плазмодий, предки которого были одноклеточными
водорослями, вследствие паразитического образа жизни утратил хлоро¬
филлы и стал животным.
Во-вторых, совершенство биологических техно-
„ Методы
логий привело к тому, что на смену традиционным классификации
методам классификации организмов, основанных на организмов
сравнительной морфологии — визуальных признаках,
выявляемых (в зависимости от размера объекта) с помощью глаза, лупы,
светового и, наконец, электронного микроскопа, пришли методы клас¬
сификации, основанные на сравнении отдельных генов и целых геномов.
Системы, основанные на сравнении геномов, названы филогенетически¬
ми, так как во внимание принимается не морфологическое сходство, ко¬
торое может быть вызвано приспособлениями неродственных организмов
к жизни в одинаковых условиях (по образному выражению нашего вы¬
дающегося микробиолога Г. А. Заварзина — «пространством логических
возможностей»), а общностью происхождения. Так вот геномные сравне¬
ния показали, что многие организмы, традиционно относящиеся к грибам,
имеют различное происхождение.
Поэтому есть три способа отграничения организмов, называемых гри¬
бами.
1. Традиционный (расширительный). Английс¬
кий миколог Дэвид Хуксворт дал такое опре- посо ь| 0ТГРД”ИЧ*НЛИЯ
_ - организмов,
деление грибам: «Грибы — это организмы, являющихся грибами
которые изучают микологи». В XVIII-XIX ве¬
ках ученые в европейских университетах произвели разграничение
организмов, аналогичное тому, о котором было сказано при упо¬
минании любителей собирания грибов; давайте это разграничение
и будем соблюдать. При таком подходе к грибам надо относить раз¬
нообразные организмы, которые не привлекли внимания ботаников
и зоологов: от формирующих тонкие разветвленные нити почвооби¬
тающих бактерий — актиномицетов, до амебоподобных животных —
миксомицетов, заглатывающих поверхностью тела клетки бактерий.
2. Филогенетический (ограничительный). К грибам следует относить толь¬
ко те наиболее многочисленные таксоны, которые имеют общее про¬
исхождение и родственные связи. Остальные «грибоподобные» орга¬
14
Глава 1. Кто такие грибы
низмы следует из грибов исключить или, по крайней мере, упоминать
с приставкой «псевдо
3. Эколого-физиологический (компромиссный). Из комплекса признаков
выбрать один или несколько фундаментальных, присущих организ¬
мам данной группы и отсутствующих у организмов из других групп.
Важнейший признак в строении и функционировании клетки — нали¬
чие или отсутствие ядра. По этому признаку все организмы разделяют
на прокариот и эукариот. К прокариотам относятся бактерии и археи,
к эукариотам — ядерные организмы — от примитивных водорослей
до человека. Наиболее фундаментальным физиологическим признаком
является способ утилизации энергии и, в связи с ним, место ор¬
ганизмов в круговороте веществ. По соотношению этих признаков
организмы разделяют на эколого-физиологические группы, приве¬
денные в таблице.
Автотрофы
(способны к синтезу органических
веществ из неорганических)
Гетеротрофы (питаются готовыми
органическими веществами)
хемотрофы
фототрофы
осмотрофы
зоотрофы
используют для
синтезов энергию
химических
реакций
используют для
синтезов энергию
солнца
всасывают
органику
из окружающей
среды
заглатывают
органические
субстраты
Прокариоты
хемосинтезирую¬
щие бактерии
и археи
цианобактерии
(синезеленые
водоросли)
многие бактерии
и археи
миксобактерии
Эукариоты
растения
грибы
животные
Примечание: Как любая биологическая классификация, приведенная схема услов¬
на и отражает не единственный, а лишь наиболее распространенный тип питания
той или иной группы организмов с многочисленными исключениями. Например,
насекомые кровососы имеют осмотрофный тип питания, они всасывают органи¬
ческие соединения, находящиеся в крови. Также поступают и тли, всасывающие
соки растений.
Исходя их приведенной схемы, грибы можно охарактеризовать как
эукариоты, питающиеся осмотрофно.
Данный тип питания наложил определенные черты на морфологию
и метаболизм (обмен веществ) грибов. Поскольку они всасывают пита¬
тельные вещества из субстрата всем телом, их тело (таллом) должно быть
§ 1. Определение
15
устроено таким образом, чтобы максимальное число клеток имело бы кон¬
такт с субстратом. Поэтому самой распространенной жизненной формой
грибов является система сильно разветвленных нитей — гиф, называемая
грибницей или мицелием, которая погружена в субстрат, будь то почва
или ткань растения (хотя таллом у многих видов всегда, а у некоторых —
в определенных условиях жизни одноклеточный — дрожжеподобный). Та¬
кая форма тела наиболее пригодна для максимальной оккупации субстра¬
та. А поскольку большинство органических соединений находится в фор¬
ме высокомолекулярных полимеров (белков, полисахаридов, нуклеиновых
кислот и т. п.), не способных проникать в клетку через ее покровы, грибы
выделяют в субстрат активные гидролитические ферменты-деполимеразы,
которые расщепляют полимеры на олиго- и мономеры, транспортирую¬
щиеся в клетки. У нас пища попадает в кишечник, и в ходе длительного
путешествия по кишкам содержащиеся в ней органические полимеры раз¬
лагаются ферментами, которые выделяются в просвет кишечника, после
чего продукты разложения всасываются через стенки кишок. Грибы, на¬
оборот, выделяют гидролитические ферменты не внутрь, а наружу, то есть
грибная гифа — это кишка, вывернутая наизнанку. Такой тип питания
называют внекишечным пищеварением.
Внекишечное пищеварение подразумевает необ¬
ходимость решения еще одной проблемы — транс¬
порта продуктов питания в клетки из окружающей
среды. С какой стати вещества, разрушенные гриб¬
ными ферментами, должны проходить через клеточные покровы внутрь?
Есть три способа транспорта питательных веществ из окружающей среды
в грибную клетку.
1. Движение по градиенту концентрации. Если в разных частях сосуда,
разделенного проницаемой перегородкой, находится растворы како-
го-либо вещества, скажем сахара, в разных концентрациях, то моле¬
кулы сахара будут двигаться из отсека, в котором его концентрация
выше, в отсек с менее концентрированным раствором, а молекулы
воды — двигаться в обратном направлении, чтобы в конце концов
концентрации растворов выровнялись. Чтобы концентрация сахара
в субстрате, окружающем грибной мицелий, была выше, чем в самом
мицелии, сахар, проникший в растворенном состоянии в грибную
клетку, под действием ферментов быстро трансформируется в дру¬
гие соединения: шестиатомный спирт манит, дисахарид трегаллозу,
животный крахмал гликоген и другие, так что раствор сахара в окру¬
жающий среде все время остается ниже, чем в мицелии.
2. Грибы научились с помощью определенных химических реакций раз¬
вивать высокое тургорное давление, с помощью которого растворенные
в воде органические вещества всасываются в клетки, то есть грибная
гифа не только кишка, но и насос. В среднем осмотическое давление,
развиваемое грибной гифой, равняется 2,5 атмосфер (равно давлению
16
Глава 1. Кто такие грибы
Рис. 4. Искривление гиф гриба Ботритис при прохождении через плотный субстрат
(рис. Miyoshi, 1895; Money N. R, 2007): а), г) рыхлый слой желатины; б) плотный
слой коллодия; в) гифа гриба
в хорошо накачанной велосипедной шине). Поэтому клетки всех ми-
целиальных грибов покрыты прочными оболочками, иначе, клетки
могли бы лопнуть. Высокое давление необходимо грибам не только
для всасывания питательных веществ из окружающей среды* но и для
продвижения в плотном субстрате. Ведь грибы развиваются не только
в рыхлой почве (хотя глинистые почвы — тоже достаточно плотный
субстрат), но и в таких чрезвычайно плотных субстратах, как дре¬
весина. Плодовые тела шампиньона, живущего в городских почвах,
в процессе развития взламывают асфальт; кончик гифы многих гри¬
бов — паразитов растений — разрывает восковой слой на поверхно¬
сти листьев и наружную стенку эпидермальных клеток. Интересно,
что плотная полисахаридная стенка, покрывающая грибные клетки,
не только защищает клетки от разрывов наружной мембраны вслед¬
ствие высокого внутриклеточного тур горного давления, но и ослаб¬
ляет возможности передачи этого давления на окружающий гифы
субстрат. Поэтому при прохождении через плотные слои клеточная
стенка разрыхляется, что позволяет усилить давление протопласта
на субстрат, но одновременно приводит к искривлению и ветвлению
гиф, как это видно на рис. 4.
3. С мембраной клеток грибов, как, впрочем, и других организмов, свя¬
заны специальные белки-переносчики, которые специфически транс¬
портируют из клеток наружу и в клетки из окружающей среды высо¬
комолекулярные соединения.
§ 2. Система грибов
Организмы, изучаемые микологами, не являются единой группой,
ни по сравнительным признакам, ни по происхождению. На приведенном
ниже рис. 5 изображена система эукариотных организмов в виде цветочка,
§ 2. Система грибов
17
Рис. 5. Филогенетическое дерево эукариот. Закрашены черным цветом — водоросли и их
бесцветные производные; на светлом фоне — грибы и псевдогрибы (Keeling P. J., 2004)
каждый лепесток которого — это огромная группа, империя, включающая,
как и всякие уважающие себя империи, несколько царств. Организмы,
о которых пойдет речь в этой книжке, попадают в несколько империй,
расположенных в самых разных участках биоты.
Есть такая шутливая шарада: «первый слог —
„ Три независимые
совсем лошадь, второй — не совсем лошадь, а все группы грибов
вместе — совсем не лошадь». Отгадка — коньяк.
Организмы, изучаемые микологами, относя к трем большим, независимо
эволюционирующим группам, которым разные систематики присваива¬
ют ранги выше отделов (царства, империи). Совсем грибы — царство
истинных грибов (эумикота) — включено в империю одножгутиковых
(Uniconta), надцарство заднежгутиковых. В этом таксоне находятся два
царства — грибы и тканевые животные. Не совсем грибы (псевдогри¬
бы) — это отдел оомицетов, который включен в империю Chromalveolata,
царство Chromista, или, как его чаще называют, царство разножгутиковых
(гетероконта), или соламенотрубчатых (страменопила), и совсем не гри¬
бы — слизевики (Myxomycota). Настоящие слизевики (царство миксомико-
та) родственны грибам; сетчатые слизевики (отдел лабиринтуломикота)
родственны оомицетам, паразитические слизевики (отдел фитомиксины
или плазмодиофоромицеты) переведены в группу, родственную многим
18
Глава 1. Кто такие грибы
простейшим животным (царство Церкозоа); наконец, акразиевые мик¬
сомицеты ((акразиомикота) оказались родственниками эвгленовых водо¬
рослей и попали в царство Эвгленобионта. Оставим пока слизевики в покое
и рассмотрим вкратце особенности псевдогрибов и настоящих грибов.
Разножгутиковые включают большое число водорослей (золотистые,
диатомовые, бурые), имеющих специфический набор пигментов, окраши¬
вающих их клетки в буро-желтые тона, и два гетероморфных жгутика —
перистый и гладкий.
По ряду молекулярных, биохимических, цитологических и морфоло¬
гических признаков к ним примыкают грибоподобные бесцветные орга¬
низмы, для которых характерно гетеротрофное питание (как для сапро-
трофов или паразитов) и отсутствие пигментов. Таким образом, название
«фикомицеты» (грибоводоросли), используемое в старых системах грибов
для видов, подвижные клетки которых имеют гладкий и перистый жгути¬
ки, оказалось отчасти верным.
В связи с несомненным родством этих организмов с гетероконтными
водорослями долгое время считали, что оомицеты — древнейшая группа
гетероконтных организмов, возникшая до того, как эти организмы приоб¬
рели в качестве эндосимбионта одноклеточную красную водоросль, пре¬
вратившуюся со временем в хлоропласт. Однако оказалось иначе. Сиквенс
ДНК (определение последовательности оснований ДНК) фитофторовых
оомицетов, проведенный в начале нашего века, показал, что в геноме фи¬
тофторы присутствует около тысячи генов, предположительно полученных
от красных водорослей или цианобактерий (а цианобактерии дали начало
хлоропласту красных водорослей), из них более трети оказались идентич¬
ными диатомовой водоросли талассиозира. Эти гены контролируют синтез
ферментов, обслуживающих работу митохондрий и хлоропластов. Таким
образом, оомицеты — это в действительности бывшие водоросли, утра¬
тившие хлоропласты за ненадобностью в связи с переходом к паразитизму.
А отсюда следует, что паразитизм оомицетов первичен по сравнению с пи¬
танием мертвым субстратом.
Гетероконтные псевдогрибы составляют несколько
омиц ты классов; важнейший из них — класс оомицеты. Это вод¬
ные и наземные псевдогрибы. Водные оомицеты — сапролегнии живут
в пресных водоемах. Если зачерпнуть в стакан воды из пруда или озера
и бросить в него приманку — муравьиное яичко (куколку), дохлую муху
или конопляное зернышко, то через несколько дней приманка покроет¬
ся нежным белым пушком. Это — мицелий сапролегниевого оомицета
(рис. 6). Сапролегнии питаются не только дохлыми мухами, но могут за¬
ражать рыбью икру, мальков и ослабленных взрослых рыб, поэтому они
сильно вредят хозяйствам, разводящим рыбу, а также аквариумистам.
Среди наземных оомицетов много паразитов высших растений. Ми¬
целий паразитических оомицетов находится внутри ткани растения, а ор¬
ганы спороношения — пучки спорангиеносцев с зооспорангиями на кон-
§ 2. Система грибов
19
а)
Рис. 6. Сапролегния: а) мицелий без перегородок; б) спорангий с выходящими
двужгутиковыми зооспорами; в) оогоний с яйцеклетками (оосфермами); г) опло¬
дотворение оогония отрогами антеридия (З.Э. Беккер, 1963)
цах — высовываются через устьица, образуя белый налет с нижней стороны
листа, где расположено большинство устьиц. Поэтому вызываемые ими
болезни растений называют ложными мучнистыми росами. Важнейшие
из них — фитофтороз картофеля и томатов, мильдью винограда, ложная
мучнистая роса лука, огурца й других растений (рис. 7,8). Несмотря на пол-
Рис. 7. Оомицеты — возбудители болезней растений; фитофторозы: вверху —
поражения клубней и листьев картофеля; Внизу слева — поражение плодов
томата (Жизнь растений, т. 2, 1976); справа — фитофторозная корневая гниль
эвкалиптов в Австралии (Podger F.D., 1972)
20 Глава 1. Кто такие грибы
Рис. 8. Оомицеты — возбудители болезней растений; ложные мучнистые росы: слева —
ложная мучнистая роса подсолнечника; а) нижняя сторона листа с белым спороноше-
нием паразита, высовывающимся из устьиц; б) отставание в росте сильно пораженных
растений (Беляев И. М., Горленко М. В., Дьяков Ю.Т.и др., 1970); в) ложная мучнистая
роса (мильдью) винограда. (Fry W.E., Grunwald N.J., 2010)
ное отсутствие родства между паразитами растений из, класса оомицетов
и настоящими грибами поражает большое сходство между этими груп¬
пами в морфологии инфекционных структур и физиологии взаимоотно¬
шений их с растениями-хозяевами. Полный геномный сиквенс несколь¬
ких видов рода фитофтора позволил установить причины удивительного
конвергентного сходства между биотрофным питанием фитопатогенных
оомицетов и настоящих грибов. При использовании филогенетического
метода в сочетании с сравнительным геномным анализом в геноме фи¬
тофтор был обнаружен ряд генов, возникших в результате горизонтального
переноса из генома фитопатогенных грибов аскомицетов. Продукты этих
генов участвуют в осмотрофном питании растительными клетками, т. е.
в биотрофии. Таким образом, фитофторный геном (а, возможно, и геном
возбудителей ложных мучнистых рос) — мозаичный, включающий гены,
полученные от красных водорослей, анцестральных (предковых) бесцвет¬
ных гетероконт и фитопатогенных аскомицетов.
Самые многочисленные как по видовому составу,
Истинные грибы Так и По численности, истинные грибы. Они отлича¬
ются от описанных групп псевдогрибов по структуре генома, химическому
составу (в клеточной стенке большинства видов нет целлюлозы, но есть
хитин — полимер аминосахара хитобиозы), метаболизму (конечный про¬
дукт метаболизма азота не аминокислоты, а мочевина), ультраструктуре
(плоские, а не трубчатые, выросты внутренней мембраны митохондрий —
кристы) и морфологии. Поскольку наиболее примитивные представите¬
§3. Как и где возникли грибы
21
ли эумицетов (хитридиевые грибы) размножаются подвижными клетками
с одним гладким жгутиком, полагают, что грибы произошли от бесцветных
флагеллят7 в частности, от широко распространенной в пресных и морских
водах группы воротничховых жгутиконосцев. По молекулярным филоге¬
ниям истинные грибы образуют единую группу с тканевыми животными,
у которых подвижные клетки (например, человеческие сперматозоиды)
имеют один задний гладкий жгутик, конечным продуктом азотного ме¬
таболизма является мочевина, структурный полисахарид (у членистоно¬
гих) — хитин и тому подобные структурные и биохимические сходства
с грибами. Так что грибы скорее можно назвать скоромной, а не постной
пищей, и, следуя новым филогенетическим данным, священнослужите¬
лям стоит исключить их из списка продуктов, рекомендуемых в пост.
§3. Как и где возникли грибы
Итак, грибы и тканевые животные имели общего предка. Каков этот
предок, где он жил и как был устроен, никто не знает. Близки к живот¬
ным и грибам одноклеточные организмы воротничковые жгутиконосцы,
живущие в морях и пресных водах. Некоторые из них, в частности мор¬
ской хищник Моносига, могут под влиянием химических выделений со¬
путствующих микроорганизмов переходить из одноклеточного состояния
в колониальное. Считали, что воротничковые жгутиконосцы — исходная
группа, давшая начало и животным, и грибам. Но сейчас уже так не счи¬
тают, рассматривая их как древнюю группу, параллельную, а не предковую
примитивным грибам и животным. Конечно, высока вероятность того, что
предки грибов давно вымерли, и в связи с тем, что отпечатки однокле¬
точных организмов плохо сохраняются (а правильнее — почти не сохра¬
няются), мы никогда о них ничего не узнаем. Но помимо воротничковых
жгутиконосцев есть и другие кандидаты в предшественники грибов и сре¬
ди некоторых живущих вокруг нас микроорганизмов.
Эти одноклеточные простейшие являются об¬
лигатными (обязательными) паразитами различных р р
животных — от морских беспозвоночных до человека. Их клетки имеют
наиболее примитивные черты организации среди всех известных эука¬
риот: 1) самый маленький геном: ДНК имеет бактериальные размеры;
2) отсутствие митохондрий; 3) отсутствие интронов (некодируемых участ¬
ков гена, которые вырезаются в процессе синтеза белка — трансляции);
4) рибосомы, устроенные по типу прокариотных, а не эукариотных рибо¬
сом); 5) отсутствие копий генов, кодирующих рибосомальные типы РНК,
что свойственно для прокариот, но не для эукариот. Конечно, некоторые
черты примитивизма могут быть вызваны паразитическим образом жизни,
ведь паразитам не нужно многое из того, что необходимо свободно живу¬
щим организмам — они это необходимое забирают у хозяина. Но такие
структурные особенности, как число и структура рибосомальных генов,
22
Глава 1. Кто такие грибы
есть явный признак первичного примитивизма, а не потерь в процессе па¬
разитизма. Имея примитивные черты в строении клетки, микроспоридии
отличает весьма сложный, уникальный аппарат размножения и заражения
своих хозяев. Споры попадают в организм животного чаще всего при по¬
едании загрязненного ими субстрата. Внутри тела своей жертвы в споре
формируется зародыш, лишенный каких-либо покровов, эдакий амебо-
ид, и длинная полярная трубка, которая выбрасывается из споры и через
отверстие которой амебоид проникает в новую клетку хозяина. Полная
расшифровка последовательности оснований ДНК (сиквенс ДНК) микро¬
споридий показал их наибольшую близость к грибам, в частности, нали¬
чие грибных генов, кодирующих ферменты, которые участвуют в синтезе
хитина. Поэтому микроспоридии стали считать примитивными грибами.
Но как строение клеток, так и тип питания микроспоридий имеют мало
общих с грибами черт. Возможно, микроспоридии перешли к внутрикле¬
точному паразитизму в тот древний геологический период, когда не было
высших растений и содержание кислорода в атмосфере было очень низ¬
ким (поэтому митохондрии были для них излишней роскошью). В от¬
личие от примитивных водных грибов и псевдогрибов-оомицетов, виды
которых паразитируют на водных беспозвоночных, грибах и водорослях,
микроспоридии описаны только как паразиты животных. Может быть,
попадание микроспоридий в растения (например, передача их при пита¬
нии растениями зараженных ими беспозвоночных животных) и привело
к возникновению из микроспоридий организмов, называемых грибами.
Криптомицеты — ярчайший пример того, что
Криптомицеты -
г может быть достигнуто современными технологиями,
созданными молекулярной биологией. Их огромное биологическое раз¬
нообразие и широкая встречаемость в почвах, пресных водоемах и морях
на всех континентах были описаны раньше, чем их увидели, а увидели
тоже благодаря молекулярным технологиями. Дело в том, что эти ор¬
ганизмы не культивируются в искусственных условиях, поэтому об их
существовании узнали лишь после того, как стали использовать опреде¬
ленные последовательности ДНК, похожие на ДНК-последовательности
грибов, но все-таки отличающиеся от них, в качестве проб на общие
препараты ДНК, выделенные из разных источников. Затем к этим по¬
следовательностям ДНК (праймерам), или к комплементарным им после¬
довательностям РНК, пришили молекулу красителя и с помощью этого
метода (метода микрофишей) увидели, что представляют собой крипто¬
мицеты. Они оказались мелкими одноклеточными микроорганизмами,
образующими в своем жизненном цикле как подвижные клетки с одним
гладким задним жгутиком (как грибы и животные), так и неподвижные
клетки, прикрепляющиеся к клетками других более крупных организмов,
в частности к диатомовым водорослям. Только об одном криптомицете
было известно раньше этих анализов ДНК. Это паразит водных грибов
Розелла, который относили к самым примитивным грибам — хитридио-
§ 4. Сколько видов грибов живет на Земле
23
мицетам. Одноклеточная Розелла живет и размножается внутри мицелия
водных грибов, причем в отличие от настоящих грибов она не может са¬
мостоятельно синтезировать хитин, а использует заготовки хитина гриба-
хозяина для построения своей клеточной стенки. Что криптомицеты —
разнообразная группа организмов, родственная грибам и, по-видимому,
более древняя, чем грибы, не вызывает сомнения, но являются ли они
предками грибов или это параллельно развивающаяся группа организмов,
имеющая общего с грибами предка, не известно.
Итак, можно предполагать, что грибы возникли, во-первых, в море,
и, во-вторых, их предками были внутриклеточные паразиты морских бес¬
позвоночных животных. А вот на сушу они вышли, как предполагают,
из пресных мелководных озер вместе с предками наземных растений —
зелеными водорослями, под их покровами как их паразиты и симбионты.
И они имели уже мицелиальную структуру, без которой им бы не выжить
вне воды.
§ 4. Сколько видов грибов живет на Земле
К настоящему времени микологи описали около
100 ООО видов грибов. Это в три с лишним раза мень¬
ше, чем число описанных наземных растений, включая мхи и папорот¬
ники. Однако по прогнозам ученых число описанных наукой растений
составляет примерно 90 % от истинного числа обитающих на земле, а чис¬
ло описанных грибов — только 5 %. Таким образом, по прогнозируемому
числу видов грибы значительно превышают все растительноподобные ор¬
ганизмы вместе взятые. На чем основаны эти прогнозы?
1. Число ежегодно описываемых новых видов.
Для растений это единицы, а для грибов — в среднем около двух
тысяч.
2. Соотношение числа видов растений и видов грибов, развивающихся на
растениях {живых и мертвых).
В Великобритании, где достаточно подробно исследована как флора,
так и микобиота, на каждом виде растения описано около десятка видов
грибов (соотношение растения/грибы —- около 1:10). В Индии, где флора
исследована гораздо лучше, чем микобиота, соотношение растения/грибы
почти обратное. В тропических лесах, как полагают, существует огромное
число (порядка 270 000) еще неизвестных видов грибов, паразитирующих
на растениях. Так, в Панаме на небольшом участке леса за один сезон
работы микологов было описано 75 видов сумчатых грибов, обитающих
на листьях деревьев, из которых 59 видов (79 %) — новые для науки.
На пальмах в последние годы описано 1580 видов грибов, из них новых
для науки 75%.
24
Глава 1. Кто такие грибы
3. Грибы, обитающие на насекомых.
Есть большие группы грибов, целиком связанные с насекомыми (па¬
разиты и симбионты насекомых). Описание их биоты практически только
началось. Ожидается по крайней мере удесятерение числа описанных ви¬
дов.
4. Грибы экстремальных мест обитания.
Совсем недавно началось исследование видового состава грибов, оби¬
тающих в болотах, содовых озерах и солончаках, льдах Арктики и Антарк¬
тиды. Первые исследования показывают наличие в некоторых из описан¬
ных мест огромного числа новых видов грибов.
5. Водные грибы.
Древнейшие грибы — хитридиомицеты, как полагают, вышли в прес¬
ные воды и на сушу из вод морских, но их выделение из мест обитания
в море методически очень сложно, поэтому они очень плохо исследова¬
ны. В последние годы появились молекулярные методы регистрации тех
или иных видов организмов без их выделения в чистую культуру. Для
этого из образца изучаемого субстрата выделяют тотальную ДНК и за¬
мечательным приемом, получившим название полимеразно-цепной реак¬
ции (его разработчики получили за это Нобелевскую премию) определяют
присутствие интересующих групп организмов. Дискриминация ДНК раз¬
ных организмов напоминает компьютерное определение стоимости товара
на основании нанесенного на нем штрих-кода в кассовых аппаратах. Ис¬
пользование подобных методов показало, что в глубоководных участках
океана обитает большое число видов хитридиевых грибов, не известных
микологам.
6. Виды-двойники.
Поскольку человек более 80 % информации получает с помощью зре¬
ния, системы растительного и животного мира традиционно построены
на различиях морфологических признаков (если бы собаки строили свои
системы живых организмов, то в их основу были бы положены различия
в летучих веществах, улавливаемых носом; полицейские собаки, например,
могут различить запах пота любых двух человек, кроме однояйцевых близ¬
нецов). Однако легко говорить о различиях морфологических признаков,
когда их (этих признаков) очень много, как, например, у высших растений
или насекомых. Но чем проще устроен организм, тем меньше у него при¬
знаков, на основании которых можно произвести дискриминацию одного
вида от другого. Как писал русский альголог (специалист по водорослям)
Н. А. Гайдуков «в одном цветке скрытосеменного растения нам известно
больше признаков, чем во всем теле низшей водоросли». То же самое
можно сказать и в отношении таких грибов, как дрожжи. Для крупных
организмов, включая большинство животных, растений и грибы макро-
мицеты, большое значение имеют визуальные признаки. Для видового
§ 4. Сколько видов грибов живет на Земле
25
определения микроскопических грибов применяют световые микроскопы.
Затем были созданы различные модели электронных микроскопов, позво¬
лившие увидеть сложнейшие детали клеточного строения, которые также
стали использовать для дискриминации видов. Однако независимо от тон¬
кости изучаемых признаков, концепция вида у грибов долгое время была
основана исключительно на морфологических и ультраструктурных разли¬
чиях между экземплярами и размахе изменчивости этих признаков. Толь¬
ко в последние 20-30 лет к определению видового статуса грибов стали
прилагать биологическую концепцию вида, основанную на эксперимен¬
тальном скрещивании отдельных экземпляров. Она была разработана для
высших животных и растений, в определении вида которых входит такое
понятие, как репродуктивная изоляция, то есть неспособность получения
плодовитых гибридов при скрещивании разных видов. Близкородствен¬
ные виды могут скрещиваться, но их гибриды стерильны (всем хорошо
известен мул — гибрид осла и лошади), а отдаленные вообще не способны
к скрещиванию. Экспериментальные скрещивания грибов показали, что
многие экземпляры одного и того же морфологического вида репродук-
тивно изолированы друг от друга, то есть представляют собой «биологиче¬
ские» виды или виды-двойники. Например, всем известен осенний опе¬
нок, плодовые тела которого, растущие на убитых его мицелием деревьях,
являются главным объектом осенней грибной охоты. Ранее его описывали
как один вид, признаки которого сильно варьируют в зависимости от роста
на разных породах деревьев, погодных условий и проч. С помощью скре¬
щиваний показано, что под этим названием существует более десятка не¬
зависимых видов с перекрывающимися морфологическими признаками.
Наличие морфологически сходных биологических видов широко распро¬
странено среди грибов и также значительно увеличивает число их видов.
Глава 2
Мицелиалъный
образ жизни
§ 1. Грибной мицелий и его роль
У древнейших грибов (хитридиомицетов), живущих
в водной среде, мицелия нет, их тело состоит из от¬
дельных, часто активно плавающих вследствие наличия
жгутиков, клеток. В водной среде мицелиальная струк¬
тура тела не имеет особых преимуществ перед иными
типами строения. Преимущества возникают только при
переходе на землю и существовании в твердых субстра¬
тах, таких как почва или тело растений. Перечислю эти
преимущества:
1. Максимальная оккупация субстрата потомком од¬
ной споры.
2. Транспорт веществ, нередко на большие расстоя¬
ния.
3. Способность преодолевать воздушные полости, про¬
низывающие почву.
4. Спосрбность преодолевать участки, бедные пита¬
тельными веществами.
5. Морфогенетические возможности мицелия: образо¬
вание плодовых тел, стром, ризоморф и т. п. Функ¬
циональная дифференциация мицелия.
О некоторых из перечисленных функций мицелия
и пойдет разговор в дальнейшем.
Однако сначала надо сказать несколько слов об
одном классе грибных молекул, без которых грибы не
смогли бы не только выйти из воды на сушу, но и во¬
обще существовать в воздушной среде. Это гидрофоби-
ны, низкомолекулярные (содержащие около 100 амино¬
кислот) белки, секретируемые (выделяемые) на поверх¬
ность мицелия. Гкдрофобины способны к самосборке
§ 1. Грибной мицелий и его роль
27
(кристаллизации) и образованию белковой пленки тол- Гидрофобины
щиной 10 нанометров на границе гидрофобной (водо- обеспечили
отталкивающей) и гидрофильной (смачиваемой водой) жизнь на суше
поверхностей. Гидрофобная сторона пленки построена
параллельными палочками. Образование пленки сопровождается быстрым
и очень сильным падением поверхностного натяжения воды. В различ¬
ных отраслях науки и практики часто применяют поверхностно-активные
химические вещества, которые располагаются на поверхности погружен¬
ных в воду частиц того или иного материала и не позволяют отдельным
частицам слипаться, так что образуется равномерная суспензия — смесь
нерастворимых в воде частиц. Гидрофобины — наиболее активные поверх¬
ностно-активные вещества среди известных природных и синтетических
соединений. Всем известно, что мелкие предметы, погруженные в воду,
не могут подняться на поверхность без затраты значительной энергии,
ибо выдвижению из воды препятствует поверхностное натяжение, со¬
здающееся водной пленкой. Как было сказано, молекулы гидрофобинов
на границе вода-воздух полимеризуются, образуя собственную пленку,
которая снижает поверхностное натяжение воды и позволяет гифе под¬
няться из гидрофильной среды в гидрофобную (рис. 9). Без гидрофобинов
грибной мицелий не мог бы выйти из воды и существовать в наземных
условиях. Гидрофобины обнаружены у всех изученных аско- и базидиоми-
цетов, а также у одной из самых древних групп наземных грибов — гло-
меромицетов. Интересно, что мицелиальные бактерии (актиномицеты),
обитающие в почве, также синтезируют поверхностные белки — Чапли¬
ны, химически отличные от гидрофобинов грибов, но также полимеризу-
ющиеся на границе вода—воздух и снижающие поверхностное натяжение
воды. Однако функции гидрофобинов не ограничены только выходом
из водной среды в воздушную. Щцрофобины формируют гидрофобную
пленку и на поверхности спороносцев и спор, благодаря которой споры
сохраняют свои структуры во влажной среде и могут распространяться
в воздухе (не набухают). Щцрофобины обеспечивают также функциони¬
рование крупных плодовых тел грибов макромицетов. Их ложная ткань
(сплетение гиф — плектенхима) пересечена системой многочисленных
воздушных каналов, через которые происходит воздухообмен. У мутантов,
0
Рис. 9. Механизм подъема гифы гриба
из воды в воздух (Wessel J. G. Н., 1999).
Гифа, находящаяся в воде, выделяет в ок¬
ружающую среду молекулы белка гидро-
фобина, которые на границе вода—воздух
полимеризуются (соединяются друг с дру¬
гом и образуют пленку, снижающую по¬
верхностное натяжение воды
28
Глава 2. Мицелиальный образ жизни
не образующих гидрофобины, во влажных условиях каналы закрываются
вследствие взаимодействия гидрофильных поверхностей гиф, поступление
воздуха во внутренние части плодовых тел нарушено, и сами плодовые
тела коллапсируют.
§2. Строение и рост
Итак, жизненная форма большинства грибов — мицелий, представля¬
ющий собой систему более или менее сильно разветвленных нитей (гиф).
У более древних и, следовательно, более примитивных грибов и псевдо¬
грибов мицелий не разделен на перегородки (неклеточный) и образует
единую ветвящуюся трубку. Перегородки у них образуются только при
повреждении какой-то части мицелия (чтобы не вытекло все содержимое,
надо заделать брешь) или при формировании органов размножения. У бо¬
лее продвинутых сумчатых грибов (аскомицетов) весь мицелий разделен
перегородками на отдельные камеры, которые микологи называют клетка¬
ми. Однако клетки аскомицетов имеют много отличий от клеток высших
организмов (растений и животных), поэтому такое название, конечно, чи¬
стая метафора. Клетки растений и животных, как правило, одноядерные,
причем деление ядра (митоз) синхронизировано с делением клетки (ци¬
токинезом), так что после удвоения материнской клетки каждая дочерняя
получает по одному ядру. Клетки могут быть склеены друг с другом, со¬
ставляя единый орган или ткань, но биологически взаимно обособлены
(у животных) или соединены тонкими белковыми нитями — плазмоде-
смами (у растений). У большинства сумчатых грибов клетки, во-первых,
многоядерные, причем число ядер в одной клетке может быть различным,
и, во-вторых, перегородки между клетками имеют отверстие, через кото¬
рое из одной клетки в соседнюю могут мигрировать с очень приличной
скоростью, от одного до сорока сантиметров в час, не только химические
соединения, включая крупные макромолекулы, но и внутриклеточные
органеллы — митохондрии и ядра. Поэтому, если в организме животно¬
го клеточные взаимодействия и, следовательно, регуляция деятельности
клеток, составляющих различные ткани и органы, осуществляется с помо¬
щью химических или электрических сигналов, то мицелий грибов — это
единый организм, взаимодействие между отдельными частями которого
осуществляется вследствие миграции из клетки в клетку пищевых, регу¬
ляторных и информационных молекул. И только у высших базидиальных
грибов (но не у всех) клетки строго одно- или двухъядерные, и поры в пе¬
регородках в течение большей части жизни затянуты перфорированной
мембраной, пропускающей химические соединения, но не ядра.
Важнейшие процессы, сопровождающие рост
и развитие колоний мицелиальных грибов, — это
удлинение кончиков гиф, ветвление и слияние гиф (анастомозы). Все ми-
целиальные грибы растут верхушечными (апикальными) клетками. Апи¬
кальные клетки сумчатых грибов варьируют по длине и числу ядер, в то
§ 2. Строение и рост
29
время как остальные клетки имеют одинаковые размеры и 3-4 ядра. Ядра
в апикальной клетке находятся в состоянии активного деления с синхро¬
низированным митотическим циклом, при котором во всех ядрах клетки
отдельные этапы деления протекают одновременно. В остальных клетках
митозы не происходят (интерфазные ядра) до тех пор, пока не произойдет
ветвление, и клетка станет не апикальной. И тогда в ней начинается актив¬
ная деятельность. Из внутренних частей клетки к периферии направляют¬
ся пузырьки, содержащие углеводы — заготовки для строительства новой
клеточной стенки. Их содержимое выплескивается на поверхность клетки,
где встроенные в оболочку ферменты собирают из них полисахариды —
хитин и глюканы, из которых построена клеточная стенка. Скорость,
с которой строится клеточная стенка, потрясает воображение. Напри¬
мер, паразиты растений, псевдогрибы из рода Фитофтора размножаются
подвижными клетками — зооспорами, покрытыми плотной мембраной,
но не имеющими полисахаридной стенки. Перед прорастанием зооспора
теряет жгутики и покрывается оболочкой (инцистируется), а затем из нее
вырастает гифа, в которую переходит содержимое цисты. Когда капля
с зооспорами попадает на лист, то зооспоры в капле плавают неопреде¬
ленно долгое время, пока случайно не столкнутся с поверхностью листа,
ибо ростку, возникшему в удалении от поверхности, может не хватить
питательных веществ, чтобы дорасти, и он погибнет. Поэтому сигналом
для инсцистирования зооспоры является ее контакт с твердой поверхно¬
стью. Если пробирку с водой, в которой плавают зооспоры, энергично
встряхивать в течение одной минуты, то все находящиеся в ней зооспоры
мгновенно потеряют жгутики и покроются полисахаридными клеточны¬
ми стенками. Этот прием используют исследователи для синхронизации
прорастания зооспор фитофторы. Также быстро протекает и строитель¬
ство клеточной стенки в апикальных клетках мицелиальных грибов. На¬
пример, гифа сумчатого гриба Нейроспора вытягивается со скоростью
16 микрометров (микрон) в минуту, каждые 30-60 минут формируется
и выдвигается вперед новая клетка, а вся колония растет со скоростью
10-14 см в сутки. Для обеспечения растущего с такой скоростью кон¬
чика требуется ежеминутный поток, состоящий из 37 тысяч пузырьков,
наполненных строительным материалом, который выходит из фабрики
полуфабрикатов, называемой комплексом Гольджи, должен достичь рас¬
тущей верхушки и слиться с наружной мембраной. Движение пузырь¬
ков со строительными материалами осуществляется по внутриклеточным
железнодорожным путям, называемым микротрубочками и микрофила-
ментами. Вблизи верхушки клетки рельсы заканчиваются, и пузырьки
поступают на «железнодорожный узел», называемый немецким словом
«spitzenkorper», или «верхушечное тело» (большинство терминов в био¬
логии образованы английскими словами, этот — редкое исключение).
Здесь происходит распределение отдельных пузырьков по необходимым
участкам растущего кончика гифы. Достигнув мембраны клетки, пузырек
30
Глава 2. Мицелиалъный образ жизни
сливается с ней и выливает содержимое на поверхность. А встроенные
в клеточную стенку ферменты соединяют прибывшие строительные ма¬
териалы по правилам, записанным в генетическом коде клетки. В резуль¬
таты этого процесса апикальная клетка вытягивается, и ее удлинившаяся
верхняя часть с перешедшими туда после деления дочерними ядрами от¬
деляется перегородкой. Для обеспечения процессов синтеза необходимых
высокомолекулярных соединений и транспорта их к периферии клетки
необходимо иметь много различных химических веществ и запас энергии.
Однако электрофизиологические эксперименты (погружение микроэлек¬
тродов в мембраны клеток нейроспоры) показали, что в верхушечных
клетках ее мицелия практически отсутствует мембранный потенциал, то
есть верхушечная клетка, в отличие от ее расположенных ближе к центру
соседок, не способна всасывать химические соединения из окружающей
среды. Изолированная от мицелия верхушечная клетка не может расти
и делиться. Рост начинается только при наличии фрагмента из трех -
четырех клеток. Клетки, примыкающие к верхушечной, имеют активный
мембранный потенциал, более высокий, чем у клеток других исследо¬
ванных организмов, который обеспечивает создание протонного насоса,
необходимого для активного поступления метаболитов из окружающей
среды в клетку, а также для мощного тока веществ в направлении верху¬
шечной клетки. Вот для чего нужны отверстия в перегородках.
Ну, хорошо. Вещества для построения клеточ-
Кто-то работает,
кто-то ест нои стенки и ДРУ1*™ элементов клетки притекают в
верхушечную клетку из соседних через поры в пе¬
регородках, а откуда берется необходимая энергия? Оказывается, грибы
нашли простой выход. Плазматическая мембрана внутренних клеток ми¬
целия для создания протонного насоса буквально нашпигована фермента¬
ми, освобождающими энергию, запасенную в аденозинтрифосфате (АТФ).
На площади мембраны, равной 1 квадратному сантиметру, расположено
несколько сотен миллиардов молекул этих ферментов, и каждая молекула
ежесекундно выбрасывает 200-300 протонов, создавая направленный на¬
ружу электрический ток. Протонный насос нейроспоры потребляет около
трети всей энергии, производимой клеткой. Это — плата за возможность
активного всасывания веществ из окружающий среды и транспорта их
по мицелию. Так вот, как сказано выше, верхушечная клетка лишена про¬
тонного насоса (зачем тратиться на активное всасывание веществ из среды
роста, если они поступают из внутренних клеток), поэтому вся энергия
клетки идет на вытягивание и деление материнской клетки на две до¬
черние. Получается разделение функций: одни клетки питаются, другие
растут и делятся. При разделении верхушечной клетки на две только
нижняя приобретает мембранный потенциал, необходимый для создания
протонного насоса, причем, чем дальше от периферии к центру распо¬
ложена клетка, тем ее потенциал выше. Так что мицелий — это единый
организм, участки которого имеют различные функции.
§ 2. Строение и рост
31
Ветвление гиф происходит следующим образом: в отдельных клетках
активизируется синтез гидролитических ферментов — хитиназ и глюканаз,
локально разрушаются основные полисахариды стенки. Поврежденный
участок клеточной стенки размягчается и под действием внутриклеточно¬
го тургорного давления выпячивается. Материнская клетка превращается
в апикальную, в ней начинаются митозы и синтез полуфабрикатов —
заготовок для дальнейшего вытягивания и укрепления клеточной стенки
выроста. После перехода в него разделившихся ядер вырост отделяется
оболочкой от материнской клетки, которая успокаивается, а отросток от
нее приобретает свойства апикальной клетки.
Наконец, еще одно важное свойство колонии мицелиальных гри¬
бов — анастомозы гиф. Анастомозы происходят при контакте соседних
гиф в процессе разрастания колонии или при встрече двух колоний од¬
ного вида гриба. Внутри колонии сливаются не концы гиф, а растущие
навстречу боковые ветви, которые выделяют вещества, вызывающие под¬
растание встречных гиф друг к другу. При встрече двух колоний сливаются
апикальные клетки их лидирующих гиф или апикальная клетка одной ко¬
лонии и внутренняя клетка другой. Первый этап анастомоза — слипание
контактирующих поверхностей. Наружный слой клеточной стенки гри¬
бов построен из глюканов, разветвленные веточки которых при контакте
клеток сцепляются и вызывают склеивание поверхностей. Затем клеточ¬
ная стенка в месте контакта растворяется под действием внутриклеточных
ферментов и содержимое клеток сливается.
Анастомоз соседних гиф арми¬
рует колонию, делает ее более проч¬
ной, интегрирует отдельные ее ча¬
сти в единое целое (рис. 10). Если
же сливаются клетки разных коло¬
ний, то возможен обмен ядрами и
их миграции в противоположных
направлениях. Уже говорилось, что
клетки многих грибов многоядерны
и в перегородках между клетками
у большинства грибов имеются от- Рис 10 Анасгомозы межДу гифами
верстия, через которые ядра могут оомицета фитофтора (71 А. Кузовни-
мигрировать из одной клетки в дру- ковой, Ю. Т. Дьякова, 1973)
гую. Если ядра в анастомозирую-
щих клетках двух колоний генетически различаются, то в одну клетку
могут попасть разные ядра. Такое состояние разноядерности (гетерокари¬
оза) — еще одна особенность грибов. Гетерокариоа увеличивает приспосо¬
бительные возможности грибов, так как генетически различные ядра несут
информацию о синтезе продуктов, способствующих выживанию в изме¬
няющихся условиях жизни.
32
Глава 2. Мицелиальный образ жизни
§3. Питание
Выше было сказано, что грибы, во-первых, не умеют синтезировать
органические вещества из неорганических, поэтому извлекают их из жи¬
вых или мертвых органических субстратов. Во-вторых, для прохождения
через клеточные покровы высокомолекулярные полимерные соединения
должны быть предварительно разбиты ферментами на отдельные фраг¬
менты — олиго- или мономеры. Поэтому мицелий выделяет в окружа¬
ющую среду набор необходимых для расщепления субстрата ферментов.
Так как полимерных органических веществ в природных субстратах ве¬
ликое множество, то и ферментов должно выделяться адекватное чис¬
ло. Но этого не выдержит энергетика никакой клетки. Поэтому синтез
и выделение ферментов у грибов находится под сложнейшим контро¬
лем. Во-первых, это — субстратная индукция. Если в субстрате (будь
то яблоко или колба с искусственно приготовленной питательной сре¬
дой) есть распространенный у растений полисахарид пектин, то в клетках
гриба будут синтезироваться и выделяться наружу ферменты, расщепляю¬
щие пектин. Если пектин удалить из субстрата, синтез соответствующих
ферментов прекратится. Это первый шаг к претворению в жизнь ло¬
зунга, провозглашенного когда-то JI. И. Брежневым: «Экономика должна
быть экономной». Во-вторых, для разрушения большинства биополиме¬
ров, в частности, того же пектина, грибы имеют набор разных ферментов,
воздействующих на разных химические связи в макромолекуле и расщеп¬
ляющих ее с образованием различных продуктов. В-третьих, синтез одного
и того же фермента может контролироваться семейством генов, причем
каждый изофермент имеет некоторые отличия от других изоферментов
данного семейства, например, оптимально работает при разных темпера¬
турах или разных показателях кислотности среды. Это позволяет в разных
условиях выбирать оптимальное средство для превращения в пищу всего,
что окружает мицелий. Наконец, имеется еще один существенный ре¬
гуляторный механизм, делающий экономику грибной клетки еще более
экономной, — катаболитная репрессия. Она заключается в том, что на¬
личие в субстрате энергетически более выгодного (легче усваиваемого)
соединения препятствует образованию ферментов, необходимых для рас¬
щепления энергетически менее выгодного соединения. Если в субстрате
имеет только полисахарид пектин, гриб, оккупировавший этот субстрат,
будет производить и выделять в среду ферменты, разрушающие пектин
и превращающие его в глюкозу. Но если к пектину добавить глюкозу, то
эти ферменты перестанут синтезироваться. Все очень понятно: для чего
тратить энергию для синтеза новых белков и расщепления сложных ве¬
ществ, если имеется уже готовый конечный продукт. Зная этот механизм,
один американский профессор лечил томаты, зараженные грибом фуза-
риум, опрыскивая их сахарным сиропом. Паразит бросал, тяжело дыша,
расщеплять полисахариды зараженного растения и переключался на пред¬
ложенную ему дармовщину.
§ 4. Что можно сделать из разветвленных ниток
33
Некоторые грибы — узкие специалисты, выде¬
ляющие определенный, ограниченный набор фер¬
ментов, необходимых для расщепления полимеров
только одного типа, например, трутовые грибы, об¬
разующие копытообразные плодовые тела на деревьях, имеют очень актив¬
ные ферменты, разрушающие целлюлозу и лигнин — полимеры, из кото¬
рых построены стенки клеток древесины, а малосимпатичные грибы, вы¬
зывающие дерматомикозы (поражение ногтей, кожи, волос, перьев млеко¬
питающих и птиц), образуют ферменты, разрушающие кератин — белок,
из которого построены покровы животных. Но большинство грибов менее
разборчиво и образуют разнообразные ферменты, позволяющие оккупи¬
ровать и использовать различные субстраты.
§ 4. Что можно сделать из разветвленных ниток
Выше было сказано, что основная функция мицелия — питательная,
и его устройство, напоминающее вывернутую наизнанку кишку, обеспе¬
чивает оптимальные условия для адсорбционного питания. Но «не хлебом
единым» сыт не только человек, но и гриб, телу которого необходимы
вариации, обеспечивающие адаптацию к изменяющимся условиям окру¬
жающей его действительности. И для обеспечения адаптаций гифы мице¬
лия могут образовывать структуры, совершенно на них не похожие. Есть
детская игра, в которой из квадрата белой бумаги, складывая его опре¬
деленным образом, можно получить различные фигурки. Так поступают
и грибы, образуя из нитей структуры, на нити совсем не похожие.
Иногда оболочки отдельных клеток мицелия, а у не¬
которых грибов и всех клеток, утолщаются и темнеют,
так как в них откладывается темный пигмент — меланин.
После этого весь мицелий или участки с такими клетками разваливаются,
и образуются покрытые прочными оболочками споры, способные дли¬
тельный период сохраняться в почве или в растительных остатках. Такие
споры называют хламидоспорами. После наступления благоприятных для
жизни условий (например, прекращения зимних морозов или летней за¬
сухи) хламидоспоры прорастают ростковой трубкой и формируют новый
мицелий.
Многие грибы поступают по-другому. Их гифы в отдельных участ¬
ках мицелия плотно сплетаются, клетки укорачиваются, и образуются
клубочки, называемые склероциями. У многих грибов склероции быва¬
ют структурированы: их внутренние части построены клетками с богатым
питательными веществами содержимым и с тонкими оболочками. Эти
клетки предназначены для питания прорастающего склероция. Наружные
участки склероция состоят из более мелких клеток, имеющих толстые,
окрашенные меланином оболочки. Такой «коровой» слой клеток защи¬
щает склероции от возможных повреждений. У многих грибов склероции
34
Глава 2. Мицелиальный образ жизни
Проводящие
и поддерживающиеся
структуры
достигают очень больших (конечно, в грибных масштабах) размеров, обра¬
зуя черные шарики размером с горошину или фиолетовые рожки длиной
несколько сантиметров (рис. 25, стр. 89). А у некоторых — черные мелани-
зированные гифы оплетают субстрат, в котором они находятся, превращая
его в один большой склероций. Все садоводы знают плодовую гниль яблок,
которую вызывает гриб монилия. Яблоко сначала буреет, его содержимое
размягчается, и оно падает с дерева. Еще находясь на ветке, или на земле
оно чернеет, становится жестким и целиком превращается в склероций.
В таком виде гриб переносит зимние морозы.
Хламидоспоры и склероции играют огромную роль в жизни грибов.
Например, в экстремально жаркое и сухое лето 2010 года содержание
влаги в лесных почвах Средней полосы России упало до критического
минимума, и вегетативный мицелий потерял жизнеспособность. Однако
после того, как осенью пошли сильные дожди, наблюдалось массовое
плодоношение грибов. Такое явление стало возможным только благодаря
наличию в почве хламидоспор и склероциев, в форме которых эти грибы
пережили тяжелый период засухи.
Кто хоть раз гулял в лесу, не мог не обратить
внимание на огромные деревья, от основания до
верхушки покрытые копытообразными плодовы¬
ми телами трутовых грибов. Мицелий находится
внутри дерева, а плодовые тела выходят на поверхность через различ¬
ного рода ранки в коре дерева. В трубоч¬
ках или складках, покрывающих нижнюю
поверхность плодовых тел, формируются
споры, которые после созревания высы¬
паются и распространяются воздушными
токами. Тонкие гифы мицелия диаметром
в несколько микрон, конечно, не могут
обеспечить единство тела гриба на протя¬
жении десятков метров высоты дерева, да
еще в такой плотной среде, как древесина,
где приходится продвигаться буквально
с помощью локтей («локтями» служат, ко¬
нечно, ферменты). Поэтому гифы древо¬
разрушающих грибов срастаются боковы¬
ми стенками и образуют различного рода
пленки и тяжи настолько толстые, что хо¬
рошо видны в разрушенных участках дре¬
весины. Через центральные гифы в таких
шнурах транспортируется вода и элемен¬
ты питания по всему мицелию. Избытки
насосанной воды в виде капель отклады¬
ваются на плодовых телах. А у осеннего
Рис. 11. Ризиморфы осеннего
опенка в пораженной древесине
(£. П. Кузьмичев с соавт., 2004)
§ 4. Что можно сделать из разветвленных ниток
35
опенка, который не только является любимым и всегда обильным объек¬
том тихой охоты, но и разрушительным паразитом различных пород де¬
ревьев, в пораженной древесине обнаруживаются толстые разветвленные
мицелиальные шнуры, в стенках наружных клеток которых откладывает¬
ся темный пигмент меланин. Благодаря этому тяжи становятся прочны¬
ми, как проволока (рис. 11). Выходя из корней зараженного дерева, они
способны расти в почве и заражать корни соседних здоровых деревьев.
Кроме того, в их клетках происходят такие же химические реакции как
в клетках насекомых светлячков и многочисленных светящихся морских
организмов. Поэтому когда в народе говорят, что гнилушки светятся, на¬
до понимать, что светится не пораженная древесина, а находящиеся там
тяжи (ризоморфы) опенка.
Многие грибы паразитируют на различных расте- . .
Инфекционные
ниях и животных, и для заражения своих хозяев они ^ структуры
выработали различные приспособления.
Грибы-паразиты растений разделяют на две большие группы — не-
кротрофов и биотрофов. Первые могут извлекать питательные вещества
только из мертвых клеток и тканей растений, поэтому они убивают расти¬
тельные клетки своими токсическими выделениями и распространяются
по мертвой зоне, как паразиты теплого трупа. Зараженная ткань разру¬
шается, ее клетки распадаются, и образуется бесформенная гниль. Все
дачники знают серую гниль садовой земляники, при которой поверхность
плода покрывается серым пушистым налетом спор гриба, а ткань плода
под налетом сгнивает. По-иному ведут себя биотрофы. Они извлекают пи¬
щу из содержимого живых клеток, гибель зараженной клетки означает для
них гибель гриба, находящегося в ней. Не случайно растения выработали
оригинальный способ защиты от биотрофных паразитов — гибель зара¬
женных и окружающих их клеток, названная реакцией гиперчувствитель¬
ности — одной из форм программируемой гибели клеток или апоптоза.
Таким способом растение, ценой гибели отдельных участков своего тела,
освобождается от инфекции. Для входа в кладовую, называемую расти¬
тельной клеткой, паразит должен преодолеть два барьера: полисахаридную
клеточную стенку и окружающую содержимое клетки мембрану. Первый
барьер биотрофные грибы преодолевают с помощью своих ферментов,
в частности, пектиназ, о которых мы говорили ранее. В стенке образуется
отверстие, через которое инфекционная гифа входит в контакт с мембра¬
ной. Но мембрана — не стенка, ее разрывать, как это делают некротроф-
ные паразиты нельзя, иначе клетка потеряет структурную целостность,
погибнет и перестанет представлять ценность для биотрофного паразита.
Поэтому инфекционная гифа очень аккуратно, интеллигентно надавли¬
вает на мембрану, заставляя ее прогибаться внутрь, инвагинировать. Об¬
разуется мешок, внутри которого располагается отросток инфекционной
гифы, названный гаусторием. У разных грибов формируются гаустории
различной формы — от простого расширения кончика инфекционной
36
Глава 2. Мицелиальный образ жизни
Рис. 12. Гаусторий возбудителя ржавчины в клет¬
ке льна (Hartman D., 2007): НМС — материн¬
ская клетка гаустории; ehm — экстрагаустори-
альная мембрана — участок наружной мембраны
(плазмалеммы) растительной клетки, вдавленной
внутрь клетки гаусторием; Н — гаусторий
гифы (рис. 12) до сильно разветвленного
кустика, который так и называют — арбу-
скулой, то есть деревцом по латыни. Та¬
ким образом, гаусторий паразита находит¬
ся внутри клетки, но вне ее протопласта.
Как же он питается? Ранее я рассказывал,
как в апикальной клетке грибной гифы
для строительства клеточной стенки к поверхности устремляются мем¬
бранные пузырьки, заполненные строительным материалом. Пузырьки
сливаются с наружной мембраной и выливают содержимое на ее поверх¬
ность, где с помощью ферментов собирается новая стенка. То же происхо¬
дит и с зараженной клеткой растения. Оголенный, лишенный клеточной
стенки участок плазматической мембраны, окружающий гаусторий, во¬
пиет: «одень меня, заделай брешь», посылая об этом сигналы в клетку.
Оттуда устремляются пузырьки с углеводами, которые, сливаясь с мем¬
браной, выливают содержимое на ее поверхность. Но вместо того, чтобы
пойти на строительство клеточной стенки, содержимое пузырьков пере¬
хватывается гаусторием, получающим полноценное углеводное питание.
§5. Дрожжи
Дрожжи — это дрожжи, а не мицелий, особая жизненная форма
грибов. Однако, во-первых, дрожжевые грибы произошли от мицелиаль-
ных, поэтому их можно рассматривать как форму мицелиальных грибов,
адаптированную к определенным условиям жизни.
с° еннь,е Во-вторых, есть грибы, которые в любых условиях
гриоы дрожжи ✓
жизни формируют мицелии (или умирают, если из¬
менение условий заходит за границы дозволенных для жизнеобеспечения);
есть грибы, которые в любых условиях жизни остаются дрожжами, а есть
виды, которые на изменение условий жизни реагируют сменой мицели-
альной формы на дрожжевую. Это явление получило название «мице-
лиально-дрожжевой диморфизм». Следовательно, в геноме грибов очень
давно, на первых этапах их жизни на суше, возникли две программы
развития — мицелиальная и дрожжевая, причем переключение развития
с одной программы на другую регулируется условиями жизни. У толь¬
ко мицелиальных и только дрожжеподобных грибов одна из программ
оказалась утраченной, а у мицелиально-дрожжевых — сохранились обе
§5. Дрожжи
37
программы. Какие же условия жизни вызывают переключение программ
развития у диморфных грибов?
Прежде всего — состав среды их обитания. 1де в
природе растут дрожжи? Там, где много простых са- Среда 0 lfTaHH?
харов — моносахаров глюкозы, фруктозы, дисахарида дрожжей
сахарозы и прочих — на повреждениях коры деревьев весной, когда из мест
повреждения вытекают растительные соки (например, березовый), нака¬
чиваемые корнями, на гниющих ягодах и фруктах, в которых под воздей¬
ствием собственных ферментов или ферментов паразитирующих на них
бактерий и грибов происходит разрушение полисахаридов и накопление
простых сахаров. Вероятно, для такого простого питания готовыми моно-
и дисахарами дрожжевая жизненная форма подходит лучше, чем мицели-
альная. Это правило, по-видимому, характерно и для диморфных грибов.
Многочисленные плесневые грибы из семейства мукоровых образуют ми¬
целий внутри различных органических субстратов, а на поверхности суб¬
стратов формируются спороношения — тонкие, длинные белые ножки
с черными головками, внутри которых находятся, споры. Но на богатых
простыми сахарами субстратах мицелий некоторых мукоровых грибов рас¬
падается на отдельные клетки, которые размножаются как дрожжи.
На выбор стратегии жизни диморфных грибов влияет и газовый со¬
став среды. Дело в том, что большинство мицелиальных грибов, как и мы,
для разложения глюкозы используют кислород в процессах, называемых
дыханием. С помощью кислорода происходит полное окисление молеку¬
лы глюкозы, которая через ряд промежуточных соединений превращается
в углекислый газ и воду. Образующаяся при распаде глюкозы энергия
идет на проведение синтезов, необходимых для жизни клетки, или со¬
храняется в соединениях, запасающих энергию, и расходуется по мере
необходимости. Поскольку для дыхания кислород абсолютно необходим,
то в отсутствии кислорода большинство грибов, так же, как и мы, погибает.
Но некоторые мицелиальные грибы и все дрожжи обладают и альтерна¬
тивным способом разложения глюкозы, названным брожением. При бро¬
жении распад глюкозы неполный, и наряду с углекислым газом образуется
не вода, а этиловый спирт. Благодаря этим свойствам дрожжи использу¬
ют в хлебопечении (углекислый газ поднимает тесто) и виноделии (спирт
превращает растительные соки в вина). Для брожения кислород не нужен,
поэтому они могут утилизировать сахара и в бескислородной (анаэроб¬
ной) среде. В связи с этим некоторые мицелиальные мукоровые грибы
в бескислородной среде превращаются в дрожжи.
Для некоторых грибов сигналом для смены жиз-
„ ^ Разные стратегии
ненных стратегии являются изменения способов пи- одного гриба
тания. У некоторых групп грибов, например, сум¬
чатых (тафриновых) и базидиальных (головневых), паразитическая фаза
представлена мицелием, который биотрофно развивается внутри пора¬
женных растений; вне растений эти грибы могут жить на мертвом орга¬
0
38
Глава 2. Мицелиальный образ жизни
ническом субстрате, но только в дрожжевом состоянии. А некротрофные
паразиты растений из рода вертициллиум, наоборот, живут в почве в ми-
целиальном состоянии и питаются растительными остатками, а в зара¬
женных растениях находятся в сосудах, по которым от корней к листьям
и цветкам идет поток воды с растворенными минеральными веществами,
и там гриб существует в форме отдельных почкующихся клеток.
Многие грибы, паразиты позвоночных животных и человека, диморф¬
ны. Они живут в почве в мицелиальном (плесневом) состоянии и питаются
сапротрофно растительными остатками. Если через ранки или естествен¬
ные отверстия они попадают в организм человека, то превращаются там
в дрожжи. Таким образом, мицелиальная стадия у них непаразитическая,
а паразитируют они в дрожжевой. Причем переход к дрожжевому состоя¬
нию регулируется температурой тела (37 °С). При повышенной темпера¬
туре в клетках синтезируются так называемые белки теплового шока, ко¬
торые относятся к группе регуляторных белков, влияющих на экспрессию
генов и обеспечивающих приспособление организма к жизни в условиях
повышенной температуры.
Дрожжевой образ жизни принципиально отличается от мицелиально-
го. Мы его рассмотрим на примере дрожжей-сахаромицетов, которые ис¬
следованы более полно, чем другие виды, во-первых, вследствие их прак¬
тической важности, а во-вторых, потому что на сахаромицетных дрожжах
проводится много исследований фундаментального
Отличия
дрожжевой формы характера, и сделаны важные для клеточной и мо-
от мицелиальной: лекулярной биологии открытия. Жизнь дрожжевой
питание клетки отличается от жизни мицелия прежде все¬
го в особенностях обмена веществ (метаболизма).
Дрожжи в отличие от мицелиальных грибов имеют слабые гидролитиче¬
ские ферменты, поэтому сложные полисахариды им не по зубам. Зато
они могут извлекать энергию из простых сахаров даже в условиях дефи¬
цита кислорода в реакциях брожения. Отличаются дрожжи от мицелиаль¬
ных грибов и по химическому составу. Полисахариды клеточной стенки
мицелиальных грибов представлены двумя основными компонентами —
хитином, обеспечивающим прочность стенки, и глюканами. У сахароми¬
цетных дрожжей очень мало хитина в стенке, поэтому их стенки более
эластичные, что важно для обеспечения процесса почкования, и преобла¬
дающий полисахарид не полимер глюкозы глюкан, а полимер ее изомера
маннозы — маннан. Причем у диморфных грибов перестройка состава
клеточной стенки при переходе из одного морфотипа в другой осуществ¬
ляется чрезвычайно быстро.
Наконец, совершенно по-разному осуществля-
Отличия ется т и деление у мицелиальных грибов, как
дрожжевой формы /
от мицелиальной: было сказано, рост поляризован, то есть растут
рост и делятся только верхушечные (апикальные) клетки.
После деления в них ядер происходит разделение
§5. Дрожжи
39
верхушечной клетки на две путем врастания (впячивания) внутренних
слоев оболочки. У дрожжей рост клетки диффузный, может произойти
в любом месте, причем выделены определенные белки, которые локали¬
зуются в мембране, обеспечивают транспорт к мембране ионов кальция,
обозначая место возникновения почки. В этом месте начинают проте¬
кать два процесса. Во-первых, выпячивание (не прогибание внутрь, как
у мицелиальных грибов, а именно выпячивание) оболочки с образованием
растущей припухлости. Во-вторых, митотическое деление ядра, образова¬
ние двух одинаковых ядер и транспорт одного из них в зону почки. Когда
почка увеличится до необходимых размеров, начинается синтез содержа¬
щего азот полисахарида хитина, которого, в отличие от мицелиальных
грибов, в клеточной стенке дрожжей очень мало, и строительство хити¬
новой пластинки, отделяющей почку от материнской клетки. Затем место
соединения почки с материнской клеткой заделывается нормальной для
дрожжей (нехитиновой) стенкой. Хитиновая пластинка становится не¬
нужной, и в работу вступает фермент, разрушающий ее. После отделения
почка дорастает до размеров материнской клетки и сама начинает поч¬
коваться. Продолжительность всего цикла от почки до почки составляет
около 100 минут. Понятно, что в благоприятных условиях, на сахаристых
субстратах, происходит взрывоподобный рост биомассы дрожжей.
Из рассказанного процесса почкования дрожжей вы¬
текает еще одна особенность размножения дрожжей. При размножение
делении одноклеточных организмов, размножающихся и старение
делением, возникают две физиологически одинаковые
клетки, то есть бактериальная клетка бессмертна, ибо ни об одной об¬
разующейся делением клетки нельзя сказать, что это мама, а это — дочка.
На самом деле, все не совсем так, ибо так называемые факторы ста¬
рения могут неравномерно распределяться между дочерними клетками,
но об этом позже. У дрожжей материнская клетка и почка физиологиче¬
ски различны. Почка омоложена, а материнская клетка с каждым актом
почкования стареет. Было уже сказано, что перед отделением почки от¬
деляющая ее хитиновая пластинка растворяется, но не целиком: на месте
отделения почки от материнской клетки остается хитиновое кольцо —
рубчик, или детское место, видное в электронно-микроскопических пре¬
паратах (рис. 13). Оказалось, что у дрожжей, как и у наших женщин,
имеется определенный репродуктивный период. Через 25-30 почкований
клетка прекращает почковаться и погибает. В любой дрожжевой попу¬
ляции 50 % клеток составляют не почковавшиеся девственницы, 25 % —
клетки, прошедшие одно почкование, 12,5 % — клетки, образовавшие две
почки, и т. д. Многократно почковавшиеся (более 20 раз) «старухи» состав¬
ляют ничтожную часть популяции. Так что демографический состав дрож¬
жевых популяций по качеству гораздо выше демографического состава
популяций людей, особенно в наше время, когда доля работающих долж¬
на кормить такую же по численности долю неработающих пенсионеров.
40
Глава 2. Мицелиальный образ жизни
Рис. 13. Электронно-микроскопическиие снимки почкующихся дрожжевых клеток.
На поверхности клетки видны хитиновые кольца, указывающие на места почкования.
П — почка (Alexopolous C.J. et al., 1996 и Carlig M.J. et a!., 2006)
Хотя сахаромицетные дрожжи не диморфны, в их геноме сохранена
память о возврате к мицелиальному типу роста. В условиях азотного го¬
лодания у них образуется псевдомицелий: клетки вместо овальных ста¬
новятся вытянутыми, они почкуются не где попало, а, как и положено
мицелиальным структурам, строго апикально, и почки не отделяются от
материнской клетки, вследствие чего образуются цепочки из дрожжевых
клеток. Такой тип роста, по-видимому, позволяет лучше осваивать бедный
питанием субстрат.
§6. Грибы-экстремалы
Грибы живут рядом с нами, большинство из них предпочитают сход¬
ные с нами условия жизни. Наиболее комфортная для их роста температу¬
ра находится между 20 °С и 30 °С, им нравится высокая влажность воздуха
и чтобы пища была не слишком кислая, но и, не дай бог, не щелочная.
Однако есть масса грибов-экстремофилов, для которых наши «средние»
условия жизни совсем не оптимальны. Они живут, и хорошо живут, в усло¬
виях, которые по нашим понятиям вообще являются пограничными для
возможности существования.
Многие грибы активно развиваются при крайне
1 низких температурах. Плесневение продуктов, храня-
ЭКСТрбМаЛЫ
щихся в холодильниках, — обычное явление. А есть
большая группа грибов — паразитов растений, которые развиваются при
температуре, близкой к нулю. Они вызывают плесневение растений во вре¬
мя зимних оттепелей и при весеннем таянии снега и получили название
снежных плесеней. Эти грибы сильно вредят посевам озимых злаков, цве¬
точным культурам, клеверу и другим полезным растениям. Но мало этого...
Грибные структуры находят в почвах зоны вечной мерзлоты в Сибири
§6. Грибы-экстремалы
41
и даже в шурфах, проделанных в ледниках Антарктиды, причем они со¬
храняют там свою жизнеспособность тысячелетиями. А есть и грибы-тер-
мофилы,. которые отлично себя чувствуют при температурах до 50 °С. Так
что температурные траницы, обеспечивающие возможность существова¬
ния разных грибов, огромны и уступают только прокариотным бактериям.
Но на то они и бактерии: крайне примитивное строение клеток и быстрое
размножение создают условия для чрезвычайно широких адаптаций.
Много лет назад, будучи аспирантом, я сидел в од-
м „ Кислотные
ной комнате с сотрудницей, которая выращивала в чаш- экстремалы
ках Коха на жидкой питательной среде колонии гриба
черного аспергилла. Я заметил, что она не стерилизует в автоклаве пита¬
тельные среды перед их засевом, и спросил, почему она не делает этого?
Она ответила, что перед посевом гриба подкисляет среду до pH 2, и при
такой кислотности никто, кроме черного аспергилла, не растет. Такой уро¬
вень кислотности имеет сантинормальный раствор серной кислоты или
неразбавленный сок лимона! Так что среди грибов есть чемпионы мира
по устойчивости к кислотам. Кстати, микологи, чтобы при выделении
грибов из почвы подавить рост бактерий, подкисляют питательную среду.
Но не надо думать, что все грибы ацидофилы (любят кислую среду). Есть
среди них и алкалофилы, развивающиеся в щелочных почвах и водоемах.
На земле имеется несколько мест, в которых расположены щелочные со¬
довые озера. Их pH близок к абсолютному максимуму — 11-12 единиц!
Крупнейший наш микробиолог Г. А. Заварзин считал, что такими были во¬
доемы на ранних этапах эволюции земли, в докембрийские периоды, когда
планета была заселена только безъядерными клетками бактерий и архей,
а ядерные организмы еще не возникли. Поэтому население содовых озер
может отражать прокариотное население ранних этапов эволюции биоты
земли. Оказалось, однако, что в существующих ныне содовых озерах и вы¬
сохших (содовых солончаках) живут не только прокариоты, но обитают
и водоросли, и многочисленные грибы, и растения солянки, и даже рачок
Артемия салина. Выделенные оттуда грибы могли расти на питательной
среде, имеющей показатель pH 10 и даже 11 единиц! Недавние иссле¬
дования показали, что в таких условиях обитают новые для науки виды,
роды и даже семейства грибов. Поскольку кислотность внутриклеточного
содержимого у них, как и других организмов, близка к нейтральному пока¬
зателю (pH около семи), разница между наружными и внутриклеточными
показателями кислотности может составлять 4 единицы pH. Поэтому жи¬
вущие в таких условиях организмы выработали механизмы поддержания
клеточного кислотного гомеостаза с помощью наличия активного насоса,
выбрасывающего из клетки ионы натрия, наличия специальных осмоти¬
ческих вакуолей (внутриклеточных мембранных мешочков), участвующих
в процессах осморегуляции, и других особенностей в строении и обмене
веществ этих грибов.
42
Глава 2. Мицелиальный образ жизни
§ 7. Грибы-поденки и грибы-секвойи:
продолжительность жизни разных грибов
Общепринято представление о грибах, как об эфемерных организмах,
которые в массе высыпают перед удивленным взглядом при наступлении
благоприятных условий и также быстро исчезают, когда эти условия за¬
канчиваются. Это касается как грибов образующих плесени, так и шля¬
почных грибов, растущих в лесах. На самом деле мы наблюдаем не появ¬
ление и гибель самих грибов, но лишь их спороносных структур и спор.
А сам мицелий скрыт в том или ином субстрате и сведений о его возрасте
и продолжительности жизни мы так просто получить не можем. Но экспе¬
рименты с мицелиями разных грибов показывают, что продолжительность
их жизни, как говорят англичане, life span, может различаться в тысячи
раз. Есть грибы, которые, как некоторые насекомые-поденки, живут всего
несколько дней, а есть виды грибов, жизнь которых продолжается, подоб¬
но жизни могучих деревьев — секвой — тысячелетиями.
Выше говорилось о почковании дрожжей, клетки которых после
25-30 почкований погибают. Следовательно, дрожжевая клетка стареет,
причем вредные для клетки продукты «факторы старения», накапливаю¬
щиеся в течение жизни, неравномерно распределяются между материн¬
ской клеткой и почкой. Материнская клетка, как бы жертвуя собой, остав¬
ляет факторы старения у себя, тем самым омолаживая почку. Однако при
накоплении «факторов старения» выше определенного уровня, асиммет¬
рическое их распределение становится невозможным, вследствие чего до¬
черние клетки становятся такими же старыми, как материнская. По этой
причине при последних почкованиях дрожжей образующиеся почки уже
не почкуются, ибо оказываются такими же старыми, как и их мамы. Тип
старения, характерный для дрожжевых клеток, называют репликативным
старением.
Для изучения особенностей и продолжительности жизни мицелиаль¬
ных грибов их выращивают в ростовых трубках, которые представляют
собой стеклянные трубки, длиной до метра, один или оба конца которых
отогнуты и не запаяны. В стерильную ростовую трубку заливают слой
стерильной агаризованной питательной среды, застывающей после охла¬
ждения, с одного края среды вносят мицелий гриба, и отверстие в трубке
закрывают стерильной ватной пробкой. В таких трубках можно изучать
скорость и характер роста грибов в течение длительного периода. Более
полувека назад французский миколог Ризе при выращивании в росто¬
вых трубках обнаружил у сумчатого гриба Подоспора, живущего в навозе
травоядных животных, признаки старения и умирания мицелия. Вообще,
обитающие в навозе, так называемые копротрофные грибы — это уди¬
вительная группа, не перестающая изумлять микологов своими формами,
особенностями физиологии, парадоксальными жизненными циклами. Не¬
вольно приходят на память стихи Ахматовой: «Когда б Вы знали, из какого
§ 7. Продолжительность жизни разных грибов 43
сора растут стихи, не ведая стыда». Мы еще будем к ним возвращаться.
А сейчас возвратимся к грибу Подоспора. Лет тридцать после обнаружения
у этого гриба старения никто не мог не только установить причину этого
явления, но и не имел подходов к исследованию данного феномена. Затем
на совершенно иных объектах было установлено, что большинство генов
эукхариотных организмов имеют мозаичную структуру, то есть состоят
из транслируемых участков — экзонов, которые участвуют в матричном
синтезе белков — трансляции, и нетранслируемых участков — интронов,
которые вырезаются до трансляции. При использовании соответствую¬
щих технологий было обнаружено, что в ДНК митохондрий Подоспоры
один из интронов гена, кодирующего синтез важного фермента цитохро-
моксидазы, после вырезания образует циклические структуры, которые
самореплицируются (размножаются) в цитоплазме. Поэтому такие интро-
ны называют мобильными. Затем эти нитроны могут опять встраиваться
в разные места митохондриальной ДНК и вызывать вырезание соседних
участков (делеции). Эго приводит к нарушению деятельности важнейших
энергетических фабрик клетки — митохондрии, что внешне выражается
как старение и смерть клеток мицелия. Дальнейшие исследования этого
интереснейшего гриба показали, что мобильные митохондриальные ни¬
троны — не единственный и даже не главный механизм старения. Глав¬
ным механизмом старения у этого гриба, как и у дрожжей, по-видимому,
является репликативное старение. Выше было сказано, что все мицели¬
альные грибы растут путем деления верхушечной (апикальной) клетки.
При делении верхушечной клетки на две, та, что образуется на верхушке,
становится дочерней, а соседняя с ней со стороны мицелия — материн¬
ской. В результате асимметричного распределения «факторов старения»
верхушечная дочерняя клетка всегда остается молодой, остальные клет¬
ки, чем глубже они расположены к центру колонии, тем они старше. Так
вот у Подоспоры мы имеет перевернутую картину: при делении клеток
материнской становится верхушечная, апикальная клетка, поэтому волна
стареющих клеток движется не от центра к периферии, а, наоборот, —
от периферии к центру. Поэтому край колонии гриба погибает от старения
клеток, а центр — от голодания. А что же мобильные интроны? Они что,
не играют никакой роли. Нет, они играют важную роль, ускоряя гибель
стареющих клеток. Их роль аналогична роли «контрольного выстрела»
в работе модной сейчас профессии киллера.
С другой стороны, грибы могут формировать гигант¬
ские колонии, превышающие все другие организмы на ри ы"
земле протяженностью как в пространстве, так и во време¬
ни. Многие видели на лугах, а иногда и в лесах, так называемые ведьмины
кольца — одновременное образование большого числа плодовых тел шля¬
почного гриба, расположенных по периметру довольно большой окружно¬
сти. В центре кольца находился первичный мицелий, который распростра¬
нялся в почве в разные стороны примерно с одинаковой скоростью. Через
44
Глава 2. Мицелиальный образ жизни
некоторое время на окончаниях мицелия формировались плодовые тела,
показывающие, какую площадь заняла колония. Из года в год колония
может разрастаться, а диаметр колец увеличиваться. Однако чемпионом
по площади, занимаемой одним грибным индивидом, являются не поч¬
вообитающие грибы, а паразит деревьев — осенний опенок. Пучки его
мицелия, покрытые толстой меланизированной оболочкой (ризоморфы,
рис. 11), выходят из корней зараженного дерева и могут, распространяясь
в почве, заражать корни соседнего дерева. Таким способом очаг заражен¬
ных опенком деревьев медленно распространяется. При фотографирова¬
нии лесного массива с самолета, очаг выглядит как более светлый участок
леса среди зеленого массива (вследствие того, что в листьях пораженных
деревьев происходит распад хлоропластов), так что можно определить его
площадь. Есть простые молекулярные методы, позволяющие отделить ми¬
целий опенка, принадлежащий одному экземпляру, от мицелиев других
экземпляров и оценить годовую скорость роста очага. Используя комплекс
этих методов, в лесных почвах США был обнаружен индивидуум осеннего
опенка, который распространился на площади в несколько гектар, имел
вес мицелия 10 тонн и возраст около 1500 лет, т. е. это — самый большой
и самый старый организм на земле. Его временная генетическая стабиль¬
ность охватывает 10 миллионов митотических генераций!
§ 8. Что такое грибная особь
Что такое отдельный организм или особь у большинства животных
(человека, вороны или мухи) всем понятно. В написанной в XI веке книге
«Изборник», содержащей, в частности, изложение на славянском языке
упоминавшегося труда Иоанна Дамаскина «Диалектика», сказано: «Подо¬
бает знать, что святые отцы ипостасью, атомом и лицом называли одно
и то же — существующее само по себе, состоящее из существа и слу¬
чайностей». Очень интересное определение. Во-первых, лицо, особь —
это нечто неделимое, в славянском переводе «несекомое», конечный этап
классификаций; во-вторых, важнейшие свойства особи в определении
Дамаскина и современных генетиков совпадают — особь состоит из су¬
щества или, как говорят современные биологи, наследственности (не¬
которой суммы признаков, присущих всем представителям следующей,
более высокой иерархии в классификационных схемах — виду) и слу¬
чайностей, или, по-современному, изменчивости, благодаря которой нет
двух абсолютно одинаковых людей, волков, коней, собак и т. п. (кроме
однояйцевых близнецов). Однако же у многих низших животных и рас¬
тений понятие особи или организма как «несекомого» далеко не всегда
однозначно. Например, если проводится исследование какого-то лесно¬
го массива, скажем, дубравы, там каждый дуб можно считать отдельной
особью, ибо он вырастает из семени, находящегося внутри желудя. С по¬
мощью разного рода наблюдений, можно обнаружить индивидуальные
§ 8. Что такое грибная особь
45
различия между дубами в лесу. В частности есть метод анализа ДНК,
названный методом «отпечатков пальцев», который позволяет отличить
отдельные особи с такой же точностью, как применяемый в криминали¬
стике метод идентификации каждого человек по рисунку отпечатка его
пальца. Но представим себе другой лесной массив. С одного дерева тополя
срезали 100 веток и укоренили их на определенной территории, так что
через несколько лет возникла тополевая роща. Можно ли считать каждое
дерево этой рощи отдельной особью? Ведь все они происходят из участков
тела одного дерева и генетически абсолютно идентичны. Таких ситуаций
в мире растений сколько угодно. Всем дачникам знакома садовая земляни¬
ка. Ее сорта размножают побегами-усами, кончики которых укореняются
и образуют новый кустик. Если усик, соединяющий материнский и до¬
черний кустики отрезать, получится два экземпляра, но на самом деле,
один из них — не индивидуальный организм, а продолжение второго.
У грибов с определением особей также много трудностей. Мухомо¬
ры, растущие на каком-то участке леса, это не отдельные особи, а плодо¬
вые тела, а особью является мицелий, протяженность которого в почве,
как было сказано в предыдущем разделе, может быть значительной. Еще
больше проблем ставят «плесневые» микроскопические грибы. Напри¬
мер, чтобы определить встречаемость какого-либо вида гриба в почве,
берут стерильно пробы почвы, разводят навеску почвы в определенном
объеме стерильной воды, тщательно перемешивают почвенную болтушку
и маленькие дозы ее равномерно распределяют по поверхности чашек
Петри с агаризованной питательной средой. Из разнообразных выросших
колоний микроорганизмов выделяют такие, которые по внешнему виду
и микроскопическим признакам соответствуют изучаемому виду. Но число
выросших колоний — это не число особей в грамме почвы, так как коло¬
ния может вырасти из одной одноклеточной споры, имеющей одно ядро,
но может образоваться и из обрывка мицелия, состоящего из десятков
клеток с тремя-четырьмя ядрами в каждой. Поэтому подсчет указывает
не на число особей, а на число «колониеобразующих единиц» (КОЕ).
Знаменитый английский биолог Р. Докинз дал определение организ¬
му, как единице, берущей свое начало в результате нового акта размно¬
жения, осуществляемого через «бутылочное горлышко» одноклеточной
стадии развития. «Бутылочным горлышком» популяционные генетики на¬
зывают ситуацию, при которой вследствие сужения числа особей одной
популяции или клеток одной особи в новые организмы и популяции попа¬
дает случайно только малая доля геномов или ядер из исходно огромного
их числа. Прохождение через стадию «бутылочного горлышка» приво¬
дит к разбиению генетически гетерогенной популяции или генетически
гетерогенного организма на множество различных, но генетически гомо¬
генных клонов. Определение Докинза не делает различий между клетками,
возникшими в результате полового или бесполого размножения. Исходя
46
Глава 2. Мицелиальный образ жизни
из этого определения можно дать, применительно к разным грибами, три
определения особи.
1. Одноклеточные дрожжи. Каждая дрожжевая клетка — отдельная
особь, однако передача ее генома следующему поколению (через поч¬
кование) происходит без промежуточной многоклеточной стадии, сле¬
довательно, здесь нет эффекта бутылочного горлышка, и все потомство
дрожжевой клетки генетически идентично, пред¬
ставляя собой единый клон. Такие клоны, по-ви-
димому, могут достигать гигантских размеров. Бы¬
ло бы ошибочно изучать клональную структуру пе¬
карских дрожжей Сахаромицес церевизия, так как это «одомашненные
грибы» (как бывают домашние растения и животные), крайне редко встре¬
чающиеся в «диком виде», и на их генетическую структуру влияет человек,
занимающийся селекцией. Но есть «дикорастущий» родственник пекар¬
ских дрожжей, Сахаромицес парадоксус, природные экземпляры которого
были исследованы в лаборатории одного из лучших специалистов в обла¬
сти генетики дрожжей Г. И. Наумова. Было проведено сравнение экзем¬
пляров, собранных в Европе и передней Азии (Испания, Нидерланды, Да¬
ния, Финляндия, Киев, Крым, Москва, Санкт-Петербург, Эстония, Литва,
Новгород, Воронеж, Татарстан, Азербайджан, Узбекистан) и на Дальнем
Востоке (Владивосток, Сихоте-Алинь, Япония и др.). Оказалось, что все
экземпляры внутри первой и второй групп генетически очень близки меж¬
ду собой, но между группами наблюдались значительные различия. Зна¬
чит, по-видимому, авторы исследования имели дело с двумя гигантскими
клонами, каждый из которых имеет протяженность в сотни и тысячи
километров. Размножающаяся сходным способом (правда, не почковани¬
ем, а делением) гигантская по численности и широте распространения
бактерия кишечная палочка, обитающая в кишечнике каждого человека,
по результатам обследования пациентов с разных континентов также об¬
разует всего несколько клонов.
2. Мицелиальные микромицеты. У них особью является мицелий,
изолированный от других мицелиев того же вида и образующий споры
бесполого размножения из одной или нескольких клеток. Здесь, несмотря
на отсутствие полового процесса, возможен эффект бутылочного горлыш¬
ка, так как мицелиальный индивид содержит множество ядер, и в неко¬
торых ядрах могут возникать мутации. К тому же, вследствие анастомозов
возможен обмен ядрами с другими индивидами и возникновение гетеро-
кариотичных клеток. Если в клетки разных спор перейдут генетически
различные ядра, возникнет гетерогенное потомство. Однако и для этих
грибов не только возможно, но и характерно клональное размножение,
при котором потомство генетически идентично друг другу и родительско¬
му индивиду. Определить границы индивидуумов для этих грибов мето¬
дически очень трудно, а зачастую невозможно, так как индивидуальные
§ 9. Есть ли у грибов иммунитет?
47
мицелии живут короткий период времени и быстро распадаются на споры.
Поэтому, как и для первой группы, у них обычно определяют число и раз¬
мер клонов, которые также могут быть гигантскими. Например, на всей
территории Сибири от Урала до Сахалина в посадках картофеля парази¬
тирует только один клон оомицета Фитофторы.
3. Базидиальные макромицеты (шляпочные, трутовые грибы). У этих
грибов ситуация с определением индивидуума наиболее близка к живот¬
ным. Особью является индивидуальный долгоживущий, часто многолет¬
ний мицелий, который образуется в результате скрещивания, и границы
которого можно установить молекулярными методами. Размножение осу¬
ществляется одноядерными (в большинстве случаев) спорами, возникаю¬
щими, как и у высших растений и животных, в результате редукционного
деления ядра материнской клетки и, вследствие этого, генетически отли¬
чающимися друг от друга. Поэтому в полной мере проявляется эффект
бутылочного горлышка, и каждый потомок генетически индивидуален.
§ 9. Есть ли у грибов иммунитет?
Иммунитетом называют способность организма распознавать чужие
клетки или макромолекулы и создавать условия для их нейтрализации.
Конечно, использовать для этих целей специфические антитела грибы
не умеют, такие фигуры высшего пилотажа появились только у позвоноч¬
ных животных, а вот простейшие иммунные реакции присущи и растени¬
ям, и даже грибам.
Выше было рассказано о том, что при встрече двух колоний мице¬
лиальных гриба, встречные гифы могут сливаться, после чего происходит
взаимная миграция ядер навстречу друг другу и возникают разноядерные
(гетерокариотические) клетки. Как микологи установили это? Путем сра¬
щивания колоний на питательных средах. В почве обитает много видов
грибов из рода Аспергиллюс. Один из видов этого рода генетики широко
используют как модельный объект. Его споры окрашены в темно-зеленый
цвет, поэтому колония на питательной среде в период спороношения так¬
же темно-зеленая. Из одного случайно выделенного индивида генетики
университета шотландского города Глазго получили много мутантов, спо¬
ры которых окрашены в ярко-зеленый, желтый цвет или даже бесцветные
(белые). Когда желтоспоровые и белоспоровые мутанты сращивали, воз¬
никали колонии с пестрыми цепочками спор — одна цепочка спор белая,
другая — желтая. Значит, клетки мицелия после анастомозов содержали
смесь ядер, а в одноядерные споры случайно (по правилу горлышка бу¬
тылки) переходили или ядра одного мутанта (дающие желтые споры), или
другого (споры белые). В дальнейшем в исследовательских лабораториях
разных стран было проделано множество исследований по генетике это¬
го аспергилла, причем все эти исследования (а выпущено более тысячи
48
Глава 2. Мицелиальный образ жизни
научных публикаций), были проведены на одном индивиде, выделенном
из почвы в университете Глазго. Когда же стали сращивать мутанты, по¬
лученные у разных природных индивидов, то оказалось, что разноядер¬
ные клетки не образуются, то есть генетически различные индивидуумы
обладают какими-то механизмами, запрещающими взаимную миграцию
ядер. Это явление получило название гетерокарионной или вегетатив¬
ной несовместимости (в отличие от половой несовместимости, например,
невозможности половых скрещиваний между разными видами). Вегета¬
тивная несовместимость присуща практически всем исследованным гри¬
бам и псевдогрибам. У большинства видов она проявляется следующим
образом: гифы двух колоний грибов после встречи сливаются, как им
и положено, но затем слившиеся клетки (а часто, и соседние с ними)
буреют и погибают. На границе двух колоний на питательной среде об¬
разуется темная линия из погибших клеток, получившая название барраж
(по-французски — заграждение). Генетические исследования показали,
что в грибных геномах имеются гены, влияющие на результаты анастомо¬
зов между двумя разными особями. Большинство этих генов встречаются
в двух аллельных состояниях (например, Аиа, ВиЬ,Си с), хотя об¬
наружены гены, имеющие три состояния. Причем нормальный анастомоз
и взаимная миграция ядер возможны лишь между индивидуумами, у ко¬
торых все гены вегетативной несовместимости находятся в одинаковом
аллельном состоянии. Если же какой-либо один ген у двух индивидуумов
находится в разных состояниях (например, у одного имеется ген С, а у дру¬
гого — с), то слившиеся клетки погибнут. А поскольку таких генов может
быть до десятка и больше, то вероятность того, что все они у двух случайно
встретившихся природных особей будут одинаковыми, очень мала.
1. В основе вегетативной несовместимости лежит именно узнавание
«чужого», ибо чем по большему числу генов несовместимости различа¬
ются контактирующие мицелии, тем острее протекает гибель слившихся
гиф. А степень различий по генам несовместимости отражает и общие
генетические различия между особями.
2. Микологи считают, что причиной широкого распространения веге¬
тативной несовместимости среди природных экземпляров грибов является
защита от так называемой «эгоистичной ДНК». Клетки всех организмов,
в том числе и грибов, содержат в ядре и в цитоплазме большое число генов,
которые не принимают никакого участия в синтезе необходимых клетке
белков и других процессах, обеспечивающих функционирование и адапта¬
ции организма. В последние годы стала общепринятой точка зрения на эти
Вегетативная
несовместимость —
простейший меха¬
низм иммунитета
Почему же вегетативную несовместимость мож¬
но считать простейшим механизмом иммунитета?
Тому есть несколько причин, из них укажем на три,
не требующие для своего объяснения привлечения
сложных для восприятия научных аргументов.
§ 9. Есть ли у грибов иммунитет?
49
гены, как на «эгоистичные», назначение которых — лишь собственная ре¬
пликация (размножение). Некоторые из них могут покрываться белковой
оболочкой, и в таком случае они называются вирусами. Клетки многих
грибов содержат множество вирусов, геном которых отличается от гено¬
мов клеточных организмов. У последних функции хранения информации
и ее реализации в синтезе белковых молекул разделены между двухцепо¬
чечной молекулой ДНК (двойной спиралью) и одноцепочечными моле¬
кулами РНК. Геном вирусов грибов представлен двухцепочечными моле¬
кулами РНК. Эти вирусы весьма вредоносны. Они меняют морфологию
зараженных особей, снижают способность к размножению и резко снижа¬
ют способность грибов-паразитов к заражению растений и размножению
в них. Эти вирусы передаются от зараженной особи к здоровой через ана¬
стомозы, поэтому вегетативная несовместимость — надежный механизм
защиты грибов от вирусной инфекции, то есть фактор иммунитета грибов
к вирусам. Ярчайшей иллюстрацией роли вегетативной несовместимости
в защите от вирусной инфекции является история рака каштанов в Европе
и Северной Америке. Возбудитель болезни — сумчатый гриб — был заве¬
зен в Европу и Америку из Восточной Азии и вызвал массовую гибель дере¬
вьев. Его споры, попадая на ствол дерева, прорастают и через ранки в коре
распространяются внутри дерева. Гриб вызывает поражение коры вплоть
до делящихся клеток камбия, подавляет функционирование проводящих
сосудов и гибель дерева (рис. 14 а). Более полусотни лет назад французские
ученые обнаружили экземпляры гриба-паразита, отличающиеся от обыч¬
ных по форме и окраске колоний и неспособные заражать каштаны. Их на¬
звали гиповирулентными (слабовирулентными). Было обнаружено, что ес¬
ли рядом с колонией гиповирулентного экземпляра выращивать обычный,
вирулентный, то после контакта мицелиев он также становится гиповиру-
лентным. Значит какой-то фактор, вызывающий гиповирулентность пере-
Рис. 14. Рак коры каштанов: а) пораженный ствол; б) несовмести¬
мая реакция взаимодействия штаммов возбудителя болезни гриба
эндотия (барраж) (фото Н. Н. Гринько)
50
Глава 2. Мицелиальный образ жизни
дается через анастомозы гиф и распространяется по мицелию вирулентно¬
го экземпляра. Установили, что этот фактор — двухцепочечная вирусная
РНК. После этого была разработана технология биологической защиты
каштанов путем заражения их гиповирулентными штаммами. Технология
оказалась эффективной. Но когда такую технологию стали использовать
в США, выяснилось, что она не дала ожидаемого эффекта. Причина не¬
успеха — широкое распространение вегетативной несовместимости в аме¬
риканских популяциях гриба паразита. При слиянии гиф искусственно
вносимых гиповирулентных особей с природными вирулентными проис¬
ходил барраж и вирус не переходил в клетки вирулентной особи (рис. 14 б).
3. Вегетативная несовместимость сопровождается быстрым побуре-
нием и гибелью слившихся и примыкающих к ним клеток. Исследование
процессов гибели показало, что это — одна из форм апоптоза — програм¬
мируемой смерти клеток, а апоптоз — один из важнейших механизмов
иммунного ответа млекопитающих.
Но функции вегетативной несовместимости грибов не ограничивают¬
ся только защитой от эгоистичных генов. Биологов давно волнует, каким
образом происходит симпатрическое видообразование, то есть возникно¬
вение нового вида внутри одной популяции. Ведь если у отдельных особей
возникнет новый ген или набор генов, то при скрещиваниях с носителями
старых генов, которые составляют подавляющее большинство популяций,
новые гены будут разбавлены старыми аллелями и после серии скре¬
щиваний вытеснены из генома потомков первоначального их носителя.
Поэтому считается, что видообразование происходит при географическом
отделении части особей от основной популяции. Вследствие эффекта бу¬
тылочного горлышка основатели новой колонии могут нести измененный
набор генов, который передадут своим потомкам. Жизнь в изменившихся
условиях будет способствовать отбору новых мутаций, которые в прежних
условиях жизни были бесполезными или даже вредными и не фиксирова¬
лись в потомстве. Однако симпатрическое видообразование — это факт,
подтвержденный многочисленными наблюдениями. Зоологи А. В. Марков
и А. М. Куликов высказали гипотезу, что скрещиванию особей зарожда¬
ющего внутри популяции вида с доминирующими в популяции особями
препятствуют мутации в клетках иммунной системы, приводящие к тка¬
невой несовместимости и гибели гибридов (подобно тому, как погибают
донорские органы, перенесенные хирургическими методами в другой ор¬
ганизм). Отсюда следует, что иммунный механизм узнавания чужого иг¬
рает решающую роль в видообразовании без географической изоляции.
Вегетативная несовместимость — важнейший механизм Бидлабраэлвания
во многих группах грибов. Огромное число грибов (десятки тысяч видов)
не имеют полового процесса и размножаются исключительно бесполы¬
ми спорами (подробнее об этом будет рассказано ниже). Единственный
способ обмена генами этих грибов — анастомозы гиф. Вегетативная несов¬
§ 9. Есть ли у грибов иммунитет?
51
местимость, как простейший механизм иммунного ответа, препятствует
миграции ядер между генетически «чужими» организмами и создает гене¬
тически изолированные островки внутри большой исходной популяции.
Например, один из видов широко распространенного рода Фузариум мо¬
жет жить в почве, питаясь растительными остатками, но способен также
заражать корни молодых растений, поселяться в их сосудах и вызывать
увядание. В одном из исследований были выделены в культуру сотни
экземпляров этого гриба, как из почвы, так и из пораженных растений
хлопчатника. Грибы, выделенные из почвы, быстрее росли на питательно
среде, а грибы, выделенные из растений, успешнее заражали растения
в опытах с искусственным заражением. Но что самое удивительное: все
экземпляры из каждой группы были совместимы между собой (нормально
анастомозировали), но несовместимы с экземплярами из противополож¬
ной группы (формировали барраж). Следовательно, внутри исходного вида
происходит эволюция в двух направлениях — в направлении усиления па¬
разитических свойств и в направлении более эффективного разложения
почвенной органики. А для того, чтобы гены, обеспечивающие эти про¬
тивоположные свойства, не перемешивались, произошел отбор мутаций
генов вегетативной несовместимости, обеспечивающих запрет на мигра¬
цию ядер между особями из двух субпопуляций.
Глава 3
Как грибы
размножаются
В начале перестройки в СССР был показан по те¬
левидению телемост между советской и американской
телеаудиториями, который проводили Фил Донахью и
Владимир Познер. Одна из наших дам тогда заявила, что
в СССР секса нет, чем вызвала улыбки у американцев.
Если бы был проведен телемост между грибами, кото¬
рые микологи называют дейтеромицетами, и грибами,
скажем, базидиомицетами, то первые заявили бы, что
«у нас секса нет», и были бы правы, а вторые бы сказали:
«У нас секс есть», — и тоже были бы правы. Но если бы
они, как и дейтеромицеты, стали утверждать, что у них
секса нет, то они тоже были бы правы. Словом, как
в анекдоте про раввина, к которому пришли с тяжбой
две женщины. Выслушав доводы первой, он сказал: «Ты
права», — и она ушла довольная. Когда вторая женщина
привела противоположные доводы, он ей тоже сказал:
«Ты права». После ее ухода присутствующий при беседе
прислужник сказал: «Как же так, Ребе, ведь они один
и тот же эпизод описывали совершенно по-разному?».
На что раввин ответил: «И ты прав». Поскольку в основе
наших рассуждений лежит не мнение отдельного равви¬
на, пусть даже очень умного, а мнение многих ученых
микологов, основанное на многочисленных экспери¬
ментах, это противоречие надо рассмотреть подробнее.
§1.0 том, как размножаемся мы
и наши меньшие братья
Прежде чем вернуться к нашим грибам, надо ска¬
зать несколько слов о способах размножения у разных
групп организмов. У млекопитающих, включая человека,
во всех клетках, составляющих тело, имеется двойной
§1.0 том, как размножаемся мы и наши меньшие братья 53
набор хромосом, то есть каждая хромосома пред¬
ставлена двумя копиями, полученными от папы и от
мамы. Клетки, содержащие двойной набор хромо¬
сом, называются диплоидными. Почти все пары хромосом (их называют
аутосомами) одинаковы по внешнему виду и содержат одни и те же гены,
которые расположены в одних и тех же участках хромосомы (правда, они
могут по-разному влиять на проявление того или иного признака, напри¬
мер ген в одной хромосоме делает человека черноглазым, а такой же ген
во второй копии хромосомы делает его носителя голубоглазым). Но одна
хромосома, называемая половой, у двух родителей различается по разме¬
рам, числу и расположению генов. У человека имеется 22 пары одинаковых
аутосом и одна пара половых хромосом, называемых X- и Y-хромосомами.
На них, кроме прочих, находятся гены, которые регулируют преимуще¬
ственный синтез мужских или женских половых гормонов, а те, в свою
очередь, управляют образованием первичных (строение половой системы)
и вторичных (развитие молочных желез, рост волос на разных участках те¬
ла и прочее) половых признаков. Если в ядрах находятся две Х-хромосомы,
то формируется женский фенотип, если же половые хромосомы различны
(X и Y), то формируется мужской фенотип. Во всех соматических клетках
при их делении число хромосом в ядрах удваивается, так что в материнской
и дочерней клетках ядра сохраняют двойной набор хромосом. Такой спо¬
соб деления ядер называют митозом. И только в клетках половой системы
перед делением ядер хромосомы не удваиваются, а в виде одиночных ко¬
пий расходятся по дочерним ядрам. В результате такого редукционного
деления (называемого мейозом) образуются клетки с единичным набо¬
ром хромосом (гаплоидными ядрами). Эти клетки называются гаметами.
В женском организме гаметами становятся крупные неподвижные клет¬
ки — яйцеклетки, а в мужском — мелкие сперматозоиды, подвижные
вследствие наличия жгутика. При половом процессе сперматозоиды под¬
плывают к яйцеклетке, один из них сливается с ней, после чего сливаются
ядра, восстанавливается двойной набор хромосом, и в оплодотворенной
яйцеклетке (зиготе) начинаются клеточные деления и формирование пло¬
да. Какой пол будет у ребенка, определяет случай. Ведь в ядрах и мужских,
и женских гамет содержится только по одному набору хромосом. Значит,
будет одинаковое количество аутосом и по одной половой хромосоме —
или X, или Y. Поскольку диплоидные клетки женского организма имели
ХХ-хромосомы, все образующиеся яйцеклетки также содержат ее, а 50%
мужских гамет будут иметь X и 50 % — Y-хромосому. Если произошло сли¬
яние яйцеклетки со сперматозоидом, имеющим Х-хромосому, в зиготе бу¬
дет две Х-хромосомы, и родится девочка, а если с яйцеклеткой соединился
сперматозоид, имеющий Y-хромосому, то родится мальчик. Так что му¬
жья, ругающие своих жен за то, что они рожают одних девчонок, должны
пенять на себя, ведь пол определяет не женская гамета, а мужская. Чтобы
закончить с этим вопросом, еще одно замечание. Разнообразие признаков
54
Глава 3. Как грибы размножаются
в потомстве одной супружеской пары вызвано тем, что в процессе мейоза
в одну гамету могут попасть случайно часть аутосом, полученных ребен¬
ком от мамы, а часть аутосом, полученных от папы. Поскольку каждая
аутосома содержит тысячи генов, возможны различные их комбинации
у потомства. Межхромосомные рекомбинации получили название менде¬
лизма, по имени открывшего это явление (правда, у растений, а не у жи¬
вотных) августинского монаха Грегора Менделя. Кроме этого, в процессе
мейоза пары одинаковых хромосом плотно соприкасаются друг с другом,
и в это время происходят обмены участками между разными хромосомами
(кроссинговер). В результате таких обменов возникают внутрихромосом-
ные рекомбинации, названные морганизмом по имени открывшего это
явление у плодовой мушки дрозофилы ученого Томаса Моргана.
У растений формы полового процесса более раз-
Как размножаются ^ ~
нообразны. Есть виды растении, у которых мужские
рЭ^ТсНИл л . м v
(тычиночные) и женские (пестичные) цветки нахо¬
дятся на разных экземплярах, то есть имеются мужские и женские расте¬
ния. Пыльца, образующаяся на мужских экземплярах, попадает на пестики
женских, там гаплоидные клетки после ряда превращений образуют не¬
подвижные (в отличие от животных) спермии, которые сливаются с яйце¬
клетками. У некоторых видов тычиночные и пестичные цветки находятся
на одном растении. Все, наверное, помнят, как выглядят растения ку¬
курузы: на верхушке находится соцветие тычиночных цветков, а пазухах
листьев — початки пестичных. У большинства видов цветки обоеполые
(содержат и тычинки, и пестики). Перекрестное оплодотворение обяза¬
тельно только для двудомных видов, а у однодомных и обоеполых воз¬
можно как перекрестное, так и самооплодотворение (хотя имеется много
механизмов, запрещающих последнее). Кроме того многие растения спо¬
собны размножаться вегетативными органами (картофель — клубнями,
тюльпаны — луковицами, пырей и осот — корневищами).
У низших эукариотных организмов, к которым
относятся водоросли, простейшие животные и гри¬
бы, разнообразие форм и способов размножения
увеличивается во много раз.
Во-первых, разнообразна форма гамет. Уже го¬
ворилось, что у высших растений и животных муж¬
ская гамета мелкая, покидающая орган, в котором
она сформировалась, — гаметангий, и называется сперматозоидом или
(если не имеет жгутиков) спермием; а женская —■ крупная, богатая пи¬
тательными веществами, не покидающая гаметангия, называется яйце¬
клеткой. Половой процесс такого типа называют оогамией. У низших эу¬
кариот гаметы, формирующиеся на разных особях, в простейшем случае
морфологически не различаются, а половой процесс называется изогами¬
ей. Обычно гаметы снабжены жгутиками и покидают клетку, в которой
были сформированы. У одноклеточных организмов — это вегетативная
Как размножаются
грибы и водоросли
Разнообразие
гамет
§1.0 том, как размножаемся мы и наши меньшие братья 55
клетка, а у многоклеточных — одна из клеток, превратившаяся в гаметан-
гий. Слияние изогамет происходит вне родительских организмов. А если
нет морфологических различий между гаметами, нельзя определить, какая
из них мужская, а какая — женская, то нет и полов. Возникает интересная
ситуация — половой процесс есть, а полов нет. Такие организмы могут
сказать: «У нас секса нет», — и будут правы. А могут сказать: «У нас секс
есть», — и тоже будут правы.
Половые различия между гаметами начинаются при возникновении
гетерогамии, при которой обе гаметы подвижны, также покидают гаме-
тангий и сливаются вне родительского организма, но имеют морфоло¬
гические отличия. Одна из них — крупная клетка, имеющая богатый
набор питательных веществ, малоподвижная, вялая, вскоре после выхода
из гаметангия останавливается. Это — женская гамета. Вторая — мел¬
кая, активно плавающая с помощью жгутиков — мужская. Она находит
женскую гамету и сливается с ней. Дальнейшая эволюция привела к воз¬
никновению знакомой нам оогамии, при которой женская гамета вообще
неподвижна и называется яйцеклеткой.
Наконец, у многих низших эукариот гаметы вообще не образуются,
а сливаются прямо гаметангии (гаметангиогамия), или просто вегетатив¬
ные клетки (конъюгация, соматогамия), или даже просто ядра в клетке.
Вот в каких причудливых формах может проявляться половой процесс.
Во-вторых, у низших эукариот более разнооб-
разны, чем у высших, жизненные циклы. Как уже ЖИЗНеншх°циклов
было сказано, у высших растений и животных веге¬
тативные клетки содержат в ядрах двойной набор хромосом (диплоидны),
а при формировании гамет проходит мейоз и ядра возникающих гамет
имеют один набор хромосом, то есть они гаплоидны. При сливании га¬
мет (в зиготе) восстанавливается диплоидное состояние. Такой жизненный
цикл называют диплоидным. У видов низших организмов наряду с дипло¬
идным циклом встречается и противоположный ему — гаплоидный, при
котором вегетативные клетки тела гаплоидные, а при слиянии гамет, гаме-
тангиев или вегетативных клеток образуется зигота с диплоидным ядром.
Эта зигота обычно покрывается толстой оболочкой и может выдерживать
неблагоприятные условия жизни (похолодание, высушивание и прочее).
Перед ее прорастанием происходит мейоз, и вегетативные клетки восста¬
навливают гаплоидный ядерный статус. У многих видов жизненный цикл
еще более экзотичен. В отдельных клетках диплоидного организма (спо¬
рангиях) происходит мейоз, но образующиеся после этого гаплоидные
споры не сливаются друг с другом, а прорастают в новый организм, все
клетки которого содержат гаплоидные ядра. В отдельных клетках этого
гаплоидного организма (гаметангиях) вследствие дробления без редукции
числа хромосом (митозов) образуются гаметы, которые сливаются. Дипло¬
идная зигота дает рост новой вегетативной особи. Такой жизненный цикл
называют гаплодиплоидным, ибо при его протекании происходит смена
56
Глава 3. Как грибы размножаются
двух типов вегетативного организма —* гаплоидного и диплоидного. И хо¬
рошо еще, если они морфологически не отличаются, так как очень часто
они настолько не похожи друг на друга, что ученые описывали отдельные
стадии жизненного цикла одного организма, как разные виды и даже роды.
В-третьих, у низших организмов наряду с по-
Размножение без
ловым размножением широко представлено беспо-
полового процесса к к к
лое, при котором внутри одноклеточного организ¬
ма или специальных клеток многоклеточного (спорангиев) вследствие се¬
рии митозов образуется большое число спор, имеющих такой же ядер-
ный статус, как и материнский организм. Каждая спора после выхода
из спорангия прорастает в новый организм, неотличимый от родитель¬
ского ни по внешнему виду, ни по числу хромосом в ядре.
Как видно, низшие эукариотные организмы — это полигон разно¬
образнейших способов размножения, на котором Господь Бог по имени
«Естественный Отбор» отбирал и стандартизировал элементы, наиболее
подходящие для размножения высших организмов, включая человека. Ка¬
ковы же были движущие силы отбора, почему было отброшено бесполое
размножение и выбран только один тип полового — оогамия? Давай¬
те порассуждаем. При наличии бесполого размножения половой процесс
не может с ним конкурировать, так как уступает ему и по числу образую¬
щегося потомства, и по простоте его создания. Кроме прочих, достаточно
лишь одного аргумента. При бесполом размножении каждая особь остав¬
ляет потомство (так, летом в колониях тлей все особи самки, рожающие
самок), а при половом — 50% особей в популяции (самцов) непродук¬
тивны. Тем не менее, бесполое размножение по мере эволюции все более
и более уступало место половому и хотя встречается у некоторых растений,
насекомых и даже рептилий, однако у высших позвоночных животных
(млекопитающих) описано только один раз — в Новом Завете. Но чудеса
здесь не обсуждаются. Значит, кроме создания потомства (размножения)
половой процесс несет и какие-то иные функции, роль которых по мере
биологической эволюции возрастала.
Итак, рассмотрим эволюцию способов раз-
Эволюция способов Л Г
4 множения. У древнеиших организмов — прокари-
размножения „ гг
отных бактерий полового процесса нет, они раз¬
множаются исключительно вегетативно. И хотя написанная полвека на¬
зад книга двух выдающихся французских микробиологов, один из которых
был Нобелевским лауреатом, называлась «Секс и генетика бактерий», так
на то они и французы. Не было полового размножения и у первичных
одноклеточных эукариот. Затем появляется половой процесс (не пол, а по¬
ловой процесс) в простейшей своей форме — слияние двух вегетативных
особей. Поскольку организмы тогда были одноклеточные, возник процесс
их слияния. Это было, конечно, не размножение, так как в результате
слияния происходит не увеличение, а наоборот, уменьшение числа особей
(было две, стала одна). Значит, первичная функция полового процесса
§1.0 том, как размножаемся мы и наши меньшие братья 57
никак не была связана с размножением. Можно предположить, что од¬
на из первичных функций — защитная, поскольку зигота покрывается
толстой оболочкой и может выдерживать наступление неблагоприятных
для жизни внешних условий. Эта функция сохраняется длительное время,
вплоть до возникновения плодов высших растений, покрытых толстыми
защитными оболочками, таких как орех, костянка и прочие. Однако важ¬
нее другое.
Одновременно со слиянием клеток возникли два важнейших процесса.
Во-первых, слияние их ядер, возникновение в них двойного набора хромо¬
сом — диплоидности; во-вторых, возникновение нового механизма рас¬
хождения хромосом — мейоза, при котором не только восстанавливается
исходное гаплоидное состояние ядер, но и могут происходить обмены
хромосомами и их участками, то есть генетическая рекомбинация. По-ви¬
димому, эти изобретения природы сыграли выдающуюся роль в эволюции
организмов на земле.
Половое размножение (не процесс, а именно размножение) начинает¬
ся с возникновением гамет, когда внутри материнской клетки образуется
большое число подвижных клеточек, сливающихся друг с другом. В про¬
стейшем случае, при изогамии, все гаметы морфологически одинаковы.
А поскольку гаметы, образовавшиеся в одном гаплоидном организме, ге¬
нетически одинаковы (клоны одной клетки), то при слиянии их друг
с другом теряется смысл полового процесса, как источника генетической
рекомбинации. Поэтому возник генетический механизм выбора партнеров
для слияния. На одной из хромосом возник ген (или группа сцепленных
генов), получивший название локуса спаривания. Этот локус у разных
особей может быть представлен одним из двух отличающихся друг от дру¬
га (аллельных) состояний, которые обычно называют «плюс» и «минус».
В гаплоидном (имеющим один набор хромосом) организме может при¬
сутствовать только одно состояние (или «плюс», или «минус»), поэтому
все гаметы одного организма имеют одинаковый знак. А гаметы приоб¬
рели свойство не обращать внимание на тех, которые имеют одинаковый
с ними знак, но стремиться к спариванию с гаметами противоположно¬
го знака. Это приводит к тому, что способность к взаимному спариванию
приобрели только гаметы, возникшие из разных особей, и, следовательно,
повысилась вероятность генетических различий у родителей и потомков.
Итак, при изогамии есть половой процесс, есть половое размножение
и есть способ запрета на спаривание со своими братьями (сибсами). Нет
только половых (морфологических и поведенческих) различий между га¬
метами. Генетический запрет на родственные скрещивания при отсутствии
морфологических различий между гаметами получил название гетеротал¬
лизма. Пол возник с появлением гетерогамии, то есть морфологических
отличий между гаметами, которые образуют «плюс» и «минус» особи. Од¬
ни гаметы (мужские) — мелкие, активно плавающие, другие (женские) —
крупные, набитые питательными веществами. Они быстро теряют жгутики
58
Глава 3. Как грибы размножаются
и прекращают движение. Привлекательными женские гаметы для мужских
делает способность первых выделять в окружающую среду некие хими¬
ческие вещества (половые феромоны). Как видно, с древнейших времен,
начиная с простейших эукариотных организмов, женщины привлекали
мужчин, а не наоборот: женская гамета слишком ленива и малоподвижна,
чтобы гоняться за мужскими. Наконец, женские гаметы решили: «Зачем
нам вообще двигаться и подвергать себя опасности быть кем-то съеден¬
ными, если они сами прибегут к нам». И возникла оогамия, при которой
женские гаметы лишились жгутиков и в большинстве случаев вообще пе¬
рестали покидать гаметангий. Возникает вопрос, зачем это нужно? Чем
морфологическое разделение полов лучше изогамии? Ведь гетероталлизм
не менее эффективно запрещает близкородственные скрещивания, чем
половые различия. Чтобы ответить на этот вопрос, сделаем небольшое
отклонение от темы. Тем более что весь разговор в данном разделе —
отклонение от темы под названием «Грибы».
В популяционной генетике есть очень важный показатель, называе¬
мый приспособленностью, который показывает, какой вклад в виде ко¬
личества особей (генотипов) организм внесет в следующее поколение.
Приспособленность определяют произведением продуктивности (числа
произведенных потомков) на выживаемость (способность потомков дой¬
ти до половозрелого состояния и оставить потомство). Беда в том, что
одновременно увеличивать оба показателя невозможно, ибо они взаимно
несовместимы. В самом деле, чем больше потомков будет произведено, тем
меньше каждый из них получит запаса питательных веществ, тем мень¬
ше шансов на выживание он будет иметь. Конечно, возможны разумные
компромиссы, но чаще организмы эволюционируют по пути выбора раз¬
ных жизненных стратегий — стратегии размножения (получившей назва¬
ние r-стратегия) и стратегии выживания (/Г-стратегии). Такие названия
стратегий приняты в соответствии с разными показателями в математи¬
ческих уравнениях роста численности популяций. Возьмем, к примеру,
рыб. Треска мечет миллион икринок и забывает о них. Львиная доля
икринок и мальков погибает, но, согласно закону больших чисел, оста¬
ется достаточно потомков, чтобы поддерживать численность популяции
на необходимом уровне. А некоторые виды сомиков производят настолько
мало потомков, что все они помещаются во рту родителя, который содер¬
жит там мальков, пока они не подрастут. Треска — типичный г-стратег,
а сомик — К-стратег. Так вот, разделение полов — это гениальное изоб¬
ретение, при котором разные гаметы приобретают разные жизненные
стратегии. Мужские гаметы — типичные r-стратеги. Они имеют геном
(ядро) который будет передан потомству, аппарат поиска женской гаметы
(рецепторы половых феромонов) и аппарат движения (жгутики), но очень
малый запас питательных веществ. Поэтому они производятся в массе,
что гарантирует высокую вероятность достижения отдельными сперма¬
тозоидами женской гаметы и ее оплодотворения. Последняя, наоборот,
§ 2. Вернемся к нашим грибам
59
неподвижна, но набита питательными веществами, поэтому и число гамет
небольшое (расход на производство каждой гаметы увеличен). Она не по¬
кидает гаметангия, поэтому меньше рискует, зато после оплодотворения
развивающийся потомок будет снабжен необходимым питанием и спря¬
тан внутри гаметангия.
§2. Вернемся к нашим грибам
2.1. Что такое бесполое размножение грибов
Мы начали с того, что грибы дейтеромицеты не имеют секса, поэто¬
му микологи, наверное, из уважения к радостям половой жизни, назвали
их несовершенными грибами. На самом деле они очень даже совершен¬
ные, представляют одну из самых многочисленных и процветающих групп
грибов. Дело в том, что у самых древних грибов уже существовали оба ти¬
па размножения — бесполое и половое, и дальнейшая эволюция привела
к тому, что в одних группах теряло значение половое размножение, а в дру¬
гих — бесполое.
Бесполое размножение грибов претерпело длитель¬
ную эволюцию. У самых древних грибов и псевдогрибов, ^
живущих в водной среде, все тело или его отдельные размножения
участки превращаются в зооспорангии, ядра в них ми-
тотически делятся, и образуется множество зооспор, выходящих наружу
после разрыва стенки зооспорангия. После выхода на сушу такой спо¬
соб размножения становится опасным — ведь вышедшие из зооспорангия
зооспоры могут и не попасть в воду, без воды, на воздухе, жгутики беспо¬
лезны, а голенькие, не имеющие клеточной стенки, зооспоры моменталь¬
но погибнут. В разных группах адаптация к наземному образу жизни шла
разными путями, хотя и привела к одному и тому же результату. У назем¬
ных псевдогрибов оомицетов зооспорангий принял вид утолщенного, по¬
хожего на микроскопический лимон, окончания гифы. Он целиком отва¬
ливается от зооспорангиеносца и распространяется с помощью дождевых
брызг или ветра. Только попав в каплю воды, зооспорангий освобождает
зооспоры, которые вскоре покрываются оболочками и прорастают. Затем
некоторые оомицеты начали терять способность формировать зооспоры;
весь лимоновидный зооспорангий превратился в одну спору. Это нагляд¬
ный пример замены жизненных стратегий: г-стратегии на -йГ-стратегию.
Вместо десятка зооспор, выходящих из зооспорангия, всего одна спора —
сам зооспорангий, однако вероятность выживания спор на суше, где ве¬
лик риск отсутствия воды, значительно возросла. Как говорят, за все надо
платить. У одних оомицетов (из рода Фитофтора) переход от непрямого
прорастания зооспорангия зооспорами к прямому прорастанию ростком
регулируется температурой: при низкой температуре (до 15 °С) из зооспо¬
рангия выходят зооспоры, а при высоких (более 20 °С) зооспорангий цели¬
ком прорастает ростковой трубкой (гифой). Причина понятна: фитофтора
60
Глава 3. Как грибы размножаются
заражает наземные растения, на поверхности которых образуются зоос¬
порангии. Отрываясь, зооспорангии попадают не в реку и даже не в лужу
(а если и попадут, то в луже они, будучи паразитами растений, погибнут
от бескормицы), а в лучшем случае в каплю воды на листе. Там надо, чтобы
зооспоры вышли из зооспорангия, поплавали, потеряли жгутики, покры¬
лись оболочками и проросли. При низкой температуре (например, ночью)
капли долго не высыхают и времени для заражения листа проростками
зооспор достаточно, а при высокой можно и не успеть, поэтому лучше
не рисковать и прорасти сразу ростком. Спор будет меньше, но вероят¬
ность создания нового потомства — выше. А у других оомицетов (из рода
Пероноспора) произошла полная утрата механизма образования зооспор,
и зооспорангий при любой погоде прорастает целиком, как одна спора,
то есть вместо эндогенных (образующихся внутри материнской клетки)
подвижных спор возникли экзогенные неподвижные, которые микологи
называют конидиями.
У настоящих грибов эволюция к наземному образу жизни шла дру¬
гими путями. Все началось не с потери эндогенных спор, а с потери
жгутиков. У плесневых мукоровых грибов на кончиках спорангиеносцев
образуются крупные шаровидные спорангии с множеством неподвижных
спор. После расползания стенки спорангия в капле воды споры освобо¬
ждаются и прорастают. Однако многие мукоровые грибы эволюциони¬
ровали в сторону увеличения числа спорангиев на спорангиеносце с од¬
новременным уменьшением числа спор в них. Конечный итог движения
в этом направлении — односпоровые спорангии, фактически представ¬
ляющие собой конидии с двумя оболочками (собственной и оболочкой
спорангия). А подавляющее число видов наземных грибов (сумчатые и ба-
зидиальные грибы) образуют настоящие экзогенные споры — конидии.
Как же регулируется спорообразование у этих гри-
Регуляция - ,
бесполого ’ какие стимулы заставляют гифы вегетативного ми-
размножения целия превращаться в конидиеносцы с конидиями? На
эти вопросы нам ответили грибы из весьма распростра¬
ненного рода Аспергиллус, живущие в различных почвах. Пока мицелий
находится в почве и питается там органическими веществами, в частности,
растительными остатками, он не окрашен (бесцветный) и никаких спор
не образует, так как зачем тратить энергию на образование спор, если нет
никаких путей их распространения. Стимулами (индукторами) процесса
спорообразования являются два фактора: свет и движение воздуха (ве¬
тер). А где можно встретиться с этими факторами? Только на воздухе, над
почвой. Поэтому, если случайно мицелий аспергилла высунется из почвы
на ее поверхность, в нем станут происходить морфогенетические и био¬
химические изменения: вертикально вверх начинает расти толстая гифа,
лишенная перегородок — конидиеносец (для разбрасывания спор надо
повыше подняться над поверхностью почвы); в конидиеносце откладыва¬
ется темный пигмент (надо защитить его от солнечной радиации — ведь
§ 2. Вернемся к нашим грибам
61
это воздух, а не почва); кончик конидиеносца расширяется и превращается
в пузырек, вся поверхность которого вследствие почкования покрывает¬
ся выростами (конидиогенными клетками); в каждую клетку переходит
из конидиеносца по одному ядру;
в клетке происходит серия ядерных
делений и образуется длинная, рас¬
падающаяся цепочка одноядерных
спор; каждая цепочка представля¬
ет собой клон клеток, содержащих
одинаковые ядра, подобный кло¬
нам стволовых клеток в иммунных
тканях животных (рис. 15). Такая
структура позволяет на одном ко-
нидиеносце сформировать бесчис¬
ленное количество спор и эффек¬
тивно, без особых потерь, распро¬
странить их в окружающей среде.
А теперь вернемся к несовер¬
шенным грибам. Повторим, что
дает половой процесс: 1) размножение, 2) образование оболочек, защища¬
ющих зиготу от неблагоприятных условий среды обитания, и 3) генетиче¬
скую рекомбинацию потомства. Как же сохранить эти полезные свойства
полового процесса, но без него?
При бесполом размножении, как видно из рассказа об аспергиллах,
образуется у большинства видов грибов гораздо больше потомков, чем при
половом. И не только у аспергиллов. Наверное, все сталкивались на даче
с плодами садовой земляники, пораженными серой гнилью. Толстый слой
серого налета, покрывающий плод, сплошь состоит из миллионов спор,
разлетающихся при малейшем дуновении ветра. У многих подобных гри¬
бов практически весь мицелий превращается в спороносные клетки. За¬
щитные структуры грибы научились создавать и без предварительного по¬
лового процесса, вспомним хламидоспоры и склероции, описанные ранее.
А что касается создания генетической рекомбинации, то для грибов до¬
статочно одного мутационного процесса. Если предположить, что средняя
частота мутаций одного гена составляет 10 ~9 (происходит в одном из мил¬
лиарда ядер), то в популяции, содержащей 10 миллиардов спор (а таких
популяций у грибов сколько угодно), по каждому гену будет около 10 мута¬
ций. Вот вам и материал для отбора. К тому же грибы научились тому, что
не умеет больше почти никто, — генетическим рекомбинациям без мейо-
за. Как было сказано, в клетках многих грибов содержится несколько ядер,
причем могут совместно существовать ядра, полученные от разных особей
вследствие анастомозов гиф. И вот иногда происходит ошибка, и сосед¬
ние ядра в клетке сливаются с образованием диплоидного ядра. Попадая
в одноядерную спору это ядро, вследствие серии митотических делений,
Рис. 15. Аппарат бесполого (конидиаль-
ного) спороношения гриба аспергиллюс:
а) схема; б) электронномикроскопическая
фотография (Horiushi Н. et al., 1999)
62
Глава 3. Как грибы размножаются
образует диплоидный клон. А в этом клоне возможны различные гене¬
тические процессы, приводящие к генетической рекомбинации. Данное
явление получило название парасексуального (псевдополового) процесса.
Потеря полового процесса проходит через ряд
Как гри ы теряют промежуточных этапов. Например, многие грибы-
половои процесс ^ J „ к к, у
паразиты растении в течение весенне-летнего сезона
размножаются исключительно бесполым способом, а осенью происходит
половой процесс и образуется зигота, окруженная защитными покрова¬
ми, которые позволяют ей выдержать неблагоприятные условия зимы.
Таким образом, в размножении и паразитизме половые структуры вообще
не участвуют, остаются только сохранение и генетическая рекомбинация.
Но вот человек начал возделывать сельскохозяйственные культуры и изме¬
нил образ жизни не только самих растений, но и их паразитов. Приведем
два примера.
На родине картофельной фитофторы, в Мексике, паразит зимует в
почве и в клубнях диких видов картофеля. Мелкие клубни дикарей в тече¬
ние теплой мексиканской зимы в массе подвергаются микробному разло¬
жению, поэтому более надежный источник сохранения до весны — почва,
в которой могут сохраняться только половые структуры — ооспоры. По¬
этому в дикой природе половой процесс является обязательным. В полях,
на которых люди выращивают культурный картофель, споры паразита,
сформированные на листьях, заражают при уборке клубни и сохраняются
в них до посадки. Поскольку человек создает оптимальные условия, не¬
обходимые для сохранности клубней, паразит также получил комфортные
условия зимовки и обходится без необходимости тратить «электроэнер¬
гию» на прохождение полового процесса и формирование покоящихся
структур.
У грибов, вызывающих заболевание злаков — ржавчину (из-за об¬
разования коричневых пятен на листьях и стеблях), летом образуются
тонкостенный споры (урединиоспоры), которые разлетаются и заража¬
ют новые растения. Осенью, когда листья и стебли начинают высыхать,
на соломе формируются толстостенные покоящиеся споры (телиоспоры),
которые зимуют. Весной два ядра телиоспоры сливаются, мейотически
делятся, и из телиоспоры вырастает промицелий, состоящий из четырех
клеток (базидия), на которых образуются четыре гаплоидные споры. Эти
споры неспособны заразить листья ржи или пшеницы, они должны зара¬
зить листья так называемого промежуточного хозяина, разного для разных
видов ржавчины, на котором пройдет половой процесс. И только образую¬
щиеся в результате полового процесса споры (эциоспоры) заражают злаки.
Эта стратегия выработалась в дикой природе, где злаковые травы росли
в окружении других растений, а фитоценозы (сообщества растений) были
очень сложными, многочленными. Но человек начал возделывать пшени¬
цу, ячмень или овес на огромных полях, стал уничтожать сорные и другие
сопутствующие растения, так что вероятность попадания гаплоидных ба-
§ 2. Вернемся к нашим грибам
63
зидиоспор на листья промежуточного хозяина стала низкой. К тому же
крестьяне придумали во многих зонах, где зимы не слишком суровы, вы¬
севать семена пшеницы и ржи не весной, а в конце лета, чтобы всходы
сохранялись под снегом и на следующий год давали урожай более ранний
и более высокий (озимые культуры). Этим прекрасно воспользовались па¬
разиты: их летние споры (урединиоспоры) перелетали со стареющих рас¬
тений посева прошлого года на новые всходы и зимовали в их листьях под
снегом. Образование толстостенных телиоспор, формирование промице¬
лия, обязательное попадание гаплоидных базидиоспор на листья промежу¬
точного хозяина и половой процесс на его листьях оказались не нужными.
Жизненный цикл чрезвычайно упростился, зато численность урединио-
спор значительно выросла вследствие экономии «электроэнергии».
Но хватит говорить о бесполом размножении, пора перейти к чудесам
полового размножения у грибов.
2.2. Узнавание партнеров
Впервые исследования в этом направлении были проведены над весь¬
ма примитивным древним грибом под названием Аллемицес. Он живет
в водной среде и развивается на трупах водных животных, образуя на них
нежный белый пушок. Разные индивиды гриба могут формировать в ми¬
целии или гаплоидные, или диплоидные ядра, то есть этот гриб имеет
гаплодиплоидный жизненный цикл, описанный выше. На гаплоидном
мицелии образуются подвижные гаплоидные гаметы, отличающиеся друг
от друга по форме и поведению. Мужские гаметы — мелкие, чрезвычайно
подвижные, окрашены каротиноидами в оранжевый цвет. Женские гаме¬
ты более крупные и двигаются медленно. После их слияния образуется
подвижная двужгутиковая зигота (планозигота), которая через некоторое
время теряет жгутики, садится на субстрат и прорастает в диплоидную
особь. На ней формируются диплоидные подвижные споры, прорастаю¬
щие в такую же диплоидную особь (бесполое размножение), а в других
клетках после мейоза — гаплоидные подвижные споры, прорастающие
в гаплоидную особь. Таким образом, на обеих особях гриб образует пять
типов подвижных спор: два типа диплоидных — планозиготы и зооспоры,
и три типа гаплоидных — зооспоры, мужские и женские гаметы. Первые
три типа клеток должны найти подходящий субстрат и прорасти на нем,
образовать мицелий. Поэтому их вкусы чисто гастрономические — поиск
животной пищи. Если к какому-либо субстрату добавить аминокислоту
глютатион, который добавляют в фарш неизвестного состава и готовят
из него «котлеты с запахом мяса», то планозигота и зооспоры будут плыть
по направлению к этому субстрату. Что касается гамет, то у них иные пред¬
почтения. Малоподвижная женская гамета выделяет в воду некое веще¬
ство, которое заставляет биться если не сердце, то жгутик мужской гаметы,
и плыть для соединения с ней. После очень трудоемкой работы это веще¬
ство было очищено, и установлена его структура. Оно оказалось низкомо¬
64
Глава 3. Как грибы размножаются
лекулярным органическим соединением — монотерпеном, содержащим
всего десять атомов углерода, и получило названием «сиренина», по име¬
ни морских ведьм — сирен, которые своим ангельским пением увлекали
моряков в пучины. Сиренинами позже стали называть также половые
феромоны других грибов. Монотерпены, подобные сиренину аллемицес,
обычны у растений. Многие из них входят в состав различных летучих ве¬
ществ, некоторые обеспечивают специфические запахи цветков. Сиренин
легко диффундирует в воде и улавливается рецепторами, расположенными
на поверхности сперматозоида. Чувствительность его рецепторов просто
невероятная: сиренин воспринимается ими в концентрации 10““10 М!
Еще интереснее обстоят дела с регуляцией поло-
Ахлия: есть пол, „ «
нет пола? вых отношении У ДРУГ°И водной плесени — псевдо¬
гриба Ахлия из порядка сапролегниевых оомицетов.
Виды этого рода образуют белый мицелиальный пушок на растительных
и животных субстратах в пресных водоемах, могут поражать рыбью икру
и мальков, причиняя урон рыбоводческим хозяйствам. Некоторые ахлии,
как и тканевые животные, двудомны, то есть у них женские половые
органы (оогонии) формируются на одних экземплярах, а мужские (анте¬
ридии) — на других. В оогониях образуются неподвижные яйцеклетки,
которые вследствие отсутствия собственной оболочки называют оосфера-
ми, а в антеридиях не образуется ничего. Половой процесс происходит
следующим образом: кончик антеридия плотно прижимается к оогонию,
их стенки растворяются под действием гидролитических ферментов, и со¬
держимое антеридия переливается в оогоний. Ядра антеридия попадают
в лишенную оболочки оосферу и сливаются с ее ядрами (рис. 6, стр. 19).
Оплодотворенные диплоидные оосферы покрываются толстой оболочкой,
превращаясь в покоящиеся ооспоры. Однако такие действия протекают
только в том случае, если партнеры растут рядышком. Если же отдельную
особь ахлии изолировать из водоема и выращивать в гордом одиночестве,
она не будет образовывать ни оогониев, ни антеридиев, а размножаться
будет исключительно бесполым способом. В общем-то ее можно понять:
зачем тратить энергию на построение каких-то морфологических структур,
если в отсутствие партнера они все равно будут бесполезными. Но стоит
подсадить к одному экземпляру ахлии второй, как начинаются удивитель¬
ные события. Один экземпляр (женский) выделяет в окружающую среду
половой феромон, под действием которого второй начинает расти ему
навстречу и формировать антеридии. Этот феромон был давно химически
охарактеризован. Для того, чтобы его идентифицировать, было собрано
1000 литров культурального фильтрата (жидкости, в которой росли ахлии),
и из него выделено 20 мг очищенного феромона. Он назван антеридио-
лом — название не столь поэтическое, как сиренин, но гораздо более
информативное, ибо, во-первых, указывает на направление действия —
индукцию формирования антеридиев, во-вторых, окончание «-ол» любому
химику указывает на наличие в молекуле гидроксильной группы. Анте-
§ 2. Вернемся к нашим грибам
65
Ои
Рис. 16. Химическая структура полового феромона оомицета ахлии (1)
и половых гормонов млекопитающих — женского гормона эстрогена (2)
и мужского гормона андрогена (3)
ридиол принадлежит к большой группе химических соединений — тет-
рациклических тритерпеноидов — стеринов, которые играют огромную
структурную и регуляторную роль в жизни разнообразных организмов.
Интересно, что мужские и женские половые гормоны млекопитающих
также являются стероидами, только был улучшен их молекулярный ди¬
зайн (рис. 16). Как видно, природа придумала химический класс молекул,
регулирующих образование половых органов, еще в те далекие времена,
когда мы были водными плесенями.
Итак, под действием феромона антеридиола образуются антеридии,
и стерильный мицелий становится мужским. Одновременно с формиро¬
ванием антеридиев он начинает выделять в окружающую среду другой
стероидный феромон — оогониол, который, достигая мицелия партнера,
вызывает формирование женских половых органов — оогониев. В ре¬
зультате такого химического диалога мицелий с антеридиями подрастает
к мицелию с оогониями, и протекает половой процесс. Рецепторами ан¬
теридиола и оогониола являются специфические белки, расположенные
на поверхности мицелия. Их чувствительность также очень велика: анте-
ридиол проявляет активность уже в концентрации 6 * 10“12 грамм в мил¬
лилитре раствора.
Перейдем к наземным грибам, коих абсолютное большинство. Пло¬
хим хозяйкам должны быть знакомы мукоровые грибы, вызывающие го¬
ловчатую плесень (тоненькие, белые, вертикально расположенные нити,
заканчивающиеся черными головками) на остатках каши в невымытых ка¬
стрюлях. Вообще-то мукоровые грибы живут в почве, где разлагают расти¬
тельные остатки и участвуют в почвообразовательном процессе, но споры
их летают везде и, попав на питательный субстрат, развивают в нем коло¬
нии. Если колонию мукорового гриба выращивать на питательной среде
в микробиологической чашке Петри, он будет размножаться исключитель¬
но бесполым способом: на погруженном в субстрат мицелии вертикально
вырастают неразветвленные нити — спорангиеносцы, на концах которых
66
Глава 3. Как грибы размножаются
формируются темноокрашенные шары — спорангии, набитые спорами.
Но если рядом подсадить новую колонию, то в 50 % случаев начина¬
ется половой процесс. Почему в 50 процентах? Потому что эти грибы,
не имея половых различий, разделяются физиологически на два типа спа¬
ривания, названные «плюс» и «минус» Если «плюс» и «минус» мицелии
растут вблизи друг от друга, то начинаются морфогенетические процессы:
перестают формироваться спорангиеносцы со спорангиями, гифы мице-
лиев растут навстречу друг другу, их кончики становятся желтыми, вслед¬
ствие накопления пигмента бета-каротина, и отделяются от остального
мицелия перегородками. Никаких гамет, ни подвижных, ни неподвиж¬
ных, не образуется, а сливаются непосредственно кончики гиф, которые
иногда называют гаметангиями, а половой процесс — гаметангиогамией.
После слияния зигота, называемая зигоспорой, отделяется от остальных
гиф и покрывается плотной темной оболочкой. Все эти процессы про¬
текают под влиянием половых феромонов, которые у мукоровых грибов
называются триспоровыми кислотами. Если (+)- и (-)-особи выращи¬
вают отдельно, триспоровые кислоты не образуются. Если же эти особи
оказались на близком друг от друга расстоянии, то, во-первых, начинает¬
ся синтез бета-каротина, во-вторых, это соединение расщепляется в ги¬
фах соответствующим ферментом на два продукта, которые претерпевают
дальнейшие ферментативные изменения, приводящие к образованию три-
споровых кислот. Но у (+)-особи есть фермент, расщепляющий продукт,
образованный (-)-особью, но нет фермента, расщепляющего промежу¬
точное вещество, образованное внутри своего мицелия, а у (—)■-особи
есть фермент, расщепляющий вещество, образованное мицелием (+)- осо¬
би, но не своим мицелием. Поэтому при росте в отдельности триспоровые
кислоты не образуются, но при совместном росте (+)- и (-)-мицелиев
каждый как бы помогает соседу добраться до конца серии химических
реакции и образовать половой феромон.
Лучше всего изучена регуляция полового про¬
цесса у сахаромицетных дрожжей. В богатой пи¬
тательной среде они и слышать не хотят о по¬
ловом размножении и размножаются исключи¬
тельно почкованием. Но если питательные веще¬
ства в среде роста оказались исчерпанными, почкование прекращается,
и в клетках дрожжей начинаются морфогенетические изменения. По-ви¬
димому, индукция полового процесса голоданием является атавизмом, ибо
у многих предков дрожжей в результате полового процесса, как уже было
сказано, образуются покоящиеся структуры, способные сохраняться при
наступлении неблагоприятных условий жизни, в том числе при истощении
пищевого субстрата. А ученые, изучающие генетику дрожжей, для индук¬
ции полового процесса просто помещают их на среды, в которых сахар
заменен солью уксусной кислоты. В таких условиях не попочкуешься. Ядра
сахаромицетных дрожжей диплоидные. При наступления голодания они
0 Регуляция
полового процесса
у сахаромицетных
дрожжей
§ 2. Вернемся к нашим грибам
67
редукционно делятся, и внутри диплоидной дрожжевой клетки образуют¬
ся четыре дочерних гаплоидных клеточки, которые освобождаются после
разрыва маминой оболочки. На правом плече третьей хромосомы дрожжей
(а хромосом у них очень много — 15, правда каждая хромосома содержит
ДНК меньше, чем у бактерии) имеется локус, содержащий несколько тес¬
но сцепленных генов. Это — локус спаривания. Продукты входящих в него
генов регулируют протекание полового процесса. Локус спаривания имеет
два аллельных состояния, то есть входящие в него отдельные участки ДНК
различаются по порядку расположения отдельных оснований и по протя¬
женности. Эти состояния названы а и а (альфа). В диплоидных клетках
третья хромосома всегда гетероаллельна, так как одна ее копия представ¬
лена фактором а, а вторая — а. Две из четырех дочерние клетки после
мейоза получают фактор а, а две другие — фактор а. а-клетки проду¬
цируют феромон а и рецепторы феромона а, а а-клетки продуцируют
феромон а и рецепторы феромона а. Оба феромона представляют собой
пептиды (короткие белки), отличающиеся числом аминокислот (12 и 13)
и некоторыми другими химическими свойствами. Рецепторы феромонов
представляют собой белки, погруженные в цитоплазматическую мембрану
и специфически связывающиеся с одним или другим пептидным феро¬
моном. Такое связывание драматически сказывается на внутриклеточных
процессах. Оно приводит к каскадной активации ферментов протеинки-
наз. Эти ферменты присоединяют богатый энергией остаток фосфорной
кислоты (РО43) к определенным белкам (фосфорилируют белки). Фосфо-
рилированный белок становится активным и участвует в следующих этапах
каскадного переноса. В конце концов фосфорилируется особый белок —
фактор активации транскрипции, который соединяется с определенным
участком ДНК и изменяет экспрессию генов, так что синтез одних гено-
продуктов оказывается подавленным, а других — резко усиливается. Для
понимания каскадного действия протеинкиназ сравним клетку с футболь¬
ным полем. В игре принимают участие много футболистов и только один
мяч. Игроки — это разные белки, а мяч — остаток фосфорной кислоты.
Игрок, не владеющий мячом, находится в пассивном состоянии, а игрок
с мячом — в активном. Задача команды заключается в том, чтобы, пере¬
давая мяч от одного игрока к другому, приблизиться к воротам и передать
мяч нападающему шлеодору (фактору активации транскрипции), который
вколотит его в ворота.
Что же происходит в результате взаимодействия РезуЛЬтат действия
фактора активации транскрипции с участком ДНК? фактора активации
1. Прекращаются деления ядер в клетке (митозы) транскрипции на ДНК
и почкование.
2. Изменяется форма клеток: вместо сферических и эллипсовидных они
становятся грушевидными, вытягиваясь навстречу друг другу (по на¬
правлению градиента концентрации феромона). Интересно, что если
поблизости находится несколько клеток с противоположным типом
0
68
Глава 3. Как грибы размножаются
спаривания, то клетка будет вытягиваться по направлению к той, ко¬
торая образует больше феромона. Так что уже у дрожжей возникла
способность выбора партнера с большей «мужской» или «женской»
силой.
3. Синтезируются поверхностные аглютинины — химерные молекулы
(древние греки химерами называли полулюдей-полуживотных, таких
как сфинкс, кентавр и прочие), в которых белок соединен с саха¬
рами — глюкозой, галактозой, рамнозой. Эти молекулы находятся в
цитоплазматической мембране, а их разветвленные веточки сахаров
выдвинуты наружу. У а- и а-клеток эти веточки подходят друг к дру¬
гу, как ключ к замку. В результате такого соединения сблизившиеся
клетки плотно склеиваются (аглютинируют).
4. Синтезируются ферменты — деполимеразы, растворяющие клеточ¬
ные стенки в местах контакта.
5. Ядра слившихся клеток мигрируют навстречу друг другу и сливаются.
На этом половой процесс завершается, и восстанавливается дипло¬
идное состояние вегетативных клеток, прекращается экспрессия генов,
которые были активны у гаплоидных клеток, восстанавливаются процес¬
сы деления ядер (митозов) и почкования.
У большинства грибов регуляция полового процесса происходит сход¬
ными путями — образованием двух типов феромонов и рецепторов к ним;
различия касаются путей морфогенеза, приводящих к спариванию и по¬
следующей судьбе диплоидной зиготы. Некоторые принципиальные от¬
личия имеются у высших базидиальных грибов, к которым относятся
трутовики, шляпочные грибы и гастеромицеты — например, всем извест¬
ные дождевики — белые шары, которые при созревании темнеют и пылят
спорами. У них нет никаких специальных органов полового размножения,
а просто сливаются гифы растущих рядом мицелиев. Такие слияния (ана¬
стомозы) не связаны с половыми различиями или различиями в локусе
спаривания, они происходят как между гифами разных особей, так и внут¬
ри одной особи. Морфогенетические процессы, обусловленные действием
половых феромонов, возникают позднее, после слияния особей, имеющих
различия в факторах спаривания. Следовательно, морфогенетические мо¬
лекулы разных особей действуют не через окружающую среду, а внутри
слившихся мицелиев. Поэтому такие молекулы можно называть не поло¬
выми феромонами, а, как и у высших животных, гормонами внутренней
секреции.
2.3. Как грибы меняют свой «пол»
У некоторых животных (общественных насеко-
мых> Рыб и ДР*) ПРИ Дефиците особей одного пола
метод смены пиле / y
(самок или самцов) особи другого пола или рабочие
особи могут менять свой пол и восполнять утрату. Такие приспособитель¬
ные механизмы появились очень давно, у грибов и псевдогрибов. Выше
§ 2. Вернемся к нашим грибам
69
было рассказано про водного оомицета Ахлию, у которого потенциально
женский индивид образует половой феромон антеридиол, а потенциально
мужской — феромон оогониол. Если мы совместим две особи, одна из них
будет вести себя как женская, а вторая — как мужская. Но если эту жен¬
скую особь совместить с третьей особью, она может вести себя с ней, как
мужская, а третья станет женской. Таким образом, можно создать коллек¬
цию экземпляров Ахлии, которые образуют непрерывный ряд. Крайний
справа в этом ряду будет всегда мужским, крайний левый — всегда жен¬
ским, а пол остальных будет зависеть от соседа справа (по отношению
к нему он будет женским) и соседа слева, по отношению к которому
он будет мужским. Причем вовсе не факт, что крайние штаммы всегда
мужские или женские. Если расширить коллекцию штаммов, то можно
найти особи «более мужские» и «более женские», чем крайние особи на¬
шей коллекции. Такой тип полового процесса назван гинандромиксисом
(от греческих слов «гина» — женское начало и «андрос» — мужское нача¬
ло). Объясняется такая смена полов просто: любой экземпляр продуцирует
как женский феромон антеридиол, так и мужской оогониол, и все зависит
от их количественного соотношения у особей, находящихся по соседству.
Наверное, люди были очень довольны, если бы подобно водным плесеням
могли менять пол в зависимости от партнера.
Совсем уж экзотическим способом меняют «пол» сахаромицетные
дрожжи. Как было сказано, под влиянием голодания в диплоидных клет¬
ках протекает редукционное деление ядра, и образуются четыре гаплоид¬
ные споры (2 а и 2 а), которые покидают материнскую клетку и попарно
сливаются (спора а сливается со спорой а). Но если с помощью специ¬
ального прибора-микроманипулятора изолировать отдельные гаплоидные
споры до их слияния, и перенести в разные пробирки с питательной сре¬
дой, то они будут размножаться там почкованием, сохраняя гаплоидное
состояние ядер. Однако уже давно было замечено, что иногда гаплоид¬
ные клетки превращаются в диплоидные вследствие их попарных слия¬
ний. Этот факт долгое время приводил в отчаяние генетиков — ведь все
потомство одной гаплоидной споры имело одинаковый тип спаривания,
(а или а), поэтому клетки, не образуя соответствующих феромонов и ре¬
цепторов к ним, просто не могли сливаться. Причину этого странного
явления установили после того, как была исследована структура генома
дрожжей. Выяснилось, что на третьей хромосоме у них находится не толь¬
ко локус спаривания, но еще два важных локуса, названных кассетами,
один из которых находится, как и локус спаривания, на правом плече,
но дальше к концу хромосомы, а второй — на левом плече. ДНК в этих
локусах может быть идентичной или а-фактору, или а, но вследствие на¬
личия негативного регулятора транскрипции ДНК в этих локусах не экс¬
прессируется, «молчит». (Причины молчания многих генов сейчас хорошо
исследованы, и это явление используется в генно-инженерных работах.)
Однако, с помощью ряда ферментов, ДНК из одной кассеты может «пере¬
70
Глава 3. Как грибы размножаются
прыгнуть» в место, занимаемое локусом спаривания, и, вытеснив старую
информацию, заменить ее собственной. Этот процесс по-научному назы¬
вается сайт-специфической генной конверсией- Если клетка, в которой
произошла такая конверсия, имела тип спаривания а (как и все клетки
гаплоидной колонии), а в кассете находилась информация, соответству¬
ющая типу а, то после конверсии клетка приобретет тип спаривания а
и сможет спариваться с соседними а-клетками. Так постепенно все клетки
колонии приобретут способность к спариванию друг с другом? и гапло¬
идная колония превратится в диплоидную.
2.4. Сколько «полов» могут иметь грибы
Выдающийся математик и биолог, работавший в первой половине
прошлого века, Р. Фишер однажды написал: «Никакой практикующий
биолог, интересующийся вопросами полового размножения, не оказыва¬
ется перед необходимостью подробной разработки последствий того, что
было бы с организмами, имеющими три пола и более; но что же еще
ему остается, если он желает понять, почему на самом деле пола все¬
го два?». Другой, уже современный нам, выдающийся биолог Р. Докинз
в ответ на этот вопрос заметил: «Существуют ли важные факты о жизни,
которые мы плохо замечаем, просто потому что нам не хватает вообра¬
жения, чтобы представить альтернативы, которые, как фишеровские три
пола, могли бы существовать в некоем вероятном мире?». Действительно,
трудно представить себе картину, отличающуюся от всего, что мы знаем.
Мужчина и женщина у людей, самец и самка у животных, даже а и а
у дрожжей и «плюс» и «минус» у мукора — два пола или два типа спа¬
ривания, с точки зрения генетики эти различия несущественны, и иных
соотношений полов нельзя себе представить. К сожалению, оба эти свет¬
лейших ума, и Фишер, и Докинз не были знакомы с микологическими
работами, ибо еще в дофишеровские времена немецкий миколог А. Книп
обнаружил у одного гриба не два и не три, а гораздо больше «полов».
Книп исследовал базидиальный трутовый гриб, вызывающий гниль
древесины. Клетки мицелия этого гриба содержат по два ядра, получен¬
ных в результате ранее произошедшего слияния одноядерных гаплоидных
гиф разных мицелиев. Так что, хотя ядра расположены отдельно, функ¬
ционально каждая клетка мицелия ведет себя как дрожжевая диплоид¬
ная клетка. Такое состояние ядер в мицелии называют дикарион, то есть
двуядерность. На дикариотичном мицелии вырастают плодовые копыто¬
образные тела, в отдельных клетках которых происходит слияние ядер
дикариона, а затем — редукционное деление, приводящее к образованию
четырех (как и у дрожжей) гаплоидных спор. Споры высыпаются из пло¬
дового тела и прорастают гаплоидным (монокариотическим) мицелием,
который тонкий, недолговечный и, как правило, не образует плодовых тел.
Но находящиеся рядом гаплоидные мицелии могут сливаться друг с дру¬
гом, и тогда образуется мощный многолетний дикариотический мицелий,
§ 2. Вернемся к нашим грибам
71
который заражает деревья и на котором возникают плодовые тела. Скре¬
щивая друг с другом монокариотические проростки спор, полученных
из одного плодового тела (братьев и сестер, или, как их называют генети¬
ки, сибсов), он установил несколько важных правил:
1. Этот гриб (как и большинство других трутовых грибов) имеет не один
локус спаривания, как у мукоровых грибов или дрожжей, а два, ко¬
торые Книп назвал буквами А и В.
2. Скрещивание с образованием дикариотического мицелия возможно
только между монокариотическими (имеющими одноядерные клет¬
ки) гифами, в ядрах которых факторы А и В имеют различия в опре¬
деленных участках ДНК (гетероаллельны по факторам спаривания).
Например, если у одного мицелия обозначить факторы спаривания
как Ах Вх, то он может скрещиваться с индивидом, имеющим фак¬
торы спаривания АУВУ, но не с АХВХ, АХВУ и АУВХ. В выросшем
в результате скрещиваний мицелии каждое ядро дикариотической
(двуядерной) клетки будет отличаться от соседнего ядра состояни¬
ем факторов спаривания, а после слияния ядер и их редукционного
деления образуются четыре типа одноядерных гаплоидных спор: два
родительского типа (Ах Вх и АУВУ) и два — не родительского типа
вследствие рекомбинации (АХВУ и АУВУ). Поскольку скрещиваться
могут только проростки спор, различающиеся по обоим типам спа¬
ривания, проросток каждой споры может дать потомство только при
скрещивании с одним из четырех сибсов. Такой тип полового процес¬
са Книп назвал тетраполярным (в отличие от биполярного у дрожжей
и Мукора), так как мы имеем здесь дело не с двумя (как у всех извест¬
ных организмов), а с четырьмя если не полами, то физиологическими
различиями, необходимыми для успешных скрещиваний.
3. Каждый локус спаривания (и А, и В) встречается в природе не в двух
аллельных состояниях, как у других грибов и не грибов (а и а,
«плюс» и «минус»), а во многих, буквально в десятках. Поэтому ве¬
роятность того, что два случайно встретившихся в природе индивида
будут иметь одинаковые факторы спаривания, очень низка. А по¬
скольку по правилу Книпа для успешного скрещивания необходимы
у встретившихся монокариотических индивидов генетические раз¬
личия по обоим факторам, практически любые природные встречи
приводят к успешному половому процессу.
А теперь давайте напряжем свое воображение
^ Следствие много-
и подумаем, к чему приведет эта невероятная по Фи- половости грибов
шеру многополовость. Наличие двух полов или двух
типов спаривания (это неважно) не запрещает полового процесса между
близкими родственниками (братьями и сестрами). Запреты или, наоборот,
разрешения на инцест в человеческом обществе определяются морально-
этическими, а не биологическими законами. Например, в древнем Египте
72
Глава 3. Как грибы размножаются
фараон должен был обязательно жениться на родной сестре. Жизненный
опыт показал, что чем ближе родство супругов, тем выше вероятность
рождения неполноценных детей (вследствие перехода рецессивных му¬
тантных генов в гомозиготное состояние). Поэтому многие религии ввели
запрет на браки между братьями и сестрами. Однако степень жесткости
таких запретов в разных религиях неодинакова. Иудеи и христиане-ка-
толики запрещают браки только между родными братьями и сестрами,
но разрешают между двоюродными (кузенами), а православные христи¬
ане ввели запрет и на браки между двоюродными братьями и сестрами.
Одним словом, договоримся, что хотя биологических запретов на близ¬
кородственные скрещивания нет, такие скрещивания (инбридинг) хуже,
чем отдаленные (аутбридинг). Правда, такое утверждение вовсе не одно¬
значно, но примем его для дальнейших рассуждений.
Наличие двух полов (или биполярного гетероталлизма, неважно) де¬
лает вероятность (потенциал) инбридинга равным 50 % (сестра может ро¬
дить ребенка от брата, но не от сестры, а «плюс» индивид Мукора может
скреститься только с «минус» индивидом). При тетраполярном гетеротал¬
лизме согласно второму правилу Книпа потенциал инбридинга составляет
25 % (Ах Вх может скреститься с АУВУ, но не АХВУ и АУВХ). То есть ве¬
роятность инбридинга снижается вдвое.
Далее. При двуполовости, или биполярном гетероталлизме, вероят¬
ность неродственных скрещиваний (аутбридинга) составляет тоже 50%,
вследствие встречаемости двух разных полов или типов спаривания. У тру¬
товых грибов, согласно третьему правилу Книпа, каждый фактор спари¬
вания (А и В) представлен множеством аллельных состояний, поэтому ве¬
роятность успешного аутбридинга приближается к 100%. Следовательно,
система регуляции полового процесса у данной группы грибов успешно
снижает вероятность близкородственных скрещиваний и повышает веро¬
ятность отдаленных. Поэтому она делает каждый индивид уникальным.
Эта система характерна не только для трутовых грибов, но встречается
и у многих шляпочных макромицетов.
Молекулярные исследования позволили установить механизмы дей¬
ствия генопродуктов фактора спаривания у шляпочных и трутовых грибов.
Фактор В включает две группы генов (кассеты), одна из которых контроли¬
рует синтез феромонов, а вторая — рецепторов к феромонам. Кассеты фак¬
тора А кодируют образование специфических белков. Эти белки (моно¬
меры), продуцируемые разными аллелями А фактора соединяются вместе,
образуя гетеродимер. В состоянии гетеродимера эти белки становятся фак¬
торами регуляции транскрипции, то есть соединяются с определенными
участками ДНК и изменяют генную активность. Совместное действие фе¬
ромонов, рецепторов и гетеродимерных белков приводит к морфологиче¬
ским и физиологическим изменениям, происходящим после слияния гиф.
Такая система размножения привела, во-первых, к увеличению про¬
дукции полового размножения — гаплоидных спор и, в связи с этим,
§ 2. Вернемся к нашим грибам
73
во-вторых, к потере роли бесполого размножения, которое у одних видов
вообще отсутствует, а у других несет функции не размножения, а сохра¬
нения (хламидоспоры).
2.5. Морфологические структуры
Параллельно с половым процессом у большинства грибов протекают
морфогенетические процессы, и образуются специальные структуры, име¬
ющие важные функции в жизни грибов.
Выше при рассказе о половом процессе у мукоровых
грибов говорилось, что после слияния гаметангиев образу- функции
ется зигоспора, окруженная толстой оболочкой. У многих
видов эта оболочка черная, меланизированная, а у некоторых еще и снаб¬
жена выростами. Особенно впечатляют крупные зигоспоры у гриба Фи-
комицес, у которых остатки несущих гаметангии гиф и выросты в виде
оленьих рогов делают зигоспоры
похожими на панцири тевтонских
рыцарей из фильма «Александр
Невский» (рис. 17).
У сумчатых грибов-аскомице-
тов после редукционного деления
ядра зиготы образуется 4 или 8 гап¬
лоидных спор внутри клетки, кото¬
рая называется сумкой. Большин¬
ство сумчатых грибов различными
способами защищают свои сумки.
Вокруг них образуются из спле¬
тения гиф защитные образования,
называемые плодовыми телами или
аскомами (рис. 18). В клетках пло¬
довых тел многих грибов откла¬
дываются темные пигменты, дела¬
ющие плодовые тела более проч¬
ными. Некоторым грибам и это¬
го оказалось мало — их плодовые
тела погружены в плотные спле¬
тения гиф, стромы, которые у од¬
них видов мягкие, светлоокрашен¬
ные (рис. 25), а у других — черные,
жесткие, углистые. х
Плодовые тела некоторых сумчатых грибов-дискомицетов плоские,
блюдцевидные, покрытые сверху защитной пленкой мицелий. Их назы¬
вают апотециями (рис. 18 в). Дно апотеция выстлано вытянутыми верете¬
нообразными сумками со спорами. После созревания спор пленка разры¬
вается, освобождая диск плодового тела. В это время в сумках происходит
Рис. 17. Зигоспора гриба фикомицес с ос¬
татками мицелия (суспенсорами) желто¬
го цвета и черными придатками (Cerda-
Olmedo Е., Lipson Е. D., 1987)
74 Глава 3. Как грибы размножаются
Рис. 18. Плодовые тела сумчатых грибов: а) клейстотеций — замкнутое
плодовое тело с сумками, расположенными беспорядочно между гифами;
б) периотеций — плодовое тело с отверстием наверху; сумки расположены
пучком на дне перитеция, после созревания поочередно высовываются и от¬
стреливают споры; в) апотеций — открытое плодовое тело; сумки покрывают
сплошным слоем (гимением) поверхность апотеция
Функции увеличения
продуктивности и
распространения спор
взрывообразный синтез гидролитических фермен¬
тов, которые превращают полисахарид гликоген в
простые сахара. Тургорное давление в сумке резко
увеличивается, и ее оболочка разрывается там, где
тоньше, а тоньше — на верхушке сумки, и споры с силой выбрасываются
вверх, где подхватываются воздушными токами и разносятся. У некото¬
рых дискомицетов блюдцевидные апотеции сидят на тонкой ножке, тогда
они становятся больше похожи не на блюдца, а на рюмочки с длинными
ножками. Это позволяет поднять их повыше над поверхностью почвы,
чтобы споры дальше распространялись. Дальше — больше. Вместо тон¬
кой ножки возникли апотеции с толстыми и поэтому более длинными
ногами, а края диска апотеция завернулись вниз, образуя как бы шляпку
над ножкой. На самом диске образовались многочисленные морщинки
и складки. Такое строение позволило
разместить большее число сумок на по¬
верхности диска, совсем теперь не по¬
хожего на диск. Так выглядят плодовые
тела сумчатых макромицетов — сморч¬
ков и строчков (рис. 19). При таком
обилии спор, образовавшихся в резуль¬
тате полового процесса, бесполое раз¬
множение оказалось лишним.
Еще сложнее устроены плодовые
тела у базидиальных макромицетов.
Многие виды имеют всем знакомую,
поднимающуюся над почвой ножку с
шляпкой сверху. Верхняя часть шляп¬
ки образована, как и ножка, плотным
Рис. 19. Сморчки и строчки: а) смор¬
чок конический; б) строчок (Жизнь
растений, т. 2, 1976)
§ 2. Вернемся к нашим грибам
75
сплетением гиф, а нижняя часть образована «тканью», несущей гапло¬
идные споры, которые свисают на тонких ножках по четыре штуки под
материнскими клетками, называемыми базидиями. Этот слой называется
гименофором и ради него затеяны все остальные устройства плодовых
тел. Для того, чтобы на поверхности гименофора поместилось бы поболь¬
ше базидий со спорами, он имеет складочки, пластинки (как у сыроежки
или шампиньона) или тонкие трубочки, усеянные базидиями (как у белого
гриба или масленка). В отличие от сумчатых грибов, у которых слой сумок
(гимений) обращен вверх, гименофор базидиальных грибов всегда нахо¬
дится на нижней стороне плодового тела. Это и понятно. Ведь базидио-
споры находятся не внутри материнской клетки, как у сумчатых грибов,
а на тонких ножках-стеригмах, поэтому тур горное давление внутри бази-
дии не может их отбросить с такой же силой; если бы гименофор находил¬
ся на наружной поверхности плодового тела, то споры просто падали бы
на дно пластинок или трубочек. В связи с этим плодовые тела должны
решать три задачи: 1) увеличить число образующихся спор (для этого су¬
ществуют различные складки и трубочки гименофора); 2) поднять его как
можно выше над поверхностью земли (для этого вырастает длинная «но¬
га»); 3) всегда ориентировать гименофор параллельно поверхности земли
с нижней стороны плодового тела. Последнее правило хорошо иллюстри¬
ровать строением многолетних копытообразных плодовых тел трутовых
грибов. Пока дерево стоит вертикально, копыта трутовиков всегда на¬
правлены гименофором вниз (это только иллюстраторы детских сказок
рисуют иногда перевернутые трутовики на пнях). Если дерево упадет, то
имеющееся уже плодовое тело начинает уродливый рост так, чтобы новый
слой гименофора становился не перпендикулярно, а параллельно направ¬
лению ствола дерева, то есть опять же параллельно поверхности почвы.
У многих видов шляпочных (агариковых) грибов зачатки плодовых
тел имеют гименофор, покрытый защитной пленкой из сплетения гиф.
Когда плодовое тело растет и увеличивается в размерах, пленка разрывает¬
ся, чтобы освободить созревшие споры. При этом на краях шляпки оста¬
ются остатки покрывала в виде бахромы, а на ножке — в виде колечка, как,
например, у шампиньона или гриба
зонтика. А у некоторых видов кроме
такого «частного» покрывала имеет¬
ся общее, покрывающее все плодовое
тело. Когда «гриб» вырастает, общее
покрывало разрывается, и его остат¬
ки имеют вид разорванной «яичной
скорлупы», вольвы, из которой вы¬
ходит ножка плодового тела, и белых
пятен на шляпке, особенно хорошо
заметных на красной шляпке мухо¬
мора (рис. 20).
Рис. 20. Мухоморы: а) мухомор
красный; 6) бледная поганка
76
Глава 3. Как грибы размножаются
Есть еще одна удивительная группа базидиальных макромицетов — гас-
тероидные грибы. «Гастер» — по-латыни «желудок» (отсюда — гастроном).
У гастероидных грибов базидиоспоры образуются не на нижней поверх¬
ности плодового тела, а внутри него. Всем хорошо знакомы грибы дожде¬
вики. Они в незрелом состоянии имеют вид
белых шаров, покрытых толстой оболочкой.
При созревании шары темнеют, их оболочка
становится более тонкой и хрупкой, а все со¬
держимое превращается в споры, порошок ко¬
торых, как дымок, освобождается после раз¬
рыва оболочки в верхней части плодового те¬
ла. Количество спор, образующихся в одном
плодовом теле дождевика, совершенно неве-
Рис.21. Плодовое тело до- роятно. Один и видов дождевиков, головач
ждевикового гастеромицета гигантский, образует плодовые тела размером
головача (Т.Лессо, 2003) больше головы человека и весом до 20 кг
(рис. 21). В таком плодовом теле содержится
до 8 триллионов спор. Допустим для простоты, что средний вес плодового
тела головача составляет 10 кг, а число спор в нем — 1 триллион (1012).
Если бы каждая спора проросла и образовала новое плодовое тело, вес
потомков одного плодового тела в первом поколении составил бы 10 трил¬
лионов кг, то есть 1013 кг, а во втором поколении — 1026 кг. Поскольку
масса земного шара равна примерно 6.» 1024 кг, вес потомства одного
плодового тела головача во втором поколении значительно превысил бы
вес Земли со всеми ее морями, горами, ядром и магмой. К счастью для
Земли и ее обитателей, прорастает и дает потомство ничтожная доля об¬
разовавшихся спор, то есть избыточность половой продукции дождевиков
не поддается воображению.
2.6. Удивительные явления в мире грибов
Закончить эту главу мне хочется рассказом о некоторых совершенно
удивительных приспособлениях, которые были выработаны в ходе эво¬
люции грибов и направлены не на тупое увеличение числа спор, как у
дождевиков, а на мудреные способы направленной доставки спор к месту
назначения.
У польского философа и писателя-фантаста Станислава Лема есть
сборник юмористических рассказов «Звездные дневники Ийона Тихого»,
в которых космонавт рассказывает о своих полетах
трель а в цель на разд^щ^е обитаемые небесные тела и о нравах
живущих там аборигенов. В одном из рассказов описана планета, на кото¬
рой обитают огромные, размером с трехэтажный дом животные с толстой,
абсолютно непроницаемой кожей. Местные жители охотятся на них «из¬
нутри». Охотник обмазывает себя соусом, запах которого возбуждает ап¬
петит зверя, берет с собой бомбу с часовым механизмом, большую порцию
§ 2. Вернемся к нашим грибам
77
слабительного лекарства и прячется в кустах. Животное по запаху находит
его и проглатывает. В кишечнике охотник закладывает бомбу и высыпает
слабительное. В ответ на это животное выбрасывает охотника с экскре¬
ментами (это — самый опасный момент охоты, так как приходится падать
с трехметровой высоты), а затем взрывается. На вопрос Ийона, почему
надо так рисковать, нельзя ли просто спрятать бомбу, обмазанную соусом,
ему ответили, что такой способ охоты запрещен, как браконьерство.
Эта история приходит на ум при знакомстве с образом жизни одного
из интереснейших мукоровых грибов по имени Пилоболюс. Мицелий этого
гриба живет в навозе травоядных животных и питается непереваренными
растительными остатками. Из навозной кучи наружу вырастают, как и
положено мукоровым грибам, спорангиеносцы со спорангиями на конце.
Но, в отличие от своих родственников, спорангиеносец Пилоболюса за¬
канчивается утолщением в виде пузыря, на конце которого находится
небольшой спорангий, плотная оболочка которого окрашена меланином
в черный цвет. Пузырь окрашен в нежный желтовато-розовый цвет, на¬
поминающий, по определению JI. Н. Толстого, «цвет бедра испуганной
нимфы», вследствие накопления в нем светочувствительных пигментов
каротиноидов и выполняет роль линзы, фокусирующей солнечные лучи
(рис. 22). После созревания спор в спорангии, в спорангиеносце накапли-
Рис. 22. Зигомицет пилоболус: а) спорангиеносец со спорангием (Жизнь растений,
т. 2, 1976); б) механизм искривления спорангиеносца (З.Э. Беккер, 1963)
78
Глава 3. Как грибы размножаются
ваются гидролитические ферменты, запасные полисахариды превращают¬
ся в простые сахара, резко увеличивается тургорное давление и спорангий
целиком отстреливается от спорангиеносца, отлетая на несколько метров.
Если лучи солнца падают сверху, параллельно ножке спорангиеносца, то
спорангий выстреливает вертикально вверх, но если луч падает на споран-
гиеносец сбоку, то он, как показано на рисунке, изгибается в направлении
солнца и стреляет в наиболее освещенное место. А там, где светло, лучше
растет трава. Скажем, в чаще леса травы мало, так как не хватает света для
ее роста, а на полянках и лесных прогалинах трава растет гораздо гуще.
Вот и получается, что гриб стреляет в цель. В месте соединения спорангия
с пузырем спорангиеносца имеется слизистое клейкое кольцо, с помо¬
щью которого спорангий, попав на листик, приклеивается к нему. Если
мимо идет жвачное животное, будь то лось или корова, спорангий, как
охотник из сказки Лема, вместе со съеденной травой попадает в желудок,
и только там его оболочка под действием пищеварительных ферментов
разрушается, споры освобождаются и выбрасываются вместе с навозом
наружу. Таким образом, мы видим серию удивительных приспособлений:
гриб стреляет спорангием в места, где наибольшая вероятность попасть
на траву, приклеивается к ней, освобождает споры только в желудке тра¬
воядного животного и выбрасывает их с тем субстратом, который нужен
для его питания.
Спорангии пилоболюса достаточны крупные и видны невооружен¬
ным глазом в виде черных точек. Если между спорангиеносцами и свето¬
вым лучом поместить стеклянную пластинку, то можно подсчитать число
отстрелянных спорангиев. Как-то один студент биологического факуль¬
тета московского университета на летней практике, узнав о поведении
этого гриба, так им заинтересовался, что после практики остался на ка¬
никулярное время, носил из леса кучки навоза, помещал их на тарелку,
закрывал стеклянным колпаком, чтобы поддерживать высокую влажность
и занимался экспериментами с грибами. Он соорудил установку, в кото¬
рой между источником света и навозной кучей находились стеклянные
пластинки, как бесцветные, так и различных цветов — синего, красного
и прочих. Такие установки с разными светофильтрами он помещал с двух
противоположных сторон кучи и, считая число спорангиев на пластин¬
ках, пытался определить, какой свет предпочитает пилоболюс. Студенты
зоологи смеялись, говоря, что он изучает цветное зрение у грибов.
Иной способ «стрельбы в цель» применяют некоторые базидиальные
грибы. Плодовое тело гриба Круцибулюм (трубач) имеет вид раструба тру¬
бы или горна (рис. 23). К боковым стенкам его прикреплены «грибочки»,
имеющие ножки (фуникулюсы) и шляпки (перидиоли). В перидиолях со¬
держатся гаплоидные базидиоспоры. Внутри фуникулюса находится чрез¬
вычайно гигроскопичная, суперскрученная нить — корд. Когда внутрь
плодового тела в дождливую погоду попадают капли воды, нить момен¬
тально раскручивается, вытягиваясь до длины 15-20 см, и отлетает вместе
§ 2. Вернемся к нашим грибам
79
Рис. 23. Внутреннее строение плодового тела
гастеромицета круцибулюм (Alexopolus C.J. et al., 1996)
с перидиолью на расстояние до метра и больше от плодового тела. Кончик
нити имеет клейкую поверхность, с помощью которой нить приклеивает¬
ся к твердому предмету и закручивается вокруг него, прочно прикрепляя
перидиоль со спорами. Так что споры и спорангии грибов могут, как и во¬
енная техника, стрелять различными способами: и как снаряды, и как
ракеты. Главное — точность попадания в цель.
Различные насекомые имеют на своих усиках-антеннах
рецепторы, чрезвычайно чувствительные к летучим хими¬
ческим веществам. Всем хорошо известны многочисленные
ухищрения, с помощью которых цветковые растения привлекают к себе
насекомых-опылителей. В частности, ароматы цветков существуют не для
того, чтобы вызывать у нас восхищение, а для того, чтобы привлекать
насекомых. Но гораздо менее известно, что и грибы научились выделять
привлекающие насекомых ароматы.
Понаблюдайте поздней весной, в конце мая - начале июня, за побегами
одного из самых распространенных наших сорняков — осота. Отдельные
побеги его отстают в росте и имеют более мелкие листья, покрытые желто¬
ватыми пятнами. Если такой побег понюхать, можно почувствовать тон¬
кий, приятный цветочный аромат, хотя никаких цветков на таких побегах
80
Глава 3. Как грибы размножаются
нет и не будет. Эти побеги заражены ржавчинным грибом. Весенние споры
ржавчины — спермации — распространяются насекомыми-опылителями.
А цветочный запах — обман, имитация, придуманная для их привлечения.
Некоторые грибы пошли еще дальше. Ржавчина резухи (декоратив¬
ное растение из семейства капустных) образует не только летучие вещества
с цветочным ароматом, но и вызывает сильные морфологические изме¬
нения зараженного растения: увеличивается число листьев, а на сильно
вытянутых цветоносах образуются не цветки резухи, а многочисленные,
светло-желтые от грибных структур, спермогониев, листочки, внешне на¬
поминающие цветки лютика — растения, относящегося к другому се¬
мейству. Для привлечения насекомых-опылителей этот гриб имитирует
не только запах, но и форму цветка. Также поступает и другой гриб, мо-
нилия, поражающий бруснику и чернику. Ранней весной его споры зара¬
жают стебли и листья этих растений, которые приобретают запах цветков,
появляющихся гораздо позже, и отражают ультрафиолетовые лучи так же,
как и цветки растений-хозяев. Насекомые-опылители кормятся на за¬
раженных растениях сахаристым нектаром, который выделяется грибом,
и разносят его споры.
Наши вкусы не всегда совпадают со вкусами насекомых. Например,
трупный запах, отвратительный для людей, вызывает восхищение у многих
насекомых, таких как мясные мухи. И любовью насекомых к подобным
запахам также пользуются некоторые грибы. Есть замечательная группа
базидиальных грибов, относящаяся к порядку весёлковых. Гриб весёлка,
который микологи назвали Фаллюс (рис. 1 а9 стр. 11), в незрелом состоя¬
нии имеет вид яйца, прикрепленного к почве пучком гиф мицелия. Внут¬
ри «яйца» формируется ножка с шляпкой. Затем ножка начинает быстро
расти, разрывает оболочку «яйца» и может достигать длины в несколько
десятков сантиметров. Скорость роста ножки невероятна — до половины
сантиметра в минуту. Ножка покрыта зеленовато-черной, морщинистой,
слизистой шляпкой — глебой, на которой образуются базидии с базидио-
спорами и которая издает сильнейший трупный запах; нос человека обна¬
руживает этот запах на расстоянии нескольких десятков метров от гриба,
а рецепторы насекомых — на значительно большем расстоянии. Трупные
мухи слетаются со всей округи и устраивают шабаш вокруг гриба, разнося
его споры. Гриб как быстро вырастает, так и быстро разлагается. Через
сутки от него остается дурно пахнущий комочек слизи. У родственника
весёлки, гриба Диктиофора, растущего в тропических и субтропических
зонах, в добавок к «приятному» запаху из-под шляпки глебы на нож¬
ку опускается индузий — ажурная кружевная юбочка из сплетения гиф
мицелия, которая светится в темноте бело-зеленоватым светом, что так¬
же привлекает ночных насекомых (рис. 1 б). А у гриба решеточника нет
ножки, и из разорвавшегося «яйца» выходит глеба, имеющая вид ярко-
красной решетки и привлекающая дневных насекомых (рис. 1 в).
§ 2. Вернемся к нашим грибам
81
Некоторые несовершенные (потерявшие интерес к
половому размножению) грибы нашли новую для себя азад в воду
жизненную нишу: они ушли в пресные водоемы — речки, ручейки —
и разлагают там растительный опад — листья и веточки, упавшие в воду.
Зооспор, которые имеют первично водные грибы и псевдогрибы, они не
приобрели, ибо эволюция не имеет обратных путей (назад дороги нет),
но зато придумали, каким образом цепляться за опавшие листья и ветки
в водном потоке. Их конидии приобрели удивительный вид, стали круп¬
ными, многоклеточными, с различными выростами, лучами и другими
образованиями (рис. 24). Конечно, такие хонидии, отрываясь от мицелия,
разносятся водой до тех пор, пока не придут в соприкосновение с тверды¬
ми телами, находящимися в воде, заякориваются там, а затем прорастают.
Глава 4
Вторичные
метаболиты грибов
§ 1. Первичный и вторичный
метаболизм
Обмен веществ, или, по-научному, метаболизм, бы¬
вает двух типов. Первый тип — конструктивный или
пластический метаболизм (его еще называют анаболиз¬
мом), а второй — деструктивный или энергетический
(или катаболизм). Для чего нужен пластический мета¬
болизм, понятно: для увеличения размеров имеющихся
и создания новых клеток, органов и тканей в процессе
роста и развития организма. В ходе пластического мета¬
болизма создаются новые молекулы, необходимые для
жизнеобеспечения. Деструктивный метаболизм нужен
не только для разрушения отжившего. В процессе рас¬
пада сложных органических молекул высвобождается
энергия, которая запасается в специальных энергети¬
ческих молекулах (аденозин и гуанидин трифосфатах,
никотинамид дифосфате и других), имеющих остатки
фосфорной кислоты, запасающие энергию. Без этой
энергии создание новых веществ в процессах пласти¬
ческого метаболизма было бы невозможно.
Пластический метаболизм разделяют на первичный
и вторичный. Это деление условно, как условна почти
любая биологическая классификация, но оно все-таки
полезно, так отражает основные направления потоков
вновь созданных метаболитов в жизни организма.
В ходе первичного метаболизма создаются веще¬
ства, без которых не могут существовать не только целые
организмы, но и каждая клетка, их составляющая. Это
1) нуклеиновые кислоты, хранители информации обо
всех процессах, протекающих в клетках;
2) белки, функции которых чрезвычайно разнообразны:
и ферменты, и молекулы, участвующие в иммунных
§2. Токсины
83
реакциях, и скелет клеток — микротрубочки и микрофиламенты,
не только укрепляющие каркас клетки, но и создающие рельсы, по
которым идут потоки молекул внутри, а иногда и между клетками,
и которые осуществляют растягивания хромосом к полюсам клетки во
время митозов, и строительные молекулы, участвующие, в частности,
в построении клеточных покровов;
3) липиды — основной компонент мембран, покрывающих клетку и от¬
дельные ее структуры, такие как ядро, вакуоли, митохондрии, хлоро-
пласты;
4) полисахариды, которые необходимы для построения клеточной стен¬
ки у растений и грибов, отлагаются в виде запасных питательных
веществ и прочее.
В основном, но не всегда, первичные метаболиты представляют собой
полимерные, а следовательно, высокомолекулярные соединения. Посколь¬
ку важнейшие процессы и соединения, необходимые для жизнедеятельно¬
сти клетки, такие как хранение информации и ее реализация, деятельность
ферментов, построение внутриклеточных и наружных мембран и прочие
одинаковы у всех организмов, первичные метаболиты у разных групп ор¬
ганизмов сходны.
Вторичные метаболиты — это вещества не необходимые для жизни,
но облегчающие существование в определенных условиях. Например, жи¬
тели жарких экваториальных стран (негроидная раса) имеют черный цвет
кожи, а жители умеренных и холодных поясов земли (европеоидная ра¬
са) — белый. Следовательно, откладывающийся в коже темный пигмент
имеет приспособительное значение и относится не к первичным, а к вто¬
ричным метаболитам. Вторичные метаболиты, как правило (но не всегда),
представляют собой мономеры и, следовательно, являются низкомоле¬
кулярными продуктами. В отличие от первичных метаболитов, которые
сходны у организмов, относящихся к разным таксонам, вторичные мета¬
болиты гораздо более индивидуальны. А поскольку они играют важную
роль в адаптациях к условиям жизни и во взаимодействиях между орга¬
низмами в биоценозах, рассказ о жизни грибов был бы не полным без
упоминания об этих их соединениях.
§2. Токсины
Токсические метаболиты грибов, опять-таки — условно, разделяют
на три группы: вещества, токсичные для микроорганизмов (антибиотики);
вещества, токсичные для растений (фитотоксины); вещества, токсичные
для позвоночных животных и человека (микотоксины). Почему такое де¬
ление условно? Да все потому же: одно и то же вещество может быть
токсичным для двух или даже всех трех указанных групп организмов.
Поэтому мы эти группы токсических метаболитов будем рассматривать
отдельно лишь исходя из общепринятой классификации.
84 Глава 4. Вторичные метаболиты грибов
2.1. Антибиотики
Грибы сапротрофы, обитающие в почве, находятся там в сложном
микробоценозе, включающем разнообразные группы грибов, бактерий
и простейших животных. И все эти организмы имеют одну цель — оста¬
вить как можно больше потомков, а для этого им необходимо хорошее
питание. Поэтому в почвенных ценозах непрерывно происходит конку¬
рентная борьба между разными микроорганизмами за источники пита¬
ния. Конечно, такая борьба несколько ослаблена наличием групп мик¬
роорганизмов, отличающихся пищевой специализацией. Например, есть
бактерии, вырабатывающие специальный фермент нитрогеназу, который
позволяет им использовать атмосферный азот, количество которого очень
велико, для получения аммония; а из него можно уже строить и элементы
белка — аминокислоты. Такие бактерии, называемые азотфиксаторами,
не нуждаются в органических источниках азота — белках и основаниях
нуклеиновых кислот, то есть они не конкурируют с другими микроорга¬
низмами за эти источники пищи. А микроорганизмы целлюлозолитики
извлекают углерод из прочных и труднодоступных для других микроорга¬
низмов источников целлюлозы. Но первая группа нуждается в источниках
углерода, а вторая — в источниках азота, так что специализация снимает
конкуренцию лишь частично. Все почвообитающие сапротрофные мик¬
роорганизмы должны иметь широкие и острые локти для того, чтобы
оттереть конкурентов, быстрее добраться до блюда с пищей и отхватить
кусок побольше. Эти «локти» могут быть двух типов, причем у самых
активных микроорганизмов присутствуют оба типа: высокомолекулярные
ферменты, позволяющие быстрее разрушить субстрат и поглотить образо¬
вавшиеся продукты, и низкомолекулярные антибиотики, убивающие или,
по крайней мере, тормозящие рост микробов-конкурентов. Поэтому боль¬
шинство антибиотиков, применяемых в медицине и сельском хозяйстве,
получено из почвообитающих микроорганизмов — актиномицетов (ми¬
целиальных бактерий) — и грибов.
Первый антибиотик, получивший широкое
Полезные токсины —
антибиотики практическое применение, — продукт распро¬
страненного в разных почвах рода грибов Пени-
циллиум, он был обнаружен английским микробиологом А. Флемингом
в 1929 г. и назван пенициллином. В чистом виде он получен химиками
X. Флори и Е. Чейн, за что все трое ученых были удостоены Нобелевской
премии. Пенициллины и родственные им антибиотики цефалоспорины,
продуцируемые грибами из рода Цефалоспориум, подавляют образова¬
ние пептидоглюкана — основного компонента клеточной стенки многих
бактерий. Поэтому, когда бактериальная клетка находится в покое и не де¬
лится, она не чувствительна к пенициллину, но как только начинается ее
деление, содержимое растущей новой клетки остается без каркаса из стен¬
ки и погибает. Интересно, что химическое сходство с пенициллином име¬
ется у большого числа соединений, продуцируемых различными грибами
§2. Токсины
85
и бактериями. Последние этапы синтеза этих соединений осуществляется
ферментами, которые находятся под контролем трех сцепленных (распо¬
ложенных рядом друг с другом) генов. Установление структуры этих генов
у разных видов грибов и бактерий показало их удивительное сходство
и привело исследователей к парадоксальному заключению. Существует
метод расчета времени, которое прошло после расхождения сравнивае¬
мых видов, по числу замен в аминокислотах одного и того же белка или
азотистых оснований в одном и том же гене (метод молекулярных часов).
Сравнение разных генов у бактерий и грибов показало, что расхождение
между ними произошло более 1,5 миллиарда лет назад, что соответствует
времени превращения прокариотной клетки в эукариотную, однако после
сравнения генов синтеза соединений пенициллинового ряда был сделан
вывод, что они возникли у грибов гораздо позже, около 370 миллионов
лет назад. Предполагается, что эти гены появились у грибов вследствие
горизонтального переноса участка хромосомы, содержащего их кластер
(группу). Все носители этих генов (и бактерии, и грибы) встречаются
в почве, где, возможно, и произошел горизонтальный перенос группы
сцепленных генов из бактериальной клетки в грибную.
Большая группа антибиотиков, образуемых грибами, убивает другие
грибы, а не бактерии. Таковы антибиотики трихотецин и триходермин,
имеющие сходные химическую природу и механизмы токсического дей¬
ствия. Интересно, что грибы из родов Трихотециум и Триходерма, об¬
разующие эти антибиотики, развиваются не только на мертвых и живых
тканях растений, но также часто нападают на другие грибы и паразитируют
на них, то есть являются паразитами грибов (микопаразитами). Выделяе¬
мые антибиотики используются ими как орудие нападения на грибы.
2.2. Фитотоксины
Американцы, как и англичане, до появления
Макдоналдсов с фастфудами завтракали овсяной ео
кашей, поэтому овес в США выращивали на боль¬
ших площадях, несмотря на его частую поражаемость корончатой ржав¬
чиной. В 1942 г. в Америку попал сорт овса Виктория, устойчивый к аме¬
риканским расам ржавчины. Потомки сорта Виктория, имеющие его ген
устойчивости, заняли в некоторых «овсяных» штатах 80-100 % площа¬
дей, отведенных под овес. Однако вскоре новые сорта стали погибать
от неизвестной болезни, вызывающей загнивание корневой шейки и по-
бурение листьев. Из пораженных растений микологи выделили новый для
науки гриб, который назвали Кохлиоболюс виктории. Этот гриб зара¬
жал подземные части растений и выделял в зараженные растения токсин,
названный викторином, который и вызывал «ожог» листьев. Причем пора¬
жался только овес и только те его сорта, которые имели ген устойчивости
к корончатой ржавчине, перенесенный из сорта Виктория. Другие сорта
овса были устойчивы к действию токсина. Такая высокая избиратель¬
86
Глава 4. Вторичные метаболиты грибов
ность может быть обусловлена лишь тем, что продукт гена — рецептор
токсина. Если нет соответствующего гена, то нет и рецептора; в таких
растениях токсин не связывается клеткой и не может нанести ей повре¬
ждение. Другая особенность викторина — его ни с чем не сравнимая
токсичность. Проростки семян сорта Виктория погибали в растворе вик¬
торина, разведенного в миллиард раз. По отношению к чувствительным
клеткам викторин оказался одним из самых токсичных природных со¬
единений: ген сорта Виктория повышает чувствительность клеток овса
к токсину в 400 тысяч раз. Даже знаменитый колбасный яд ботулин,
вырабатываемый анаэробными бактериями, не действует в таких низких
концентрациях, как викторин. Сорт Виктория и его потомство пришлось
снять с производства и заменить другими сортами. После этого гриб исчез
также внезапно, как и появился, остались лишь его культуры в лаборато¬
риях, из которых получают викторин, ставший интересной моделью для
изучения специфичности во взаимоотношениях разных организмов друг
с другом. С помощью меченого радиоактивным йодом викторина удалось
выделить рецепторный белок, находящийся в мембранах клеток сорта ов¬
са Виктория. Им оказался фермент глициндекарбоксилаза, участвующий
в конечных этапах метаболизма азота в клетках. У человека мутации ге¬
на глициндекарбоксилазы вызывают тяжелое заболевание с летальным
исходом —* гиперглицинемию. По-видимому, к не менее тяжелому за¬
болеванию растений приводит связывание этого фермента викторином.
А гриб-продуцент викторина находят иногда в почве или на корнях ди¬
ких злаков. Предполагают, что он всегда существовал в почве и питался
растительными остатками или корнями ослабленных, стареющих диких
злаков, пока, наконец, не наступил его «звездный час» — появилось рас¬
тение, необычайно чувствительное к его метаболиту викторину.
После истории с ожогом овса токсины, обладающие, подобно викто¬
рину, высокой специфичностью по отношению к видам и сортам опреде¬
ленных растений, были обнаружены и у некоторых других видов грибов,
имеющих крупные темноокрашенные споры. Особенно много шума на¬
делал гриб, поразивший кукурузу.
В США, как известно, кукурузой заняты самые
ео ычные большие в мире площади. В центре страны с востока
болезни: кукуруза u „ .
на запад протянулись широкои полосой фермы, спе¬
циализирующиеся на выращивании кукурузы. Эта полоса получила назва¬
ние кукурузного пояса. Но многие фермы южнее и севернее кукурузного
пояса также выращивают кукурузу. На кукурузе паразитирует несколь¬
ко видов грибов, вызывающих образование пятен на листьях. Эти грибы
были хорошо известны фитопатологам, к ним были созданы устойчивые
сорта, и общие потери от всех этих болезней не превышали 2 %. Так что
причин для беспокойства как будто бы не было.
В послевоенные годы в США, как и во многих других странах, при
выращивании кукурузы использовали эффект гетерозиса, то есть получе¬
§2. Токсины
87
ние высокоурожайных гибридов между линиями, обладающими высокой
комбинационной способностью. Урожай таких межлинейных гибридов
значительно выше, чем урожай обычных сортов. Но на одном растении
кукурузы расположены соцветия как мужских цветков (верхушечные сул¬
таны), так и женских (пазушные початки), Чтобы избежать самоопыления
материнских растений, у них обрезали мужские соцветия. Это легко сде¬
лать, если растет несколько растений на огороде. Но на плантациях, зани¬
мающих тысячи гектаров, такая работа требует огромных затрат средств
и времени. У некоторых растений кукурузы, растущих в Техасе, было об¬
наружено генетическое уродство, названное цитоплазматической мужской
стерильностью (ЦМС), — недоразвитость пыльцы. Их пыльца не могла
оплодотворять собственные початки, поэтому их семена всегда были ги¬
бридные. Стерильность пыльцы была обусловлена мутацией гена, находя¬
щегося в ДНК цитоплазматической органеллы митохондрии. И вот, чтобы
не тратить средства на обрезку мужских соцветий, в сорта кукурузы путем
гибридизации стали вводить гены ЦМС. Были созданы гибриды, у кото¬
рых ядра различных сортов совмещались с цитоплазмой (митохондриями)
техасской кукурузы (Т-цитоплазмой). Использование ЦМС для получения
высокоурожайных гибридов — изящный и экономически очень эффек¬
тивный прием, поэтому в США ежегодно стали выращивать миллиарды
растений, различающихся по ядерным генам, но имеющих одинаковые
митохондрии техасского типа. Такие растения занимали 85 % всех посевов
кукурузного пояса. В середине 60-х годов прошлого века было обнаружено
поражение нескольких линий кукурузы с Т-цитоплазмой заболеванием,
называемым южный гельминтоспориоз, несмотря на наличие у них ядер-
ных генов устойчивости к этой болезни. Гриб, выделенный из пораженных
растений, оказался высокопатогенным именно для кукурузы, имеющей
Т-цитоплазму, и получил название Т-расы (в отличие от обычной О-расы).
Настоящей угрозой стало использование Т-цитоплазмы в южных штатах.
Возбудитель болезни теплолюбив и может накапливаться только в годы
с теплым летом. В 1970 г. болезнь появилась вначале во Флориде и вызвала
там массовое поражение кукурузы, в результате чего накопилось огром¬
ное число спор гриба. Тропический шторм, возникший в Мексиканском
заливе в конце июня, перенес споровое облако через Аппалачские горы
на северо-запад в штаты кукурузного пояса. В конце лета болезнь распро¬
странилась по всем центральным и восточным штатам США. Гриб вызы¬
вал ожог листьев, гниль основания стебля и даже гниль початков. Потери
от болезни составили 100 % на юге страны и 15 % в среднем по стране. По¬
скольку кукуруза занимала огромные площади (25 миллионов гектаров),
недобор урожая, равный 15 %, составил 20 миллионов тонн зерна стоимо¬
стью свыше миллиарда долларов. Американские фитопатологи измерили
потери от болезни в более доступных воображению единицах: если бы
потерянный урожай пошел на корм скоту, а не грибу, то из дополнитель¬
88 Глава 4. Вторичные метаболиты грибов
ного привеса можно было бы приготовить 20 миллиардов 100-граммовых
бифштексов (по несколько кусков мяса на каждого жителя планеты).
Откуда же возникла Т-раса? Оказалось, что ее мицелий встречается
на корнях различных диких злаков, вызывая слабые их повреждения. На
кукурузе с нормальной цитоплазмой Т-раса вытесняется О-расой и не на¬
капливается. Эта ситуация очень напоминает ту, что произошла с посевами
сорта овса Виктория. Было показано, что раса Т выделяет в среду роста
токсин, вызывающий повреждения только клеток, имеющих митохондрии
Т-типа. Этот токсин, названный Т-токсином, связывается с низкомолеку¬
лярным белком, находящимся на внутренней мембране Т-митохондрий,
и отсутствующий в нормальных митохондриях. После соединения с ток¬
сином этот белок полимеризуется (несколько молекул белка соединяются
друг с другом) и образует каналы в мембранах митохондрий, через которые
происходит утечка важных соединений, и работа митохондрий прекраща¬
ется. Ген, контролирующий синтез этого белка, был клонирован и пере¬
несен в другие организмы (кишечную палочку, дрожжи, муху дрозофилу,
табак). Все трансгенные объекты стали чувствительными к Т-токсину.
Кроме высокоспецифичных токсинов многие грибы обладают неспе¬
цифичными токсинами, которые вызывают повреждения клеток прак¬
тически любых растений, в которые попадают. Эти низкомолекулярные
соединения легко мигрируют из зоны заражения по всему растению и,
убивая клетки, подготавливают плацдармы для заселения растения пара¬
зитическим грибом — продуцентом токсина.
2.3. Микотоксины
Микотоксинами, с легкой руки нашего знаменитого миколога А. X. Сар¬
кисова, стали называть вещества микроскопических плесневых грибов,
вызывающие отравления человека и сельскохозяйственных животных. Ча¬
ще всего микотоксины попадают в организмы человека и животных с рас¬
тительной пищей, зараженной грибами, которые образуют эти вещества.
С отравлениями микотоксинами люди познакомились
Токсины о
.nrtnuuL1J очень давно, и, пожалуй, наиболее ранние свидетельства
спорыньи
таких отравлений связаны с заболеваниями эрготизмом,
бушевавшими наряду с чумой в Средние века. Эта болезнь возникает в ре¬
зультате попадания в организм токсинов гриба Клавицепс, вызывающего
заболевание ржи под названием спорынья. Гриб заражает цветки, в кото¬
рых вместо семени образуется плотный, удлиненный, темно-фиолетовый
рожок (склероций), обычно более крупный, чем зерно ржи, и поэтому вы¬
совывающийся наружу (рис. 25 а). Судьба склероциев может быть двоякой.
Если они осыпятся или попадут в партию семенного зерна, то рано или
поздно окажутся в почве, весной прорастут и образуют споры, которые
заразят цветки ржи. Если же они попадут в партию продовольственного
зерна, то будут перемолоты в порошок и, как примесь к муке, станут
составной частью хлеба. И этот путь самый опасный, ибо склероции со-
§2. Токсины
89
Рис. 25. Спорыньевые грибы: а)у б) — спорынья; а) склероции гриба в колосе
ржи; б) прорастание склероция головчатыми стромами, по периферии которых
расположены плодовые тела перитеции с сумками; в), г) — чехловидная болезнь
ежи сборной; в) строма в виде чехла на стебле; г) разрез, стромы через перитеций
с сумками; д) строма паразита насекомых кордицепса воинственного на теле
пораженной гусеницы; на верхней части стромы расположены многочисленные
перитеции (Жизнь растений, т. 2, 1976)
держат химические вещества — алкалоиды, обладающие сильным нервно-
паралитическим действием и вызывающие резкие сокращения мышц —
судороги. Поэтому заболевание получило в народе название «злые корчи».
Другой характерный симптом болезни — сильное жжение внутри всего
тела, за что средневековые врачи дали заболеванию латинское название
sager ignis — святой огонь. Заболевание сопровождается рвотой, болями
в области желудка, галлюцинациями и часто имеет летальный исход.
Две страшные болезни, как Дамоклов меч, постоянно угрожающие
населению средневековой Европы, — чума и отравление спорыньей (эрго¬
тизм). Первое письменное упоминание об эпидемии в Париже, вызванной
отравлением, содержится в хронике Реймского собора за 945 год. Автор
хроники пишет, что избежали смерти лишь немногие, кто посетил святые
места. В 994 году разразилась эпидемия эрготизма в Аквитании вблизи
Лиможа. В тот год в страшных мучениях погибло более 40 тысяч человек.
Эпидемии во Франции следовали одна за другой. За 400 лет с X до XIV ве¬
ка описаны 24 особо сильные эпидемии, сопровождавшиеся гибелью ты¬
сяч людей. Эпидемии породили огромное число пилигримов, отправля¬
ющихся помолиться святым о заступничестве перед страшной болезнью.
90 Глава 4. Вторичные метаболиты грибов
В 1090 году богатый парижанин по имени Гастон со своим сыном Гери¬
ном, заболевшим эрготизмом, забрели в небольшое селение Моте-Бор,
в котором находилась церковь святого Антония. Мощи этого знаменитого
отшельника были перенесены из египетской пустыни, где он провел в оди¬
ночестве большую часть своей столетней жизни и стал еще при жизни ле¬
гендой, в Александрию. Оттуда мощи путешествовали из монастыря в мо¬
настырь по Ближнему Востоку, пока не были перенесены французскими
пилигримами во Францию. Герин и Гастон страстно молили святого Анто¬
ния о помощи. И произошло чудо: Герин поправился, после чего его отец
пожертвовал госпиталю при церкви все состояние, а сам, согласно данно¬
му обету, постригся в монахи. Весть о чудесном выздоровлении распро¬
странилась по Франции и достигла Рима. С 1095 года папа Урбан II (автор
идеи и вдохновитель 1-го крестового похода) уполномочил монахов ордена
святого Антония лечить людей, пораженных эрготизмом, святой Антоний
был объявлен покровителем всех больных и убогих (а впоследствии и сту¬
дентов), а за болезнью на века укрепилось название «Антонов огонь».
Причина болезни выяснилась гораздо позже. В 1582 г. немецкий бо¬
таник и врач Адам Лонизер описал удлиненные твердые образования,
иногда заменяющие зерна в колосьях ржи. Он слышал, что некоторые
женщины для облегчения родов (чтобы усилить сокращения матки) съе¬
дают по три рожка спорыньи, и первый высказал предположение, что
зараженный спорыньей хлеб — причина заболевания Антоновым огнем.
А между тем эпидемии эрготизма продолжали периодически вспыхи¬
вать в разных странах. За 300 лет с1581по 1889 годы было зафиксировано
65 эпидемий: 29 — в Германии, 11 — в России, 10 — в Швеции, 4 — в Ита¬
лии, а также в Швейцарии, Финляндии, Англии, Голландии, Норвегии,
Венгрии. Например, в России во время похода Петра I на Азов в Поволжье
бушевала эпидемия эрготизма. Болезнь не обошла армию: ведь солдаты,
в основном, питались черным хлебом, а лошади — ржаной соломой. На¬
чались массовые заболевания, вызвавшие гибель солдат и падеж лошадей,
что явилось основной причиной неудачи военной кампании. Всего в тот
год в Астрахани умерло 20 тысяч человек. Очень сильная эпидемия про¬
изошла в России в 1832 г. Она охватила 30 различных районов страны,
причем в разных местах смертность от болезни составила от 11 до 66
процентов населения. В связи с широким распространением болезни ме¬
дицинский департамент в Петербурге создал специальную комиссию для
изучения ее причины.
Последнее сообщение об эрготизме в Европе, как и первое, пришло
из Франции. В августе 1951 г. в деревне Понт-сан-Эспри близ Авиньона
около 200 человек внезапно отравились хлебом. Отравление сопровожда¬
лось резкими болями в области желудка, рвотой, похолоданием конечно¬
стей, галлюцинациями. Заболели и многие домашние животные. В газетах
писали, что во Францию вернулся Антонов огонь. О причинах этой эпи¬
демии не было единого мнения. Одни считают, что это было спорыньевое
§2. Токсины
91
отравление, другие — что произошло отравление органическими фосфор¬
содержащими пестицидами, которыми опрыскивали растения для защиты
от вредителей.
Все эти события стали достоянием истории. Сейчас благодаря улуч¬
шению техники сортирования зерна и ежегодному обязательному обсле¬
дованию (апробации) состояния семенных посевов, зараженность их спо¬
рыньей резко снизилась. Но, хотя в Европе опасность отравления зара¬
женным спорыньей хлебом сведена до минимума, в ряде тропических
стран она продолжает существовать. Так, в Восточной Африке, на Абис¬
синском нагорье, кроме ячменя население использует для приготовления
хлеба несколько видов дикорастущих злаков, часто сильно пораженных
спорыньей. Процент тяжелых заболеваний с летальным исходом в этих
местах в отдельные годы бывает очень высоким.
Биологически активные вещества склероциев давно привлекли вни¬
мание врачей и химиков. Из склероциев выделено несколько алкалоидов,
вызывающих сокращение мышц и получивших применение в акушер¬
ской практике (можно только удивляться наблюдательности средневеко¬
вых женщин). Кроме того, из алкалоидов спорыньи получают некоторые
гормональные медицинские препараты. Поэтому в ряде стран созданы
хозяйства, в которых рожь специально заражают спорыньей. Такое хозяй¬
ство имеется и у подмосковного института лекарственных растений. Кро¬
ме того, было показано, что исходным веществом, из которого в клетках
склероциев синтезируются алкалоиды, является также интересное в фар¬
макологическом отношении соединение — лизергиновая кислота, или
знаменитый галлюциноген ЛСД, известный многим наркотик.
В 1960 г. английские газеты пестрели сенсационными
сообщениями о гибели индюшек на многих фермах. Всего (рл т ы ^
за этот год в Англии от острого некроза печени погибло около 100 ООО ин¬
дюшек. Так как причина болезни была неизвестна, ее заболевание назвали
индюшачьей болезнью X. Исследования, проведенные по горячим следам,
показали, что на всех фермах, где погибли индюшки, их кормили арахисо¬
вой мукой, закупленной в Бразилии. Закупленные партии были заражены
грибом желтым аспергиллом и содержали его метаболит, названный афла-
токсином (от начальных букв латинского названия гриба —- Аспергиллюс
флавус). В настоящее время у штаммов желтого аспергилла найдено целое
семейство сходных по структуре и токсичности соединений (афлатоксины
Вь В2, Gj, G2 и др.), что обычно для многих вторичных метаболитов.
Афлатоксины обладают не только остро токсичным действием, но,
даже при попадании в организм в очень низкой концентрации (менее од¬
ного микрограмма на килограмм веса животного), способны вызывать рак
печени. Из афлатоксина Bi под действием ферментов печени образуется
соединение, которое способно вызывать мутации гена, продукт которого
подавляет рост опухоли. В связи с этим афлатоксины не только сами вы¬
зывают рак, но и усиливают онкогенное действие вирусов гепатита типа В.
92 Глава 4. Вторичные метаболиты грибов
Росту и размножению продуцента афлатоксинов Аспергиллюс флавус,
как и других аспергиллов, способствует теплая, влажная погода, поэтому
он вызывает плесневение сельскохозяйственной продукции во влажных
тропиках и субтропиках. Токсичную дозу афлатоксинов можно получить
из зараженных арахиса, кукурузы и других растений, а также из молока,
мяса, яиц сельскохозяйственных животных, которых кормили зараженны¬
ми аспергиллом кормами. Так что горьковатые на вкус арахисовые орешки
надо сразу выплевывать.
Опасность ввоза сельскохозяйственной продукции, содержащей афла-
токсин, заставила разработать аналитические методы, позволяющие вы¬
явить очень низкое его содержание. Во всех пунктах, принимающих
импортную продукцию сельского хозяйства (железнодорожных вокзалах,
морских и аэропортах), созданы лаборатории, анализирующие поступаю¬
щие продукты, и если содержание афлатоксина в них превышает предель¬
но допустимую концентрацию (ПДК), стране-поставщику предъявляются
претензии, а продукт возвращается обратно.
В течение около десяти лет первой четверти
^ р XX века русские крестьяне вместо того, чтобы вы¬
ращивать рожь и пшеницу, стреляли: сначала в немцев, а затем — друг
в друга. Старики и женщины, оставшиеся в деревнях, не успевали за наше
короткое лето справиться с сельскохозяйственными работами, и многие
поля уходили под снег неубранными. Но так как кушать хочется, перези¬
мовавшие зерна собирали и пекли из них хлеб. Евшие такой хлеб люди
часто тяжело заболевали. Симптомы отравления— сильное головокруже¬
ние, боли в области желудки, рвота — были сходными с симптомами алко¬
гольного отравления, поэтому болезнь получила в народе название «пья¬
ного хлеба». Причиной отравления были токсины, производимые грибами
из рода Фузариум. Многие виды этого рода заражают различные растения,
вызывая у них разного рода заболевания. Одни виды заражают подземные
части молодых растений и вызывают корневые гнили и выпадения всхо¬
дов, другие — проникают внутрь зараженных растений и, поселяясь в со¬
судистой системе, препятствуют току воды с питательными веществами,
вследствие чего зараженные растения увядают даже в достаточно влажной
почве. Некоторые виды заражают цветки и развивают свой мицелий в ге¬
неративных органах (фузариозы колоса). В результате болезни колосковые
чешуйки часто приобретают красноватый цвет от пигментов, образуемых
грибом, а зерна становятся щуплыми и тоже красноватыми. Фузариоз
колоса особенно опасен, так как зерна содержат токсины, не разруша¬
ющиеся при хлебопечении. Большинство фузариотоксинов имеют струк¬
турное сходство и общий начальный путь биосинтеза с описанным уже
грибным антибиотиком трихотецином, поэтому их называют трихотеци-
новыми токсинами. Они ингибируют синтез белка в эукариотных клетках,
поэтому высокотоксичны для многих эукариотных организмов, включая
человека и позвоночных животных. Кроме того, трихотециновые токси¬
§2. Токсины
93
ны сходны с половыми гормонами млекопитающих и могут замещать их
в физиологических процессах, вызывая серьезные нарушения течения бе¬
ременности. В связи с этим нельзя не упомянуть о трагической истории,
вызванной внедрением новых сортов кукурузы. Белок семян кукурузы
имеет высокую диетическую ценность, но в нем очень низкое содержа¬
ние незаменимой аминокислоты лизина. А незаменимые аминокислоты
потому и называются незаменимыми, что наш организм их делать не мо¬
жет, они должны поступать с пищей. В 70-е годы прошлого века генетики
получили высоколизиновые мутанты кукурузы, которые стали широко
использоваться при создании новых сортов. Однако оказалось, что лизин
полезен не только нам, но и фузариевым грибам. Новые сорта стали силь¬
но страдать от фузариоза початков, который не только снижал урожай,
но и вызывал накопление в початках трихотециновых токсинов. Кормле¬
ние зараженными початками свиней (а кукуруза в основном используется
на корм скоту) вызывало массовые эпизоотии этих животных. Особенно
страдали беременные самки, у которых происходили выкидыши плода,
эмбриональные уродства и прочие тяжелые симптомы отравлений.
Возбудители фузариоза колоса — теплолюбивые грибы, поэтому бо¬
лезни от них развивались, в основном, в южных районах нашей страны.
При правильной агротехнике и своевременной уборке зерна они не при¬
носят большого вреда. В житнице России, Краснодарском крае, вторая
половина лета была обычно сухой, поэтому фузариоз колоса был редок.
Однако в 60-х годах прошлого века огромные площади в поймах крас¬
нодарских рек были засеяны рисом и покрыты водой. Это стало одной
из причин изменения климата в регионе: дождливые осени стали нормой,
хлеб стал полегать, а семена — плесневеть на корню. Возникли массовые
поражения колосьев фузариозом. В отдельные годы содержание фузарио-
токсинов в зерне превышало ПДК в десятки раз, так что весь урожай при¬
ходилось уничтожать; зерно не годилось даже для переработки на спирт.
Поскольку в районах Поволжья и Среднего Урала часто не хватало заго¬
товленного сена для прокорма скота, представители колхозов и совхозов
закупали солому в Краснодарском крае и тем самым не только развозили
инфекцию в новые районы, но и вызывали массовые отравления скота
фузариотоксинами. Сейчас в связи с глобальным потеплением фузариоз
колоса стал обычным даже в Ленинградской области.
В 30-е годы прошлого столетия на Украине •
получило значительное распространение заболе- тахи 0ТРИ0Т0КСИНЫ
вание лошадей. Сначала поражались ротовые органы, появлялось воспа¬
ление губ, стоматиты, некрозы гортани, риниты, конъюнктивиты. Затем
заболевали внутренние органы, наблюдались легочные кровотечения, ча¬
сто сопровождающиеся гибелью животных. Была отмечена связь заболе¬
ваний лошадей с их питанием мокрым сеном и соломой. А из мокрой
соломы выделили гриб стахиботрис. Этот гриб развивается на субстратах,
содержащих целлюлозу, которую он эффективно перерабатывает своими
94 Глава 4. Вторичные метаболиты грибов
ферментами, но только в условиях высокой влажности — не ниже 90-95 %.
Он образует очень токсичные продукты, трихотециновые токсины, сход¬
ные по строению и механизму действия с токсинами фузариевых грибов.
Поскольку для роста гриба требуется высокая влажность, подсушивание
соломистых кормов — лучшая форма защиты скота от инфекции.
Сейчас стахиботрис волнует в большей степени не ветеринаров, а са¬
нитарных врачей. Поскольку гриб может питаться чистой целлюлозой,
он развивается на обоях и других покрытиях стен во внутренних поме¬
щениях, имеющих высокую влажность воздуха, образуя темно-зеленые
налеты плесени. Его споры и обрывки мицелия, разносятся по жилым
(или рабочим) помещениям и, попадая в рот и дыхательные органы, мо¬
гут вызывать серьезные заболевания, не только вследствие аллергических
реакций на грибной белок, но и вследствие наличия микотоксинов. За¬
болевания людей, вызванные токсинами гриба стахиботрис, получили на¬
звание «синдром больного строения» (sick building syndrome).
* * *
Здесь рассказано лишь о некоторых микотоксинах. На самом деле их
известно огромное количество. Многие из них продуцируются фитопато¬
генными грибами, вызывающими различные болезни растений, например,
спорынью или фузариозы колоса, другие, как описанный выше желтый
аспергилл, вызывают плесневение сельскохозяйственной продукции или
даже изготовленных продуктов питания, таких как варенье, колбаса, сыр
и прочее. Профессор ереванского университета JI. JI. Осипян организовала
у себя на кафедре масштабные исследования заселенности грибами про¬
дуктов сельского хозяйства и наличия в них микотоксинов. Результаты
исследований были поразительными — продукция многих производств
в Армении была признана опасной, и производства были закрыты. Бо¬
лее того, многие лекарства, произведенные из лекарственных растений,
особенно полученных из тропических стран, таких как Индия, оказались
зараженными микотоксинами, так что их прием вместо лечения мог при¬
вести к опасным заболеваниям.
Во многих странах проблемы экологии и защиты окружающий среды
от ксенобиотиков, то есть веществ, созданных человеком, вошла в госу¬
дарственные программы; весьма популярными становятся принципы био¬
логического земледелия, в основе которых лежит отказ от минеральных
удобрений и пестицидов, то есть продуктов химической промышленности
при выращивании растений. К сожалению, борцы за чистоту урожаев сель¬
ского хозяйства плохо знают и биологию, и само сельское хозяйство. Отказ
от химических средств защиты растений от грибов — фунгицидов — может
привести не только к снижению урожая от грибов-паразитов, но и к на¬
коплению в урожае микотоксинов. И если химические средства защиты
растений, прежде чем попасть на рынок, проходят серьезнейшую проверку
на остротоксическое, мутагенное и канцерогенное действие, персистент¬
§2. Токсины
95
ность, миграцию в окружающей среде и прочие тесты, то действие многих
митоксинов не предсказуемо, а некоторые токсины грибов в сотни и ты¬
сячи раз опаснее разрешенных к применению фунгицидов.
Микотоксины очень коварны. В низких дозах, не вызывающих от¬
четливых симптомов отравления, он могут быть причинами повышенной
активности, чувства страха и агрессивности. Микологи, проанализировав
историю средневековой Европы, пришли к выводу, что многим войнам
предшествовали случаи массового отравления спорыньей. То есть токси¬
козы приводили к тому, что люди не могли договориться, найти компро¬
миссные решения, между ними возникали стычки, приводящие к войнам.
Да и недавней перестройке в нашей стране предшествовали несколько лет
массового заражения пшеницы фузариозом колоса. Так что утверждение
авторов гротескной телевизионной передачи, что большевики в 1917 г.
объелись грибами и устроили революцию, не столь уж дурацки бессмыс¬
ленны. Но это все-таки домыслы, не имеющие серьезных доказательств.
А несомненно то, что в низких дозах микотоксины в пищевых продуктах
могут стимулировать онкогенные заболевания. Исследования Всемирной
организации здравоохранения показали, что у населения стран, в раци¬
оне жителей которых преобладают мучные изделия, наиболее высокий
процент раковых заболеваний составляют опухоли пищеварительных ор¬
ганов, что связано как с приготовлением хлеба из зараженного зерна, так
и с плесневением хлеба в процессе хранения. В странах, население кото¬
рых питается в большей степени мясом, рыбой и овощами, преобладают
раковые заболевания иных органов.
2.4. Токсины макромицетов
Народы разных стран можно разделить на микофилов (любителей
собирать и есть дикорастущие грибы) и микофобов, которые боятся со¬
бирать и есть дикорастущие грибы. К микофобам относится, в основном,
население Западной Европы и Северной Америки, а к микофилам — насе¬
ление Восточной Европы и, прежде всего, России. При этом использовать
в пищу грибы, выращенные искусственно, например, шампиньоны, лю¬
бят и те, и другие. Микофобия развивалась у населения постепенно, как
ответ на отравления, вызванные грибными токсинами. В древнем Риме
грибы считались лакомством, деликатесом, недаром один из съедобных
видов рода Аманита (мухоморы) римляне называли цезарским грибом. От¬
менным вкусом обладает и другой представитель этого рода, растущий в
Средней России розовый шампиньон (не самый распространенный крас¬
ный, который ядовит, а съедобный, розовый), и даже, по свидетельству
тех, кто успел свидетельствовать, бледная поганка. Но в этом роде есть
много ядовитых и, в том числе, смертельно ядовитых видов. И использо¬
вание ядовитых грибов, прежде всего бледной поганки (рис. 206, стр. 75),
для решения жизненных задач и задачек, стало излюбленным занятием
дворцовых интриганов.
96 Глава 4. Вторичные метаболиты грибов
Первым известным ученым-токсикологом была
Бледная поганка _
. римлянка Локуста, которая готовила ядовитые до-
в древнем мире
бавки к пище по заказам видных особ. Для отработ¬
ки оптимальных доз яда она проводила опыты на козах. Так что она была
и основательницей экспериментальной медицины, проверяющей новые
лекарства и приемы лечения на животных. Услугами Локусты часто поль¬
зовалась другая римская дама, Агриппина. Сначала, желая выйти замуж
за овдовевшего императора Клавдия, она отравила грибами собственного
мужа. Затем она уговорила императора усыновить и сделать наследником
своего сына Нерона, а самого императора, чтобы он не передумал, также
отправила на тот свет соусом, приготовленным из бледной поганки. После
этого она отравила сына Клавдия от предыдущего брака Британика, двух
внуков императора Августа и капитана императорской гвардии Серенуса.
Последний был микофобом и не ел грибы, поэтому, зная, что капитан ест
из общего котла со своими солдатами, ей пришлось отправить на тот свет
вместе с капитаном 100 его солдат. И все это для того, чтобы расчистить
от конкурентов дорогу на императорский престол своему сыну Нерону,
который отплатил ей черной неблагодарностью: сначала он хотел ее отра¬
вить поганкой, но не смог, так как она приучила свой организм приемом
малых доз яда, тогда он приказал убить свою мать.
Другая интересная римская история, связанная с грибными токсина¬
ми, произошла в Восточном Средиземноморье. После распада империи
Александра Македонского там сформировалось несколько царств, воз¬
главляемых его генералами и их потомками. В Понтийском царстве пра¬
вили потомки македонского полководца Митридата. Один из них, Мит-
ридат VI, опасаясь, что его отравят ядом бледной поганки, стал приучать
к нему свой организм, съедая периодически небольшое, но все увели¬
чивающееся количество гриба с пищей. После того, как римская армия
во главе с Цезарем разгромила его войско и оккупировала царство, бе¬
жавший Митридат решил покончить с собой и принял большую дозу яда,
но оказался нечувствительным к нему. С тех пор возникновение устойчи¬
вости к ядовитым веществам в результате периодического приема низких
доз получило название митридатизма.
Главным токсином бледной поганки оказался
Аманитин ~ токсин
бледной поганки циклический октапептид, названный аманитином.
Октапептид — это соединение, содержащее восемь
аминокислот, циклический означает, что первая и последняя аминокисло¬
та в цепочке соединены друг с другом, образуя цикл. Такие циклические
пептиды синтезируются не на рибосомах матричным синтезом — транс¬
ляцией (на матрице информационной РНК), а с помощью специальных
ферментов — цикл аз. Аманитин подавляет действие одного из важнейших
ферментов — PH К-полимеразы, который участвует в процессе транскрип¬
ции — построения цепочки информационной РНК на матрице ДНК.
Поэтому молекулярные биологи, изучающие процессы транаскрипции
§2. Токсины
97
и трансляции, используют аманитин, как реактив, вмешивающийся в эти
процессы. А если нарушены процессы синтеза белка в клетке, то клетка
гибнет. Поэтому аманитины так токсичны.
Аманитин не только очень ядовит, но и коварен: симптомы отравле¬
ния появляются поздно — через 6-12 часов после приема грибной пищи,
когда часто бывает уже поздно принимать меры для спасения. Снача¬
ла наступает период острого заболевания желудочно-кишечного тракта,
сопровождающийся непрекращающейся мучительной рвотой, спастиче¬
скими болями в желудке, мучительной диареей, головными болями, ги¬
потонией и тахикардией. Через 2-3 суток часто боли проходят, и наступает
период мнимого выздоровления, но спустя сутки наступает период острой
печеночной недостаточности, заканчивающийся при приеме высоких доз
гриба смертью.
Аманитины очень ядовиты. LD50 (доза, убивающая половину особей,
которые приняли яд) для наиболее токсичного альфа-аманитина составля¬
ет 0,1 мг на килограмм веса особи, то есть 6-7 мг для человека со средним
весом. Содержание альфа-аманитина в бледной поганке варьирует от 0,5
до 1,5 мг на грамм веса гриба. Следовательно, при среднем весе шляпки
60 г достаточно съесть одну шляпку, что поставить свою жизнь под угрозу.
Аманитины содержатся не только в бледной поганке и некоторых род¬
ственных ей мухоморах (мухоморе весеннем), но и в других грибах. Гриб-
никам-любителям хорошо известен гриб зонтик пестрый — крупный гриб
на длинной ножке с хорошо сохраняющимся кольцом, которое не при¬
креплено к ножке, и большой зонтиковидной шляпкой, сверху покрытой
коричневыми чешуйками, а снизу имеющей крупные белые пластинки.
Шляпка гриба очень вкусная, используется для приготовления различных
изысканных блюд. Однако более мелкие родственные виды грибов тоже
образуют аманитины, хотя и в меньших, чем у бледной поганки, концен¬
трациях, но достаточных, чтобы вызывать тяжелые симптомы отравления,
вплоть до гибели.
Среди родственников бледной поганки,
грибов из рода Аманита, известно еще не- ней^“Нп“^ %£££
сколько ядовитых видов, содержащих не ама¬
нитины, а другие, менее вредоносные токсины, и поэтому не столь опас¬
ные, как она. У всем известного красного мухомора выделено токсиче¬
ское соединение мускарин — алкалоид, имеющий структурное сходство
с ацетилхолином, переносчиком (медиатором) сигнала в нервных клет¬
ках. Мускарин соединяется с рецепторами на синаптической мембране,
которые так и называются — мускариновыми рецепторами, — и повышает
ее проницаемость для ионов, что приводит к нарушениям в деятельности
нервной системы. Поэтому симптомами отравления наряду с рвотой, диа¬
реей, кишечными коликами являются потливость и нервно-психические
расстройства — эйфория, тревога, мышечная дрожь. Как это часто быва¬
ет, токсическое вещество, нареченное мускарином по названию мухомора
98 Глава 4. Вторичные метаболиты грибов
(Аманита мускария), в самом мухоморе находится в очень низкой кон¬
центрации и не токсично, а в некоторых грибах — видах волоконницы
и говорушки — концентрация мускарина в десятки раз выше, чем в му¬
хоморе, отравление этими грибами гораздо опаснее.
А чем же опасен мухомор? В красном, пантерном и желтом мухоморах
содержатся соединения иботеновая кислота, мусцимол и другие, которые
блокируют рецепторы головного мозга и вызывают повышение двигатель¬
ной активности вплоть до судорог, угнетение эмоционально-психической
деятельности мозга, галлюцинации, а в высоких концентрациях могут ока¬
зывать летальное действие. Именно этими свойствами мухомора обуслов¬
лено его применение в Средние Века норманнскими воинами-берсерками
и шаманами северных народов, которые вводили себя в кататоническое
состояние, сравнимое с безумием.
Но наиболее сильным нейротропным действием обладают псилоцибин
и родственные ему соединения, образующиеся из аминокислоты трипто¬
фана. Их образуют различные виды грибов, токсические вещества которых
вызывают нарушение деятельности центральной и периферической нерв¬
ной системы. Обладая химической структурой, сходной с серотонином,
они взаимодействуют с рецепторами серотонина в головном и спинном
мозге и вызывают возбуждение и острый психоз. Вначале возникают эй¬
фория, беспричинное веселье, нарушение координации, зрительные гал¬
люцинации. Затем беспричинный смех сменяется приступами тревоги и
ужаса, чувства агрессии и суицидального состояния. После глубокого сна
больной выздоравливает. Однако при приеме высокой дозы грибов-галлю-
циногенов возможны необратимые нарушения сердечной деятельности,
инфаркт миокарда, коматозное состояние и смерть.
Очень интересна группа сумчатых грибов
Микотоксины-мутагены:
гиромитрин макромицетов, которые сочетают отменные вку¬
совые данные с наличием опасных для здоровья
токсинов. Это сморчки и строчки (рис. 19), для которых характерно массо¬
вое появление в отдельные годы весной. Настоящие сморчки реабилити¬
рованы, ибо содержат токсические начала в очень низкой концентрации
и в процессе приготовления полностью их теряют. Надо только эти гри¬
бы отваривать и сливать воду, в которой они варились. А вот строчки
очень опасны, ибо содержат токсический продукт гиромитрин в высо¬
кой концентрации. Его летальная доза составляет 20-50 мг на килограмм
веса взрослого человека и 10-30 мг на килограмм веса ребенка. Такая
доза содержится всего в нескольких (2-5) плодовых телах. Гйромитрин
представляет собой простое азотсодержащее соединение, которое в кис¬
лой среде желудочно-кишечного тракта разлагается до образования более
токсического соединения монометилгидразина, а последний в печени пре¬
вращается в источник свободных радикалов нитрозоамин — супермутаген,
который соединяется с нуклеиновыми кислотами, включается в их цепь
и тем самым вызывает различные генетические нарушения.
§2. Токсины
99
Содержание гиромитрина в плодовых телах строчков сильно варьиру¬
ется. Отдельные экземпляры могут содержать менее 100 мг гиромитрина
в килограмме сырых грибов, но описаны экземпляры, содержащие более
1000 мг на килограмм. С чем связано разное накопление гиромитри¬
на в различных популяциях строчков, не известно. Кроме того, есть виды
строчков, никогда не накаливающие гиромитрин в опасной концентрации.
Так что сбор строчков для питания —- это игра в русскую рулетку. Гиромит¬
рин, в отличие от аманитина, — соединение нестойкое. Он имеет более
низкую температуру кипения, чем вода, и частично улетучивается при
варке. Отмечены случаи, когда в маленькой, плохо проветриваемой кухне
люди отравлялись летучей фракцией токсина в процессе варки. Частично
гиромитрин разрушается и в сушеных грибах, однако, вследствие их обез¬
воживания, концентрация гиромитрина после сушки остается опасной.
Смертельно ядовиты и некоторые виды паутинников — грибов, у ко¬
торых частное покрывало, прикрывающее пластинки, расположенные вни¬
зу шляпки, не пленчатое, как у большинства грибов, а имеет вид пау¬
тинок. Красно-оранжевые паутинники смертельно ядовиты. Их токсины,
орелланины, вызывают поражения почек, часто несовместимые с жизнью.
К тому же симптомы заболевания обнаруживаются поздно, часто через
несколько суток после приема пищи, поэтому лечение может опоздать.
Смерть взрослого человека может вызывать прием 40 г свежих грибов.
Некоторые грибы токсичными становятся
Сочетание алкоголя
только в том случае, если ими закусывают вы- и грибов может бь|ть
пивку, что у нас делается очень даже нередко. смертельно опасным!
Таковы некоторые виды навозников, в частно¬
сти чернильный гриб. Этот гриб растет на богатой органическими веще¬
ствами почве и на навозе. Он образует тонкую ножку и изящную шляп¬
ку. Молодые экземпляры, имеющие белые пластинки съедобны, но при
созревании гриб чернеет и под действие мощных ферментов быстро пол¬
ностью разрушается, так что от плодового тела остается только лужица
черной жидкости. В плодовых телах содержится соединение, названное
коприном, в котором циклический углеводород циклопропанол соеди¬
нен с аминокислотой глютамином. В желудочно-кишечном тракте коприн
включается в процесс нейтрализации алкоголя. Этот процесс в организме
осуществляется в два этапа. Сначала этиловый спирт при участии фермен¬
та алкогольдегидрогеназы превращается в ацетальдегид, а затем последний
окисляется другим ферментом — альдегиддегидрогеназой. Аминоцикло-
пропанол подавляет действие последнего фермента, вследствие чего в кро¬
ви накапливается ацетальдегид, который оказывает токсическое действие
на вегетативную нервную систему. В результате этого развивается силь¬
ная головная боль, головокружение, шум в ушах, потливость, усиленное
сердцебиение, а в тяжелых случаях — судороги и коматозное состояние.
В связи с таким специфическим действием коприна его используют в ка¬
честве средства для лечения алкоголизма.
100 Глава 4. Вторичные метаболиты грибов
Разговор о грибных токсинах нельзя завершить без обсуждения ядо¬
витых свойств одного из самых распространенных и часто собираемых
в России грибов — свинушки. Многие горожане, да и сельские жители,
собирают свинушку бельевыми корзинами, солят и едят всю зиму, да еще
и под водку, которая усиливает растворимость грибных токсинов. И вро¬
де бы ничего. Однако мало кто знает, сколько людей попадает в больницы
с симптомами отравления свинушками. К сожалению, природа токси¬
нов свинушки вызывает споры. По-видимому, за отравление этим грибом
ответственны несколько химических соединений. В отличие от вышеопи¬
санных токсинов, и, прежде всего, от токсинов бледной поганки, лишь
небольшая часть съевших свинушки людей попадает в больницы. Неко¬
торые наблюдения позволяют предположить, что симптомы отравления
проявляются, во-первых, у лиц, страдающих дефицитом ключевого фер¬
мента энергетического обмена глюкозо-6-фосфатдегидрогеназы; во-вто-
рых, у лиц, страдающих патологиями в желудочно-кишечном тракте (га¬
стрит, язва желудка и двенадцатиперстной кишки и прочее) и, в-третьих,
у лиц, имеющих нарушения в иммунной системе. Часто первое поедание
свинушек никак не сказывается на состоянии пациентов, которые стано¬
вятся пациентами больниц лишь при повторном приеме этих грибов. При
этом часто развивается гемолитическая анемия, вызванная соединением
грибного антигена с иммуноглобулинами. Образовавшийся комплекс ад¬
сорбировался на эритроцитах и вызывал их гемолиз (потерю гемоглобина).
Так что употреблять в пищу свинушки не следует никоим образом.
Описанные грибы есть нельзя, во всех справочни¬
ках они указаны как высоко или смертельно ядовитые.
Однако есть много грибов, казалось бы, вполне съе¬
добных, но вызывающих проблемы в желудочно-кишечном тракте, как
легкие, быстро проходящие, так и очень острые с тяжелыми последстви¬
ями. Проблемы связаны с наличием в мякоти смолоподобных веществ,
альдегидов и кетонов, раздражающих слизистые оболочки желудочно-ки-
шечного тракта. Многие лица имеют индивидуальную непереносимость
к отдельным соединениям тех или иных видов грибов. Да что говорить, да¬
же распространеннейший съедобный гриб, культивируемый шампиньон
двуспоровый, содержит соединения, продукты разложения которых яв¬
ляются сильными мутагенами (веществами, вызывающими мутации). Так
что грибы, несмотря на их отменные вкусовые достоинства, лучше не есть,
особенно детям, старикам и нездоровым людям.
Грибы опасны не только возможностью по ошибке использования
в пищу ядовитых видов. Есть много сообщений об отравлении видами
бесспорно съедобных грибов, причем число таких сообщений с годами
все увеличивается. Дело в том, что, будучи активными осмотрофами, гри¬
бы поглощают из окружающей среды (почвы, воздуха) не только пита¬
тельные вещества, но и находящиеся там ксенобиотики (минеральные
удобрения, пестициды, тяжелые металлы, радионуклиды), что делает их
§2. Токсины
101
употребление в пищу еще более рискованным. Вот, например, данные ис¬
следования, проведенного в Швейцарии: накопление ртути в высушенных
плодовых телах навозника, собранных в сельской местности, составило
0,51 мг на килограмм, в промышленной зоне — 3,51 мг, а в городе —
до 5,61 мг. Для дикорастущего шампиньона лугового цифры оказались
следующими: 3 мг, около 8 мг и до 30 мг. Эти цифры указывают на две
закономерности. Во-первых, грибы в городах и промышленных зонах на¬
капливают гораздо больше токсических веществ, чем грибы из сельской
местности, а во-вторых, разные виды грибов поглощают ксенобиотики
с разной интенсивностью, что также важно знать.
Наконец, плодовые тела грибов, особенно крупные, стареющие, часто
подвергаются поражению насекомыми и микроскопическими плесневы¬
ми грибами. Личинки грибных мух, «черви», не ядовиты, но некоторым
их присутствие в пище может быть неприятно; к тому же в их ходах могут
поселяться различные бактерии, среди которых есть и токсичные. По¬
этому «червивые» грибы лучше не брать. Но особенно опасны плесневые
грибы микофилы, часто развивающие на плодовых телах макромицетов.
Многие из них способны продуцировать микотоксины, о которых шла
речь в предыдущем разделе. Так что причиной отравления могут быть
не только токсины собранных плодовых тел, но и их паразитов. Поэтому
заплесневевшие грибы собирать нельзя.
В заключение можно рекомендовать читателю одинна¬
дцать правил, соблюдение которых уменьшит риск отравле¬
ния грибами.
1. Не есть грибы. Особенно это относится к детям, пожилым и нездо¬
ровым людям.
2. Если Выгне можете не есть грибы, то покупайте не дикорастущие,
а искусственно выращенные (шампиньоны, вешенки и прочие).
3. Если Вы все-таки хотите есть дикорастущие грибы, то не покупайте
их на рынке, ибо вы не знаете, в каких местах эти грибы собирали.
4. При самостоятельных сборах ни в коем случае не собирайте грибы
в городах, вдоль оживленных автомобильных дорог, около сельско¬
хозяйственных полей, вблизи заводов, свалок и т. п., ибо все вредные
вещества, попадающие из этих мест в воду, почву и воздух, в конечном
итоге окажутся в теле грибов.
5. Ни в коем случае не собирайте незнакомые Вам виды грибов, как бы
аппетитно они ни выглядели и как бы приятно ни пахли.
6. Не собирайте старые, червивые и заплесневевшие грибы.
7. Предпочтительнее собирать грибы, растущие на пнях и мертвых де¬
ревьях (вешенку, летний, осенний и зимний опенки и прочие), чем
грибы, растущие на почве, так как первые накапливают меньше ксе¬
нобиотиков, чем вторые, и, к тому же, содержат полезные для здоро¬
вья вещества, о чем речь будет вестись в дальнейшем.
102 Глава 4. Вторичные метаболиты грибов
8. При сборе гриб лучше не отрезать ножом от мицелия, а выворачивать.
Во-первых, при отрезании образуется участок поврежденной «ткани»,
через который, как через открытые ворота, могут легко попасть пато¬
генные для гриба микроорганизмы, способные повредить грибницу;
во-вторых, отрезав нижний участок ножки, Вы может ошибиться
в определении гриба, например, спутать съедобную зеленую сыроеж¬
ку со смертельно ядовитой бледной поганкой, имеющей в основании
ножки утолщение и рубцы — остатки общего покрывала.
9. Собирать грибы надо в корзинки, а не в целлофановые пакеты, в ко¬
торых плодовые тела быстро задыхаются и поражаются очень ток¬
сичными анаэробными бактериями. К тому же через щели в корзине
высыпаются споры собранных грибов, так что Вы возвращается лесу
часть грибных спор.
10. Придя домой, необходимо сразу приступить к разборке, очистке и мы¬
тью собранных грибов, не давая возможности развиваться плесеням
и бактериозам.
11. Солить грибы необходимо только в открытой посуде во избежание
развития анаэробных бактерий — клостридий, образующих смертель¬
но ядовитые токсины.
§3. Фитогормоны
Фитогормоны регулируют деление и растяжение клеток растений,
развертывание жизненного плана, то есть время перехода из одной фа¬
зы развития в другую, например, у злаков — кущение, выход в трубку,
цветение, плодоношение. Поскольку для грибов-паразитов растений рост
и развитие своего растения-хозяина небезразличны, они в ходе эволю¬
ции приобрели способность продуцировать фитогормоны и регулировать
ими обмен веществ зараженного растения в наиболее благоприятном для
себя направлении. Например, если лист пшеницы заразить ржавчинным
грибом, то в месте заражения образуется ржавое пятно из-за спор гриба,
освободившихся вследствие разрыва эпидермиса. Если после образования
пятна лист отрезать от растения и поместить в микробиологическую чаш¬
ку Петри (или блюдце с влажной фильтровальной бумагой, прикрытое
прозрачным стеклом), то через несколько дней вследствие разрушения
хлоропластов лист пожелтеет, однако вокруг ржавого пятна сохранится
кольцо зеленой ткани, зеленый островок. Следовательно, гриб выделяет
в растение фитогормоны, поддерживающие окружающие его клетки рас¬
тения в молодом, ювенильном состоянии и препятствующие разрушению
хлоропластов. Пусть погибнет все растение, но я должен быть обеспе¬
чен кормом как можно дольше! И это воистину так. Ученые помещали
зараженные ржавчиной растения пшеницы в камеры, заполненные воз¬
духом с углекислым газом, содержащим радиоактивно меченый изотоп
§3. Фитогормоны
103
углерода. В процессе фотосинтеза из углекислого газа образуется глюкоза,
распределение которой в растении можно проследить по распределению
радиоактивной метки. Оказалось, что метка концентрируется вокруг пу¬
стул ржавчины, то есть паразит образует фитогормоны, которые регули¬
руют направление питательных потоков по растению в благоприятном
для него направлении. А многие паразиты выделяют фитогормоны, кото¬
рые вызывают разрастание зараженных участков растения. Миксомицет
Плазмодиофора заражает корни капусты и других крестоцветных расте¬
ний и выделяет фитогормон индолилуксусную кислоту (ауксин), который
вызывает усиленное деление и растяжение клеток. В результате этого
на корнях зараженных растений образуется опухоль, которую называют
кила и в клетках которой находятся плазмодии паразита. Для роста опу¬
холи необходим строительный материал, поэтому в растении происходит
перераспределение потоков питательных веществ в сторону зараженных
паразитом клеток. В результате этого образуется маленький недокормыш-
кочан размером с кулак и крупные наросты на корнях, в которых жиру¬
ет паразит. А грибы-возбудители рака картофеля и пузырчатой головни
кукурузы клетки любого зараженного органа превращают в меристемати-
Рис. 26. Опухоли растений, вызываемые грибами: а) пузырчатая
головня кукурузы (Беляев И. М., Горленко М. В., Дьяков Ю.Т. и др.,
1970); б) кила капусты (С. Бредли, 2003); в) рак картофеля (Жизнь
растений, т. 2, 1976)
104 Глава 4. Вторичные метаболиты грибов
0
Как березу
превратили
в ольху?
ческие (делящиеся клетки образовательной ткани). В результате деления
клеток также образуются крупные опухоли (рис. 26).
Тафриновые грибы интенсивно выделяют фитогор¬
мон индолилуксусную кислоту, избыток которой токси¬
чен для растений. Поэтому зараженные органы и ткани
становятся уродливыми: листья персика — курчавыми,
плоды сливы и черемухи — деревянистыми и морщинистыми (кармаш¬
ки слив), ветви вишни сильно ветвятся тоненькими побегами (ведьмины
метлы) (рис. 27). Кстати, в романе В.Дудинцева «Белые одежды» описан
случай, свидетелем которого был автор этой книжки, в то время студент-
старшекурсник. Знаменитый обскурант и знахарь от науки Т.Д.Лысенко,
отрицая хромосомную теорию наследственности, придумал новую теорию
эволюции: новые виды, якобы, внезапно и сразу зарождаются в недрах
(зародышах, что ли?) старых при изменении типа питания или других
внешних воздействий. Недостаточно образованное в области биологии
население, зачарованное новизной этой теории, всюду искало примеры
превращения одних видов в другие. И некая учительница биологии, про¬
водя со школьниками экскурсию в природу, обнаружила на березе ветку,
покрытую листьями ольхи. Она срезала ветку и принесла ее в Тимиря¬
зевскую сельскохозяйственную академию, где работал академик Лысенко.
Он во время чтения публичной лекции в качестве доказательства сво¬
ей теории показал ветку и пустил ее по рядам слушателей. Ветка дошла
до присутствовавшего на лекции фитопатолога доцента А. Я. Трофимови¬
ча, который узнал в ней поражение листьев березы тафриновым грибом.
Этот гриб вызывает образование глубоких жилок и некоторое утолщение
мякоти, вследствие чего лист березы становится похожим на лист ольхи.
А в Японии рис часто болеет одним из видов рода грибов Фузариум.
Больные растения сильно кустятся, образуя много зеленой массы, но мало
семян. Японские фитопатологии выделили из зараженных растений веще-
Рис. 27. Болезни растений, вызванные тафриновыми грибами:
а) курчавость листьев персика; б) кармашки слив (Дьяков с со-
авт., 2007); в) ведьмины метлы вишни (С. Бредли, 2003)
§4. Пигменты
105
ство, которое вызывает обильное кущение, и назвали его гиббереллином,
поскольку сумчатая стадия паразита относится к роду Гибберелла. Гиббе-
реллин принадлежит к терпеноидным соединениям и по этому признаку
родственен трихотециновым токсинам фузариев, в отличие от них не ток¬
сичен, но обладает сильным фитогормональным действием, в частности
усиливает растяжение клеток и другие процессы. Затем было обнаружено,
что существует большое число сходных молекул, целое семейство гиббе-
реллинов, и что они образуются и в растениях, но, в отличие от гриба-
паразита, очень экономно, только там, где это нужно.
Наконец, некоторые фитотоксины в низких концентрациях оказы¬
вают на растения гормональное действие. Например, токсин возбудителя
рака миндаля фузикокцин влияет на работу устьиц — отверстий на поверх¬
ности листьев. Замыкающие клетки устьиц могут утолщаться вследствие
накачивания в них воды и закрывать отверстие или, наоборот, утоньшаться
вследствие откачки воды и открывать его. Состояние замыкающих кле¬
ток (тургор или плазмолиз) определяется состоянием протонного насоса,
регулирующего транспорт воды и ионов в клетку через мембрану. В эти
процессы и вмешивается фузикокцин.
§4. Пигменты
1]эибы не имеют фотосинтезирующих пигментов — хлорофиллов и
фикобилипротеинов, они не способны использовать энергию возбужде¬
ния световым квантом молекул пигмента для связывания неорганических
соединений в органические. Однако большинство грибов образует разно¬
образные пигменты, которые играют важную роль в их жизни. О некото¬
рых пигментах грибов разговор пойдет ниже.
Меланины — темноокрашенные пигменты. В рассказе меланины
И. Ильфа и Е. Петрова «Директивный бантик» описаны ва¬
риации цветов одежды, которую выпускала текстильная промышленность
СССР: «...черный, черно-серый, серо-черный, черновато-серый, серова¬
то-черный, грифельный, аспидный, наждачный, цвет передельного чугу¬
на, коксовый, торфяной, земляной, мусорный, цвет жмыха и тот цвет,
который в старину называли „сон разбойника**». В зависимости от кон¬
центрации меланинов ткани или органы окрашиваются в описанные вы¬
ше оттенки, от светло-серого до черного. У растений меланин образуется
только в результате повреждения клеток; отмершие участки листа или ино¬
го органа растения, как мы знаем, чернеют, некротизируются. А живые
органы и ткани растений черными не бывают, создание черного тюльпана,
вдохновившего когда-то Александра Дюма, до сих пор остается несбыточ¬
ной мечтой садоводов. У грибов и у животных меланизированные клетки,
ткани и органы обычны, что еще раз подчеркивает близость грибов к жи¬
вотным, а не к растениям.
106 Глава 4. Вторичные метаболиты грибов
Меланины откладываются обычно в клеточных стенках грибов и вы¬
полняют разнообразные функции. Во-первых, они повышают прочность
клеточных стенок к различных повреждающим воздействиям, как биоти¬
ческим (антибиотикам и ферментам конкурирующих микроорганизмов),
так и абиотическим, таким как очень низкие или очень, высокие темпе¬
ратуры. Поэтому покоящиеся структуры грибов — хламидоспоры и скле¬
роции — обычно темно-окрашены, меланизированы.
Во-вторых, меланины защищают клетки от радиации, как от солнеч¬
ной, так и от излучения радионуклидов. Хорошо известно, что у людей
европеоидной расы меланин откладывается в клетках кожи после пребы¬
вания на солнце, а у представителей негроидной расы, как постоянных
жителей тропиков и субтропиков, отложение в коже меланина закрепле¬
но генетически. То же самое наблюдается и у грибов. Мицелий многих
видов грибов, живущих в почве, не меланизирован, но их конидиеносцы
и конидии окрашены в темные цвета, так как образуются на поверхности
почвы и подвержены воздействию солнечной радиации. Среди грибов,
характерных для высокогорья, очень высок процент видов, у которых ме¬
ланизированы не только спороносцы и споры, но и мицелий, так как
в горах воздействие солнечной радиации более сильное, чем на равнин¬
ных местах. Украинский миколог Н. Н. Жданова проводила исследова¬
ние грибов из почв, загрязненных радиоактивными соединениями в зоне
Чернобыльской АЭС, и показала, что там преобладают виды, имеющие
меланизированные мицелии. Ультрафиолетовые и ионизирующие лучи
вызывают различные нарушения в клетках, в том числе окислительные
процессы, в результате которых образуются активные формы кислорода,
такие как перекись водорода, ОН-радикал, синглетный (возбужденный)
кислород. Все они — сильнейшие окислители, способные вызвать повре¬
ждения основных биополимеров клетки — белков и нуклеиновых кислот.
Меланины выступают как молекулярные ловушки, которые гасят актив¬
ные формы кислорода и защищают клетки от их повреждающего действия.
В-третьих, меланины очень нужны фитопатогенным грибам, мицелий
которых внедряется в растение не через ранки или устьица, а путем давле¬
ния гифы на неповрежденную поверхность, давления, приводящего к об¬
разованию разрывов в эпидермисе. Проросток споры у этих грибов закан¬
чивается расширенной нашлепкой — апрессорием, который плотно при¬
клеивается с помощью гидрофобных белков гидрофобинов к поверхности
листа. Наружная клеточная стенка апрессория толстая и темноокрашенная
вследствие отложения в ней меланинов, а участок клеточной стенки, при¬
крепленный к листу, — двухслойный. Наружный ее слой тонкий, а внут¬
ренний плотный, меланизированный. Меланин играет роль молекулярной
ловушки, поглощающей растворы из окружающей среды, но не выпуска¬
ющей их из клетки. В результате этого внутри апрессория развивается вы¬
сокое тургорное давление, которое приводит к разрыву наиболее тонкой
наружной оболочки подошвы и пробиванию клеточных покровов прорас¬
§4. Пигменты
107
тающей вниз инфекционной гифой, которая покрыта внутренним слоем
оболочки. Например, апрессорий возбудителя ожога листьев риса, гри¬
ба Пирикулярия, развивает давление, равное восьми милипаскалям; для
создания такого давления необходимо, чтобы концентрация растворов
в апрессории превышала 3 моля. Мутанты, у которых не образуются фер¬
менты, участвующие в синтезе меланина, имеют светлоокрашенные гифы,
растущие по поверхности листа, но не способные пробить его покровы.
Эти широко распространенные соединения окра¬
шивают клетки в различные оттенки оранжевого цвета. р ™ ды
В детской песенке про «оранжевое небо, оранжевое море...», по-види-
мому, все, что окружало ребенка, содержало каротиноиды. Впервые они
были выделены из корнеплодов моркови и получили от нее свое название
(видовое название моркови по-латыни — карота). Их молекулы представ¬
ляют собой линейные углеводородные цепочки, замкнутые по краям ше¬
стичленными циклами. Каротиноиды, которые состоят только из углерода
и водорода, называются каротинами, а если в их молекулах присутству¬
ет и кислород, они называются ксантинами или ксантофилами. Таким
образом, ксантины — окисленные формы каротиноидов, а каротины —
восстановленные формы. Известны также ациклические каротиноиды,
(не имеющие циклических участков по концам молекулы). Их называют
ликопинами, так как они накапливаются в плодах томата (родовое на¬
звание томата — Ликоперсикон). Каротиноиды синтезируются в клетках
растений и грибов, а животные получают их с пищей. В клетках грибов
они накапливаются в мембранных пузырьках сферосомах и выполняют
не менее разнообразные функции, чем меланины.
Каротиноиды, как и меланины, защищают клетки от фотодинамиче-
ского действия активных форм кислорода, образующихся на свету, то есть
действуют как активные антиоксиданты. Кроме того, каротиноиды са¬
ми являются светособирающими пигментами, их молекулы переходят под
действием света в возбужденное состояние и, тем самым, влияют на мор¬
фогенез. В главе 3 были описаны механизмы изгибания в направлении
света спорангиеносцев мукоровых грибов, живущих в навозе травоядных
животных. Такое изгибание вызвано накоплением в спорангиеносцах бе¬
та-каротина. У мукоровых грибов производные каротиноидов являются
половыми феромонами, регулирующими протекание полового процесса
между «плюс» и «минус» штаммами.
Каротиноиды широко используются в медицине как предшествен¬
ники витамина А, антиоксиданты, противораковые соединения. Грибы,
как активные продуценты каротиноидов, используются в микробиологи¬
ческой промышленности многих стран.
Глава 5
Грибы и растения
*
Грибы и растения — это две огромные группы не¬
родственных организмов, неразрывно связанных друг с
другом. Ферментативный аппарат грибов настроен на
разложение полисахаридов — строительного материа¬
ла и запасных продуктов растений, поэтому не только
паразиты, но даже и сапротрофные грибы (мертвояды)
питаются погибшими растениями, оставляя трупы жи¬
вотных бактериям. Большая группа грибов питается на¬
возом, но, опять же, травоядных животных, навоз ко¬
торых содержит непереваренные растительные остатки.
В основном в разложении мертвой древесины участвуют
грибы.
Связь грибов с растениями сложилась, по-видимому,
на самых ранних этапах эволюции партнеров. Наиболее
примитивные грибы паразитируют на наиболее прими¬
тивных растениях — водорослях. На остатках древней
водоросли Палеонителла, жившей более 400 млн лет на¬
зад, найдены хитридиевые грибы, паразиты и симбион¬
ты этой водоросли. А если учесть, что родственники
Палеонителлы, харовые водоросли, — прямые предше¬
ственники наземных растений, можно с уверенностью
сказать, что грибы вышли на сушу под покровом тканей
вышедших на сушу водорослей, как их паразиты и сим¬
бионты.
Девонские отпечатки первых наземных растений
свидетельствуют о наличии в их тканях грибов, по¬
добных современным грибам арбускулярной микоризы.
Молекулярные данные также свидетельствуют о древно¬
сти этих грибов. Поэтому есть серьезные основания по¬
лагать, что харовые водоросли вышли из мелководных
озер на сушу с грибным мицелием внутри тканей. Если
учесть, что у вышедших водорослей не было корневой
системы, а на суше в те времена не было почвы, можно
представить себе, что выходящие из основания тела во¬
доросли грибные гифы принимали активное участие как
§ 1. Грибы-симбионты. Микоризные грибы 109
в прикреплении к субстрату, так и в питании водоросли. В дальнейшем,
с возникновением и развитием корневой системы у наземных растений,
увеличением их видового разнообразия и возникновением почвы проис¬
ходило одновременно увеличение как видового, так и физиологического
разнообразия грибов. Некоторые грибы ушли в почву и стали сапротрофа-
ми (мертвоедами), питающимися растительными остатками и играющими
важную роль в создании почвенного плодородия. Другие, наоборот, ста¬
ли паразитами, вызывающими болезни растений. Поскольку первичными
наземными грибами, как полагают (хотя точно не знает никто), были
симбионты, с них мы и начнем.
§1. Грибы-симбионты. Микоризные грибы
Микориза (в переводе с латыни — грибокорень) — это форма расти-
тельно-грибного симбиоза, при котором мицелий гриба частично погру¬
жен в почву и частично находится в ткани корня растения. Такое строение
делает гриб посредником между корнями растений и почвой.
Древнейшей формой микоризы была арбускулярная -
микориза, вызываемая гломусовыми грибами (очень ин- р
■ - микориза
тересна эволюция взглядов микологов на эти грибы: вна¬
чале это был один род среди зигомицетов, потом его выделили в отдельное
семейство, затем их статус повысили до порядка, класса и, наконец, для
-них создали новый отдел, маленький по числу видов, но важный в эволю¬
ционном отношении). Покоящиеся споры этих грибов находятся в почве.
Под влиянием химических выделений корней растений, они прорастают,
и гифы растут в направлении корня, заражают растение и распростра¬
няются внутри корня между клетками (такой тип микоризы называют
эндотрофной, то есть питающейся внутри растения). Внедряясь в клетки,
они образуют там питательные органы гаустории, имеющие вид сильно
разветвленных кустиков или деревец, почему этот тип микоризы и назван
арбускулярной (по латыни арбускула — деревце). Арбускула существует
в отдельной клетке всего несколько дней, после чего переваривается клет¬
кой (дружба — дружбой, но если есть возможность, почему бы не пола¬
комиться другом), но вскоре образуется новая. Арбускулярную микоризу
вызывает небольшое число видов грибов (около 150), но она встречается
у огромного числа видов растений (до 300 тысяч) из разных таксонов —
от мхов до цветковых. Следовательно, гломусовые грибы малоразборчивы
в выборе корня, который подходит для заражения, хотя, конечно, есть рас¬
тения, которые они микоризуют с большим или меньшим удовольствием.
Ответ на вопрос, что получает микоризный гриб
от растения, не требует большого мыслительного на- Взаимовыгодное
пряжения — конечно, это продукты фотосинтеза, сотрудничество
прежде всего сахара. А что получает растение от та¬
кого сожительства? Оказывается, очень много.
Рис. 28. Схема, показывающая, как мицелий микоризного гриба
увеличивает площадь корневого питания зараженного растения
1. За счет выходящих из корней гиф гриба увеличивается всасывающая
поверхность и площадь питания (рис. 28). Прежде всего, улучшилось
корневое питание, поскольку увеличился объем почвы, охватывае¬
мой всасывающей поверхностью. Радиус грибной гифы на два поряд¬
ка меньше радиуса корневого волоска, поэтому поверхность корня
на единицу объема (или веса) в 100 раз меньше поверхности гифы,
т. е. для продукции одинаковой всасывающей зоны растению требу¬
ется в 100 раз больше материала, чем грибу. Кроме того, почва вокруг
корня обеднена питательными веществами вследствие всасывающей
активности корня; а с помощью грибных гиф удается преодолеть эту
обедненную зону.
2. Гриб переводит нерастворимые, труднодоступные соединения фос¬
фора в усвояемую для растений форму и снабжает ими растение.
Микориза также важна для фосфорного питания растений, как клу¬
беньковые бактерии — для азотного.
3. Растение использует биологически активные вещества, продуцируе¬
мые грибом, например, некоторые витамины.
4. Микоризованные корни имеют повышенную устойчивость к потен¬
циальным почвенным патогенам. Присутствие в тканях корня чужого
организма — гриба, вызывает ответную реакцию и синтез защитных
веществ. Микоризные грибы выработали толерантность к этим ве¬
ществам, которые поэтому слабо влияют на их жизнеспособность,
хотя и мешают слишком глубокому проникновению, а вот на нахо¬
дящихся в почве паразитов растений эти вещества могут оказывать
губительное воздействие. Таким образом, микоризные грибы стиму¬
лируют образование в корнях растений «минных полей», препятству¬
ющих заражению растений патогенными микроорганизмами. В част¬
ности микоризные грибы индуцируют у растений синтез защитных
110 Глава 5. Грибы и растения
§ 1. Грибы-симбионты. Микоризные грибы
111
фенолов — флавоноидов. Не исключено, что грибы не только акти¬
визируют фенольный метаболизм в зараженных растениях, но и пе¬
редали в растительный геном с помощью горизонтального переноса
гены важнейших ферментов, необходимых для синтеза фенолов.
5. Микоризация «научила» растения воспринимать в качестве симбион¬
тов другие почвообитающие микроорганизмы (например, клубенько¬
вые бактерии). Микоризные симбиозы гораздо более древние, чем
симбиозы растений с бактериями. Под влиянием арбускулярной ми¬
коризы у растений выработалась система рецепции аминосахаров, с
помощью которой клубеньковые бактерии прикрепляются к корню.
Генетики выяснили, что некоторые гены растений, регулирующие их
взаимоотношения с грибами и бактериями, общие.
В дальнейшем растения все больше и больше приспосабливались
к жизни вне воды, изменялась их численность, увеличивалось число жи¬
вущих вне воды видов, изменялись условия их обитания на земле, в част¬
ности появилась новая отсутствовавшая ранее субстанция — почва. Все
это наложило влияние на судьбу грибов и на их роль в жизни наземных
биоценозов. Произошли следующие события:
1. Уход от растений в сформированную к этому времени почву, переход
к питанию мертвым органическим субстратом (к сапротрофии) и
приобретение важных функций в почвообразовательных процессах.
2. Вспышка видообразования и экологическая иррадиация (расселение
в новых местностях, приобретение новых свойств и новых форм вза¬
имодействия с растениями).
3. Возникновение в каменноугольном периоде (карбоне), когда поверх¬
ность многих областей земного шара была покрыта лесами, состоящи¬
ми из древовидных папоротникообразных и голосеменных растений,
древоразрушающих (ксилотрофных) трутовых грибов. Образование
лигнинов растениями и лигнин-разрушающих ферментов грибами.
4. Паразитизм на растениях и роль грибных паразитов в формировании
сложных фитоценозов.
5. Образование новых типов микориз и их роль в жизни растений,
в частности в захвате растениями экстремальных мест обитания (вы¬
сокогорных, высокоширотных).
Подробное рассмотрение этих пунктов будет предметом дальнейшего
изложения материалов данной главы, а начнем мы с других типов микориз,
чтобы продолжить знакомство с этой важнейшей экологической группой
грибов.
В отличие от арбускулярной микоризы, эта ^
„ Эндотрофная микориза (0
встречается только у представителен одного по- вереско;ь^ (эрикоидная
рядка растений — вересковых, к которым отно- микориза)
сятся азалии, рододендроны, вереск, черника,
112
Глава 5. Грибы и растения
брусника, голубика, клюква и другие, хорошо известные кустарники и ку¬
старнички. Их корни заражают сумчатые грибы, мицелий которых зара¬
жает корни и образует там микоризные ассоциации. Этот тип микоризы
оказал огромное влияние на распространение вересковых растений. Бла¬
годаря микоризе они смогли поселяться в крайне неблагоприятных для
жизни условиях — в высоких широтах (в тундре) и больших высотах
(в высокогорье), с очень бедными, мелко залегающими почвами, холод¬
ным климатом, но где мало конкурентов: кому же захочется жить в таких
условиях? А благодаря микоризе вересковые растения могут мириться
с трудностями жизни, вернее, преодолевать их.
Это — один из самых интересных типов мико-
микориза^ ризы. Он вызывается видами базидиальных грибов,
но половая стадия у них встречается редко, обычно
формируется лишь мицелий, даже без бесполого спороношения (стериль¬
ная грибница). Гриб образует в клетках корня мицелиальные клубочки.
В чем уникальность этого типа микоризы? Дело в том, что образующиеся
при прорастании семян проростки орхидных растений не имеют хлоро¬
филла и не могут самостоятельно фотосинтезировать. Поэтому они полу¬
чают углеродное питание (сахара) от мицелия симбионта. То есть, не гриб
паразитирует на растении и получает от него продукты фотосинтеза, а
наоборот, растение паразитирует на грибе, получая от него эти продукты.
Но откуда же берет их гриб, не обладающий способностью к фотосинтезу
по определению? Оказывается, гриб ведет себя подобно слуге двух господ
из комедии Гольдони. Помимо орхидных, у которых гриб формирует эндо¬
трофную микоризу, его мицелий, выходящий из корней, заражает и другие,
не орхидные растения, в которых ведет себя менее интеллигентно, как не-
кротрофный паразит. Извлекаемые из этого растения углеводы мицелий
перекачивает в клубеньки орхидеи и делится награбленным с ростками
второго хозяина — орхидеи. В дальнейшем у растений орхидей образуется
в клетках хлорофилл, они приступают к самостоятельному фотосинтезу
и, в знак благодарности, подкармливают сахарами симбиотический гриб.
Но некоторым видам так понравилось жить на дармовщину, что они пол¬
ностью потеряли способность к фотосинтезу (бесхлорофилльные расте¬
ния) и всю жизнь живут за счет находящегося в корнях гриба, которому
приходится трудиться в поте лица, чтобы прокормить ленивого обжо¬
ру; а он может достигать немалых размеров (один вид бесхлорофилльной
тропической лианы, растущий в Юго-Восточной Азии и Северной Ав¬
стралии, образует стебли длиной до 40 м). В их защиту надо отметить, что
в тропических лесах, где распространено большинство бесхлорофилль-
ных орхидей, к их листьям проникает очень мало света, и интенсивность
фотосинтеза значительно ограничена, так что пришлось искать новые
источники питания. Бесхлорофилльные орхидеи живут не только в тро¬
пиках. Под густыми кронами ели в лесах умеренной зоны можно най¬
ти бесхлорофилльную орхидею, гнездовку, названную так из-за своего
§ 1. Грибы-симбионты. Микоризные грибы 113
корневища, обросшего множеством переплетающихся толстых корешков.
Возможно, разрастание подземных частей гнездовки — зародышевой лу¬
ковицы, корневища — результат реакции на грибное заражение. Извест¬
но много грибных болезней, возбудители которых выделяют в заражен¬
ное растение фитогормоны и вызывают разрастание зараженных тканей
(см. главу 4). Фактически, бесхлорофилльные орхидеи опосредовано (че¬
рез грибной мицелий) паразитируют на других растениях.
Этот тип микоризы получил свое название (наружное
ч * Эктотрофная
питание) вследствие того, что гриб оплетает корень рас- jT'f
„ „ микориза
тения, образуя мицелиальныи чехол, а основной мицелии
находится в почве, удаляясь от зараженного растения иногда на много
метров. Взаимодействие с тканью корня осуществляется гифами, внедря¬
ющимися в корень (сеть Гартинга), но неспособными не только распро¬
страняться в глубокие слои корня, но и развиваться внутриклеточно, как
у разных типов эндотрофной микоризы. В связи с образованием мицели-
ального чехла изменяется морфология корней, их кончики дихотомически
ветвятся, корневые волоски не об¬
разуются, так что почвенное пита¬
ние осуществляется исключитель¬
но грибными гифами. Эктотрофная
микориза формируется примерно у
6 тысяч видов растений, главным об¬
разом, древесных и кустарниковых,
и образует ее также около 6 тысяч
видов грибов-макромицетов, как
базидиальных (мухоморы, сыроеж¬
ки, подосиновики, подберезовики,
маслята и прочие), так и сумча¬
тых (сморчки, трюфели). Вне свя¬
зи с корнями растений (на искус¬
ственных средах) эти грибы могут
расти в виде стерильного мицелия,
но не могут образовывать плодо¬
вых тел, поэтому белые грибы нель¬
зя выращивать, как шампиньоны,
на грядках.
Эктотрофная микориза, поми¬
мо описанных ранее полезных для
растения качеств, имеет некоторые
дополнительные. Во-первых, она
лучше, чем эндотрофные микоризы
защищает корни от внедрения обитающих в почве паразитических микро¬
организмов. Кроме индукции защитных веществ в микоризованных кор¬
нях, имеется механическая защита в виде мицелиального чехла, окружа¬
Рис. 29. Соединение корневой системы
двух сеянцев сосны через слияния мице-
лиев микоризных грибов, оплетающих их
корни (Smith S. Е., Read D. J., 2008)
114
Глава 5. Грибы и растения
ющего корень, и химическая защита в виде антибиотических веществ, об¬
разуемых грибами-микоризообразователями. Во-вторых, распространяясь
в почве на большое расстояние, мицелий может формировать микоризу
с несколькими рядом стоящими деревьями, или же связь между этими
деревьями осуществляется через анастомозы гиф двух микоризообразо-
вателей (рис. 29). При этом возможно перетекание питательных веществ
из корня одного растения в корень другого. Этот факт доказан экспери¬
ментально с помощью меченых атомов. Таким способом осуществляется
подкормка не окрепших еще всходов, растущих рядом с материнским
растением, а весь лес, благодаря огромной сети мицелия, объединяется
в один огромный организм.
§2. Грибы-паразиты
Паразиты человека, его домашних животных и культивируемых им
растений — это почти всегда плохо. Но паразиты в природе — это не всегда
плохо для отдельного индивидуума, часто хорошо для популяции и всегда
хорошо для биоценоза или экосистемы. Начнем с верхнего уровня —
с экосистем, а затем через популяции опустимся до отдельных организмов.
Еще в середине прошлого века известный зоолог В. Н. Беклемишев
опубликовал статью под интригующим названием «Возбудители болезней
как члены биоценозов». В этой статье он доказывал, что паразиты — это
не напасть Божья, данная нам за грехи, а чрезвычайно важный компо¬
нент экосистем, без которого экосистемы не могут сохранять свою ста¬
бильность. Основными причинами стабилизации биоценозов паразитами,
по Беклемишеву, являются следующие:
1. Паразиты регулируют численность нормальных сочленов биоцено¬
зов. Тем самым осуществляется регуляция количественного состава
биоценоза. Таким способом действуют обычно летальные паразиты,
вызывающие гибель пораженных особей.
2. Они защищают сложившийся биоценоз от вторжения чуждых элемен¬
тов, осуществляя регуляцию качественного состава биоценоза. Таким
способом чаще действуют возбудители хронических болезней.
Давайте рассмотрим эти действия паразитов по порядку,
р В 30-х годах прошлого века великий советский
численности эколог Георгий Францевич Гаузе создал основопола-
членов биоценозов гающую для экологии теорию экологической ниши.
Гаузе выращивал пары видов дрожжей и инфузорий
по отдельности и совместно в пробирках, содержащих питательный суб¬
страт, и изучал кривые роста их численности. Он показал, что если два
вида имеют одинаковые пищевые потребности, то они не могут существо¬
вать вместе неопределенно долгое время; всегда, с большей или меньшей
скоростью один вид вытесняет другой. Отсюда был сделан вывод о том,
§2. Грибы-паразиты
115
что два вида организмов не могут длительно занимать одну экологическую
нишу. Это вывод получил название Закона или правила Гаузе.
А теперь вернемся к нашим растениям. Каждому известно, сколь
богаты разнообразием видов растений такие растительные сообщества
(фитоценозы), как смешанный лес и разнотравный луг. Даже вскопанные
грядки, на которых посажены овощные или цветочные культуры, если их
не пропалывать, вскоре целиком покроются всходами разнообразных сор¬
ных растений. Конечно, у одних видов корневая система залегает глубоко,
а у других — распространяется в поверхностных слоях почвы, одним тре¬
буется сильное солнечное освещение, и оли тянутся вверх, к свету, а дру¬
гие, теневыносливые, находятся внизу. Но есть много видов, имеющих
сходные требования к условиям жизни, занимающих одну экологическую
нишу. Почему же более быстро растущие или образующие больше семян
виды не вытеснят менее удачливых? Почему не работает правило Гаузе?
Потому что в сложных экосистемах весьма эффективно работает так на¬
зываемый частотно-зависимый отбор, работающий против тех генотипов,
которые накапливаются в сообществах с высокой частотой. Этот тип от¬
бора экологи поэтически назвали «Правилом белой королевы» из сказок
Кэрролла про Алису. Белая (в английской интерпретации шахматной иг¬
ры — красная) королева все время бежала вперед, но оставалась на одном
месте. Так кто же осуществляет частотно-зависимый отбор в фитоцено¬
зах? Конечно, паразиты. В каких местах чаще всего возникают эпидемии
гриппа? В больших городах с высокой концентрацией населения. В на¬
битом людьми в час пик транспорте любой чих больного человека может
заразить многих окружающих его пассажиров. В сельской местности с раз¬
реженным населением столь массового заражения не происходит. Те же
правила работают и в растительных сообществах. Чем выше концентрация
какого-либо вида растения, тем выше вероятность его заражения парази¬
тическим грибом, а поскольку, согласно Беклемишеву, паразиты, которые
регулируют численность сочленов биоценоза, имеют летальный эффект
для своих хозяев, массовое заражение приводит к массовой гибели. Это
вызывает снижение численности и разреживание популяции вида, кото¬
рый в отсутствие паразитов имел бы наибольшую конкурентоспособность
и вытеснял бы другие виды из фитоценоза. Поскольку свято место пусто
не бывает, разреженные места моментально занимаются другим, менее
конкурентоспособным, но устойчивым к данной болезни видом. Но если
это приведет к неумеренному накоплению другого вида, то его паразит
не позволит ему этого, и так далее. Таким образом, паразиты — основные
факторы частотно-зависимого отбора. А поскольку большинство парази¬
тов растений — это грибы, они явились важнейшим фактором, создавшим
богатейшие растительные сообщества.
Вот несколько примеров того, как это происходит.
В наших лесах часто встречается цветковое растение ветреница лес¬
ная. Одно растение ветреницы производит огромное количество семян.
116
Глава 5. Гоибы и растения
Однако более 20 % ее проростков гибнут от корневой гнили, вызываемой
почвообитающим грибом до появления всходов.
В производстве кормов для сельскохозяйственных животных часто
используют посевы травосмесей, включающих бобовое растение (для по¬
вышения содержания белка в корме) и злаковое, содержащее много клет¬
чатки, необходимой для пищеварения. Изучение конкурентоспособности
компонентов кормосмеси дает много фактов для интересующей нас про¬
блемы. Например, в смести, состоящей из клевера (бобовое) и райграса
(злаковое) клевер вытесняет райграс вследствие поражения последнего
корончатой ржавчиной. После опрыскивания фунгицидом, убивающим
ржавчину, райграс начинает вытеснять клевер.
Прекрасный пример того, как разреживают популяции восприимчи¬
вых растений их паразиты, являет история растения хондриллы в Австра¬
лии. Хондрилла — сорняк с желтыми соцветиями, похожими на соцветия
нашего одуванчика. Она распространена на полях в Средиземноморской
зоне, но не является злостным сорняком, так как концентрация ее побегов
невелика (никогда не встречается более 20 побегов на квадратном метре).
Случайно попав в Австралию, хондрилла размножилась там с невероятной
скоростью и стала зловреднейшим сорняком; ее концентрация достигла
200 побегов на квадратном метре. Австралийцы имеют богатый опыт борь¬
бы с незваными пришельцами с помощью их паразитов. Когда привезен¬
ные в Австралию кролики размножились так, что съели почти всю траву,
был привезен вирус миксомы кроликов, значительно снизивший их чис¬
ленность. Также поступили и с кактусом Опунция, занявшим в Австралии
многие поля; привезли из Аргентины жука — вредителя опунции. Этот
опыт вспомнили и после распространения хондриллы. На юге Франции
нашли гриб, вызывающий болезнь хондриллы — ржавчину. Удачным ока¬
залось то, что этот гриб никакие растения, кроме хондриллы, не поражает.
Его споры привезли в Австралию и заразили ими сорняк. И что интересно:
ржавчина хондриллу не уничтожила, но снизила ее численность до 20 по¬
бегов на квадратном метре, то есть, как и в Южной Европе.
Таким образом, возбудители болезней способствуют снижению агре¬
гации отдельных видов растений в фитоценозах, или (что то же самое)
увеличению дисперсности ценоза. Для того, чтобы попасть на восприим¬
чивый вид растения-хозяина, грибы в таких фитоценозах теряют значи¬
тельную часть своих спор, оседающих на поверхность устойчивых видов
растений. Поэтому чем выше видовое разнообразие фитоценоза, тем сла¬
бее поражаются болезнями его члены.
Более того, грибные паразиты способствуют
распределение^пород равномерному распределению деревьев одной по-
деревьев в лесу роды в лесах. Ботаников давно интересовало,
почему это происходит? Ведь плоды, особенно
такие тяжелые, как желуди или костянки, падают, в основном, вблизи
материнского растения, а всходы чаще обнаруживаются на некотором
§2. Грибы-паразиты
117
расстоянии. Канадские ученые провели исследования со всходами дикой
вишни. Они брали образцы почвы под старой вишней и вдали от нее,
и проращивали в этих почвах семена. Если почва была предварительно
простерилизована, то семена прорастали во всех образцах почвы одинако¬
во хорошо, но в нестерильных условиях семена хорошо прорастали только
в тех образцах почвы, которые были взяты вдали от материнского дерева,
а в почвах, взятых под деревом, проростки семян погибали. Причиной ги¬
бели проростков были почвообитающие грибы, которые взрослому дереву
не могли нанести ущерб, но вызывали гниение молодых проростков. По¬
чему такие грибы накапливались под взрослыми деревьями, тоже понятно:
большинство плодов вишни падает непосредственно под деревом (вдали
от дерева распространяются только плоды, съеденные птицами), поэто¬
му патогенным грибам всегда есть, чем полакомиться.
Спустимся на один этаж ниже и рассмотрим влия-
Эндемичные
ние паразитов на одновидовые популяции. Оказывается, болезни
грибные паразиты способствуют не только разнообразию
фитоценозов, но и генетическому разнообразию популяций отдельных
видов в фитоценозах. Существует такое понятие, как генетический поли¬
морфизм, то есть наличие в популяции двух или более фенотипов в течение
длительного времени. Частоты отдельных фенотипов могут меняться в от¬
вет на изменяющиеся условия жизни. Примером полиморфизма может
служить индустриальный меланизм бабочки березовой пяденицы. В по¬
пуляциях этой бабочки встречаются экземпляры с двумя типами окраски
крыльев: белой с пестринами и черной. Бабочка любит отдыхать на стволах
березы, покрытых лишайником гипогимнией, и окраска надкрылий свет¬
лой формы настолько похожа на поверхность лишайника, что сливается
с ним. Чернокрылая форма, наоборот, очень заметна, и поэтому склевыва¬
ется птицами. В связи с этим белокрылая форма преобладает в популяциях
пяденицы. Но в промышленных зонах, во-первых, исчезают лишайники,
не выдерживающие повышенной концентрации вредных веществ в воз¬
духе, а во-вторых, стволы самих берез покрываются копотью. И в таких
условиях белокрылая форма становится более заметной и склевывается
птицами, а чернокрылая становится доминирующей в популяциях. Это
явление получило название индустриального меланизма. Так вот, иссле¬
дование популяций диких растений показало, что все они полиморфны
по генам, определяющим их устойчивость к грибным болезням. Разно¬
образие растений по генам устойчивости вызывает встречный процесс
у паразита — возникает разнообразие его популяций по генам вирулентно¬
сти, дающим возможность поражать устойчивое растение. Таким образом,
разнообразие фитоценоза дополняется внутрипопуляционным разнообра¬
зием отдельных видов растений, входящих в фитоценоз. Вследствие этого
не только ни один вид паразита не накапливается до опасного эпидемио¬
логического уровня численности, но и ни одна вирулентная раса паразита
также не становится доминирующей над менее вирулентными расами.
118
Глава 5. Гоибы и растения
Видовое и внутривидовое разнообразие растений создает еще один
интересный феномен — обмен химическими сигналами между разными
растениями. Лет пятьдесят назад американский ученый Стаман провел
следующий опыт: он заразил кусочки клубней сорта батата, устойчивого
к грибу-возбудителю гнили, и поместил их под стеклянным колпаком вме¬
сте с не пораженными ломтиками сорта батата, восприимчивого к болезни.
Затем он заразил эти ломтики, но они оказались устойчивыми. Отсюда
однозначно вытекало, что устойчивый сорт после заражения выделил ка¬
кое-то летучее вещество, которое достигало ткани восприимчивого сорта
и сделало их устойчивыми. Это вещество оказалось этиленом — расти¬
тельным гормоном, ускоряющим созревание плодов. Сейчас установлено,
что не только батат, но практически все растения после заражения воз¬
будителями болезней или нападения вредителей издают разнообразные
химические сигналы тревоги, которые воспринимаются соседними рас¬
тениями и вызывают протекание в них защитных реакций. Чем более
разнообразен фитоценоз и чем более полиморфны популяции его членов,
тем более разнообразными химическими сетями опутаны растения и тем
лучше они вооружены в борьбе с фитопаразитами.
Спустимся еще ниже и снизойдем до индивидуальных растений. Мил¬
лионы лет они сосуществуют со своими паразитами. За эти годы проис¬
ходит постепенная притирка растения-хозяина и паразита друг к другу,
их коэволюция. В результате коэволюции растения выработали системы
химической защиты от паразитов, а последние — новые орудия напа¬
дения. Но поскольку взаимная гонка вооружений ни к чему хорошему
не приводит, а только истощает экономику, между растениями и пара¬
зитами были выработаны «соглашения»: мы даем вам возможность жить
и размножаться за наш счет, но вы будете вести себя тихо, интеллигентно,
и не брать больше того, что вам нужно, а если сможете, то и помогать
нам, оплачивая этим дом и стол. В результате такого «соглашения» воз¬
никли эндемичные (местные) болезни, которые всегда присутствуют в по¬
пуляциях, но никогда (при относительно стабильных внешних условиях)
не приводят к гибели популяции своих хозяев. Например, каждый дачник,
на участке которого растет много деревьев, страдает головной болью из-за
сорняка сныти, которую совершено невозможно вывести. Весной на мо¬
лодых листьях сныти появляются желтоватые пятна, с нижней стороны
которых заметен белый налет спороношения паразитического оомицета
Плазмопары. Заболевание интенсивно развивается каждый год, но ни¬
сколько не мешает процветанию своего хозяина.
К чему приводит коэволюция растений и парази-
спорыньевых тов’ можно показать на примере спорыньевых грибов,
грибов Жизненный цикл спорыньи похож на сказку. В колосе
ржи, которая часто поражается спорыньей, в некоторых
колосках вместо семян образуются крупные, морщинистые, фиолетовые
рожки — склероции. Спорынья поражает и многие дикие злаки, но у них
§2. Грибы-паразиты
119
более мелкие колоски и, соответственно, более мелкие склероции. Значит,
заболевание приводит к снижению семенной продукции, то есть это —
паразит, который ничего хорошего растению не приносит. Осенью скле¬
роции попадают в урожай или опадают и зимуют на поверхности поч¬
вы. О том, что происходит при попадании склероциев в муку, рассказано
в предыдущей главе. Здесь же расскажем о более естественном жизненном
пути гриба паразита. Весной опавшие склероции прорастают причудливым
образом: на каждом склероции вырастают круглые желтовато-оранжевые
головки, сидящие на длинных ножках (рис. 25 б). Эти головки называются
стромами. В каждой строме по ее периферии располагаются бутылковид¬
ные плодовые тела перитеции, направленные своими отверстиями нару¬
жу, в каждом плодовом теле находятся многочисленные вытянутые сумки,
а в них — нитевидные споры. Получается, как в сказке про смерть Кощея:
в зайце — утка, в утке — щука, в щуке — яйцо, в яйце — иголка. При созре¬
вании споры-иголки выбрасываются из сумок и разносятся ветром. Попа¬
дая на цветки ржи, они там разрастаются и образуют мицелий с бесполы¬
ми спорами — конидиями. Одновременно с конидиями мицелий образует
сладкий нектар, называемый в народе «медвяная роса», который привлека¬
ет насекомых. Перелетая с цветка на цветок, насекомые переносят на сво¬
ем теле конидии, которые заражают новые цветки. В ткани цветка ми¬
целий разрастается на обильной пище (ведь для развития семян растение
направляет в цветки большую часть продуктов фотосинтеза, выработан¬
ных листьями) и формирует склероции. При сильном поражении растение
вообще может остаться без семенной продукции. Таким образом, гриб осу¬
ществляет стерилизацию пораженных растений, не влияя на их вегетатив¬
ный рост. По-другому осуществляет стерилизацию пораженных растений
родственница спорыньи, вызывающая чехловидную болезнь кормового
злака ежи сборной. Гриб заражает не цветки, а основания листьев, и фор¬
мирует там сплетение гиф — строму, образующую чехол вокруг стебля
(рис. 25 в). Сначала в строме образуются конидии, которые могут заражать
другие растения, а затем строма буреет, и в ней формируются зимующие
плодовые тела с сумками и аскоспорами. Зараженные растения не обра¬
зуют соцветий (метелок) и, следовательно, семян. Дальнейшая эволюция
гриба привела к постепенным потерям: сначала было утрачено половое
размножение — плодовые тела, сумки и аскоспоры; затем произошла утра¬
та и конидий, остался один мицелий, живущий внутри тканей растения-
хозяина. Следовательно, было потеряно основное свойство паразитов —
способность инфекции распространяться от больного организма к здоро¬
вым — горизонтальная передача инфекции. Взамен этого гриб приобрел
способность проникать в формирующиеся семена и передаваться с ними
новому поколению, то есть вертикальную передачу инфекции. Таким об¬
разом, грибы тихо существуют в растении, извлекают из него продукты
фотосинтеза, но не снижают ни семенную продукцию, ни вегетативный
рост. Как же узнали об их существовании? Оказывается, эти эндофитные
120
Глава 5. Грибы и растения
(живущие внутри растений) грибы утратили не все, что присуще спорынье-
вым грибам, в частности токсины, описанные в предыдущей главе. Более
ста лет назад появились сообщения о странной болезни коров, которые ху¬
деют, слабеют, теряют аппетит, а иногда и погибают. Затем выяснили, что
коровы болеют после питания травой на определенных пастбищах. Анализ
травы показал, что внутри растений находится грибной мицелий, а в зара¬
женных растениях обнаружены токсины, сходные с токсинами спорыньи,
о которых уже рассказано. Эти открытия стимулировали интенсивные ис¬
следования эндофитных грибов кормовых злаков. Оказалось, что польза
их для растений с лихвой возмещает потери, вызванные тратой продуктов
фотосинтеза на их питание. Зараженные ими растения становятся ядови¬
тыми для жвачных животных, насекомых-вредителей, голых слизней и да¬
же для фитопатогенных грибов. Кроме того, зараженные растения лучше,
чем свободные от инфекции, переносят жару и засуху, более устойчивы
к зимним морозам. Вот так эндемичные паразиты постепенно эволюцио¬
нировали в сторону ослабления вредного воздействия на растения и даже
стали полезными для них, то есть приобрели свойства симбионтов.
Много видов эндофитных грибов обитает в древесных растениях. Для
некоторых из них также установлена способность защищать своего покро¬
вителя от вредных организмов. Так, эндофитные грибы, живущие в дубах
и американских соснах дуглассиях, препятствуют размножению в них на¬
секомых, которые вызывают разрастание тканей растений — образование
галлов. Некоторые грибы заселяют ткань галлов и вызывают гибель на¬
ходящихся там насекомых, другие — делают растения непригодными для
питания насекомыми.
Итак, эволюция хозяино-паразитических взаимоотношений приво¬
дит к снижению вреда, приносимого паразитами своим хозяевам, к воз-
л . никновению генотипов растений, обеспечива-
2. Защита сложившегося
биоценоза от внедрения ющих толерантность, замене острых летальных
чуждых элементов заболеваний хроническими, вялотекущими. Од¬
нако если в сложившийся фитоценоз попадает
новый вид растения, ранее живший в отдаленном регионе и не встречав¬
шийся с паразитами данного фитоценоза, он может подвергаться большо¬
му риску и просто быть уничтоженным паразитами, ибо не имеет набора
механизмов, обеспечивающих толерантность к обитающим в новом для
него регионе паразитам. Паразиты — это мощная крепость, выстроен¬
ная обитателями биоценозами и защищающая их от пришельцев, которые
потенциально могут быть опасными конкурентами для них. И это харак¬
терно не только для растений и их грибных паразитов. Экспедиционный
корпус Наполеона погиб на Антильских островах не от стрел туземцев,
ведь его составляли опытные солдаты, закаленные в войнах и оснащенные
современным (для того времени) огнестрельным оружием. Наполеонов¬
ских солдат скосил вирус — возбудитель желтой лихорадки, к которому
местные жители были толерантны, а европейцы — чрезвычайно чувстви¬
§2. Грибы-паразиты
121
тельны. Другой пример: как писал известный зоолог Гржимек, муха цеце —
это последний оплот в защите диких африканских копытных от экспан¬
сии фермеров-животноводов. Муха цеце переносит возбудителя опасной
сонной болезни, к которой африканские зебры и антилопы толерантны,
а вывезенные из Европы коровы и овцы — высокочувствительны. Фер¬
меры расчищают природные кормовые угодья диких животных, саванны,
превращая их в пастбища для скота, но его падеж от болезней задерживает
этот процесс.
То же самое происходит и в фитоценозах. В середине прошлого века
один из советских лидеров, Н. С. Хрущев, увлекавшийся экспериментами
в сельском хозяйстве, приказал засевать поля семенами конских бобов.
Конские бобы — очень хорошая кормовая культура, беда только в том,
что она ранее не возделывалась в России в больших масштабах. Когда бо¬
бами засеяли огромные площади, оказалось, что паразиты, которые слабо
развивались на дикорастущих бобовых растениях, яростно накинулись
на бобы. В частности, обитающий в почве гриб Фузариум вызвал мас¬
совое увядание и гибель конских бобов. Так что от этой новой культуры
пришлось отказаться.
А теперь перейдем к обсуждению практически очень важного вопро¬
са: если в природных фитоценозах паразиты не очень вредят, а иногда
даже помогают своим хозяевам, то почему же они так опасны для сель¬
скохозяйственных растений? Почему, скажем, оомицет Плазмопара никак
не влияет на процветание популяций сорного растения сныти, но в вино¬
градных плантациях вызывает такой урон (рис. 8 в), что если не проводить
ежегодно по десять опрыскиваний виноградников фунгицидами, то ни¬
какого урожая не соберешь и можешь сгубить плантацию? Прежде чем
ответить на этот вопрос, приведем несколько примеров губительно воз¬
действия грибных паразитов на сельскохозяйственные и лесные культуры.
2.1. Головня и мучнистая роса
Головневые грибы поражают многие травянистые растения, но нико¬
гда не бывают на древесных, причины этой избирательности растений-хо-
зяев не по родству, а по жизненным формам, не ясны. Особенно страдают
от головни злаки, поэтому в сельском хозяйстве головня наносит наиболь¬
ший урон зерновым культурам. Самые вредные — твердая и пыльная го¬
ловня пшеницы (рис. 30). Мицелий возбудителя твердой головни заражает
всходы пшеницы и распространяется внутри тканей по всему растению,
а в период цветения концентрируется в колосках, питаясь их богатым со¬
держимым (ведь растение, заботясь о потомстве, направляет туда потоки
питательных веществ). В конце сезона мицелий распадается на зимую¬
щие споры, покрытые толстыми, темно окрашенными оболочками, масса
которых замещает ткань формирующегося семени. Поэтому, если поте¬
реть пораженный колосок между пальцами, разрушив семенные чешуйки,
то пальцы будут черными от множества спор и приобретут селедочный
122
Глава 5. Грибы и растения
запах, отчего эта головня называется еще
и вонючей. То же самое происходит и во
время обмолота зерна: при отделении ра¬
бочими органами комбайна семян от по¬
ловы и мякины споры рассыпаются и
покрывают поверхность семян. Весной,
когда семена высевают в землю, споры
головни прорастают, и их ростки внедря¬
ются в ткани всходов через еще не окреп¬
шие покровы. Поскольку споры гриба зи¬
муют на поверхности семян, бороться с
ними очень просто: надо перед посадкой
Рис. 30. Головнёвые болезни обработать семена фунгицидами, убива-
пшеницы (Дьяков с соавт2007) ющими споры. Таких фунгицидов сейчас
химики насинтезировали очень много, так
что выбор основывается не только на токсичности для спор головни, но и
на экологической безопасности для организмов, не являющихся мишеня¬
ми, — фермеров, птиц, почвенных и водных организмов.
По-иному происходит заражение пшеницы пыльной головней. Мице¬
лий этого гриба, как и предыдущего, находится внутри растения и концен¬
трируется в тканях колосков, но при созревании спор полностью разруша¬
ет все кроющие ткани, так что остается лишь стержень колоса, покрытый
черным пылящим порошком (почему головня получила название пыль¬
ной). Если разлетающиеся споры попадут на цветок соседнего растения,
они там прорастают, и коротенький мицелий, состоящий всего из не¬
скольких клеток, остается зимовать в зерне. Весной, когда зараженное
зерно прорастает, с ним вместе начинает расти и зимующая в нем гифа.
Бороться с пыльной головней сложнее, чем с твердой: зачатки гриба нахо¬
дятся не на поверхности семени, а внутри него, и, поскольку большинство
используемых фунгицидов не проникает внутрь семени, паразитический
мицелий недоступен для их действия. Однако были получены фунги¬
циды внутрирастительного действия (системные фунгициды), и некото¬
рые из них оказались высоко эффективными против возбудителя пыль¬
ной головни. Они подавляют активность ферментов, необходимых для
выработки энергии в митохондриях, причем ферменты базидиомицетов,
к которым принадлежат головневые грибы, подавляются значительно бо¬
лее низкими концентрациями этих препаратов, чем ферменты растения.
Поэтому можно подобрать концентрации фунгицида, убивающие гриб,
но никак не влияющие на растение.
Кроме того, селекционеры создали много сортов пшеницы и других
зерновых культур, устойчивых к разным видам головни.
Мучнисторосяные грибы ничем не похожи на головневые. Мы при¬
выкли связывать такие понятия, как «грибы», «плесени» с сыростью.
Мучнисторосяные грибы, наоборот, развиваются на открытой для солнца
§2. Грибы-паразиты 123
Рис. 31. Мучнисторосяные болезни растений: а) пораженный
лист дуба (£. П. Кузьмичев с со авт., 2004); б) мицелий и спо-
роношение гриба-возбудителя болезни на листьях пшеницы
(фотография Г. Аветисяна)
верхней поверхности листьев или плодов, и наиболее обильно они разви¬
ваются на растениях в жаркое сухое лето. Их мицелий «ползет» по поверх¬
ности листа, пуская в клетки эпидермиса гаустории. На мицелии образу¬
ются цепочки спор (конидий), которые разносятся воздушными токами
и заражают новые растения. Мицелий и споры образуют на листьях на¬
лет белого цвета, что и определило название болезни «мучнистая роса»
(рис. 31). Особенно активна мучнистая роса в жаркое, сухое лето — тогда
вся растительность кажется обсыпаной зубным порошком. Всегда можно
наблюдать мучнистую росу на листьях птичьей гречишки (топтун травы),
молодых листьях дуба и других растений. Мучнисторосяные грибы часто
обусловливают сизоватый оттенок флоры сухих субтропиков, например,
крымской растительности. Массовое развитие мучнистой росы наносит
большой урон злакам, винограду, яблоням, крыжовнику, флоксам и дру¬
гим растениям.
Почему же эти две столь несхожие болезни рас-
0 тт Разные стратегии
тении мы рассматриваем вместе? Потому что это две воспроизводства
крайности на длинной шкале грибов-возбудителей
болезней растений. Как уже было сказано, мицелий возбудителя головни
распространяется по всему растению внутри него, а в период цветения
концентрируется в колосках, питаясь их богатым содержимым, в конце
сезона мицелий распадается на зимующие споры. Допустим, что на всхо¬
ды попала всего одна головневая спора, заразившая один росток (хотя
это предположение нереально, ибо одной спорой вызвать заражение не¬
возможно). В результате такого заражения осенью окажется зараженным
только одно растение. Допустим снова, что в нем образовался милли¬
он головневых спор, и на следующий год каждая спора заразила один
проросток. Следовательно, осенью будет поражено уже миллион расте¬
ний. Болезни, возбудители которых, подобно твердой головне, образуют
в течение года одну генерацию спор, называют моноциклическими (весь
124
Глава 5. Грибы и растения
их годовой цикл состоит из одного поколения спор). Число поражений,
возникающих при таких болезнях, не увеличивается в течение сезона
и определяется лишь числом успешных заражений.
Теперь рассмотрим мучнистую росу. При первичном заражении листа
на нем через несколько дней образуется пятно с обильным спороношени-
ем гриба на поверхности. Каждая спора, попав на лист того же или сосед¬
него растения, вызовет образование нового пятна со спорами, причем та¬
кие повторные заражения могут происходить многократно в течение лета.
Болезни, чьи возбудители подобно возбудителю мучнистой росы, образуют
несколько генераций спор в течение годового цикла развития, называют
полициклическими. Допустим теперь, как и для возбудителя головни, что
весной на лист пшеницы попала одна спора и вызвала первичное зараже¬
ние. Через неделю вокруг зараженного места образуется пятно с десятью
тысячами спор. Если каждая спора после их разлета вызовет вторичное
заражение листьев, то через две недели на поле будет уже десять тысяч пя¬
тен мучнистой росы, содержащих 100 миллионов спор (108). Если каждая
спора снова заразит восприимчивую ткань листьев, то через три недели
после первичного заражения это число возрастет до ста миллионов пя¬
тен, содержащий триллион спор (1012). Этот процесс будет продолжаться
до тех пор, пока погодные условия не перестанут способствовать развитию
болезни или на поле не останется больше восприимчивой ткани. На самом
деле скорость развития болезни, конечно, не так катастрофически высока,
так как не все образовавшиеся споры попадают на восприимчивую ткань,
которой к тому же после каждого поколения спор становится все меньше.
Но и с учетом этих поправок при полициклических болезнях количество
пораженных растений зависит не только от первичного число спор, как
при моноциклических, но и от таких показателей, как скорость образова¬
ния нового поколения спор, число жизнеспособных спор, образующихся
в каждом поколения и т. п.
Различия скорости нарастания болезни при моно- и полицикличе¬
ских болезнях можно сравнить с нарастанием вклада в банке на вложен¬
ный капитал по типу прерывистых и непрерывных сложных процентов.
Прерывистыми сложными процентами называют начисление процентов
на вложенную в банк сумму только один раз в год. Допустим, мы вложили
в банк 1000 рублей под 10 % годовых.
За первый год прибавка к вкладу составит 1000 • (1 + ^) рублей.
За второй год она увеличится до [1000*(1 + ^)](1 + ^) = 1000-(1 + ^)2.
А за десять лет — 1000 • (1 + *^)10 = 1000 • 2,5937.
Хотя большинство банков начисляет проценты на вложенный капитал
подобным образом, возникает вопрос, а почему учитывается прибавка
только один раз в год. А что получится, если учитывать прибавку каждый
месяц?
§2. Грибы-паразиты
125
Тогда через год она составит 1000 • (1 Н- щ^)12, а через десять лет —
то есть выше, чем при ежегодном начислении.
Если же делать пересчеты надбавки не ежемесячно, а ежедневно, то
увеличение вклада через 10 лет составит
10*365
А почему, собственно говоря, надо перечислять прибавку вклада еже¬
дневно? Давайте перечислять ежесекундно и посмотрим, что получим
через десять лет:
Этот размер прибавки математики обозначают показателем е, кото¬
рый вычисляется как
где п — очень большая величина. Ежесекундный пересчет прибавки —
это принцип начисления непрерывных сложных процентов.
Вернемся опять к нашим двум грибам — возбудителям головни и муч¬
нистой росы. Первый образует только одну генерацию за сезон (проростки
спор заражают всходы пшеницы, а споровое потомство образуется в конце
лета). Таким образом, рост численности возбудителя болезни и, соответ¬
ственно, рост числа пораженных растений можно рассчитывать как рост
банковского капитала при начислении прерывистых сложных процентов.
Такой рост определяется лишь тем, сколько спор образовалось на одном
зараженном растении (этот показатель равнозначен процентной ставке
и обозначается индексом г). Следовательно, пораженность в следующей
генерации (через год) Xt будет равна исходному (весеннему) числу зара¬
женных растений (Х0), помноженному на число образовавшихся на одном
растении спор (г). Возбудитель мучнистой росы образует в течение се¬
зона много бесполых генераций, причем, поскольку продолжительность
генераций разных индивидуумов не совпадает по времени и генерации
перекрываются, расчет скорости роста пораженное™ можно проводить
по уравнениям сложных процентов, то есть скорость нарастания болезни
будет не арифметической, а логарифмической (Xt = Xoert).
Наконец, рассмотрим возбудителей этих двух болезней с точки зре¬
ния их жизненных позиций. Возбудитель головни уходит от превратно¬
стей окружающего мира под покровы тканей растения и «сидит» там весь
= 1000 • 2,7181.
315 360000
126
Глава 5. Грибы и растения
вегетационный период. Его жизненная стратегия — это стратегия расчет¬
ливого реалиста. В его действиях нет осечки: споры во время обмолота
обязательно попадут на семена восприимчивого растения и будут внесены
с ними в почву; росток споры обязательно внедрится в ткань проростка
растения; находясь в растении, гриб спрятан от опасных перемен погоды
и даже ядохимикаты ему не страшны, если они не обладают внутрирас-
тительным действием. Но этот комфорт приходится оплачивать дорогой
ценой — недостаточной плодовитостью. Такая жизненная стратегия, при
которой главная цель — сохранение индивидуума, называется, как уже
было сказано, If-стратегией.
По-иному ведет себя возбудитель мучнистой росы. Он все жизнен¬
ные силы тратит не на сохранение, а на размножение, ежедневно фор¬
мируя десятки тысяч спор. Мицелий гриба находится на поверхности
листа и подвержен риску погибнуть от всевозможных причин, однако та¬
кое расположение позволяет не тратить энергию на прорыв эпидермиса
для освобождения спор, как это делают грибы, растущие внутри тканей
растения. Мелкие легкие споры отделяются от мицелия непосредственно
на месте их образования и воздушными токами разносятся на огромные
расстояния (например, зафиксирован воздушный перенос вирулентной
расы возбудителя мучнистой росы ячменя из континентальной Европы
на Британские острова). Миллионы и миллиарды спор гибнут, не по¬
пав на листья восприимчивых растений. Жизнеспособность спор теряется
уже через несколько дней после их образования, однако огромная энер¬
гия размножения обеспечивает поддержку популяции на высоком уровне
и быстрый захват новых территорий. Такая стратегия, как уже было рас¬
сказано, называется г-стратегией.
2.2. Ржавчинные грибы
На ржавчину хлебных злаков люди обратили внимание, по-видимому,
вскоре после возникновения земледельческой культуры. О ней упомина¬
ется в Ветхом Завете высокопарно-мистически: «Будет ли на земле голод,
будет ли моровая язва, будет палящий ветер, ржавчина, саранча, червь,
неприятель ли будет теснить его в земле его...». Как видим, ржавчина упо¬
минается наравне с моровой язвой, нападением неприятеля, саранчой. Ве¬
роятно, ржавчина была бичом хлебных злаков во всех странах Средиземно¬
морья. «Отец ботаники», грек Теофраст, крупнейший ученый того времени
(рубеж III и IV веков до P. X.), пишет о ржавчине в своей книге «Иссле¬
дование о растениях» сухо, но вполне по-научному: «Хлебные растения...
подвержены ржавчине больше, чем бобовые, при этом ячмень больше, чем
пшеница, и одни сорта его больше, чем другие... На местах ветреных и вы¬
соких ржавчина бывает редко или не бывает вовсе; она страшна для кот¬
ловин и мест безветренных. Появляется она преимущественно в полнолу¬
ние». Римлянин Овидий 300 лет спустя, будучи поэтом, а не ученым, в по¬
эме «Фасты» («Календарь») описывает ржавчину вполне поэтически, как
§2. Грибы-паразиты
127
результат действий злой богини Робиги, которой 23 апреля (день помина¬
ния Робиги — робигалии) следует приносить жертвы и молить о милости:
Злая Робига, щади посевы церериных злаков,
Дай им над почвой качать нежные стебли свои!
Всходам дай возрастать под сиянием звезд благосклонных
Вплоть до того, как они станут годны для серпов.
Сила твоя велика: ведь хлеб, пораженный тобою,
Грустный сочтет селянин горькой потерей своей.
О, пощади и не тронь ты всходов шершавой рукою,
И не губи урожай, мощь твоя ведома всем!
Прошли почти два тысячелетия, но ясности в
вопросе, какова природа ржавчины злаков, боль- меняющи^хоз^на
ше не стало. Ботаник Унгер считал, что пятна на
листьях, наполненные ржавым порошком, возникают в результате заболе¬
ваний, вызванных неблагоприятным условиями жизни, а сам порошок —
это «сыпь», побочный результат болезни. Во второй половине XIX века
по Западной Европе прокатились «ржавчинные процессы». Крестьяне сея¬
ли рожь, а помещики высаживали в садах и вокруг усадеб вошедший в моду
декоративный кустарник барбарис. Крестьяне стали замечать, что на по¬
лях, расположенных вблизи барбариса, листья и стебли ржи покрываются
ржавыми пятнами, семена становятся щуплыми и урожай падает. Они
стали судиться с помещиками, требуя вырубить кусты барбариса. Судьи,
как это делается до сих пор, обратились к экспертам. Но ученые-ботаники
отвергли претензии крестьян, как неграмотное суеверие. На листьях бар¬
бариса и на листьях ржи обитают совершенно разные болезни, имеющие
различные научные названия и не способные превращаться одна в другую
(тогда еще не родился академик Лысенко, провозгласивший возможность
зарождения одних видов в недрах других). И хотя некоторым опытникам,
в частности, голландскому школьному учителю, удалось искусственно за¬
разить рожь порошком, собранным с листьев барбариса, тяжбы не пре¬
кращались. И только исследования великого немецкого миколога Антона
де Бари окончательно закрыли вопрос о связи хлебной ржавчины с куста¬
ми барбариса. Де Бари показал, что пятна на листьях барбариса и на ржи
являются разными стадиями развития одного гриба, споры которого вес¬
ной заражают барбарис, а летом перелетают на злаки (рис. 32).
От ржавчинных грибов страдают не только злаковые культуры. Эти
грибы поражают огромное число видов растений, от папоротников до
цветковых. Особенно страдают сосны, которые поражаются около 20 ви¬
дами ржавчинных грибов. Многие виды образуют многолетний мицелий
в стволах сосен и постепенно приводят их к гибели. В Центральной Рос¬
сии, особенно в городах, почти все сосны страдают суховершинностью —
высыханием верхушки деревьев. Главная причина этой болезни — пора¬
жение ржавчинными грибами.
128
Глава 5. Грибы и растения
Рис. 32. Ржавчина пшеницы: а) пятна на листьях промежуточного
хозяина барбариса; б) пораженные стебли пшеницы
Ботаники разделяют виды сосны на несколько групп. Одна группа
включает виды с пятью находящимися в одном пучке хвоинками (а не с
двумя, как у нашей обычной сосны). Пятихвойные сосны образуют мало
смолы и в связи с этим имеют более светлую и мягкую древесину, пред¬
ставляющую большую ценность для деревообрабатывающей промышлен¬
ности. Как полагают, они возникли еще в мезозойской эре, свыше 100
миллионов лет назад, ца исчезнувшем материке Берингия, соединявшем
Евразию с Северной Америкой (там, где теперь Берингово море). Оттуда
пятихвойные сосны распространялись двумя потоками. Первый путь —
на восток: Аляска, Юкон, острова Франклина, Гренландия. На этом пути
возникла веймутова сосна, образующая сосновые леса в северных штатах
США и Канаде. Второй путь — на юго-запад: в Сибирь, Китай, Афга¬
нистан, Турцию, Балканы, Альпы. На этом направлении возникли си¬
бирская сосна (сибирский кедр) и встречающаяся в Альпах европейская
или кедровая сосна. Ржавчина, поражающая пятихвойные сосны, возник¬
ла, скорее всего, в Центральном Китае, когда исчез мост, связывающий
Азию с Америкой. Поэтому паразит эволюционировал параллельно с ази¬
атскими, а не с американскими видами сосны. Параллельная эволюция
паразита и хозяина приводит, как было уже сказано, к их взаимному
приспособлению, в результате чего ржавчина, хотя и поражает азиатские
сосны, в частности, сибирский кедр, но развивается медленно и не вызы¬
вает гибели деревьев.
Как и возбудитель ржавчины злаков, гриб, поражающий сосну, для
прохождения полного цикла развития должен менять хозяина — перехо¬
дить с сосны на смородину, на листьях которой возникают пятна со спо-
роношением в виде столбиков. У нас гриб вредит смородине сильнее, чем
сибирским кедрам, и защищают от него не сосну, а смородину.
Сто лет назад, в результате массовых рубок сосновые леса в ряде
североамериканских штатов сильно поредели. Начались искусственные
посадки веймутовой сосны. В одном из штатов для посадок сосны не¬
обходимо было закупить ее сеянцы. Анализ рынка показал, что выгоднее
покупать сеянцы в Европе, чем в соседнем штате. Поэтому большая партия
§2. Грибы-паразиты
129
сеянцев, закупленная в Европе, была
высажена на востоке США. И вскоре
сработало «правило Балды»: «Не го¬
нялся бы ты, поп, за дешевизною».
Некоторые сеянцы оказались пора¬
женными ржавчиной (рис. 33), и бо¬
лезнь начала свое трагическое шествие
на запад, оставляя позади себя мерт¬
вые леса.
Выяснилось, что заражение здо¬
ровых сосен ржавчиной возможно
только через листья смородины. Без
смородины новые очаги болезни не
возникают. Поэтому началось плано¬
мерное уничтожение кустов дикой
смородины. Было уничтожено около
полумиллиарда кустов, но эффект от
этого мероприятия оказался недоста¬
точным: ведь на одном квадратном
сантиметре площади пораженного ли¬
ста смородины образуется более 4-х
миллионов спор, каждая из которых,
попав на хвоинку, может вызвать по¬
ражение сосны. Поэтому даже единичных, случайно оставшихся в лесу
кустов смородины достаточно для поддержания эпидемии. Было испы¬
тано другое средство — опрыскивание больных лесов противогрибными
антибиотиками, главными образом, с помощью авиации. Обработка лесов
на площади более 200 тысяч гектаров показала, что, во-первых, внутрен¬
няя инфекция не уничтожается, а многолетний мицелий гриба находится
внутри стволов и ветвей, и, во-вторых, используемый антибиотик высоко¬
токсичен для теплокровных животных и человека. Поэтому от химических
обработок пришлось отказаться. В ряде районов главным способом борь¬
бы с заболеванием стала обрезка пораженных ветвей и выкорчевывание
стволов. В одном только 1967 году обрезали и выкорчевали 1,2 миллиона
деревьев. Весь этот комплекс мероприятий позволил остановить распро¬
странение болезни, но какова же была цена этого?
Очень интересна история замечательной культуры — кофейного де¬
рева — и роль ржавчинной болезни в ней. Родина кофейного дерева —
Абиссинское нагорье в Эфиопии. Около тысячи лет назад арабы освои¬
ли культуру кофе и объявили кофейный напиток священным. В Европу
кофе попало через Турцию. Долгое время арабы сохраняли монополию
на выращивание кофе и, как любые монополисты, продавали его по бас¬
нословным ценам. В начале XIX века мусульманин из Индии Муслим
во время хаджа в Мекку согрешил против Аллаха: выкрал семь кофейных
Рис. 33. Пузырчатая ржавчина
стеблях веймутовой сосны
(Е. П. Кузьмичев с соавт., 2004)
130
Глава 5. Грибы и растения
зерен, привез их внутри бамбукового посоха в Индию и вырастил из них
деревца. Так кофе проникло через ближневосточный кордон и Индию.
Оттуда зерна кофе были перевезены на остров Цейлон, где условия для
его выращивания оказались наиболее благоприятными. Поскольку в это
время Индия и Цейлон были английскими колониями, англичане через
мощнейшую Ост-Индскую компанию развернули кипучую деятельность.
Компания предоставляла кредиты жителям Англии, пожелавшим стать
плантаторами на Цейлоне, и держала в своих руках всю торговлю кофе.
За короткий срок Цейлон превратился в «кофейный остров». В 1835 г. ко¬
фейные плантации на острове размещались на площади всего 200 гектаров,
а в 1870 занимали уже 200 тысяч гектаров, то есть за 35 лет площади, за¬
нятые кофейными деревьями, выросли в тысячу раз! В 60-х годах XIX века
Ост-Индская кампания ежегодно экспортировала в Европу 50 миллионов
килограмм кофе. Для ухода за плантациями и сбора урожая из Южной Ин¬
дии на Цейлон были вывезены тысячи рабочих. Кофе стал национальным
напитком англичан. Однако кофейная идиллия получила трагический ко¬
нец: в 1868 г. на Цейлон попала ржавчина кофе, вызывающая массовое
опадание листьев. Пораженные растения погибают через несколько лет.
Бедствия, вызванные ржавчиной, трудно поддаются описанию. Урожай
упал с 45-50 до 3 миллионов килограммов в год. В 1880 г. английский Во¬
сточный банк объявил себя банкротом, разорив 417 плантаторов. Многие
индийские рабочие вернулись голодать на родину. В 1890 г. были вы¬
рублены последние кофейные деревья на Цейлоне. Английский ботаник
М. Уорд, направленный на Цейлон для изучения болезни, дал ее возбу¬
дителю название «вастатрих», то есть опустошительная. Поскольку свято
место пусто не бывает, сельскохозяйственный вакуум, вызванный ржав¬
чиной на Цейлоне, постепенно стал заполняться чайными плантациями.
Если в конце кофейного бума на Цейлоне было всего 500 гектаров чайных
плантаций, то уже в 1880 г. чай занял 140 тысяч гектаров. С 90-х годов
XIX века национальным напитком англичан становится чай. А отзвуком
кофейных событий XIX века после обретения Цейлоном независимости
стали гражданские войны в государстве Шри Ланка между местным насе¬
лением сингалами и потомками вывезенных из Индии рабочих тамилами.
А что же кофе? Оно спряталось от ржавчины за океан — в Южную
Америку. Отсутствие там ржавчины обеспечивало высокие стабильные
урожаи. Океан казался непреодолимым препятствием для паразита. Од¬
нако, к сожалению, только казался. Если во второй половине XIX века
наблюдалась экспансия ржавчины на восток, то во второй половине про¬
шлого столетия она двинулась на запад. В 1955 г. ржавчина была обнару¬
жена в Либерии, в 1962 г. — в Нигерии. Для западного полушария это было
еще не страшно, так как севернее 20° южной широты господствующие
ветры дуют из океана вглубь африканского континента. Однако в 1966 г.
страшная болезнь появилась в Анголе, а с юга Африки ветры дуют в сто¬
рону океана. Поэтому уже через три года ржавчина попала в Бразилию.
§2. Грибы-паразиты
131
При скорости ветра 20 км в час, расстояние 7000 километров, разделяю¬
щее Анголу и Бразилию, споры ржавчины могут преодолеть за 15 дней.
Появление ржавчины вызвало в Бразилии шок. В 1971 г. урожай кофе
в штате Парана упал с 9 до 1,2 килограмма с одного дерева. За сто лет,
прошедших со времени Цейлонской трагедии, сельскохозяйственная на¬
ука и практика многому научилась, поэтому в Бразилии не повторилась
цейлонская трагедия, но след в бразильской экономике ржавчина все-
таки оставила. В 1972 г. страна импортировала 15 тысяч тонн фунгици¬
дов и с их помощью восстановила урожай до прежнего уровня. Срочно
начались селекционные работы, приведшие к созданию устойчивых и то¬
лерантных сортов кофе. Однако все это делается не бесплатно, поэтому
международный кофейный консорциум резко повысил цены на кофе, что
ощутили на себе миллионы любителей ароматного напитка.
2.3. Фитофторовые оомицеты
Наверное, нет среди болезней растений другой столь трагически из¬
вестной, как фитофтороз картофеля (рис. 7 а, б, стр. 19). Эта болезнь ока¬
зала роковое влияние на судьбу целой нации. Даже сейчас, когда болезнь,
казалось бы, обуздали, она время от времени дает о себе знать опустоши¬
тельными эпифитотиями. Недаром описавший возбудителя знаменитый
ботаник Де Бари дал ему название фитофтора, что значит «пожиратель
растений», а саму болезнь разные народы окрестили «картофельной гни¬
лью» и «картофельной чумой».
Картофель было завезен в Европу из Америки. Вскоре после появ¬
ления он стал в странах Западной Европы основным продуктом питания
бедных слоев населения. Можно в связи с этим вспомнить картину Ван
Гога «Едоки картофеля», на которой изображена трапеза фламандских
шахтеров. Картофельная болезнь появилась почти одновременно в США
и Западной Европе (1842-1843 гг.). В 1844 г. болезнь была зарегистрирова¬
на во Франции, в Бельгии, Англии и Шотландии, но сухое лето не способ¬
ствовало ее распространению. Однако 1845-1846 годы стали драматиче¬
ской вехой в судьбе народов северо-западной Европы. В июле 1845 г. кар¬
тофель уже был поражен на всей территории Фландрии и прилегающих об¬
ластях Франции и Голландии. Августовские газеты заполнили статьи о но¬
вой болезни, которая перешла в Люксембург и по долинам Роны и Рейна —
в Швейцарию. В середине августа наблюдали массовое заболевание карто¬
феля в окрестностях Парижа. Положение становилось настолько серьез¬
ным, что члены Французской академии наук были отозваны из отпусков.
Болезнь тем временем распространилась по
южной Англии. Первое сообщение о картофель- ^трагедия ХсГв.
ной гнили в Ирландии было опубликовано в Дуб¬
линской вечерней газете 6 сентября. Вначале болезни не придали большого
значения, но при уборке картофеля в октябре выяснилось, что большин¬
ство клубней гнилые. Это вызвало панику. В следующем году болезнь
0
132
Глава 5. Грибы и растения
распространялась в Ирландии со скоростью 50 миль в неделю на восток
и северо-восток.
Картофельная болезнь принесла в Европу голод и нищету. Особенно
сильно она отразилась на судьбе Ирландии. В 1845 г. население Ирлан¬
дии составляло около 8 миллионов человек. Из них для шести миллионов
картофель составлял около половины пищевого рациона, а 2 миллио¬
на питались почти исключительно картофелем. Лишившись его, люди
потеряли единственный источник существования. Очевидцы описывают
селения, жители которых походили на обтянутые кожей скелеты. Смерть
косила людей с такой скоростью, что их не успевали хоронить. Вот, на¬
пример, характерное для того времени объявление в дублинской газете:
«Деревообрабатывающая фабрика на Королевской улице, как мы пола¬
гаем, самое крупное предприятие подобного рода в стране, обеспечивает
ежедневной работой в течение последующих шести или восьми месяцев
с утра и до ночи одновременно от 16 до 20 пар пильщиков для распилки
досок для гробов». Вид гниющих растений вызывал у населения ужас,
который отражен в художественных произведениях той эпохи, например,
в повести швейцарского писателя Иеремии 1отхельфа «Кэтти бабушка»:
«И вот при свете факела увидела Кэтти на всех кустах картофеля ужасную
черную чуму. И чем дольше она глядела, тем, казалось, чернее станови¬
лись пятна страшной болезни. Отчаяние овладело тогда старой женщиной
и, опустившись на землю, она горько заплакала».
За голодом последовали его неминуемые спутники — инфекционные
болезни. Началась массовая эмиграция оставшихся в живых ирландцев
в Англию и, особенно, в США. Толпы голодных людей атаковали отплы¬
вающие из Ирландии суда, бросая землю, дом, а иногда и близких людей.
К 1851 году после нескольких лет картофельной болезни население Ир¬
ландии уменьшилось на 2 миллиона человек. «Что значит золотуха, —
писал Ф. Энгельс, — в сравнении с тем голодом, который постиг в ре¬
зультате болезни картофеля Ирландию и который свел в могилу миллион
питающихся исключительно или почти исключительно картофелем ир¬
ландцев, а два миллиона заставил эмигрировать за океан!» Ирландские
колонии в Америке, ирландские почтальоны и ирландские полицейские,
ирландские политические деятели, подобные семейству Кеннеди — все
они потомки умирающих от голода ирландцев. В 1995 году, через 150 лет
после начала ирландской трагедии, в столице Ирландии Дублине было
проведено очень представительное научно-практическое совещание под
названием «Фитофтора-150».
В XX веке появились сообщения о поражении фитофторозом томатов.
Сейчас во многих районах болезнь вредит томатам сильнее, чем картофелю
(рис. 7 в).
Развитие фитофтороза в еще большей степени, чем развитие ржавчи¬
ны, связано с погодными условиями. Возбудитель болезни размножает¬
ся подвижными зооспорами, для существования которых требуется влага
§2. Грибы-паразиты
133
в виде дождя или обильных рос. В теплую сухую погоду развитие болезни
прекращается, однако, при смене теплых дней холодными ночами, что
часто бывает в августе, влага конденсируется в виде росы, и массовое по¬
ражение растений становится неизбежным. Поэтому в народе говорят, что
помидоры на кустах коричневеют и сгнивают вследствие того, что на них
легла роса, то есть ставят телегу впереди лошади. Чтобы плоды не сгнива¬
ли, их срывают зелеными и оставляют дозревать в помещениях. Вследствие
фитофтороза Россия стала страной «вечнозеленых помидоров».
Благодаря интенсивной селекционной работе, в мире создано боль¬
шое число сортов томата и картофеля, имеющих разную степень устой¬
чивости к фитофторе. Синтезированы высокоэффективные фунгициды
и разработаны технологии их применения. Однако паразит не перестает
удивлять картофелеводов. Более 100 лет картофель в Европе размножал¬
ся исключительно бесполым способом. Микологи, работавшие на родине
возбудителя болезни, в Мексике, установили, что там он имеет половой
процесс и образует зимующие ооспоры, которые сохраняются в почве.
Для протекания полового процесса необходимо слияние особей, имею¬
щих разные типы спаривания, которые названы А1 и А2. Исследование
европейских особей показало, что все они имеют тип спаривания А1 и по¬
этому не могут вступать в половой процесс и формировать ооспоры. Да
они, вроде бы, и не нужны; ведь имеется комфортный способ зимовки
в зараженных клубнях. Однако после того, как в Мексике в результате ра¬
боты международного коллектива ученых были разработаны эффективные
меры борьбы с фитофторозом, эта страна из импортера стала мировым
экспортером картофеля. С мексиканскими клубнями в страны Старого
Света попали и особи возбудителя, имеющие тип спаривания А2. Возник¬
ли ооспоры, которые приобрели важное эпидемиологическое значение:
паразит получил возможность зимовать не только в клубнях, но и в поч¬
ве и в семенах зараженных плодов томата. В связи с этим болезнь стала
появляться на томатах в более ранние сроки, чем на картофеле (до этого
первичным источником инфекции были картофельные клубни, поэтому
на томатах болезнь появлялась в результате перелета спор с листьев кар¬
тофеля), и повысилась ее вредоносность.
Фитофторы поражают не только картофель и томат. Многочисленные
виды этого рода вызывают серьезнейшие заболевания древесных растений.
Есть фитофторы, поражающие яблони, пальмы и гевеи (каучуковые дере¬
вья). Фитофтора, поражающая коричное дерево на островах Индонезии,
попав в Австралию, вызвала там массовую гибель эвкалиптов (рис. 7 г).
А недавно появился новый, неизвестный ранее ученым вид фитофторы,
который вызвал болезнь, названную «внезапная гибель дубов». Под ее
действием начали усыхать целые дубравы в США, Канаде, Западной Ев¬
ропе. Поражает она не только дубы, но и рододендроны, калину и другие
деревья и кустарники. Беда и в том, что этот возбудитель, как и мно¬
гие другие виды фитофтор, крайне сложно выделить в чистую культуру,
134
Глава 5. Грибы и растения
поэтому установить природу внезапной болезни деревьев почти невоз¬
можно. Сейчас и не выделяют. Просто создали диагностические системы
для установления присутствия разных видов фитофтор в субстрате моле¬
кулярными методами.
2.4. Почему так происходит
Этих примеров, наверное, достаточно, чтобы показать, что грибные
болезни сельскохозяйственных растений приводили к огромным финан¬
совым потерям, изменению географии размещения отдельных культур и
даже к массовой гибели населения. Возникает вопрос: почему же в ди¬
кой природе болезни мало вредят и даже иногда помогают растительным
популяциям и сообществам, а в сельском хозяйстве очень сильно вредят
и никогда не помогают. Какие регуляторные механизмы нарушил человек
своей аграрной политикой и технологиями? Их много, поэтому остано¬
вимся на главных.
Сама пахотная культура привела к изменениям в химическом со¬
ставе и физических свойствах почвы, что не могло не сказаться на их
обитателях. Вспашка вызвала увеличение аэрированно-
Что наделала
сти и уменьшение связанной воды в почве, а внесение
агротехника и
минеральных удобрений повысило содержание в ней до¬
ступных элементов питания — азота, фосфора, калия. Это привело к сдви¬
гу в соотношении разных форм микробного населения почвы. В частно¬
сти, увеличилось содержание в почве сумчатых и несовершенных грибов
и уменьшилось содержание их антагонистов — базидиомицетов и про¬
кариот бактерий и актиномицетов. Многие несовершенные грибы, рас¬
пространенные в почвах, питаются растительными остатками, но могут
поражать корни старых и ослабленных растений. Увеличение их числен¬
ности в пахотных почвах привело к ускорению эволюционных процессов,
протекающих в их популяциях, усилению факторов вирулентности к рас¬
тениям. Бич сельского и лесного хозяйства — корневые гнили, вызываю¬
щие массовую гибель всходов различных сельскохозяйственных и лесных
культур, — это побочный продукт пахотного земледелия.
Вторая важнейшая причина — сужение генетического разнообразия
растений. Это вызвано, прежде всего, малым числом культур, занима¬
ющих основные посевные площади в мире. Пять наи-
Одноо разие — более распространенных видов растений, которые вы¬
ращивают на более 50 % пахотных земель, это злаки
(пшеница, рис, кукуруза, ячмень, сорго, африканское просо — миллет),
имеющие общих вредителей. Например, в Великобритании на 73 % пахот¬
ных земель выращивают пшеницу и ячмень. Причем агроэкономическое
районирование мировых территорий привело к тому, что районы, наибо¬
лее подходящие по почвенно-климатическим условиям для той или иной
культуры, целиком заняты этой культурой. Центральные штаты в США це¬
ликом заняты под кукурузу и сою (эта зона и называется кукурузным поя-
§2. Грибы-паразиты
135
Рис. 34. Остров Мауи (Гавайские острова). Вид с самолета
на плантации сахарного тростника (ГВент, 1972)
сом), на целинных землях в СССР выращивали только пшеницу, поливные
земли в республиках советской Средней Азии были заняты непродоволь¬
ственной культурой — хлопчатником, что создавало проблемы снабжения
многочисленного местного населения продовольствием. Очень показа¬
тельны в этом отношении Гавайские острова. Вот как описывает остров Ла¬
наи известный чешский этнограф Мариан Стингл: «Ланаи — не край тури¬
стов. Это край ананасов. Ананасов и только. На центральной равнине ост¬
рова от горизонта до горизонта заполняют все пространство бесконечные
ряды столь привольно чувствующих себя здесь растений... Сегодня Ланаи
населяют практически одни ананасы. Около трех тысяч человек, разме¬
стившихся на острове, почти без исключения прислуживают ананасам, ра¬
ботая на компанию, которая их выращивает». А соседний остров Мауи —
это одна огромная плантация сахарного тростника (рис. 34). К этому следу¬
ет добавить, что все эти огромные плантации заняты практически одним,
наиболее урожайным по результатам предварительного испытания сортом.
Таким образом, многовидовые природные фитоценозы заменены однови¬
довыми, а генетически полиморфные популяции отдельных видов расте¬
ний — генетически однородными чистолинейными сортами. Как мы уже
136
Глава 5. Грибы и растения
говорили, паразиты в биоценозах, подобно пружине, раздвигают растения
друг от друга в слишком загущенных участках путем выпадения отдельных
пораженных особей. Люди с помощью химических средств защиты сжима¬
ют эту пружину, но стоит ослабить давление на популяции паразитов, как
пружина разжимается, возникают массовые эпифитотии и гибель посевов.
Еще одна причина массовых болезней — за-
Каждыи должен жить
на родине тогда все 303 вместе с растительным материалом паразитов
будет спокойно в новые районы. Как уже было сказано, мест¬
ные растения в процессе длительной совместной
эволюции со своими паразитами выработали механизмы устойчивости
и толерантности к ним. Попавшие в новые районы паразиты с яростью
накидываются на местные растения, у которых нет факторов устойчивости
к ним. Выше было рассказано о трагической ситуации, вызванной попа¬
данием завозом в Северную Америку возбудителя ржавчины пятихвойных
сосен. Другой пример: полвека назад в Европу попал оомицет, возбудитель
голубой плесени листьев табака. А так как лист — это орган, ради которого
табак выращивают, ущерб от болезни был очень высокий. Селекционеры,
прочитав в книжках, что родина табака — Южная Америка, стали искать
устойчивые к болезни формы в коллекциях южноамериканских Табаков,
но ничего не нашли. Потом устойчивые формы были найдены среди ав¬
стралийских Табаков. Дело в том, что после раскола Южного проматерика
Гондваны в мезозое на Южную Америку, Африку и Австралию, большин¬
ство предков современных видов табака оказались в Южной Америке,
но несколько видов попало в Австралию, где на них стал паразитировать
возбудитель голубой плесени. Вывезенные оттуда устойчивые виды табака
скрестили с культурными сортами; так были получены сорта, устойчивые
к голубой плесени. Для защиты местных сортов сельскохозяйственных
и лесных растений от попадания чуждых паразитов во всех странах созда¬
ны карантинные службы, которые составляют периодически пересматри¬
ваемые списки карантинных объектов, запрещенных к завозу, и проверяют
завозимые растительные материалы на возможное присутствие в них этих
объектов.
В последние годы появилась еще одна причина, усилившая пора-
женность растений болезнями и вредителями. Селекционерам стало мало
создавать высокоурожайные сорта растений. Им захоте-
для качества лось еще и созДавать соРта с улучшенным химическим
составом. Например, по убеждению врачей-диетологов,
высокое содержание крахмала в зерне злаков — не столь уж ценное каче¬
ство. Было бы лучше за счет крахмала повысить содержание в зерне белка.
При этом белок нужен не всякий, а с высоким содержанием незаменимых
аминокислот, которые клетки человека синтезировать не могут и должны
получать из продуктов питания в готовом виде. Так, в селекции сортов ку¬
курузы широко используют мутанты с повышенным содержанием лизина
§3. Грибы-сапротрофы
137
в зерне. Однако как было сказано, выращивание высоколизиновых сортов
кукурузы создало новую проблему — фузариоз початков.
Наконец, еще одна причина массовых болезней растений — отрав¬
ление воздуха и почвы продуктами хозяйственной деятельности — ксено¬
биотиками, которые ослабляют естественные
механизмы, обеспечивающие сопротивление ^"вс^бьмо^ь? хорошо
растений патогенным организмам. Например,
было замечено, что на вершинах Аппалачских гор в США стали поги¬
бать сосны. Как выяснилось, причиной их гибели стали кислые дожди,
содержащие продукты промышленных производств, которые приводи¬
ли к гибели микоризных грибов. А микориза, как было сказано выше,
не только необходима для питания древесных растений, но и защищает их
от поражения фитопатогенными почвенными грибами. А массовая гибель
австралийских эвкалиптов от корневой гнили, вызванной фитофторой, за¬
тронула, в основном, леса, которые испытывали высокую рекреационную
нагрузку, или, проще говоря, вытоптанные многочисленным гуляющим
населением. В таких лесах почва уплотняется, в ней уменьшается содержа¬
ние воздуха, и от недостатка кислорода происходит отмирание корешков,
которые подвергаются нападению возбудителей корневых гнилей.
Как писал известный зоолог и паразитолог В. JI. Контримавичус, «кон¬
фликт между стабилизирующим экосистемы механизмом, каковым явля¬
ются паразиты, и человеческой деятельностью — вероятно, наиболее дра¬
матический конфликт человечества с законами существования биосферы».
0
§3. Грибы-сапротрофы
Ранняя весна. Только что под лучами весеннего солнца сошел снег,
и обнажилась прошлогодняя пожухлая трава. И вдруг, как будто внезапно,
она оказалась пронизанной тонкими, как паутина, нитями, кое-где пере¬
ходящими в ватообразные хлопья, и все: в течение нескольких дней трава
исчезла, место для роста молодой, весенней зелени освобождено. Про¬
шлогоднюю траву уничтожили сапротрофные грибы так называемого «ве¬
сеннего аспекта», или «снежные плесени». Это целый комплекс различных
грибов, среди них есть и зигомицеты, и аскомицеты, и базидиомицеты.
Объединяет их то, что они, с одной стороны, слабо конкурентоспособны
и быстро вытесняются другими, более конкурентоспособными видами,
а с другой — способны, в отличие от других грибов, расти при очень
низких положительных температурах, близких к нулю. То есть эти грибы
толерантны к холодовому стрессу и относятся к большой группе стресс-
толерантных организмов. Вот они и уходят от конкуренции в свою, сво¬
бодную от других, более теплолюбивых, грибов, нишу. А в природе они
выполняют вполне достойную роль: уничтожение прошлогодней травы
и растительного опада. В теплые зимы с оттепелями, когда отдельные,
прогреваемые солнцем, участки поля освобождаются от снега, они могут
138
Глава 5. Грибы и растения
развиваться на всходах зимующих под снегом озимых злаков, вызывая
их гибель. Весной на поле образуются «проплешины». В оправдание воз¬
будителей снежной плесени можно заметить, что они честно выполняли
свои экологические функции, и не предполагали, что люди будут высевать
семена не весной, а осенью.
А что с точки зрения химика представляет собой прошлогодняя трава?
Углерод, связанный в органических полимерах, главным образом, в цел¬
люлозе. То же самое представляют собой и опавшие в воду мертвые листья
и веточки, разлагаемые вторично водными грибами, о которых было ска¬
зано в предыдущих главах. Пгобальная экологическая роль сапротрофных
грибов — участие в круговороте углерода на земле. Через тело грибов про¬
ходит 2/3 связанного углерода, формирующегося на суше, причем на входе
мы имеем стойкие высокомолекулярные полимеры углерода — целлюлозу
и лигнин, а на выходе — простейшее химическое соединение углерода —
углекислый газ. В связи со сказанным обратимся к бухгалтерии.
Баланс углерода на земле складывается из следующих показателей:
1. Первичная продукция (ПП) — весь углерод, ассимилированный рас¬
тениями.
2. Нетто-первичная продукция (НПП) — ПП минус дыхание, то есть
часть ПП, перешедшая в биомассу растений.
3. Нетто-экосистемная продукция (НЭП) — НПП минус потери в де-
струкционных процессах в течение года.
4. Нетто-биомная продукция (НБП) — устойчивый углерод, длительно
пребывающий в резервуаре.
Давайте начнем с последнего показателя.
Болота Очень емкий резервуар длительного хранения углерода —
болота. В болотах Российской федерации сохраняется 113,5 ги¬
гатонн (Гт) углерода. Это — огромная величина, ибо одна гигатонна —
это миллиард тонн (109). И это самый стойкий резервуар, ибо анаэроб¬
ные (без доступа воздуха) условия препятствуют массовому размножению
микроорганизмов. Ежегодный импорт углерода болотной растительностью
составляет 20 грамм на квадратный метр болот, а эмиссия в виде летучих
веществ — 0,1-2,5 г. Основная роль в круговороте углерода болот выпа¬
дает на долю прокариот (бактерий и архей), многие из которых успешно
развиваются в анаэробных условиях, причем на выходе из их тела образу¬
ется не только углекислый газ, но и его восстановленное соединение —
метан. Однако исследования последних лет показали, что и грибы явля¬
ются постоянными обитателями болот, хотя их вклад в потоки болотного
углерода пока не известен.
Почва Другой резервуар — почва, в которой в целом по России
сохраняется 235-366 Гг углерода. Среди различных форм, в ко¬
торых содержатся органические соединения (от опавших листьев и веток
до свободных молекул), наиболее устойчивая форма — почвенный гумус,
§3. Грибы-сапротрофы
139
состоящий из очень стойких полимеров — ароматических соединений.
В его разложении принимают участие многие бактерии и грибы. Их отно¬
сительная роль пропорциональна количественному содержанию. На пло¬
щади, равной одному гектару, в лесной почве общая биомасса живых ор¬
ганизмов, включая дождевых червей и других беспозвоночных животных,
а также всевозможных микроорганизмов, составляет 497 килограммов,
из них на грибы приходится 454 кг, или более 90 %. Протяженность гриб¬
ного мицелия в почве в разы и даже на порядки превышает протяженность
корней всех растущих на поверхности почвы растений. В одном кубиче¬
ском сантиметре почве содержится, в среднем, 800 метров мицелиальных
гиф, а ступня человека покрывает около полумиллиона километров тесно
расположенных гиф.
Поступление углерода в почву происходит вследствие ежегодного
опада растительного материала. Основные формы содержания углерода в
опаде — целлюлоза и лигнин. Грибы активно разлагают эти соединения и,
вследствие этого, принимают важнейшее участие в почвообразовательном
процессе. В процессе разложения полимерных соединений образуются
органические кислоты, которые мицелий выделяет в окружающую среду.
Кислоты воздействуют на минералы, вызывая их частичное растворение
(выветривание) и перемешивание с почвой. Таким образом осуществля¬
ется процесс первичного почвообразования и увеличение толщи почвы
на подстилающих породах.
Еще одна важная роль грибов в почве — образование и разложение
гумуса. Почвенный гумус представляет собой смесь многих органических
соединений, в которой преобладают гуминовые кислоты и полисахариды.
Они образуются двумя путями, причем оба пути связаны с химической де¬
ятельностью грибов. Во-первых, как продукты разложения растительного
лигнина, стойкого полимера, который способны разлагать только грибы,
о чем подробнее будет сказано ниже. Во-вторых, из темного пигмента
меланина, который накапливается в клеточных стенках гиф, хламидоспор
и склероциев у многих почвообитающих грибов.
Почвенный гумус очень устойчив к разложению и может сохраняться
сотни и тысячи лет. Некоторые почвообитающие грибы и бактерии спо¬
собны разлагать гумус и использовать продукты разложения для питания
в качестве источников углерода и азота. Но поскольку при разложение
гуминовых кислот до 60 % азота не используется в питании микроор¬
ганизмов, а переходит в растворимое состояние, продукты разложения
гумуса служат важным источником азота в корневом питании растении.
Именно поэтому почвы, содержащие высокий процент гумуса называют
плодородными.
Наконец, третий важнейший резервуар — древесина,
в которой запасено 34,4 Пг углерода. И здесь грибам при¬
надлежит не только решающая, но почти исключительная роль. Углерод
древесины фиксирован, в основном, в виде двух высокомолекулярных
140
Глава 5. Грибы и растения
полимеров — целлюлозы и лигнина. Целлюлоза — самый большой по весу
биополимер. Она представляет собой длинные линейные цепочки молекул
глюкозы, соединенные в пучки водородными связями. Каждая отдельная
водородная связь непрочна, но когда таких связей тысячи, разорвать пучок
на отдельные молекулы, которые можно будет затем расщеплять на ко¬
роткие фрагменты, также трудно, почти невозможно, как невозможно
было Гулливеру разорвать путы, которыми его привязали лилипуты сво¬
ими тончайшими в масштабах людей, но многочисленными веревками.
Некоторые бактерии и многие сумчатые и базидиальные грибы облада¬
ют набором ферментов, которые способны поэтапно разлагать целлюлозу.
Второй по численности — полимер ароматических веществ лигнин. Его
масса составляет 25 % от общей фотосинтетической продукции. В лигни¬
не содержится больше углерода, чем в целлюлозе и других полисахаридах.
Но его разрушение — еще более трудная задача, чем разрушение цел¬
люлозы. При производстве бумаги из целлюлозы лигнин накапливается
как побочный продукт. А поскольку он очень стоек, и, к тому же, весьма
токсичен, терриконы лигнина не поддаются утилизации и лишь создают
экологически опасную обстановку. Практически только грибы облада¬
ют ферментами лигнин-пероксидазами, способными разрушать лигнин.
И важнейшая роль в разрушении лигно-целлюлозного комплекса древе¬
сины принадлежит базидиомицетам, главным образом, трутовым грибам,
образующим мелкие однолетние или огромные многолетние плодовые те¬
ла на пораженных стволах и ветках.
Если оторвать кору от лежащих на земле, пораженных трутовыми
грибами стволов деревьев, то можно увидеть, что гниль древесины при
поражении разными видами грибов различна. Некоторые грибы вызывают
так называемую белую гниль (рис. 35с), при которой древесина становится
белой с черными прожилками и разрушается с образованием длинных
продольных волокон. Эти грибы разрушают главным образом лигнин,
но не целлюлозу. При поражении другими видами древесина, наоборот,
не светлеет, а буреет, и распадается на короткие кусочки. Это — бурая гниль
(рис. 35 б), возбудители которой более интенсивно разрушают целлюлозу,
оставляя лигнин почти не тронутым. Но большинство трутовых грибов
могут разрушать оба полимера и вызывают смешанную гниль.
Итак, что делают эти грибы? В России годичное депонирование уг¬
лерода в древесные растения составляет 240-245 миллионов тонн, в том
числе в древесину (стволы, скелетные ветви, корни) — 45 миллионов
тонн, т. е. 20-30 %. Грибы принимают активное участие в балансе как эко-
системной, так и биомной продукции. Ежегодно в лесах образуется детрит,
углеродная емкость которого зависит от двух показателей: 1) годичного
поступления опада (листьев) и отпада (веток); 2) скорости его деструкции.
Ежегодное поступление детрита в лесах нашей холодной климатической
зоны составляет около 33 кг на гектаре лиственных лесов и 45 кг на гектаре
хвойных, а время разрушения этого количества детрита грибами состав-
§3. Грибы-сапротрофы
141
Рис. 35. Гнили древесины, вызванные трутовыми грибами: а) белая гниль,
вызванная настоящим трутовиком; б) бурая гниль, вызванная окаймленным
трутовиком (Е. П. Кузьмичев с соавт., 2004)
ляет соответственно 10 и 18 лет. Таким образом, скорость поступления
в 10 и 18 раз превышает скорость разложения. Но в разложении участвуют
не только трутовые грибы. Многие шляпочные базидиомицеты относятся
к группе подстилочных сапротрофов, которые также принимают важное
участие в разложении лесной подстилки — детрита. В наших лесах часто
встречается грибочек говорушка воронковидная, с воронковидными пло¬
довыми телами телесного цвета, имеющими пластинки, сбегающие вниз
по ножке. Если плодовое тело говорушки осторожно вынуть из почвы,
то вместе с мицелием, на котором образовалось плодовое тело, из поч¬
вы будут извлечены прошлогодние листья, оплетенные мицелием. Таких
грибов много, и они принимают активное участие в разложении опада.
Затем на смену базидиальным макромицетам приходят микроскопические
плесневые грибы и бактерии, а завершают процесс разложения почвенные
беспозвоночные животные — насекомые, черви и др.
Большая часть (до 80 %) продуктов распада превращается под дей¬
ствием грибов в углекислый газ и лишь от 15 до 35 % идет на построение
мицелия. Таким образом, грибы завершают углеродный цикл, начатый
фотосинтезирующими растениями. Переход углерода в углекислый газ
сопровождается освобождением других элементов (азота, фосфора, серы,
кальция и др.), связанных в полимерах растительных клеток.
Наконец, грибы, главным образом, базидиальные, — единственные
природные первичные деструкторы древесины, то есть нетто-биомной
продукции. Процесс этот очень медленный, зависит от толщины стволов
и ветвей, видов грибов, климатических условий. Например, до полного
разложения крупных стволов сосны тсуги проходит около 3 тыс. лет, а мел¬
ких веток — от двух до 20 лет. В среднем релаксация углерода в древесине
составляет 500-600 лет. Следовательно, природный круговорот углерода
142
Глава 5. Грибы и растения
в лесных массивах сопровождается относительно медленным накопле¬
нием связанного углерода в результате фотосинтетической деятельности
растений и во много раз более медленным его освобождением в виде уг¬
лекислого газа в результате деструкционной деятельности грибов.
Быстрее всего связанный углерод освобождается при хозяйственном
или случайном (в результате лесных пожаров) сжигании древесины. Но
поскольку более чем в 90 % случаев причиной пожара является человек
со спичками, то и большинство лесных пожаров случайными не назовешь.
Таким образом, люди в результате своей деятельности выводят из мед¬
ленного оборота огромные массивы древесины, приводят их к быстрой
минерализации, несравнимой с естественной скоростью связывания, и со¬
здают тем самым глобальные экологические проблемы, подобные глобаль¬
ному потеплению.
Глава 6
Лишайники:
два в одном
§ 1. Что такое лишайник
Говоря о взаимоотношениях грибов с растениями,
нельзя не остановиться на самом интегрированном спо¬
собе таких взаимоотношений — на лишайниках. Прав¬
да, в лишайниках грибы взаимодействуют не с высшими
растениями а, за редкими исключениями, с зелеными
и (или) синезелеными водорослями, но зеленые водо¬
росли, живущие в лишайнике, и высшие растения — эти
двоюродные сестры, ибо произошли от единого предка
и имеют много общего в строении хлоропластов, наборе
пигментов, химическом составе клеточной стенки, за¬
пасных питательных веществ и прочее. А синезеленые
водоросли или, как их сейчас называют, цианобактерии,
имеют еще более глубокие связи с растениями, так как
в незапамятные времена дали начало хлоропластам, со¬
держащимся в клетках и зеленых водорослей, и высших
растений.
Лишайники — самые странные организмы на зем¬
ле. Это грибы, в талломе (теле) которых живут водо¬
росли. Поскольку гриб не съедает находящиеся в нем
клетки водоросли, а даже заботится о них, поскольку во¬
дорослевые клетки живут в лишайнике неопределенно
долгое время, а лишайники — одни из самых долго¬
живущих обитателей планеты, они живут сотни, а не¬
которые виды — и тысячи лет, и поскольку после ги¬
бели водоросли гибнет и весь лишайник, сожительство
гриба и водоросли рассматривают как симбиоз. Однако
симбиоз этот самый (опять слово — самый) своеоб¬
разный. К. А. Тимирязев назвал лишайник растением-
сфинксом, подразумевая этим двойственную природу
организма. Но у сфинксов, кентавров и иных мифо-
144
Глава 6. Лишайники: два в одном
логических химер отдельные части тела имеют четкие морфологические
черты составляющих его организмов: голова человека, а тело льва или
лошади; голова и тело женщины, а хвост — рыбий и т. п. Ту же картину
мы видим и у биологических химер — симбионтов. Растение, вступившее
в симбиотические отношения с микоризными грибами или клубенько¬
выми бактериями, приобретает некоторые морфологические изменения
в строении корней, но в остальном остается таким же растением. Ми¬
коризованную или немикоризованную сосну любой наблюдатель, даже
не будучи ботаником, назовет сосной, также как клевер, имеющий на кор¬
нях клубеньки с бактериями или не имеющий их, остается таким же
клевером. Беспозвоночные животные, содержащие в теле водорослевых
симбионтов (зоохлореллы, зооксантеллы), меняют окраску вследствие на¬
личия в клетках водорослей хлоропластов, но и только. А рак-отшельник,
который сажает на свой хитиновый панцирь актинидию, вообще стано¬
вится сфинксом или кентавром — одна часть симбиотической ассоциации
остается раком, а другая — актинидией. В лишайнике же изменено все,
что может быть изменено: форма тела, окраска, физиология, образ жизни.
Поэтому не удивительно, что долгое время ботаники считали лишайники
единым организмом; двойственная их природа была обнаружена только
около полутора сотен лет назад. Но и после этого более сотни лет лишай¬
ники не помещали внутри грибов или водорослей, а выделяли в отдельную
группу (таксон), придавая им статус отдела или царства. Несмотря на то,
что лишайники образованы смешиванием разных, далеких друг от дру¬
га организмов, причем виды лишайников возникали неоднократно при
смешивании разных видов грибов с разными видами водорослей, то есть,
несмотря на полифилетическое происхождение лишайников, у ботаников
не поднималась рука разбить единую группу лишайников, поместив от¬
дельные их виды среди видов грибов, составляющих «тело» лишайника.
И только не более полусотни лет назад поднялась рука микологов, ко¬
торые назвали лишайники лихенизированными грибами и разместили их
виды внутри различных групп грибов. В самом деле, сосна, корни кото¬
рой вступили в симбиоз с микоризным грибом, продолжает называться
сосной, почему же гриб, в теле которого живут водоросли, надо называть
не грибом, а как-то иначе? Так что логика в этом есть. Но была логика
и у тех ученых, которые выделяли лишайники в отдельную от грибов и во¬
дорослей группу организмов. Ведь несмотря на различное происхождение
и огромные морфологические различия, все лишайники имеют какие-то
общие черты, позволяющие наблюдателям отделить лишайники от не-
лишайников. По-видимому, симбиоз грибов и водорослей, независимо
от конкретных видов этих организмов, приводит к сходным, как гово¬
рят биологи, канализованным направлениям в изменениях морфологии,
физиологии и биохимии слагающих лишайник организмов, изменениях
настолько глубоких, что отдельно друг от друга они уже существовать
не могут, а если и существуют, то совсем не так, как в лишайнике.
§3. Как выглядят лишайники
145
§2. Обитатели лишайниковой «коммуналки»
Сейчас описано около 20 тысяч видов лишайников (пардон, лихени-
зированных грибов), что составляет около четверти всех описанных ви¬
дов грибов. Поскольку в свободном состоянии лихенизированные грибы
не живут, единственный способ, позволяющий определить принадлеж¬
ность их к той или иной группе грибов, — это изучение морфологии
плодовых тел, образующихся на поверхности таллома многих лишайни¬
ков (рис. 36 г, д, з). Оказалось, что 98 % всех лишайников образуют аско-
мицеты и только меньше одного процента лишайников, встречающихся
в тропиках, сформировано базидиальными грибами. Это означает, что
лихенизировано около половины всех известных сумчатых грибов. Неко¬
торые лишайники не формируют плодовых тел, но для установления их
родства с нелихенизированными грибами пришли на помощь молекуляр¬
ные методы геносистематики.
Второй компонент лишайника — способная к фотосинтезу водоросль,
названная фотобионтом. Число видов водорослей, живущих в лишайни¬
ках, составляет менее одного процента от числа лихенизированных грибов,
следовательно, один фотобионт может формировать ассоциации со мно¬
гими микобионтами. В отличие от грибов, лихенизированные водоросли
могут жить и без лишайника, однако и среди них есть высоко специализи¬
рованные виды; более 40 % лишайниковых микобионтов ассоциированы
с водорослями рода Требуксия, которые в свободном состоянии встреча¬
ются крайне редко. Надо отметить, что видовая и даже родовая принад¬
лежность лихенизированных водорослей определена лишь у небольшого
числа видов лишайников. Дело в том, что жизнь в лишайнике настолько
изменяет морфологию и поведение водорослей, что сопоставить их со сво¬
бодно живущими родственниками удается далеко не всегда. Среди фото-
бионтов по числу видов преобладают зеленые водоросли, на втором месте
находятся синезеленые, или цианобактерии (в отличие от остальные водо¬
рослей синезеленые не имеют ядра и иных покрытых мембраной органелл
и относятся к прокариотам, поэтому их и называют цианобактериями).
Возможны и двойные симбиозы. У ряда лишайников кроме первичного
зеленого фотобионта на поверхности таллома образуются бородавочки —
цефалодии, в которых находятся клетки вторичного фотобионта — ци¬
анобактерии, причем у некоторых видов в цефалодиях находятся не один,
а несколько видов цианобактерий. Если к этому прибавить, что в талломах
многих лишайников живут и нефотосинтезирующие бактерии, то лишай¬
ник действительно можно уподобить «коммуналке» со многими жильцами.
§3. Как выглядят лишайники
Самые примитивные лишайники имеют вид белой или желтоватой
пудры, которой посыпаны камни или кора деревьев. Такой порошковид¬
ный таллом называют лепрозным. Он образован нитями микобионта, опле-
146
Глава 6. Лишайники: два в одном
Рис. 36. Морфология лишайников: а), б) накипные лишайники ризокарпон и гема-
томма на камнях; а) листоватый лишайник пельтигера на почве; г) листоватый лишай¬
ник ксантория (стенная золотянка) на коре с блюдцевидными апотециями в центре
таллома; д) кустистый лишайник кладина; е) кустистый лишайник цетрария (исланд¬
ский мох); ж) кустистый лишайник кладония (олений мох); з) кустистый лишайник
Нейропогон с апотециями на вершинах веточек; и) кустистый лишайник уснея (ма¬
ралий мох) свисающий с ветки (Жизнь растений, т.З, 1977)
§ 3. Как выглядят лишайники
147
тающими отдельные клетки водоросли. Более сложно устроены накипные
(рис. 36 я, б) лишайники, которые выглядят как разноцветные пятна на ка¬
менистом субстрате или коре деревьев. Именно по виду накипных лишай¬
ников, напоминающих пятна лишая на теле животного, эти организмы
и получили свое название. Их таллом или полностью врос в субстрат,
так что наружу выступают только плодовые тела — апотеции, или раз¬
вивается на поверхности субстрата, но настолько плотно сросся с ним,
что его нельзя отделить от субстрата, не повредив. У листоватых лишай¬
ников таллом всегда находится на поверхности субстрата и имеет вид
плоских морщинистых розеток или более или менее широких лопастей
(рис. 36 в, г). Он прикреплен к субстрату или многочисленными пучками
гиф, расположенными на нижней поверхности таллома, или одним цен¬
тральный пучком. Листоватые лишайники часто обрастают кору деревьев,
которая покрывается серым мохнатым покровом, или образуют круглые
розетки на земле, коре, камнях, заборах, крышах домов и т. п. Еще бо¬
лее разнообразны кустистые лишайники, таллом которых не распростерт
по субстрату, как у листоватых, а поднимается над ним. Их талломы име¬
ют вид одиночных выростов, заостренных на конце, подобно шилу, или,
наоборот, имеющих расширенный конец, напоминающий горн или трубу
(рис. 36 д), или сильно разветвленных изящных кустиков, веточки кото¬
рых утончаются к концам (рис. 36ж). Такие сильно ветвящиеся кустистые
лишайники, окрашенные в светлые, серовато-зеленоватые тона, являются
доминирующими «растениями» в некоторых фитоценозах, в частности,
в сосновых борах и тундрах. В геоботанических классификациях фито¬
ценозов имеется растительное сообщество, названное «бор-беломошник»,
в котором вместо травяного покрова почва покрыта кустистыми лишай¬
никами из рода Кладония, названными «белым мхом». Всем известны
также лишайниковые пустоши в тундре, где почва на протяжении тысяч
километров покрыта кустистыми лишайниками, называемыми «оленьим
мхом» (виды рода Кладония) и «исландским мхом» (виды рода Цетрария).
Некоторые кустистые лишайники, растущие на стволах и ветвях де¬
ревьев, не поднимаются над субстратом, а наоборот, свисают с него. Такие
«вислянки», «бородатки» (рис. 36 и) обычны в хвойных и лиственных лесах.
Таким образом, лишайники по своему внешнему виду не имеют ни¬
чего общего ни с грибами, ни с водорослями, а больше всего напомина¬
ют мхи (особенно кустистые и листоватые лишайники). Поэтому многие
их виды называются «белый мох», «олений мох» и т. п. Но, в отличие
от мхов, лишайники никогда не бывают окрашены в травянисто-зеле¬
ный цвет. Окраска талломов лишайника может быть палево-коричневой,
почти белой, ярко-желтой, серо-зеленой, даже черной, но никогда не яр-
ко-зеленой, так как цвета эти обусловлены пигментами, которые образует
гриб, а не водорослевые клетки. И только в период беспрерывных дождей,
когда внутри лишайника происходит массовое размножение водорослей,
талломы зеленеют.
148
Глава 6. Лишайники: два в одном
Итак, разные виды лишайников различаются, во-первых, строением
таллома, то есть бывают накипными, листоватыми, кустистыми, и, во-вто¬
рых, субстратной приуроченностью, то есть заселением камней, почвы,
коры деревьев, оголенной древесины и т. п. Более того, щелочные кар¬
бонатные породы и нейтральные (гранит, кварц) заселяют разные виды
лишайников.
§ 4. А что у них внутри?
У самых примитивных по строению накипных лишайников и внутри
все примитивно: беспорядочно расположенные грибные гифы оплетают
беспорядочно расположенные клетки или цепочки клеток водорослей.
Устроенные так талломы называют гомеомерными. Уже некоторые накип¬
ные, а также все листоватые и кустистые лишайники имеют гетеромерные
талломы, разделенные на четко различающиеся слои. Наружный слой, на¬
зываемый коровым, образован параллельными рядами плотно прижатых
друг к другу пигментированных гиф гриба, которые к тому же выделя¬
ют на поверхность клейкие полисахариды. Под корой располагается зона
фотобионта — слой таллома, в котором находятся водоросли, а далее —
сердцевина, образованная бесцветными, не столь плотными, как в коро-
вом слое, и не упорядоченными гифами гриба. У листоватых лишайников
(но не у всех) таллом снизу ограничен нижней корой, из которой вы¬
ходят пучки гиф, прикрепляющих таллом к субстрату. Интересно, что
гифы корового слоя гидрофильны, то есть смачиваются водой, и выделя¬
ют на поверхность разбухающие в воде полисахариды, а гифы сердцевины,
наоборот, гидрофобны, то есть водой не смачиваются. На их поверхности
кристаллизуются гидрофобные белки — гидрофобины. Благодаря таким
свойствам гифы мицелия, слагающие таллом, не слипаются, между ними
сохраняются воздушные полости, обеспечивающие воздухообмен внутри
лишайника, поступление воды и углекислого газа.
Жизнь в лишайнике наложила существенные черты на морфологию,
физиологию и обмен веществ как гриба, так и водоросли. В клетках грибов
синтезируются разнообразные вторичные метаболиты, которые живущие в
свободном состоянии родственники лихенизированных грибов не образуют.
Это жирные кислоты, лактоны, поликетиды, антрахиноны, фенилпропано-
иды и другие производные фенолов. Они обусловливают окраску, защи¬
щающую слой водорослей от действия прямых солнечных лучей, а также
обладают антибиотическими свойствами и, тем самым, защищают лишай¬
ники от заражения различными микроорганизмами. А недавно у лишай¬
ников были обнаружены и присущие многим плесневым грибам микоток¬
сины, описанные в главе 4, которые, по-видимому, также имеют защитные
функции. Однако, как и у других организмов, защитные свойства лишай¬
ников относительны; обнаружен ряд видов грибов, специализирующихся
на поражении лишайников, их так и называют: лихенофильные грибы.
§ 5. Как они размножаются
149
Изменились строение и функции отдельных групп гиф в мицелии ли-
хенизированных грибов. В зоне развития фотобионта образуются специ¬
альные движущие гифы гриба. Они окружают клетку водоросли и, вслед¬
ствие интенсивного деления клеток, образуют вокруг нее пучок, сдавли¬
вающий водоросль. После это начинается интенсивное деление мицелия
в направлении периферии таллома, в. результате чего гифы, образующие
сердцевину таллома, раздвигаются, и клетка водоросли помещается в то
место, где условия для фотосинтеза наиболее благоприятны. То есть внутри
этого неподвижного, медленно растущего, полумертвого на вид организма
идет непрерывная работа — веточки мицелия ищут водорослевые клетки
и тащат их куда надо.
§ 5. Как они размножаются
Водоросли внутри лишайника коренным образом изменили способы
размножения. Они утратили половой процесс, а также потеряли способ¬
ность образовывать зооспоры, что характерно для большинства живущих
в воде водорослей. В самом деле, что делать зооспорам внутри лишайни¬
ка? Размножаются они или делением клеток, или же внутри материнской
клетки образуется несколько дочерних клеток, похожих на материнскую,
только маленьких, называемых автоспорами.
Грибы в лишайнике, как и свободно живущие аскомицеты, образуют
на поверхности таллома плодовые тела, в которых формируются сумки
с аскоспорами, а у некоторых видов и споры бесполого размножения —
конидии. Однако в отличие от водорослей, автоспоры которых не поки¬
дают лишайник, споры грибов, выброшенные из таллома на какой-либо
субстрат, прорастут, но, при отсутствии рядом лежащих и подходящих
для образования нового таллома водорослей, погибнут. Правда, пророст¬
ки их спор имеют приспособления для поиска водорослей. Боковые ве¬
точки их проростков становятся тонкими и длинными, их перегородки
образуются только у окончания веточки. Они служат для поиска клеток
водоросли и называются ищущими. После того, как ищущая гифа най¬
дет водорослевую клетку, она превращается в охватывающую — образует
боковые ответвления, которые охватывают клетку, оплетают ее, начиная
формирование нового слоевища лишайника. Но, конечно, такой способ
размножения лишайника ненадежен. Ведь ищущая гифа может и не най¬
ти подходящую для формирования лишайника клетку водоросли. И тогда
проросток погибнет. К счастью, многие лишайники придумали и иные
способы размножения. В отдельных участках таллома водоросли начи¬
нают бурно размножаться. Это вызывает раздражение микобионта, гифы
которого в зоне деления водорослей начинают расти, сильно ветвиться,
так что веточки мицелия оплетают делящиеся клетки водоросли и ока¬
зывают сильное давление на коровой слой. В результаты этого в коровом
150
Глава 6. Лишайники: два в одном
слое возникаю разрывы, обнажающие порошок соредий — клеток водо¬
росли, оплетенных гифами гриба. Соредии разлетаются и образуют новые
талломы лишайников. У некоторых видов лишайников на поверхности
таллома образуются не соредии, а изидии, при образовании которых клет¬
ки корового слоя не разрываются, а выпячиваются, образуя небольшие
выросты, подобные бородавочкам. Отрываясь от таллома, они разносятся
ветром и служат началом для образования новых лишайников.
§6. Как они питаются и где живут
Кухня, в которой готовится пища для лишайников, — фотобионты.
Фотосинтетическая деятельность водорослей обеспечивает поступление
в лишайник органического углерода, а если в качестве первичного или вто¬
ричного (в цефалодиях) фотобионта в лишайнике живут цианобактерии,
то они умеют еще и связывать атмосферный азот, превращая его в амино¬
кислоты. Какими же способами гриб отбирает эти вещества у водоросли?
Грибы-паразиты высших растений или убивают клетки своих хозяев фер¬
ментами и токсинами и затем всасывают содержимое (некротрофное пита¬
ние), или, пробивая клеточную стенку, образуют в клетке хозяина вырост
мицелия — гаусторий, который всасывает содержимое живой клетки (био-
трофное питание). В последнем случае клетка растения тоже погибает, хотя
и не так быстро, как в первом. Для лишайника такой грубый способ отъе¬
ма неприемлем. Водоросли в талломе лишайника должны по возможности
не умирать, а жить и размножаться. Поэтому только самые примитивные
лишайники образуют мелкие внутриклеточные гаустории, разрушающие
клеточную стенку фотобионта. У большинства видов лишайников гаусто¬
рии не образуются вообще и питание осуществляется поглощением выде¬
лений из клетки фотобионта. У некоторых видов расширенное окончание
гифы вступает в контакт со стенкой водоросли, в результате чего стенка
водоросли вдавливается, но не разрушается. Лихенизированные грибы по¬
ступают хитрее, чем фитопатогенные. Они вынуждают клетки водорослей
не накапливать продукты фотосинтеза в качестве внутриклеточных запас¬
ных веществ — крахмала, липидных капель, цианофициновых зерен, как
это делают свободно живущие водоросли, а выбрасывать их в окружаю¬
щую среду. Живущие в свободном состоянии водоросли теряют в окружа¬
ющую среду около 10 % продуктов фотосинтеза. Водоросли в лишайнике
теряют до 80 % углерода из соединений, созданных в результате фотосин¬
теза. Цианобактерии выделяют в полости лишайника водорастворимые
полимеры глюкозы — глюканы, которые разрушаются ферментами гриба
и всасываются в его клетки. А зеленые водоросли теряют углерод в фор¬
ме сахароспиртов — рибита, сорбита или эритрита. А гриб превращает
растворимые мобильные углеводы, выделяемые фотобионтом, в манит,
гликоген и трегалозу, то есть вещества, отсутствующие в водорослевых
клетках. Это приводит к падению в пространстве, окружающем клетки
§ 6. Как они питаются и где живут
151
водорослей, концентрации углеводов и выбросу новой их порции, кото¬
рая также быстро уходит в черную дыру мицелия. То же самое происходит
и с аммонийным азотом, производимым цианобактериями.
Итак, водоросли в лишайнике используют столько продуктов фото¬
синтеза, сколько необходимо для жизнедеятельности и размножения; из¬
быток не откладывается про запас, а отдается грибу. Гриб, в свою очередь,
защищает своего симбионта от неблагоприятных биотических и абиоти¬
ческих воздействий и снабжает его водой и минеральными веществами,
которые мицелий поглощает из воздушной среды и субстратов, на кото¬
рых растет лишайник. Такой пищевой баланс создает высокую экономию
в расходовании продуктов питания. Лишайники — самые экономные ор¬
ганизмы на земле. Они могут быть отличной иллюстрацией афоризма:
«Счастлив не тот, кто много имеет, а тот, кто в малом нуждается». Содер¬
жание хлорофилла на единицу массы лишайника примерно в 5 раз меньше,
чем у высших растений, и, соответственно, менее активен процесс фото¬
синтеза. Поэтому лишайники, с одной стороны, очень медленно растут,
всего на несколько миллиметров в год, а с другой — могут существовать
в экстремальных условиях, там, где большинство других организмов су¬
ществовать не могут. Они заселяют голые скалы, пустыни, приполярные
области. Интересно в связи с этим сравнить места обитания лишайни¬
ков и высших растений. В тропиках, на острове Ява, описано 600 видов
лишайников и 9600 видов высших растений, то есть соотношение лишай¬
ников и растений (лишайниковый коэффициент, ЛК) составляет 0,06.
Это и понятно: условия влажных тропиков очень благоприятны для суще¬
ствования растений, которые не оставляют места для медленно растущих
лишайников. В умеренной зоне, в Германии, число видов лишайников со¬
ставляет почти половину (ЛК = 0,48) от числа видов растений: 1200 видов
лишайников и 2600 растений. Еще севернее, в Исландии, зафиксировано
одинаковое число видов лишайников и растений — по 600. На аркти¬
ческом острове Шпицберген живут 450 видов лишайников и только 130
видов растений (ЛК равен 3,5), а в Антарктиде описано 350 видов лишай¬
ников и всего 2 вида растений, то есть ЛК составил 175. То есть по мере
продвижения от экватора к полюсам лишайниковый коэффициент увели¬
чивается почти в 30 000 раз!
Как видно, лишайники приспособились к существованию в очень жест¬
ких климатических условиях и при очень скудном снабжении пищевыми
ресурсами. Однако приспособиться к отходам человеческой деятельности
они, в отличие от многих других организмов, оказались не способны.
Кислые дожди, наличие в воздухе и в водяных парах окислов азота, озона,
фенолов, ионов тяжелых металлов, органических соединений и прочих
ксенобиотиков очень быстро приводят талломы лишайников к гибели.
Большие города, промышленные зоны превратились в лишайниковые пу¬
стыни. Предполагают, что есть несколько факторов, способствующих вы¬
сокой токсичности поллютантов (загрязнителей воздуха) в окружающей
152
Глава 6. Лишайники: два в одном
среде для лишайников, причем наложение этих факторов друг на друга
усиливает чувствительность лишайников. Основные факторы следующие:
1. Лишайники — долгожители (индивидуальный таллом может вегети¬
ровать 1,5-2 тысячи лет). Поэтому они подвергаются вредным воз¬
действиям круглый год в течение длительного времени.
2. Медленное отмирание таллома не позволяет освобождаться от ток¬
сических соединений в нем.
3. У лишайников отсутствуют кутикула и устьица, они, как и все грибы,
впитывают поллютанты всем телом.
4. При подсыхании таллома (а лишайники могут выдерживать длитель¬
ное высушивание без потери жизнеспособности) концентрация ток¬
сических веществ, попавших в него, увеличивается до летальной.
§ 7. Какую роль играют лишайники
в биосфере и в биоценозах
Тундряные кустистые лишайники являются основной пищей для тра¬
воядных животных, прежде всего северных оленей, особенно зимой, когда
никаких других растений в тундре просто нет. Миграция этих животных
по тундре вызвана поисками богатых лишайниками пастбищ. Причем
олени не съедают весь кустик до грунта, они осторожно объедают толь¬
ко верхние его веточки. Такое питание — замечательное приспособление
животного; ведь вследствие медленного роста лишайников, объеденное
до почвы пастбище на долгие годы оставалась бы пустыней. Поэтому,
несмотря на то, что такой экономный способ кормления вызывает не¬
обходимость обходить большие участки тундры для насыщения, стра¬
тегически он является единственно верным. Вспоминаются наблюдения
писателя Сент-Экзюпери над поведением африканской лисицы фенека,
которая питалась улитками, живущими на редко расположенных в пусты¬
не кустарниках. Фенек никогда не съедал всех улиток, сидящих на кусте,
а оставлял часть популяции нетронутой, хотя из-за этого для насыщения
ему приходилось обходить огромную территорию. Если бы он съел всех
улиток, куст перестал бы служить ему кормовой базой, а при таком спо¬
собе питания оставшиеся улитки успевали размножиться до следующего
посещения. Кроме северных оленей, ягели потребляют и другие дикие
травоядные животные, живущие в тундре, а также домашний скот — сви¬
ньи, коровы, овцы. А маралы объедают талломы кустистого лишайника
уснея, которые, как спутавшиеся бороды, свисают с ветвей деревьев. По¬
этому в Сибири этот лишайник называют маральим мхом.
Известный микробиолог Г. А. Заварзин считал, что лишайники игра¬
ли решающую роль в первичном освоении суши животными. Дело в том,
что по данным палеонтологии животные вышли на сушу значительно
раньше, чем растения, и, поскольку животные не способны создавать
органические соединения из неорганических, для их существования тре-
§ 7. Какую роль играют лишайники в биосфере и в биоценозах 153
буется внешний источник питания. Наличие на суше цианобактериальных
матов, состоящих из фотосинтезирующих синезеленых водорослей (ци¬
анобактерий) и нефотосинтезирующих бактерий, количественно не было
способно обеспечить первичную наземную фауну питанием, во-первых,
потому, что среди первичных животных было немало достаточно крупных
экземпляров, требующих большого количества пищи, а, во-вторых, суще¬
ствует такое понятие, как пищевая пирамида, в которой первичное звено
пищевой цепи — растения продуценты — должно иметь значительно
большую биомассу, чем вторичное звено — травоядные животные, а по¬
следние — большую биомассу, чем третичное звено — хищные животные,
и т. д. Поэтому, по мнению Г. А. Заварзина, первичными продуцентами
на земле до Девонского периода, то есть полмиллиарда лет назад, когда
суша еще не была заселена растениями, были лишайники. Однако, гео¬
логических пород этого периода, в которых были бы идентифицированы
лишайники, почти не найдено. В Южном Китае, в фосфоритах, возраст
которых датируется 600 млн лет, обнаружены отпечатки, идентифициро¬
ванные как лишайник. Они состоят из тонких гиф гриба (предположи¬
тельно, гломеромицета) и одноклеточного фотобионта. Наиболее древний
отпечаток несомненного лишайника относится к Девонскому периоду, то
есть около 400 млн лет назад. Этот лишайник, названный винфернацией,
состоял из гиф микобионта (предположительно — зигомицета), не имею¬
щих перегородок, и нитчатых цианобактерий. Современные лишайники
образованы сумчатыми и базидиальными грибами, которые появились
на суше в одно время с растениями. Но есть один организм, ныне живу¬
щее живое ископаемое — геосифон. Это небольшой, размером от одного
до нескольких миллиметров пузырек, грушевидной формы, с заякоренны¬
ми в почву гифами. Внутри пузыря находятся не только присущие грибу
органеллы, но и нити цианобактерии. Геосифон относится к гломеромице-
там — группе гораздо более древней, чем сумчатые и базидиальные грибы.
Не исключено, что предки геосифона были более крупными, чем ныне
живущие экземпляры, как это характерно для многих древних животных.
Возможно, лишайниковые ассоциации типа винфренации или геосифона,
сформированные древними грибами зигомицетами и гломеромицетами,
занимали в палеозое большие площади, формировали, подобно совре¬
менным тундровым экосистемам, лишайниковые пустоши, но с выходом
на сушу тканевых растений грибы нашли более благоприятные условия
существования колонизацией крупных, обеспечивающих лучшую защиту
и питание растений. Как реликт от тех времен, когда на суше не бы¬
ло иных фотосинтезирующих организмов, кроме цианобактерий, остался
лишь сильно редуцированный геосифон.
Описанный сценарий, конечно, гипотетичен, а вот почвообразова¬
тельная деятельность лишайников, говоря стилем Остапа Бендера, — ме¬
дицинский факт. Они первыми колонизируют субстраты, непригодные
для жизни других организмов, например, скалы. Гифы лихенизированного
154
Глава 6. Лишайники: два в одном
гриба выделяют в субстрат активные соединения, такие как органические
кислоты, и вызывают локальное размягчение горных пород. По микро¬
трещинкам гифы гриба проникают внутрь породы, разрушая их. А отми¬
рающие талломы лишайников создают запас органических соединений,
на которых поселяются сначала микроорганизмы, а затем и растения.
§ 8. Возможные пути использования лишайников
Когда евреи, воодушевленные призывами пророка
использование Моисея, бежали из Египта и много лет бродили по пу¬
стыне в поисках земли обетованной, они начали уми¬
рать от голода. Но однажды, когда голод стал невыносимым, они обнару¬
жили на земле непонятный налет, похожий на крупу. Впервые увидев его,
люди удивленно стали спрашивать друг у друга: «Манна?» (В переводе
на русский — «Что это?») Манна (так и назвали этот налет) оказалась
съедобной, ее мололи, толкли в ступах, варили в горшках или пекли
из нее лепешки. Многие годы странствий, говорится в книге Библии
«Исход», каждое утро люди собирали принесенную ветром или упавшую
с неба манну, «небесный хлеб», что помогало им прокормиться в пусты¬
не. Описанная в Ветхом завете «Манна небесная», как полагают, — это
лишайник аспицилия, называемый за столь важную роль, сыгранную им
в истории, лишайниковой манной. Он обитает в пустынях, степях, за¬
сушливых горных областях. Таллом аспицилии, имеющий вид серых или
бурых комочков, может подхватываться ветром и переноситься на далекие
расстояния, после чего они могут оседать в низменных местах, образуя
наносы. Конечно, никакой лишайник не мог бы прокормить целый на¬
род, кочующий по пустыне, но Библия полна и не такими чудесами. Ведь
прокормил же через тысячу лет после этого события Иисус Христос тремя
хлебами тысячу страждущих. И если это чудо имело место, то лишайнику
мы обязаны возникновением трех великих мировых религий — иудаизма,
христианства и ислама. Кроме упомянутого в Библии эпизода, пищевое
использование лишайников весьма ограничено; в некоторых националь¬
ных кухнях их используют как добавки к мучным изделиям, в качестве
ароматизаторов, для приготовления соусов и т. д.
Использование ^Же ^ыло сказано» лихенизированные гри-
биологически бы образуют большое число вторичных метаболи-
активных веществ тов “ лишайниковых веществ, многие из которых
обладают разнообразными и хозяйственно важными
свойствами. Многие лишайниковые вещества оказались активными анти¬
биотиками, подавляющими рост стафилококков и других болезнетворных
бактерий. На их основе созданы даже лекарственные препараты, продаю¬
щиеся в аптеках. Таков германский препарат «Эвозин», который получают
из растущего на стволах деревьев листоватого лишайника эвернии сли¬
вовой, называемого также «дубовый мох». Эвозин рекомендуют и для
§ 8. Возможные пути использования лишайников
155
лечения кожных заболеваний, как неинфекционных (фурункулез, вол¬
чанка), так и инфекционных, вызванных грибами-дерматофитами из рода
трихофитон. Смесь эвозина со стрептомицином рекомендуют для лечения
туберкулеза. А в Японии из кустистого лишайника уснея, таллом которого
свисает с ветвей, получен антибиотический препарат «Уснин». Натриевая
соль усниновой кислоты под названием препарата «Бинан» была создана
в Советском Союзе и применялась в конце прошлого века. Некоторые
лишайниковые вещества обладают также противоопухолевым действием.
Изучение лишайников, растущих на деревьях, привело к любопытному на¬
блюдению: древесина под колониями лишайников обычно не загнивает.
Из этого наблюдения были сделаны выводы о возможности применен™
усниновой и физодиевой кислот для защиты древесных построек от дре¬
воразрушающих грибов. Показаны также более широкие возможности ис¬
пользования лишайниковых веществ, в частности, в фитопатологии — для
защиты растений от бактериальных и вирусных заболеваний. А содержа¬
ние в лишайниках ароматических веществ и эфирных масел делает их весь¬
ма перспективными для использования в парфюмерной промышленности.
Все это замечательно, если бы не одно но. Даже заготовки многих
видов лекарственных растений вызывают серьезные возражения у бота¬
ников, так как могут подорвать нормальную численность их популяций,
привести к переводу этих видов в категории «редких» и даже «исчезаю¬
щих». Поэтому были разработаны агротехнические методы искусственного
выращивания и даже получения биомассы клеток и тканей в фермен¬
терах для редких растений, обладающих важными фармакологическими
свойствами, таких, например, как женьшень. Что же говорить о таких
медленно растущих организмах, как лишайники. Заготовки сырья в объ¬
емах, которые оправдали бы затраты на заготовку, получение препарата,
его рекламу и продажу, однозначно в короткие сроки подорвали бы чис¬
ленность их популяций, ведь даже олени так не поступают. А на грядке,
как огурцы, их не произведешь. Поэтому лишайники — объекты будущих
биотехнологий. А пока — в ряде лабораторий ведутся исследования их
физиологии. Беда в том, что грибы, составляющие лишайник, не могут
расти в искусственных условиях. В лаборатории американского ученого
Ахмаджана впервые удалось разделить лишайник на отдельные компо¬
ненты и в одной пробирке выращивать гриб, а в другой — водоросль.
Гриб рос чрезвычайно медленно, но все-таки рос. После этого, совмещая
отдельные компоненты, Ахмаджан вновь ресинтезировал целый лишай¬
ник. Эти исследования открыли путь для работ в области биотехнологии
лишайников. Если водоросли так влияют на обмен веществ грибов, что
они начинают образовывать несвойственные им биологически активные
вещества, то, может быть, надо создавать искусственные ассоциации раз¬
личных видов грибов и водорослей, включая биотехнологически удобные
виды грибов (быстро растущие на дешевых питательных средах) и живу¬
щие в составе лишайников водоросли, и подбирать такие компоненты,
156
Глава 6. Лишайники: два в одном
которые приводили бы к получению полезных продуктов. Таковы совре¬
менные направления в экспериментальной лихенологии. Но лишайники
могут использоваться не только в биотехнологиях, но и для других на¬
правлений научной и практической деятельности.
_ „ Уже говорилось, что лишайники чрезвычайно
0 Лихеноиндикация
^ чувствительны к присутствию поллютантов в воздухе.
Специальные исследования показывают, что чувствительность разных ви¬
дов лишайников не одинакова. При повышении в воздухе концентрации
вредных веществ первыми исчезают кустистые лишайники, затем — ли¬
стоватые и последними — накипные. Лишайники, живущие на деревьях,
более чувствительны, чем живущие на почве и на камнях. Наконец, име¬
ются виды, имеющие повышенную чувствительность к отдельным химиче¬
ским загрязнителям — окисиду серы, окислам азотам, соединения фтора и
другим. Эти данные предложено использовать для оценки степени загряз¬
ненности среды обитания людей. Лихеноиндикация окружающей среды —
важное подспорье в работах по экологическому мониторингу. Во многих
крупных городах мира, включая Москву, созданы карты, на которых отме¬
чены районы, имеющие разную степень загрязненности: от лишайниковых
пустынь, где лишайники вообще не встречаются, до чистых зон с высо¬
ким видовым разнообразием и численностью лишайников. Для создания
таких карт производят маршрутные обследование территорий с определе¬
нием видового состава лишайников и оценкой обилия отдельных видов.
„ Лишайники — медленно растущие организмы.
0 Лихенометрия ~
^ г Ежегодный прирост диаметра слоевища накипных и
листоватых лишайников у разных видов колеблется от нескольких мил¬
лиметров до десятых долей миллиметра. К тому же они долгожители;
возраст отдельных талломов может достигать нескольких тысяч лет. Эти
их свойства используют геологи и археологи для установления возраста
пород. Метод получил название лихенометрии и заключается в измере¬
нии диаметров наиболее крупных слоевищ определенного вида лишайни¬
ка и установлении его возраста. Чаще всего для этих целей используют
долгоживущий и широко распространенный на каменистых субстратах
накипный лишайник ризокарпон. Метод этот только кажется простым.
На самом деле скорость роста лишайника может зависеть не только от его
генетической природы, но и от ряда привходящих факторов, например,
с северной или южной стороны он находится, как менялась температура
и влажность в разные годы и прочее. Кроме того, скорость роста лишайни¬
ка, как и других организмов, зависит от возраста: молодые талломы растут
значительно быстрее, чем старые. Поэтому созданы сложные системы
уравнений, устанавливающих более объективные связи между диаметром
талломов и возрастом лишайников. Тем не менее, метод лихенометрии
очень популярен у геологов, географов и даже археологов. С его помо¬
щью, в частности, был уточнен возраст каменных идолов острова Пасхи.
Глава 7
Грибы и животные
В предыдущих главах было рассказано о древней¬
ших связях грибов с растениями, как главными источ¬
никами питания. Однако не все грибы вегетарианцы.
Многочисленные виды грибов в своем питании связа¬
ны с различными животными, начиная от простейших
одноклеточных амеб (есть группа грибов, так и называ¬
емая: амебофаговые грибы) и кончая млекопитающими.
Поскольку вся книга задумывалась как отдельные очер¬
ки, а не систематическое изложение предмета, в дан¬
ной главе будет рассказано о взаимоотношениях грибов
лишь с несколькими группами животных.
§ 1. Грибы и нематоды
Великое множество круглых червей нематод, ви¬
довое разнообразие которых изучено столь же недоста¬
точно, как и грибов, населяет воды, почву, растения,
то есть субстраты, в которых живут и грибы. Их встре¬
чи происходит постоянно, что не могло не привести
к противостояниям, которые завершились однозначной
победой грибов. В отличие от зоофаговых и фитофаго¬
вых нематод, которые широко распространены и при¬
носят значительный вред растениям и животным, мы
почти ничего не знаем о фунгифаговых. Зато хорошо
известные нематофаговые грибы — паразиты и хищни¬
ки нематод. Паразиты нематод известны во всех боль¬
ших группах (отделах, классах) грибов и псевдогрибов.
Древние псевдогрибы оомицеты из рода гаптоглос-
са образуют зооспоры, которые садятся на нематоду, по¬
крываются оболочкой и образуют стреляющую клетку,
из которой с силой выбрасывается острый гарпун, про¬
бивающий покровы нематоды. Через образовавшуюся
ранку паразит прорастает в тело нематоды и питается
ее содержимым. Верхом на нематодах гаптоглоссовые
158
Глава 7. Грибы и животные
оомицеты попали из моря на сушу, так
как их виды поражают как морских,
так и почвообитающих нематод. По¬
скольку наземные нематоды стали по¬
вреждать корни растений, ученые счи¬
тают, что оомицеты перешли из не¬
матод в растения и после некоторых
мутационных изменений стали их па¬
разитами.
Интересны и своеобразны также
орудия нападения на нематод насто¬
ящих несовершенных грибов, которые
называют нематофаговыми грибами.
Они живут в почве и могут, вроде бы,
прекрасно обходиться без нематод,
питаясь растительными остатками и
почвенным органическим материалом.
Но, как и многие люди-вегетарианцы,
они только делают вид, что не нуж¬
даются в мясе. На самом деле, стоит
только нематоде проползти мимо это¬
го мелкого хищника, как она оказыва¬
ется в его лапах. У некоторых видов нематофаговых грибов орудием охоты
являются капканы, представляющие собой свернутые в кольцо гифы ми¬
целия. Если нематода случайно заползет внутрь такого ловчего кольца,
то кольцо от контакта с ее телом сжимается и начинает выделять фер¬
менты, разрушающие покровы нематоды, которые постепенно растворяют
все ее тело (рис. 37). Другие грибы охотятся на нематод с помощью раз¬
нообразных выростов мицелия — «гарпунов», липких шариков, «ловчих
сетей» и прочих. При контакте с телом проползающей мимо нематоды,
они прилипают к ней и выделяют пищеварительные ферменты. Орудия
нападения на нематод имеются и у некоторых видов древоразрушающих
базидиомицетов. Они живут в древесине, где полно углерода (надо только
иметь ферменты для его усвоения), но почти нет азота. Поэтому заползшая
в зону роста мицелия нематода — это лакомый кусочек, «конфетка» для
гриба. У всем известной вешенки, мицелий которой растет в древесине,
а съедобные плодовые тела продают в магазинах, на мицелии образуются
выросты, похожие на электрические лампочки. Долгое время не было по¬
нятно, зачем они нужны грибу, так как они не отваливаются бт мицелия
и не прорастают, как споры. Затем выяснили, что лампообразные выросты
покрыты клейкой слизью, которая прилипает к телу проползающих рядом
нематод и притягивает их к мицелию.
Рис. 37. Нематофаговый гриб, поймав¬
ший в свои ловчие кольца нематоду
(З.Э. Беккер, 1963)
§ 2. Грибы и насекомые
159
§2. Грибы и насекомые
Взаимоотношения грибов с насекомыми очень сложны, так как, во-
первых, в тесном контакте с насекомыми живет огромное число видов гри¬
бов и псевдогрибов, от примитивнейших одноклеточных микроспоридий
до высоко организованных видов сумчатых и базидиальных грибов, и, во-
вторых, между грибами и насекомыми наблюдается широчайший спектр
взаимодействий — от жестокого паразитизма до кооперации и симбио¬
зов, причем между разными формами существует множество переходов.
Начнем с паразитов.
Целый класс грибов получил название энтомофто-
ровые (насекомоядные) грибы, потому что болыиин- ак
ство его представителей — паразиты насекомых. Эти
грибы поражают насекомых из разных отрядов — и бабочек, и мух, и са¬
ранчу, и тлей, и цикад, причем поражение всегда приводит к быстрой
(в течение нескольких дней) гибели насекомого. На его теле, которое
меняет окраску и во влажных условиях раздувается, а в сухих — высы¬
хает и мумифицируется, через разрывы кутикулы высовываются щетинки
конидиеносцев, которые, подобно пушечным залпам, выстреливают ко¬
нидиями. На теле одного пораженного насекомого может образоваться
до нескольких миллионов конидий. Густой белый пушок, образованный
слоем конидий вокруг мертвого насекомого, — характерный признак по¬
ражения энтомофторовыми грибами (рис. 38 а). Конечно, стрельба на рас¬
стояние нескольких сантиметров может достичь цели только в том случае,
если насекомые находятся рядом друг с другом. Но эти грибы имеют
приспособления для повышения эффективности заражения. Их конидии,
не достигшие тела насекомого, прорастают не гифой, а новым, коротким
конидиеносцем, который выстреливает вторичную конидию. Если и она
не попала в цель, образуется третичная конидия, и такие двойной, трой-
Рис. 38. Энтомофторовые грибы: а) зараженная муха с окружающим ее белым
налетом из разбросанных спор (Carlig M.J. et al., 2006); б) два зараженных
кузнечика, вцепившиеся в стебель (Pell J.K. et al., 2001)
160
Глава 7. Грибы и животные
ной, четверной и так далее прыжки продолжаются до тех пор, пока кони¬
дия не попадет на тело насекомого или пока не истощится полностью запас
пластических веществ в конидии. Но и этого мало. Находящийся в теле
насекомого гриб меняет его поведение. Зараженные насекомые становятся
более подвижными, разлетаются и тем самым распространяют инфекци¬
онное начало. Пораженные особи саранчи, как и различных видов мух, за¬
бираются на верхушки растений, плотно закрепляются там судорожно све¬
денными ногами, и поднимают вверх голову (рис. 38 б). Такая поза привле¬
кает к ним других насекомых, которые инфицируются конидиями. Из тела
тлей и гусениц выдвигаются специальные гифы гриба (ризоиды), кото¬
рые прикрепляют пораженных насекомых к субстрату. При этом гусеницы
приобретают характерную позу, свешиваясь вниз головой. В теле поражен¬
ных насекомых обнаружен новый белок, который влияет на их поведение.
Попав на поверхность тела насекомого, конидия прорастает, и ее
росток начинает выделять ферменты, растворяющиеся кутикулярный
покров. Через образовавшееся отверстие гифа проникает внутрь и рас¬
пространяется по всему телу. Затем она распадается на отдельные участ¬
ки — гифальные тела, из которых формируются высовывающиеся наружу
конидиеносцы. Конидии, попавшие на поверхность почвы, сохраняют
способность к отбрасыванию вторичных конидий в зависимости от тем¬
пературы от нескольких дней до нескольких месяцев. А в теле погибшего
насекомого образуются покоящиеся споры, покрытые толстыми оболоч¬
ками, которые сохраняют жизнеспособность длительное время и являются
источниками первичного (весеннего) заражения насекомых.
Многочисленные виды сумчатых и несовершенных грибов также па¬
разитируют на насекомых. Особенно интересны среди них спорыньевые
грибы. Я уже рассказывал об этих грибах, которые образуют высокоток¬
сичные алкалоиды и вызывают заболевания злаковых культур, а внутри
некоторых видов кормовых трав растут эндофитно и приносят своим хозя¬
евам больше пользы, чем вреда. Но удивительные свойства спорыньевых
грибов этим не исчерпываются.
В старинном китайском трактате, написанном в VII веке, упомина¬
ется странное существо, обитающее в горах Тибета, под названием «Дун
чунг ся цао», что в переводе означает «зимой — червяк, летом — трава».
В мае-июне, когда земля в горах еще не освободилась полностью от снега,
в Тибетское высокогорье направлялись целые семьи с нехитрым скарбом
на поиски червяка-травы, обладающего, как считалось, колоссальными
целебными свойствами. Такие экспедиции направляются в Тибет и сей¬
час. Исследования, проведенные в США, Японии, Южной Корее и других
странах, подтвердили высокий терапевтический эффект этих странных
существ, описанный в старинных китайских книгах. В 1999 г. один кило¬
грамм воздушно-сухого материала «червяка-травы» стоил в Китае $1250,
а в США - $2200.
§ 2. Грибы и насекомые
161
Что же представляет собой это удивительное существо? Оно выглядит
как толстая оранжевая или серая проволочка, длиной несколько сантимет¬
ров, высовывающаяся из почвы. Если осторожно раскопать землю вокруг,
можно обнаружить, что она выходит из трупа насекомого, находящегося
в почве. «Проволочку» сформировал гриб, который заразил насекомое,
образовал в его теле гифы и высосал все содержимое, оставив лишь обо¬
лочку тела, а затем выдвинул на поверхность вырост. Как и описанные
ранее головчатые выросты, формирующиеся на рожках-склероциях его
родственницы спорыньи, китайский «червяк» представляет собой плотное
сплетение гиф — строму, верхняя часть которой, подобно миниатюрно¬
му наперстку, сплошь утыкана углублениями — отверстиями плодовых
тел, перитециев. Разные виды насекомоядных грибов из рода Кордицепс
и близких ему родов образуют на теле насекомого стромы, одиночные или
многочисленные, короткие толстые или длинные нитевидные (рис. 25 д).
Разлетающиеся аскоспоры, попадая на тело насекомого, прилипают к не¬
му, прорастают, разрушают кутикулу с помощью выделяемых проростком
ферментов и внедряются, внутрь. Многие виды кордицепсовых грибов
образуют и конидиальные стадии, так что заражение новых насекомых
возможно проростками конидий.
Энтомопатогенные грибы являются важным фактором, регулирую¬
щим численность популяций массовых видов насекомых. Процент боль¬
ных особей многих видов насекомых в отдельные годы и в некоторых по¬
пуляциях может достигать 50 и даже 90 %. Как рассуждал большой знаток
энтомопатогенных грибов Б. А. Борисов: «Если бы в какой-то местности
вспыхнула страшная инфекция (чумы, сибирской язвы и т. п') и унесла
жизни даже 1-2 человек из каждых ста, то это, несомненно, вызвало бы
панический ужас: „эпидемия". Чем же тогда назвать гибель порядка 40%
белокрылок на растениях огурца в теплице площадью 3200 кв. м., где
общая численность вредителя достигает 100 млн особей, т. е. на поря¬
док превышает население Москвы?» Этот пример показывает, что мерки,
применяемые при оценке людских эпидемий и эпизоотий насекомых,
различны. Генерация у людей (продолжительность времени от рождения
до деторождения) составляет в среднем более 20 лет, а у многих насеко¬
мых в продолжительность одного летнего сезона укладывается несколько
генераций. У одной пары людей редко бывает более пяти детей, дожи¬
вающих до половозрелого возраста, а потомство насекомых составляет
десятки, сотни и даже тысячи особей. Поэтому массовая гибель какой-
либо изолированной популяции людей от болезни чревата полным ис¬
чезновением этой популяции, что и происходило в истории человечества.
У насекомых же потомство единичных выживших особей после окончания
эпизоотии может быстро восстановить популяцию до исходной числен¬
ности. К тому же эпизоотии насекомых не бывают слишком длительными
и постепенно затухают вследствие многих факторов. Во-первых, уменьше¬
ние численности насекомых-хозяев снижает как количество образуемого
162
Глава 7. Грибы и животные
инфекционного начала, так и вероятность попадания его на живых на¬
секомых; во-вторых, рост численности энтомопатогенного гриба часто
сопровождается ростом численности и его врагов. Так, по наблюдениям
упомянутого выше Б. А. Борисова, в первый год эпизоотии калиново¬
го листоеда (жука, который продырявливает листья калины), вызванной
энтомопатогенным грибом Боверия, у 2 % погибших жуков мицелий Бо-
верии был поражен грибом гиперпаразитом (сумчатый гриб Меланоспора,
паразитирующий на мицелии Боверии), а на третий год пораженными ока¬
зались мицелии из 98 % больных жуков. Поэтому в естественных условиях,
в природе, этомопатогенные грибы, как и другие природные регуляторы
численности насекомых, не могут длительное время держать численность
вредных для сельского хозяйства насекомых на уровне ниже экономиче¬
ского порога вредности. Однако эти грибы давно заинтересовали специа¬
листов по защите растений от вредителей, которые разработали технологи
накопления спор энтомопатогенных грибов и заражения ими популяций
вредных насекомых на полях и в теплицах. Первый, кому пришла голову
такая идея, был наш великий соотечественник И. И. Мечников, который,
работая в Одесском университете, создал лабораторию по размножению
грибов-паразитов насекомых. Сейчас лаборатории, в которых размножают
споры различных энтомопатогенных грибов, работают во многих странах.
А как обстоит дело с лекарственными свойствами кордицепсовых
грибов? Китайцы разработали технологию получения их стром и без насе¬
комых, выращивая на рисе и других природных субстратах, что, конечно,
удешевило стоимость продукции и, главное, несколько остановило варвар¬
ское уничтожение природных популяций этого гриба. Кроме того, было
показано, что не только китайский высокогорный гриб обладает лечебны¬
ми свойствами; полезны стромы и других кордицепсовых грибов, в част¬
ности обычного в средней России кордицепса воинственного, крупные
стромы которого вырастают на пораженных гусеницах. А платить беше¬
ные деньги сомнительным фирмам, продающим грязные препараты кор¬
дицепса под звучными названиями «Императорский гриб» и т. п. не стоит.
Энтомофторовые и кордицепсовые грибы — это жестокие паразиты
насекомых. Зараженные ими организмы обычно погибают. Между тем на
поверхности и даже внутри тела насекомых часто живут разнообразные
грибы, которые практически не влияют на их жизнеспособность и пло¬
довитость. Взаимоотношения подобных грибов с насекомыми имеют са¬
мые разные формы — от слабого паразитизма до симбиозов, приносящих
взаимную пользу партнерам. Иногда подобные взаимоотношения приоб¬
ретают совершенно фантастические формы, как, например, у септоба-
зидиальных грибов —■ родственников паразитов растений, вызывающих
ржавчинные болезни. Септобазидиальные грибы тесно связаны с группой
насекомых, называемых щитовками. Щитовки выбирают удобное местеч¬
ко на стволе или ветке дерева, устраиваются там, погружают свой ротовой
аппарат в растительную клетку и остаются в таком состоянии всю жизнь,
§ 2. Грибы и насекомые
163
посасывая питательные вещества из растительных клеток. На поверхно¬
сти их тела откладывается восковой панцирь-щиток, откуда и возникло
соответствующее название. И только их личинки, которых энтомологи
назвали бродяжками, расползаются из места своего рождения в поисках
новых удобных «диванов», на которых можно, подобно Обломову, прове¬
сти в комфорте остаток жизни. Так вот, базидиоспоры септобазидиальных
грибов, попав на тело щитовки, прорастают, и их ростки внедряются
в насекомое, но не убивают его. Вокруг насекомого образуется мощное
сплетение гиф — многолетний мицелиальный мат, с многочисленными
камерами внутри. Зараженные щитовки под защитой этого плодового тела
гриба дольше живут, чем свободноживущие щитовки, но не размножаются.
Это не беда, так как под защиту мицелиального мата заползают незаражен-
ные бродяжки, способные в дальнейшем к размножению, так что внутри
камер плодового тела может находиться множество щитовок, как заражен¬
ных, так и здоровых. Их детки, выползая из-под плодового тела в поисках
новых мест обитания, разносят на своем теле споры гриба. Как же можно
назвать взаимоотношение септобазидиального гриба и щитовок? Насеко¬
мое высасывает растительный сок. Гифы гриба в его теле забирают часть
питательных веществ на построение мицелиального мата. То есть гриб
паразитирует непосредственно на насекомом и косвенно — на растении,
которым питается насекомое. Но и насекомое в обиде не остается, так
как находит в сплетении гиф своего паразита дополнительную защиту.
Не только грибы едят насекомых, но и некоторые
- Как насекомые
насекомые питаются грибами. Их так и называют: едят грибы
насекомые-микофаги.
Есть виды жуков, которые заражают только крупные плодовые тела
трутовых грибов, растущие на пораженных этими грибами деревьях. Неко¬
торые виды двукрылых насекомых откладывают яйца в ткань шляпочных
(агариковых) грибов. Этих насекомых называют грибными мухами или
грибными комариками. Их личинки делают ходы в шляпках и ножках за¬
раженных грибов. Такие «червивые» грибы очень часто встречаются среди
плодовых тел маслят, рыжиков, сыроежек и многих других видов съедоб¬
ных грибов и «портят кровь» любителям тихой охоты. Но есть виды съе¬
добных грибов, у которых никогда не встречаются «червивые» плодовые
тела. Таковы, например, известные всем грибниками лисички. На Руси их
за это называли «жидовский гриб» (сразу оговорюсь, что в старину на Ру¬
си слово «жидовин» означало не оскорбительную кличку, а обозначение
национальности или религиозной принадлежности; вспомним хотя бы
знаменитое высказывание одного из основателей Киево-Печерской лав¬
ры святого Феодосия Печерского: «Аще ли видишь нага или голодна или
зимою, или бедою одержима, аще ли то будет жидовин, или сарацин, или
болгарин, или еретик, или латинянин, или от всех поганых (то есть языч¬
ник) — всякого помилуй и от беды избави, аще можеши»). Дело в том,
что в священной книге иудеев Торе запрещено есть животных, которые
164
Глава 7. Грибы и животные
не покрыты шерстью, чешуей или перьями. А таковы не только свиньи,
но и личинки многих насекомых, поэтому, из опасения съесть невзначай
«червяка», грибы для правоверного иудея находятся под запретом. А ли¬
сички собирать и есть можно, так как они не червивеют.
Есть насекомые, которые приспособились к питанию дрожжами. Это
виды мелкой плодовой мушки дрозофилы. Все мы сталкивались с эти¬
ми мушками, которые летят на перезрелые, забродившие фрукты и овощи
и в массе размножаются на них. Но питается дрозофила не разлагающейся
мякотью плодов, а размножающимися в мякоти дрожжами. В предыдущих
главах было рассказано, что дрожжи в массе размножаются на сахари¬
стых субстратах. В созревших фруктах и овощах ферменты деполимеразы
разлагают запасенные полисахариды до простых сахаров. Процессы рас¬
щепления усиливаются при заражении плодов фитопатогенными грибами
и бактериями, которые выделяют в зараженную ткань собственные депо¬
лимеразы. Всегда находящиеся на поверхности плодов и листьев (эпифит-
ные) дрожжи начинают питаться в таких плодах и быстро накапливают
большую биомассу. Она-то и привлекает мух. Поскольку и дрозофила,
и дрожжи — это удобные объекты, с помощью которых генетики изуча¬
ют закономерности наследственности и изменчивости, пищевая модель
муха дрозофила — сахаромицетные дрожжи привлекла внимание ученых.
Ее, в частности, детально исследовали сотрудники кафедры генетики Ле¬
нинградского университета под руководством С. Г. Инге-Вечтомова. Были
получены мутанты дрожжей, не съедобные для дрозофилы. Их анализ
показал, что у мутантных клеток блокирован синтез эргостерина. Накап¬
ливающийся в клетках грибов эргостерин имеет структурное сходство с че¬
ловеческим холестерином, накопления которого в виде бляшек на стенках
сосудов так боятся пациенты поликлиник. Как и холестерин, эргостерин
откладывается в мембранах грибных клеток и усиливает их прочность,
а также оказывает регуляторное действие на процессы роста и разви¬
тия. Дрозофилы, как и многие другие насекомые, не умеют синтезировать
ни эргостерин, ни родственные ему молекулы стеринов. Но он им не толь¬
ко нужен, но и просто необходим. Дело в том, что тело насекомых покрыто
жестким наружным скелетом, состоящим из хитина. Взрослые особи насе¬
комых поэтому не растут. А вот их личинки расти обязаны, но хитиновый
скелет им мешает растягивать свое тело. Поэтому в процессе роста они
должны периодически сбрасывать хитиновый скелет, линять. Наверное,
обитатели квартир, заселенных тараканами, видели маленьких белых та¬
раканчиков, «деток». Белые они потому, что сбросили старый хитиновый
покров, а новый еще не успел сформироваться. Наступление времени
линьки и сам процесс регулируются стероидными «гормонами линьки»
эктизонами, которые в клетках дрозофилы синтезируются из эргостерина,
а последний поступает в организм насекомого с дрожжами. Нет в дрожжах
эргостерина, нет и эктизонов, а нет эктизонов, личинки погибают вслед¬
ствие неспособности к росту. Кстати, опыты с дрожжами и дрозофилами
§ 2. Грибы и насекомые
165
объяснили, почему не «червивеют» лисички. Анализ показал, что в их пло¬
довых телах содержится в несколько раз меньше эргостерина (на грамм
сухого веса) чем в шампиньонах, вешенках и других съедобных грибах.
Но, конечно самые изощренные способы питания грибами придумали
муравьи. Люди на заре цивилизации перешли от охоты на животных и
собирательства растений к приучению и разведению животных и посевам
растений. То же самое сделали и муравьи. Животноводческими фермами
для них служат колонии тлей. При питании растительным соком тли
не полностью переваривают сахара, большой процент которых находится
в отходах. Эти-то сахаристые отходы и привлекают муравьев, которые
переносят тлей поближе к муравейнику, ухаживают за ними, защищают
их от хищников.
А объекты земледелия у муравьев — грибы.
Виды муравьев в подавляющем большинстве — хищники, питаю¬
щиеся насекомыми и другими мелкими животными. Однако в Южной
и Центральной Америке живут многочисленные группы муравьев-листо-
резов, которые своими мощными челюстями срезают лист и, забрасывая
его «за плечи», как рюкзак, несут в муравейник. Принесенные листья
они используют не для питания; другие, более мелкие, муравьи разреза¬
ют их на мелкие кусочки и пережевывают до пастообразного состояния
и на этот компост высаживают мицелий специального вида гриба. Этот
гриб, относящийся к роду Леукоагарикус, — родственник шампиньона,
который выращивают люди. Причем этот вид гриба не растет свободно,
только в гнездах муравьев. Молодые матки, покидающие муравейник для
основания нового, прихватывают с собой самое ценное — кусочек этого
гриба. Для заражения мицелием муравьи используют листья розоцветных
и цитрусовых растений.
Некоторые муравьи-листорезы строят самые крупные в мире мура¬
вейники, диаметром до 10 метров и глубиной до 6 метров, включающие
около 2000 камер объемом до 20 кубометров. В этих камерах и устраива¬
ются компостированные грядки, на которых муравьи выращивают грибы.
За всю жизнь одна крупная колония выкапывает около 40 тонн почвы. Му¬
равейники листорезов не только самые крупные, но и самые населенные:
зрелые колонии состоят из нескольких миллионов особей. И питаются
они исключительно грибами, которые выращивают. Можно представить,
какое огромное количество листьев надо собрать, принести и пригото¬
вить. Поэтому муравьи-листорезы во многих местах являются бичом для
садоводов. Земледельческая культура муравьев находится на высоком тех¬
ническом уровне. Они приготавливают «листовой компост», удобряют его
своими экскрементами, засевают грибницей, пропалывают и ведут борьбу
с «сорняками». Для этого они вносят в компост мицелий одного из видов
мицелиальных бактерий — актиномицетов, — который образует антибио¬
тик, подавляющий рост многих микроорганизмов, но не ядовитый для
166
Глава 7. Грибы и животные
гриба, которым засеяны грядки. Так что и применению антибиотиков
в сельском хозяйстве муравьи научились гораздо раньше людей.
Сбор урожая также своеобразен. Муравьи подгрызают выросты ми¬
целия (примордии), из которых в дальнейшем должны формироваться
плодовые тела. На местах погрызов гриб образует утолщения диаметром
до полумиллиметра — стафилы или, как их назвал первый исследователь
муравьиных садов Ф. Мюллер, грибные кольраби. Этими стафилами му¬
равьи выкармливают личинок. Такой способ питания весьма энергетиче¬
ски эффективен. Грибы перерабатывают растения, как уже было сказано,
гораздо эффективнее, чем насекомые; грибной мицелий содержит много
питательных веществ — белка, липидов, более усвояемых, чем раститель¬
ные, углеводов.
Некоторые виды муравьев строят маленькие муравейники, в камерах
которых выращивают не мицелиальные грибы, а дрожжи. Интересно, что
они используют для приготовления компостов не растения, а фекалии
и измельченные шкурки убитых гусениц.
«Грибоводством» занимаются и другие насекомые — некоторые тер¬
миты, жуки короеды, комарики.
Наконец, широко распространены такие взаимо-
Как они помогают отношениям между грибами и насекомыми, при ко-
ДРУг другу торых никто никого не ест, но и те, и другие полу¬
чают взаимную пользу.
Многие грибы используют насекомых для распространения своих
спор. В главе, посвященной размножению грибов, было рассказано, как
спороносные органы грибов выделяют сладкую ароматную жидкость или
летучие вещества, обладающие трупным запахом, для привлечения тех или
иных насекомых, распространяющих споры на своем теле. А есть и мно¬
гочисленные случаи того, как грибы и насекомые коэволюционировали
в направлении получения взаимной выгоды от совместного воздействия
на субстраты, представляющие обоюдный интерес. В качестве примера
таких сложных взаимоотношений приведу историю появления и распро¬
странения голландской болезни вязов.
После 1910 г. Европа жила в напряженном ожидании войны. План
германского генерального штаба — план Шлиффена — предусматривал
быстрый разгром Франции путем удара через нейтральную Бельгию на Па¬
риж. Через несколько лет армия генерала фон Клюка осуществит этот
маневр и будет остановлена только под Парижем. А пока французы, хотя
и отнеслись к плану Шлиффена несколько легкомысленно, но все-таки
решили укрепить свои северо-восточные границы и наняли для рытья
окопов во Фландрии несколько сот китайских рабочих. Вскоре разрази¬
лась первая мировая война, а еще через несколько лет в Европе начали
погибать ильмовые деревья. Впервые болезнь была обнаружена в Гол¬
ландии и поэтому получила название голландской болезни вязов. Листья
на пораженных деревьях увядали, ветви засыхали и все дерево погибало.
§ 3. Грибы и жвачные животные
167
Возбудителем болезни оказался сумчатый гриб Офиостома ульми, который
приникал в сосуды ксилемы дерева и выделял там токсические соедине¬
ния, в частности, пептид церотоульмин. Токсины нарушали транспорт
питательных веществ по сосудам и отравляли все дерево. Ослабленное
дерево заселяли жуки короеды, которые не способны заразить здоровые
растения (поэтому энтомологи называют их «вторичными вредителями»).
Жуки делают ходы в коре в виде галерей с многочисленными разветв¬
лениями, напоминающие типографские знаки, потому этих насекомых
и называют типографами. В этих галереях жуки спариваются и отклады¬
вают яйца. Из яиц вылупляются личинки, которые питаются древесиной,
а из них — жуки. Мицелий гриба офиостома прорастает в ходы типогра¬
фов и образует там свои споры — конидии. Молодые жуки, выходящие
на поверхность дерева, несут на своем теле споры гриба. Их питание более
поэтично, чем их деток — цветками вязов. При питании они заносят в по¬
врежденные ткани споры гриба, ростки которого проникают внутрь ветвей
и начитают свое губительное воздействие на дерево. Через несколько лет
дерево ослабляется настолько, что становится доступным для поражения
короедами. Цикл развития гриба, жука и болезни завершается. Таким об¬
разом, жуки короеды могут поражать только ослабленные деревья, гриб
ослабляет его и подготавливает для заселения жуками. А жуки выносят
споры на поверхность и заражают ими здоровые растения. Таково удиви¬
тельно скоординированное действие двух разбойников, убивающих вязы.
Откуда же и каким образом попала в Европу голландская болезнь вя¬
зов? На этот счет есть несколько предположений, но в этой книге следует
остановиться на предположении не самом аргументированном, но самом
романтическом. Микологи решили, что гриб, попавший в Европу, родом
из Азии, так как азиатские виды ильмов устойчивы к болезни. Поиски
путей проникновения гриба в Европу заставили вспомнить о китайцах,
рывших каналы и окопы во Фландрии: свой скарб они привезли в де¬
ревянных корзинках, видимо, зараженных жуками-короедами. А дальше
все стало понятно. Кстати, с зараженными досками болезнь из Европы
попала в Северную Америку, где на новых видах ильмов и в новых клима¬
тических условиях произошло быстрое видообразование, сформировался
новый вид, названный Офиостома ново-ульми, гораздо более агрессив¬
ный, чем его европейский предшественник. Америка не осталась в долгу
перед Европой и опять-таки с лесоматериалами вернула гриб, к тому же
значительно более вредоносный, в Англию, где за пять лет страна поте¬
ряла от голландской болезни 6,5 миллионов деревьев.
§3. Грибы и жвачные животные
Тело жвачного животного и, прежде всего, самой знакомой нам и са¬
мой изученной — коровы — это сложное биотехнологическое производ¬
ство, на входе которого находятся малокалорийные и трудно усвояемые
168
Глава 7. Грибы и животные
соломистые корма, а на выходе — мясо, молоко и богатый азотом на¬
воз. Причем, производство это очень экономное, достаточно сказать, что
даже конечный продукт расщепления белка (мочевина) не удаляется пол¬
ностью, а частично циркулирует в организме, давая материал для обра¬
зования аминокислот. Конечно, собственными силами все эти сложные
процессы обеспечить невозможно, поэтому организм коровы и, главным
образом, ее желудочно-кишечный тракт набит симбиотическими мик¬
роорганизмами, которые своими ферментами расщепляют полисахариды
кормов (прежде всего, целлюлозу или клетчатку) и передают продукты
расщепления корове для дальнейшей утилизации. Долгое время считали,
что расщепление клетчатки в рубце коровы осуществляют исключительно
целлюлозолитические бактерии. Однако лет 30 назад в рубце были обна¬
ружены совершенно удивительные и высоко специализированные грибы,
относящиеся к древнейшей группе водных и наземных грибов, хитриди-
омицетам, которые обитают только там. Они представляют собой микро¬
скопические одноклеточные организмы, тело которых имеет вид пузыря,
прикрепленного к внутренней стенке рубца разветвленной гифой — ризо-
мицелием. В период размножения содержимое клетки-пузыря распадается
на множество зооспор, имеющих пучок жгутиков (у некоторых видов —
один жгутик), которые, поплавав в содержимом рубца, садятся в новом
месте, теряют жгутики, покрываются оболочкой и прикрепляются с помо¬
щью ризомицелия (рис. 39). Грибы эти образуют мощнейшие ферменты,
разлагающие целлюлозу, и играют главную роль в разрушении грубых цел¬
люлозных волокон корма. Их ферментативный аппарат давно привлекает
внимание биотехнологов. Беда только, что они являются строгими анаэ¬
робами, не терпят кислорода и погибают даже при очень низком его со¬
держании, так что их очень сложно культивировать. Поэтому в их клетках
даже нет митохондрий — важнейших подстанций для выработки энергии
при участии кислорода. Митохондрии были утрачены за ненадобностью
после того, как предки этих грибов вошли в симбиотические отноше¬
ния с жвачными животными. А произошло это событие очень давно, так
как в рубце евроазиатских коров, североамериканских бизонов, южно-
Рис. 39. Внутренний эпителий же¬
лудка коровы с многочисленными
мешковидными талломами хитри-
диального гриба неокаллимастикс,
прикрепленными с помощью ризо¬
мицелия (Bauchop Т., 1985)
§ 3. Гоибы и жвачные животные
169
американских лам, африканских антилоп и даже австралийских кенгуру
обитают близкие виды хитридиевых грибов. Когда-то земля представляла
собой единый материк Пангею. Затем произошел ее раскол и разделе¬
ние на северную часть — Лавразию и южную Гондвану. После этого обе
часть продолжали раскалываться, так что из единой Гондваны возникли
современные Африка, Южная Америка и Австралия. Значит, предок всех
жвачных животных, обитающих на разных материках, приобрел симбио¬
тические грибы хитридиомицеты еще до начала раскола материков.
В одном грамме содержимого рубца находится около миллиарда кле¬
ток бактерий, около 10 миллионов клеток грибов хитридиомицетов и око¬
ло миллиона клеток простейших животных. Кроме строго симбиотических
анаэробных бактерий целлюлозолитиков и грибов хитридиомицетов в руб¬
це находится и транзитная микрофлора, попадающая туда, в основном,
с кормами. Некоторые ее представители также участвуют в разложении
корма. В рубце протекают различные микробиологические процессы:
— во-первых, переваривание поли- и олигосахаридов (клетчатки, крах¬
мала и др.);
— во-вторых, расщепление белков и небелковых соединений азота;
— в-третьих, сбраживание молочной, янтарной и других органических
кислот;
— в-четвертых, расщепление липидов, гидратирование и изомеризация
жирных кислот.
Конечно, важнейший процесс, протекающий в рубце, — разложение
клетчатки, которая переваривается на 60-70 %. Основные продукты ее
разложения — органические кислоты: уксусная, пропионовая, масляная,
валериановая, а также газы — углекислота, метан и водород.
В этих сложных микробиологических процессах животное, конечно,
также принимает участие, во-первых, снабжением микроорганизмов пи¬
тательной средой: кормами и мочевиной, поступающей через клетки ки¬
шечника и со слюной; во-вторых, в рубце созданы условия термостата
(постоянная температура) и поддерживается одинаковый уровень кислот¬
ности pH, благодаря буферным свойствам слюны. А это очень важные
условия для химической деятельности микроорганизмов в любом биотех¬
нологическом процессе.
Долгое время было не очень понятно, каким способом коровы переда¬
ют анаэробные симбиотические микроорганизмы — бактерии и грибы —
телятам. Ведь хитридиевые грибы быстро погибают в воздухе. Сейчас
считают, что в процессе вылизывания теленка коровы передают ему сим¬
бионтов изо рта в рот.
Глава 8
Эти удивительные
создания миксомицеты
После разговора о взаимоотношениях грибов с жи¬
вотными сделаем небольшое отступление и расскажем
об удивительной группе живых существ, которые вели¬
кий ботаник Антон де Бари назвал мицетозоа или по-
русски грибоживотными.
По латыни их называют миксомицетами (слизи¬
стыми грибами), а по-русски — слизевиками. Такое на¬
звание они получили в связи с тем, что их вегетативное
тело представляет собой голый комочек цитоплазмы,
окруженный плотным белковым слоем и цитоплазмати¬
ческой мембраной и содержащий много ядер. Размеры
этого комочка у разных видов могут составлять как де¬
сятки микрометров (тысячных долей миллиметра), так
и десятки сантиметров. Некоторые слизевики способны
подобно амебам ползать по субстрату в поисках лучших
условий для жизни. Так что признаки миксомицетов
сближают их с простейшими животными. Да и питание
относимых к слизевикам видов смешанное, как осмо-,
так и зоотрофное, ибо наряду с всасыванием они погло¬
щают целые клетки бактерий и дрожжей. Отнесение их
грибам чисто традиционное: так повелось исторически,
что о них преподают студентам в курсах микологии,
и публикуют результаты исследований в научных ми¬
кологических журналах. Ранее к слизевикам относили
разнообразные организмы, имеющие слизистое много¬
ядерное тело. После появления инструментальных, осо¬
бенно молекулярных методов исследований, было выяс¬
нено, что многие их виды висят на очень удаленных друг
от друга ветках дерева, называемого биотой, и не имеют
никаких родственных связей друг с другом.
§ 2. Паразитические слизевики, или фитомиксины 171
§ 1. Сетчатые слизевики,
или лабиринтуломицеты
Эти организмы живут в морях и являются внутриклеточными парази¬
тами морских растений и животных, а также салротрофами, участвующи¬
ми в разложении морского детрита. Лабиринтуломицетами их называют
в связи с тем, что их клетки продуцируют наружу полисахариды, застыва¬
ющие в виде слизистых трубочек, внутри которых, как в лабиринте, клетки
передвигаются с помощью жгутиков. Так что ничего общего с истинны¬
ми слизевиками они не имеют. А принадлежат эти организмы к группе
гетероконтов, то есть родственны описанным в первой главе оомицетам.
Паразитические лабиринтуломицеты вызывают иногда массовую гибель
водорослей и морской травы зостеры, заросли которой покрывают дно
морских мелководных участков. А сейчас эти организмы заинтересовали
диетологов, так как содержат много омега-3-полиненасыщенных жирных
кислот, весьма популярных в данный период развития диетологии.
§2. Паразитические слизевики,
или фитомиксины
Это небольшая группа организмов, паразитов растений. Они являют¬
ся внутриклеточными паразитами растений и образуют плазмодии в за-
ражешшх клетках. Наиболее известный их представитель — возбудитель
килы капусты и других крестоцветных растений. Находящиеся в клетках
зараженных корней плазмодии выделяют фитогормоны, которые застав¬
ляют растения направлять потоки питательных веществ не в листья, фор¬
мирующие кочан, а в корни. Происходит усиленное разрастание и деление
зараженных клеток, которое приводит к образованию опухолей на кор¬
нях — киле (рис. 26 в, стр. 103). Поэтому зараженное растение образует
маленький кочан и крупные, распухшие корни. А плазмодий паразита
распадается на отдельные споры, покрытые плотными оболочками и осво¬
бождающиеся в почву после разрушения корней. Эти споры могут десят¬
ки лет лежать в почве без движения, а для их прорастания необходимы
корневые выделения крестоцветных растений. Под их влиянием споры
преобразуются в зооспоры, которые плывут на «запах», достигают кор¬
невых волосков, покрываются оболочкой и выдвигают свой «бандитский
обрез» — трубку, через которую с силой выбрасывается острое хитиновое
копьецо, пробивающее клеточную стенку корня. Через образовавшуюся
брешь содержимое споры переливается в клетку корня и формирует пер¬
вичный плазмодий. Так что эти организмы по образу жизни и типу пи¬
тания ближе к грибам, чем к истинным слизевикам: они содержат хитин,
их плазмодий не способен к активному движению, они питаются только
осмотрофно, как грибы, и не могут заглатывать отдельные клетки. Вред
172 Глава 8. Эти удивительные создания миксомицеты
от них заключается не только в том, что они снижают урожай (как воз¬
будитель килы капусты) или его качество (как возбудитель порошистой
парши картофеля), но и в том, что на своем теле они могут переносить
от больных растений к здоровым вирусы растений, как при заражении
злаков фитомиксинами.
§3. Настоящие миксомицеты,
или миксогастриды
К ним относят организмы, большинство видов которых в вегетатив¬
ной стадии представлены талломом (плазмодием), содержащим большое
число диплоидных ядер, лишенным клеточной стенки и живущим внутри
субстрата (гнилой древесины, коры, опавших листьев). Плазмодий питает¬
ся зоотрофно, заглатыванием клеток бактерий, дрожжей, амеб и грибных
спор. Плазмодий активно движется по направлению к источнику пищи
(обладает трофотаксисом), а также уползает в более темные и влажные
места. Если на дно чашки поместить кусочек коры, а на подставке, распо¬
ложенной сверху, свесить вниз на кору полоску фильтровальной бумаж¬
ки, помещенную другим концом в воду, то плазмодий выползет из коры
по фильтровальной бумажке навстречу току воды. После периода питания
у плазмодия резко меняются вкусы, что связано с активизацией опре¬
деленных генов, ранее молчащих, и синтезом новых белков. Плазмодий
выползает наружу и буквально на глазах начинает превращаться в спо¬
роносящие структуры. Спорангии слизевиков могут быть одиночными,
сидячими или на ножках, или плазмодий преобразуется в группу споран¬
гиев. Иногда спорангии срастаются боковыми стенками, а у некоторых
видов весь плазмодий покрывается общей оболочкой, превращаясь в спо-
рокарп. И все эти структуры окрашены в яркие, удивительно разнообраз¬
ные цвета. Эти мелкие организмы — прекрасное украшение наших лесов
(рис. 40), только большинство сельских жителей ходят в лес с чисто потре¬
бительскими целями — собирать грибы, ягоды, лекарственные растения,
и не обращают внимания на такую мелочь, как споровместилища миксо-
мицетов, а горожане и того хуже — ходят в темных очках и с воткнутыми
в уши наушниками, так что вообще ничего не видят и не слышат. Внут¬
ри споровместилищ происходит мейотическое деление ядер и образуются
гаплоидные споры, покрытые прочными, часто шиповатыми, оболочка¬
ми, и различного рода нитевидные структуры, которые при изменении
влажности воздуха изгибаются и разрыхляют споровую массу, не давая
ей слежаться, и даже участвуют в разбросе спор. Оболочка высыпавшихся
спор лопается и из них образуются зооспоры, которые быстро теряют жгу¬
тики и превращаются в амеб. Амебы попарно сливаются (причем только
«плюс» амебы с «минус» амебами, при этом сливаются и их ядра, так что
в результате полового процесса восстанавливается диплоидность). Дипло¬
идные амебы разрастаются и превращаются в плазмодий, ядра которого
§ 3. Настоящие миксомицеты, или миксогастриды 173
Рис. 40. Плодовые тела миксомицетов: а) цератиомикса (златослизь); б) икогаля
(волчье вымя); в) арцирия; г) трихия; д) дидерма; е) стемонитес; ж) физарум; з) лео-
карпус (Жизнь растений, т. 2, 1976)
многократно делятся митотически. Таким образом, в вегетативной стадии
слизевики сходны с простейшими животными, а в стадии спороноше-
ния — с грибами.
Помимо органов спороношения миксомицетов сближает с грибами
и наличие у них покоящихся структур. При неблагоприятных условиях
жизни, таких как длительная засуха, холода, отсутствие пищи, содержи¬
мое плазмодия уплотняется, твердеет и он превращается в склероций,
который может долгое время сохраняться, а затем опять превращаться
в жизнеспособный плазмодий. В одной лаборатории сообщалось о скле¬
роции миксомицета фулиго, пролежавшем в гербарии 20 лет и сохранив¬
шем способность формировать плазмодий.
Ученых давно интересовало движение плазмодиев, „ _
Как движется 0
как плазмодии воспринимает внешнии сигнал и откли- nil„uftn -
ПЛаЗМОДИИ
кается на него, каковы механизмы, заставляющие плаз¬
модий ползти в нужном направлении. В институте биофизики Российской
академии наук есть даже лаборатория немышечных движений, в которой
в качестве модельного объекта изучают миксомицет Физарум многого¬
ловый. Его ярко-желтый или зеленоватый плазмодий иногда развивается
на гименофорах шляпочных грибов (с нижней стороны шляпок). Головки
спорангиев венчают извилистые ножки, плотно прижатые друг к другу,
отсюда и возникло название «многоголовый». Его исследуют в лаборато¬
риях разных стран. Разработаны методы получения полного цикла разви¬
тия этого миксомицета в лабораторных условиях и получено множество
мутаций. Этот миксомицет вошел в книгу рекордов Гиннеса, как самая
174
Глава 8. Эти удивительные создания миксомицеты
крупная в мире клетка, так как удалось вы¬
растить плазмодии диаметром в полтора-два
метра и толщиной несколько сантиметров!
В плазмодии физарума постоянно осу¬
ществляются автоколебательные возвратно¬
поступательные токи цитоплазмы. Они обу¬
словлены периодическими сокращениями
белка актомиозина, получающего энергию от
молекул аденозинтрифосфата — универсаль¬
ного хранителя энергии в живых клетках.
Тот же самый белок — актомиозин является
источником мышечного движения позвоноч¬
ных животных. Так что природа экономна в
выборе средств. Внутри плазмодия находится
сеть трубчатых элементов, подобных веноз¬
ным сосудам, с помощью которых осуществ¬
ляется транспорт питательных веществ и распространяются сигналы, по¬
лучаемые от окружающего плазмодий мира (ползти туда, где темно, сыро
и есть что покушать). В покоящемся плазмодии сеть трубочек образует,
подобно круговой шнуровке, концентрические круги. Когда плазмодий
начинает двигаться, трубчатые элементы изменяют свою ориентацию и ра¬
диально расходятся от центра плазмодия к периферии (рис. 41). Вперед
по направлению движения плазмодий подобно амебам выпускает ните¬
видные псевдоподии с луковицеподобными окончаниями, а задняя часть
скелетирована сетью толстых тяжей. С этими плазмодиями проведено
множество интереснейших опытов.
Если предварительно выдержанный на голодном пайке
Буриданов о
и. плазмодий поместить в сосуд, дно которого выстлано увлаж-
плззмодии
ненной фильтровальной бумагой, а поблизости положить
кусочек пищи (например, овсяных хлопьев), то плазмодий будет полз¬
ти к пище, окружит ее и поглотит. Если поместить два источника пищи
вблизи от разных сторон плазмодия, то он будет тянуться в разные сто¬
роны, однако он, как бы мысленно, должен оценить свои возможности.
Во-первых, для того, чтобы эффективно поглотить пищу, она должно быть
окружена достаточно большой массой тела, так что этой массы должно
быть достаточно для абсорбции питания из двух источников. Во-вторых,
при разнонаправленных движениях участков одного плазмодия должна
сохраняться связь между ними. Таким образом, возможность растяжения
плазмодия к разным источникам пищи лимитирована его массой. И толь¬
ко в том случае, если пищи очень много, плазмодий принимает решение
разбиться на две части.
Если же вокруг плазмодия на разном расстоянии от него расположить
несколько источников пищи, причем не по прямой, а вдоль ломаной
линии, то плазмодий не будет ползти ко всем сразу, а сначала освоит бли-
Рис. 41. Сеть трубочек
в движущемся плазмодии
физарума (Hickey D. S.,
Noriega L. A., 2008)
§ 3. Настоящие миксомицеты, или миксогастриды 175
Рис. 42. Движение плазмодия физарум
к пищевым приманкам (Adamatzky, 2009)
жаишии источник, затем поползет к следующему ^
/ А~\ ^ Как физарум
и так далее (рис. 42). Созданы математические мо- проходит лабиринт
дели, свидетельствующие о том, что он выбирает
оптимальные направления движения. Следовательно, одноклеточный фи¬
зарум обладает элементами рассудочной деятельности. А как проверяют
интеллект у животных и даже у человека? Одним из широко распро¬
страненных тестов является способность ориентироваться в лабиринте.
Вот японские ученые и поместили плазмодий физарума у входа в умень¬
шенную копию лабиринта, который применяют для проверки интеллекта
и памяти у мышей, а на выходе положили корм. Плазмодий начал расти
в направлении приманки, причем, когда его выросты попадали в дорожки,
заканчивающиеся тупиком, возникали ответвления, и нити росли в другом
направлении. Наконец, плазмодий нашел приманку. Но самое интересное
случилось после того, как этот плазмодий перенесли в новый, такой же
лабиринт с приманкой, расположенной на том же месте. Плазмодий сразу
стал расти правильной дорогой, не заходя в тупики. То есть плазмодий
физарума мало того, что находит самые короткие пути по направлению
к пище, он еще и обладает памятью. Обучение и память — свойства мозга
или, по крайней мере, нервной системы. Заменитель мозга физарума —
трубчатые элементы плазмодия, которые подобно «ногам» позволяют ори¬
ентироваться в окружающей среде. Они легко распадаются и собираются
вновь при изменении условий жизни или стоящей перед плазмодием зада¬
чи. О наличии у плазмодия памяти свидетельствуют и другие интересные
эксперименты. Например, плазмодий физарума ползет по субстрату при
комнатной температуре со скоростью около 1 сантиметра в час, но при
подсушивании субстрата скорость роста уменьшается. В поставленных
опытах плазмодий кратковременно подсушивали (а затем снова увлажня¬
ли) с интервалами в один час. После этого через каждый час в течение
двух-трех часов рост плазмодия замедлялся, несмотря на прекращение его
подсушивания. В опытах, где изменяли интервалы подсушивания, изме¬
нялись и интервалы последующей задержки роста без подсушивания. То
есть плазмодий обладает не только памятью о перенесенных неприятных
176
Глава 8. Эти удивительные создания миксомицеты
ощущениях, но и биологическими часами, которые, кстати, работают и
у настоящих грибов, например, у сумчатого гриба нейроспоры.
Таким образом, физарум обладает всеми необходимыми свойствами,
которые должен иметь робот: сенсорами, воспринимающими сигналы
из окружающей его среды; компьютером, способным рассчитать опти¬
мальную траекторию движения; механизмом движения, обеспечивающим
продвижение к намеченной цели; наконец, способностью к обучению.
Поэтому миксомицетов ждет большое будущее как выдающихся объектов
для бионики, биофизики, биотехнологий. Будут ли они от этого счастли¬
вее, не знаю, но нам они могут помочь в решении многих задач. Например,
та же группа японских ученых, которая помещала плазмодий в лабиринт,
положила его на карту Японии в точку, отмеченную как город Токио,
а на места, где расположены другие крупные города, поместила кусочки
пищи. Через некоторое время нити плазмодия протянулись к этим местам.
В результате этого получилась почти точная копия железнодорожной сети
вокруг Токио. Подобные опыты проведены с географическими картами
в Великобритании и в Испании. Авторы этих работ считают, что, уни¬
кальные способности миксомицета помогут в будущем не только понять,
как строить инфраструктуру с улучшенной архитектурой, но и создавать
наиболее эффективные и быстрые информационные сети.
Другая исследовательская группа попыталась использовать удиви¬
тельные способности физарума для управления движением робота. Был
создан шестиногий робот, каждая нога которого через систему датчи¬
ков была связана с одним из шести кусочков плазмодия, расположенных
по окружности на равном расстоянии друг от друга. Робот имел также
светоулавливающую систему, соединенную с сосудом, в котором находи¬
лись плазмодии. Падение света под определенным углом достигало одно¬
го из шести плазмодиев, который, как известно, уползает от света. Его
движение передавалось связанной с ним ноге робота, которая двигалась
до тех пор, пока плазмодий не уползал в темноту и не останавливался. Та¬
ким образом, под влиянием плазмодиев, робот убегал от света. Несмотря
на примитивность этого создания, предсказанная эффективность его ра¬
боты вселяет уверенность в возможности разработки гораздо более сложно
устроенных механизмов, выполняющих разнообразные задачи.
§ 4. Клеточные миксомицеты:
диктиостелиды и акразии
Клеточные миксомицеты — это своя сказка. Они не
Социальные ^
амебы образуют гигантских многоядерных клеток, как истинные
миксомицеты, а представляют собой небольшие одноядер¬
ные амебы, способные к амебообразным движениям с помощью псевдо¬
подий и заглатыванию клеток бактерий. К тому же, в отличие от на¬
стоящих слизевиков, они не имеют полового процесса. В лабораториях,
§ 4. Клеточные миксомицеты: диктиостелиды и акразии 177
а эти организмы волнуют множество ученых из разных стран, по по¬
верхности чашки Петри с плотной (агаризованной) питательной средой
размазывают взвесь бактериальных клеток (обычно — сенной палочки) и
после того, как поверхность чашки покрывается сплошным слоем бакте¬
рий, помещают на чашку амеб. Они питаются бактериальными клетками
и размножаются делением. В общем, амебы как амебы. Но когда исчер¬
пываются пищевые ресурсы, начинается самое интересное: отдельные их
экземпляры приобретают способность выделять хемоаттрактанты (веще¬
ства, привлекающие других амеб), названные акразинами (циклический
аденозинмонофосфат, фолиевую кислоту, дипептид глорин). Кроме ат-
трактантов эти клетки образуют вещества, подавляющие, во избежание
хаоса, способность выделять аттрактанты соседними клетками. Эти во¬
левые клетки-организаторы получили название центров агрегации. На их
зов близлежащие амебы начинают двигаться со всех сторон в направ¬
лении центров агрегации. Движение осуществляется очень согласовано,
синхронно: амебы ползут со всех сторон к центру в течение полутора
минут, а затем 2-3 минуты отдыхают. В результате сползаний через не¬
которое время вместо равномерного распределения амеб по поверхности
чашки возникает несколько центров агрегации, в которых собираются все
амебы. Там они начинают наползать друг на друга и формировать холмик,
псевдоплазмодий, который способен, как настоящий плазмодий, пере¬
двигаться по поверхности чашки. В общем, социальное поведение амеб
оказывается таким же, как и у людей: когда жизнь сытая и комфортная,
социалистические идеи никому не интересны, все замыкаются по своим
особнякам и фазендам и встречаются только по делам бизнеса или для
развлечений. А вот когда плохо, люди сбиваются в группы, «чтоб не про¬
пасть поодиночке», как сказал поэт, появляются центры агрегации в виде
харизматичных партийных лидеров и агитаторов.
Затем начинаются морфогенетические изменения, которые у акрази-
вых и диктиостелиевых слизевиков протекают по-разному, что не удиви¬
тельно, ибо они принадлежат к разным царствам, и их похожесть вызвана
лишь адаптациями, вызванными сходными условиями жизни. У боль¬
шинства акразиевых наползающие друг на друга несколько тысяч амеб
образуют столбик, поднимающийся над субстратом на высоту около од¬
ного миллиметра, на верхней части которого, подобно ветвям дерева,
образуются цепочки спор (клетки покрываются оболочками). Образуется
изящное, почти микроскопическое деревце, у некоторых видов, напри¬
мер, у розовой акразии, ярко окрашенное.
У диктиостелиевых образующиеся из псевдоплазмодия плодовые тела
имеют более совершенную форму вследствие четких структурных разли¬
чий между клетками, образующими ножку и споры (у многих акразиевых
таких различий нет). Многие наползающие друг на друга амебы образу¬
ют довольно длинную (около пяти миллиметров) и тонкую, сужающуюся
к концу ножку, покрытую плотным целлюлозным покровом. Клетки, со-
Рис. 43. Диктиостелий дисковидный, жизненный цикл (Olive, 1956)
ставляющие ножку, увеличиваются в размерах и отмирают, превращаясь
в лишенные содержимого пузыри (вакуолизируются). На такие клетки
наползают следующие амебы, также превращающиеся в пузыри. Клетки,
поднявшиеся на самый верх ножки, не погибают, а покрываются оболоч¬
кой и превращаются в споры. У самого известного вида — дисковидно¬
го диктиостелия — на верху ножки образуется головка из спор, покры¬
тая тонкой целлюлозной оболочкой (рис. 43). Таким образом, на наших
глазах развертывается интереснейшее событие — превращение отдельно
живущих индивидуальных клеток в единый организм. Нас поражает син¬
хронность полета с одновременными резкими поворотами стаи галок или
синхронность движений в воде косяка мелких рыб, но в обоих случаях
каждый член стаи сохраняет пространственную индивидуальность, под¬
чиняясь лишь каким-то общим для всех командам. Здесь же маленькие
амебы индивидуальность теряют полностью. Более того, многие из них,
которые составят ножку, должны погибнуть в процессе морфогенеза.
Для чего же нужны эти превращения? Социальные
объединяться? амебы живут в богатой органикой почве и питаются, как
уже было сказано, бактериями. Они, как и настоящие
слизевики, имеют аппарат улавливания «запаха», исходящего от потенци¬
альной пищи, и аппарат, позволяющий двигаться в нужном направлении
с помощью псевдоподий, то есть обладают трофотаксисом. Но в почве пи¬
ща амеб — клетки бактерий, дрожжей или других, более мелких, амеб —
§ 4. Клеточные миксомицеты: диктиостелиды и акразии 179
распределена очень неравномерно, лоскутно, ибо их много в местах, где
имеется их собственная пища (растительные и животные остатки) и очень
мало там, где пищи нет. Поэтому исчерпав место, богатое бактериальными
клетками, амебы должны найти новое такое место, но оно может быть
на таком расстоянии, что его будет невозможно ни уловить, ни доползти
до него. Поэтому при исчерпании запасов пищи, находящейся «в шаго¬
вой доступности», надо менять стратегию поиска. Так поступают не только
эти амебы. Мицелии шляпочных грибов живут в почве, но образуют над
почвой плодовые тела со спорами, причем для обеспечения распростра¬
нения спор на более дальние расстояния спороносная ткань поднимается
над почвой с помощью длинной ноги. Плазмодии настоящих слизевиков
выползают из своих субстратов на поверхность и там, на воздухе, фор¬
мируют споры. Также и маленькие диктиостелии при исчерпании пищи
переходят к социальному поведению, образуют единый организм — нож¬
ку, поднимающую на поверхность головку спор. Что это дает? Во-первых,
вегетативные клетки не могут быть покры¬
ты полисахаридными оболочками, ибо они
в таком случае не смогли бы заглатывать
пищу с помощью пиноцитоза (амеба на¬
ползает на бактериальную клетку, окружает
ее и в собственном мембранном пузырь¬
ке проталкивает внутрь клетки, где перева¬
ривает) (рис. 44). Такое питание возможно
только в том случае, если клетки «голень¬
кие», не покрытые оболочками. А клетки,
не имеющие прочной оболочки, недолго¬
вечны и быстро погибают в неблагопри¬
ятных условиях. Образующиеся в головке
споры покрыты целлюлозными оболочками и могут длительное время
сохраняться без пищи и воды. Во-вторых, поднятые на несколько мил¬
лиметров над поверхностью земли споры могут распространяться на дли¬
тельные расстояния порывами ветра, токами воды и на теле различных
животных — насекомых, нематод, да и прилипнув к подошвам обуви.
В процессе морфогенеза клетки, из которых образуется ножка, поги¬
бают, а клетки, превращающиеся в споры, остаются, естественно, живыми.
Каким образом осуществляется сортировка амеб, составляющих псевдо¬
плазмодий, на смертников и основателей новых колоний? Генетических
различий между ними не обнаружено, даже в клоне, возникшем в результа¬
те размножения одной клетки основательницы, плодовое тело нормально
дифференцируется на ножку с мертвыми клетками и головку с живыми
спорами. Однако обнаружено, что на дифференциацию клеток влияют
условия их жизни. Например, при смешивании амеб, получавших обиль¬
ную пищу, с амебами, которых держали на голодном пайке, голодные обра¬
зовывали ножку и умирали, а сытые превращались в споры. Также клетки,
Рис. 44. Пиноцитозный способ
питания: Б — бактериальные
клетки; П — псевдоподии; ПП —
пиноцитозный пузырек
180
Глава 8. Эти удивительные создания миксомицеты
которые начали голодать перед или во время деления, с высокой вероят¬
ностью погибают, формируя ножку, а клетки, которые попадают в условия
голодания после деления, чаще становятся спорами. С точки зрения стра¬
тегии выживания такой отбор понятен: клетки, богатые питательными
веществами, имеют больше шансов сохраниться в неопределенных усло¬
виях, в которые они могут попасть после освобождения из спорангия, чем
бедные клетки. Но каков механизм такого отбора, пока неизвестно.
Превращения многих индивидуальных малень-
Диктиостслии ~
простейшие модели °Рганизмов в единый большой организм привле-
развития зиготы кает огромное внимание исследователей к диктио-
стелиям. Они стали излюбленным объектом многих
лабораторий. Дело в том, что мы довольно много знаем уже об окружа¬
ющем нас мире, вплоть до отдаленных галактик, но очень мало — о соб¬
ственном организме. Каким образом из одной оплодотворенной клетки
(зиготы) в результате серии делений образуются такие непохожие друг
на друга органы и ткани, как печень, легкие, глаза, кожа и прочее. Эту
проблему, проблему генетики онтогенеза, афористично сформулировал
наш знаменитый генетик Н. В. Тимофеев-Ресовский: «Почему в должное
время, в должном месте создается должное?». Вот диктиостелии и мо¬
гут быть простейшими моделями для ответов на поставленные вопросы.
Некоторые считают, что с них все началось, то есть образование многокле-
точности проходило через те же этапы, что и у них. Конечно, есть прин¬
ципиальное различие: у социальных амеб единый организм, разделенный
на функционально различные отделы, возникает вследствие объединения
одинаковых вначале клеток, а у многоклеточных организмов — вслед¬
ствие дробления одной первичной клетки. Но, возможно, все началось
с объединения, которое было первичным по сравнению с дроблением.
Есть и более частные проблемы. Например, единичные клетки (амебы)
ведут себя подобно лейкоцитам позвоночных животных: они имеют ана¬
логичный механизм движения, хемотаксис и способность к заглатыванию
бактерий вследствие фагоцитоза. Еще одна проблема — величина тела
организмов. Почему мышь останавливается в росте, достигнув десятка
сантиметров, а тело слона достигает нескольких метров? У диктиостелий
соотношение длины ножки и размеров споровой головки определяется не¬
обходимостью, во-первых, создать как можно больше спор, и, во-вторых,
поднять их как можно выше над субстратом. Одновременно выполнение
обоих условий невозможно, так как на очень длинной ноге очень крупные
головки спор согнут ножку и упадут. Значит должен быть достигнут некий
оптимальный компромисс между длиной ножки и размером головки. Он
достигается в том случае, если плодовое тело формируется из примерно ста
тысяч амеб. Как же они знают, сколько их нужно? Оказывается, есть груп¬
па из шести белков, которые осуществляют подсчет числа амеб, необходи¬
мых для образования псевдоплазмодия и формирования плодового тела.
И есть белок, регулирующий продукцию этих белков. Если с помощью
§4. Клеточные миксомицеты: диктиостелиды и акразии 181
генетических манипуляций выключить синтез этого белка регулятора, то
произойдет перепродукция белков — счетчиков клеток. У таких генети¬
чески измененных клеток сначала возникает нормальный агрегат из ста
тысяч клеток, который затем распадается на маленькие псевдоплазмодии,
состоящие из пяти тысяч клеток. Из них формируются миниатюрные
плодовые тела с короткими ножками и мелкими головками. Если же, на¬
оборот, выключить работу гена, который необходим для синтеза хотя бы
одного из белков счетчиков, то псевдоплазмодии, наоборот, оказываются
составленными из большего, чем надо, числа амеб, и образуются плодовые
тела с длинными ножками и крупными головками, под тяжестью которых
тонкие ножки сгибаются и споры оказываются на субстрате.
Геном диктиостелий полностью секвенирован. Оказалось, что он ко¬
дирует синтез около двенадцати с половиной тысяч белков. Сравнение
их белков и нуклеиновых кислот с другими организмами показало, что
диктиостелии возникли очень рано, вскоре после расхождения первичных
эукариотных организмов на две ветви — растения и животных — и на¬
ходятся на второй ветви, причем отошли на нее раньше, чем отделились
друг от друга грибы и животные.
Глава 9
Грибы и человек
Грибы окружают человека дома, на работе и на от¬
дыхе. Поэтому и связи людей с грибами многообразны
и могут быть как положительными, так и отрицатель¬
ными. В этой главе будут приведены сведения о том,
как человек борется с вредными грибами и использует
полезные.
§ 1. Как грибы портят нам кровь
1.1. Грибы — возбудители болезней
полезных растений
Об этих грибах много было сказано в главе 5. Оста¬
ется лишь добавить немного о том, к чему в истории
человечества приводили массовые заболевания расте¬
ний, эпифитотии, и как люди борются с ними.
Разрушительные грибные эпифитотии приводили
к следующим последствиям:
1. К огромным материальным потерям. В главе 4 опи¬
сано поражение кукурузы в США Т-расой гри¬
ба Кохлиоболус. В результате страшнейшей эпи¬
фитотии потери американских фермеров только
в 1970 году превысили миллиард долларов.
2. К массовой гибели населения от голода и сопро¬
вождающих голод болезней. В главе 5 приведены
данные об эпифитотии фитофтороза картофеля в
Ирландии в середине XIX века, приведшей к гибе¬
ли миллиона жителей этой страны и эмиграции в
Америку полутора миллионов. К еще более страш¬
ным последствиям привела эпифитотия гельмин-
тоспориоза риса в 1942 г. в Бенгалии. Неурожай
риса, практически единственной продовольствен¬
ной культуры этой части Индии, заставил местное
население покидать деревни и уходить в города —
Калькутту и Дакку, где также было нечего есть. А анг-
§ 1. Как грибы портят нам кровь
183
лийскому правительству (Индия в то время была английской коло¬
нией) было не до помощи голодающим — оно было занятой войной
с немцами и японцами. В конце года все дороги были усеяны трупами
голодающих — погибло более двух миллионов человек.
3. К замене зон выращивания важных сельскохозяйственных культур.
В главе 5 было уже рассказано, как под влиянием эпифитотии ржав¬
чины основное производство кофе переместилось из юго-восточной
Азии (острова Цейлон) в Южную Америку.
4. К изменению ландшафтов на больших территориях. В главе 2 упо¬
миналось о возбудителе рака коры каштанов грибе Крифонектрия
паразитика. Через несколько лет после попадания гриба в Северную
Америку произошло почти 100 %-е поражение каштанов, усыхание
и гибель деревьев выше пораженного места. Снятие апикального до¬
минирования вследствие гибели верхушки дерева вызвало пробуж¬
дение спящих почек в прикорневой части дерева. Но отрастающие
побеги, достигнув 15-летнего возраста, вновь поражались, и их вер¬
хушки усыхали. В результате прошедших эпифитотий каштановые ле¬
са в восточных штатах США превратились в кустарниковые пустоши.
5. К массовым отравлениям людей микотоксинами. В главе 4 было по¬
дробно рассказано об отравлениях населения токсинами спорыньи,
фузариотоксинами и другими вторичными метаболитами грибов.
Конечно, терпеть подобные бесчинства грибов люди не могли, и вот
уже более ста лет множество ученых — микологов и фитопатологов, гене¬
тиков, биохимиков и химиков — разрабатывает способы защиты растений
от болезней, а многочисленные фирмы во многих странах мира заняты
производством устойчивых к болезням сортов растений и химических
средств их защиты.
Начиная с 20-х годов прошлого столе-
тия, великий русский ученый Н. И. Вавилов елек1^астТн^болмней
стал собирать в разных странах, которые, со¬
гласно его учению, были генетическими центрами формирования тех или
иных видов культурных растений, семена местных сортов и видов и приво¬
зить их в Ленинград, в институт растениеводства, который он возглавлял.
Семена выращивали на опытных полях и оценивали выросшие растения
по разным признакам, в том числе по устойчивости к болезням. Затем
Вавилов стал посылать в экспедиции по сбору семян своих сотрудни¬
ков, и вскоре этим стали заниматься ученые всего мира. В результате
подобных работ были собраны огромные коллекции видов и примитив¬
ных, местных сортов растений, и выделены среди них формы, устойчивые
к болезням. Затем генетики определили, сколькими генами определяется
устойчивость исследуемого образца растения к болезни, и на каких хро¬
мосомах эти гены расположены. Были разработаны методы передачи этих
генов в культивируемые сорта. Так селекционеры создавали устойчивые
184
Глава 9. Грибы и человек
сорта сельскохозяйственных растений, которые занимают сейчас большие
посевные площади. Однако наряду с несомненными успехами, селекция
устойчивых сортов знала и сокрушительные провалы. Дело в том, что
скорость эволюции растений (а селекция, по выражению Н. И. Вавилова,
это эволюция, направляемая человеком) несравнима со скоростью эволю¬
ции грибов. В год растение дает одну генерацию семян (при выращивании
в фитотронах с регуляцией температуры влажности и освещенности можно
получить в году несколько поколений отдельных видов растений), а гриб
образует десятки генераций. Одно растение образует десятки, редко —
тысячи семян, а гриб в одном пятне формирует сотни тысяч спор. Поэто¬
му редкие генетические изменения, маловероятные у растений, обычны
у грибов. И очень часто наблюдались ситуации, когда устойчивый сорт,
на создание которого ушли годы, вскоре после начала коммерческого вы¬
ращивания поражался новым штаммом гриба-паразита.
На помощь селекции пришла молекулярная биология. Во-первых, ис¬
следования молекулярных механизмов взаимоотношений гриба и растения
позволили установить, какие молекулы паразита (их называют элисито¬
рами) узнаются устойчивым к нему растением, какова структура рецеп¬
торов этих элиситоров, какими путями сигнал тревоги передается в ядро
растительной клетки и как изменяется экспрессия ядерных генов, на¬
конец, какие вещества, убивающие паразита, начинают синтезироваться
в зараженной клетке. Во-вторых, были разработаны методы генной инже¬
нерии, которые позволили переносить в целевое растение гены не толь¬
ко из близкородственных видов (это удавалось и с помощью скрещива¬
ний), но и из любых организмов, где желательные гены были найдены —
из бактерий, грибов, животных. То есть молекулярная биология позво¬
лила создать методы конструирования нужного метаболизма растения.
Простой пример: гриб склеротиния вызывает загнивание многих расте¬
ний (белую гниль). Он часто развивается на корнеплодах моркови в виде
белого ватообразного мицелия, покрывающего загнивший участок кор¬
ня. Еще в XIX веке Антон де Бари установил, что токсином этого гриба,
убивающим клетки растения, является простое соединение — щавелевая
кислота. У одного вида бактерий был обнаружен фермент оксалатдегид-
рогеназа, разрушающий щавелевую кислоту. Ген, кодирующий этот фер¬
мент, был перенесен генно-инженерными методами в маленькое растение
резуху (Арабидопсис), удобное для генетических исследований. Генномо-
дифицированный Арабидопсис стал продуцировать фермент и приобрел
устойчивость к белой гнили. Так что за генно-инженерными растениями
будущее. Без них человечество, численность которого растет, а посевные
площади постоянно изымаются под несельскохозяйственное использова¬
ние, не прокормить. А страх перед генно-модифицированными растени¬
ями обывателей подогревается не научными данными об их вреде (таких
данных нет), а, как всегда, политическими (не пойдем на поводу у амери¬
канцев, которые хотят испортить генофонд русских людей) или экономи-
§ 1. Как грибы портят нам кровь
185
ческими (кому же химические фирмы станут продавать пестициды, если
растения будут устойчивыми к болезням и вредителям) мотивами.
Другой путь снижения вредоносности фитопато¬
генных грибов — уничтожение их с помощью фунги- Химия приходит
цидов, химическая защита растений. История химиче- на помощь
ской защиты растений от болезней прошла несколько
этапов.
Первый этап — использование в качестве фунгицидов неоргани¬
ческих веществ и, прежде всего, королевы фунгицидов в прошлом —
бордоской жидкости, введенной еще в XIX веке французом Милларде
для защиты виноградников от болезни мильдью, вызываемой оомицетом
плазмопара. Действующее начало бордоской жидкости — ион меди в со¬
ставе медного купороса (сернокислой меди). Поскольку медный купорос
вследствие кислой реакции ожигает листья, его нейтрализуют известью.
Кроме меди высокую токсичность для грибов проявляют и другие метал¬
лы, поэтому в состав многих препаратов, применяемых для обеззаражива¬
ния семян от поверхностной грибной инфекции, входила ртуть. Ртутные
препараты были отменены по экологическим соображениям, вследствие
накопления токсичной для многих животных и микроорганизмов ртути
в окружающей среде, а медные — вследствие дороговизны меди и ее солей.
Следующий этап связан с открытием фунгицидных свойств многих
органических соединений. Эти вещества обладают широким антигрибным
спектром действия и низкой токсичностью для растений и теплокровных
животных. Их применение позволило сократить расходы дефицитных ме¬
таллов для нужд сельского хозяйства и упростить процессы приготовле¬
ния и применения рабочих растворов фунгицидов. Открытие все новых
классов органических фунгицидов вызвало создание мощной химической
промышленности, связанной с их производством, в которой заняты де¬
сятки тысяч человек и которая ежегодно производит и продает продукцию
стоимостью сотни миллионов долларов.
Третий этап связан с открытием фунгицидов системного действия.
Дело в том, что используемые на первых этапах органические и неорга¬
нические фунгициды, в сущности, фунгицидами не являются. Ведь ионы
ртути или меди токсичны для любых клеток, будь то клетки гриба или
растения, они убивают грибы, но не обработанные ими растения толь¬
ко потому, что растительная ткань защищена от проникновения в нее
препарата кутикулярным покровом. А отсюда следует, что такого рода
фунгициды бессильны против проникшего внутрь растительной ткани
грибного мицелия, ибо не могут проникнуть вслед за ним. Это создает не¬
обходимость проводить опрыскивания раньше, чем произойдет заражение
растения, и многократно в течение лета повторять обработки. Системные
фунгициды способны проникать в растение, мигрировать в нем и, вслед¬
ствие селективного действия на структуры гриба, но не растения, убивать
грибной мицелий.
186
Глава 9. Грибы и человек
Селективность системных фунгицидов обусловлена специфической
токсичностью для отдельных звеньев обмена веществ, которые присут¬
ствуют у грибов и отсутствуют у растений. Например, обязательный ком¬
понент клеточных стенок многих грибов, хитин, отсутствует у растений.
Антибиотик полиоксин D препятствует работе фермента хитинсинтетазы,
которая складывает длинную цепочку хитина из отдельных молекул-моно-
меров, поэтому полиоксин проникает в растение убивает в нем мицелий
гриба, но не оказывает токсического действия на растительные клетки.
Как уже было сказано в предыдущих главах, в мембране грибов откла¬
дывается эргостерин, который отличаются структурно и от фитостеринов
растений, и от холестерина млекопитающих. В частности, один из эта¬
пов биосинтеза эргостерина предусматривает отрезание метильной груп¬
пы (деметилирование) от 14-го атома углерода молекулы-предшественни-
ка. Это процесс осуществляется специальным ферментом. Синтезировано
много химических соединений, которые ингибирует данный фермент и,
тем самым, ставит блок на пути образования молекулы эргостерина. Эти
соединения стали основой многих фунгицидов, причем не только сель¬
скохозяйственного использования; они входят в состав многих мазей,
продаваемых в аптеках в качестве средств борьбы с грибковыми заболе¬
ваниями ног, например, ламезила. Очень популярны группы химических
препаратов, связывающиеся с белком тубулином, из которого построе¬
ны микротрубочки клеточного цитоскелета, растягивающие к полюсам
хромосомы в процессе деления клетки, и вдоль которых осуществляется
транспорт многих веществ в клетках. Соединение этих препаратов с ту¬
булином грибов и некоторых беспозвоночных животных осуществляется
в концентрациях, на несколько порядков более низких, чем с тубулином
растений и позвоночных животных. Значит, можно подобрать для защи¬
ты растений такие концентрации препарата, которые будут токсичны для
грибов, но не для растений. Созданы также препараты, блокирующие
синтез грибного меланина. Как уже неоднократно было сказано, меланин
откладывается в клеточных стенках многих грибов, а у некоторых фи¬
топатогенных грибов для внедрения в растение необходима меланизация
инфекционной гифы; белый, немеланизированный мицелий растет по по¬
верхности листа, но не внедряется внутрь. А ряд химических соединений
блокирует ключевой фермент синтеза белков РНК-полимеразу, причем
только у оомицетов, поэтому такие соединения используются для борьбы
с возбудителями фитофторозов и ложных мучнистых рос.
Однако системные фунгициды имеют серьезный недостаток, вытека¬
ющий из самой их сущности, — специфические мишени в клетке гриба.
Изменение структуры мишени в результате мутации может привести к
тому, что мишень, при сохранении своих биологических функций пе¬
рестанет связываться с фунгицидом. Например, фунгицид беномил (одно
из его коммерческих названий — фундазол) активно связывается с белком
тубулином у грибов, однако есть масса организмов, имеющих в клетках
§ 1. Как грибы портят нам кровь
187
тубулин, но устойчивых к действию беномила вследствие того, что для
их связывания требуется гораздо более высокая концентрация беномила.
Значит и у грибов возможна мутация в гене, контролирующем синтез
тубулина, которая снизит возможности связывания и сделает гриб устой¬
чивым к фунгициду. Таких устойчивых к беномилу штаммов грибов было
обнаружено очень много, вследствие чего препарат после нескольких лет
использования потерял свою эффективность. Поэтому применять систем¬
ные препараты надо осторожно, с использованием специальных стратегий,
направленных на то, чтобы не допустить или хотя бы снизить скорость
накопления резистентных штаммов в грибных популяциях. Системные
фунгициды химические фирмы выпускают только в смеси с контактными,
которые неспецифичны и направлены на уничтожение штаммов грибов,
резистентных к системному компоненту смеси. Требуется также непре¬
рывный поиск новых классов химических соединений, обеспечивающий
частую смену системных фунгицидов.
Широкое использование фунгицидов, как и других Опасность
химических средств защиты растений (инсектицидов,
ХИ МИЧсСКИХ
токсичных для насекомых-вредителей, гербицидов, уби- средств защиты
вающих сорные растения), может представить опас¬
ность для самих растений (вследствие фитотоксичности, образования ожо¬
гов на листьях), потребителей растений человека и сельскохозяйственных
животных (вследствие наличия остатков пестицидов в продукции) и окру¬
жающей среды (вследствие миграции в почве, воде и воздухе и отравления
организмов немишеней — полезных микроорганизмов, насекомых и про¬
чих). Поэтому в обществе развито отрицательное отношение к химической
защите растений, подогреваемое такими группировками, как зеленые.
Пятьдесят лет назад была опубликована книга американской жур¬
налистки Рейчел Карсон «Безмолвная весна», в которой, на основании
большого собранного материала, был сделан вывод, что, сохраняя ны¬
нешние темпы химизации сельского хозяйства, мы рискуем стать свиде¬
телями страшной весны, когда не зацветет ни один цветок и не запо¬
ет ни одна птица. Книга была написана настолько ярко и убедительно,
что произвела впечатление разорвавшейся бомбы. Горожане стали отка¬
зываться покупать обработанную ядохимикатами продукцию, фермеры
перестали использовать пестициды, федеральное правительство засыпали
бесчисленными запросами, были созданы правительственные комиссии
из компетентных специалистов. Автор книги подверглась жестким на¬
падкам со стороны представителей фирм, занимающихся производством
пестицидов, ученых-энтомологов и фитопатологов. Знаменитый амери¬
канский фитопатолог Дж. Хорсфолл обвинил Карсон в разрушении аме¬
риканского сельского хозяйства. Что же мы видим, спустя полвека после
публикации книги Карсон? Сельское хозяйство не разрушено, произ¬
водство и применение пестицидов продолжается. Но, под влиянием об¬
щественной критики, и в том числе книги Карсон, а также вследствие
188
Глава 9. Грибы и человек
возникших в последние годы новых знаний и технологий ученые и произ¬
водители пестицидов стали более осторожными. Предлагаемые химиками
препараты стали подвергаться гораздо более разнообразным испытани¬
ям, прежде чем получали разрешение к производству, а, следовательно,
стоимость одного препарата от первичного синтеза до выпуска на рынок
выросла на порядки.
Наконец следующий этап в применении химических средств защиты
растений от грибных болезней — использование соединений, которые не
убивают грибы, то есть не фунгицидов, как таковых, а веществ, которые
вмешиваются во взаимоотношения между растением и грибом-парази¬
том, повышают потенциал устойчивости первого или снижают потенциал
патогенности второго. Выше говорилось о химических соединениях, вме¬
шивающихся в синтез грибного меланина. Они не тормозят рост гриба
и не уменьшают интенсивность его спороношения, но не позволяют ему
внедряться в ткань растения.
«Современный интерес к системным фунгицидам внушил мысль о том,
что могут быть найдены материалы, усиливающие врожденные механиз¬
мы устойчивости к болезням. Такие открытия будут ускорены при лучшем
понимании механизма опознания паразитом устойчивого хозяина». Это
написал 40 лет назад известный английский фитопатолог и генетик Питер
Дэй. Сейчас достигнуты большие успехи в понимании механизмов опо¬
знания паразитом хозяина и хозяином паразита, и ряд фирм за рубежом
и в России производит коммерческие препараты, иммунизирующие расте¬
ния, делающие их устойчивыми к поражению фитопатогенными грибами.
Таким образом, селекция и химическая защита растений, начинавшиеся
как совершенно различные по методологии и задачам направления в сель¬
скохозяйственной науке, в настоящее время сливаются в один поток —
усиление потенциала природной устойчивости растений к болезням.
1.2. Грибные болезни домашних животных и человека
Скажем несколько слов о грибных болезнях сельскохозяйственных
и домашних животных. Наиболее широко распространены поражения по¬
кровных тканей — различного рода лишаи. Эти болезни вызывает группа
малосимпатичных грибов, имеющих фермент кератиназу, которая разла¬
гает кератин, белок, из которого построены покровы животных — кожа,
волосы, когти, перья птиц. Поэтому их называют дерматофитами (дер¬
ма — по-гречески кожа). Грибы дерматофиты разделяют на три группы:
геофильные, зоофильные и антропофильные. Многие виды этих грибов
(геофильные) живут в почве, как сапротрофы, и разлагают кератинизо-
ванные остатки трупов животных — перья, кожу, шерсть. От них, по-
видимому, произошли и зоофильные виды, которые с мертвых живот¬
ных перешли на живых. Так возникли зоофильные и антропофильные
виды, которые мигрируют внутри популяций животных и людей. Мно¬
гие сельскохозяйственные животные (коровы, лошади) очень страдают
§ 1. Как грибы портят нам кровь
189
от дерматомикозов. Пораженные места вызывают сильный зуд, животные
расчесывают их до кровавых ран, через которые в организм проника¬
ет бактериальная инфекция. Больные животные снижают привес, удои
молока. Лечение животных также было чрезвычайно болезненным и ма¬
ло эффективным. Корочки, покрывающие ранки отдирали, и оголенную
ткань смазывали антисептиками. Спасение для больных животных пришло
из института экспериментальной ветеринарии, где работал выдающийся
миколог А. X. Саркисов. В его лаборатории впервые в мире были получены
противогрибные сыворотки, нейтрализующие антигены важнейших видов
зоофильных грибов. Инъекция сывороточных антител больным животным
приводила к их излечению. Работы А. X. Саркисова получили высочайшую
оценку у ветеринаров и животноводов во всем мире.
Некоторые зоофильные инфекции могут переходить и на людей.
В сельской местности источниками инфекции человека служат сельско¬
хозяйственные животные, а в городах — домашние (собаки, кошки). Это
позволяет плавно перейти к грибным болезням человека.
Грибы могут вызывать заболевания людей различными способами.
Во-первых, вследствие попадания в организм токсических продуктов об¬
мена веществ — микотоксинов, о которых уже было рассказано. Во-
вторых, вследствие попадания в дыхательную систему грибных спор, со¬
держащих вещества (главным образом, белки), которые воздействуют на
иммунную систему и вызывают аллергию. Часто в жилых комнатах на обо¬
ях или оштукатуренных стенах развиваются темно-зеленые налеты пле¬
сени. Налеты вызваны грибом стахиботрис, споры которого могут вызы¬
вать у жильцов серьезные аллергические заболевания. Споры плесневых
грибов могут адсорбироваться на частицах пыли и с ней попадать в ды¬
хательные пути. Поэтому важнейший инструмент борьбы с плесневыми
грибами в помещениях — пылесос. Особенно опасны профессиональ¬
ные аллергические реакции на грибные споры — заболевания работников
на производстве грибных антибиотиков, выращивании вешенки и т. п.
Наконец, болезни человека, вызванные паразитическими грибами,
разделяют на поверхностные и глубокие микозы. Поверхностные мико¬
зы или дерматофитии вызывают грибы, описанные выше у животных.
Они способны разлагать белок кератин, из которого построены покро¬
вы позвоночных животных, и вызывают поражение кожи, волос, ногтей.
Человека поражают антропофильные виды дерматофитов, хотя возможно
поражение зоофильными, споры которых переходят от пораженного жи¬
вотного к человеку. Давление проростка споры в стыки между клетками
и выделение им ферментов позволяют гифе продвинуться в эпидермисе
до зернистого слоя, где его встречают факторы иммунитета — макрофа¬
ги, антитела и прочие. Поэтому распространение инфекции ограничено
лишь поверхностными клетками. Однако вследствие высокой контагиоз-
ности инфекции, то есть легкому переходу спор от их носителя к здо¬
ровому человеку не только при непосредственном контакте, но и через
190
Глава 9. Грибы и человек
обсемененные спорами вещи, воду в бассейнах и т.п., заболевание имеет
характер эпидемий. Сейчас носителями инфекции является более 20 % на¬
селения. Дерматофитии чаще всего развиваются на пальцах ног и приносят
носителям неприятные ощущения — зуд, покраснения, потливость и не¬
приятный запах ног, но они не смертельны. Есть старая шутка о том, как
кожный врач хвалит свою профессию: «Мои пациенты никогда не подни¬
мают меня ночью с постели, никогда не умирают и никогда не выздорав¬
ливают». Однако сейчас доля правды в этой шутке сильно уменьшилась.
Во-первых, появились в продаже препараты, способные, при правильном
и регулярном их применении не только остановить развитие инфекции,
но и полностью ее подавить. А во вторых... Выше было сказано, что раз¬
витие грибов дерматофитов ограничено покровными тканями и останав¬
ливается после контакта с активно функционирующими клетками внут¬
ренней ткани вследствие наличия линии обороны из иммунных клеток.
Все это происходит так при заражении здорового, как говорят, иммун¬
нокомпетентного организма. Но если у человека произошли нарушения
в иммунной системе, то в линии его обороны возникают бреши, через
которые инфекция из поверхностной может перейти в разряд глубокой,
и тогда, как сейчас говорят, мало не покажется. А поломки иммунной
системы могут быть вызваны разнообразными факторами. Это и вирус¬
ные болезни, такие как СПИД и гепатит, и ксенобиотики в окружающей
нас среде обитания, такие как выбросы химических производств, про¬
дукты сгорания хлорорганических веществ, и психологические стрессы,
и некоторые микотоксины, и ряд лекарственных препаратов. Например,
для купирования опухоли раковых больных подвергают облучению или
химиотерапии. Оба действия приводят к цитотоксическому эффекту, то
есть гибели раковых, а с ними и иммунных клеток. Поэтому глубокие ми¬
козы часто поражают людей в больницах, а большинство людей, больных
СПИДом, погибает от грибных болезней.
Таким образом, возбудителей глубоких микозов можно разделить
на две группы: 1) грибы, исторически связавшие свою судьбу с человеком,
вернее, его внутренними органами, и выработавшими механизмы защиты
от его иммунной системы; и 2) грибы, в принципе, не патогенные, пи¬
тающиеся различными мертвыми субстратами, например, растительными
остатками в почве, но способные развиваться в теле человека, который
вследствие поломок в иммунной системе не может сопротивляться ин¬
фекции. К последней группе может быть отнесено огромное число видов
грибов, лишь бы они а) хорошо росли при температуре 37 ° С (температу¬
ре тела человека) и б) обладали набором ферментов, разрушающих зара¬
женные ткани. Эти грибы называют оппортунистическими, так как они
способны приспосабливаться к различным условиям жизни, как парази¬
тическим, так и сапротрофным.,Сейчас список грибов оппортунистов на¬
считывает около 800 видов, и он постоянно пополняется новыми видами.
§ 1. Как грибы портят нам кровь
191
Серьезные заболевания человека могут быть вызваны
дрожжевыми грибами. Очень широко распространены кан- грибы
дидозы, возбудители которых — дрожжи из рода Кандида.
Это аскомицетные (сумчатые) дрожжи, утратившие половую стадию и раз¬
множающиеся исключительно почкованием. Наиболее патогенный вид —
белая кандида — распространен повсеместно и обнаруживается в смывах
со слизистых оболочек у большинства населения. Он вызывает различные
заболевания, такие как воспаление десен — стоматит, поражение половых
и детородных органов, воспаление легких и т. д. Иногда кандидозы прояв¬
ляются в виде язв на различных участках кожи. Хотя возбудители болезней
обладают механизмами, позволяющими нейтрализовать иммунный ответ
(например, их клетки не перевариваются, будучи поглощенными мак¬
рофагами), но все-таки наиболее опасно, часто с летальным исходом,
протекают заболевания у людей, страдающих иммунодефицитом.
Крайне опасны также базидиомицетные дрожжи из рода криптокок-
кус. Они вызывают криптококковый менингит — заболевание с высокой
степенью вероятности летального исхода, требующее длительного, иногда
пожизненного поддерживающего лечения. Как и кандиды, криптококки
обладают механизмами противодействия иммунной защите зараженных
организмов, их клетки окружены полисахаридной капсулой, защищающие
их от железных объятий рецепторными молекулами макрофагов, притя¬
гивающих к себе и поглощающих чужеродные клетки. Но, в еще большей
степени, чем кандидозы, рост криптококковых инфекций связан с ослаб¬
лением иммунного статуса. В частности, от 5 до 15 % (а в тропических
странах — до 30 %) ВИЧ-инфицированных умирает от криптококкоза.
Другая группа патогенных для человека грибов — ди-
морфные грибы. Они живут в почве в мицелиальной форме грибы
и питаются растительными остатками. Конидии и обрывки
мицелия этих грибов с частицами почвы попадают в дыхательные органы
человека и домашних животных (в частности, лошадей), где прорастают
и вызывают первичное заражение. Возможен перенос спор гриба на черда¬
ки жилых домов животными, которые поражаются в местах естественного
обитания — летучими мышами, птицами. Массовые распространения ко¬
нидий происходят с пыльными бурями, при проведении строительных
работ, исследовании пещер. В теле человека конидии прорастают и осу¬
ществляют первичное заражение. Но при повышенной температуре (37 °С)
в клетках гриба синтезируются так называемые белки теплового шока, ко¬
торые регулируют переход из мицелиального типа роста в дрожжевой (ми-
целиально-дрожжевой диморфизм). Так что патогенной фазой этих грибов
является дрожжевая. Эти грибы вызывают эндемические респираторные
микозы. Эндемические, значит местные, распространенные не повсемест¬
но, как дерматофиты или кандидозы, а только в определенных местах.
Например, бластомикоз и гистоплазмоз встречаются только в некоторых
районах Северной и Южной Америки и, возможно, Африки. Эндемичной
192
Глава 9. Грибы и человек
0
0
зоной гистоплазмоза считается бассейн рек Миссисипи и Огайо в США.
Основной тип заболеваний, вызываемых этими грибами, — поражение
дыхательных органов — бронхиты, воспаление легких. Но у больных,
имеющих нарушения в иммунной системе, возможны и более серьезные
заболевания, такие как поражения костей и т. п.
Наконец среди оппортунистических грибов боль-
Мицелиальные ^ %
ГрИ£ь| шинство потенциальных возбудителей болезней чело¬
века — мицелиальные грибы. Многие из них представ¬
ляют собой обычные тривиальные виды грибов, живущих в почве и обра¬
зующих плесени на продуктах питания, на отсыревающих стенах и других
местах. Например мукоровые грибы, особенно виды рода Ризопус, кото¬
рые вызывают плесневение сочных плодов (садовой земляники, малины)
на грядках и остатков каши в кастрюлях у плохих хозяек, у людей с ослаб¬
ленной иммунной системой, больных сахарным диабетом, использующих
в больших дозах антибиотики или получивших длительное лечение пред-
низолоном, могут вызывать тяжелые, часто летальные заболевания. При
риноцебральном мукоромикозе инфекция через нос поражает глаза, вы¬
зывает лицевой паралич и поражение мозга. При отсутствии лечения чело¬
век в течение недели может умереть. Возможно также поражение легких,
кровеносных сосудов, желудочно-кишечного тракта. К сожалению, врачи
терапевты ранее редко сталкивались с микозами, поэтому при нынешнем
увеличении числа обращений с недомоганиями, диагностируют болезни
как более привычные вирусные или бактериальные и прописывают непра¬
вильные схемы лечения, что усиливает тяжесть последствий заражения.
В связи с ростом числа людей, имеющих повреждения в иммунной
системе, увеличивается число заболевших и мицелиальными грибами из
повсюду встречающегося рода Аспергиллюс. Наиболее расположены к
заболеваниям аспергиллезами больные СПИДом, лейкемией, нарушени¬
ями функций печени, вызванными алкогольной зависимостью и другие.
В общем, одна из самых частых причин смерти ВИЧ-инфицированных —
грибные болезни, ибо для людей, имеющих нарушения в иммунной си¬
стеме, грибы оказываются более агрессивными патогенами, чем бактерии
и вирусы, болезни, вызываемые ими, протекают очень быстро и трудно
поддаются лечению. Это и понятно: грибы, как и мы с вами, имеют эука¬
риотические клетки, в отличие от прокариотических бактерий или вообще
неклеточных вирусов. Большинство противобактериальных антибиотиков
тем то и хороши, что они подавляют синтез белка или клеточной стенки у
бактерий, но не у эукариот. Поэтому не только люди устойчивы к антибак¬
териальным антибиотикам, но и грибы, А антибиотики, убивающие гри¬
бы, высокотоксичны и для человека, поэтому их применение ограничено.
И наконец, нельзя обойти и большую группу одно-
Микроспоридии клеточных паразитов — микроспоридий, поскольку
с недавних пор выяснилось, что они тоже грибы, только очень древние.
В предыдущих главах было рассказано, как микроспоридии паразитируют
§ 1. Как грибы портят нам кровь
193
на насекомых. Но они могут вызывать болезни не только у членистоно¬
гих, но и у позвоночных животных, рыб, птиц и млекопитающих, включая
человека. Предполагают, что они перешли к паразитированию на позво¬
ночных животных и человеке от инфицированных насекомых-паразитов
этих животных. Ранее заболевания человека, вызываемые микроспори¬
диями, отмечались в редчайших случаях у людей с врожденным имму¬
нодефицитом, но сейчас, на фоне иммунодефицита, вызванного ВИЧ-
инфекцией или иными причинами, микроспоридии стали вызывать тя¬
желейшие диареи с летальным исходом, воспаления слизистых оболочек
и другие болезни.
1.3. Грибы — агенты биокоррозии
Грибы не только вызывают губительные болезни у человека, у живот¬
ных, которых он содержит, и у растений, которые он выращивает. Они так¬
же принимают активное участие и в разрушении материалов и изделий, со¬
зданных трудами рук человеческих. Коррозией сначала называли разруше¬
ние металлов под влиянием атмосферных или химических воздействий —
осадков, кислот, высокой температуры, то есть русским аналогом термина
коррозия является ржавчина. Затем термин коррозия получил расширен¬
ное толкование и стал применяться к разрушению не только металлов,
но и других материалов. И, наконец, появился термин «биокоррозия»,
которым стали обозначать повреждения материалов и изделий, вызван¬
ные жизнедеятельностью различных организмов, — от налетов на стенах,
образуемых водорослями или грибами, до погрызов архивных документов
мышами. И здесь грибы оказались активнейшими участниками нападения.
Самые древние постройки и изделия людей были изготовлены из при¬
родных материалов — дерева, камня, костей, поэтому грибам не надо было
перенастраивать свой ферментативный аппарат для их заселения и ис¬
пользования в качестве пищевого субстрата. Уже было рассказано о дре¬
воразрушающих грибах, их ферментах и их роли в жизни леса. Не будь
этих грибов лес постепенно до макушек деревьев покрылся бы опадаю¬
щими ветками. Так что древоразрушающие грибы с честью выполняют
свою экологическую миссию. Но они не знали, что люди будут из сруб¬
ленных, то есть мертвых деревьев, делать постройки и изделия. Для них
мертвое дерево — субстрат, который надо развалить на составные ком¬
поненты и скушать. Поэтому древоразрушающие грибы — бич деревян¬
ных построек. Особенно агрессивна группа базидиомицетов, названная
домовыми грибами. Пучки их сросшихся гиф, белые шнуры, пронизы¬
вают доски и бревна, на поверхности которых образуются плодовые тела
в виде лепешек, а на них — огромное число базидиоспор, коричневой
пылью покрывающих окружающие предметы. Особенно быстро разру¬
шение протекает в местах с высокой влажностью и застойным воздухом
(при отсутствии проветривания). От домовых грибов страдают деревян¬
ные бани, полы в домах, особенно если в стенах фундамента не сделаны
194
Глава 9. Гэибы и человек
большие продухи — отверстия для вентилирования воздуха. Часто разру¬
шаются деревянные сваи мостов, так как в участках на границе вода —
воздух создаются идеальные условия для развития домовых грибов. Ско¬
рость разрушения древесины увеличивается из-за варварских подсечек —
сбора смолы перед рубкой деревьев. Смола содержит терпены — ценные
продукты, используемые в хозяйстве, но эти терпены обладают сильными
антибиотическими свойствами и транспортируются в места механическо¬
го повреждения или заражения (раны в стволах сосны или ели заливаются
смолой). Истощенное сборами смолы дерево становится беззащитным пе¬
ред грибным нападением и быстро разрушается.
Древоразрушающие грибы наносят огромный урон, как отдельным
гражданам, разрушая их деревянные дома и бани, так и народному хо¬
зяйству в целом. Из-за них пришлось заменять деревянные телеграфные
столбы на цементные и деревянные железнодорожные шпалы на хуже пру¬
жинящие бетонные. Это, конечно, дорого, но восполнимо. Однако грибы
могут привести и к трагическим, невосполнимым потерям. Жемчужина
деревянного зодчества, красивейший Покровский собор на острове Кижи
(рис. 45 а) сейчас закрыт для посещения. Причина — плачевное состоя¬
ние бревен, из которых построен собор, и необходимость их реставрации.
И пока не ясно, сумеют ли фирмы, взявшиеся за эту работу, не загубить
собор окончательно.
В Подмосковье, возле платформы «Сенеж» по Ленинградской же¬
лезной дороге, расположена лаборатория по консервации древесины. Ее
сотрудники — микологи, химики и технологи — разрабатывают рецепты
химических обработок, защищающих древесные изделия от разрушения
грибами, и рекомендации по созданию оптимальных климатических усло¬
вий, в которых должны находиться эти изделия. На полигоне лаборато¬
рии — большом земельном участке — испытывают обрубки из древесины
различных пород, наполовину заглубленные в грунт, которые пропитаны
разными химикатами и заражены различными видами древоразрушающих
грибов. Сотрудники лаборатории ведут многолетние наблюдения состоя¬
ния древесины на полигоне.
Менее трагично поражение грибами каменных сооружений. Каза¬
лось бы, как продвигаться мицелию в этом твердом субстрате и что там
есть? Но есть группа грибов, причем группа очень древняя, которые раз¬
виваются на камнях. Их мицелий продвигается в расщелинах между ча¬
стицами камня, питается очень экономно питательными веществами, рас¬
творенными в каплях воды, и выделяет органические кислоты и другие
агрессивные соединения, разрушающие породу. Развитие грибов стимули¬
руют питательные вещества помета птиц, часто поселяющихся на зданиях.
В результате деятельности этих грибов в камне образуются неровности,
каверны, полости, часто окрашенные в черный или темно-зеленый цвет
грибными пигментами (рис. 45 6). Старые, тысячелетние памятники ар-
§ 1. Как грибы портят нам кровь 195
Рис. 45. Грибные повреждения памятников, имеющих историче¬
скую, культурную и культовую ценность: а) Покровский собор на
острове Кижи; б), в) повреждение плесневыми грибами белокамен¬
ной лестницы в палате бояр Романовых (http://www.morozostop.com/)
хитектуры — церкви, замки — в течение своей долгой жизни могут быть
подвержены серьезным разрушениям.
Постепенно люди начали создавать новые, не существующие в природе,
материалы. Одним и первых таких материалов была бумага, которая сыг¬
рала важнейшую роль в развитии цивилизации, ибо столетия оставалась
главным хранителем информации. Поскольку бумага — это чистая цел¬
люлоза, она была быстро освоена грибами-целлюлозолитиками. Поэтому
196
Глава 9. Грибы и человек
пришлось разрабатывать технологии хранения документов, препятству¬
ющие развитие на них грибов, физические и химические способы кон¬
сервирования бумаги. В крупных библиотеках — Государственной биб¬
лиотеке в Москве, Публичной библиотека имени Салтыкова-Щедрина
в Санкт-Петербурге и других созданы специальные лаборатории, где ра¬
ботают микологи.
Тысячелетия существует и живопись, причем древние живописцы ис¬
пользовали для создания красок только природные материалы — мине¬
ралы, масла, яичный белок. Все эти материалы были легко освоены раз¬
нообразными грибами. Поэтому защита произведений искусства — кар¬
тин, фресок, икон — важная отрасль прикладной микологии (рис. 45 в).
Во всероссийском институте художественной реставрации имени Грабаря
работает мощная лаборатория, изучающая грибы, растущие на живопис¬
ных работах и методы защиты полотен.
А в последнее столетие возникло огромное число новых искусствен¬
ных материалов: вместо кожи — ее заменители, вместо стеклянной посу¬
ды — пластмасса и т. п. Микологам приходится выяснять причины помут¬
нения стекол в линзах, повреждения эмульсии на кинопленках, плесневе-
ния строительных материалов, расползания искусственной кожи на сиде¬
ниях в автомобилях, повреждения электроизолирующих материалов про¬
водов и прочее, и прочее. Грибные повреждения могут быть причинами
не только материальных потерь, но и аварий, приводящих к человеческим
жертвам. Например, несколько видов грибов питается углеводородами.
Когда они развиваются в баках с топливом, это еще не очень страшно: ну
образуются хлопья мицелия на дне, но когда попадают в тонкие трубки,
по которым топливо подается в мотор, и забивают хлопьями мицелия
просвет трубки или фильтры, все может окончиться аварией. Хорошо,
если это автомобиль, а если самолет или ракета? И такие случаи бывали.
Поэтому микологи работают и в институтах, занимающихся разработкой
рецептов авиационного топлива и присадок к нему.
§ 2. Как люди используют грибы
Чрезвычайная экологическая пластичность и конкурентоспособность,
умение приспосабливаться к различным климатическим условиям и пи¬
щевым субстратам, ферментативная активность и образование множества
вторичных метаболитов грибов не только вызывает головную боль, но за¬
ставляет придумывать способы использования уникальных способностей
грибов, думать о том, как заставить эти способности служить не во вред,
а на пользу человечеству.
2.1. Пищевые биотехнологии
Первыми объектами пищевых биотехнологий явились дрожжи. Они
дали человечеству два продукта, без которых развитие цивилизации было
§ 2. Как люди используют грибы
197
бы невозможным — хлеб и вино. Говорят, что при-
^ Дрожжи в пищевои
готовление кислого дрожжевого хлеба придумали промышленности
древние египтяне, которые научились хранить за¬
кваску, превращающую сырую плотную массу размешанной в воде муки
в воздушное тесто. А полученные с помощью дрожжей алкогольные на¬
питки — виноградные вина и хлебное пиво — были известны еще раньше,
ибо с забродившим соком люди сталкиваются постоянно с древнейших
времен и до сих пор. В библии описано состояние алкогольного опьяне¬
ния одного из первых людей — патриарха Ноя. Раньше для приготовления
вин люди использовали то, что дано самой природой — дикие дрожжи,
постоянно обитающие на поверхности плодов. Затем началась селекция
штаммов винных дрожжей, так как для виноделов желательно, во-пер¬
вых, чтобы они быстро и эффективно сбраживали сахаристые субстраты,
и, во-вторых, были бы устойчивы к повышенным концентрациям спир¬
та в среде. Процесс брожения приводит к накоплению спирта, который
ядовит для всех микроорганизмов, в том числе и дрожжей. Поэтому при
достижении определенной концентрации спирта процесс брожения оста¬
навливается и дрожжи начинают выпадать в осадок. В связи с этим на¬
туральные (сухие) вина содержат низкую концентрацию спирта (10-12 %)
и долго не хранятся (закисают) после вскрытия. Для увеличения содер¬
жания в них алкоголя вина надо закреплять спиртом. Создание устойчи¬
вых к более высоким концентрациям спирта штаммов дрожжей — одно
из важных направлений их селекции. Таковы, например, хересные дрож¬
жи, придающие натуральным винам хересам довольно высокую крепость.
Сейчас мировое производство дрожжей для хлебопекарной и винодельче¬
ской промышленности составляет миллионы тонн.
Значительно позже плесневые грибы стали использовать для приго¬
товления острых сыров. В магазинах часто можно увидеть сыр рокфор,
срез которого покрыт мраморной сеточкой мицелия зеленой плесени и сыр
камамбер, содержащий белую плесень. Эти сыры имеют специфический
вкус и очень популярны у любителей. Однако злоупотреблять ими все-та¬
ки не стоит, так как среди биологически активных веществ пенициллов,
которыми заражают сыры в процессе их созревания, есть отнюдь не ней¬
тральные для здоровья соединения.
Широко используются в пищевой промышленности и ферменты ми¬
целиальных грибов. Во второй главе книге было рассказано о пектолити-
ческих ферментах — пектиназах, которые грибы выделяют в среду роста
для расщепления пектина — межклеточного цемента, склеивающего от¬
дельные клетки в ткань. При приготовлении плодовых и овощных соков
и вин хлопья пектина выпадают в осадок, часто составляющий треть всего
объема сока. Грибные пектиназы широко используют для расщепления
пектина и осветления соков. Пектиназы и другие ферменты грибов —
а-амилазу, целлюлазы, протеазы и др. применяют для мацерации рас¬
198
Глава 9. Грибы и человек
тительного субстрата при приготовлении масел, кофе, осветления соков
и вин, получения мясного и рыбного белка, сыра.
А с недавних пор диетологи рассказали нам, что растительные масла
не всегда полезны, необходимо, чтобы они содержали омега-3 полинена-
сыщенные жирные кислоты, имеющие как можно больше двойных свя¬
зей в молекуле. У растений таких жирных кислот практически нет, но их
много в рыбьем жире и в клетках водных псевдогрибов — лабиринту-
ломицетов и оомицетов. Так что эти организмы, в недалеком прошлом
рассматриваемые как враждебные силы, паразиты растений, с которыми
надо вести беспощадную борьбу, скоро, возможно, станут предметами лю¬
бовного культивирования.
Ну и, конечно, нельзя обойти вниманием промыш-
Выращивание - -
грибов ленное культивирование съедобных грибов, производ¬
ство которых составляет миллионы тонн и непрерывно
растет. Рост производства съедобных грибов вызван не только ростом
численности населения, но также уменьшением количества и ухудшени¬
ем качества естественных мест обитания дикорастущих съедобных грибов.
Вырубки лесов и загрязнение их продуктами деятельности человек при¬
вели к тому, что во многих районах мира собирать грибы стало просто
небезопасно, так как грибы вследствие особенностей своего адсорбцион¬
ного питания накачивают в плодовые тела ядовитые вещества из воды,
почвы и воздуха.
Человечество пытается приручить грибы очень давно. В странах Юго-
Восточной Азии древоразрушающий гриб сиитаке возделывали еще до
рождества Христова, то есть более двух тысячелетий назад. Большими
любителями грибов были древние римляне. Ранее уже рассказывалось
о том, как широко использовали они ядовитые грибы в дворцовых интри¬
гах. Но если бы они не любили грибную кухню, игры отравителей были бы
невозможны. В романе «Сатирикон» Гая Петрония Арбитра (умер в 65 году
нашей эры) при описании пира у Тримальхиона находим строки: «Да вот
только на днях он написал в Индию, чтобы прислали семян шампиньо¬
нов». Таким образом, можно предположить, что уже в Римской импе¬
рии предпринимались попытки выращивания шампиньонов. В 50-х годах
новой эры грибы размножали путем закапывания их в землю в места,
аналогичные их естественному произрастанию, которые затем поливали.
А в Китае и Корее уже в VI столетии н. э. выращивали иудино ухо или
аурикулярию уховидную.
Грибы, обладая специфическим вкусом и прекрасным запахом, поль¬
зуются заслуженной славой деликатесных продуктов. Но они не только
вкусны, но и питательны.
Наиболее богаты свежие грибы белками; правда, усваиваются они ор¬
ганизмом всего лишь на 40 процентов. Из углеводов в грибах содержится
глюкоза, специфический грибной сахар — трегаллоза (микоза), а также
гликоген — животный крахмал. Кроме того, в свежих грибах много ми-
§ 2. Как люди используют грибы 199
Рис. 46. Плодоношение шампиньонов на стеллажах:
a) Carlig М. J. et al., 2006; б) фото Гарибовой Л. В.
неральных веществ — калия, фосфора, серы, магния, натрия, кальция,
хлора. Пищевая ценность грибов повышается еще и от того, что в них
немало различных витаминов: провитамин А, витамины группы В, вита¬
мин С. Много в грибах витамина Д, а витамина РР столько же, сколько
в говяжьей печени.
Сейчас из всего разнообразия грибного царства культивируют в про¬
мышленных масштабах для пищевых целей чуть больше десятка видов.
Первенство принадлежит шампиньону двуспоровому, объемы мирового
производства которого превышают 1,5 млн тонн в год. Современные сор¬
та шампиньона образуют при плодоношении сплошной слой плодовых
тел на субстрате, которым покрывают стеллажи в помещении (рис. 46). Их
урожай может за две волны плодоношения достигать 20 и более килограмм
на квадратный метр субстрата, а в течение года производят пять-шесть
замен и засевов субстрата (на производственном жаргоне — оборотов).
Поскольку помещения, в которых выращивают шампиньоны, оборудуют
многоярусными стеллажами, можно легко подсчитать, что как по обще¬
му урожаю, так и по выходу белка с единицы площади продукция грибов
во много раз превышает продукцию и растениеводства, и животноводства.
200
Глава 9. Грибы и человек
Выращивание шампиньонов — сложный микробио-
Шампиньоны
логическии процесс, это не огурцы на грядках; современ¬
ные шампиньонницы напоминают скорее заводские цеха, нежели тепли¬
цы. Для создания субстрата (компоста), в котором будет расти грибница,
требуется навоз только от определенных пород животных (конский на¬
воз, полученный от городских, а не от сельских лошадей или птичий
помет), в нем необходимо определенное (не больше и не меньше) со¬
держание азота, он должен быть простерилизован или, по крайней мере,
пропарен. После зарастания компоста мицелием компост покрывают сло¬
ем нестерильной почвы и снижают температуру в помещении, так как
плодоношение индуцируется пониженными температурами и почвенны¬
ми микроорганизмами. Все эти манипуляции, во-первых, требуют нали¬
чия высококвалифицированного персонала, который должен тщательно
соблюдать все технологические режимы, и, во-вторых, удорожают произ¬
водство. Поэтому шампиньоны никогда не станут дешевым продуктом,
они всегда останутся деликатесом.
Вешенка Несколько проще выращивать древоразрушающие грибы,
среди которых в нашей стране массово выращивают только ве¬
шенку. Конечно, по вкусу вешенка уступает шампиньону. Недаром гриб¬
ники в лесу ее не собирают. Один знакомый биолог назвал ее «про¬
должением пня». Но ее производство менее хлопотно, чем производство
шампиньона, а в переработанном (соленом и маринованном) виде она
очень даже вкусна. Разработаны технологии выращивания нескольких ви¬
дов вешенки, среди которых есть виды, образующие удивительно краси¬
вые плодовые тела лимонно-желтой или нежно-розовой окраски (рис. 47).
Их поставляют в рестораны, на приемы, банкеты. Но наиболее крупно-
тоннажное производство — выращивание вешенки устричной, имеющей
серую поверхность плодовых тел. Поскольку древоразрушающие грибы
растут на деревьях, содержащих целлюлозу и лигнин, вешенку выращива¬
ют на таких субстратах, как древесные опилки или стружки, солома злаков
и т. п. Этими субстратами (после их стерилизации или пропаривания) за¬
полняют большие пластиковые мешки с многочисленными перфорациями
по бокам, создают определенную влажность в субстрате и в помещении
и культивируют при повышенной температуре. Перед началом плодо¬
ношения температуру в помещении снижают, так как для плодоношения
многих сортов вешенки необходим Холодовой шок. Через отверстия в меш¬
ке вырастаю пучки (сростки) плодовых тел (рис. 47). Таким образом, при
выращивании ксилотрофных (древоразрушающих) грибов помимо полу¬
чения продукта питания решается еще одна важная задача — утилизация
отходов лесной промышленности (опилки) и сельскохозяйственного про¬
изводства (солома), а также пищевой и легкой промышленности. Можно
использовать и другие отходы. Например, при получении подсолнечно¬
го масла остаются горы лузги, которую не знали, куда девать. Сейчас
за подсолнечную лузгу грибоводы готовы платить маслобойным заводам
§ 2. Как люди используют грибы
201
Рис. 47. Плодоношение вешенки в мешках:
а), в) устричной; б) розовой (фото Л. В. Гарибовой)
202
Глава 9. Грибы и человек
деньги, так как она оказалась прекрасным субстратом для вешенки —
остатки масла в лузге стимулируют рост мицелия. Хорошими субстратами
для вешенки оказались очесы хлопкового волокна, остающиеся при про¬
изводстве тканей на текстильных фабриках, отходы производства пеньки
и прочее. Наконец, остающийся после выращивания плодовых тел суб¬
страт обогащен грибным мицелием, в нем произошло разложение значи¬
тельной части высокомолекулярных неусвояемых биополимеров — цел¬
люлозы и лигнина, поэтому он, после дополнительного компостирования
может быть использован как удобрение или в качестве добавок в корм
скоту и птице. Таким образом, культивирование ксилотрофных грибов —
безотходное многофункциональное производство, при котором осуществ¬
ляется превращение отходов в целевые продукты. Ко всему этому следует
добавить, что ксилотрофы, в отличие от шампиньона, содержат ряд по¬
лезных для здоровья соединений. Но об этом будет рассказано позже.
Шиитаке В° многих странах, особенно на востоке, выращивают
много других видов ксилотрофных грибов. Среди них первое
место по объему производства занимает японский гриб сиитаке (или, как
его чаще называют у нас, шиитаке). Его плодовые тела очень ароматны,
гораздо вкуснее, чем вешенки, и очень полезны для здоровья. Японцы го¬
ворят, что если каждый день съедать по одному сырому грибу сиитеке, то
у вас будет старость без склероза. Этот гриб более требователен к субстра¬
ту, чем вешенка, тем не менее, его мировое производство даже превышает
производство последней. В меньшем объеме выращивают и опята — лет¬
ний и зимний.
При культивировании вешенки и некоторых других ксилотрофов воз¬
никают проблемы, связанные со здоровьем работников. Плодовые тела
шампиньона имеют частное покрывало — пленку, покрывающую пла¬
стинки гименофора с нижней стороны шляпки. При созревании спор эти
пластинки разрываются, но, согласно технологическому регламенту, надо
срывать молодые плодовые тела шампиньонов с закрытым гименофором.
Так что в процессе выращивания и сбора грибов работающие не контакти¬
руют со спорами. А плодовые тела вешенки не имеют частного покрывала
и образуют огромное число спор, которые легко высыпаются и образу¬
ют взвесь в воздухе. Их вдыхание приводит к тяжелым аллергическим
реакциям, поэтому хозяйства по выращиванию вешенки по санитарным
требованиям надо располагать вдали от жилых домов, а работы по уходу
за грибами и их сбору должны проводиться в респираторах, защищающих
дыхательные органы от спор. Генетики и селекционеры многих стран давно
ведут работы по созданию бесспоровых сортов вешенки. В последние годы,
благодаря получению мутаций, нарушающих процессы мейоза, на рынке
появились бесспоровые сорта вешенки, так что проблему можно считать
решенной (размножают вешенку и другие грибы, как картошку, не спо¬
рами/семенами, а вегетативными органами — мицелием/клубнями).
§ 2. Как люди используют грибы
203
Помимо твердофазного культивирования, то есть
- / Культивирование
выращивания мицелия на плотных субстратах (ком- J uul,fllllJ(1 ua
. МИЦсЛИЛ Hd
посте, соломе) и получения плодовых тел, широкое жидких средах
распространение получило глубинное культивирова¬
ние мицелия на жидких питательных средах в ферментерах. Это позволя¬
ет, во-первых, значительно расширить ассортимент съедобных грибов —
кроме базидиомицетов таким образом можно выращивать и более быст¬
ро растущие мицелиальные сумчатые грибы и даже дрожжи. Во-вторых,
при таком способе выращивания значительно увеличивается выход ко¬
нечного продукта. Конечно это не плодовые тела, а мицелий, грибок
нельзя зацепить вилкой, но из мицелия можно создавать грибные пасты,
суповые концентраты, добавлять его в качестве заменителя мяса в колба¬
су или котлеты. Грибной запах, как и питательная ценность, при таком
способе выращивания сохраняются. Беда в том, что глубинное культиви¬
рование имеет ряд отрицательных моментов. Во-первых, возникают те же
проблемы аллергических заболеваний, что и при выращивании вешенки;
во-вторых, как уже было сказано, вследствие иммунодефицита многие
непатогенные грибы становятся патогенными (оппортунистические ин¬
фекции), и это может произойти также с грибами, рекомендуемыми для
выращивания в пищевых или кормовых целях; в-третьих, требуется очень
тщательное соблюдение технологических режимов на всех стадиях про¬
изводства, что, к сожалению, не всегда соблюдается, и срабатывает так
называемый «человеческий фактор». В качестве примера можно привести
историю с производством так называемого белково-витаминного концен¬
трата (БВК), который добавляли в корм скоту. Для его получения было
предложено использовать непатогенные виды дрожжей рода Кандида, ко¬
торые, во-первых, хорошо растут на тяжелых углеводородах, а это — почти
единственное, что у нас пока имеется в избытке, и, во-вторых, быстро на¬
капливают биомассу. Поскольку углеводороды обладают канцерогенными
свойствами, технология предусматривала такой режим питания дрожжей,
при котором на выходе они были бы полностью утилизированы. Кроме то¬
го, пищевая биомасса должна состоять из убитых клеток дрожжей. Но, как
писал JI. Н. Толстой о первой кавказской войне, «гладко было на бумаге,
да забыли про овраги». Что же получилось? Вследствие высокой концен¬
трации дрожжевых клеток в воздухе, возникли аллергические заболевания
у работников производства и жителей близлежащих домов. Мало этого.
Не все циклы выращивания приводили к полному исчезновению углеводо¬
родов в концентрате. Далее. Часть клеток дрожжей в концентрате остава¬
лись живыми. И, наконец, вследствие нестерильного производства (а сте¬
рилизация сильно удорожает работу) в концентрате оказывались поми¬
мо клеток непатогенных дрожжей клетки высокопатогенного вида белой
кандиды. В результате этого иногда после подкормок БВК наблюдались
эпизоотии кандидоза у рыб в прудовых хозяйствах и птиц на птицефермах.
С учетом всех этих факторов от производства БВК пришлось отказаться.
0
204
Глава 9. Грибы и человек
Так что несоблюдение технологических требований при твердофазном
выращивании плодовых тел грозит низкими урожаями и, в крайнем слу¬
чае, разорением производителей, а несоблюдение технических требований
при глубинном культивировании грозит более серьезными последствиями.
Конечно, из выше сказанного не следует, что от глубинного культивирова¬
ния грибов и других микроорганизмов следует отказаться. На этом приеме
основано большинство микробных технологий. В данном случае речь шла
лишь о пищевых технологиях, в которых необходимо повышенное вни¬
мание к соблюдению прописанных режимов.
2.2. Медицинские биотехнологии
пенициллину, за открытие которого и получение очищенного препарата
Флеминг, Флори и Чейн получили Нобелевскую премию. Пенициллин
спас больше человеческих жизней, чем все остальные лекарства вместе
взятые. С его появлением стало возможным лечить болезни, ранее считав¬
шиеся абсолютно летальными, такие как, например, перитонит и гангрена.
Потребность в пенициллине вызвала необходимость, во-первых, строить
большое число производств антибиотика, во-вторых, разработать методы
повышения выхода антибиотика путем оптимизации условий выращива¬
ния (состава питательных сред, регулирования температурного режима
и прочее) и, в-третьих, провести селекционные работы по получению
штаммов грибов, производящих больше пенициллина. В результате таких
селекционных работ, проведенных, в частности в СССР под руководством
С. И. Алиханяна, были получены генетически измененные штаммы гриба,
повысившие выход пенициллина во много раз.
Через несколько лет после массового использования пенициллина,
его эффективность начала падать в связи с появлением резистентных
к пенициллину штаммов стафилококков и стрептококков. В большинстве
случаев их устойчивость к действию пенициллина была обусловлена об¬
разованием фермента пенициллазы, способной связываться с молекулой
пенициллина и вызывать его деградацию. Это заставило усилить поиск
новых антибиотиков, увенчавшийся созданием нового антибиотика цефа-
лоспорина, выделенного из гриба, относящегося к роду Цефалоспориум.
Цефалоспорин имел такую же активную группу, как и пенициллин, и был
сходен с ним по механизму действия на бактерии, но хуже связывался пе-
нициллазой, то есть убивал устойчивые к пенициллину штаммы бактерий.
Однако бактерии приспособились и к разрушению цефалоспоринов, что
заставило ученых создать полусинтетические пенициллины, к молекулам
которых, полученных микробным синтезом, были привешены с помощью
химических реакций радикалы, препятствующие связыванию с бактери¬
альным ферментом. Так что соревнование между скоростью и возможно-
Первая революция:
антибиотики
Химические соединения, полученные из гри¬
бов, совершили две революции в фармакологии и
медицине. Первая революция произошла благодаря
§ 2. Как люди используют грибы
205
стями бактериальной эволюции и разнообразием грибных антибиотиков
вкупе с изобретательностью мозга человека продолжается. И до сих пор,
несмотря на открытие и применение в медицине десятков антибиотиков,
полученных из почвообитающих мицелиальных бактерий, актиномице-
тов, пенициллины и цефалоспорины остаются наиболее востребованными
антибиотиками.
История второй революции может быть сюже- „ ^
, ~ Вторая революция: 0
том для художественного фильма. В 70-х годах про- иммуносупрессоры
шлого века группа микробиологов из швейцарской
фармацевтической фирмы Сантос поехала отдыхать в Норвегию. Но за¬
нятия наукой — это как пристрастие к алкоголю, они не отпускают тебя,
где бы ты ни был и чем бы ни занимался. Вот и эти ребята не только лю¬
бовались фиордами, но и брали образцы почвы из мест, в которых бывали.
Дома, в лаборатории, они выделили из норвежских почв микроорганиз¬
мы и начали их исследование. Один из выделенных грибов, относящий¬
ся к роду Толипокладиум, показал слабую антибиотическую активность,
которая была ниже, чем у коммерческих штаммов актиномицетов и гри¬
бов, поэтому его хотели отбраковать. Но в процессе исследований этого
штамма у него был обнаружен побочный эффект — способностью вызы¬
вать иммуносупрессию, избирательно ингибируя высвобождение интер¬
лейкина-2 из Т-4 лимфоцитов. Интерлейкины — это белки, относящиеся
к группе иммунных молекул цитокинов, которые образуются в иммун¬
ных клетках — макрофагах, В- и Т-лимфоцитах. С помощью цитокинов
иммунные клетки обмениваются информацией о появлении в организме
чужеродных бактерий, вирусов, молекул или даже целых тканей и орга¬
нов, после чего принимается решение о способах нейтрализации чужого.
Иммуномодулятор из гриба Толипокладиум был очищен. Он оказался
циклическим пептидом, содержащим 11 аминокислот, замкнутых в коль¬
цо, и получил название циклоспорина. В отличие от линейных пептидов
(белков), молекулы которых синтезируются на матрице (информацион¬
ной РНК), копирующей структуру ДНК какого-либо гена, и в состав
которых входят сотни и тысячи копий двадцати аминокислот в разных со¬
четаниях, циклические пептиды синтезируются с помощью специальных
ферментов, циклаз, и состоят всего из нескольких аминокислот (5-10),
отличающихся по структуре от 20 тривиальных аминокислот, составляю¬
щих белки. Циклоспорин, ингибируя один из цитокинов (интерлейкин-2),
вмешивается в ход молекулярного диалога между иммунными клетками
и подавляет реакции тканевой несовместимости. А как мы знаем, хирур¬
ги давно разработали методы пересадки органов, позволяющие заменить
плохо работающие почки, сердце, печень и другие на здоровые органы,
полученные от живых или только что погибших доноров. Однако даже
при блестяще проведенных операциях пациенты долго не жили из-за того,
что пересаженные органы отторгались и некротизировались вследствие их
иммунных отличий от собственных тканей и органов. Поэтому возникла
206
Глава 9. Грибы и человек
идея: совмещать хирургические операции пересадки органов с обработ¬
кой циклоспорином. Опыты на животных дали блестящие результаты,
и в 1983 г. было получено разрешение на клиническое применение цикло¬
спорина. Создать такой препарат можно было только при огромной вере
в разработку и при большой финансовой поддержке руководством фирмы
и правительством. Что же касается энтузиазма и веры в препарат, то мож¬
но отметить, что разработчик препарата Дж. Борел апробировал препарат
на себе еще до того, как были проведены исследования его токсичности.
Циклоспорин сделал фирму Сантос одной из богатейших в мире, так
как все клиники мира, проводящие операции по пересадке органов, по¬
купали у нее циклоспорин. И тут норвежское правительство спохватилось
и предъявило фирме иск: ведь продуцент был выделен из норвежской
почвы и, следовательно, был собственностью Норвегии, так что фирму
Сантос попросили делиться. На самом деле грибы-продуценты циклоспо-
ринов обитают не только в Норвегии. Недавно студентка Московского
университета Т. Дарханова выделила коллекцию грибов из почв Бурятии
и в лаборатории известного микробиолога, специалиста в области синтеза
антибиотиков М. В. Бибиковой, нашла в этих почвах несколько штаммов
грибов-продуцентов циклоспоринов.
Для чего же нужен циклоспорин почвенным грибам? Ведь не для того,
чтобы ингибировать иммунные реакции у позвоночных животных. Оказа¬
лось, что нужен, но, конечно, не для этого. Цжбы из рода Толипокладиум
паразитируют на насекомых и образуют на их трупах крупные оранжевые
стромы, описанные в предыдущих главах. Так вот, циклоспорин оказал¬
ся токсичным для насекомых, то есть он — фактор патогенности, одно
из орудий нападения гриба на насекомых.
Также в 80-е годы в клиническую практику был введен и другой
продукт жизнедеятельности грибов — специфический ингибитор синтеза
холестерина ловастатин, который, вместе с его полусинтетическими ана¬
логами статинами, широко используются в клинике для лечения заболе¬
ваний, связанных с отложением холестериновых бляшек на сосудах.
Открытие циклоспорина и статинов стимулировало поиск фармако¬
логических соединений, не обладающих антибиотической активностью, то
есть эти грибные препараты открыли новое направление в фармакологии
природных соединений — лечение не только инфекционных, но и функ¬
циональных болезней. По данным М. В. Бибиковой, начиная с 80-х годов
прошлого века, природные продукты оказались превалирующими в полу¬
чении истинно новых структур с различными биологическими активно¬
стями, в открытии принципиально новых направлений, в создании основ
для получения полусинтетическнх соединений с улучшенными свойства¬
ми. Из 868 введенных за этот период лекарственных средств только 389
было получено в результате химического скрининга. А из полученных для
лечения функциональных заболеваний препаратов более половины проду¬
цируют грибы. Причем поиск продуцентов из разнообразных природных
§ 2. Как люди используют грибы
207
субстратов (почвы, воды, растений, насекомых и т. д.) является важным
направлением в фармакологических исследованиях.
Кроме лекарственных веществ, грибы используют для получения ви¬
таминов и антиоксидантов, которые также продают в аптеках, а не в га¬
строномах, хотя и называют пищевыми добавками. В главе 3 было рас¬
сказано, что половые феромоны мукоровых грибов образуются в резуль¬
тате разложения бета-каротина, поэтому при встрече мицелиев (+) и (-)
штаммов в гифах накапливаются каротиноиды. Это свойство используют
для получения каротиноидов из некоторых видов мукоровых грибов. Для
максимального получения бета-каротина в ферментеры помещают мице¬
лии (или споры) (+) и (-) штаммов в определенных, экспериментально
подобранных соотношениях. Огромное количество каротиноидов обра¬
зуется также в мицелии сумчатого гриба Нейроспора. А мелкий (имеет
самый маленький размер ДНК среди всех эукариот), поганенький гриб
Эшбия, паразитирующий на томате, хлопчатнике, цитрусовых и других
южных растениях, образует огромное количество витамина В2 (рибофла¬
вина) и с удовольствием делится им с людьми при промышленном про¬
изводстве.
А несколько десятилетий назад микологи обратили внимание на ба-
зидиальные макромицеты — шляпочные и трутовые грибы, развивающи¬
еся на древесине. Собственно говоря, обратили внимание на них, как
и на многое другое, древние китайцы и японцы. Но в недавние годы
и европейская медицина признала наличие у них важных лекарственных
соединений. Было показано, что они образуют высокомолекулярные моле¬
кулы полисахаридов — соединенные друг с другом определенным образом
молекулы глюкозы и других простых сахаров, которые обладают иммуно¬
модулирующими (повышают иммунитет) и противораковыми свойствами.
Не только полисахариды базидиальных грибов нашли применение
в медицине. Все настоящие грибы откладывают в клеточной стенке длин¬
ный линейный полимер аминосахаров хитин. Кроме грибов много хитина
образуют членистоногие животные — ракообразные и насекомые. После
добычи и переработки крабов на берегах Дальневосточных морей скап¬
ливались горы крабовых панцирей, от которых не знали, как избавить¬
ся. Сейчас многочисленные фирмы борются за панцири крабов, вернее
за хитин, из которых они состоят. Когда запасы крабовых панцирей при¬
близились к исчерпанию, обратили внимание на пчеловодов, которые
каждую весну выгребают из ульев массу погибших за зиму пчел. Но чле¬
нистоногие все равно не удовлетворяют потребностей в хитине, границы
использования которого увеличиваются. Поэтому начались работы по по¬
лучению грибного хитина. Ведь при различных грибных биотехнологиях,
будь то получение антибиотиков, ферментов, витаминов, органических
кислот и прочего остается биомасса мицелия, обогащенная хитином. Так
что грибы — легко возобновляемый источник хитина, который использу¬
ется как прекрасный адсорбент, более эффективный, чем активированный
208
Глава 9. Грибы и человек
уголь; хитиновые повязки, накладываемые на раны или ожоги, способ¬
ствуют ускорению заживления и снижают риск загноения ран. Кроме
медицинского использования хитин нашел широкое применение в сель¬
ском хозяйстве. Выше было сказано, что полисахариды базидиальных
древоразрушающих грибов повышают иммунитет человека. А хитин по¬
вышает иммунитет у растений к их болезням. Дело в том, что хитин
находится в клеточных стенках грибов — паразитов растений, из хитина
построены стилеты ротового аппарата насекомых и нематод, питающихся
растениями, поэтому растения выработали способность узнавать хитин,
как чужеродное вещество, как сигнал об опасности, и реагировать на не¬
го активизацией работы отдельных генов, продуцирующих вредные для
паразитов вещества. Поэтому обработка растений обрывками молекулы
хитина повышает иммунные свойства. Таким образом, через хитин, мы
плавно перешли от медицинских биотехнологий к сельскохозяйственным.
2.3. Сельскохозяйственные биотехнологии
В сельском и лесном хозяйстве широко используется
Микоризация - ^
микоризация — обработка семян перед посевом препа¬
ратом, содержащим споры или мицелий микоризных грибов. О значении
микоризных грибов в жизни растений было рассказано в главе 5. Семена
сельскохозяйственных растений обрабатывают возбудителями арбускуляр-
ной микоризы — гломусовыми грибами, а лесных пород — мицелием воз¬
будителей эктотрофной микоризы — базидиомицетов. Микоризация рас¬
тений является одним из элементов так называемого адаптивного земледе¬
лия, которое предполагает использование природных биотических связей
растения с микроорганизмами для получения максимальных эффектов,
таких как экономное питание, экологическая пластичность и перенесение
неблагоприятных погодных условий (засухи, жары или, наоборот, холода),
защита от разнообразных болезней и вредителей.
Другое направление сельскохозяйственных био-
регуляторы роста технологий — получение и применение фитогормо¬
нов, продуцируемых грибами. Хотя растения сами
вырабатывают фитогормоны, но вырабатывают их только в нужное для
растения время, в нужном для растения месте и в количестве, не превы¬
шающем необходимого минимума. Но наши потребительские требования
к растению не всегда совпадают с их собственными требованиями, а ино¬
гда и противоречат им. Простой пример — для яблони важно, чтобы ее
плоды после созревания побыстрее сгнивали и освобождали семена, а нам
нужно, чтобы они как можно дольше не портились. Поэтому созданы тех¬
нологии, предусматривающие обработку растений или их отдельных орга¬
нов фитогормонами, регулирующими процессы роста и развития, в част¬
ности, увеличивающими размеры плодов, ускоряющими или, наоборот,
замедляющими их созревание и т. д. Основными поставщиком фитогор¬
монов в этих технологиях являются грибы. В главе 4 было рассказано, как
§ 2. Как люди используют грибы
209
грибы-паразиты растений научились регулировать потоки питательных ве¬
ществ и ростовые процессы растений в своих интересах, значит надо пере¬
хватывать у грибов щ фитогормоны и регулировать ими эти процессы в на¬
ших интересах. К тому же многие грибы несложно и относительно недоро¬
го культивировать в ферментерах и получать большое число различных со¬
единений, в том числе и фитогормонов, то есть решать многоцелевые зада¬
чи, получая в одном производстве и ферменты, и гормоны, и хитин. Кроме
того, метаболиты некоторых грибов, отсутствующие у растений, также об¬
ладают свойствами фитогормонов и могут быть использованы для регуля¬
ции жизненно важных процессов. Например, упомянутый в главе 4 фито¬
токсин фузикокцин не только регулирует работу устьиц, но и стимулирует
рост корней, ускоряет прорастание семян. В лаборатории Г. С. Муромцева
микросодержание фузикокцина было найдено и в незараженных растени¬
ях, то есть фузикокцин — природный регулятор роста растений. Сейчас он
производится на микробиологических заводах для обработки им растений.
Еще одно важное направление использования
грибов в сельскохозяйственных биотехнологиях — ри ная за“*и™
- - „ растении
борьба с вредными для сельскохозяйственных и лес¬
ных растений организмами. В предыдущих главах было рассказано, как
много грибов паразитирует на растениях, насекомых, нематодах. Но по¬
скольку есть много видов сорных растений, насекомых и нематод, на¬
носящих урон полезным растениям, давно возникла мысль использовать
патогенные для них грибы в качества средства, снижающего их числен¬
ность. Были созданы технологии массового размножения спор грибов,
патогенных для сорных растений и вредных насекомых, и обработки ими
посевов и лесных массивов.
В 80-х годах прошлого века был настоящий бум по использованию
этих технологий. Биологический метод защиты растений рассматривался
как безопасная альтернатива химический защиты растений с помощью
синтетических пестицидов. Были созданы микогербициды (грибы, уни¬
чтожающие сорняки), микофунгициды (грибы, паразитирующие на гри¬
бах) и микоинсектициды (грибы, паразитирующие на насекомых). В США
возникли фирмы, специализирующиеся на размножении фитопатогенных
грибов — микогербицидов.
Однако в последние десятилетия появились новые
технологии, которые оказались экономически более вы- Организмы
годными, чем обработка растений спорами патогенных
для вредителей грибов. Например, давно известна патогенная для на¬
секомых бактерия — тюрингская бацилла, которую размножали в фер¬
ментерах и клетками которой опрыскивали с помощью самолетов поля
сахарной свеклы против свекловичного долгоносика и кедровые леса про¬
тив сибирского шелкопряда. Затем было выяснено, что основной фактор
патогенности этой бациллы — белки, названные дельта-токсинами, кото¬
рые специфически связываются с рецепторами в кишечнике насекомых
210
Глава 9. Грибы и человек
и образуют поры в клеточных мембранах, после чего клетки погибают.
В геноме бактерий были локализованы гены, ответственные за синтез
токсинов, которые методами генной инженерии перенесли в целевые рас¬
тения. Токсин стала вырабатывать не бактерия, которую надо размножать
и разбрызгивать над посевами, а трансгенная картошка, которая перестала
быть съедобной для колорадского жука. Использование таких трансгенных
растений экологически безопаснее, чем применение пестицидов и даже
обработки посевов бактериями. Обработка посевов, как химическими ин¬
сектицидами, так и патогенными для насекомых бактериями приводит
к гибели не только вредных насекомых, но и полезных — опылителей,
пчел, насекомых, паразитирующих на вредителях. А на трансгенных рас¬
тениях будут погибать только насекомые, питающиеся непосредственно
этими растениями, то есть мы получаем самонаводящееся оружие, стре¬
ляющее точно по цели. Также пришлось закрыться некоторым фирмам
по производству микогербицидов. Дело в том, что в 70-х годах прошлого
века был запатентован гербицид глифосат, который подавлял активность
фермента, необходимого для синтеза ароматических аминокислот — фе¬
нилаланина, тирозана и триптофана. Все три аминокислоты входят в со¬
став различных белков и поэтому абсолютно необходимы. Кроме того,
с помощью различных ферментов в клетках из них производятся многие
небелковые соединения, очень важные для жизни организмов — фено¬
лы, флавоноиды и прочие. В отличие от растений и микроорганизмов
животные этого фермента не имеют, да он им и не нужен, так как не¬
обходимое количество ароматических аминокислот животные и человек
получают с пищей. Поэтому глифосат чрезвычайно слабо токсичен для
животных. Но он вызывает гибель всех растений без разбору, и вредных,
и полезных. Поэтому им надо пользовать очень осторожно — до появле¬
ния всходов полезных растений. Дачники иногда глифосатом (его продают
под названием раундап) смазывают листья сорняков, выросших на цветоч¬
ных клумбах, с помощью кисточки. Затем у некоторых микроорганизмов
и некоторых устойчивых к глйфосату растений был обнаружен другой
фермент, который также участвует в синтезе ароматических аминокис¬
лот, но не чувствителен к глйфосату. 1Ьн, контролирующий синтез этого
фермента с помощью методов генетической инженерии был перенесен
в сою, кукурузу, хлопчатник и другие растения, что позволило обрабаты¬
вать их глифосатом в период вегетации: чувствительные сорняки погибали,
а устойчивые трансгенные растения оставались неповрежденными. Этот
метод защиты растений от сорняков оказался более эффективным, чем
применение микогербицидов. Но биологические методы защиты расте¬
ний имеют важное значение: а) в странах, где законодательно запрещено
выращивание генномодифицированных растений, для защиты растений,
не имеющих генетической вставки; б) в теплицах; в) в целях уничтожения
посевов незаконно выращиваемых наркотических растений и во многих
других случаях.
§ 2. Как люди используют грибы
211
2.4. Промышленные биотехнологии
Трудно перечислить все отрасли народного хозяйства, в которых мог¬
ли бы быть использованы грибы. Их них получают органические кислоты,
ферменты, полисахариды и т.д. Поэтому остановлюсь лишь на двух на¬
правлениях.
Производство из древесины целлюлозы, а из нее
бумаги — чрезвычайно энергозатратное и экологи- бумажная
чески грязное производство, ибо приходится иметь промышленность
дело с одними из самых высокомолекулярных, слож¬
но устроенных и стойких природных соединений — целлюлозой и лигни¬
ном. Древесину для получения целлюлозы обрабатывают горячей серной
кислотой (проводят «варку» целлюлозы), а отходы, называемые «суль¬
фитным щелоком», содержат различные соединения серы, загрязняющие
окружающую среду. Кроме того при производстве целлюлозы накаплива¬
ются горы чрезвычайно стойкого соединения — лигнина. Поэтому есть
много предложений по использованию древоразрушающих грибов и их
ферментов в целлюлозном производстве. Например, многие базидиаль-
ные грибы производят фермент лигнин-пероксидазу, разрушающий лиг¬
нин. Поскольку эти грибы медленно растут, и их культивирование пред¬
ставляют определенные трудности, лигнинпероксидазные гены генноин¬
женерными методами были перенесены в технологически более удобные
виды грибов, например, в дрожжи. А из полисахаридов, составляющих
основу сульфитных щелоков, с помощью дрожжей можно получать эта¬
нол (древесный или гидролизный спирт), необходимый не столько для
производства алкогольных напитков, сколько для получения различных
полимерных материалов и биотоплива. Таким образом, мы перешли к рас¬
смотрению другого важного направления промышленных биотехнологий.
Этанол, полученный из растительного сырья, рас-
* с Получение
сматривается сейчас как экологически безопасная и воз- биотоплива
обновляемая альтернатива продуктам переработки нефти.
Из двух основных моносахаров, находящихся в гидролизатах целлюлозы
(сульфитных щелоках), — глюкозы и ксилозы — сахаромицетные дрожжи
могут превращать в этанол только глюкозу Поэтому были сконструирова¬
ны штаммы дрожжей сахаромицетов, в которые из других видов дрожжей
и из бактерий вставили гены, необходимые для сбраживания ксилозы. Это
была очень остроумная и изобретательная работа, ибо надо было не только
перенести в дрожжевой штамм гены многоэтапного процесса сбраживания
ксилозы, но и избавиться от катаболитной репрессии (описанной в гла¬
ве 2) промежуточными продуктами сбраживания. Использование таких
рекомбинантных штаммов дрожжей привело к более полной переработке
сульфитных щелоков и более высокому выходу спирта.
Другой путь получения биотоплива — использование жирных кис¬
лот определенного состава для добавки к дизельному топливу — создание
«биодизеля». Природные растительные масла или животные жиры при
0
212
Глава 9. Грибы и человек
взаимодействии со спиртом разлагаются с образованием глицерина и мо-
ноалкиловых эфиров жирных кислот, которые могут служить заменителя¬
ми дизельного топлива. При этом необходимы определенные требования
к химической структуре природных жирных кислот. В частности, в семенах
крестоцветного растения рапса и некоторых других масличных растений
содержатся жирные кислоты, которые дают высокий выход «биодизеля».
В Германии в конце прошлого века возник бум производства семян рапса
для извлечения из них масла для дизельных моторов. Однако через не¬
сколько лет произошло отрезвление — большинство посевных площадей
оказались занятыми непродовольственной культурой, а кушать хочется
даже сильнее, чем водить автомобили и трактора. И тогда обратили вни¬
мание на быстро растущие низшие организмы — водоросли и грибы, как
на источники жирных кислот для топлива. Выращивание водорослей в от¬
крытых водоемах возможно только в районах с теплой солнечной погодой.
В СССР такие установки были созданы в Среднеазиатских республиках.
А установки, основанные на использовании тепла теплоэлектростанций
(ТЭЦ), наталкиваются на большие трудности, которые пока не удается
преодолеть. Однако многие грибы в своем мицелии накапливают боль¬
шое количество жирных кислот требуемого состава. Учитывая быстрый
рост грибов, возможность выращивать их на дешевых средах, включая
коммунальные, сельскохозяйственные и промышленные отходы, а также
наличие большого числа хозяйственно важных побочных продуктов (хити¬
на, ферментов и проч.), использование грибов для получения «биодизеля»
очень перспективно. В лаборатории Е. П. Феофиловой в институте мик¬
робиологии РАН было показано, что оптимальный состав жирных кислот,
необходимых для этой цели, и максимальное накопление их в мицелии
происходит в мукоровых грибах зигомицетах, и разработаны технологии
производства из них заменителей дизельного топлива.
2.5. Экологические биотехнологии
Как сказал персонаж повести Дж. К. Джерома по поводу необходи¬
мости финансировать похороны тещи, «во всяком хорошем деле есть своя
отрицательная сторона». Перефразируя это высказывание, можно сказать,
что во всяком плохом действии грибов есть своя положительная сторо¬
на. Уже было неоднократно сказано, что грибы обладают уникальным
набором ферментов, позволяющих осваивать (разрушать и использовать
в пищу) разнообразные субстраты, как природные, так и рукотворные. По¬
этому давно возникли соображения по поводу использования грибов для
разрушения веществ, которые являются отходами производства, ни для
чего уже не нужны и только загрязняют среду. Например, коммунальные
отходы больших городов (и не только больших, да и не только городов)
скапливаются на огромных свалках, сжигать которые опасно не только
из-за риска возникновения пожаров, но и вследствие накопления токси¬
ческих продуктов сгорания полимерных материалов. Поэтому утилизация
§ 2. Как люди используют грибы
213
отходов часто становится не менее дорогим мероприятием, чем производ¬
ство веществ, в конечном итоге попавших в отходы. И здесь на помощь
приходят грибы. Если есть виды грибов, которые растут на углеводородах,
то почему бы не использовать эти грибы для разрушения нефтяных пле¬
нок на воде, образовавшихся после крушения танкеров, разливов топлива
в местах его хранения, вокруг пунктов заправки и прочих местах?
Важной проблемой стала утилизация полимерных материалов, ко¬
торые все шире входят в быт и в промышленность. Таких материалов
в природе не было, поэтому они оказались очень стойкими по отношению
к процессам естественного разрушения. Сейчас микробиологи выращива¬
ют микроорганизмы не в дорогих стеклянных чашках Петри, а в пласти¬
ковых чашках разового пользования (после выращивания чашки не моют,
а выбрасывают в помойку). В лесах Амазонии недавно обнаружили гриб,
способный быстро разлагать полимерные материалы. Это было установ¬
лено, когда вдруг заметили, что колония гриба съела чашку, в которой ее
выращивали.
Можно создавать композиции из сменяющих друг друга видов гри¬
бов для постепенно разложения городских свалок, так, чтобы в конечном
итоге получить субстраты, пригодные для внесения удобрений под сель¬
скохозяйственные растения или для других целей.
Даже невероятные адсорбционные свойства грибного мицелия люди
стали использовать во благо, применяя грибы вместо фильтров, задержи¬
вающих вредные вещества. В зоне Чернобыльской АС грибы, как было
установлено украинским микологом Н. Н. Ждановой, поглощают радиоак¬
тивные вещества из почвы, очищая тем самым местность вокруг мицелия.
Созданы также технологии помещения на почву в местах между промзо¬
ной и жилым комплексом мицелиальных матов, которые будут поглощать
вредные выбросы и создавать зону, благоприятную для роста древесной
и кустарниковой растительности (рис. 48).
Итак, области использования грибов почти также разнообразны, как
и сами грибы, и, к тому же, непрерывно расширяются. В связи с этим,
исследование видового разнообразия грибов, создание коллекций грибов,
описание новых видов, их местообитаний и экологии, разработка меро¬
приятий по спасению редких видов от исчезновения — это не только чисто
научная задача, но возможность вовлечения их в хозяйственное использо¬
вании на благо человечества, ибо возможности, которые дают нам грибы,
неисчерпаемы. В главе 5 я рассказывал о грибах эндофитах, живущих
внутри злаковых трав и защищающих эти травы своими токсинами от по¬
едания животными. Недавно было установлены, что эти грибы обладают
сильнейшими антибиотиками, подавляющими рост других видов грибов.
Но эндофитные грибы не желают расти на искусственных питательных
средах, образуют лишь карликовые колонии. Значит, задача заключает¬
ся в том, чтобы подобрать вещества, стимулирующие рост этих грибов.
А может быть, легче методами молекулярной биологии найти в геноме
214
Глава 9. Грибы и человек
Рис. 48. Технология создания зеленой зоны между населенным пунктом (слева)
и промышленным предприятием (справа) с помощью матов из мицелия
шляпочного базидиального гриба (P.Stamits, 1995)
§ 2. Как люди используют грибы
215
этих грибов уникальные гены и перенести их в быстро растущие виды
грибов? А в главе 2 были упомянуты грибы экстремалы из содовых солон¬
чаков, растущие на щелочных питательных средах. Из них были выделены
разнообразные ферменты — расщепляющие белки протеазы, расщепляю¬
щие жиры липазы и другие, активность которых сохраняется в щелочных
растворах. Такие свойства чрезвычайно важны при создании детергентов,
стиральных порошков и иных моющих средств, в кожевенной, пищевой
и легкой промышленности, косметологии и других производствах. И со¬
всем уж удивительный пример. На еловых шишках в лесу часто можно
увидеть мелкие изящные шляпочные грибочки. Их так и называют: шиш-
когрибы (по латыни — стробиломицеты). У одного из них было выделен
антибиотик, довольно простой по химической природе (производное ак¬
риловой кислоты), который назвали стробилурином. Исследования этого
антибиотика показали, что он подавляет одно из звеньев дыхательной
цепи в митохондриях грибов и псевдогрибов. Такой широкой спектр дей¬
ствия стробилурина на разные фитопатогенные организмы вызвал к нему
огромный интерес. Оказалось также, что это соединение не только уби¬
вает грибы, но и повышает устойчивость растений к разных возбудителям
болезней — вирусам, грибам и бактериям. Поэтому на рынок поступили
многочисленные производные стробилурина, как выделенные из грибов,
так и созданные в результате химических синтезов. Первый стробилурин
был описан в 1993 г., а к 2010 г. различные производные стробилуринов
стали производиться на суммы в многие миллионы долларов и по стоимо¬
сти продаж превзошли все иные фунгициды вместе взятые. Так что грибы
еще долго будут удивлять нас, причем не только огорчать, но и радовать.
Глава 10
Грибы — модели
в научных исследованиях
Поступательное движение естественных наук вооб¬
ще и биологии в особенности — движение, сопровожда¬
ющиеся кардинальными и не очень изменениями в по¬
нимании устройства окружающего мира и нас самих —
происходит вследствие трех событий, или, говоря цер¬
ковным языком, ипостасей: возникновения новой идеи,
нового объекта и нового метода или прибора. Написа¬
ны книги о том, к каким революционным открыти¬
ям привело совершенствование приборов визуализации
мелких объектов — от лупы Левенгука до современных
электронных и конфокальных микроскопов. Гениаль¬
ная идея о том, что используя особенности редупли¬
кации (самоудвоения) нуклеиновых кислот можно их
многократно амплифицировать, то есть получать мил¬
лионы копий участка ДНК, интересующего исследова¬
теля, привело к созданию и совершенствованию прибо¬
ров, с помощью которых это можно делать практиче¬
ски в любой лаборатории. Использование этой полиме-
разно-цепной реакции революционизировало процессы
исследования геномов разных организмов и открыло су¬
ществование не только вокруг нас, но и внутри нас тем¬
ной биологической материи — огромного числа микро¬
организмов, которые не культивируются на искусствен¬
ных питательных средах и о существовании которых мы
не имели никакого представления.
О том, как революционизировали развитие биоло¬
гии новые объекты исследований, также написаны це¬
лые книги. Мендель установил закономерности насле¬
дования признаков благодаря использованию удачного
объекта — гороха, а затем он стал проводить сходные
опыты с другим видом растения — ястребинкой, но
опыты оказались неудачными, потому что объект не
удовлетворял поставленным перед ним задачам. Гений
Глава 10. Грибы — модели в научных исследованиях
217
Моргана выразился не только в том, что он был блестящим эксперимен¬
татором, но и в том, что он ввел в науку изумительно удобный объект —
плодовую мушку дрозофилу. Немецкий физик Макс Дельбрюк, увлек¬
шись лекциями великого русского генетика Н. В. Тимофеева-Ресовского,
стал заниматься изучением природы гена и ввел в исследования новый
объект — бактериофага, чрезвычайно простое устройство которого дало
возможность проводить анализ генома. Бактериофаг вместе с Дельбрю¬
ком разделили славу создателей молекулярной биологии. Кстати, после
бактериофага Дельбрюк обратился к зигомицетному грибу Фикомицес,
который, правда, в отличие от бактериофага, не стал «экспериментальной
лягушкой». Среди растений не было объектов, равных дрозофиле, поэто¬
му исследования биологии растений отставало от исследований биологии
животных и, конечно, микроорганизмов. Но вот нашли маленькую кресто¬
цветную травку — Арабидопсис, которая легко культивируется на искус¬
ственных средах, проходит полный жизненный цикл от семени до семени
за месяц с небольшим и имеет малюсенький для растений геном, в сто
раз меньший, чем геном пшеницы. И дело быстро пошло. Сейчас больше
половины статей в научных журналах, излагающих работы по экспери¬
ментальной ботанике, посвящены исследованиям арабидопсиса.
К сожалению, модельные и практически важ-
ные объекты биологии — это не одно и то же. Осо енности ^ °® 0
т, - t как научных моделей
И бактериофаг, и арабидопсис, и дрозофила мало
интересны людям с практической точки зрения. Это — не пшеница, не ме¬
доносная пчела, не корова или свинья. Счастливое исключение из этого
противопоставления — гриб — сахаромицетные дрожжи. Сейчас при ис¬
следовании дрожжей сделано столько фундаментальных открытий, что
они затмили даже знаменитую дрозофилу. А ведь дрожжи, к тому же, —
важный объект, без которого не обходится ни хлебопечение, ни виноделие.
Вот мы и перешли к грибам, среди которых внимание экспериментальной
биологии привлекают не только дрожжи. Например, о таких объектах, как
описанные в главе 8 слизевики Физарум и Диктиостелиум, ученые не мог¬
ли даже мечтать, пока не исследовали их свойств. Чем же интересны грибы
для биологов не микологов? Они интересны следующими особенностями
своего строения и образа жизни.
1. Большинство грибов — микроорганизмы, хорошо растущие на
простых питательных средах, к ним, как и к бактериям, можно применять
стандартные микробиологические методы исследований. Это означает, что
если один ученый открыл какое-либо явление и подробно описал в своей
статье, как он это сделал, то другие ученые могут в точности повторить
его опыты.
2. В отличие от бактерий грибы — типичные эукариоты, их клетки
устроены также, как и клетки других эукариот, включая человека, по¬
этому установленные у грибов закономерности можно экстраполировать
на клетки высших эукариот.
218
Глава 10. Грибы — модели в научных исследованиях
3. Грибы имеют наименьший по размеру (среди других эукариот)
геном, что облегчает изучение их генетики. Например, сахаромицетные
дрожжи имеют 15 хромосом, но каждая хромосома по своему размеру
меньше, чем ДНК бактерии кишечной палочки. С помощью специальных
методов разделения в электрическом поле хромосомы грибов можно отде¬
лить друг от друга, выделить и изучить. А недавно была сконструирована
искусственная хромосома, которая нормально реплицировалась в дрож¬
жевых клетках.
4. Многие грибы в вегетативном состоянии гаплоидны, что облег¬
чает получение рецессивных мутаций (в диплоидных клетках рецессив¬
ные мутации маскируются доминантным немутантным аллельным геном),
а вследствие многоядерности их клеток летальные мутации, которые вы¬
зывают гибель клетки, можно сохранять в состоянии гетерокарионов.
5. У ряда грибов можно исследовать индивидуальные продукты мейо¬
за (проводить тетрадный анализ), что значительно увеличивает возможно¬
сти гибридологического анализа. В предыдущих главах было рассказано,
что клетки дрожжей сахаромицетов имеют диплоидные ядра, а после мей¬
оза внутри клетки образуется четыре гаплоидных ядра и, соответственно,
четыре аскоспоры. Их можно осторожно изолировать в отдельные пробир¬
ки с питательной средой и изучать, как расщепляются различные признаки
родителей в потомстве. Например, диплоидные дрожжи всегда гетерози¬
готны по локусу спаривания, поэтому в потомстве индивидуальной дрож¬
жевой клетки всегда потомство расщепляется по локусам спаривания на 2а
к 2а. У мицелиальных сумчатых грибов диплоидное ядро перед образова¬
нием аскоспор в сумках кроме двух делений мейоза претерпевает еще одно
митотическое деление, так что образуется восемь аскоспор и расщепление
по типу спаривания составит 4А к 4а. Когда Мендель изучал расщепление
гибридных семян сортов гороха с красными и белыми цветками, у не¬
го во втором гибридном поколении соотношение двух типов растений
никогда не было точным (3 с красными цветками: 1 с белыми цветка¬
ми), потому что от первого деления диплоидного ядра до образования
цветков и семян проходило огромное число митотических делений ядер,
что приводило к отклонениям от точного воспроизведения соотношения
двух типов ядер, образующихся в процессе мейоза. Поэтому приходилось
применять статистические методы, доказывающие, что отклонения экс¬
периментально полученных показателей расщепления от предполагаемых
не превышает заданный предел случайности. А у грибов при образовании
потомства вообще не происходит дополнительных митозов (как у дрож¬
жей и базидиомицетов) или протекает только одно митотическое деление
(как у мицелиальных аскомицетов), поэтому расщепление всегда точно
соответствует расчетному. Это и называется тетрадным (или при наличии
восьми аскоспор) октадным анализом. С помощью тетрадного анализа
можно быстро отличить сумки, в исходном диплоидном ядре которых
Глава 10. Грибы — модели в научных исследованиях
219
не было обмена участками хромосом, кроссинговера (соотношение раз¬
ных ядер в октаде 4 темных: 4 светлых), от сумок, в которых образование
спор сопровождалось обменами участков хромосом (соотношение темных
и светлых спор 2:4:2 или 2:2:2: 2) (рис.49). А по соотношению числа
кроссоверных и некроссоверных сумок можно картировать ген, то есть
определять его расстояние от участка, разделяющего хромосому на два
плеча — центромеры. Таким же образом можно картировать не только
гены, влияющие на окраску спор, но и любые иные, например, контроли¬
рующие такие важные признаки, как синтез тех или иных ферментов или
вирулентность по отношению к растениям, животным, человеку. Только
в этих случаях надо не просто смотреть на веер сумок из раздавленного
перитеция, а изолировать по 8 спор из индивидуальных сумок и изучать
свойства выросшего мицелия.
6. У грибов обнаружено чрезвычайное разнообразие генетических и
гормональных систем регуляции обменов генетической информацией (ге¬
терокариоз, гетероталлизм, моно- и диэцизм, гинандромиксис, димиксис
и диафоромиксис, парасексуальный процесс). Грибы, вероятно, были по¬
лигоном, на котором природа испытывала и отбирала эти системы.
Ниже приведен далеко не полный список выдающих¬
ся открытий, сыгравших серьезную роль в развитии науки * ри ные>>
^ ^ открытия
и практики, которые были сделаны при работе с грибами.
1. Обнаружение первого антибиотика пенициллина и установление
его структуры и механизма токсичности. За это исследование английский
ученый Ф. Флеминг и его соавторы получили Нобелевскую премию.
2. Исследование механизма формирования спор у сумчатых грибов
и разработка теории тетрадного анализа у дрожжей и нейроспоры. Раз¬
работанный в этих работах метод генетического анализа привел к значи¬
тельным научных открытиям, приведенным ниже.
3. Открытие молекулярных механизмов рекомбинации. Как уже было
сказано, если какой-либо признак, например, светлая или темная окраска
аскоспор, контролируется одним геном, то из восьми спор, образовав¬
шихся в одной сумке, четыре споры обязательно будут светлыми и четыре
темными. Однако при изучении большого числа гибридных сумок было
обнаружено, что иногда, очень редко, но обязательно возникают отдель¬
ные сумки с аномальным соотношением спор, например, 5:3, 6:2 и даже
7:1 (рис. 49). Поскольку такие аномалии наблюдали многие исследовате¬
ли, необходимо было найти им объяснение. И английский миколог Робин
Холлидей чисто умозрительно, рисуя разные варианты на бумаге, опубли¬
ковал гипотезу, объясняющую происхождение аномалий в расщеплении, в
самом престижном научном журнале Nature. Он предположил, что наряду
с обменом участками хромосом (двухцепочечных молекул ДНК) в процес¬
се мейоза, который (такой обмен) называется кроссинговером, возможно
220
Глава 10. Грибы — модели в научных исследованиях
Рис. 49. Гибридные сумки из раздавлен¬
ного перитеция гриба нейроспора. Были
скрещены штаммы, имеющие светлые и
темноокрашенные аскоспоры. 1 — сумки,
сформированные без рекомбинации ди¬
плоидного гетерозиготного ядра (расщеп¬
ление 4 темных : 4 светлых споры); 2,
3 — сумки, сформированные в результа¬
те кроссинговера (расщепления 2:4:2 и
2 : 2 :2: 2); 4 — сумки, сформированные
в результате конверсии (расщепление 2:6)
частичное расплетание молекул ДНК, однонитевые разрывы и обмены
однонитевыми участками двух хромосом. А затем нерекомбинирующая
нить в этом участке вырезается, и брешь заделывается информацией,
находящейся в другой хромосоме. Такой тип обмена получил название
конверсии. После публикации статьи Холлидея, ученые начали прово¬
дить эксперименты по ее проверке. Сначала работали с грибами, затем
перешли на исследования более простых (и, стало быть, более удобных)
организмов — бактерий и бактериофагов. Основные положения гипоте¬
зы Холлидея блестяще подтвердились. Были изолированы и ферменты,
осуществляющие отдельные этапы конверсии. Таким образом, изучение
генетики сумчатых грибов дало начало для выяснения молекулярных ме¬
ханизмов рекомбинации всех организмов.
4. Получение биохимических мутантов нейроспоры и создание но¬
вого научного направления — биохимической генетики. Сумчатый гриб
нейроспора способен самостоятельно синтезировать аминокислоты, вита¬
мины, азотистые основания и прочие органические соединения, необхо¬
димые для жизнедеятельности клетки. Поэтому нейроспора может расти
на простой питательной среде, содержащей только источник энергии —
сахар (сахарозу или глюкозу) — и минеральные соли. Такие среды на¬
зывают минимальными. Однако у нейроспоры, как и у других грибов,
можно выделить мутанты, не растущие на минимальной среде вследствие
повреждения гена, на матрице которого производится фермент, управля¬
ющий синтезом какого-либо необходимого вещества. Подобные мутанты
называют биохимическими. Есть методы, позволяющие довольно быстро
определить, какого вещества не хватает тому или иному мутанту для нор¬
мального роста. Если недостающее вещество добавить к минимальной сре¬
де, мутант будет расти на такой среде. Синтез любого конечного продукта
(аминокислоты, витамина и т. п.) из отдаленного предшественника прохо¬
Глава 10. Грибы — модели в научных исследованиях
221
дит в несколько этапов, причем для превращения одного предшественника
в другой, расположенный ближе к конечному продукту, необходим опре¬
деленный фермент. Биохимики давно это предполагали, но конкретно
ничего не знали, ни о промежуточных продуктах, ни о воздействующих
на них ферментах, ни о генах, контролирующих эти процессы. И вот два
американских ученых, генетик Дж. Бидл и миколог Э. Тейтум, придума¬
ли, как это узнать. Они получили у одного штамма нейроспоры большое
число мутантов, нарушающих синтез какого-либо вещества, например,
аминокислоты метионина, и предположили, что среди них могут быть по¬
вреждения разных генов, необходимых для синтеза метионина. А затем эти
мутанты стали сращивать попарно и высаживать на минимальную среду
(без метионина). Как было рассказано во второй главе, в клетках многих
грибов, в том числе нейроспоры, содержится несколько ядер, поэтому при
слиянии двух штаммов возможно совмещение в клетках ядер, полученных
из разных мицелиев. Если два штамма имели повреждения в одном гене, то
совмещение одинаковых ядер не приведет к росту на минимальной среде,
а если были повреждены разные гены, то промежуточный продукт, ко¬
торый один мутант не может трансформировать, будет трансформирован
ядром второго мутанта, и, наоборот, продукт, накапливающийся во втором
штамме, пойдет в дело с помощью фермента, образуемого первым. Проис¬
ходит то же самое, что делают при монтаже поврежденных кинолент: если
в двух лентах повреждены разные кадры, то можно их вырезать и скле¬
ить так, чтобы из двух поврежденных в разных местах лент сделать одну
неповрежденную. Компенсация разных повреждений в диплоидных или
гетерокариотических клетках называется генетической комплементацией.
Таким способом все мутанты были разделены на комплементационные
группы. Далее. Допустим, что синтез какого-либо конечного продукта
проходит через четыре этапа, и переход одного промежуточного этапа
в другой (А в В, В в С и С в D) контролируется отдельным ферментом. Если
мутация нарушает синтез фермента, контролирующего переход вещества
А в В, то в питательной среде будет накапливаться вещество А. У другой
группы мутантов будет накапливаться вещество В и так далее. Сочетание
генетического анализа с биохимическим позволило провести исследова¬
ние путей синтеза многих аминокислот, витаминов и других первичных
метаболитов. Таким образом, исследования, проведенные на нейроспоре,
привели к созданию новой науки — биохимической генетики. В частно¬
сти, возникла основополагающая концепция «один ген — один фермент».
Бидл и Тейтум за свои работы получили Нобелевскую премию.
5. Обнаружение генетической рекомбинации без полового размноже¬
ния (при митозах) и разработка теории парасексуального процесса. Геро¬
ем этих исследований стал гриб аспергилл, у которого группа английских
микологов обнаружила интересное явление. В его вегетативных (не поло¬
вых, как положено, а именно вегетативных) многоядерных клетках иногда,
по ошибке, два ядра сливаются и возникает среди множества гаплоидных
222
Глава 10. Грибы — модели в научных исследованиях
ядер одно диплоидное. Если клетка гетерокариотическая, то возможно
слияние генетически разных ядер. Была разработана методика быстрого
выделения спор, имеющих гетерозиготные диплоидные ядра, и выращены
диплоидные штаммы гриба. Исследования с ними показали, что у неко¬
торых подобных штаммов возникает генетическая рекомбинация, которая
в норме происходит только при половом процессе. Были установлены все
этапы генетической рекомбинации при бесполом размножении, которые
получили название парасексуального (псевдополового) процесса. Факти¬
чески было обнаружено новое, доселе неизвестное, явление.
6. Анализ генетического контроля ядерного цикла дрожжей. Все эу¬
кариотические клетки, от дрожжевой до человеческой, претерпевают сход¬
ные циклические изменения. В течение одной части цикла ядерное ДНК
находится в активном состоянии и управляет биохимическими процесса¬
ми, протекающими в клетке. Эта стадия называется интерфазой. Во вто¬
рой стадии, называемой митозом, происходят сложные процессы удвоения
ДНК и расхождения двух копий каждой хромосомы из центра клетки к ее
полюсам. Отдельные этапы митоза получили специальные названия —
профаза, метафаза, анафаза и телофаза. И наконец, третья стадия, при
которой в центе клетки образуется перегородка и из одной материнской
клетки образуются две дочерние, называется цитокинезом. Как осуществ¬
ляется регуляция этих процессов, почему у разных организмов они про¬
текают с разной скоростью, какие вещества заставляют ядра переходить
из одной фазы ядерного цикла в другую, долгое время было не только не¬
известно, но, главное, никто не понимал, как это можно установить. Ведь,
как было рассказано несколькими строками раньше, для того чтобы изу¬
чать какие-либо вещества или процессы, надо получить мутации, выклю¬
чающие синтез этого вещества или исключающие этот процесс из какой-то
стадии. Однако все этапы ядерного цикла настолько важны для существо¬
вания клетки, что любая мутация летальна, то есть вызывает гибель клетки.
Американский миколог J1. Хартвелл первым догадался, как это сделать. Он
получил полторы тысячи температурочувствительных мутантов дрожжей.
Такие мутанты нормально растут при оптимальной для роста температуре,
но погибают при повышенной (рестриктивной) температуре. Поэтому их
можно поддерживать при нормальной температуре, но, перенося на по¬
вышенную, вызывать гибель клетки. Такие мутанты называют условно¬
летальными. Скрещиваниями друг с другом были выделены комплемен-
тационные группы мутантов и показано, на какой стадии прекращает
протекание ядерного цикла та или иная мутация. Полный цикл сахаро-
мицетных дрожжей от почки до почки составляет примерно 100 минут.
Хартвелл наложил на 100-минутный циферблат полученные им мутации
и установил, какие белки влияют на переход клетки от предыдущей фазы
к последующей, за что был удостоен Нобелевской премии. Он показал,
что переход от одной стадии к другой в клеточном цикле определяется по¬
сменным образованием двух типов белков — короткоживущих циклинов
Глава 10. Грибы — модели в научных исследованиях
223
и долгоживущих протеинкиназ. И что особенно важно, дальнейшие иссле¬
дования показали, что у всех организмов, включая человека, клеточный
цикл регулируется такими же белками, как и те, что у дрожжей обнару¬
жил Хартвелл. А поскольку клеточный цикл является важнейшим, самым
фундаментальным элементом жизни любого организма, регулирующие его
протекание белки чрезвычайно консервативны и мало изменяются в ходе
эволюции. Показано, например, что если с помощью генноинженерных
манипуляций удалить один из жизненно необходимых генов и заменить
его гомологичным геном человека, то претерпевшая эту операцию дрож¬
жевая клетка будет нормально жить и почковаться. Таким образом, мож¬
но сказать, что между простейшим эукариотным организмом, дрожжами
и венцом творения, человеком, больше общего, чем различного.
7. Обнаружение прионных белков у дрожжей и гриба подоспора.
Прионы — это удивительная группа белков, у которых произошли струк¬
турные изменения. Они воздействуют на такой же, но нормальный, не¬
измененный белок, вызывая его изменения. В клетках накапливаются из¬
мененные белки, которые кристаллизуются и образуют очень токсичные
амилоидные формы. Кроме того эти измененные белки вызывают усиле¬
ние собственного синтеза (активацию транскрипции собственного гена).
Прионы называют инфекционными белками, так как, попадая в чужую
клетку, они также вызывают модификацию нормального белка в нем, про¬
воцируя серьезные заболевания, такие как губчатая энцефалопатия (бе¬
шенство) коров, болезнь скрепи (вертячка овец), болезни Кройцфеддта—
Якоба и куру человека и другие. Это — страшные болезни, с высоким
летальным исходом, изучение их чрезвычайно важно и очень сложно. По¬
этому, когда в лаборатории Ленинградского генетика С. Г. Инге-Вечтомова
были обнаружены странные мутанты дрожжей, синтезирующие не менее
странные белки, а американские генетики установили, что данные белки
имеют свойства прионов, это было расценены, как счастливое открытие,
которое позволит изучать природу прионных белков на такой простой мо¬
дели, как сахаромицетные дрожжи. Недаром в московском кардиоцентре
есть лаборатория, сотрудники которой занимаются молекулярной гене¬
тикой дрожжей, в частности, амилоидными белками. Дрожжи не болеют
инфарктом миокарда. Но закономерности клеточной биологии, исследо¬
ванные у дрожжевой клетки, могут помочь в лечении инфаркта. Затем бел¬
ки со свойствами прионов французскими микологами были обнаружены
и у мицелиального гриба подоспора. Так что приходится снова повторить:
грибы не перестают удивлять нас и, по-видимому, еще долго не перестанут.
Словарь основных терминов
Агар (агар-агар) — смесь полисахаридов, получаемая из морских красных
водорослей. В водных растворах образует плотный студень. Использу¬
ется для приготовления твердых питательных сред при выращивании
бактерий и грибов, а также для приготовления кондитерских изделий
(например, мармелада).
Аденозинтрифосфат (АТФ) — химическое соединение, состоящее из азоти¬
стого основания аденина, сахара рибозы и трех остатков фосфорной
кислоты, в которых запасена энергия химических связей. Энергия
АТФ используется для проведения различных процессов, протекаю¬
щих в клетках.
Акразины — вещества, стимулирующие агрегацию амеб акразиевых и
диктиостелиевых миксомицетов и образование ими псевдоплазмодия.
У диктиостелид основной акразин — аденозинмонофосфат.
Актиномицеты — мицелиальные бактерии. Образуют более тонкий, чем
у грибов, мицелий, с клетками, имеющими прокариотное строение.
Алкалофилы — организмы, живущие в щелочной среде.
Аллели — гены диплоидных организмов, расположенные в одинаковых
местах (локусах) хромосомы, сходные по протяженности, структуре
и функциям (фенотипам). Диплоидный организм, два изучаемых ал¬
леля которого одинаковы, называют гомозиготным, а при различии
аллелей организм называют гетерозиготным. У гетерозиготного орга¬
низма один аллель (доминантный) может подавлять фенотипическое
(внешнее) проявление существования второго (рецессивного) аллеля
(например, цветки гороха имеют красную окраску как у гомозиготы
по «красному» аллелю, так и у гетерозиготы, один из аллелей которой
не окрашивает цветки, они остаются белыми). Возможна и частич¬
ная доминантность, при которой у гетерозиготных растений цветки
розовые.
Анастомозы — слияние гиф мицелия. Встречаются А. между гифами одного
или разных мицелиев.
Анизогамия — тип полового процесса, при котором гаметы имеют морфо¬
логические различия (разделяются на мужскую и женскую). Если обе
гаметы подвижны, но одна (мужская) мелкая, активно плавающая,
Словарь основных терминов
225
а вторая (женская) более крупная, малоподвижная, половой процесс
называют гетерогамией, а если женская гамета неподвижна (не имеет
жгутиков), половой процесс называют оогамией.
Антеридий — мужской гаметангий.
Антеридиол — половой феромон сапролегниевых оомицетов, который ин¬
дуцирует образование антеридиев мужскими особями.
Апикальный рост — верхушечный рост за счет деления апикальной клетки.
Апотеций — тип плодового тела аскомицетов, в котором сумки располага¬
ются открыто на поверхности, называемой диском апотеция.
Апрессорий — орган, с помощью которого гифа паразитического гриба
(или проросток его споры) плотно прикрепляется к поверхности ли¬
ста. Имеет вид подошвы или нашлепки.
Аск — см. сумка.
Аскома — см. плодовое тело.
Аскоспора — гаплоидная спора сумчатых грибов, формирующаяся внутри
специальной клетки (сумки, или аска) и освобождающаяся пассивно
вследствие разрушения оболочки сумки или активно вследствие уве¬
личения тургорного давления внутри сумки.
Аскомицеты (сумчатые грибы) — самый крупный по числу видов отдел гри¬
бов, у членов которого после слияния клеток в результате полового
процесса диплоидные ядра редукционно делятся в специальных спо¬
рангиях, называемых асками, или сумками, где формируется восемь
или (реже) четыре аскоспоры.
Аутбридинг — неродственное скрещивание.
Ацидофилы — организмы, живущие в кислой среде.
Базидиома — см. плодовое тело.
Базцдиомицеты — отдел грибов, у которых: 1) основная стадия жизненно¬
го цикла представлена мицелием с двуядерными клетками (дикарио-
ном); 2) в специальных клетках (базидиях) два ядра сливаются и затем
мейотически делятся с образованием четырех гаплоидных ядер; 3) га¬
плоидные ядра мигрируют в окончания выростов базидии (стеригм);
4) окончания выростов расширяются и отделяются перегородками,
образуя четыре базидиоспоры.
Базидиоспоры — гаплоидные споры базидиомицетов, образующиеся в ре¬
зультате мейоза и располагающиеся на выростах специальных клеток,
базидий.
Барраж — гибель сливших (анастомозировавших) клеток двух мицелиев,
имеющих разные гены несовместимости.
Белки теплового шока —* регуляторные белки, образующиеся при повы¬
шенной температуре (у грибов регулируют различные изменения,
в частности, переход из мицелиального строения к дрожжевому).
226
Словарь основных терминов
Биотрофный паразит — паразит, извлекающий питание и энергию из жи¬
вых клеток хозяина.
Биполярный (однофакторный) гетероталлизм — тип регуляции полового
процесса, при котором сливаться разрешено особям, имеющим ал¬
лельные различия в одном локусе (факторе) спаривания.
Брожение — неполная утилизация глюкозы с образованием в качестве
конечного продукта спирта, а не углекислого газа, образующегося
при дыхании. Осуществляют дрожжи.
Вегетативная несовместимость — гибель слившихся клеток штаммов гри¬
бов, различающихся (гетероаллельных) генами вегетативной несов¬
местимости.
Видообразование — возникновение нового вида вследствие мутационных
изменений старого или межвидовых скрещиваний. Различают алло-
патрическое (при географическом разделении двух популяции одного
вида) видообразование и симпатрическое (без географического разде¬
ления) видообразование.
Гамета — гаплоидная половая клетка.
Гаметангий — клетка, в которой формируются гаметы.
Гаметангиогамия — слияние специализированных половых клеток (гаме-
тангиев) без формирования в них гамет.
Гаплоидные ядра — ядра, содержащие одинарный набор хромосом (каждая
хромосома представлена только одной копией).
1Ьстеромицеты — группа базидиальных грибов, имеющих замкнутые пло¬
довые тела.
Гаусторий — вырост инфекционной гифы грибов-паразитов растений,
пробивающий клеточную стенку, но окруженный наружной мембра¬
ной пораженной клетки. Через гаусторий осуществляется транспорт
питательных веществ из зараженных клеток хозяина в мицелий па¬
разита.
Ген — единица наследственной информации организма; участок ДНК,
несущий информацию о синтезе одной субъединицы белка.
Геном — совокупность генов организма, включающая гены ядра (ядерный
геном), митохондрий (митохондриальный геном) и хлоропластов (у рас¬
тений) (хлоропластный геном).
Геносистематика — система классификации организмов, основанная не на
морфологическом сходстве и различии, а на филогении, т.е. сходстве
и различии их геномов, определяемом с помощью сиквенирования
определенных генов.
Гетерогамия — см. анизогамия.
Гетерозигота — зигота, диплоидные ядра которой имеют аллельные разли¬
чия в одном или нескольких генах.
Словарь основных терминов
227
Гетерокариоз — ситуация, при которой в одном талломе или даже в клетках
таллома находятся генетически различные ядра.
Гетероконтные клетки — подвижные клетки, снабженные морфологиче¬
ски различными жгутиками, один из которых (более длинный) пе¬
ристый, а другой — гладкий. Организмы, имеющие гетероконтные
клетки (водоросли, псевдогрибы) объединяют в царство гетероконт,
или страменопил.
Гетеромерный таллом — таллом лишайников, имеющих упорядоченное
внутреннее строение (наружный слой — кора, слой, содержащий
клетки водоросли, сердцевинный слой и прочие).
Гетероталлизм — раздельнополость, выраженная физиологическими раз¬
личиями при отсутствии морфологических различий и обусловлен¬
ная аллельными различиями в одном или двух локусах спаривания.
В первом случае гетероталлизм называют биполярным, или однофак¬
торным, во втором — тетраполярным, или двухфакторным.
Шиений — спороносный слой, состоящий из спороносных элементов (су¬
мок, базидий) и стерильных структур (парафиз).
Пшенофор — структура плодового тела базидиомицетов, на которой фор¬
мируются базидии (часто имеет вид пластинок или трубочек).
Гифа — ветвящаяся нить, структурная единица мицелия.
Глеба — внутренняя спороносная часть плодового тела гастеромицетов.
Гшоканы — полисахариды, полимеры глюкозы, соединенные друг с другом
различными способами и образующие линейные или разветвленные
цепи. Частные случаи глюканов — крахмал и целлюлоза.
Гольджи комплекс — система внутриклеточных пузырьков, в которых про¬
исходит модификация белков, синтез полисахаридов и формирование
клеточной мембраны.
Гомеомерный таллом — таллом лишайников, не имеющий упорядоченного
внутреннего строения.
Дезоксирибонуклеиновая кислота (ДНК) — гигантская молекула, состав¬
ленная чередованием четырех азотистых оснований, соединенных
с сахаром дезоксирибозой и остатком фосфорной кислоты (четырех
нуклеотидов). Структурная прочность молекулы обеспечивается спи¬
ральной скрученностью двух цепей ДНК, соединенных водородными
связями. Основной хранитель информации. В процессе транскрипции
двойная спираль частично расплетается, и на одной из цепей с помо¬
щью специальных ферментов синтезируется комплементарная цепь
матричной (информационной) РНК.
Дейтеромицеты (несовершенные грибы) — грибы (главным образом, сумча¬
тые) частично или полностью утратившие половой процесс.
228
Словарь основных терминов
Дикарион — грибной мицелий, клетки которого содержат по два ядра,
полученных, как правило, от двух родительских клеток в результате
предшествовавшего полового процесса.
Зигомицеты — отдел грибов, большинство видов которых имеют нераз¬
деленный перегородками (неклеточный) мицелий, споры бесполого
размножения, формирующиеся внутри спорангиев, и половой про¬
цесс гаметангиогамию с образованием покоящихся зигот (зигоспор).
Зооспора — подвижная спора, имеющая один (у настоящих грибов) или
два (у псевдогрибов оомицетов) жгутика.
Зооспорангий — клетка, в которой формируются зооспоры.
Изидий — форма вегетативного размножения лишайников; имеет вид ко¬
ралловидного выроста на поверхности таллома, который может отла¬
мываться и разноситься ветром.
Изогамия — тип полового процесса, при котором сливаются морфологи¬
чески неразличимые гаметы.
Инбридинг — близкородственное скрещивание.
Интроны — нетранслируемые (не участвующие в синтезе белка) участки
гена, которые вырезаются после транскрипции (синтеза информаци¬
онной РНК на матрице ДНК) перед трансляцией (синтезом белка
на матрице информационной РНК).
Кариогамия — слияние ядер.
Каротиноиды — желто-оранжевые пигменты, по химической природе тет¬
ратерпены, содержащие 40 атомов углерода и замкнутые на концах
в шестичленные циклы. Каротиноиды, содержащие только атомы
углерода и водорода, называются каротинами, а каротиноиды, имею¬
щие также один или несколько атомов кислорода — ксантофиллами.
Кератин — белок, из которого построены покровы птиц и млекопитающих
(кожа, волосы, перья).
Кератиназы — ферменты грибов дерматомицетов, разрушающие кератин.
Представляют собой щелочные сериновые протеиназы, способные
разлагать кератин, коллаген и эластин.
Клеточный цикл — чередование стадий жизни клетки: 1) интерфаза, в тече¬
ние которой в клетке не происходят внешние изменения, а ДНК хро¬
мосом активно участвует во внутриклеточных синтезах; в конце ста¬
дии интерфазы происходит удвоение ДНК и образование копий каж¬
дой хромосомы; 2) митоз, в ходе которого копии хромосом выстра¬
иваются друг напротив друга в центре ядра, к определенным местам
на хромосомах прикрепляются микротрубочки, которые растягивают
их к полюсам, после чего образуются оболочки двух новых ядер; 3) ци¬
токинез — деление клетки и образование двух дочерних клеток. Гены
и их продукты (белки), влияющие на переход клеточного цикла из од¬
ной стадии в другую, были впервые найдены у дрожжей. У большин¬
Словарь основных терминов
229
ства грибов митоз не сопряжен с цитокинезом, поэтому клетки мно¬
гоядерны, а некоторые грибы (многие зигомицеты) вообще не имеют
перегородок в мицелии, то есть у них цитокинез не протекает.
Клон — генетически идентичное потомство организма. Чаще всего обра¬
зуется в результате бесполого или вегетативного размножения (у гри¬
бов — спорами, у растений — клубнями, луковицами и т.д.).
Конидия — неподвижная (не имеющая жгутиков) грибная спора, форми¬
рующаяся не внутри спорангиев, а непосредственно на мицелии или
на его выростах — конидиеносцах.
Конверсия генов — тип половой рекомбинации, при которой ген, нахо¬
дящийся в одной хромосоме, вырезается и заменяется информацией
аллельного гена из другой хромосомы. Впервые обнаружена у сумча¬
тых грибов.
Кроссинговер — наиболее распространенный тип внутрихромосомных ре¬
комбинаций, при котором происходит обмен участками двух гомо¬
логичных (полученных от разных родителей) хромосом в процессе
мейоза.
Ксенобиотики — неприродные химические вещества, продукты деятель¬
ности человека.
Ксилотрофы — грибы, растущие на живых и мертвых деревьях и разлага¬
ющие древесину.
Кутикула — наружный покров тела. У растений состоит из эфиров жирных
кислот, погруженных в аморфный полимер воск.
Локус — участок хромосомы, занимаемый геном или группой функцио¬
нально связанных генов.
Макромицеты — грибы, формирующие крупные, хорошо различимые не¬
вооруженным глазом плодовые тела.
Мейоз — тип деления диплоидного ядра, при котором происходит редукция
(уменьшение вдвое) числа хромосом, так что дочерние ядра получают
одинарные (гаплоидные) наборы хромосом.
Меланины — темные пигменты. У большинства грибов представляют со¬
бой поликетиды — конденсированные ароматические кольца.
Микориза — грибокорень; симбиотическое образование, включающее ми¬
целий гриба и корень растения. По степени проникновения грибного
мицелия в ткани корня микоризы разделяют на эндотрофные, прони¬
кающие в глубокие слои корня, и эктотрофные, в которых развитие
мицелия ограничено клетками корового слоя.
Микромицеты — грибы, не образующие видимых невооруженным глазом
плодовых тел. Многие из них называют плесневыми грибами.
Микротрубочки — элементы внутриклеточного скелета; полые трубочки
диаметром около 25 нанометров, образованные в результате полиме¬
230
Словарь основных терминов
ризации белка тубулина. Обеспечивают расхождение хромосом к по¬
люсам при делении ядер, входят в состав жгутиков, участвуют во внут¬
риклеточном транспорте веществ.
Микрофиламенты — элементы внутриклеточного скелета; представляют
собой нити белка актина, обеспечивают внутри- и межклеточный
транспорт веществ.
Митоз — деление ядер без редукции числа хромосом (см. мейоз). Перед
началом митоза происходит удвоение числа хромосом, так что каждая
хромосома получает вторую копию. Благодаря сложному механизму
выстраивания копий хромосом в центре (экваторе) ядра напротив
друг друга и растягивания их к полюсам с помощью микротрубочек
осуществляется точное распределение одинакового набора хромосом
по дочерним ядрам.
Митохондрии — энергетические фабрики клетки. Мелкие органеллы, по¬
крытые двумя мембранами. Внутренняя мембрана имеет впячивания,
называемые кристами, на которых находятся окислительные фермен¬
ты. Образующаяся в результате расщепления глюкозы пировиноград-
ная кислота (пируват) в митохондриях через серию окислительных
процессов превращается в углекислый газ, а образующаяся в резуль¬
тате окисления энергия запасается в образующихся в митохондриях
молекулах АТФ, производство которых и является основной «целью»
работы митохондрий. Митохондрии имеют собственную молекулу
ДНК и рибосомы, поэтому часть структурных и ферментных бел¬
ков они образуют сами, а часть получают из цитоплазмы клетки.
Мицелиально-дрожжевой диморфизм — наличие в геноме некоторых грибов
двух программ развития: программы растущего мицелия и программы
почкующихся клеток. Переключение программ осуществляется в ре¬
зультате воздействий условий жизни или совмещения определенных
ядерных генов при гибридизации.
Мицелий (грибница) — совокупность гиф, образующих таллом, гриба.
Мутации — изменения структуры гена в результате замены отдельных ос¬
нований ДНК (нуклеотидов) (точковые мутации) или потери участка
гена (делеции), вставки дополнительного участка (инсерции) и про¬
чих структурных изменений. Мутации разделяют на биохимические
(нарушается синтез какого-либо соединения), морфологические (из¬
меняется внешний вид организма), летальные (приводят к гибели
организма), условно-летальные (вызывают гибель лишь при опреде¬
ленных условиях, например, при повышенной или, наоборот, низкой
температуре).
Некротрофный паразитизм — тип паразитизма, при котором паразит пред¬
варительно убивает клетку хозяина и питается содержимым мертвых
клеток.
Несовершенные грибы — см. дейтеромицеты.
Словарь основных терминов
231
Облигатный паразитизм — тип паразитизма, при котором паразит не может
вегетировать вне тела своего хозяина и не растет на природных или
искусственно приготовленных средах (вне хозяина могут сохраняться
лишь покоящиеся структуры). Облигатные паразиты, как правило,
биотрофы.
Онтогенез — история развития организма от оплодотворенной клетки
(зиготы) до старения и умирания.
Оогамия — форма полового процесса, при которой женские гаметы (яйце¬
клетки) неподвижны.
Оогоний — гаметангий, в котором формируются яйцеклетки.
Оогониол — половой феромон, регулирующий образование оогониев у са-
пролегниевых оомицетов.
Оомицеты — псевдогрибы; организмы, имеющие, подобно грибам, мице-
лиальное строение и абсорбционное питание, но относящиеся к цар¬
ству гетероконтных организмов, или страменопил.
Ооспора — покоящаяся спора (зигота), образующаяся в результате оогам-
ного полового процесса.
Парасексуальный процесс — уникальный способ генетической рекомбина¬
ции, присущий некоторым грибам и оомицетам. Первый этап П. п. —
гетерокариоз — наличие генетически различных ядер в многоядерных
клетках. Второй этап — диплоидизация — случайное слияние гене¬
тически различных ядер и формирование гетерозиготных диплоли-
дов. Затем возможны рекомбинации (смешивание генов, полученных
от разных ядер) в результате протекания некоторых процессов (гап-
лоидизации диплоидных ядер, митотического кроссинговера и др.).
Перидиоль — вместилище небольшого числа базиоспор внутри базидиомы
у некоторых гастеромицетов.
Перитеций — плодовое тело аскомицетов, имеющее вид бутылки без гор¬
лышка или, наоборот, с вытянутым горлышком. На дне перитеция
располагается пучок сумок с аскоспорами, которые после созревания
активно выбрасываются через выводное отверстие.
Пиноцитоз — поглощение мелких клеток (бактерий, дрожжей и прочих)
и вирусных частиц более крупными клетками, не имеющими поли¬
сахаридной клеточной стенки. Крупная клетка наползает на более
мелкую, обволакивает ее своей наружной мембраной, которая раз¬
рывается и замыкается в пузырек. Находящаяся в пузырьке клетка
оказывается внутри хищника и переваривается. Пиноцитоз — основ¬
ной способ питания амеб и миксомицетов. Для вирусных частиц это,
наоборот, способ проникновения внутрь клеток хозяина и размноже¬
ния в них.
Плазмодий — тип таллома настоящих миксомицетов. Содержит много ядер,
не имеет жесткой полисахаридной клеточной стенки и способен к из¬
232
Словарь основных терминов
менению формы и амебоидным движениям с помощью выростов —
псевдоподий. Размеры П. могут варьироваться от нескольких милли¬
метров до более метра.
Планозигота — зигота, способная к активному плаванию вследствие того,
что подвижные гаметы сохраняют жгутики некоторое время после
слияния.
Плектенхима — ложная ткань, образованная не в результате деления кле¬
ток в разных плоскостях, а в результате сплетения гиф мицелия.
Из плектенхимы построены плодовые тела и склероции грибов.
Плодовые тела — специальные структуры сумчатых и базидиальных гри¬
бов, сложно устроенные и часто крупные, внутри или на поверхности
которых происходит кариогамия ядер дикариона, мейоз и формиро¬
вание аско- и базидиоспор. П. т. аскомицетов называют аскомами,
а базидиальных грибов — базидиомами.
Прионы — инфекционные белки, способные модифицировать собствен¬
ные белки организма, в котором они находятся, и стимулировать их
синтез. Вызывают серьезные болезни животных и человека. Белки
со свойствами П, обнаружены у грибов.
Прокариоты — самые древние организмы, не имеющие ядра и других по¬
крытых двойной мембраной органелл (митохондрий, хлоропластов),
хромосом, митоза, полового процесса и т. д. Разделяются на две импе¬
рии — бактерии и археи, которые различаются строением клеточной
стенки, химическим составом мембран и другими признаками.
Протеинкиназы — ферменты, присоединяющие к белкам богатые энергией
остатки фосфорной кислоты. Являются сигнальными молекулами,
с помощью которых внешние сигналы передаются в геном и изменяют
активность отдельных генов.
Псевдоплазмодий — скопление амеб, вегетативная фаза клеточных мик¬
сомицетов (акразий, диктиостелий). От настоящего плазмодия отли¬
чается тем, что отдельные клетки не сливаются, а сохраняют свою
индивидуальность.
Рибонуклеиновая кислота (РНК) — молекула, отличающаяся от ДНК 1)
заменой сахара дезоксирибозы на сахар рибозу; 2) заменой одно¬
го из азотистых оснований; 3) одноцепочечной структурой (только
у некоторых вирусов встречаются двухцепочечные молекулы РНК).
В клетке имеется несколько типов РНК: матричная, или информа¬
ционная (мРНК), построенная по ДНК-матрице, транспортные РНК
в числе 20 вариацйй, каждая из которых связывается с определенной
аминокислотой и транспортирует ее к мРНК; рибосомальные РНК,
из которых построены рибосомы — швейные машинки, сшивающие
в определенном порядке аминокислоты в процессе трансляции.
Ризомицелий — тонкие выросты таллома некоторых хитридиомицетов и
оомицетов, прикрепляющие таллом к субстрату.
Словарь основных терминов
233
Ризоморфы — длинные, крупные мицелиальные тяжи, в стенках наруж¬
ных клеток которых откладывается темный фермент меланин. Выходят
из корней зараженных осенним опенком деревьев и могут, распро¬
страняясь в почве, заражать корни соседних здоровых деревьев.
Сиквенирование — определение последовательного расположения амино¬
кислот в белках или азотистых оснований в нуклеиновых кислотах —
ДНК и РНК. Осуществляется с помощью специальных приборов —
сиквенаторов.
Сиренины — половые феромоны женских гамет, улавливаемые с помощью
специфических рецепторов сперматозоидами и определяющие на¬
правление их движения. Обычно — низкомолекулярные, легко диф¬
фундирующие в водной среде соединения.
Склероций — покоящаяся структура грибов, плотное сплетение гиф, раз¬
мером от долей миллиметра до десятков сантиметров. У некоторых
грибов склероции структурированы: наружные клетки мелкие, с тол¬
стыми оболочками, в которых отложен темный пигмент меланин,
внутренние — крупные, тонкостенные, богатые питательными ве¬
ществами. У других видов склероции более мелкие, с однородным
содержимым.
Соредии — порошковидная структура, обнажающаяся на поверхности ли¬
шайника вследствие разрыва корового слоя, состоит из клеток во¬
доросли, оплетенных гифами гриба, и предназначена для расселения
лишайника.
Спора — одно- или многоклеточное образование, служащее для размно¬
жения низших эукариот (споровых организмов). Подвижные споры
называют зооспорами, неподвижные споры грибов, образующиеся от¬
крыто, называют конидиями.
Спорангий — споровместище, вместилище зооспор называют зооспоран¬
гием.
Страменопилы (соломинотрубчатые) — царство организмов, имеющих ге-
тероконтные клетки — зооспоры и гаметы — со сложноустроенным
перистым жгутиком, трубчатые кристы митохондрий. Включают мно¬
гочисленные виды водорослей, окрашенных в буро-желтоватые тона
и псевдогрибов.
Строма — сплетение гиф, на поверхности или внутри которого располо¬
жены плодовые тела или структуры бесполого размножения.
Сумчатые грибы — см. аскомицеты.
Таллом (слоевище) *— вегетативное тело прокариот и низших эукариот.
В отличие от тела высших организмов, не дифференцировано на ор¬
ганы (листья, стебли и т. п.).
Тетраполярный (двухфакторный) гетероталлизм — тип регуляции полового
процесса, при котором сливаться разрешено только особям, имею¬
234
Словарь основных терминов
щим генетические различия в двух не сцепленных локусах, называ¬
емых А и В. Характерен только для высших базидиальных грибов
(трутовых, шляпочных, гастеромицетов).
Транскрипция — синтез молекулы информационной РНК на матрице ДНК;
считывание информации с ДНК на РНК.
Трансляция — синтез первичной структуры (линейной цепи) молекулы
белка на матрице информационной РНК согласно правилам генети¬
ческого кода (определенное сочетание трех из имеющихся в молекуле
РНК четырех нуклеотидов соответствует одной из двадцати молекул
аминокислот белка); считывание информации с РНК на белок.
Тубулин — белок, из которого построены микротрубочки.
Факультативный паразитизм — способность организма существовать как
в паразитическом состоянии, так и на мертвом субстрате. Факульта¬
тивные паразиты обычно имеет некротрофный тип питания.
Феромоны — регуляторные молекулы, влияющие на поведение организма
через окружающую среду. Половые Ф. регулируют развитие половых
органов грибов и направление движения сперматозоидов; Ф. кле¬
точных миксомицетов акразины регулируют образование псевдоплаз¬
модиев.
Филогения — история развития вида, его генетические связи с другими,
живущими сейчас и вымершими видами. На основании изучения
филогении строят филогенетические деревья, обозначающие точки
расхождения видов друг от друга и длину ветвей.
Фунгициды — химические соединения, убивающие грибы и применяемые
для защиты растений, животных и человека от грибных болезней.
Хитин — линейный полимер глюкозы, каждая молекула которого содержит
аминогруппу и остаток уксусной кислоты. Откладывается в клеточ¬
ных стенках грибов и образует наружный скелет у членистоногих.
Благодаря жесткости своих молекул, X. заменяет грибам целлюлозу.
Хитридиомицеты —* единственный отдел настоящих грибов, виды которого
имеют в жизненном цикле подвижные клетки с один гладким задним
жгутиком. Талломы одноклеточные, амебоидные, могут прикреплять¬
ся к субстрату ризомицелием. Древнейшая группа грибов.
Хламидоспоры — покоящиеся споры грибов, покрытые толстыми, часто
меланизированными оболочками. Обычно образуются вследствие
распада мицелия на отдельные клетки.
Центромера — участок хромосомы, располагающийся в более или менее
центральной части и разделяющий хромосому на два независимо ре¬
комбинирующих плеча. К центромерам прикрепляются микротру¬
бочки, растягивающие гомологичные хромосомы к разным полюсам
при делении ядра.
Словарь основных терминов
235
Цефалодии — выросты различной формы на поверхности талломов ли¬
шайников, в которых наряду с грибными гифами находятся нити
цианбактерий.
Цианобактерии (синезеленые водоросли) — прокариотные организмы, име¬
ющие фотосинтезирующие пигменты (хлорофилл А и др.) и способ¬
ные, подобно растениям, к фотосинтезу, сопровождающемуся выде¬
лением кислорода (оксигенному фотосинтезу). Дали начало хлоро-
пластам всех водорослей и наземных растений.
Цитокинез — разделение материнской клетки на две дочерние путем врас¬
тания оболочки в центре клетки (у животных и грибов) или образо¬
вания в центре материнской клетки целлюлозной пластинки (у рас¬
тений).
Экзоны — участки гена, с которых производится считывание информации
о последовательности аминокислот в белке в процессе трансляции.
Соединяются друг с другом после вырезания интронов.
Эндосимбиоз — форма симбиоза, при которой один симбионт живет внут¬
ри другого. Внутриорганизменный эндосимбиоз характерен для не¬
которых грибов, мицелий которых живет внутри растений. Внутри¬
клеточный симбиоз аэробных бактерий и цианобактерий с первичной
ядерной клеткой привел к образованию митохондрий и хлоропластов.
Эндофитный рост — рост мицелия фитопатогенного или эндосимбиотиче-
ского гриба внутри растения.
Эргостерин — специфический стерин грибов, имеющий химические от¬
личия от холестерина позвоночных животных. Включается в состав
мембран грибов, а также обладает регуляторной активностью.
Эукариоты — организмы, имеющие покрытые двойной мембраной орга-
неллы (ядра, митохондрии, хлоропласты), хромосомы и аппарат их
разделения (митоз), половой процесс.
Яйцеклетка — неподвижная женская гамета.
Источники иллюстраций
1. Беккер 3. Э. Физиология грибов. М.: Изд.-во Московского ун-та, 1963.
2. Беляев И. М, Горленко Л/. А, Дьяков Ю. Г., Лекомцева С. #., Успен¬
ская Г. Д. Болезни и вредители полевых культур: альбом. М.: Сель-
хозиздат, 1970.
3. Белякова Г. А., Дьяков Ю. Г., Тарасов К. Л. Ботаника. Т. 1. Водоросли
и грибы. М.: Академия, 2006.
4. Ботаника. Курс альгологии и микологии / Под. ред. Ю. Т. Дьякова.
М.: Изд-во Московского ун-та, 2007.
5. Бредли С. Защита растений. М.: Кладезь-Букс, 2003.
6. Вент Г. В мире растений. М.: Мир, 1972.
7. Галактионов В. Г. Иммунология. М.: Академия, 2004.
8. Жизнь растений. М.: Просвещение. Т. 2. Грибы / Под ред. проф.
М. В. Горленко. 1976. Т. 3. / Под ред. проф. М. М. Голлербаха. 1977.
9. Кузовникова-Виштынецкая Т. А., Дьяков Ю. Г. Гетерокариоз у Phytoph-
thora infestans. 1. Цитологические исследования. Микология и фито¬
патология. 1973. 7. С. 479-485.
10. Кузьмичев Е. П., Соколова Э. С., Мозолевская Е. Г\ Болезни древесных
растений. Т. 1. М., 2004.
11. Лесото Т. Грибы. ACT, 2003.
12. Adamatzky A. From reaction-diffusion to Physarum computing. Nat Corn-
put. 2009. 8. P. 431-447.
13. Alexopuolos C./., Mims C. W., Blackwell M. Introductory Mycology. Wiley,
1995.
14. Bauchop T Rumen anaerobic fungi and utilization of fibrous feeds. In
Biotechnol. And Recombinant DNA Technol. in the Animal Production
Industries in Australia. Armidall. Australia. 1985. P. 118-123.
15. Carlie M.J., Watkinson S. C.f Gooday G. W. The Fungi. Acad. Press., 2001.
16. Cerda-Olmedo E.y Lipson E. D. Phycomyces. Cold Spring Harbor. 1987.
17. de Hoog G. S., Guarro /., GeneJ., Figueras M. J. Atlas of Clinical Fungi.
CBS. Spain. 2000.
18. Eilenberg J. E. Biology of fungi from order Entromophthorales. Univ.
Copenhagen. Denmark, 2002.
Источники иллюстраций
237
19. Fry W. Е., Grunwald N.J. Introduction to Oomycetes. The Plant Health.
2010.
20. General Mycology. Textbook. Ser. for 21st century. Higer education Press.
2001. (На китайском языке).
21. Hardham A. R. Ceil biology of fungal and oomycetes infection of plant // Bi¬
ology of Fungal Cell. The Mycota. VIII. Springer-Ferlag, 2007. P. 252-289.
22. Horiuchi Fujiwara Л/., Yamashita S., Ohta A., Takagi M. Proliferation of
intrahyphal hyphae caused by disruption of csmA, which encodes a class V
chitin synthase with a myosin motor-like domain in Aspergillus nidulans.
J. of Bacteriology. 1999. 181. P. 3721-3729.
23. Keeling P. J. Diversity and evolutionary history of plastids and their hosts.
American J. Botany. 2004. 91. P. 1481-1493.
24. Money N. P. Biomechanics of invasive fungal growth. In «Biology of Fungal
Cell» The Mycota. VIII. Springer-Ferlag, 2007. P. 2347-2349.
25. Olive L. S. Mycetozoa: a revised classification // Botanical. Rev. 1970. 36.
P. 59-87.
26. Podger F. D. Phytophthora cinnamomi, a cause of lethal disease in indige¬
nous plant communities in western Australia. Phytopathologia. 1972. 62.
P. 972-981.
27. Poggeler S., Nowrousian M., Kuck U. Fruiting biology development in as-
comycetes. In The Mycota. I. Springer-Verlag, 2006. P. 325-355.
28. Read £., Smith S.E. (editors). Mycorrhizal Symbiosis. Elsvier Sci, 2011,
29. Rumpho M. Summer E. Manhart J. R. Solar-powedred sea slugs,
mollusc/algae chloroplasr symbiosis. Plant Physiology. 2000. 123. P. 29-38
30. Stamets P. Mycelium Running. The Speed. Press. 2005.
31. Wessels, J. G. Fungi in their own right. Fungal Genet. Biol. 1999. 27.
P. 134-145.