











































































































































































































































Автор: Максимов А.А. Ердаков Л.Н.
Теги: материальные основы жизни биохимия молекулярная биология биофизика биология
Год: 1985




Текст
А. А, МАКСИМОВ
Л.Н.ЕРДАКОВ
ЦИКЛИЧЕСКИЕ ПРОЦЕССЫ
В СООБЩЕСТВАХ ЖИВОТНЫХ
АКАДЕМИЯ НАУК СССР
СИБИРСКОЕ ОТДЕЛЕНИЕ
БИОЛОГИЧЕСКИЙ ИНСТИТУТ
А. А. МАКСИМОВ
Л. Н. ЕРДАКОВ
ЦИКЛИЧЕСКИЕ ПРОЦЕССЫ
В СООБЩЕСТВАХ ЖИВОТНЫХ
биоритмы, сукцессии
Ответственный редактор
д-р’биол. наук проф. И. В. Стебаев
НОВОСИБИРСК
ИЗДАТЕЛЬСТВО «НАУКА»
СИБИРСКОЕ ОТДЕЛЕНИЕ
1985
УДК 577.49.591.553
Максимов А. А., Е р д а к о в Л. Н. Циклические про-
цессы в сообществах животных (биоритмы, сукцессии).— Ново-
сибирск: Наука, 1985.
В монографии с помощью традиционных методов анализа времен-
ных рядов подтверждено существование циклов разной периодичности
в жизнедеятельности популяций животных. Сообщества мелких млеко-
питающих показаны как полициклические системы. Это дало возможность
по-новому подойти к современной теории сукцессий в биоценозах и пред-
ставить их как элементы природной цикличности. По данным многолет-
них отловов животных разработаны методы биологической индикации
циклов: смена доминирования видов в! сообществе, вспышки численности
отдельных видов. ^Предлагаются численные методы анализа таких мате-
риалов. Выявлена связь популяционной цикличности с изменениями при-
родной среды. Все это позволило считать анализ циклов в сообществе
одним пз реальных подходов к прогнозированию биологических процес-
сов в природе.
Книга рассчитана на биологов, преподавателей, специалистов сель-
ского и лесного хозяйства, работающих в области защиты растений, ме-
дицинских зоологов.
Рецензенты В. Г. Мордкович, Ю. Г. Швецов
2005000000— 909248J_85_iv
042(02)—85
(g) Издательство «Наука», 1985 г.
М
ВВЕДЕНИЕ
Эколог,, работающий в течение ряда лет на одном и том же участке,,
не может не обратить внимание на большую разногодичную измен-
чивость видового состава животного населения. Некоторые виды мо-
гут в текущем году вовсе выпасть из сборов материала или встре-
чаться крайне редко, другие же, которые в предыдущие годы были
случайными в сборах или отсутствовали, оказываются многочислен-
ными. Разовое фаунистическое обследование района может создать
превратное представление о его животном населении. Поэтому по
материалам одного полевого сезона невозможно судить о фауне дан-
ного района, соотношении отдельных видов в населении, доминантах.
Меняются по годам не только виды, господствующие в сообще-
ствах, но и весь биоценоз, происходит глубокая перестройка его.
В то же время существует повторяемость тайих изменений. Домини-
рующий вид одного, года уже был доминантой несколько лет назад,,
а еще через несколько лет вновь увеличивает свою численность.
Некоторые виды животных отмечаются в уловах не ежегодно, а с не-
которой ритмичностью. При длительных наблюдениях обнаружи-
вается, что большая часть происходящих в биоценозе изменений
цпклична.
Проблема цикличности в глазах эколога, анализирующего ди-
намику биологических процессов и ставящего целью их долгосроч-
ное прогнозирование, приобретает принципиальное значение. При-
знавая цикличность процессов, происходящих в природе, и зная
тенденцию их развития, можно заблаговременно прогнозировать
исследуемый биологический процесс. Разработка экологических ос-
нов долгосрочного прогноза — это,, по сути дела, поиск и исследо-
вание цикличности в местных биогеоценозах [Максимов, 1978].
Многолетняя и сезонная цикличность в экологииеских и хроно-
биологических работах изучается редко. Известную трудность пред-
ставляет сбор данных по сезонным ритмам. Длительные наблюде-
ния, чаще всего непрерывны^ многолетние, дают возможность по-
лучить оценки лунйых и месячных режимов жизнедеятельности
животных. Среди немногих экспериментальных исследований та-
ких длиннопериодических ритмов можно назвать работы М. Фин-
германа [1964], К. Хауншельда [1964], Гвиннера [1984а, б]. Су-
ществует и другой, непрямой, способ изучения сезонной и много-
летней цикличности, когда исследователи анализируют ритмы с бо-
3
лее короткими периодами. Так, Г. Гаттерманн [Gattermann, 1978}
для оценки сезонной ритмичности исследовал суточную активность
кроликов. Соотношение и мощность суточных, недельных и месяч-
ных колебаний жизнедеятельности животных от сезона’ к сезону
изменяются. При изучении цикличности численности животных
есть смысл рассматривать не только сезонные циклы, но и более
высокочастотные (месячные; двухнедельные, недельные и пр.). Зна-
чительное число работ по изучению сезонных изменений сделано
при анализе суточной периодичности [Grodzinsky, 1963; Смирнов,
1968; Sawicha-Kapusta, 1969; Калабухов, 1969; Stebbins, 1972;
Thompson, 1977]. Так можно получить значительное число интерес-
ных сезонных характеристик изучаемых процессов,, но нельзя вы-
яснить, например, длину периода исследуемого ритма.
Годовые ритмы у млекопитающих начали изучать относительно
недавно [Pengelley, Fischer, 1963; Gwinner, 1981а, б]. Обзор данных
по цирканным ритмам животных и растений [Berthold, 1974] пока-
зывает, что зти циклы имеют протяженность от 7 до 15 мес (обычно
10—13). Периоды у 54 видов животных были следующими: меньше
года у 19, год у 19 и длиннее года у 16 видов. В некоторых опытах
длительностью до 4 лет цикл повторялся 4—5 раз, причем у отдель-
ных видов он варьировал мало, а у других от года к году значитель-
но различался (до нескольких месяцев). Цирканный ритм у них был
проявлен не строго. Пока не ясно, насколько широко среди живот-
ных и растений распространены цирканные ритмы.
Среди многолетних периодических процессов более'всего изучен
ны циклы на уровне популяций. Обычно основнре, внимание в таких
исследованиях уделялось механизмам цикличности, периоды же ус-
танавливались, как правило, приблизительно. Четырехлетняя цик-
личность установлена для некоторых популяций грызунов с помо-
щью длительных наблюдений. Так, П. Р. Грант [Grant, 1976], изу-
чая в течение И лет хомячков рода PeromyscUs в 35 км к югу от Мон-
реаля, установил для них 5-летние колебания численности. За этот
же срок там же для рыжпх полевок цикличности не установлено.
По ,32-летним данным в- Лапландском заповеднике численность ры-
жих и красно-серых полевок закономерно повышается и понижается
с периодом в 4—5 лет [Семенов-Тян-Шанскпй, 1970]. Трехлетияя
цикличность описана для популяций обыкновенной полевки из
Болгарии [Старка, Герасимов, 1977], 4-летняя — для пашенной
полевки из Финляндии [Myllymaki, 1974]. Для полевок рода Mic-
rotus в тундре описаны Л- и 6-летние ритмы изменений численности
[Batzli, 1981]. Литературный обзор'по этому вопросу дан А. А. Мак-
симовым [1984а]. i ,
Интересны работы, рассматривающие авторегуляторные меха-
низмы цикличности в популяциях. В одной из них описан демогра-
фический принцип регулирования численности. Разная функци-
ональность генераций полевок с помощью изменений уровня выжи-
ваемости обеспечивает цикличность плотности популяций длиной
в 4 года у рыжих полевок [Wiger, 1982]. Иногда цикличность попу-
ляций грызунов сопоставляется с цикличностью их кормовой базы.
4
Для четырех видов грызунов из финской Лапландии: полевки-эко-
номкп, пашенной и красно-серой полевок, а также норвежского лем-
минга найдены 5—6-летние циклы численности. Они сопоставлены
с показателями цветения основных кормовых растений. Ритмы гры-
зунов всех четырех видов оказались синхронными, а ритмы расте-
ний — кратными им — 3-летними. Предполагается связь изменений
численности с динамикой кормовой базы -[Tast, Kalela, 1971]. Су-
ществуют биологические ритмы с очень большими периодами. Дак,
у 42-летнего ритма предполагают связь с периодом обращения пла-
нет [Dewey, 1982].
Предположив определенную цикличность в популяции, иссле-
дователи обычно начинают искать ее причины и возможные эколо-
гические’ толкования. Работ по выяснению, расчету и доказательству
тех или иных многолетних циклов в популяции мы не' нашли. Сколь-
ко-нибудь строго установленные периодические изменения в попу-
ляциях неизвестны. Во всяком случае для популяций пока не опи-
саны наборы периодических составляющих их жизнедеятельности
со значениями от нескольких суток до многих лет.
Наблюдая многие годы сообщества мелких млекопитающих Ба-
рабинской низменности (Западная Сибирь), мы использовали мето-
ды, позволившие количественно оценить различные виды природной
цикличности сообществ. Это дало нам возможность рассматривать
природные циклы различной длительности (от 2 сут до 12 лет). Цик-
лические процессы присущи не тол,ько популяциям отдельных ви-
дов, но ЙГ биоценозам, так как динамические процессы в сообще-
ствах являются суммарным выражением динамики популяций от-
дельных видов. На формирование циклических процессов в сооб-
ществах влияют также факторы, привносимые вновь складывающи-
мися межпрпуляционными СВЯЗЯМИ.
Смены сообществ, или сукцессии, прежде всего связываются с
именами ботаника Ф. Клементса [Clements, 1904, 1916, 1Q36 | и зо-
олога Ч. Адамса [Adams, 1908, 1915]. В Советском Союзе большое
внимание сукцессиям уделял акад. В. Н. Сукачев — основополож-
ник учения о биогеоценозах. В настоящее время исследование сук-
цессий-признается одним из ключевых и актуальных в экологии.
Особенно большие успехи в этом плане достигнуты и фитоценологии
[Александрова, 1964; Работнов, 1972; Раменский, 1971; Куркин,
1976; Разумовский, 1981].
Сукцессиям на примере беспозвоночных животных посвящены
работы И. В. Стебаева [1963], Н. М. Черновой [1977], В. Г. Морд-
ковича [1976]. У -позвоночных они1 изучены еще очень слабо. Со
времени издания сводки по экологии Д. Н. Кашкарова [1945], ко-
торый писал, что «вопрос о сукцессии в отношении животных компо-
нентов биоценоза не разработан» (с. 266), положение, по сути.дела,
мало изменилось. Материалы по многолетним сменам в сообществах
млекопитающих разобщены и не являлись объектом специальных
исследований [Ивантер, 1975; Семенов-Тян-Шанский, 1970; и др.].
Экологические сводки [Наумов, 1963а; Макфедьен, 1965; Одум,
1975; Риклефс, 1979; Дажо, 1975; и др.] при описании сукцессий
5
основываются на геооотанических, почвенно-зоологических и гид-
робиологических материалах.
Перед экологией наземных позвоночных животных стоит задача
восполнить пробел в исследовании сукцессий, проследить их раз-
витие в сообществах через динамику пойуляций отдельных видов,
которая у этих животных достаточной полно изучена.
Помимо накопления и анализа фактического материала по сме-
нам животных сообществ необходима также оценка возможности
приложения излагаемого в литературе учения о сукцессиях к сооб-
ществам позвоночных животных. При этом важно исходить из не-
обходимости того, чтобы общие положения учения о сукцессиях бы-
ли бы приемлемыми для разных направлений экологии, т. е. удов-
летворяли бы требованиям как фитоценологов, так и зоологов раз-
ного профиля. В настоящее время развиваемые в литературе кон-
цепции сукцессий мало приемлемы для экологов, интересующихся
этими процессами в сообществах позвоночных животных. Сукцес-
сии в последних опосредованы влиянием целого ряда промежуточ-
ных факторов, среди которых особенно важную роль играют прояв-
ляющаяся 'через климат разногодичная изменчивость кормовых
(растительных и животных) ресурсов, несходство разных лет по ем-
кости П защитности мест обитания животных и др. Очевидно, имен-
но в опосредованных взаимоотношениях климата с биогеоценозами
и надо искать общие признаки сукцессий фито- и зооценозов.
В предлагаемой читателю книге на материалах по мелким на-
земным позвоночным излагается взгляд на экологические 1 сукцес-
сии как на циклический биологический процесс. Методологической
основой экологических сукцессий послужило общее представление
о цикличности биологических процессов в природе [Максимов,
1978, 1979а, 1982а,б, 1984а,б].
Авторы считают, что развиваемая ими концепция экологиче-
ских сукцессий носит полемический характер. Она не альтернатив-
на по отношению к распространенным в настоящее время представ-
лениям о сукцессиях как последовательных, идущих в одном направ-
лении и необратимых сменах одного сообщества другим и заверша-
ющихся климаксом.
Наша точка зрения основывается на представлении, что видо-
вые перестройки сообществ включают и процессы, имеющие законо-
мерно повторяющийся циклический характер, отражая соответ-
ствующую ритмику среды существования биоценозов.
Из существующих в природе типов сукцессий мы вычленяем, та-
ким образом, те, которые протекают в циклически изменчивых усло-
виях среды. Видовые перестройки соответствующих сообществ дол-
жны неизбежно отражать эту изменчивость, т. е. проходить цикли-
чески, подчиняясь колебаниям природного фона. При этом мы не
1 Экологической сукцессией сообщества мелких млекопитающих мы назы-
ваем закономерные перестройки его видовой структуры во времени под влияни-
ем циклов внешней (например, смена фаз увлажненности). и внутренней (био-
ценотические отношения) среды.
настаиваем на тезисе, что развитие всех биогеоценозов происходит
циклически (хотя и не отрицаем такой возможности). Циклы природ-
ной среды имеют разные особенности, несходные временные интер-
валы, значительная их часть еще вовсе не изучена.
Цикличность в сообществах мелких млекопитающих (грызуны,
землеройки) описывается нами на примере лесостепной зоны Запад-
ной Сибири (Барабинская низменность). Достаточно детальная ис-
ходная экологическая изученность Барабы [Сообщества..., 1978;
Структура..., 1976], включая также исследование процессов и ме-
ханизмов динамики луговых биогеоценозов [Куркин, 1976], служит
основанием для того, чтобы считать этот район хорошим полигоном
для постановки и решения наших задач.
При исследовании проблемы экологического, прогнозирования
(применительно к животным сообществам Барабинской низменно-,
сти) наше внимание привлек местный 11-летний цикл обводненности'
территории. Оказалось, что в пределах каждого 11-летнего цикла
увлажненность Барабы сменяется дважды. В ходе смен влажной и
сухой фаз происходят крупные перестройки в биогеоценозах: изме-
няется общая обводненность территории,' колеблется по фазам уро-
вень воды в болотах, речках и малых озерах, это отражается на ус- >
ловиях существования растительных и животных сообществ [Мак-
симов и др., 1979; Природные циклы..., 1982; Панадиадп, 1953;
Куркин, 1976].
Первоначально 11-летняя цикличность привлекла наше внима-
ние, поскольку с влажными фазами этого цикла была установлена
регулярная повторяемость вспышек массового размножения водя-
ной крысы, являющейся в лесостепи Западной Сибири в эти перио-
ды злостным вредителем зерновых посевов и главным источником
туляремии среди сельского населения. Одновременно установлена
связь смен фаз увлажненности с 11-летним циклом солнечной актив-
ности. Именно на этом разработаны экологические основы долго-
срочного прогнозирования вспышек массового размножения водя-
ной крысы [Максимов, 1959, 1966, 1971, 1977 и др.]. Одиннадцати-
летняя цикличность в сменах фаз увлажненности Барабы — это
природный цикл, влияние которого на динамику биогеоценозов и
было исследовано авторами. Выяснялось, может ли 11-летний цикл
служить основой для прогнозирования других биологических про-
цессов в животных сообществах Барабы, есть ли между ними при-'
чинно-следственная связь. Речь идет, следовательно, о поисках ме-
тодов долгосрочного прогнозирования динамики биогеоценозов, ос-
нованных на наличии характерной природной цикличности.
Выявленная 11-летняя цикличность послужила основой для ис-
следования разногодичных смен функционирования животного на-
селения Барабы.
Показательными оценками биоценоза в разные фазы обводнен-
ности могут служить: перестройка видовой структуры сообщества,
смена доминантов, динамика численности. В этих исследованиях
представляет интерес анализ варьирования как сообщества в целом,
так и популяций фоновых видов при изменении обводненности. Нас
7
интересовала реконструкция биоценоза во влажный и сухой пери-
оды. Важно было найти индикаторы перестройки сообществ, т. е.
структуры, наиболее четко реагирующие на' степень заболоченности
. территории, объяснить биологический смысл наличия в биоценозах
подобных структур.
Среди щричин, вызывающих перестройку в животном населении
।, Барабы, имеются не только климатические, но и' биоценотические,
которые в совокупности приводят к модификации сообщества по ти-
пу ^влажного или сухого варианта. । i ।
[Из биоценотиФеских факторовг наше внимание привлекло, в
частности, влияние вспышек массового размножения водяной кры-
сы (Arvicola terrestris Б.) на видовую структуру животного населения.
Водяная крыса,- будучи околоводным зверьком, очень чувствитель-
на к степени обводненности! территории. Она может рассматривать-
ся как вид с наиболее четкой экологической зависимостью от смен
.фаз увлажненности.1 В периоды? вспышек этот грызун не может не
влиять на популяции видов, |свя:занных с ним биоценотическими от-
ношениями. Было интересно выяснить, сказываются ли вспышки
массовых размножений водяной крысы на видовом разнообразии на-
селения грызунов и землероек, динамика каких видов сообщества за-
висит от состояния Очагов водяной крысы.
Для выполнения поставленных задач был необходим анализ ма-
териалов по циклам жизнедеятельности отдельных видов и их по-
пуляций и выяснению связи этой цикличности с ритмами природной
среды и биоценотическими отношениями.
. . Существование характерных? природных циклов, как показы-
вает опыт работы в Западной Сибири,— типичное природное явле-
ние. Нами исследованы сезонные и многолетние циклы, а также
экологические сукцебсии в разных биогеоценозах. Сделана попытка
доказать наличие цикличности в развитии биоценозов и обосновать
подчиненность экологических сукцессий в сообществах колебаниям
природного фона. Для этого было важно: 1) представить объектив-
ные доказательства цикличности природной среды исследуемого
биогеоценоза, которая определяет регулярную изменчивость усло-
вий существования животных сообществ; 2) выяснить, существуют
лц у популяций животных в исследуемом регионе видовые и попу-
ляционные циклы динамики, и охарактеризовать их связь с 11-лет-
ней цикличностью'природного фона, биоценотическими и другими
факторами; 3) рассмотреть, как влияют смены фаз увлажненности
в 11-летних циклах на изменение видовой структуры животного на-
селения; 4) оценить роль вспышек массового размножения домини-
рующих видов в изменении структуры сообществ животных.
Цикличность и сукцессии, в экологических системах рассматри-
ваются как взаимосвязанные процессы. Повторяемость серий сук-
цессии обусловливает многолетнюю цикличность в биоценозе, се-
зонные и другие краткосрочные циклы в свою очередь включены в
каждую серию сукцессии. Таким образом,, циклическим процессам
уделено столько же внимания, сколько иодному из них—сукцессии.
Исследование влияния природных циклов на биогеоценозы вы-
I
I
8
ходит далеко за пределы изучения биологических процессов. Послед-
ствия смен климатических фаз имеют важное значение для сельско-
хозяйственного производства. От них зависит качество урожая от-
дельных культур, продуктивность лугов, обилие сорняков, успех
мелиоративных , мероприятий, нашествие вредителей, развитие бо-
лезней сельскохозяйственных животных и др. Как следствие этогоJ
экономика хозяйств не может быть'сходной в разные фазы измен-
чивости природной среды. ; : i ’ '
Смена фаз увлажненности в закономерно повторяющихся.цик-ж
лах влияет и на медицинское .состояние местного населения. Так,
во влажные периоды в биоценозах1 развиваются процессы, приводя-
щие к появлению интенсивных эпизоотий туляремии и, как след-
ствие этого, к -осложнению эпидемической обстановки [Максимов,,
1979а]. В этом проявляется лесная, опосредованная через климат,
зависимость биоценозов от циклов солнечной активности и других
циклических процессов.1 ' . ! 1
। •i’ | -И* . / *
, '* . 1
Для исследования циклов в биогеоценозах Западной Сибири ав-
торы использовали многолетние сборы материалов, которые прово-
дились сотрудниками лаборатории экологического прогнозирова-
ния Биологического института СО АН СССР. В этих сборах и их
первичной обработке помимо авторов активное участие принимали
канд. биол. наук А. С. Николаев, В. Е. Сергеев, И. Н. Глотов,
мл. науч. сотр. В. В. Панов, В. В. Салтыков, О. В. Григорьев. При
подготовке рукописи в печать большую помощь оказали Е.. П. Мерз-
лякова, А. А. Переберин, Н. Л. Чубыкина, Н. А. Шевченко. Ав-
торами учтены ценные замечания д-ра биол. наук, проф. И. В. Сте-
баева, кандидатов биол. наук В. Г. Мордковича и Ю. Г. Швецова.
Определение насекомоядных проведено В. Е. Сергеевым,
В. В. Пановым и Л. Н. Ердаковым, консультации при определении
этой группы животных получены от Б. С. Юдина и В. Е. Сергеева.
Грызуны определены участниками, сборов, помощь и консультации
получены от Л. И. Галкиной и Н. Ф. Шушпановой. Всем указанным
лицам авторы глубоко благодарны.
Работа . над текстом между авторами распределялась следующим
образом. Главы V и VII написаны А. А. Максимовым, II, III, IV —
Л. II. Ердаковым, остальные разделы написаны совместно. ।
Г л а в a I ' 1 , ! .
МЕТОДЫ АНАЛИЗА ЦИКЛИЧНОСТИ^
В ПОПУЛЯЦИЯХ МЕЛКИХ МЛЕКОПИТАЮЩИХ
И ОБЪЕМ ИСПОЛЬЗОВАННОГО МАТЕРИАЛА
ОЦЕНКА ПЕРИОДИЧНОСТИ
С ПОМОЩЬЮ СПЕКТРАЛЬНОГО АНАЛИЗА
Данные наблюдений обычно представляются в виде измерений через
определенные интервалы времени, подсчета количества каких-либо
дискретных элементов в единицу времени, it. е. временные ряды.
На основании их можно делать заключения о той или иной периодич-
ности наблюдаемого процесса или явления. В настоящее время су-
ществует единственный метод, имеющий хорошую теоретическую
базу, с помощью которого можно достаточно обоснованно рассчиты-
вать величину периода наблюдаемого процесса. Речь идет об ана-
лизе временных рядов. Он широко исполь&уется в технике, физиче-
ских науках и экономике, а в последние годы все чаще находит при-
менение в биологии и медицине. Один из важных видов анализа вре-
менных рядов — спектральный анализ, разделяющий их на раз-
личные частотнее составляющие.
' В основе спектрального анализа лежит построение по экспери-
ментальным данным оценок функции спектральной плотности, даю-
щей распределение дисперсии изучаемого временного процесса по
частоте. Появление на какой-нибудь частоте острого, хорошо выра-
женного пика обычно трактуется как присутствие в процессе ритми-
ческой составляющей с данным периодом.
Поскольку различные участки временного ряда обнаруживают
сходство только в их осредненных свойствах, ряды описывают с по-
мощью вероятностных законов или моделей. Таким образом, пред-
полагается, что возможные значения временного ряда в данный мо-
мент времени описываются с помощью случайной величины
и связанного с нет! распределения вероятностей. Тогда наблюдаемое
значение временного ряда в момент t рассматривается как одно
из семейства значений, которые могла бы принять случайная вели-
чина X(f) в момент t.
Важнейшее предположение о временных рядах заключается в
том, что соответствующий случайный процесс является стационар-
ным и может1 быть адекватно описан с помощью младших моментов
его распределения вероятностей. Младшие моменты включают в се-
бя среднее значение, дисперсию и преобразование Фурье ковариа-
ционной функции — спектр мощности.
10
Качественно стационарный ряд — это такой ряд, который нахо-
дится в статистическом равновесии в том смысле, что он не содержит
никаких трендов, тогда как нестационарный ряд таков, что его свой-
ства изменяются во времени. Ряды, встречающиеся на практике,
обычно трех видов: 1) проявляющие свойство стационарности в те-
чение долгого времени; 2) достаточно стационарные в течение корот-
ких периодов; 3) явно нестационарные (их видимые свойства непре-
рывно изменяются). ’ ।
Г. Дженкинс и Д. Ватте 1)1971 ] замечают, что, хотя для описания
случайного процесса с1 помощью его спектра необходимо предпола-
гать стационарность, на . практике предположение стационарности
не представляет серьезной проблемй. Это происходит, поскольку
спектр отделяет вклады во временной ряд, которые можно приписы-
вать разным частотным полосам. Если нестационарный ряд харак-
теризуется присутствием большой мощности на низких частотах,
а представляющая интерес информация — на высоких, то можно
отфильтровать нестационарные низкочастотные компоненты и ис-
пользовать' оставшийся ряд. С практической' точки зрения неверо-
ятно, чтобы какой-нибудь ряд оставался стационарным бесконечно
долго. Важен вопрос, запись какой длины можно взять для анализа,
не нарушая при этом предположения о стационарности. . )
Этими основными предварительными соображениями । примене-
ния спектрального анализа мы руководствовались при обработке
наших данных. Временные ряды различной длины получаются при
учетах попадаемости мелких млекопитающих в разные орудия лова.
Мы анализируем попадаемость в канавки и отдаем себе отчет, что
экологическая характеристика «попадаемость в канавки» есть функ-
ция двух переменных: численности и подвижности. В настоящем ис-
следовании не стоит вопрос о разделении этих переменных, и мы счи-
таем попадаемость интегральным показателем изменения числа жи-
вотных, т. е. считаем, что выбранная характеристика дает представ-
ление об изменении численности. Наши задачи сводятся к установ-
лению длин возможных периодических составляющих процесса
изменения численности. Предполагается, что спектральный анализ
дает возможность установить периодичности от сезонных и месяч-
ных до многолетних (предельная длина «сшитого»1 ряда 15 лет).
Вопрос о стационарности исучаемого периодического процесса
мы решаем, разделяя ряды на участки, в которых сохраняются ус-
тойчивые колебания попадаемости. Основной участок, подвергае-
мый анализу,— вегетационный период в'биоценозе. В это время mho-i
гократно происходят рождение и расселение молодых животных,
часть которых совершает характерные, строго привязанные ко вре-
мени миграции по биотопам. Временной ряд, взятый за этот период
(100 членов ряда), рассматривается для каждого вида и, кроме того,
анализируется «сшитый» ряд из вегетационных периодов многих се-
1 «Сшитым» называют спектр, посчитанный из временного ряда, имевшего
пропуски данных. В нашем случае отсутствуют материалы за 1975 г., и вре-
менной ряд составлялся без них. В этом месте массива при счете ставится спе-
циальная карта п программа учитывает разрыв —«сшивает».
И
‘ зонов. Правомочность использования «сшитого» спектра обсужда-
лась в литературе [Мерсер, 1964].
Поскольку спектральный анализ как способ обработки биологи-
ческих данных применяется пока еще нечасто, мы подробно опишем
использованные нами программы. Основной упор при этом сделаем
на способах применения наших программ и возможности получения
новой информаций! с их помощью. Описание самих программ и ал-
горитмов, на основе| которых они составлены,' не входило в нашу
задачу. Тем более, что алгоритмы и сам метод' спектрального ана-
лиза традиционны, а программы составлены по 'книге Г. Дженкинса
и Д. Ваттса [1971], которая является [подробным руководством по
способам применения спектрального анализа.
Данные ежедневных уловов в канавки представляют собой вре-
менные ряды, обработка которых производилась на ЭВМ по програм-
мам спектрального анализа для выявления скрытых (колебаний. Со-
ставленные программы адаптированы к нуждам хронобиологическо-
го изучения животных [Ердаков и др., 1981; Ердаков, 1984]. Хотя
сама процедура счета сложна и обычно осуществляется' на вычисли-
тельной машине, параметры счета задает биолог. Г. Дженкинс и
Д. Ватте [1971] советуют перед теме как начать считать, эмпириче-
ски определить параметры счета. К ним относятся: шаг суммиро-
вания данных, длина автокорреляционной функции, форма и шири-
на корреляционного окна. Обычно, ищут оптимальное соотношение
шага, длины автокорреляционной функции и ширины окна. Причем
длина автокорреляционной функции подбирается в пределах длины
имеющихся наблюдений и зависит от шага (интервала суммирова-
ния данных). Так,! если у нас основа счета — ежедневные уловы,
то нижний (предел шага — 1 день. При таком шаге в наблюдениях
в течение вегетационного периода автокорреляционная функция не
должна превышать! 90, т. е. количества дней наблюдений. Если мы
зададим такой шаг и такую длину автокорреляционной функции,
то на спектре получим распределение гармонических составляющих
по мощности и самая высокочастотная из них будет длиной в 2 сут.
Форма корреляционного окна может быть избрана любая в со-
ответствии с обычно применяемыми при спектральном анализе
I [Дженкинс, Ватте, 1971]. Мы применяем окно Бартлета. Сравнение
спектров, полученных на наших материалах при разных формах
окон, не дало существенных различий. >
| Программы для счета данных были составлены таким образом,
чтобы биолог мог менять параметры, добиваясь наибольшей точно-
сти расчетов, учитывая при этом характер материала и удобную фор-
му выдачи; результатов машиной. Так, известно, что спектральная
функция получается путем косинус-преобразования автокорреля-
ционной функции исходного ряда, предварительно помноженной на
сглаженную функцию (корреляционное окно). При использовании
большинства известных корреляционных окон на спектре появля-
ются ложные пики. Поэтому перед счетом нужно выбрать пригодную
ширину корреляционного окна. ।
Наша программа позволяет провести предварительную процеду-
ру «стягивания окна», что дает в руки биолога сразу несколько гра-
12 I
фпческих изображений спектра, среди которых он может выбрать
одно и остановиться на избранной ширине окна. Соображения для
выбора определяются компромиссом между уровнями детализации
и шума. Пики правильной экспоненциальной формы с максимально
узким основанием — это некоторая гарантия точности оценки пе-
риодичностей, появившихся на спектре. В то же время со стягивани-
ем окна в какой-то момент на спектре появляется шум в виде лишних
(ложных) пиков,! кцторые могут оказаться многочисленными, мощ-
ными и исказить всю картину.
Процедура стягивания окна, по рекомендации Г. Дженкинса и
Д. Ваттса [1971], проводилась следующим образом. Рассчитывался
спектр одного и того же процесса при 1/8, 1/4, 1/2 й 1 длины авто-
корреляционной функции. Полученные спектры уравнивали <между
собой и отбирали наиболее устойчивый. Это был спектр, на котором
количб|Ство и расположение пиков не изменились или почти не изме-
нились в сравнении с предыдущим. .После всех предварительных
процедур нахождения периметров счета мы остановились на следую-
щих: длина автокорреляционной функции 72 для годичных и 360
для многолетних рядов, шаг суммирования данных — 1 сут для го-
дичных и 5—10 — для многолетних данных, окно Бартлета с шири-
ной, равной длине автокорреляционной функции/ i
Кроме того, для отдельных разделов обработка проведена тра-
диционными методами. Морфологические признаки сравнивали с
помощью дискриминантного анализа [Кендалл, Стюарт, 1976] и
статистических оценок [Плохинский, 1961].
ОБЪЕМ И РАСПРЕДЕЛЕНИЕ ИСПОЛЬЗОВАННОГО МАТЕРИАЛА
Для удобства описание материала мы приводим по отдельным ста-
ционарам. Время сбора на всех стационарах было унифицировано
и ограничено только вегетационным1 периодом — с июня по сен-
тябрь каждого года. Проанализированы данные по 8 видам насеко-
моядных и 14 видам,грызунов. Материалы по насекомоядным вклю-
чают данные о бурозубках: обыкновенной (Sorex araneus L.), круп-
нозубой (5. daphaenodon Thomas), арктической ’(5. arcticus Kerr),
равнозубой (5. isodon Turov), средней (5. caecutiens Lax.), малой
(5. minutus L.), крошечной; (S, minutissimus Zimm.); водяной (обык-
новенной) куторе (Neomys fodiens Pennant). Материалы по мышевид-
ным грызунам включают данные о водяной крысе (Arvicola terrestris
L.), полевкам: йолевке-экономке (Microtus! oeconomus Pall.), узко-
черепной (M. gregalis Pall.), темной, или пашенной (М. agrestis L,),
обыкновенной (/If. arvalis Pall.), красной (Clethrionomys rutilus Pall.),
красно-серой (Cl. rufocanus Sunderv.), рыжей (Cl. glareolus Schreb.);
обыкновенному хомяку (Cricetus cricetus 1л), Нолевой мыши; (Apode-
mus agrariu's Pall.), мыши-малютке (Micromys minutus Pall.), лесной,
или северной, мышовке (Sicista betulina Pall.), степной, или южной,
мышовке ’ (5. subtilis Pall.), джунгарскому хомячку (Phodopus sun-
gorus Pall.)2. 1 '
2 Латинские названия животных приведены в соответствии с Каталогом
млекопитающих СССР [1981].
13
Численность* мелких млекопитающих по годам,
1950 | 1965 1966 1967 1968 1969 1970
Вид . Отработано
10200 1380 5320 7080 5460 6165 6330
1 ! 2 3 4 6 7 8
Мышовки
лесная . 167 21 „ i 31 20 44 86 71
1,63 1,52 0,58 0,28 0,80 1,39 1,12
степная 0 3 6 3 4 12
1 : 0,21 0,11 0,04 0 0,06 0,18
Полевая мышь 167 216 26 108 22 68 68
i 1,63, 15,65 0,48 1,52 0,40 1,0.7 1,07
Мышь-малютка 64 30 4 45 .8 22 45
1 0,62 2,17 0,07 0,63 0,14 0,35 0,71
Обыкновенный 14 3 20 65 25 7 2
хомяк . 0,13 0,21 0,37. 0,91 0,45 0,11 0,03
Джунгарский , 2 . 14 5 2
ХОМЯЧОК 0 0 0 0,02 0,25 0,08 0,03
Полевки
красная 53 36 10 63 52 53 41
0,51 2,60; 0,18 О’, 88 0,95 0,85 0,64
рыжая 66 о 0 ' 25 '25 32 17
' 0,64 0,35 0,45 0,51 0,25
' красно-серая 19 1 i 21 11 31 3
0 1,37 0,01 0,29 0,20 ' 0,50 0,04
Водяная крыса 1048 . 25, 15 46 14 17 37
Полевки 10,27 1,81 0,28 0,64 0,25 0,27 0,58
экономка 64 8 44 213 72 44 182
0,62 0,57 0,82 3,00 1,31 0,71 2,87.
i темная 122 23 10 93 33 •20 57
1,19 1,66 0,18 1,33 0,60 0,32 0,90
узкочерепная 43 6 15 117 116 95 105
0,42 0,43 . 0,28' 1,65 2,12 1,54 1,65
Кутора i 1139 11,16 .° , 0 ,! 5 0 0 1 0,01 0
Бурозубки [ 1
обыкновенная ; 520 163 145 519
5,09 11,81 2,35 8,19
крупнозубая 71 \ ' 22 6 5
, 0,75 1,59 0,09 0,07
арктическая о 0 1_ 8 35
0,12 0,55
средняя 247 0 • 7 37
2,42 0,11 0,58
14
данные отловов на Ровенском стационаре Таблица 1
1971 1972 1973 1974 .1978 1979 1980 J 1981, | 1982
цйлиндро-суток
4850 3995 2340 4490 . 4175 3915 4110 4717 4940
9 Ю 11 1 12 . ) .13^ 14 ! , 15 16 17
19 0,39 0 137 2,82 104 : 2,14 16 0,32 •о . 48 0,98 18 0,37 3 0,26 203 4,18 41 0,84 127 2,53 143 2,94 7 0,14 311 6,41 ~ 15 0,30 17 0,35 31 0,63 21 0,52 0 22 , 0,55 6 0,15 . 24 0,60 0 9 0,22 1 0,02 2 0,05 818 20,47 6 0,15 0 11 0,27 0 49 1,22 29 0,72 2 0,05 3 0,07 46 1,96 0 234 10,0 43 1,83 2 1 0,08 ' °! 46 . ! т> 5 0,21 10 0,42 180 7,69 9 0,38 4 0,17 0,42 : 81 3,46 410 17,52 И 0,47 13 0,55 14 0,59 32 0,71 0 309 6,88 331 52 1,24 ' 0 168 4,02 . 50 1,-49 25 0,59 0 72 1,72 ' 54 1,29 0 78 1,86 ; 130 3,11 3 0,07 57 1,36 9 0,21 279 6,68 10 0,23 17 0,40 17 0,40 66 1,68 0 • 282' ' 7,20 73 1,86 4 0,10 3 ’ ; 0,07 88 2,24 53 1,35 0 84 2,14 33 0,84 14 . 0,35 12 0,30 20 0,51 366' 9,34 12 0,30 9 0,22 6 0,15 t 68 1,65 0 256 6,22 48 1,16 13 0,31 0 70 1,70 110 2,67 0 362 . 8,80 44 1,07 22 0,53 12 0,29 . 4 0,10 335 ' 8,15 23 0,55 12 0,29 8 0,19 12 0,25 i ‘ 0 7.4 ‘‘ 1,56 ’38 0,80 1 0,02 1 0"Ti2 23 0,48 56 1,18 0 166 3,52 29 0,61 6 0,12 10 0,21 31 0,65 215 4,55 18 0,38 2 0,04 16 0,33 38 0,76 . i 0 292 5,91 26 0,53 0 2 ’ 0,04 18 1 0>л'' lj й 3 W о,об 33 0,67 3 0,06 3 0,06 0 182 3,68 18 0,36 5 0,1 31 0,63
7,37 0 0 0 . 168 3,74 85 1,89 7 0,15 40 0,89 86 1,91 23 0,51 96 2,13 > 56 1,24 1692 37,68 342 7,61 16 0,35 22 0,48
15
1 2 •ч 14 6 7 8
малая (1 С крошечная . f> 87 0,85, ' >°! (| . 75 , 5,43, 0,07 ; i — — 22 0,35 |! 9 \ 0,14 78 1,23 5 0,07
* В табл.! li—4 в числителе—’число пойманных ;зверьков, в’ знаменателе — их отно
. ' i 1 О ; 1 ' I !
1 Ровенский стационар расположен в с.'Ровенское Каргатского
района Новосибирской области. (Подробное 'описание его приводится
в литературе [Сообщества...,- 1978]. Регулярные исследования на
атом стационаре ведутся с 1965 г. За предыдущие годы "использова-
ны также данные сборов 1950 г. [Максимов, 1959а]. Наблюдения за
изменениями в сообществе мелких млекопитающих осуществлялись
с помощью 10ловчих ! канавок: с цилиндрами.
За весь период работы на Ровенском стационаре отработано
74 5201 цилиндро-суток (табл. 1). Зазто время: поймано 20 276 мел-
ких млекопитающих, । из них грызунов — И 481 зкз., землероек —
riJ:73Q5 экз. Сбор материала осуществляли И. Н. Глотов (1965—1971),.
-УН. Ердаков (1968-1972, 1974-1982), А. С. Николаев (1967—
С $72), В. Е. Сергеев (1974)^ ! . । ii
, . / Потюкановский стационар расположен на' юге Северного района
у в верхнем течении р. Кама. Исследования проводились в 1976—
1981 гг. с использованием 13’ловчих канавок. Кроме них для отло-
ва- использовали плуговую борозду, проходившую по краю луга и
старой залежи. Цилиндры располагались1 через 10 м, длина борозды
в разные годы варьировала от 200 .до 380 м.
За 7 лет исследований отработайо ,31 548 4илинДро-суток и от-
ловлено - 16 246 экз. мелких млекопитающих (10 115 грызунов и
6131 насекомоядных). Кроме того, другими способами (капканы,
живоловушки и т. д.) [Отловлен еще 1191 |экз. мелких млекопитаю-
щих- Таким образом, | количество исследованных животных соста-
вило 17 437 зкз. । ।
Видовой состав исследованных животных представлен в табл. 2.
Сбор материалов проведен А. С. Николаевым, В. В. Пановым.
Усть-Ургульский стационар, расположен в Верх-Красноярском
сельсовете Северного района. ; Исследования проводились с 1978
по 1982 г. включительно. Отлов зверьков велся: на пяти 100-метро-
вых плуговых ловчих бороздах с 10 цилиндрами каждая.
За 5 лет сборов отловлено 9666 зверьков, из них грызунов —
6964 и землероек — 2702 (на 19 060 щил.-сут). О видовом составе
отловленйых особей дает представление табл. 3. Материал собран
А. А. Максимовым и В. В. Салтыковым: (1978—1982 гг.), В. В. Па-
новым (1982 г.).
Коноваловский и Орловский стационары расположены на терри-
тории Пихтовского сельсовета Колыванского района, д. Орловка
16
Окончание табл, t
9 10 11 12 13 14 1-5 16 17
73 13 23 49 .( 222 ; 63 . 137 38 : 158
1,50 0,32 1 0,98 1,09 ; 5,31 1,60 - 3,33 0,80( 3,2
5 1 1 !° 1 S А : : 31. 7 ’ ! 9 И
0,10 0 0,11 ! 0,07 ! 0,12 • ' 1 0,16 0,19 0,22
-!------------ i ;i 'Л. . ' ' ' L
сительная численность на ЛОО цилРсут. Прочерк означает отсутствие данных.
1 ; ' < ।
находится в 15 км от д. Коноваловка. Исследования проведены в ок-
рестностях д. Коноваловка в 1974—1978 гг. и в окрестностях д, Ор-
ловка,— в)1979—1980 гг. На стационаре в д. Коноваловка1исполь-
зованы данные отлова в одну борозду. Она находилась на право-
бережье р. Баксы, состояла! из 28 цилиндров и проходила по краю
колка! и луга, разнотравнойу лугу и краю болота. За болотом —
поле, ежегодно засеваемое яровой пшеницей. ।
На Орловском стационаре отлов мелких! млекопитающие прове-
ден на 10 ловчих канавках (табл. 4). Материал собирали А. А. Мак-
симов (1974 г.), Н. П. Сасов (1975 г.), И. Н. Глотов (1977—1980 гг.),
Л. Н. Ердаков (1976 г.), В. Е. Сергеев (1976, 1978-1980 гг.). ,
Черновский стационар расположен в Кыштовском районе в 30 км
севернее районного центра Кыштовка. Исследования проведены в
1973—1974 гг. в окрестностях с Черновка,. в 1973 г. сборы были
в основном рекогносцировочными, а в 1974 г. осуществлялись с
9 ловчих канавок. ,1
На 2460 цил.-сут поймано 1482 зверька, из них1— 823 грызуйа
и 659 насекомоядных. Грызуны представлены следующими видами
(экз.): водяная крыса 142, полевки — экономка 28,| узкочерепная
26, обыкновенная 10, темная 20, красная 196, красно-серая 5, ры-
жая 47, обыкновенный хомяк 1,'полевая мышь 254, мышь-ма-
лютка 35, лесная мышовка 59; из насекомоядных: кутора —
21, бурозубки: обыкновенная — 523, крупнозубая—5, арктиче-1
ская — 43, средняя — 19, малая — 47, крошечная —!.1 зкз. Мате-
риал собран А. С. (Николаевым. , - I -
Крещенский стационар расположен на севере Убинског.о района.
Мышевидные грызуны и землеройки изучались в 1977 г. Использо-
вали ловчие борозды с цилиндрами. На 2495 цил.-сут отловлено
1623 экз., из них 367 грызунов и 1256 землероек. Видовой состав
был следующим (экз.): водяная крыса 38, полевки — экономка 89,
узкочерепная 55, темная 1, обыкновенная 1, красная 22, красно-се-
рая 2, рыжая 26; хомяк 9; мыши —» полевав 7, мышь-малютка 39,
азиатская лесная 1; лесная мышовка 77, кутора 8; । бурозубки"—
обыкновенная 1056, крупнозубая 43, арктическая 11, средняя 21,
малая 103, крошечная 11, равнозубая 3. Сборы проводили А. А. Мак-
симов и В. В. Салтыков.
Исходные материалы сборов мелких млекопитающих по Ровен-
скому (1950—1974 гг.), Коноваловскому (1974—1976 гг.), Потю-
2 А. А. МАКСИМОВ, Л. II. Ердаков 1 1.7
.-z Таблица 2 Численность мышевидных грызунов и землероек по годам, данные отловов на Потюкановском стационаре
Вид 1976 1977 1978 1979 1980 1981 1982 ,,
Отработано цилиндро-суток
7118 6285 . 6850 : 4668 3577 2950 100
। Водяная крыса | Полевки экономка ; । ; ' 1. ! узкояеррпная темная ! и обыкновенная 1 | красная 'красно-серая 1 рыжая Обыкновенный хомяк Полевая мышь Мышь-малютка Лесная мышовка Кутора 1 Бурозубки ’ (: обыкновенная: крупнозубая 1 арктическая^ 1. , средняя малая [ крошечная равнозубая 137 1,92 512 7,19 155 2,18 60 0,84 ' 22 0,13 51 0,72 6 0,08. 32 0,45 0 43 0,60 18 . 0,251 159 2,23 17 0,24 1032 14,50 360 5,05 63 0,88 3 0,04 66 0,93 8 0,11 2 0,03 181, 2,88 658' 10,47 373 '5,93 49 0,78 1 0,02 36 0,57 1 4 0,06 10 0,16 0 40 0,64 32 0,51 50 0,80 5 0,07 858 13,65 253 4,02 52 0,82 16 0,25 146 2,32 19 0,30 4 0,06 66 0,96 207 3,02, 97 : 1,42 24 0,35 2 0,03 41 0,60 1 0,01 25 0,36 0 , 36 0,53 3 0,04 138 2,01 7 0,10 208 3,04 9 0,13 116 1,69 19. 0,28 204 2,98 5 0,71 2 0,03 55 .1,17 . :9 0,19 4 0,08 8 0,17 0 9 0,19 16 0,34 1 0,02 . 0 157 3,36 . 15 0,32 109 2,34 16 0,34 279 5,98 4 0,08 8 0,17 24 0,51 64 1,37 0 0 3354 93,76 77 2,15 12 0,33 54 ' 1,51 0 10- 0,28 - 0 25 • 0,70 4 0,11 103 2,88 124 3,47 69 1,93 154 4,30 214 5,98 1 0,02 6 0.16 31 0,86 271 7,57 1 0,02 0 976 33,08 9 0,30 2 0,06 1 0,03 0 0 0 0 0 37 1,25 9 0,30 НО 3,72 326 11,05 97 3,28 3 0,10 6 0,20 28 0,95 159 5,39 О 0,10 0 0 0 : ' 1 0 1- ‘ 0 1 0 ' 1 'О' 86 2 5 0 0 14 0 0 0 1 0 0
18
Таблица 3
Численность мышевидных грызунов и землероек по годам, данные
отловов на Усть-Ургульском стационаре
1978 , 1979 1980 1981 1982
Вид Отработано цилиндро-суток
3720 3120 4150 3860 4610
Водяйая крыса 0 23 260 3843 (I 356 ‘
Полевки ' 0,74 6,27 99,56 7,72 ,
экономка 15 ~ : 32 75' 58 8
— .
' 0,40 1,03 1,81 1,5 0,17
узкочерепная 51 59 . 27 15 4
1,37 1,89 0,65 0,39 w
темная 3 12 74 19 4
0,08 0,38 1,78 0,49 ~0fi9
обыкновенная 2 1 '
0 ’0^06 0 о; оз1 0
красная 32 60 46 it 9
— 1 — — —
0,86 • 1,92 1,11 0,28 0,19
красно-серая 2 73
0,05 2,34 и 0
рыжая 48 67 77- 37 24
1,29 2,15 1,86 0,96 0 52
Хомяк обыкновенный 0 0 0 7 2
0,18 0,04
Полевая мышь 185 442 166 74 54
4,97 14;17 4,00 1,92 1,17
Мышь-малютка 8 42 38 19 4
-
0,22 1,35 0,92 0,49 0 02
Лесная мышовка 152 102 122 60 57
4,09 3,27 2,94 Т55 1 24 -
Кутора г 2 17 ; 289 166 1
0,05 0,54 6,96 4,30 ' 0 02
Бурозубки , 1
обыкновенная 55 311 265 672
1,48 9,97 6,39 17,40
крупнозубая 3 16
♦ 1 0,08 ! 1, 0,41 । !
арктическая 14 0,38 73 2,34 1 0,02 37 0,96 —
средняя 12 152 21 36
0,32 4,87 : 0,51 0,93 ।
малая 94 84 59 33 1
2,52 2,69 1,42 0,85
крошечная 4 5 2
0,11 0,16 0,05 0
2 2
равнозубая 0 0,06 0,05 0 » i*
2*
18
Таблица 4
Численность мышевидных грызунов и землероек по годам, данные отловов, на
Коноваловском и Орловском стационарах
Вид - Коноваловка Орловка
197,4 ] 1975 | 1976 | 1977 1978 1979 1980
> Отработано цилиндро-суток 1
1 ! 1539 | 1080 .| 2016 | 2630 3540 3530 4175
• : 1 2 Г 3 4 • ' . • 5i , 6 7 8
Лесная мышонка i 30 1 1 ! ! 22 31 43 12 4 ;->
i 1,96 1 0,09 1,09 : 1,18 1,21 0,34 0,09
Мыши ' 1 i !
полевая i 92 18. 86 128 79 *93 14
6,01 0,74 4,27 4,87 2,23 2,63 0,33
лесная 1 1 0,
। 0 0,09 0 0 i 0 0
Мышь-малютка 31 2 i 9 28 12 22 1
^Домовая мышь 2,03 0,18 0,45 ' 1,06 0,34 0,62 0,02 1
1 ! 0, ii<'1 -
0 0,09 0 0 0 0
Хомяк обыкновенный 5 5
0 ! 0 . 0 • 1 0 0 0,14 0,12 »
Полевки
рыжая 22 0 8 6 ‘ 18 1 0
1,44 0,40 0,23 0,51 0,03
красная । 19 13 41 4 2 5 4
1,24 1,20 ,2,03 0,15 0,06 0,14 0,09
красно-серая 6 3 3 3 .3 5 7
0,39 0,28 0,15 0,11 6,08 0,14 0,17
il Водяная крыса , : 1 38 5 30 35 8 85
2,48 0 0,25 1,14 0,99 0,23 2,04
Полевки ’ ' - ' i 1
узкочерепная । ( 57г 14 76 44 22 1 0
3,73 1,30 3,77 1,67 0,62 0,03
обыкновенная ' ; ; 1 24 429 20 .1 34 19 23 •
1 j 1 1 I . ' • 0,07, 2,22 6,40 - 0,76 0,96 0,54 0,55
темная !ы. !i . 21 ' ,17 104 ' 12 6 23 37 i
1 , • । ; 1,37 1,57 ’5,“16 , 0,46 0,17 0,65 0,89
экономка п 62 26 . 108 , 32 1 15 2 4 л.
! 1 |! ! •4,05 2,411 5,36 ! Г,22 1 0,42 0,06 0,09 ''
Крот Сибирский | 0 , . ,1 ; ' 0,05 i ,5 0,19 5 ! 0,14 0 0
Бурозубки ।
обыкновенная ' 143 114 252 371 341 192 46
9,35 10,56 12,50 14,11 9,63 5,44 1,16
крупнозубая 1131 2 i ' 15 9 22 1 5
8,56 0,19 0,74 0,34 0,62 0,03 0,12
20
Окончание табл.
4
1 2 3 4 5 6 7 8
арктическая s, 3 j 4 39 12 j |29 7 2
fr ; __. 0,20 1 0,37 i 1,93 ; 0,46 i i 0,82 0,20 0,05
средняя j , и i|... 2 3 ,J 2 , < .JL 'i 20 26 2
_ j 0,13 0,28 ., 0,10 0,19 0,56 0,74 . 0,05
малая i 12 ' 12 '61 4 41.1 50 i: 18
0,78 . 1,11 0,30 0,15 1,16 1,42 0,43
крошечная i ; 2 ' • 0,13 j 0 2 . 0,10 ° И 1 o •2 0,06 o -
равнозубая । 6 ’ ; 3 ! 4 61 i3 4
0,15 ! 0,15 0,17 ; 0,08 0,09
Б'утора 9- 6 1 6 - 2 ' 7 11
0,59 0,56 0,05 - 0,23 • 0,06 0,20 0,26
!
каковскому (1976 г.), Черновскому (1974 г.) .стационарам уже Опуб-
ликованы [Сообщества..., 1978]. В этой же сводке дается более пол-
ное описание! стационаров и их размещение Па территории Барабы.
Нами проведены предварительные расчеты некоторых периоди-
ческих составляющих климата по данным Каргатской метеостанции
за 5 лет (1968—1973 гг.). Выяснилось, что для элементов климата
существуют устойчивые «короткие» ритмы: атмосферной (относи-
тельной) влажности — 5- и 17-суточные; осадков — 9- п 12-суточ-
ные (в среднем 10-суточные) и температуры воздуха — 7- и 20-су-
точные. Эти данные мы использовали для сравнения с циклами по-
падаемости животных в ловушки, считая, что короткие циклы у жи-
вотных соответствуют таким ритмам элёментов климата.
Все эти материалы положены в основу анализа циклических про-
цессов в популяциях и сообществах Барабы. Привлечение дополни-
тельных данных для описания биоценотических процессов оговари-
вается в соответствующих главах книги. ; , 1
! i i 'i r?'j , и ' i I ; i •
1 । Г л а в a II
, ' ~ i . - . i ьч i .
1 ПОПУЛЯЦИОННАЯ цикличность
i И ЕЕ; ТЕРРИТОРИАЛЬНЫЕ РАЗЛИЧИЯ
' . ' 1
. I. . ' i : I
В этой главе обсуждаются оценки биологической цикличности гры-
зунов и насекомоядных, основанные На анализе отловов животных.
Как уже говорилось выше, попадаемость в канавки — функцйя двух
переменных: подвижности и численности. Обе переменные представ-
i
21
I ii
ляют собой сложные биологические процессы со многими циклично-
стями. Так, изменение численности происходит циклично в соот-
ветствии с появлением очередных генераций (приблизительно 40-
суточный цикл у полевок), возможно, имеются циклы миграций и
гибели. Вполне .вероятно, что увеличение численности путем имми-
граций тоже циклично, ибо пришельцы появляются в связи с избы-
точной плотностью в каких-то других местах. Аналогично можно
рассматривать и подвижность животных,. Все это наводит на мысль
о существовании волн подвижности и волн численности с разной
длиной периодов. Такое предположение сразу яге подтверждается
полученными- данными. Как мы уже отмечали, спектры попадаемо-
сти в канавки животных любого вида содержат множество пиков в
разных-полосах частот. Весь этот комплекс ритмов выделяется для
наземных позвоночных животных 1 впервые и его интерпретация
очень сложна из-за новизны способа обработки и представления ма-
териала.
। Множество пиков на спектрах существенно затрудняет анализ,
,потому что большинство этих мощных периодичностей описывают
колебания значений неизвестной нам функции. Наиболее логичный
выход при такой ситуации — попытка именовать пики на спектре,
присваивая им экологические, этологические или физиологические
значения. Это возможно в специально поставленных опытах, а так-
же при анализе большого количества литературных сведений по это-
логии, экологии и физиологии мелких млекопитающих. Цели же на-
ших исследований не столь широки, поэтому мы ограничимся рас-
смотрением отдельных периодичностей, а именно:
а) связанных, а возможно, и наведенных периодичностью клима-
тических воздействий (см. гл. I). Кроме того, 2—3-суточную рит-
мичность, подвижности мы относим за счет экологических факторов:
поиска пищи, охраны территории;
б) имеющих явный экологический смысл: например, 40—50-су-
точная цикличность связана с выходом молодых, 10—12-суточная —
с циркадной или лунной периодичностью, 90—100-суточная — с ми-
грациями (у водяной крысы) и др.;
в) обусловленных фазами обводненности Барабы. Это 4—5- и
10—12-летние периоды,;которые могут проявиться на спектрах, вы-
численных по многолетним данным отловов мелких млекопитающих.
Второй важный вопрос, который необходимо решить прежде,
чем мы перейдем к обсуждению спектров, заключается в выборе спо-
соба описания соответствий цикличности фазам увлажненности кли-
мата. Прямое решение этого вопроса состоит в. отборе только тех
ритмов, периоды которых соответствуют фазам увлажненности.
1 Спектральный анализ успешно применен- для выявления скрытых пери-
одических составляющих динамики численности котиков на о. Тюлений ’ [Ко-
лесник и др., 1978]. Описаны сезонные колебания численности разных демогра-
фических групп животных, периоды таких колебании были 8—12 сут, до 1 мео
и 1—2 мес. Применен он и для выяснения циклов численности обыкновенной
полевки в Центрально-Черноземном заповеднике [Николов, 1980].
22
В этом случае из обсуждения будут исключены своеобразные сезон-
ные и 2—3-летние ритмы жизнедеятельности популяций мелких
млекопитающих. Второй способ описания фаз увлажненности состо-
ит в учете и сравнении изменений коротких периодичностей (недель-
ных, сезонных и .пр.). Часть из них может изменять свою мощность
в зависимости от фазы обводненности или совсем исчезать на.спект-
рах. Вполне вероятны изменения соотношений- периодических со-
ставляющих в зависимости от фазы, т. е. сам вид спектра сезонных
периодичностей жизнедеятельности животного может быть своеоб-
разным маркером той или иной фазы обводненности территории, на
которой обитает этот биологический вид., :
Исходя из незначительного в сравнении с задачами исследования
промежутка времени, в течение которого*собирался материал, мы
будем использовать все возможности для интерпретации результа-i
тов в аспекте связи с фазами увлажненности, описанными для Ба-
рабы [Природные циклы..i., 1982]. Это значит, что будут выделены
ритмы соответствующих периодов, сгруппированы спектры внутри
каждой фазы и сравнены состав и мощность их гармонических со-
ставляющих.
Прежде всего нас в этой связи интересует, существует ли в сооб-
ществе некоторая группа популяций с хорошо проявленными изме-
нениями численности и подвижности в ответ на изменение увлажнен-
ности территории. Для этого было проведено сравнение спектров
ритмов у всех симпатрических популяций в различные фазы ув-
лажненности. Выделены группы видов насекомоядных и грызунов,
имеющие в своей внутрисезонной динамике множество цикличных
процессов, связанных с изменением численности и подвижности
животных. Эти группы видов отличаются от остальных тем, что внут-
рисезонная цикличность у них изменяется в зависимости от фазы
увлажненности.
i ЦИКЛЫ ДЛИНОЙ ДО ГОДА
В ПОПУЛЯЦИЯХ НАСЕКОМОЯДНЫХ И ГРЫЗУНОВ
В жизнедеятельности мелких млекопитающих одновременно могут
быть прослежены циклы разной длины. Мы старались выявить эти
циклы, узнать их характеристики на разных фазах обводнения тер-
ритории региона. Выполнение этой задачи было осложнено, отсут-
ствием данных о биологической функциональности многих выделен-
ных циклов. Некоторые из них понятны экологам (например,. 40-су-
точная периодичность попадаемостй серых полевок), но большин-
ство пока не расшифровано, их объяснение— задача будущего.
Насекомоядные. Популяции бурозубок в большинстве своем из-
меня4от внутрисезонную цикличность в соответствии с фазой увлаж-
ненности.
Для крупнозубой бурозубки представление о внут-
рисезонной цикличности в сухую фазу удалось получить по данным
23
I oo 4824 12 6 . 4 3 ' 2 <*>4824 12 S 4 3 c 2
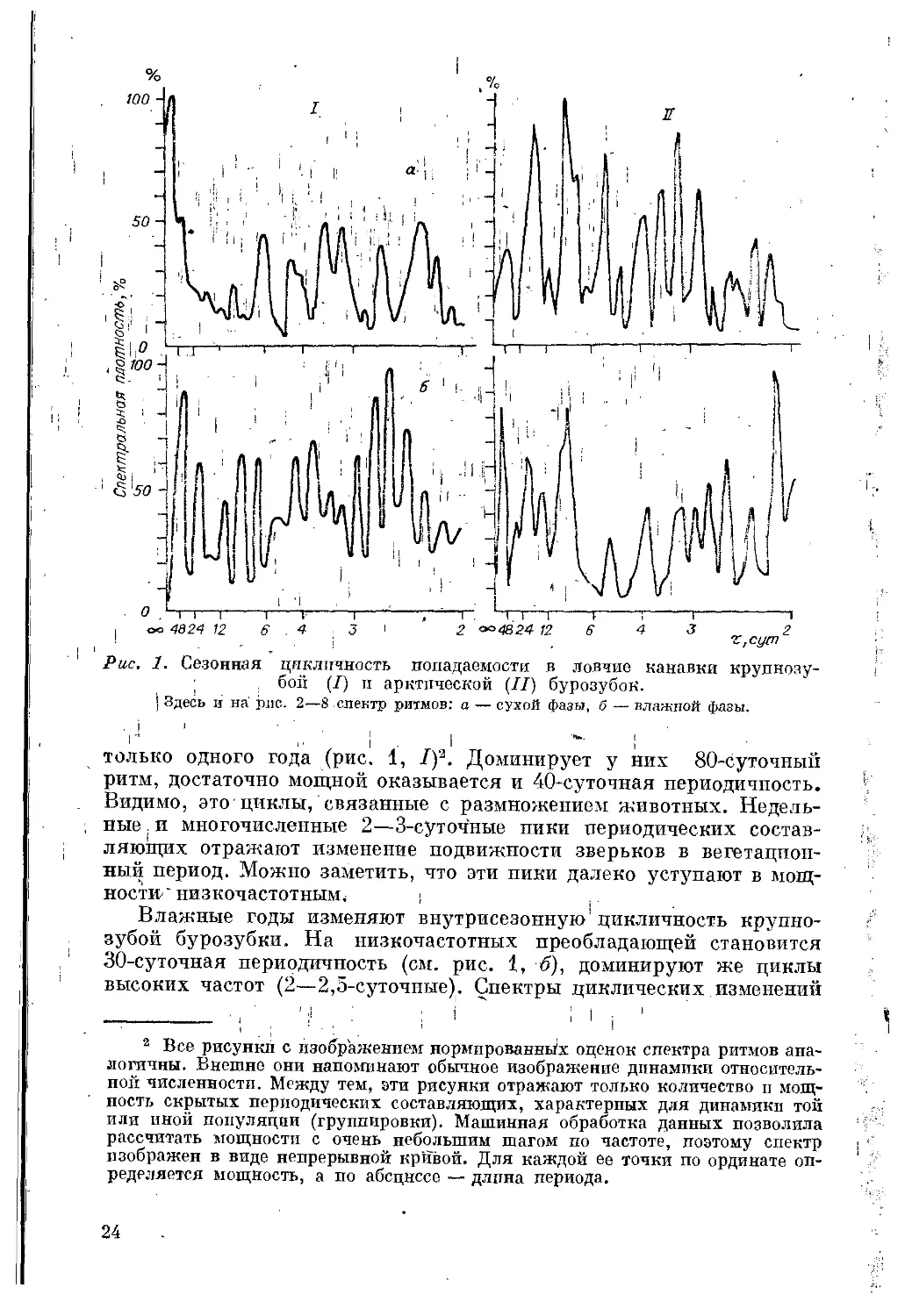
Рис. 1. Сезонная цикличность попадаемости в ловчие канавки крупнозу-
; бой (/) и арктической (II) бурозубок.
Здесь и на рис. 2—8 спектр ритмов; а — сухой фаза, б — влажной фазы.
। I । >
только одного года (рис. 1, /)* 2. Доминирует у них 80-суточнып
ритм, достаточно мощной оказывается и 40-суточная периодичность.
Видимо, это циклы, связанные с размножением животных. Недель-
ные . и многочисленные 2—3-суточные пики периодических состав-
ляющих отражают изменение подвижности зверьков в вегетацион-
ный период. Можно заметить, что эти пики далеко уступают в мощ-
ности^' низкочастотным. < .
Влажные годы изменяют внутрисезонную! цикличность крупно-
зубой бурозубки. На низкочастотных преобладающей становится
30-суточная периодичность (см. рис. 1, б), доминируют же циклы
высоких частот (2—2,5-суточные). Спектры циклических изменений
; ' . ’ L . : ' 1 i '
2 Все рисунки с изображением нормированных оценок спектра ритмов ана-
логичны. Внешне они напоминают обычное изображение динамики относитель-
ной численности. Между тем, эти рисунки отражают только количество и мощ-
ность скрытых периодических составляющих, характерных для динамики той
или иной популяции (группировки). Машинная обработка данных позволила
рассчитать мощности с очень небольшим шагом по частоте, поэтому спектр
изображен в виде непрерывной кривой. Для каждой ее точки по ординате оп-
ределяется мощность, а по абсциссе — длина периода.
24
в сухую и влажную фазы заметно различаются не только мощ-
ностью, но и количеством периодических составляющих. Во влажные
1оды хорошо проявленных циклов функционирования популяций
этого вида становится больше. , । I । , ’
Внутрисезонная цикличность^ арктической б|у р о'з у б-
к и мало зависит1, от фаз увлажненности .территорий.: Спектры по-
падаемости этихрверьков в! канавки всегда имеют множество мощ- i
пых пиков во всех шцлосах частот. Тем не менее, сравнивая спектры
сухого и влажного годов; (см, ,рис<1,!//, а, б) можно заметить разли-
чие в соотношении' мощностей' гармонических составляющих.
В сухой; год iпреобладают пики средних щастот, — от 24’до.З сут
(см. рис. 1,//, о). Влажный год характеризуется'спектром, । где до-
минируют низко- и высокочастотные составляющие (см. >рис’ ф, //,
б). Во влажные годы проявлены 80-, 20-и 8-*суточные периодичности,
а наряду, с ними — 2—-2,5-суточные; , ; '
Внутрисезонная цикличность ю б ь( к н о'в е, н н oi й ! буро-
зубки претерпевает наглядные изменения в зависимости от фазы!
увлажненности территории. В сухие годы значительную мощность
на спектре имеют пйки 80- и 9,6-суточиой цикличности. Соотноше- i
ние мощностей основных периодических составляющих, .попадаемо
сти в канавки обыкновенной бурозубки следующее: 1
Влажная фаза
80 241
73,57+12,63
Период, сут
Мощность, %
Период, сут
Мощность,- %
Период, сут
Мощность, %
6,8
19,43+4,57
2,7
16,86±3,49
Период, сут
Мощность, %
80
100±0
58,86+12,78
4,0
17,29+5,24
2,4
19+7,79
'Сухая фаза 1 ;
48 !
25+3,55
12
14,71+4,89
!3,4
21+6,92
9 9
17,14+3
30
28,5±6,03
9,6
। 12,43+3,27
3,0
23±7,71
9,6 .
6,5+1,06
Спектр периодичностей попадаемости зверьков в" это время беден ,
мощными пиками. Высокочастотные ’ пики почти не I проявлены
(рис. 2, I, а). Г ; : н , i I
В годы обводненности территории усиливается подвижность жи-
вотных и наряду |С циклами,, существующими в сухие годы, прояв-
ляется множество других в высокочастотной области., Спектр в это
время отличается многочисленными, но маломощными пиками < во ।
всех полосах частот (см.; рис. 2, I, б). i ; । и
Кроме различий в числе проявленных цикличностей между фаза-
ми существует значительная разница в мощности'одинаковых перио-
дических составляющих. Так, 80-суточная периодичность,, связан-
ная, очевидно, с размножением зверьков, особенно четко проявляет-
ся в сухие годы (t = 2,09; Р — 0,92). Различаются у бурозубок, и
циклы подвижности длиной 9,6 сут. Во влажные сезоны они выра-
жены сильнее (t = 1,76; Р = 0,86)!
Сухая фаза климата существенно воздействует на внутрисезон-
ную цикличность в популяции малой бурюзубки. В это вре-
мя доминируют 80-суточные и месячные циклы, число пиков на
25
i;,cym
Рис. 2. Сезонная цикличность попадаемостп в ловчие канавки обыкновен-
; ной (/) и малой (Л) бурозубок.
спектрах невелико (см. рис. 2, II, б). Среди коротких циклов по
мощности выделяются недельные и 2,7-суточные, которые, очевидно,
отражают изменения подвижности животных. Соотношение мощ-
ностей основных периодических составляющих попадаемости в ка-
навки малой бурозубки следующее:
Влажная фаза
Период, сут- 80 24 12 6,8
Мощность, % 84+9,62 45,57+8,59 49,71±12,6 45±11,36
:Период,.сут 4,8 3,4 • 2,8 2,5 2,2
Мощность; % ’ 37,71+12,15 40,43±12,4 36,43+7,46 36,71+10,07 53,57±11,65
।
’’ Сухая .фаза
Период,' сут • 80 30 9,6
Мощность, % 85+10,84 74±17,02 21,5+11,7
Период, сут 6,1 4 2,7 2,1
Мощность, % 41,5±27,3 23,5±15,25 52,5-Ь- 33,69 31,5±16,67
26
Во влажный период число мощных пиков на спектрах заметно,
возрастает (см. рис. 2, II, б), но 80-суточный ритм по-прежнему
остается мощным. Количество пиков увеличивается в связи с усиле-
, нием и усложнением подвижности животных. В это время проявлено
6 мощных периодических составляющих, связанных с подвижностью,,
периоды которых составляют от 2,2 до 6,8 сут.
Кроме несовпадения ряда периодических составляющих во влаж-
I ную и сухую фазы, существует еще и заметное различие в мощности
сходных гармоник. Так, цикличность подвижности (периоды 2,1 и
2.2 сут) во влажные годы проявлена сильнее,, чем в сухие (£ = 1,24;
Р = 0,73). , . ...
, На спектрах попадаемости сре.диих бурозубок , во
J влажную и сухую фазы присутствует большое число периодических
составляющих (рис. 3, I, б). Только в соотношениях их мощностей
обнаруживаются некоторые различия. В сухие годы при домини-
рующих 2,-3—4-суточных циклах подвижности имеется 80-суточная
1 периодичность изменений. Соотношение мощностей основных перио-
дических составляющих попадаемости в канавйи средней бурозубки
следующее:, । /
Влажная фаза
Период, сут 48 16 12 ; 6,8
Мощность, % 40+12,2 31,2+8,35 $5,6+11,1 66,6±16,9
Период, сут 4,8 4 3 2,5
Мощность, % 44,8±10,6 50+11,42 49,4+15,4 42,8±15,3
Период, сут 2,4 2,2 .
Мощность, % 33,6±9,9 60,0+13,6 Сухая фаза
Период, сут 80 24 16 9,6
Мощность, % 72,5± 19,5 57,5+19,5 57,5+5,32 43,5±9,6
Период, сут 8 5,3 4,0 2,7
Мощность, % 61±7,8 42±4,9 89+6,32 55±3,5
Период, сут 2,3 ।
Мощность, % 52,54+25,2
По количеству преобладают низкоч:астотные гармонические со-
ставляющие — от недельных до многомесячных.
Во влажные годы резко возрастает число короткопериодических
составляющих (от недели до 2 сут),, т. е. подвижность зверьков
усложняется по структуре. Высокочастотные ритмы преобладают,
однако достоверное изменение мощности при сравнении влажной
фазы с сухой наблюдается только у 16-суточной периодичности
(t = 2,65; Р — 0,95), которая больше проявлена в сухие годы.
Каких-либо заметных тенденций в изменении попадаемости буро-
зубок, зависящих от фазы увлажненности, не обнаружено. Каждая
популяция экологически специфична и на изменение j влажности
местообитаний, видимо, отвечает «включением» различных адапта-
ционных механизмов. Общим для этой группы животных является
обязательность изменений циклических характеристик при переходе
от одной фазы увлажненности к другой. Весьма вероятно, что во
влажные годы число периодических составляющих у всех видов бу-
27
«4824 12 6 4 3 '2<*>4824 12 6. 4 3- 2
. ! i i i; i cym
Puc.\3i Сезонная цикличность попадаембсти в ловчие канавки средней бу-
розубки (I) и лесной мыпювки (II).
•Ь I » ‘ . I н -
розубок возрастает, особенно это проявляется в области высоких
частот спектра; Обычно растут число и мощность пиков, соответст-
вующих 2—7-суточным циклам. ! 1 i
.Грызуны. У большинства!видов' грызунов изменяется внутрисе-
зонная цикличность численности и подвижности. в соответствии с
фазами увлажненности. В каждом конкретном случае (у каждой по-
пуляции); это вызвано разными причинами: изменением числен-
ности в ейлу размножения (циклы в 40 и 80 сут), миграций (60—
90 -сут); изменением подвижности (20-, 7-суточная, а также 5-, 3-
и 2-суточная, цикличность). Все. эти виды цикличности в популяции
образуют сложную систему взаимосвязанных колебаний. В сухие
годы такая система определенным образом адаптпро|вана! к экологи-
ческим требования^! вида. При переходе к влажной фазе происходит
ее перестройка, изменение мощностей и соотношений периодических
составляющих, т! ,е. адаптация к новым условиям. Видимо, поэтому
спектры, отражающие.пэту полипериодическую структуру жизнедея-
тельности популяций во: влажный и сухой периоды, оказываются
различными. , I
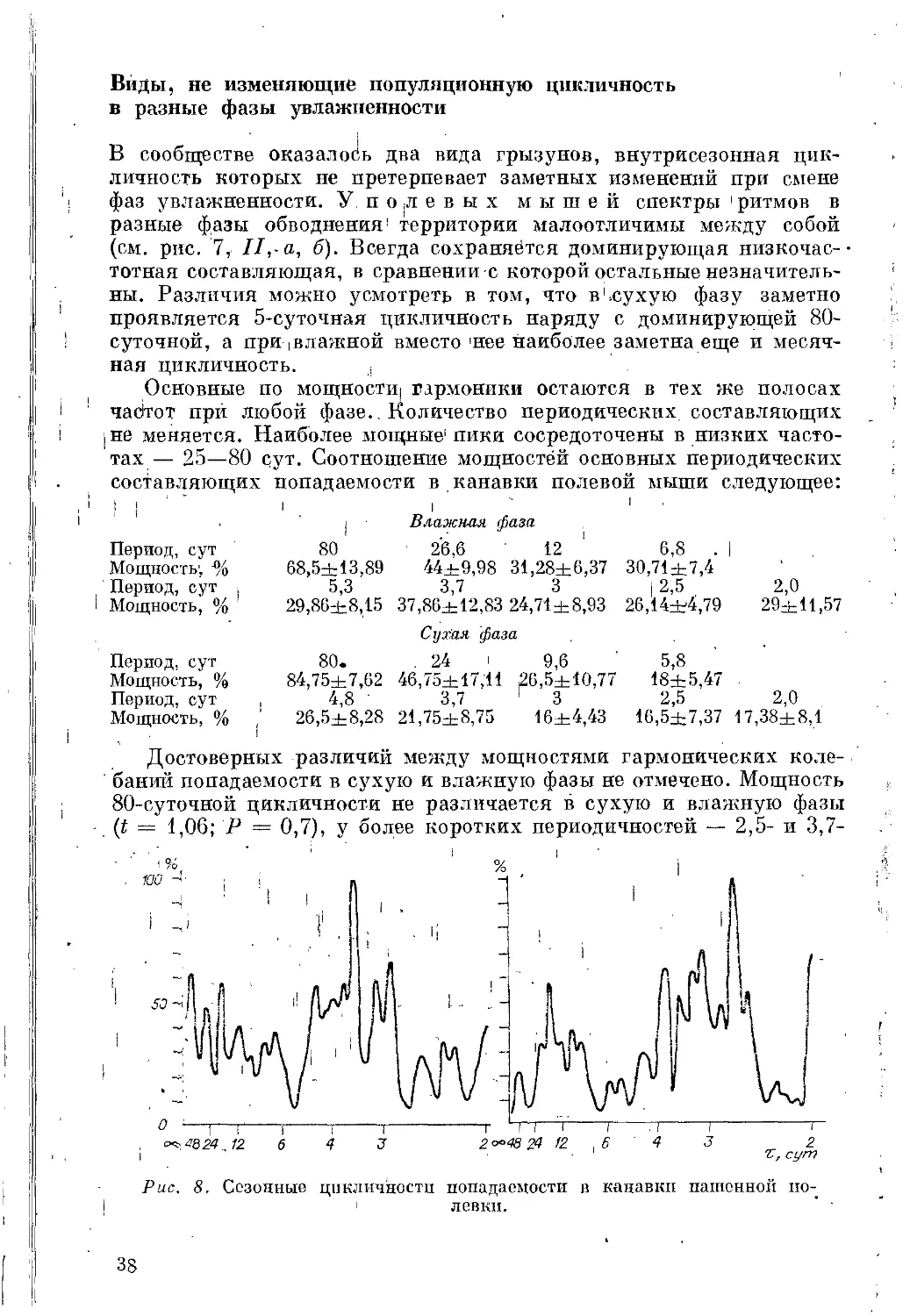
В сухие годы у лесной м ы ш о в к и преобладают 1,5-ме-
сячные и 24-суточные циклы (см. рис. 3, II, а). Первые можно от-
28
нести за счет периодичности выхода молодых: и изменения подвиж-
ности, а вторые вызваны, вероятно, миграциями и расселением.
В периоды обводненности наблюдается несколько иная цикличность.
Соотношение мощностей основных периодических составляющих
попадаемости в канавки лесной мышовки следующее:'
। ! - ! Влажная, фаза ; . 1 ,,
Период, сут Мощность, % Период, сут Мощность, % ' 80- . : 1 76,6±11,3 6,8 39,4±7,5 ' L 24 1 12 . ! л i.39,22±12,3' 42,3±7,3 4 , ' 3,4 ' 2,8 1 :: 2,4. 1! . • 1 2,1 56,3±Ю,7 1 .36,1±11,5 27,1±8,4 ; 49,2±8,9 ,
। ,5 1! 1 Сухая фаза . i
Период, сут | Мощность, % Период, сут Мощность, % 80 49,8± 23,7 8 1 58,5±16,6 1 jr 48 - 24 ! 12 ' i , ' 42,5±21,8 \ 61,3±13,3 ; 40,8+14,5^ i ; | 37 27' 23 2 ,57,5±7,7 \ 41,3±18,7 41>3±4,6 32,5±19,7
Доминируют периодические составляющие длиной около 3 мес.
Они, по-видимому, кратны длине обводненности. В эту же влажную
фазу у животных особенно хорошо проявляется 7-суточная, а также
3—4-суточная ритмичность (см. рис.'З, П, \б\.
Различия коротких ритмов по фазам увлажненности определяют-
ся не только соотношением мощностей периодических составляющих,
но и отсутствием отдельных из них в ту или иную климатическую фа-
зу. Так, во влажную фазу у этого вида нет 48-суточной цикличности.
Что же касается соотношения мощностей пиков, то в сухую фазу у
мышовки значительно мощнее 80-суточная составляющая (t = 2,32;
Р ~ 0,95), а во влажную — 2—2,5-суточные (t = 1,48; Р = 0,80).
У м ы ш и - м а’ л ю т к и внутрисезонная цикличность попадае-
мости в канавки хорошо различима по фазам увлажненности террито-
рии. В ।сухие годы спектр цикличностей этих грызунов имеет мень-
шее количество мощных периодических составляющих, чем во влаж-
ные (рис. 4, 7, б). Соотношение мощностей основных периодических
составляющих попадаемости в канавки мыши-малютки следующее:
h 1 ' Влажная фаза ( ~ .
Период, 1 1‘
сут ! 80" ' 24 ! 12 8
Мощность, % 82,22±12,67 43,67±9,62 40,55±9,13 30,89±11,81 ‘ !
Период, сут 6. 3,4 2,7 ' 2,2 12
Мощность, 0/0 37,33±10,0 33,22±10,16 1 28,33±10,71 1 ' ( 31±7,5 23±'5,2t
Период, сут ' 80 1 Сухая! 24 фаза 8 rС,/;1 i
Мощность, % 84,5±14,37 63,75±13,5 37,0±7,14 ' iJI ; ,
Период, сут , 4 3 н 2,2 ' .. / i
Мощность, % 49,5±21,5 25± 10,33 50,75±16,4 - 1 !
29
Рис. 4. Сезонная цикличность попадаемостп в ловчие канавки мышп-ма-
люткп (/) п обыкновенного хомяка (II).
Многочисленные пики во влажный период очень изменчивы по
мощности. Постоянно мощен только 80-суточный пик. В сухие же
: годы большой мощностью обладают как 80-суточный и околомесяч-
ный пики, так и средне- и высокочастотные. Даже на нашей малой
выборке проявляются заметные различия их’мощностей. Так, силь-
нее проявлена околомесячная периодичность в сухие годы (t = 1,21;
Р — 0,75). Цикличность подвижности с периодом около 2 сут также
сильнее проявляется в сухие сезоны.
Сравнивая наиболее влажный год влажной фазы с самым засуш-
ливым сухой (см. рис. 4,1, а,, б), можно заметить крайнее проявление
тенденции роста числа пиков на спектру. Большое число пиков,
особенно в высокочастотной области спрйтра, во влажный год сви-
детельствует о неустойчивости цикличности. Вполне вероятно, что
многие периодические составляющие наводятся внешними факто-
рами. /
Для обыкновенного хомяка характерна различия
в распределении п мощности коротких циклов, зависящие от той
или иной фазы увлажненности. Прежде всего, в сухой период на его
спектрах попадаемостп несколько меньше пиков. С ростом обвод-
30
ненности число их увеличивается. Цмеется некоторое несовпадение
полос частот во влажные и сухие годы, которым соответствуют мощ-
ные периодические составляющие (см. рис. 4, II, а, б). Соотношение
мощностей основных периодических составляющих попадаемости в
манавки обыкновенного хомяка следующее:
।
Влажна! г фаза- ' .: , •
Период, сут [48 ! 24 ( 16 9,6 : 6,8
Мощность, % 73±11,2 44,25±18,7 40,5±8,95 60,75+13,66 58±7,89
Период, сут 4,8 3,7 ' ; ^3,2 , [ 2,5 • } *
Мощность, % 51,75±8,24 33±6,07 54,25±15 ’ 62±8,93. 56,5±;14,4
Период, сут 80 Сухая фаза 24 : 12 '8
Мощность, % ; 76±6,45 47^75+10,8 35,5^18,71 51,75±15,94
Период, сут 6,1 4,4 2,5 ... 2
Мощность, % 64+14,98 75,75±13,18 44±1'2,5 61,75+19,37
Характерно и то^ что наибольшая мощность гармоник в сухие
годы проявлена как на низких (2—3-месячные), так й на высоких
(5—6 суточные) частотах, а во влажные годы большая мощность
соответствует средним и высоким частотам. Кроме того, имеются
различия в мощностях пиков и в совпадающих периодичностях.
В полосе [частот даже при такой непредставительной выборке
(4 влажных и 4 сухих сезона) есть значительное преобладание по
мощности гармоник с периодом около 5 сут (4,8 и 4,4) в сухую фазу
(t — 1,55; Р = 0,81), а периодичностей длиной в 2,5 сут — во влаж-
ный период (t — 1,18; Р = 0,72).
Таким образом, обыкновенный хомяк относится к животным, пе-
рестраивающим- цикличность жизнедеятельности в зависимости от
фазы увлажненности территории. i [
У красных полевок, наблюдается значительная измен-
чивость цикличности попадания в канавки в отдельные годы, но она
не маскирует различий, связанных со сменой фаз увлажненности.
В период большой обводненности, спектр попадаемости в канавки
значительно отличается от такового в период иссушения территории.
Это можно увидеть при сравнений спектров в очень сухой год и в
очень влажный (рис. 5, I, а). Особенно заметно снижение мощности
высокочастотных ритмов во влажный год. Во влажную фазу по мощнос-
ти преобладают более низкочастотные циклы: 80-суточные, недельные,
а в сухую — 4- и 2-суточные. Соотношение мощностей основных
периодических составляющих попадаемости в канавки красной
полевки следующее
Период, сут,
Мощность,, Й
Период, сут '+ J
Мощность, % !s
I „
Влажная, фаза
24 12
•' 35,25±9,35,• 25,12±7|,1Р
! 3 ,2,5 !
80'
69,4+7,1
РР 3,7.
.41,5±12,26 42,63±9,1 \ г-40,75±9’,68 35,.13±7,1-
*;! •• 1, । ' СухаЯ'Сфаза.; '
Р, 80. + ЦР ' W1;-’; ; 12 •
52+14,42 59,6^4,51 51,4±9,9 _
1 !»! |4,8; ....4 Д 3 'r2,7fi'
48±12,54 / 58±11,;01 '46'8^61 i’l 40 2±8,8
6,8
46,5+13,16 27,37±6,24
2,3
80
4,4
' 2~ ' ’
1 32,5±7,0
6Л г .
25,0+7,45 38,8±10,28 1
— 2 2
50,4±15,1
8i
, Период, сут 1
ч Мощность, °о
, +Период,,сут,
' .Мощность, %! !,
<; *3 8 , "! ! I , 1 ' ! । । -iL , I .
! И, , При этом в сухую фазу 12-и24-суточные периодические состав-
1 ляющие достоверно более; мощные, чем во влажную (i = 2,16;
у;, ;;1,96; Р = 0,95; О-,92)у тогда как;80-суточная’периодичность, связан-
ii' пая' скорее щсего; с изменением численности и миграциями этих поле-
• вок, лучше выражена во влажные водыi(i — '1,1; Р == 0,71). Наблю- у
. , дается, и -некоторое несовпадение1 периодов, i Boi' влажную фазу от-| -
11 сутствУют обычные,' для сухой 8- и 4-суточные периоды, а в сухую —'
;2,5т и 2,3-суточные, имеющие заметную мощность’ во влажную фазу, ; о
' т. е. в различные по обводненности территории фазы, имеются отли-
.. чия в характерных, сезонных циклах попадаемости.
1 ' >. L ' г
# > 1 ! 1 Р
, МД
1 . £0
100 -
О'
100-
На
Т. сит
0
°° 4624 12
И, сит
Рис. 5. Сезонна!я цикличность попадаемостп в ловчие канавки красной (I)
и рыжей (II) полевок.
32
У рыжих полевок в разные фазы увлажненности наблю-
даются различные внутрисезонные циклы. Основное отличие спект-
ров состоит в. том, ч:то при увлажнении климата число циклов под-
вижности снижается за счет «выпадения» цикличностей длиной в 12
и 5,3 сут. Заметно отличие и в распределении мощностей одних и
тех же периодических составляющих. В сухой период наряду с мощ-
ными низкочастотными ритмами (80 сут, неделя) наблюдаются и зна-
чительные высокочастотные — 2,4-суточные. Соотношение мощностей
основных периодических составляющих попадаемости в канавки
рыжей полевки следующее: .
1 Влажная фаза
Период, с^т Мощность, % Период, _сут t । Мощность, %' 80 ' 71;5+15,46 3,4 55,75+16,61 24 51,25+11,25 2,8 23,75+3,7 4 ! 32+7,35 2,2 18,5+3,71
Сухая фаза ।
Период, сут Мощность, % Период, сут Мощность, % 80 ' 98,3+1,36 5,3 . 87,33+3,21 24 45,33+22,66 4,4 50+2,37 12 28,33+2,72 3 31,67+5,45 2,4 51,66+21,39
При повышении обводненности такие ритмы (2,8—2,2 сут) хотя
и устойчивы, но маломощны; 80-суточные пики имеются на спектрах
ритмов попадаемости в обе фазы, но различны по мощности. В сухой
период они заметно выше (t = 1,73; Р — 0,88), так же как и 4-суточ-
ные (7 = 2,33; Р = 0,95). Трехсуточные ритмы, наоборот, оказы-
ваются мощными во влажную фазу (7 = 1,38; Р = 0,81). Различия в
мощности и количестве периодических составляющих в разные фазы
хотя и имеют значительную вариабельность от сезона к сезону, хо-
рошо различимы на спектрах (см. рцс. 5, II, а, б).
Внутрисезонная цикличность жизнедеятельности узкочереп-
ной полевки заметно различается в зависимости от фазы об-
водненности территории. Во влажные годы эти зверьки имеют огра-
ниченное число циклов, среди которых доминируют периодичности,
связанные с размножением и выходом молодых, 48- и 12-суточные.
Соотношение мощностей основных периодических составляющих
попадаемости в канавки узкочерепной полевки следующее:
Период, сут
Мощность, %
Период, сут
Мощность, %
80
50,88+13,56
6,1
45,88+8,48
Период, сут
Мощность, %
Период, сут
Мощность, %
'48
66,8+19,9
4,8
44,4±10,01
Влажная фаза
48 12
62,38+9,59 63,88+10,89
37 * 27 22
38,63+7,73 44+9,94 47,25+4,06
Сухая фаза
2А 12 '8 6,1
49,8+5,1 65,4+13,37 54,6+13,31 68,2+10,19
3,7 2,8 2,3 2,1
. 52+6,41 67,2+5,14. 85,6+6,96 70,9+11,85
В сухие годы число пиков на спектре резко увеличивается, и до-
минируют по мощности циклы, связанные с изменением подвиж-
ности, 2—3- и 6-суточные.
3 Л А. Максимов, Л. Н. Ердаков
33
&=48 Z4 12 '6 j ,4 2 ^>48 24121 8 ' n 4 / . 2
4 - - | 1 . г, сут I
Puc. 6. Сезонная цикличность попадаемости в ловчие канавки узкочерепной
полевки (/) и полевки-экономки (II).
; ‘ ’
Близкие по длине периодичности в'разные фазы различаются по
мощности1., Так, недельная цикличность заметно лучше проявлена в
сухие годы (t — 1,68; Р = 0,88). Короткие циклы длиной от 2 до
4 сут всегда проявлены лучше в сухую фазу: 3,7 сут (£ = 2,86; Р =
= 0,95); 27-2,,8 сут (t = 2,07; Р = 0,94); 2,3 и 2,1 сут (t = 4,76
и 1,89; Р = 0,99 и 0,92). ,•
i Таким образом^ в сухие годы для узкочерепной полевки харак-
Черны мощные ритмы подвижности в полосе 'высоких частот, мало
проявляющиеся во влажную фазу. Иллюстрацией резкого различия ‘
цикличности в соответствии с фазами увлажненности могут быть
спектры, взятые в наиболее сухой и .влажный годы |(рис. 6, I, а, б), л
, ~У п о ,л е в о к - э к о н о (м о к ’количество мощных пиков на -
спектре ритмов попаддемости одинаково в сухую и влажную фазу. ।
Тем не менее внутрисезонная цикличность их меняется в зависи-
мости от обводненности территории. Максимальная мощность перио-
дических составляющих во влажные годы сосредоточена в средних
и высоких частотах, от 2,5 до 12 сут. Соотношение мощностей основ-
ных периодических составляющих попадаемости в канавки полевки-
34
экономки следующее:
Влажная фаза
Период, сут 80 24 12 8
Мощность, % 65,3+9,5 50+8,3 68,1±10,6 62,63+5,91
Период, сут 4 3,4 2,5 2,2 2,0
Мощность, % 72,8±6,2 62,5ф7,2 54±6,11 60,75+9,34 29,75+8,8
Сухая фаза i
Период, сут 80 24 : 1,2 • /• 8 .
Мощность, % 77±18,1 41,6±13,91 35+8,88 42+12,25
Период, сут 5,3 4 3,4 . 2,7 ! 2,3
Мощность, % 41,4±5,98 36(6+10,18 36+12,8.1*- 30,4±8,5 : 34±8,5
В сухие же годы наиболее, проявлены циклы с длинными перио-
дами до 80 !сут. Последнее, видимо,1 связано с экологией этих поле-
вок и отражает смену местообитаний, происходящую в начале и кон-
це лета, т. е. с периодичностью 80—90 сут. Во влажные годы, оче-
видно, наблюдаются только мелкие миграции, потому что вся тер-
ритория сильно обводнена. При сравнении спектров внутрисезонной
цикличности влажного и сухого годов (см. рис. 6, II, а, б) видны зна-
чительные различия в соотношениях мощностей периодических со-
ставляющих. При увлажнении климата на спектрах наблюдается
Рис. 7. Сезонная цикличность попадаемости в ловчпё канавки водяной кры-
сы (7) и полевой мыши (II).. |
3*
35
много мощных пиков, среди которых доминирует 12-суточный. Высо-
кочастотные ритмы проявлены значительно сильнее, чем в сухую фа-
зу. Большинство периодичностей, имеющих аналогии в сухие годы,
во влажные достоверно более мощны. Так, 12- и 8-суточные ритмы
мощнее во влажные годы (t — 2,39 и 1,52; Р — 0,95 и 0,84). Ритмы
подвижности с периодами от 2 до 4 сут, как правило, лучше проявле-
ны во влажные годы: 4-суточные — t — 3,03; Р = 0,99; 3,4-суточ-
ные — t — 2,5; Р — 0,95. Периодичности 2,7' сут сухой фазы зна-
чительно уступают по мощности 2,5-суточным влажной (£ = 2,3;
Р = 0,95). В сухие же годы наблюдается абсолютное доминирование
• > 80-суточного цикла, а остальные сильно уступают ему по мощности
(см. рис. 6, II, а, б).
'. Спектр ритмов попадаёмости в .канавки в течение сухой фазы у
водяной крысы, напоминает «белый шум», который на спект-
ре обычно проявляется в виде одинаково мощных пиков во всех по-
лосах частот.. Подвижность животных в такие годы не имеет устойчи-
. вых доминирующих ритмов, и большая часть периодических состав-
ляющих не повторяется из года в год. Типичный пример спектра
ритмов попадаемости водяных крыс получен на материале отлова
1966 г. (рис. 7, I, б). Это как раз середина сухой фазы, и на спектре
видны мощные высокочастотные составляющие с периодом 2 сут.
Они и указывают на то, что спектр неустойчив. Во влажные годы
отловы водяной крысы имеют следующие устойчивые периодические
составляющие: 80, 12, 8 и 6 сут. Соотношение мощностей основных
периодических составляющих попадаемости в канавки водяной кры-
сы следующее:
Влажная фаза
Период, сут Мощность, % Период, сут Мощность, % 80 г 70,5+11,99 ' 6,1 , 33,5+6,73 24 40,5+9,08 , 3,7 38+9,32 8 46,75+6,9 2,5 44,88+ 3,65 . 2 2 45,63+6,68
Сухая фаза
Период, сут 80 24 12 8
Мощность, % : 54,2±10,01 49+11,43 56,4+5,78 72,2±7,75
Период, сут 6,1 ' 4,8 4,0 3,4
Мощность, % 84,4+7,08 59,8+8,51 46,6+10,12 70+9,79
Период, сут 2,8 • 2,5 2,2
Мощность, % 66+8,88 55+6,62 ! . 50+11,49
Эти ритмы ।имеют очевидный экологический смысл: длинные пе-
риоды связаны с размножением зверьков, а короткие, возможно,
отражают попадаемость при разных погодных условиях.
Несколько изменяется обычная картина ритмичности передвиже-
ния особей в период вспышки. Вспышка приходится на влажную
фазу. В это время внутрисезонные ритмы водяных крыс имеют до-
минирующую 90—100-суточную периодичность, что, по-видимому,
36
означает массовую смену местообитаний, происходящую верной и
осенью. Этот ритм особенно отчетливо проявлен при высокой чис-
ленности, в моменты депрессии он отсутствует.
Различия внутрисезонной цикличности водяной крысы во влаж-
ную и сухую фазы очень велики. Прежде всего они касаются коли-
чества мощных периодических составляющих. Во влажный период
на спектрах ритмов значительно меньше пиков, чем в сухой
(см. рис. 7, I, а, б). Доминируют во влажный период низкочастотные
циклы длиной в 1 и 2,5 мес. В сухую фазу в подвижности водяных
крыс наблюдается множество циклов с небольшой длиной периода.
Доминируют среди них недельные и 3-суточные. Экологическая
интерпретация этого такова: во влажные периоды происходит не-
уклонный рост численности и основные передвижения связаны с рас-
селением молодых (20—40 сут) и сезонными миграциями на гривы
(80—90 сут). В сухое время зверьки ищут оптимальные стации, по-
этому подвижность высока в течение всего вегетационного периода.
Величина пиков одинаковых периодических составляющих в
разные фазы увлажненности чаще всего достоверно различается^
В сухие годы по сравнению с влажными мощнее ритмы с периодом в
8 сут — t = 2,45; Р = 0,95; недельные различаются еще сильнее
t = 5,21; Р = 0,99 и 3—4-суточные тоже отличаются достоверно —
t = 2,37; Р = 0,95; t = 2,2; Р = 0,95. Это возрастание мощности
в сухие периоды связано с поисками подходящих стаций по мере
пересыхания болот.
Сезонные ритмы с периодом около 80 сут сильнее проявлены во
влажные годы (t = 1,04; Р = 0,66), когда отмечается массовый,
хорошо выраженный переход с болот на гривЫ. Летом в болотах
достаточно воды и водяные крысы успешно размножаются, пред-
принимая за. вегетационный период в зависимости от погоды обычно
5—7 подвижек. Осенью же вся масса зверьков выходит на гривы,,
определив тем самым мощный миграционный ритм. В сухие годы
часть болот пересыхает, многие зверьки выходят на гривы раньше
обычного и даже размножаются там. Поэтому осенняя миграция бы-
вает выражена гораздо'слабее.
Резюмируя вышеизложенное, можно сказать, что грызуны, как
п бурозубки, изменяют цикличность своей жизнедеятельности в за-1
висимости от фазы увлажненности территории. Это проявляется в
достоверных изменениях мощности и значений периодических состав-
ляющих на спектрах их ритмов. В то же время какой-либо оиреде-1 t
ленной закономерности в изменениях цикличности, присущей всем 1
видам грызунов, нами не отмечено. Число мощных сезонных ритмов
у них в отличие от бурозубок не возрастает во влажную фазу, не за-
метно и преобладания в это время высоких частот на спектрах. Ви-
димо, у грызунов приспособления к изменению влажности связаны
с экологической спецификой вида. Это не противоречит современ-
ному учению о различных уровнях адаптации. Вполне естественно,
что сухолюбивые формы реагируют на избыточную для них влаж-
ность совсем не так, как влаголюбивые.
37
Виды, не изменяющие популяционную цикличность
в разные фазы увлажненности
В сообществе оказалось два вида грызунов, внутрисезонная цик-
личность которых не претерпевает заметных изменений при смене
фаз увлажненности. У п о левых мышей спектры 1 ритмов в
разные фазы обводнения' территории малоотличимы между собой
(см. рис. 7, II,-а, б). Всегда сохраняётся доминирующая низкочас-•
тотная составляющая, в сравнении с которой остальные незначитель-
ны. Различия можно усмотреть в том, что в'.сухую фазу заметно
проявляется 5-суточная цикличность наряду с доминирующей 80-
суточной, а при (Влажной вместо !нее наиболее заметна еще и месяч-
ная цикличность. (
Основные по мощности, гармоники остаются в тех же полосах
частот при любой фазе.. Количество периодических составляющих
не меняется. Наиболее мощные' пики сосредоточены в низких часто-
тах— 25—80 сут. Соотношение мощностей основных периодических
составляющих попадаемости в канавки полевой мыши следующее:
Период, сут Мощность; % Период, сут । Мощность, % । Влажная фаза 80 26,6 12 6,8 . ! 68,5*13,89 44*9,98 31,28*6,37 30,71*7,4 5 3 3 7 3 | 2,5 2,0 29,86*8,15 37,86*12,83 24,71*8,93 26,14*4,79 29*11,57
Период, сут Мощность, % Период, сут Мощность, % Сухая фаза 80. . 24 । 9,6 5,8 84,75*7,62 46,75*17,11 26,5*10,77 18*5,47 , 4,8 ' 3,7 1 3 2,5 2,0 26,5*8,28 21,75*8,75 16*4,43 16,5*7,37 17,38*8,1
Достоверных различий между мощностями гармонических коле-
баний попадаемости в сухую и влажную фазы не отмечено. Мощность
80-суточной цикличности не различается в сухую и влажную фазы
(t = 1,06; Р = 0,7), у более коротких периодичностей — 2,5- и 3,7-
о ~—г—г—г------।‘—।---------г Н~т т i । г
оч>4824 12 б 4 J 2 <*48 24 12 , 6 ' 4 8 2
Т, сут
Рис. 8. Сезонные цикличности попадаемости в канавки пашенной по-
левки.
38
суточной тоже (t = 1,04; Р = 0,66 и t = 1,1; Р ~ 0,7). В фазу об-
воднения сокращается площадь биотопов, пригодных для полевой
мыши, так как увеличивается количество заболоченных мест. Поэто-
му возможно, что расселение зверьков, с территорий размножения
проходит менее интенсивно, чем в сухие !годы, и 80-суточная циклич-
ность при, увлажнении климата,!выражена слабее. ,; 1; ।
Внутрисезонные циклы у пашенных полевок не разли-
чаются выгоды разной увлажненности11(рис. 8)1.' Набор . периодич-
ностей остается тем' же самым. Соотношение ^мощностей основных
периодических составляющих попадаемости в канавки пашенной по-
левки следующее: - ' . > ( : ,1
Влажная | фаза
Период, сут Мощность, % ; Период, сут । ! 1 80 33,3±9,7 4,4 ' 24 58,4±4,1 3,2 1 9,6 1 43±12,5 2,8 ! 6,1 | 58,5±15,8 2,5 I 2,2
Мощность, % |43,6±4,9 63,1±13,2 61,5±10,3 60,8±10,8 48,8± 15,7
Сухая /фаза.
Период, сут , 80 24 1 9,6 6,1
Мощность, % 55±13,7 63,8±13,1 ' 51±16,1 42(4±15,6
Период, сут ... 4,4 3,2 2,8 2,5 2,2
Мощность, % 35,2±16,1 54,2±12,9 . 60±5,8 44,2±16,3 ..-47,8+16,6
Сохраняется то же соотношение мощностей, во всяком случае
достоверных различий в мощности в разные фазы ни одна полоса
частот не содержит. Наиболее проявлены околомесячные циклы,!
видимо, связанные С размножением, а таггже недельные и 2-суточ-
ные, обусловленные скорее всего изменением подвижности жи-
вотных.
В сухую фазу 80-суточная цикличность мощнее, чем во влажную,
но зти различия недостоверны (t = 1,29; Р — 0,7). | Возможно, они i
объясняются тем, что в сухие годы численность пашенной (Полевки
несколько выше, чем во влажные, следовательно, при расселениях
в конце лета в такие годы попадаемость в канавки также .немного
выше. , i
! .. -- | 1 ’ ; , Г "
Таким образом, мы выяснили,^ что в таких1 таксономически и эко-
логически несходных группах, как грызуны и бурозубки, в зависи-
мости от фазы увлажненности изменяется сезонная цикличность по-
пуляций. Выраженность этих изменений может быть различна1, как
в той, так и в другой группе существуют видьц различий сезонной ,
цикличности которых в зависимости от разных фаз увлажнения нам
установить не удалось. Для землероек характерна 2—2,5-мйсячная
сезонная цикличность. В сухие годы такие циклы оказываются са-
мыми мощными, во влажные яге — сильнее выражены более корот-
кие периодичности (2—8 сут). Число скрытых периодических] состав-
ляющих во влажной фазе больше, чем в сухой. У грызунов’обычны
80-суточные и месячные циклы. Во влажной фазе они доминируют
по мощности, а в сухую фазу часто оказываются доминирующими
в жизнедеятельности отдельных популяций короткие периодичности
39
(2—6-суточные). Адаптация насекомоядных и грызунов, судя по из-
менениям сезонной цикличности, в популяциях происходит видо-
специфично, и универсальных циклов, проявляющихся к определен-
ным изменениям среды, сразу у многих сосуществующих популяций
не отмечено.- *
МНОГОЛЕТНИЕ ЦИКЛЫ В ПОПУЛЯЦИЯХ МЕЛКИХ МЛЕКОПИТАЮЩИХ
Анализ временных рядов — дисциплина статистическая, и ее методы
широко используются в разных отраслях науки и техники. Его мож-
но применять^ случае больших объемов данных. При.расчетах внутри-.
! сезонных циклов использованы ряды, включающие до 1,5 тысяч
данных (там испбльзован ежесуточный материал). Для расчета мно-
голетних цикличностей длина ряда, естественно, сокращается за
счет увеличения шага суммирования данных, что сказывается на точ-
ности полученных результатов. Тем не менее, используя спектраль-
; ный анализ, мы можем с некоторой уверенностью выделить циклы с
периодами др 20 лет. Нас же особенно интересуют периодические
составляющие длиной в 9—13 лет и кратные им 4—4,5-летние. Пер-
: вые соответствуют периодичности увлажнения территории, а вторые
равны одной из фаз увлажненности..Известно, что сухая фаза длится
5—7 лет, а влажная — 4—5 лет [Максимов и др., 1979].
Описание многолетней цикличности в популяциях мелких млеко-
питающих удобно проводить в крупных эколого-таксономических !
! группах. Между видами в этих группах правомерно сравнивать на-
боры периодических составляющих. Одна из крупных групп —
насекомоядные, в наших данных они представлены 8 видами бурозу-
бок. Грызунов мы разделили на следующие группы: 1) мышовки,
мыши и хомяки; 2) рыжие лесные полевки; 3) серые полевки и водя-
ная крыса. По трофическим признакам эти группы представляют со-
бой градацию потребителей от высококалорийного до клетчаткового
корма. ; . . _ ,
Насекомоядные. Рассчитанные для разных вйдов насекомоядных
частотные спектры (рис. 9, 10) имеют от одной значительной перио-
дической составляющей (у крупнозубой бурозубки) до многочислен-
ных пиков (у крошечной). Только у двух видов бурозубок спектры
имеют заметные пики в области интересующих нас частот — циклич-
ностью с периодом около 10 лет обладают малая и арктическая буро- .
зубки. Эта цикличность по мощности не является доминирующей на
спектрах бурозубок, но оказывается столь же хорошо выраженной,
как и годовой цикл (см. рис. 9). . ।
Обе бурозубки широко распространены в Барабе и представляют Q
наиболее варьирующие по численности популяции насекомоядных.
С увлажнением территории численность арктической бурозубки воз-
растает, а малой снижается, Т. е. оба вида заметно реагируют па фа-
зы увлажненности, что отмечено при чисто биологическом анализе
популяций Барабы [Сообщества..., 1978].
40
%
Т, гпВ
Рис. 9. Спектры многолетней цикличности в популяциях арктической
(слева) и малой (справа) бурозубок из окрестностей с. Ровенское.
Рис. 10. Спектры многолетней цикличности попадаемостп в ловчие канавки
бурозубок.
а — средняя, б — обыкновенная, в — крошечная, г — крупнозубая.
Можно отметить большую общность спектров ритмов этих живот-
ных. Доминируют на них пики 1,5—1,8 лет. Очень мощными оказы-
ваются 3,5—4,3-летние, соответствующие продолжительности фаз.
I Следующие по мощности — годовые циклы,1 а затем 0,1-летние
(см. рис. 9).,‘|Дакое подобие спектров! возможно при значитель-
ном разделении экологических!' ниш животных. Hei обсуждая, здесь
экологию бурозубок, :можно объяснить этот феномен особенностями
'цикличности. Дело^в, томО'Что эти бурозубки находятся чаще всего в ।
'противофазе; по численности — арктическая! предпочитает влажные
! ! годы, а малая —Icyxnei. Такая предпочитаемость является достаточ-
ной для разделения видов и хорошо объясняет подобие спектров их
многолетней цикличности! , ; .
i Три вида бурозубок не имели ;на, спектрах многолетней циклич-
. 1ности гармонической составляющей в 10—13 лет. Самым низкочас-
1 1 тотным у них оказался пик, соответствующий периоду 7—8 лет
1 (см. рис. 10, а — в). У обыкновенной бурозубки (см. рис. 10, б) чис-
ло пиков на спектре меньше, чем у!двух других видов. У нее есть
! "только низкочастотные ритмы, самый маломощный из которых —
годовой. Этот вид во всех районах Барабы доминирует среди буро-
। ; зубок, а чаще всего и среди всех мелких млекопитающих. Он отно-
сителйно стабилен, т. <е. всегда богат особями, поэтому особенных!
колебаний численности обыкновенной бурозубки отметить удается'
редко. Исключение составляет период вспышки массового размноже-
ния водяной крысы. В такие годы численность обыкновенных буро-
зубок резко падает. Видимо, наиболее низкочастотную составляющую
этим бурозубкам наводят колебания численности в популяции водя-
, ной крысы. _ , |
Т', Средних и крошечных бурозубок1 в Барабе всегда мало и в лю-
'бую фазу увлажненности они сохраняют незначительную числен-
ность. Кроме циклов, характерных и для обыкновенной бурозуб-
ки, они имеют на спектрах и более высокочастотные пики, указы-
вающие на колебания их i попадаемости в ловушки в течение года
(см. рис. 10, а, в). , .|| 1 1
: Своеобразен спектр ритмов крупнозубой бурозубки. Он, как и у
обыкновенной (см. рис. 10, б, г), содержит небольшое количество пе-
риодических составляющих. У этого вида численность также колеб-
лется слабо, он занимает 3—4-е место що численности среди земле-
рое^ в период депрессии у водяной крысы, в годы же ее предпика и
пика оказывается весьма малочисленным (подробнее см. гл. V и
. 'табл! 40). Видимо, поэтому наиболее,мощная периодичность у вида
^составляет около 7 лет, т. е. в 2 раза длиннее доминирующего пе-
риода обыкновенной бурозубки (нм. рис. 10, г). <
I Таким образом, мы рассмотрели скрытые периодичности, свойст-
, венные бурозубкам в биоценозах Барабы. Большинство популяций
I этих, животных, 'испытывают циклические процессы (изменение чис-
ленности, подвижности). 7—8-летней длительности. Такая перио-
дичность значительно отличима от рассматриваемых нами 11-летних
циклов. Правда, па спектрах всех изученных видов существуют пи-
ки, кратные 11-годичным циклам,— 3—4-летние. Эта цикличность
. может быть приспособлением к сухим и влажным фазам климата.
42
Рис. 11. Спектры многолетней цикличности в популяциях лесной (а) и степ-
ной (б) мышовок п попадаемости в канавки полевой мыши (е) п мышп-малют-
ки (г).
i 'I
Два- вида землероек наряду с многолетними циклами имеют и
такие, период которых близок к периодичности увлажнения терри-
тории. Видимо, эти виды особенно хорощо адаптированы к циклам
Барабы. Они изменяют численность синхронно с фазами обводнения,,
тогда как у остальных - видов настройка колебаний происходит с
помощью । осцилляций, кратных 111-летним циклам.
Грызуны. Среди животных первой группы! степная мйшовка на
спектре ритмов жизнедеятельности имеет пик, соответствующий 13-1
летней цикличности (рис. 11, а). Оба >вида — степная и леснай мы-
шовки — испытывают колебания численности с периодами, кратны-
ми 11—13-летнему циклу увлажненности территории. Количество не-'
рподических составляющих у степной 'мышовки больше, чем у лесной
(см. рис. ,11, й, б), видимо, это связано с ее относительно редкой
встречаемостью в биоценозах.
Периодическую составляющую, близкую к продолжительности
цикла увлажнения, территории, имеют на спектрах оба вида мышей
(см. рис. 11, в, г). Причем именно эта цикличность у них оказывает-
ся наиболее мощной среди низкочастотных. Видимо, популяции
мышей хорошо адаптированы к смене фаз увлажненности террито-
рии Барабы. Мы отнесли полевую мышь к видам, цикличность кото-
43
। Рис. 12. Спектры многолетней цикличности в популяциях джунгарского хо-
мячка (а) и обыкновенного хомяка (б).
рых независима от фаз обводненности (см. выше). Видимо, она, имея
довольно устойчивый спектр высокочастотных ритмов, не зависящий
от фаз увлажнейности, обладает одновременно низкочастотным, синх-
ронным с циклом изменения метеорологических условий региона.
Хомяки имеют ритмы, близкие к периодичности увлажнения тер-
ритории. Наиболее редкий из них (джунгарский) обладает 11-летней
цикличностью, которая н доминирует на его спектре (рис. 12, а).
С помощью ее популяция хорошо адаптируется к ритму обводнения
территории. Данная популяционная цикличность позволяет зверьку
увеличивать численность точно в фазы иссушения.
Обыкновенный хомяк обладает почти 20-летней цикличностью,
но наиболее сильно у него проявлена 5—6-летняя периодичность,
кратная циклам обводненности территории (см. рис. 12, б). Грызун,
видимо, адаптирован в какой-то мере к этим циклам.
Таким образом, обычные для Барабы грызуны — потребители
калорийного корма, как правило, имеют среди своих периодических
составляющих цикличность, соответствующую периоду обводнен-
ности территории. Вероятно, это важная популяционная адаптация,-
без которой виду трудно создать значительную численность особей.
Вторая группа грызунов — три вида рыжых полевок. По трофи-
ческим потребностям они занимают промежуточное положение между
потребителями высоко- й низкокалорийного корма. Эти грызуны со
смешанной диетой обитают в закрытых биотопах и имеют большое
таксономическое родство, что, видимо, и обусловливает сходные
спектры их ритмов, делая их еще более похожими (рис. 13, а — в).
Все виды этой группы обладают цикличностью, соответствующей пе-
риодам обводненности (см. рис. 13, а). В Барабе было отмечено, что
рыжие лесные полевки значительно изменяют численность в соот-
ветствии с фазой увлажненности [Сообщества..., 1978]. Эта циклич-
ность и отражена на спектре, где самая мощная гармоника — 13-
летняя. Изменения численности — наиболее частый и заметный от-
44
Рис. 13. Спектры многолетней цик-
личности в популяциях полевок.
а — красной, б — рыжей, в — красно-се-
рой.
вет популяций на изменение
условий среды. Численность—1
интегральный показатель «бла-
гополучия» популяции. Адапт
тиров анность популяций к это-
му природному циклу дости-
гается наличием 3—4-летних ритмов жизнедеятельности, кратных
периоду обводнения территории. На спектрах этих полевок по
мощности они занимают обычно второе место.
У красно-серой полевки тоже есть 3—4-годовая периодичность,
но она маломощная (см. рис. 13, в). Отмечается та же закономер-
ность, что и у предыдущих групп мелких млекопитающих: чем мень-„
ше численность животного, ’ тем больше значительных пиков на
спектре его ритмов. Красно-серая полевка —- наиболее редкий вид
рассматриваемой группы. j ,
Таким образом, у наиболее многочисленных и обычных для дан-
ной местности грызунов со смешанной диетой существует адаптиро-
ванность к многолетним циклам обводненности. На спектрах ритмов
они имеют .соответствующую природному процессу 13-летнюю гар-
моническую составляющую и дополнительно кратные ей циклич-
ности.
Наиболее массовые виды в Барабе — представители третьей
группы: серые полевки и водяная крыса. Если эти полевки имеют
популяционную цикличность, соответствующую фазам увлажнен-
ности, то она, безусловно, отразится на валовых уловах зверьков.
В этом случае биотические изменения будут служить индикатором
многолетних метеорологических процессов. При отсутствии такой
цикличности уловы будут ацикличны. Хорошо известны примеры про-
43
ia — пашенная полевка; б — водяная крыса; в — полевка-экономка; г — узкояерепная
- полевка.
! ч н
гноза климатических явлений, исходя из вспышек численности в
популяции одного вида {Максимов и др., 1979].
При расчете спектров ритмов изменений численности многолет-
ние периодические составляющие выделены у трех видов: пашенной,
узкочерепной полевок и водяной крысы (рис. 14, а — в). Пашенная
полевка имеет примерйо 13-летнюю цикличность численности. В су-
хие годы ее численность несколько возрастает, а во влажные незна-
Г чительно снижается, тогда с 7-го м!еста по численности в сообществе
она передвигаете^ на 8-е [Сообщества..., 1978]. Колебания эти ।не-
велики по мощности, но устойчивы (правильный экспоненциальный
пик на спектре с узким основанием). Наиболее проявлены у этих
полевок 3—4-летние колебания. Такие ритмы для популяций поле-
вок давно известны [Матюшкин и др., 1972; Кошкина, 1969; Batzli,
1981]. Они 'кратны 11-летним циклам и, следовательно, способст-
вуют дополнительной, т. е.- большей, адаптированное™ популяции
пашенной полевки к многолетним циклам увлажненности терри-
.тории.
Для водяной крысы характерна примерно 10-летняя цикличность
численности в Барабе. На рассчитанном для нее спектре есть боль-
шой пик, соответствующий именноютой части (см. рис. 14, б). Наибо-
46
лее мощная гармоническая составляющая спектра водяной крысы,
как и у прочих полевок, 3—4-летняя. Она также кратна многолетне-
му циклу обводненности и, видимо, является добавочной адаптацией
к нему. Пиков на спектре водяной крьщы немного, как и у всех
зверьков, обладающих в ,Барабе большой численностью. i
Узкочерепная полевка имеет доминирующую периодичность дли-
ной около |l[3,4i года. Эта цикличность помогает синхронизации коле- •
баний популяционных' и обводненности территории. Дополнительно
на спектре присутствуют пики 5,5- и 3—4-летние, щричем последние
достигают значительной мощности (см. рис.. 14, а),' Такие колебания
численности могут совпадать по фазе! с многолетними колебаниями
обводненности и тем самым тоже быть адаптивными для популяции.
При анализе' изменений численности узкочерепной полевки в связи '
с фазами увлажненности уже отмечалось, что эта' полевка, будучи
лабильным видом среди мелких млекопитающих, значительно из-
меняет свою численность в соответствии с фазой увлажненности.
[Сообщества..., 1978]. Это можйо считать ^хорошей адаптированно- .
стью популяции. вида к многолетним колебаниям условий среды. '
У двух видов 'серых полевок нет многолетней цикличности, соот-
ветствующей фазам обводненности территории. Но у полевки-эко-
номки имеется; гармоническая составляющая, соответствующая поло-
вине периода обводненности. Она может быть приспособлением имен-
но к циклу обводненности местности. Во всяком случае, у полевок
описаны периодичности, адаптированные к какому-либо процессу и
при этом кратные ему по величине периода [Daan, Slopsema, 1978;
Jost, 1973]. Наиболее мощная периодичность у этих зверьков —
обычная для полевок: 3—4-цетняя (см. рис. 14, в). Наличие'такой
периодичности может служить адаптацией к 10—13-летнему циклу
природных условий. । ’
Таким образом, все рассмотренные популяций на спектрах изме-
нения численности имеют хорошо выраженные пики, соответствую-
щие различным по значению периода многолетним циклам. При
сравнении спектров мелких млекопитающих можно заметить, что
количество значительных по мощности периодических составляющих
(пиков) находится в обратной связи с численностью данного вида
животных. Фоновые, многочисленные (богатые особями) виды на
спектрах ритмов имеют Небольшое । число мощных гармонических
составляющих. I I . i
Для каждой трофической группы грызунов были посчитаны сред-
ние мощности разных периодических составляющих их ритмов чис-
ленности (табл. 5). У потребителей высококалорийного корма пре-
обладают периодичности в 1,5—2 года, на втором месте стоят 10-лет- _
ние циклы, видимо, связанные с обводненностью территории. В этой
группе представлены в основном сухолюбивые ]впды, и для их успеш-
ного проживания в БараЦе нужна адаптированность к большой
влажности, которая здесь периодически наблюдается. При отсутст-
вии такой адаптации во влажные периоды численность представите-
лей этой группы снижается. Следовательно, колебания влажности
и наводят этот один из ^наиболее мощных ритмов численности в дан-
ной группе животных. Важными ритмами жизнедеятельности попу-
47
Таблица 5
Средние мощности периодических составляющих (в единицах спектральной плотности) численности грызунов разных
трофических групп
Группа потреби- телей. Статистиче- ская оценка* Период в годах
19,7 13,1 9,84 6,56 5,61 4,36 3—4 1,5—2. 0,9—1,1 0,15 0,1 0,055
Калорийного \м 0 30,0 38,1 16,1 20,0 22,2 32,8 66,1 33,6 22,7 22,1 11,1
корма т — 12,5 13,3 5,8 9,4 7,9 9,8 9,6 9,6 3,3 7,4 2,5-
и / — 124,7 104,5 0 141,5 106,7 89,9 43,6 86,0 43,2- 66,1 22,5
Смешанного м 0 40,0 25,0 0 21,2 33,5 48,5 35,5 42,5 25,3 22,2 17,5
корма т — 17,3 13,7 — 10,2 14,3 3,6 12,5 15,3 6,3 8,3 5,2
- и — 105,3 134,0 — 117,5 104,7 48,1 86,2 - 88,5 60,9 91,4 73,1
Клетчаткового м 15,8 27,5 25,0 26,3 20,4 22,5 51,6 28,8 12,8 . 11,9 12,8 12,1
корма 5,6 9,7 9,7 9,6 7,5 8,3 11,0 7,3 1,2 1,2 1,5 1,9
U 122,2 121,5 134,0 126,2 126,9 127,1 74,0 87,5 32,0 36,1 / 40,6 57,1
* М — средняя, т — ошибка средней, ц — коэффициент вариации,
ляций являются и мощные годовые, 3—4- и 13,1-летние периодич-
ности (см. табл. 5).
Рыжие лесные полевки > образуют обособленную эколого-таксоно-
мическую группу зверьков, питающихся смешанным кормом, при-
чем наиболее концентрированный корм употребляет красная полев-
ка, а наиболее грубый (клетчатковый) — красно-серая. В группе
рыжих лесных полевок самую большую мощность имеет цирканная
периодичность изменений численности (см. табл. 15) и почти та-
кую же — 13-летняя. Эти зверьки не менее, чем потребители кон-
центрированного корма, адаптированы к ритмам обводненности тер-
ритории. Такая адаптация лесных полевок отмечена в описаний био-
топических комплексов мелких.млекопитающих Барабы [Сообщест-
ва..., 1978]. Хорошая адаптация этой группы грызунов к фазам ув-
лажненности удивительна, потому что рыжие полевки стенотопны,
обитают исключительно в колках. Колки же по микроклимату ста-
бильнее других биотопов, они хорошо задерживают влагу, т. е.
местообитания рыжих полевок термо- и гидростатированы, и циклы
обводненности в них наименее проявлены, чем в прочих биотопах.
По-видимому, кроме метеорологических факторов, оказывающих,
влияние на многолетние колебания численности мелких млекопитаю-
щих в биоценозах Барабьг; существуют еще и чисто биологические
колебания, могущие также служить осцилляторами 10—13-летней
цикличности. Возможно, через эти биотические колебательные сис-
темы осуществляется передача колебаний условий среды к популя-
циям. Передача колебаний осуществляется скорее всего через попу-
ляции грызунов — обитателей открытых биотопов. Важным меха-
низмом такой передачи служит смена доминантой в сообществе и
особенно специфика Барабы, из-за которой доминант всегда наибо-
лее многочислен во всех (даже не свойственных ему) местообитаниях.
В этом случае периодические появления больших масс, скажем, се-
рых полевок в колках, являются фактором, поддерживающим коле-
бания численности аборигенного населения колка. Серые полевки
должны особенно сильно испытывать периодичность сменй фаз ув-
лажненности. ।
Наиболее выражены в цикличности численности серых полевок
и водяной крысы периодические составляющие длиной в 3—4 года
(см. табл. 5). Приблизительно одинаковы по мощности колебания
численности с периодами 13, 10 и 6,5 лет. Они 4- следующие по
выраженности после 3—4-летних, т. е. у серых полевок хорошо про-
явлен (лучше, чем в остальных трофических группах грызунов,
п лучше, чему бурозубок) весь набор периодических составляющих,
синхронизированных с периодичностью увлажненности Барабы.
Остальные периодические составляющие на спектрах этих животных
заметно уступают! по мощности перечисленным.
Наиболее выраженные периодичности численности (оценка по
попадаемости в ловчие канавки) у бурозубок — циклы от 0,4 года
до 2 лет, а у грызунов 1—3-летние. Такие периодические составляю-
щие динамики популяций не только наиболее частые, но и наиболее
мощные по своим проявлениям.
4 А. А. Максимов, Л. Н. Ердаков
49
ТЕРРИТОРИАЛЬНЫЕ ОСОБЕННОСТИ ЦИКЛИЧНОСТИ
Рассматривая цикличность в популяциях мелких млекопитающих^
можно предположить как.наличие, так и отсутствие .синхронности
; в движениях их численности и изменениях подвижности на большой |
1 территорий. Как известно, мелкие млекопитающие населяют верх-
!!НийрСлойрпочвы,. т. ;е. существуют!в, некоторой изоляции от обычно
1 (измеряемых: климатических факторов (температура и влажность
1 “ воздуха, ветер и tj п.):. Микроклимат же их: местообитаний измере-
। ,нию и тем более длительному прослеживанию не всегда поддается.
, 'Ориентируясь,ра некоторую связь климата: й микроклимата, можно
предположить, что длительные, многолетние,, циклы у одинаковых
। видов из разных географических точек должны совпадать. Это тем
выраженнсе; чем шире распространяется 10—13-летПяя цикличность.
Совпадать или,: во всяком случае, оказываться близкими по про-
должительности на разных стационарах должны, очевидно, и такие
периодичности, длина которых обусловлена устойчивыми видовыми
i характеристиками — сроком беременности, числом генераций за
репродуктивный период, сезонными миграциями,- Остальные виды
i цикличности, связанные, например, с передвижениями в зависимости
от । локальных перемен климата, с сокращением численности из-за
биотического воздействия или неожиданной । климатической катаст-
рофы (засуха на небольшой площади региона), могут в значительной
мере не совпадать у животных одного и того же вида из разных
; - географических точек., - . " ~
Изучаемые нами популяции обитают в одном регионе (Барабе) на
значительном расстоянии. Причем между с. Усть-Ургулька и Потю-
каиовоИоно заметно меньше, чем от обоих этих пунктов до с. Ровен-
ское (см. гл. I). Для достоверной количественной оценки соотноше-
ний периодических составляющих число пунктов недостаточно, '
а сроки работ на них весьма коротки. Поэтому мы проведем только
1 качественный анализ соотношений мощностей периодических состав-
ляющих у каждого вида животных в разных пунктах наблюдения.
Насекомоядные. У бурозубок по грубой оценке наибольшее сов-
падение. цикличностей на разных стационарах — 87,5% (у крошеч-
ной бурозубки)^ а наименьшее —20% (у куторы). У наиболее мас-
, нового вида — обыкновенной бурозубки — цикличности совпадают
1 в'37,5%; примерно та же доля совпадений у крупнозубой и аркти-
ческой бурозубок. Фоновый-, барабинскпй вид — малая бурозубка
имеет значительную долЮ| совпадений; цикличности —66,7'%, и при- -
мерно; столько же — средняя; Эти два вида, а1 также (особенно) кро-
; шечная бурозубка/ видимо, весьма синхронно изменяют численность .
и подвижность'на значительной территории Барабы. ы
У подавляющего большинства видов (у 6 из 7 изученных) в райо-
не i с. Потюканово устойчиво проявляется 13-летняя цикличность
{табл!-6). Не прослежена она только у крошечной бурозубки, что
может быть связано не с отсутствием ее, а с постоянной малой чис- :
ленностыо этого вида в районе исследований. Преобладание во вре- :
меннбм ряду нулей вместо значащих цифр может создавать замет- -
50
Таблица 6
Циклы численности бурозубок в разных географических точках
Вид Место Период в годах
13,1 ' 9,84 7,87 6,56 4, 36 3,4-М 1,7-2,3 1,4-1,”6 1-1,3 0,7-0,9 0,15 0,055
Бурозубки: — — -
обыкновенная р — — + — , — + — + + — — —
П • 4- — — 4- - — — — . + — — • - + —
'У — — 4- — — _ — 4- — -и — — —
малая р — + — — - - -1- -и — — +
п - - — — + - - — + + + . —
У . —- — 4- + -и -- + . -и-. —
крупнозубая р — — — +' — + . 4- — —.
и — -и •— —. — - — + — — —
У - —• • — — + - + - — +
арктическая - + — — 4- — + - + — + ' + —
п + — — — — + — , -— 4- . -ь. -F —
У - — — . — — -1- — -н 2— . — , + . +
.средняя — р п 4- 1 — + 1 1 — 4- 4- + 1 + 4- 4- 4- + +
У - — — — —. — 4- —. — ——- + . — +
крошечная - р — — 4- —— — — + 4- + 4- + 4-
— п — + —~ — — — -1- + — 4- +
Кутора У р — -— + . Нет данных + —- + —
: п -4 — — — — — 4- — .+ — —
— у- — — — — — — + 4- - —
Примечание. Здесь и в табл. 7: Р — Ровенское, II — Потюканово, У — Усть-Ургулька.
ный «шум» при расчетах периодических составляющих, и имеющиеся
циклы невозможно будет выявить.
Рассматривая все виды цикличности, можно выяснить, какие из
них наиболее характерны для бурозубок. Мы считаем таковыми те
периодические составляющие, которые встречаются наиболее часто
у всех видов и па всех стационарах. По этому принципу самые харак-
терные цикличности сосредоточены в полосе частот 1,4—1,6 года
(см. табл. 6). Почти столь же часты циклы с периодами 1,7—2,3 и
0,5—0,6 года. Все эти периодические составляющие средней длитель-
ности, видимо, отражают обычную .динамику численности популяций
в сообществе землероек, периодичность их размножений и миграций.
Многолетние циклы длительностью в 10—13 лет, соответствующие
периодичности обводнения на спектрах( бурозубок, фиксируются
редко. Среднее положение по частоте встречаемости занимают циклы
протяженностью в 3—4 года, кратные фазам увлажненности и также
могущие служить адаптацией к многолетним колебаниям климата.
Исходя из. этого рассмотрения, можно предполагать, что’ наибо-
лее важными для жизнедеятельности популяции являются циклич-
ности, встречающиеся часто и у многих видов. Циклы же, синхрон-
ные многолетним процессам с нечетко выраженным периодом, более
редки. Очевидно, это связано с трудностью выработки такой попу-
ляционной адаптаций зверьками с коротким, относительно этой
цикличности, сроком жизни особи. Кроме того, неточная периодич-
ность процесса затрудняет приспособление к нему, и, видимо, ком-
промиссный путь — выработка нескольких циклических процессов
с близкими периодами (10, И, 13 лет), а наряду с этим наличие
кратных цикличностей в 1/2, 1/3, 1/5 периода многолетнего цикли-
ческого процесса. Это предположение объясняет причины поли-
периодичности колебаний численности мелких млекопитающих, на-
личие у одной и той же популяции мощных гармонических состав-
ляющих, близких друг другу по частоте, и приспособленность бу-
розубок к многолетней природной цикличности.
Грызуны. Циклические изменения относительной численности в
популяциях грызунов, по-видимому, в значительной мере связаны с
определенными местообитаниями. Нетрудно заметить, что животные
। одного и того» же вида, взятые в различных географических точках,
отличаются по многим периодическим составляющим между собой
сильнее, чем грызуны разных видов, но из одного района (географи-
ческой точки). Так, многолетние циклы водяной крысы в Ровенском
имеют период 9,84 года, в Потюканове — 13,1 года, а в Усть-Ургуль-
ке — 19,7 и 6,5 лет (табл. 7); В то же время в Ровенском 6 видов
грызунов из рассмотренных 10 имеют цикличность 13,1 года, а в
Потюканово 5 видов грызунов из рассмотренных 8 имеют периоди-
ческую составляющую численность 9,84 года. Эта особенность рит-
мов численности свидетельствует в пользу наведения тех или иных
периодичностей (особенно низкочастотных) внешними воздействия-
ми. Другими словами существуют внешняя среда местообитания,
довольно одинаковая для всех обитающих видов грызунов, и частные
экологические особенности у популяций, которые заметно диффе-
52
Таблица 7
Циклы численности грызунов в разных географических точках
СП
со
Вид Место 13,1
19,7
Мыптовка лесная в — >
п — —
У — —
Мышь полевая р — +
п — —
У — —
Мышь-малютка р * ч-
п —. —.
У — —
Хомяк обыкновен- р . —
ный п
- У
Полевки
красная р — +
п — —
У — —
рыжая р — +
и —-!— ——
У, — —
Водяная крыса р — ——
II — ч-
У + —
Полевки
узкочерепная р — +
п — —
У — —
пашенная р — +
II — —
У — —
экономка р —
п — —
У - с. —
Период в го
9,84 6,56 5,61 4,36 3-4 1
. — + — +
— — — Ч-“Г
— •h —. — -т
- — — — — ч-
+ — — — —
— —. — —
—- — — — ч-
+ — — —. ч-
— — — ч- —
— + — -—
Нет данных
Нет данных
— - - — — . ч-
+ — — — ч-
—.. — — + —
— —- — —• ч-
— — + —• —
— — ч- —
+ — ‘ ч-
— — — ч-
— + — ч-
— — -h — ч-
+ — -— — —
+ — — ч-
— - - — ч-
— - — •— —
—• — -- + —
— + — — +
— — — — ч-
.
I I + I++I II I I I I I I
ренцированы друг от друга разнокачественностыо своих требований
к среде. Если бы ритмы численности обусловливались внутренними
причинами, то они были бы более видоспецифичны и различия между
популяциями одного вида в разных точках оказались бы меньше,
чем между популяциями разных видов из одной местности.
В то же время более короткие ритмы, особенно начиная с перио-
дов в 1,5—2 года, повторяются для многих виддв независимо от
стационаров, где-проводились наблюдения. Видимо, начиная с этих
- периодических составляющих и к более высоким частотам усили-
'вается роль эндогенной компоненты в появлении колебаний числен-
ности. ; । i !
Чаще .всего спектру ритмов численности грызуна присуще 4—
5 м!ощных периодических составляющих (см. табл. 7). Во всяком •
случае, у большинства исследованных нами грызунов наблюдалось
именно такое количество пиков в диапазоне периодичностей от 20 ,
лет до 0,055 года. При сравнении трех сообществ, как правило, пары
одинаковых популяций из Потюканово и Усть-Ургульки по частот-
ным характеристикам численности оказываются более сходными, чем
любая-из них с >соответствующей из с. Ровенское. Это обусловлено
географической близостью пунктов Усть-Ургулька и Потюканово.
' Оценивая общность периодических составляющих численности
для каждого вида грызунов по; всем трем пунктам, можно заметить,
что наибольшее сходство имеют популяции полевки-экономки
(см. табл. 7). Наиболее выражено различие у популяций рыжей по-
левки (32,2% общности). У подавляющего большинства популяций
грызунов различие в составе периодичностей оказывается большим,
чем сходство. Исключение, кроме полевки-экономки, составляют
популяции лесной мышовки. Это1 может быть еще одним указанием
на диктат среды, большей зависимости, циклических процессов в ,
популяции от внешних условий, а не от .внутренних популяционных '
механизмов.
Чаще всего ритмы численности у грызунов имеют периоды 2 го-
да, 1 год, 6—8 мес и 3 года. Эти периодичности есть у всех грызунов
во всех популяциях. Заметно проявлены трехлетние циклы у поле- ’
вок, причем у водяной крысы такая цикличность численности не толь- !
ко имеется во всех популяциях, но и доминирует (см. табл. 7). Циклы
с короткими периодами у грызунов менее мощные, чем с длинными. .
Очень редко на спектре у грызунов наблюдается 20-летний цикл
(гораздо реже, чем у бурозубок) и 5—6-летние‘. Цикличность числен-
hocthj соответствующая обводнению территории, выражена хорошо
и является обычной для их спектров. Различия, как уже указыва- ,
лось, могут состоять в том, что в одной географической точке лучше
проявлен 13-летний цикл, а в другой — 10-летний.
Несмотря на хорошо выраженные различия цикличности в раз-
ных точках, ‘ подавляющее большинство популяций имеет 10—13-
летнюю ритмичность, т.'е. их колебание соответствует циклам обвод- .
ненности Барабы. Различия значений периода в разных точках ре-
гиона определяются особенностями локального климата. Так, в райо-
54
не с. Ровенское лучше проявлена 13-летняя цикличность, а около
с. Потюканово — 10-летняя (см. табл. 7).
Различия ритмической организации популяций проявляются не
только в разных географических точках, но и у разных трофических
и таксономических групп мелких ^млекопитающих. Наиболее сильны
они между; бурозубками^ и грызунами; Хорошо различима циклич-
ность в популяциях потребителей концентрированных, смешанных й
низкокалорийных кормов. ‘ । , I [
Метод спектрального анализа, использованный для 1 выявления
скрытых колебаний численности, оказался наиболее удобным из
известных нам. Он позволил выявить весь набор колебаний числен-
ности (популяции, получить соотношение мощностей', периодических
составляющих, что, в свою очередь, помогло широкому межвидовому ।
и межпопуляционному сравнению ритмичностей, определению наи-
более мощных и устойчивых среди них.
Выявление полипериодического процесса колебаний численности
в популяциях мелких млекопитающих дает возможность построения!
краткосрочных, а возможно, и долгосрочных прогнозов изменений
численности этих животных. Первым шагом такого прогнозирования
может быть математическое моделирование изменений в полипериоди-
ческой системе, скажем, в [связи с положением колебаний, далеких
по частоте. Для приближенного прЬгноза в таких моделях можно ис-
пользовать полученные нами, спектры ритмов жизнедеятельности!
барабинских популяций. I
' ; ' ' i i Глава III
[ ! 1 ' I 1
ЦИКЛИЧНОСТЬ В БИОТОПИЧЕСКИХ ГРУППИРОВКАХ
МЕЛКИХ МЛЕКОПИТАЮЩИХ
1 ! I ' ; । ' I
ОСОБЕННОСТИ ЦИКЛОВ В НАСЕЛЕНИИ МЕЛКИХ МЛЕКОПИТАЮЩИХ
ОСНОВНЫХ БИОТОПОВ! БАРАБЫ
Биотопические Группировки грызунов Барабы уже описаны [Сооб-
щества..., 19781. Нас интересовала цикличность изменений числен-
ности грызунов в различных биотопах. Для оценки циклов мы ис-
пользовали суммарные данные попадаемости грызунов в канавки в
том или ином биотопе. Ежегодно в каждом из выделенных биотопов
получали временной ряд суммарной [посуточной попадаемости зверь-
ков и анализировали его с точки зрения цикличности,
Предполагалось, что сообщество грызунов в каждом местооби-
тании изменяется от Рода к году. Происходящие изменения в струк-
туре сообщества грызунов отражаются на мощности'и соотношении
периодов их экологической цикличности. Для анализа циклов раз-
ной длины использовали материалы по трем массовым размножениям
водяной крысы (с. Ровенское). Одно из них было в 1961 г. в конце
55
влажной фазы; второе — в 1972 г. в середине следующей влажной
фазы и третье — в 1980 г., когда начавшееся в 1979 г. увлажнение
местообитаний уже в 1981 г. сменилось на участке работ сильной за.
сухой. Анализ цикличности в биотопических группировках проведен
на материале одного стационара — Ровенского.
Наши наблюдения цикличности включают два значительных от-
резка времени. Первый характеризует изменения цикличности после
массового размножения 1961 г. и до массового размножения 1972 г.
! : Второй.— цикличность, установившуюся после предыдущего массо-
. вого размножения в ^последний год сухой фазы — 1978. Сообщество
, р это время стабильно и представляет собой последний год короткой
сукцессии, развившейся из пионерного сообщества 1973 г., с 1979 г.
начинается нарушение сообщества, подъем численности водяной кры-
сы и ее пик в 1980 ч\ То есть начало новой сукцессии с 1981 г., когда
пионерное 1 сообщество грызунов развивается на остатках предыду-
щего в1 совершенно отличных от 1973 г. условиях. Если 1973 г. был
влажным, то 1981 г. характеризовался сильной засухой, которая
продолжалась и в последующий год. Эти различия должны были
отразиться на цикличности в биотопических группировках и, кроме
того, изменить ход сукцессии, повлиять на ее масштаб, продолжи-
тельность и видовую структуру серий. Поэтому мы рассматривали
биотопическпе группировки отдельно в период с 1966 по 1972 г. и в
период с 1978 по 1982 г.
При расчетах ритмов попадаемости грызунов в канавки замет-
\ ные пики могут проявиться на любой частоте спектра. Экологический
или иной смысл хорошо проявленной цикличности (пика) зависит
от ее периода. Так, нам известны приблизительные сроки беременно-
сти полевбк и выкармливания ими молодых. Обычно*беременность
длится 20 дней, и примерно столько же мать кормит детенышей до
ухода их из гнезда. Стало быть, периодичность в 40—50 сут связана
с выходами молодых, т. е. с изменением численности зверьков. Водя-
ная крыса, например, совершает ежегодные миграции весной с по-
лей на болота, а осенью обратно. От весенней миграции до осенней
проходит около 80—90 сут. Эта цикличность относится к изменениям
численности прй перераспределении зверьков в местообитаниях. Для
полевок известна 2—5- и 7-суточная цикличность, связанная с цир-
кадной организацией их ритмов, территориальным и пищевым по-
ведением. ;
Приведенные примеры позволяют.грубо разделить пики на спек-
трах по их экологическому значению. Низкочастотную цикличность
(30-суточные ритмы и больше) отнесем к циклам численности, а вы-
сокочастотную (от 2 до 15 сут) — подвижности. Средние частоты в
функциональном отношении неясны и могут быть отнесены как к
численности, так и к подвижности. Эти цикличности вносят некото-
рую неопределенность в наш анализ, но, пока не накоплены более
точные данные, с нею приходится мириться.
1 В данном случае пионерное — оставшееся после вспышки численности
водяной крысы. Оно бедно видами и именно с него начинается новая серия
сукцессии.
56
Болото. Биотоп «болото» представляет собой наиболее обыч-
ный, широко распространенный биотоп района исследований. Су-
ществует в нескольких различных вариантах: кочкарниковые зай-
мища в виде обширных низин, поросших тростником; прибрежные
широкие полосы со слабым кочкарником, насыщенные водой, где на
толстом слое мокрого торфа растет только низкая ’зеленая осочка;
низинные болота, разделенные гривами, и солонцы, в мокрые годы
превращающиеся в покрытые водой болота; колкц в низинах, где во-
да подчас не сходит до глубокой осени и хорошо выражен кочкарник.
Линии ловчих цилиндров в годы исследований стояли по кочкарнико- ч
выл болотам, на солонцах, где держалась вода (заболоченные солон-
цы), однако чаще всего канавки закладывались на участках, соот-
ветствующих, зонам перехода от расположенного выше луга к болот-
ной низине пли к небольшому озерку. : ;
В 1966 г. на болоте отлавливали (в зависимости от характера
болота) 4—5 видов грызунов (табл. 8). Из видов, обычных для района
исследований, здесь отсутствовали степная мышовка, джунгарский
хомячок (за все годы наблюдений эти виды не встречались на боло- •
тистых местообитаниях), мышь-малютка, красно-серая полевка и
полевая мышь. Доминировала полевка-экономка. Близки ей по чис-
ленности (обилию) были обыкновенный хомяк, водяная крыса' и -па-
шенная полевка, лесная мышовка. Общая численность грызунов ос-
тавалась низкий.
Землеройки составляли около половины всех отловленных мел-
ких млекопитающих. Численность землероек, как и грызунов, низ-
кая. Численность землероек нарастала к осени. Она максимальна в
конце августа. Грызуны имеют один заметный подъем попадаемости
во второй половине июля.
Группировка грызунов в этом местообитании сохраняла низкую
численность с одним подъемом попадаемости, и это отмечается на
спектре их цикличности (рис. 15) — наиболее низкочастотный пик
соответствует 30-суточпому ритму. В то же время подвижность гры-
зунов была значительной. Высокочастотные составляющие в этом
году на болоте доминируют над всеми прочими. Это прежде всего не-
дельные и 12-суточные, связанные с погодными подвижкайи, а так-
же двухсуточные, обусловленные, видимо, этологическими причина-
ми: обследования территории, защита участков, смены мест кормеж-
ки. Значительное число мощных пиков, особенно в высокочастотной
области, свидетельствует б том, что циклическая система сообщества
в биотопе «болото» в соответствующем году неустойчива. Преоблада-
ли перемещения зверьков по местообитанию, происходило освоение
его —«обживание». Численность же оставалась низкой и к концу
вегетационного периода, по данным отловов, несколько уменьшилась.
В 1967 г. на болотах (основном биотопе Барабы) встречались от
6 до 10 видов грызунов. В этом биотопе многочисленны даже такие
«сухолюбы», как обыкновенный хомяк, полевая мышь, мышь-малют-
ка, узкочерепная полевка. При средне!! численности, когда домини-
рует полевка-экономка, перечисленные виды встречаются довольно
часто. Плотность насекомоядных здесь обычно в 2—3 раза превышает
57
СП 00 Изменение населения мелких млекопитающих биотопа «болото» Таблица 8
— Число видов Грызунов Землероек О —
Год Доминант (субдоминант) грызунов землеро- ек на 100 цил.-сут, ' ' ill 1 ' 1 на 100 цил.-сут % 1 iBcero на : цил.-сут Д 1 i! 1 Подъемы попадаемости в - цилиндры — —
1966 Полевка-экономка (хомяк, водя- ная крыса, мышовка) — 4-5 2,85 57,0 2,15 43,0 5,0 Вторая половина ИЮ- ЛЯ
1967 Полевка-экономка (обыкновенный хомяк, узкочеренная полевка) . 6-10 12,54 зМ 28,10 .69,2 40,65. -- Конец июля ' --
1968 Полевка-экономка (узкочереп- ная, темная полевки) 3—7 4,83 74,3 - 1,67 5,7 6,50 - Начало июля
1969 Узкочерепная полевка (полевка- экономка, полевая мышь, обыкно- венная бурозубка) 5—7 1-4 5,13 69,1 2,30 30,9. 7,43 Конец июня, нача- ло сентября
1970 Полевка-экономка (обыкновен- ная бурозубка, узкочерепная по- левка, малая бурозубка) 5—6 3-5 8,26 49,3 ' 8,58 50,7 16,84' Стабильна _ - ,
1971 Водяная крыса (обыкновенная бурозубка, узкочерепная полевка, полевка-экономка, темная полевка) 6-10 5 13,04 74,1 4,64 25,9 17,68 Конец ' июля, нача- ло сентября
1972 Водяная крыса (обыкновенный хомяк, северная мышовка, обыкно- венная бурозубка) 4-7 2 23,10 95,4 1,11 Г 4,6 24,21 Конец июня, нача- ло августа
В сред- нем Полевка-экономка (водяная кры- са, обыкновенная бурозубка, узко- черепная полевка) 3-10 .<• V "' 3-5 12,78 63,3 7,37 36,7 20,15 Конец июня, июля, начало сентября
Рис. \15.' Спектры ежегодной цикличности численности и подвижности
грызунов биотопа «болото».
Серия сукцессии: I —годы: а — 1966, б — 1967, в — 1968, г — 1969, д — 1970, е — ।
1971, ж — 1972. 11 — годы: а — 1978, 6 — 1979, в — 1983, г — 1981.
плотность грызунов. В 1967 г. доминировали полевка-экономка и
близкие ей по численности узкочерепная полевка и обыкновенный
хомяк. Отсутствовали рыжая полевка и джунгарский хомячок. Чис-
ленность грызунов в течение лета была высокой и стабильной, не-
значительное увеличение наблюдалось в конце июля. Насекомоядные
млекопитающие также имели значительную численность, в 2 раза
выше, чем у грызунов.
Продолжающееся увлажнение местообитаний в Барабе обусло-
вило характер цикличности, общий с -таковым предыдущего года.
Также на спектре ритмов грызунов по мощности доминировала вы-
сокочастотная составляющая (см. рис. 15, I, б). Значительны 5- и-
l'9-суточные-^циклы подвижности. В данном году наблюдается рост
численности и появлялся низкочастотный 120-суточный цикл, крат-
ный 40-суточному ритму выхода новых генераций у полевок. Эта
цикличность обладает заметной мощностью и является второй по ве-
личине на спектре (см. рис. 15).
В 1968 г. отмечено 10 видов грызунов, в разных стациях заре-'
гистрировано от 3 до 7 видов. Доминировали полевка-экономка и уз-
кочерепная полевка. Для последней заболоченные солонцовые луга
и небольшие гривки . между болотами — обычные местообитания.
Третьим по численности видом была темная полевка, а за ней — во-,
дяная крыса. Все прочие виды встречались в ничтожно малом коли-
честве. Интересно, что 5-е и 6-е место по численности занимали два
неболотных вида — мышь-малютка и северная мышовка.
Землеройки в этом году на болоте встречались в небольшом чис-
ле: два небольших подъема численности — в конце июня и августа.
У грызунов был только один подъем попадаемости — в начале июля.
Данный год завершал сухую фазу в Барабе. Сезонная циклич-
ность численности (120 сут) была доминирующей, продолжался рост
численности грызунов в биотопе. Из ритмов подвижности преобла-
дали, как обычно, недельный и двухсуточный (см. рис. 15). На спект-
ре в этом году количество крупных пиков заметно сократилось по
сравнению с 1966 г. Спектр изменился в соответствии с тенденцией,
наметившейся в 1967 г., на нем различимы три области с проявлен-
ными мощными пиками (120-, 7- и 2-суточные).
В 1969 г. в зависимости от расположения канавок было отмече-
•но от 5 до 7 видов грызунов и от 1 до 4 видов землероек. Из грызунов
не встречали темную полевку, из землероек — среднюю бурозубку и
кутору.
Доминировали узкочерепная полевка (доминант года) й полевая
мышь (см. табл. 8). Многочисленными в уловах были мышовка, водя-
ная крыса и мышй-малютка. Все прочие виды отлавливали в еди-
ничных экземплярах. Численность насекомоядных была небольшой,
хотя и немного превышала уровень предыдущего года. Наиболее
часто отлавливали обыкновенную бурозубку. По численности она
уступала лишь доминирующим видам грызунов. Второй по числен-
ности из землероек была малая бурозубка.
У грызунов в течение летнего периода было два подъема подвиж-
ности — в конце июня и начале сентября, причем у доминантов был
60 ;
выражен только июньский подъем. У насекомоядных отмечены два
максимума подвижности — в начале июля и середине августа.
В этом году начался цикл обводненности территории. Однако,
как и в 1968 г., на спектре были те ню три области с доминирующими
пиками (ом. рис. 15). Проявленный 80-суточный цикл свидетельству-
ет о двух подъемах и двух спадах численности, что подтверждается
данными отловов (подъемы в начале июня и в середине июля и спады
в конце июня и конце августа). Наиболее мощный пик в зоне низких
частот 30-суточный, он, вероятно, связан с изменением численности
и подвижности. Самыми мощными оказываются циклы подвижности,
особенно 2-суточные. Спектр в общем характеризует начало измене-
ний картины циклов грызунов в биотопе «болото».
В 1970 г. доминировала полевка-экономка, но так как кочкарни-
ковые болота в районе исследования обычно окружены солонцовыми
лугами (на солонцах и разделяющих займища гривах), то вторым по
значимости доминантом оказывается узкочерепная полевка. Неред-
ко встречаются здесь темная полевка и мышь-малютка. В кочкарни-
ковом болоте обычно присутствует не более 5 видов грызунов, а так-
же туда заходят около 5 видов, обитающих в этом районе. Постоянное
население биотопа: экономка, узко черепная, темная полевки, водя-
ная «крыса, мышь-малютка или полевая мышь.
Численность как аборигенов, так и пришельцев все лето довольно
стабильна в сравнении с другими биотопами. Общее число насе-
комоядных животных близко числу отловленных за летний пе-
риод грызунов. В болоте зарегистрированы 6 _ видов землероек.
Доминирует обыкновенная бурозубка. В порядке убывания числен-
ности идут малая, арктическая и средняя бурозубки. Крупнозубая и
крошечная бурозубки единичны. В каждом местообитании встреча-
лось от 3 до 5 видов землероек.
Характерно, что водяная крыса в этом биотопе ловилась до сере-
дины июля, потом ее‘численность резко спала, но поднималась в
других биотопах.
Динамика попадаемости грызунов в целом в течение лета равно-
мерно высокая, но тем не менее небольшие повышения отмечаются
в начале июля, середине августа и начале сентября. В начале авгу-
ста зарегистрирован спад, обусловленный миграцией водяной кры-
сы. Несмотря на высокую численность грызунов в районе исследо-
вания, на болотах она все же заметно меньше, чем в других биотопах.
Наиболее выражен подъем численности землероек в конце авгус-
та. Это относится к обыкновенной, малой и арктической бурозубкам.
У обыкновенной бурозубки (обычном доминанте среди землероек)
небольшие подъемы создают общую картину изменения численности
насекомоядных за сезон. Отмечены они в начале июня, первых де-
кадах июля и августа. У арктической и средней бурозубок нараста-
ние численности идет с начала августа. ***
Общий рост численности в 1970 г. был на фоне усиливающейся
вегетации в связи с повышением влажности. На спектре циклично-
стей имелись небольшие по мощности, но устойчивые циклы числен-
ности — 120- и 40-суточные (см. рис. 15). Доминировал же 5-суточ-
61
ный цикл, и имелись значительные 2-суточные цикличности. Повы-
шение подвижности животных обусловлено перераспределением тер-
ритории между зверьками разных видов, увеличением потока миг-
рантов. . ! ; ।
В 1971 г. в разных болотах насчитывалось 6—10 видов грызунов.
Полностью отсутствовали рыжая и красно-серая полевки, южная
мышовка и джунгарский хомячок. Два последних вида отсутствовали '>
во всех биотопах района, j Доминировала водяная крыса (доминант
года), вторым по численности видом была узкочерепная полевка, за
нею — полевая мышь, экономка и темная полевки. Характерно, что
в том же порядке убывания численности эти виды следуют во всем "
районе.. Значительное место в отловах занимала мышь-малютка, >
прочие виды были малочисленными.
Видовой состав землероек на болотах в рассматриваемом году был
представлен 5 видами." Не было крошечной бурозубки и куторы, до-
минировала по численности обыкновенная бурозубка. Далее в по- •
рядке убывания численности можно расположить малую, крупно*
зубую,, арктическую и среднюю бурозубок. По численности обыкно-
венная бурозубка уступала только доминанту года — водяной кры-.
се, а малая бурозубка, кроме того,— другим широко распространен- “
ным видам: узкочерепной и темной полевкам, полевке-экономке, по- >
левой мьЕши и мыши-малютке. i
Для грызунов характерны дйа подъема попадаемости в цилинд- s
ры — в середине июля и в конце августа. |Общая численность гры-
зунов несколько превышала уровень 1970 г. У всех землероек на-
блюдался подъем численности в конце июня, у обыкновенной буро-
зубки, кроме того, в конце июля и середине августа, а у средней,
арктической и малой -Н только в середине августа. . , ;
В 1971 Г. наблюдался значительный рост численности водяной '
крысы. На сильно обводненных болотах она практически вытесняла ~
прочие виды. Формированию на болотах моновидовой группировки i
грызунов способствуют как биотические, так и абиотические при-
чины. Как следствие обеднения видового состава, упрощается и
спектр ритмов населения грызунов (см. рис. 15). Он отражает в ос-
новном цикличность растущей популяции водяной крысы. Поэтому
наиболее мощные цикличности находятся в низкочастотной области.
Но существуют небольшие пики в обычных 7- и 2-суточных полосах
частот (см. рис. 15), отражающие цикличность подвижности. Ос-
новной же процесс в биотопе «болото»— изменение численности’
грызунов. .
Одинаковые изменения численности у полйвки-экономки и уз-^
кочерепной полевки. Казалось бы, противоположные по требованиям
к среде зверьки однотипно реагируют на многие факторы среды, а их
Зависимость от болот подтверждает такую связь. Полевка-экономка
живет во влажных местах, избегая открытой воды. В сильно увлаж-
ненные годы увеличивается площадь, занятая открытой водой, и сни-
жается численность зверьков. То же отрицательно воздействует и на
популяцию сухолюбивой узкочерепной полевки. В сухие годы при .
сокращении площадей открытых водоемов создаются дополнительные '
02
благоприятные условия для полевки-экономки и узкочерепной по-
левки.
В 1972 г. началась вспышка массового размножения водяной
крысы. Ее численность достигла максимума, что привело к измене-
нию видового состава грызунов и на болоте. Своеобразие года выра-
зилось в обеднении видового состава на болоте,;здесь отмечалось от
4 до 6'видов грызунов^, и все они имели 1 ничтожную численность.
Редко встречались | обыкновенный хомяк,; лесная мышовка, а побли-
зости От границ болот— узкочерепная полевка. Резко снизилась и
численность -землероек (см. табл.; 8); На болотах зарегистрировано i
только । два вида —• обыкновенная и крупнозубая бурозубки. Все
эти мелкие млекопитающие попадались в виде единичных экземпля- !
ров, практически ж|е в биотопе ловился один вид — водяная крыса!. ’
Спады и подъемы численности обусловлены были только переме-
щениями юсобей; доминирующего вида. Подъемы отмечены в конце
июня и первой половине августа. Наиболее глубокий спад попадае-
мости — в начале сентября (водяная крыса мигрировала с болот, и
они стали малонаселенными). Наибольшая численность насекомояд-
ных была в первой половине лета, после чего они отлавливались в !
ёдиничных экземплярах. > ; *
По данным отловов, на болотах абсолютно преобладала водяная
крыса. Число зверьков в течение всего сезона непрерывно нараста-
ло. Доминирующая цикличность — 80 сут. Она кратна цикличности
появления новых генераций у этого грызуна и в то же время отража-
ет ритм изменения местообитаний — зто срок между весенними и
осенними миграциями зверьков. Остаются заметными по мощности
циклы подвижности, прежде всего 2—3-суточные, ио к ним добавля-
ются 16—20-суточные ритмы (см. рис. 15). Спектр бедей мощными
периодическими составляющими и потерял облик, характерный для
многовидового биотопа «болото», когда мощные ритмы группирова-
лись в трех полосах частот. От обычного сообщества, сформировав-
шегося на болоте после вспышки 1961 г., опять ничего не осталось и
все оно представлено одним видом. Причем популяция водяной кры-
сы, сформировавшаяся на болоте, по энергетическим характеристи- I
кам очень напоминает пионерную группировку. Она наращивает
значительную биомассу,' и нарушается имевшееся здесь соотношение
продукции гетеротрофов и автотрофов в сторону увеличения первых.
Резюмируя описание динамики населения биотопа «болото», сто-,
ит отметить его постоянные элементы.: Доминирует обычно полевка- -
экономка, далее (по вероятности доминирования) следуют.водяная .
крыса, обыкновенная бурозубка и узкочерепная полевка (см. табл. 8).
Сравнительно высокую численность имели полевая мышь, обыкно-
венный.хомяк, северная мышовка, мышь-Малютка. и темная полевка.
Прочие виды обычно малочисленны, и в отдельные годы совсем не
встречаются в этом биотспе (исключая малую бурозубку). На болоте
отмечалось до 11 видов грызунов, все 7 видов землероек. К постоян-
ным обитателям болота из них можно отнести только 5—6 видов гры-
зунов и 3—4 вида землероек. Соотношение численности грызунов и
насекомоядных колеблется в широких пределах, в среднем это со-
63
Рис. 16, Спектры сезонных (/) и многолетних.///) ритмов численности и под-
вижности грызунов биотопа «болото» из разных серий сукцессии Барабы.. ’
I Годы: а — 1966—1972, б — 1978—1981. .
отношение 65 к 35%, но грызуны составляли в отловах от 30 до 95%,
а насекомоядные — от 4 до 69%. Подъем попадаемости обычно при- ,
ходится на конец июня и начало сентября, реже появляется еще один
i подъем — в конце июля.
i Если рассмотреть общий спектр ритмов в период 1966—1972 гг., -
то преобладающими по мощности оказываются низкочастотные рит-
мы с периодом более года! (рис. 16). Хорошо выражены и ритмы под- „
вижности с периодами около 2 и 10 сут. Изменив шаг суммирования, ,
детализируем спектр в полосе от 10 сут до бесконечности (см. рис. 16).:
Можно видеть, что хорошо проявлена цикличность у грызунов в
этом биотопе в 3,3 года, т. е. подавляющий состав населения боло-
та — полевки, ибо именно им характерны циклы численности такой
протяженности. Хорошо выражен 2-летний и годовой ритмы (см.
рис. 16). Причем последний при уточнении оказывается цирканным с
периодом около 1,04 года. Заметны на спектре ритмы размножения
40 сут и хорошо выражены обычные для болотных грызунов ритмы
подвижности — 20-, 13- и 10-суточные.
Перерыв в слежении за сообществом продолжался до 1978 г.
(см. гл. II). Далее исследования проводились уже в пределах сле-
дующей серии экологической сукцессии после вспышки массового
размножения, водяной крысы и разрушения структуры сообщества в
। 1972 г.
За небольшой отрезок времени наблюдений (с 1978 по 1982 г.),
мы оценили основные ритмы грызунов биотопа «болото» из другой
серии экологической сукцессии. Она начиналась при ином фоне абио-'
тических факторов и ином пионерном составе сообщества. Заключи-
тельные годы этой серии 1978—1979.
В 1978 г. доминирующим циклом в сообществе грызунов был 80-
суточный, т. е. численность их имела один минимум и один максимум
за вегетационный период (см. рис. 15). Наиболее проявленные циклы
подвижности: недельные, трех- и двухсуточные заметно уступали по
мощности циклу численности. Среди болотных грызунов абсолютно
преобладала полевка-экономка, а по обсыхающим краям болот в
массе селилась полевая мьйпь, доминирующая в сообществе в этом
году. ' ' ; ,
В 1979 г. (пик увлажненности территории) население болот быст-
ро менялось!. Сократилась численность полевки-экономки, возрос-
ла — водяной крысы. Преобладающей цикличностью стлала 120-
суточная (см,, рис. 15). Заметных циклов очень мало; спектр ритмов
грызунов, рассматриваёмого местообитания имел немного пиков, осо-
бенно большими среди них были недельные и 4-суточные.
В 1980 г., несмотря назначавшееся иссушение болот, продолжа-
лась вспышка массового размножения водяной крысы. Численность
грызунов на болоте резко возросла. На спектре в это время домини-
рует 40-суточный цикл численности и кратный ему 120-суточный.
Хорошо проявлены обычные здесь циклы подвижности- грызунов 5-
и 2-суточные (см. рис. 15). Но вспышка развивалась в иных условиях,
чем в 1972 г., и цикличности сообщества, а также их мощность изме-
нились. Циклы подвижности остались похожими, только несколько
различными по количествуш мощности, циклы численности совершен-
но различались (см. рис. 15).
В 1981 г. сообщество было бедно видами. Наиболее заметным на
болоте был 80-суточный цикл численности и кратный ему 20-суточ-
ный (см. рис. 15). Средней мощностью обладали многочисленные
циклы подвижности — от 2-недельного до 2-суточного (см. рис. 16).
Видовое разнообразие на болоте возрастало, и, видимо, следствием
этого был рост числа циклов в группировке грызунов.
Средние данные о цикличности биотопической группировки за
этот промежуток времени отличаются от 1980 г. Так, доминирующим
по мощности среди сезонных оказывался 80-суточный, а второй по 1
величине — 40-суточный цикл численности. Незначительно по мощ- -
пости в сравнении с ними выглядят циклы подвижности (см. рис. 16).
Рассчитав отдельно низкочастотную полосу спектра, видим, что до-
минирует по мощности цикличность длиной в 1,3 года, мощен 4-лет-
ний ритм численности и мало уступает ему 40-суточная цикличность.
Пз циклов подвижности, какй на предыдущем спектре (см. рис. 16),
наиболее заметна 2-недельная периодическая составляющая. Таким
образом, и низкочастотные ритмы болотной группировки грызунов
изменились со времени предыдущей вспышки массового размножения.
Другими словами, в это время существовало качественно новое сооб-
щество с иными видами цикличности и с другим .числом циклов.
Колок. Этот биотоп объединяет в себе несколько колков различ-
ной разреженности: от очень разреженных насаждений (с прогали-
нами) до густых с обильным подлеском и подрост ом л Канавки в кол-
ках обычно делают в центре, нигде не доходя ближе 5 м к границе
(опушке). Физиономическая картина населения колка описывается
нами по суммарным отловам во всех колках.
А. А. Максимов, Л. Н. Ердаков
65
В 1966 г. в этом биотопе отловлено 11 видов грызунов, хотя обыч-
но там встречается 7—8 видов (табл. 9). Отсутствовали в колке в этом
-году джунгарский хомячок ц рыжая полевка (их не было и в других
биотопах). (Доминировали полевка-экономка, красная полевка и
мышовка; близки к |ним По1 численности узкочерепная и темная по-
; левки. jBce прочие: виды отлавливались в единичных экземплярах.
^Землеройки составляли незначительную часть отловов всех мелких
млекопитающих. ' . ! . ., > '-i 1 '
। Попадаемость в цилиндры в течение сезона была равномерно
низкой, без заметных подъемов. Некоторым спадом отличалось нача-
ло августа, когда зверьков было особенно мало.
Общая численность грызунов,в биотоперколок» мало, изменялась
на протяжении вегетационного (периода. Об этом свидетельствуют
данные отловов и спектр ритмов численности и подвижности зверь-
। ков В рассматриваемом местообитании (рис. 17). Цикл изменений
численности (период 60, сут) на спектре один из самых маломощных.
! Наиболее: выражены перемещения животных внутри колков, домп-~
нирует 2-суточная цикличность, а та!кже циклы подвижности, навс-,
(денные, видимо, погодными факторами с пери'одичностью 17 и 5 сут.
В колках из 1967 г. обычными были 10 видов грызунов, т. е. все
х виды грызунов района, исключая южную, мышовку. Доминировал
абориген колка — красная полевка. Близки к ней по частоте попа-, '
даемости в канавки были полевая мышь и полевка-экономка.. Общее
число отловленных землероек разных видов превышало суммарное
количество отловленных грызунов1 (
Подъем численности населения колков отмечен во второй полови-
не июля, причем он характерен 'как для грызунов, так и для пасе-;
комоядных. Численность всех мелких млекопитающих в течение лета
сохранялась высокой. Интересно, что лесные полевки в сумме не со-
ставляют и трети от численности всех прочих грызунов колка.
Заметно изменяется набор периодических составляющих числен-
ности и подвижности в 1967 rj. Численность животных быстро растет
и ее цикличность Хорошо проявляется в виде мощного 80-суточного
пика на спектре (см. рис. 17). Доминирует в этом году кратный цик-
лу численности 20-суточный ритм. Вполне возможно, что он синхро-
нней ритму температуры воздуха (cmj гл. I). Заметную мощность име-
ют и циклы подвижности грызунов, в порядке убывания мощности
сти они 12,7-. 4- и 2,5-суточные. Эти цикличности, кроме 2,5-суточ-
ных, соизмеримы обычным для регион^, ритмам температуры воздуха
и цикличности осадков. , , , ; । '
В 1968 г. л колках присутствовали ?—10(вйдов грызунов. Доми-
нирующее положение занимала группа лесных полевок. Абсолютно
преобладала красная полевка, далее в порядке! убывания численно- ..
сти — рыжая и красно-серая. Достаточно многочисленными были
темная полевка и полевка-экономка. В разреженном колке и по пе-
риферии их заметную (численность имела ' узкочерепная полевка.
Нередкой была также северная мышовка. Численность землероек
была крайне низкой. Подъем ее наблюдался в конце августа — на-
66
Таблица 9
Распределение численности населения млекопитающих биотопа «колок»
Год L Доминант (субдоминант) Число видов Грызуны Землеройки Всего на 100 цйл.-сут Подъемы попадаемости в :- цилиндры .... ’ Л
грызунов • землеро- elf 1 на 100 . ци^д.-сут 1 "%' на 100 дил.-рут
1966 . - S Полевка-экономка — (мышовка, красная полевка)^ и ? 5,14 79,4 "1,33 20,6 6,47 __ — Спад в начаЛе августа
1967_ Красная полевка (полевая мышь, полевка-экономка) _ - 10 - ? 14,20 44,6 17,66 55,4 / 31?86 “ Копециюля
1968 ; Красная полевка (рыжая, красно- серая, темная полевки, полевка-эко- номка) ~ 9-10 ? 5,19 82,9 1,07 17,1 6,26 - июня, начало августа
1969 Красная полевка (обыкновенная бурозубка, рыжая, красно-серая полевки)’ “ 11 2-3 13,27 71,4 5,30 28,6 "18,57 " Середина июня, нача- ло августа • —
1970 Красная .полевка (обыкновенная бурозубка, полевка-экономка, мы- шовка, полевая мышь) - 10 4-5 12,27 41,9 17JJ ^58,1 29,41 Конец июня, начало августа
1971 Водяная крыса (обыкновенная бурозубка, полевая мышь, малая бу- розубка, красная полевка) — 10 . 1 14,60 42,6 19,80 57,4 ’ 34,40 Начало "сентября
1972 Водяная крыса (обыкновенная бурозубка, мышовка, крупнозубая бурозубка) 7 4 17'53 83,3 3,51 16,7 21,04 - Конец августа
В сред- нем Красная полевка (обыкновенная бурозубка, полевка-экономка, по- левая мышь) - 8—10 4—5 13,84 55,3 11,18 44,7’ 25,02 _ Конец июня, начало сентября. Спад в нача- ле августа
I I I I I I III!! I I I
°° 4824 12 8 4. 3 2 0=4824 12 8 4 3£j
Puc. 1/’.Спектры ежегодной цикличности численности и подвижности
. ! - грызунов биотопа «колок».
Уел. обозн. см. рис. 15.
«шра/nvnd в «иди ou-суточпого пика малой мощности. Наиболее
проявлена периодичность передвижений 3, 5-суточпая, 2-недельная
и 24-суточная. Последние два вида цикличности скорее всего связаны
с периодичностью температуры и влажности (соизмеримы с ними по
длине периодов). Спектр ритмов этого года содержит много мощных
пиков (см. рис. 17), т. е. цикличность жизнедеятельности грызунов
(ее проявления) возрастает в сравнении с предыдущими годами. По
данным отловов, в этом году разнообразнее становится и видовой
состав грызунов. 4 i: г - ' 1
В 1969 г. в колках отмечены все виды грызунов, кроме южйой
мышовки и джунгарского хомячка. По сравнению с 1968 г. население
колка имело большее видовое разнообразие и численность особей.
Среди грызунов доминировали красная полевка, рыжая и красно-
серая полевки. Двум последним по численности не уступала и лес-
ная мышовка. Много ,было полевых мышей и полевок-экономок.
Нужно отметить, что отсутствовавшие в колке грызуны были мало-
численными и в других биотопах. Общая численность землероек была
выше, чем в 1968 г. (5 видов). Среди них доминировала обыкновенная
бурозубка. По численности она превосходила доминирующую среди
грызунов красную полевку. Второй по обилию вид землероек — ма-
лая бурозубка. Крупноэубая, арктическая и средняя бурозубки
единичны, крошечная бурозубка и кутора не зарегистрированы. ;
В разных колках за сезон встречались 2—3 вида землероек.
У всех мелких млекопитающих отмечено нарастание численности
к осени, примерно с середины августа. Небольшой подъем ее наблю-
дается и в середине июня, за исключением отдельных малочисленных
видов бурозубок, которые появляются в уловах только в конце лета.
Рост численности животных сопровождается более проявленной
цикличностью. Так, в 1969 г. в колке за вегетационный период чис-
ленность заметно возросла и на спектре цикличности жизнедеятель-
ности (попадаемости) грызунов этого местообитания доминирует пик
с периодом 120 сут, кратный 40-суточной периодичности появления в i
местообитаниях новых генераций полевок (см. рис. 17, а). В этот год
началось повышение влажности в регионе, с чем, вероятно, связан
второй по мощности пик на спектре. Во всяком случае, по своему зна-
чению он соизмерим с цикличностью влажности. Хорошо проявлены
в этом году обычные для колка циклы подвижности грызунов дли-
ной в 7,5 и 2 сут. Видовой состав населения;грызунов колка несколь-
ко сократился, и, видимо, это повлияло на вид спектра ритмов жиз-
недеятельности — число значительных по мощности пиков стало
меньше, чем в 1968 г. ; -
В 1970 г. в порядке доминирования население грызунов было
представлено следующими видами: крйсная полевка, полевка-
экономка, лесная мышовка, полевая мышь, узкочерепная полевка.
Как всегда зарегистрировано около 10 видов грызунов. Отмечается ;
i
69
следующая закономерность: чем более разреженный колок, тем боль- '
1 шая роль в нем доминанта региона; чем колок гуще, тем заметнее
' преобладание аборигенов колка. Так, в этом году в разреженном кол-
ке доминировали полевка-экономка и узкочерепная полевка, тогда
как в (соседнем густом —..красная полевка и лесная мышовка. Отно-
сительная численность риелки» млекопитающих в'колке более высо-
' кая в сравнении с прочими биотопами; района.1 Г .
। Всего в этом биото!пе зарегистрировано 6 видов землероек^(в раз-
। i ных колках от 4 до 5, видов), Отсутствовала только кутора. Домини-
; .ровала: обыкновенная бурозубка, численность которой более чем в
‘5 раз превышала таковую красной полевки —Доминанта среди гры-
; зунов! Далее землеройкй пр убыванию: численности располагаются в
। следующем порядке: средняя,' малая, ;арктичбская, крупнозубая и
крошечная бурозубки. ./I ' Ь j J 1
За летний-сезон у грызунов наблюдалось два. подъема попадаемо-
•сти в цилиндры — в конце июня и конце августа, а у насекомояд- •
ных — три подъема с пиками численности — в середине июня, июля;
, I и начале августа. По общему числу попаданий за лето землеройки
! превосходили грызунов. 1 : । i
i |Болыпим числом (видов и Отсутствием выраженных доминантой
| для сообщества грызунов । биотопа «колок» характерен 1970' г.
;(см. табл;. 9)."Численность всех видов, населяющих колок, росла, хотя
. и очень медленно. На Спектре цикличности численности грызунов;
этого года особенно много мощйых пиков |(см. рис. 17). Заметен пик
ритма численности 48-суточный^ он значителен по мощности, хотя
ш несравним с доминирующими. Доминируют циклы подвижности
I 7-, 5-, 4-, 2,5- и 2-суточные. Эти ритмы сравнимы с периодичностями
н основных абиотических (климатических) факторов и, видимо, отра-
жают как обычные перемещения грызунов на своих территориях, так
. и их погодные Подвижки. u , i '
В 1971 г. в колке обитало около 10 видов-грызунов (см. табл. 9)л...
Не было видов,, не (Отмененных в ртом году в Барабе вообще;—
Ц южной мышовки и: джунгарского хомячка'. Население колКа, как
; ' обычно, многочисленно по количеству1 видов и особей. Землеройки
Н по суммарному количеству не уступают грызунам. В каждом из коЯ-
I ков зарегистрировано по 6 видов землербек. Общее (максимальное)
число видов для; региона 7, т. е. встречаются все виды, свойственные
, лесостепной'подзоне Западной Сибири. . 1 | j ।
Среди грызунов в полке доминирует водяйая крыса, среди насе-
комоядных — обыкновенная бурозубка, по численности превышаю-
| ;щая водяную крысу в 3 раза. Среди всего населения колка она — до-
минирующий вид. Далее в порядке убывания численности следуют-
полевая мышь; малая бурозубка, красная и темная^ полевки, средняя
бурозубка. У прочих вйдов численность значительно ниже. Рыжие:
। лесные полевки превосходят по количеству особей любой вид грызу-
нов, но значительно уступают обыкновенной бурозубке. Вновь под-
тверждается, что доминант района (среди грызунов) многочислен не
только в своем, но и в любом биотопе [Сообщество..., 1978].' >
Усилилась общая обводненность; территории, и в 1971 г. начался
подъем численности водяной крысы. Видовое разнообразие в колках
70 Г
заметно снизилось в сравнении с 1970 г. Все это отразилось на цик-
лах численности и подвижности грызунов и, как следствие, на кон-
фигурации спектра (см. рис. 17). Значительный рост численности
наблюдался только у Нодяной крысы, которая, доминируя в колках,
тем самым повышала общую численность грызунов. На спектре до-
минирует пик, соответствующий 1;20-суточной цикличности.
Очень невел!ики । по мощности i пики подвижности животных 17|-,
3- и 2,5-суточные ,(см. рис. 17). По цикличности уловов в канавки
этот год резко отличается от всеХ| предыдущих. ! I
В 1972 г .(была вспышка численности водяной крысы. В разре-
женных колках отмечено 7 видов грызунов, тогда как в густых-3—
4 вида. Второй iпо численности вид — северная, мышовка встреча-
лась в 10—15' раз реже доминанта (в’одяной крысы). Все остальные
виды грызунов отмечены единично. Насекомоядных было мало, хо-
тя по численности среди всех мелких млекопитающих обыкновенная
бурозубка занимала второе место после водяной крысы, а численность
крупнозубой1 бурозубки была близка таковой северной мышовки.
Общее число видов землероек в биотопе 4. В разных колках зареги-
стрировано по 3 видащтих животных. I ।
Динамика попадаемости грызунов определялась только водяной
крысой, поэтому наблюдался один подъем численности во второй по-
ловине августа. Именно в это время водяная крыса окапывается у
границ колков. У землероек два подъема численности: наиболее за-
метный во второй половине июня, второй — в начале августа, при-
чем численность обыкновенной бурозубки во второй половине лета
держалась примерно на одном уровне. . ।
В текущем году не отмечены вовсе такие аборигены колка, как
рыжая и красно-серая полевки. Красная полевка отловлена ,в тече-
ние сезона в 2 экз. , '
В этот год, год вспышки массового размножения водяной крысы,
на спектре доминирует пик, соответствующий 80-суточной циклич- ,
пости изменения численности грызунов. Изменение численности в
1972 г.— самый мощный процесс в биотопической группировке жи-
вотных, причем численность росла yiодного вида, а все остальные
либо совершенно ^отсутствовали в уловах, либо единичны. Так что
спектр циклов населения колков в этот год представлял, ио сути,
спектр ритмов численности водяной крысы. По виду он схож с прош-
логодним, но лучше проявлены циклы подвижности. Естественно,.что
быстрый рост плотности популяции ведет к стрессу животных, тер-
риториальным конфликтам, т. е. цикличность их передвижений по
территории-становитсЯ| хорошо выраженной, и преимущество имеют
ритмы высокой частоты. Так и на спектре наиболее мощные из ритмов
подвижности — 2,5—2-суточные. Имеются и ;незначительные пики
погодных цикличностей передвижений водяных крыс с периодами в
13 и 5 сут (см. рис. 17). ' - ।
Таким образом, в колке из грызунов обычно доминирует красная
полевка, либо полевка-экономка, либо полевая мышь (последняя
значительно реже). Обыкновенная бурозубка часто доминант не
только среди землероек, Но и среди всего животного населения
I
71
Рис. 18. Спектры; сезонных/.-!) и многолетних (ZJ) ритмов численности пподвпж-
: ности грызунов биотопа «колок».
Усл. обозн. см. рис. 16. ,
бпотопа «колок». В колках отмечается 8—10 видов грызунов и 4—
'7 видов землероек. Как правило, грызуны и насекомоядные в сумме
близки по числу отловленных особей. Каждые три года землеройки
оказываются многочисленнее, чем грызуны,.а в промежутках их
численность может быть очень низкой. Биотоп «колок»— один из на-
иболее населенных биотопов района исследований.
ДпнамикА попадаемости в ловушки в колке своеобразна: подъемы
чаще всего приходятся на конец июня и начало сентября, спад —на
начало августа, ) ,
Оценивая общий вид спектра цикличности численности и подвиж-
ности грызунов в биотопической группировке «колок» за 1966 —
1972 гг., можно заметить, щто на нем доминируют низкочастотные
циклы (рис. 18). Основные процессы, происходившие в этой серии
сукцессии,— рост численности и формирование ее цикличности. Из
ритмов подвижности заметной мощностью обладают 20-, 4- и. 2-су-;
точные, ©вязанные в основном с цикличностью температуры и влаж-
ности. Рассматривая отдельно низкочастотный спектр цикличности
уловов в этой биотопической группировке, можно заметить, что до-
минирует здесь 9, 8-летний цикл, связанный, видимо, с циклично-
стью обводнения территории. Хорошо проявлен и по мощности бли-
зок к доминантному цирканный ритм численности грызунов, причем
его период несколько меньше годового (около 0,85 года). Существует
среди доминирующих 2,5-годовая цикличность (см. рис. 18).
Из ритмов подвижности наиболее заметен 15-суточный, скорее
всего связанный с цикличностью влажности, и 25-суточный, завися-
щий, видимо, от температурного ритма, (см. гл. I) характерного для.
Барабы.
i
72
Кризис териокомплекса в колке, наступивший в 1972 г. и привед-
ший к тому,'что среди грызунов там осталась только водяная крыса,
начал собой новую серию сукцессии в этом биотопе. Она развивалась
при пониженной влажности, й за 6 лет здесь сформировалась груп-
пировка грызунов, заметно отличающаяся от аналогичного года
(1969) предыдущей серии экологической сукцессии.
Набор цикличностей псшадаёмости животных в канавки также
был своеобразен в 1978 г. Наиболее выраженный процесс в биотопе
«колок»— рост численности грызунов. Качественно группировка
сильно изменилась. Среди аборигенов- прошлой’серии всегда доми- .
пировала красная полевка и значительную численность имела крас-
но-серая полевка. В этой же серии постоянный доминант — рыжая
полевка, а красно-серая в уловах отсутствует. Наиболее выраженная
цикличность 120-суточная. Из ритмов подвижности (все они мало-
мощны) .выделяются недельный и 2-суточный (см. рис. 17). Недель-
ный синхронен обычному здесь температурному циклу, а 2-суточиый
связан, видимо, с передвижениями грызунов по территории.
В наборе цикличностей грызунов «колка» в 1979 г. видимйх из-
менений нет (см. рис. 17). Остается доминирующей 120-суточная не--
риодичность изменения численности, и на спектре; заметны пики 5-
и 3-суточных изменений подвижности. Сообщество в этом году, как и в
1977 г., отличалось максимальным разнообразием видов (есть даже
такие редкие, как джунгарский хомячок), и цикличности их популя-j
ций синхронны. Последнее и обеспечивает небольшое число пиков на
спектре.
В 1980 г. продолжалось массовое размножение водяной крысы.
Цикличность ее размножения, как и полевок, примерно 40-суточная^
и этот пик появляется на спектре. Доминирование водяной крысы в
колках привело к перераспределению в них численности грызунов,
массовым передвижениям зверьков, потере синхронности в! их цик-
личности. Доминирующими на спектре • становятся ритмы подвиж-
ности с периодами 14 и 9 сут (см. рис. 17). Число пиков на спектре
увеличивается, короткие 3- и 2,5-суточные периодичности подвижно-
сти становятся более проявленными.
В регионе идет нарастание засухи, и йспышка численности водя-
ной крысы в 1981 г. прекращается. Численность ее в колках резко
снижается, но не замечено рорта плотности популяций других гры- .
зунов. Население колка бедно видами. > Наиболее проявлен в это
время 27-суточный ритм;, близкийпо длине периода температурному,
циклу. Мощные пики отражают цикличность подвижности грызунов :
в это время, но численность их не возрастает. С уходом водяной кры-
сы из колков в середине лета численность зверьков становится вовсе
незначительной. 1
Рассматривая в средней цикличность грызунов в' биотопической
группировке «колок», хорошо видны отличия в ее цикличности по
сравнению с аналогичным сроком в предыдущей серии. Прежде все- ,
го, основная цикличность в колке 240-суточная (см. рис. 18). Все
прочие значительно уступают ей по мощности. Аналогов такой цик-
личности нам не известно, и для объяснения ее функциональности
I
73
пока нет данных. Судя по спектру, основные виды цикличности со-
I общества грызунов колка превышают по длине периода 240-суточ-
ную. Изменив шаг суммирования, получим спектр низких частот для
I этого сообщества (см. рис. 18). Цикличность 240-суточная (0,8 года)
на нем действительно по. мощности незначительна. Основную дис-
персию! включает периодичность длиной 3,3 года. Многими авторами
циклы такой;длины описаны как1 характерные для грызунов (Аве-
; тисян;'197О; Starka,.' Gepasimov, 1971]. Существует мнение, что эти ’
циклы у_грызунов синхронны; с ритмами вегетации растений [Tast,
! Kalela, 19^1 1j Моркио отметить и цирканную составляющую циклич-
ности населения колков, она невелика по мощности и ее период не
точно соответствует годовому, а равен 1,2 дода.
Сравнивая средние наборы цикличностей в сообществе биотопа
«коЛок» в двух разных сериях,сукцессии, можно заметить их сильное
различие (см. рис. 17). Пионерный состав в колке сильно изменялся
после каждой вспышки численности водяной крысы, и, кроме того,
! годы экологической серии сукцессии! 1973—1978 пришлись на сухую
;фазу. Совокупность различий привела к тому, что низкочастотные
1 циклы во,второй из рассмотренных серий сократились, в количестве, .
из трех остался только 3,3-летний («полевочий») цикл. Стал маловы-
раженным доминантный в сообществе цирканный ритм,1 и его период
значительно отклонился от годового. ii
( , Таким образом, в цикличности жизнедеятельности экологической
группировки трызунов колка от серии к серии сукцессии наблюдайт-
ся изменения. Значительные различия имеются в количестве циклов,
шх, мощности и соотношении. Ежегодно картина цикличности в сооб-i
ществе меняется. Обычно в первые годы каждой серии и в периоды
нарушений в сообществе наблюдается большое число мощных пиков’
во многих полосах частот. Со стабилизацией сообщества, приближе-
нием его к климаксовой стадии (т. е., годы с максимальным видовым*
разнообразием) возрастает согласованность цикличностей его попу-
ляций, и число, пиков на спектре ритмов численности уменьшается.
Поле. Биотоп «поле» объединяет в себе три возможных варианта
поля с различными культурами. Обычно они были заняты пшеницей,1
i овсом или кукурузой. Ловчие борозды закладывали на середине по-
лей’ и, они ежегодно работали во все фенофазы (весенняя па-
хота, всходы, развитие растений, 'уборка урожая, стерня, । осей-’
няя вспашка). Таким образом, динамика попадаемости мелких мле- '
I копитающих в этом биотопе отражает летне-осенний цикл биоцено-
за !«поле». ' г; 1 1 * i.
. --j Ловчие цилиндры вкапывали на полях, когда высота выходов до-
стигала 5—8iCM. Почва в зто Кремя остается свежевспаханной с на-
рушенным почвенным слоем,, Дернина только начинает образовывать-
ся. Тем не менее с самого начала установки цилиндров постоянно, ,
отлавливались Зверьки 6—7 iвидов. . I -
В 1966 г, численность зверьков была невелика, видовое.разно-
образие — 6—7 видов. Доминировали полевая мышь, обыкновенный
хомяк, полевка-экономка. Нередкими были лесная мышовка и во-
।
дяная крыса. Насекомоядные млекопитающие 2 составляли незначи-
тельную долю от всего количества отловленных животных (13,4%),
и распределение их во времени (динамика попадаемости в цилиндры)
было довольно равномерным. . . j , •
Плотностй популяций грызунов к осени йовышалась. До середи-
ны лета в рассматриваемом биотопе было отловлено только три вида
грызунов! полевая мышь, । обыкновенный ।хомяк и лесная мышовка.
К концу лета рисло видов увеличилось., Таким- образом, ; осенний'
подъем попадаемости млекопитающих объясняется не только ростом
плотности популяций,! но и увеличением числа популяций? В связи
со своеобразными условиями это1*о биотоца' последнее- соображение ।
справедливо и для всех последующих лйт. Развитие посевов от весны
к осени неуклонно повышает привлекательность пол!я для все боль-
шего числа особей различных популяций. * 1 1
Численность грызунов на полях в_.1966 г. возросла и к концу ве-
гетационного периода почти удвоилась. На спектре ритмов грызунов
этой группировки имеется мощный 120-суточный цикл изменения i
численности (рис. 19). Доминируют ритмы подвижности. Это прежде
всего 20-суточный, синхронный температурному ритму в Барабе,
и 5-суточный,jсинхронный ритм-у влажности (cmi гл. I). Кроме того,
имеется и 2, 2-суточная цикличность, связанная, скорее всего, с тер-
риториальными перемещениями зверьков. Мощных гармонических
составляющих на спектре немного.
На полях,в 1967 г. регистрировались 6—7 видов грызунов, но
из них только 3—4 имели, достаточно высокую численность. Прочие
отлавливались единично. Доминировали полевка-экономка, узко-
черепная полевка, полевая мышь. Преобладание полевки-экономки
в этом не свойственном для нее биотопе объясняется тем, что она бы-
ла самой многочисленной среди грызунов района. Общая высокая чис-
ленность вида может быть причиной доминирования даже в малопри-
годных для него местообитаниях. Узкочерепная полевка в этом году
также имела в районе исследований высокую численность. Полевая
мышь — обычный обитатель полей, мышь-малютку встречали только
однажды. Обыкновенный хомяк и водяная крыса обычны и в равной
мере отмечены в отловах на полях. , ;
; Число отловленных за сезон насекомоядных почти в ,2 раза превы-
шало число грызунов (136 против 82). Обыкновенная бурозубка от-
личалась высокой относительной ^численностью । среди всех насеко-
моядных (51,1—76(4% по данным отловов 1969—1972 гг.) и в 1967 г.
4 могла быть дбминирующим видом среди всех обитателей рассмат-
риваемого биотопа. । •
Количество грызунов и насекомоядных постепенно уменьшается
к осени. Небольшой подъем попадаемости грызунов отмечен в кон-
це июля. Для насекомоядных ; животных, характерны два незначи-
тельных подъема третья декада июня и июля.
В 1967 г. число животных возросло. Увеличилась мощность
120-суточной цикличности (см. рис. 19), и появляется пик около
2 Видовая принадлежность землероек в 1966—1968 гг. не устанавливалась,
i
75
Рис. 19. Спектры ежегодной цикличности ^численности и подвижности грызу-
нов биотопа «поле»,
Усл. обозн. см. рис. 15,
40-суточной периодичности численности. Доминируют по-прежнему
циклы подвижности, особенно велика мощность циклов, синхрон-
ных с изменением погодных факторов — 20-, 8- и 5-суточных. Сухая
фаза приближается, к концу, и начинает возрастать обводненность
территории. Наблюдается и рост численности. Перегруппировки
грызунов на полях происходят всегда независимо от фазы увлаж-
ненности и серии сукцессии, .Поля летом осваиваются грызунами,
и на спектрах их цикличности всегда есть > 2-суточные ритмы.
Как и прежде, в 1968“г. на полях отлавливались! зверьки 6—7
видов. В! конце июня наблюдалсц первый1 летний подъем численно-
сти (отловов), а в конце- августа — второй. Обращает на себя вни-
мание крайне малая численность насекомоядных животных, они
попадались на полях в единичных экземплярах в течение всего' лет-
него сезона. В то же время численность грызунов немного уступает
уровню 1967 г. В биотопе доминировали узкочерепная полевка,
северная! мышовка, обыкновенный хомяк. Нередко в отловах отме-
чался джунгарский хомячок. Другие виды встречались редко. В пер-
вой декаде августа попадаемость зверьков в цилиндры заметно сни-
жается. При невысокой общей численности отмечено большое число ।
значимых корреляций между популяциями видов. ;
Ежегодно в группировке грызунов биотопа «поле» прослежива-
ется 120-суточный пик. Таково примерное время между распашка-
ми поля — весенней и осенней. Имеется этот пик и на спектре рит-
мов 1968 г. По мощности он уступает многим высокочастотным цик-
лам. Хорошо выражены и многочисленны периодичности подвижно-
сти животных. Есть месячная ритмичность, мало уступают ей в
мощности ритмы, связанные с погодными факторами. Доминируют
ритмы 2-суточной подвижности грызунов. Большое число циклов
в этом году обусловлено целым рядом факторов, в том числе и тем,
что это год. переходный от сухой фазы климата к влажной. Возрас-
тают численность и видовое разнообразие животных.
Число зарегистрированных видов на ролях в 1969 г. возросло
до 11. Доминировали полевая мышь, узкбчерепная полевка. Часто
встречались представители таких видов, как мышь-малютка и лес-
ная мышовка. Прочие виды грызунов отмечены редко, а обычные
для поля полевка-экономка и обыкновенный хомяк в этом году со-
вершенно отсутствовали. Необычна в районе исследований попада- ;
емость значительного числа южной мышовки, которая в регионе
встречается, как правило, редко.
В течение летнего сезона численность грызунов плавно возра-
стала. Более заметный ее подъем отмечен в начале сентября. Как
обычно, др середины лета Зверьков на полях почти не было, и толь-
ко со второй половины лета попадаемость грызунов в канавки стала
заметно расти. !
Численность насекомоядных животных в рассматриваемом! био-
топе мала. Среди них доминировала обыкновенная бурозубка. Еди-
нично отлавливались крупнозубая, малая и крошечная бурозубки.
В первой половине лета землеройки на полях практически отсут-
77
г
ствовали. Их появление здесь, как и для грызунов, отмечается со ;
второй половины лета — первой декады августа.
| В 1969 г. видовое разнообразие грызунов на поле оставалось вы- f
i соким, но особого роста численности не наблюдалось. Видимо,1 по-
этому 60-суточный1 пик изменений' численности очень небольшой. <
h ”Н; На спектре циклов этого года преобладают по мощности пики рпт-
1 !мов подвижности (cmj рис., 19)< Они так же, как и в прошлом году,''
обусловлены в основном переходом, климата региона от сухой фазы ,
к влажною Доминирует на спектре 3,4-суточная цикличность) под- )'
вижности (см. рис. 19), [обусловленная передвижениями грызунов \
) л !по гнездовым территориям) проходом мигрантов из других место-
! обйтаний. Количество1 циклов в этом году заметно меньше, чем в
I 1968 г., видимо, возрастает синхронность жизнедеятельности у раз- .<
пых видов- грызунов. , - 1
Как уже отмечалось, биотоп «поле» описывается нами как некий ?
средний показатель нескольких конкретных полей. Население био-
' топа может оказаться весьма разнообразным в зависимости от про-
израстающих культур и от того, [какими биотопами данное поле <
. окружено. Естественно, что поле, расположенное среди колков, бу- и
дет иметь население, отлйчное от поля на границе с болотом.
I i ' | Три поля, обследованные нами в 1970 г., имели сообщества гры-
1 зунов, состоящие соответственно - из। 2, 5 и 10 видов. Основной со-
став сообщества биотопа в порядке доминирования был следующий:
полевая мышь, i лесная ।мышовка, узкочерепная полевка, полевка-
экономка (если поле окружено болотами) и мышь-малютка. Эти ви-
ды отмечались !на полях ежегодно. В начале августа появлялась ,
. водяная крыса. Красно-серая, рыжая полевки) а также обыкновен-
I- ный хомяк в этом году отсутствовали. I
Население испытывало два подъема численности — в конце июля
и в начале сентября. Это связано как! с рождаемостью и миграция- 1 ;
ми, так и, с фенологией грызунов. Кроме того, на: свежевспаханном
: роле грызуны обычно не поселяются. ! ।
1 ; Численность землероек в биотопе «Йоле» в 1970 г. была достаток- <
но высокой, но не (меньше, чем у грызунов. Среди насекомоядных
(доминировала обыкновенная бурозубка и была многочисленнее до-
минирующей среди грызунов полевой мыши. Таким образом, обык-
новенная бурозубка в текущем году была доминантой среди всех ? *'
мелких млекопитающих биотопа‘«поле». Второй по численности вид '
среди землероек — малая бурозубка. Менее распространена аркти-
ческая бурозубка, крупнозубая и средняя— отлавливались редко.
В) первой половине лета (июнь) землеройки в полях крайне ред- '-
ки. Количество их возрастает в июле, когда поля приобретают вид
«закрытых биотопов» [Юдин, 1962], т. е. высота посевов увеличивает
их защитные свойства. Наиболее заметны подъемы численности зем-
лероек здесь во второй декаде июля и первой декаде августа. К осени
i численность землероек по&тепенно снижалась. В этом году возросла
увлажненность полей, снизилось видовое разнообразие, но увеличи-
лась численность грызунов. В соответствии с этим доминировала
- цикличность численности с периодом 80 сут, большой мощностью
обладала и месячная цикличность (см. рис. 19).
78 - .
Следующей по мощности была 2-суточная ритмика подвижности.
Рост численности, вызывает повышение передвижек грызунов, свя-
занных с территориальностью. Погодные же передвижения в 1970 г.
были небольшими. На спектре имеется пик, близкий по периоду к
цикличности влажности ;'(4 сут) и к температурной цикличности
(8 сут). Количество циклов, уменьшилось, видимо, потому, что.син-
хронность функционирования популяций сохранилась., к этой, од-
ной из последних лет серии, а число видов, несколько сокра-
тилось. - , । i 1, ' ~~ ; - у
В 1971 г. на-"полях ।встречалось в среднем 9—10 видов грызунов
(на некоторых1 полях до И). Отсутствовали только виды, не отме-
ченные в этом году совсем,— южная мышовка и джунгарский хомя-
чок. Кроме грызунов,! на|полях отлавливали все виды землероек,^
встречающиеся, в северной лесостепи,— 6 видов представителей ро-
да Sorex и кутора. Среди грызунов доминировали полевая мышь,
мы’шь-малютка,1 а также узкочерепная полевка и водяная крыса.
Нередко в отловах была и темная полевка. Прочие виды встречались
единично. J ; i , ,
Насекомоядные занимали примерно четвертую часть всех отлов-
ленных за сезон в цилиндры мелких млекопитающих (57 экз. из
230 отловленных зверьков). По общей численности за сезон иссле-
дований, насекомоядных меньше, чем в 1970 г., но численность их
достаточно высока. Среди землероек доминировала обыкновенная
бурозубка. По числу .попаданий в ловушки в течение сезона она
близка к домйнантам среди грызунов — полевой мыши и мыши-
малютке. Меньше попадали в канавки средняя и малая бурозубки.
Арктической, крупнозубой, крошечной бурозубок и куторы в этом
году поймано несколько экземпляров. ,
Плотность грызунов все лето оставалась довольно равномерной
и1 высокой, но к концу его отмечен'значительный подъем числа осо-
бей. Небольшой подъем подвижности грызунов отмечался также и
в конце июня, В первой половине лета редки мышовка, лесные по-
левки, темная полевка, мышь-малютка и обыкновенный хомяк.
К концу лета численность снизилась только*у узкочерепной полев-
ки. Увеличение-числа особей землероек заметно было с третьей* де^
кады июля и продолжалось до середины августа, после чего числен- I
ность Снизилась. Это прежде всего касается обыкновенной бурозуб-
ки. Для менее многочисленных, чем обыкновенная, средней и малой
бурозубок!, наибольшая подвижность наблюдалась в конце авгу-
ста — начале сентября,. । I
Подъем,численности водяной крысы начался; в 19711 ,,i*. В это1 же
время продолжалось дальнейшее обводнение территории. Сильный,
рост численности грызунов на полях обусловил хорошо проявлен-
ную цикличность численности.; Доминирует характерная для гры-
зунов «поля» 120-суточная цикличность (см. рис. 19). Почти такой
же большой мощностью обладает и лунная цикличность у зверьков
(~ 30-суточная). Велика и синхронная с циклами влажпбсти 5-су-
точная периодичность подвижности. В 1971 г. особенно часты пере-
движения грызунов, связанные с погодными условиями, в частно-
7Э
сти с ритмом изменения влажности. Как обычно, хорошо проявлены
2-суточные циклы подвижности.
В связи со вспышкой численности водяной крысы в 1972 г. ви-
довой состав мелких млекопитающих во всех биотопах был значи-
тельно обеднен. Так, совершенно отсутствовали рыжая, темная,
узкочерепная полевки и полевка-экономка, которые юбычны для
этого биотопа. Доминирующая на полях водяная крыса по । числен-
ности превосходила сумму особей популяций всех прочих видов гры-
, зунов этого биотопа более чем в 3 раза. За ’ней по численности сле-
довали полевая мышь, обыкновенная и крупнозубая бурозубки,
: i северная мышовка, еще меньше численность красной полевки, мыши-
малютки и малой бурозубки.
Коль скоро динамика численности зверьков в биотопе в значи-
тельной степени зависит- от доминанта, то на полях наблюдалось
увеличение числа зверьков в начале августа (водяная крыса мигри-
ровала на поля). На это же время приходился и максимум попада-
. |вмости землероек. ’ -
В; 1972, г. спектр ритмов грызунов в биотопе «поле» еще более
упростилсй. Заметной численностью (13,09 особи на 100 цил.-сут.)
! обладала только водяная крыса. Наиболее выражена характерная
' для животных в этом биотопе посуточная цикличность. Заметны
циклы подвижности, связанные с изменением влажности— 5- и
17-суточные, а также температурные 7-суточные. Впервые за время
наших наблюдений совершенно не проявлены 2-суточные ритмы
, подвижности, обычно характерные для поля.1
! Подводя итог рассмотрения динамики популяций грызунов и
насекомоядных в биотопе «поле», мощно отметить, что обычными
доминантами здесь являлись полевая мышь и обыкновенная буро-
зубка. Большую численность популяций имели узкочерепная полев-
ка, лесная мышовка, полевка-экономка (в случаях, когда поля при-
мыкают к болоту) й водяная крыса. Число видов грызунов, отмечен-
ных на поле,— 7—8, хотя значительную плотность всегда имеют
3—4 вида. В отдельные годы на полях встречащтся все 7 видов зем-
лероек, свойственных району исследований. Наиболее распростра-
нена обыкновенная бурозубка, реже достаточно высокую плотность
популяций имеет малая бурозубка, а в редкие годы и кутора. На
полях; можно встретить и всех грызунов, свойственных району ис-
следований.
Численность мелких млекопитающих колеблется в значительных
пределах (та!бл. 10). Насекомоядные в среднем составляют четвер-
тую часть всех отловов на поле. Выявлена интересная тенденция, по
которой, несмотря на преобразование сообщества животных биотопа
«поля», каждые два года численность насекомоядных приближается
к половине всех отловов (1967, 1970;и 1973 гг.). Увеличение попада-
емости зверьков в цилиндры приходится обычно на конец июня
(вселение грызунов на зазеленевшее поле), конец июля (выход мо-
лодых и их расселение у грызунов и насекомоядных) и начало сен-
тября (последний помет, начало устройства на зиму, последние ми-
грации).
. 80
Таблица 10
А, А. Максимов, Л. H. Ердаков
Распределение численности населения млекопитающих биотопа «поле»
Год - Доминант (содоминант) Числа видов Грызуны Землеройки Всего на 100 цил.- сут Подъемы попадаемости в цилиндры
грызунов землеро- ; ек на 100 цил.-сут % на 100 цил.-сут %
1966 . ... 1» Полевая мышь (обыкновенный хомяк, полевка-экономка) 6-7 ? 3,88 87,3 0,56 J.2,7 4,44 [Конец„июля и авгу- ста '
1967 [Полевка-экономка(узкочереп- ная полевка, полевая мышь) 7-8 ? 8,37 37,7 13,87 62,3 22,24 - Конец июля
1968 Узкочерепная полевка (южная мышовка, обыкновенный хомяк) 6-7 ? 5,48 90,2 0,60 9,8 6,08 KoHeij июля и авгу- ста
1969 Полевая мышь (узкочсрспная полевка, мышь-малютка, обыкно- венная бурозубка) - 10-11 1—4 5,11 78,7 1,38 '21,3 6,49 - Начало сентября
1970 Полевая мышь (северная мьппов- вка, узкочерепная полевка, обык- новенная бурозубка, полевка- экономка) 9-11 3-4 10,57 53,1 9,34 46,9 1 — 19,91 Конец июля, начало сентября
1971 Полевая мышь (мышь-малютка, обыкновенная бурозубка, узкоче- репная полевка, водяная крыса) — 9-10 5-6 20,22 76,1 6,37 23,9 36,59 Конец июля, начало сентября
1972 Водяная крыса (полевая мышь, обыкновенная бурозубка) 5-6 2—3 14,95 89,8 1,70 10,2 16,65 . Начало августа
В сред- нем Полевая мышь _ (узкочерепная - полевка, мышь-малютка, полевка- экономка, обыкновенная бурозуб- ка) 7-8 3-4 11,57 67,3 5,64 32,7 17,21 . - Конец июня, июля, начало сентября
Рис. 20. Спектры сезонных и многолетних ритмов численности и подвижности,.
। . si грызунов биотопа «поле». , , [ •,
, 11 Усл. обозн. см. рис. 16. ' г J 11
Рассматривая цикличность населения полей на многолетнем ма-
териале всей этТин серии, можно видеть (рис.1 20), что большой мощ-
ностью обладает только ; годовая цикличность. Все более высокие л
частоты не содержат на1 спектре заметных пиков. Увеличив шаг, ?
рассчитали низкочастотную часть. Наиболее выражен по мощности
9,8-летний цикл, видимо синхронизируемый цикличностью обвод-
нения региона. Очень невелик по мощности и плохо выра-е
жен цирканный пик (см. рис. 20), отражающий цикличность чуть ;
больше годовой — 1,1 года. Еще менее выражены, чем годовой/
ритмы подвижности зверьков — месячные и' ,2-недельные. Таково
распределение цикличности в серии 1966—1972 |гг., ;
Заключительные годы следующей серий прослежены с 1978 г.
Это последний год сухой фазы. Спектр циклов грызунов биотопа «по-,
; ле» изобилует мощными пиками и йапоминает поэтому спектр «шу-
ма» (см. рис. 19). Видимо, в этом году численность и подвижность
грызунов изменялись хаотично, и флуктуации ее носили случайный
характер. В такие годы|, наблюдая изменения попадаемости грызу-
нов,-легко прийти к выводам X. Гарольда и В. Е. Говарда [Harold,
Howard, 1981] р [том,'что периодов! в численности и подвижности
। грызунов, нет, и все эти флуктуации полевок совершенно случайны.
• Нарушения обычной цикличности [ в популяциях iнаблюдаются на
репрессии численности. В1 это время уловы в канавки снижаются и
получаемый временной ряд становится'[менее полным. ,
В 1979 г. численность! грызунов заметно возросла в сравнении
с предыдущим годом. Началось обводнение территории — при по-
вышении влажности Бараба «оживает*». Циклы животных стали бо-
лее выраженными (см. рис. 19). Наибольшей мощностью обладает
в этом году 80-суточная цикличность численности. Второй по мощ-
ности 17-суточный цикл подвижности, синхронный с изменениями
влажности в регионе, и почти столь же значителен 20-суточный
82
ритм, видимо, наведенный температурными циклами. Из маломощ-
ных также проявлен 7-суточный температурный и 5-суточный ритм
влажности. Хорошо выражена территориальная подвижность, ее i
2-суточная цикличность имеет значительный пик на спектре (см.
рис.. 19).. ! iL ' I । । ip > .
Вспьнйца численности водяной крысы, .'начавшаяся в 1980 г., по-
влияла на цикличность грызунов: в биотопической группировке «по-
ле»1. Почти ^половину (47,5%) грызунов .в уловах составляла водя-
ная'крыса. .Наиболее проявленной цикличностью численности б,ыла
8-месячная.1 Остальные циклы связаны с подвижностью животных.।
Из них 26,6- и 7,7-суточные синхронны температурным ритмам, хо-
рошо проявлен ритм территориальных передвижений грызунов .
2-суточный (см. рис. 19). Спектр этой вспышки численности водяной ।
крысы вполне своеобразен и не похож на вспышку 1972 г., когда во- ।
дяная крыса составляла в уловах 87,3% от зверьков в биотопе «поле». >
Начавшаяся в 1981 г. засуха сильно повлияла не только на во-
дяную крысу, но и на все сообщества грызунов на полях. Числен-
ность их здесь снизилась в 3 раза по сравнению с 1980 г. Скудность
отловов повлияла и на оценки цикличности. Как и в 1978 г., спектр ;
их имеет множество высокочастотных |пиков (см. рис. 19), хотя на I
нем остается 240-суточная цикличность. Доминирующей становится ,
2-суточная ритмичность .перемещений । по территории и 10-суточная
ритмичность,, сходная по периоду с цикличностью осадков. Была
особенно хорошо проявлена синхронность подвижности и осадков.
В среднем за рассмотренные годы этой серии сукцессии домини-
рующей была 2-летняя цикличность численности. Но в отличие от
предыдущей серии были хорошо проявлены циклы, связанные с по-
годными изменениями подвижности, с территориальными переме-
щениями грызунов (см. рис. 20, а). Рассчитав низкочастотные циклы
сообщества «поле» в.этой серии, мы получили все тот же доминирую-
щий 2-летний цикл. Цирканная цикличность проявлена мало, дли-
на периода несколько больше года — 1,02 года. Значительную мощ-
ность имели 40-суточная цикличность численности и' 17-суточная —
подвижности (см. рис. 20, б). Последняя связана, видимо, с циклом
влажности в Барабе. Рассмотренные характерные для этой серии ви- 1
ды цикличности в биотопической группировке грызунов хорошо от-
личимы от совокупности, циклов в предыдущей серии. Снова при-
ходится признать, что действие абиотических факторов имеет еже-
годное своеобразий и,, кроме того, каждый год меняется экологиче-
ская структура!, на которую действуют эти факторы. Отсюда наблю-
даемые ежегодные особенности цикличности и значительные разли-
чия в сериях сукцессии.
Луг. Ловчие канавки заложены на Лугах, которые располагают-
ся либо на слабо возвышенной равнине между колками, либо около
озер. В понижениях вокруг них обычно располагаются болота с коч-
ками и без открытой воды. Увлажнение лугов грунтовое неустойки-.
вое, проективное покрытие составляет 60—90%, задернованность —
15—24%. Почвы — луговой солончак, при обилии воды многие лу-
га оказываются залитыми водой в течение почти всего летнего се-
6*
83
зона. На лугах нередки солонцы и солончаки, только между кол-
ками на возвышениях луга без солонцов, с хорошей задернован-
ностью и разнообразием травянистой растительности.
В 1966 г. население грызунов было представлено 6—7 видами
(табл. И). Не встречены лесные полевки и джунгарский хомячок.
Доминировала по частоте попадаемости в ловчие цилиндры полевка- -
экономка (она же доминировала в этом году по. сумме отловов за
весь сезон во всех биотопах). Значительную плотность (часть отло-
вов) имели обыкновенный хомяк и узкочерепная полевка. Прочие
грызуны были отловлены в единичных экземплярах. Численность
зверьков в регионе низка, изменение (динамика) попадаемости гры- С
зунов в канавки носило «синусоидальный характер», т. е. начиная
:! с первых чисел июня (каждую вторую декаду) отмечен подъем по-
падаемости, и так до 10 сентября. Число пойманных землероек по-
вышалось к jосени. ' '
! Восстановление экологической группировки грызунов в 1966 г.
на лугах после очередной вспышки проходило в условиях сухой фа-
зы. Поэтому темпы его были невелики. Виды малочисленны (6,3
особи на 100 цил.-сут), видовое разнообразие мало (7). Увеличение
грызунов на лугу незначительно к концу вегетационного периода.
Наиболее значительный по мощности цикл — 3,5 сут (рис. 21, а).
Видимо, это ритм перераспределений, связанных с территориаль-
ностью, Второй по величине пик подвижности зависим от влажно-
сти, он совпадает пощлине периода (5 сут) с ритмом увлажнения.
Имеется заметный по мощности 40-суточный цикл изменения чис-
ленности. Количество мощных пиков на спектре велико, что свя-
зано с общей малой численностью зверьков.
На лугах в 4967 г. отсутствовали лесные полевки и отмечалось
8—10 видов грызунов, причем, хотя и доминировала полевка-эко-
номка, но на солонцеватом и разнотравном лугу между колками
преобладала по плотности'узкочерепная полевка, а полевка-эконом-
ка имела большую численность в сумме во;всех вариантах луга. Во-
обще численность грызунов в этом году была высокой, а такие виды,
как темная полевка, узкочерепная полевка, обыкновенный хомяк
и мышь-малютка, были сравнимы по числу отловленных особей с
доминантом. На разнотравном лугу в значительном числе появля-
лась водяная крыса. Плотность популяций грызунов на разнотрав- ’
ном лугу была выше, чем на прочих вариантах луга. Насекомоядные
по количеству многократно превосходили грырунов.
Относительная численность (число особей на 100 цил.-сут) пой-
' । манных на лугу зверьков колебалась в незначительных пределах,
хотя можно отметить два ее подъема — в конце июня и августа.
Эту' картину колебаний дают грызуны. Насекомоядные по числен-
ности плавно убывали к концу лета.
Спектр цикличности в 1967 г. содержит меньше, мощных пиков.
Совершенно отсутствует на нем цикл изменения численности (см.
рис. 21,.а). Заметны и устойчивы ритмы подвижности, и, судя по .
длине периодов, это передвижения, связанные с цикличностью по-
годы. Наиболее мощный ритм синхронен 20-суточной температур-
84
Таблица И
Распределение численности населения млекопитающих биотопа «луг»
Год Доминант (содоминант) Число видов Грызуны Землеройки Всего на 100 цил.-сут Подъемы попадаемости в цилиндры
грызунов землеро- ек j 1 На 100 цил.-сут % н а 100 цил.-сут 1 %
1966 Полевка-экономка (обыкновенный хомяк, узкочерепная полевка) 6-7 ? 3,19 71,7 0,97 28,3 4';i6 Начало июня, середи- на июля --
1967 Полевка-экономка .(узкочерепная полевка, темная полевка, лесная мы- шовка) 8-10 ? 19,42 38,4 32)40 61,6 51,82 Конец июня, конец ав- густа
1968 Узкочерепная полевка (полевка- ,экономка) 6-7 ? 4,77 75,8 1,52 24,2 6,29 Начало июня и сен- тября
1969 Лесная мышовка (обыкновенная бурозубка, узкочерепная полевка, полевка-экономка) 7-8 4-5 5,19 61,4 3,28 38,6 ' 8,47 _ - Конец июня, начало сентября -
1970 Полевка-экономка (узкочерепная полевка, водяная крыса, обыкновен- ная бурозубка) 7-9 4-5 13,39 55,2 10,89 44,8 24,28" Конец июня, начало августа
1971 Узкочерепная полевка (водяная крыса, обыкновенная бурозубка) 6-8 3—5 15,8 76,4 * 6,97 23,6 22,77 Конец июня, начало сентября
1972 Водяная крыса (обыкновенная бу- розубка) 3-4 3 23,8 87,0 3,57 13,0 27,37 Конец июня, начало сентября
В сред- нем Полевка-экономка (узкочерепная полевка, обыкновенная бурозубка, водяная крыса) 6-8 3-5 12,2 66,6 8,87 33,4 ‘ 20,74 ' Конец июня, начало сентября
Рис. 21. Спектры ежегодной цикличности численности и подвижности
I грызунов биотопа «луг».
! Усл. обозн. см. рис, 15.
ной цикличности, второй по величине также близок к недельному
температурному циклу, 17- и 5-суточные ритмы подвижности, про-
явленные на спектрах, соизмеримы по длине с циклами влажности
в Барабе. Перемещения, связанные с этологическими причинами —
территориальностью зверьков, имеют' соответствующие пики на
спектре циклов 2,5- и 2-суточных, правда; очень небольшой мощ-
ности. I ; ! , ; ( ,
В 1968 г. на лугу отмечено 6—7 видовгрызуновПолностью от-,
сутствовали рыжая полевка и южная мышовка, последней не было1
ни в одном из биотопов. Доминирует узкочерепная полевка, близки
к ней по плотности популяции полевки-экономки и обыкновенного
хомяка. Нередки были темная полевка и лесная мышовка. Прочие |
виды встречались редко. 1 । ।
Насекомоядных поймано меньше, чем узкочерепных полевок,
что необычно для луга. По сравнению с 1967 г. их численность рез-
ко упала. Подъемы попадаемости зверьков приходились на начало
июля и сентября^ । ;
Общее снижение численности грызунов на лугу в 1968 г. не от-
ражается существенно на их видовом разнообразии. В' этом году
население луга имеет хорошо проявленную 120-суточную циклич-'
ность численности (см. рис. 21, а). Из пиков подвижности преобла-
дают два — оба отражают цикличность, связанную с погодными ус-,
ловиями. Это 7-суточный ритм, соизмеримый с температурным, И '
10-суточный, соизмеримый с цикличностью осадков. Остальные ви-
ды цикличности в этом году у биотопической группировки грызунов
на лугу ^проявлены незначительно.
Численность мелких млекопитающих в 1969 г. оставалась низ-
кой. Полностью! отсутствовали на лугу обыкновенный хомяк, юж-
ная мышовка, рыжая и красно-серая полевки. Доминировали се-:
верная мышовка, узкочерепная полевка и полевка-экономка. До-
статочно часто отлавливались мышь-малютка, полевая мышь, тем-
ная и красная полевки. Таким образом, на лугу были обычными 7—
8 видов грызунов. । j, , 1 । h’
Численность землероек в этЬм году на лугах Незначительно пре-
вышала уровень 1968 г. При общей малой численности мелких мле-
копитающих на лугу обыкновенная |буро|3убка;— наиболее распро-
страненный вид (почти вдвое превышала численность; доминантов
среди грызунов). Единично .встречались арктическая, средняя,
малая и крошечная бурозубки., В разных вариантах луга за летний ,
сезон встречались 4—5 видов землероек. г q ;
Для динамики численности грызунов в этом биотопе характерны
подъемы попадаемости в канавки в и(оне и начале сентября, а также
глубокий спад ее к концу июля. Интересно участие доминануов
в динамике плотности населения грызунов. Среди них лесная мышов-
ка многочисленна к осени, а полевки (узкочерепная и экономка) —
в начале лета. Они и дают два подъема активности у населения луга.
Насекомоядные имеют только один подъем, активности в августе, при-
чем рост: попадаемости в канавки начинается в начале этого месяца,
а максимум численности отмечается в конце.
87
В 1969 г. начинается новый цикл обводнения территории, что
влияет па население грызунов луга. Численность зверьков невели-
ка, нет хорошо выраженного доминирования какого-либо вида (см.
рис. 21). Последнее свидетельствует о том, что сообщество стабилич
зирбвано. Доминирующая цикличность в> этом году (48-суточная),
видимо, отражает ритм численности. Среди ритмов! подвижности
преобладают 17- и 10-суточные, связанные с цикличностью темпе-
ратуры и осадков.-Имеется на спектре и 3,5-оуточный ритм передви-
жений грызунов. Общее; количество мощных пиков на спектре не-
велико, вероятно, изменения в популяциях происходят довольно
синхронно. н. ।
. В 1970 г. биотоп «луг» объединял три характерных луга района,
исследований: солонцовый, разнотравный между колками и разно-
травный на берегу, озера. На лугу недалеко от берега озера абсолют-
но преобладала । полевка-экономка. Ее попадаемость много выше,
чем ।у прочих членов сообщества мелких млекопитающих. Высока
;здесь была и плотность насекомоядных, отмечено 7 видов грызунов;
и 4 вида .землероек. На солонцовом лугу преобладали насекомояд-
ные (4 вида), обыкновенная и арктическая, средняя и малая буро-
зубки. Из грызунов в порядке убывания: полевка-экономка, водя-
ная крыса,, лесная мышовка, узкочерепная >полевка в-мышь-малют-
ка. На разнотравном лугу между колками также преобладали на-
секомоядные (5 видов землероек, в порядке уменьшения численно-
сти: обыкновенная, срёдняя, арктическая, малая и крошечная бу-
розубки). Из грызунов присутствовали полевка-экономка, узко-
черепная полевка, полевая мышь и мышь-малютка.
Луга населены большим количеством грызунов, но на них отсут-
ствовали обыкновенный хомяк, джунгарский хомячок, а южная мы-
шовка и красно-серая полевка отловлены по одному экземпляру.
Насекомоядных почти столько же, сколько грызунов. Доминиро-
вала обыкновенная бурозубка, которая в 1,5 раза превышала чис-
ленность доминанта .среди грызунов — полевую мышь. Далее в по-
рядке убывания численности шли малая, арктическая и средняя
бурозубки. За весь период отловлено только две крошечные буро-
зубки.
Подъемы попадаемости на лугу у грызунов и землероек отмече-
ны дважды в” течение лета: в июне и первой половине августа. Оба/
подъема легко объяснимы циклами размножения; второй подъем^1
имеющий большую продолжительность,— результат еще и мигра-
ций грызунов и землероек.
В 1970 г. наблюдалось увеличение численности, !чему способство-
вало общее увлажнение территории. Цикличность численности в
этом году была небольшой по мощности, длина ее периода — 120 сут
(см. рис. 21, а). Доминировал ритм подвижности, синхронный с од-
ним из циклов влажности — 5-суточным. Хорошо проявленной бы-
ла 20-суточная ритмичность подвижности. В этом году в сообществе
грызунов луга вновь отсутствовали высокочастотные ритмы.
В 1971 г. в луговых биотопах не отмечено 5 видов грызунов, два
из которых отсутствовали в районе работ — южная мышовка и
88
джунгарский хомячок. Обыкновенный хомяк, рыжая и красно-се-
рая полевки обитали в других биотопах. Таким образом, на лугах
встречено б—8 видов. Доминировали узкочерепная полевка, водя-
ная крыса, обычной была темная полевка, редкими — красная по-
левка и северная мышовка.. Численность землероек была намного
ниже, чем в 1971 г., и в суммарных отловах эти зверьки заметно ус-
тупали грызунам. Среди землероек доминировала обыкновенная
бурозубка, кстораях'по численности значительно отставала от узко-
черепной полевки и, водяной крысы. Снизилась .численность малой
бурозубки, арктическая, средняя и крошечная —были редки. В раз-
ных вариантах луга встречались от 3 до 5 видов землероек.
За сезон отмечено два подъема попадаемости зверьков в канав-
ки: в конце июня и августа. Отмечалось снигйёние попаддрмости в!
первой половине июля.
В 1971 г. появляются и даже становятся доминирующими по мощ-
ности высокочастотные циклы (см. рис. 21). Дальнейшее увеличение
увлажненности территории, начинающийся подъем численности во- ’
дяной крысы способствовали уменьшению стабильности сообщества.
Об этом свидетельствуют снижение видового разнообразия и уве-
личение числа мольных пиков на спектре цикличности сообщества.
Рост численности обусловливает заметное проявление цикличности
численности, на спектре есть очень мощный пик 120-суточной пери-
одичности. Преобладание водяной крысы приводит к увеличению
мощности 5-суточной цикличности, синхронной 5-суточным ритмам
увлажненности Барабы (см. гл. I). В связи с появлением на лугу
мощной популяции водяной крысы происходят массовые террито-
риальные перегруппировки; грызунов (ритмы 2- и 2,2-суточные)
и вытеснение частп видов из этого биотопа (уменьшение видового
разнообразия).
Вспышка-численности водяной крысы в 1972 г. сильно изменила
как количество, так и соотношение популяций мелких млекопитаю-
щих на лугах. Грызунов отмечено всего 2—3 вида, насекомоядных —
3 вида. Доминировала водяная крыса. Второй по численности была
обыкновенная бурозубка, далее следуют крупнозубая и малая бу-
розубки, и единично встречающиеся на лугах noneiBKa-экономка п
лесная мышовка: Численность грызунов за счет массового вида
была высокой. В 1972 г. водяная крыса составлял^ около 86% от
всех видов грызунов биотопа «луг». Стабильность была резко нару-1
шена, видовое разнообразие снижено до крайних пределов. Произо-
шло нарушение биотопической группировки «луг», после чего .нача-
лось восстановление сообщества, т. е. началась новая серия сукцес-
сии. Спектр ритмов, рассчитанный для грызунов «луга» (см. рис. 21,
а), представляет собой набор цикличностей всего одной популяции —
водяной крысы. Наиболее мощный у нее в 1972 г. ритм изменений
численности с периодом 48 сут. Хорошо проявлены циклы террито-
риальных перераспределений, связанных с разделом площади лу-
га. Это 2,5—2-суточные цикличности большей мощности (см. рис. 21,
а). Выражены на спектре ритмов пики погодных подвижек (5- и
89
7-суточные), но в сравнении с ритмами численности и территорйаль-
1 ности они незначительны по мощности. Г
Таким образом, на лугах обычно вылавливали 7—9 видов грызу-
1; нов , и 3—5 видов насекомоядных. Среди грызунов доминировала
либо полевка-экономка, либо узкочерепная полевка, среди земле-
i в । роек — обыкновенная^бурозубка, нередко,по численности превыша-
тощая доминантовсредйгрызунов. В .годы, вспышки размножения
водяной крысьпойа доминировала, а видовой состав млекопитающих
j । ’значительно обеднялся. Необходимо; отметить, что среди встречаю-
щихся на «лугу»; грызунов постоянно высокую численность имели
1 ; только три из них, из насекомоядных два —, обыкновенная и малая
(бурозубки. Дроме названных видов доминантов достаточно часто в '
। этом биотопё .встречалась темная полевка. Насекомоядные в среднем
j за- все годы исследований составляли примерно четверть всех отлов-
। ленных! млекопитающих. К статике луга можно отнести также не- - '
; которое ! постоянство; подъемов попадаемости, зверьков в летне-осен-
i ний период. । • 1 •1 | 1 • i
. i 1 Численность и соотношение попадаемости грызунов й землероек
на «лугу».заметно варьировали (см. табл. ,14). Вариации эти можно ;
• i объяснить|анализируя природную и метеорологическую обстанов-
ку района,.но; достаточно полный анализ возможен только с привле-
чением биотической обстановки (отношения между популяциями и
внутренние ресурсы популяций). 1 . । . н j
Рассмотренная । серия экологической сукцессии позволяет отме-
тить,’что разные ее годы отличаются основными видами циклично-
сти. Причина этого заключена в изменении природно-экологической
I обстановки в биотопе «луг». Средние данные о ритмах в этой группи-
ровке показывают, что основные по мощности периодические про-
цессы имеют низкую частоту., из высокочастотных заметны циклы ;
48-суточный численности и 3,4- и 2,1-суточные подвижности. Наибо-
лее же выражен на «лугу» 9,8-летний цикл (рис. 22, а), связанный (
Рис. 22. Спектры сезонных и многолетних ритмов численности и подвижности.^
грызунов биотопа «луг».
Усл. обозн. см. рис. 16.
i ' si. t
90
с фазами обводненности территории Барабы, второй по мощности —
2-летний. Цирканного ритма изменений жизнедеятельности в этой
экологической группировке и в этой серии сукцессии не проявлено'
Следующая серия рассмотрена нами, как обычно, с лет, пред- , ।
шествующих вспышке численности водяной крысы. Сообщество в
ЭТО время Сформировано И, УСТОЙЧИВО. j | I ' ij
'В 1978 г. заканчивалась’ фазаi!иссушения территории^ числен-
ность грызунов была средней (16—17!особей'на 100 цил,’-сут)3видовое ,
разнообразие высокое.’ На” спектре можно выделить' три области,
в которых распределены'особенно мощные цикличности (см. рис.^21,'
б). По мощности доминируют "ритмы территориальных । перераспре- >
делений грызунов на «лугу», периодичность их 2—3 сут. Вторая
группа мощных1 пиков, показывает цикличность1 грызунов, синхрон- . |
ную с «малыми» циклами температуры и влажности .в Барабе — 7- и
5-суточная, периодичность. В третьей группе сосредоточены пики i
цикличности размножения|80-_и 40-суточные^, а также цик 20-суточ- ;
ного ритма подвйжности; также синхронный с аналогичным по дли-
не .периода'температурным. р , । I 1 ! ।
В 1979 г1 количество мощных пикой на спектре заметно снижа-
ется (см. рис. 21, б). Рост численности зве1рьков обусловливает за-r I
метную мощность пика численности с периодом 120 сут. Увеличение
количества зверьков на «лугу» ведет к территориальным перегруп- j
пировкам, й следующая по мощности цикличность — 2,3- и 2-суточ-
ные ритмы подвижности. Заметны на спектре 7-суточный ритм по-
движности,- связанный с температурным циклом, а также 10-суточ-
иый, обусловленный । периодичностью осадков. Сообщество грызу-,
нов стабильно, численность водяной крысы еще сравнима с чис-
ленностью других грызунов. <
При вспышке численности водяной крысы в 1980 г. сообщество
грызунов оставалось достаточно многочисленным, водяная крыса
составила около 50% общей численности по лугу. Спектр циклич-
ности грызунов в этом году беден ритмами. Доминирует цикличность ।
с периодом около месяца (см. рис.| ,21, б); Выражена 2-недельная и J
3-суточная ритмичность подвижности. Обычных ритмов витом году’
связанных сщогодными и территориальными передвижениями, зверь- I
ков, не отмечается. Отсутствуют и циклы изменений численности,
хотя количество водяных крыс К|0сенИ'на лугу нарастает. Возмож- '
но, такая необычная картина развития вспышки численности на h i
«лугу» и перестройка сообщества^вязаны с начавшейся засухой.
Луга выгорают, травостой развивается не в полной мере. Растения,
не достигнув ц’половины своего размера, ^начинают цвести. Обычная
цикличность в сообществе нарушена. । 1
Засуха продолжалась и даже усилилась в 1981 г. Наблюдалась
депрессия численности всех видов грызунов. С середины лета резко ’
снизилась численность водяной крысы. Луговое сообщество грызу- ,
нов стало малочисленным, состоящим в основном из водяной кры-
сы, численность ее к осени также снизилась. Вспышка численности
водяной крысы не достигла своего обычного размаха, а популяции-
прочих* грызунов биотопа «луг» при спаде численности водяной кры-
сы не вышли из-состояния депрессии. । 1
91
Спектр цикличности сообщества содержит много мощных пиков
(см. рис. 21, б), видимо, это следствие «шума» из-за малой числен- ,
ности и преобладания нулей во временном ряду. Сообщество, не-
: смотря на слабую вспышку численности водяной крысы, оказыва-
; ется разрушенным как вспышкой, так и сильной 2-летней засухой.
Начинается новая серия сукцессии на лугу.
1 Таким образом, для периода с 1979 по 1981 г. характерны! 120-су-
точная цикличность численности и' ритмы/подвижности, синхрон-
ные цикличности! погодных факторов (см. рис. 22, I, б). Обычно в
это время у грызунов проявлена и 2-суточная цикличность" подвиж-
ности. Из низкочастотных ритмов численности по мощности преоб-
: ладают 3-летние, ,а среди ритмов подвижности наиболее проявлены
; !20-суточные (см. рис. 22, II,б). Спектры сезонных и многолетних,
ритмов содержат пики большого количества почтй одинаково, мощ-
ных ритмов, что свидетельствует о их нестабильности в этой серии
! сукцессии. Видимо,; дестабилизирующее влияние на биотопическую*
группировку оказала сильная двухлетняя засуха, прервавшая да-
же вспышку массового размножения водяной крысы. \
1 Опушечная полоса. Эти пограничные местообитания,'между кол-
ком и лугом, колком и полем, колком и болотом выделены нами ус-;...
ловно. Часто население этого биотопа разнообразно и многочислен-
! но. Здесь, как правило, !ловятся все виды зверьков, обитающих в
районе исследования. В разных вариантах биотопа были установ-
-лены четыре канавки, сборы из которых дали нам многолетнюю кар- ;
тину динамики млекопитающих.
В 1966 г. здесь совершенно отсутствовали все лесные рыжие по-
левки ц джунгарский хомячок. Полевка в этом году была редка да-
же/в колках, а джунгарский хомячок повсеместно отсутствовал. До- ,
минировали полевая мышь, лесная мышовка и- полевка-экономка.
Плотность популяций была низкой, отмечено 8 видов грызунов. На-
- секомоядные составляли менее четвертой части отловов всех мелких
млекопитающих.
В течение лета у грызунов и землероек было два подъема числен-
ности — в начале июня и сРнтября. ;
Численность грызунов в опушечных полосах в 1966 г. была низ-»
; кой при незначительном видовом разнообразии. Доминировали три
। вида цикличности. Наибольшей мощностью обладала 2-суточная*
. цикличность территориальных перемещений грызунов (рис. 23,1 I,
а). Следующими по мощности были циклы подвижности,, связанные
с погодными факторами,— 6—8-суточные. Значительным был пик
48-суточной ритмичности численности.
В 1967 г. повсёместно на опушечных полосах отлавливали 9—11
видов грызунов. Как правило, преобладали полевка-экономка, по-
левая мышь и часто встречалась мышь-малютка. Кроме того, пять '
обитающих здесь видов (обыкновенный хомяк, темная, красная, ..
узкочерепная полевки и водяная крыса) имели значительную и
примерно одинаковую плотность, популяций. Плотность землероек
была высокой, они составляли более половины всех уловов. Дина-
мика их попадаемости без заметных подъемов и спадов. То же самое
отмечено и для грызунов.
92
co 4824 12'8 <7 J 20*4824 12 Ь 4 о
Рис. 23, Спектры ежегодной цикличности численности и подвижности
грызунов биотопа «экотон».
Усл. обозн. см. рис. 15.
В 1967 г. заметно изменяются основные циклы (см. рис. 23, I, а).
Доминирующим становится 24-суточный ритм, синхронный преоб-
ладающему температурному циклу. Остальные' периодические со-,'
ставляющие, заметно «уступают (ему по (мощности и приблизительно'
' равны между собой!* 'Среди* них несколько выделяются 10-суточный, \
связанный, s видимо, с осадками, all также высокочастотные j 4- и 2-&
s суточные щйклы,. имеющие этологическую, природу. Совершенно от-
сутствуют цикличности (численности, грызунов, хотя численность их
возросла в сравнении/с 1966 г. почти*в 3 раза, i । i , К ?
। ,В 1968 г. в экотоне на границе!колка и поля, колка и выгона
присутствовали, все!'виды грызунов, обитавшие в районе исследова-
йий в этом сезоне: 11 и 9 видов соответственно! Землеройки состав^-,
ляли около пятой части всех отловленных мелких млекопитающих:
Доминанты были выражены очень слабо^ выделялась тоЛько уз-
|КОчерепная полёвка, i|Общий порядок доминирования следующий:
узкочерепн^я! полевка, северная мышовка. - । у
: Можно отметить три подъема! попадаемости у грызунов в течение
сезона —I в начале июня, июля и сентября, а также глубокий спад
в конце июля. У насекомоядных наблюдалось некоторое увеличение
числа особей ю сентябрю. < 1 ,
(И 'В этом году-ф экотоне наблюдалось заметное падение плотности
грызунов, хотя видовое* разнообразие продолжало возрастать. На-
ибольшей мощностью в это время обладают цикличности! подвижно-
сти, свйЗанные1 с погодой. .Доминирует 5-суточный ритм, близкий
циклу влажности-воздуха. Большая мощность и у 24-суточного,
I^синхронного, вероятно, температурной цикличности (см. рис. 23, в).
В этом году в экотоне проявлен ритм численности. Он 2-месячный,
такая периодичность была описана для нескольких видов грызунов,
причем этот ритм присущ как активности, так и многим физиологи-
ческим функциям (даже такому экстерьерному показателю, как
масса тела) [Morosovsky е. j;a.,| 1980].
•Число видов, отмеченных в экотоне в 1969 г., было большим^
, Так, на границе колок — по!ле, колокД- выгон было, как 1й в 1968 г,
: 11 и 9 видов соответственно. В первом случае доминировала полевая
мыщь, во втором — узкочерепная полевка. Отсутствовала только
।южная мышовка —। самый редкий в этом году вид в районе. Обобщая
все" варианты населения опушечных полос, можно заметить, что вез-
де доминировала полевая мышь, а' численность северной мышовки
!и узкочерепной полевки была одинаковой (2—3-е место) и мало от-
личалась от числа особей полевой мыши. Многочисленными были
темная и красная; полевки. 1 j г ,
(Насекомоядные составили примерно .третью часть всего количе-
ства отловленных !млекопитающих. В каждом варианте биотопа
встречалось от 3 до15 видов землероек. Общее число видов 7, т. е.
отмечены все виды землероек, свойственные Северной Барабе.
В экотоне доминировала обыкновенная бурозубка, которая по чис-
ленности занимала первое место среди всех видов мелких млекопи-
тающих. Остальные виды землероек отлавливались единично (1—
3 экз. за время наблюдений).
94
У грызунов в течение летнего сезона отмечен только один подъем
попадаемости в цилиндры — конец августа — начало сентября.
У насекомоядных, вероятно, таких подъемов два. Уровень числен-
ности в июле несколько падает и остается почти без изменений до
середины августа, затем следует заметный подъем ее и вновь спад
к осени. , ! ; 1 , /
Для 1969 й. характерно ।начало обводненця территории, что по-
влекло'за собой изменение цикличности в Экологической группи-
ровке экотона. На 'сей раз доминирует ритм численности с периодом^
120 сут (см.,рис.!|23, I, г). Численность в этот год несколько возросла
у всех видов. Четко проявленного доминанта по численности нет.
Заметны на спектре пики 20-, 7-, 5-суточных циклов, связанных с по-
годными изменениями подвижности зверьков. Столь же| значителен
по мощности и 2-суточный цикл Территориальных перемещений -жи- ..
вотных. I - I I”
В 1970 г. в.экотоне обитало сравнительно мало грызунов| — 6—7
видов, а уровень их численности заметно уступал таковому преды- ।
дущего года. Отсутствовали красно-серая полевка, южная мышов-
ка и обыкновенный хомяк; i Все перечисленные виды были крайне
малочисленными и во всех других биотопах. Доминировали на сты-
ке биотопов полевка-экономка, красная полевка, узкочерепная по-'
левКа. Часто встречались темная полевка и полевая мйшь. Прочие
грызуны отмечены в малом числе.
Насекомоядные в отловах составляли почти 2/3 всех зверьков и
по численности значительно (почти в 2 раза) превосходили уровень
1969 г. В разных вариантах рассматриваемого биотопа встречалось
по 2—4 вида землероек. Всего на опушечных полосах зарегистри-
ровано в этом году 5 видов. Везде.доминировала обыкновенная буро-
зубка, численность которой превосходила не только численность
видов-доминантов, но и всей группы грызунов в целом. Второе место
по численности вслед за обыкновенной бурозубкой и узкочерепной
полевкой занимала малая бурозубка.
Попадаемость в цилиндры грызунов в течение лета была доволь-
но равномерной. Отмечалось лишь небольшое повышение ее в нача-
ле августа. Изменения в попадаемости землероек определялись чис-
ленностью ведущих видов —-обыкновенной | и малой бурозубок.
Подъем попадаемости начался со второй половины июня и достиг
максимума в середине июля. Очередные подъемы — в начале и кон-i1
це августа. ~ ! j ;
В 1970 г. на спектре циклов число мощных пиков возрастает.
Доминируют циклы, соизмеримые| с погодными,— 5-, 10- и 20-су- ;
точные (см. рис. 23, /„ Д). Проявлена и цикличность численности,
ее период составил 120 сут, т. е. по сравнению с прошлым годом этот
цикл менее мощный. Хорошо выражены на спектре высокочастот-
ные ритмы. 2,4—2 сут,1 связанные с территориальным передвиже-
нием грызунов. _ •
В 1971 г. на границе колка и поля доминировали водяная крыса
и полевая мышь, встречались все виды, отмеченные в районе иссле-
дования. По краю колка и выгона доминировала узкочерепная по-
I
I
95
левка и появилось много особей темной полевки. Численность зем-
лероек лишь незначительно превышала прошлогоднюю. В этом го-
ду в рассматриваемом биотопе «экотон» зарегистрировано макси-,
мальное для района исследований число видов землероек — 7, в
разных вариантах биотопа — от 6 до 7 видов. В среднем в экотоне
среди грызунов доминировала водяная крыса, среди насекомояд-
ных—обыкновенная бурозубка. Последняя была доминантой ио
отношению ко всем другим видам мелких млекопитающих экотойа.
Далее по степени доминирования идут узкочерепная полевка, по-
левая ’мышь, мышь-малютка, темная полевка, красйая полевка,,
малая бурозубка. Прочие виды грызунов и землероек в биотопе:
редки, .
У грызунов два подъема попадаемости в цилиндры — конец ию-
ня и начало сентября, у землероек также два подъема численности —
конец июня и середина августа. В эти периоды поймано в] Цилиндры
максимальное число зверьков. Численность землероек, оставаясь
высокой, незначительно снижалась к сентябрю.
С 1971 г. начался подъем численности водяной крысы. Снизилйсь.
видовое разнообразие и устойчивость сообщества, повысилась мощ-
ность высокочастотных периодических составляющих — домини- '
рует 2-суточный цикл подвижности. Цикличность численности изме-
няет период и становится 80-суточной, пик ее — второй по мощно-
сти на спектре (см. рис. 23 * I, е). Наименее проявлены пики ритмов
подвижности, связанные с погодными изменениями, из них силь-
нее выражены 20- и 7-суточные ритмы, соизмеримые по периоду с
температурными циклами.
В 1972 г. во всех вариантах экотона наблюдалось безусловное
преобладание водяной крысы, заметная численность была у поле- -
вой мыши и обыкновенного хомяка. Зверьки прочих видов ловились
очень редко. Таким образом, на опушечных полосах в год вспышки
численности водяной крысы в зависимости от вариант^ биотопа ока-
залось всего 5—6 видов грызунов.
Цо сравнению с 1971 г. в экотоне снизилось как число видов зем-
лероек, так и их обилие. В разных вариантах биотопа отлавливали
от 2 до 4 видов, а всего зарегистрировано 5 различных представите-
лей этого семейства. Доминировала среди них обыкновенная буро-
зубка, далее следовали крупнозубая, арктическая, средняя и малая
бурозубки1 были редкими. , . •
Порядок доминирования населяющих экотон мелких млекопи-
тающих был следующий: водяная крыса, обыкновенная бурозубка,
полевая мышь, обыкновенный хомяк, круцнозубая бурозубка.
У грызунов в течение сезона был один подъем попадаемости —
в начале августа. Он вполне соответствует времени смены стаций
у водяной крысы. Для землероек характерны два подъема численно-
сти — в конце июня и в первой половине августа.
Вспышка массового размножения в 1972 г. привела к полному
доминированию водяной крысы в экотоне (около 86%), поэтому
спектр ритмов здесь отражает набор цикличностей, характерных
для одного вида — водяной крысы. Доминирует 120-суточная пе-
I
96
риодичность численности (см. рис. 23, I, ж). Она отражает массовые
сезонные миграции грызунов. Остальные циклы, выражены гораздо
меньше. Среди них преобладали 17- и 10-суточные, близкие по пе-
риоду цикличности влажности воздуха и осадков.
Подводя итог рассмотрения населения экотона, необходимо от-
метить доминирование среди грызунов полевой мыши,, узкочерепной
полевки и водяной крысы. Во всех случаях, как и во всех других
биотопах, среди землероек доминировала обыкновенная бурозубка,
которая нередко была доминантой и среди всех обитающих в биото-
пе мелких млекопитающих. Число видов в биотопе: грызунов — от
от 3 до 11, землероек— от 2 до 7. Чаще всего здесь присутствуют
все виды, свойственные району, и только в годы вспышки числен-
ности водяной крысы число их значительно уменьшается. Среднее
соотношение числа отловленных грызунов к насекомоядным 63,7
и 36,3%, хотя в отловах грызуны колеблются от 40 до 89%, а насе-
комоядные — от 17 до 59% (табл. 12). Общая численность грызунов
в этом биотопе по сравнению с другими высокая. Численность зем-
лероек здесь в среднем уступает таковой биотопа «колок».
Общая динамика плотности сообщества мелких млекопитающих'
здесь выражена нечетко. Обычно подъемы подвижности наблюда-
ются в начале июня, июля и августа. Нередки спады численности
в конце июля.
В период 1966—1972 гг. для экотона характерен месячный ритм
изменения численности (рис. 24, 7, а). В литературе описан такой
же по длительности ритм изменения физиологических функций,
массы тела и активности у желтобрюхих сурков [Ward, Armitage,
1981]. Среди низкочастотных ритмов, характерных для этой биото-
пической группировки грызунов, наиболее выражен 5-летний цикл
численности. Ритмы такой длины описаны для грызунов рода Рего-
mysciis П. Р. Грантом [Grant, 1976]. Ранее цикличность такой же
длины в горах Финской Лапландии описана у 4 видов грызунов: по-
левки-экономки, темной и красно-серой полевок, а также норвеж-
ского лемминга [Tast, Kalela, 1971]. Этими же авторами отмечена
полная синхронность ритмов у описанных видов. Вторая по Мощно-
сти цикличность в этой группировке грызунов — 2,5-летняя (см.
рис. 24, II, а). Ее описаний в литературе не встречено. Значитель-
ный пик соответствует более чем годовой цикличности численности,
часто описываемой для разных видов грызунов [Heller, Poulson,
1970; Walker, Haskell, 1980]. Прочие циклы в сообществе очень ма-
ломощны.
Следующий интервал времени наблюдений — с 1978 до 1981 г.
В нем доминировал 240-суточный ритм изменений численности (см.
рис. 23, II, а). Заметными, по мощности были пики погодных подви-
жек (20-, 7- и 5-суточные). Высокочастотные компоненты спектра —
2-суточные ритмы — были проявлены менее всех прочих, т. е. спектр
ритмов численности и подвижности грызунов в экотоне был доволь-
но устойчив.
Дальнейшее развитие сообщества в 4979 г. привело к усилению
240-суточной цикличности численности грызунов (см. рис. 23, II, б).
7
А. А. Максимов, Л. Н. Ердаков
97
о 00 Л. -— Распределение численности населения млекопитающих «экотона» - Таблица 12
— — - Число ’ПИДОВ Грызуны Землеройки - -
Год Доминант (содоминант) грызунов землеро- 1 ек . 1 на 100 цил.-сут % ; на ЮО цил.-сут % Всего на 100 цил.'-сут Подъемы, попадаемости в • цилиндры
—
1966 Полевая мышь (лесная мышовка, полевка-экономка) 4—7 ? 5,0 86,2 1,21 17,8 6,21 Начало июня и сен- тября
1967 Полевка-экономка (полевая мышь, мышь-малютка) 9-10 ? 17,62 48,41 18,78 53,6 . 36741)?- Нет !
196'8 Узкочерепная полевка (красная полевка, лесная мышовка) 10-11 ? 7,79 83,6 1,51 16,4 9,30 Начало июня, июля, сентября. Спад — в кон- це июля
1969 Полевая мышь (обыкновенная бу- розубка, лесная мышовка, узкоче- релпая полевка) 10-11 3-5 11,49 73,5 _ 4,18 26,5 15,67 Начало сентября —
1970 Полевка-экономка (обыкновен- ная бурозубка, узкочерепная полев- ка, малая бурозубка) 5-8 2—4 5,79 40,3 8,62 59,7 14,41 Начало августа
1971 Водяная крыса (обыкновенная бу- розубка, узкочерепная полевка, нолевая мышь)__ И 6-7 17,60 62,1 10,80 37,9 ' 28,40 Конец июня и августа
1972 - Водяная крыса (обыкновенная бу- розубка, нолевая мышь, обыкновен- ный хомяк, крупнозубая бурозубка) 3-4 2-4 24,82 89,4 2,93 10,6 27775 - Начало августа
В сред- нем Полевая мышь (обыкновенная бу- розубка, узкочерепная полевка, во- дяная крыса) 3—11 2—7 15,16 63,7 8,65 ”36,3 \ 23,81 Начало июня, июля, сентября. Спад р конце июля. .
" о.
О ~г~\ | | —| Г~ “T~I i - I , '
oo <8 24 12 i 4 J 2 V“! 2 ' 0,1 0,05 0,03,
'10,1-ym Г,,:‘од
Puc. 24. Спектры сезонных п многолетних ритмов численности и подвижности
! грызунов биотопа «экотон».
Усл. обозн. см. рис. 16. ’ |
Из высокочастотных остался связанный с влажностью 5-суточный
ритм подвижности и весьма незначительный по мощности 2,4-суточ-
ный цикл территориальных перемещений животных. Численность
возрастала равномерно, значительного доминирования не наблю-
далось ни у одной популяции, кроме водяной крысы. Численность
ее в экотоне достигла 14,1 особи на 100 цил.-сут.
В 1980 г. началась вспышка численности водяной крысы и видо-
вая структура сообщества грызунов ib экотоне резко изменилась. ! j
Цикличность в этой группировке также стала иной (см. рис. 23,
II, е). Доминирующим стал цикл численности длиной в 60 сут. Близ-
кими по мощности оказались ритмы, связанные с погодными воздей-
ствиями — 20- и 5-суточные. Число мощных пиков на спектре зна-
> чптельно возросло, т. е. устойчивость сообщества снизилась.
J Дальнейшее развитие вспышки в 1981 г. и сильная засуха, на- I,
чавшаяся еще осенью 4980 г., совершенно изменили вид'спектра цик- ;
лов численности и подвижности.; Доминирующим стал 10-суточный
цикл, связанный с осадками (см, рис. 23, II, г). На спектре имеется
значительное число пиков большой мощности, т. е. «шум», создава- j
емый малой численностью (наблюдается резкое падение ее во всех '
популяциях грызунов), возрастает. Сообщество после пика числен-
ности водяной крысы и в связи с сильной засухой оказалось в глубо-
кой депрессии. (Видовое разнообразие в нем резко уменьшилось.
К осени в экотоне наблюдалось малочисленное, небогатое видами
пионерное сообщество грызунов.
В среднем в этой серии сукцессии, как и в предыдущей, домйнп- .
ровала 240-суточная цикличность численности (см. рис. 24, I, 6)t
но были заметны пики циклов подвижности, связанные с погодными '
колебаниями: 5- и 10-суточные колебания влажности и 20-суточные
изменения температуры. На более низких частотах доминировал по
мощности 3,3-летний цикл; 3-летняя цикличность часто упомина-
ется в литературе. Совсем недавно таковая описана для полевок ро-
99
|| да Microtus в тундре [Batzli, 1981]. Кроме того, для группировки
J । грызунов в экотоне оказался характерным небольшой по мощности
лунный цикл — 29,5 сут.
! Биотопические группировки популяций обладают собственными
; характерными колебаниями численности и определенными, прису-
|||, щими им, циклами. Эти, циклические изменения могут быть ответом
ц!| ; сообщества на совокупность факторов;конкретного местообитания и'
II I отличаться от цикличности составляющих его популяций. Бйотопи-
, ческая группировка — это экологичёскп сложившийся набор видов
|| в конкретном местообитании, для которого характерны устойчивость
м ' видового-состава, его разнообразие и смена доминирования популя-
। I, ций. Все эти характеристики цикличны. Биотопическая группиров-
11 / ка должна рассматриваться как подсистема большой экологической,
г । j системы;— биоценоза. .
1 Каждый биотоп характеризуется своеобразием изменений труп-:
I I пировки мелких млекопитающих по численности и особенностям ее
|| I цикличности,,^ числу видов, соотношению доминантой, распределе-
;|1! i нию во времени максимумов попадаемости животных в ловушки.
|,|| I j | Наиболее длинные циклы (от 240 сут до 2 лет) доминируют среди
|| I |. внутрисезонных в биотопах: поле, луг, экотон. На болотах и в кол-
: ках доминирующая цикличность в течение, сезона короче — обычно
11, ; 80-суточна1я. Видимо, эти различия связаны с особенностями микро-
I'lll , климата местообитаний. Так на болотах в окр. с. Ровенское-в тече-
Ь ние 80 сут уровень воды изменяется от 50—70 см над поверхностью- •'
/ц болота в мае до 1,5—2 м ниже уровня поверхности в августе.
j,l|i , Преобладающие по мощности многолетние циклы биотопиче-
I ских группировок также! различаются. Наиболее длинные отмечены
|Т ; в колках, на лугах и полях (9,8 лет). В то же время в биотопе «боло-
||,'! 1 то» и экотоне циклы иные. Они не превышают 3—5 лет. Скорее все-
го, это определено видовым составом грызунов в группировках;
I в болотах преобладают полевки с цикличностью 3—4 года; в экото-
J нах — мыши с цикличностью 3—5 лет.
й I . Цикличность в биотопических группировках соизмерима с ус--
I, , тановленными для Барабы 11-летними циклами увлажненности тер-
l| I рптории. Эти циклы описаны как приблизительно равные 11 годам/
। ! обычно же они варьируют от 9 до 13 лет, т. е. 9,8-годпчный цикл^
/|| проявленный। на спектре биотопических группировок, вполне соот-
| ,|| ветСтвует режиму обводнения территории; 3—5-летняя цикличность
/Ц кратна основному циклу (11-летнему). Она присутствует на спектре
I почти каждой из описанных группировок. Нужно заметить, что цик-
I личности группировок своеобразны по набору ритмов и не совпадают
II', со спектрами -любой из входящих в группировку популяций. Сте-
!| пень их соответствия нуждается в отдельном анализе.
/ I " ;
СООТВЕТСТВИЕ ЦИКЛОВ БИОТОПИЧЕСКОЙ ГРУППИРОВКИ
I И ДОМИНИРУЮЩЕГО В НЕЙ ВИДА
। | Любая биотопическая группировка в Центральной Барабе насчи-
I , тывает обычно 8—10 видов грызунов [Сообщества..., 1978]. Каждая
' популяция видов испытывает определенные и различные по периоду
100
колебания численности и подвижности особей. Группировка же этих
популяций в биотопе имеет собственный набор ритмов, отличаю-
щихся от колебаний любой из популяции по периоду и мощности.
Видимо, существует некоторая аналогия с колебательными систе-
мами, описываемыми в! физике. Известно из физики,, что система вза-
имосвязанных колеблющихся элементов имеет период собственных
колебаний, который больше по периоду и меньше по частоте, чем
у любого из составляющих эту систему элементов. , Это верно в том
случае, когда все осцилляторы равны по своим характеристикам
(например, цепь, собранная из одинаковых колебательных контуров).
Бпотопическая группировка отличается от такой идеализированной
системы тем, что ее осцилляторы (популяции) отличаются по своим
характеристикам, а особи этих популяций имеют разную долю в на-
селении группировки. Некоторые виды многочисленны и домини-
руют. Кроме того, на группировку, испытывающую колебания чис-
ленности особей, оказывает воздействие внешняя среда. Ее элемен-
ты (характеристики) могут быть синхронизаторами ритмов популя-
ций, затягивающими их периоды агентами. Наконец, сама биото-
пическая группировка с большой натяжкой, может считаться систе-
мой взаимосвязанных осцилляторов. Связи между популяциями
совместно обитающих грызунов описаны в литературе еще недоста-
точно. Наибольшее приближение к такому описанию достигнуто в
исследовании И. А. Шилова [1977].
Таким образом, рассмотрение биотопической группировки гры-
зунов как системы взаимосвязанных колебательных единиц сильно
идеализируется. Мы предполагаем ее как модель, рассчитывая, что
у нее общие с реальной группировкой свойства.
Рассмотрение аналогий цикличности доминирующего вида и со-
общества, в которое он включен, мы проводим на небольшом числе
примеров. Доминирование может иметь разную степень и опреде-
ляется его показателем [Одум, 1975]. У нас таким показателем слу-
жит численность. Доминант — наиболее многочисленный вид в от-
ловах. Он может составить от 20 до 90% всех пойманных в ловчие
канавки животных. Предполагается, что чем больше доля особей ,
вида в сообществе, тем сильнее воздействие этой популяции на цик-
личность сообщества. Крайний случай — доминант составляет 100 %
населения биотопа, и1 тогда ритмы численности биотопической груп-
пировки будут идентичны ритмам одного вида животных.
В 1967 г. среди грызунов в окр. с. Ровенское доминировала по-
левка-экономка. Наибольшее проявление доминирования было па
молях сельскохозяйственных культур и опушечных полосах. В . этих
биотопах численность ее в отловах составляла 32,7 и 23,6% от сум-
марной соответственно. Сравнивая спектры колебаний численности
и подвижности доминанта и биотопической группировки грызунов
в целом (рис. 25, а, б), можно заметить, что совпадение периодов
мощных периодических составляющих у них невелико. В биотопе
«поле» спектры ритмов имеют много мощных пиков, но совпадение
периодов наблюдается только в трех полосах частот: 48-, 8,5- и 4-
суточные периодичности. Мощность этих пиков у группировки и до-
101
%
: Рис. 25. Соответствие спектров колебаний численности п подвижности '1,;
| у биотопической группировки грызунов - п доминирующего вида в •
- ! ней — полевкп-экономки в 1967 и 1970 гг. '
Биотопы: а —cm.:, 1987 г.; б — экотон, 19S7 г.; в—болото, 1970 г. I—труп-
! , пировка грызунов, 2 — полевка-экономка.
' I
минаита всегда различна. Еще менее схожи спектры I в экотоне:,
Здесь совпадают 10- и 4-суточные колебания, но их мощность значи-
тельно различается. ,
Если возрастает степень доминирования и доминант составляет».
половину общей численности грызуновг спектр его более сходен с
102
^>4824 12 6 4 3 I 2
Рис. 26. Соответствие спектров колебания числен-
ности и подвижности у биотопической группиров-
ки грызунов и доминирующего вида в ней — водя-
ной крысы в 1972 г.
Биотопы: а — колок, б — экотон, в — поле. 1 — группиров-
ка грызунов, 2 — водяная крыса.
общим спектром биотопической группировкп. Так, в 1970 г. полев-
ка-экономка доминировала в сообществе. Особенно сильно это было
проявлено в болотах, где она составляла 51,17% всех отловов. Срав-
нение спектра ритмов доминанта и сообщества (см. рис. 25, е) пока-
зывает, что периоды основных по мощности пиков на этих спектрах
совпадают, это 5-, 4-, 2,8-, 2,3-суточные пики. Кроме того, совпада-
ют периоды пиков 60-, 10-, 3,5-, 2,1-суточных ритйов, хотя они и
неодинаковы по мощности. , ।
Наиболее проявлено доминирование в районе исследований у во-
дяной крысы. Во время вспышек массового размножения этот вид
нередко составляет более половины. зверьков в отловах. Особенно
хорошо .такая высокая степень доминирования была проявлена в
1972 г. во многих биотопах. Так, в «колке» водяная крыса занимала
88,38% отловов грызунов. Спектры колебаний ее численностп и ко-
лебаний численности всего сообщества колка очень похожи (рис. 26,
а). Доминирующие пики (48 сут) и 3,7-суточная периодичность, зна-
чительная по мощности, есть на обоих спектрах. Оба спектра похо-
жи и имеют по одному доминирующему пику в низких частотах.
В опушечных полосах доля водяной, крысы в отловах грызунов
составляла 86,74%. Спектры ритмов сообщества и доминанта здесь
еще более схожи, чем в «колке». На спектрах по периоду и мощности
совпадают практически все доминирующие и значимые пики (см.
102
рпс. 26, б). Только в области высоких частот можно видеть либо не-
которое расхождение их по близким полосам частот, либо несоот-
ветствие в мощности.
Особенно полное совпадение наблюдалось в биотопе «поле».
Здесь доля водяной крысы в отловах составляла 82,38%. Спектры
же биотопической группировки и вида-доминанта совпадают прак-
тически полностью во всех полосах частот (см. рис. 26, в).
Таким образом, предположение о близости спектров циклично-
сти численности и подвижности грызунов доминирующего вида и
биотопической группировки, в которую он входит, подтверждается.
Близость спектров очевидна (см. рис. 25, 26), но степень доминиро-
вания должна быть не .менее 50% попадаемости в отловах. Прй1
меньшей доле валовой численности усмотреть подобие спектров
трудно. -
При изучении биологических ритмов часто проводят описание
всей колеблющейся системы по . одной из ее функций. Активность
животного оценивают по его бегу в колесе «моционнике», т. е. толь-»
ко по локомоторной активности [Де Курси, 1964; Раусон, 1964].
Иногда проводят некоторое сравнение соответствия ритмов отдель-
ных функций животного (локомоций, питания и пр.), т. е. циклич-
ности активности (состояния бодрствования), и приходят к выводу
о подобии этих колебательных процессов и о возможности описать
систему на основе наблюдения одной из ее характеристик [Gatter-
mann, 1978]. Исходя из сделанных нами сравнений, такое описание
не всегда возможно. Даже если численность доминанта составляет
половину от всего сообщества, описание цикличности только по не-
му будет неточным (см. рис. 25).
Сравнивая спектры цикличности доминанта и сообщества, можно
видеть значительное их соответствие. Между тем, подобие спектров
не прямо зависит от доли доминанта в сообществе. Об этом свиде-
тельствуют данные по ритмам водяной крысы в 1972 г. (см. рис. 26).
Частота встречаемости в отловах водяной крысы из разных биотопи-
ческих группировок отличалась мало,, а соответствие спектров —
значительно. Имеется даже тенденция уменьшения общности спект-
ров р ростом доли доминанта в уловах. Доля возрастает в порядке
биотопов «поле», экотон, «колок», а в обратном порядке увеличива-
ется общность спектров ритмов.
Таким образом, повышенная численность грызунов какого-либо
вида может влиять на характер цикличности биотопической группи-
ровки. Спектр ритмов такой группы грызунов может даже стать по-
хожим на- спектр вида-доминанта. Подобие спектров ритмов числен-
ности и подвижности у сообщества и доминирующего в нем вида
не прямо пропорционально росту численности последнего. Зависи-
мость между ними, видимо, более сложная. Предположительным
механизмом этого явления может быть канализированная адаптация,,
свойственная сразу многим видам, мелких млекопитающих [Уод-
дингтон, 1970]. Она проявляется в ответ на одинаковое изменение
среды. Такие адаптации для серых полевок известны, например
блокирование беременности при избыточной плотности популяции
[Mallory,, Cltilow, 1977].
104
Глава IV
ВЛИЯНИЕ ПРИРОДНОЙ цикличности
НА ВИДОВОЕ РАЗНООБРАЗИЕ
И РИТМЫ ЧИСЛЕННОСТИ МЕЛКИХ МЛЕКОПИТАЮЩИХ
‘ ВОЗДЕЙСТВИЕ МНОГОЛЕТНЕЙ ЦИКЛИЧНОСТИ СРЕДЫ
НА ВИДОВОЕ РАЗНООБРАЗИЕ МЕЛКИХ МЛЕКОПИТАЮЩИХ
i (йа примере полевок)
После! анализа различных видов цикличности, характерных для по-
пуляций и биотопических группировок,, рассмотрим воздействие
этой цикличности на видовое разнообразие в сообществе животных.
Считается, что экосистема в ходе сукцессии увеличивает свою ста-
бильность. По современным представлениям с устойчивостью ее
тесно связано видовое разнообразие, рост которого усиливает ста-
бильность биоценоза [Pimlott, 1969; . McNaughton, 1970; Bezzel,
Reichhoff, 1974]. Эта точка зрения вошла в большинство учебников
экологии и подтверждается следующими положениями: среда с ус-
тойчивым климатом благоприятна для высокой специализации видов
и возникновения узких экологических ниш. Видов будет больше
в том из двух местообитаний с одинаковым диапазоном доступных
ресурсов, в котором каждый вид использует небольшую часть всех
ресурсов [Пианка, 1981].
Некоторыми авторами оспаривается однозначная связь видового
разнообразия и стабильности сообществ [Whittaker, 1972; Buzas,
1972]. Общепринятому мнению о том, что стабильная среда способст-
вует увеличению видового разнообразия, противоречат примеры,'
в которых этому помогает цикличность среды. Так, наличие дня и
ночи дает возможность сосуществовать дневным и ночным живот-
ным — потребителям одного пищевого ресурса. Сезонность климата
также приводит к увеличению разнообразия видов.
В природных сообществах существуют 1 и описаны кроме сезон-
ных еще и многолетние колебания условии среды. Как правило, по-
следние не имеют строгого периода и значительно менее выражены,,
чем сезонные. В настоящем разделе на одном примере рассмотрим,-
как многолетние нестрого периодические колебания условий среды
могут служить фактором, увеличивающим видовое разнообразие.
Кроме того, покажем, что цикличность условий среды облегчает со-
существование симпатрических видов.
Прямое сравнение стабильного и цикличного по климату сооб-
ществ не разрешит нашу задачу. Подобрать совершенно одинаковые
сообщества, различающиеся только по одному признаку — наличию
цикличности, невозможно. Это соображение вынуждает обратиться
к другим, менее прямым способам доказательств значения циклич-
ности среды в увеличении видового разнообразия. Одним из них
105
I • является комплексное экологическое рассмотрение группы родст- ;
I венных видов, обитающих в сходных по составу сообществах. Причем
। многолетняя цикличность среды имеется только в одном из iHhi.
! Смысл такого рассмотрения состоит в выяснении того, помогает ли
; цикличность среды сосуществованию родственных видов, т. е. спо-
собствует ли увеличению видового разнообразия. 1 i ’
Для решения наших задач были взяты три таксономически! и
экологически близких вида серых полевок, обитающие на одной тер-
[ ! ритории !(Бараба) в изменчивой природной обстановке. Многолетние
колебания условий среды ярко выражены и подтверждены анализом
метеорологических и биоценотических, данных [Сляднев, 1965;
. Куркин, 1976]. Мы хотим показать, что видовое разнообразие может '
увеличиваться за Счет, цикличности изменений среды. Например,,
в случае стабилизации внешней среды в Барабе количество видов
серых полевок здесь, скорее всего, уменьшилось бы. Доказательство
этого проводится с помощью комплексного анализа экологически^
ниш видов, т. е. сравнения исследуемых полевок по всем важнейшим
экологическим характеристикам: пищевая и биотопическая специа-
лизации; суточная и сезонная цикличность жизнедеятельности, мно-
голетняя динамика численности. Сильное разделение экологических
ниш по любой из этих характеристик позволит сосуществовать ви-
цам н,а одной территории. Не менее важно и сравнение морфологи-
ческих черт, так как совокупность таких различий у близких видов
определяется разным |их образом жизни.
По каждой из основных характеристик и по комплексной (мор-
фологической) все рассматриваемые виды довольно близки. Тем не
менее именно благодаря сумме некоторых различий в отдельные го-
ды пз-за природной цикличности условий среды получает преиму-
щество то один, то другой вид, остальные же в это-время имеют под-
чиненное значение. Если бы подобных колебаний не было, например,,
в Барабе установился бы степной климат и соответственно ландшаф-
ты (в этом направлении, по мнению К. А. Куркина, и развивается
бараблпский биоценоз), то виды полевок, тяготеющие к |Лесам и бо-
лотам, исчезли бы и остались только степные1, т. ег видов стало бы
меньше. Для подтверждения этого мы сравниваем сообщество Бара-
бы и близкое ему в пойме среднего течения р. Оби, в котором от-
сутствует один из трех видов, тяготеющих к остепнепным участкам.
I : ' ' , S
Черты сходства серых полевок ! 1
! " ' ’ . . .
:Виды серых полевок обычно плохо различимы, и определение их про-
водится по комплексу морфологических и краниологических призна-
ков, что само по себе говорит об их большом экологическом сходст-
ве. Об этом писал С. С. Шварц [1980]: «Род,,Серые полевки11 насе-i
ляет все ।ландшафтные зоны, нашей планеты. Тем не менее роду
может быть дана достаточно четкая экологическая характеристика.
Ото мелкие, способные к подземному образу жизни полевки, обла-
I
106
Таблица 13
Соотношение длины и массы тела взрослых полевок из с. Ровенское * [Со-
общества..., 1978]
j ( Вид ! 1- 1 Самец Самка
'I п ! М±т - п •М±т
! Д и н а, \т. Ал а, мм.
, ' । Узкочерепная ' 37 98,4±2,'58 ~ 25 । 96j5±3,ll
Пашенная ' i 30 1 103,3+2,32 28 l! 97,4±2,26,
Экономка 1 151 117,7±1,22 ’ 135 1110,3+1,13
1 । Масса тела-, г i Ь 11
Узкочерепная ( 37 . 26,7±1,07 25 ' 24,5±1,10
Пашенная 1 । 30 29,1±1,52 28 31,7±0,95
Экономка I ,151 49,9+1,21 135 33,4±0,97
* На 95%-ном уровне значимости неразличимы самцы узкочерепвой и па- и
шейной полевок по массе In длине тела, а также самки узкочерепной и пашенной по- и
левок — по длине тела и самки пашенной полевки и лолевки-экономкп — по мас-
се тела. । । 1 . • ’ ।
. \
дающие комплексом морфофизиологических приспособлений, обес-.
печивающих использование в качестве основного источника жизни
корма низкой калорийности: с развитой химической терморегуля-
цией, обладающие совершенной системой регуляции интенсивности
размножения, позволяющей быстро восстанавливать численность
после ее депрессии» (с. 246). , ;
Три рассматриваемых вида серых полевок: узкочерепная, па-
шенная! и полевка-экономка очень похожи морфологически ц эколо-
гически по общим не детализированным характеристикам. Такм
малоразличимы в |Барабе экстерьерные । показатели животных
(табл. i'13). Беременность у зверьков этих видов длится 20 дней,,
половозрелость наступает через 20—30 сут после рождения. Плодо-
витость составляет 5—7 эмбрионов на беременную самку. Размно-
жаются 'ОНИ| с (середины мая| по сентябрь. Зверьки всех1 трех видов
могут быть отловлены одновременно в одном и том же биотОпе. Хотя
они имеют известную биотопическую приуроченность, тем не менер
на пиках численности грызуны любого из этих видов оказываются
доминирующими во всех биотопах. Для всех трех видов характерны^
циклические изменения численности. В питании зверьков преобла-
дают вегетативные части'растений, что в । большой мере определяет
сходство ритмов их суточной активности. Форма е'е ;у всех полевок
полифазная. Только сравнение по комбинациям признаков позво-
ляет найти различия между этими видами полевок [Dienske, 1979].
Естественно, что при пристальном изучении любой из перечислен-
ных характеристик проявляются межвидовые различия. Еще более
тонкое исследование дает возможность найти различия между попу-
ляциями внутри каждого вида. Различия для них так же характер-
ны, как и общность. В данном случае общность оказывается боль-
107
Динамика среднесезонных отловов трех видов полевок.
Декада отлова,
впц. 1 ‘ 2 з . 4
Экономка 1,39±0,81 1)б2±0,49 2,06±0,66 2,26±0,79
Узкочерепная i 1,45±0,86 2,34±1!,13 . 2,36+0,89 1,91+0,37
Пашенная 1,43±1,21 1,44±0,68 0,74^=0,36 : 0,75±0,34
шей, чем различия. Это свидетельствует о значительном совмещении
экологических ниш рассматриваемых видов и1 соответственно об,-
имеющейся между ними конкурентной напряженности. По-видимо-
му, она могла бы привести к полному иёключению какого-то вида,из
рассматриваемых, но в Барабе этого не происходит,
г 'А
Разделение жизнедеятельности серых полевок во времени
Различий в сезонной цикличности рассматриваемых видов полевок
в литературе не описано. Известно, что все они размножаются в
один и тот же период года [Сообщества..., 1978], а сроки беремен-
ности п периоды вскармливания молодых у этих грызунов близки..
Все это говорит в пользу общности сезонных циклов у всех трех
популяций.
По нашим данным, при сравнении внутрисезонной динамики
отловов в канавки у зверьков всех трех видов в окр. с. Ровенское
в течение сезона размножения была одинаковая поцадаемость
(табл. 14).
Данные подекадных отловов у всех трех видов в большинстве
случаев достоверно не различаются. Сравнивая относительную чис-
ленность в одинаковые декады, можно видеть, что у полевки-эконом-
ки по этой характеристике нет достоверных средних различий с
узкочерепной полевкой, а у той, в свою очередь,— с пашенной.
Достоверные различия средней численности имеются только в пя-
той и шестой декадах между отловами полевки-экономки и пашенной
полевки (см. табл. 14). У всех трех видов грызунов наблюдается не-
которое нарастание численности в летние месяцы и снижение ее
в начале и конце лета.
Если обратиться к анализу > количества и мощности периоди-
ческих составляющих ритмов активности у этих видов полевок, рас-
считав их спектры (рис. 27, а), можно, заметить некоторые различия.
Даже при беглом сравнении обнаруживается, что у полевки-эконом-
ки в отличие от остальных невелика мощность суточного пика
(см. рис. 27, а, б). У всех трех видов имеются мощные периодиче-
ские составляющие длиной менее суток. Расположены же они у каж-
дого вида в специфической полосе частот. Самые короткие периодич-
ности на спектре ритмов узкочерепной, а самые длинные — на спект-
ре полевки-экономки (см. рис. 27).
108
Таблица 14
в окр. с. Ровенское, особей на 100 цил.-сут
с 1 июня
5 6 7 >8 9 10
з;о5±1,и 3,46+1,89 , 2,41+0,98 1,35+0,34 1,"11+0,47 1,35+0,68
1,89+0,39 1,29±0,26 ' 2,00+0,49 1,18+0,31 1,08+0,29 1,05+0,19 '
- ! 0,55+0,24 0,61±0,24 0,98+0,39 0,75+0,39 0,86+0,34 0,77+0,34
Сравнение хронограмм у этих видов не дает оснований для опи-
сания различий в формах их суточной активности (см. рис. 27, б).-
У всех трех видов форма активности полифазная и активность круг-
лосуточная. Причем у них одинаково велика вариабельность подъе-
мов и спадов активности и, следовательно, вероятность того, что в
природе зверьки этих видов часто активны одновременно.
! Разделение по биотопам и динамика численности
Наибольшие экологические различия для этих видов касаются ха-
рактера их биотопической приуроченности [Громов, Поляков, 1977].
Рассматривая верность разным биотопам \ можно, однако, за-
метить, что все три вида тяготеют к экотону и избегают полей
(табл. 15). В остальных местообитаниях происходит дифференциров-
Т аб лица 15
Верность биотопам у серых полевок в окр. с. Ровенское
Вид Индекс верности биотопам
Поле Луг Колок Болото ' Экотон
Узкочерепная • -0,10 0,58 —0,5 -0,15 0,01
Пашенная —0,40 -0,10 0,37 —0,01 0,18
Экономка ; -0,35 0,42 -0,38 0,21 0,10
ка этих видов. В то же время замечено, что при большой численности
любого из трех видов полевок он расселяется практически по всем
биотопам [Сообщества..., 1978] и везде доминирует. Несмотря на
имеющиеся в литературе данные о биотопической приуроченности
этих грызунов!, ни один из рассматриваемых видов не описывается
как стенотопный- [Громов, Поляков, 1977]. Это тоже подтверждает
экологическую пластичность и общность местообитаний у данных
полевок. । ।
Наблюдая динамику численности зверьков, можно заметить, что
периодически каждый из видов доминирует среди прочих. Преобла-
дание его численности обычно длится 1—2 года (табл. 16). Видимо,,
это время доминирующая популяция имеет какие-то преимущества,.
1 Индекс верности рассчитан- по формуле из книги «Сообщества мелких
млекопитающих Барабы» [1978].
109
I
Рис. 27. Спектры (/) и хронограммы (II) ритмов активности серых полевок.
а пашенная; б — полевка-экономка; в — узкочерепная полевка (вертикальными чер-
тами на графиках помечены доверительные интервалы при Р — 0,05).
Таблица 16
Смена доминирования численности у серых полевок в окр. с. Ровенское, особей
на 100 цил.-сут
: Вид , Год наблюдения
1,866 1967 1968 1969 1970 । 1971 1972-1973 | 1974
Пашенная! 0^2 1,41 0,53 0,23 .1,071 2,36: Вспышка; чпс- 1 0,39
Экономка 1,10 3.21 о;бо 0,56 i 2,70 0,87 ленности во- s ! 2,05
Узкочерепная ; 0,27 2,18 1,50 i 1,36 1,97 1,75 дяной крысы j 2,51
может быть, более подходящие для нее условпя среды в сравнении с
двумя другими.. Через 1—2 года лучшие условия наступают для дру-
гого вида серых полевок, и он, в свою очередь, становится домини-
рующим по численности. На монотонность этой смены доминирования j
серых полевок в Барабе влияют только вспышки численности водя-
ной крысы. В период такой вспышки все прочие виды грызунов на-
ходятся в глубокой депрессии, что отмечено в 1972—1973 гг. Мно-
голетние изменения численности у всех трех видов довольно син-
хронны. Доказательство большого, сходства динамики численности
у этих видов полевок в Барабе сделано при анализе экологической
структуры сообщества мелких млекопитающих [Ердаков и др.,,
1980], т. е. влияние на численность внешней среды в основном оди-
наково, различия же видовых требований проявляются только в сме-
не доминирования. !
Смена доминирования выражается в попеременном достижении
видами высокой численности при улучшении условий местообитания.
Иными словами, при большой общности экологических требований
животных периодически среда становится оптимальной то для одно- ,
го, то для другого.вида, что является для каждого из них поддержи-
вающим фактором. Если природной многолетней цикличности нет,
то одному из видов всегда лучше, чем прочим.
Для сравнения с Барабой использовали данные изменений чис-
ленности и смены доминирования у серых полевок в пойме среднего
течения р. Оби (табл. 17). Можно заметить, что здесь всегда домини-
рует полевка-экономка. Пашенная полевка уступает ей в числен-
ности, а временами совсем отсутствует. Популяция полевки-эконом-
ки возобновляется в пойме за счет мигрантов с коренного берега
, Таблица 17
Доминирование по численности серых полевок в пойме среднего течения р. Оби
в разные годы, экз.7100 цил.-сут
Вид 1957 .1958 1962 1967 1968 1970 I' 1971 1972 1975 1977
Экономка 1,36 1,09 9,36 9,81 28,36 17,59 23,67 23,52 1,61 11,25
Пашенная 0 0,18 0 0,06 0 7,79 1,35 0,06 .0,91 0,85
Примечание. Данные'приводятся из работы А. А. Максимова с соавторами
[1981]. В 1989 г. особенно большой и продолжительный разлив вызвал глубокую депрес-
сию численности, поэтому сборы этого года мы не приводим.
111
(узкочерепной полевки здесь нет совсем, хотя имеются луга, пригод-
ные для ее обитания). Видимо, условия поймы всегда благоприятст-
вуют полевке-экопомке и всегда малопригодны для пашенной.
Разделение по морфологическим (краниологическим) признакам
5 Сравнивали результаты девяти измерений черепа у исследуемых ви-
дов (табл. 18). Предполагалось, что различия в образе жизни могут
проявиться при сравнении краниологических признаков. Для мно-
гих из них описана связь с различными проявлениями жизнедеятель-'
ности [Воронцов, 1967; Башенинд, 1977]. Различия между видами-
оценивали с помощью.дискриминантного анализа. При сравнении же
отдельных признаков у разных видов использован критерий Стью-
дента. - ।
По совокупности всех девяти признаков виды хорошо различимы
по критериям Фишера и Стъюдента. Но по такому же количеству
признаков не хуже различимы и представители разных популяций
одного и того же вида (см. табл. 18).
Рассматривая различия по отдельным признакам, можно видеть,
что особенно сильны они в размерах скуловой ширины, межглазнпч-
ного расстояния и высоты черепа. По-видимому, здесь сказывается
своеобразие биотопического распределения. Утолщение черепа (уве-
личение ширины) обычно связывают с норной деятельностью [Бате-
нина, 1979]. Именно эта деятельность заметно различается в зависи-
мости от биотопа. На болотах норы редки, а наиболее интенсивная
роющая деятельность у грызунов проявлена на лугах.
Попарные сравнения признаков часто не дают достоверных раз-
личий не только у зверьков различных популяций (см. табл. 18),:
но и у полевок разных видов. Даже различия в длине диастемы и
зубного ряда (признаки, которые связывают с особенностями пита-
ния грызунов) достоверно неразличимы у пашенной и узкочерепной,
а также экономки и пашенной полевок (см. табл. 18).
Таким образом, по краниологическим признакам межпопуляцион-
ные различия полевок сравнимы с межвидовыми. Это свидетельству-
ет о большом сходстве рассмотренных видов и их значительной эко-
логической и морфологической пластичности.
Известно, что небольшие различия в сумме способствуют эколо-
гической изоляции, которая снижает конкурентную напряженность
и допускает симпатрию [Солбриг О., Солбриг Д., 1982]. Так, при
постоянном общем сходстве рассмотренных видов по каждой из ха-
рактеристик обнаруживаются и некоторые различия.
Наибольшие различия у этих видов сказываются в биотопической
приуроченности, которую нетрудно сопоставить с многолетними
циклическими процессами среды, описанными для Барабы. Периоды'
иссушения, когда преобладают остепненные территории, сменяются
периодами увлажнения и остепненные участки вновь заболачивают-
ся, т. е. в разные годы создаются оптимальные биотопы для одного из
видов. Но они не сохраняются настолько долго, чтобы виды, нахо-
112
Таблица 18
Сравнение краниологических показателей взрослых самцов из популяций раз-
ных видов серых полевок из окр. с. Ровенское (Р) и поймы р. Оби (0) (подчерк-
нуты значения t, удовлетворяющие 0,05 уровню значимости)
Признак ! > * 1 Популя- ция ‘ п t Популя- ция п t , Популя- ция п t
Место : п ; t
Длина 1. общая i р О в е н С К О е
р 58 1 10 1 39 3 101
О 41 1,12 2 8 2,08 3 10 1,16 2 6 1,74
2. кондилобазальная р О 58 41 0,35 -1 2 10 8 0,86 1 3 39 10 0,78 3 2 10 6 0,38
Ширина 3. скуловая р О 58 41 1,25 1 2 10 8 0,29 1 3 39 10 0,48! 3 2 10 6 5,26
4. межглазничная р О 58 41 4,84 1 2 10 8 0,42 1 3 39 10 1,57 3 2 10 6 7,28
5. Мозговой отдел р О 58 41 2,55 1 2 10 8 0,45 1 3 39 10 1,01 3 2 10 6 6,41
6. Носовой отдел р О 58 41 1,63 1 2 10 8 1,71 1 3 39 10 1,03 з 2 10 6 0,05
7. Высота черепа р О 58 41 3,24 1 2 10 8 2,04 1 3 39 10 2,59 3 2 10 6 4,46
8. Длина верхнего ряда зубов р О 58 41 1,46 1 2 10 8 7,62 1 3 39 10 0,53 3 2 10 6 3,87
9. Длина диастемы р О 58 41 3,93 1 2 10 8 7,07 1 3 39 10 1,80 СО 10 6 0,68
Анализ по 9 признакам Критерий Ствюдента 9,34 17,9 5,63 18,65
» Фишера 1,16 1,41 0,57 1,48
Примечание. 1 — полевки-экономки, 2 — узкочерепные, 3 — пашенные.
дящиеся в неоптимальных условиях, совсем исчезли на этой терри-
тории. Видимо, поэтому сохраняется весь набор видов (в данном
случае три).
В той же ситуации в пойме р. Оби при схожем составе сообщества
никогда не наступает фазы иссушения территории (река стабилизи-
рует влажность в пойме). Видимо, поэтому там отсутствует узкоче-
репная полевка, так как для нее никогда не наступает биотопиче-
ского оптимума.
Анализ наших данных позволяет сделать вывод, что сосущество-
вание трех видов серых полевок в Барабе возможно в основном
благодаря многолетним колебаниям условий среды. Следовательно,
такие колебания могут быть фактором, позволяющим сосуществовать
близким видам и тем самым поддерживающим видовое разнообразие.
8 Л. А. Максимов, Л. II. Ердаков
ИЗ
Напрашивается предположение, что причины симпатрип объясняют-
ся не только биологическими свойствами, позволяющими близким
видам одновременно сосуществовать на одной территории, но и упо-
рядоченным внешним воздействием на комплекс их популяций.
’’ . i I ; - j i ' < 1 ; ' i
ЦИКЛИЧНОСТЬ ИЗМЕНЕНИЯ СОСТАВА . р !. ' ! 1 д
И ЧИСЛЕННОСТИ НАСЕЛЕНИЯ МЕЛКИХ МЛЕКОПИТАЮЩИХ
В МНОГОЛЕТНЕЙ СУКЦЕССИИ ЭКОСИСТЕМЫ БАРАБЫ t
Н I J "[ [ ! , ' 'll
В процессе развития сообщества приобретают известную автономность
и независимость от окружающих условий, подгоняя свою жизнь не
к флуктуациям внешней среды, а вырабатывая собственные эндоген- .'
ные ритмы. Механизмами этих преобразований служат изменения-
внутренней структуры сообществ: увеличение видового разнообра-
зия, усиление специализации видов (расхождение по j экологическим
нишам). В ходе сукцессии усиливается стратификация и пространст- [
венная гетерогенность; сообществ [Чернова, 1977].
Одна р пз основных закономерностей сукцессии — волнообраз-
ность процесса. Колебательный характер имеют изменение соотно-,
тлений и численность видов, сезонная и сукцессионная динамика тес- ;
но переплетены. Волнообразностй развития комплекса беспозвоноч- (
ных показана, например,; при первичном почвообразовании И. В. Сте- ;
баевым [1963, 1968]. I г
Численность животных — хороший показатель того, насколько i ;
благоприятны для вида условия обитания. Состав видов определя-
ется численностью. Отсутствие вида, недавно обитавшего в данной
местности, свидетельствует о снижении его численности и несовер-
шенстве методов отлова. Например, водяная крыса — обычный и
часто доминирующий представитель грызунов Барабы — в 1982 г. >.
на Ровенском стационаре почти не улавливалась канавками (см. s
табл. 1). Животные в небольшом количестве были сосредоточены в ,
стациях переживания — по берегам некоторых озер. Отсутствие
особей в уловах 2 говорило об исключительно малой численности ,
вида. I
Сообщество грызунов — система, объединяющая все биотопиче- -
;ские группировки животных. Хотя оно состоит из таких группировок и й
^обладает их свойствами, но имеет и собственные признаки. В случае
анализа колебательных процессов предметом исследования становят-
!ся периоды собственных колебаний в сообществе. Оценка такой пе-
риодичности проведена нами по валовому ,сбору всех видов грызу-
нов во всех биотопах в период серии с 1966 по 1972 г. и окончания
следующей серии сукцессии — с 1978 по 1981 г. 1 '
2 Соблюдая форму описания, мы ежегодный видовой состав сообщества бу-
дем определять числом видов, фиксированных ловчими канавками, считая его
дополнительной характеристикой при оценке цикличности численности сооб-
щества.
i
I ' '
114 if
В 1966 г. в сообществе было И популяций грызунов. Отсутст-
вовали в отловах только рыжая полевка, обычный для Барабы вид,
и редкий, здесь джунгарский хомячок. Этот обедненный видов'ой сос
тав характерен для 5-го года после пика численности'водяной крысы.
Число циклов численности в этом гбду невелико (рис. 28, I, а), спектр
их беден пиками. Хорошо выражен ритмсс периодом в,60 сут, он до-
минирует. Этот ритм был наиболее характерными колках. Заметные
недельные и 2-суточные периодичности подвижности животных де-
лают спектр ритмов этого года для валовой численности похожим па
цикличность попадаемости грызунов в экотоне. > ' ! i
Количество видов в отловах 1967 г. было максимальным для|э!б- 1
го района1 Барабы. В колках появляется в заметном количестве ры-
жая полевка. Число мощных пиков на спектре численности и под-
вижности грызунов возрастает в сравнении с 1966 г. (см. рис. 2)8,•
I, б). Цикличность численности животных в этом году не прояви-
лась. Наиболее низкочастотный 24-суточный ритм характерен для
подвижности, обусловленной хоДом|>климатических факторов. Близ-
кие по мощности 2- и 1-недельные [циклы также характеризуют под-
вижность зверьков. Появляются заметные высокочастотные состав-
ляющие подвижности. Все эти циклы подтверждают пространственное
перераспределение грызунов. Численность зверьков возрастала, но
не циклично — сборы в течение вегетационного периода фиксирова-
ли постоянный ее рост от весны к осени. Цикличность функциониро-
вания сообщества в этом году по набору периодических составляю-
щих более всего похожа на таковую группы грызунов в колках.
В 1968 г. снизилось видовое разнообразие грызунов. Из обычно-
го набора видов в отловах отсутствовала южная мышовка. Спектр
ритмов сообщества имеет значительное число периодических [состав- 1
ляющих (см. рис. 28, I, в). Прежде всего для него характерен мощ- ।
ный пик с периодом 80 сут. По величине доминирует месячный цикл.
Из прочих ритмов заметные мощности у 4—5- и 2,5-суточных. В об-
щих чертах спектр напоминает прошлогодний. Из биотоцических
группировок близким набором периодических составляющих с1 похо-
жим соотношением мощностей обладает сообщество грызунов, биото-
па «колок». . И ' ' '
Основные цикличности численности [и подвижности грызунов из-
менились в 1969 г. Цикл численности .становится 60-суточным
(см. рис. 28, I, г). Из ритмов подвижности йо мощности преобладают [
2-недельный и 4-суточный. Изменения спектра в сравнении с преды-
дущим годом невелики, основная мощность [сместилась в 2-недель-
ную полосу .частот, 3—4-суточные ритмы, [мало изменились и сокра-
тилась мощность 2-суточных периодических составляющих. Спектр
ритмов почти такой же, как в группировке грызунов биотопа «поле».
Видовое разнообразие в этом году было максимальным — 13 видов
мышевидных грызунов. С этого года началось нарастание увлажнен- ,
ности Барабы. । [ 1
В 1970 г. на фоне продолжающегося увлажнения территории
видовое разнообразие в сравнении с предыдущим годом не измени-
лось. Цикличность сообщества изменилась сильно. На спектре от-
[
8*
115
Рис. 28. Спектры ритмов численности и подвижности сообщества гры-
зунов в окрестностях с. Ровенское.
i Усл. обозн. см. рис. 15,
сутствуют пики циклов численности длиной 40 сут и более, нет'и
самых коротких 2-суточных ритмов (см. рис. 28, I, д'). Общая карти-
на цикличности более всего напоминает соотношение ритмов в биото-
пах «луг» пли «болото» в этом же году (см. гл. III). Отсутствие мощ-
ных пиков в высокочастотной области спектра свидетельствует о том,
что спектр стал более устойчивым. Преобладающие ритмы сообщест-
ва в этом году 4-суточные, значительной мощностью обладают 6,- i
и 24-суточные; > i
Начавшийся подъем численности водяной крысы в 1971 г. по-
влиял на видовое разнообразие, число видов сократилось до 11.
В отловах отсутствовали джунгарский хомячок и южная мышовка —
грызуны остепненных ландшафтов. На сокращение числа видов мо-
жет влиять не столько доминирование водяной крысы, сколько вся
природно-климатическая обстановка, в частности заболачивание лу-
гов в низинах. ч
Резко изменился вид спектра в сравнении с предыдущими года-
ми. Доминирует околомесячная периодичность подвижности зверь-
ков (см. рис. 28, I, е). Хорошо проявлен пик 120-суточной циклич-
ности численности. Из высокочастотных заметной мощностью обла-
дает 2,5-суточная периодичность. Характерной для 1971 г. была
большая схожесть спектров цикличности во всех биотопических
группировках и по сумме отловов. ;|
В 1972 г. значительное обводнение территории совпало со вспыш-
кой массового размножения водяной крысы, что привело к значи- 1
тельным диспропорциям соотношений видов в сообществе грызунов <
и сильному сокращению видового разнообразия. Кроме отсутствовав- :
ших в уловах в 1971 г. перестала попадать в канавки пашенная полев-
ка. Остальные виды, исключая водяную крысу, были в сильной де-
прессии. Так, отловы на 89% составлял один вид — водяная крыса. i
Обычные доминанты сообщества: узкочерепная полевка — 1,2% от
всех пойманных , зверьков; полевка-зкономка — 0,65, полевая
мышь — 2,4% (см. табл. 1). - .
Таким образом, развитие этой серии сукцессии закончилось разру- >.
шением сообщества. Этому способствовал сам процесс развития его/ (
нарушающий видовые соотношения — нарастание численности во-
дяной крысы и депрессия прочих видов. Кроме того, такие условия
среды, как сильная обводненность территории, становились экстре-
мальными по отношению к подавляющему числу видов. После раз-
рушения сообщества на следующий год начала развиваться новая !
серия сукцессии. Пионерное сообщество состояло из 11 видов при
доминировании полевой мыши. '
Цикличность в .сообществе заметно изменилась в сравнении с
1971 г. Наибольшая мощность на спектре у колебаний подвижности — 7
3—5-суточных (рис. 29, I, а). Заметной мощностью,обладает циклич-
ность численности, период ее равен 48 сут, что соразмерно проме- '
жуткам времени между выходом и началом самостоятельной жизни
У молодых.водяных крыс. Значительные пики на спектре образованы :
2-суточными периодичностями. Это говорит, с одной стороны, об ин-
тенсивности территориальных перемещений грызунов, а с другой — f
117
Г
Рис. 29. Спектры цикличности в сообществе грызунов в разных сериях сукцес-
1 . i 1 1 сии-
j г 1 I ' Усл. обозн. см. рис. 16,
И' ' ] 1 ;
о неустойчивости спектра ритмов. Периодические изменения числен-
ности и подвижности животных всего" сообщества по виду спектра ;
ритмов; близки к луговым и i полевым биотопическим (группировкам,-
что связано с доминированием водяной крысы во всех биотопах. । J
В целом, рассчитывая циклические процессы, присущие этой
серии сукцессии, оказывается, что доминирующие сезонные циклы ’
сосредоточены в низкочастотной области спектра (см. рис. 29, I, а),,
незначительную мощность имели 2.0- и 5-суточные циклы подвпж-
; ности грызунов. При уточнении периодов цикличности в низкочас-
тотной области мы получили два доминирующих — 9,83 и 2,6 года.
Первый из них близок по значению колебаниям обводненности тер-
ритории Барабы. Такая1 периодичность составляет около 10 лет‘и
. । оцисаиа ниже (см. гл. V). Периодичность в 2,6 года соизмерима с
3-летними циклами изменений численности у полевок, неоднократно ,
^писанными в литературе 1 [Batzli, 1981;' Starka, Gerasimov, 1971; ..
' Старка, Герасимов, 1977; Матюшкин и др., 1972; Кошкина, 1969;
[Костенко, 1966], а также с 3-летними ритмами вегетации растений ;
i;! iTast, Kalela, 197'1]. ; i, > - [
ii i Спектры, рассчитанные для сообщества в этой, серии букцессии,-
похожи на полученные при расчетах колебаний численности в био- '.
топическйх группировках «поле», «луг», «экотон» (см. гл. III).
Следующая серия сукцессии началась в 1973 г..;Восстановление ,
сообщества грызунов и его стабилизацию мы наблюдали с 1978 г.
до следующей вспышки 'массового размножения в 1980 г. Видовое
разнообразие грызунов снизилось, в сравнении с предыдущей серией;
и составляло в 1973| г. 11, в 1974 г. 10 видов. Таким оно оставалось и .
к концу серии. i .
В 1978 г. сообщество насчитывало 10 видов мышевидных грызу-
нов. В уловах отсутствовали степная мышовка, джунгарский хомя-
118
чок и красно-серая полевка. Численность зверьков в этом году была
большой, п доминировало сразу несколько видов (см. табл. 1). Спектр !
циклов численности и подвижности грызунов насчитывал в этом году
много мощных пиков. Доминировала,цикличность с периодом около
недели (см. рис. 28, II, б). Значительная мощность была у 60-суточ- .
ной цикличности численности, которая присуща многим физцоло-
гичерким функциям животных, ia также их активности [Morosovsky,-
Melhyk, 1980]. Заметная мощность пиков 2:,5-сутдчных ;периодич«-
ностей. На спектре, такий образом, выделяются три области с мощ-
ными пиками, что. делает dro похожим на спектр 1968 г. Биотопиче-
ская группировка грызунов «луга» имеет спектр, ^близкий по набору
и соотношению мощностей периодических составляющих. '
В 1979 г.,возросло разнообразие видов, в уловах появился'джун-
гарский хомячок. Еще сказались последствия сухой фазы, несмотря
на наступившую влажную фазу. Численность животных изменялась
мало (сохранялась высокой).
Спектр ритмов численности и подвижности грызунов подобен
прошлогоднему, но мощных пиков на нем стало меньше (см. рис. 28,:
II, б). Доминирует 2-месячная цикличность численности, а следую-
щий по величине пин соответствует примерно недельным ритмам. I
Заметную мощность имеет 2-суточный ритм подвижности. Смещение
мощности в сторону низкпх частот и уменьшение числа крупных пи-
ков говорят о стабилизации, устойчивости спектра. Это последний
год перед вспышкой массового размножения водяной крысы, т. е.
наиболее устойчивый год данной серии сукцессии (общепринятое
мнение, что сообщество развивается в сторону наибольшей устой-
чивости). ।
В 1980 г. произошла очередная вспышка массового размножения
водяной крысы. По интенсивности она значительно уступала вспыш-
ке 1972 г. Водяная крыса доминировала в сообществе мелких мле-
копитающих, по субдоминанты (полевая мышь, обыкновенная буро-
зубка) мало уступали ей по численности. Доля водяной крысы в от-
ловах была незначительной в сравнений с предыдущей вспышкой —
36,1%. Нарушения в структуре сообщества были небольшими, коли-
чество! видов сократилось (не было в отловах джунгарского хомяч-
ка, а также степной мышовки и красно-серой полевки) и стало |Та-
ким же, как в 1978 г.
Заметно преобразился спектр ритмов в сообществе. На нем со-
вершенно не проявлены низкочастотные компоненты (см. рис. 28, i
II, в), он напоминает спектр грызунов 1967|Г. Доминирует бколоме-
сячная цикличность. Все прочие составляющие (их много) имеют зна-
чительную мощность и почти равномерно представлены на спектре —
от 2-недельной др 2-суточпой. Вид этого спектра близок к спектрам
группировож этого года в биотопах «экотон», «луг» и «поле».
К новой серии! сукцессии мы относим 1981 г. Видовое разнооб-
разие повышается в сравнении с 1980 г. незначительно, в отловах
появился джунгарский хомячок. ’ |
Спектр ритмов грызунов в этом году имеет два мощных пика, со-
ответствующих периодам в 10 и 2,2 сут (см. рис. 28, II, г). Пиков,.
119
соответствующих циклам численности (период 40 сут и более), на
спектре нет. Мощная 2-суточная цикличность свидетельствует о не-
устойчивости полипериодической структуры сообщества, что соот-
ветствует и нашим предположениям о начале формирования сооб-
i щества новой серии сукцессии. Похожие спектры ритмов имеют в
этом году биотопические группировки грызунов «экотона»'и «поля»
(см. гл. III). ; . ;
Оценивая вею серию сукцессий^ мы рассчитали для нее спектр
ритмов. Его высокочастотные составляющие маловыражены, некото-
рой мощностью обладают 5—10-суточные и месячные. Основную же
мощность имеет окологодовая цикличность (см. рис. 29, I, б). Доми- -
нирует по величине пик окологодовой цикличности. Рассчитывая и ..
уточняя количество и величину низкочастотных циклов в этой серии,, ,
видим, что ‘доминирует 3-летняя периодичность численности (см. -
рис. 29, II, б), более крупных по периоду циклов не обнаружено.
Достаточно хорошо выражен пик цирканной цикличности, а из высо-
кочастотных — 10-сутоЧный. Имеются на спектре 2-недельный и око- . .
помесячный ритмы.
Сравнивая спектры циклов в разных сериях сукцессии сообщест- -
ва грызунов, легко заметить их значительные различия. Они опре-
делены многими причинами. Прежде всего, каждая серия начинает-
ся своеобразным пионерным составом сообщества. Хотя серии обыч-
»но!начинаются после вспышки массового размножения водяной кры-
' сы, интенсивность вспышки, а часто и ее продолжительность бывают
различны. И, наконец, серия сукцессии тянется от 5 до 10 лет, в те-
чение которых климатические воздействия различны. Так, в первой
из рассмотренных серий вспышке соответствовала большая обвод- >
ненность территории, продолжавшаяся еще два года после нее. Во
второй же серии вспышка совпала с началом засухи, которая в те-
чение 2 лет после нее усугублялась. Засуха в Барабе обычно вызы-
вает депрессию численности грызунов, поэтому вспышка размноже- .
ния водяной крысы на этот раз не полностью была реализована.
Все это влияет на цикличность сообщества. Общие циклы в рас-
.смотренных сериях — приблизительно трехлетние и месячные. Во
второй серии гораздо больше ритмических составляющих? и при доми-
нирующей 3-летней хорошо выражена цирканная и 10-суточная.
Серии сукцессий имеют больше различий в своих периодических
составляющих, чем сходству. Каждая из них своеобразна, потому что
во время своего развития имеет неповторимый набор внутренних
структур и внешних воздействий.
Глава V
I
ЦИКЛИЧНОСТЬ ПРИРОДНОГО ФОНА
! ВСПЫШЕК ЧИСЛЕННОСТИ ВОДЯНОЙ КРЫСЫ
И ПЕРЕСТРОЕК ВИДОВОЙ СТРУКТУРЫ
,' . МЕЛКИХ МЛЕКОПИТАЮЩИХ
ХАРАКТЕРИСТИКА ЦИКЛИЧЕСКОЙ ИЗМЕНЧИВОСТИ СРЕДЫ
Смена фаз увлажненности Барабы] в 11-летних циклах
Трудность исследования смен фаз увлажненности Барабы объясня-
ется тем, что разные фазы не проявляются четко по суммам сезонных
и годовых атмосферных осадков. Они характеризуются прежде всего
степенью увлажненности территории и температурным режимом:
в периоды влажных, прохладных фаз все кочкарниковые болота,
лесные колки, различные понижения рельефа, низинные окраины
больших озер, мелкие плоские озера, речки заполняются водой.
Из-за обилия ^мелких водоемов появляется много комаров, слепней.
Влажные периоды сменяются сухими теплыми, и обстановка в Ба-
рабе резко отличается: низины обсыхают, уменьшается количество
гнуса, наблюдается размножение нестадных саранчовых [Куркин,
1976].
Механизм регулярно повторяющихся и сменяющих друг друга
многолетних .колебаний уровня грунтовых вод в северной лесостепи
Западной Сибири, видимо, тот же, что и описанный А. В. Шнитни-
ковым [1957] для внутривековых колебаний уровня воды в озерах,
его мы здесь специально не рассматриваем.
Смену фаз увлажненности Барабы в 11-летних циклах, а именно
их хронологию и длительность отдельных фаз, удалось расшифро-
вать по анализу вспышек массового размножения водяной крысы
[Максимов, 19826]. Здесь речь идет, следовательно, о методах ана-
лиза изменений среды с помощью биоиндикации. . i
Основой для анализа послужило то, что «болотный тип вспышек»,,
когда водяная крыса приносит большой вред зерновым культурам,
наблюдается в периоды высокой, а «озерный тип вспышек»— в пе-
риоды низкой обводненности территории Барабы [Максимов, 1959а,.
б, 1974, 1977]. По развитию этих типов вспышек массового размно-
жения водяной крысы и была выявлена хронология смен влажной
и сухой фаз. Эта цикличность к настоящему времени прослежена с
1925 по 1982 г. (рис. 30). За этот период по сменам болотных и озер-
ных вспышек массового размножения водяной крысы выявлена сле-
дующая хронология влажных прохладных и сухих теплых фаз:
влажных фаз было шесть (1926—1929, 1936—1940, 1946—1950, 1957 —
1961, 1969—1973, 1979—1981 гг.), сухих пять (1930—1935, 1941 —
1945, 1951—1956, 1962—1968, 1974—1978 гг.).
Обнаружена характерная связь развития влажных и сухих фаз
с показателями солнечной активности, выраженными числами Воль-
121
Рис. 30. Хронология.смен фаз увлажненности Барабы в 11-летних циклах за »
1925—1983 гг., установленная по биологическим материалам (по А. А. Мак-
симову [1977] с доп.). ।
1 — солнечная активность; 2 — влажная фаза; з — сухая фаза; 4 — годы болотных вспы-
шек водяной крысы.
фа; влажные фазы в предыдущие 50 лет совпадали с высокими пока-
зателями, а! сухие — с низкими [Максимов, 1966, 1971, 1977; Макси-
мов, Абашкин, 1976; Максимов и др., 1979]. Связь вспышек массово-
го размножения водяной крысы и солнечной активности опосредова-
। на через климат.
Все это послужило1 основой для долгосрочного прогнозирования ;
в Барабе периодов, неблагополучных по вспышкам массового размно-
жения водяной крысы и туляремии [Максимов, 1975, 1976]. Одно-;
1 временно было подтверждено наличие цикличности климата в лесо-
степной зоне Западной Сибири, что представляет большой интерес
для различных отраслей народного хозяйства [Максимов, Абашкину
1976; Максимов и др., 1979; Максимов, 1979а, б, 1981а, б].
При поисках объективных критериев разделения сухих и влаж-
ных фаз по гидрометеорологическим характеристикам и солнечной
активности А. Г. Сытин предложил использовать билинейную функ-
цию относительно среднегодового стока и среднегодового числа Воль-
фа. В качестве прогнозирующего признака оказалось возможным
указать узкий интервал переходных значений для выбранного ин-
декса WQ. Если индекс меньше | определенного числа, то мы имеем
дело с сухой фазой, если больше, то с влажной. Так, для Венгеров-
ского района Новосибирской области (р. Тартас, с. Венгерово) этот
интервал выражается числами: верхняя граница — 1108,7; ниж-
няя — 813,1. Индексы увлажненности рассчитаны для р. Каргат
(с. Гавриловское), р. Тартас (с. Венгерово), р. Тара (с. Малокраснояр-
ское) и р. Омь в'г. Куйбышеве [Максимов, Сытин, 1979; Сытин,
Максимов, 1982].*
122
В связи со сменой фаз увлажненности рассчитаны многолетние
колебания гидротермического режима Барабы [Токарев, 1982 ].' С по-
мощью нормированного отклонения дан анализ запасов продуктив-
ной влаги на глубине 20 и 50 см под основными сельскохозяйственны-
ми культурами за послёдние 20—30 лет. Установлено уменьшение
увлажненности в сухую фазу1 и повышение — во влажную. Просле-
жена некоторая периодичность увлажненности полей в многолетнем
аспекте, выделены 2,5-годовые, 5-летние и 11—12-летние циклы ув-
лажненности угодий, занятых зерновыми культурами [Николаев,;
Богомолова, 1982]. i । i ;
На основании анализа данных болотно-стоковых постов (с. Коно-
валовка Колыванского и с. Потюканово Северного районов) в водном
режиме болот (уровень грунтовых вод и продолжительность обвод-
ненности) на качественном уровне выделена цикличность, несколько
запаздывающая по отношению к фазам 11-летнего климатического
цикла. Эта цикличность обусловлена динамикой климата и, в свою
очередь, определяет структуру и динамику болотных биоценозов
[Панов, 1982].
Рассмотрены взаимосвязь брйкнеровской и 11-летней циклично-
сти климата и интенсивность проявления влажных и сухих периодов;
Приводятся примеры влияния смен увлажненности на сельское,
лесное, рыбное и охотничье хозяйства, а также в регулировании сис-
тем скрещивания в популяциях некоторых видов млекопитающих
[Абашкин, 1982].
Смена влажной климатической фазы сухой сопровождается адек-
ватными изменениями в растительности в направлении олуговения
болот, остепнепия, лугов, широки?.! распространение.м за счет солон-
цово-солончаковых лугов солонцеватых степей,- мезофитизацией за- ;
болоченных колков и другими изменениями, которые можно отнести |
к экзоэндодинамическим сменам — сукцессиям [Куркин, 1976; Ва-
гина, 1982]. . .
На основании изучения многолетней динамики пушно-промысло-
вых млекопитающих Барабы установлена, связь между колебаниями
и.х численности и циклическими изменениями местного климата.
В динамике большинства видов пушных зверей существенное значе-
ние имеет характер фаз увлажненности в 11-летнпх1 климатических
циклах [Поляков, 1982]. ' * (
Рассмотрены циклические изменения гидрометеорологических
факторов в связи с колебаниями воспроизводства окуня в оз. Чаны.
Выявлена 11- и 36-летняя цикличность биологических показателей
популяции окуня, хорошо согласующаяся с 11-летними периодами
солнечной активности и 33-летними брикнеровскими циклами [Вос-
кобойников, 1982].
В связи с циклическими изменениями уровня воды исследованы
колебания численности язя в оз. Чаны. В динамике естественного
воспроизводства этого вида выявлены 2- и 4-летние циклы; в динами-
ке вылова — 10—13-летние и, вероятно, близкие к 30 годам циклы;
в динамике общего вылова рыбы — 11-летние и брикнеровские цик-
лы [Крайнов, 1982].
123
Получены данные о резком изменении населения паразитических
гамазовых клещей на территории Барабинской низменности при
смене сухой и влажной фаз в 11-летних циклах. Это связано с изме-
нениями населения зверьков-прокормителей [Давыдова, Глотов,,
1982]. Выявлены колебания серологического фона мелких млекопи-
тающих в отношении туляремийного микроба в связи с различной
увлажненностью Барабы [Федосов, Максимов, 1982].
Изучены динамика и структура заболеваемости туляремией среди
населения Барабы в зависимости от фазы увлажненности территории <
в 11-летних циклах. Показано, что в пределах зоны высокой вредо-
носности водяной крысы в Барабе существуют циклически повто-
ряющиеся изменения в динамике, уровне и структуре заболеваемо-
сти населения туляремией. Основные мероприятия по вакцинации
населения в Барабе должны проводиться в заключительные годы су-
хой фазы с целью предупреждения заболеваний в наиболее опасные
по туляремии влажные годы [Букин, 1982].
Все эти исследования подтверждают наличие в Барабе многолет- !
ней природной цикличности с периодом около 11 лет и являются ос- ,
новой для уточнения и изучения цикличности в структуре и жизне-
деятельности биогеоценоза региона.
Приводим дополнительные свидетельства и доказательства су-
ществования на севере лесостепной зоны Западной Сибири смен фаз
увлажненности в 11-летних циклах, привлекая главным образом ма-
териалы по динамике численности водяной крысы. Эти материалы
вместе с тем характеризуют и биотические последствия описываемо-
го явления.
Период развития болотной вспышки
массового размножения водяной крысы как показатель
длительности фазы большой увлажненности Барабы
На территории лесостепной зоны Западной Сибири, составной ча-
стью которой является Барабинская низменность, выявлена и окон-
турена площадь, на которой развивались во влажную фазу болот-;
ные вспышки массовых размножений водяной крысы. В этой зоне
вредоносности [Максимов, 1967] вспышка может продолжаться до
4—6 лет. Период с появления первых локальных очагов размноже-
ния водяной крысы до исчезновения последних интенсивных очагов
на этой территории можно лишь условно принимать за период разви-
тия влажной фазы обводненности. Для расчета продолжительности
влажной и сухой фаз приходится привлекать дополнительные дан-
ные. Особенно важно уточнить методы обнаружения начала развития
влажной фазы, так как последний ее этап, когда происходит смена
влажной фазы на сухую, фиксируется достаточно четко по биологи-
ческим и климатическим показателям.
Ниже анализируется продолжительность влажных и сухих фаз
в Барабе на основе развития вспышек массового размножения водя-
ной крысы в 1957—1962, 1970—1973 и 1979—1982 гг. (табл. 19);
Массовое размножение водяной крысы в первый год развития
124
Таблица 19
Ход развития и продолжительность болотных вспышек массового раз- .
множения водяной крысы в Барабе
Период раз- вития вспыш- ки Площадь, охваченная очагами* (тыс. га) в год
1-й 2-й 3-й 4-й 5-й 6-й
1957—1962 138 ; 1056 1620 1350 2019 340
1970-1974 122 503 2600 1230 45 —
1974-1982 600 1500 1000 50 । — • —•
* Учитывается вся площадь угодий, в 'пределах которых размещались оча-
ги водяной крысы [Максимов, 1967 J.
вспышки (такими были 1957, 1970 и 1979) не несет еще черт типично
«болотных вспышек». Характерными в этом отношении были очаги,
проявившие себя в 1979 г. на севере Убинского района. Они носили
черты, переходные от сухой к влажной фазе. Наряду с зимовками
водяных крыс в обсохших болотных понижениях (что типично для
сухой фазы) оюенью 1979 г. отмечались зпмовочные поселения водя-
ной крысы на,лугах, полях, в разреженных колках и на пустошах
(что характерно для влажной фазы).
I Появление первых очагов «болотного типа» (осенний выход водя-
ной крысы на различные типы лугов и поля) следует считать началь-
ным этапом развития в Барабе очередной влажной фазы. Очаги
имеют еще локальное распространение.
Год, когда отмечается резкое сокращение площади и интенсивно- '
сти болотной вспышки и завершающийся полным ее угасанием,
обычно характеризует собой начало развития сухой фазы. Такими
были в Барабе 1962 и 1974 гг., это относится и к 1982 г., который так-
же был сухим, типичным уже для сухой фазы. Очаги водяной крысы ।
еще проявляли себя не только в обводненных стациях переживания,
но и на заросших сорняками пустошах, по окраинам кочкарниковых
болот, на посевах многолетних трав, особенно и полосе, прилегаю--
щей к займищам; 1982 г. следует считать в целом по Барабе годом
завершения вспышки, но уже относящимся к сухой фазе.
Если период интенсивного проявления влажной фазы рассчиты-
вать с учетом сделанных замечаний, то продолжительность влаж-
ной фазы в Барабе была равна: в первом случае (1957—1961 гг.)'
.5 годам, во втором (1969—1973 гг.) — 5 годам, в третьем (1979—
1981 гг.) — 3!годам. ।
Сокращение периода развития вспышки массового размножения
водяной крысы в 80-х годах мы объясняем тем, что текущая влажная
фаза увлажненности Барабы в 11-летнем цикле совпадает с регрес-
сивной фазой озер в 35-летнем цикле. Последнее привело к меньшей
и более краткой общей заболоченности территории Барабы.
Очевидно, именно такой была и вспышка в середине 30-х годов
(1936—1940 гг.), когда влажная фаза в 11-летпем цикле приходилась
125>
I
иа очень низкий внутривековой уровень воды в озерах. Ретроспек-
тивный анализ этой вспышки всегда был сложен из-за недостаточных ’
данных в Дитературе [Максимов,. 1959, 1971]. Сейчас ясно, что дан-
ная вспышка была кратковременной.
Наиболее интенсивными и имеющими наибольшую площадь рас-
пространения! в Барабе и в целом в лесостепи Западной Сибири бы-
вают болотные вспышки массовых размножений водяной крысы при
i трансгрессивной фазе озер. Такой была вспышка в конце 40-х годов,
по развитию которой была установлена зона вредоносности этого
вида в Западной Сибири [Максимов, 1967]?
Если ориентироваться на гидрологический .прогноз уровенного
режима озер, данный А. В. Шнитниковым и предсказывающий оче-
редное: повышение уровня воды в озерах Западной Сибири с конца
второй половины 80-х годов, то очередная вспышка массового раз-
множения водяной крысы должна быть более интенсивной и продол- ;
жительной, чем предыдущая. . >
Помимо большей скоротечности наблюдался сдвиг пиков к нача-
I лу вспышки: в 1957—1962 гг. пик отмечался на 5-й, в 1970—1974 гг.—
на 3-й и в 1979—1981 гг,— на 2-й годы влажной фазы (см. табл. 19). ।
; Наибольшую площадь очаги вредителя занимали в 1961, 1972 и
1 1980 гг. Два последних года в пределах соответствующих влажных
фаз отличались особенно неблагоприятными агроклиматическими ,
условиями, в результате чего в эти годы урождй яровых не созревал
до глубокой осени (1972 г.) и убирался для кормовых целей (1980 г.) :
В период вспышки 1957—1962 гг. таким неблагоприятным в агрокли- •
. магическом отношении был 1960 г., который предшествовал пику
вспышки [Максимов, Черанева,। 1979].
।: Приведенные материалы по развитию в Барабе «болотных» вспы- 1
щек массового размножения водяной крысы позволяют уточнить
длительность отдельных фаз увлажненности.1, <
। । . >
Перемещение очагов вспышки как показатель
неодновременности развития фазы большой увлажненности
При большой I однородности Северной Барабы в ландшафтном отно-
шении (характер местности, общность хозяйственного использова- .
ния, условия существования животных, пх видовой состав и др.)
популяции водяной крысы на этой территории неодновременно реа-;
; гнруют на смену фаз обводненности. Характерным показателем этого.
служат значительные- территориальные различия в сроках развития г
^очагов массового размножения. В пределах влажной фазы они воз-
никают и развиваются в разных районах Северйой Барабй неодно-
таременно.1
В ходе развития вспышки очаги массового размножения как бы ;
перемещаются по «зоне вредоносности». Прекратив свое существова-
ние в одном месте, они'возникают в другом; близкорасположенные
локальные очаги сливаются, а затем снова распадаются на отдель-
ные пятна, которые исчезают к концу | вспышки [Максимов и др.{
1965, 1976; Максимов, 1977]. .
<26
Рис. 31. Передвижение пиков кассового размножения водяной крысы
в Северной Барабе с 1979 по 1981 гг.
i 1
1 — 1979 г., 2 — 1980 г. (очаги затухли в 1981 г.), 3 1980 г. (очаги, действующие
в 1981 г.), 4 — 1981 г.. 5 —граница зоны вредоносности водяной крысы.
Пики массового размножения водяной крысы в Северной Бара-
бе в 1979—1982 гг. возникали неодновременно в разных местах.
Вспышка началась с появления нескольких локальных очагов на;
севере Убинского района (рис. 31). В 1980 г. очаги широко распро-
странились.по Центральной Барабе, а в 1981 г. они особенно прояви-
лись на западе и востоке Барабы. Заметна также тенденция смещения
очагов к северу от прежних мест их дислокации. 1
Неодновременность развития очагов массового размножения во-
дяной крысы в Северной Барабе может иметь разные причины. Не
исключена возможность того, что неодновременная реакция вида на
изменение условий существования объясняется наличпем большого
числа популяций этого вида в Барабе. Тогда зоны одновременного
развития вспышки характеризуют границы отдельных популяций.
Оставляя этот вопрос для будущего анализа, обратим внимание
на расшифровку обсуждаемого явления в связи с развитием фаз
увлажненности, а также дальнейшего обоснования и характеристи-
ки самих фаз.
Для долгосрочного прогноза развития вспышки массдвого раз-
множения водяной крысы важно прежде всего установить причины
территориальной приуроченности первичных очагов и факторы, ко-
торые определяют направление их последующего перемещения. То,,
что первые очаги водяной крысы появились на севере Убинского
района, возможно, связано с обилием здесь стаций переживания
этого вида в сухой период (мелиоративные каналы, различные низи-
ны со светлухо-вейниковым покровом и др.). Здесь существует более
127
к
4 -
к-\
5-
2
"1
О
Рис. 32. Многолетние из-
менения годового стока.
а — р. Каргат (с. Гаврилов-
ское); б — р. Омь (г. Куй-
бышев); в — р. Тартас
(с. Венгерово); г — р. Тара
(с. Малокрасноярское). К —
модульный коэффициент
стока.
гп
к -
пТк
нТ\.
2
тесная преемственность развития вспышки от условий, типичных
для сухого периода (озерных, озерно-болотных), до типичных
для влажного периода. Следовательно, начало развития очередной
вспышки больше оснований ожидать на участках, где сочетаются
озерные (регрессивная фаза озер) и болотные очаги. Следует учиты-
вать также и сроки завершения предыдущей вспышки на конкрет-
ном участке. Вероятно, там, где она завершилась раньше в преды-
дущий период, популяции лучщё подготовлены для развития очеред-
ной вспышки.
Следующее возможное объяснение причин первичной локализа-
ции очагов и их последующего перемещения по зоне вредоносности
заключается в неодновременном развитии фазы увлажненности на
территории Барабы. Мы не исключаем того, что движение очагов в
Барабе идет вслед за перемещением зоны большой обводненности.
Появление интенсивных очагов водяной крысы служит показателем
того, что на данном участке в этом году проявляется пик увлажнен-
ности.
Сказанное выше о несинхронном развитии обводненности тем
более вероятно, что здесь имеются бассейны различных рек (Чулым,,
Каргат, Омь, Тартас, Тара и др.), где.условия развития увлажнен-
ности могут быть неодинаковыми.'
Высказанное предположение отчасти подтверждают материалы 7
по стоку рек как показателю обводненности болот, за счет которых
питаются первые (рис. 32). Характерно, что если в Убинском(р. Омь)
«и Каргатском (р. Каргат) районах большой сток отмечался в послед-
428
Рис. 33. Многолетняя динамика уровня грунтовых вод в Северном
районе'(скважины 1—4) и в Убннском районе (скважины 5—8).
Климатические фазы: 9 —сухая, 10 — влажная. ,
ние годы лить в 1979 г., то в Северном, Венгеровском (р. Тартас)
и Кыштовском (рч Тара) районах он регистрировался в течение двух
лет (1979—1980 гг.). Это благоприятствовало более интенсивному
развитию в последних районах вспышки массового размножения
водяной крысы по сравнению с Кар.гатским районом (см. рис. 32).
Возможно, это объясняет и различия в сроках достижения пика
в центральной и западной частях Барабы, о чем.писалось выше. То,
что1 пик обводненности в Северном районе был в 1980 г., подтвержда-
ет динамика, грунтовых вод по четырем профилям в этом районе
(рис. 33). Наиболее высокий уровень грунтовых вод в последнюю фа-
зу увлажненности в Северном районе был в 1980 г. В Убинском райо-
не начало развития влажной фазы регистрировалось раньше и весь
период высокой обводненности был более растянут по времени Т
Этому соответствует п хронология’ развития местного очага водяной
крысы, интенсивная вспышка которого началась здесь в 1979 г.,,
продолжалась в 1980 г., в 1981 г. очаги уже прекратили свое
развитие.
Обращает на себя внимание характерная цикличность в разви-
тии пиков увлажненности Барабы и их неодновременное проявление
на территории (см. рис. .33).
1 Показательный график изменчивости уровня почвенно-грунтовых вод по
Убинскому району с пиками в 1948—1949 и 1958—1959 гг. имеется в работе
К. А. Куркина |1976]. Автор пшпет, что уровень почвенно-грунтовых вод от-
ражает в интегральной форме 11-летнпй климатический цикл.
q
А. А. Максимов, Л. II. Ердаков
1А»
Приведем дополнительные материалы по характеристике фазы
высокой обводненности в 1979—1981 гг. в Серерном районе.
В с. Усть-Ургулька .уже в 1978 г. отмечались признаки роста забо-
, лоченности 'территории, однако. дороги на дальние уч!астки земле- !t
пользования местного колхоза были в этом году еще хорошо проез-
жими; в, 1979, 1980 и 1981 гг. ,, отличавшихся наибольшей^заболочен-
ностью, вплоть до июля 1982 г. дороги стали непроезжими для авто- ’
машин. , • , |
О'Динамике обводненности территории можно судить также по '•
уровню воды в цилиндрах на ловчих,бороздках. Одна из них, рас-
положенная рядом с заболоченным низинным колком, наиболее про-.'
должительный срок была залита водой в 1979 г. (до конца июля). ; '
В 1980 и 1981 гг. эта бороздка использовалась для отлова зверьков-’,
с 20-х чисел итбля, а в 1982 г.— уже. с 8 июня. |
В 1981 г. кочкарниковые болота к середине лета Сильно обсохли, J
но травостой на лугах был густой и сочный. Водяная крыса, поки-
нув!с середины лета болота, осела на таких лугах и к осени их силь-
но изрыла. ।
Развитие очагов массового размножения водяной крысы на Устр-
Ургульском участке, было, однако, несинхронно с описанной выше
динамикой заболоченности территории. Последнее отражают резуль-
таты отлова водяной крысы на ловчих бороздках (на 100 цил.-сут):
1978 г, - 0; 1979 г, —0,74;. 1980 г.- 6,27; 1981 г, - 99,56; 1982 г.— ;
7,72. Водяная крыса достигла Здесь пика численности в 1981 г., т. е. 1
лишь на 3-й год с начала развития в Барабе влажной фазы. •
В Северной Барабе влажная фаза с 1979,г. развивалась с некото-
рыми особенностями из-за того, что она в своем 11-летнем цикле со-
четалась в этот период, как уже упоминалось выше, с регрессивной
фазой озер во впутривековом цикле. Такая ситуация предусматрива-
лась в экологическом прогнозе развития данной вспышки [Макси-
мов, 1977]. Описываемое своеобразие проявилось в общем снижении?
интенсивности и площади, охваченной повышенной увлажненностью.- -
Однако то обстоятельство, что последняя фаза высокой увлаж-
ненности Барабы совпала с регрессивной фазой уровенного режима
озер, имело и некоторые неблагоприятные последствия в проявле-
нии вредной деятельности водяной крысы: отмечался интенсивный
выход вредителя в некоторых хозяйствах поздней осенью па поля
озимых культур, что не отмечалось при предыдущих вспышках;
усиление вредной деятельности водяной крысы на лугах и лесных .
посадках. Это было связано с ранним выхЬдом грызунов на луга,
сильно изрытые за лето. К осени здесь создавались неблагоприятные-
условия для зимовки. Водяная крыса — влаголюбивый вид, и не-
достаток сочцых кормов из-за обсыхания болот вынуждал зверьков
искать его на полях. Заселению озимых всходов способствовала и
талая земля' поздней осенью 1980 и 1981 гг.
Завершение фазы увлажненности, судя ,по предварительным дан-
ным, на территории Северной Барабы наступает неодновременно^
В этом отношении был характерным 1981 г., когда в Новосибирской
области наблюдалась засушливая погода] которая привела к резко-
130
му снижению урожая, в тО время как на севере Барабы в этот год,
завершающий влажную фазу, были высокие урожаи Зерновых-
культур.
Характерно, что общая засуха 1981 г. проявилась на показате-
лях урожая зерновых культур.в Северной Барабе лишь в централь-
ных районах (Каргатский, Убинский, Чулымский), т. е. на той тер-
ритории, где началось^ массовое размножение водяной крысы.
На западе и востоке Барабы в 1981 г. влажная фаза все еще про-
должалась, по крайней мере до середины лета, что подтверждается
наличием пика численности водяной крысы. То, что лишь часть Се-
верной Барабы подверглась влиянию общей засухи, подтверждает
своеобразный климатический режим этого' региона. В 1981 г. Север-
ная Бараба еще находилась в фазе повышенной увлажненности, и
лишь 1982 г. характеризовался всеми признаками перехода к оче-
редной, сухой фазе.
., । ’ , . - ! • '
Развитие вспышки массового размножения водяной крысы
। |на локальных участках
1 как показатель степени увлажненности территории
Требует анализа явление неполного совпадения сравнительно про-
должительной длительности (3.—5 -лет) влажной фазы на конкрет-
ном участке территории и сравнительной краткости периода пика
численности водяной крысы (1 год). ।
Кратковременность периода пика подтверждается п при анализе
данных по отлову водяных крыс. Вспышка на локальном участке
характеризуется одним пиком с предшествующей низкой числен-
ностью и последующим ее резким спадом. Последнее нами иллю-
стрируется на примере развития четырех пиков массового размно-
жения водяной крысы на Ровенском, Усть-Ургульском, Потюка-
новском стационарах (рис. 34).
Начало заметного подъема численности свидетельствует о том,
что в следующем году может быть пик. На, стационаре в Потюканово-
этот перепад выражался следующими показателями отлова водя-
ной крысы (на 100 цнл.-сут): 1978 г.— 0,96; 1979 г.— 1,17; 1980 г,—
93,76 (нарастание: 11,2; 8,1); на Ровенском стационаре: 1969 г.—
0,27; 1970 г.— 0,58; 1971 г,— 4,18; 1972 г.— 20,47 (нара-
стание: 2,0; 7,2; 4,8). При следующей вспышке;- 1978 г,— 1,86;
1979 г.— 2,14; 1980 г.— 8,80 (нарастание:. 1,1;- 4,1).
После пика отмечается резкий спад, численности популяции.
На Ровенском стационаре в обоих случаях он наблюдался в год пе-
рехода от влажной к сухой фазе, когда в середине лета кочкарнп-
Ковые болота начинали резко обсыхать. Такими были здесь 1973,
1981 гг. и завершающий год ырм вспышке в 1946—1950 гг. [Макси-
мов. 19591. Эго же наблюдалось на Потюкановском стационаре
(1981 г.) На Усть-Ургульском стационаре резкий спад численности
водяной крысы наблюдался в 1982 г., т. е. в год перехода от влажной
к сухой фазе.
8*
131
Рис. 34. Продолжительность периода нарастания, пика и спада численности во-
дяной крысы на конкретных участках в пределах влажной фазы (экз./
/100 цил<сут).
а; б — с. Ровенское, в — Усть-Ургулька, г — Потюканово.
Таким образом, чем позже на участке развиваются очаги массово-
го размножения, тем больше они смещаются к концу влажной фазы
в целом по Барабе.
На остальной территории Северной Барабы, потенциально опас-
ной по болотным, вспышкам массового размножения водяной крысы,
но не охваченной вспышкой в предыдущие годы, очаги с наступле-
нием сухой фазы не реализуются.
Описанный характер быстрого достижения пика и последующе-
го резкого спада численности популяции, т. е. кратковременность пе-
риода вспышки массового размножения, объясняет причины, по ко-
торым водяная крыса на конкретном участке проявляет себя как
вредитель сельского хозяйства чаще всего лишь в течение одного
года.
Тот факт, что влажная фаза длится 3—5 лет, а пик вспышки
массового размножения на локальном участке — всего 1 год, пред-
ставляет и общий биологический интерес в исследовании механиз-
мов динамики численности животных. Речь идет о том, что вспышка
массового размножения бывает короче периода развития благопри-
ятной для нее обстановка, и крах популяции часто происходит тог-
да, когда на данном участке еще имеется оптимум условий для су-
ществования вида. Последнее, однако, относится к очагам, возник-
шим в начале развития вспышки. Судьба очагов, возникших позже*
иная. Нередко они прекращают существование, не достигнув пика
(«срезанный пик»). Такова в Барабе судьба всех очагов, проявляю-
щих себя во второй половине влажной фазы. Очередная сухая фаза,
препятствующая высокому воспроизводству вида, наступает рань-
ше, чем популяция достигла пика. Это одна из причин часто наблю-
дающихся низких пиков численности. '
При характеристике года пика численности водяной крысы сле-
дует уточнить, о каком пике идет речь. Пик может быть полностью
132
реализованным, когда после его завершения природная обстановка
остается прежней, т. е. благоприятной для интенсивного размно-
жения популяции. Причиной краха популяций может быть в этом
случае, например, эпизоотия, сочетающаяся с каким-либо иным не-
благоприятным для популяции фактором. Пик может быть не пол-
ностью реализованным, когда численность, не достигнув кульми-
нации, снижается. Этот вариант обычен в Барабе, и его наиболее
частая' причина — наступление сухого года, когда болота пересы-
хают и условия существования водяной крысы резко ухудшаются.
Поскольку пик численности продолжается один год, он хорошо
отличим по величине от года предпика и года, следующего за пиком1.
Судить о том, достигла ли численность кульминации или пик был
«срезан», можно по его высоте. Так, уровень численности водяной
крысы на Ровенском стационаре в 1980 г. есть основания считать
не очередным пиком, который здесь ожидался в 1981 г., а, видимо,
предпиком. Наступившая в 1981 г. в Каргатском районе засуха не
позволила реализоваться вспышке в полной мере[
Вместе с тем уровень численности водяной крысы на Усть-Ур-
гульском стационаре в 1981 г. был очень высоким и носил все чер-
ты полностью реализовавшегося пика (см. рис. 34, в), хотя здесь
с середины лета болота резко обсохли и размножение водяной кры-
сы прекратилось. Пик полностью реализовался за счет интенсив-
ного размножения водяной крысы весной и в первую половину лета.
Сходство структурных перестроек
в сообществе мелких млекойитающих,
связанное с цикличностью увлажненности территории
и вспышками массовых размножений водяной крысы
Для анализа экологической сукцессии сообществ, а также цикли-
ческих процессов удобно использовать видовую структуру сообще-
ства. Изменение численности в популяциях разных видов одного
сообщества происходит различно, в зависимости от их экологиче-
ских требований. Виды группируются, скажем, по зеленоядности и
одновременно по отношению к влажности местообитания. Распре-
деление видов в сообществе по их требованиям к среде (параметрам
экологических ниш) мы называем экологической структурой сооб-
щества. При сходстве популяций любой из характеристик ни-
ши они объединяются в группы (зеленояды, гидробионты, лесные
виды и т. п.).
Численность — обобщенный показатель, она изменяется под вли-
янием большого числа действующих на популяцию факторов. Оди-
наковый ход численности у двух популяций свидетельствует об их
экологической близости. Мерой близости динамики численности
двух видов может быть коэффициент корреляции. Корреляционные
связи между видами дают представление о близости требований к
среде у разных видов, и совокупность таких связей описывает эко-
логическую структуру сообщества [Ердаков и др., 1980].
133
Экологическая структура сообществ определялась для каждого
года серии сукцессии, начиная с 1968 г. Рассчитывалось сходство
динамики численности видов в-течение вегетационного периода.
Каждая популяция за этот период в несколько этапов рамножения
увеличивает число особей, когда этому благоприятствуют условия
среды. Естественно предполагать, что связь (корреляционная) ме-
жду динамиками популяций говорит о примерно одинаковых воз-
можностйх роста численности. Изменение роста, потеря Связей оз-
начают флуктуации в сообществе, способствующие-нарушению эко-,
логической структуры. Для сообщества грызунов в Барабе в период
с 1968 по 1972 г. характерна определенная структура связей.между *
видами (рис. 35). j I
После массового размножения в 1961 г. вплоть до 1968 г. еже- -
годно в сообществе были 1—2 устойчивые корреляционные связи
между особенно многочисленными видами. В 1968 г. группа видов| •
уже была представлена в виде некоторой экологической структуры
(рис.. 36, а). Малочисленные популяции водяной крысы, полевой
мыши и рыжей полевки не имели корреляционных связей с числен-
ностью других популяций сообщества. '
| Значительная плотность — наиболее частое, но не единствен- ,
I ное условие корреляционной связи между динамиками популяций.
Так, в 1969 г. статистическая структура сообщества грызунов была
довольно полной, но отсутствовали связи у красной и темной поле-
вок. Оба вида имели малую численность. В то же время многочис-
ленный вид — лесная мышовка — не был связан, как обычно,, с
красной полевкой, а малочисленные виды — пашенная полевка и
водяная крыса — имели высокую корреляционную связь изменения
численности (см. рис. 36, б).
В 1970 г. экологическая структура сообщества была полной.
Начавшийся цикл обво'дненйя территории положительно повлиял;
на рост численности всех видов грызунов, и корреляция изменений
численности наблюдалась между всеми популяциями (см. рис. 36, в) Л
С ростом численности водяной крысы в 1971 г. начались нару-
шения экологической 'структуры сообщества мелких млекопитаю-
щих. Пропала связь насекомоядных с полевой мышью/хотя числен-
ность последней была велика, нарушилась связь их и с лесной мы-
шовкой (см. рис. 36, г). У последней уже в 1971 г. в период размно-
жения водяной крысы наступила, депрессия. Вспышка численностй
водяной крысы в 1972 г. привела к потере всех корреляционных свя-
зей между популяциями мелких млекопитающих (см._ рис. 36, д).
Таким образом, в период между вспышками массового размноже-
ния водяной крысы происходит восстановление сообщества, и сло-
жившаяся экологическая структура разрушается очередной вспыш-
кой. Восстановление экологической структуры сообщества происхо-
дит долго. В рассмотренной серии сукцессии на это понадобилось
почти 6 лет. Только перед самой вспышкой размножения водяной
крысы структура' сообщества становится полной. Можно предполц-
жить, что каждая серия сукцессии знаменует приблизительно оди-
наковый цикл экологической структуры сообщества. По мере вос-
134
Рис. 35. Значимые корреляционные связи между видами и группами мелких
млекопитающих в Барабе.
Рис. 36. Структура экологических
связей мелких млекопитающих.
а — на 7-й год после вспышки массового
размножения водяной крысы .(1968); б —
на 8-й год после вспышки (1969); в — за
2 года до новой вспышки (1970); г — за
1 год до вспышки (1971); 0 — отсутствие
связей в год вспышки (1972). 1 — полевая
мышь; 2 — водяная крыса; 6 — красная
полевка; 7 — северная (лесная) мышовка;
8 — темная (пашенная) полевка; е ры-
жая полевка. ,
Рис. 37. Изменение по годам индекса общности видового состава
мелких млекопитающих в экологической сукцессии.
। А, Б — серии сукцессии.
становления его после очередной вспышки постепенно складыва-
ется структура и к следующему подъему численности становится на-
иболее полной и устойчивой. Начавшийся подъем численности водя-
ной крысы нарушает структуру, а вспышка следующего года унич-
тожает ее полностью1. Здесь при разборе динамики экологической .
структуры сообщества с определенностью видна взаимосвязь сук-
цессий и цикличности. Внутри многолетнего цикла смены домини-
рования происходит экологическая сукцессия. Она приводит к по-:
явлению устойчивого сообщества, разрушаемого началом очеред-
ного цикла.
Иллюстрация -цикличности развития сообщества и связанной с
нею сукцессии подтверждает одно из современных представлений
о динамике экосистем — представление о широком распространении
и разномасштабности сукцессий, когда в рамках отдельных ценозов
постоянно осуществляется множество видов различных сукцессий
разной продолжительности и масштабов [Чернова, 1977].
Дополнительные сведения, подтверждающие результаты анали-
зов экологической структуры сообщества, мы получили при рассмот-
рении общности видового состава в разные годы и в разных сериях
сукцессии. Это 'часто используемая характеристика рассчитывалась
нами с помощью индекса Жаккарда (цит. по [Песенко, 1982])..
Оценив общность 'видового состава, можно сравнивать не только
различные! сообщества, но и изменение одного в разные годы иссле-
дований. Изменение видового состава в сообществе происходит под
влиянием разных экологических причин. Обычно такие различия
определяются ходом сукцессии.
Мы рассмотрели изменения видового состава сообщества грызу-
нов в районе с. Ровенское за ряд лет и отметили цикличность такого
рода изменений (рис. 37). Большой общностью по этому признаку
обладают соседние годы в серии, такая 2-летняя цикличность про-
является, в частности, и в сохранении доминанта. Обычный срок дб-
136
минирования в местном сообществе грызунов 2 года. Внутри серии
отмечены 5- и 7-летние циклы. В серии сохраняется значительная
общность видового состава. Она разрушается в год вспышки числен-
ности водяной крысы. При расчетах в 1972 г. сообщество оказалось
совершенно своеобразным по видовому составу.
Включив в анализ результаты работ в 1973 и 1974 гг., мы рассчи-
тали общность видового состава сообщества предыдущей серии сук-
цессии (см. рис. 37). Оказалась, что-высокая общность только у со-
обществ 1965 и 1973 гг. Высокая общность нередко обнаруживается '
в разные годы смежных серий, особенно при депрессии и в пере-
ходные периоды. Это требует дальнейшего исследования.
Таким образом, рассмотрев общность видового состава в разное
годы и в различных сериях сукцессии, мы пришли к выводу, что
в каждой серии существует несколько циклов развития сообщества.
Обычные периоды таких циклов 2- и 5-летние'. Вполне вероятны
9-летние цикличности, объединяющие сообщества разных серий по
признаку общности видового состава,
ПЕРЕСТРОЙКА ВИДОВОЙ СТРУКТУРЫ СООБЩЕСТВ
В СВЯЗИ СО СМЕНОЙ ФАЗ УВЛАЖНЕННОСТИ
В БАРАБЕ В 11-ЛЕТНИХ ЦИКЛАХ
Видовой структурой сообщества мы называем ранжированный по
численности состав популяций. Видовая структура сообщества в
каждый сезон может быть представлена в виде градации населения
этого сообщества от доминантов до самых малочисленных видов.
В отличие от экологической видовая структура не предполагает
подобия в динамике численности у видов или близости по этому при-
знаку. Виды, близкие по видовой структуре, могут быть совершенно
различны по экологической. Для описания видовой структуры до-
статочно иметь сведения о суммарной численности видов за время
наблюдения. Виды, обладающие большой численностью (превосхо-
дящие по ней всех остальных), обычно называются экологическими
доминантами [Одум, 1975]. Нами под доминированием понимается
величина относительной численности популяции. Виды-доминанты
и субдоминанты в каждом конкретном году иллюстрируют своеобра-
зие биотической и в какой-то мере абиотической обстановки в сооб-
ществе.
।
Видовые аспекты в населении мышевидных грызунов
Мы представляем видовую структуру 2 как следствие «ответа» сооб-
щества на сложившуюся природную обстановку. Смена доминантов
свидетельствует об изменениях экологической ситуации. Ретро-
2 В этом разделе речь идет о суммарной (общей) видовой структуре в лю-
бой из рассматриваемых точек. Подробно о видовой структуре сообществ раз-
личных биотопов в разные годы исследований см. гл. III.
137
спективные оценки, сделанные по смене доминантов в разные годы,
. могут помочь в определении тенденций развития рассматриваемого
биогеоценоза. Эти;соображения и легли в основу нашего анализа. ,
। Видовая структура грызунов рассмотрена по 37-годовым выбор-
кам (Ровенское — 16 лет, Потюканово — 7, Усть-Ургулька — 5,;
Кбноваловка (Орловка) — 7, Черновка — 1, Крещенка — 1). До-
минантами среди грызунов были: полевая! мышь — 14 раз, водяная 1
крыса ,— 11, полевка-экономка- — 8, : узкочерепная полевка — 2, 'л
мышь-малютка — 1, обыкновенная полевка — 1. Наиболее обыч- 1;
ными субдоминантами _были: лесная мышовка — 10 раз, полевая \
мышь — 6, полевка-экономка — 4, узкочерепная полевка — 5,- >“
красная полевка — 3, водяная крыса —-2, темная полевка — 2,
мышь-малютка — 2, обыкновенный хомяк, обыкновенная полевка, ?
рыжая полевка — по одному разу. ।
В поды, когда] дОминантом была полевая мышь, субдоминантами
были: лесная мышовка — 3 раза, красная полевка — 3, водяная
крыса — 2, полевка-экономка — 2, темная полевка — 1, узкочереп-
ная полевка — 1, рыжая полевка — 1, мышь-малютка — 1. "В годы
доминирования водяной крысы субдоминантами были: полевая
'мышь — 5 раз, лесная мышовка — 2, темная полевка — 1, узкоче-
репная полевка — 1, мышь-малютка — 1, обыкновенный хомяк — 1. ' .
<В годы доминирования полевки-экономки субдоминантами были: >
лесная мышовка — 4 раза, узкочерепная полевка — 3, обыкновен- ,
ная полевка — 1. При доминировании узкочерепной полевки суб- i;
доминантами по одному разу были лесная мышовка и полевка-эко- .
номка. У мыши-малютки в год доминирования субдоминантой была
полевая мышрь, . а у обыкновенной полевки,— полевка-экономка. ’
В перечне доминантов и субдоминантов есть общие виды. Так,
у водяной крысы субдоминантам была полевая мышь (5 раз), а у •,
полевой мыши — водяная крыса (2 раза)..
Если ориентироваться на вид-доминант и учитывать частоту со--
пряженных с ним субдоминантов, можно сказать, -что для Барабы
наиболее характерны следующие видовые аспекты грызунов, в ко-
торых роль доминанта и субдоминанта может взаимно меняться: ,
водяная крыса + полевая мышь — 7 случаев; полевая мышь -f-
лесная мышовка — 3; полевая мышь + красная полевка — 3; по- ‘
девая мышь -J- полевка-экономка — 2; полевка-экономка. -1- лесная :
мышовка — 4; полевка-экономка узкочерепная полевка — 4; по- '
левка-экономка + обыкновенная полевка — 2. *
Характерно, что, исключая полевую мышь, i водяная крыса не
имеет постоянных субдоминантов, ими могут быть те или иные ма- %
Исчисленные виды. ।
Выявлено 25 случаев из 37 годовых выборок (67,5%), когда со-,
хранились описанные выше 7 годовых аспектов,- что говорит об их
i типичности для Барабы. Однако при этом варианте выделения ви-
довых аспектов остались вне анализа 12 выборок. '
С целью более полного обобщения собранного материала сдела—
; на попытка выделить видовые аспекты по трем первым видам в со-’
обществе грызунов. Так, при анализе аспекта водяная крыса + по-.
138
экз. на ЮОц.сут
1 10 7 3 4 2 1 7 9 4 2 1 4 2 7 3 6 4
2 5 6 8 9 53 8 6 11 7 5 8 19 8 5
•2 7 9 4 11
\ 1 3 8 6 14
| Рис. 38. 1-й годовой аспект грызунов Барабы: водяная крыса — полевая
МЫЩЬ. I
Ровенское: а — 1950, б —. 1971, в — 1972, г — 1973, д — 1978, е — 1979, ж — 1980,
з — 1981; Потюканово: и —> 1979, к — 1980, л — 1981; Усть-Ургулька: м —, 1980, н —«
1981, 0 — 1982; Коноваловка: п — 1979; Орловка: р — 1980; Черновка: о — 1974.
1 — полевая мышь; 2 —• водяная крыса; 3 — полевка-экономка; 4 —. узкочерепная по-
левка; 5 — мышь-малютка; 6 — красная полевка; 7 — северная (лесвая) мышовка; « —
темная (пашенная) полевка; 9 — рыжая полевка; 10 —. красно-серая полевка; 11 —«
обыкновенный хомяк; 12. — южная (степная) мышовка; 13 —. джунгарский хомячок; 14 —<
обыкновенная полевка.
левая мышь заметно, что эта пара нередко представлена в числе пер-
вых трех видов, причем она перемежается постоянно меняющимся
третьим видом. Такое -объединение видов по двум общим в первой
ранговой тройке и взято нами в основу выделения видовых аспектов
13S
экз.оа'ЮОц- сут
\4 1 3 10 8 11 12\ 9 3 6 4 13 12 1 4 6 5 10 7 9 4 3 в’ \
7 \6 9 5 2 13 \1 7 5 8 2. 11 10\ 7 9 3 8 10 6 5 2 14
Рис. 39. 2-й годовой аспект грызунов Барабы: полевая мышь + лесная мы-
шовка.
Ровенское: а — 19ff9, б — 1982; Усть-Ургулька: в — 1978, г — 1979. Остальные усл. обозн.
см. рис. 38.
грызунов, характеризующих животные сообщества мелких млеко-
питающих Барабы.
Ниже перечислены видовые аспекты мышевидных грызунов в
Барабе, в основу выделения которых положен принцип наличия
двух общих видов в первой ранговой тройке данного годового набо-
ра при условии, что один из этих двух видов представлен как доми-
нант.
Видовые аспекты следующие:
1. Водяная крыса -J- полевая мышь; представлен в 37 выборках
16 раз. Для этого сочетания весьма характерно присутствие( лесной
мышовки в качестве третьего (реже четвертого) компонента (в 9 слу-
чаях из. 16). К этому аспекту мы присоединяем также годовые наборы
экз. на 100 а, сут
9 10 11
2 12 13
5 8 6 10 7 "1б\ 3 1 6 5 2'
14 4 1 5 15 14 8 4 7 9 10
г 14 9 10'3 4 8 15
5 8 6 \ 7 5 9 11 10 14 ,
Рис. 40. 3-й годовой аспект грызунов Барабы: полевка-экономка -J- узкочереп-
ная полевка + лесная мышовка.
Ровенское: а — 1966, в — 1967, в —• 1968, г — 1970; Потюканово: д — 1976, е — 1977, ж —
1978; Коноваловка: з — 1974, и — 1975, к — 1976, л — 1977, л —• Крещенка, 1977. Осталь-,,
ные усл. обозн. см- рис. 38.
140
экз.наЮОц. сут
1 з а ти ч it 7 ч ы / iu ю о 14 з о о i ~ у , в \
6 2 1 3 12 5 6 3 2 8 12 1 5 7 11 10 4 5 6 \
Рис. 41. 4-й годовой аспект грызунов Барабы: мышь-малютка + пб-
। левая мышь.
Ровенское: а — 1965, б — 1974; в — Орловка, 1979; г — Потюканово, 1982. Усл.
обозн. см. рис. 38.
видов, где доминантом была водяная крыса при иных, чем в выше
указанных случаях, субдоминантах. В нашем примере такой вариант
представлен лишь в одном случае (с. Орловка, 1980 г.), где субдоми-
нантами были темная и обыкновенная полевки (рис. 38).
2. Полевая мышь (чаще всего доминант) Ц- лесная мышовка. За-
регистрирован в четырех случаях (рис. 39). Близок к первому аспек-
ту, но в нем среди трех- лидирующих видов отсутствует водяная кры-
са. Руководящей формой этого аспекта служит лесная мышовка.
3. Характерно присутствие в первой тройке во всех случаях в ка-
честве руководящей формы полевки-экономки (чаще всего доминант),
узкочерепной полевки (чаще всего) и лесной мышовки (часто). Такой
вариант отмечен в 12 случаях. В ряде случаев вместо узкочерепной
полевки среди основных членов сообщества представлены темная и
обыкновенная полевки (рис. 40). ’
4. В первой тройке присутствует (иногда доминант) мышь-малют-
ка (руководящая форма). Характерно присутствие также полевой
мыши — 4 случая (рис. 41).
Из выделенных четырех видовых аспектов грызунов в Барабе
чаще, всего наблюдается 1-й (водяная крыса + полевая мышь). Он
представлен в период наших работ (37 годовых выборок) в 45,9%;
2-й аспект (полевая мышь -j- лесная мышовка) отмечен в 10,8% слу-
чаев; 3-й аспект (полевка-экономка + узкочерепная полевка + лес-
ная мышовка) — в 35,1% случаев; 4-й аспект (мышь-малютка 4-
полевая мышь) — в 10,8% случаев.
i
Характеристика видового состава сообщества
в сухую и влажную фазы
Большая изменчивость природной обстановки, вызванная сменой
влажной и сухой фаз, создает преимущества в условиях существо-
вания то для одного, то для другого вида, приводя к перегруппиров-
кам видового состава сообществ. В табл. 20 приводится ранговая
141
Таблица 20
Частота доминирования различных видов грызунов в сухую (с) и влажную (в) фазу
Вид Фаза 1 Место в сообществе (ранг)
1-е; 2-е 3-е 4-е. 5-е 6-е 7-е 8-е 9-е 10-е 11-е 12-е. 13-е
Полевая 'мышь । с 5 'о 3 1 ol 11 0 1 1 '0 0 0 0 . >
1 I в 3 5 5 0 0 0 0 0 0 0 0 0 0
Водяная крыса с 0 0 1 1 3 0 1 0 2 2 0 0 0 .
в 10 2 1 0 10 0 0 0 0 0 б 0 0
Полевка-экономка С 4 3 1 1 4 1 1 1 0 0 0 0 0 г
I В 0 0 0 2 2 1 5 2 0 0, 1 О’ 0 0 о .
Узкочерепная полевка с 2 2 2 1 2 1 0 1; 1 0 0 0
в 0 1 0 0 2- 0 3 Г’ 5 1 0 0 0
Мышь-малютка ; G . 0 0 1 1 1 1 1^ 5 б ! 1 0 1 0 '
В 0 1 0 4 3 2 1 1 0 1 - 0 0 0
Красная полевка с 0 1 1 1 2 3 1 0 0 1 ' 1 0 0 . г
в 0 '1 0 2 1 4 1 3 0 0 0 0 0 ,
Лесная мышовка с 0 .5 1 2 0 1 1 0 1 0 1 0 •0 .
Полевки^ в 0 2 4 0. 4 0 1 1 I 0 0 0 0 0
пашенная с 0 0 2 1 2 0 2 2 2 0 1 0 0
в 0 0 0 2 0 4' 0 4 1 0 0 0
, рыжая с 0 1 0 1 0 4 1 0 •3 0 1 0 ' 0
i в 1 0 0 2 2 2 1 ’1 0 2 2 0 0 о
красно-серая с 0 0 0 0 0 0 2 .0 1 |3 ' 4 0 1 г
в 0 0 1 0 0 1 0 0 1 0 3 2 0
Обыкновенный хомяк с 0 0 0 1 1 0 1 1 0 0 3 0 о .
В 0 1 :0 0 0 0 0 0 3 5 2 0 0 е
Степная мышовка с 0 0 0 0 0 0 0 0 1 1 0 2 1
в 0 0 0 0 0 0 0 0 0 0 1 з. 1
Джунгарский хомячок с> 0 0 0 0 0 0 0 0 0 2 0 1 2
i 1 в1 0 0 0 0 0 0 0 0 0 0 2 2 3
Обыкновенная полевка с 1 г 0 1 0 0 1 0 0 0 0- 1 0
Мыши ' 1 в. 0 0 0 0 0 0 0 0 0 1 0 0- 0 .
лесная с 0 0 0 0 0 0 0 0 0 1 0 0 1 .
в 0 0 0 0 0 0 0 0 0 0 0 0 0
домовая: с 0 б 0 0 0 '0 0 0 0 ' 0 0 0 о
в* 0 0 0 0 1 0- 0 0 0 . 0 0 0 0 0
характеристика различных видов грызунов Барабы по относитель-
ной численности в сухую и влажную фазы. Показано место, которое
занимал по численности данный вид в отловах в соответствующий
Год влажной и сухой фаз. 'i
Влажная фаза характеризуется по сборам в окр. с. Ровенское.в.
1950, 1971, 1972, 1973, 1979, 1980 и в 1981 гг., с. Усть-Ургулька —
в 1980, 1981 и в 1982 гг., с. Потюканово — в 1979, 1980 и 1981 гг.
(рис. 42). Сухую фазу мы характеризуем по сборам в окр. с. Ро-
венское в 1965, 1966, 1967, 1968, 1969, 1982 гг.-, с. Коноваловка — в
142
.мзнаЮОц. cyw
\ 20,47
I2 7, 9, 3 4 \2" 1 ,5 3 9 10 13\ 11'7 6 3 .9- \1 7 5 4 9 11 13'
I 7 ! К 5 .6 11 4 в 6'7 11 12 \2 \ 1^4 5 Ю 12[- 2 10 3 8 12 ,
е
з
.29.46
7'3 13 12. -1 6 5 8 4 13 \2 9 13 '2
6 5 9 8 11 10 \2. 9. 7 3 11 10' 12x1 5 6 4 11.12\ 19 8
\127,53 " " "
8
\ 9 3,56
1 3 8 Ь 14 I
I м' й ’ н
h
4 11 17 10 7 3 4
3 8 5 < 2 5 6 8 .
7 9 4 \2
8 6 11 \
4 '
8\
Рис. 42. Видовые структуры населения грызунов во влажную фазу увлажнен-
। ности Барабы.
Ровенское: а — 1950, б —1 1971, а —1972, г" — 1973; д — 1979, е — 1980, ж — 1981; Усть-
Ургулька: а — 1980,: и — 1981, к — 1982; Потюканово: л — 1979, м — 1980, »•— 1981. Ос-
тальные усл. обозн- см. рис. 38.
1975,! 1976, 1977 и 1978 гг., с. Крещенка — в 1977, с. Усть-Ургуль-
ка — в 1978 г. (рис. 43). । j
Обнаруживается четкое разграничение видов, которые были до-
минирующими в разные фазы^ увлажненности Барабы. В сухую фазу
доминантами в ценозах грызунов были полевки: экономка, узкоче-
репная и обыкновенная, а субдоминантами, помимо перечисленных
видов, также лесная мышовка и красная полевка. Наиболее типичны
видовые аспекты, включающие полевку-экономку, узкочерепную по-
левку, лесную мышовку, полевую мышь. • i ।
Во влажную фазу типичным доминантом была лишь водяная кры-
са. Полевая мышь в равной степени является многочисленной как во
влажную, так и в сухую фазу. Интересно, что если во влажную фа-
зу этот вид всегда занимает одно из лидирующих мест в биоценозе,
то в сухую может значительно варьировать в ряду сообщества гры-
зунов.
С помощью критерия %2 проверена достоверность различия поло-
жения полевой мыши в структуре сообщества в зависимости от
143
экз.наЮОц. сут
6-i
о L
г
7 3 12
L7 11 4 12 10 I 4 в 6 5 10 12
д
\4 6 8 11 2 10
; J____7 9 1 13 5
4 13 10 8 11 12\1 1 5 8 2 11 10.
7_:.6. 9 5 2 13 \ 9 3 6 4 13 12 J
I 3
2 14 9 10
5 8 6
\3 8 6 10
' 14 4 1 5 15
к
I 7 14 9 5 10
6
7 5 9 11 10 14
7 9 3 8
Рис. 43. Видовые структуры населения грызунов в сухую фазу ув-
лажненности Барабы.
Ровенское: а — 1965, б — 1966, в — 1967, г — 1968, д — 1969, е — 1982; Коно”
валовкя: ж — 1975, з — 1976, и — 1977, к — 1978; Крешенка — л — 1977; Усть-
Ургулька: м — 1978. Остальные усл. обозп. см. рис. 38.
фазы увлажненности. На 5%-ном уровне значимости различий не -
обнаружено. Водяная крыса многочисленна лишь во влажную фазу,
в сухую редка, в спектре доминирования занимает.от 3—4 -го до-10-
го места. Это подтверждалось при расчетах достоверности различий
численности в зависимости от фазы увлажненности. С 99%-ной ве-
роятностью (критерий %2) можно сказать, что во влажную фазу водя- .
ная крыса занимает ведущие места в сообществе.
У полевки-экономки прослеживается обратная тенденция: бу-
дучи многочисленной в сухую фазу, она уступает место другим ви-
дам во влажную фазу, занимая по численности места от 4-го до 8-го.
Примерно то же отмечается и у узкочерепной полевки, редкой во
влажные периоды. Мышь-малютка более четко выраженный, чем» уз-
кочерепная полевка, влаголюбивый вид. Она чаще встречается во
влажные годы, чем в сухие.
Редкие в Бцрабе виды мало экологически валентны, и у таких
из них, как обыкновенный хомяк, джунгарский хомячок, степная .
мышовка и обыкновенная полевка, хорошо выражена зависимость
численности от фазы обводненности территории. Все они в сухую фа-
144
зу достоверно (Р = 0,01) меняют место в сообществе ближе к доминан-
там, а во влажную смещаются к самым малочисленным.
Обращает на себя внимание и разный характер гистограмм в су-
хую и влажную фазы (см. рис. 42, 43). В сухую фазу лишь в одном
случае отмечается яркое преобладание по численности вида-доми-
нанта (полевая мышь, с. Ровенское, 1965 г.) и в двух случаях не столь
заметное ее преобладание (с. Ровенское, 1982 г.; с. Коноваловка,
1977 г.). В одном случае (с.. Усть-Ургулька, 1978 г.) было два вида-
доминанта — полевая мышь и лесная мышовка, но доминирование их
над остальными видами не было особенно резким. В остальных вось-
ми случаях резкого доминирования какого-либо одного вида, не на-
блюдалось, и ранговые значения отдельных видов были более или
менее сглаженные (с. Ровенское, 1967, 1968 гг.) или с почти равными ,
показателями численности (с. Коноваловка, 1976 г.: с. Крещенка,.
1977 г.). В четырех случаях (с. Ровенское, 1966, 1969 гг.; с. Конова-
ловка, 1975, 1978 гг.) численность всех видов грызунов вообще была
низкой и сообщество находилось как бы в подавленном состоянии
(см. рис. 43). . . . -
Совсем иной'ранговый порядок численности у видов в сообществе
во влажную фазу. Они отражают большие перепады численности
отдельных видов, яркое доминирование одних видов в сообществе,
подавленное состояние других. Из 13 рассматриваемых случаев
(см. рис. 42), характеризующих влажные фазы, .в шести отмечается
резкое преобладание водяной крысы (с. Ровенское, 1960, 1972 гг.; -
с. Потюканово, 1980, 1981 гг.; с. Усть-Ургулька, 1981, 1982 гг.),
в двух случаях четко доминировала полевая мышь (с. Ровенское,
1973, 1979 гг.), в одном доминантом выступали оба эти вида (с. Ро-
венское, 1980 г.), в остальных трех случаях доминирующая роль во-
дяной крысы (с. Ровенское, 1971, 1981 гг.; Усть-Ургулька, 1980г .)
была не столь ярко выражена, но все же четко прослеживалась. В од-
ном случае подобным доминантом была полевая мьйиь (с. Потюкано-
во, 1979 г.) Такое положение в ценозе более характерно для периода
подъема численности водяной крысы. При ярко выраженном доми-
нировании ее (а в некоторых случаях — полевой мыши) численность,
остальных видов грызунов в сообществе во влажную фазу в значи-
тельной степени подавлена. ,
На рис. 44, 45 и табл. 21 те же данные приводятся для насекомояд-
ных; Ранги относительной численности различных видов землероек
в сухую фазу мы характеризуем по сборам в окрестностях с. Ровен-
ское в 1965, 1969, 1982 гг.; с. Коноваловка — в 1975, 1976, 1977 и
1978 гг.; с. Усть-Ургулька — в 1978 г.; с. Крещенка — в 1977 г.;
а влажную фазу по сборам: с. Ровенское — 1950, 1971, 1972, 1973,
1979, 1980, 1981 гг.; с. Потюканово — 1979, 1980, 1981 гг.; с. Усть-
Ургулька — 1980, 1981 гг.; с. Черновка — 1974 г., т. е. в годы пред-
пика, пика и первого года после пика массового размножения водя-
ной крысы.
Во влажные фазы в сообществе землероек бывает многочисленна,
кутора. Большая численность во влажные фазы средней и малой бу-
Ю А. А. Максимов, Л. И. Ердаков 145 ,
I
:-'3 hgIOuj,. сна
, ;/ • ' I j..
Рис. 44. Видовые структуры населения землероек во влажную,фазу увлаж-
i , ненности Барабы. ;
Ровенское: а — 1950, 6 — 1971, в — 1972, в — 1973, Э — 1979, е — 1980, ж — 1981; -
Потюканово: а,— 1979, и— 1980, к — 198Г, Усть-Ургулька: л — 1980, м — 1981; Чер-
। новка: н — 1974.
Бурозубки: 1 — обыкновенная, 2 — крупнозубая, 4 — малая,’ 5 — арктическая, в —
средняя, 7 — крошечная, 8 — равнозубая; з — кутора.
лс 100 ц. сут
2 Т]
б
в
:h
д
в
14,11
h'
н
..'
-I
О ' I 1 , 11—1,1..; j . felT~>—» ГТт-Н . I I—Н-^-, . Г~1—Гч .-_та
4 4 2 7 3 56\14 7 56~2 3\1 4 62 7 5 1 4 3 5 6 2\1 5 2 4 8 6 7 3\1 5 23 6 4 8
8 -!
I 42^2
'3
о
14 52 68 3
142 6 57 33\41 5672 3
Рис. 45. Видовые структуры
населения землероек в сухую
фазу увлажненности Барабы.
Ровенское: а — 1965, 6 — 1969,
в — 1982; Коноваловка: г — 1975,
Э — 1976, е — 1977, ж — 1978; а —
Крещенка, 1977; и — Усть-Ургуль-
ка, 1978. Остальные усл. обозн.'
см. рис. 44.
Таблица 21
Частота доминирования различных видов землероек в сухую (с) и влажную (в)
фазы
Вид ' Фаза Место в сообществе (ранг)
1-е 2-е . 3-е 4-е 5-е 6-е 7-е 8-е
Бурозубки ' s обыкновенная' ; 8 1 0 0 0 0 0 0
в 10 - 2 1 0 0 i О' 0, О'
крупнозубая с’| 0 0 4 1 2 iO. '3 ! - О' 0
В 0 1 1 2 1 2 6 1 0 1
Кутора с 0 i 0 . 1 1 1 0 . 5 1
в 3 • 2 3 - 2 0 1 2 j О'
Бурозубки малая i 1 6 0 ' 1 0 1. 0 0
в 0 8 3 1 1 0 0 0
арктическая с о 2 . 2 2 1 2 2 0
в 0 0 1 У1 6 1 0 0
средняя 1 . с 0 0 1 2 3 1 1 ! 0
в 0 0 4 5 3 1. 01 0
крошечная с 0 0 1 1 । 1 1- 0
в 0 0 0 0 1 3 8 0
равнозубая с 0 0 . 0 • 0 1. 1 1 1
в 0 0 0 0 0 1 0 4
розубок, хотя последняя, как и обыкновенная бурозубка, бывает
многочисленна и в сухую фазу. Более многочисленны в сухую фазу
арктическая, крошечная и равнозубая бурозубки. К сожалению, на-
ши выборки невелики, и доказать достоверность различия домини-
рования в зависимости от фазы обводненности не удалось.
В заключение приводим данные о видовой структуре населения
грызунов и землероек в 1976 и 1977 гг. на Потюкановском стациона-
ре. Эти годы в Северной Барабе характеризуют сухую фазу, одна-
ко в отношении данного, стационара их следует выделить из характе-
ристики сухой фазы и рассмотреть отдельно. Причина в том, что в
Рис. структуры населения грызунов (Z) и землероек
(II) на Потюкановском стационаре в 1976 (а) и 1977 гг. (б).
Усл. обозн. см. рис. 38 (для Г) и рис. 44, (для II).
10*
147’
Распределение видовых аспектов населения
Село 1950 Сухая фаза Влажная фаза
1965 J1966 1967 1968 1969 1970 1971 1972 1973
Ровенское Потюканово ‘ У сть-У ргулька Коповаловка (Ор- ловка) Черновка । 1 I {рещепка 1-П 4-й 3-й 3-й 3-й 2-й i I 3-й 1-Й 1-й 1-й
отличие от Ровенского и Усть-Ургульского участков здесь сочетают-
ся очаги, дающие подъемы численности водяной крысы как во влаж-
ную (болотные вспышки), так и в сухую фазу (болотно-озерные
вспышки). Как следствие этого, для .видовой структуры населения ,
экз. на 10Сщ. сут
Рис. 47. Видовые структу-
ры населения грызунов (II)
и землероек (I) в послед-
ний год депрессии числен-
ности водяной крысы. •
Ровенское: а — 1970, б — 1978,
в — Потюканово, 1978; г —
Усть-Ургулька, 1979; 0 — Ор-
ловка, 1979. Остальные усл.
обозн. см. рис. 38 (для IP 11
рис. 44 (для Г).
Г
148
Табл и ц а 22
грызунов по годам в разные фазы увлажненности
Сухая фаза Влажная фаза
1974 . 1975 j 1976 j 1977 1978 1979 1980 1981 | 1982
4-й ' 1-й 1-й 1-й 1-Й 1 j 2-й
- З-й , ! 3-й З-й 1-й 1-н 1-й | 4-й
! ' ’ i j 5 2-й 2-й 1-й' 1-й ! 1-Й
З-й З-Й j -З-й j З-й 1-й 4-й 1-й I
1-й
1 З-й
i 1 i 1
грызунов водяная крыса может быть обычным представителем во
влажную и сухую фазы.
Водяная крыса в 1976 и 1977 гг. в сообществе грызунов в с. По-
тюканово занимала соответственно четвертое и третье места, что, как
было показано, не типично для сухой фазы в других районах Бара-
бы (рис. 46). В остальных случаях набор видов грызунов на Потю-
кановском стационаре в 1976 и 1977 гг. оставался характерным для
сухой фазы: доминантом была полевка-экономка (как и в 1978 г.).
Многочисленными были узкочерепная полевка— типичный сухолю-
бивый вид, а также лесная мышовка.
Характерным для сухой фазы был и видовой набор землероек.
Лидирующее положение среди них в 1976 и 1977 гг. занимали обык-
новенная, крупнозубая, а также малая и арктическая бурозубки
(рис. 46).
Для Барабы выше выделены четыре наиболее характерных видо-
вых аспекта грызунов. 1-й — водяная крыса + полевая мышь;
характерно присутствие в качестве третьего — четвертого вида в
ценозе — лесной мышовки. Наличие в нем водяной крысы свидетель-
ствует, что такой набор доминантов характерен для влажной фазы.
Этот аспект наблюдался в окрестностях с. Ровенское в 1950, 1971,
1972, 1973, 1978, 1979, 1980 и 1981 гг.; с. Потюканово — в 1979,
1980, 1981 гг.; с. Усть-Ургулька — в 1980, 1981, 1982 гг.; с. Коно-
валовка (Орловка) — в 1978, 1980 гг.; с. Черновка — в 1974 г.
Близок к нему 2-й выделенный аспект — полевая мышь -1- лес-
ная мышовка. Отличает его отсутствие среди лидирующих видов во-
дяной крысы, но в ряде случаев этот вид в биоценозе появляется,
характеризуя, преддверие наступления очередной, влажной фазы.
Этот аспект наблюдался в с. Ровенское в 1969, 1982 гг. и в Усть-
Ургульке — в 1978 и 1979 гг.
Для сухой фазы типичен 3-й аспект — полевка-экономка (чаще
всего доминант) + лесная мышовка, характерно присутствие среди
первых трех видов узкочерепной полевки. Этот аспект наблюдался
в окрестностях с. Ровенское в 1966, 1967, 1968, 1970 гг.; с. Потюка-
ново — в 1976, 1977, 1978 гг.; с. Коноваловка (Орловка) — в 1974,
1975, 1976, 1977 гг.; с. Крещенка — в 1977 г.
149
4-й аспект — мышь-малютка (иногда доминант) -J- полевая .
мышь — характерен для сухой фазы. Отмечен в с. Ровенское в;
1974 г.; с. Коноваловке — в 1979 г.; с, Потюканово,— в 1982 г. '
Выделенные, 1-й и 2-й видовые аспекты грызунов характеризуют
период влажной, ,3-й и 4-й — сухой фазы. Исключение составляют
•годы, переходные1 от одной фазы к другой (табл. 22). ।
По годовому аспекту сообщества можно говорить не только о том,
с влажной или сухой фазой мы имеем дело в текущем году, но й о ка-
ком периоде этих фаз (начало, пик или его завершение) идет речь./
I. • । . . ;
ВЛИЯНИЕ ВСПЫШЕК МАССОВОГО РАЗМНОЖЕНИЯ ГРЫЗУНОВ
НА ВИДОВУЮ СТРУКТУРУ СООБЩЕСТВ МЕЛКИХ МЛЕКОПИТАЮЩИХ
. I ’ ' ч Г ' .
Виды, которым свойственны вспышки у
массового размножения в Барабе
Для Северной Барабы характерны вспышки массового размножения
водяной крысы, во время которых, вид становится исключительно,
многочисленным и лидирующим по численности в сообществе. В годы ,
вспышек водяная крыса наносит вред сельскому и лесному хозяйст-
ву, влияет на сообщества мелких млекопитающих.
Возникает вопрос, каким видам, помимо водяной крысы, свой-
ственны в Барабе вспышки массовых размножений. Выделение нами/
таких ле1т, как «год куторы», «год полевки-экономки» и другие, может
ли служить подтверждением того, что и у этих видов в Барабе сущест-
вуют вспышки массовых размножений?
Выявление в 'барабинском зооценозе видов, для которых свойст-
венны вспышки массовых размножений, представляет интерес и я
том отношении, что позволяет рассмотреть материалы по их динами-;
ке в связи со сменами фаз увлажненности. Если в период влажной
климатической фазы в сообществе животных имеется вид, которому
присущи вспышки массовых размножений и который тем самым >в.
значительной степени определяет структуру животного населения в
течение ряда лет (см. ниже), то имеется вероятность, что такой вид
существует и для сухой фазы, когда водяная крыса ввиду своей ма-
лочисленности не влияет на другие виды. ,/
Исследования видовой структуры и динамики животного населе-
ния Барабы, многолетние материалы по численным соотношениям .
отдельных |видов в сообществе мышевидных грызунов и землероек
позволяют подходить дифференцированно к оценке значения видов
барабинского сообщества в рассматриваемом биологическом про-'
цессе. ’
Очевидно, при выделении видов, которым свойственны вспышК®
массовых размножений, надо прежде всего выяснить, можно ли при
этом ориентироваться на признак доминантности. Таких видов ,.:в>
Барабе среди мышевидных грызунов пять (кроме водяной крысы)^
а среди землероек три (табл. 23). Из грызунов это полевая мышь,(14
раз была доминантом из 37 исследованных годовых спектров); полов- !
150
। , Таблица 23
Частота доминирования грызунов (число годовых выборок 37) и насекомояд-
ных (32) по численности
Место в сообществе
Вид' !'. .. о О) <33 Отсутствова- ли в отловах
! 1 ! - -1 л 00 а
Г рызуны
Полевая мышь1 Г 141 71 8 3 '0 2 1 1 1 1 0 0 .0 —
Водяная крыса ! п! 2; 5 4 3 1 2 1 3 2 0 0 - 3
Полевка-экономка ! , ! 8! 4’ 1 3 4 2 7 4 1 1 0 0 2
Узкочерепная полевка; 2i 5! 5 о 5' 2 3 2 6 1 1 0 ' 2
Мытль-малютг;а г 2 2 5 4 5 3 8 1 3 2 0 1 - 1
Обыкновенная полевка 1 1! 1, 1 2 0 0 1 0 1 0 4 2 1 24
Насекомоядные
Бурозубки , 1
обыкновенная I 27| | 4' 1 0 0 0 0 0 0 0 0 0 —
малая L 2: 17j 8 3 1 0 1 0 0 0 0 0
Кутора i 31 21/6 i ) i 5, 5 2 8 0 01 0 о 1 1
ка-экономка (8), узкочерепная полевка (2), мышь-й1айютка (1) и обык-
новенная полевка (1); из землероек обыкновенная бурозубка (27 раз
из 32 годовых выборок), кутора (3), малая бурозубка (2).
Расвмотрим каждый из перечисленных видов.
Обыкновенная полевкавид, редкий в Барабе.
Из 37 годовых выборок в 24 вообще отсутствовал (см. табл. 23). По
максимальной относительной численности (6,40 экз./100 цил.-сут)
обыкновенная полевка средн мелких млекопитающих стоит на 9-м
месте (табл. 24). Доминантом среди грызунов она была всего один
раз, причем на локальном участке (с. Коноваловка, 1976 г.). Ее до-
минирование в сообществе грызунов при сравнительно низком уров-
не численности других видов было выражено слабо.
На том же стационаре один раз в 1975 г. обыкновенная полевка в
сообществе грызунов была субдоминантой (см. рис. 40). Этот год от-
личался еще более 'низкой численностью грызунов, чем 1976 г.
Так же, как и обыкновенная полевка, всего один раз была до-
минантом и 2 раза субдоминантой мышь — малютка (см.
табл. 23). Этот вид в отличие от предыдущего, обычен для Барабы и|
всегда, за .очень редким исключением, присутствует в населении Мел-
ких зверьков. Год доминирования мыши-малютки (1974 г.) на Ро- I
венском стационаре (см. рис. 45)- отличался наибольшей ее уловис-
тостью (7,37 экз. на 100щил.-сут) за весь период наблюдений в Бара-
бе (см. табл. 24). Максимальный перепад численности по сравнению !
с предыдущим годом равнялся 10,8 (см. табл. 25).
В [1980 и 1982 гг. на Потюкановском стационаре, когда мышь-
малютка была субдоминантом, численность ее составляла 3,46 и
2,0 экз./100 цил.-сут, т. е. незначительна по сравнению с численно-
стью видов, следующих за ней в сообществе. Мышь-малютка чаще
151
Таблица 24
Максимальная относительная численность грызунов и землероек, эг.'з. на 100 цил.-сут (год)
Вид Ровенское Иотюк аново Усть-Ургулька Коповаловка Орловка Макси- мальная Место в раду
Лесная мышовка 1,96(1973) 3,72(1981) 4,09(1978) 1,96 (1974) 0,34 (1979) 4,09 13-е
Полевая мышь 15,65(1965) 3,36(1979) 14,17(1979) 6,01 (1974) 2,63 (1979) 15,65 4-е
Мышь-малютка 7,37(1974) 3,46(1980) 1,35(1979) 2,03 (1974) 0,62 (1979) - 7,37 8-е
Полевки —
красная . 3,74 (1974) 0,72 (1976) 1,92 (1979) 2,03-(1976) 0,14 (1979) 3,74 14-е
р ыжая 2,67 (1980) 0,45 (1976) 2,15 (1979) 1,44 (1974) 0,03 (1979) . 2,67 15-е
Водяная, крыса 20,47 (1972) 93,76 (1980) 112,87 (1981) 2,48 (1974) 2,04 (1980) - 112,87 1-е
Полевки
экономка 3,11 (1978) 10,47 (1977) 1,81 (1980) 5,36 (1976) ’ 0,09 (1980) 10,47 7-е
пашенная 2,53 (1971) 1,61 (1980) 1,78 (1980) 5,16 (1976) 0,89 (1980) 5,16 11-0
узкочерепная 2,94 (1971) 5,93 (1977) 1,89 (1979) 3,77 (1976) 0,03 (1979) 5,93 10-е
обыкновенная - — 0,12 (1976) 0,06 (1979) 6,40 (1976) 0,55 (1979) 6,40 9-е
Кутора 11,16 (1950) 11,05 (1981) 6,96 (1980) 0,59 (1974) 0,26 (1980) 11,16 6-е
Бурозубки
обыкновенная 37,68 (1974) 37,72 (1976) 17,46 (1981) 14,11 (1977) 5,44 (1979) 37,72 2-е
крупнозубая 7,61 (1974) 13,16 (1976) 0,41 (1981) 8,56 (1974) 0,1g (1980) 13,16 5-с
арктическая ; 0,65 (1965) 2,30 (1976) 2,34 (1979) 1,93 (1976) 0,26 (1979) 2,34 16-0
средняя 2,42 (1950) 1,71 (1978) 4,87 (1979) 0,56 (1978) 0,74 (1979) 4,87 12-о
малая 5,43 (1965) 18,36 (1978) 2,69 (1979) 1,16 (1978) 1,42 (1979) 18,36 -3-е
крошечная 0,19 (1981) 0,45 (1978) 0,16 (1979) •0,13 (1974) 0,06 (1979) 0,45 17-е
райнозубая — — • ' 0,17 (1978) 0,09 (1980)
всего занимала 4—9-е места в сообществе (см. табл. 23). Это свиде-
тельствует о значительной устойчивости ее положения в барабинском
сообществе грызунов.
На Ровенском стационаре за период с 1965 по 1982 г. наблюдался
лишь один высокий подъем численности мыши-малютки (1974 г.),;
и циклы динамики в этот период плохо прослеживались.
Узкочерепная полевка дважды (37 годовых выборок)
была доминантом (в 1968 !щ 1969 гг., с. Ровенское), в обоих случаях
при низком уровне численности этого и все£ остальных видов сооб-
щества (см. рис. 40, 43). Подобная картина наблюдалась в Ровенском
в 1971 г., когда узкочерепная полевка была субдоминантой (см.
рис.|42), численность ее составляла (2,94 экз./100 цил.-сут),. в с. По-
тюканово в 1977 г. — 5,93 (см. табл. 24). Наибольший перепад ее чис-
ленности (пятикратный) наблюдался в Ровенском в 1973—1974. гг.
(табл. 25).
Анализ многолетних данных Ровенского стационара (1965 —
1982 гг.) позволяет предположить отсутствие больших подъемов чис-
ленности у узкочерепной полевки в Барабе.
Полевка-экономка отмечена в Барабе в качестве до-
минанта в 8 годовых выборках (из 37). Были годы, когда этот вид от-
сутствовал в отловах (см. табл. 23). Показательно, что в годы, когда
эта полевка была доминантом, в двух случаях (с. Потюканово, 1976,
1977 гг.) она абсолютно преобладала в сообществе грызунов (см.
рис. 46), и численность достигала соответственно 7,36 и 10,48 экз./
/100 цил.-сут. Об устойчивости высокого подъема, численности сви-
детельствует то, что эти годы на одном стационаре смежные. Наиболь-
шие перепады на подъеме численности в два смежных года (см.
табл. 25) отмечены в с. Потюканово в 1979 и 1980 гг. (11,3), когда
численность йолевки-экономки была сравнительно невысокой.
Динамике численности полевки-экономки в Барабе свойственна
цикличность (см. рис. 14, в), которая., подчиняется сменам фаз ув-
лажненности.
Наиболее частый доминант и субдоминант среди грызунов в Ба-
рабе полевая мы ш ь. Из 37 годовых выборок на первых трёх
местах по рангу относительной численности на нее приходится
78,3% случаев (см. табл. 23). По максимальной относительной чис-
леннос!ти полевая мышь после водяной крысы среди грызунов стоит
на втором месте (см, табл. 24). Максимальная ее численность наблю-
далась в 1965 г. в с. Ровенское (15,65 экз./100 цил.-сут) и с. Усть-
Ургулька в 1979 г. (14,17 экз-./ЮО цил.-суд). Наибольший перепад
численности полевой мыши отмечался в Ровенском между 1972 и
1973 гг. и был 18-кратным (соответственно 0,55 и 10,0 экз./100 цил.-
сут) (см. табл. 25). . I
Полевую мышь, как и полевку-экономку, мы относим к видам,
имеющим ярко выраженную динамику численности, с большими
подъемами численности, в периоды которых Она становится домини-
рующим видом в сообществе грызунов. Ее динамика, судя по мате-
риалам Ровенского стационара, имеет примерно 6—7-летние циклы.
15.3
' 7
Таблица 25
Максимальные перепады относительной численности у грызунов и насекомояд-;
пых, экз. на 100 цил.-сут (год)
i Вид ; Ровенское ! I U Потюканово i Усть-Ургульиа Макси- ' мальный перепад
_ ’ 1 Н Лесная мышовка 1 L; i ! Ь,5^-1,Ьб. (1972-1973) i JO,19—1,92 1 (1979—1980) । i । Г 1 '-4,97—14,17 . : 10,1' !|- '
i Полевая мышь I 0,55-10,00 0)53—3,36 ; 18,0 {
(1972-1973) (1978-1979) ] (1978-1979)
Мышь-малютка' 1,83—7,37 0,32-3,46 0,22—1,35 1 40,8
(1973—1974) (1979-1980) (1978-1979)
1 Полевки V' • i 10,86—1,92 (1978-1979)
красная . :1 Ч 0,22—1,96. 1 .. .4 i ,8>9 ;
- . '!) Ч (1972-1973)
рыжая | - 0.21—1,89 1,29—2,15 , 9,0
(1973-1974) (1978—1979) 1 к
Водяная крыса 4,18—20,47 1,17—93,76 6,27—112,87 ' 80
(1971—1972) । (1979—1980) (1980-1981). 1 '> -
Полевки <
экономка 0,71—2,87 0,19—2,15 , 0,40-1,03 11,3
(1969—1970) (1979-1980) (1978—1979)
темная 0,90—2,53 0,71—1,61 ; 0,38—1,78 9,4
(1970-1971) (1979-1980) (1979—1980)
узкочерепная 0,42-2,13 2,18—5,93 1,37—1.89, 5,0 К
— '1 (1973—1974) (1976-1977) (1978—1979) 1
Бурозубки 14,3 11.
обыкновенная 1,22-17,52 (1972—1973— 13,65-18,72 (1977—1978) 1,48-9,97 (1978—1979)
1 : л
1974
крупнозубая 0,47—7,61 0—0,41 16,2 ,
(1973—1974) (1980-1981)
малая1 0—3,46 0,34-4,30 ' 0,54-6,96 12,8
(1972—1973) (1979—1981) (1979-1980)
арктическая 0,35-1,23 2,32—18,36 2,53-2,69 . 8,0 \
(1969—1970) (1977—1978) (1978-1979)
1 Кутора , 0,05—0,55 —. 0,54-6,96 13,0 у
(1972-1973) — t (1979-1980)
Бурозубки к. к'
i средняя 1 0,07—0,59 0,32—4,87 15,2 Ч:
(1972-1973) 1 ' . (1978-1979)
1 крошечная 0-0,11 0,11—0,16
(1973—1974) r 1 (1978-1979) i' ‘
равнозубая — 1 0—0,06
ii « (1978-1979)
' 1 -X Обращает внимание, что последний ниц. численности в Ровенском,1 : ' Потюкановском и Усть-Ургульском стационарах был в один 1979 г.Ч
Наиболее частый доминант и субдоминант среди землероек в Ба- )
рабе обыкновенная б у р о з у б к а. Из 32 годовых выборок
в 27 случаях она была доминантой среди землероек, 4 раза субдоми-
154
нантом и только 1 раз занимала 3-е место (см. табл. 23). По макси-
мальной относительной численности она также занимает 2-е место
после водяной крысы (37,68 зкз./100 цил.-сут, с. Ровенское, 1974 г.;
37,72 — в 1976 г., см. табл. 24). Численность обыкновенной-буро-
зубки в Bapai6e испытывает большие колебания, которые прослежи-
ваются по материалам трех стационаров.
М а. л а 1я бурозубка — типичный субдоминант в сообщест-
ве землероек Барабы, чадце всего (в 78,1% случаев) занимает 2-е и
3-е места по относительной численности среди мелких- млекопитаю-
щих (см. табл. 23). По максимальной относительной численности в со-
обществе грызунов и насекомоядных она стоит на 3-м месте, наи-
большая ее численность, превысившая таковую предыдущего года
почти в 8 раз,[наблюдалась в с. Потюканово в 4978 г,— 18,36 экз./)
100 цил.-сут (см. табл. 24). В указанном году ее высокая численность
отмечалась также в Ровенском и Усть-Ургульке. Многолетние коле-
бания численности выражены у малой бурозубки в Барабе достаточно
резко. '| ।
К у .т о р а — последний вид в сообществе мелких млекопитающих
Барабы, играющий в некоторые годы роль доминанта. За период
наших исследований кутора доминировала трижды и 2 раза была
субдоминантом. Как видно из табл. 23, для нее характерны 3—7-е
места по численности (81,2% случаев). В сообществе землероек Ба-
рабы она занимает 6-е место (см. табл. 24). Наибольшее ее количество
наблюдалось в с. Ровенское в 1950 г. (11,16 экз./100 цил.-сут) и
с. Потюканово в 1981 г. (11,05).
Колебания численности куторы в Барабе выражены четко и, как
показано выше, хорошо согласуются со сменой фаз увлажненности
и динамикой водяной крысы. Наибольший перепад численности это-
го вида за два смежных года (1979—1980) отмечался в Усть-Ургуль-
ке (см. табл. 25) и был почти 13-кратным i (соответственно 0,54 и
6,96 экз./100 цил.-сут).
Поскольку в литературе нет единого мнения, что следует понимать
под «вспышкой массового размножения», это понятие нуждается в
уточнении. При этом необходимо опираться на материалы, собран-
ные в конкретных районах, и на объекты исследований с разными
особенностями массового размножения.
Мы при определении понятия «вспышка массового размножения»
ориентируемся на развитие этого процесса у популяций водяной
крысы в Западной Сибири [Максимов, 1984а]. Что же касдется дру-
гих видов| то их доминирующее положение в сообществе 'в данном
году не может служить подтверждением наличия вспышки массового
размножения. Пример этого — обыкновенная полевка, которая по
одному разу была доминантом и субдоминантом.. Однако это редкий
в Барабе вид с общим низким уровнем численности. Ее доминиро-
вание в сообществе грызунов наблюдалось на локальном участке и
при общей незначительной численности других видов. Это не позво-
ляет говорить о вспышке массового размножения у данного вида.
По этой же причине нет оснований относить к видам, дающим в Ба-
рабе вспышки массового размножения, мышь-малютку и узкочереп-
155
ную полевку, которые редко доминируют в барабпнских сообщест-
вах грызунов.
Таким образом, даже если данный вид бывает часто доминантом,
ато не служит признаком его причастности к группе видов, которым
свойственны вспышки массовых размножений. В Барабе есть виды
(например, 'обыкновенная бурозубка), которые почти всегда являют-
ся доминантами. ' ।
Для вида, характеризующегося вспышками массовых размноже-
ний, более типично, когда его место в видовых рангах численности''
изменяется по годам от доминанта до почти полного исчезновения в
зооценозе. Именно такой характер изменения численности типичен
для водяной крысы. i 1
Поэтому при оценке уровня пика численности особенно важно
учитывать величину ее перепада. Перепад характеризует потенци-
альные особенности воспроизводства у популяций данного вида, его
возможность реализовать эту способность.
При вспышках массовых размножений всегда есть элемент вне-
запности, почему они и именуются «вспышками». В результате такой •
внезапности у местных жителей часто создается: впечатление о на-
шествии данного вида. На примере водяной крысы выше было пока-
зано, что описываемый элемент внезапности связан именно с резкими
перепадами численности.
По наблюдениям на Усть-Ургульском стационаре, между 1980 г.
(предпик) и11981 г. (пий) разница в относительной численности водя-
ной крысы соответственно равнялась 6,27 и 112,87 экз./100 цил.-
сут, т. е. было 18-кратное увеличение, что, однако, далеко не пре- ,
дел, так кай в районе стационара уже с середины лета 1981 г. болота
сильно пересыхали и темп размножения был снижен. Если исклю-
чить 1978 г., когда на этом стационаре водяные крысы в ловчие ка-
навки вовсе не отлавливались, и взять за отчетную точку 1979 г.
(0,74 экз./ЮО цил.-сут), то численность водяной крысы в 1981 г. .
возросла в 152,5 раза. На Потюкановском стационаре в 1979 г. (пред-
пик) п в 1980 г. (пик) относительная численность равнялась соответ- •
ственно 1,17 и 93,76 эяз./100 цил.-сут, т. е. численность возросла в
80 раз. । z
У рассмотренных выше видов в период наших работ максималь-
ные перепады численности составляли 5—18 раз (см. табл. 25).
Очевидно, что 10—20-кратный рост численности у перечисленных
видов не может служить достаточным основанием для утверждения,
что мы имеем дело со вспышками массовых размножений. Это тем .
более вероятно, что подобные перепады численности существуют в
Барабе и у других видов с более низкой максимальной относитель-
ной численностью. Например, у крупнозубой бурозубки перепад
численности равен 16,2, у средней бурозубки — 15,2 и т. д.
Подобные подъемы численности можно оценивать как типичные
многолетние флуктуации. Взятые выше для анализа виды, домини-
рующие в сообществе мелких млекопитающих, не относятся поэтому . '
к тем, для которых характерны в Барабе вспышки массовых размно- <
жений. ’
156
Показатели относительной численности водяной крысы в годы
массовых размножений на порядок превышают максимальные показа-
тели относительной численности наиболее многочисленной из земле-
роек — обыкновенной бурозубки. Это дает основание к видам, для
которых характерны вспышки массовых размножений, относить
в Барабе лишь те, у которых численность при пике возрастает не
менее чем на два порядка.
Явление вспышек массового размножения должно рассматривать-
ся в процессе динамики численности популяций данного вида как
логическое завершение прохождения разных фаз. Поэтому непре-
менным условием цри определении понятия «вспышка массового раз-
множения» должно быть представление о цикличности этого процес-
са, т. е. повторяемости вспышек во времени в связи с циклической
изменчивостью природной среды обитания популяций вида [Макси-
мов, 1978, 1984а]. Колебания этой среды определяют отдельные фа,-
зы динамики и последовательность их смен. Вспышка массового раз-
множения является закономерным следствием многолетнего цикла
динамики, кульминацией ее.
Продолжительность циклов в динамике численности непостоянна
у того или иного вида и не является его неизменной характеристикой
как эволюционно сложившаяся особенность. Частота и амплитуда
вспышек у популяций, как это показано на примере водяной крысы
[Максимов, 1977], различны для разных ландшафтных типов оча-
гов массового размножения.
Однако наличие цикличности в динамике численности вида пе
может служить определяющим признаком того, что цикл приведет к
вспышке, поскольку цикличность свойственна и видам, у которых
вспышки массового размножения отсутствуют. Необходимо разли-
чать вспышки массовых размножений и обычные флуктуации чис-
ленности. ' ।
Под вспышкой массового размножения следует понимать лишь
необычно высокие подъемы численности вида, при которых его зна-
чение выходит за рамки внутрибиоценотпческих отношений и вид
становится объектом большого хозяйственного значения,, которое
воспринимается как из ряда вон выходящее явление, например как
неожиданное бедствие (когда вид вредный) или нашествие (напри-
мер, массовые миграции полезных пушных зверьков) и т. д. Послед-
няя оговорка необходима, так как вид может представлять большое
хозяйственное значение и при мало изменчивом среднем или даже
низком уровне, численности. Вспышки массовых размножений харак-
терны лишь для тех видов, экология которых особенно полно соот-
ветствует существующим в настоящее время циклам изменчивости
природной среды. <
В лесостепной зоне Западной Сибири, как показано выше, необыч-
но интенсивным вспышкам массовых размножений водяной крысы
оптимальным образом соответствуют фазы обводненности террито-
рии в 11-летних циклах. Надо признать, что именно влажная фаза
могла бы способствовать в Барабе развитию вспышек массовых раз-
множений ряда видов мелких млекопитающих, но этому препятст-
157
вует водяная крыса. Последняя — единственный в Барабе вид, пол-
ностью реализующий потенциальные возможности своего воспро-
изводства. ,
‘Из-за большой конкурентной способности водяной крысы, очень
высокой|ее численности во влажные фазы и широкого биотопического
распространения, охватываюшеРо в годы вспышек болота, луга, по-
ля и лесные угодья, возможности развития вспышек массовых раз-
множений у (других 'видов, близких по экологии к водяной крысе
(например, полевка-экономка, кутора), в Барабе практически исклю-
чаются. Что касаетсй сухой фазы в 11-летнем цикле, то виды, для
которых в Барабе условия сухой фазы были бы оптимальны для
существования, отсутствуют. Динамика таких «сухолюбивых» видов,”
как узкочерепная полевка, не । выходит за рамки обычного уровня
флуктуаций численности. В условиях большой заболоченности тер-
ритории Барабы, что отражается и на лугах, ее стации распростра-
нены весьма ограниченно. '
Для каждого вида, характеризующегося вспышками массовых
размножений, существует своя цикличность природной среды, спо- •
собствующая в некоторые годы исключительно высокой плодовитости*
когда численность популяций перекрывает обычные для нее уровни.,
Это те необычно высокие уровни численности, когда вид приобретает
большое значение во внутрибиоценотических отношениях и начина-
ет играть роль вредителя сельского или лесного хозяйства, массово-
го, источника инфекций, служит объектом интенсивного охотничьего
промысла и др. Резко повышается его значение и в естественных био-
ценозах как консумента. <
Признаками вспышки массового размножения являются и такие 5
особенности, как широкое географическое распространение очагов
массового размножения (в пределах сходного природного ландшаф-
та), перемещение на территории и во времени пиков в ходе развития ’
вспышки. Последнее характеризует длительность развития благо-<
приятной для вида фазы природного цикла, что описывалось выше
на примере вспышек массового размножения водяной крысы.,
Перестройка видовой структуры сообщества
мелких млекопитающих
в связи с массовыми размножениями водяной крысы
I Смена фаз увлажненности Барабы в 11-летних циклах характеризу-
ется существенными различиями в структуре населения мелких мле-
копитающих. Влажной фазе соответствует одна, сухой фазе — дру-
гая видовая структура населения грызунов и насекомоядных. Но по-
мимо смены природного фона на разногодичные изменения структуры;
сообществ влияют также и внутрибиоценотические отношения, из
которых особое внимание привлекают регулярно повторяющиеся в 4
Барабе вспышки массовых размножений водяной крысы. •
Долгосрочное прогнозирование вспышек массового размножения
водяной крысы основывалось на ландшафтной типизации очагов,
каждому из которых свойственны свои типы вспышек и условия их
I-
158 ;
развития. «Болотные вспышки» массового размножения водяной
сы в Барабинской низменности прогнозировались по их связи с вта-Ы'
ными климатическими фазами в 11-летних циклах при высокой сол-
нечной активности [Максимов, 1966, 1971, 1977]. Вызванные сменой
фаз увлажненности перестройки видовой структуры , животного на-
селения являются так£ке следствием регулярно повторяющихся в
указанном цикле вспышек массового размножения водяной крысы.
Это сложный взаимообусловленный процесс: 1
Видовая структура сообщества мелких млекопитающих рассма-
тривается в связи со следующими фазами динамики численности 'во-
дяной крысы: последним годом депрессий, совпадающим с заверши-!
ющей стадией сухой; фазы в 11-летнем цикле, фазой нарастания
численности, ее пика, последующим спадом,и первым годом низкой:
численности.
Фазу низкой численности водяной крысы перед подъемом, отра-
жают материалы сборов в Ровенском в 1970 и 1978 гг.,‘ в ;Потюкано-
ве — в 1978 г., в Усть-Ургульке — в 1979 г,; фазу подъема числен-
ности и предпика: Ровенское — 1971, 1979 гг., Потюканово —
1979 г.,'Усть-Ургулька — 1980 г.; фазу пика: Ровенское — 1972,
1980 гг., Потюканово — 1980 г., ,Усть-Ургулька — 1981 г.; фазу
спада численности после пика: Ровенское — 1950, 1973, 1981 гг.,
Потюканово — 1981 г., Усть-Ургулька — 1982 г.; первый год фазы
низкой численности водяной крысы: Ровенское — 1974 г. Использо-
ваны также данные по другим участкам Барабы, характеризующие
отдельные годы перечисленных фаз динамики: - Коноваловка —
1974 г.; Черновка — 1974 г.; Крещенка — 1977 г.; Орловка — 1979,
1980 гг. .
Сменяются и доминанты в указанные фазы цикла динамики чис-
ленности водяной крысы (табл. 26). 1 '
В населении мелких млекопитающих Барабы имеется сравни-
тельно немного видов, которые бывают доминантами в многолетних
сукцессиях. Из мышевидных грызунов это водяная крыса, полевая
мышь, полевка-экономка, узкочерепная полевка, мышь-малютка,
а из землероек — обыкновенная и малай бурозубкщ кутора. Из
этих видов в годы подъема, пика и спада численности водяной крысы
доминируют водяная крыса, полевая мышь и все '‘три перечисленные
выше вида землероек, а в год пика доминантом из грызунов остается
лишь водяная крыса, а из землероек — обыкновенная и малая бу-
розубки. - i
Рассмотрим перестройку структуры животного населения Ъарабы
в соответствии с четырьмя фазами динамики численности водяной
крысы. ' .
Структура сообщества в последний год фазы депрессии числен-
ности водяной крысы, предшествующий началу подъема. В конце
сухой фазы при низкой численности водяной крысы (1970 г.), когда
она занимала среди грызунов 8-е место (рис. 47, /), на Ровенском ста-
ционаре доминантом была полевка-экономка, а субдоминантой —
узкочерепная полевка. Третье и четвертое места по численности за-
нимали соответственно лесная мышовка и полевая мышь. Среди
159
Смена доминантов в сообществе мелких млекопитающих в ходе
Место наблюде- ний Период влаж- ной фазы 1 Фаза
- низкая численность подъем численности I
Ровенское, Каргатским район 19701-.19.73 1 1979-1981 1970, обыкновенная бурозубка, полевка-эко- номка 1973, обыкновенная 1971, обыкновенная бурозубка, водяная . крыса 1979, обыкновенная -
Потюканово, Северный рай- он Усть-Ур- гулька, Север- ный район Коновалов- ка, Колыван- ский район Черновка, Кьнптовский , район 1946-1-1950 1979—1981 10(79—1981 1969—1973 1969—1973 бурозубка, полевая мышь - ‘- 1978, обыкновенная бурозубка, полевка-эко- номка 1979, обыкновенная 1 бурозубка, полевая мышь бурозубка, нолевая . мышь , 1979, ’ обыкновенная бурозубка, полевая i МЫШЬ '1 । । 1980, кутора, водя- ная крыса j . .!> 1:У
Орловка, Колыванскип район 1979-1981 1 1979, обыкновенная бурозубка, полевая мышь •1980, обыкновенная । j 4 бурозубка, водяная ; С крыса 1
землероек в этом году высокая численность отмечалась у обыкновен- ’
ной бурозубки. Субдоминантом с более низкой численностью была
малая бурозубка (см. рис. 47, I).
При следующей вспышке аналогичным на Ровенском стационаре
был 1978 г. Доминантом среди грызунов была полевая мышь, а суб-..
доминантом — полевка-экономка. Третье место в сообществе гры-
зунов занимала водяная крыса, численность которой в этом году бы-
ла невысокой и почти равной численности красной подовки, запи- '
мавшей 4-е место. Среди землероек явно преобладающими были обык-
новенная и малая бурозубки (см. рис. 47, I, II).
На Потюкановском стационаре в 1978 г. доминантом среди гры-•-
зунов была полевка-экономка, субдоминантами — лесная мышовка
и узкочерепная полевка (см. рис. 48). Среди землероек по численно-’
•сти преобладали обыкновенная и малая бурозубки (см. рис. 47, I). .
На Усть-Ургульском стационаре за 2 года до пика численности
водяной крысы (1979 г.) роль доминантов играли полевая мышь,
особенно многочисленная в этом году, и лесная мышовка. Узкоче-
репная полевка переместилась с 3-го (1978 г.) на 6-е место. По-преж-
пему многочисленными оставались лесные полевки. Численность
полевок-экономок несколько увеличилась по сравнению с предыду-
• 160
Т а б л и ц а 26 развития вспышки массового размножения водяной крысы (
динамики ч
дик ; ; спад численности-(1-й год после пика) низкая численность (2-й год после пика)
1972, обыкновенная бу- розубка, водяная крыса 1973, : обыкновенная бурозубка, полевая ! МЫШЬ I 1974, , обыкновенная , ‘ бурозубка, мышь-ма- i лютка /
1980, обыкновенная 'бу- розубка, водяная крыса 1981, обыкновенная бурозубка, водяная крыса 1982, обыкновенная 1 бурозубка, полевая. । мышь
— 1950, кутора, водя- ная крыса । —
1980, малая бурозубка, водяная крыса 1981, кутора, водя- ная крыса 1982, обыкновенная । .' бурозубка, . полевая f мышь 1 <•'
1981, обыкновенная бу- i розубка, водяная крыса 1982, водяная крыса
— — 1974, обыкновенная бурозубка, полевая мышь
, — — 1974, обыкновенная I- бурозубка, полевая ।' мышь
’И ~ —
щим годом,, и в отловах на бороздах появилась водяная крыса
(см. рис.; 47, II).
Среди землероек при преобладающей численности обыкновенной
бурозубки обычными становятся средняя бурозубка и еще немного-
численная кутора. Таким образом, в 1979 г. в отличие от 1978 г.,
характеризующегося «сухолюбивым» составом сообщества млеко-
питающих, появились представители «влаголюбивого» комплекса
(см. рис. 47, Г). :
В Орловке за 2 года до пика численности водяной крысы (1979 г.)
при одинаковых показателях численности разных видов доминантой
была полевая мышь, субдоминантами — темная полевка, мышь-
малютка, обыкновенная полевка. Водяная крыса среди грызунов в
этом году занимала 6-е место. Среди землероек при доминанте — обык-
новенной бурозубке субдоминантами были малая, а потом средняя,
бурозубки (см., рис. 47). I
Структура сообщества в фазу подъема численности водяной кры-
сы (предпик). В год предпика (1971 г.) доминантой у грызунов в ок-
рестностях с. Ровенское была водяная крыса, а субдоминантами со
сходной численностью — узкочерепная полевка и полевая мышь.
Незначительно от них отставали темная полевка и мышь-малютка
и А, А. Максимов, Л. Н. Ердаков
161
зкз.наЮОЦ. ci/Я?
S
Рис. 48. Видовые структуры населения грызунов (II) и з!емлероёк (I)
в фазу подъема численности и предпика водяной крысы.
Ровенское: а — 1971, б —• 1979, в — Потюканово, 1979; г — Усть-Ургулька, 1980;
О — Орловка, 1980. Остальные усл. обозн. см. рис. 38 (для II) и рис. 44 (для Г).
. ; ii 1 ' ,
(рис. 48, II). Структура сообщества землероек оставалась такой же,’
как в ^предыдущем году (обыкновенная и малая бурозубки) (см.
рис. 48, Г). В аналогичной ситуации на этом стационаре при сле-
дующей вспышке (1979 г.) доминантом была полевая мышь, на второе
место вместо полевки-экономки (ее численность была низкой) вышла
красная полевка. Водяная крыса в этщм году занимала третье место,
лишь' немногим уступая по численности красной полевке (см. рис. 48).
Из землероек лидирующее положение удерживали обыкновенная и
малая бурозубки (см. рис. 48, /).|
В окрестностях с. Потюканово в год предпика (1979 г.) также
доминировала полевая мышь, а водяная крыса была субдоминантой.
Узкочерепная полевка и полевка-экономка были малочисленными
(см. рис. 48, II). Среди землероек в этом году, как и в предыдущем,
доминировала обыкновенная бурозубка, а Субдоминантой была ма-
лая бурозубка. Положение в сообществе куторы несколько упрочи-
лось, если в 1978 г. она занимала 5-е, то в 1979 г.-— 4-е (см.
рис. 48, I).
162
J :. 11 i J . ;
«Рис. 49. Видовые структу-
ры населения грызунов (Z)
1 и землероек (11} при пике
численности водяной кры-
сы.
' Ровенское: а — 1972J б — 19801
-в — Потюканово, 11980; г — ।
Усть-Ургулька, 1981. Осталь-
ные усл. обозн. см. рис. 38
(для 1) и рис. 44 (для II). \
В предпиковом, 1980 г.„ впервые на Усть-Ургульском стационаре
доминантом стала водяная крыса, оттеснив полевую мышь и лесную
мышовку соответственно на 2-е и 3-е места. В этом году увеличилась
численность полевки-экономки и темной полевки. Такой сухолюби-
вый вид, как узкочерепная полевка, переместился на 9-е место
(см. рис. 48, II). ( . 1
В год предпика численности водяной крысы место дбминйнта в
сообществе, занимала кутора, сместившая на второе место среди зем-
лероек обыкновенную бурозубку (см. рис. 48, I).
В Орловке в 1980 г. доминантом среди грызунов была водяная
крыса, а «субдоминантом — темная, а затем обыкновенная „полевки..
Среди землероек доминировала обыкновенная бурозубка1^ на 2-м
месте была малая бурозубка, на 3-м — кутора (см. рис. 48, I).
Таким образом, водяная крыса становится доминантом еще при
сравнительно низкой численности. Последнее подчеркивает ее боль-
шую конкурентоспособность в сообществе млекопитающих. 1
Структура сообщества при пике численности« водяной крысы. В
год пика (1972) численность водяной крысы на Ровенском стационаре
резко возросла, этот вид стал- хорошо, выраженным доминантом,-
Субдоминантами с низкой численностью были обыкновенный хомяк,
полевая мышь и северная мышовка. Подавлена была в этом году чис-|
ленность бурозубок обыкновенной (доминант) и крупнозубой (суб-
доминант) (рис. 49, I, II). «
При следующей вспышке (1980 г.) водяная крыса на этом стацио-
наре также была доминантом, оттеснив полевую мышь на 2-е место.
Лидирующее положение потеряла и красная полевка, уступив место
рыжей (см. рис. 49, Г).
11*
163
У землероек изменений в доминировании в год пика численно-
сти водяной крысы на Ровенском стационаре- не наблюдали; роль
доминанта и субдоминанта по-прежнему играли обыкновенная и ма-
лая бурозубки (см. рис. 49; II). t \
В год-пика массового размножения водяной крысы на Потюка-
новском стационаре (1980 г.) этот вид был также четко выраженным
доминантой, а? субдоминантами;; были мышь-малютка и полевая
мышь. Следующие места занимали полевка-экономка, лесная мышов-
ка, темная цолевка. Узкочерепная полевка., как и в предыдущем го-
ду,занимала 9-е место. Среди землероек в этот год по-прежнему наи-
более многочисленными были малая и обыкновенная бурозубки.
^Обращает внимание резкое возрастание численности куторы, заняв-
шей в сообществе землероек 3-е место.
. В год пика на Усть-Ургульском стационаре (1981) численность
(и доля в отловах) водяной крысы резко возросла. Однако роль суб-
доминантов по-прежнему удерживалась за полевой мышью и лесной
мышовкой. Обычными были полевка-экономка лесная и темная по- ,
: левки. Численность узкочерепной полевки оставалась низкой
(см. рис. 49, /). При преобладающей среди землероек численности
обыкновенной бурозубки .по-прежнему высокой была численность
куторы. Оба эти вида, как и в предпиковый год, преобладали в труп- .
пе землероек (см. рис. 49, II).
Структура сообщества, при спаде численности водяной крысы
(первый год после пика). В первый год после: пика (1950 г.) доми- -
нантом среди грызунов на Рфвенском стационаре была водяная кры-
са, субдоминантами — полевая мышь и лесная мышовка (рис. 50,
I). Среди землероек отличалась высокой численностью кутора, а суб-
доминантой была обыкновенная бурозубка (см. рис. 50, II).
При вспышке массового размножения водяной крысы на Ровен-
ском стационаре bi 70-х годах доминантой среди грызунов в первый
год после пика (1973 г.) была полевая мышь, водяная крыса занимала
второе место. Численность других видов была заметно подавлена.
Характерно резкое возрастание численности обыкновенной буро-
зубки, а субдоминантой среди землероек ’ становится кутора (см.
рйс. 50, II).
1 Сходной с предыдущим годом остается структура лидирующих
видов мелких млекопитающих на Ровенском стационаре в 1981 г.:
водяная крыса — доминант, полевая мышь — субдоминант, на 3-м
месте — рыжая полевка. Среди землероек в этот год резко преобла-
дала обыкновенная бурозубка. Субдоминантами были малая буро-
зубка и кутора (см. рис. 50, II).
В первый год спада численности водяной крысы (1981 г.) на По-
тюкановском' стационаре на фоне доминирующего положения этого
вида в ценозе возросла численность лесной мышовки, ставшей суб-,
доминантой, как и в конце сухой фазы (1978 г.). Полевая мышь удер-
живалась на 3-м месте, а численность остальных видов в этот год со<
цратилась. Кутора оттеснила малую и обыкновенную бурозубок на
2-е и 3-е места (см. рис. 50, II).
164
Рис. 50. Видовые структуры населения грызунов (Z) и землероек (ZZ) в фазу
спада численности водяной крысы (первый год после пика численности).
Ровенское, а — 1950, б — 1973, в — 1981; г — Потюканово, 1981; д — Усть-Ургулька,
1982; е — Черновка, 1974. Остальные усл. обозн. см., рис. 38 (для 1) и рис. 44 (для II).
На Усть-Ургульском стационаре в 1982 г. (пик был в 1981 г.)
доминантом оставалась водяная крыса, а субдоминантами при низ-
кой численности — лесная мышовка и полевая мышь (см. рпс. 50).
В качестве дополнительного материала приведем данные отлова
зверьков в 1974 г. в окрестностях с. Черновка на севере Кыштовского
района. Пик водяной крысы здесь был отмечен в предыдущем году.
Преобладающими по численности в 1974 г. были три вида грызунов:
165
эка. на 100 ц. сут
I
.8
86,0 I
□ г I
I "4-
I I И
i! 6 ,6. 3 2. 8 12
' 9i '7 10113
.1 7 . 5 ,8 . 2.11 10
9 3 Чб" 4 13 12 ।
в
i|5 2 7 8 10 1 9 8
1 4 5.i 9 6 14\ 5 6 । , ;
' ’ j f137.68 Д
i;
О ГТН-Н.
! -1 23 4'6 5'7
1243567
14,0.
' '1 >' I , Ji ' ,7
Рис. 51. Йидовые 'структуры ';
11 населения грызунов (Z) и зем-
' ле роек (II) в фазу низкой чпе- ё
ленности водяной крысы (2-й
год после пикаг численности). *
' Ровенское, а — 1974, б\— 1982; j-
в — Коноваловка, 19741; г — Потю- '
, наново, 1982. Остальные [усл.
J |Обозн: <ж. рис. 38 (для Г) и рис. 44
8Г (для /I). .
а
е
1 4
1 9 I 8
лИ" . I; ! ' J \ ! г' ! 1 1 У
полевая мышь (доминант), красная полевка (субдоминант) и на 3-м
месте — водяная крыса. Численность лесной мышовки (быпа значи-
I тельно ниже и она находилась на 4-м месте.. Из землероек резко пре-
обладала обыкновенная бурозубка. На 2-м и 3-м местах по численно-
сти были малая и арктическая бурозубки (см. рис. 50, II).
Структура ' сообщества при низкой численности водяной крысы
. (второй год после пика). В 1974 г. на второй год после пика числен-
। ности водяной крысы этот вид на Ровенском стациойарев сообществе
грызунов переместил ей на 7-е место (рис. 51), а доминировали мышь-
малютка и полевая мышь (субдоминант). Среди землероек домини-
рующую роль играли обыкновенная, крупнозубая (субдоминант!) бу-
нрозубки. Кутора в сообществе |землероек. в этот год занимала 3-е
i место (см; рис. 51, II). . । !•
Г \ В 1982 г. на второй год после пика численности водяная крыса на
Ш Ровенском стационаре была малочисленной Ли занимала в ценозе
йрызунов 9-е место. Доминантой среди грызунов была полевая мышь,
субдоминантой — ^ыжая полевка. Среди землероек наиболее много-
। , численными были обыкновенная и малая-бурозубки ;(см. рис. 51).
। ' На Коноваловском стационаре водяная !крыса в 1974 г. занимала
4-е место; Доминировали в сообществе грызунов полевая мышьи
i полевка-экономка. Среди Землероек первые два'места по численности
занимали । обыкновенная и крупнозубая бурозубки, на 3-м — малая
бурозубка (см. рис. 51, I, II). •• у '
На Потюкановском стационаре второй год после пика характери-
зуют сборы 1982 г.। Водяная крыса в отловах на плуговых бороздах
отсутствовала. Доминантой была полевая мышь, субдоминантой —
i мышь-малютка. Среди землероек многочисленной в этом году была
1С>6
обыкновенная бурозубка, малочисленной — малая бурозубка. При
рассмотрении этих данных следует учитывать кратковременность
периода работ и незначительный объем материалов.
. ' ! , . i : J . Л,1 -
! ! i । । • Прогностические Возможности перестроек
! . 1 , й структуры сообщества
‘ ' Л - I ', ‘ Н' > ( ;
j I* * j * ' j _ l! 1 j r - j' 1 i h1 '1 ‘ t
Обзор многолетних данных (см. предыдущий'-раздел) показал, что за1
21 года до пика численности водяной крысы в период1, переходный от ; i
сухой к влажной фазе^численность этого вида в сообществе грызунов
обычно низкая. Она Занимает соответственно 3-е, 4-е,, 6-е, 8-е и 9-е i
места. В; двух случаях доминантом были; полевка-экономка и в
трех — полевая !мДшь, а' субдоминантами— полевка-экономка, уз-
кочерепная, темная полевки и в двух случаях — лесная мышовка.
Из землероек наиболее многочисленной оставалась обыкновенная
бурозубка, а субдоминантами в четырех случаях была малая бурозуб-
ка и в одном — средняя. Весьма характерно наличие среди видов с
высокой численностью узкочерепной полевки, которая занимала в
рассмотренных случаях 2-е, 3-е, 5-е, 6-е, |и 41-е места.
Численности отдельных видов грызунов обычно i соизмеримы.
Лишь в одном случае (с. Усть-Ургулька, 1979 г.) перепады числен-
ности! доминанта и субдоминанта носили ярко выраженный харак-
тер. В большей степени в этом отношении различалась численность
доминантов1 и субдбминантов у землероек.
В Год начавшегося подъема численности водяной крысы, пред-
шествующий пику, этот вид в трех случаях уже занимал ведущее
место в сообществе грызунов!. В двух других случаях эту роль вы-
полняла полевая мышь. Субдоминантами в двух случаях были также
водяная крыса и полевая мышь,, по одному случаю — узкочерепная,
темная и красная полевки. Из землероек в год предпика численности
водяной крысы в четырех случаях лидирующее положение занимала
обыкновенная бурозубка и в одном случае — кутора, а субдоминан-
тами в четырех случаях были малая бурозубка и в одном — обыкно-
венная, поменявшаяся местами с куторой. ! ।
Характерно сокращение численности узкочерепной полевки.
Та же тенденция прослеживается и у мыши-малютки, лесной мышовки
и лесныХ| полевок. Численность темной полевки в этом году становит-
ся выше по сравнению с предыдущим годом. Проявляется четкая
тенденция роста численности куторы, которая в год предпика водя-
ной крысы' перемещается с 5—7-го на 1—3-е места. Численность до-
минантов. в год предпика значительно больше контрастирует с ос-
тальными видами, но это доминирование в половине случаев связано
с полевой мышью. I ! '
В год пика водяная крыса во всех случаях становится ярко вы-
раженным доминантом, а роль субдоминанта выполняют пблевая
мышь (в двух случаях), мышь-малютка и обыкновенный хомяк.
Узкочерепная полевка по-прежнему имеет низкую численность, за-
нимая среди грызунов 5-е, 8-е, 9-е п 10-е места. Для сообщества обыч-
167
но характерен большой разрыв в численности вида-доминанта с дру-
гими видами.
Из землероек доминантом является обыкновенная бурозубка, j
а субдоминантами — малая, крупнозубая и кутора. ' >
В первый год после пика численности водяной крысы в четырех
: из шести случаев этот вид занимал по-прежнему, ведущее место по
численности, изредка।уступая его полевой мыши. Субдоминантами
в двух случаях были полевая,мышь и северная мышовка и по одному,
случаю — водяная крыса и красная гполевка. Лолевка-зкономка и,
узкочерепная полевка характеризовались низкой численностью, за-
нимая в сообществе от 5-го до 8—9-го места.. Несколько выше в эти
годы численность лесной мышовки, играющей уже роль субдоминан- -
та. Превалирующая численность водяной крысы как'доминанта про-
должает , быть еще четко выраженной. 11 ! ;
! Из землероек роль доминантам трех случаях играли обыкновен- г
ная бурозубка и в двух случаях-кутора, а субдоминантами были в
трех случаях малая бурозубка, по одному случаю кутора и обыкно-
венная бурозубка. 1 1
Второй, год после пика численности водяной крысы знаменует
собой начало периода депрессии этого вида. Численность водяной;
крысы сокращается, она в структуре' населения грызунов переме- .
щается на 4—9-е места; В одном случае (с. Потюканово, 1982 г.)
она отсутствовала в сборах. : 1
Характерно увеличение численности полевок: экономки и узко-
черепной, мыши-малютки, т. е. видов, которые были малочисленными
f в годы пика водяной крысы. Соотношения численности грызунов
сглаживаются, нет такого резкого контраста между видами-доминан-
тами и субдоминантами, с одной стороны, и остальными — с другой.
Проведенный анализ вскрывает важную роль водяной крысы в
формировании видовых структур животного населения Барабы. На
вопрос, в течение скольких лет должно учитываться влияние мас-
сового размножения водяной крысы на структуру животного населе-
ния, можно ответить, что водяная крыса, будучи самым конкуренто-
способным видом в исследуемом зооценозе, подавляет своей числен-?
ностью и повсеместным распространением другие виды в течение трех
лет: предпика, пика и первого года после пика. Лишь после этого
'периода восстанавливается видовая структура населения. Но при-
ходится учитывать, что зта новая структура в .значительной степени
является следствием предшествующей вспышки массового размно-
, жения водяной крысы, так как включает ряд видов, которые, будучи
экологически близкими к водяной крысе, вынуждены уйти от ей
пресса и другую «временную нишу». .
Здесь уместен сравнительный анализ видовых перестроек живот-
I ного населения Северной Барабы и поймы р. Оби. В пойме р. Оби,
временным репером для анализа перестроек видовых структур нами
. принят год особенно высокого и продолжительного разлива, который
, приводил к краху пойменное сообщество мелких млекопитающих и к
последующему его восстановлению в годы, следующие за высоким;
168
Рис. 52. Схема формирования видового состава животного населения в Ба-
рабе в связи со сменой фаз увлажненности в 11-летних циклах и с вспышка-
ми массовых размножений водяной , крысы.
разливом. Выявлены определенные ряды сукцессии в период между
высокими разливами [Максимов и др., 1981].
В животных сообществах Барабы такой точкой'отсчета, или ре-
пером, разногодичной изменчивости видовых структур может быть
принят год "пика численности водяной крысы в период влажной фазы
в 11-летнем цикле.
За 11-летний цикл, в период которого происходит смена фаз кли-
матической увлажненности, видовые сообщества сменяют- друг дру-
га, меняются видовое разнообразие и численные соотношения отдель-
ных видов. В этих процессах прослеживается определенная после-
довательность, обусловленная циклами природного фона и регу-;
лярно повторяющимися вспышками размножения водяной крысы.
Проведенный анализ свидетельствует о том, что при краткосроч-
ном прогнозировании вспышек массового размножения водяной кры-
сы и определении года, когда на данном участке следует ожидать пик
ее численности, целесообразно привлекать также- и материалы по
видовой структуре населения мелких млекопитающих. Эти данные
можно использовать для определения фазы динамики численности
водяной крысы, так как определенные структуры животного населе-
ния мелких млекопитающих характерны не только для фазы пика, но
и для фазы спада численности водяной крысы и ее депрессии.
Рассмотренная перестройка видового доминирования мелких
млекопитающих отражает смену фаз увлажненности территории в
169
11-летнем цикле, но во влажную фазу приобретают важное значение
также и массовые размножения водяной крысы — вида с высокой
конкурентной способностью И[ ‘широкой эврибионтностью. Многие
виды сообщества вступают в жесткую конкуренцию с доминирующим
видом, их численность в; сильной степени становится зависимой от
! \ внутриценотических -отношений.., ; ; J'i"' М
Следовательно, «для некоторых!,видов смена фаз j увлажненности
является фактором :изменчивости их, численности (виды, , типичные
' для влажной-фазы; виды,! типичные для сухой фазы). Для других
видов главной причиной изменения численности служит не смена фаз
! природного климатического цикла, йа бйоценотические отношения.
; Существуют виды, малозависимые от смён фаз увлажненности и вспы-
шек: массового размножения водяной крысы (рис. 52).
5 ' । 1 ! | ' ' ! ' | ‘ i । > 1 у
: Значение некоторых видов мелких млекопитающих , 1 н
( как эдификаторов. : фаз । динамики численности водяной крысы
fl I ' i I i : Н I I ’ II . I I ‘I
ё 1 * I 1 | '’ll" а
1 В годовых [пёрестройках животного населения Барабы участвует
14 видов мышевидных грызунов и 8 видов землероек. Ниже (табл. 27)
показано, какое место в ряду этих перестроек занимал каждый вид в
разные фазы динамики численности .водяной крысы. (
Водяная крыса занимала 1-е место среди грызунов (была доми-
нантом) — 11 раз, субдоминантом — 2 раза; 3-е место — 3 раза;
4-е и 7, 9-е места —;по 2 раза; 6-е — 1! раз. Она становилась доми-
1 яантом в сообществе грызунов всегда в гёды пика своей численности;
в трех случаях из пяти — в годы подъема численности (предпика),
IB четырех случаях — в । годы спада численности после пика; субдо-
минантом была- по одному разу в год предпика и после пика числен-
ности. । ;
При поисках видов-эдификаторов развития вспышек массового
размножения водяной крысы не все указанные в табл. 27 виды могут
I быть использованы для этой цели. Сказанное прежде всего относится
; к редким и, постоянно малочисленным .видам. Из грызунов это преж-
де всего, южная мышовка, джунгарский хомячок, обыкновенная по-
левка, а из землероек — крошечная и равнозубая бурозубки.
В этот! список мы условно; включаем также и обыкновенного хомяка.
' За рассмотрейный период он был редким в сборах, однако в одном .
случае (с. Ровенское, 1972, г.) при общей низкой его численности
вышел.в субдоминанты. Это процзошло в год массового размножения
I водяной крысы. Значение этого1 вида как возможного эдификатора
Н прогнозирования вспышек массового размножения водяной крысы
нуждается в;: дальнейшем исследовании. 1 i ! !
। Из такого анализа можно ।исключить также виды, постоянно до-
минирующие в сообществе, независимо от уровня численности во- л
дяной крысы. К таким видам мы относим обыкновенную бурозубку и
полевую мышь. Из 22 рассмотренных выше случаев, характеризую-
щих два года перед [пиком, пик и два года после пика численности
водяной крысы, обыкновенная бурозубка была в 18 случаях доми-
170
нантом, в трех случаях — субдоминантой и в одном случае занимала
3-е место в ряду перестроек (см. табл. 27).
Из 24 рассмотренных случаев полевая мышь по численности
10 раз ^занимала место доминанта, 6 раз — субдоминанта, 5 раз ____
3-е место и только; в трех случаях — 4-е или 0-е. р
Полевая' «мышь ! И обыкновенная бурозубка среди лидирующих
по численности видов ; барабйнского ^сообщества могут рассматри-;,.
ваться как существующие вне видимого пресса водяной крысы. , i
При! поисках эдификаторов вспышек массового размножения во- •*'
дяной крысы из'.представителей животного сообщества ! могут ’ быть J
исключены виды,'хотя и обычные для него, но занимающие (с неко-
торыми исключениями) постоянно среднюю часть ряда и не переходя-, .
щие в,число доминантов и субдоминантов. У этих видов слабо] прояв-
ляется рёакция на резкие изменения численности, водяной крысы.
К ним могут быть отнесены полевки: темная, рыжая и красно-серая,
а из землероек — арктическая и средняя бурозубки. , ।
I При ,выборе эдификаторов особое внимание привлекают виды с
резко, изменяющейся численностью. Они имеют как очень высокую'
(доминируют), так и| очень низкую , численность, |а в некоторые
годы нередко и вовсе отсутствуют в отловах. Эти виды следующие:
из землероек — кутора, а также крупнозубая и малая бурозубки, ।
из грызуров — экономка, узкочерепная, красная,,полевки, мышь-;
малютка и лесная мышовка. Именно за счет этих, обычных и много-
численных в некоторые годы видов и происходят в основном резкие,
четко обнаруживаемые перестройки в населении сообществ в годы
нарастания, пика и спада численности водяной крысы.
Из землероек особого внимания заслуживает кутора, обитающая
в сходных с водяной крысой околоводных биотопах. Это типичный
стенотоп, эволюционно связанный с водной средой обитания, отлича-
ющийся в то же время высокой экологической валентностью и эври-
фагией [Сообщества..., 1978]. Когда ।численность водяной крысы в
болотах возрастает, на условия существования куторы это не может
не сказаться. Вытеснение ее идет из-за конкуренции не за корм,
а за места обитания. Характерно, что в годы пика । численности
водяной крысы кутора занимает ;в ранговом порядке (см. табл, 27) I
по одному разу 2-е, 3-е, затем 6-е и 7-е кеста. .Наиболее характерны
для нее 3-е 'и последующие места, которые .отражают положение
вида в различные годы в периоды нарастания и спада численности
водяной крысы. Кутора становится доминантом в сообществе, земле-
роек ;чаще в год, предшествующий пику, и в первый год после пика ,
численности водяной крысы. Это говорит о том, что она вынуждена .
уходить От конкуренции с водяной крысой во временную нишу, ко-
торая тяготеет к периферии периода* массового размножения водяной .
крысы, т. е. к тем годам влажной фазы и большой обводненности бо-
лот,! когда численность водяной крысы еще не достигла пикового
уровня или уже снизилась после пика.
В сухие фазы 11-летнего цикла вместе с сокращением численно-
сти водяной крысы снижается и численность куторы. Перепады чис-!
ленности между отдельными1 годами поэтому весьма значительные.
171
Место вида в ряду перестроек видовой структуры населения мелких млекопита
Вид Место по
1-е 2-е , 3-e .. 4-e 5-e ।
i 1 : 2 . , И : з 4 ir ' ; 5 6
Полевая мышь' 1,1. п, и, ii, in, ш, II, 111,(111, I, И —
Водяная крыса' IV, V,' IV, V, V, i II, II, III, iv, : v, IV ; И, IV! v ; IV, IV I, II, IV I,iV
* ' 1 ,1 1 1 > ! ' Полевка-экономкаН III,1 III, IV, IV, IV, IV, f II, III I,I t I, V 1, III, III, - n, IV,
Узкочерепная полевка Мышь-малютка -' 1 V I, II । III, V I, V • I V v , ; 1 II, II, v I, III, IV I, II, IV,
Красная полевка n, iv V IV, IV III, IV, v,v I, I, IV
Лесная мышовка 1 I,i Ij, IV, IV I, II, III, V III, IV II, II,
S - у- Полевки i темная I, II ' IV, IV, V , i • II, IV III, III, ! I V
ч красно-серая '! — ' ,1, II, , II —
- ; • 1 рыжая Обыкновенный хомяк ’ Г - V ,111, IV, V I, JI, IV Ill, IV,
— III — IV II
172
Таблица 27
ющих Барабы в годы развития вспышки массового размножения водяной крысы
доминированию
6-е 7-e .8-e . 9-e 10-e: 11-e 12-e 13-e
7 8 ' 91 101 11 12 13 14
I I,V ' I, V - . —
IV П, П, II, III IV, IV ' I, III, IV, II I .. — • *
I i III, IV, IV IV, IV, V II, II, II. Ill, IV I, III I
1 III, IV, IV I, III I, II I, II IV — — —
1, II, II, III, IV, V II, II III, III, IV, I V ’ —
V, II I, IV II, V — — — —-
II, III, III, IV1 V I, I, II, II, EV, V in, IV, V, IV I, IV —
IV I, III V, V I, I. I, II, II, III, III, IV II V
I, II,v I, III, v , — I, IV, II II, III — — -—
— I I, III, III, IV II, II, III, III, IV, IV IV, IV, V IV I
173
1 2 3 4 15 6 -1
Южная мышовка ;' । . i , . Джунгарский 1хОмя- чок ii Обыкновенная полев- ка । I, Бурозубки ; ' । 4 обыкновенная 4 Л ' ,Т . J , -'i г ' 1 ’ ! 1 ' ” i крупнозубая • Кутора ' 1 i ’ _ ' Бурозубки ; малая i i i ! 1 i арктическая ( < ! ! <; > средняя' v :i! крошечна^ ! i : - i ! 1 i - 1 i ' 1 равнозубая , I! • 1 П р'л мычание после пика); V — низкая 1 1 i, I, i, i, ii, II, II, V, III, III, III,i! IV,t iiv,v, v,' iv, I, IT • I i II, IV,.jIV ! i 4 ’ - 1 i Г ' ' '1 I — 4 аза в л численность ( ’! 1 ' I 4 [' । - 'h II, III, IV - ' I ' ’ I bJ in, v, v-' i ' III, IV I, I, I, II, II, II, v, III, IV, IV, IV, 1, II f ! 1 , i ' I ' ; - , ; Г 1 ; 1 И' ! ! Й- ? зной численно 2-й год после п 5 ; ; 1 >1 '! (!' —1 ill ‘ I ili !h . ; IV । , 1 !' i in II, II, ill, IV, V I, II, Ilf, IV, V I, IV ' i i 1 i, i, ii, и, in, IV, I, V i ! 11 } ' СТИ ВОДЯНОЙ ' H ика). В графах iii ' !i i I, II, iv, II,v II, V," | IV ; V, IV I,,i,i II, , in, in, i i, и, in, in, IV, IV 5 ‘ 1' рысы (пере фазы поста — 1 г г ; 1. I, II, IV I, I, I III II, II,' III. III, IV, IV, V III, IV, ’ V, IV; I, п, V II д подъемок влеиы в по-
174
Окончание табл. 27
7 8 9 10 11 . 12 13 14
1 и ’ — i';\. 1 “ ! !--i I ' ill, V II, IV,1 iv, v . I, ' II, III
1 II 1 ( 1 ! I ' i:— ' ' 7 ' 1 i V, p- II, IV , i 1 I,; I,!il, . Ill, Vi IV
i : ’ . -' / j ' ! I, ,11, III, V I, V i r I —i
'| 1 1. 'I 1 I — 1
I, П„ III, III, IV, IV, IV 1 - ! 1 / ! • П, I ; — 1 'l i 1 i - । i ! 1 1 : 1 -Л
I, III I, II, III, V — — — — - —
— 1 — — — — —
I II, V, V, IV 1 I, II, IV — — 1 i ' ! — I “
I II, V II 1 ~~~ i i j । , : 1
I, I, II, III, IIV 1 ‘ i I, II, II, III, v, v, III, I, III, IV, IV, IV, IV — i • ! I 1 J i 1 i
II, I I I, II, III, III, IV, IV 1 i . 1 — — —.
численности); II —-фаза подъема численности; III — пик; IV — спад численности (1-й год'
рядке годовых выборок.
.1 1
175
Так, в Усть-Ургульке в 1980 г. (предпик водяной крысы) числен-
ность куторы по сравнению с предыдущим годом возросла почти в
13 раз, а в 1981 г. (пик водяной крысы) вновь снизилась, и этот вид в
ряду землероек переместился с 1-го на 2-е место.
Приведенные данные свидетельствуют о том, что кутора являет- '
ся хорошим эдификатором развития в Барабе влажных фаз и может
использоваться в качестве признака наступления массового размно-
жения водяной крысы: Появление куторы в отловах, когда числен-
1 ность водяной крысы еще низкая, говорит о приближающейся вспыш-
ке массового размножения последней. Когда численность куторы
возрастает п она начинает занимать; в сообществе землероек 3-е и
последующие места, это является-признаком начала, подъема чис-
ленности водяной крысы. Если же кутора выходит в сообществе
землероек на место доминанта или субдоминанта, это свидетельство
начала развития пика численности водяной крысы. Такое же состоя-
ние численности этого вида после пика может быть дополнительным
свидетельством, что вспышка водяной крысы уже завершается.
Крупнозубая бурозубка, для которой также характерна большая
изменчивость численности в годы развития вспышек массового раз-
множения водяной: крысы,— эвритопный вид, тесно связанный с
влажными участками. Она отдает предпочтение приболотным участ-
кам займищных низин и опушкам березово-осиновых лесов. Цикличе-
ские изменения климатической обстановки в Барабе оказывают суще-
ственное воздействие на ход и интенсивность размножения крупно-
зубой бурозубкп. Ее плодовитость в условиях обводнения выше, чем
в сухие периоды [Сообщества...,.1978].
Указанные особенности экологии крупнозубой бурозубки сбли-
жают этот вид с водяной крысой и делают динамику ее численности, ->
сопряженной с зтим видом грызуна. Будучи по численности 4-м ви-
дом в сообществе землероек Барабы, крупнозубая бурозубка ни в
одном случае не играет роль доминанта в годы пиков численности
водяной крысы. В такие годы она занимала, по одному разу 2-е и
3-е места! и два раза — 6-е место (см. табл. 27). На подъеме и в пер-
вый год спада численности водяной крысы крупнозубая бурозубка
занимала 4-е и последующие места и лишь на второй год после пика
численности водяной крысы начинала играть роль субдоминанта в
сообществе землероек. ! ,
То, что численность этого вида, тяготеющего к влажным место-
обитаниям, очень изменчива в годы нарастания, пика и спада числен-
: ности водяной крысы и то, что в годы вспышек водяной крысы круп-
нозубая бурозубка занимает чаще всего с 4-го по 6-е места и никогда
не бывает доминирующим среди землероек, подтверждает связь ди-
намики ее численности с водяной крысой. Видимо, она тоже может
служить эдификатором размножения водяной крысы.
Малая’ бурозубка отличается высокой экологической валент-
ностью и тяготеет к местообитаниям с влажным микроклиматом: .
приболотным участкам и лесным биотопам [Сообщества..., 1978].
Численность ее в годы развития массовых размножений водяной
крысы менее изменчива, чем у куторы и крупнозубой бурозубки, и в
176
сукцессионном ряду занимает места не ниже 5-го (см. табл. 27).. .
Лишь в одном случае она доминировала среди землероек в год пика
численности водяной крысы (с. Потюканово, 1980 г.). Чаще же все-
го в годы нарастания, пика и спада численности водяной крысы ма-
лая бурозубка была субдоминантом среди землероек и занимала по
численности 2-е и 3-е места. • 1
В более зависимом, чем землеройки,'положении от водяной крысы
находятся полевка-экономка, узкочерепная и красная полевки,
а также мышь-малютка. В годы высокой численности водяная крыса. I
вытесняет эти виды из их типичных местообитаний, конкурируя так-;
же и по трофике. Это касается не только зеленоядных полевок, но и .
мыши-малютки, так как водяная крыса зиму проводит на засоренных -
лугах и полях — типичных стациях этого вида. Характерно, что все
четыре вида никогда не бывают доминантами в годы пиков числен-
ности водяной крысы, а субдоминантом лишь в одном случае была
мышь-малютка (см. табл. 27). В периоды роста, пика и спада числен-
ности водяной крысы для всех указанных видов грызунов характерно
положене в ряду ниже 4—5-го места.
В особенно острые конкурентные отношения с водяцой крысой
вступает полевка-экономка, наиболее близкий по экологии вид.
Она бывает доминантой и субдоминантом в сообществе мышевидных
грызунов в условиях Барабы лишь при низкой численности водяной
крысы, а при пике последней занимает место не выше 4-го (см.
табл. 27).
Сказанное иллюстрирует график изменения относительной чис-
ленности водяной крысы и полевки-экономки с 1976 по 1981 г., со-
ставленный по данным Потюкановского стационара (рис. 53). Пик
численности полевки-экономки наблюдался здесь в 1977 г., когда
численность водяной крысы была еще низкой. В 1980 г. когда в
районе с. Потюканово отмечена вспышка массового размножения
водяной крысы, численность полевки-экономки была подавлена.
Сходные соотношения численности обоих видов наблюдались в
1978—1981 гг. на Усть-Ургульском стационаре (см. рис. 53).
Особенно интересны в этом отношении многолетние материалы
Ровенского стационара. В период сухой фазы при депрессии числен-
ности водяной крысы полевка-экономка может давать два помета и
достичь пика численности (1967 и 1970 гг.). В годы массового размно-
жения водяной крысы численность полевки-экономки снижается
(1972, 1980 гг.). Будучи влаголюбивым видом, полевка-экономка
вынуждена занимать временную экологическую нишу, приуроченную
к сухой фазе.
Наличие в сообществе Бафабы такого конкурента, как водяная
крыса, с ее интенсивными вспышками массовых размножений, ска-
зывается и на размахе колебаний численности полевки-экономки.
Наибольшие перепады численности у полевки-экономки наблюдали
в с. Ровенское в 1966—1967 гг. (увеличение в 3,5 раза), с. Усть-
Ургулька в 1978—1979 гг. (увеличение в 2,5 раза) и с. Потюканово
в 1977—1978 гг. (снижение в 3,5 раза). Это говорит о сравнительно
небольших колебаниях численности полевки-экономки в Барабе.
1- А. А. Максимов, Л. H, Ердаков
177
Рис. 53.' Изменение относительной численности в.одяной крысы (а — в)иполев-
; ; ; > , ки-экономки (г — е). ;
'' , ", А — Потюканово, Б — Усть-Ургулька, В,— Ровенское. . '
Так же, как кутора, полевка-экономка может служить одним из
наиболее надежных эдификаторов развития вспышек массового1 -
размножения водяной крысы. Снижение ее численности и доли учас-
тия в населении мышевидных грызунов свидетельствует о наступаю-
щем в Барабе размножении водяной крысы.
В большой зависимости от последней находится узкочерепная по-
левка, которая, будучи, типично луговым видом, страдает от нее в
178
1
годы пиков, когда водяная крыса в массе заселяет различные типы
лугов, в том числе и стации узкочерепной полевки. В условиях мас-
сового обитания водяной крысы на лугах поселения узкочерепной
полевки исключаются. Она вытесняется из своих стаций, размноже-
ние ее подавлено. (Характерно, что в годы подъема и пика числен-
ности {водяной крысы узкочерепная пойевка среди грызунов занима-
ет чаще всего 9—10-е места. Ее доля в населенйи грызунов несколь-
ко повышается в годы! спада и депрессии численности водяной крысы
(см. табл. 27). । ।
В годы нарастания численности водяная крыса интенсивно за-
селяет и стации красной полевки — различные лесные участки, ста-
вя и этот вид в зависимое положение и обусловливая снижение
доли ее участия в животном сообществе.
Проведенный анализ показал, что массовое размножение водя-
ной крысы сказывается на видовой структуре сообщества мелких
млекопитающих. В годы высокой численности водяная крыса засе-
ляет все болота и доминирует в них. Часть видов под влиянием кон-
курентной напряженности несколько изменяет свои экологические
требования. В сухую климатическую фазу, когда численность водя-
ной крысы низкая и она в основном приурочена к стациям пережи-
вания, другие виды увеличивают свою численность. Поскольку ба-
рабинское сообщество мелких млекопитающих характеризуется в
разные годы 11-летнего цикла преобладанием того или иного вида,
можно ли в нем по примеру восточного календаря выделять и име-
новать отдельные годы, ориентируясь на вид-доминант?
Если исключить такие постоянно встречающиеся и многочислен-
ные виды, как полевая мышь и обыкновенная бурозубка,!то особенно
четко в Барабе проявляют себя «годы водяной крысы». Такие годы в
целом по Барабе растягиваются на весь период влажной фазы, про-
являясь то в том, то в другом месте. Высокой численностью и пре-
обладанием в сообществе характеризуются не только год пика водя-
ной крысы, но и, хотя и в меньшей степени, год предпика и следую-
щий за ним год. В барабинском сообществе мелких млекопитающих
четко обнаруживаются также «год куторы» и «год полевки-экономки».
Как было, показано выще, имеются определенные |3акономерности
в распределении таких лет в 11-летнем цикле в зависимости от фазы
увлажненности и фазы динамики численности водяной крысы: «год
куторы» в пределах влажной фазы предшествует или следует за го-
дом пика, «год полевки-экономки» более типичен для сухой фазы,
когда численность водяной крысы находится в депрессии.
Эти данные нуждаются в дополнительном подтверждении мате-
риалами по нескольким 11-летним циклам, но уже сейчас в качестве
предварительного вывода можно сказать, что наблюдаемая в кон-
кретном году видовая структура населения зверьков и появление
среди лидирующих по I численности видов руководящих форм можно
использовать как для краткосрочного, так и для долгосрочного био-
логического прогнозирования.
179
Одновременность и сходство перестроек
животных сообществ в ходе развития вспышки
массового размножения водяной крысы
Анализ по разным биотопам. Выше рассмотрены, перестройки ви-
довой структуры ^животного населения в периоды разных уровней
численности водяной, крысы. Возникает вопрос, захватывают ли
зти перестройки определенные биотопы' или распространяются и од-
новременно охватывают различные участки территории в пределах
района работ? ! •
Для анализа взяты пятилетние материалы по Усть-Ургульскому
стационару, где 100-метровые ловчие канавки с 10 цилиндрами
каждая охватывали, помимо типичных летних местообитаний водя-
ной крысы у кочкарниковых болот и заболоченного колка, также два
типа :луга и поле. Население мелких млекопитающих в 1978 г. (по-
следний год сухой фазы, водяная крыса в отловах отсутствовала),
характеризовалось доминирующим положением в сообществе поле-
,вой мыши, лесной мышовки, а из землероек малой и обыкновенной
бурозубок и было типичным для всех исследованных биотопов'
(рис. 54).
В 1979 г. (первый год влажной фазы, водяная крыса еще мало-
численна) разные биотопы характеризовались также сходным насе-
лением зверькбв. Везде чаще всего превалировали полевая мышь и
лесная мышовка. Происшедшая в этом году смена домйнантов и суб-
доминантов в сообществе землероек одновременно коснулась всех
угодий, где отлавливались зверьки (рис, 55).
В 1980 г., когда начался рост численности водяной крысы (год
предпика) и она стала доминантом в сообществе, это проявилось опять
же сходным образом во всех 5 биотопах. Полевая мышь и лесная
мышовка почти во всех биотопах заняли места субдоминантов. Сре-
ди, землероек доминантами во всех биотопах стали кутора и обыкно-
венная бурозубка (рис. 56). :
В год пика численности водяной крысы (1981) последняя занима-
ла по-прежнему лидирующее положение в сообществе и это косну-,
лось всех исследованных биотопов. Но произошли одновременно и
некоторые изменения в биотопическом размещении других видов
зверьков. На болотах в субдоминанты вышла полевка-экономка,
а полевая мышь переместилась на 3-е место. На лугу и поле полевая
мышь осталась на 2-м месте. Почти везде была весьма Многочисленной
и лесная мышовка. Что касается землероек, то, как и в прошлом году,.
лидирующее положение во всех биотопах занимали обыкновенная
бурозубка и кутора (рис. 57). 1 ;
В первый год после пика (1982) водяная крыса играла во всех 5
биотопах роль доминанта, среди двух следующих по численности в
четырех случаях были лесная мышовка и полевая мышь и лишь в
пятом случае один из этих видов сменил ранг численности и вышел на
4-е место (рис. 58).
Рассмотренные закономерности свидетельствуют о том, что видо-
вая перестройка структуры сообществ в ходе развития вспышки
180
экз.на 100и,, сут
1
Рис. 54. Видовые структуры животного населения в различных бпо-
•топах Усть-Ургульского стационара, 1978 г.
1 — грызуны (обозн. см. рис. 38); 2 — землеройки (обозн. см. рис. 44); а —акочкар-
виковое болото; б — заболоченный колок; в — луг с порослью осины; г — разно-
травный луг среди леса; д — ноле.
I . .
массового размножения водяной крысы протекает на конкретном
участке в целом сходно и охватывает одновременно разные биотопы.
Это особенно четко обнаруживаете^ в нарастании численности
и повышении лидирующей роли в сообществе водяной крысы, что
проявляется синхронно в разных местообитаниях. Своеобразие био-
топов, сказывающееся в облике животного населения, проявляется
не в видах-доминантах и субдоминантах, а в видах, стоящих в рангах
численности на последующих местах, которые, собственно, и харак-
теризуют видовое своеобразие ценоза данного биотопа. '
Описанная общность разных биотопов по доминантам и субдоми-
нантам связана с их миграционной активностью в условиях высокой
численности и с тем обстоятельством, что виды, доминирующие
в сообществе, всегда отличаются большей эвритопностью. В барабпн-
ском ценозе последнее особенно свойственно водяной крысе. При сни-
жении ее численности и особенно в годы депрессии животное населе-
ние биотопов становится более специфичным.
181
Рис. 55. Видовые
структуры животного ।
населения в различ-
ных биотопах Усть- .
Ургульского стацио-
। нара, 1979 г.
Усл. обозн. см. рис. 54.'?
!
I •-
- ! I. 1 . _ - Г
1 , ;Отмеченное выше сходство видовых структур животного сообще-
ства в ходе развития вспышки массового размножения водяной кры-
сы важно и в методическом отношении, так как позволяет суммиро-
вать данные отловов в разных биотопах конкретного стационара и
представлять их в годы массового размножения водяной крысы в4
единстве, ^которое и характеризует аспект данного года. (
Анализ в разные циклы вспышек. Есть ли сходство в видовой
структуре животного населения, если сравнивать соответствующие
182
экз.на ЮОц.сут
Рис'. 56. Видовые струк-
туры животного населе-
ния в различных биото-
пах Усть-Ургульского
стационара, 1980 г.
Усл. обозн., см. рис. 54.
фазы динамики чис-
ленности водяной
крысы на одном и
том же стационаре
при разных вспышках ее массового размножения? |
Для ответа на этот вопрос мы используем данные по ровенс-
кому стационару, где прослежено развитие вспышки в 1970—1972
и в 1978-1981 гг, .
Годами-аналогами по динамике численности водяной крысы
здесь были 1970 и 1978 (последний год низкой численности водяной
крысы). Они сходны по видам-доминантам и субдоминантам в сооб-
ществе землероек .и по полевке-экономке, которая была в 1970 г,
доминантой, а в 1978 г.— субдоминантом (см. рис. 47). ।
Годы 1971 по 1979 (предпик) сходны по доминантам и!субдоми-
нантам у землероек. В группе трех первых по численности видон
грызунов два (водяная крыса и полевая мышь) были общими (см.
рис. 48).
В годы 1972 и 1980 (пик) в рангах численности первые три вида
землероек были общими в обоих случаях. У грызунов из первых трех
видов два (в том числе доминант) были общими (см. рис. 49).
183
Рис. 57. Видовые структу-
ры животного населения, в
различных биотопах Усть-
Ургульс'кого стационара,
1981 г.
Усл. обозн. см. рис. 54.
Спад численности во-
дяной крысы на сле-
дующий год после пика,
на Ровенском^ стацио-
наре был в годы-ана-
логи: 1950, 1973, 1981.
Общими для всех трех
лет среди лидирующих
по численности трех
видов землероек были обыкновенная бурозубка и кутора, а для двух
лет также и малая бурозубка. У грызунов общими для трех лет бы-
ли полевая мышь и водяная крыса, а для двух лет также и северная
мышовка (см. рис. 50).
Условия существования. биоценозов Северной Барабы при раз-
ных циклах вспышек имеют отличительные черты, и последнее не мо-
жет не накладывать в каждом случае своеобразный отпечаток на
структуру животных сообществ и ее изменчивость. И все же при-
веденные материалы свидетельствуют о большом сходстве видовых
аспектов животного населения на одном и том же участке при ана-
логичных фазах динамики численности водяной крысы. Можно, сле-
довательно, говорить о повторениях видовой структуры населения
мелких млекопитающих в пределах 11-летних циклов. Временными
реперами для повторного развития сходных видовых структур в Ба-
184
Рис. 58. Видовые структуры грызунов в различных биотопах Усть-Ургуль-
ского стационара, 1982 г.
Усл. обозн. см. рис. 38, 54.
рабе служат смены фаз увлажненности в 11-летних циклах и соот-
ветствующие им фазы динамики численности водяной крысы.
Анализ по разным районам Барабы. Есть ли сходство в перестрой-
ках стуктур животного населения, если сравнивать соответствующие
фазы развития вспышек массового размножения водяной крысы в
в разных районах Барабы?
Для ответа на этот вопрос ниже рассмотрен видовой состав насе-
ления грызунов и землероек по годам-аналогам, характеризующим
соответствующие фазы динамики численности водяной крысы на
разных стационарах Барабы (табл. 28).
Последний год низкой численности водяной крысы после депрес-
сии. Это состояние в животном населении Барабы характеризу-
ют отловы зверьков: в с. Ровенское — 1970, 1978 гг.; с. Потюкано-
во — 1978 г.; с. Усть-Ургулька — 1979 г.; с. Орловка — 1979 г.
В четырех из. пяти анализируемых случаев все три лидирующие по
численности вида землероек (обыкновенная, малая и средняя буро-
зубки) были для всех стационаров общими (в пятом случае средняя
бурозубка стояла на 4-м месте). Из грызунов на трех стационарах
были общими полевая мышь, полевка-экономка и северная мышовйа,
а на двух стационарах — узкочерепная полевка. Наиболее сходными
по видовому составу были ценозы в Ровенском в 1970 г. и в Потю-
канове в 1978 г. (см. табл. 28).
Фаза подъема численности (предпика): Ровенское — 1971,
1979 г..; Потюканово — 1979 г.; Усть-Ургулька — 1980 г.; Орлов-
ка — 1980 г. Среди первых трех видов по численности общими во
всех 5 случаях были обыкновенная и малая бурозубки, в трех —
кутора и в двух — средняя бурозубка. Аналогичными по землерой-
кам были видовые аспекты в Ровенском в 1971 г. и в Потюканове в
1979 г.; в Ровенском в 1979 г.; в Усть-Ургульке в 1980 г.; в Орловке
в 1980 г. По грызунам общими для всех случаев была водяная крыса
и для четырех лет — полевая мышь (см. табл. 28).
185
Таблица 28
Общность видового состава доминирующих видов мелких
млекопитающих в разные фазы динамики численности
водяной крысы । ।
Год ; i , Место : - 1 Землеройкф : । - ( Грызуны '
! 'i ’L i 1
! 1970 ' __ - ! ; 1 Ровенское , 1 1 1, 4, 6, 5 । I .3, 4, 7
1978 » l,i 4, 5, 6 'J 1„ 3, 2
. 1978 Потюканово ; ; i | 1, 4, 6 I 3, 7, 4
! 1979 Усть-Ургулька J 1, 6, 4 ! 1, 7, 10 1
1979 . Орловка ! j 1, 4, 6 1 1 1, 8, 5
1 - 1 1 . ! - II! । 1 '' 1
1971 т-» ! ’' । Ровенское j 1, 4, 6 2, 4, 1
1979 ' » 1. 1, 4, 3 1, 6, 2
1979 Потюканово 1 । 1, 4, 6 1 ’ 1, 2, 10
, 1980 Усть-Ургулька । 3, 1, 4 2, 1, 7
1 1980 . i Орловка 1, 4, 3 2, 8, 14
1 пР 1 1
'1972 Ровенское i ‘,1, 2, 4 2, 11’ 1
1980 » '1, 4, 2 2, 1, 9
i ‘ 1980 Потюканово s 4, 1, 3 2, 5, 1
1981 Усть-Ургулька 1, 3, б 2, 1, 7
! , . IV |
. 1950 Ровенское 3,1,6 2, 1, 7
1 1973 » 1, 3, 4 1, 2, 7
1981 “» 1, 4, 3 2, 1, 9
1981 Потюканово- 3, 4, 1 2, 7, 1
1974,[ Черновка 1, 4, 5 ' 1, б, 2
1982’ Усть-Ургулька Нет данных 2, 7, 1
г V 1
।. 1974 Ровенское * 1, 2, 3 5, 1, б
1974 Коновалов!» 1, 2, 4 1, ,3, 4
1982 Ровенское 1, 4, 6 1,19, 7
1982 Потюканово i 1, 4 1, 5
Примечание. I— последний год низкой численности;.
! II — предпик; III — пик; IV — спад численности (1-й год после
' пика); V — спад численности (2-й год после пика).- 1—8 — виды ,
; . землероек (усл. обозн. см. рис. 44), 1—14 —виды грызунов (усл.
1 обозн. см. рис. 38).
; ; 1 .
Фаза пика численности: Ровенское — 1972, 1980 гг.; Потюкано-
во —t 1980 г.; Усть-Ургулька — 1981 Р. (см. табл. £8). Общей для
всех лет была обыкновенная бурозубка, для трех случаев — малая
бурозубка, для.двух лет — крупнозубая! бурозубка и кутора. У гры-
зунов во всех случаях были общими в первой тройке водяная крыса
и полевая мышь (см. табл. 28). ;
Фаза спада численности (1-й год после пика): Ровенское — 1950,
1973, 1981 гг.; Потюканово — 1981 г.; Усть-Ургулька — 1982 г.;
I . . ''/
186
Черновка — 1974 г. Общими во всех случаях были обыкновенная
бурозубка, в четырех случаях кутора и малая бурозубка, а из грызу-
нов во всех случаях — полевая мышь и водяная крыса и в четырех
случаях — северная мышовка (см. табл. 28).
Фаза спада численности (2-й год после пика): Ровенское — 1974,
1982 гг.; Коноваловка — 1974 г.; Потюканово — 1982 г. Общими для
землероек во всех случаях были обыкновенная бурозубка, в трех слу-
чаях — малая и в двух — крупно зубая бурозубки., а для грызунов
во всех четырех случаях — полевая мышь и в двух — мышь-малютка
(см. табл. 28). ।
Из рассмотренных материалов следует,> что перестройки видовой
структуры населения грызунов и землероек в ходе развития вспыш-
ки массового размножения водяной крысы имеют много общих черт
в различных, районах Барабы. Эти перестройки видовой структуры
животного населения в Барабе происходят неодновременно, как и
вспышки массового размножения водяной крысы. , । - ,
Можно сказать, что в аналогичные фазы динамики численности
водяной .крысы видовая структура животных сообществ в разных
частях Барабы очень сходна| по видам-доминанТам и субдоминантам.
В порядке предварительного вывода можно сказать, что карта Бара-
бы, на которой оконтурень) территории предпика, пика,_ 1-го и 2-го
годов спада численности водяной крысы, отражает площади весьма
сходных видовых структур животного населения.
В методическом отношении этот вывод представляет несомненный
интерес, так как позволяет по характеристике очагов водяной крысы
судить о населении мелких млекопитающих соответствующей тер-
ритории. Вместе с тем при обзорах и прогнозах развития вспышки
массового размножения водяной крысы ।полезно щзменить или до-
полнить существующую систему картирования очагов. Помимо
принятого деления очагов на прогрессирующие, действующие, про-
горевшие, потенциально опасные [Максимов, 1967], можно диффе-
ренцировать их также и на следующие категории: предпик числен-
ности водяной крысы, пик, спад численности (1-й год после пика),
низкая численность (2-й год после пика). Эти градации будут отра-
жать одновременно и площади сходных) видовых структур животного,
•населения. Ч i 1
Таким образом, географические особенности в перестройке видо-
вых структур сообществ выявляются через анализ вспышек массо-
. вых размножений водяной крысы. । ;
. Проведенный анализ позволяет дифференцировать отдельные фазы
динамики численности водяной крысы в 11-летнем цикле на две кате-
гории применительно: 1) к конкретному участку (локальный очаг)
и 2) ко всей зоне вредоносности вида. Если на конкретном участке фа-
зы предпика, пика, спада численности'водяной крысы длятся по од-
ному году и затем несколько лет продолжается депрессия числен-
ности, то в целом в зоне вредоносности вспышка массового размно-
жения длится 3—5 лет, соответственно продолжительности влажной
фазы.
187
Именно в этом плане надо рассматривать и территориальную ди-
намику перестройки видовой структуры животного населения.
Продолжительность существования отдельных видовых аспектов
определяется, с одной стороны, длительностью влажной и сухой фаз
в 11-летнем цикле и, с другой (в! пределах влажной фазы) — динами-
кой численности водяной жрысы.
В заключение еще раз обратим внимание на описанное выше
сойпадение вспышек массового размножения водяной крысы с влаж-
ными климатическими фазами. ; Развитие же локальных очагов в
пределах влажной фазы подчиняется своим законам, поэтому пики
вспышек в пределах 1 зоны вредоносности не совпадают во времени.
Именно этим экологическим особенностям в развитии и перемещении
по территории очагов массового размножения водяной крысы под-
чиняются и перестройки видовых структур животного населения..
В пределах конкретного года видовые спектры мелких млекопитаю-
щих в Барабе поэтому различны, но они имеют общие черты на всей
площади со сходной фазой динамики численности водяной крысы. '
9 I
Г л* а в а VI \ г
ЦИКЛИЧНОСТЬ ПРОДУКТИВНОСТИ СООБЩЕСТВА
МЕЛКИХ МЛЕКОПИТАЮЩИХ 1
Описанные выше видовые перестройки сообществ мелких млекопи-
тающих, ритмы которых подчинены сменам фаз увлажненности и
влиянию вспышек массового размножения водяной крысы, представ-
ляют интерес с точки зрения изменчивости биологической продук-
тивности барабинского jсообщества животных. Поскольку мы не про-
водили абсолютных учетов численности, а имеем только относитель-
ные данные, мы оценивали продуктивность сообщества, применяя
расчеты относительной биомассы. При наличии средних масс особей
каждого вида можно сосчитать относительные биомассы, значения
которых и использовать как сравнительные оценки, сопоставляя
продуктивность сообществ в каждом вегетационном периоде. Сравт
нительной биомассой мы называем произведение среднестатистиче-
ской массы особей каждого вида на их относительную численность.
При расчетах средних масс особей разных видов мы использовали
все возрастные и половые группы в том соотношении, в котором они
отлавливались канавками в сезоны работ (табл. 29).
В -исследуемом сообществе, состоящем из двух, по-разному спе-
циализированных групп животных, преобладали землеройки. В от-
1 В период с-1973 по 1977 г. сбором и обработкой материалов по продуктив-'
пости сообществ мелких млекопитающих в окр. с. Ровенское занимались
Л. Н. Ердаков и В. Е. Сергеев.
188
Т а б .т [ п ц а 29
Средняя масса животных из разных популяций видов
мелких млекопитающих в окр. с. Ровенское
Вид п М±т > и । Ч ’
Полевая мышь : 50 18,3±0,84 5,91
Мышь-малютка 38 i 6,1+0,17 1,09
Хомяк обыкновенный 26 303,3+20,6 105,03
Джунгарский хомячок Полевки рыжая ‘ 26 1 23,6* 15,9±0,7 3,5 ! S
красная 27 18,8+0,7 3,4 ।
красно-серая 20 23,1±0,06 0,3 ' .
Водяная крыса 50 106,8+0,9 7,0
Полевки экономка 38 30,4+1,9 11,5 к 7,79 ' *
пашенная 22 30,2+1.7
узкочерепная 27 24,7+1,13 5,89
Северная мышовка 30 7,0±0,31 1,67
Водяная кутора Бурозубки. ' , обыкновенная 31! 13,9* 7,4+0,4 2,11 ' :
крупнозубая 9 5,7+0,8 2,3 :
арктическая . 17 6,2+0,27 0,88
средняя 20 4,0+0,22 1,02
малая 26 3,1+0,17 0,86
крошечная 9 1,99+0,14 0,42
Обыкновенная полевка 23,4* 7 1 i
* Данные из книги «Сообщества мелких млекопитающих Ба-
рабы» [1978].
дельные годы преимущественная численность этой группы в сообще-
стве была настолько очевидной, что доминирующий вид (Sorex агапе-
us) составлял более половины от всех мелких млекопитающих (имен-
но такая ситуация сложилась в 1974 г.). Между тем, биомасса земле-
роек в барабинском сообществе ничтожна, она составляет в среднем
1/10 часть общей биомассы (табл. 30). Размах колебаний численности
и биомассы в популяциях сообщества велик, что может говорить о
малой стабильности этой системы. Но последнее возможно только
при одной оговорке: колебания должны быть неупорядочены (сто-
хастические). Наши же наблюдения позволили предполагать в изу-
ченном сообществе некоторую упорядоченность, на основании кото-
рой даже при таком большом варьировании характеристик систем
можно говорить о ее стабильности и высоком уровне организации.
Рассмотрим группу грызунов. Она наиболее богата видами и
составляет, как мьгуже заметили, основную массу населения.
Обычно в наших широтах основную долю биомассы продуцируют
2—3 доминирующих вида грызунов (табл. 31). Особенностью сооб-
щества являются относительно мелкие размеры составляющих его
189
/
Таблица 30
Относительная численность и биомасса мелких млекопитающих в сообществах Барабы
_____ — Сухая фаза | Влажная фаза Сухая фаза.< | - Влажная фаза
Сообщество - _ Биомасса _ —- — Численность ~
— кг/100 цил.-сут 1- - кг/100 цил.-сут 0/ /0 особь/100 цил.-сут' % особь/100 цил.-сут %
Землеройки Грызуны 0,02—0,13 - 0,17—0,73' "14,3 __ 85,7 0,014—0,21. 0,16-10,83 1,96 98,04 3,2 -15,8 . 3,4-28,2 37,5~ 62,5 .. 2,38—24,9 5,6-107,4 19,4 80,6
В целом. . . 0,22—0,84 юо,о- 0,22-11,03 100,00 10,93-45,25 ~~йо,о' 13’72—132,2 100,0
Таблица 31
Грызуны, создающие в совокупности основную биомассу в районе исследований
Вид _ 1966‘ Год наблюдений
1967 1968 1969 1970 1971 1972 1973 1974 1978 1979 1980 1981 1982
Полевая мышь Обыкновенный хомяк ь + ' + + , - + + + + ь + +
Водяная крыса t +- + 4- ь + ’ + + + + +- -+
Полевка-экономка Узкочерепная полевка 4- + + + + + + — — + .
особей. Тем не менее масса особей заметно различается, и зверьки ,,
одного вида могут превосходить по этому параметру зверьков друго- ।
го вида на порядок и более. Так, соотношения массы водяной крысы
п лесной мышовки или обыкновенного хомяка и узкочерепной по- *
левки различаются более чем на порядок (см. табл. 29). Отсюда i 1
двойственный характер возможных изменений биомассы. Она может )
увеличиться с ростом численности животных, а также благодаря пе- 1
регруппировке видов при условии возрастания числа крупных осо-
бей, несмотря на- то, что сообщество в целом испытывает депрессию.
Таким образом, динамика численности видов в сообществе грызу-
нов может и не^отражать динамику биомассы, хотя обычно (и по лите- ;
ратурным данным) не вызывает сомнений взаимообусловленность ;
этих характеристик. Убедительной иллюстрацией могут служить
уловы животных на Ровенском стационаре в 1973 и 1974 гг., когда i'.
численность была соответственно 25,12 и 26,18 особей на 100 цил.-сут. '<
Биомассы грызунов в это же время составляли 1,13 кг и
0,5 кг/100 цил.-сут. Наблюдалась парадоксальная зависимость: ;
ic возрастанием численности биомасса упала почти вдвое. Объясни- :
ется это тем; что нарастание численности в 1974 г. происходило за I
счет мелких форм грызунов при резком сокращении крупных. ;
Обратим внимание на изменение продуктивности сообщества t. J
за время исследований и на возможность его интерпретацйп в рам- }
ках • экологической сукцессии. Прежде всего опишем сам процесс.
। . К 1966 г.2 относительная биомасса грызунов возросла, но к 1968 г. i
вновь снизилась. Это период окончания засушливой фазы в Барабе. (
Ориентируясь на литературные данные [Глотов, 1969], заметим, что \ ' t
в 1965 г. состояние сообщества по численности и видовому составу
сопоставимо с 1967 г. ,
Влажный период, начавшийся в 1969 г., сначала не повлиял на
изменение биомассы. В 1970 г. стала заметна тенденция к возраста-
нию биомассы грызунов, которая в 1972 г. резко, увеличилась. За-
метим, что увлажненность региона увеличивается, достигая апогея
в 1972 г. Окончание влажного периода знаменуется уменьшением
биомассы, причем уже в 1-й год влажной фазы (1979) биомасса до- !
стигает уровня 1967 г. (579,8 и 500,8 г соответственно). Б конце су-
хой фазы биомасса грызунов сохраняется такой же, как в конце
предыдущей (1967), т. е. довольно низкой—0,65 кг. С ростом увлаж-
ненности она начинает возрастать, достигая максимума в 1980 г. ;
(1,3 кг/100 цил.-сут). :
. Картина изменений биомассы за ряд лет дает основание^ предпо- • 1
дожить, что в барабинском сообществе грызунов имеется своеобраз- .
ная многолетняя цикличность продуктивности, связанная с фазами
увлажненности региона. । Ь
Динамика продуктивности (сообщества землероек сходна с тако- ' '
вой грызунов по размаху колебаний. Относительные численность
и биомасса этих животных при депрессии на порядок отличаются от
значений тех же характеристик при подъеме численности. Сообще- 1
2 В расчетах мы пользуемся данными, приведенными к одним единицам
и взятыми за строго одинаковые отрезки времени.
191
ство землероек в сравнении с грызунами имеет большее постоянство
структуры, и основу биомассы в нем создает популяция постоянно до-
минирующего вида — обыкновенной бурозубки. В отдельные годы
заметную долю в общей биомассе (но несравнимую с доминантой)
создают водяная кутора (в 1973 г. она составила 21,5% биомассы)
и крупнозубая бурозубка (в 1974 г.— 12%). Прочие члены сообще-
ства не составляют и 15% валовой его продукции.
В период исследований у зейлероек отмечено, два подъема про-
дуктивности. В 1967 г.—кратковременный подъем, ограниченный
. всего одним вегетационным периодом. Уже на следующий год заме-
тен спад продуктивности сообщества землероек. Происходит он
.синхронно с грызунами, но депрессия у насекомоядных затягивает-
ся на пять лет. Если первый спад (1968 г.), возможно, обусловлен
крайним иссушением территории, то дальнейшая депрессия на фоне
общего повышения обводненности могла быть вызвана только общим
повышением численности грызунов. Если исходить из точки зрения
о независимости рассматриваемых сообществ, причины депрессии
приходится искать в неблагополучном для землероек абиотическом
фоне. Но лучше всего считать так: 1968 г,— очень сухой и неблаго-
приятный для всех мелких млекопитающих; 1969 г.— увеличение
увлажненности, почва глубоко просушена, но влажные напочвенные
подстилка и опад создают убежища для землероек, растет и числен-
ность грызунов; 1970 г.— влажность । продолжает расти, бурная ве-
гетация, численность грызунов стремительно растет, увеличивается
и численность землероек, но незначительно, продукция их увеличи-
вается в 3 раза по сравнению с прошлым годом; 1971 г.— продол-
жает расти численность грызунов, большая обводненность террито-
рии, продукция землероек остается на уровне прошлого года и даже
слегка снижается; 1972 г.— продукция грызунов достигает максиму-
ма за период наблюдений, а землеройки сокращают свою биомассу
до минимума.
Только в 1973 г. при сохраняющемся обводнении территории,
но в условиях резкого спада продуктивности сообщества грызунов,
землеройки наращивают свою биомассу на порядок в сравнении с-
прошлым годом, а в 1974 г., когда началась сухая фаза при продол-
жающемся спаде продуктивности грызунов, землеройки еще вдвое
увеличили биомассу. ; : . J
Такие же изменения претерпевает биомасса землероек и в сле-
дующую фазу увлажненности. Она минимальна в конце сухого
периода и в 1978 г. составляет лишь 0,06 кг/100 цил.-сут. В первый
год повышения влажности (1979) происходит еще некоторое, незна-
чительное, снижение биомассы у этих зверьков. С дальнейшим
ростом влажности неизбежно начинается увеличение биомассы земле-
роек — в 1980 г. они уже составили 0,07 кг/100 цил.-сут. В то же время
в окрестностях с.Ровенское наступает засуха во второй половине
1980 г. Масса землероек под влиянием этой засухи снова начинает
снижаться и при непрерывной засухе падает до 0,05 кг/100 цил.-сут.
Таким образом, при различных погодных условиях и ситуациях
обводненности территории продуктивность сообщества землероек
192
может быть сходной. Единственно непротиворечивая связь на протя-
жении периода исследований сохранилась между продуктивностью
землероек и грызунов. Эти процессы находятся между собой в проти-
вофазе, чего нельзя сказать о численности этих групп животных,!
Численность землероек может возрастать с ростом численности гры-
зунов, но биомасса их изменяется в обратной пропорциональности
биомассе грызунов, за исключением периодов общей депрессии мел-
ких млекопитающих в регионе. , j
Рассмотрим связь динамики продуктивности землероек с фазами'
увлажненности Барабы. На основании имеющихся материалов
можно предполагать, что эти процессы связаны обратной зависимо-
стью, хотя имеются и исключения. Прежде всего, это рост продуктив-
ности землероек в последний год фазы обводненности и низкая про-
дуктивность 'в 1966 г. (засушливая фаза). Но как мы уже отмечали
выше, есть экологические причины, объясняющие это.
По-видимому, управление жизнью популяций происходит не-
только абиотическими факторами, которые не всегда могут быть
ведущими. Действительно, начавшаяся благоприятная для повыше-
ния продуктивности землероек обстановка тормозится, а потом и
сводится на нет ростом продуктивности грызунов. В то же время ис-
сушение территории не может серьезно повлиять на продуктивность
землероек, снизить ее, потому что одновременно снижается биомасса
грызунов, и землеройки получают дополнительный и, вероятно, бо-
лее сильный стимул для своего развития.
Разбор динамики продуктивности наблюдаемых сообществ за
период исследований позволяет не только уяснить особенности функ-
ционирования каждого из них. Как можно заметить, существует
связь соотношений продуктивности грызунов и землероек, ,на осно-
вании которой можно сделать некоторые выводы о всем сообществе
мелких млекопитающих Барабы..
Увеличение биомассы на единицу потока энергии, происходящее
в конце каждого периода повышенной влажности, является нормой
для цикличного функционирования наблюдаемой системы.
Для выяснения хода экологической сукцессии необходимо сравнение
нескольких аналогичных фаз и расчета порядка происходящих изме-
нений.
В нашем распоряжении для сравнения имеются материалы Ро-
венского стационара за 1950 и 1973 гг. Эти годы аналогичны по эко-
логической ситуации в сообществе мелких млекопитающих 1981 г.,
они сразу следуют после пика численности водяной крысы (1-й год
спада численности). Первые два года приходятся на влажную фазу
в целом по Северной Барабе.
Соотношение биомассы мелких млекопитающих в сообществе в
годы со сходной климатической и экологической обстановкой в
окрестностях с. Ровенское было следующим (г): ,
Год Грызуны Землерой- ки Общая
1950 1256,3 109,0 1365,3
1973 1130,5 187,5 1318,0
1981 469,9 49,3 519,2
13 а. А. Максимов, Л. Н. Ердаков
193
t
Как видно из приведенных данных, биомасса грызунов и землеро-
ек с каждым новым циклом снижается. Рассмотрение продуктив-
ности сообществ в разные фазы увлажненности территории, затруд-
„ненб инерционностью изменений этих сообществ. Ведь наступление'
очередной фазы знаменуется очень постепенным увеличением марки-
рующей характеристики (влажности). Это обусловливает постепен-
ную перестройку сообщества и неотличимость первого года влажной
фазы от последнего сухой. Когда же!мы рассчитываем средние’ха-
рактеристики продуктивности, то в каждой выборке у нас присут-
ствуют все годы анализируемой фазы. Это обеспечивает трансгрессию-
выборок и нечеткость различий средних значений. В то же время
’при использовании только ярко ^выраженных лет । из; разных фаз
увлажненности (пренебрежение йри расчетах пограничными годами).
мы существенно теряем представительность наших и без того неболь-
ших выборок.^ Сознавая эти трудности анализа, мы тем не менее ис-
пользовали для расчетов все имеющиеся в нашем распоряжениидан-
ные по каждой фазе (табл. 32).
Оценивая продуктивность сообщества по средним значениям био-
массы, можно видеть различия в зависимости от фазы увлажненности
территории Барабы (см. табл. 32). Так, в окрестностях с. Ровенское
биомасса грызунов достоверно возрастает в фазу обводнения (t =
= 2,42; Р = 0,95). Землеройки в этом районе не изменяли биомассы
-по фазам увлажненности и тем самым стабилизировали продуктив-
ность сообщества в целом.1 Видимо, поэтому общая биомасса не име-
ла достоверных ^различий в сухую и влажную фазы (£ = 1,93; Р <Z 1 "
<0,95).
Насекомоядные, как указывалось выше, обычно выступают в
противофазе к циклам продуктивности грызунов. Особенно хорошо-
заметно это при сравнении биомассы землероек в разные фазы увлаж-
ценности территории из окрестностей д. Коноваловка. Здесь био- ‘
масса насекомоядных достоверно возрастала во время сухой фазьг
(i = 3,95; Р = |0,99). Стабилизировали же всю систему мелких мле-
копитающих здесь грызуны, и общее1 значение биомассы также было
неразличимым в разные фазы (£ = 2,31; Р = 0,95).
Таким образом, продуктивность сообщества мелких млекопитаю-
щих в Барабе испытывает циклические изменения каждые 5—6 лет*
а на! 9—11-й год цикла обводнения она достигает максимума. В ходе
этой цикличности общая! относительная биомасса животных изменя-
ется по спирали, т. ё.) несмотря на повторяющиеся колебания,
максимальная продуктивность ib период, наших работ снижалась с
каждым 11-летнпм циклом. ’
Продуктивность сообщества мелких млекопитающих Барабы ин-
тересно сравнить с другим сообществом, близким по видовому соста- -
ву. Для такого сравнения наиболее подходит сообщество поймы
Среднего течения' р. Оби. Видовой состав мелких млекопитающих ч
здесь тот же, что и в Барабе, но цикличность природных факторов
иная. Циклы в пойме подчинены разливам реки. Сукцессии здесь
проходят по типу катастрофических, но одинаковый видовой состав
194 ;
Таблица 32
Относительная биомасса (г/100 цил.-сут) и численность (особь/100 цил.-сут)
мелких млекопитающих в разные фазы увлажненности
. < 1 Сухая фаза Влажная фаза
Сообщество1; | 1 М±т : 1 а и «I 1 i1 О и
! 418,9+78,7 РовеАское { , ! J i;
Грызуны 8 1 222,7 53,1 8- 1024'7+237,6 672,1 . 65,6
Землеройки 5 i , 1 118,7±58,5 130,9 110,3 8 76,2± 181,4 j ( 58,2 76,4
В' целбм 5 ! 569,1+143,5 320,8 56,4 8. 1100,9+236,0 667,5 60,6
1 Коноваловка
Грызуны 51 449,9±110,1 246,3 54,7 2 235,8+73,4 103,7 43,9
Землеройки 5 113,9+7,6", 17,1 1 15’° 2 33,7+18,8 । 26,6 78,9
В целом 5 563,7+115,1 257,3 i 45,6 2 269,5+54,5 .. i 1 77,1
Усть-Ургулька i
Грызуны 1 207,1 1 4 3301,5+2509,7 1 5019,4 152,0
Землеройки 1 23,8 1 „ 1 3 160,2+24,0 ) . 41,6 ' 25,9
В целом 1 230,9 1 “ I- 3 | ,4284,3+ 3374,8 [ 5845,4 ! 136,4
должен обеспечивать черты общности между этими сообществами
[Максимов и др., 1981]. f
Величину биомассы грызунов в пойме среднего течения р. Общ
так же как и в Барабе, в значительной степени определяет числен-
ность водяной крысы. Высокая плотность популяции ее в 1968 и
1972 гг. обусловила высокие показатели биомассы в эти годы. В пой- i|
ме кроме своеобразия динамики численности сообщества, связанной
с разливами реки, существует собственная динамика продуктивности |
сообщества грызунов. Продуктивность сообщества здесь зависит не
от доминирующего вида и не от численности видов, а от численности
водяной крысы. Последняя — индикатор продуктивности сообщества ; I
•мелких млекопитающих поймы. : I । ' | '
Размах варьирования относительной биомассы грызунов в пойм®
заметно уступает таковому численности (табл. 33). Биомасса в пой-
ме более стабильна, чем в Барабе, но средняя । продуктивность пой-
менного сообщества почти вдвое ниже барабинского. В то же время
по относительной численности рассматриваемые сообщества вполне
сравнимы. Соотношения численности и биомассы в них различны по
вариабельности. В пойме р. Оби численность варьирует намного
сильнее, чем в Барабе.
Последнее можно объяснить разнообразием структур ^сообщества.
В пойме меньше разнообразие видов, лучше выражен доминант, т. е.
система проще устроена и в силу этого менее равновесна. Возможности
13*
195
Таблица 33
Относительная численность и биомасса мелких млекопитающих сообщества
• поймы Оби [Максимов и др., 1981]
Сообщество \ Биомасса (предел колебаний) Численность (предел колебаний)'
кг/100 цил.-сут , % : особь/100 цил.-сут ' . % .
Землеройки 0,072 (0,001—0,02) : 7,66 8,6 (0,1—23,0) 32,19
Грызуны 0,868 (0,073—2,164) 92,34 18,11 (1,22—40,42) 67,81
В целом 0,94 (0,075—2,182) 100,00 26,61 (2,03-46,51) 100,00
; 1 4
компенсирования падения численности вида-доминанта^ в пойме прак- .
тически отсутствуют. Дополнительный размах колебаниям числен-
ности грызунов поймы придают разливы реки.
Биомасса землероек в пойме в сравнении с таковой грызунов
незначительна (см. табл. 33) и колеблется несколько меньше. В,то
время .как биомасса грызунов может от года к году изменяться
на два § порядка и более, биомасса землероек изменялась между эк-
стремумами в период наших наблюдений на один порядок. В связи с
небольшими различиями массы тела у представителей разных видов
биомасса землероек больше] чем у грызунов, зависит от численности
зверьков. Размах варьирования численности насекомоядных в пой-
ме намного больше, чем в Барабе, что связано с особенностями гидро-,
логического режима р. Оби. Нельзя не отметить, что и вариабель-
ность численности этой группы животных в пойме больше, чем у гры-
зунов. Это лишний раз подтверждает, что разлив реки — более
сильный элиминирующий фактор для землероек, чем для грызунов.
Этот же фактор наиболее сильный в своем воздействии на сообщество,;
нежели многолетние фазы увлажненности Барабы. Именно поэтому
численность, продуктивность животных, особенности их доминиро-
вания; и многие другие экологические показатели в этих сообществах
различаются. Общность видового состава, напротив, обусловливает
элементы подобия в сообществах. Так, соотношение продуктивности
насекомоядных и грызунов в пойме оказывается таким же, как во
влажную, фазу в Барабе. Соотношение численности этих групп жи-
вотных в пойме такое же, ;каК в сухую фазу в Барабе^
i
Глава VII
- КОНЦЕПЦИЯ ЭКОЛОГИЧЕСКИХ СУКЦЕССИИ
КАК ЦИКЛИЧЕСКОГО БИОЛОГИЧЕСКОГО ПРОЦЕССА
j СУЩЕСТВУЮЩИЕ ПРЕДСТАВЛЕНИЯ
ОБ ЭКОЛОГИЧЕСКИХ СУКЦЕССИЯХ
' I
Биогеоценотический процесс происходит непрерывно и приводит к
изменению компонентов в сообществе, т. е. процесс состоит в том,
что каждое воздействие одного компонента на другой нарушает ус-,
тановившийся тип, взаимоотношений, создавая новкй. Таким обра-
зом все время идет нарушение одних взаимоотношений и создание
новых — непрерывная перестройка биогеоценозов. Влияние на со-
общества извне происходит постоянно, поэтому биогеоценотический
процесс никогда не останавливается. Он все время приводит к смене
(сукцессии) одних биогеоценозов другими. С течением времени про-
цесс этот может замедлиться, но не прекратиться [Сукачев, 1972].
Эволюция направляется естественным процессом сукцессии, при-
сущим всем экосистемам, в сторону, задаваемую сукцессией и назы-
ваемую возрастанием зрелости. Эволюция идет в трм же направле-'
нии, что и сукцессця. По данным Р. Маргалефа [Margalef, 1968],
представление о сукцессии в экологии должно занимать такое же
место, как представление об эволюции в общей биологии. С энерге-
тических позиций сукцессия — такое неустойчивое состояние сооб-
щества, которое характеризуется неравенством двух показателей:
общей продуктивности (П) и энергетических трат всей системы
(П/Д) [Odum,, 1969]. В устойчивой, стабильной системе эти показа-
тели почти равны (П/Д близко 1). Количество первичной продукции,
создаваемой автотрофами, полностью расходуется на разных трофи-
ческих уровнях. 1
Экологи признают, что у сукцессии существует определенное
направление, но мнения расходятся, когда надо формулировать ра-
бочее определение или даже некоторые категории или общие черты
этого направления. Заметными среди используемых критериев сук-
цессии является тенденция к увеличению: 1) биомассы, 2) стратифи-
кации структуры, 3) сложности экологических взаимодействий и
4) видового разнообразия. Иногда предлагается также максимиза-
ция общей продуктивности. Другим часто используемым принципом
является увеличение стабильности сообщества, т. е. уменьшение ча-
стоты колебаний п большая независимость от условий среды. Во мно-
гих случаях сукцессию связывают с географическими структурами,
и тогда становится очевидной важность локальных климатов для
различных стадий.
Основным направлением увеличения стабильности сообщества
считается изменение экосистемы в сукцессиях. Сообщества приобре-
тают известную степень автономности и независимости от окружаю-
щих условий, подчиняя свою жизнь не флуктуациям внешней среды,
197
а ‘вырабатывая собственные эндогенные .ритмы. Механизмами этих
преобразований служат направленные изменения внутренней струк-' '
туры ценозов: увеличение видового разнообразия, усиление специа-
лизацни видов (расхождение по экологическим нишам)! В ходе сук-
' цессии усиливаются стратификация и, пространственная гетероген-
ность сообщества. , “ ! ;- ] । ' : !
Видовое разнообразие сообществ :по современным экологическим .
.представлениям тесно связано со стабильностью экосистем.. Обще- 1
признано,- что наиболее стабильные системы1 отличаются и наиболь- •
шим видовым .разнообразием [Pimlott, 1969; McNaughton, Wiolf,
1970; Bezzel, Reichholf,, 1974]. Эта стабильность считается, с одной
стороны? следствием высокой предсказуемости абиотических изме-
। нений [Slobodkin, Sanders, 1969};. ас другой — функцией самого
разнообразия, т. е. сложности возникающих биотических взаимо-
связей [Whittaker, 1969; Pimlott, 19691. С помощью математических
моделей показано, что большое разнообразие группировок зависит
} от времени существования местообитания, оно увеличивается в ходе
!эволюции сообщества [Whittaker, 1972]. Увеличение видового раз-
нообразия сопровождается низкой численностью и спорадическим
распространением каждого вида [Recher,; 1971 ].
Некоторыми авторами ставится под сомнение однозначная связь
'видового разнообразия И| стабильности сообществ [Whittaker, 1972;
Buzas, 1972]. Основанное'чаще всего на «здравом смысле» положе1- •
ние о взаимосвязи устойчивости и разнообразия проверялось на ма-
। тематических моделях. Оказалось, что с возрастанием сложности
модельных сообществ1 устойчивость йх снижается [Мау, 19731. Хо-
тя общепринято [Одум, 1968], что в ходе сукцессии разнообразна
должно нарастать по мере приближения к устойчивому состоянию.
Имеется ряд исследований' [Whittaker,. 1969 ], которые показывают^
что на поздних, климаксовых, этапах сукцессии видовой состав мно-
гих группировок несколько обедняется в сравнении с промежуточ-
ными стадиями. По Н. М. Черновой [1977], 'наибольшим видовым
разнообразием ,и наиболее равномерным соотношением численности
отдельных видов характеризуются средние, а не климаксовые этапы
сукцессий. Это же ранее Отмечал А. М. Гиляров [19691, а позднее — .
Ю..И. Чернов [1971;]'и С. И. Ананьева [1977]. .
...Скорость сукцессий сообществ бывает различной в зависимости-
от того, чем они обусловлены. Медленные Фукцессйи связаны со сме-
' ной ландшафтов, (изменениями ^климата. Бол»© быстрые сукцессии
обусловлены геоморфологическими процессами, еще быстрее про- -
исходят смены; при активном воздействии человека. !В некоторых
[классификациях сукцессий [Лавренко, 1959; Сукачев, 1972; Чер-
нова, Былова; 1981 ] наряду с долгосрочными указаны и быстро про-
текающие.» Их называют катастрофическими сменами, и они обычно
(для фитоценозов.) происходят в результате сильного и внезапного,
воздействия извне. ।
Темпы сукцессий на разных этапах неодинаковы, они, кроме
внутренних законов, определяются еще и внешними условиями.
.Сукцессия в современном понимании — это не любая последователь- .
198
I
пая смена организмов, а закономерный процесс развития сообщества,
связанный с неизбежным изменением им окружающей среды,- Там»
где физические условия .экстремальны или подвержены глубоким из-
менениям, организм не в, состоянии преобразовать среду. В таких
случаях сукцессии кратки и обрывочны. Если восстановление сооб-
щества начинается после его полного или частичного уничтожения
в результате катастрофических воздействий (массовое размножение
вида, разлив реки), то, мы имеем дело с кратковременной Сукцессией
сообщества [Сукачев,! 1972 ]. । . г .
[Новый тип сообщества возникает в том случае, когдаIизменения
всех или большинства его компонентов стаЬги столь значительными,
что сообщество стало качественно другим. Группа экзогенных сук-
цессий включает в себя кроме климатогенных еще и смены, связан-
ные с процессами расселения растений или животных, когда новый
для данной местности вид, расселяясь,'внедряется в сообщество, вы-
тесняет некоторых аборигенов и этим меняет все сообщество в целом-
(зооселектоценогениый процерс) [Сукачев, 1944]. Сукцессии, имею-
щие локальный характер, обычно внезапно начинаются и быстро
протекают, будучи вызванными какими-либо коротко, но сильно
действующими причинами. После внезапного уничтожения или
сильного нарушения сообщества на остатках его । относительно бы-
стро развивается производный биогеоценоз. Не исключена возмож-
ность того, что часто следующие (хотя бы и с промежутками) одно-
родные смены погоды отражаются на биогеоценозе так сильно и вы-
зывают такие изменения, что в течение нескольких периодов приво-
дят уже к сукцессиям. 1
Описаны еще сезонные, пли поточные, сукцессии. В ряде случа-
ев последовательная смена стадий развития сообщества точно совпа-
дает с течением вегетационного сезона, начинаясь весной и обрыва-
ясь в начале зимы. Погодные сезонные условия оказываются в роли
факторов, запускающих сукцессию, позволяющих ей начаться, но
в последующем отдельные этапы зависят только от предыдущих и
являются их следствием. Факторы среды не могут определить пре-
емственность биологических циклов, а являются лишь необходимым
условием для ее осуществления. Сукцессия протекает так, что ее
завершающие этапы наступают осенью при исчезновении физиче-'
ских условий. । ,
Биоценозы, не являясь категорией застывшей, неизменной,- не-
прерывно обновляются, представляя собой динамическую систему.
Несмотря на наличие определенного постоянного ядра, сообщество
в целом перестает существовать и сменяется другим. Это и будет
сукцессия. Есть авторы, защищающие идею ритма в упорядоченной
смене сообществ [Кашкаров, 19331, хотя это и затрудняет,, по мнению i
других, дифференцировку сукцессий и циклических изменений.
Качественным отличием одного сообщества от другого (сменяю-!
щего первое) будет прежде всего доминирование в них различных
экологических типов. Так, Адамс [Adams, 19081 прямо говорит -- •
о сукцессии, как об изменении доминирования некоторых видов
или ассоциаций. Обычно вначале имеет место слабое увеличение ко-
199
личества особей одного типа и соответственно уменьшение другого
(других). Их соотношение может меняться, пока первый не станет
>, доминирующим. Процесс смены, как правило, не ограничивается
одним изменением доминанта, в него вовлекаются обычно все или
большинство членов сообщества, и соотношение их плотностей изме- •
пяется. Что касается скорости смен сообществ, скорости сукцессии,
то наиболее быстрой она будет во временном водоеме, разлагающем-,
ся трупе животного, настое сена. Скорость их измеряется временем
достижения устойчивого состояния или полного исчезновения суб- '
страта [Grahams, 1925L ; ’ ' ’
Этот же процесс в том или ином масштабе происходит всюду,;
•он (процесс сукцессии) универсален для всех времен, т. е. следует \ (
признать, что в любом сообществе сукцессия идет постоянно [Кашка-
ров, 1933]. Более всего характер сукцессии, зависит от климатиче^-
ских прйчин (спорадическая засуха, обводненность). Они уничто-
жают первоначальное население биоценоза, после чего ряд перестро-
ек создает новую структуру сообщества с новым доминантом или
иным соотношением обитающих здесь видов (потеря видов, приобре-
тение).
Словом «сукцессия» обозначают само явление смены, конкретные:
же ее проявления в каждый отдельный момент времени называют
рядом, или серией. Под термином «ряд» подразумевается развитие
формации от начала ее появления до стабилизации. Упор на струк-.
туру сообществ позволяет различать смену аспектов в нем и сукцес- -
сию. Понятие «доминирование животного» включает зависимость от i
него других животных и способность процветать, несмотря на по-
стоянные колебания его количества [Вистол, 1913, цит. по Кашка-
ров, 1933]. Чаще всего разбирают первичные сукцессии, где, напри-
мер, по мере того, как обнаженная скала освобождается ото льда,
происходит постепенное заселение ее животными, причем одни виды . .
сменяются другими вполне закономерно. Наибольшее внимание
экологов привлекают вопросы зависимости между изменениями фа-
уны и сукцессией растительности, а также стабилизация и крите-
рии климаксового Сообщества [Heirston, 1959; Lindeman, 1942]. ;
Не вызывает сомнения, что для глубокого понимания механизмов -
сукцессий необходимо предварительное детальное изучение струк-
туры сообществ [Макфедьен, 1965].
Существует мнение, что если факторы, действующие извне, пре-
кращаются или их воздействие исчезает, то данное сообщество может
вернуться в прежнее состояние. В этом случае говорят об обратимых
изменениях и относят их не к сукцессиям, а к циклам. Колебания,
таким образом, отделяют от сукцессий, считая последние направлен-
ными процессами в отличие от ритмических (циклических). Между
тем, признается и взаимосвязь сукцессий и циклических процессов.
Так, Р. Маргалеф [Margalef, 1968] замечает, что колебания всегда
присутствуют в сообществах, но различаются по значимости. Они
сильны в молодых экосистемах и менее выражены в зрелых. Еще
более очевидной становится связь сукцессий и циклических процес- ,
сов в результате исследований Н. М. Черновой [1977]. Автор рас- .
200
смотрела сукцессии населения микроартропод в растительном опа-
де. Появление опада циклично, и многолетнюю сукцессию при
его разложении можно представить в виде колебательного процесса.
Разнообразие же серий сукцессий при разложении органических
остатков создается за счет разного экологического состава пионер-
них группировок и неравномерности прохождения отдельных эта-
пов. Одна из основных закономерностей сукцессий — волнообраз-
ность процесса. Колебательный характер имеют и изменения соот-
ношения видов, сезонная и сукцессионная динамика видов’тесно
переплетены. Волнообразность развития комплекса почвенных бес-
позвоночных показана и при первичном почвообразовании И. В. Сте-
баевым [1963, 1968]. Спиральный ход преобразований на всех ста-
диях сукцессии обусловлен как ритмом смены генераций, так и от-
ражением сезонной динамики видов. По-видимому, такой ход посту-
пательных движений — одна из наиболее общих черт динамики со-
обществ любого масштаба.
Другим примером взаимосвязи сукцессий и циклов является
концепция циклического климакса. Примером его могут служить
изменения Чапарраля в Калифорнии. Климаксный фитоценоз здесь
кустарниковый, он подвержен естественным пожарам, после каждо-
го из них быстро восстанавливается доминирующий кустарник
[Одум, 1975]. Огонь уничтожает не только зрелую растительность,
но и продуцируемые ею ингибиторы, т. е. снимает аллелопатическое
воздействие. Таким образом, наблюдается циклический климакс,
контролируемый взаимодействием пожара и аллелопатии. Пиро-
генные сукцессии — распространенное явление в лесной зоне [Fox,
1982; Gockbnrn е. а., 1981]-, и все они — процессы циклические.
Климаксный зрелый лес это одна из серий, следующий этап сук-
цессии — пожар и развитие очередного пионерного фитоценоза.
Макдональдом в 1965 г. (цит. по [Одум, 1975, с. 344]). описана
цикличность сукцессии в еловом лесу, в котором происходят пе-
риодические вспышки массовых размножений вредителя и зрелые
деревья погибают. Ель и ее вредитель образуют естественную са-
моподдерживающуюся систему.
; КОНЦЕПЦИЯ ЦИКЛИЧЕСКИХ СУКЦЕССИЙ
Концепция экологических сукцессий как циклического биологи-
ческого процесса выдвигается и обосновывается для тех биогеоцено-
зов, которые существуют в условиях многолетней цикличности при-
родной среды. Регулярная изменчивость природного фона ставит
местные биогеоценозы в несходные условия при разных фазах при-
родного цикла, ведет к видовым перестройкам сообществ и опреде-
ляет их ритм. Это повторяющаяся (цикл за циклом) изменчивость
среды обитания биоценозов — естественная форма их суп^ествова-
ния и развития. Виды, их популяции и сообщества эволюционно фор-
мировались в этой ритмике природной среды и адаптировались к ней
адекватными перестройками видовых структур и образа жизни.
201
Понятие «природный цикл», которое берется в основу анализа
динамики биогеоценозов,— это понятие не климатологическое, а
'экологическое; оно более широкое, так как включает в себя как
внешний, чаще всего гидроклиматический фактор, изменяющий сре-
ду обитания организмов, так и последствия его влияния на биогео-
‘ ценозы [Максимов,, 19846]. Последствия влияния природного цикла
на биоценозы заключаются в iмногократной повторяемости перестро-
ек их структуры. Сообщества, .формируясь, сменяют друг [друга/
подчиняясь фазам природного цикла, и сама их смена (сукцессия) .
оказывается । циклической. i 1 1
Циклическая сукцессия , является биологическим следствие^
природно-климатического, внешнего по отношению к биоценозу
фактора. Система взаимосвязанных, синхронизированных колеба-
ний бибценоза и его природно-климатического окружения опреде- .
1 (ляется нами как «природный цикл». । '
I ! Примеры! природных циклов. .. Биогеоценозы характеризуются
; Своеобразными природными циклами или их сочетанием. Рассмот-
! ренные выше материалы по Барабинской низменности Западной'
Сибири, где четко1 прослеживается в среднем' 11-летняя цикличность
в животных сообществах, связаны со сменами фаз увлажненности
территорий IB той же ритмичности. Смена влажных и сухих фаз в '
11-летнем цикле отражается на всем лесостепном биоценозе, приво-
дит к сложным видовым его перестройкам и влияет одновременно на '
сельское хозяйство й другие отрасли народного хозяйства [Природ-
! ные циклы...^ 1982]. i'
Лесостепным биогеоценозам Западной Сибири свойственны при-
родные цикличности. Циклические сукцессии характерны также и
для других зональных и интразональных биогеоценозов. В том, что
это явление имеет широкое распространение, убеждает наличие раз-
ных типов циклической изменчивости- природной среды. Например,;
в Западной Сибири, помимо названного выше лесостепного, выде- V.
ляются следующие природные циклы.
О з е р н Ы|й внутривековой природныйцикл.
! 'Уровень воды в озерах Западной Сибири подвержен внутривековьгй
колебаниям, закономерности которых детально изучены А. В. Шнит- i
никовым [1950, 19571, 1969]. В результате анализа различных фи-
зико-географических явлений, обобщения обширных литературных
I данных за период с конца XVII в. до середины текущего столетия
выявлено семь полных циклов во внутривековых колебаниях уровен-
ного режима озер продолжительностью от 29 до 45 лет. Отмечено]
при этом, что подъем уровня озер происходит относительно быстро^,
начинаясь обычно резко и в, короткий период (2—3 года), после
чего темп подъема в зависимости от размера бассейна на 2—6 лет
замедляется. Далее следуют краткий период неустойчивого макси-
мума (1—3 года) и продолжительный, медленный спад (12—20 лет;
и более) до периода самого низкого неустойчивого уровня, продол-
жающегося примерно 6—8 лет. 1
Амплитуда колебаний уровней по высоте составляет‘для круп-Г
ных озер с речным питанием (типа оз. Чаны) около ±2—2,5 м, а для ,
202
соленых, рапных, озер — около ±1,2—1,5 м. Установлено нали-
чно зависимости состояния уровня озер от условий распределения
атмосферных осадков и температуры воздуха во времени; их сочета-
ние на рассмотренной территории в условиях периодической ув- '
лажненностп формирует ритмы, продолжительность которых за про-
шедшие 25 лет не выходила за пределы 20—47 лет.
Сопоставление между । собой циклов различной интенсивности
осадков с колебаниями уровня озер показало наличие достаточно
хорошо выраженной зависимости уровня от осадков, причем уро-
вень озер запаздывает по отношению к изменениям количества ат-
мосферных осадков на 2—4 года [Шнитников, 19571. t '
Регулярный процесс колебаний уровня озер в Западной Сибири
в интервалах от 29 до 45 лет приводит к такой- же регулярной из-
менчивости условий существования сообществ озерных , котловин,
формируя своеобразный для этих биоценозов озерный внутривеко-
вой природный цикл.
Для экологов представляет особый интерес смена растительных
и животных сообществ с начала развития регрессивной фазы уро-
венного режима озер. Постепенное освобождение из-под воды об-
ширных прибрежных тростниковых займищ, оседание на дно спла-
вин приводят к разительным преобразованиям местных биогеоцено-
зов, вызывают характерные видовые перестройки растительных и
животных сообществ, определяют динамику численности ряда видов
животных, вызывая у некоторых из них вспышки массовых размно-
жений, оказывают большое влияние на сельское, охотничье-промыс-
ловое, рыбное и другие отрасли озерного хозяйства степной и лесо-
степной зон [Формозов, 1934; Иоганзен, 1939; 1951, 1956; Воронов,
1947; Шнитников, 1950; Максимов, 1959а, 1977; Вагина, 1962; Мак-
симов и др., 1974; Фолитарек, 1976].
Исследование циклических сукцессий биогеоценозов озерных
котловин в связи с внутрпвековыми колебаниями уровенного режи-
ма озер еще не привлекало к ^себе специального внимания, хотя в
литературе по этому вопросу;,уже накоплен большой материал, тре-
бующий анализа и обобщения. Приходится учитывать длительный
интервал смен фаз озерного природного цикла, вызывающий необ-
ходимость преемственности прп многолетних стационарных иссле-
дованиях. Трудности исследования озерных сукцессий возникают
из-за большого разнообразия типов озерных котловин и несходных!
путей их зарастания [Смиренский, 1950]. Биоценозы разных озёр
реагируют несходно на колебания уровня воды. Отсюда разнообраз-'
ные варианты циклических сукцессий в пределах озерного природ-
ного цикла. ; ।
Это пример, когда один и тот же природный цикл'создает боль-
шое разнообразие циклической изменчивости в условиях существо-
вания биогеоценозов и смен их сообществ.
Пойменно-речной природный цикл. Регулярно
повторяющиеся разливы в поймах рек создают характерную цик-
лическую изменчивость условий существования пойменных биоце-
нозов. Высокий разлив преобразует биотопы и биоценозы, многие
203
виды бывают подавлены или даже вовсе исчезают в этот год в пойме.
В последующие после разлива годы ценозы восстанавливаются, но
очередной высокий и продолжительный разлив вновь приводит их
к исходному состоянию. Годы между высокими разливами характе-
ризуются промежуточными видовыми структурами населения.
Имеется большая литература, посвященная влиянию разливов
на растительные и животные сообщества [Максимов, 1974; Герман
и др., 1981 ]. х
Сукцессии животных сообществ в связи с разливами изучались
в Западной Сибири в пойме р. Оби [Сукцессии..., 1981]. Речные пой-
мы имеют особенно динамичные биоценозы. После каждого высо-
кого и продолжительного разлива происходит почти полное обнов-
ление в пойме сообщества мелких млекопитающих. Наблюдаются
типичные циклические сукцессии. Основные сообщества, разруше-
нием которых начинается каждая сукцессия, отличаются друг от
друга соотношением плотности ^популяций мелких млекопитающих,,
количеством составля!ощих видов и местом в ряду сукцессии. ,
Лесной природный цикл. Смена лесных сообществ
при естественном возобновлении леса после лесных пожаров и р)т-
бок стала классическим примером экологических Сукцессий, и это-
му явлению посвящено много работ [Сукачев, 1972; Наумов, 1963а,.
б; Одум, 1975; ,Работнов, 1978; Риклефс, 1979; Grange, 1949].
Здесь важно подчеркнуть, что естественное возобновление хвой- '
ного леса на гарях, вырубках — это типичный пример циклической
сукцессии, не всегда находивший правильную трактовку в литера-
туре. К этой категории смены лесных сообществ В. Н. Сукачев не
применял термин «сукцессии». Он пишет, что «для циклических из-
менений лесных биогеоценозов характерно периодическое возвра-
щение их в состояние,- практически однородное с исходным, хотя,,
понятно, абсолютного тождества между ними нет... Такие изменения
нельзя называть сукцессиями...» [Сукачев, 1972, с. 383].
। Между тем, после естественного .восстановления исходного хвой-
i ного леса (цикл равен примерно 80—90 годам), его последующего
старения,; развития стволовых вредителей, образования сухостоя,,
Очередной и неизбежный лесной пожар вновь приведет к повторным
рядам той же сукцессии с промежуточными растительными форма-1
( циями травянистых лугов, кустарников и лиственных лесов. Это.
естественный циклический процесс смен сообществ в вековом цикле.
Сукцессиям лесных сообществ посвящено большое число работ.
Они требуют анализа и обобщения с позиций представлений о цик-
личности этого биологического процесса. ,
* *'
Мы приходим к выводу, что циклический характер сукцессий —
это типичное и широко распространенное явление в жизни биогео-
‘ ценозов. Природные циклы — свойство зональных (ландшафтных)
экосистем, они отражают колебания природной среды, характерные. ;
204
для данного зонального или интразоиального (например, пойма)
типа ландшафта. Им подчиняются сообщества биотопов, типичных
для ландшафтных зон. Сукцессии в биогеоценозах возникают не в
результате влияния внешних причин (сукцессии, связанные с лес-
ными пожарами, здесь не являются исключением, так как описыва-
емый процесс типичен для лесного биоценоза в целом), а в ре-
зультате естественных процессов, свойственных самим биогеоценозам.
Циклические сукцессии зональных сообществ имеют разные рит-
мы в соответствии с местными ритмами природной среды, которые
различаются также по вызывающим их причинам, территориально-
му охвату (зона, тип ландшафта, биотоп) и экологическим послед-
ствиям (изменчивость биологической продуктивности, хозяйствен-
ное и медицинское значение популяций и сообществ при смене до-
минантов, развитие различных других биологических процессов).
Местные биоценозы не одновременно включаются в ритм насту-
пающей фазы природного цикла, а некоторые сообщества в преде-
лах природного региона могут в данный отрезок времени и вовсе
не включиться в его ритм. Это связано с географическими особен-
ностями местности и, в частности, с неодновременным и несходным
в разные циклы развитием на территории гидроклиматического фак-
тора, вызывающего данный природный цикл (см. гл. V). Поэтому
развитие того или иного биологического процесса, характерного
для данного биогеоценоза, может занимать более узкий интервал
времени, чем длительность соответствующей фазы природного цик-
ла [Максимов, 1984а].
Сложность исследования циклических сукцессий состоит в том,
что, помимо влияния смен фаз природного цикла на весь биоценоз,
популяции каждого вида имеют, кроме того, свою циклику, связан-
ную с размножением животных и подчиненную экологическим фак-
торам, включая внутрибиоценотические связи. В результате цикли-
ческой изменчивости окружающей среды популяции каждого вида
занимают свою, наиболее благоприятную экологическую нишу.
Циклические сукцессии в разных биогеоценозах существенно раз-
личаются и по ряду других признаков. Различия связаны прежде
всего с несходными внешними факторами, вызывающими ритмы.
Это обнаруживается даже при большой общности животного населе-
ния, например в Барабе и в пойме р. Оби.
В биоценозах мелких млекопитающих Барабы очередное мас-
совое размножение водяной крысы нарушает структуру сообщества,
‘обедняет его видовой состав. После этого из пионерной группировки
развивается новое сообщество, видовое разнообразие и стабильность
которого в течение некоторого времени и в результате прохождения
нескольких серий сукцессии увеличиваются. Образовавшееся сооб-
щество разрушается новой вспышкой численности водяной крысы,
которая подготавливается как внешними причинами (многолетняя
цикличность обводнения территории), так и функционированием
самого сообщества. Здесь уместно вспомнить замечание Одума [1968]
о том, что сукцессия есть результат жизнедеятельности самого
ценоза.
205
Пойменные биоценозы, в частности сообщества мелких млекопита-
ющих, развиваются по той же схеме, но цикличность изменений здесь
зависит не от смен фаз увлажненности, а от разливов реки [Макси-
мов и др1981]. Последние играют роль «пускового ^механизма»
очередной стадии сукцессии, а, количество серий зависит от числа
лет между высокими паводками, После очередного такого паводка
начинается заселение освободившейся от воды территории пионер-
ной группировкой мелких млекопитающих-мигрантов с коренного»;
•берега. Наблюдается несколько серий сукцессий, с каждой из ко*
торых нарастает устойчивость и увеличивается видовое разнообра-
зие в сообществе. । , j , ' «'
. Если в Барабе сообщества возвращаются к исходной структуре;
' примерно через 8—10 лет, то в пойме Оби с 1966 по 1977 г. в сооб-
ществе грызунов наблюдали четыре серии сукцессии с 1966 по 1978 г;
с 3 рядами, с 1969 по 1972 г.— с 4, с 1973 по 1974 ту— с 2 и с 1975—
по 1977 г.— с 3 рядами. Для землероек характерны три серии с 1966 '
I по 1968 г. с 3 рядами, с! 1969 по 1972 г,— р 4 и с 1973 по 1977 г.—,
с 5 рядами [Максимов и др., 1981]. . । ,
Сукцессий в Барабе и пойме р. Оби различаются по степени За-
вершенности. В Барабе чаще, чем в пойме, сукцессии носят завер-
, шенйый характер, потоку что смена фаз увлажненности, не имеет
'таких резких1 перепадов, как, разлив, и влияние ее на биоценозьг
проявляется не столь катастрофически. Последнее накладывает
отпечаток на глубину депрессии численности в сообществах мелких
млекопитающих, которая в пойме носит особенно выраженный ха-
. рактер. i
.Эти различия касаются также зависимости численности отдель-
I ных видов oti гидроклиматических условий: в Барабе численность -
некоторых видов не столь резко зависит от смен фаз увлажненности,,
как в пойме, где большие разливы имеют сильное воздействие на
сообщество. Все это ! сказывается на различии сукцессионных про-
цессов, биологической продуктивности и характере ее‘изменчивости
н разных сообществах.
Неодинаковы при разных природных циклах исходные точки
(реперы)| для расчета стадий сукцессий. Они зависят от внешнего;,
1 ^фактора, , вызывающего смену условий^,существования сообществ,,
1 и от глубины его влияния на последние. При циклических сукцес-
сиях выбор ^акого репера для отсчета биологического процесса
весьма условен, тйк как сам процесс цикличен. Такой выбор может
зависеть, в частности, от цели работы. .
Для пошиы среднего течения р. Оби в качестве репера мы избра- _ -
ли год очень высокого разлива, с которого начинается возобновле-
ние ценоза наземных позвоночных в пойме после их почти поголов-
ной гибели [Максимов и др.^ 1981]. В лесостепном ландшафте Бара-
бы таким естественным репером может служить год наивысшего-
уровня грунтовых вод на конкретном участке. Прп проведении зоо- .
логических исследований удобным и четким экологическим репе-
ром в Северной Барабе является год пика массового размножения
водяной крысы. 1
206
При внутривековых колебаниях уровня воды в озерах отсчет био-
логических процессов можно вести с года максимального' уровня
воды, наблюдая сообщества по мере обнажения прибрежной зоны
озер, оседания сплавин и др.
' 1 ! <
I ;. - • i i 11 ДИСКУССИЯ
•D - i
Выдвигаемая концепция экологических сукцессии как циклическо-
го биологического- процесса ставит в разряд .дискуссионных неко-
торые положения учения о сукцессиях, излагаемого в- литературе.
Современйое, учение о сукцессиях акцентирует внимание на вос-
становлении элементарных сообществ после их разрушения. Разли-
чают первичные сукцессии, когда живыми организмами осваиваются
незаселенные места, и вторичные, если зти процессы происходят
в стациях, ранее уже заселенных, но лишившихся обитателей. Ана-
лизируются пути восстановления исходного сообщества и достиже-
ния им состояния наибольшего соответствия с окружающей физиче-
ской средой (климакс). Анализируемые. причины разрушения сооб-
ществ носят чаще всего локальный, сторонний по отношению ко все-
му биоценозу характер-и относятся к отдельным участкам арены
жизни. Весьма типичны примеры сукцессий, приводимые в эколо-
гических сводках. Так, у Ю. Одума [1975J примеры сукцессий сле-
дующие: сукцессии растений и некоторых беспозвоночных на песча-
ных дюнах озера, а также на небольших прудах; после отступления
воды; изменение видового состава птиц на заброшенных пахотных
землях; сукцессии растений на заброшенных проселочных дорогах;
на искусственных прудах и подпружных озерах;'.при обрастании
днищ судов и причалов усоногими ракообразными и другими сидя-
чими морскими организмами; в сенном отваре, гниющих стволах
деревьев, трупах животных,- фекалиях; при вторичных стадиях об-
работки сточных вод, чрезмерном выпасе, а также на ранних этапах
колонизации островов, на вышедших на поверхность водьь скалах,
лавовых потоках и.т. д. (см. также [Макфедьен, 1965; Дажо, 1975;
Чернова, 1977; и др.]). , ।
Во всех приведенных случаях речь идет не об естественных фак-
торах сукцессионного развития, типичного для сообществ или био-
геоценоза в целом, а о различного рода локальных, причинах, разру-
шающих то или иное элементарное сообщество. Таких причин, раз-
рушающих конкретное сообщество, может быть невероятное множе-
ство, так же как и самих элементарных сообществ. ; ;
Процессы, обсуждаемые в качестве примеров сукцессий, пред-
ставляют, таким образом, частные случаи развития биоценозов, по-
скольку различные причины, вызывающие гибель или разрушение
исходного сообщества, характерные, но не единственные и не глав-
ные в развитии биогеоценозов.
При таком биоценологическом анализе остается нерешенным во-
прос о механизмах развития сообществ при естественных процессах,
свойственных данной природной среде, когда кроме различных раз-
207
рушающих причин, захватывающих элементарные комплексы, дей-
ствуют и созидательные процессы, отражающие развитие зональных
сообществ. В. Д. Александрова [1964] по поводу подобных катастро-
фических смен справедливо пишет, что «фактор, вызывающий смену,
чужд растительному сообществу, его действие никак не связано с
предшествующим развитием сообщества, как правило, он является
ио отношению к данному сообществу внешним и случайным» (с. 303).'
Легко видеть, что данное направление учения о сукцессиях изу-
-«. чает, по сути дела, аномальную ситуацию, отвечая на вопрос, что
произойдет, если данный биоценоз или его Часть будут уничтожёны.
При подобного рода биоценологических исследованиях внимание
обращено не на процессы развития, присущие самим биогеоценозам,
и не на поиски механизмов, управляющих их динамикой, а на про-
цессы, имеющие для общего развития биогеоценозов не столь суще-
! ственное значение. Естественнее сосредоточить внимание на про-
цессах, присущих системе, а не случайно ее возмущающих., Таковые
существуют. Так,. Ю. Одум [1968] предполагает, что Сукцессия
есть результат’ жизнедеятельнЬсти самого ценоза.
Сказанным мы, однако, не хотим снизить самостоятельное тео- ’
ретическое и практическое > значение исследований смен сообществ. .
Оно, как пишет В. Д. Александрова [1964], «выявляет закономер-
ности, по которым сменяются растительные сообщества при воздей-
ствии на них тех или иных разрушающих факторов, и поэтому дает
возможность предсказать, что произойдет с растительностью, на-
пример, в результате затопления в связи со строительством плотины,
в результате пожара той или иной интенсивности, в результате раз-
вевания песков вследствие выбивания их скотом или неумело про-
изведенной вспашки» (с. 304). Что же касается механизмов сукцес- ?
сий биоценозов, не разрушаемых по каким-либо причинам, а обус-
ловленных более общими зональными причинами и характеризую-
щими процессы «саморазвития» сообществ, то этот аспект остается
еще малоисследованным. -.......
На вопрос, происходят ли'сукцессии в устоявшихся сообществах,
сторонники классического учения о сукцессиях отвечают утверди-
тельно, но объясняют эти смены причинами, разрушающими есте-
ственную среду обитания сообществ, т. е. не обусловленными гене-
зисом данного природного комплекса.
То же относится и к возможности циклического развития сооб-
ществ, которое не отрицается, но может иметь место, согласно имею-
щимся взглядам, в результате полного разрушения сообществ и его
; последующего восстановления [Одум, 1975, с. 344; Риклефс, 1979,
с. 372—373]. Характерен перечень причин демутаций (возврат к со-
стоянию, близкому к исходному), приводимый Т. А. Работновым
[1973] для фитоценозов: 1) значительные отклонения от средних
метеорологических и (или) гидрологических условий; 2) пожары;
3) массовое размножение растительноядных животных (грызунов,
насекомых); 4) ураганы, лавины и др.
По В. Д. Александровой [1964], последовательные частные сме-
ны (сукцессии) происходят при заселении растительностью первич-
208
но (речная отмель и др.) или вторично (заброшенная пашня и др.)
свободной территории; при изменении растительностью условий
своего существования (отложение торфа, снегозадержание, приво-
дящее к выщелачиванию почвы), а также при постепенном измене-
нии условий местообитания благодаря внешним по отношению к
растительности причинам (постепенный-выход луга из пойменного
режима, систематически проводимое сенокошение или выпас), т[ е.
речь идет о нарушенных , в той или иной степени биоценозах.
Особняком в этой группе сукцессий стоят гологенетические сме-
ны, которые происходят под влиянием внешних для фитоценоза при-
чин, когда последние, по В. Д. Александровой,, действуют не грубо-
разрушающим образом, -а постепенно, иногда весьма медленно. ;Но
под гологенезом В. Н. Сукачев [1954] имел в виду!не рецептные (сов-
ременные) смены, которые являются объектом нашего рассмотрения,
а процесс изменения растительного покрова вследствие изменения
более общего крупного единства, например всей географической сре-
ды или отдельных ее частей: атмосферы, литосферы п т, п. (напри-
мер, климатогенпые сукцессии, вызванные постепенным изменением
климата). ,
Естественная текущая । изменчивость биогеоценозов в стабили-
зированных, ненарушенных условиях еще не привлекала к себе,
по- крайней мере у зоологов, должного внимания. Такое односто-
роннее развитие учения о сукцессиях, уход от анализа общих при-
чин их развития, явное увлечение элементарными сообществами не
может свидетельствовать о полноте разработки данной проблемы.
Изучение животных сообществ в зональной । среде обитания убе-
ждает, что они имеют свои законы сукцессионного развития, недо-
статочно отраженные в существующем учении о сукцессиях.
Концепция циклических сукцессий ставит задачей вскрыть ме-
ханизмы динамики биогеоценозов и смены их сообществ на уровне
естественно идущих процессов развития. Речь идет о механизмах
сукцессий, эволюционно свойственных биогеоценозам, в целом и рас-
пространяющихся на зональные фаунистические комплексы и их
ландшафтные биотопические группировки. Сукцессии биогеоцено-
зов этого ранга связываются нами с цикличностью среды их суще-
ствования, свойственной дайному природному региону.
В развитие учения об экологических сукцессиях авторы настоя-
щей книги предлагают наряду с сукцессиями при возобновлении
нарушенных сообществ в качестве самостоятельной категории выде-
лять сукцесии в уже сложившихся, зональных (ландшафтных) со-
обществах, в результате характерных для биогеоценозов естествеп1
ных процессов, связанных с генезисом их становления и развития.
Понятие о климаксе.’Сукцессия, согласно общераспространенным
взглядам, достигает климакса, когда завершается формирование
наиболее устойчивого сообщества, находящегося в равновесии с ок-
ружающей физической средой. Последнее оценивается как стабили-
зированное, самоподдерживающоеся сообщество [Adams, 1908; Cle-
ments, 1916, 1936; Александрова, 1964; Одум, 1975; Риклефс, 1979;
Разумовский, 1981; и др.].
На. а. Максимов, Л. H. Ердако» 209
-А
Критика взглядов о климаксе как финальной стадии дипамиче- :
ского равновесия между биоценозами и физико-географическими ус- ,
ловиями дана в трудах, Н. П. Шумова [19636], В. Н. Сукачева
[1972] и др. «Ясна,— пишет Н. П.; Наумов,— неправильность взгля-
да на «клийакс», или завершающее сообщество, как неизбежный ко-
нец сукцессионного ряда. Многочисленные наблюдения показали,'
что; сукцессия никогда не завершается полным восстановлением ра-
' ; нее бывшего; на ртом месте,(сообщества, а кроме того, 'любое из звень-
ез сукцессий может; устойчиво, существовать,! т. е. превратиться; в
«климакс», если сохраняются благоприятна® для этой фазы усл о-
; вия:..»| (с. , 123). I । , ! ।
- .Теория'климакса ‘детально разработана в фитоценологии [Алек-
сандрова,111964; Разумовский, 1981],’.что же касается позвоночных
ЖИВОТНЫХ, ЭТОТ вопрос; ПОЧТИ НС ИЗуЧбН. . I . ! ]
Рассмотрим учение io ,'климаксовых 1!сообществах как заключи-
I тельной стадии в) ряду изменчивости с Позиций концепции о цикли-
ческих сукцессиях у позвоночных животных., '
Если понятие «климаксовое! сообщество», полностью соответству-
ющее; и эволюционно приспособленное к данным физико-географи-
ческим условиям, связать с существующими ныне биогеоценозами,
то практически это। тундровые, лесотундровые, лесные, лесостепные ,
и другие , зональные, ландшафтные, биотопические комплексы рас-
тительности и .животного населения) с которыми эколог имеет дело
в своей повседневной работе. •
Применительно к этим стабилизированным в ходе эволюции эко-
I системам и выдвигается концепция циклических сукцессий. При-
родный цикл распространяется в пределах зональной территории,
охватывает; характерные для нее биотопы с их сообществами. По-
скольку природный цикл включает разные, по крайней мере две1
основные фазы, закономерно сменяющие друг друга, сукцессионный
ряд каждой фа'зы должен рассматриваться как наиболее. соответ-_
i ствующип ей; , , . _ ( к ... -
। Пример этого — бйрабинсйий । биоценоз; d Характерной для него
; сменой оухой и влажной фаз,(Каждой фазе соответствует определен-
ная видовая структура животных сообществ. Поскольку рассмотрен-
ный нами 'комплекс, будучи типично лесостепным, соответствует
местному климату, можно признать его за. завершающее сообщество,
однако положение егр временное; преходящее. При очередной сме-
не фазы увлажненности оно вновь заменяется другой серией сукцес-
сйи, соответствующей уже новой фазе увлажненности.;
(Поэтому нет оснований какую-либо стадию или;рйд сукцессии
считать промежуточным пли| завершающим сообществом. Смена фаз
природного цикла — это, характерный для данного региона процесс,,
• жаждая фаза которого — естественное следствие природной циклич-
’ ности, к которой приспособился местный биоценоз. >
Развиваемая концепция не дает1 оснований считать отдельные ря^
ды сукцессий как сообщества, не соответствующие существующему
состоянию условий среды. В. ходе смены природных циклов форми-
руются сообщества, в1 наибольшей степени соответствующие возни-
210
кающим новым условиям. Собственно завершающим сообществом
лесостепного биогеоценоза является комплекс, типичный для дан-
ного зонального ландшафта и объединяющий видовые структуры
обеих фаз увлажненности. ।
Когда обсуждается концепция о сукцессиях как циклическом
процессе, то правильнее говорить не о том,-что «климаксовый био-
ценоз ... максимально защищен от возможных1 нарушений со стороны
(внешних факторов» [Дажо, 1975, с. 287 ], а о том, что он эволюционно
приспособлен к изменчивости внешних факторов, являющихся средой
обитания сообществ. ; ' ~ ' I
। Существует ряд признаков, подтверждающих, что. I зональные
биогеоценозы! адаптированы к определенным природным циклам.
Эти адаптации проявляются, прежде вседо, в наличии в зональных* ।
биогеоценозах видовых комплексов, экологически связанных с раз-
ными фазами местного цикла. Так, у позвоночных животных наряду
! с видами автохтонами, четко проявляющими свои экологические и
поведенческие адаптации к одной из фаз природного цикла, суще-
ствует комплекс видов, который характерен для другой ландшафтно-
географической зоны и получающий преимущественное развитие в
период другой фазы природного цикла. Ранее это было отмечено для .
тундр [Максимов, 1947, 1959< 1974]. Подобные явления характерны
и при смене растительных, в частности лесных, формаций в вековом .
цикле, когда промежуточные серии лиственных пород в хвойной тай-
ге являются характерной стадией в ряду сукцессий после пожаров и
рубок.
О сообществах животных во влажную и сухую фазы увлажнен-
- ности в лесостепной зоне Западной Сибири, отражающих эти явле-
ния, говорилось выше.
Здесь важно подчеркнуть, что в типичных зональных раститель-
ных и животных сообществах постоянно присутствуют виды другого
зонального биогеоценоза, и это явление можно трактовать как адап-
тивный механизм к циклическим колебаниям среды. .
У представителей О|боих комплексов существуют, ( помимо’того,
различного рода экологические и морфологические адаптации, напри-
мер способность ряда видов растений длительно переживать неблаго-
приятные условия в стадии зачатков (жабрей, тростник), наличие у
животных стаций переживания и т. д. у *
Прав В. Г. Мордкович [1976], утвержда!я, что сукцессионная ,
мобильность биоценоза, т. е. возможность быстрого замещения эко-,
элементов в сообществе, является одним из главных механизмов его/
устойчивости к колебаниям абиотических факторов с]реды. i jy
В своем современном статусе ни к какому гипотетическому
климаксовому состоянию зональные сообщества не «стремятся»,
потому что существующие в настоящее время они и есть климакс о-
вые. К этому «стремятся» лишь разрушенные сообщества, но они
должны оцениваться в биоценологии вне ранга явлений, определя-
ющих естественную динамику биогеоценозов, т. е. процессов, эво-
люционно им присущих.
Поэтому с излагаемых здесь позиций вряд ли правильно ставить,
вопрос (например, в отношении Барабинской лесостепи), «стремит-
14*
211
ся» ли к климаксовому состоянию местный биогеоценоз? Сообщества
лесостепной Барабы в ходе исторического развития уже достигли
стадии, соответствующей ныне существующим зональным физико-
географическим условиям. Эти сообщества и среда их обитания фор-
мировались совместно. В ряде случаев смена фазы очередного при-
родного цикла (например, начало развития сухой фазы в 11-летнем
цикле) могла быть ошибочно принята за общую тенденцию развития
'зонального ландшафта.
Сказанное вовсе не юзначает( что развитие барабинского биогео-
ценоза, как. и любой другой зональной экосистемы, прекратилось. :
Дальнейшие изменения зональных биогеоценозов происходят, и,'
есйи при этом исключить антропогенные’ причины их коренного п-
необратимого преобразования (этот аспект, общий для всех биогео-
ценозов,‘Здесь не рассматривается), то сукцессии-могут происходить
в результате «саморазвития» отдельных биотопов. Примером этого
может служить зарастание озерных котловин, все стадии которого г
(от открытых плёсов до рямов) хорошо прослеживаются в лесостепи
Западной Сибири. Но эти смены (возможно, также циклические)-
происходят в пределах того же зонального; типа ландшафта.
' Трансформация биогеоценозов возможна также при последующем
историческом развитии всего зонального природного комплекса,
связанного с коренными изменениями климата, геоморфологии и др.
Такие гологенетические смены, как пишет В. Д. Александрова
[1964], никогда не прекращаются, поскольку никогда не останавли-
вается естественный процесс развития ландшафта.
Смены сообществ в результате, во-первых, разрушения суще-
ствующих биотопов, во-вторых, циклов природной среды', в-третьих,
.внутривековой естественной трансформации биотопов, а также,
Ьз-четвертых, как следствие исторических преобразований климата и
рельефа — это разные категории и уровни экологических сукцессий,
требующих к себе равноценного внимания экологов.
Поэтому вопрос о длительности завершающего, климаксового,
Сообщества должен рассм<атриваться по-иному. Р[ Дажо [1975] пи-
шет, что «климаксовый биоценоз остается неизменным в течение вре-
мени, равного «нескольким человеческим жизням». Такая оценка
может быть уточнена, если иметь в виду, завершающие сообщества,
свойственные ландшафтно-географической зоне и ее ландшафтным:
подразделениям. Здесь определенные, стабилизированные струк-
туры сообществ существуют так долго, как долго в процессе истори-
ческого формирования .существуют зональные ландшафты с их био-
ценозами. ; 1 1
Совершенно точно заключение В. Н. Сукачева [1972], что кли-
макс может оцениваться как конечная стадия развития биогеоцено-
за, пока остается неизменным климат. Этот вывод можно относить
как к историческим, так и к современным условиям. Примеры, рас-
смотренные выше (смена фаз увлажненности Барабы в 11-летних
циклах, влияние разливов на пойменные биогеоценозы, внутривеко-
вые колебания уровня озер) —- это климатические смены, имеющие
циклы разной длительности.
212
Из рассмотренных выше примеров циклических сукцессий тре-
буют оговорки пирогенные сукцессии сообществ в лесном (напри-
мер, хвойно-таежном) биогеоценозе, причиной которых являются лес-
ные пожары и рубки. Климаксовая стадия их хвойный лес, по-
скольку он в наибольшей степени; соответствует климату лесной
зоны. Промежуточные ряды сукцессий (травянистый луг, кустарни-
ки, лиственный лес) — временные формации, не соответствующие
зональным условиям. Пожар в лесу, приводящий к циклической и
взаимосвязанной сукцессии всего лесного биогеоценоза, может оце-
ниваться как естественный фактор, присущий лесному сообществу
и обусловливающий его саморазвитие. Старение хвойного леса,
образование сухостоя, деятельность лесных вредителей неизбежно
вызывают гибель перестойного леса. Пожары как естественная при-
чина гибели и последующей сукцессии в этом процессе закономерны. -
। Длительность сукцессионных периодов. Излагаемые в книге
взгляды на сукцессии как циклический процесс отвечают на вопрос
о длительности их существования. Обычно они классифицируются на
вековые, длительные быстрые и катастрофические. Мы же под дли-
тельностью /сукцессий понимаем длительность соответствующего при-
родного цикла, т. е. периоды развития сукцессий могут измеряться
от геологических эпох до очень кратких, исчисляемых несколькими
годами.
«Стратегия» сукцессии как быстро протекающего процесса в
своей основе, как пишет Ю. Одум [1975], сходна со «стратегией»
длительного эволюционного развития биосферы. Биогеоценозы, или
экосистемы, в ходе своего саморазвития по мере климатических и
геологических перемен и преобразований природной среды претер-
певают изменения, сменяя друг друга.
Геологические эпохи, характеризующиеся сменами фаун, носят
характер очень длительных климатических циклов — от ледниковой
фазы через фазу умеренного климата к засушливой (ксеротермиче-
ской) фазе, после чего климат вновь возвращается к исходной ледни-
ковой фазе [Личков, 1960]. Для экологов особый, интерес представ-
ляют краткие природные циклы, результаты которых прослежива-
ются текущими наблюдениями. Смены фаз увлажненности лесостеп-
ной зоны Западной Сибири в 11-летних циклах являются примером
такой цикличности.
В пределах ландшафтно-географической зоны природные циклы
могут сочетаться и накладываться друг на друга. Так, для междуре-
чий Барабы характерно сочетание 11-летнего (болотного) и 35:лет-
него (озерного) природных циклов,-что накладывает отпечаток и на
динамику биоценозов [Максимов, 1977]. >
Оценка значения экзогенных факторов в динамике биогеоценозов.
В излагаемых в литературе взглядах движущим фактором сукцес-
сий признаются внутрибиоценотические отношения. Так, по Ю. Оду-
му [1975], «сукцессии происходят в результате изменения физиче-
ской среды под действием сообщества, то есть сукцессия контроля- 1
руется сообществом» и «замещение видов в сукцессиях вызывается
темг что популяции, .стремясь модифицировать окружающую среду,
создают условия, благоприятные для других популяций» (с. 324).
По Р. Дажо [19751, сукцессии являются результатом изменений,
вносимых в .среду обитания самими сообществами (см. также [Рик-
лефс, Д979г, Разумовский, 1981; Разумовский и др., 1983].
Остановимся на трактовке (обсуждаемого ^вопроса Н. М. Черно-
вой И 974 J.; Ссылаясь; на Ю. Одума [Odum, 1989], М. М. КаМшилова
. [1970 ], i Ё, Н. Мишустина! [19561,; она пишет: «Накопленные к на-
стоящему ' времени сведения убеждают, что природа сукцессий —
взаимосвязи внутри сообщества. Физическое окружение определяет
характер сукцессии или вызывает ее^ начало) не является ее причи-
ной и механизмом». И далее: «сукцессии в .современном понимании —
это не любые последовательные смены механизмов, а закономерный
процесс развития сообщества, связанный с неизбежным изменением
им окружающей физической среды. Чтобы могла осуществиться сук-
цессия, сам ценоз* должен играть определенную , роль; в управлении 1
средой» (с. 11). Эта точка зрейия- находит отражение также у А. Г. Во-
Аронова [1963*]J «В постепенном формировании фитоценоза и затем в J
смене сложного фитоценоза* другим ।играют основную роль внутрей-
ние осрбеннбсти сообщества» (с. 277). !
I Между тем развитие , биогеоценотйческого процесса не должно
определяться только теми факторами, о которых пишут указанные
авторы и которые так или иначе относятся к категории внутренних,
связанных с, самими биогеоценозами, их жизнедеятельностью или
1 влиянием растительности и животного мирада также микроорганиз-
мов на другие компоненты биогеоценоза.i
Приходится также учитывать, что средообразующая роль поз-
воночных животных в условиях, например лесостепной зоны Запад-
ной Сибири, чрезвычайно! м!ала и, проявляется лишь по влиянию на
почвенную фауну (роющая деятельность водяной крысы в годы вспы-
шек массового размножения). , . .
При циклических сукцессиях* не биоценозы определяют смену >
среды обитания (хотя и этот аспект не отрицается, например при лес-
ных сукцессиях, вырванных пожарами,'и др,), а именно изменчив'ость
физической среды является фактором,, обусловливающим смену це-
нозов в разные фазы природного цикла. Значение среды обитания не
может ограничиваться только ролью внешнего толчка или повода,
к, возникновению перестройки сообществ. । **. .
Не совсем правильно оценивать внешние (экзодинампческие)1
силы как случайные по отношению к ходу развития'ценозов [Воро-
нов, 1963 J. Физико-географические условия существования био-
геоценозов изменчивы, и рассматриваемая здесь цикличность этой ,
изменчивости выступает важнейшим фактором, влияющим на усло-
вия существования и развития их. i
Поэтому не бесспорно утверждение С. М. Разумовского [1981],
что в случае экзогенной смены внешний фактор не вызывает прямого
уничтожения сообществ, а лишь создает условия для его конкретного
вытеснения другим сообществом. Спорность этого положения осо- „
бенно четко проявляется на примере сообществ позвоночных живот-
214
ных в речных поймах, где каждый цовый ряд циклических сукцес-
сий начинается на остатках уничтоженного сообщества. В лесостеп-
ной зоне смена фаз увлажненности! также резко изменяет биотопы
предыдущего сообщества, делает невозможным, его существование.,'
. Факторы внешней среды, I определяя условия существования биоце-
нозов и их регулярную изменчивость во времени, должны оценивать-
ся в. качестве вызывающих направленные перестройки в биоценозах ।
И ИХ ритмы. И • * (i ' I ! ' I.
В изменившихся условиях при смене,, доминантов’большое’!зна-:
чение начинают ; приобретать внутрибибценотические; отношения.
Их роль возрастает и становится, ведущей в пределах- сменившейся, ,
фазы природного цикла. Экологически это легко1 объяснймо и на-1
ходит отражение в литературе [Воронов, 1983; и др.].; i " I (
Изменившаяся среда обитания создает обстановку,- благоприят-
ную лишь для определенной части сообщества. Получая благоприят- i
ные условия для существования, размножения й1 территориального'
распространения, эта часть биоценоза начинает играть ведущую роль
: в сообщество, заменяя в нем те популяции, для которых изменив-
шиеся условия стали непригодными. ih
Очевидно,- при такой смене сообществ для части видов, которым,
изменившиеся условия благоприятны, но они менее конкурентоспо-
собны, чем доминирующие, играют роль и внутрпбпоцепотпческпе|
отношения, в том числе влияние доминирующих видов на физиче-
скугощреду обитания. Последнее более полно исследовано на приме-
ре водяной крысы. Особенно большой трансформации в годы массо-
• вого размножения этого вида подвергаются^ лесные луга,’ зарастаю-
щие жабреем. Такие луга вновь восстанавливаются и становятся 1
пригодными для сенокосов лишь через два года после завершения > ;
вспышки размножения водяной крысы. , II. ’
Приводимые материалы по ,барабинскому биогеоценозу под-
тверждают эту точку зрения. Сложные перестройки животного насе-
ления, имеющие циклический характер, в северной лесостепи Запад- j
ной Сибири подчиняются многим факторам, из которых можно вы-
делить ДНа ведущих: смены фаз увлажненности территории в 11-
летних циклах и вспышки массового размножения водяной крысы!
Первые определяют общий облик животного населения по влажному
и сухому вариантам,! а вторые вносят существенные коррективы в
структуру населения и численность отдельных; видов и тем1’самым ;
существенно влияют на видовой состав не только вл'ажной, но и су-. «
хой фазы. . । ; ,
Поэтому сукцессии можно рассматривать как результат сово-
купного влияния на биогеоценозы. экзогенных (изменчивость при-
родного фона) и эндогенных (внутрибиоценотические отношения) . ,
факторов. Более уместно говорить, что цикличность природного фо-
I на — это не внешний фактор, а естественная среда существования ,
биоценоза. 1 । । 1 .. ' 1
О соотношении понятий «сукцессия» и «цикличность». В ли- .
тературе о сменах сообществ понятие «сукцессия» нередко противог
поставляется понятию «цикличность». Неоднократно высказывалось
215
i
мнение, что цикличность происходящих в сообществе изменений ука-
зывает на1 отсутствие сукцессии в нем. Так, В. Н. Сукачев [1972]
отличал собственно сукцессии от циклических изменений, когда
; биоценозы возвращаются в исходное состояние (хотя оговаривался,
что и последнем случае абсолютного тождества может и не быть)..
. Собственно же к сукцессиям он Относил процессы, при которых про-
. исходит смена типов биоценозов (чаще всего, необратимые смены).
Поскольку "понятие i «цикличность»! связано с изменчивостью
1 природной среды, к той же категории взглядов относится и утверж-
дение С. М. Разумовского [1981, 19831, что «смены, вызываемые жиз-
1 i1 недеятельностыо самого сообщества, следует называть сукцессиями,
i а смены под влиянием факторов, внешних по отношению к сообще-
ству, — экзогенными сменами» (с. 231).
: Такое противопоставление понятий «сукцессия» ^и «цикличность»
1йы считаем неправомочным, поскольку сукцессия — это биологпче-
: ский процесс, а цикличность — лишь временная его характеристика;
। сукцессии могут быть как циклическими, так и1 нециклическими.
На наш взгляд, целесообразно под термином «сукцессия» пони-
! мать общее явление многолетних последовательных смен раститель-
Д ных и животных сообществ. В ходе жизнедеятельности биогеоценоза
; такие смены могут быть связаны с разными причинами и иметь зако-
номерный необратимый и обратимый (циклический) характер. Боль-
шое разнообразие типов сукцессий, включая текущие циклические,
должно найти отражение, в классификации сукцессий. ,
Рассмотрим термин «флуктуация» (колебание). В советской гео-
ботанической литературе он ставится в один ранг с термином «сук- ;
г цессия». Флуктуации относят к динамике фитоценозов, а сукцессии —
[ к динамике растительного покрова [Александрова, 1934; Работнов,
;1972] По В. Д. Александровой [1964], «смены растительного покрова
имеют место, когда на каком-либо участке территории (или аквато- :
। рии) существовавшая здесь растительность сменяется другой, отли-
чающейся, от предыдущей существенными признаками: составом эди-
фикаторов, доминантных и детерминантных или характерных видов,
а также продуктивностью, типом йруговорота вещества и энергии, i :
; ритмикой сезонного. развития, характером погодичных флуктуаций <
и т. п.» (с. 301). Это определение понятия «сукцессия» как процесса
. может быть полностью отнесено и к, описанным выше, сменам живот- ,
ного населения. 1 - '
I; Под термином «флуктуация» Т. А. Работнов [1972] понимает;
i «ненаправленные, различно ориентированные, или циклические,
[изменения! их от года к году или в течение краткосрочных климати-
ческих или иных циклов,, завершающихся iвозвратом к исходному
। или, что более точно, близкому к ним состоянию» (с. 100).
Этот термин, отражающий разногодичную изменчивость фитоце-
нозов, вполне характеризует описанный выше биологический про-
цесс в сообществах животных, который мы именуем как циклические
сукцессии. *;
Такие противоречия в терминологии связаны с тем, что «флукту-
ации» и «сукцессии» — это термины разного порядка: флуктуации
216
отражают лишь изменчивый, колеблющийся характер развития сук-
цессий, Флуктуация — явление, характерное как для-растительного
покрова, так и для фитоценозов, а также для животного населения и
зооценозов. Эти колебания в ходе динамики находят выражение в
сменах видового состава, в положении отдельных видов в сообще-
ствах, в колебаниях биологической продуктивности и в других про-
явлениях жизнедеятельности растительного и животного 'мира.
Во многих случаях флуктуации в сменах растительного покрова и
животного населения носят явно выраженный циклический харак-
тер, например,; когда колебания природного фона характеризуются
определенной ритмикой.
, Должно существовать одно понимание термина «экологическая
сукцессия» как явления последовательных в течение ряда (многих)
лет смен растительных и животных сообществ. Эти смены могут быть
необратимыми или обратимыми (циклическими), но последнее отра-
жает. лишь характер развития сукцессий во времени. Такое разно-
образие процесса может найти отражение в классификации описыва-
емого явления.
Оценка явления цикличности; В общераспространенных пред-
ставлениях о сукцессиях часто игнорируется тот факт, что среде >
существования сообществ может быть свойственна циклическая из-
менчивость как закономерный процесс. Этой форме направленной
изменчивости биогеоценозов не уделяется должного внимания. Явле-
ние цикличности природного фона обычно не ускользает от внима-
ния исследователя, но оно не оценивается как естественный, регуляр-
ный процесс, который определяет закономерную повторяемость в сме-
не сообществ. ।
Так, среди циклических, обратимых изменений биоценозов
В. Н. Сукачев [1972] рассматривает таковые в течение суток, в раз-
личные сезоны года, в зависимости от погодных изменений, а также
циклические изменения, связанные с естественным процессом возоб-
новления и роста древесной и другой растительности. При класси-
фикации сукцессий В. Н. Сукачев учитывает и климатогенные сук-
цессии биогеоценозов, но включает их в группу гологенетических
(необратимых). ; ,
Таким образом, сукцессии сообществ в регулярно и циклически
изменяющейся среде обитания биоценозов в этой классификации не
находят себе места. Между тем, цикличность природной среды может
служить внешним синхронизатором колебаний сообществ, т. е. на-
водить характерный ритм. (|р
Б!ез учета цикличности природной среды нельзя вскрыть меха-
низмы непрерывных изменений в развитии биогеоценозов. Не может
быть правильным исходное допущение, что общие условия окружения
биогеоценозов не меняются, что природа должна рассматриваться как
косное явление и что приток солнечной энергии неизменен,[Сукачев,
19721. - ,
Биоценоз Существует в условиях циклически изменяющейся при-
родной среды, обусловленной в том числе и циклами солнечной ак-
тивности. Эти колебания условий существования сообществ являются
217
.«нормальным» состоянием и источником их развития. Циклический
характер изменений должен, по мнению Р. Риклефса [19791, найти
отражение в концепции климаксного сообщества. Климакс, по его
, , мнению, это «динамическое состояние, самовозобновляющееся по
1 ^составу,;даже если это достигается в результате регулярных циклов
, изменений» (с. 374). । > ।
. Сукцессии как обратимый процесс.. Требует уточнения тонка зре- i
ния'на сукцесбии как на «направленный процесс» '[Дажо, ;1975], иду- ,
щий с разной степенью, ускорения, но «в одном направлении» (с. 28).
! Последнее оценивается' как основной экологический принцип сук-
i'ii цессий [Риклефс, 1,979]. Сукцессиям даются следующие определения:
' как «процессы в ра'стительности, состоящие в необратимой перестрой-
ке фитоценозов и ведущие,к замене их другими» ! [Шенникюв, 1964,
с.'3151; как «многолетние,направленные смены» фитоценозов во вре-
j мини [Воронов. 1933, с? 269]; как «необратимые, направленное, т. е.
Происходящие в определенном , направлении, изменения раститель-
, кого покрова, проявляющиеся в смене одних фитоценозов другими»
i .[Работное, 1978, с. 252]; Необратимость изменений, по Т. А. Работ-
। , нову [1972];— это «очень важный признак сукцессии» (с. 96).
, Точка зрения о необратимости сукцессий отражается и в тех
формулировках, в- которых подчеркивается постепенный характер
замены одних сообществ или организмов другими [Реймерс, Ябло-
1Ков,. । 1982]; «последовательная смена одного биоценоза другим»
'Кащкаров, 1945], и что «любая сукцессия линейна: каждая ее ста-
дия, (при отсутствии внешних помех) всегда! сменяется одной и той яге
следующей стадией» [Разумовский и др., 1983].
Обратимость условий существования биогеоценозов рассматри-
। вается л'ишь для, того случая, когда влияние экзогенных факторов
'' прекращается, i что приводит к исходной структуре сообществ. По-
казательно,, что подобные обратимые ,смены относятся к разряду «ка-
тастрофических», т. е. внезапно действующих, фактически не пред-
сказуемых явлений.^ !. i . ...
; I Характерно в, этом! отношении следующее положение В. И. Су-
качева [1972]? «Когда данный лесной биогеоценоз саморазвивается
или изменяется под, влиянием развития других более общих или со-
i седнпх единств, то его сукцессии необратимы. Если же факторы,
действующие извне# 'изчезаюц или их воздействие прекращается, то
данный биогеоценоз может вернуться практически, в свое прежнее
,! состояние? В этом случае мощно говорить об обратимых изменениях.
Только первые изменения биогеоценозов^мы^будем называть сменами
(сукцессиями),, вторые же будем именовать циклическими (периоди-
ческими) изменениями» (с. 375). Периодический возврат фитоценозов
к состоянию, близкому к исходному, Т. А. Работнов [1973] относит
не кI.сукцессиям, а к особой форме изменчивости растительности —
демутацпям.1 ; । . I
Между тем,-' процесс Циклических сукцессий связан, с обратимыми
сменами сообществ, с возвратом к прежнему, хотя и не полностью
. тождественному состоянию биогеоценоза. Обратимость среды оби-
i тания биоценозов как результат цикличности должна рассматри-
। i
218 -1
ваться в качестве регулярного, постоянно существующего в при-
роде явления, приводящего к таким же регулярно повторяющимся
и обратимым сменам сообществ. 1
Явление обратимых преобразований биогеоценозов, связанных
с возвращением среды к исходному состоянию, косвенно подтверж-
дается и наличием в существующих классификациях сукцессий раз-
личного рода оговорок. Так, по. отношению к сменам лесных био-
геоценозов В. Н. Сукачев .[1972} шипел: «Чаще всего такие измене-
ния биогеоценозов оказываются необратимыми, но в некоторых
случаях, спустя 'то более, | то менее длинный промежуток времени^
в связи с устранением причины, вызывающей iсмену биогеоценозов
восстанавливается биогеоценоз исходного типа. Тогда динамический
процесс имеет обратимый характер» (с. 383). И дальше: «Однако надо,
иметь в виду, что в ряде случаев наблюдаемое изменение биогеоце-
нозов! нелегко отнести к одному из основных подразделений динами1-
кп, т.. е. к сукцессиям или не к сукцессиям. К такому случаю, напри-
мер, можно отнести изменение биогеоценоза, связанное с изменением
погоды, т. е. климатических условий в различные годы (погода, пли
погодная динамика)'. Если; изменений Климата затягиваются на не-
сколько лет, следующих друг за другом, то изменения в биогеоценозе
становятся। столь значительными и настолько относительно устойчи-
выми, что возникают основания говорить уже о появлении лесного
биогеоценоза другого типа... Вфтом случае можно и должно говорить
о происшедшей сукцессии лесных биогеоценозов. Однако дать впол-
не четкие, абсолютные критерии для установления того, появился ли
новый тип лесного биогеоценоза или нет, часто бывает трудно»
- [Там же, с. 3851. 1Ь, ! ; 1 .
Нечеткость признака «необратимости» _в определении понятия
«сукцессия» видна и из следующего примера. Т. А. Работнов [1972]
не считает возможным относить к сукцессиям циклическую измен-
чивость растительности в пределах 11-летних циклов солнечной
активности. Ссылаясь на материалы К. А. Куркина; по Барабе, где
цикличность климата вызывает значительные обратимые изменения
в растительности, Т. А. Работнов колеблется, относить .ди эти изме- .
нения к флуктуациям или к сукцессиям. Автор. приходит к выводу,
что «условно можно относить обратимые изменения, если они кратко-
срочны, к флуктуациям, а протекающие в течение длительного пери-
ода времени — к сукцессиям» [Там же, с. 97}, Такая неопределеп- ; j
ность является следствием нечеткости в классификации описываемо- '
го процесса, принятой в геоботанике, и большой^ условности исполь-
зуемых терминов, о чем писалось выше. ; '
Эти трудности с классификацией смен растительности и биоцепо-1
I зов легко преодолеваются, если их рассматривать как циклические
сукцессии, какими они по существу и являются. Необратимые и об-
ратимые смены сообществ отражают лишь разные этапы развития ;
одного и того же процесса. На каком-то из них, очень большом по, 1!
периоду, на фоне направленных последовательных смен в биогеоце- !
ноз'ах, связанных с климатогенными,, антропогенными^ и другими ’
.факторами изменчивости, смены с короткими периодами приобрета-
219
ют обратимый циклический характер, соответствующий флуктуаци-
ям среды. Обратимые смены существуют, повторяясь из цикла в
цикл.
Развиваемые в книге взгляды на понятие «сукцессия» как обра-
тимый циклический процесс близки к тому, о чем писал Н. П. На-
умов [196361, .относивший обратимые смены к сукцессионным про-
цессам. Встречающиеся в литературе свидетельства о повторяемости
сукцессий в природе также подтверждают налагаемые взгляды на
сукцессии как циклический, процесс. Так, А. Г. Воронов [1963] пи-
шет, что «в одном и.том же месте однотипная серия может повторять-
ся неоднократно, если повторяются нарушения растительного по-
крова1 или его уничтожение» (с. '269). Смену фаз увлажненности в
рассмотренном выше примере лесостепцого биогеоценоза можно
трактовать и как причину, нарушающую растительный покров.
В. Г. Мордкович [1966] в работе, посвященной сукцессиям живот-
ного населения почв, свидетельствует о неоднократной повторяе-
мости всегб ряда развития биогеоценозов, что «позволяет предпола-
гать, что процесс развития аккумулятивных ландшафтов в южной
Сибири шел скачками, с неоднократными перерывами и повторени-
ями» (с. 90).. .
Выше указывалось, что сообщества в ходе циклических преобра-
зований возвращаются к прежним соотношениям видов. В барабин-
ском сообществе мелких млекопитающих повторяемость населения
особенно четко проявляется в аспектах доминирующих видов (см.
гл. V). Однако полного тождества не бывает, иначе процесс не носил
бы характер поступательного развития и был бы замкнутым.
В рассмотренных нами примерах экологических сукцессий, свя-
занных с циклами среды обитания сообществ, каждый ряд сукцессий
при общем сходстве имеет свои особенности.
Полноту тождественности соотношений видов в пределах смежных
циклов;не может быть и потому, что изменчивость природного фона,
а следовательно, и условий существования сообществ никогда не
бывает аналогичной. Каждый природный цикл, будучи в общих чер-
тах похожим на предыдущий, вместе с тем несет и свои неповторимые
черты. Этот процесс не периодический со строгими интервалами,
ащиклический: повторение биологических процессов идет не по зам-
кнутому циклу (кругу), а по спирали.
Уместно в связи с этим привести замечание Н. М. Черновой
[1977], которая пишет, что «наиболее общей особенностью изучен-
ных сукцессий являетсй спиралевидный характер поступательного
изменения сообщества, что, по-видимому, можно отнести к универ-
сальным законам протекания всех сукцессий» (с. 181). ;Спиралевид-
ный характер развития сукцессий является фактически признанием
того, что сукцессии — это циклический процесс.
Мы приходим к выводу, что эволюционное развитие сообществ
осуществляется, в том числе и через непрерывные их циклические
смены. Циклически обратимые преобразования — это элементы на-
правленного развития биогеоценозов. Из последующих, сменяющих
220
друг друга, биологических, циклов формируется общий поступатель-
ный процесс развития экосистем.
Смена типов биогеоценозов как показатель сукцессии. В совре-
менной литературе широко распространено представление, что сук-
цессия — это смена одного типа биоценоза (фитоценоза) другим
[Мильков, 1970; Викторов, 1964; Сукачев, 1972]., Обсуждается воп-
рос, с какого момента этих изменений становится возможным гово-
рить о происшедшей смене биогеоценозов, или, точнее, о возникнове-
нии биогеоценоза другого, нового типа.,В. Н. Сукачев [1972] отвеча-
ет на это так: «Новый тип биогеоценоза возникает в том случае, ког-
да изменения всех или большинства его компонентов стали столь
значительны, что биогеоценоз стал качественно! другим, что суще-
ственно изменился и характер обмена веществом и энергией -как
внутри биогеоценоза (между его компонентами), так и между ними
и другими явлениями природы» (с. 381). .;
Концепции цикличности сукцессий такая точка зрения может и
не противоречить, например, при вековых сукцессиях в лесных био-
ценозах. Смена таежного сообщества последующими сукцессионными
рядами (кустарники, лиственный лес, хвойный лес) действительно
отражает смену одного типа биоценоза другим.
Но не для всех биогеоценозов сукцессию надо понимать как по-
добную смену типов сообществ. В рассматриваемых выше случаях
(лесостепь, пойма, озерные котловины) циклические перестройки
сообществ происходят в предёлах того же зонального типа биогео-
ценоза; происходит лишь перестройка доминирования видов, при
которой изменяется положение! и других компонентов в сукцессион-
ном ряду очередного сообщества. Так, в Барабе в сухую фазу 11-
летнего цикла в сообществе мелких млекопитающих заметную роль
начинают играть виды более ксерофитных местообитаний (например,
узкочерепная полевка), хотя эти виды являются характерными пред-
ставителями того же лесостепного биогеоценоза. Между тем, эти
смены сообществ в пределах данного типа биогеоценоза есть все ос-
нования относить к качественно иным, так как каждая такая смена
связана с глубокими ^экологическими изменениями в жизни сооб-
ществ. ; : . !
Выявляемые нами особенности сукцессий в сообществах живот-
ных аналогичны процессам, происходящим в фитоценозах, где, по
словам Л. Г. Раменского [1971], «нет двух лет за весь период, в кото-
рые состав растительности был бы одинаков» (с. 25). При чрезвычай-
ной сложности, многомерности и неповторимости аномалии траво-
стоя вычисление для него среднего состава за какой-либо период вре-
мени, по-словам Л. Г. Раменского [1971], не имеет смысла: это будет
фикция, лишенная какого-либо нормативного значения. «Следует,—
добавляет цитируемый автор, — не вычислять величины уклонений
от воображаемой нормы, а сравнивать реальные смены соседних лет
и стараться проникнуть в их закономерность» (с. 26).
О том, что сукцессии происходят в пределах одного типа биоцено-
за, вокруг «некоторого постоянного ядра», писал Д. Н. Кашкаров
[1945]. По его словам, «между биоценозами, сменяющими друг дру-
V
221
га во времени, границ нет. Но некоторые стадии развития количе-
ственно п качественно отличны от других. Качественным отличием
биоценозов будет прежде всего доминирование в них различных
экологических типов» (с. 263). Д. Н[ Кашкаров в этомщопросе раз-
делял точку зрения Ч. Адамса [19081, который.писал, что сукцес-
сия — это изменение* доминирования некоторых видов пли ассоциа-
ций над доминированием других (цпт. по [Кашкаров, 1).
В современной литературе понятие «сукцессия» подобным обра-
зом трактует С. *М. Разумовский [1981], понимая под сменой сооб-
ществ «прежде всего замену одних господствующих видов другими»
(С. 6). . . ; - ; ; . . Д .
Подводя итог дискуссии, заметим, что точка зрения на сукцес-
сию как циклический процесс чаще всего не противоречит многим
основным положениям уже сложившегося учения об экологической
сукцессии. В то же время понятие «циклическая сукцессия» позво-
ляет объяснить более широкий круг природных явлений, и мы счи-
таем возможным ^иёполъзовать! его для 'последующих - экологических
разработок^ : > ! ; । -
Таким образом, сукцессия, если ее не сопровождают ландшафт-
ны'е изменения, либо 'горообразовательные процессы, оказывается
сама по себе элементом (частью,1 этапом) какой-либо долговременной
природной цикличности. Точного повторения серий не происходит
в связи с различием их пионерных группировок. Йюбйя серия сукцес-
сии развивается обязательно циклично, для пёе характерны суточ-
ные, месячные, сезонные и другие виды колебаний, т. е. в каждой
с?рии й во всей сукцессии проявляется .одно из фундаментальных
свойств живой природы — Цикличность большинства происходящих
в ней процессов. ; ; । । .
' 'i ii-
h , 7 ' i 1 I , jHi 'заключение
_ i । I j ।'
। । i i ' 1 . !
Мцоголетнйе наблюдения сообществ мелких млекопитающих в Ба-
рабе и одновременный анализ"природно-климатической обстановки
в этом регионе позволили выявить характерные природные циклич-
ности. Применение спектрального анализа для обработки материа-
лов способствовало уточнению значений различных циклов п!соот-|
ношения пх мощностей. । 1 ,
Использование этогометода обработки данных! для уточнения
природной многолетней цикличности в популяциях мелких млеко-
питающих дало возможность выявить Скрытые колебания чис-
ленности. Другими способами выявить эти колебания, тем бо- '
лее уточнить значение периодов и соотношение мощностей различ-
ных периодических составляющих, можно только очень приблизи-
тельно. При расчете и построении спектров ритмов численности
многие из полученных пиков (периодов) имеют явный биологиче-
ский смысл и уже обсуждались разными исследователями, но каю
предположения пли феномены, выделенные только па качествен-
ном уровне. Спектральный анализ,позволил оценить их количест-
венно п принес новые биологические' сведения (уточнение длины
периодов, соотношение мощностей, количество периодических про-
цессов, видоспецпфпчность набора гармоник и пр.). Кроме того, по-
лученные спектры несут значительное число пиков заметной мощ-
ности, биологическая информация ц । которых мала( или отсутст-
вует вовсе. Это наряду I с точными количественными оценками
и легкостью/ получения спектров может послужить одним из сти-
мулов дальнейшего анализа циклических структур в популяциях.
В разных районах Барабы создаются своеобразные природно-
климатические условия, определяющие продолжительность циклов
численности в популяциях мелких млекопита'Ющпх. Природпо- j1
климатические условия и обусловливают > эти колебания численнос-
ти. В меньшей степени они вызываются биотическими причинами— ,
циклическим воздействием на популяции какого-либо доминирую-
щего по численности вида. Так, периодически.резко увеличивает плот-
ность популяции водяная крыса, тем самым являясь осциллято-
ром многолетних, колебаний <в сообществе мелких млекопитающих. (
В еще меньшей степени в поддержании и осцилляции ритмов дне- .
ленпостп играют роль внутрипопуляционные механизмы. В пользу
такого заключения говорит значительный популяционный полимор- .
физм цикличности численности.
223
Подавляющее число популяций имеют циклические колебания
численности примерно с 11-летним периодом, что соответствует мно-
голетнему циклу обводненности Барабы. Наличие па спектрах рит-
мов численности мелких млекопитающих мощных пиков, соответст-
вующих цикличности в 9—13 лет, подтверждает реальность такой
многолетней цикличности. . Многие мелкие млекопитающие кроме
',9—13-летней периодичности; имеют кратные ей гармонические сос-
.. тавляющйе ; численности с периодом 6 лет и 3—4 года. Такие ритмы
1 могут быть адаптированы к смене фаз увлажненности территории
Барабы. !
Все серые полевки имеют на спектрах’ритмов своей численности
! лики, соответствующие 3-летней цикличности. Именно такой дли-
ны циклы отмечены разными наблюдателями как характерные для
серых полевик. На нашем материале о помощью спектрального ана-
лиза у всех исследованных видов серых полевок доказано сущест-
вование пиков соответствующей частоты. Как правило, у этих ви-
дов жцвотных 4—3-летние периодические составляющие оказывают-
ся наиболее мощнымш среди прочих низкочастотных ритмов.
Выявленные с ; помощью спектрального анализа цикличности
в популяциях мелких млекопитающих могут иметь различный био-
! логический смысл. Сами по себе сезонные;и многолетние циклы —
это приспособления млекопитающих к цикличности природной сре-
ды. Соотношение мощностей, специфичное для вида, несет на себе
!отпечаток хронобиологических адаптаций. Это может касаться не
только видов, но и таксонов более высоких!panroBj Так, полевки
и мыши имеют 3—4-и 5-летние цикличности, но у первых наиболее
мощны 3—4-летние, а у вторых—5-летние. Следовательно, времен-
ные характеристики экологических ниш животных могут опре-
делять не только значения периода, но и мощность циклов.
Годовые ритмы мелких млекопитающих выражены на спектрах
в виде значительных по мощности пиков. Мощность этих периоди-
ческих составляющих*, .уточнение их положения на спектре и вооб-
1 ще особенности цирканных ритмов у млекопитающих — отдельная
болыпа,я; проблема изучения природной цикличности.
Биотопические ’ комплексы грызунов Барабы имеют собственные
колебания с различной периодичностью. Наиболее распространен
цирканный цикл,! характерный не только для всех биотопических
группировок,; но и щля сообщества в целом. Внешними синхрони-
; заторами такого цикла выступают, вйдимо, смена времен года и свя-
занные с нею фотопериодический и температурный ритмы. Длят всех
группировок характерны 2,5-и 3,3-летние цикличности, которые
обусловлены популяционными ритмами серых полевок, обычно
доминирующих среди грызунов в регионе. И, наконец,! частый в био-
' топических группировках и у сообщества в целом 9,8-летний ритм,
видимо, связан с (фазами увлажненности Барабы.
Нередко оценка процесса проводится по одной из его учиты-
ваемых функций. Мы оценивали цикличность жизнедеятельности по
ритмам численности. Можно отметить, что процессы жизнедеятель-
ности в популяциях и сообществах грызунов полицикличны. По дан-
224
ным отловов животных можно выделить имеющиеся периоды и их
значение оценить с помощью спектрального анализа.
Цикличность природной среды влияет на условия существования
биоценоза, что позволяет говорить об общем значении ее з жизни зо-
нального ландшафта. Установленная на примере лесостепной зоны
Западной Сибири цикличность смены сообществ не является особен-
ностью только этого природного региона. Она свойственна также и
другим зональным (интразональным) биогеоценозам, что подтвер-
ждается данными исследования сукцессий в пойме р. Оби, в озерных
котловинах степной; и лесостепной зон. I j
Цикличность— это! характерная jj особенность зональных биоце-
нозов!, для которых регулярная изменчивость природного фона яв-
ляется средой существования и условием развития. Исследование
смен населения в сообществах мелких млекопитающих на между-
речьях лесостепной зоны Западной Сибири показало, что перестрой-
ки видовой структуры подчиняются определенным временным за-
кономерностям и этот процесс цикличен. Выяснилось, что в преде-
лах сходных фаз природного цикла видовые структуры по общим-
признакам повторяются. Последнее особенно четко регистрируется
по господствующим в сообществах видам и субдоминантам. Этот
процесс смены сообществ мелких млекопитающих подчиняется су-
ществующим здесь сменам фаз обводненности территории в 11-лет-
них циклах и вспышкам массового размножения доминирующе-
го вида.
При классификации экологических сукцессий целесообразно
принимать во внимание наличие в природе характерных цикличе-
ских сукцессий, понимая под ними обратимые видовые перестройки
сообществ в пределах данного биогеоценоза, подчиненные местным
циклам природной среды, адекватно изменяющим условия сущест-
вования сообществ, биоценотические связи, численность отдельных
видов и популяций, порядок доминирования видов в сообществах,
их биологическую продуктивность.
Поскольку циклы природной среды— общее явление для всего
биогеоценоза, имеется возможность исследования сукцессий не только
фито-и зооценозов, но. и всего биоценоза в целом. В этой цельности
подхода и состоит принципиальная сторона выдвигаемой концеп-
ции, которая позволяет трактовать и изучать) экологические сук-
цессии в биогеоценозах как единое взаимосвязанное явление. Ориен-
тировка на циклы природной среды помогает биоценологам в пони-
мании путей развития! > подсистем исследуемогб- биогеоценоза.
Именно в этом осуществляется, экологический, системный подход
к анализу биологических процессов в природе.
Один из возможных причинно-обусловленных путей, формиро-
вания указанных биологических ритмов может быть выражен сле-
дующей последовательностью явлений: ритмы солнечной и другой!
космической цикличности— циклические колебания климата—
природные циклы в биогеоценозах— биологические процессы в
сообществах.
i
5 А. А. Максимов, Л. Н. Ердаков
225
Таким образом, на основании наблюдений и расчетов мы выде-
ляем конкретные виды природной цикличности барабнпского биогео-
ценоза. Это колебания передвижений животных с характерной перио-
дичностью от 3 до 20 сут. Они связаны с территориальным поведе-
нием, цикличностью активности и синхронизируются локальными
климатическими условиями местообитаний? Хорошо проявлены по-
пуляционные циклы изменений численности с -цериодами от 3 до
6 лет, характерные для многих видов ; грызунов ] Одним из таких
циклов динамики численности является повторяемость вспышек
массового размножения водяной крысы, цикличность которых на-
ходится в пределах' от 7 до 11 лет. Она соизмерима по значению с
выделенными нами циклами в биотопических группировках |жи-
цотных— 9,8-летний и, кроме того,, с описанным для1 Барабы
, 14-летним циклом увлажненности территории. Последний является
ь'«одновременно и циклом (сукцессии животных сообществ,, характер-
ным для' Барабы. I ! ' । ' i
Важным i следствием цикличности iявляется то, что .видовые пе-
рестройки сообществ изменяют биологическую продуктивность био--
геоценозов.-Изучение, колебательных процессов может Служить прин-
ципиальной основой для прогнозирования изменчивости Природ-
ных ресурсов в интересах различных отраслей народного хозяйст-
ва. । Изучение экологических сукцессий, т. е. смен растительных
и животных сообществ в ходе их развития, тес по связано с из-
менчивостью биологических ресурсов и поэтому относится к осо-
бенно (актуальным, проблемам биоценологии.................
Выявленные цикличности в изменениях численности популяции,
комплекса или сообщества наряду с прагматической ценностью
(прогнозирование) имеют еще и теоретическое значение.; । Прежде
всего представляется вероятным, .что таким сложным системам,
как популяция и сообщество, внутренне присуща колебательная
природа. Выделение основных цикличностей в жизнедеятельнос-
ти их может иметь, значение при моделировании процессов, проис-
ходящих в1 популяциях, а возможно, они могут. стать основой при
разработке методов управления этими 'процессами.|
I I ! г: '' । i ’ ; . 1 i > I. ’ I > 1C ; .
! ..I .J >. H i , i I ' . I. ।
1 ‘ lv j/ ' - + И НГ' l| ;
! ' j '
” j I ! . L - ' i - I
' I " • I L - I H? л -t . ! " -
’ 'J < 1 ih . r Hl' ч -
.......г H '! " 1 '
i ' ! i I II! i I I .
. I -. 1 !
J ЛИТЕРАТУРА
Абашкин С. А. Климатическая цикличность и влияние ее на некоторые отрасли
хозяйственной деятельности человека и биологические процессы в Бара-
бе.— В кн.: Природные циклы Бйрабы и их хозяйственное значение.
Новосибирск: Наука, 1982, с. 55—65.
Аветисян О. Р. Основные факторы, обусловливающие численность обыкновен-
ной полевки и принципы прогноза.— В кн.: Труды1 НИИ' защиты растет
ний АрмССР.; Т. 1. Ереван, 197-0, с.' 365—384. ,
Александрова В. Д.‘ Изучение смен растительного покрова.— В кн.: Полевая
геоботаника. Т. 3. Л.: Наука, 1964, с. 300—447.
Александрова В. Д. Классификация растительности..— Л.: Наука, 1969.— 275 с.1
Ананьева С. И. Закономерности микробиотопического распределения Collem- |
bold в пятнистой тундре Западного Таймыра в связи с динамикой расти-
тельного покрова.— Зоол. журн., 1971, т. 50, вып. 6, с. 817—825. 1
Башенина Н, В. Пути адаптации мышевидных грызунов.— М.: Наука, 1977.—
355 с..
Букин В. Н. Эпидемиология туляремии в Барабе при разных фазах увлажнен-
। ности территории в 11-летцих циклах.— В кн.: Природные циклы Бара-
бы и их хозяйственное значение. Новосибирск: Наука, 1982, с. 120—127.
Вагина Т. А. Динамика [развития растительного покрова Барабинской низмен-
ности,— В кн;: Природные циклы Барабы и их хозяйственное значение.
। Новосибирск: Наука, 1982, с. 65—78. • ,. i
Вагина Т. А. Луга Барабы. Ч. 1.— Новосибирск: Наука, 1962.— 198 с.
Воронов А. Г. О колебаниях уровня озер Кустанайской области Северного Ка-
захстана.— Изв. Всесоюз. геогр. о-ва, 1947, т. 79, вып. 5, с. 523—536.
Воронов А. Г. Геоботаника.— М.: Высш, школа, 1963.— 374| с.
Воронцов Н. Н. Эволюция пищеварительной системы ^грызунов (мышеобраз-
ные).— Новосибирск: Наука, 1967.— 240 с. j . . ' ,
Воскобойнйков В.| А. Влияние периодических колебаний уровня воды на вос-
производство окуня оз. Чаны,— В кн.: Природные циклы Барабы и их
хозяйственное значение. ’ Новосибирск: Наука, 1982, с. 99—105.
Гвиннер Э. Годовые ритмы: общая персйектива.— В кн.:-Биологические ритмы
Т. 2. М-: Мир, 1984а, с. 44—55. i и
Гвиннер Э. Цирканнуальные системы,— Тамижё, 19846, с. 55—81. .
Герман В. А., Максимов А. А., Черанева М. Г. Указатель литературы по реч-,
ным ноймам.— В кн.: Сукцессии, животного населения в биоценозах пой-’
мы реки Оби. Новосибирск: Наука, 1981, с,. 175—258. . ;
Гиляров М. С, Соотношение размеров и численности почвенных беспозвоноч-
ных,- Докл. АН СССР, 1944, т. 43, № 6-, с. 283-285. I .
Гиляров А. М. Индекс разнообразия и экологическая сукцессия.— Журн. общ.
биол., 1969, т. 30, № 6, С.; 652—657. , । .
Глотов И. Н. Распространение и численность мышевидных^ грызунов.— В кн.:
Биологическое районирование Новосибирской области. Новосибирск:
Наука, 1969, с. 33—58. ।
Громов И. М., Поляков И. Я. Полевки (Mierolinae).— В кн.: Фауна СССР. Мле- ’
копитающие. Т. 3, вып. 8. Л.: Наука, 1977.— 504 с.
Давыдова М. С.,.Глотов И. Н. Изменение чидозого состава и численности пара-
зитических гамазовых клещей в Барабинской низменности при смене
А
влажной и сухой фаз в 11-летних циклах.— В кн.: Природные циклы Ба-
рабы и их хозяйственное значение. Новосибирск: Наука, 1982, с. 116—
120.
Дажо Р. Основы экологии.—М.: Прогресс, 1975.— ’415 с. I
Де Курси П. Сдвиг фазы ритма активности у грызунов под влиянием внешних
синхронизирующих факторов.— В кн.: Биологические часы. М.: Мир,
1964, с. 91—103. i ’ . '
Дженкинс Г., Ватте Д. Спектральный анализ и его!приложения.— М.: Мир,
1971.—316 с. \
Ердаков Л. Н. Организация ритмов активности грызунов,— Новосибирск: На-
। ука, 1984.— 180 с.
Ердаков Л. Н., Максимов А. А., Рябко Б. Я. Экологическая структура сооб-
щества мышевидных грызунов (на примере Западной Сибири).— Изв.
СО АН СССР, 1980, с. 55-62. f ,
Ердаков Л. Н., Путилов А. А., Тарновский А. В. Свободнотекухцие ритмы ак-
тивности трех видов грызунов,— Экология, № 5, 1981, с. 71—77.
Ивантер Э. В. Популяционная, экология мелких млекопитающих таежного се-
веро-запада СССР.—Л.: Наука, 1975.— 246 с.
Иоганзен Б. Г. О современном состоянии уровня барабинскпх озер,—Изв.
Всесоюз. геогр. о-ва, 1939, т. 71, № 7, с. 1034—1042. ।
Иоганзен Б. Г. Колебания уровня равнинных озер Западной Сибири как при-
чина многолетних изменений состава и численности их рыбного населе- ,
ния.— В1 кн.: Вторая научная конференция Томского ун-та (тезисы
докл.). Томск, .1951, с. 118—122. .
Иоганзен Б. Г. К изучению типов колебания оз. Чаны.— Тр. Том. ун-та, 1956',
т. 133, с. 55—66.
Калабухов Н. И. Периодические изменения в организме-грызунов,—Л.: HajT-
ка, 1969,— 248 с.
Камшилов М. М. Взаимные отношения между организмами и их роль в эволю-
ции.— Журн. общ. биол., 1953, т. 20, № 5, с. 370—379.
Камшилов М. М. Биотический круговорот.— М.: Наука, 1970.— 157 с.
Каталог млекопитающих СССР/Под ред. Громова И. М., Барановой Г. И. Л.: I
Наука, 1981.— 455 с. ,
Кашкаров Д. И. Основы1 экологии животных.—Л.: Медгиз, 1933,— 600 с.
Кашкаров Д. Н. Основы экологии животных. — iM..: Учпедгиз, 1945.— 383 с.
Кендалл М. Дж., Стюарт А. Многомерный статистический анализ и временные
ряды,— М.: Наука, 1976.— 723 с.
Колесник Ю. А., Тимофеева А. П., Помыкалов В. Г., Чупахина Т. И. Корреля-
ционный и спектральный анализ динамики численности морского котика
острова Тюлений (Охотское liope).— Зоол. жури., 1978, вып. 1, с. 115—
124. . i
Корсаков Г. К.-. Смиренский А. А: Зарастающие водоемы и их использование
для ондатроводства,—М.: ,Заготиздат, 1956.— 136 с.
Кошкина Т. В. Прогнозирование численности лесных полевок по экологиче-
ским показателям состояния популяций,—Бюл. МОИП. Отд. биол.,
1969, т. 74, № 2, с. 5—16. 1
Крайнов В. М. Колебания численности язя оз. Чаны в связи с цикличными из-
менениями уровня воды.— В кн.: Природные циклы Барабы и их хо-
зяйственное значение1. Новосибирск: Наука, 1982, с. 105—111.
Куркин К. А. Системные исследования динамики (лугов.—М.: Наука, 1976.—
280 с. . "
Куркин К. А., Стебаев И. В. Вспышки массового размножения нестандартных
саранчовых в Барабе и ее влияние на луговую растительность.— Бюл.
МОИП. Отд. биол., 1959, т. 64, вып. 1, с. 51—60.
Лавренко Е. Основные закономерности растительных сообществ и пути их изу-
чения.— В кн.: Полевая геоботаника. Т. 1. М.— Л.: Изд-во АН СССР,
; 1959, с. 13—75.
Лпчков Б. Л. Природные воды Земли и литосферы.— М.— Л.: Изд-во АН СССР.
I960:— 164 с. .
Максимов А. А. О двух фаунистических комплексах в биоценозе тундр.— Реф.
науч,- исслед. работ АН СССР за 1945 г. М., 1947, с. 252.
228
Максимов А. А. Фауна млекопитающих в природных очагах Западной Сибири
и роль водяной крысы как основного эпидемически опасного вида грызу-
на в этих очагах,— В кн,: Водяная крыса и борьба с ней' в Западной
Сибири. Новосибирск: Кн. пзд-во, 1959а, с. 27—237.
Максимов А. А. О кормовых связях в биоценозах тундры,— В кн.: Тр. Биоло-
гического ин-та СО АН СССР. Вып. 5, зоологический. Новосибирск,
19596, с. 85—90.
Максимов А. А. Цикличность массовых размножений водяной крысы в Запад-
ной Сибири в сопоставлении с солнечной активностью.— В кн.: Вопросы .
зоологии. Томск; изд. Том. ун-та, 1966, с. 1216—217.
Максимов А. А.'Зона вредности водяной крысы в Западной Сибири, методы уче-
та численности и прогноза.— Новосибирск: Наука, 1967.— 59 с.
Максимов А. А. Динамика численности и ритмы эпизоотий у грызунов в сопо-
ставлении с циклами солнечной активности.— В кн.: Влияние солнечной
активности на атмосферу и биосферу Земли. М.: Наука, 1971, с. 61—74.
Максимов А. А. Структура и динамика биоценозов речных долин.— Новосиг t
бирск: Наука, 1974,— 260 с. । i .
Максимов А. А. Практический опыт использования данных по солнечной актив-
ности для долгосрочных прогнозов вспышек массового размножения гры-
зунов.— В кн.: Физико-математические и биологические проблемы дей-
ствия электромагнитных полей и ионизации воздуха, Т. 1. М.: Наука,
1975, с. 91—95.
Максимов А. А. О прогнозе туляремии в лесостепной зоне Западной Сибири.— t
В кн.: Природноочаговые антропозоонозы (тезисы докладов). Омск, 1976,
с. 104—105. . .
Максимов А. А. Тппы вспышек и прогнозы массового размножения грызунов
। (на примере водяной крысы).— Новосибирск: Наука, 1977.— 189 с.
Максимов А. А. Цикличность массовых размножений животных — основа дол-'
госрочного прогнозирования.— Экология, 1978, № 6, с. 5—13.
Максимов А. А, Биоценоз и проблема прогнозирования биологических про- ,
цессов в природе,— В кн.: Экологическое прогнозирование. М.: Наука,
1979а, с. 7—13.
Максимов А. А. Циклы Барабы,— Газ. «Сов. Сибирь», 19796, 18 ноября. ,
Максимов А. А. Требуется нестандартный подход. Учитывать 11-летние природ-
ные циклы в сельском хозяйстве Барабы.— Газ. «Колос Сибири», 1981а,
№ 11 (409), 15 марта.
Максимов А. А. Долгосрочное экологическое прогнозирование.— Газ. «За на-
уку в Сибири», 19816, № 13 (994) от 26 марта, с. 4.
Максимов А. А. Соотношение между учением о факторах динамики численности
животных п проблемой долгосрочного прогнозирования вспышек мас-
сового размножения.—Экология. 1982, № 1, с. 38—45., , t
Максимов А. А. Исследование смен фаз увлажненности территории лесостепной •
зоны Западной Сибири в 11-летних циклах (к истории вопроса)..— В кн.:
Природные циклы Барабы и их хозяйственное значение. Новосибирск:
Наука, 19826", с. 6—24. .
Максимов А. А. Многолетние колебания численности'животных, нх причины
' и прогноз.— Новосибирск: Наука, 1984а,— 250 с.
Максимов А. А. Колебания биологической продуктивности животных сообществ
в смей с природной цикличностью.— В кн.; Вид и его. продуктивность
в ареале. Ч. 2. Свердловск, 19846, с. 32—33.
Максимов А. А., Абашкин С. А. Климатические циклы Барабы. — Газ. «За на-
уку в Сибири»,' 1976 г., 4 марта.
Максимов А. А., Ердаков Л. Н., Сергеев В. Е., Салтыков В. В. Сукцессии на-
селения землероек и грызунов в пойме среднего течения Оби.— В кн.:
Сукцессии животного населения в биоценозах поймы реки Оби. Ново-
сибирск: Наука, 1981, с. 5—63. .
Максимов А. А., Леонов Ю. А., Глотов И. Н. и др. Мышевидные грызуны и зем-
леройки в озерных очагах геморрагической лихорадки 'Северной Кулун-
ды и Барабы. — В кн.: Биологическая и эпизоотологическая характери-
стика очагов омской геморрагической лихорадки Западной Сибири. Но-
восибирск: Наука, 1974, с. 6—24.
229
Максимов А. А., Понько В. А., Сытин А. Г. Смена фаз увлажненности Барабы
(характеристика и прогноз). Новосибирск: Наука, 1979.— 64 с.
Максимов А. А., СаИбв Н. П., Абашкин С. А., Мерзлякова Е. П. Массовое раз-
множение водяной крысы в Барабе, в 1971—1974, гг,— В кн.: Вспышка 1
размножения; водяной крысы. Новосибирск: Нау^ка, 1976, с. 9—25. *
Максимов А. А., Сытин А. Г. К методике прогноза смен фаз увлажненности ле-
состепной зоны Западной Сибири в 11-летних циклах в связи с проблемой
ондатроводства.'—; В кн.:, Проблемы; ондатроводства. Киров, ' 1979, с. <
С. 166 —168. | | , : , .
! Максимов А.; А., Фолптарек С. С., Барбаш Л. А. и др. Обзор массового раз- 1
i |множения водяной крысы- в лесостепной зоне Западной Сибири в 1956 —
; 1962 гг.— В кн.: Животный мир Барабы. Новосибирск: Изд-во СО АН
СССР, 1965, с. 5-28. , |
Максимов А. А., Черанева М. Г. Значение организационно-хозяйственных ме-
роприятий по защите зерновых посевов от водяной крысы.— В кн.: Хи- ,
' мизация и защита растений. ;Новосибирск; ,1979, с. 98—107. #
Макфедьен Э. Экология щивотпых.'— М.: .Мир!, 1965,— 375; с. , i '
1 Матюшкин Е. Н., Смирнов Е. И,, Сизова Г. ЕЕ-Территориальные'группировки I
(сообщества)^ мышевидных грызунов Среднего Сихотэ-Алиня.— В кн.:
! Материалы к познанию флоры и фауны СССР.,Вып. 47 (62). М., 1972,
i ’ С. 116 — 148. T j" , । , .
Межжерин В; А. ^Динамика численности животных и построение прогнозов.— | (
.»« Экология,:1979, № 3, ,с. 5 —12. : ! /
; Мерсер Д. Аналитичёсйие методы исследования периодических процессов, за-
i маскированных случайными флуктуациями.—В кн.: iБиологические ча-I Л
сы., М.: Мир, 1964,, с. 126—153. ; !
МильковАФ, Н. Словарь-справочник по физической-географии.— М.: Мысль, £•'
1970.- 344 с. ; i о ; и । '
Мордковпч В. Г. Сукцессии животного Населения почв в аккумулятивных ланд-
шафтах степной Сибири.— В кн.: -Проблемы почвенной зоологии. М.: ;
Наука, 1966, с. 89—90. , j
Мордкович В. Г. Динамика составами численности мезогерпетобпя как показа- Г)
, тель сукцессионного процесса.— В кн.: Структура, функционирование I
п эволюция системы биогеоценозов Барабы. Т. 2. Новосибирск: Наука, 9
; 1976, с. 401—416. ' 1 , , . .
1 Наумов ЕГ. П. Экология животных.— М,: ВысшОшк.') 1963а.— 618 с.
Наумов Н. П. Органический мир как|целое,— В кн.: Очерк Диалектики живой и
; природы. М’,, 19636,. с.| 100—133, , i,
Николаев А. С., Богомолова И. Н. Запасы продуктивной; влаги под основными ,,;
; .; [ сельскохозяйственными, ;культурамп Северной Барабы и их многолетние 1
! ' изменения.— В кн.: П^йрбдные циклы БАрабАии их хозяйственное зна-
чение. Новосибирск; Наука, 1982, с. 33—40.- , ! ; :
Николов X. М. Статистическое определение цикличности' в динамике .численно- '
। , - сти обыкновенной по.цевки в луговой степи Центрально-Черноземного за- • (
' поведника.—В кн.; Материалы 12. конференции молодых ученых биоАа- ।
i i! Н ка МГУ. М.(:'Изд-во МГУ;,'1981ф с. 10—13;. I. - । ,, - , Й';’
Одум Е. -Экологий,—; М.: , Просвещение, , 19,68.— 167 с. 1 ..
Одум Ю; Основы Экологйи.— М.: .Мир, 1975.— 740 с. , ' • )
ПанадиадИ 'А.;Д. Барабинская низменность,— М.: Географиздат, 1953.— 231 с, [,
Песенко Ю. А. Принципы и методы количественного аналйза в фаунистических
исследованиях.— М.: Наука, 11982,— 285 ю. । ’
Панов В. В. Водный режим болот Северной Барабы (по материалам болотнс-
। стоковых постов).— В кн.: Природные,циклы Барабьци их хозяйственное j
значение. Новосибирск: Наука, 1982( с. 41—46. '
Пванка Э. 1 Эволюционная (Экология.—М.: Мир, 1981.— 399 с. ,1
Плохинский И. А. Биометрия.— Новосибирск: Пзд-во СО АН СССР, 1961.— )
362 с. I . , . i!
Плохинский П. А. Биометрия.— М.: Пзд-во МГУ, 1970.-г 362 с. • !
Поляков Е. Ф. Многолетние изменения численности охотппчье-промысловых
животных в Барабе в связи со сменой фаз увлажненности.— В кн.: При-
родные циклы Барабы и их хозяйственное значение.— Новосибирск:
Наука, 1982, с. 78—99.
Природные циклы Барабы и их хозяйственное значение.— Новосибирск: Нау-
ка, 1982.— 152 с.
Работнов Т. А. Изучение флуктуаций (разногодичной! изменчивости) фитоцено-
зов.— В кн.: Полевая геоботаника. Т. 4. Л.: Наука,' 1972, с. 95—136.
Работнов! Т. А. Факторы устойчивости наземных фитоценозов.— Бюл. М01Ш.
Отд. биол., 1973, т. 78, вып. 4, с. 67—76.
Работнов Т. А. (Фитоценология.— М.: Изд-во МГУ, 1978.— 384 с.
Разумовский С. М. Закономерности динамики биоценозов.— М.: Наука, 1981.—
' 231 с. I .
Разумовский С. М., Рыбаков Л. Б., Тихомирова А. Л. Закономерности сукцес-
сий биоценозов Подмосковья (на примере Приокско-Террасного заповед-
ника).—-В кн.: Региональный экологический мониторинг. Мп Наука,
1983, с. 230—242. । . [ I
Раменский Л. Г. Избранные работы. Проблемы п методы изучения раститель-
ного-покрова,—Л.: Наука, 1971.— 334 с.
Раусон К’. Влияние температуры тела на ритмы активности млекопитающих,—
В кн.: Биологические часы. М.: Мир, 1964, с. 182—196.
Реймерс И. Ф., Яблоков А. В. Словарь терминов и .понятий, связанных с охра-
ной живой природы.— М.: Наука, 1982.— 144 с.
Ряклефс Р. Основы общей экологии.— М.: Мир, 1979.— 424 с.
Семеиов-Тян-Шанекий О. II. Цикличность в популяциях лесных йолевок.—
Бюл. МОИН. Отд. бпол., т. 75, вып. 2, 1970, с. 11—-26.
Смиренскпй А. А. Водные кормовые и защитные растения в охотничье-промыс-
ловых хозяйствах. Вып. .1.— М.: Заготиздат, 1950.— 135 с.
Смирнов П. А. Флора Прпокско-Терраспого государственного заповедника.—
В кн.: Труды Приокско-Террасного’ заповедника. Вып. 2.— М., 1958.—
248 с.
Смирнов П. К. Эколого-физиологическое исследование некоторых видов гры-
зунов.— Л.: Наука, 1968.— 133 с.
Сляднев А. П. Засухи на юго-востоке Западной Сибири, их признаки п повто-
ряемость.— В кн.: География Западной Сибири. Новосибирск: Зап,-
Сиб. кн. изд.-во, 1965, с. 123—146.
Солбриг О., Солбриг Д. Популяционная биология и эволюция.— М.: Мпр,
1982.— 484 с.
Сообщества мелких млекопитающих Барабы,—Новосибирск: Наука, 1978.—
230 с. |
Старка Ф., Герасимов С. Числена динамика и зонп на вредност на обикновена-
та полевка (Microtus arvalis Pall.) в Бъелгария.— Екология (НРБ), 1977,
т. 3,, ср 79—9,1. t . । , ! • j
Стебаев И. В. Изменения животного населения почв в ходе их развития на скалах
и на рыхлых продуктах выветривания в лесолуговых ландшафтах .Юж-
ного Урала,— Pedobiologia,, 1963, Bd 2, Н. 4, S. 265—309. , ।
Стебаев И. В. Пространственная структура населения беспозвоночных степных
котловин, юга Сибири в связи о особенностями структуры их почвенного
покрова.— В кн.: Доклады спб. почвоведов к IX Международному кон-
грессу. Новосибирск: Наука, 1968, с. 45. ,
Структура, функционирование и эволюция системы биогеоценозов Барабы.—
Новосибирск: Наука, Т. 1, 1974.— 306 с.; Т. 2, 1976.— 495 с.
Сукачев В. Н. О принципах генетической классификации в биоценологии.—
। Журн. общ. биол., 1944, т.г 5, К» 4, с. 213—226. ,
Сукачев В. Н. Некоторые общие теоретические вопросы фитоценологии. —
В кн.: Вопросы ботаники. Вып. Ij— М..— Л.: Изд. АН СССР, 1954, с.
с. 291—309. I
Сукачев В. Н. Избранные труды. Т. 1,—Л.: Наука, 1972.— 418 с.
Сукцессии животного населения в биоценозах, поймы реки Оби,— Новосибирск:
Наука, 1981.— 263 с.
231
Сытин А. Г., Максимов А. А. Физико-статистическая обусловленность смен фаз
увлажненности лесостепной зоны Западной Сибири в 11-летних циклах
по осадкам и стоку рек.— В кн.: Природные циклы Барабы и их хозяй-
’ ствснное значение. Новосибирск: Наука, 1982, с. 46—54.
Токарев В. М. Многолетние колебания гидротермического режима Барабы,—
Там же с. 24—33. . с
, Уоддингтон К. X. Основные биологические концепции.— В кн.: На пути к тео-
ретической биологии. 1. Пролегомены. М,: Мир, 1970, с. 11—38. •
Федосов Н. А., Максимов А. А. Колебания серологического фона мелких млеко-
питающих в отношении туляремийного микроба в связи с различной ув-
лажненностью Барабы.— В кн.: Природные циклы Барабы и их хозяй-
ственное значение.. Новосибирск: Наука, 1982, с.. 141—145.
Фиигерман М. Приливные ритмы у морских организмов.— В кн.: Биологиче-
ские часы. М.: Мир, 1964, с. ,667—682. ,
Фолитарек С. С. Некоторые вопросы охраны и прогнозирования природы.—
В кн.: Охрана и преобразование природы лесостепи Западной. Сибири.
Новосибирск: Наука, 1976, с. 3—70.
Формозов А. И. Озерная лесостепь и степь Западной Сибири как области мас-
сового обитания птиц,— Бюл. МОИП. Отд. биол. 1934, т. 43, вып. 2,
с. 25,6-285. i> , . i
Хауншельд К. Лунные ритмы.,— В кн.: Биологические часы. М.: Мир, 1964,
,с. 682—691. 1
Чернов Ю. И. О некоторых индексах, используемых при анализе структуры жи-
вотного населения суши.— Зоол. журн., 1971, т. 50, вып. 7, с. 1079—1092.
Чернова Н. М. Экологические сукцессии при разложении растительных остат-
ков.— М.: Наука,1 1977.— 200 с. ,
Чернова И. М., Былова А. М. Экология.— М.: Просвещение, 1981.— 255 с.
Численко Л. Л. О размерной структуре пелагиали мирового океана.— Журн.
общ. биол., 1968, т. 28, № 5,, с. 529—540.
Численко Л. Л. О размерной структуре мировой фауны млекопитающих.—
, Бюл. МОИП. Отд. биол., 1969, т. 74(1), с. 5—13.
Шварц С. С. Экологические закономерности эволюции.— М.: Наука, 1980,—
277 с.
Шенников А. П. Введение в геоботанику.— Л.: Изд-во ЛГУ, 1964,— 447 с.
, Шилов И. А. Эколого-физиологические основы популяционных отношений жи-
вотных.— М.: Изд-во МГУ, 1977.— 261 с.
Шнитников А. В. Внутривекбвые колебания уровня стетаых озер Западной Си-
бири и Северного Казахстана.— В кн.: Труды лаборатории озероведения
1 АН СССР. Т.-1. М„ 1950.— 129 с.
Шнитников А. В. Изменчивость' общей увлажненности материков северного
полушария.— Зап. геогр. о-ва СССР, т. 16, М.— Л.: Изд-во АН СССР,
1957, . с. 1-337. ;
Шнитников А. В. Внутривековая изменчивость компонентов общей увлажнен-
ности.— Л.:^Наука, 1969,— 244 с.
Юдин Б. С. Экология бурозубок (род Sorex) Западной Сибири.— В кн.; Труды
Биол. ин-та. Вып. 6. Новосибирск, 1962. с. 33—134.
Adams Ch. С. The ecological succession of birds.— The Auk, V. 25, N 2, 1908,
p. 109-153. . , .
Adams Ch. C. An ecological study of Prairie and forest invertebrates.— Bull, of
; the Illinois State, Laboratory of Natural History, 1915, v. 9, art. II.
j Alee W., Emerson A., Park P. e. a. Principles of animal ecology.— Philadelphia:
W. B. Saunders Company, 1949.— 837 p.
Batzli G. O. Population and energeties of small mammals i,n tundra ecosystem. —
In: Tundra ecosystem: a comparetive analysis. Cambridge Univ. Press.,
i 1981, p. 377—396. .
Berthold P. Beivielen Tieren innerer Jahres — Kalender.— Umschan Wiss. and
Techn., 1975, Bd. 75, N 22, 700 S.
Bezzel E., Reichholf J. Die Diversity als Kriterium zur Bewendung der Reichhal-
tigneit von Wasserfogel-LebensrKnmen.— J. Ornitol., 1974, v. 115, N 1.
Buzas M. A. Patterns of species diversity and explanation.— Texon, 1972, v. 21,
N 2-3.
232
i
Clements F. E. The development and .structure of vegetation.— Bot. Surv. Neb-
raska, 1904, v. 7, p. 75—81.
Clements F. E. Plant succession: an analysis of the developmlent of vegetation.—
Curnegie Inst. Washington, Publ., 1916, N 242. p. 1—512.
Clements F. E. Nature and structure of the climax, 1936, v. 24, N 1, p. 253—284.
Cockburn A., Braithwaite R. \V., Lee A. The response of the heathrat, Pseudomys
shortridgei, to pyrie succession: a. temporally dynamic life-history strate-
gy.— J'.. Anim. Ecol.,: 1981, v. 50, N 3, p. 649—666. ,
Daan S., Slopsema S. Short term rhythms in foraging behaviour of the common
vole, Microtus arvalis.— J. Comp. Physiol., 1978, A 127, N 3', p. 215—217.
Dewey E. R. The 42-year rhythm.— Cycles, 1982, v. 33, N 6, p. 195—196.
Dienske H. The importance of Social interactions and habitat in competition bet-
ween Microtus agrestis and 71/. arvalis.— Behaviour, ,b. 71, N 1—2, 1979,
p. 1-125. i.
Fox B. J. Fire and mammalian secondary succession in an Australian coastal he-
ath.—Ecology, 1982, v. 63, N 5, p. 1332—1341.
Harold H., Howard W. E. A 19-year study of microtine population fluctuations
using time series analysis. — Ecology, 1981, v. 62(11), p. 930—937.
Gattennann R. Zur Biorhythmik des Kaninchene (O. cuniculus (L.) f. domestica). —
J. Mitt: Circadiane rhythmen der lokomotorischen Akti vital, dor Fritter
und Wasseraufnahme, der Harnabgabe und der Kdrpergewichts, 1978. Zool.
Jb. Physiol., Bd 82, S. 214—233.
Grahams S. A. The felled trunk as an ecological unit.— Ecol. vol., v. 1, N 4, 1925,,
p. 397-412. ’ >- ‘
Grange W. B. The way to game abundance.— N. Y.— L.: Charles Schriner’s sons,
1949.— 85 p. !
Grant P. R. An 11-year study of small mammal populations at Mont. St. Hilaire,
Quebec.— Can. J. Zool., 1976, v. 54, N 12, p. 2156—2173. I
Grodzinsky W. Sezonowe smiany w rytmice dobowej aktywnosci drobnych gry-
, zoni.— Ecol. Polska, 1963, Ser. B.,- t. 9, N 1, s. 3—17.
Heirston N. G. The soil zoology of a Belgian Oak woods.— Ecology, v. 40, 1959.—
168 p.
Heller П. C., Poulson T. L. Cireannian rhythms. 1.1-Endogenous and exogenous
factors controlling reproduction and hibernation in chipmunks (Eutamias)
and ground squiriels (Spermophilus).— Comp. Biochem. and Physiol.,
1970, v. 33, N 2, p. 357—383.
Herbert R. H. The origin of species diversity in ecological communities.— Taxon,
1972, v. 21, N 2-3.
Jost V. Das Aktivitatsmuster der schermaus Arvicola terrestris (L.) — Zool.
Jahrb., 1973, Abt. 1, Bd 77, N 1, S. 98-106.
Lindeman R. The trophic dynamic aspect of ecology.— Ecology, 1942, v. 23,
p. 399—418. ;
MacArthur R. H. Fluctuations of animal populations and a measure of community
stability.—Ecology, 1955, v., 36, p. 533—536.'
Mallory F. F., Clulow F. V. Evidence of pregnancy failure in the wild meadow
vole, Microtus pennsylvanicus.— Can. J. Zool., 1977, v.>55, N 1, p. 1—17.
Margalef R. Perspectives in ecological theory.— L., 1968.— 112 p.
May R. M. Stability and complexity in model ecosystems. Princeton Univ. Press,
Princeton. N. Y., 1973.— 28 p.
McNaughton S. I., Wolf L. L. Dominance and the nicl in ecological sistems.—
Science, 1970, v. 167, N 3915, p. 131—139. :
Morosovsky N., Melnyk R. B., Lang K. e. a. Infradian cycles in dormice (Glis
.glis).— J. Comp. Physiol., 1980, A 137, N 4, p. 315—339.
Myllvmaki A. Outbreaks and damage by the field' vole Microtus agrestis (L.) in
Finland 1945—1973.— Eppo Pubis., 1974, C, N 31, p. 13—22.
Odum E. The strategy of ecosystem development.— Science, 1969, v. 164, N 3877,
p. 262-270.
Pengelley E. T., Asmundson S. M. Free — running periods of endogenous circan-
nian rhythms in the golden mantled ground squierel, (5-itellus lateralis.—
Compar. Biochem and Physiol., 1969, v. 30, N 1, p. 177—183.
Pimlott D. П. The value of diversity.— Trans. N. Amer. Wildlife and Natur.
Resources Conf., 1969, v. 34.
Recher II. F. Bird species diversity: a review of the relation between species num-
ber and environment.— Quart. Ecol., 1971, v. 6.
Sawicha-Kapusta K. Annual fat cycle of field mice Apodemus flavicollis (Mel-
chior, 1834).—Act. itheriol.,. 1968,, v. 13, N 19, p. 329—339.;
Slobodkin L. B., Sanders H., L. On the contribution of environmental predictabi-
lity-to species diversity-Brockhavenj—In; Sympos. Biol. V. 22. N. Y.,
i ; 1-969. j । J , . ; \ .
Stelibins L.i L. Seasonal and latitudinal valuations in circadian!rhythms of red-
! backrd vole.— Arctic, 1972, v. 25, Ni3, p. 216—224., I
StrakaE., Gerasimov ,S. Correlations between some climatic factors and the abun-
dance of Microtus\ arvalis in Bulgaria.— Ann. Zool. fenn., 1971, v. 8, N 1,
' p. 113—116. i ' ; i ' . '
1 Tansley A. G. Succession: the concept and its values.— Proc. Intern. Congr. Plant.
1 i . । Sci., 1929, v.' 1. i i s i $.. • I
Tast J., Kalela ,0. Comparisons^ between rodent icycles and plant production in
. Finnish Lapland.— Suomalais. tiedeakat. toiinituks, 1971, Ser. A, IV,
N 186,—,14 p. i ।
। Vestal A. G. Anlassociational study of Illinois sand prairie.— Bull. ill. stale Lab.
i' j Nat. Hist,; 1913, v. 10, p., 1—96. , ; । i
Ward J. M,, Armitage К. B. Circannual rhythms'of food consumption, body mass
and metabolism ini yellow-bellied marmots. — Comp. Biochem. and Phy-
siol., 1981, A69, N 4,. p. 621—626;
Walker; J. M., Haskell E.-H.,; Berger R. J., Heller H. C. Hibernation and ciicannu-
al rhythms of, sleep.— Physiol. Zool.j, 1980, v.i53, N 1, p. 8—1,1. ।
; Whittaker R. H., Evolution of diversity in plant communities,— Broakhaven
........Sympos; Piol. '1969, v. 22, pj 81—85. I i
Whittaker R. H. Evolution and measurement of species diversity.— Taxon, 1972,
v. 21, N 2-3, p. 213-251. ,
Wiger R. Role of self regulatory mechanisms in cyclic population of Clethrionomys
i , with special reference to Cl.i glareolus: hypothesis.— Oikos, 1982, vi. 38,
; N 1, p. 60—71. I I ' ’ '
'i ' 'l ' ' i i, ’ - ' - '
‘ J' - /' 4 ' ;
- - '
. ’ , I ! I!
!И > 1 ; i ' ! 'd । ! 1 - - ' ' I
.i ,f . 3 , hi о I - - li
li
Введение 1
ОГЛАВЛЕНИЕ
।
Глава I? Методы анализа цикличности в популяциях мелких млекопи-
тающих и объем использованного материала . . . 10
I Оценка периодичности с помощью спектрального анализа . . —
1 .. Объем и распределение использованного материала............ 13
Глава II. Популяционная цикличность и ее территориальные различия 21
Циклы длиной до года в популяциях насекомоядных и гры-
зунов . . . . .................. . ।.............. 23
Многолетние цйклы в популяциях мелких млекопитающих 4(1
Территориальные особенности цикличности................. 50
Глава III. Цикличность в биотопическпх группировках мелких млекопи-
тающих ....................... . ........ ......................... 55
Особенности циклов в населении мелких млекопитающл|х ос-
новных биотопов Барабы ................................ —
Соответствие цпктов биотопической группировки и доминп-
» рующего в ней вида............................... 100
Глава IV. Влияние (природной цикличности на видовое разнообразие и
, ритмы численности мелких млекопитающих . .,.......... 105
Воздействие многолетней цикличности среды на видовое раз-
нообразие мелких млекопитающих (на примере полевок) —
Цикличность изменения состава и численности населения мел-
ких млекопитающих в многолетней (сукцессии экосистемы
Барабы............................................. 114
- I ; ,! |
Глава V. Цикличность природного фона вспышек численности водяной
। крысы и перестроек видовой структуры мелких млекопитающих 121
Характеристика циклической изменчивости среды . —
' ! I ; ;
Перестройка видовой структуры сообществ в связи со сменой
фаз увлажненности в Барабе в 11-летнпх циклах 137
Влияние вспышек массового размножения грызунов на видо-
вую структуру сообществ мелких млекопитающих 150
I
Глава VI. Цикличность продуктивности сообщества мелких млекопитаю-
щих .............................................................. 188
Глава VII. Концепция экологических сукцессий как циклического биоло-
гического процесса ... 197
Существующие представления об экологических сукцессиях —
Концепция циклических сукцессий.........:................ 201
Дискуссия........................................... . 207
Заключение ......................... . I . . ? ................ 223
Литература..................................................... 227
Анатолий Александрович Максимов
Лев Николаевич Ердаков
ЦИКЛИЧЕСКИЕ ПРОЦЕССЫ
В СООБЩЕСТВАХ ЖИВОТНЫХ
биоритмы, сукцессии
Утверждейо к печати 1
Биологическим институтом
СО АН CCGP
Редактор издательства В. В. Волковинцер
Художественный редактор И. В. Краснокутская
Технический редактор Г. Я. Герасимчук
Корректор С. В. Блинова
ИБ № 23674
Сдано в набор 04.01.85. Подписано в печать 03,10.85. МН-01588. Формат 60 X OO’/ie-
Бумага типографская № 3. Обыкновенная гарнитура. Высокая печать. Усл печ. л. 15.
Усл. кр.-отт. 14,5. Уч.-изд. л. 18.9. Заказ № 537. Тираж 1400 экз. Цени 3 р. 20 к.
Ордена Трудового Красного Знамени издательство «Наука», Сибирское отделение.
630099, Новосибирск, 99, Советская, 18.
4-я типография издательства «Наука». 630077, Новосибирск, 77, Станиславского, 25.