






































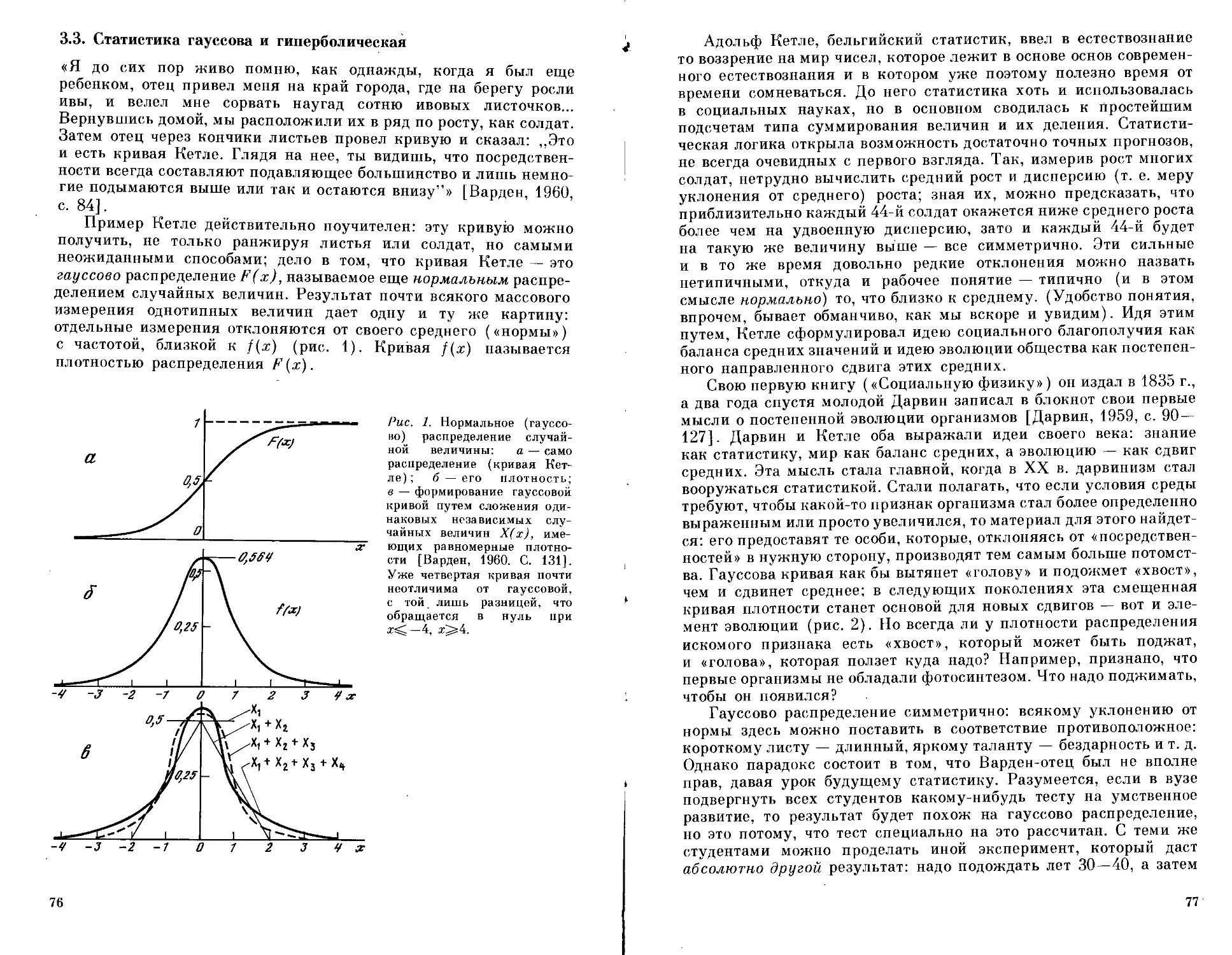
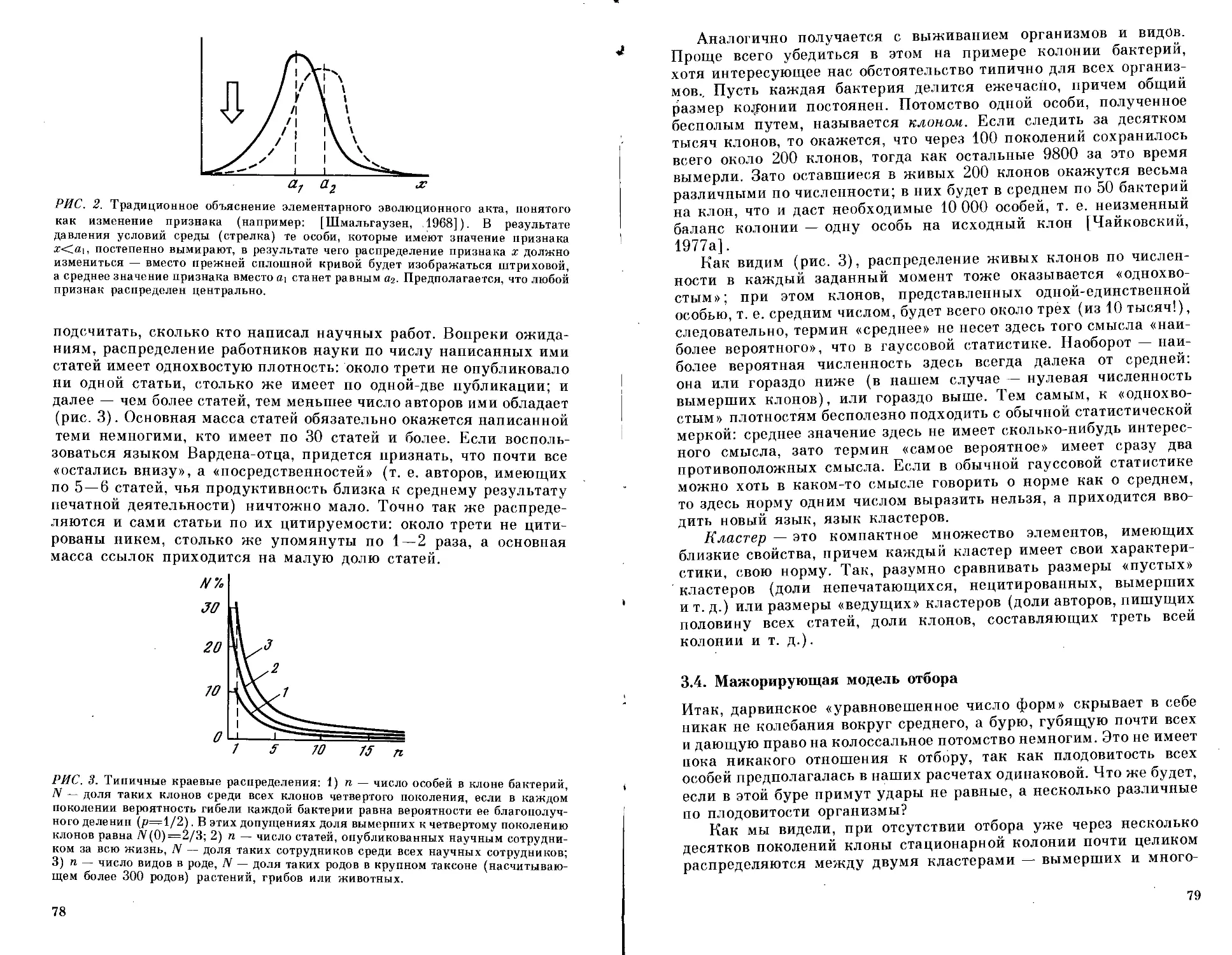





































































































Текст
Ю.В. Чайковский
Элементы
ЭВОЛЮЦИОННОЙ
ДИАТРОПИКИ
Академия наук СССР
Институт истории естествознания и техники
Ю.В. Чайковский
Элементы
ЭВОЛЮЦИОННОЙ
ДИАТРОПИКИ
Ответственный редактор
доктор философских наук
И. А. АКЧУРИН
6
МОСКВА
«НАУКА»
1990
ББК 15.1
Ч 12
Рецензенты
кандидат биологических наук В. В. БАБКОВ
кандидат философских наук В. А. ШУКОВ
Редактор издательства Е. А. ЖУКОВА
Чайковский Ю. В.
Ч 12 Элементы эволюционной диатропики.— М.: Наука,
1990,- 272 с.
ISBN 5-02-008086-1
С позиций системного подхода С. В. Мейена и Ю. А. Урманцева, а также
на базе историко-методологического анализа проблемы разнообразия форму-
лируются основы диатропики — общего учения о разнообразии в строении
материи на разных уровнях ее организации. Показано, как сформулирован-
ные общие закономерности организации разнообразия могут применяться
при исследовании методологических оснований проблемы научного прог-
ноза — на примере экологического прогноза. Автор доказывает, что проб-
лема экологического кризиса требует для своего решения учета закономер-
ностей эволюционной диатропики.
Для философов, науковедов, методологов, биологов, экологов.
„ 0301040100-164 <OQft 1т
4 ~042(02)—90 25~1990 П КаТЯЛОГ
ISBN 5-02-008086-1
© Издательство «Наука», 1990
ПРЕДИСЛОВИЕ
Диатропика (от греч. бштоолос — разнообразный, разнохарак-
терный) — наука о разнообразии, т. е. о тех общих свойствах сход-
ства и различия, которые обнаруживаются в больших совокупно-
стях объектов. О разнообразии писали многие философы начи-
ная с древности, в конце XVII в. Г.-В. Лейбниц включил это по-
нятие в свою философскую систему в качестве одного из основ-
ных, но общего учения о разнообразии не существовало до самого
недавнего времени — до последних работ Сергея Викторовича
Мейена (1935 — 1987), блестящего палеоботаника и геолога, эво-
люциониста и методолога, трагически ушедшего из жизни в самом
расцвете сил. Именно Мейену принадлежит мысль интегрировать
то общее, что до него уже было найдено в конкретных разнообра-
зиях (химических элементов, геологических толщ, биологиче-
ских видов, языков и т. д.) и что он называл рефренной структу-
рой разнообразий.
Данная книга посвящена анализу основ и становления диатро-
пики, но не во всех ее аспектах (что в одной книге вряд ли воз-
можно) , а в основном, в эволюционном. Занимаясь четверть века
проблемой развития, прежде всего биологического, я постепенно
пришел к убеждению, что эволюция прежде всего — процесс пре-
образования разнообразия. Это относится, например, к биологи-
ческой и языковой эволюции. Самодовлеющий характер феномена
разнообразия стал понятен мне в ходе бесед с Мейеном в 1976 —
1978 гг., Мейен поразил меня знанием истории науки — не оби-
лием известных ему имен, дат и фактов, а пониманием того, что все
наши концепции находятся в становлении и являются звеньями,
соединяющими науки прошлого и будущего. Эта позиция ра-
дикально отличалась от господствовавшей тогда (и едва поко-
лебленной сейчас) убежденности натуралистов, что они, обладая
новейшими приборами, вот-вот найдут долгожданную окончатель-
ную истину. Мейен преподал нам яркий урок активной методоло-
гии, т. е. методологии, прямо помогающей строить новую теорию.
Насколько мог, я постарался этот урок использовать.
Беседы с Мейеном обратили меня к той мысли, что путь к по-
строению работоспособной теории эволюции (не только биоло-
гической) лежит через историко-методологический анализ проб-
лемы разнообразия. В качестве исходной для современной диа-
тропики мне представляется работа Мейена [Меуеп, 1973], отчасти
кратко пересказанная по-русски [Мейен, 1974]. Ясно проведенная
3
им мысль о наличии у формы и структуры своих закономерностей,
не сводимых к функции, сближает его направление исследований
со структурализмом, который в гуманитарных науках разраба-
тывался тогда уже полвека, а недавно прокламирован и в биоло-
гии группой «биоструктуралистов», которую основал в Осаке зо-
олог Атухиро Сибатани [Sibatani, 1986]. В отношении биологи-
ческого эволюционизма главная задача виделась мне, до бесед с
Мейеном, в выявлении верного соотношения между выводами ос-
новных школ — ламаркизма, дарвинизма и номогенеза; Мейен же
показал, что прежние школы изучали не столько эволюцию (исто-
рический процесс), сколько наличное разнообразие, лишь снабжая
свои умозрительные эволюционные схемы подходящими примера-
ми из палеонтологии. Сам же он строил эволюционные модели
(в геологии и биологии) как механизмы преобразования конкрет-
ных разнообразий в историческом времени, используя весь корпус
известных ему данных по ископаемым организмам (растениям),
а не подбирая примеры для иллюстрации априорных схем.
Феномен разнообразия, как и феномен эволюции, не знает дис-
циплинарных границ, поэтому в книге пришлось коснуться многих
предметов. Это потребовало консультаций с людьми разных про-
фессий и взглядов, которым я выражаю искреннюю благодарность.
Имена их приведены в соответствующих местах книги. Особо хочу
отметить большую пользу от бесед на общенаучные темы с физио-
логами И. А. Аршавским, А. В. Григорьевой и Б. П. Ушаковым,
генетиками В. И. Корогодиным и Л. И. Корочкиным, микробиоло-
гами Г. А. Заварзиным и Н. П. Кирилловой, зоологами Н. И. Аб-
рамсон и А. С. Раутианом, эмбриологами Л. В. Белоусовым и
Е. Ш. Ярмолинской, теоретиками широкого профиля Г. Г. Мален-
ковым и С. В. Чебановым, системологом Б. И. Кудриным, истори-
ками В. В. Бабковым и М. X. Реммель. Однако консультации, из-
бавив от явных конкретных ошибок, не могли гарантировать меня
от других существенных промахов, поскольку наивно полагать,
что всякий станет, отвечая на мой вопрос, вникать во все тонкости
проблематики, занимающей не его, а меня. Поэтому я старался
свести материал наук, с которыми менее знаком, к минимуму и
нигде не подавать свои выводы в качестве общепринятых. Многих
интереснейших вопросов, например вопроса об иммунологическом
разнообразии, я решил пока не касаться вовсе, кроме тех мест,
где без них изложение теряет цельность.
Анализ разнообразия невозможен без математики, но горький
опыт предшественников заставляет отказаться от обычных прие-
мов обращения с нею. Достаточно вспомнить блестящие работы
В. Вольтерры и Р. Фишера по математике популяций (1920-е гг.):
хотя эти математики ясно писали о чисто модельном характере
своих построений, годных лишь в качестве частных аргументов
в пользу тех или иных принципиальных возможностей, однако
биологи восприняли эти работы в основном как главное и надеж-
ное обоснование вполне конкретных биологических концепций.
В результате прекрасное начинание принесло биологии, пожалуй,
4
больше вреда, чем пользы. Поэтому я старался уделять главное
внимание смыслу исходных понятий и конечных результатов, что-
бы читатель мог сам оценить тот круг явлений, к которым могут
быть приложены приводимые мною теоремы и формулы.
Хотя в книге использованы соответствующие выводы многих ав-
торов, но почти все они касаются проблемы разнообразия лишь
косвенно. Большую роль здесь сыграл для меня семинар «Исследо-
вание форм органического разнообразия», работавший в Москов-
ском обществе испытателей природы (МОИП) в 1978—1982 гг.,
где в дискуссиях по моим докладам был впервые несколько про-
яснен статус диатропики. Предлагаемая книга в основном является
результатом 12-летнего труда в этом направлении. Хотя она в зна-
чительной мере основана на моих опубликованных статьях, однако
при ее написании остро встал вопрос о стиле изложения, поскольку
статьи были адресованы разным кругам специалистов, тогда как
книга предполагается доступной читателю как целое. Пришлось
облегчить стиль изложения настолько, насколько это вообще
возможно в научной монографии, заменяя громоздкий специаль-
ный материал всюду, где это возможно, простыми объяснениями
и ссылками на подходящие руководства и обзоры. Ввиду этого спи-
сок литературы (по необходимости краткий) содержит прежде
всего работы, по которым можно найти остальную литературу.
Для краткости опущены многочисленные ссылки на справочную
литературу (за исключением сведений из редких изданий). Недо-
статок места заставил также отказаться от изложения полемиче-
ского материала, который всюду, где это не мешает пониманию
сути дела, заменен ссылками на соответствующую литературу,
прежде всего обзорную.
Как я стараюсь показать в книге, диатропика имеет собствен-
ную исследовательскую программу и ставит целью не конкурен-
цию с другими теориями на ниве объяснения фактов, а подготовку
будущих практических рекомендаций. Создание теории эволю-
ции — дело будущего, возможный результат работы многих лиц, но
уже в этой книге можно кое-что сказать о том, чего стоит ждать
от нее, а чего — нет.
Хочу поблагодарить тех, кто помог мне при подготовке руко-
писи к публикации: мою жену Н. П. Кириллову, методологов
А. А. Печенкина, В. А. Чудинова и В. А. Шукова, а также директо-
ра Института истории естествознания и техники Н. Д. Устинова и
сотрудников В. Г. Арутюнян, Т. М. Гаврилову и В. С. Кухарчука,
много сделавших, чтобы данный труд увидел свет. Наконец, пони-
мая, что книга является эскизом чего-то будущего, буду рад лю-
бым замечаниям — особенно в отношении изложения работ Мейе-
на, которое я вряд ли всюду смог дать вполне адекватно мыслям их
автора.
Часть I. УПОРЯДОЧЕННОСТЬ РАЗНООБРАЗИЯ
Глава 1. ВВЕДЕНИЕ
1.1. Феномен разнообразия
Вряд ли найдется явление, которое было бы так же хорошо из-
вестно и так же плохо понимаемо, как разнообразие. Все мы знаем,
что нет двух одинаковых людей, что «о вкусах не спорят», что хо-
рош тот учитель, который ориентируется не на «среднего ученика»,
а на каждого в отдельности, что нельзя «покупать кота в мешке»,
а следует проверять именно покупаемый экземпляр — знаем, но
редко делаем должные выводы. Наоборот, часто утверждаем неле-
пости. Например, говорим собственным детям (отнюдь не желая их
обманывать): «все другие умеют, почему же ты...», словно не
знаем, что одни умеют, а другие — нет. По-видимому, человек не
может принять решение, не упрощая ситуацию, вот он и стре-
мится обойти разнообразие. Однако обойденные проблемы не
исчезают, а лишь накапливаются, и настает момент, когда не
замечать их больше нельзя.
Под разнообразием того или иного множества будем понимать
совокупность отношений различия и сходства между элементами
этого множества. Множество же, образующее некоторую целост-
ность, называют системой. Отношения между элементами, обес-
печивающие целостность, называют системообразующими, а от-
ношения различия и сходства мы назовем диатропическими *.
Эти два типа отношений во многом пересекаются, но друг друга
не покрывают. Так, множество зеленых предметов не образует
системы, однако о его разнообразии можно говорить содержа-
тельно — например, сопоставить зеленым растениям зеленых на-
секомых и задать вопрос о приспособительности окраски. Осветим
кратко разные взгляды ученых на разнообразие, отсылая читателя
к работам [Мейен и др.. 1977; Чайковский, 1985, 19876, 19886, в,
19896; Система..., 1988] за соответствующими ссылками.
Простейшим способом отразить (а вернее, обойти) разнообразие
всегда было усреднение. Мы привыкли усреднять все, будто нас
окружают не реальные объекты, а средние величины. Часто гово-
рят, например, что «программа рассчитана на среднего ученика»,
хотя лучше бы сказать наоборот: «средний ученик» придуман для
составления программы (которая получается при этом прими-
тивна для одних, непосильна для других, зато почти всем скучна).
Другой способ обойти разнообразие предложен физикой: раз-
* Здесь и далее полужирным шрифтом выделены понятия, вводимые автором.
6
нообразие попросту выносится за рамки анализа как нечто случай-
ное, в лучшем случае второстепенное. Вот как писал об этом один
из ведущих физиков: «Мир'очень сложен, и человеческий разум
явно не в состоянии полностью постичь его. Именно поэтому че-
ловек придумал искусственный прием — в сложной природе мира
винить то, что принято называть случайным,— и таким образом
смог выделить область, которую можно описать с помощью прос-
тых закономерностей. Сложности получили название начальных
условий, а то, что абстрагировано от случайного,— законов при-
роды. Каким бы искусственным ни казалось подобное разбиение,.,
но лежащая в основе такого разбиения абстракция... позволила
создать естественные науки» [Вигнер, 1971. С. 9].
Оба приема сходны тем, что вместо самого разнообразия рас-
сматривают соотнесенную с ним норму. Только в первом случае ее
понимают как среднее, а во втором — как общее. Естественно, что
в обоих случаях индивидуальное отклонение от нормы рассмат-
ривается как случайное и тем самым обычно как маловажное.
О норме речь пойдет в главе 5.
Третий прием, наоборот, выводит случайность на первый план
и состоит в утверждении, что разнообразие в основном хаотично и
о нем нечего сказать, если не описывать по очереди свойства каж-
дого элемента множества. Так, в 1968 г. генетик Ф. Леритье увидел
в природе «довольно-таки хаотическое обилие видов животных и
растений». В главе 2 мы покажем, что это расхожее мнение просто
неверно, а сейчас напомним ту мысль Вильгельма Оствальда, хими-
ка и натурфилософа, что «разнообразие представляется нам бес-
порядочным» лишь постольку, поскольку нам «нет дела др дей-
ствовавших здесь законов» [Оствальд, 1903. С. 72].
Четвертый прием, казалось бы, противоложен третьему в том
отношении, что приписывает разнообразию полную упорядочен-
ность,— каждый элемент занимает определенное место в системе.
Таким предстает разнообразие химических элементов согласно
периодическому закону, и таким же декларировано разнообразие
организмов в адаптационных учениях — ламаркизме, жоффруизме
и особенно в дарвинизме. Эти учения утверждают, что каждое
наследуемое свойство организма является приспособлением к опре-
деленным качествам среды; но упорядоченность при таком взгляде
на организмы ничем не отличаются от неупорядоченности по Ле-
ритье, так как место каждого элемента в системе приходится
задавать перечислением, а не общим правилом.
Согласно пятому взгляду, свойства разнообразия определяются
тем, что они необходимы для существования своего множества.
В космологии такой взгляд именуется антропным принципом
[см.: Балашов, 1988], а в биологии его разделяют дарвинисты.
Ошибка здесь в том, что наблюдаемый феномен просто отождест-
вляется с его причиной.
В предлагаемой книге развивается совсем иной взгляд, а имен-
но — что разнообразие имеет свои собственные законы, достаточно
общие и существенные, но не формальные и не строго однознач-
7
ные. Разнообразие — не хаос, но и не космос: несмотря на общую
упорядоченность, фундаментальную роль в нем обычно играет
случайность. Она — как бы цемент, скрепляющий здание, как
материальных объектов, так и представлений о них (см. главы 3,
4). Эти общие положения иллюстрируются в книге в основном
биологическими примерами.
Вся наша жизнь требует разнообразия. Если бы оно вдруг исчез-
ло, жизнь потеряла бы все ориентиры, а, может быть, стала
бы вообще невозможной. Представьте себе, чем бы были без разно-
образия такие занятия, как, например, селекция новых пород,
искусство, спорт или суд? Кому кого судить, когда все одинаковы?
Но феномен разнообразия всеобщ, и, как указывает системолог
Ю. А. Урманцев, между любыми двумя системами существует как
некоторое сходство (именуемое в теории систем изоморфизмом),
так и некоторое различие — полиморфизм (если только системы
неквантовые). Есть общие свойства разнообразий, не зависящие
от природы элементов, образующих данные множества. Прежде
всего это — параллелизм, т. е. тенденция к сходству, обнаружи-
ваемая между рядами объектов.
Подробно понятием «ряд», исходным для диатропики, мы зай-
мемся в 2.4, а сейчас отметим, что диатропика оперирует им так же,
как опытные и наблюдательные науки — понятием факта. И так же
как факт не имеет смысла вне объясняющей схемы, так и ряд бес-
смыслен для диатропики без сопоставления с другим рядом, зато
ряды для сопоставления могут быть взяты из самых далеких обла-
стей знания. Диатропика оперирует не отдельными фактами, а их
рядами, сопоставление которых — ее элементарная операция.
Само слово «ряд» неудачно, поскольку ассоциируется с построе-
нием объектов в одну линию, каковой может и не быть, но слово
уже прижилось (в основном благодаря «гомологическим рядам»
Н. И. Вавилова), и менять его поздно. Заметим, что ряд может не
обладать никакой целостностью и потому не быть системой.
Он — элементарная упорядоченность (организованность) изу-
чаемого множества.
Образно говоря, диатропика занимается теми различиями меж-
ду сходным и теми сходствами между различным, на которые до
сих пор обращали мало внимания. Поэтому тот тип рядов, который
наиболее очевиден и известен как периодичность свойств, нас будет
интересовать лишь в порядке сравнения. (Кроме периодической
системы элементов существуют периодические таблицы элементар-
ных частиц, метаболических путей и т. п.). Нас в большей мере бу-
дут интересовать параллелизмы, но не настолько полные, чтобы
создавать периодические таблицы,— именно на таких параллелиз-
мах строятся многие науки, в частности биологическая система-
тика («... Имеется проникающая всю систематику повторяемость
сходных форм» [Любищев, 1968, с. 24]), хотя роль параллелизмов
не всегда осознается явным образом. В качестве других примеров
параллелизмов укажем на типологическое сходство генетически
далеких языков (этот пример подсказал мне Г. Г. Маленков);
8
описание одним и тем же математическим аппаратом феноменов
разной природы (например, механические, электрические и кон-
центрационные колебания); «сравнительный метод» — познание
эволюции какого-либо класса объектов с помощью анализа налич-
ного разнообразия объектов этого класса. Уже из этих примеров
видно, что должны существовать некоторые общие закономер-
ности организации разнообразий. Каковы они?
1.2. Проблема разнообразия. Познавательные модели
О разнообразии думали всегда, и от древности до нас дошли соот-
ветствующие афоризмы: «о вкусах не спорят», «каждому свое»,
«мы судим о себе по помыслам, а о других по действиям», «разно-
образие радует» (varietas delectat). Дошли и важные обобщения.
Так, с Востока в Грецию пришла идея видеть во всяком разнообра-
зии комбинацию небольшого числа первичных сущностей («сти-
хий»). Под влиянием Востока находился и Пифагор, около 530 г.
до н. э. выступивший с мистико-числовым пониманием природы.
Основу миропорядка у него составила гармония чисел: Единица
(монада) символизировала общность и целостность, Двоица
(диада) — расчленимость и изменчивость, Троица (триада) —
все имеющее начало, середину и конец, Четверица (священная
тетрактида) — гармонию, как музыкальную, так и космическую.
(Анализ см. [Гайденко, 1980, гл. 1]). Пифагорейцы, а за ними
и другие, признавали 4 стихии — огонь, воздух, воду и землю, трак-
туя их через пару оппозиций, например: теплое — холодное,
легкое — тяжелое. Эмпедокл (V в. до н. э.), хотя и считал эту чет-
верку основой «всех вещей», рассматривал еще и другую оппози-
цию: любовь — ненависть. Такая дихотомичность познания стала
обычной, и Аристотель в «Метафизике» приводил, ссылаясь на
пифагорейцев, уже 10 оппозиций, служивших для ориентации
в конкретных разнообразиях.
У Гиппократа (ок. 400 г. до н. э.) темпераменты и здоровье
людей рассматривались как следствие соотношения «четырех
соков». Это воззрение продержалось 2 тыс. лет и повлияло на вы-
работку представлений о гормонах. В Гиппократовом сборнике
сходство и различие между родителями и детьми приписано сме-
шению мужского и женского «семени» в разных пропорциях, при-
чем «семя» образуется путем сбора частиц ото всех органов тела.
Натурфилософ и знаток античности В. П. Карпов, видя здесь сход-
ство с генетикой (частицы — гены, смешение — рекомбинация),
восклицал даже: «Факты гибнут, теории остаются» [Гиппократ,
1936, с. 257]. В справедливости этого парадокса мы не раз убе-
димся, только надо уточнить, что, согласно нынешней термино-
логии [Холтон, 1981], сохраняются не сами теории, а их темы.
Такова, например, тема рождения разнообразия из однообразия,
одинаково трактуемая и в древнейших космогониях (разделение
первородного хаоса на основные стихии), и в новейшей теории
Большого взрыва (разделение вещества и энергии и появление
9
различных элементарных частиц по мере охлаждения Вселенной,
вначале однородной [Девис, 1985]).
Кроме идеи попарных оппозиций, в древности при познании
разнообразия использовалась и идея ряда. Так, в восточных систе-
мах нередко присутствовал пятый первоэлемент — эфир, отчего
вся пятерка выступала как ряд. Платон (начало IV в. до н. э.) тоже
признавал пять первоэлементов (по числу правильных много-
гранников), и именно он дал им название «стихии» (от греческого
стойхос — ряд). Латинское слово «элемент» тоже означает ряд —
его считают производным от ряда букв I, т, п. Кстати, аналогия
с буквами проводилась в древности далеко: разнообразие свойств
материи некоторые мыслители объясняли сочетанием элементов
так же, как разнообразие слов — сочетанием букв. Мы тоже будем
пользоваться подобными аналогиями, поскольку языковые при-
меры иногда проще биологических, а законы разнообразия доста-
точно общи.
Понимание разнообразия через комбинацию (членов ряда или
членов оппозиции) имело далекие последствия. Оно дало импульс
к развитию, с одной стороны, атомизма, с другой — формальной
логики. Анализ оппозиций лег в основу классификаций Нового
времени, а венцом этой традиции явился дарвинизм, противопоста-
вивший свойства, полезные для выживания, всем прочим и декла-
рировавший эволюционный процесс как накопление полезных
свойств.
Познание разнообразия в античности опиралось не только на
комбинаторику, но и на метод группировки сходных объектов.
Аристотель, к примеру, собрал в группы известные ему 500 видов
животных, основав таким образом систематику. Он же, выявив
то общее, что нашлось в каждой группе, основал морфологию (хотя
сам этот термин придумал только И.-В. Гете).
Грубо говоря, систематикой называют науку о различиях орга-
низмов, а морфологией — о сходствах, тогда как диатропика долж-
на включать их синтез (см. главу 2). Более двух тысяч лет биоло-
гия была по преимуществу наукой о сходствах и различиях орга-
низмов, но последние сто лет систематику и морфологию считают
как бы второсортным знанием. Вместо пары сходство—различие
стала доминировать пара структура —функция, и интерес к раз-
нообразию надолго угас. Лишь сейчас он вновь пробуждается, по-
скольку угрожающе убывает разнообразие организмов, а в крити-
ческие моменты всегда полезно вспомнить, что же о данной теме
говорили в прошлом.
Еще у орфиков * Зевс, творя мир, восклицал: «Как мне сделать,
чтобы все вещи были едины и раздельны?» Платон полагал, разви-
вая идеи орфиков и пифагорейцев, что все виды произошли от лю-
дей путем деградации, так что от самых достойных произошли
другие люди, от менее достойных — звери и так далее, вплоть до
растений. Эта мысль, изложенная им в диалоге «Тимей», носила
этический характер и в таком смысле логична, как логично и
* Орфизм — религиозное течение в Греции, возникшее в VI в. до н. э.
10
построение Аристотеля, фактически расположившего животных в
ряд от зверей до губок. Однако только идеи линейного ряда недоста-
точно для упорядочения разнообразия, и у Платона мы видим еще
идею рода. Род (genns) означал у него группу объектов, обладаю-
щих некоторой общей сущностью, и определить род объекта зна-
чило познать его сущность (например, для Аристотеля все птицы
были одним родом). Но сами роды тоже нуждались в упорядочении,
и здесь мы находим у Платона идею параллели между микрокосмом
и макрокосмом (строением организма и строением природы).
Бог создал, согласно Платону, мир по образу такого живого сущест-
ва, «которое объемлет все остальное живое по особям и родам как
свои части» (см.: [Эгертон, 1978]).
Здесь воображаемое существо объединяет в себе роды, т. е.
реализует высшую сущность. Гораздо прозаичнее аналогичное
объединение сделано у Аристотеля: низшие роды объединены в
высший род, например ящеры, змеи, черепахи и крокодилы (четы-
ре низших рода) объединены у него в род, ныне именуемый реп-
тилиями [Аристотель, 1937, с. 213]. Объединение групп в группы
более высокого ранга ныне обозначается термином иерархия
(см. 1.3).
Итак, мы имеем три типа упорядочения — линейный, парал-
лельный, иерархический. Этой тройки достаточно, чтобы сформу-
лировать проблему разнообразия', в чем причина сходств'и разли-
чий между объектами и как их естественно упорядочивать?
Для нынешнего биолога обычен такой ответ: разнообразие орга-
низмов в точности соответствует разнообразию их экологических
ниш, т. е. условий существования. Генетик А. Мюнтцинг в 1961 г.
писал: «Если бы среда повсюду была совершенно одинаковой, то
Земля, вероятно, была бы населена одним-единственным видом
живых организмов». Хочется спросить беспечного автора: чем бы
этот вид питался и кто бы разлагал его отходы?
Еще удивительнее, что уверенность в экологической обуслов-
ленности разнообразия мирно уживается в систематике с почти
полным отрицанием всякой экологии при конкретном упорядоче-
нии организмов: принято считать, что «естественная система»
должна отражать историю происхождения организмов, и только.
Разумеется, наука систематики состоит при этом, в основном,
из нескончаемых споров и перегруппировок.
Идея «единого и раздельного», как и идея макрокосма-
микрокосма, развивалась и в эпоху Возрождения. Джордано Бруно
в 1584 г. писал: «...Всякая возможность и действительность, кото-
рая в начале как бы свернута, объединена и едина, в других ве-
1цах развернута, рассеяна и умножена», а «вселенная есть все то,
чем она может быть, сообразно развернутому, рассеянному и разли-
ченному способу» [Бруно, 1949, с. 242 — 243]. Через 125 лет Лейб-
ниц развил Пифагора. Платона и Бруно: «...Каждая монада не-
обходимо должна быть отлична от другой. Ибо никогда не бывает
в природе двух существ, которые были бы совершенно одно как
другое», но «это многоразличие должно обнимать многое в еди-
11
ном и простом». Поэтому «всякую часть материи можно предста-
вить наподобие сада, полного растений, и пруда, полного рыб. Но
каждая ветвь растения, каждый член животного, каждая капля
его соков есть опять такой же сад и пруд» [Лейбниц, 1982, с. 414,
425]. Здесть мы подошли к нашему главному пункту, к диатропи-
ческой познавательной модели.
Понятие познавательной модели введено А. П. Огурцовым
в 1980 г. для обозначения приема организации и истолкования
конкретного материала учеными разных специальностей, но сход-
ного миропонимания (подробнее см. [Чайковский, 19886]). Для
науки Нового времени наиболее характерны пять таких моделей:
1) схоластическая (трактующая природу как текст, как шифр),
2) механическая (природа как машина, как часы), 3) статисти-
ческая (природа как баланс средних величин), 4) системная
(природа как организм) и 5) диатропическая (природа как сад,
как ярмарка). В разные эпохи господствуют разные познаватель-
ные модели, но все они присутствуют в науке почти всегда.
Схоластическая модель господствовала в средние века и в эпоху
Возрождения, одним из последних ее защищал в XVII в. Галилей,
выразив свой пифагорейский идеал тезисом о «двух священных
книгах», одна из которых — Откровение, а другая — книга при-
роды (см.: [Ахутин, 1988]). Уйдя на периферию науки, модель
вернулась, вновь оказалась господствующей в одной из современ-
ных дисциплин — в генетике, и физик Джон Бернал в 1967 г. пи-
сал: «Жизнь перестает быть таинством и практически становится
чем-то вроде головоломки, некоей тайнописью, шифр которой
можно разгадать» (анализ см.: [Мейен и др., 1977]).
Понимание мира как механизма пришло в XVII в. с Декартом
и Ньютоном и завершилось в мировоззрении, именуемом «Лапла-
сов детерминизм». Однако хотя в начале XIX в. современники
Лапласа и полагали доказанным, что мир есть механизм, в это
время, как мы теперь знаем, мир уже завоевывала другая мо-
дель — статистическая. Родилась она еще в XV в., вместе с идеей
бухгалтерского баланса, но долгое время лишь обслуживала меха-
ническую модель, породив идею законов сохранения в физике,
торгового баланса — в экономике, баланса природы — в биологии,
равновесия властей — в политологии и т. д. Ее самодовлеющий
характер стал выясняться в середине XIX в., когда баланс стали
явно трактовать как результат игры разнородных случайностей,
когда в 1859 г. одновременно выступили Чарлз Дарвин со стати-
стическим пониманием микроэволюции и Джеймс Клерк Макс-
велл со статистической теорией газов [Чайковский, 1983, 1990],
В начале XX в. говорили уже о «новом статистическом мировоз-
зрении», и в качестве курьеза можно напомнить попытку тракто-
вать даже принцип наименьшего действия в механике как ста-
тистическую процедуру отбора данного движения среди малых
вибраций [Богданов, 1913, с. 56].
В настоящее время сложилось понимание того факта, что
принцип наименьшего действия — феномен системный (см.:
12
[Чайковский, 19886]). Понимание общности системных феноме-
нов, идущее от А. А. Богданова [1913], привело к возрождению
прежних трактовок мира как организма (например, у Дж. Лавлока
и Э. Янча). Это понимание становится популярным в связи
с глобальноэкологической ориентацией нынешней науки (см.:
[О современном статусе..., 1986]). Можно уверенно сказать, что
наука переориентируется с механико-статистического понимания
мира на системное.
Что касается пятой, диатропической, модели, то она еще не
осознана научным сообществом, хотя отдельных высказываний,
подобных высказыванию Лейбница о саде и пруде, в литературе
встречается немало. Поскольку прокламированный Лейбницем
поиск «единого во многом» до сих пор не реализован, поскольку
до сих пор «познание жизни ,,спотыкается” о противоречие еди-
ного и многообразного» [Карпинская, 1984, с. 80], поскольку со-
кращение разнообразия начинает осознаваться как трагедия,—
постольку за пятой моделью мне видится большое будущее. Если
Галилей и Кеплер видели мир как книгу, Ньютон и Лаплас — как
часы, Дарвин и Максвелл — как баланс случайностей, а Вернад-
ский и Янч — как организм, то ученые XXI в., возможно, захотят
представить его себе как сад, вслед за Лейбницем, который опери-
ровал свободно едва ли не всеми пятью моделями. Именно во
взаимосвязи моделей, а не в замене одной на другую, видится мне
понимание природы.
Наоборот, во всякой смене моделей содержится определенный
просчет.
Поскольку каждая хронологически следующая модель не
только вносит в мировоззрение ученых нечто новое, но и отри-
цает что-то, признанное моделью, господствовавшей до нее, то она
неизбежно выступает в качестве отрицания отрицания каких-то
положений той модели, которая господствовала прежде, чем сей-
час отрицаемая модель. Поэтому при смене моделей происходит
возвращение (на более высоком научном основании) в науку не-
которых тем, казалось бы навсегда из нее ушедших (диалекти-
ческая спираль познания).
Третья модель, будучи отрицанием отрицания первой, за-
имствовала у нее расчленяющий характер познания, основанный
на символах (буквах, словах, признаках); а четвертая, явившись
отрицанием отрицания второй, заимствует у нее функционализм:
пытается всякий объект понять утилитарно — как необходимый
для работы чего-то целого. Это — шаг вперед по сравнению с преж-
ним делением объектов на полезные и вредные, но все-таки поиск
утилитарного понимания вряд ли всюду оправдан. Мы теперь со-
гласны признать, например, что мухи не только досаждают нам,
но и нужны природе; даже — что нужны ей разнообразные мухи.
Однако мы ничего не можем сказать о том, зачем мухи столь разно-
образны, зачем одних только дрозофил — свыше тысячи видов.
По-видимому, разнообразие нельзя понять целиком, если быть
в плену прежних моделей, способных лишь фиксировать проблему
13
разнообразия, но не решать ее. Диатропическая познавательная
модель — отрицание отрицания статистической; обеим свойствен-
но внимание к рядам, демонстрирующим какие-то однородности.
Только если статистическая модель видела всюду баланс и усред-
нение, то диатропической ближе сопоставление и обобщение, т. е.
выявление свойств, общих элементам множества. Единство разно-
образия может быть не только утилитарным, но и, например,
эстетическим. Для обоснования этого надо вернуться назад, к пер-
вой модели.
Первым четырем моделям приблизительно соответствуют че-
тыре типа причин, перечисленные у Аристотеля *, поскольку со-
ответствие природы тексту устанавливается формально, поскольку
толчок предшествует движению, а баланс сил сопутствует ему
и поскольку всякая система мыслится как целесообразная. На-
оборот, пятая модель стоит особняком, ибо в ней преобладают со-
поставления и заключения по аналогии. Здесь следует вспомнить
пятый тип причины, который ввел философ и богослов Франсиско
Суарес [Suarez, 1600, с. 616], основатель «второй схоластики»,—
уподобляющую причину, causa exemplaris (лат. exemplaris — по-
добный, служащий образцом). Этот тип у Суареса отличен от
формальной причины тем, что предполагает сознательное уподоб-
ление. Он был забыт, когда заглох интерес к схоластическим по-
строениям вообще и к построениям «по образу и подобию» в осо-
бенности, но без него аналогия как побудительная причина выпа-
дает из схемы причинно-следственных объяснений. Запишем,
в какой модели какая причина преобладает:
в схоластической — формальная (cansa formalis)
в механической — действующая (cansa efficiens)
в статистической — материальная (causa nialeiialis) **
в системной — конечная (causa finalis)
в диатропической — уподобляющая (causa exemplaris).
Теперь легко сформулировать связь первой и пятой моделей:
диатропическое обобщение ближе к формальному соответствию
схоластов, чем к материальному уравновешиванию или усредне-
нию статистиков (второй виток спирали познания). Зато третья
и пятая близки в том плане, что в обеих существенно выявление
тенденций.
* Когда, например, биллиардный шар катится в лузу, то формальной причиной
движения являются законы механики, действующей — удар кия, материаль-
ной — плоскость стола, а конечной — замысел игрока. Действующая причина
предшествует своему следствию, конечная проявляется в результате действия,
хотя существует до него, а остальные причины существуют во времени парал-
лельно со своими следствиями [Любищев, 1982, с. 151].
** Точнее, эта причина преобладает в описательной статистической модели, где
модельными объектами служат весы и бухгалтерский баланс. В более развитой
математической статистической модели (где модельный объект — игральные ко-
сти) приходится, кроме причинной связи, говорить еще и о случайной связи,
которую прежде иногда именовали случайностной причиной - causa occasionalis.
О ней см. у Лейбница [1982].
14
В рамках проблемы разнообразия организмов можно спросить
о причине несоответствия разнообразя видов и разнообразия усло-
вий жизни. Почему «простая функция может выполняться чрезвы-
чайно разнообразными органами, а на один и тот же тип органов
могут быть возложены самые разнообразные функции»? [Мейен
и др., 1977, с. 119]. В сущности, с довольно близкого вопроса на-
чал эволюционные размышления молодой Дарвин. В колледже он
воспринял тогдашнее естественнобогословское решение проблемы
разнообразия (каждый вид сотворен для своей среды) как прием-
лемое, но вот, путешествуя, обнаруживал массу противоречий:
вид мог встречаться вдоль тысячи миль побережья, но отсутство-
вать на соседнем острове: соседние виды могли сильно разли-
чаться. Понемногу Дарвин стал объяснять разнообразие видов не
отдельными творениями, а медленной эволюцией и расселением.
При этом статистическая модель (уже достаточно обычная тогда
в описательных науках) побудила его рассматривать организм
как набор признаков. Затем Дарвин «принял молчаливое допуще-
ние. что каждая деталь структуры, за исключением рудиментар-
ных, служит какой-то специальной, хотя еще и невыясненной
цели», в чем ему содействовали «труды Уильяма Пэйли по ес-
тественному богословию» [Малкей, 1983, с. 180]. Желание разло-
жить целостность на признаки привело его и его современников
к пониманию всякой системы как итога баланса противоборству-
ющих сил. Ко дню публикации «Происхождения видов» такое по-
нимание системы было в Англии достаточно популярно. Так, фи-
лософ и сподвижник Дарвина Герберт Спенсер видел мозг как
«усредняющий интересы жизни — физические, интеллектуаль-
ные, моральные, социальные», а парламент — как «усредняющий
интересы различных классов общества...» [Spencer, 1860, с. 118].
Для этого мыслителя «форма каждого вида организмов — резуль-
тат усредненной игры внешних сил, действовавших в течение его
эволюции как вида...» [там же, с. 96].
Позже Спенсер, отметив «недостаточность отбора» (нехватку
движущего фактора в схеме Дарвина), выделил пару тенденций,
общую для всей природы (дифференциацию — переход из одно-
родного состояния в неоднородное и интеграцию — соединение
объектов в системы), и охарактеризовал жизнь как прогрессивное
приспособление внутренних отношений к внешним потребностям
[Спенсер, 1870]. Таким образом, разнообразие у Спенсера —
результат дифференциации, интеграции и приспособления (а не
одного приспособления, как принято в дарвинизме). Это было
хотя и большим шагом вперед, тем не менее не было решением
проблемы: по Спенсеру, развитие возможно лишь к состоянию
равновесия внутреннего и внешнего (анализ см.: [Виндельбанд,
1913, с. 373]) — таков дефект, неизбежный для любой статисти-
ческой модели. Для анализа процессов саморазвития она не
годится.
Если английский философ понимал разнообразие через его
становление, через историю, то Оствальд трактовал его чисто ста-
15
тически, как имманентное свойство материи. Цель Оствальда
была далека от проблемы разнообразия, как мы ее сформулиро-
вали выше,— он даже не дал разнообразию отдельного названия,
воспользовавшись одним и тем же словом Mannigfaltigkeit как
для обозначения разнообразия (свойства множества), так и самого
этого множества (которое русская переводчица Оствальда обо-
значала отдельным термином «многообразие» [Оствальд. 1903]).
Цель Оствальда заключалась в построении энергетического пони-
мания природы, где энергией назывался инвариант любых пре-
образований материи, т. е. строилась чисто статистическая (балан-
совая) модель, что было ошибкой, поскольку разнообразие инва-
риантом не является [Чайковский, 1985, с. 164]. Однако именно
Оствальд понял, что описываемый объект надо сначала отнести
к какому-то типу упорядочения (Ordnungstypus), который он на-
звал рядом (Reihe) [Ostwald, 1902, с. 137], а затем уже проводить
измерения и сравнения.
Если бы наука пошла тогда за Оствальдом, это позволило бы
избежать обычной ошибки one рационалистов и инструмента-
листов — определять объекты чисто функционально и исследовать
их чисто измерительными средствами, не задумываясь об их сход-
ствах и различиях с другими объектами.
Оствальд напомнил, что Аристотель «признавал в каждой вещи
нечто неизменное, названное им субстанцией, и, кроме того, раз-
ного рода переменное, которому подходило имя акциденции»
(т. е. случайного, несущественного), и сделал в связи с этим
очень важное замечание: при образовании общих понятий «надо
выявлять инвариантную часть или сторону, причем восприятие
зависит от того, с какой стороны мы рассматриваем объект»
[там же, с. 146]. Здесь Оствальд близко подошел к понятиям ради-
кала по Вавилову и дополнительности по Бору (см. гл. 2), но ре-
шительного шага не сделал, а наоборот, заявил, что энергия яв-
ляется одновременно и высшей субстанцией, и акциденцией.
Однако постоянное и переменное не следует обозначать единым
термином; то, что может изменяться без изменения энергии, я бы
назвал (именно в терминах Оствальда) разнообразием, только
вместо его двусмысленного Mannigfaltigkeit здесь лучше подошло
бы Verschiedenartigkeit или Verschiedenheit.
Оствальд выразил надежду, что разнообразие явлений жиз-
ни «не содержит ничего не выразимого энергетически» [там же,
с. 317], а затем уверенно заявил, что «чрезвычайно разнообраз-
ные внешние формы организмов могут быть объяснены физико-
химически, как форма кристаллов нашатыря определяется внеш-
ними добавками» [там же, с. 346—347]. В сущности, такая пози-
ция оставалась типичной в течение последующих 80 лет, но разве
разнообразие, например, морозных узоров на стекле (излюбленный
пример А. А. Любищева) определяется добавками? А ведь узоры
тоже допускают классификацию. Разве добавками определяются
цветные узоры на крыльях бабочек, узоры, которые «не вписыва-
ются в систему чешуек и жилок, а ведут себя, как рисунок на набив-
16
ном ситце»? [Мейен и др., 1977, с. 117]. По-моему, лучше бы
сказать, что закономерности окраски и внешней формы организ-
мов — акциденции той субстанции, которая задается эко-физио-
логйческими закономерностями.
В общем, вводя новое фундаментальное понятие, важно соблю-
сти такт и не пытаться подвести под него все — иначе оно утеряет
смысл, как случилось с «энергией» Оствальда, «естественным
отбором» Дарвина и многими другими. Чтобы с разнообразием
не случилось то же, надо помнить, что диатропика — лишь аспект
рассмотрения, правда, рассмотрения всего на свете. Аспект со-
стоит в отказе анализировать отдельные факты (это — задача
других дисциплин), в готовности рассматривать только их ряды.
Прежде чем заняться этим, напомним кратко вехи становле-
ния наук, бравшихся за упорядочение органического разнообразия.
Изложение будет основано (до конца гл. 1) на материале книг
В. В. Лункевича [1960], И. И. Канаева [1963, 1966], Л. Я. Бля-
хера [1976] и А. А. Любищева [1982], и только на материал,
в них отсутствующий, будут даваться ссылки в тексте.
1.3. Систематика.
От первых реестров до Линнея и Жюссье
Средневековые компиляторы видели науку преимущественно в соз-
дании систем, но почти не смотрели на природу. Наоборот, у сред-
невековых правителей сады и зверинцы были в такой же моде,
как и у древних, однако научной классификацией в них не зани-
мались; только в 1490 г. возник первый научный ботанический
сад — при университете в Кёльне. В 1533 г. в печати появились
«Подлинные изображения трав» Отто Брунфельса, лишенные сим-
волизма и фантастики; здесь впервые изображались рядом неко-
торые виды, сходные по строению. А в 1538 г. в «Новом травнике»
Иеронима Бока содержалось уже нечто такое, что позже назвали
попыткой естественной классификации. До Бока умели, вслед за
древними, различать только деревья, кустарники, травы, водные
растения, т. е. так называемые жизненные формы (биоморфы);
они могут не соответствовать сходству в строении растений (так,
кроме водорослей в воде иногда растут и папоротники, и цветко-
вые). Бок, еще не предлагая классификации, уже предпочитал
располагать при описании рядом растения, сходные по строению
цветка и плода, чем фактически впервые выявил некоторые важ-
ные группы (например: губоцветные, крестоцветные и сложно-
цветные), признаваемые естественными и в наши дни. Естествен-
ными — значит обладающими существенным сходством по глав-
ным свойствам. Позже, в XVII в., существенное сходство было
осознано как общий план строения (см. 1.4).
Сходные формы недостаточно только располагать рядом —
надо еще уметь как-то обозначать сходство, и уже у Брунфельса мы
находим такой прием: большой и малый подорожники он именует
Plantago major и Plantago minor. Вряд ли он мог подумать, что
17
это — бинарная номенклатура, т. е. обозначение организма двумя j
терминами: с большой буквы — рода, с малой — вида этого рода.
Впервые сознательно группировал виды (в том числе и зоологиче-
ские) в роды энциклопедист Конрад Геснер в 1550-е годы, но еще
более ста лет не удавалось объединить в родах все известные виды,
так что многие виды описывались по-прежнему по алфавиту
названий.
Роды также нуждались в упорядочении, и здесь наметилось
три главных приема. Геснер располагал все сущее в единый ряд —
от минерала до человека (в XVIII в. — до ангелов, что называлось
«лестницей существ»). Растения он поместил между минералами
и низшими животными. В 1586 г. Андреа Чезальпино (Цезальпин)
упорядочил растения по наличию и сложности цветка и плода.
Это был проблеск морфологического подхода к систематике,
приведшего позже к идее прогресса как усложнения строения
(см. 1.4). «Две идеи связаны с „лестницей", одна — упорядо-
чивающая группы согласно их уровню развития, и другая — !
непрерывность форм природы» (отсутствие резких скачков), ,
причем обе идеи «перешли в двадцатый век» [Stevens, 1984,
с. 174].
Другой прием продолжал линию Бока и состоял в группировке
близких родов в группы, почему-либо казавшиеся исследователю
естественными. В 1623 г. Каспар Баугин расположил растения
по 12 «книгам», которым, однако, не дал названий. Для той же цели
сближения Пьер Маньоль в 1689 г. ввел понятие «семейство» (fa-
milia) . «Эти семейства состояли из групп сходных организмов, для
которых, однако, могло не быть никакого особого диагностического
признака» [там же, с. 171]. Не следует путать семейство по
Маньолю с нынешним (с таксоном, состоящим из родов и вклю-
чаемым в состав отряда).
Формальное объединение родов явилось третьим приемом. ]
Фактически это делал еще Баугин, а в конце XVII в. ботаник
Джон Рэй дал таким объединениям названия: он группировал
роды в «старший род», а те в свою очередь — в «главный род»;
платонов принцип рода как сущности понемногу уступал принципу
иерархии.
Греческим словом «иерархия» («власть священника») в сред-
ние века обозначали тот тип управления, когда прихожане под-
чинены священнику, священники — епископу, епископы — мит-
рополиту, а митрополиты — патриарху (папе). Позже иерар-
хией стали называть последовательное подчинение вообще, причем
иерархический принцип казался единственно возможным спо-
собом упорядочения, став логической основой классификации.
Его и узаконил применительно к познанию природы Карл Линней
в 1735 г. в «Системе природы».
Согласно этому принципу все организмы, «отличающиеся друг
от друга не более, чем дети от родителей», составляют один вид
(иногда — разновидность); каждый вид относится к определен-
ному роду и т. д. В наше время в систематике обязательны следую-
18
щие таксоны: вид, род, семейство, отряд (у ботаников — порядок),
класс, тип (отдел) и царство. Кроме обязательных, часто вводят
дополнительные таксоны от подвида до надцарства.
Роль Линнея для диатропики огромна, но не вполне совпадает
с тем, что обычно о нем пишут. Дело в том, что чаще всего ему при-
писывают три фундаментальных новшества: иерархический прин-
цип классификации, бинарную номенклатуру и отнесение че-
ловека к отряду обезьян. Однако эти положения были уже хорошо
известны до Линнея, который, наоборот, в какой-то мере проти-
вился их введению.
Во-первых, он понимал недостатки иерархического метода
классификации и, признавая его удобным для искусственной
процедуры адресации, хотел видеть естественную систему (под-
робнее см. гл. 6) не в виде древа (иерархии), а в виде сети или
карты, где соседство групп означало бы их существенное сходство
(«сродство»). Эти группы в чем-то соответствовали семействам
Маньоля, и в «Философии ботаники» (1751) он назвал их семей-
ствами.
Во-вторых, бинарную номенклатуру (обозначение организма
через его род и вид) не надо путать с биномиальной номенклатурой
(обозначение его парой слов). Бинарной номенклатурой пользо-
вался еще с 1596 г. Баугин, но как вид, так и род он часто харак-
теризовал несколькими словами. В этом ему следовал и Линней,
считавший пары слов не научными, а бытовыми названиями
(nomina trivialia), каковые дал лишь в одном труде [Linne, 1753],
да и то не в тексте, а на полях (см.: [Бобров, 1958]). Nomina
trivialia заимствовались в основном из литературного латинского
языка, поэтому Линней, писавший по-латыни, не мог рассматри-
вать их как научные термины. Он лишь уточнял их, закрепляя
за определенными родами с помощью диагнозов. Так, слово Bo-
letus означало у римлян и белый гриб, и цесарский гриб [Ains-
worth, 1976], и шампиньон, и просто гриб, а Линней закрепил его
за белым грибом (и остальными грибами этого рода). Иногда он
придумывал родовые названия, например, назвал в честь Маньоля
(Magnol) магнолию. Только когда латынь отжила свой век, no-
mina trivialia Линнея стали научными терминами, биномиальной
номенклатурой.
В-третьих, человека включали еще и до Линнея не только
в один отряд с обезьянами, но иногда и в один род с орангутангом
и шимпанзе [Козо-Полянский, 1937]. Однако в «Системе при-
роды» положение человека от издания к изданию все более обособ-
лялось, так что в 12-ом (последнем, какое он правил сам) описание
просто начато с человека, без всякого разговора о его положении
в системе [Linne, 1766, с. 12]. Вскоре Линней потерял дееспособ-
ность, и мы не знаем, кто выделил в последующих изданиях че-
ловека в особое царство — он или его редактор, но ясно одно:
логика эволюции взглядов Линнея привела именно к идее царства
человека, которая господствовала после этого в науке почти сто лет.
Наконец, Линней вовсе не дал исчерпывающе полной класси-
19
фикации, основы нынешней — достаточно сказать, что пальмы
описаны им вне общей системы [Linne, 1753, т. 2], зато вклю-
чены в особую систему из семи семейств. Шесть из них — естест-
венные (в понимании Маньоля): грибы, водоросли, мхи, папорот-
ники, злаки и пальмы [Linne, 1751, § 78, с. 37], тогда как седьмое
(остальные цветковые) названо просто Plantae (растения) и было
с другими семействами несопоставимо.
Величие Линнея видится в другом. Он первый показал, что
разнообразие действительно можно в основном упорядочить
с помощью единого формального принципа. Дав всем известным
организмам номенклатуру, Линней не только завершил 200-лет-
нюю работу ботаников, не только перенес ее достижения в зооло-
гию, но и достиг заветной цели средневековых энциклопедистов.
Недаром в основе у него лежала средневековая идея всепроникаю-
щей иерархии. Каждый организм получил у Линнея точный
адрес в системе, причем ни один вид не имел двух адресов, и ни по
одному адресу не числилось двух видов (выражаясь нынешним
языком, он избежал как синонимии, так и гомонимии). Никому
ни до, ни после Линнея этого не удавалось, и дело, конечно, не толь-
ко в том, что Линней был великий систематик, айв том, что он на-
меренно сузил свою задачу. Прекрасно понимая, что идеальная
цель систематики — в уяснении взамосвязи строения всех орга-
низмов, т. е. в построении естественной системы, он фактически
построил удобную адресацию, искусственную систему. Линней
выбирал диагностические признаки не формально, а на основе ин-
туитивного восприятия группы как целого, что и выразил своим
знаменитым афоризмом: «Не признаки задают род, но- род за-
дает признаки». Поэтому его система достаточно хорошо отражала
«суть вещей» (анализ см.: [Любищев, 1972]), хотя главной
целью здесь была адресация. После Линнея искусственную систему
(адресацию) перестали отличать от естественной, выражающей
«сродство» групп. Если Линней понимал, что «безошибочное раз-
личение» возможно только путем «самовластной дихотомии»,
дающей «ключ классам» [Linne, 1751, с. 98, § 154], т. е. искусствен-
ной системой, то вскоре стало всеобщим убеждение, что буду-
щая естественная система примет форму древа и сама собой вы-
полнит роль системы адресов.
В 1759 г. королевский садовод Бернар Жюссье отобразил
(как бы развивая традицию Геснера) естественные сходства ра-
стений в форме соседства их на расположенных в одну линию
грядках сада Трианон близ Версаля. Его племянник Антуан
Жюссье нашел эту последовательность похожей на «ряд химиче-
ского сродства», которым пользовались тогда химики, и счел глав-
ной задачей ботаников и зоологов построение таких рядов [Jussieu,
1778, с. 197], тогда как естественную классификацию он в то время
считал недостижимой, подобной философскому камню [тамч же,
с. 178]. Ряд он разделил пополам (на однодольные и двудольные),
а каждую половину — на классы, классы — на порядки и т. д.,
т. е. сопоставил ряд и древо (иерархию). Древо служило здесь лишь
20
для сохранения привычного способа адресации, главной же
была идея сродства (близости в целом), но с тех пор стало приня-
тым называть естественной всякую иерархическую систему, если
она использует идею сродства. После Дарвина и Геккеля сродство
стали трактовать как генетическое родство, но в остальном ни-
чего не изменилось: все трудности формального разбиения были
упрятаны в интуитивную процедуру поиска близости, которую
каждый понимает по-своему, отчего системы то и дело пересмат-
риваются, и конца этому не видно. Непрестанный пересмотр неко-
торые (напр.: [Тахтаджян, 1987]) считают вполне приемле-
мым фактом, другие считают, что надо лишь найти «истинные»
признаки родства, и система будет окончательна. В этом отно-
шении важно напомнить одну работу времен Линнея.
В книге Мишеля Адансона (1764) «семейства» растений по-
строены формально, по совокупности сходств, причем все признаки
сходств приняты равноценными. В XX в. адансонова системати-
ка возродилась, только теперь каждому признаку стали припи-
сывать свой вес, благо, вычислительные средства теперь это по-
зволяют. Сторонники этой «нумерической систематики» уверены,
что строят окончательную истинную систему (напр.: [Смирнов,
1969]), однако их схемы оказались не только эфемерны, но подчас
и противоречили друг другу. А ведь главная трудность была от-
мечена давно [Максимович, 1827, с. 41] и состоит не в сложности
обсчета свойств, а в том, как отразить целостность объектов клас-
сификации, ибо сродство — характеристика целостная. Поэтому
сейчас, когда господствуют идеи системности (о системной позна-
вательной модели см. 1.2) следует мысленно вернуться назад,
к А. Жюссье, объединившему две традиции понимания сродства —
линейную (Платон — Геснер —Цезальпин) и иерархическую
(Аристотель—Рэй—Линней). Сейчас, через 200 лет, этот путь
многим видится исчерпанным, но, по-моему, альтернативу ему
следует искать вовсе не в русле идей Адансона, а скорее на пути
линнеевой «географической карты», что мы покажем в гл. 6.
1.4. Морфология.
От Везалия до Бэра и Геккеля
Сходство между всеми птицами (или между всеми цветковыми)
очевидно и с глубокой древности использовалось практически.
Основные сходства были давно выявлены даже между очень да-
лекими формами. Так, Аристотель провел сравнение между воло-
сами зверей, перьями птиц, щитками пресмыкающихся и чешуей
рыб (чисто морфологический результат), хотя не брался раз-
бивать тех же зверей хотя бы на самые очевидные группы (т. е. мор-
фология обгоняла его систематику). В средние века факт морфоло-
гической общности млекопитающих был в сущности общепризнан
врачами: обучение анатомии человека проводилось вплоть до
XVI в. на тушах домашних животных (обычно свиней). Даже
знаменитая книга «О работе человеческого тела» Андреа Везалия
21
(1543) в значительной мере была основана на данных о живот-
ных — автор смог вскрыть только несколько человеческих тру-
пов, за что едва избежал казни.
Нам не понять тех людей: вскрытие трупа врачом возмущало
их не меньше, чем нас возмущают проводившиеся в старину опыты
с рассечением живого человека; в то же время созерцать внутрен-
ности,. вываливающиеся на эшафот, считалось вполне поучитель-
ным. (Вспоминается гравюра времен Возрождения: на площа-
ди — два столба, к ним за ноги привязан обнаженный атлет, кото-
рого два других атлета распиливают двуручной пилой через пах.
И все мышцы даны в напряжении, с явным и любовным знанием
внешней анатомии.) Впрочем, легче ли будет понять нас нашим
потомкам?
Анатомия Везалия была кинематической: каждый орган рас-
сматривался с точки зрения его работы. Эта установка, ставшая
позже довольно обычной, была очень полезна для нужд медицины,
но препятствовала установлению сходства между органами, ра-
ботающими различно, т. е. в нынешних терминах — установлению
гомологий. Поэтому морфология, т. е. учение о самых общих
структурных сходствах, развивалась медленно. Хотя в XVI в. в ее
рамках делалось довольно много: было описано сходство скеле-
тов человека и птицы, выявлены сходства некоторых групп бес-
позвоночных и т. п., а в XVII в. Аврелий Северино даже писал о
едином «прототипе» всех животных,— но эти идеи мало повлия-
ли на научную мысль в целом, вплоть до начала XIX в., до работ
зоолога Жоржа Кювье.
Главная установка, отличающая морфологов от систематиков,^
поиск «единого во многом» — сформулирована немецким бо-
таником Иоахимом Юнгом (умер в 1657 г.), а в начале XVIII в.
стала известной благодаря философской системе Лейбница. В наше
время понемногу утверждается старый взгляд на природу как
на экологическое единство, но столь же старый взгляд на нее как на
морфологическое единство понимается пока с трудом, поэтому
о нем следует сказать подробнее.
Еще в 1659 г. в Англии натуралист Томас Виллис писал, что
идея единства природы видна в общности строения животных,
например, в наличии жесткого скелета: рак тоже имеет скелет,
но живет внутри него, потому что у него все наоборот, включая
перемещение задом наперед, это — позвоночное наоборот.
В 1744 г. английский анатом и врач Александр Монро писал
в своем «Опыте сравнительной анатомии», что изучение разнооб-
разия животных дает возможность понять человеческое тело.
Так, наличие ключиц у обезьян и белок он связывал с преимущест-
венно вертикальным положением тела. (В этом приеме легко
заметить продолжение кинематической анатомии Везалия.)
Через тридцать лет такой антропоцентрический взгляд на мор-
фологию (и на разнообразие) поддержал другой английский
анатом и врач, Джон Хантер, добавивший, что у низших живот-
ных, у которых органы проще, легче оказывается понять их строе-
22
ние и функции, аналогичные человеческим. Что же касается чи-
стых натуралистов, то мысль о единстве строения была им важна
сама по себе, как указание на какой-то фундаментальный закон
природы. Например, Жорж-Луи Бюффон, один из основателей
французской зоологической науки, писал в 1753 г. о едином «прото-
типе»: «...Модель прекрасного как бы рассыпана по всей Земле,
так что в каждом климате пребывает лишь часть ее». Здесь про-
тотип — не простая исходная форма, давшая начало сложнейшим
(как мы привыкли считать, читая руководства по эволюции),
а, наоборот, нечто более сложное и совершенное, нежели любой
конкретный организм; прототип здесь понят как совокупность
принципиальных возможностей. В 1786 г. ту же идею высказал
поэт и натурфилософ Иоганн Вольфганг Гёте, который полагал, что
есть основная форма, «с помощью которой природа как бы всегда
только играет и, играя, вызывает многообразнейшую жизнь». Фи-
лософ Жан-Батист Робинэ в 1766 г. придал идее прототипа мате-
риальный смысл: «Все существа... представляют естественные ва-
риации прототипа... Он их порождает подлинным образом, путем
развития». Это развитие (что позже много раз отмечалось) может
носить сходный характер как в онтогенезе, так и в филогенезе,
к чему мы еще вернемся.
Естественно, вставал вопрос: сколько таких прототипов сущест-
вует? Самые смелые фантазеры видели всего один, даже растение
мысля как обращенное ртом в почву и половыми органами вверх.
Что же касается прототипа всех животных, то его признавал даже
такой серьезный морфолог, как Этьен Жоффруа Сент-Илер, ви-
девший (как бы в развитие идеи Хантера) в панцире насекомого
тот же позвоночник, только вместивший внутрь себя все органы
туловища. Кювье, признавая единство всех позвоночных, утверж-
дал все же, что существуют четыре не сводимых друг к другу типа
животных: позвоночные, членистые (включая насекомых), мягко-
телые (в основном — моллюски) и лучистые (или радиаль-
ные — медузы, морские звезды и т. и.). Когда в 1830 г. выясни-
лось, что школа Жоффруа претендует также на единство плана
позвоночных и мягкотелых, то между Кювье и Жоффруа возник
знаменитый спор.
Расхожее мнение о нем как о споре эволюциониста (Жоффруа)
с противником эволюции (Кювье) неточно. Два великих морфолога
спорили на старую, как мир, тему — о способе упорядочить разно-
образие. Грубо говоря, Кювье защищал Платонову идею неизмен-
ных сущностей, а Жоффруа — аристотелеву идею постепенности
изменений (приведшую к «лестнице существ»). Поэтому оказа-
лись несовместимы их взгляды на разнообразие и на возможности
его изменения: Кювье отрицал возможность постепенного перете-
кания одной формы в другую, тогда как Жоффруа видел его всюду.
Новое и старое причудливо перемешалось в их взглядах: «новатор»
Жоффруа защищал идею о раке, якобы живущем внутри своего
позвоночника, а «консерватор» Кювье предвосхитил Дарвина, по-
ставив во главу угла связь строения организма с условиями су-
23
ществования (в 1928 г. В. И. Вернадский даже говорил об эволю-
ционизме Кювье —Дарвина).
Сколько бы ни видеть типов в природе — один, четыре или
много, — все равно встает вопрос о том, как упорядочить организмы
внутри и вне типа. Многие пытались, вслед за Цезальпином, вы-
страивать виды от простых к сложным, так что возникла проблема
общего описания феномена сложности, объективных критериев
усложнения. Что рыба проще человека — всем интуитивно ясно,
но чем ящерица сложнее лягушки? У лягушки 3 способа дыха-
ния, кожные железы, а у ящерицы есть амнион, т. е. оболочка,
препятствующая высыханию яйца, сложнее устроены сердце, че-
реп и мозг. Для науки нужны общие правила — иначе о каждом
случае будут идти бесконечные споры. Такой критерий и предло-
жили независимо в 1827 г. зоологи Анри Мильн-Эдвардс и Карл
Бэр: более сложны (или, что то же — выше организованы) те, у ко-
го можно указать большее число различных по строению и функци-
ям органов. При этом Мильн-Эдвардс делал акцент (вслед за Гёте)
на «разделение труда между частями тела», а Бэр — на дифферен-
цировку (обособление) органов в ходе онтогенеза (при развитии
яйца во взрослый организм). Бэру в 1828 г. удалось показать, что
четыре типа Кювье являются четырьмя типами онтогенеза,
что само по себе явилось огромным морфологическим дости-
жением.
Критерий Мильн-Эдвардса —Бэра довел в 1853 г. до совершен-
ства Генрих Брони, ботаник, зоолог и палеонтолог, первый критик
и переводчик Дарвина в Германии. Дифференцировку он понимал
не только как разделение работы между органами и усложнение
самих органов (например, четырехкамерное сердце у птиц и зве-
рей) или как концентрацию органа (например, появление голов-
ного мозга вместо разбросанных по телу нервных ганглиев),
но и как появление новых элементов симметрии тела, уменьшение
числа одноименных органов, проникновение их в глубь тела и,
наконец, как усложнение онтогенеза. Он предложил систему
«четырех царств природы» (небесных тел, минералов, растений
и животных), но у него, в отличие от Линнея, она была основана
на «силах», действующих в этих царствах, и на симметрии тел. Он
ввел пятый тип животных — бесформенные, или зоофиты, — в ко-
торый включил губок и одноклеточных; это было некоторой натяж-
кой (далеко не все одноклеточные бесформенны), неизбежной,
когда всему разнообразию навязывается один-единственный
принцип.
Итак, в середине XIX в. появилась возможность серьезно срав-
нивать организмы и органы по степени их развития. Однако вы-
яснилось, что, выстраивая ряд организмов в порядке усложнения
одного органа, мы чаще всего видим, как вдоль этого ряда ка-
кой-то другой орган упрощается или даже исчезает. Так, у пти-
цы — четырехкамерное сердце, зато у ящерицы есть зубы — кто из
них совершеннее? (После Дарвина можно спросить: почему птицы
вытеснили зубастых первоптиц? Разве зубы не нужны в борьбе за
24
существование?) Понятие морфологического прогресса не удава-
лось, кроме отдельных случаев, приложить к эволюции, поэтому
прогресс было долго принято понимать просто как один из аспектов
описания разнообразия органов.
И вот Мильн-Эдвардс прямо сформулировал закон разнообра-
зия: «Разнообразие в проявлениях и экономия в средствах исполне-
ния составляют, по-видимому, главные стремления природы». При
этом он имел в виду разнообразие как «основных типов», так и
«степеней совершенства» в пределах каждого типа и, наконец,
«легких органических видоизменений» в пределах одного уровня
организации.
Завершить представление о «типе организации» удалось зооло-
гу Ричарду Оуэну, который в 1846 г. заменил его понятием «архе-
тип». Для этого ему пришлось уточнить все сказанное до него о
функциональном, структурном и внешнем сходстве с тем, чтобы
разделить понятия «аналогия» (внешнее и функциональное
сходства) и «гомология» (соответствие органов, не исчезающее
«при всевозможных вариациях формы и функции» органа). Так,
рог и бивень аналогичны, а бивень и зуб гомологичны. Тогда
архетипом оказывается система гомологичных органов.
Понятие гомология ввел в биологию Э. Жоффруа, причем в
двух различных смыслах. В 1824 г. он назвал гомологией повторе-
ние органов одной особи, а в 1825 г.— сходство путей индиви-
дуального развития. Он заимствовал понятие у «немецкой филосо-
фии», но уже Оуэн не мог прокомментировать, что оно означало,
и «что-то неопределенное» [Owen, 1855, с. 25] оставалось всегда
в трактовках этого основного для нашей темы понятия [Чайков-
ский, 1987а]. Когда А. Н. Северцов напомнил, что у Оуэна ис-
следование гомологий «носило геометрический характер» [Север-
цов, 1949, с.19], это уже мало кому было понятно. Попробуем крат-
ко объяснить основную идею Оуэна.
Он исходил из «позвоночной теории черепа» Лоренца Окена,
появившейся за 40 лет до него (и развивавшей идею прототипа),
согласно которой череп позвоночного состоит из четырех видоиз-
мененных позвонков. Оуэн сформулировал понятие идеального
позвонка, состоящего из 11 типов элементов, которые на практике
могут представлять собой как фрагменты реального позвонка, так
и отдельные кости. Так, в качестве примера «естественного типич-
ного позвонка» Оуэн приводил костно-хрящевое кольцо, состоящее
из обычного позвонка, пары ребер, фрагмента грудины и соединя-
ющих их хрящевых элементов. Такого же сорта «позвонки» он
видел в черепе, плечевом и тазовом поясах и т. д. Эти «позвонки»
образуют (выражаясь нынешним языком) несколько гомологи-
ческих рядов. Тут-то и начинались сложности.
Проще всего с таким очевидным рядом, который образуется,
например, серией позвонков одного позвоночника,— его Оуэн
обозначил термином серийная гомология. К ней же относится, на-
пример, ряд из трех фаланг одного пальца. Не многим сложнее
то, что Оуэн назвал частной гомологией,— соответствие между
25
однотипными органами животных разных видов: между плавни-
ком, лапой и крылом, между чешуей, шерстью и перьями и т. д.
Здесь, однако, возможны трудные для классификации случаи,
например, гомология четырех костей жаберного аппарата рыб че-
тырем косточкам слухового аппарата наземных позвоночных.
Это соответствие впервые доказал Жоффруа, чем вызвал восторг
столь строгого критика, как Кювье. Сам Жоффруа понимал, что
возможность подобных гомологизаций означает наличие каких-то
общих (мы бы сейчас сказали — системных) отношений в органи-
зации строения тела животных, что и привело его к идее единого
типа всех животных — идее, которую Кювье разгромил. Гораздо
плодотворнее был подход Оуэна, рассуждавшего только о морфоло-
гическом единстве типа позвоночных: он попросту решил перечис-
лить части, из которых состоит этот тип, и составить из них архетип
позвоночных. Таковыми частями были у него «идеальные позвон-
ки», если шла речь об архетипическом скелете.
Только зная архетип можно в полной мере говорить о гомоло-
гии, т. е. о том месте, которое занимает данная часть в системе,—
такова главная мысль Оуэна. С чем, например, сопоставить чью-то
мандибулу (нижнюю челюсть) ? До построения архетипа сопостав-
ление возможно только в рамках частной гомологии, т. е. с другими
мандибулами; поняв же архетип, можно сопоставлять одноимен-
ные элементы «идеальных позвонков», а именно: элемент гема-
пофиз в одном «позвонке» образует часть мандибулярной кости,
в другом — клювовидную кость (коракоид) плечевого пояса,
в третьем — часть реального позвонка, в четвертом — подвздош-
ную кость и т. д. Вот это-то место реальной кости в идеальном,
архетипическом, скелете Оуэн и назвал общей гомологией кости,
полагая, что тем же образом можно упорядочить не только кости,
но и все части тела.
Итак, у Оуэна мы видим три вида гомологии — общую, частную
и серийную. Разумеется, многие случаи было трудно отнести к
определенному ее виду. Так, соответствие жаберных и слуховых
костей вначале воспринималось как частная гомология, но в дей-
ствительности его нельзя установить без представления (хотя бы
интуитивного, как у Жоффруа) об архетипе скелета головы, сле-
довательно, его правильнее назвать общей гомологией. Сам Оуэн
писал, что, кроме простых серий, бывают сложные, общегомологи-
ческие: «Если же части не могут быть гомологами... иначе, чем
в общем смысле» (в смысле соответствия одному архетипу), «как,
например, мандибула, коракоид, подвздошная кость и т. д., то я даю
такой серии частей, относительных или повторяющихся, имя
гомотипов. Затылочная кость гомотипна клиновидной, но с другой
стороны, когда говорят, что затылочная повторяет в своем позвон-
ке... клиновидную или тело теменного позвонка или первого шей-
ного позвонка и последующих, то указывают на серийную гомо-
логию» [Owen,1855, с. 30].
Понять это было нелегко (еще сложнее писал Брони), поэтому
биологи охотно перестали читать подобных авторов, когда появи-
26
лась более простая, хоть и бедная содержанием, морфологическая
схема Эрнста Геккеля, ставившая целью объединить морфологию
с эволюционизмом Ламарка и Дарвина.
«Происхождение видов» Дарвина появилось в 1859 г. и расколо-
ло ряды морфологов. У Дарвина морфологические вопросы затра-
гивались лишь в самом общем виде, и многие морфологи (включая
Бэра, Мильн-Эдвардса, Бронна и Оуэна) заявили, что дарвинский
механизм (накопление мелких изменений) не объясняет процес-
сов, связанных с изменением типа организации. Другие увидели
в архетипе попросту предковую форму, родоначальника таксона, и
на этом основании приняли дарвинизм. Поворотом в судьбах мор-
фологии явилась огромная «Общая морфология» Геккеля, вы-
шедшая в 1866 г.
Следуя идеям симметрии Бронна, Геккель развил целую новую
дисциплину — проморфологию, учение о формах организмов. Это
была блестящая попытка завершить линию Кювье. С другой сторо-
ны, следуя широко понятой идее общего предка, он построил «есте-
ственную систему организмов», в которой выводил все из амеб,—
явное продолжение линии Жоффруа (единства типа). Система Гек-
келя стала образцом на целое столетие, и до сих пор многие видят
цель морфологии в выявлении предковых форм (хотя построенные
при этом конкретные системы организмов неизменно рушатся —
«факты гибнут, теории остаются»). Наоборот, проморфология ока-
залась почти не у дел, и сам Геккель отошел от нее. «Историческая
морфология пожрала конструктивную»,— сказал по этому поводу
Любищев [1982, с. 202]. Через сто лет все поменялось: от родослов-
ного древа Геккеля почти ничего не осталось, зато вновь ожила
проморфология, чтущая его как основателя [там же, с. 209]. Это
и понятно: синкретическое признание взаимоисключающих точек
зрения нежизнеспособно, поэтому научному общественному мне-
нию остается колебаться между альтернативами, пока не будет
найден синтез.
На учение Геккеля живо откликнулся старый Бэр. Допуская
в принципе идею эволюции, он резко возражал против смешения
морфологического сходства с историческим родством. Для него это
смешение - «разрастающийся сорняк в вопросе о трансмутации»,
т. е. об эволюции. Выводить разнообразие из единой предковой
формы Бэр считал ненаучным, в чем, кстати, был близок Дарвину,
который свою схему постепенной эволюции никогда не пытался
приложить к становлению какого-либо архетипа.
1.5. Комбинативность.
От Парацельса до школы Вавилова
Парацельс — алхимик, врач, фантазер, мыслитель и практик, по-
служивший связующим звеном между средневековой и новой нау-
кой, в 1530-е годы размышлял о разнообразии. Он воскресил
гиппократову идею образования зародыша слиянием материнских
и отцовских частиц, поступающих ото всех частей их тел. Он
27
полагал, что внешние условия, в которых развивается организм,
не формируют его сами, но лишь реализуют те или другие его
качества, потенциально имеющиеся у зародыша данного вида.
(Вот где источник позднейших мыслей о прототипе.) Тем самым,
разнообразие организмов — это прежде всего разнообразие их по-
тенций (заданных прототипами), а потом уже — условий суще-
ствования. Эта простая мысль, проходя через всю историю биоло-
гии, всегда понималась с большим трудом.
Парацельс сравнивал природу с аптекой, где Бог — главный
провизор, дав этим афоризмом начало целой веренице сравнений.
Лейбниц сравнивал мир с часами, позже мир видели как фабрику,
как систему уравнений, как организм... А самый организм видели
как механическую машину (Декарт), как химическую фабрику,
как ЭВМ. «Мысль — не более, как ток электричества по проводам
нервов», — писал Томас Гексли, один из первых дарвинистов;
вслед за ним мышление уподобляли работе парламента, телефон-
ной станции, ЭВМ и даже проводили параллель между памятью
и голографией. Каждое такое объяснение казалось очень убеди-
тельным — пока не проходила мода на соответствующую новинку
техники. Это нам надо постоянно иметь в виду: будем стараться
не путать моду с выявлением чего-то постоянного и важного.
С другой стороны, всегда есть опасность упустить важную
мысль только потому, что она выражена в ненаучной, с нашей
нынешней позиции, форме. Парацельс, например, высоко ценил
астрологию, а ее развитие наук отвергло. Стоит, однако, вдуматься
в его рассуждение о человеке как «микрокосме», чтобы понять,
что Парацельс в своих фантазиях (врач «постигает пульс небесного
свода, физиономию звезд, хиромантию в минералах, дыхание в
ветрах, лихорадку в землетрясениях») искал общие законы приро-
ды, те, которые много позже Лейбниц характеризовал как «един-
ство в многообразии».
Парацельс решил, вопреки Библии, что каждая раса «имела
своего Адама». (Вопрос стоял тогда очень остро: обязательно ли
считать полноправными людьми вновь открытое население Аме-
рики, не упомянутое в Библии? Несмотря на папскую буллу 1512 г.,
объявлявшую всех людей потомками Адама и Евы, конкистадоры
обращались с туземцами, как правило, до абсурда жестоко.) Вопрос
о независимом происхождении рас обсуждается, с перерывами,
до сих пор.
Первым собрал сведения о заселении Америки Мэттью Хэйл,
правовед, богослов и натурфилософ, и пришел к заключению,
что здешние высшие животные, равно как и люди, произошли
от мигрантов из Старого Света, причем их своеобразие — результат
накопления изменений (возникших случайно или под влиянием
местных условий жизни) и скрещиваний измененных форм друг
с другом [Hale, 1677, с. 196 —199]. Что касается низших животных,
то их он считал попросту порождением сочетаний «семенных
молекул», витающих в земле, воде и воздухе [там же, с. 274].
Он же провел замечательную аналогию между скрещиванием орга-
28
низмов и взаимовлиянием западных языков [там же, с. 197].
Значительно дальше продвинулся физик и философ Пьер Луи
Мопертюи [Maupertuis, 1756]. Он писал, что комбинацией частиц
отцовского и материнского «семени» можно объяснить не только
сходство ребенка с обоими родителями, но и феномены уродства
(как недостаток или избыток частиц определенного типа), и про-
исхождение новых видов, и, естественно, — обычное разнообразие
особей внутри вида. Эти «частицы» не были для него абстрактной
натурфилософской идеей — он пришел к ним на основе реальных,
пусть и скромных, наблюдений. В 1747 г. он напоминал, что
великий Ньютон видел целесообразность (божественный замысел)
в орбитах планет (они движутся по почти круговым орбитам все
в одну сторону) и в строении животных — в их «единообразии»
и «чудесной, полной пользы, организации». Но, возражал Мопер-
тюи, «если единообразие, наблюдаемое нами во многом, является
доказательством, то не опровергается ли это доказательство беско-
нечным разнообразием, которое мы наблюдали во многом другом?»
Так, некоторые видят целесообразность в складках кожи носорога
(иначе она не могла бы сгибаться), но забывают при этом о черепа-
хе, панцирь которой действительно не сгибается. Можно, правда,
искать целесообразность в строении организма как целого, но зачем
сложнейшие формы и инстинкты даны насекомому, «которое по-
жирает первая птица»? Для Мопертюи явно односторонни и фило-
софы, которые ищут целесообразность только в природе как целом,
и Лейбниц (мир просто невозможно сделать лучше), и Мальбранш
(мир создан простейшим из возможных способов), и Поп («все, что
существует, хорошо»), и Декарт (мир попросту движется по зако-
нам механики). Сам он предлагал искать «первичные зако-
ны»— те, которым подчиняется природа, но которые обычно не
удается найти в феноменах, «являющихся только слишком слож-
ными следствиями этих законов». Существует общий закон творе-
ния, такой, что «движение животных, произрастание растений,
вращение звезд являются только его следствиями». Это — принцип
наименьшего действия, открытый самим Мопертюи, одна из основ
механики [Мопертюи, 1959].
Здесь ясно выражена важная мысль: для понимания явлений
надо выявить в чистом виде те элементы, комбинация которых
(«слишком сложные следствия») создает разнообразие; так, еди-
ный набор частиц наследственности (через 150 лет их назвали
генами), комбинируясь, может вызывать к жизни, согласно Мопер-
тюи, все разнообразие организмов. Пусть мы не считаем (как счита-
ли первые генетики), что организм сводится прямо-таки к набору
генов, но так не думал и Мопертюи: его «частицы» наделены
атрибутами целостности — «сродством» и «памятью».
Бюффон включил понимание «частиц» Мопертюи в свою кон-
цепцию, которую усвоил Эразм Дарвин, дед Ч. Дарвина. (В 1868 г.
Ч. Дарвин назвал ее пангенезом, а еще через 20 лет ботаник
Гуго де Фриз в статье «Гипотеза внутриклеточного пангенеза»
дал набросок основ генетики.) Бюффон ничего не добавил к идее
29
«частиц» Мопертюи, но его идею комбинативности он блестяще
развил как морфолог, отметив, что она приводит к параллелизмам,
к повторению свойств от группы к группе: у одного конца группа
млекопитающих сходна с птицами (летучие мыши), у другого — с
рыбами (киты, тюлени), в ее середине обезъяны сходны с челове-
ком. С другой стороны, группа птиц как бы протягивает ветвь
в сторону млекопитающих - - это нелетающие птицы. С удивитель-
ной проницательностью Бюффон писал: «...Ветви, которые прости-
раются к другим отрядам, всегда довольно коротки и образуют
лишь очень маленькие роды»; эти редкие и малые роды важны,
однако, тем, что должны «дать понять философам, что она [приро-
да] не может быть сжата оковами наших классификаций...» (По-
следние слова — упрек Линнею, системы которого он не призна-
вал.) От этих «ветвей» всего шаг до рефренов и периферии (см.
гл. 2).
Интересный поворот той же мысли мы находим у Мильн-Эд-
вардса, в его «законе повторений»: природа повсюду скупа на
первичные элементы, повсюду применяя одни и те же конструкции
по многу раз, — это и многочисленные реснички инфузории, и
сходства планов строения позвоночных, и идентичность плана
руки и ноги. Все это он выводил из более общего «закона эконо-
мии». Экономия на новшества — это, по существу, переосмысление
принципа наименьшего действия Мопертюи. Такое же переосмы-
сление, но уже со ссылкой на Мопертюи, произвел и Дарвин:
«...Взгляд, согласно которому первоначально были созданы немно-
гие формы или лишь одна,., согласуется с философской аксиомой
Мопертюи о „наименьшем действии”» [Дарвин, 1951, с. 107].
Другой подход к комбинативности составило изучение парал-
лельных рядов. Еще Баугин в своей «Описи растительного мира»
фактически регистрировал параллели форм. Так, род Plantago
(подорожник) он разделил на три группы видов: широколистные
(Р. latifolia), узколистные (Р. angustifolia) и водные (Р. aquatica),
причем среди водных (вероятно, обозначавших нынешний род
Littorella) тоже выявил широколистную и узколистную формы,
а также выявил форму stellata (сверкающая) как в составе Р. lati-
folia, так и в составе Р. aquatica [Bauhin. 1623, с. 189 — 190].
К большинству названий видов Баугин давал списки синонимов,
из которых видно, что уже тогда параллелизм свойств вызывал
путаницу в названиях. Чтобы ликвидировать ее, Линнею пришлось
постулировать иерархию диагностических признаков, например,
он полагал, что число тычинок цветка определяет класс, а пести-
ков — порядок в этом классе. Тем самым Линней искусственно
изобразил логическим древом то, что само по себе (естественно)
было таблицей: порядок точнее мог быть задан пересечением стро-
ки (число тычинок) и столбца (число пестиков). В результате
параллелизм выпал из понятийных рамок систематики.
В XIX в. внимание к параллельным рядам возникало не раз,
но оставалось уделом отдельных авторов. Так, Иоганн Якоб Берн-
гарди в 1834 г. отмечал, что растения, в других отношениях
30
несходные, дают сходные ряды типов листа. Изложив его мысль,
Дарвин в 1868 г. добавил: гладкокожий персик выведен из немно-
гих форм бархатистого, но «между сортами бархатистых и гладких
персиков наблюдается замечательный параллелизм» — общие ва-
рианты цвета плода и его мякоти, характер косточки, цветков,
листьев и т. д., хотя «признаки каждого сорта гладкого персика
получены отнюдь не из соответствующего сорта бархатистого»
[Дарвин, 1951, с. 716].
В 1825 г. шведский ботаник Элиас Фрис предложил систему
растений, в которой отмечались сходства между таксономически
далекими растениями, демонстрирующими сходные «степени раз-
вития», например, между однодольными и двудольными. Ботаник
М. А. Максимович (более известный ныне как энтузиаст украин-
ской словесности) значительно развил мысли Фриса. В своих
выводах он писал: «В природе везде двойство: свет без двойства
являться не может». Это «двойство» он видел во всяком познании,
призывая идти попеременно то от высших форм к низшим, то
наоборот; то от общих свойств организмов к частным, то наоборот;
то от органов к плану строения, то наоборот; то от анатомии к
физиологии, то наоборот. Выискивая «двойства» (взаимодополни-
тельные аспекты - сказали бы мы сейчас, зная принцип дополни-
тельности Бора), он отметил замечательную мысль Фриса: если
группировать виды по «сродству» (т. е. по гомологии) в столбцы,
то образующиеся при этом строки (соседние виды, образующие
разные столбцы) тоже будут образовывать сходства — «аналогии»
| Максимович, 1827]. Через 93 года Н. И. Вавилов назвал этот
феномен гомологическим рядом наследственной изменчивости. Он
показал, что один и тот же видовой комплекс может встречаться
в различных родах настолько закономерно, что его можно предска-
зывать. Так, изучая злаки Памира, он заметил, что характерные
признаки разновидностей пшеницы удивительно повторяются у
разных форм ржи. Следовательно, решил Вавилов, видовое разно-
образие упорядочено: «Мутации, идущие как бы случайно в разных
направлениях, при объединении их обнаруживают общий закон».
Теперь вопрос о параллелизмах приобрел совсем новый аспект:
откуда они берутся — создаются требованиями среды независимо,
реализуются за счет свойств, унаследованных от общего предка,
прямо передаются от вида к виду подобно инфекции, или виды,
эволюционируя, могут развиваться не как попало, но только в
рамках ограниченного числа логических возможностей (как гово-
рил Дарвин, в рамках «природы организма») ? Два таксона могут
как бы двигаться на разных этажах, но по одинаковым коридорам.
Дарвин писал: «...Члены одного и того же класса, независимо
от их образа жизни, сходны между собой по общему плану органи-
зации» [Дарвин, 1939, с. 623], причем «гомологичные части склон-
ны изменяться одинаковым образом» [Дарвин, 1951, с. 697].
Тогда же (1868) палеонтолог Эдуард Коп, пытаясь истолковать
параллелизмы у ископаемых, писал, что родовые признаки могут
меняться со временем без обязательного изменения видовых [Фи-
31
липченко, 1977]. В целом же, как констатировал Любищев, вопро-
сы параллелизма мало занимали Дарвина и его последователей,
поскольку не гармонировали ни с его теорией, ни со взглядами
эпохи.
После работы Вавилова некоторые стали смотреть на систе-
матику и генетику (а частично и на морфологию) как на науки о
комбинациях. Среди них оказались такие крупные наши биологи,
как ихтиолог Л. С. Берг, гистолог А. А. Заварзин, энтомолог
А. А. Любищев, палеонтолог Д. Н. Соболев и генетик Ю. А. Филип-
пенко. Последнему принадлежит, кстати, первая обзорно-теоре-
тическая статья о «законе Вавилова», где сформулированы три
вида параллелизмов — генотипический, обусловленный сходными
генами и наблюдаемый обычно у близких видов, анатомический,
«вытекающий из одинаковых возможностей развития», и гисто-
логический, «который выражается в сродстве гистологической
структуры одинаково функционирующих частей у очень далеких
в смысле своего систематического положения организмов» [Фи-
липпенко, 1924, с. 255]. Верный своему времени, автор предлагал
применять термин «комбинация» только к генотипическому
параллелизму [там же, с. 254], поскольку только для генов мог
представить механизм комбинирования. (Впоследствии стало ясно,
прежде всего — на основе работ Мейена, что комбинироваться
могут любые факторы, ответственные за параллелизм.) Зато он
как бы предвосхитил наше нынешнее системное понимание срод-
ства (которое будет изложено далее, в 9.3) — это видно из того,
сколь необычно для тогдашней биологии он употребил подчеркну-
тое мной слово «сродство». На эту замечательную работу Филип-
пенко мое внимание обратил В. В. Бабков.
В наше время понемногу выясняется, что познание разнообра-
зия прежде всего состоит именно в выяснении законов парал-
лелизма (пользуясь аналогией — плана коридоров, общего для
разных этажей), и важное место среди них занимает комбинатив-
ность. Только узнав эти и прочие законы, по которым упорядочено
разнообразие живого, можно всерьез заняться проблемами эволю-
ции, которая предстает при этом как процесс исторического пре-
образования разнообразия — экосистем, таксонов, организмов
и составляющих их структур.
Глава 2. РАЗНООБРАЗИЕ, РЯДЫ И ТЕНДЕНЦИИ
Основная цель этой главы — познакомить читателя с диатропиче-
ской концепцией С. В. Мейена в той мере, в какой это необходимо
для понимания данной книги. Поэтому, в частности, опущен геоло-
гический материал, бывший для него основным [Мейен, 1977],
зато привлечены другие примеры, на мой взгляд, более доступные
большинству читателей, а также проведен ряд историко-методоло-
32
гических параллелей. Мейен не оставил единого изложения своей
концепции, однако прочел первый вариант предлагаемого ниже
текста и сделал много письменных замечаний, которые я постарал-
ся учесть со всею возможною полнотой.
2.1. О биологической грамматике
Учения о различиях (систематика), о сходствах (морфология)
и о комбинативности форм (например, элементы генетики) вошли
в общее употребление с итервалами примерно в сто лет (первая
треть XVIII, XIX и XX вв. соответственно), хотя и начали свое
развитие в одно время (вторая четверть XVI в.) и не испытывали
недостатка в талантливых исследователях. Очевидно, уже эти три
круга проблем достаточно различны по степени трудности их усво-
ения научным сообществом, тогда как сейчас у нас речь пойдет
о более трудном, о четвертой линии познания разнообразия — об
упорядочении материала, даваемого первыми тремя линиями, о
биологической грамматике '. Введение этого термина должно
подчеркнуть сходство познавательных процедур: если общность
словоформ рыбе и яме мы легко устанавливаем как «дательный
падеж единственного числа 2-го склонения существительных», то
нельзя ли так же обойтись, например, со сходными формами
описанных Дарвином персиков или Вавиловым — злаков?
Трудность усвоения связана с тем, что первые три линии поз-
нания никак не назовешь последовательными ступенями — углуб-
ляясь в познание разнообразия, все время приходится опираться,
как при спуске в горную расщелину, то на одну стену, то на другую,
то на удачный выступ спереди или сзади. Чтобы понять это, напом-
ним, как пользуются системой организмов.
Положим, вы поймали птицу И хотите определить ее вид. Ход
и успех работы радикально зависят от того, что вы заранее знаете.
' Прочтя это место в первом варианте рукописи, Мейен записал следующее: «Мне
не очень нравится такое разбиение: систематика — различие, морфология —
сходство. Со времен Жюссье систематика строится по конгрегационному прин-
ципу, т. е. по сходству; мы идем путем последовательного объединения сходного.
Кроме того, я считаю морфологию (отчасти) систематикой частей, причем здесь
действует (как показал Е. С. Смирнов) тот же конгрегационный принцип. Ком-
бинативность же не рядоположена этой паре, она указывает на один аспект
распределения признаков в системе. Твоя четвертая линия — результат соедине-
ния систематики и морфологии (это типология в моем понимании)». Здесь все
верно, но противоречия между нами я не вижу. Все разрешается, по-моему,
взятым у Мейена в скобки словом «отчасти»: у меня речь идет о другой «части»
классификационной науки. Ведь систематика тоже лишь отчасти строится по
сходствам — уже тот факт, что ни один объект не может остаться вне системы,
говорит о том, что первичен здесь принцип разбиения заданного множества (см.
главу 6), а принцип объединения вторичен. Если же рассматривать только раз-
граничительный аспект систематики (вольно говоря, систематику по Линнею) и
только объединительный аспект морфологии (вольно говоря, морфологию по
Юнгу), то вне их останется именно комбинативность, точнее — учение о паралле-
лизмах, которое в этом смысле им рядоположено. На базе этой тройки и нужен
синтез, даваемый далее схемой, которую Мейен называл типологией, а я — биоло-
гической грамматикой.
2 Ю. В. Чайковский
33
Прежде чем взять определитель птиц, вы должны быть уверены,
что поймали именно птицу, а это предполагает какое-то знаком-
ство с морфологией: не все птицы летают, летать могут не только
они и т. п. Далее, к какому отряду она принадлежит? Если вы взя-
ли определитель впервые, то с удивлением обнаружите, что раз-
личение даже отрядов (не говоря уже о меньших таксонах) идет
по признакам, ничего вам не говорящим, не имеющим отношения
к обыденному опыту. Например, принадлежность к отряду во-
робьиных определяется по характеру щитков на задней стороне
цевки (части ноги, от которой отходят пальцы). Пусть вам уже уда-
лось установить, что птица относится к роду синиц семейства
синицевых отряда воробьиных (формально доказать это совсем
непросто для начинающего), и осталось только определить ее
вид. Читаем: «Вдоль средней линии груди и брюха проходит чер-
ная или черноватая полоска»,— если это найдено, перед нами
Синица большая; если нет, то надо читать дальше: когда на за-
шейке имеется светлое пятно, то надо смотреть на концы крою-
щих перьев (если концы белые, то это Синица московка, если
пет — читать дальше), когда же пятна нет, то надо смотреть на цвет
спины, и т. д.
Какой кропотливый 400-летний труд скрыт за этими диагно-
зами! 2 Однако, определяя вид по дихотомической таблице, мы не
познаём самых его существенных свойств («завершающих раз-
личий»), мы ведем себя, словно комиссия в музее, сверяющая
инвентарные бирки ярких полотен и равнодушная к самим полот-
нам. Ведь большая синица — птичка яркая, а московка — грязно-
ватая. И все-таки, вопреки великому Аристотелю, мы почти всегда
доходим, следуя дихотомии, до определенного вида. Почему? Да
просто потому, что всякая дихотомическая таблица делается теперь
по готовой системе, а сама система была придумана без дихотомии;
систематик, желая упорядочить известное ему разнообразие (на-
пример, воробьиных), следует линнееву афоризму (не признаки
задают род, но род задает признаки), т. е. пользуется не формаль-
ным ключом, а интуицией. Только создав хорошую систему, можно
начинать ее формализацию — хотя бы составляя дихотомиче-
скую таблицу,— и недаром Ламарк достиг впервые в этом успеха
лишь в год смерти Линнея. В то же время при Аристотеле никакой
писанной системы 3 не существовало, дихотомию рассматривали
как путь ее построения, и ничего, разумеется, не получалось.
2 Удобная дихотомическая форма (если да, то читай одно, если нет — другое)
пришла в систематику только в 1778 г. с «Французской флорой» Жан-Батиста
Ламарка, хотя предлагали ее еще до Линнея (например, Мартин Йорений в
1710 г. [Максимович, 1827, с. 17]); ее прекрасно знал и Аристотель, который пи-
сал, что путем дихотомии «приходишь к последнему признаку, но не к завер-
шающему различию и виду» и что поэтому «тем, кто применяет дихотомию,
невозможно дойти до какого-нибудь отдельного животного» [Аристотель, 1937,
с. 47].
3 Мейеп не раз подчеркивал, что в основе классификационных построений очень
долго лежали бытовые («кухонные») таксономические и морфологические пред-
ставления, что без них невозможно начать работу [Мейен, 1977, с. 29].
34
На этом простом примере можно убедиться в очень важном
факте: познание разнообразия состоит как в его интуитивном
уяснении, так и в его упорядочении, причем интуитивное пони-
мание и формальное упорядочение невозможны друг без друга.
Да, дихотомия Ламарка построена на системе растений Линнея,
но ведь и система стала возможна потому, что еще давным-давно
были выявлены и закреплены словами разговорного языка такие
дихотомии, как живой —неживой, животное — растение, муж-
ской — женский, наземный — водный, и многие другие. Не менее
четки и важны были также деления организмов на части (напри-
мер, корень, стебель, лист, цветок, плод и т. п.). Долиннеевы систе-
матики уже исходили из этих и подобных делений (морфологиче-
ских, физиологических, экологических и т. и.) как данных. Наука о
разнообразии должна строиться последовательными шагами:
каждый успех морфологии позволяет улучшить систематику,
и обратно. Такой прием называется итеративным.
Одна из первых задач диатропики — сделать итеративный
характер познания разнообразия явным, эксплицировать итера-
тивные процедуры. Это давно проделано в математике, где извест-
но, что итеративная процедура либо приводит к определенному
пределу, либо оказывается бесконечным блужданием. Предел
(данной процедуры на данном множестве) является инвариантом
множества в том смысле, что конечная точка процедуры не зави-
сит от того, с какой точки множества итерации начаты. Встает
задача выявления структурных инвариантов разнообразия орга-
низмов. В других науках подобные инварианты успешно выявляют-
ся 4, а в биологии для их выявления естественно обратиться
к вавиловским рядам. Почему открытия с их помощью так редки?
Почему определители не делаются в форме таких таблиц, в которых
все таксоны одного ранга были бы представлены по единым пра-
вилам, пусть и с исключениями, подобно таблицам склонения (на-
борам падежных форм) или спряжения (наборам личных и вре-
менных форм) в языке?
Выяснение законов «биологической грамматики» едва начато.
Здесь дело сдвинулось только после переосмысления закона Ва-
вилова: «в законе Вавилова в его обычной формулировке обраща-
ется внимание на повторность отдельных признаков от таксона
к таксону, но опущена повторность в правилах преобразования
варьирующих признаков», причем «запомнить порознь все разно-
видности и относящиеся к ним самостоятельные термины невоз-
можно (это все равно, что запоминать порознь все склонения
(правильнее: падежные формы,— Ю. Ч.) каждого существитель-
4 Такова в этнологии структура мифа, которая сохраняется неизменной при всех
вариациях, происходящих во времени и пространстве,— на это обратил внимание
еще Александр Гумбольдт в 1814 г. [Гумбольдт, 1963, с. 293], а в XX в,— этно-
лог Клод Леви-Строс. В филологии примерами инвариантов являются грамма-
тические категории (падеж, время и т. д.): любое русское существительное при-
надлежит шестипадежному ряду, именуемому склонением (если пренебречь ру-
диментарными падежами), любой русский глагол — трехвременному трехлично-
му ряду, именуемому спряжением, и т. д.
2*
35
ного, не познакомившись с правилами склонения) » [Мейен, 1978,
с. 500—501]. Добавим, что падежи, когда их много, не просто
перечисляются (как в латыни и русском), а упорядочиваются. Та-
кова система локативов (местных падежей) в дагестанских языках.
Слова обычно образуют большие группы грамматически сход-
ных форм (таких, как бал и пол, где все падежные формы иден-
тичны), т. е. подчиняются единой парадигме. Однако встречаются
слова, к исходной форме которых не подобрать пары, сходной
и грамматически, и по звучанию конца. Так, существительному
хрящ нет созвучного ему существительного в том же числе и па-
деже (оканчивающегося на -ящ). Зато, просклоняв хрящ, найдем,
что его производным формам созвучна группа слов (того же скло-
нения) типа хвощ', немного хуже соответствует праща — в ходе
развития языка слово попало в другое склонение. Кроме них, мно-
го разрозненных сходств — совпадений той или иной падежной
формы с отдельными словоформами других частей речи (см.
табл. 1).
Таблица 1. Пример фонетического параллелизма (совпадения ударных концов)
между падежными окончаниями и прочими словоформами
Падеж Данное Сходные по склонению Несходные части речи
тельное существительные словоформа и ее исходная форма
им.-вин. хрящ — [пращ *] горящ * (краткое причастие от гореть)
РОД. хряща хвоща праща ** таща (деепричастие от тащить)
дат. хрящу хвощу пращу ** трещу (1-е лицо от трещать)
твор. хрящом хвощом пращом * [общо] (наречие от общий)
пред. хряще хвоще праще вотще * (наречие от тщетный)
ИМ. хрящи хвощи пращи ищи (повелит, форма от искать)
род. хрящей хвощей пращей кощей (именит, падеж)
дат. хрящам хвощам пращам овощам (дат. падеж от овощ)
твор. хрящами хвощами пращами щами (творит, падеж от щи)
пред. хрящах хвощах пращах натощах * (наречие от тощий)
* архаическая форма * * несовпадение падежей (ныне праща — женского рода). Прямые скобки означают неполное созвучие ударного слога
В языках, не имеющих падежей, ту же закономерность легче
всего увидеть на глагольных формах. Так, французские глаголы
devenir, revenir, souvenir, tenir, venir и т. п., образующие свою
небольшую группу спряжения: venir—venant — viens, vient—ve-
nu—vins, сходны с существительными: инфинитив сходен c avenir
и souvenir, настоящее время — с soutien и revient, активное прича-
стие — с revenant и tenant, пассивное причастие — с avenu и
revenu, простое прошедшее — с vin и devin. Есть сходства с при-
лагательными (avenant, tenu), местоимением tien, междометием
tiens и т. и.
36
Подобным же образом изменяются признаки биологических
видов, родов и т. и., но если в ботанике нередки довольно полные
параллельные ряды, то для зоологии более характерны разрознен-
ные сходства. Возьмем для примера тех же синиц. Собственно си-
ницы (род Parus семейства Paridae) образуют в фауне СССР 12 ви-
дов, но термин «синица» применяется и к другим воробьиным:
ремезу, длиннохвостой синице, усатой синице и др. Уже это гово-
рит о параллелизме: птицы других родов и даже семейств имеют
характерный «синичий» облик. Однако параллелизм наблюдается
гораздо шире. Почти все признаки, отраженные в названиях синиц,
представлены и у других воробьиных (см. табл. 2), но, как и в при-
мерах со словами, здесь нет таксономических границ для проявле-
ния разрозненных сходств. Так, многие птицы различных отрядов
хохлаты (крохали, турухтан, чибис и др.), а характерная полоска
вдоль щеки, давшая видовое название усатой синице (из семейства
толстоклювых синиц отряда воробьиных), наблюдается и в отряде
дятловых, у сирийского пестрого дятла (на этот факт мое внимание
обратил Е. Н. Панов). Параллелизм не ограничивается классом
птиц; например, хохлатыми называются некоторые виды млекопи-
тающих. Естественно, что в пределах отряда воробьиных паралле-
лизм полнее.
Таблица 2. Пример параллелизма внешних обликов организмов в рамках отряда
Синицы (отряд воробьиных) Другие воробьиные
хохлатая (Parus cristatus) рыжешейная (Р. rufonuchalis) ремез (Remiz pendulinus) толстоклювые синицы (семейство) в том числе: усатая (Panurus biarmicus) длиннохвостая (Aaegithalos caudalus) Свиристель, Жаворонок хохлатый Овсянка рыжешейная Овсянка-ремез Камышовка толстоклювая некоторые овсянки Личинкоед серый; сороки (отчасти)
Оставляя профессионалу-орнитологу доведение таблицы 2 до
максимального сходства с таблицей 1, заметим, что обе таблицы
и так проявляют одно существенное сходство — пересечение рядов.
С одной стороны, слово хрящ входит в обычный ряд существитель-
ных 1-го склонения, а с другой — в ряд слов на -ящ, причем на пе-
ресечении этих рядов слово хрящ — единственное. Аналогично,
хохлатая синица — единственный вид на пересечении ряда синиц с
рядом хохлатых птиц. При этом сама принадлежность объекта ряду
может определяться как одним признаком {-ящ; хохлатость), так
и комплексом признаков (принадлежность 1-му склонению; ком-
плекс признаков рода синиц). Как мы увидим, наиболее интересны
пересечения комплексных рядов. Не раз высказывалась идея
(В. М. Шимкевичем, Н. И. Вавиловым, А. А. Еленкиным и др.),
что саму систематику следует строить так, чтобы таксон (раса,
37
подвид, вид, род и т. д.) задавался пересечением рядов. В некото-
рых случаях это действительно удобно, но в любом случае надо
знать, возможно ли это в принципе,— чтобы понимать структуру
разнообразия.
2.2. Мероно-таксономический анализ (типология по Мейену)
Главная идея анализа состоит в следующем: чтобы познавать раз-
нообразие, обращаясь итеративно то к различиям, то к сходствам,
надо уметь различия и сходства классифицировать сопряженным
образом. В 2.1 мы видели, что объектами итерации служат систе-
матика и морфология, поэтому их следует представить единым
(удобным для итерации) образом. Н. Н. Страхов [1858], зоолог,
более известный сегодня как публицист славянофильского направ-
ления, отметил, что сравнительная анатомия классифицирует
органы так же, как зоология классифицирует организмы. Развивая
эту мысль, Мейен пришел к фундаментальному выводу: гомологи-
зация (классификация частей) и классификация организмов
являются двумя логически самостоятельными процедурами, но не-
явная связь между ними существует — она существует итеративно'.
морфология обеспечивает систематику «признаковым простран-
ством и данными о соотношении признаков у разных объектов»
[Мейен, 1978, с. 496], а систематика дает морфологии пространство
объектов, допускающих соответствующие гомологизации. Эту связь
Мейен решил сделать явной.
В основу он положил понятие мерона. Мерой (от греческого
peQOg — часть) — это «класс частей». Когда мы говорим, напри-
мер, о конечности, это в равной мере может относиться к руке, ноге
или лапке насекомого, а это значит, что мы умеем мысленно расчле-
нять тела животных, выделяя части естественным образом. При
этом слово «конечность» означает результат мысленного объеди-
нения таких частей разных организмов, которые обладают некото-
рыми очевидными общими свойствами,— это подвижные придатки
тела, имеющие собственную мускулатуру и служащие для передви-
жения или хватания (да простит мне морфолог некоторое упро-
щение!). Мерой «конечность» — не конкретная рука или лапка,
а множество всевозможных конечностей. Организм может обла-
дать органами, частями тела, свойствами, признаками, но не меро-
нами — ими обладают только группы объектов. Понятие мерона
обобщает понятие гомологии по Оуэну. Часто, выявляя мерой,
мы называем некоторый таксон систематики (например, мерой
«позвоночник» характеризует подтип позвоночных в типе хордо-
вых, а мерой «цветок» — отдел цветковых растений), но порою ме-
рой объединяет в одну группу организмы, которые нельзя признать
таксоном, например: мерой «пищеварение» объединит животных с
насекомоядными растениями.
Предложено учение о меронах, мерономия, где основным яв-
ляется понятие об архетипе. Мы видели, что для определения пти-
цы надо заранее знать, что такое птица, а это значит не только
38
уметь выявлять характерные для птиц мероны (определенная
форма тела, клюв, крылья, перья и т. д.), но и уметь мысленно
складывать из них обобщенный образ (архетип) птицы, т. е.
знать, какое сочетание комбинаций меронов делает животное пти-
цей. Если сам организм можно мысленно расчленять на части тела
и говорить, что организм из них состоит, то архетип состоит в этом
смысле из меронов.
Однако организмы можно сравнивать не только по частям их
тел, но и по всем их качествам: по способности летать или образо-
вывать брачные пары, по цветным узорам на теле, по роли в экоси-
стеме и даже по размерам. Архетип получается куда богаче, чем по
Оуэну, и строить его нужно по-новому. Прежде его изображали
простеньким рисунком, содержащим только самые общие свойства
таксона. Например, архетип позвоночного — это очень схематич-
ный скелет, правда, с очень сложными позвонками (см. рис. 22 в
кн.: [Бляхер, 1976]), столь же схематичные органы пищеварения
и т. д., т. е. «инвариантная для всех членов таксона совокупность
меронов» [Мейен, 1977, с. 29]. Будем называть архетип-рисунок
архетипом по Оуэну. Архетипом по Мейену будем называть «ар-
хетип в более широком смысле», когда «гомологизированные, то
есть расклассифицированные и ставшие меронами части организ-
мов данного таксона в сумме составляют архетип таксона» [Мейен,
1978, с. 497]. Это обширная таблица, где каждая строка — мерой,
а каждый столбец — возможная в принципе для данного таксона
особь. Для позвоночных она, например, включает строку «конеч-
ности», в которой друг за другом записаны всевозможные значения
мерона: отсутствие, плавник, ласт, лапа и т. д., причем каждый вид
конечности может иметь разные степени развития. Грубо говоря,
архетип по Оуэну — пересечение множеств признаков, а по Мей-
ену — их объединение. В терминах модальной логики [Целищев,
1977, с. 5 -9| архетип по Мейену близок к понятию модельной
структуры (системы), а по Оуэну - общезначимому утверждению
в ней.
Чаще всего эволюция — это реализация нового столбца, но
прогрессивная эволюция всегда включает появление новой строки
(и связанных с нею новых столбцов). До появления птиц в таблице
позвоночных не было, например, строки «перья» или, если таблица
менее детальна,— в строке «роговые покровы» были пусты клетки,
соответствующие всевозможным перьям. Вообще, большинство
клеток таблицы пусто всегда (поскольку особь задается комбина-
цией признаков, а мерой — самими признаками), зато число строк
и столбцов в ней необозримо, и нет смысла выписывать ее в явном
виде. Она — средство теоретического анализа, подобная матрицам
в математике, которые можно изучать, не выписывая самих таблиц
в явном виде.
Если положить головами в одну сторону рыбу, лягушку, яще-
рицу, воробья и крысу, то легко увидеть ряды сходных частей:
ряд черепов, ряд глаз, ряд желудков и т. д. Их сходство объясняют
общностью происхождения всех позвоночных, но вопрос не так
39
прост: ведь сюда можно подложить и рака, и жука — параллелей
будет меньше, но все-таки много. Если здесь и можно говорить об
общем предке, то уж у него-то не было ни мозга, ни сердца, ни раз-
дельнополости — всего, чем сходны высшие животные. Эта общая
тенденция к появлению сходных свойств и есть параллелизм.
Параллелизмы бывают самые разные. Если взять много разных
видов рыб, птиц, насекомых и т. д., то легко увидеть параллели в
окрасках — не только покровительственные или контрастные, но и
просто сходные рисунки. Скелет руки поразительно сходен со ске-
летом ноги, хотя рука и нога всегда имели отдельных «предков» —
переднюю и заднюю лапы. Когда удается обследовать много тысяч
членов одного вида, открывается еще один морфопараллелизм —
уродства. В них часто реализуются признаки, нормальные для дру-
гих видов, и это важно для осознания морфологического единства
природы. Палеонтолог Д. Н. Соболев считал, что «почти для каж-
дого из известных типов аномалий можно подобрать соответствую-
щие примеры видообразования» [Соболев, 1924, с. 161]. В 1935 г.
П. Н. Кренке обосновал на ботаническом материале правило: при-
знак, являющийся уродством у данного вида, обычно нормален в
каком-то ином виде [Мейен, 1978].
Обычно у миндаля плод сухой, а у персика — окруженный мя-
систым околоплодником, но изредка бывает наоборот, причем не-
обычная форма может существовать и как индивидуальная вариа-
ция, и как устойчивая раса. Эта пара видов проявляет целый ряд
сходств изменчивости, и его можно сопоставить с падежным рядом
существительных одного склонения. Как и в грамматике, ряды
сходств довольно полны для близкородственных видов (персик и
миндаль обычно включают в один род) и довольно разрозненны для
форм далеких; но как и в грамматике, определенных границ для
проявления сходств нет. Кренке назвал этот феномен законом род-
ственной изменчивости [Корона, 1987, с. 50 — 53]. Вне ботаники
прекрасным примером того же закона является мутационная из-
менчивость дрозофил, почти одинаковая у многих видов этого
рода.
Это оказалось общим правилом: чем значительнее отклонение
от нормы, тем реже оно наблюдается и тем дальше таксономически
вид, у которого соответствующее свойство можно встретить в каче-
стве нормального. Такой безобидный дефект, как нарушение пиг-
ментации, дает признаки другой расы (белый негр, описанный у
Мопертюи); а уже «заячья губа» не встречается в качестве нормы у
человека и обезьян, зато обычна у лемуров (низшие приматы).
Очень редкие дефекты людей (нечто вроде чешуи) нормальны для
низших позвоночных, а самые редкие (и тяжелые) уродства прихо-
дится искать в качестве нормы еще дальше; так, две головы на вд-
ном теле и одна голова на два тела возможны как норма у некото-
рых червей. Отметим, что дело здесь не в атавизме; черви предками
людей не считаются. Не было и рогов у предков лошадей, а рогатые
лошади иногда рождаются. (Примеры сообщены мне С. В. Мейеном
и А. С. Раутианом). Без «грамматики биологии» было почти невоз-
40
можно сказать, что общего есть между всевозможными паралле-
лизмами, в ее же свете видно, что все они подчиняются простой схе-
ме: у разных таксонов общие мероны. Этой формуле можно придать
вполне грамматический вид, и недаром раздел грамматики, изуча-
ющий строение слов, тоже называется морфологией. Слово делится
на основу и окончание, которое, изменяясь, указывает грамматиче-
скую форму (падеж, время и т. д.). Изменяя окончания, мы полу-
чаем разные словоформы, т. е. термин «слово» выступает как род,
в котором отдельные словоформы являются видами (диалектные
варианты их выступают в качестве рас). Основа может состоять из
корня и аффиксов (приставки и суффикса), и группа слов с общим
корнем играет роль семейства. Слова можно группировать по-раз-
ному — просто в алфавитном порядке (обычные словари), в алфа-
вите окончаний (рифмовники), по частоте употребления (частот-
ные словари), по смыслу (словари синонимов), по происхождению
(этимологические словари) и т. и., и какой бы способ ни выбрать,
обнаружится параллелизм слов — потому что другие, не исполь-
зованные для классификации признаки будут заявлять о себе на
каждой странице. Так же и в биологии: по каким бы правилам ни
создавать систему, другие, неучтенные закономерности выступят
как параллелизмы.
Если в опытных и описательных науках исходным является
понятие факта, то в диатропике — понятие ряда 5. И так же, как
факт не имеет смысла (а подчас и места) вне объясняющей схемы,
так и ряд бессмыслен без сопоставления (параллели) с другими
рядами. Тем самым параллелизм выступает исходным теоретиче-
ским элементом диатропики. С его помощью строятся архетипы.
Архетип по Оуэну сворачивает разнообразие в единый образ, архе-
тип по Мейену максимально разворачивает его с целью показать
пределы возможного.
Мейен предложил видеть в каждом таксоне как архетип по
Оуэну (т. е. нечто коренное), так и изменчивое множество, причем
это последнее оказывается упорядоченным, как упорядочены па-
дежные или личные формы (подробнее см. 2.4). Эту упорядочен-
ность можно наблюдать на таксонах всех уровней — от подвида
(бархатистые и гладкокожие персики) до надцарства (водоросли
повторяют свойства цианобактерий), но мы поясним ее на одном
примере — характере изменений в пределах мерона «конечность».
Этот мерой вообще может отсутствовать в архетипе таксона
(губки, черви, растения), а может принимать самые различные зна-
чения. Если говорить только о парных конечностях позвоночных и
избегать деталей, то обнаружится следующая картина. Позвоноч-
ные делятся на пять классов: рыбы, амфибии (земноводные), реп-
5 Слово «ряд» надо понимать широко, как латинское ordo, означающее и ряд и по-
рядок. Так, бытует выражение «падежный ряд», хотя, например, в языках Во-
сточного Дагестана лучше видеть систему таблиц: одни лишь локативы (падежи,
соответствующие вопросам где, куда, откуда), образуют здесь до 8 серий (верх,
низ, глубь и т. д.) по 3 падежа [Магометов, 1965, с. 117], т. е. таблицу. Порядок
может означать не только линейную, но и любую организованность.
41
тилии (пресмыкающиеся), птицы и звери (млекопитающие). Ме-
рой же принимает девять основных значений: простой плавник —
только у рыб; плавник-ласт (с суставчатым скелетом) — встре-
чается у всех пяти классов (кистеперые рыбы, ископаемые амфи-
бии и рептилии, пингвины и киты); ласт (для плаванья и пол-
занья по земле) — у вымерших амфибий и рептилий, у черепах и
млекопитающих (ластоногие); рыбы, как ни странно, ползают с
помощью простых плавников; ползателъная лапа — у многих ам-
фибий, рептилий и некоторых млекопитающих, часто снабжена
плавательной перепонкой; «планер» (лапа или плавник с летатель-
ной перепонкой) — у некоторых рыб, лягушек, ящериц и мелких
зверей; крыло для активного полета — у вымерших птицеящеров,
у птиц и летучих мышей; как редкое исключение — у клинобрю-
шек (рыбок из семейства харциновых) — наблюдаются элементы
активного полета; нога для хождения и стояния без касания брю-
хом земли — у некоторых вымерших амфибий, у рептилий, птиц
и зверей; хватательная лапа — известна у всех классов, кроме рыб;
рука —- только у приматов.
В этих параллельных рядах наблюдаются одни и те же тенден-
ции. Как у всякого мерона, у конечности бывают разные степени
выражения: полное отсутствие, рудимент (зачаток, невидимый
снаружи или видимый, но бесполезный), слабое развитие, полное
развитие. Во всех классах мы видим тенденцию к реализации са-
мых разных степеней выражения конечностей: полное отсутствие
наблюдается у рыб (семейство угрей — отсутствует передняя или
обе пары), у амфибий (семейство червяг) и рептилий (змеи); ру-
дименты бывают у пещерных рыб, многих хвостатых амфибий и
ящериц, у китов (задняя пара) и у некоторых птиц (передняя
пара). Оформленные, но совсем слабые конечности тоже хорошо
известны у всех пяти классов, так что можно выстроить пять парал-
лельных рядов по степени их развития (о других тенденциях
см. 2.5).
«То, что разные члены рядов изменчивости объединяются од-
ной тенденцией, одним правилом преобразования, дает возмож-
ность предсказывать и целенаправленно искать недостающие
члены. Эту повторяющуюся, подчиненную одному правилу преоб-
разования последовательность состояний мерона назовем рефре-
ном»,— писал Мейен [1978, с. 501]. Как и сами мероны, рефрены
могут быть выявлены в любой теории классификации.
Лучше всего обследованы языковые рефрены — формы спря-
жения глаголов, склонения существительных и т. д. Неполнота ря-
дов характерна и здесь — как для отдельных слов («сутки» не
бывают в единственном числе, «рожь» — во множественном, хоть
это и вынуждает строить громоздкие обороты речи), так и для це-
лых классов (в современном русском языке сослагательное накло-
нение утратило формы времени). И все же именно благодаря реф-
ренам грамматика является полезным для всех инструментом —
они помогают лучше понимать язык при меньшей затрате сил:
вместо заучивания каждой словоформы, мы запоминаем исходное
42
слово и то правило, по которому изменяется данная группа слов
(о биологических рефренах см. 2.4).
Еще в 1862 г. Герман Гельмгольц, блестящий физик и физиолог,
обращал внимание на необходимость организации знания по грам-
матическому образцу. Он говорил: «С помощью хорошего словаря
гимназист может сделать теперь кое-что в объяснении классиков,
что должно быть так трудно было Эразму, несмотря на начитан-
ность долгой жизни», так как требуется хорошая систематизация,
«чтобы не потеряться безнадежно в лабиринте учености». Системы
«представляют, так сказать, основной фонд научного достояния че-
ловечества, проценты с которого пускаются в хозяйство» [Гельм-
гольц, 1866, с. 15 —16]. Он указывал, что хорошая система — не
просто каталог, а инструмент, способный к предсказанию. Об этом
писал и Мейен: возможность восстанавливать значения неизвест-
ных меронов на основании знания архетипа он называл типологи-
ческим прогнозом, возможность которого выводил из того свойства
природы, которое называл типологической упорядоченностью (в
наших терминах — мероно-таксономический прогноз).
У всякой птицы мы ожидаем найти перья, у каждого организ-
ма — ДНК с четырехбуквенным кодом. И дело здесь не в происхож-
дении их от общего предка, о котором обычно ничего не известно,
а в том, что без этих меронов мы не в состоянии сконструировать
соответствующие архетипы. «Все современные птицы имеют перья,
а ископаемые, для которых перья неизвестны... немыслимы без
них»,— пишет зоолог А. С./Раутиан [1978, с. 109]. Аналогично,
единый архетип организма немыслим для нас без мерона «ДНК».
Однако, как и всякий прогноз, типологический прогноз может не
оправдаться: даже универсальность генетического кода у некото-
рых одноклеточных нарушена (см. гл. 6).
Итак, биологическому языку предложено нечто вроде граммати-
ки: словом в ней служит мерой, осмысленной фразой — архетип (и
соответствующий ему таксон), грамматическим правилом — реф-
рен, а связным текстом может служить или таксон более высокого
ранга, или экосистема, или, к примеру, последовательность сменя-
ющих друг друга видов — смотря о чем идет речь. Точнее, пока что
намечен аналог грамматической морфологии (теории построения
слова), а аналог синтаксиса, т. е. теорию связи меронов в архетип
(по Оуэну) еще предстоит создать. Здесь уместно вспомнить, что,
по В. Н. Беклемишеву [1964, с. 20], «разделение целого на части
и описание частей составляют предмет тектологии; синтез целого,
обратное его построение из частей, составляет предмет архитекто-
ники». Архитектоника и должна в будущем явиться искомым син-
таксисом.
2.3; Ядро и периферия
Зоолог Карл Бэр в 1825 г. говорил, что всякий крупный таксон име-
ет ядро из характерных форм (таковы, например, воробьи и сини-
цы среди воробьиных, кошки и собаки среди хищных) и перифе-
43
рию из сравнительно малочисленных форм, у которых облик дан-
ного таксона выражен нечетко, образуя ряд переходных форм,
сближающих этот таксон с другими [Бэр, 1959, с. 401—404]. Тако-
вы, например, ворона среди воробьиных и морской зверь калан сре-
ди хищных. О том же, что и Бэр, вскоре писал Максимович [1827,
с. 41], и в этом легко увидеть развитие идей Бюффона. В 1840 г. ло-
гик и историк науки Вильям Уэвелл сформулировал на этой основе
принцип классификации: «Класс установлен твердо, хотя и не ог-
раничен точно» (см.: [Мейен, 1978]). Напомнив это, вернемся к
«биологической грамматике».
Кювье недаром брался восстановить облик животного по не-
скольким зубам — одни свойства огранизмов влекут за собой дру-
гие, и понятие архетипа призвано выразить эту взаимосвязь. Дело
в том, что строки таблицы резко неравноправны: сравнительно не-
многие почти не содержат пустых клеток (признак присутствует
почти у всех особей) и характеризуют ядро таксона по Бэру, а в боль-
шей части строк пустые клетки преобладают — эти строки означа-
ют периферические признаки, т. е. признаки, характерные для пе-
риферических форм. Чтобы строить естественную систему, надо
разумно вырезать из большей таблицы меньшую, т. е. так выбирать
столбцы, чтобы у них было свое ядро (почти сплошь непустые
клетки в некоторых строках). Так, ядро класса птиц образуют ле-
тающие формы, имеющие крепкие маховые перья и киль (гребень
грудины), но ядро отряда страусов — нелетающие птицы без киля,
с мягкими декоративными перьями на месте маховых. И все же
страус останется среди птиц при любом способе классификации,
так как перья, клюв вместо зубов, цевка, способ прикрепления че-
репа к позвоночнику и многие другие его признаки принадлежат
классу птиц.
Возможность строить систему связана с тем, что разнообразию
свойственна типологическая упорядоченность по Мейену или, что
то же, с тем, что ядро обычно сильно превосходит размерами
периферию. Бэр пояснял это на классе млекопитающих, которые в
большинстве ходят на четырех ногах, мало или совсем не покидают
землю и голова их направлена вперед. Это — ядро класса, значи-
тельное большинство его видов, «собственно четвероногие», тогда
как периферийные виды могут быть похожи на рыб (киты), на
птиц (летучие мыши), двигаться вертикально (обезьяны) и т. д.
Добавим, впрочем, что изредка (у некоторых крупных таксонов)
ядро выявляется плохо, не превышает периферии. Тогда классифи-
кация оказывается затруднительной, независимо от подходов тех
или иных систематиков. Среди цветковых такие таксоны издавна
заслужили эпитет «crux botanicorum» (тяжкий крест ботаников),
например — порядок Salicales (ивовые), число видов которого ко-
леблется от 400 до 700 в различных системах, причем почти все они
приходятся на один род Salix (ива). Такой же «стих» являли собою
все бактерии, пока бактериологи не отказались от иерархических
принципов систематики. Однако для высших многоклеточных ядро
обычно таково, как говорил Бэр.
44
К ядру зверей Бэр отнес хищных, а к их ядру — кошек и собак.
Рассмотрим ныне живущие виды собак чуть подробнее, чтобы
выявить логику ядра и периферии. В семействе Canidae (псовые,
или собачьи) ядром («центральным родом») все признают род
Canis (собака), а ядром рода — вид Canis lupus (волк), но в осталь-
ном это семейство подвергается непрерывной перегруппировке
(хотя новых его представителей в природе не обнаруживают с
1914 г.). Причина — «промежуточные формы, которые не позво-
ляют проводить четких границ между ними, постулировать их дис-
кретность» [Волк, 1985, с. 21]. В 1880 г. Томас Гексли выявил
в семействе два параллельных ряда форм — Thooida (волкообраз-
ные) и Alopecoida (лисицеобразные), отличающиеся строением че-
репа и глаза (у волкообразных зрачок круглый, у лисицеобраз-
ных — вертикальный, эллиптический). В частности, у южноаме-
канских псовых есть и свои «волки» и свои «лисы», хотя эта груп-
па произошла, как все признают, от своего общего предка. Вывод
Гексли остался без внимания, и в начале XX в. установилась очень
дробная и вычурная классификация семейства: 54 его вида были
разделены на 4 рода, из которых 3 имели по 1 виду, зато род Canis
делился на 13 подродов, объединенных в 3 группы — волков, шака-
лов и лис [Hunde, 1908]. Затем число видов уменьшили до 35 —
37, но о структуре семейства до сих пор договориться не могут:
число родов то сокращают до 8, то увеличивают до 15.
Признаки видов семейства были изучены с помощью ЭВМ
[Glutton-Brock е. а., 1976] и оказалось, что волк (вместе с динго и
домашней собакой) — периферическая группа. Компактное ядро
семейства образовано двумя тесно сомкнутыми субъядрами —
группами лисиц и шакалов (к последней на некотором отдалении
примыкает волк). Ближе всего к середине пространства признаков
оказались две внешне сходные лисицы — тибетская песочная
(Vulpes ferrilata) и перуанская пустынная (Dusicyon sechurae).
Переоткрытие параллелизма не побудило авторов к существенному
пересмотру системы Canidae — они лишь уменьшили число родов
с 14 до 9. Характерно, что для перегруппировки пришлось привлечь
английские бытовые названия видов псовых. Однако в 1978 г. по-
явилась новая классификация. Ее автор использовал в основном те
же материалы, но повторил построения прошлого века почти без
вариаций. Он предложил вернуться к широкому пониманию рода
Canis — 8 подродов и 29 видов,— включающему упраздняемый им
род Vulpes. (Тем самым обыкновенной лисе предложено бинарное
название Canis vulpes, и мы возвратились на 220 лет назад,
к Линнею). Остальные 6 родов семейства вновь содержат по 1 виду
[Van Gelder, 1978]. Такое блуждание будет бесконечно, если не
задумываться о теоретической базе (итеративной процедуре).
Как видим, детализация исследований не привела ни к един-
ству мнений, ни к их устойчивости, ни даже к существенно новым
вариантам системы. Аналогичные примеры можно приводить из
всех областей систематики. Среди млекопитающих «нельзя, пожа-
луй, указать ни одной группы, которая не нуждалась бы в полной
45
ревизии» [Павлинов, Россолимо, 1987, с. 4]. И авторы этих слов
ревизуют: без объяснений ликвидировали отряд ластоногих, вклю-
чив его виды в качестве двух отдельных надсемейств в состав
«естественного» отряда хищных. («Естественность» авторы пони-
мают в весьма искусственном смысле, который носит название
кладистического и оценку которому дал в свое время Любищев
[1968]; он указал, в частности, что согласно кладизму надо бы от-
нести крокодилов к птицам, чего никто, разумеется, не делает).
Единственным поводом послужила ссылка на работу, отмечавшую
параллелизм моржей с псовыми, а тюленей — с куньими. Тот факт,
что с любой архетипической (целостной) точки зрения морж бли-
же к тюленю, чем к собаке, для авторов несуществен, и сходство
между ластоногими вообще в их «системе» не отражено. Как гово-
рил Любищев, отказ теоретизировать — худший вид теоретизи-
рования: привычка игнорировать параллелизмы приводит на деле
к полной зависимости исследователей от любого частного паралле-
лизма, к невозможности выявить ядра таксонов.
При перетасовках часто страдают бинарные названия видов:
пользуясь разными трудами, не всегда можно понять, об одинако-
вых или различных видах идет речь — если только не быть специ-
алистом именно по данному семейству. В отношении известных
животных и растений спасают, как ни парадоксально, их бытовые
названия, не меняющиеся столетиями,— вопреки расхожему тези-
су, что они вносят путаницу, устраняемую только ученой латынью.
(К сожалению, огромное большинство мелких организмов бытовых
названий не имеет). Так, виды фауны СССР, в прошлом
относимые к роду Canis, значатся в общей сложности под 16 родо-
выми названиями, хотя самих родов ни в один момент не насчи-
тывалось более пяти. Песец носил 4 родовых и 3 видовых на-
звания, причем нынешнее (Alopex lagopns) является комбинацией
из третьего по времени родового и первого видового. И здесь, как
видим,— блуждание вокруг старого.
Эти подсчеты оказались возможными потому, что недавно
впервые вышла сводка синонимов [Павлинов, Россолимо, 1987].
Она не содержит никаких описаний или объяснений (опять-таки
рассчитана только на узких специалистов), охватывает едва ли ты-
сячную долю всей мировой фауны и все-таки заняла 18 авторских
листов. Ясно, что подобная сводка для всех видов животных, ра-
стений, грибов и микробов просто немыслима, да и бессмысленна,
если не обуздать словотворчество натуралистов. Таксономическая
чехарда особенно недопустима сейчас, когда спасение экосистем
требует экологических обобщений, охватывающих самые разные
группы организмов и значительные промежутки времени.
В принципе задача стабилизации названий не так уж сложна,
и ответ дан в сущности самим Линнеем: не надо путать естествен-
ную систему с адресацией (см. 1.3). От адреса требуются просто-
та и постоянство, тогда как сам таксон может быть устроен сложно,
а взгляды систематиков на «естественность связей» видов в нем
могут то и дело меняться. Даже если допустить, что когда-нибудь
46
4 споры систематиков утихнут, то и тогда одной системой не обой-
I тись: в силу наличия ядра и периферии естественная система не
может быть удобна для адресации. Ведь даже нечеткие виды долж-
ны адресоваться четко. Методолог С. С. Розова пишет, что «удобное
для нахождения расположение материала» несовместимо в одной
системе с «выявлением внутренних системных отношений» [Тео-
рия и методология..., 1983, с. 14].
Сравнение таксонов с адресами обнаруживает факт, неизвест-
ный систематике организмов,— адрес не может заменить имя.
(Если человек, переехав на другое место жительства, меняет и имя,
то найти его крайне трудно.) Латинское название вида сможет слу-
жить ему именем только тогда, когда перестанет использовать-
ся в качестве адреса, т. е. если оно не будет меняться при переносе
вида в другой род. В отношении других таксонов систематика так
и делает: перенос рода в другое семейство, семейства — в другой
отряд и т. д. не обязано отражаться в их названиях. Бинарное ла-
тинское название удобно как имя вида, если не придавать совпа-
дению первых слов у разных видов таксономического значения.
Так, обыкновенную лису удобно навсегда назвать Canis vulpes L. *,
а ее принадлежность определенному роду обозначать отдельно.
Например, можно писать: вид Canis vulpes L. из рода Vulpes (точ-
но так же, как сейчас пишут: род Vulpes из семейства Canidae).
До Линнея это допускалось: Баугин мог, например, в составе рода
Plantago назвать один вид Plantaginella palustris (Подорожничек
болотный); он счел это более точным и выразительным, чем преж-
нее Plantago aquatica minima [Bauhin, 1623, с. 190]. Можно сде-
лать и по-другому: закрепить за словом, означающим род, опре-
деленное окончание (точно так же, как окончание -idae означает
семейство животных, а -асеае — семейство растений, грибов или
микробов), но не включать такие слова в имя вида. Это имя не
должно меняться, даже если вид будет признан подвидом или,
наоборот, разбит на несколько видов, как мы не перестаем называть
«динго» австралийскую дикую собаку, хотя идут споры о ее видо-
вом статусе.
В сущности, специалисты не так уж далеки от принятия такого
принципа. Так, в монографии, написанной по просьбе Междуна-
родной ассоциации таксономии, признано, что «система номенкла-
туры, в которой названия изменяются при изменении классифи-
кации, как справочное пособие имеет очевидные недостатки»
[Джеффри, 1980, с. 34]. В ее защиту приведен единственный аргу-
мент: общее родовое название у близких видов удобно, так как ука-
зывает на их большое сходство. Этот аргумент сохранит полную си-
лу и при навсегда закрепляемых бинарных названиях. Понятие не-
изменных названий уже существует в таксономии (nomen conser-
vandum), и осталось только договориться о применении его к имени
определенного организма (голотипа), а не только к названию
таксона.
* Символ L. означает, что это бинарное название дано Линнеем.
47
Отделив имя от адреса, легко обнаружить, что в центре споров
систематиков лежит именно вопрос о соотношении ядра и пери-
ферии таксона. Так, род Canis «в узком смысле» (волк-(-ближай-
шие 5 — 8 видов) это и есть ядро рода, понятого широко. А сам род
Canis «в широком смысле» — ядро семейства Canidae. (Те, кто хо-
тят выявить это ядро более четко, склонны вводить особый так-
сон — подсемейство Caninae.) Замечательно, что границы ядра
рода Canis все понимают одинаково, а спорят только о том, какие
формы внутри него считать отдельными видами. Поэтому мы мо-
жем уверенно записать строение рода в форме:
группа «lupus» (волк, домашняя собака, динго и подвиды вол-
ка, иногда выделяемые в виды) — внутреннее ядро рода Canis;
группа «thos» (койот, шакалы, включая эфиопского6) —
внешнее ядро рода Canis;
группа «dusicyon» (южноамериканские лисицы, что приблизи-
тельно соответствует роду Dusicyon — около 10 видов) — внутрен-
няя периферия рода;
остальные Caninae — внешняя периферия рода.
Аналогично можно очертить ядро и периферию семейства:
ядро — подсемейство Caninae, т. е. род Canis в его широком пони-
мании. Характерно, что Ван Гельдер, строя систему Canidae, как
раз и пользуется расстояниями видов (в признаковом пространст-
ве) от вида Canis lupus, т. е. описывает как бы слои ядра и перифе-
рии. Следует только оговориться, что волк в качестве центра неуда-
чен: центром семейства естественнее считать общую периферию
родов Canis (узкого) и Vulpes, где расположены тибетская и перу-
анская лисицы.
Тот факт, что периферии пересекаются, очень важен: если ядро,
как бы его ни понимать, может совпадать с каким-то таксоном, то
периферия таксоном быть не может, поскольку ее виды отличаются
друг от друга больше, чем от видов соответствующих ядер, и како-
го-либо своего ядра (а только оно и оправдывает введение таксона)
в ней найти не удается. К сожалению, иногда систематики собира-
ют периферические виды в недолговечные таксоны (см. гл. 6).
Важно и то, что между ядром и периферией нет четкой границы,
что, однако, по Уэвеллу, не лишает этих понятий строгости.
Ядро и периферия существуют объективно, а вовсе не являют-
ся следствием плохого знания объектов. Поэтому, например, един-
ственный способ определить прочно место енотовидной собаки со-
стоит в признании, что она — нечеткий род между семействами
псовых, енотовых и виверровых. А для адресации ее удобнее оста-
вить там, где она уже признана,— среди псовых, на их внешней
периферии.
И наконец, отметим, что ядро и периферия — не особенность
биологических таксонов, а общее явление природы, одно из прояв-
лений естественной типологической упорядоченности (по Мейену),
свойственной группам как материальных объектов, так и абстракт-
6 Canis siinensis. Он имеет ряд лисьих черт [Glutton-Brock е. а. 1976, с. 149], и его
иногда выделяют в особый подрод [Волк, 1985, с. 24].
48
ных понятий. Например, в Солнечной системе известны два прави-
ла, упорядочивающие' движения небесных тел. Первое — правило
планетных расстояний (закон Тициуса—Боде), которое гласит, что
по мере удаления от Солнца планетные орбиты располагаются всё
реже и реже и что так же располагаются вокруг планет орбиты по-
чти всех спутников 7. Второе правило — однонаправленность:
почти все тела вращаются вокруг своих осей и обращаются по ор-
битам в ту же сторону, в какую вращается Солнце. Хотя эти прави-
ла лежат в основе любой теоретической модели Солнечной системы,
ряд авторов отрицает их познавательную роль, поскольку не все
тела им подчинаются. В сущности, предмета для спора нет:
каждое правило имеет обширное ядро (тела, подчинающйеся ему с
достаточной точностью) и скромную периферию [Чайковский,
19876].
Другой пример являет система русских падежей. Обычно их
называют 6, с помощью которых упорядочивается подавляющее
большинство (ядро) именных словоформ, но есть еще как минимум
3 падежа. Так, например, звательный падеж соответствует форме
именительного (кто, что), но означает не подлежащее, а обраще-
ние; кроме архаических словоформ (Батюшки! Боже! Владыко!),
есть и вновь возникшие, используемые в устной речи специально
при обращении: бабусь, браток, Вась, земёля и т. п. Этот падеж
(как и другие архаизмы) нужен не только для понимания изрече-
ний вроде «Врачу, исцелися сам», но и для выявления парадигмы
системы склонения в широком смысле, т. е. ее архетипа по Мейену.
(Таблицу архетипа для четырех индоевропейских языков см.: [Бо-
городицкий, 1910]. Парадигмой склонения в узком смысле имену-
ют конкретный ряд вроде пол — стол, пола — стола..., т. е. реф-
рен.) Тем самым, если не ограничиваться вариациями «кто, что»,
а упорядочивать сами ряды словоформ, то выявляется периферия
падежной системы. (О других малозаметных русских падежах см.:
[Иванов, 1983, с. 268; Трубецкой, 1987, с. 72], а также: [Пялль и
др., 1962, с. 93].)
Наконец, сложный пример — гомология по Оуэну (1.4). Мало
кто принимал ее в полном объеме, да и Оуэн защищал ее скорее как
идею, чем как анатомический факт. Однако место ее вполне ясно,
если признать у этого понятия ядро — то, что вошло в учебники
(соответствие руки и ноги, руки и крыла, двух позвонков), и пери-
ферию — те примеры, о которых долго спорили и не договорились.
Они не могут быть поняты сами по себе, так как в каждом из них
мало от гомологии (плечевой пояс не похож на позвонок), но на-
ходят свое место в общем ряду гомологичных объектов, где на од-
ном конце — почти полная идентичность (два соседних позвонка),
а на другом — полное отсутствие сходств. Поскольку переходы в
7 Каждой орбите как бы соответствует свой дискретный энергетический уровень.
По одному уровню могут двигаться несколько мелких тел (пояс астероидов, тро-
янцы, некоторые спутники). После открытия у планет-гигантов систем колец и
связанных с ними мелких спутников правило можно уточнить: оно действует
снаружи от этих систем, т. е. вне пяти планетных радиусов. (Вблизи кольца
Солнца небесных тел попросту нет.)
49
этом ряду достаточно плавны, мы можем искать общий механизм,
их порождающий, понимая, что его действие в одних случаях доми-
нирует, в других — маскируется действием других механизмов,
а на пограничных объектах (в смысле границ таксона «гомоло-
гия») вообще исчезает.
2.4. Параллельные ряды
Параллельных рядов мы уже не раз касались, и в 2.2 были сфор-
мулированы три основных свойства параллелизма: он выражается
правилом «у разных таксонов — общие мероны», вызван невоз-
можностью использовать для классификации сразу все существен-
ные сходства объектов и фиксируется он наличием у разных рядов
общего для них рефрена. Теперь нам надо показать общебиологи-
ческую роль параллелизмов и как ими пользоваться.
Совокупность объектов образует ряд. если она обладает каким-
то общим свойством. Это свойство может быть либо заключенным
в каждом объекте (например: ряд стульев), либо задавать способ
построения ряда (например: ряд четных чисел 2, 4, 6...), либо со-
поставлять одну совокупность с другой (например: англо-русский
словарь). Параллелизм может наблюдаться между разными рядами
объектов. Поясним это на примерах, даваемых филлотаксисом (по-
рядком расположения листьев на стебле) *.
Листья могут быть собраны в одном месте стебля, образуя розет-
ку, или располагаться вдоль него регулярно — либо мутовками
(т. е. группами), либо поочередно. Если в мутовке только 2 листа,
то листья называются супротивными. Тип листорасположения
может быть общим свойством крупного таксона (напрмер, в се-
мействе губоцветных листья всегда мутовчаты и почти всегда су-
противны, а в семействе гвоздичных — почти всегда супротивны,
но иногда очередны), однако может меняться от вида к виду в одном
семействе и даже роде, а иногда — ив рамках одной особи (так.
у можжевельника в нижней части обычно очередное, а в верхней —
мутовчатое расположение). Изредка листорасположение бывает
беспорядочно (например, у женьшеня). Самое же интересное в
том, что очередные листья располагаются вдоль стебля по спирали
(вообще спиральная симметрия растениям свойственна во многом)
через равные промежутки, так что между двумя листьями, распо-
ложенными на одной вертикали, помещается одно и то же число
листьев, характерное для данного вида. Спиральное расположе-
ние свойственно, например, всему семейству злаков, причем у них
всех (кроме одного австралийского рода) на виток спирали прихо-
дится два листа.
Здесь естественно выявляются разные типы параллельных ря-
дов. Так. для большинства губоцветных и валериановых типично
супротивное расположение листьев, хотя эти семейства принадле-
жат разным порядкам. Это — пример параллельных рядов с общим
свойством.
* Литературу можно найти по книге С. В. Петухова [1981].
50
Бук, вяз, липа, рожь и некоторые другие растения имеют по 2
листа на виток, хотя далеки таксономически. Столь же далеки друг
от друга виноград, ольха, орешник, осока и другие растения, имею-
щие по 3 листа на виток. Многие растения имеют по 5 листьев на 2
витка (вишня, дуб, слива и др.) и по 8 на 3 витка (барбарис, мали-
на, тополь и др.) — это, по-видимому, самые частые типы спира-
лей. Гораздо реже встречаются 13 листьев на 5 витков (миндаль,
облепиха) и, как исключение, известны примеры, когда 21 лист
приходится на 8 витков (Linimi регеппе — лен многолетний). По-
следовательность дробей
«1=2/1, а2=3/1, й3=5/2, «4=8/3, «5=13/5, «6=21/8 ...(*),
где числители и знаменатели образованы числами Фибоначчи
(каждое число — сумма двух предыдущих), сходится ко квадрату
золотого сечения [Петухов, 1981, с. 12]. Дальнейшие члены (я?=
=34/13 и т. д.) на листьях просто не удается измерить, так как со-
седние члены (*) слишком мало различаются, но, например, на
шишках они известны.
Сказанное дает возможность указать параллельные ряды, пост-
роенные по общему закону. Так, представители порядка буко-
вых — бук («|), ольха («2) и дуб («з) имеют с розоцветными —
вишня (аз), малина («4) и миндаль («5) — то общее, что являются
членами последовательности (*). Заметим, что сама эта последова-
тельность — хороший пример рефрена.
Чаще, однако, рефрен не удается выразить в виде математиче-
ской формулы, и приходится описывать его путем непосредствен-
ного сопоставления параллельных рядов. Так, если рассмотреть
два порядка цветковых — норичниковые и сложноцветные, то в
них обоих обнаружатся разные формы как мутовчатых, так и оче-
редных расположений, и. кроме того, есть розеточные формы (по-
дорожники, чертополох бесстебельный). Тем самым, можно постро-
ить два параллельных ряда.
Именно так. непосредственно сопоставлением,' построены и
ряды Вавилова. В каждом таксоне Вавилов различал радикал, т. е.
совокупность свойств, его определяющих, и переменную часть, т. е.
свойства, различные у разных представителей таксона. Если груп-
пировать таксоны по близким радикалам, то получится обычная
систематика, но их можно сближать и по сходным переменным ча-
стям. Два вида одного рода Вавилов представлял формулой
(а—]—6—[—с—]—...), Л2(я+б-|-с-[-...),
где L: — видовые радикалы, а а, Ь, с... — общая для них перемен-
ная часть. Недостаток одного из слагаемых в одном из рядов позво-
лял Вавилову успешно искать новые формы растений, но это было
лишь начало пути. Мейен предложил (развивая мысли Кренке и
Урманцева) более общее понимание параллелизма: дело здесь не
столько в повторности самих признаков, сколько в повторности
«законов преобразования» этих признаков. Другими словами, Мей-
51
ен предложил расматривать не столько те простейшие случаи,
когда а, Ь. с — признаки конкретных особей, сколько более общую
ситуацию, когда a={ai, «г---} — последовательность признаков,
связанных некоторой постепенностью переходов, причем у кон-
кретной особи может проявляться один или несколько признаков ц.
Примером может служить последовательность листорасполо-
жений (*): обычно растение данного вида имеет один тип а» но
иногда (например, у можжевельника) в разных частях растения
наблюдаются разные типы. Другие примеры (расчленения листа,
жилкования листа и др.) приведены у Мейена [Меуеп, 1973]; здесь
они могут быть записаны как b={bi,b2,...} с=={с;, С2,...} и т. д.
Естественно, что прогнозировать недостающий признак а. по на-
личному ряду а можно гораздо увереннее, чем у Вавилова, где по
признакам а и b прогнозировался с. Однако нам здесь важнее дру-
гое: радикал-рефренная структура таксона допускает эффектив-
ную свертку (компактное описание) информации о нем, дает воз-
можность увидеть в нем систему, а не случайный набор.
Таблица 3. Пример морфологического межклассового рефрена животных (после-
довательность основных состояний мерона «парная конечность» — для всех
главных классов позвоночных)
класс значения мерона
отсутст- вие или невидимый снаружи зачаток развита только слабая передняя пара суставча- тые плав- ники или ласты орган для
полза- ния планирую- щего полета активного полета
костные рыбы мурена и др. китовид- на, угри и др. кистеперые морской нетопырь летучие рыбы клинобрю- шка
амфибии червяга сирен пек. ископа- емые батра- хоморфы саламан- дры и др. яванская лягушка
рептилии змеи и безногие ящерицы хирот морские черепахи и др. черепа- хи, кро- кодилы И др. ногокрыл (ископа- емый) летающие ящеры (ис- копаемые)
птицы киви (нет передней пары) — пингвины некото- рые ви- ды * СОВИНЫЙ попугай И др. большин- ство видов
звери киты, ла- мантин, дюгонь (нет задней па- ры) киты ламантин, дюгонь (передние) калан (задние) кроты, тюлени и др. шерстокрыл рукокры- лые (лету- чие мыши И др.)
* Ползание, т. е. перемещение по субстрату с касанием его брюхом, имеет у птиц различные
формы. Пингвины часто скользят брюхом по снегу, хотя обычно ходят на лапах.
Очень коротконогие длиннокрылые птицы (стрижи, фрегаты), случайно оказавшись
на ровной поверхности, могут лишь ползти в поисках уступа. Перемещение лазающих
птиц по веткам, стволам, стенам и т. д. тоже иногда называют ползанием (попугаи,
клесты и др.), причем «поползень... в ползании превосходит всех прочих птиц» [Брем,
Россмесслер, 1867, с. 407].
52
Поясним сказанное тем же примером парных конечностей по-
звоночных, о котором шла речь в 2.2. Сведем в таблицу 3 то, что
касается изменения конечности от полного отсутствия до пригодно-
сти .к активному полету. Для пяти классов позвоночных получим
5 параллельных рядов, которые мало соответствуют последователь-
ности исторических событий, но именно этим и интересны. Киты
не произошли прямо от рыб, тем более не произошли они от рыб,
лишенных задних (брюшных) плавников, но кит до того похож
на одну мелкую глубоководную рыбку, что ее так и называют —
китовидка (Cetomimus). Еще более похожи друг на друга хвостатая
амфибия сирен и ящерица хирдт, равно как безногая амфибия
червяга похожа на змею.
При этом наблюдается сходство целых комплексов свойств.
Так, китовидка — черная, с голой кожей, огромными головой и
ртом, с малюсенькими глазками, не имеет брюшных плавников.
Назвать сходство кита и китовидки независимым приспособле-
нием — значит не сказать ничего, поскольку условия их обитания
различны предельно. Аналогично, нет смысла толковать как неза-
висимое приспособление крохотные, но вполне развитые «ручон-
ки» сирена и хирота: другие виды в тех же условиях несут по
4 крохотных лапки или вообще безноги. Зато и здесь налицо
целый комплекс сходств, в том числе неожиданных: и сирены,
и хироты живут только на юге Северной Америки, несут по 3—
4 (сирены) и по 3—5 (хироты) пальцев на лапке и составляют
семейства по три вида в двух родах каждое. (Правда, сейчас
хиротов отдельным семейством не считают, но это, естественно,
не умаляет их параллелизма с сиренами, а является лишь одним
из примеров той таксономической нестабильности, о которой мы
говорили в 2.3). Словом, сравнение сиренов и хиротов заслуживает
особого исследования: сам феномен — мелкие, слабые, но вполне
развитые передние конечности при отсутствии задних — назовем
хиротностыо.
Вывод из этого следует тот, что кроме сходств, порождаемых
общностью происхождения и функций, есть еще сходства чисто
диатропической природы. Обычно их связывают с еще неизвест-
ными законами формообразования, но такую догадку следует при-
нимать с одной оговоркой: речь идет здесь не о путях, клеточной
дифференцировки, а об общих законах построения форм. Эту
разницу Мейен любил пояснять таким примером: листья высших
растений демонстрируют те же ряды форм, что и талломы сифо-
новых водорослей, хотя последние не разделены на отдельные
клетки. Спектр возможных вариантов формообразования достаточ-
но широк, чтобы обеспечить наблюдаемое разнообразие, но все же
он ограничен, и это выражается в параллелизмах. Если этого не
учитывать, то неизбежны курьезы вроде упомянутого в 2.3 исчез-
новения ластоногих из системы млекопитающих.
Всеобщность феномена параллелизма привела Мейена к
уверенности, что делить свойства таксона на радикал и рефрен
можно лишь условно, так как признак, входящий в радикал.
53
столь же изменчив в смысле диапазона возможных изменений,
но только их гораздо реже удается наблюдать. Это различие частот
носит качественный характер и дает возможность практически
выявлять радикал, т. е. обеспечивает «естественную типологиче-
скую упорядоченность», как ее понимал Мейен. В качестве приме-
ра изменчивости радикала он напоминал давние указания Д. Н. Со-
болева, который отмечал, например: «Аномальное увеличение
числа частей также может стать нормальным отличительным
признаком какой-либо группы ... У коров, овец и коз в виде ано-
малии вместо одной пары рогов появляются иногда две пары...
и если бы этот признак сделался стойким, то возникла бы четырех-
рогая порода вроде азиатской четерехрогой антилопы (Tetra-
ceros cfiiadricorriis). которая, можно думать, так и возникла»
[Соболев. 1924. с. 161]. Количество рогов входит в радикал, поэто-
му четырехрогость явно нарушает радикал копытных. Столь же
явным нарушением радикала червей является наличие двух лен-
товидных тел при одной головке у Triplotaenia mirabilis [там же,
с. 162]. Редкость таких фактов не снимает вопроса о путях и при-
чинах их появления, точно так же, как редкость алмаза не делает
его менее важным и интересным для кристаллографии, чем графит
[Меуеп, 1973. с. 206, 233]. Редкие варианты указывают на рамки
возможного, что впервые использовал для эволюционной схемы
Мопертюи (1756) и что ныне выступает в виде четкого терато-
таксономического параллелизма: то. что водном таксоне выступает
как уродство, в другом является нормой.
Выявление принципиальных рамок формообразования приво-
дит к возможности эволюционного прогноза — только не в том
смысле, какой обычно вкладывают в этот термин. Появляющиеся
изредка в литературе фантастические изображения организмов
будущего (человек с огромной головой на недоразвитом теле и т. п.)
вряд ли представляют научный интерес, но то внимание, которое
им уделяет критика, свидетельствует о некоем прогностическом
вакууме, вызывающем беспокойство, пусть и неосознанное, эволю-
ционистов. Ведь способность к прогнозу обычно приводится в
качестве основного критерия работоспособности теории. Поэтому
важно, что анализ рефренов и образуемых ими параллельных
рядов позволяет отсекать некоторые мысленные варианты эволю-
ции как практически невозможные. А это — первое условие для
возможности прогнозировать будущее развитие природы. Так,
мы можем уверенно сказать, что невозможен «ангел» (позвоночное
с третьим поясом конечностей), поскольку эта тенденция в соответ-
ствующих параллельных рядах отсутствует нацело. Однако иногда,
очень редко, мы в подобных негативных прогнозах все же будем
ошибаться. Например, способность к планирующему полету у
позвоночных обеспечивается конечностями (иногда и перепонками
между ними — ногокрыл, шерстокрыл) и хвостом, а другой воз-
можности, казалось бы, нет. Однако в действительности существует
род летучих ящериц (Draco), у которых «планер» натянут на
выростах ребер. Запрет, казалось бы наложенный рефреном мерона
54
«конечность», преодолен в рамках другого параллелизма, о кото-
ром мы скажем в 2.5. Подобные казусы не должны обескураживать,
так как сопровождают прогнозы во всех науках.
Наконец, коснемся кратко множественного параллелизма.
Он имеет место между параллельными рядами А, В, С, D,... тогда,
когда один и тот же рефрен связывает любые два ряда (А, В),
(А, С), (A, D), (В, С) и т. д.; т. е. когда можно говорить о параллели
между параллелизмами. В 1857 г. геолог и зоолог Луи Агассиц опи-
сал 5 типов таких рядов: 1) геологическая последовательность
появления организмов, 2) их таксономическое положение, 3) их эм-
бриональное развитие, 4) их «структурная градация» (т. е. слож-
ность строения, о которой мы говорили в 1.4) и 5) их географиче-
ское распространение [Agassiz, 1857]. И сами ряды, и их сходства
между собой были обоснованы здесь далеко не одинаково (что
отметил сам автор), и в литературу вошло лишь обыкновение
сопоставлять первый, третий и четвертый ряды, получившее (после
«Общей морфологии» Геккеля) название метода тройного парал-
лелизма.
До Агассица «теорией параллелизма» называлось сопоставле-
ние реально наблюдаемых сходств зародышей высших животных
со взрослыми формами низших и, на этой основе, гипотетический
вывод в пользу идеи эволюции организмов. Вот что писал об этом
Агассиц: «Столь поразительно сходство молоди высших животных
со взрослыми индивидами низших типов, что многими авторами
было допущено, что все высшие животные проходят, в ходе своего
роста, через фазы, соответствующие постоянной конституции
низших классов. Эти допущения, будучи результатом несовер-
шенных исследований, даже стали основой системы философии
Природы, которая представляет всех животных как различные сте-
пени развития нескольких примитивных типов» [там же, с. 118].
В своем отрицании примитивного эволюционизма Агассиц, фор-
мально говоря, был прав, так как общность законов развития сама
по себе не указывает на общность происхождения, но он, против-
ник самой идеи эволюции, был бы, наверное, сильно удивлен,
если бы узнал, что именно множественный параллелизм, им откры-
тый, дал впоследствии первое серьезное доказательство эволюции.
Это произошло через сто лет, когда данные по нынешним и вымер-
шим таксонам были спроецированы на палеогеографическую карту
(см. 7.3).
Замечательно и то, что обратиться к единой схеме параллелизма
Агассица побудила междисциплинарная параллель — возмож-
ность представить правило планетных расстояний (о котором шла
речь выше) той же последовательностью (*), что и филлотаксис.
Для него эта параллель была свидетельством божественного
замысла [там же, с. 130], для нас же — общности системных
законов.
Наиболее наглядный пример множественного параллелизма
дают те же падежные парадигмы. Родственные слова могут скло-
няться совсем по-разному (например, дочь и дочка) или даже
55
относиться к различным архетипам склонения (стул и немецкое der
Stuhl), зато параллелизмы (пол — стол и т. д.) являют собой
самое тесное сходство склонений. Однако главный интерес пред-
ставляют параллели между параллелизмами. Во-первых, все
русские слова склоняются по единому архетипу, ядро которого
шестипадежно (см. 2.3); но, во-вторых, для уяснения его струк-
туры надо знать, что в основе его лежит древняя (санскритская)
восьмипадежная парадигма [Богородицкий, 1910], общая для
славянских (кроме болгарского, где падежи почти утеряны)
и балтийских языков индоевропейской семьи. Парадигма здесь
поразительно устойчива (хотя отдельные падежи в отдельных
языках утрачены), что не мешает ей проявлять параллели с языка-
ми других семей (алтайской, угро-финской и др.).
Например, в самом устойчивом языке семьи — литовском,
наряду с обычными падежами в некоторых диалектах встречаются
множественные локативы — падежи, основанные не на вопросах
что, чего, чему..., а на вопросах где, куда, откуда', для их описания
потребовались термины угро-финской лингвистики [Грамматика...,
1985, с. 90]. В русском языке что-то похожее видно в рамках
родительного падежа. В нем слились функции двух древних
падежей — собственно генитива, означавшего принадлежность
(вкус чая), и аблатива, означавшего отъединение (отказался от
чая), но кроме того, он выполняет функцию партитива, или частич-
ного падежа (немного чаю), свойственного, например, финскому
языку.
Как и биологи, лингвисты склонны объяснять подобные сход-
ства ссылкой на общее происхождение и заимствование (так ска-
зать, скрещивание); казалось бы, ссылка носит эволюционный
характер, но она весьма туманна как раз в эволюционном смысле.
Почему диалекты воспроизвели не иноязычные окончания, а сам
локативный принцип, чуждый данной семье? Почему это произош-
ло в зоне весьма слабых межсемейных влияний? Почему пери-
ферийные падежи в литературном русском языке путаются
(на столе, но на. полу) и в то же время выявляются четко в диалек-
тах [Иванов, 1983, § 170]? Последнее естественно, если считать
периферийные падежи вымирающими, но Н. С. Трубецкой [1987,
с. 214] характеризовал их как новации. Эти и подобные вопросы
аналогичны биологическим и носят отчетливо диатропический
характер: всюду речь идет о рефренах, частых в одних таксонах,
но редких в других 8. Историческое движение вдоль данного рефре-
на возможно в любую сторону. Параллели с далекими языками —
это именно параллели, подчас возникающие как безо всякого
контакта их носителей, так и вне родственных связей. Допускать
8 Точнее, рефрены меронов «индоевропейский падеж» и «угро-финский падеж»
совпадают отдельными значениями в отдельных таксонах (языках). Совпадения
часты, если принадлежат ядрам таксонов (эстонские падежи терминатив и
абессив соответствуют русскому родительному: до чая и без чая [Пялль и др.,
1962, с. 95]) и редки, если принадлежат перифериям или ядру и периферии (упо-
мянутые примеры диалектов).
56
сохранение диалектов с праиндоевропейских времен столь же
абсурдно, как допускать гендвухголовости, унаследованный всеми
животными от безголового предка. Вернее сказать, что и в лингви-
стике, и в биологии мы видим феномен транзитивного полимор-
физма, эволюционного феномена, о котором речь пойдет в 7.3.
2.5. Другие тенденции
Обычно за рефреном (правилом преобразования мерона) стоит
какая-то тенденция (массовая, не всегда четкая, но в каком-либо
смысле бесспорная закономерность), хотя в рамках того же рефре-
на часто есть нечто далеко не массовое и даже совсем редкое. Так,
рефрен, обрисованный таблицей 3, демонстрирует тенденцию
превращения лап в ласты и в крылья, но он же дает примеры ред-
костей — элементы активного полета у рыб, хиротность (отсут-
ствие задних конечностей при наличии передних) у некоторых
амфибий и ящериц; эти редкости в совокупности также образуют
тенденцию — редкое в одном таксоне нормально в другом (правило
Кренке). А пустые клетки таблицы свидетельствуют о возмож-
ностях, вообще не реализованных известными на сегодня организ-
мами. Для упорядочения разнообразия надо у каждого рефрена
выявить по возможности все тенденции. Сделаем это для рефрена
таблицы 3.
В 1957 г. Брони сформулировал «закон террипетного развития»
(le loi du developpement terripete), т. e. развития, устремленного к
земле. Согласно Бронну, морфологический тип «стоит тем выше,
чем более преобладают в нем наземные классы и отряды» (подроб-
нее см. [Чайковский, 1984]). Более того, по мере перехода от
типично морских к типично наземным организмам внутри крупно-
го таксона тоже наблюдается усложнение строения организмов.
В этом состоит террипетная тенденция организации разнообра-
зия. В частности, она видна на конечностях: всякое возвращение
в воду (превращение конечностей в ласты и плавники) приводит
к упрощению внешней морфологии конечностей, а выход из
воды — к ее усложнению. Таким образом, террипетная тенденция
обратима. Продолжая мысль Бронна, можно заметить, что переход
с суши в болото и воду, с поверхности под землю, из толщи вод на
дно — все это вызывает редукцию (упрощение, уменьшение, утра-
ту) конечностей, преимущественно задних; если остается одна
пара конечностей, то передняя. Среди рыб мы видим это у угрей
(зарываются в дно), у водных форм, принадлежащих наземным
классам,— у ихтиозавров, китов, ламантинов. Очень характерен
и калан: у этого прибрежного морского зверя спереди еще лапы,
а сзади уже ласты. Хотя у современных амфибий и рептилий
ласт нет (кроме некоторых черепах), однако тенденция ясно видна
и у них: у подземных, болотных и водных форм уменьшаются или
утрачиваются лапы. В обоих классах есть небольшие группы
(хвостатые амфибии сирены и ящерицы хироты), сохранившие
крошечные, но вполне развитые передние лапки. Объяснить их как
57
приспособление никто и не пытается, так что рефрен можно в
этой его части уверенно трактовать как чисто диатропический, а не
экофизиологический феномен. Донный угорь, глубоководная кито-
видка, болотный сирен, подземный хирот, стремительный морской
хищник дельфин, неповоротливый прибрежный ламантин и поеда-
тель планктона гренландский кит — все они встали в один ряд,
реализовав в рефрене мерона «конечность» состояние «хирот-
ность». Это состояние расположено на периферии всех пяти клас-
сов, но может характеризовать ядра меньших таксонов (например,
отряда китообразных). Будучи редким в двух классах (амфибий и
рептилий), это состояние характеризует в рамках всего подтипа
позвоночных ясную тенденцию (входящую как часть в террипет-
ную тенденцию).
Оказывается, что разнообразие конечностей позвоночных скла-
дывается за счет наложения небольшого числа таких тенденций
разного уровня. Обычно они проявляются совместно и создают
впечатление невозможности упорядочить разнообразие, однако их
в принципе можно выявить в качестве отдельных. В частности,
для конечностей характерна тенденция к упрощению строения их
скелета по мере усложения их функций: «Пятипалая конечность
развивалась не путем усложнения, а путем упрощения из более
богато расчлененной конечности», так что нормальная пятипалая
конечность — итог развития из 7 — 10-лучевой структуры кистепе-
рых рыб, причем и у нынешних наземных форм наблюдаются
«остатки лишних лучей — элементы, лежащие впереди первого
пальца... и элементы, лежащие позади пятого пальца» [Шмаль-
гаузен, 1938, с. 170—172]. В этом можно видеть «концентрацию»
в смысле Бронна (см. 1.4) и «олигомеризацию» в смысле В. А. До-
геля (см.: [Канаев, 1966]). Существенно, что обратное превраще-
ние лапы в ласт (упрощение функции и внешней морфологии)
вновь увеличивает число фаланг пальцев, а иногда и число самих
пальцев (ихтиозавры), т. е. эта тенденция тоже обратима.
Не включаясь в давний спор о том, насколько прогрессивна
олигомеризация сама по себе, отметим только, что это — само-
стоятельная тенденция, не имеющая отношения к террипетной
(при которой преимущественно сохраняется передняя пара).
Олигомеризация может заходить так далеко, что будет уже озна-
чать явный морфофизиологический регресс — например, полная
редукция ног (а заодно — органов чувств и т. д.) у многих реп-
тилий; и при этом редукция задней пары отстает, так что един-
ственная рудиментарная пара здесь всегда — задняя. Здесь есть свои
закономерности. В частности, порядок утраты в точности противо-
положен тому, по какому шло эволюционное приобретение со-
ответствующих структур [Северцов, 1945, с. 474, 496]. Характерно,
что А. Н. Северцов, отказавшись объяснить этот класс фактов по
Ламарку или по Дарвину, т. е. как чем-то полезных организмам,
ограничился выводом, что редукция здесь является самостоятель-
ным «процессом, который, однажды начавшись, ведет к-полному
исчезновению редуцирующегося органа» [там же, с. 500]; тем
58
самым он констатировал тенденцию, и притом необратимую. Заме-
чательно, что «функционирующие органы, несмотря на то, что они
исправно функционируют, все же зачастую редуцируются» [там
же, с-. 498].
Заметим на будущее (см. 4.4), что Любищев называл такую
редукцию рутипизацией (утерей способности к новизне) и видел
причину ее в однообразности (рутинности) использования органа.
В качестве примера он приводил сохранение на ноге лошади лишь
одного пальца: это называют приспособлением к бегу, но ведь
другие животные бегают подчас не хуже на обычных лапах, зато
остальные функции лапы копыто выполнять не может.
Третья тенденция исследуемого рефрена проявляется в том,
что в ходе эволюции наземных позвоночных регулярно наблюдался
подъем головы и связанный с этим перенос тяжести тела на заднюю
пару конечностей. Это приводило к появлению прямосидения,
прямохождения, а затем и летания (при этом, в частности, репти-
лии эволюционировали в птиц). Тенденция видна еще у лягушки,
но лучше всего выражена у птиц и у человека. В данном процессе
неотъемлемой частью являлась, как ни странно, необратимая
редукция. Так, первоптица (археоптерикс) имела зубы и свобод-
ные пальцы на крыльях (которые тем самым могли выполнять
некоторые функции лап), но птицы утратили эти свойства и притом
необратимо: утеря способности к полету не возвращает крылу
свойств лапы, а наоборот — завершает редукцию крыла.
Комбинацией этих трех тенденций — террипетной, олиго-
полимеризационой и подъема головы — разнообразие конечностей
позвоночных, по-видимому, в основном и упорядочивается. Каждой
тенденции, когда она доминирует, соответствует свой пример
нулевого значения мерона (утраты органа): первой — сирен
(нет даже следов тазового пояса, но есть передние лапки), вто-
рой — змеи (нет даже следов плечевого пояса, но часты рудименты
задних лап), третьей — киви (нет передних конечностей, но есть
неуклюжие ноги).
Кроме трех тенденций, общих для позвоночных, есть еще част-
ные тенденции, выявляемые в рамках рефренов меньших таксонов.
Так, мерой «плавники костистых рыб» проявляет югулярную
тенденцию — по мере снижения функциональной значимости
брюшных плавников они сдвигаются вперед, вплоть до горла
(то есть вперед от грудных), вырождаются (в декоративные лучи
и т. п.) и подчас исчезают. Линней выделил горлопёрых (Jngiila-
res) в особую группу, а позже югулярность иногда рассматривали
как параллелизм [Бобрецкий, 1891]. Тем самым исчезновение
брюшных плавников может означать как отсутствие террипетно-
сти, так и последнюю ступень югулярности, смотря в какой мор-
фологический ряд ставится данный вид.
Югулярная тенденция замещает у рыб тенденцию подъема
головы и отчасти противоположна ей: плавание головой вверх
освоили многие рыбы (вогмер, морской конек, сельдяной король
и др.), но брюшные плавники при этом не развиваются, а наоборот,
59
югулируют. Югулярности места нет у всех активно ползающих
рыб, даже у тех видов, которые не пользуются при ползании
брюшными плавниками. Причина этого сохранения бесполезных
органов видится мне в том, что активное дижение по поверхности
являет (в сравнении с пассивностью и с зарыванием в субстрат)
террипетность, а она связана с развитием конечностей. Если так,
то переход рыб к ползанию означает не специальное приспособле-
ние к какой-то экологической нише, а преобладание террипетной
тенденции над югулярной. Самые неуклюжие из ползающих рыб,
разучившись плавать, ничего еще не получили взамен, например,
морской нетопырь Ogcocephalus (бывший Malthe).
Разнообразие любых объектов только кажется бесконечным,
а в действительности часто видно, как оно довольно просто упоря-
дочено и образуется за счет комбинаций небольшого числа тенден-
ций. Приведу пример из психологии, который понадобится нам в
главе 12. Это шесть тенденций, или типов мотиваций (жизненных
установок), которые ввел в науку немецкий психолог Эдуард
Шпрангер [Spranger, 1914]: теоретический — стремление к позна-
нию, экономический — стремление к выгоде, эстетический —
стремление к впечатлению и самовыражению, социальный —
стремление к общению и любви, политический (я бы назвал его
чиновным) — желание подчинять и подчиняться, религиозный —
поиск высшего смысла жизни. У отдельного человека обычно пре-
обладает один-два типа мотивации, но никогда не бывает, чтобы
присутствовал лишь один. Можно спорить о том, насколько пере-
чень полон (я бы ввел седьмой тип: животный — стремление
сохранить себя и потомство, но кто-то может сказать, что само-
сохранение — частный случай выгоды, а для кого-то забота о
потомстве — высший смысл жизни), однако несомненно, что
тенденции очень удобны для ориентировки в разнообразии. Ста-
новится ясным, например, что политэкономия описывает не само
поведение людей, а лишь те его аспекты, в которых доминирует
второй тип мотивации (подробнее см.: [Чайковский, 1988в]).
Желая дать определение царствам животных и растений, фило-
соф Анри Бергсон пришел к выводу, что нет не только ни одного
признака, но и ни одного комплекса признаков, позволяющего
четко различить царства, зато указал на четко выраженные у них
«тенденции к развитию» [Бергсон, 1909, с. 94]. Например, для
растений характерно усвоение углерода из атмосферы, тогда как
хищничество (хотя и представлено насекомоядными растениями)
не получило здесь развития, будучи животной тенденцией. Опреде-
ление таксона через его тенденцию основано на выявлении ядра
таксона. Мы воспользуемся этим в главе 6.
Качественная тенденция, допускающая исключения, играет в
описательных науках ту же роль, какую играет математически
выразимый закон в науках точных. Более того, к ней приходится
прибегать и в самых, казалось бы, точных науках — в тех случаях,
когда закономерность налицо, но носит нечеткий характер. Таковы,
например, три качественные тенденции в организации разнообра-
60
зия планет и спутников Солнечной системы [Чайковский, 1985;
19876]. В этом отношении описательные науки дают урок точным.
В частности, закон больших чисел (3.6) является тенденцией, что
важно для понимания философских оснований теории вероятно-
стей: вероятностные законы нечетки не только тем, что выпол-
няются лишь на массовом материале, но и тем, что даже на нем
выполняются почти всегда, а не всегда. Для традиционной мате-
матики это — изъян, который она пытается, не всегда удачно,
преодолеть (анализ см.: [Чайковский, 1989а]), диатропика же ищет
качественные методы работы с подобными объектами как тако-
выми.
Тенденция может проявляться не только во времени. Так, для
последних 100 млн лет эволюции характерно наступление цвет-
ковых растений, но в северной флоре до сих пор господствует
хвойный лес. Это — одна из тенденций в пространстве. Другая
состоит в том, что разные группы обладают различными преиму-
щественными способами освоения новых зон обитания. Например,
птицы освоили водную среду не так, как звери. С водой связаны
сотни видов птиц: из 33 отрядов (более 8600 видов) 5 — чисто
водные (170 видов) и 4 — водные и околоводные (200 видов),
но только 1 отряд (состоящий из одного семейства пингвинов —
17 видов) обладает подлинно водным обликом (сигарообразное тело
и ласты). В прошлом сходным обликом обладала еще бескрылая
гагарка. В других же отрядах собственно водных форм нет, и более
того, бывают водные виды, совсем лишенные характерных водных
черт строения. Самый яркий пример (его любил повторять Люби-
щев): оляпка (водяной воробей) плавает, ныряет и бегает по дну,
не имея даже перепонок на пальцах. Почти так же удивителен
зимородок. Совсем другую картину мы видим у плацентарных
млекопитающих [Томилин, 1977, с. 141 — 143]: здесь имеются три
водных отряда — китообразные, ластоногие и сиренообразные
(к ним относится ламантин) — причем все их 108 видов обладают
водным обликом. Всего же плацентарных меньше, чем птиц:
около 4 тысяч видов в 17 отрядах. Даже отдельные представители
неводных отрядов (например, калан из хищных) имеют водные
черты строения. Только у белого медведя перемена в строении
сильно отстает от образа жизни. Очевидна тенденция: птицы легче
других подстраивают поведение (вспомним, что только птиц можно
обучить имитации человеческой речи), но довольно жестко ограни-
чены в изменении строения. Плацентарные, наоборот, легко изме-
няют как строение, так и привычки. Этим они резко отличаются
от сумчатых, у которых только 1 вид из 250 — водяной опоссум —
связан, и то частично, с водой.
Змее летать не на чем, да и в истории всего отряда змей ничто не
говорит о такой возможности. Однако и летающие змеи бывают —
в Индонезии. Там кто только не летает! Мало того, что там летает
самое крупное из рукокрылых млекопитающих (калонг — размах
крыльев до 1,5 м), но только там летают амфибии и рептилии
(последние, так сказать, «залетают» частично и на материк).
61
О «летающем драконе» (ящерице) мы уже говорили, но в Индо-
незии летает и другая ящерица (из гекконов) — на перепонках,
тянущихся от носа до хвоста и захватывающих обе пары лап; при
этом, как бы специально для иллюстрации параллелизма, точно
так же летает (и только здесь) млекопитающее — шерстокрыл.
Наконец, здесь летает лягушка (растопыривая огромные лапы с
перепонками) — единственная летающая амфибия вообще. После
такого перечня как-то не столь удивительно узнать, что несколько
видов древесных ужей совершают здесь 10-метровые прыжки-
перелеты между ветвями, а для одного вида известно даже, как он
добивается устойчивости в полете: растопыривая костные щитки и
втягивая живот, он превращается в нечто вроде уголковой балки.
Почему все они летают (точнее — планируют) именно здесь
(в далеком прошлом рептилии планировали повсюду), никто
не знает, но поразительно, что в сходной по климату зоне Цен-
тральной Америки вместо страсти к полету царит страсть висеть на
хвосте - здесь цепкохвосты и амфибии, и рептилии, и звери. Того
же сорта явление (хоть и помельче) — «хиротность» амфибий и
рептилий на юге Северной Америки, о которой мы уже говорили.
Этот географический стиль (термин Любищева) пусть и непоня-
тен в привычных терминах приспособления, но самый факт нали-
чия ряда свидетельствует о тенденции. Таких рядов можно указать
много.
Феномен географического стиля характерен не только для
биологии. Так, для языков Европы характерна тенденция к редук-
ции падежной системы с востока на запад, причем она захватывает
языки разных семей с очень разными структурами падежной
парадигмы, так что объяснить это прямым заимствованием (скре-
щиванием) можно далеко не всегда. Языки пограничья Восточной
Европы и Азии многопадежны: наибольшее (в Евроме и в мире)
число падежей (более 40) имеют языки Восточного Дагестана
(кавказская семья), к северу от них калмыцкий язык демонстри-
рует наибольшее число (11) падежей монгольской семьи, далее к
северу господствует угро-финская семья (инода более 20 падежей).
От Каспия и Урала к Карпатам и Балтике данные семьи.уступают
место индо-европейской (до 7 падежей); затем, к Пиренеям и
Британии, падежи редуцируются в рамках этой семьи, так что в
португальском и валлийском практически отсутствуют. (То же де-
монстрирует другая широтно ориентированная семья — кавказ-
ская.) Тенденция в целом несомненна, хотя в ирландском языке
4 падежа, а в румынском 3. Она столь же несомненно сопряжена
с тенденцией к усложнению глагольных конструкций, хотя в самом
многопадежном языке (табасаранском) глагольные конструк-
ции — отнюдь не самые простые [Взаимодействие лингвистиче-
ских ареалов, 1980, с. 208—209].
Таксономические тенденции могут быть не проще географи-
ческих. Почему треть видов отряда кукушкообразных — гнездовые
паразиты, тогда как среди других птиц это редкость? Почему две
группы насекомых (палочники и клопы-хищнецы) почти сплошь
62
состоят из видов, имитирующих чью-то окраску, тогда как у других
насекомых это бывает редко? Почему в одном отряде динофлагел-
лят (панцырных жгутиконосцев) и только в нем встречаются почти
все типы клеточного деления? Почему сумчатые избегают воды?
Ответов на эти вопросы пока нет, но, как мы увидим в главе 3, рез-
кая неравномерность имеет свои причины, а здесь заметим, что
неравномерность задает тенденцию сама по себе, без выявления
рефрена.
2.6. Гомология и аналогия взаимодополнительны
Вавилов назвал свои ряды гомологическими (ссылаясь на гомоло-
гические ряды в химии и на Дж. Майварта [Вавилов, 1987, с. 21,
51]), но какова их связь с понятием гомологии? И с каким именно
из многих значений этого термина? А можно ли назвать гомологи-
ческими те более широкие ряды, которые описаны после Вавилова?
Дарвин называл подобные ряды аналогическими, т. е. не связы-
вал с гомологией и даже как бы противопоставлял ей. Прямо
противопоставлять их стало обычным после Геккеля, без сомнений
объяснявшего параллелизм независимым приспособлением. Таким
способом он объяснял, например, параллелизм между отрядами
сумчатых и плацентарных зверей [Haeckel, 1866, с. CXLIII].
Произошло смешение трех понятий — гомологии, наследования
и приспособления. Как справедливо отмечал в 1956 г. немецкий
морфолог А. Ремане [см.: Канаев, 1966, с. 117], нельзя путать
объяснение гомологии (как эволюционного феномена) с определе-
нием самого понятия «гомология» (носящего структурный харак-
тер и возникшего до идеи его эволюционного или физиологического
толкования). Обычно гомология противопоставляется аналогии —
так поступал еще Оузн .(см. 1.3), затем Дарвин и Геккель, а Карл
Гегенбаур, один из последних приверженцев Оуэна, уточнил дан-
ное противопоставление афоризмом: «Аналогия для физиолога —
то же, что гомология для морфолога» (подробнее см.: [Чайков-
ский, 1987а]).
В начале XX в. эта мысль поляризовалась в двух противополож-
ных тезисах, когда морфолог-ботаник К. Гёбель утверждал: «Мор-
фологично то, что еще не может быть понято физиологически»,
а его ученик В. Тролль — наоборот: «Морфологично то, что вообще
нельзя понять физиологически» [Канаев, 1966, с. 133]. Зажатая
между этими антитезами, проблема гомологии не могла быть
решена — хотя бы потому, что феномен параллелизма лежал как
бы в другой плоскости. Прежде всего — считать ли параллельные
ряды аналогиями (как Дарвин) или гомологиями (как Вавилов)?
Гёбель еще в 1880-х годах считал их гомологиями [Stevens,
1984, с. 188]. В 1928 г. гистолог А. А. Заварзин писал, что морфо-
логия, пользующаяся понятием гомологии для выяснения род-
ственных связей, «становится в тупик перед сходствами, часто
поразительными, которые существуют между органами только
лишь аналогичными». Их «приходится рассматривать как случай-
63
ность, как некоторую игру природы и обозначать самый факт
ничего не объясняющим термином конвергенции». Если же рас-
смотреть параллелизмы («конвергенции») в их совокупности,
то выясняется, «что они являются выражением каких-то законо-
мерностей, не менее важных, чем даже те, которые лежат в основе
учения о гомологических частях» [Заварзин А. А., 1986, с. 80—81].
Заварзин развил целое учение о параллелизме в строении тканей,
где объяснял его общностью тех функций, которые выполняют
ткани в разных органах и организмах. Однако многие параллелиз-
мы, как мы видели, этим не объясняются.
В 1930-е годы морфолог М. М. Новиков четко указывал, что
вопрос о соотношении гомологии и аналогии шире, чем вопрос о
связи формы с функцией, и, в частности, связан со статусом, припи-
сываемым феномену параллелизма [Бляхер, 1976, с. 201].
Ссылаясь на «Номогенез» Л. С. Берга, Заварзин утверждал,
что формы, имеющие различное происхождение, «в своем развитии
не могут беспорядочно варьировать, а эволюционируют в одном
более или менее определенном направлении, почему в различных
параллельных рядах и развиваются конвергирующие признаки.
Таким образом, с этой точки зрения даже многие гомологические
части обязаны своим сходством конвергенции», на что в частности
указывает закон Вавилова [Заварзин А. А., 1986, с. 81].
Конвергентное понимание гомологии звучит странно, но именно
так понимал ее Оуэн: он, в отличие от Жоффруа, выводившего
гомологию из сходства развития зародышей, видел в гомологии
лишь соответствие самих структур. И совсем отказаться от пони-
мания Оуэна никто не пытается, ибо некоторое различие путей
развития часто наблюдается даже там, где гомология несомненна
для всех (включая строгих последователей Геккеля). Наиболее
наглядно это видно на дроблении (первых стадиях развития мно-
гоклеточного зародыша).
У высших животных господствуют два типа дробления — спи-
ральное у первичноротых (в частности, у моллюсков и членисто-
ногих) и радиальное у вторичноротых (в частности, иглокожих и
позвоночных), но ярко выражен «параллелизм эволюции радиаль-
ного и спирального типов» [Преснов, Исаева, 1985, с. 62], так что
приходится то и дело говорить о «вторичных утратах», «вторичных
приобретениях», «сборных и неоднородных группах» и т. д.,
т. е. трудности здесь в точности те же, что и в любой систематике.
Сходство типа дробления с таксономическим типом может оказы-
ваться чисто номинальным, так что например, по дроблению мле-
копитающие сходны с плоскими червями, а рыбы — с осьминогами.
Наоборот, с другими позвоночными и даже некоторыми млеко-
питающими дробление у человека несходно. Тем самым, налицо
различие ранних стадий развития всех органов, которые гомоло-
гичны у человека со всеми позвоночными; а для некоторых пер-
вичноротых такое различие еще очевиднее. Различие дробления
осьминога и улитки могло бы поставить под сомнение все гомоло-
гии моллюсков, но никто в них не сомневается.
64
В свете сказанного проясняется «понимание гомологизации как
классификации частей» [Мейен, 1978, с. 496], а не как путей их
развития. Те части, кторые принято гомологизировать в руковод-
ствах (рука и нога, перо и чешуя и т. п.), это — элементы радикала
таксона по Вавилову (2.4). Но ведь радикал отличается от рефрена
лишь малой частотой вариации, а не ее возможными пределами.
Поэтому гомологизировать элементы параллельных рядов можно
с тем же основанием, что и другие регулярно реализуемые призна-
ки организмов, в частности — функциональные признаки с не-
функциональными (чисто структурными). Грань между гомоло-
гией и аналогией стирается. Гомологизировать приходится не
только основные свойства организмов (принадлежащие радикалу
таксона), но и все регулярно встречающиеся признаки: «Принад-
лежность меронов одному классу означает их гомологию» [Мейен,
1977, с. 30]. Более того, понятие архетипа по Мейену (2.2) под-
разумевает, что гомологизировать в принципе приходится не толь-
ко наличные, но и принципиально возможные свойства — на этом-
то и основаны были прогнозы Вавилова. Но если любые сходства
могут, с некоторой точки зрения, оказаться гомологией, то что же
такое аналогия? Очевидно — то, что рассматривается с другой
точки зрения, что дополняет основной набор сходств. Если какой-то
набор сходств признан основным, т. е. особи, им обладающие,
отнесены к одному таксону, то этот набор признается первичным
(гомологичным), а все остальные сходства — вторичными (анало-
гиями или параллелями). Так, весь комплекс псовых свойств ено-
товидной собаки признан достаточным для включения ее в семей-
ство псовых; при этом все енотовые и виверровые ее черты призна-
ются параллелизмом. Это обосновано серьезными морфологиче-
скими соображениями, имеется ископаемый материал для срав-
нений, и все же в 1959 г. зоолог С. Фрешкоп предлагал отнести
ее к енотовым. Предложение не было принято [Van Gelder, 1978],
но налицо тот факт, что она внешне очень похожа на енота и «не
имеет тесного сродства ни с какими другими псовыми» [Glutton-
Brock е. а., 1976, с. 165]. Так что в принципе гомология и аналогия
могут меняться местами, и мы вправе говорить о взаимодополни-
тельности этих понятий (в смысле Бора).
Этот вывод звучит странно для всех, кто привык связывать
гомологию с наследованием от общего предка и полагать поэтому,
что в принципе гомология — нечто более важное, чем аналогия,
и что в будущем всякая гомология будет установлена или отвергну-
та окончательно. Сказанное выше приводит к другой точке зрения:
общий предок — лишь один из источников гомологии, источник
частной гомологии. Поскольку феномен множественного парал-
лелизма носит всепроникающий характер, то механизм, его обеспе-
чивающий, тоже всеобщ, и Вавилов справедливо связал его с
гомологией. Столь же справедливо Оуэн назвал однотипность
органов (вроде позвонков) серийной гомологией', онтогенез носит
характер серийного производства. Два соседних позвонка сходны,
как две детали, сошедшие с одного станка, а далекие позвонки
3 Ю. В. Чайковский
65
могут быть совсем несходны, и все же они гомологичны, принад-
лежат одной серии, созданы в принципе одним и тем же онто-
генетическим механизмом. Таковы же принципиальные основания
для гомологизации глаз позвоночных не только между собой, но и
со всеми типами камерных глаз — моллюсков, кольчатых червей,
медуз [Заварзин А. А., 1986, с. 81]. Это — свойство гомологии как
понятийного таксона (см. 2.3). Стоит вспомнить Дарвина, говорив-
шего о гомологии как о свидетельстве родства, а не как о следствии
родства [Чайковский, 1987а]. В главе 9 мы увидим, что современ-
ный термодинамический эволюционизм рассматривает «подлин-
ную гомологию» как общность процессов, а не предков.
Итак, ряды сходств — следствие общности законов развития,
и более важные (для данного исследования) сходства обычно
именуются гомологиями, а остальные — аналогиями. Такой подход
позволяет снять одно старое противоречие, отмеченное Мейеном
[Меуен, 1973, с. 244]. Он напомнил, что часто гомологические ряды
объясняют как результат сходных мутаций в сходных генах.
Действительно, простейшие примеры так объяснить можно (на-
пример, альбинизм всегда вызван нехваткой пигмента), но если
связывать ряды с родством вообще, то придется допустить, напри-
мер, что «общий предок» высших растений обладал способностью
к реализации всех типов расположения листьев вдоль стебля,
хотя не имел ни листьев, ни стебля, а значит — и генов, управ-
ляющих их развитием. Мы еще почти не знаем, в чем состоят эти
законы, но видим, что действие их можно описать, выявляя общие
закономерности. Точно так же, как грамматика сформулирована и
успешно применяется, хотя нет еще определенных представлений
о законах развития языка и лингвисты спорят даже о самом факте
развития. Да и геология успешно использовалась для поиска руд
задолго до появления сколько-нибудь убедительных теорий рудо-
образования.
Грамматика, когда начинаешь ее изучать, подчас кажется
надуманной и излишней, и многие призывали «писать как слы-
шится», однако фактически без грамматики никто не пишет. (Как
отметил М. В. Арапов, малограмотный пишет согласно своей соб-
ственной грамматике, мучая себя и других.) Всякая наука рано или
поздно составляет себе общепринятую «грамматику»; недалеко,
вероятно, то время, когда систематик, предлагая объединить
какие-то виды в один род, будет предъявлять таблицы их «склоне-
ния», т. е. единообразно описанной изменчивости.
Глава 3. РАЗНООБРАЗИЕ, СЛУЧАЙНОСТЬ И ОТБОР
ЗЛ. Случайность и закономерность
Что значит — объяснить явление? Для одних — вывести его из
очевидных положений, для других — найти его механизм, для
третьих (особенно для биологов) — понять его полезность, для
66
1 четвертых — указать ряд, к которому принадлежит явление, тогда
’ как для пятых достаточно, чтобы явление не вступало’ в явные
I противоречия с другими, привычными (и в этом смысле понят-
I ными); То, что не нашло приемлемого объяснения, чаще всего
трактуется как случайность (см. цитату из Вигнера в 1.1) и выпа-
дает из дальнейших рассуждений, как выпал, например, географи-
ческий стиль (2.5). На это выпадение обращали внимание многие
методологи, например, А. А. Любищев и Имре Лакатош. Всякая
1 наука начинается с упорядочения некоторых данных, до этого
казавшихся хаотическими, а наука о разнообразии ставит целью
упорядочить сами эти процедуры частных упорядочений, в ходе
которых из случайного выявляется закономерное. Поэтому необхо-
дим анализ категорий «случайность» и «закономерность».
< Ранее мы говорили о шести типах случайности, имеющих отно-
шение к нашей теме [Чайковский, 1985]:
А. Случайность как непонятая закономерность.
Б. Случайность как скрещение несогласованных процессов,
j В. Случайность как уникальность.
Г. Случайность как неустойчивость движения.
Д. Случайность как относительность знания.
J Е. Имманентная случайность.
(Седьмой тип, Ж, будет введен в 3.2). В той же статье была
приведена литература, на которой основан (если не оговорено
иное) следующий краткий анализ. О типах А и Б следует заметить
только, что к ним относятся псевдослучайные явления, т. е. явле-
I ния, механизм которых в принципе может быть описан без понятия
случайности. Сейчас, после работы Мейена [1984], правильнее
характеризовать тип Б как скрещение несогласованных тенденций
(процесс — это тенденция во времени). В качестве одного из глав-
j ных источников разнообразия Мейен указал мероно-таксономи-
ческое несоответствие (опираясь на работы Ю. А. Урманцева);
( для нас это значит, что распределение меронов по таксонам и
наполнение таксонов меронами в определенной степени случайны.
В прошлом ученым было свойственно видеть во всех явлениях
только случайности типа А и Б, но нынешняя наука понимает
случайность шире. Сложнее характеризовать другие типы. Так, тип
В может иметь в основе тот же механизм, что и типы А и Б, но,
наблюдая явление только однажды, убедиться в этом невозможно.
Тип В бывает важен для понимания разнообразия, поскольку оно,
как правило, уникально: тогда как сами объекты появляются и
исчезают многократно, их разнообразие при этом не повторяется,
а только все полнее разворачивается (ни планета, ни вид орга-
( низмов, ни язык, ни поэт не появляются дважды). Выявить, что в
। каждом таком разнообразии случайно, а что закономерно, нельзя
! без сопоставления с другими разнообразиями. Случайность типа Г
известна всем на примере стока воды из ванны: симметричный
поток сквозь отверстие в дне неустойчив (несмотря на симметрию
I уравнения и начальных условий), и реализуется водоворот, кото-
j
J 3* 67
рый закручивается вправо или влево. Случайность типа Г реали-
зуется, например, в играх (тасовка карт, бросание кости или мо-
неты — здесь случайность вызвана нерегулярностью отображения
множества начальных точек движения во множество конечных).
Она же лежит в основе механизма естественного отбора. В детерми-
нированных системах обычно возникает случайность типов Б и Г
[Заславский, 1984].
Достаточно очевидно, какую роль типы случайности А —Г игра-
ют в структуре любого достаточно богатого разнообразия: случай-
ность возникает за счет сложности, неповторимости и нестабиль-
ности процессов, его породивших. Короче, случайность типов
А—Г является в основном отражением истории множества, разно-
образие которого исследуется. Наоборот, случайность типа Д
относится, как правило, не к истории, а к наличной структуре
множеств. Об относительности знания говорят в разных смыслах,
и едва ли не первым, кто связал ее с проблемой разнообразия, был
Иоганн Кеплер, который в 1595 г. писал, что истинное «заключе-
ние, выведенное из ложных посылок, является случайным; а его
внутренняя ложность немедленно обнаруживается, как только его
применяют к иному объекту, чем тот, для которого оно было выве-
дено» [Kepler, 1938, с. 15]. Именно для того, чтобы преодолеть эту
случайность, полезно исследовать не отдельные факты, а их ряды.
Другой смысл относительности знания связан с принципом
дополнительности. Первоначальная формулировка принципа —
в форме соотношения неопределенностей — прямо связала случай-
ность с относительностью знания, но еще не касалась разнообразия.
Однако Нильс Бор, истолковав неопределенность как частный
случай дополнительности и придав последней общефилософскую
интерпретацию, отмечал возникновение взаимодополнительных
пар (аспектов) в самых разных областях знания. Такова взаимо-
дополнительность точек зрения, терминологий, наук, культур
и т. д. Тогда же Любищев, развивая так называемую концепцию
пробабилизма писал, что в процессе познания метрическая точ-
ность достигается в ущерб общности (достоверности) знания об
объекте и наоборот. Либо надо характеризовать разнообразие как
некое качественное целое (при этом каждый элемент описывается
не своими признаками, а местом в системе), либо детально (в том
числе метрически) описывать отдельные элементы и мириться с
тем, что свойства разных элементов выступают друг относительно
друга как случайности.
Приведем пример. Всем известен тезис Дарвина, согласно кото-
рому эволюция движется за счет естественного отбора, в нем
никто не сомневается (и никогда не сомневался), если понимать
его в самом общем смысле — как апробацию на жизнеспособ-
ность, которой среда подвергает организмы. Правда, в таком виде
тезис многих не удовлетворяет своей расплывчатостью, но оказы-
вается, что уточнить его можно только ценой уменьшения досто-
верности (отсюда и бесчисленные споры о роли отбора в эволю-
ции) ; наконец, если описать феномен отбора математически точно,
68
то такое описание нельзя будет приложить ни к одному природ-
ному процессу, поскольку оно оперирует с немногими наугад
взятыми параметрами организма так, словно они — единственные
существенные для его судьбы свойства (см. 3.4).
Можно уточнить мысль Любищева специально в аспекте анали-
за разнообразия: то, что достоверно с одной позиции исследования,
может быть сомнительным, т. е. в некотором смысле случайным,
с другой (дополнительной по Бору) позиции. Так, общеизвестен
спор о том, случайно ли то разнообразие мутаций, из которого
естественный отбор должен черпать материал, и разрешается этот
спор достаточно просто: если наблюдать за отдельными организ-
мами популяции, то каждая мутация предстает возникшей случай-
но (вне связи с конкретными условиями существования именно
данного экземпляра); однако при анализе механизма мутации
(т. е. в эксперименте с препаратами, выделенными из клеток
большого числа особей) она предстает как цепь детерминирован-
ных физико-химических процессов, в которых случайность (стати-
стическая неопределенность) играет роль только подчиненную.
Неточность процедуры познания неизбежна уже по той причине,
что даже исчерпывающее теоретическое описание, проведенное с
одной позиции, окажется набором более или менее случайных
фактов с какой-то другой (в примере с мутацией таковыми оказа-
лись популяционная и биохимическая позиции исследования).
В биологии случайность часто противополагают целесообраз-
ности, в действительности же эта пара понятий тоже взаимо-
дополнительна. Еще Бэр в 1876 г. обращал внимание, что различ-
ные целесообразные поведения являются друг для друга случай-
ностями, когда не объединены в общую систему, обладающую
своей собственной целесообразностью [Берг, 1977, с. 68]. Действи-
тельно, всякая попытка точно указать место каждого элемента
и цель, которую он себе ставит (или ему ставят), приводит
к толкованию всего разнообрзия элементов как хаоса; наоборот,
при трактовке всей системы как целесообразного целого, ее разно-
образие оказывается понятным, хотя далеко не каждый элемент
можно трактовать при этом как целесообразный. Так, трудно
назвать целесообразным патогенный микроб, уничтожающий свою
жертву (и себя вместе с нею); но целесообразность биосферы
в целом этим не умаляется: она не может существовать без
избыточности — как в отношении числа видов и индивидов, так
и в отношении их разнообразия. Без этого она не была бы само-
достаточна.
Если во всех случайностях типов А—Д оставалась хотя бы
абстрактная возможность видеть псевдослучайность, то в некото-
рых явлениях современное научное мировоззрение такой надежды
нам не оставляет, и приходится говорить об имманентной случай-
ности. Как математическое понятие она существует безусловно:
приходится признать имманентно случайной последовательность
знаков любого неконструктивного числа (числа, для записи кото-
рого в принципе не существует алгоритма). Оказывается, что почти
69
все действительные числа неконструктивны, т. е. «случайны по
Колмогорову», и если рассматривать этот математический факт
как модель реальности, то возникает мысль о преобладании в при-
роде имманентно случайных процессов над детерминированными
и псевдослучайными.
«Случайность по Колмогорову» из всех многочисленных опре-
делений случайности заслуживает наибольшего внимания. Приня-
то считать, что закономерности массовых случайных явлений
должны устанавливаться теорией вероятностей, и часто пишут
даже (например, в недавней «Математической энциклопедии»),
что случайно то событие, которое происходит с некоторой вероят-
ностью. Однако о вероятности можно говорить лишь в тех случаях,
когда многократно происходят однотипные события и вся случай-
ность сводится к отсутствию видимого порядка их появления; тогда
случайность можно описывать через частоту. Мы же в этой книге
должны прежде всего говорить о связи случайности со сложностью,
о системах, в которых выявление каких бы то ни было частот —
не главное в понимании их структуры и функции. Примером
подобной системы является текст: чтобы выразить его суть, его
надо понять. Анализ частот появления в нем букв и слов, если
и может при этом играть роль, то только подчиненную.
После появления в 1963 г. статьи А. Н. Колмогорова «О табли-
цах случайных чисел» математики стали рассматривать связь
случайности со сложностью в аспекте теории информации: слож-
ность последовательности знаков, записанных по определенному
алгоритму, оценивают по минимальной возможной длине этого
алгоритма. При этом максимально сложной называется последова-
тельность, которую нельзя выразить короче, нежели записав ее
целиком. Такую последовательность знаков и называют случайной,
что связывают с интуитивным пониманием случайности как отсут-
ствия алгоритма. Полученные здесь результаты (например: после-
довательность знаков почти любого действительного числа случай-
на) важны для выявления оснований теорий информации и вероят-
ностей, но в других отношениях дают мало и могут даже заводить
в познавательный тупик. Да, обоснованием применения вероят-
ностных, а не каких-либо иных методов к массовым цифровым
данным действительно может служить тот факт, что у наугад
взятой последовательности чисел нельзя ожидать наличия жесткой
закономерности — алгоритма. Но откуда берется сама посылка
«наугад»? Ведь окружающий мир предлагает нам ряды чисел,
образованные не наугад, а в результате совокупного действия
определенных законов; мы, извлекая из этих рядов конечные
выборки, пытаемся угадать порядок их построения, но не всегда
можем найти его (ведь закон обычно проявляется не жестко,
а демонстрирует ядро и периферию — см. 2.3). Далее, об алгорит-
ме можно говорить, когда задан алфавит, на котором тот записан,
но его выявление уже предполагает понимание структуры изучае-
мого множества. Так, прежде чем упорядочить звезды с помощью
диаграммы Герцшпрунга —Рессела, мы должны выявить спект-
70
ральные классы звезд, так как сам по себе спектр конечным
алфавитом не задается; а выявив классы, мы обнаруживаем, что
распределение спектр —светимость неслучайно: оно обладает не-
четкой, но очевидной закономерностью (тенденцией).
Еще нагляднее уже упомянутый пример с текстом. Возьмем
важный для нас содержательный текст и сократим его настолько,
чтобы из него уже нельзя было изъять ни одной буквы без ущерба
для смысла. Текст окажется идеалом информативности (и в этом
смысле — неслучайности), но он же будет «случаен по Колмого-
рову». Здесь тип Е при ближайшем рассмотрении оказался ти-
пом Д. «По сути дела, высокое разнообразие может трактоваться
и как порядок и как беспорядок» [Левич, 1980, с. 51].
В ходе неформальной по своему существу процедуры выявле-
ния алфавита исследователь выражает свое, в основном интуитив-
ное, понимание сложности исследуемой системы, а вместе с тем —
и ее случайности. Для дальнейшего, уже более или менее форма-
лизованного изучения случайности остается обычно только самый
поверхностный уровень ее проявления — тот, который связан с
повторяемостью и вероятностью. (Однако есть ли на деле достаточ-
ная повторяемость? Типично в этом отношении понимание биологи-
ческой эволюции как процесса естественного отбора удачных мута-
ций: если буквой считать нуклеотид, то длина минимального
осмысленного текста оказывается много больше числа актов чтения
(поколений), а если буква — возможный ген, то даже перечень
букв будет не короче длины возможных текстов. Здесь можно
говорить о случайностях типа В, но не о частотах и вероятностях.)
Тогда закономерность случайных явлений получается в ходе
усреднения. Нас же, как уже отмечено в 1.2, больше будет интере-
совать та информация, которую можно извлечь из обобщения '.
Простейшими обобщениями являются ряд и тенденция.
Всякий конкретный пример случайности может быть, в прин-
ципе, с развитием знаний, отнесен к одному из типов псевдо-
, случайности: даже радиоактивный распад, обычно приводимый
в качестве примера чисто самопроизвольного (а потому имманент-
ного объекту) процесса, демонстрирует некоторую управляемость
(нейтрон, например, быстро распадается вне ядра). Поэтому
необходимо особо проанализировать тип Д — возникновение слу-
чайности в системе знаний.
Согласно представлениям, которые сформулировал философ
Вильгельм Виндельбанд на грани XIX и XX вв., каждая наука
имеет свою номотетику, т. е. общие правила, упорядочивающие
в немногих словах большое число фактов, и свою идиографию,
т. е. описание конкретного материала. Виндельбанд утверждал, что
науки сильно различаются по соотношению в них номотетического
и идиографического компонента, и он же заметил, что само разгра-
1 Существенной особенностью диатропики (в отличие от формальной теории
систем) является акцент на итеративность процедуры обобщения и возможное от-
сутствие формальных выражений.
71
ничение номотетики и идиографии зависит от позиции исследова-
теля (анализ и литературу см.: [Беклемишев, 1925]). Рассмотрим
вопрос подробнее.
3.2. Номотетико-идиографическое чередование
Рассмотрим любое частное утверждение, хотя бы, например:
«этот стол — деревянный». Фраза, чисто идиографическая, указы-
вает на частный конкретный факт, но несет в себе в свернутом
виде мощную номотетическую информацию. Ведь слово «стол» —
символ принадлежности к некоторому абстрактному множеству
столов, указание на номотетику. Еще мощнее свертка информации
в слове «деревянный», которое указывает на все предметы, сделан-
ные из дерева, причем дерево может быть любое; т. е. проведена
еще и свертка по всем видам деревьев, тогда как, например, слово
«дуб» уже означает свертку по всем видам и экземплярам, принад-
лежащим к данному роду. Но более всех абстрактно слово «этот»,
формализующее не предметы или их качества, а отношение их
к говорящему. Оказывается, идиографический факт формулирует-
ся как пересечение номотетик.
Отметив это, можно перейти к вопросу о том, что номотетично,
а что идиографично в той или другой научной дисциплине. Это
зависит от - позиции исследователя, целей, которые он ставит,
и средств, которыми он располагает. Так, для математика «Курс
геометрии» — номотетический текст, а «Курс практической систе-
матики высших растений» — сугубо идиографический, и мало кто
замечает, что для ботаника ситуация прямо противоположна.
Действительно, геометрия для него, как правило,— набор теорем,
которые нельзя усвоить иначе, чем заучив их, и наличие доказа-
тельств не упорядочивает для него текст, но только загромождает
(надо заучивать еще и их); наоборот, инструкции «Практической
систематики» упорядочивают для него море растений, стоящее
перед глазами, дают метод свернуть необозримое множество при-
знаков в компактное множество диагнозов. Словом, номотетично
то множество, в котором данный исследователь видит номотетику.
В этом отношении интересно вспомнить следующее определе-
ние полезности информации: информация тем полезнее для вос-
принимающего, чем более она увеличивает его информирован-
ность. Другими словами, информация бесполезна как для того,
кто ее уже знает, так и для того, кто не может ее понять
[Шрейдер, 1965].
Приняв, что слово — это имя множества и тем самым указы-
вает номотетику, получаем следующую циклическую схему постро-
ения понятий, общую как для разговорной и научной речи, так
и для самой процедуры познания. Как уже говорилось, любой
конкретный идиографический факт задается пересечением номо-
тетик. Из идиографических фактов как из элементов строится
всякая идиография, т. е. всякое конкретное описание, составляю-
щее первичный материал науки и практической деятельности.
72
Другими словами, идиографии выступает как объединение идио-
графических фактов. Наконец, номотетика оказывается пересече-
нием идиографий. Из того факта, что номотетика и идиографии
задают друг друга, следует, в частности, что в разговорном языке
нет каких-либо основных слов, через которые определяются осталь-
ные; наоборот, язык определяет сам себя, зато через него опреде-
ляются искусственные языки. Поэтому в основе всякой формаль-
ной теории лежит словесное описание, причем почти всякое слово
разговорного языка может выступать как категория 2. Очевидно,
что понятия номотетики и идиографии взаимно дополнительны —
точно так же, как понятия гомологии и аналогии. Нетрудно также
увидеть сходство между номотетико-идиографическим и мероно-
таксономическим чередованием в процессе познания соответствую-
щих разнообразий.
Теперь можно сформулировать понятия случайности и законо-
мерности, удобные для наших целей. Именно, закономерностью
назовем всякую возможность выявить и именовать некоторое номо-
тетическое множество (т. е. множество, элементы которого обла-
дают существенным для наблюдателя общим свойством). Тогда
случайностью естественно назвать выбор элемента (или ряда эле-
ментов) из какого-то множества (неважно, номотетического или
нет), если наблюдатель не может характеризовать этот выбор
в номотетических терминах. Другими словами, случайность раз-
нообразия можно характеризовать как такой взгляд на него, при
котором мы отказываемся исчерпывающе характеризовать отдель-
ные объекты, но можем, хотя бы в принципе, характеризовать
их множество. Удобно ввести седьмой тип случайности:
Ж. Случайность как произвольный выбор.
Он тесно связан с типами Б, В и Д, но не сводится к ним.
Вопрос о том, имеет ли данное множество номотетику (т. е.
закономерно) или не имеет (т. е. случайно), бессмыслен, пока
не задан аспект, в котором желательно видеть упорядочение: ведь
любую идиографию можно расчленить так, что получатся отдель-
ные идиографические факты, а можно так, что получатся элемен-
ты, входящие в некоторые номотетики. Если же достаточно раз-
нообразное множество не с чем сопоставить, то оно обычно высту-
пает как идиография — ведь номотетика создается пересечением
различных идиографий. Такое понимание случайности разнообра-
зия характеризует процесс познания, а не само познаваемое яв-
ление, и обычно для многих наук. Даже в такой точной науке,
как небесная механика, где царят формулы, работа началась
с упорядочения.
Для первобытного человека звезды расположены беспорядочно,
но уже в глубокой древности было установлено, что все они,
за исключением пяти планет, неподвижны на небесном своде. Тем
самым понятия «звезда» и «планета» породили два ряда. В древ-
ности же разнообразие наблюдений планет упорядочили в систему
2 Это частный случай «категориальной когерентности» по Н. Гартману.
73
кругов. Коперник заново упорядочил эти круги, положив центром
Солнце (точнее, положив центр рядом с Солнцем; подробнее см.
[Чайковский, 19876]), Кеплер упростил эту систему, заменив
большое число кругов небольшим числом эллипсов (т. е. найдя
более простые ряды для известных наблюдений), после чего Нью-
тон, объяснил их, найдя формулу закона тяготения. В биологии
до формул дело доходит редко, поэтому трудно решить, какой
из возможных рядов лучше («естественнее»), но упорядочение
наблюдений проводится столь же неукоснительно.
Не следует объявлять какое-либо множество имманентно слу-
чайным, так как вас может опровергнуть любой, кто укажет в нем
ряд, вами не замеченный. Даже для простейших множеств —
числовых последовательностей — нет способа практически устано-
вить, что они случайны. Например, последовательность знаков
числа л случайна в том смысле, что ни один статистический
критерий не поможет предсказать (хотя бы в вероятностном
смысле) (н-|-1)-й знак по п предшествующим знакам. Статистик
вправе назвать эту последовательность случайной, но это — слу-
чайность типа А и только, ибо любой знак числа л вычисляется.
Тем более нельзя говорить, что какой-то опыт доказывает случай-
ность данного биологического явления (например, мутации). Един-
ственная бесспорная случайность, о которой можно всерьез гово-
рить при понимании ее как неупорядоченности, возникает в тех
ситуациях, где вообще нельзя выстроить ряд — когда различных
объектов слишком мало. Если, например, считать жизнь возникшей
от единственной первобактерии, то это возникновение можно
считать случайным, но если допустить, что первых организмов
было два, то сразу обнаруживается пара богатых параллельных
рядов (наследственная память, ее считывание, энергетика, мембра-
ны, размножение и т. д.), так что неслучайность налицо. Хирот-
ность сирена и хирота (2.5) тоже можно было считать случайным
совпадением, пока они не встали в общий рефрен. Как видим,
номотетико-идиографический подход оставляет случайности очень
скромное место.
И все же без идеи случайности разнообразие анализировать
невозможно. Дело в том, что при исследовании разнообразий
часто бывает необходим один частный атрибут случайности —
независимость исследуемых событий. Если вам надо, например,
выяснить, действительно ли долгожительство является существен-
ной тенденцией разнообразия горцев, то самый простой выход —
переписать возраст всех горцев и установить процент долгожите-
лей. Если же всех переписать невозможно, то необходимо органи-
зовать выборку, в каком-то смысле случайную, и есть довольно
общий прием: обработать какой-нибудь обозримый ряд, заведомо
составленный без связи с возрастом (например, брать список
жителей каждого десятого поселения из алфавитного списка посе-
лений) . Вообще, основной источник фактически удобной случай-
ности (тип Б) — в том, что разные способы упорядочения одного
и того же разнообразия — например, по возрасту и по алфавиту
74
адресов -- можно считать несопоставимыми (хотя доказать это и
невозможно).
Другой частый источник фактической случайности — неполно-
та комбинаторики. Если виды одного рода различаются только
десятью признаками, причем ни один признак не влечет обязатель-
но другого, и они принимают всего по два значения (есть или
нет), то даже тогда возможно 210, т. е. более тысячи видов.
А поскольку чаще всего род включает менее десяти видов (как
и семейство обычно включает менее десяти родов и т. д.), то
фактически реализованной мы видим небольшую долю комбинаций
и можем, в зависимости от вкуса, или считать их случайными,
или строить для них «родословные древа», или искать «запреты
на определенные комбинации».
У двух голубоглазых родителей может родиться черноглазый
ребенок — в результате мутации. Случайна ли она? Да, случайна
в том смысле, что мы не можем предсказать, произойдет ли она
у членов данной голубоглазой пары (как не можем предсказать,
что встретим сегодня на улице знакомого), но из этого никак
не следует, что мутация возникает без определенной причины
(как нельзя считать, что наши знакомые без цели бродят по ули-
цам) . Мутация — результат очень сложной комбинации реакций
(каждая из которых течет по четким законам), подчас несопо-
ставимых друг с другом в таком же смысле, как несопоставимы
наши намерения с намерениями знакомого, которого мы встречаем
на улице. Замечательно, что Дарвин, ничего не зная о механиз-
мах наследственности, понимал случайность наследственных из-
менений именно так: «Хотя каждое изменение должно иметь
собственную возбуждающую причину и хотя каждое из них под-
чиняется закону, мы все-таки так редко можем проследить в точ-
ности соотношение между причиной и следствием, что нам хочется
говорить о вариациях, как о проявляющихся произвольно. Мы даже
можем назвать их случайными...» [Дарвин, 1951, с. 770].
Это очень существенно: дарвинская установка означает готов-
ность искать закономерность там, где с виду есть только случай-
ности. Наоборот, у позднейших авторов мы чаще видим беспред-
метный спор — случайна ли та изменчивость, которая поставляет
материал для эволюции? Разумеется, она случайна в одних смыс-
лах и неслучайна в других. Эту мысль легко пояснить техническим
примером. При решении задач на компьютере часто требуются
таблицы случайных чисел, и для этой цели придуманы специаль-
ные короткие программы, всего в 5-7 арифметических действий.
Естественно, никакой случайности в работе программы нет, но
в результате последовательной ее работы получается таблица, со-
держащая миллионы чисел, и любой статистический критерий под-
твердит, что таблица — случайная.
Как говорится, статистике часто принадлежит первое слово,
но последнее — никогда. Она часто наводит на размышления,
но никогда сама не дает ответов, поскольку занимается лишь
явлениями, а не их внутренней сутью. Первое слово и у нас
будет принадлежать статистике.
75
3.3. Статистика гауссова и гиперболическая
«Я до сих пор живо помню, как однажды, когда я был еще
ребенком, отец привел меня на край города, где на берегу росли
ивы, и велел мне сорвать наугад сотню ивовых листочков...
Вернувшись домой, мы расположили их в ряд по росту, как солдат.
Затем отец через кончики листьев провел кривую и сказал: „Это
и есть кривая Кетле. Глядя на нее, ты видишь, что посредствен-
ности всегда составляют подавляющее большинство и лишь немно-
гие подымаются выше или так и остаются внизу”» [Варден, 1960,
с. 84].
Пример Кетле действительно поучителен: эту кривую можно
получить, не только ранжируя листья или солдат, но самыми
неожиданными способами; дело в том, что кривая Кетле — это
гауссово распределение F(х), называемое еще нормальным распре-
делением случайных величин. Результат почти всякого массового
измерения однотипных величин дает одну и ту же картину:
отдельные измерения отклоняются от своего среднего («нормы»)
с частотой, близкой к /(ж) (рис. 1). Кривая f(x) называется
плотностью распределения F (х).
Рис. 1. Нормальное (гауссо-
во) распределение случай-
ной величины: а — само
распределение (кривая Кет-
ле); б — его плотность;
в — формирование гауссовой
кривой путем сложения оди-
наковых независимых слу-
чайных величин Х(х), име-
ющих равномерные плотно-
сти [Варден, 1960. С. 131].
Уже четвертая кривая почти
неотличима от гауссовой,
с той. лишь разницей, что
обращается в нуль при
—4, z^4.
76
Адольф Кетле, бельгийский статистик, ввел в естествознание
то воззрение на мир чисел, которое лежит в основе основ современ-
ного естествознания и в котором уже поэтому полезно время от
времени сомневаться. До него статистика хоть и использовалась
в социальных науках, но в основном сводилась к простейшим
подсчетам типа суммирования величин и их деления. Статисти-
ческая логика открыла возможность достаточно точных прогнозов,
не всегда очевидных с первого взгляда. Так, измерив рост многих
солдат, нетрудно вычислить средний рост и дисперсию (т. е. меру
уклонения от среднего) роста; зная их, можно предсказать, что
приблизительно каждый 44-й солдат окажется ниже среднего роста
более чем на удвоенную дисперсию, зато и каждый 44-й будет
на такую же величину выше — все симметрично. Эти сильные
и в то же время довольно редкие отклонения можно назвать
нетипичными, откуда и рабочее понятие — типично (и в этом
смысле нормально') то, что близко к среднему. (Удобство понятия,
впрочем, бывает обманчиво, как мы вскоре и увидим). Идя этим
путем, Кетле сформулировал идею социального благополучия как
баланса средних значений и идею эволюции общества как постепен-
ного направленного сдвига этих средних.
Свою первую книгу («Социальную физику») он издал в 1835 г.,
а два года спустя молодой Дарвин записал в блокнот свои первые
мысли о постепенной эволюции организмов [Дарвин, 1959, с. 90 —
127]. Дарвин и Кетле оба выражали идеи своего века: знание
как статистику, мир как баланс средних, а эволюцию — как сдвиг
средних. Эта мысль стала главной, когда в XX в. дарвинизм стал
вооружаться статистикой. Стали полагать, что если условия среды
требуют, чтобы какой-то признак организма стал более определенно
выраженным или просто увеличился, то материал для этого найдет-
ся: его предоставят те особи, которые, отклоняясь от «посредствен-
ностей» в нужную сторону, производят тем самым больше потомст-
ва. Гауссова кривая как бы вытянет «голову» и подожмет «хвост»,
чем и сдвинет среднее; в следующих поколениях эта смещенная
кривая плотности станет основой для новых сдвигов — вот и эле-
мент эволюции (рис. 2). Но всегда ли у плотности распределения
искомого признака есть «хвост», который может быть поджат,
и «голова», которая ползет куда надо? Например, признано, что
первые организмы не обладали фотосинтезом. Что надо поджимать,
чтобы он появился?
Гауссово распределение симметрично: всякому уклонению от
нормы здесь можно поставить в соответствие противоположное:
короткому листу — длинный, яркому таланту — бездарность и т. д.
Однако парадокс состоит в том, что Варден-отец был не вполне
прав, давая урок будущему статистику. Разумеется, если в вузе
подвергнуть всех студентов какому-нибудь тесту на умственное
развитие, то результат будет похож на гауссово распределение,
но это потому, что тест специально на это рассчитан. С теми же
студентами можно проделать иной эксперимент, который даст
абсолютно другой результат: надо подождать лет 30—40, а затем
77
РИС. 2. Традиционное объяснение элементарного эволюционного акта, понятого
как изменение признака (например: [Шмальгаузен, 1968]). В результате
давления условий среды (стрелка) те особи, которые имеют значение признака
z<ai, постепенно вымирают, в результате чего распределение признака х должно
измениться — вместо прежней сплошной кривой будет изображаться штриховой,
а среднее значение признака вместо at станет равным а?. Предполагается, что любой
признак распределен центрально.
подсчитать, сколько кто написал научных работ. Вопреки ожида-
ниям, распределение работников науки по числу написанных ими
статей имеет однохвостую плотность: около трети не опубликовало
ни одной статьи, столько же имеет по одной-две публикации; и
далее — чем более статей, тем меньшее число авторов ими обладает
(рис. 3). Основная масса статей обязательно окажется написанной
теми немногими, кто имеет по 30 статей и более. Если восполь-
зоваться языком Вардена-отца, придется признать, что почти все
«остались внизу», а «посредственностей» (т. е. авторов, имеющих
по 5 — 6 статей, чья продуктивность близка к среднему результату
печатной деятельности) ничтожно мало. Точно так же распреде-
ляются и сами статьи по их цитируемости: около трети не цити-
рованы никем, столько же упомянуты по 1—2 раза, а основная
масса ссылок приходится на малую долю статей.
РИС. 3. Типичные краевые распределения: 1) п — число особей в клоне бактерий,
А? — доля таких клонов среди всех клонов четвертого поколения, если в каждом
поколении вероятность гибели каждой бактерии равна вероятности ее благополуч-
ного делении (р=1/2). В этих допущениях доля вымерших к четвертому поколению
клонов равна N(0)=2/3; 2) п - число статей, опубликованных научным сотрудни-
ком за всю жизнь, N — доля таких сотрудников среди всех научных сотрудников;
3) п — число видов в роде, — доля таких родов в крупном таксоне (насчитываю-
щем более 300 родов) растений, грибов или животных.
78
Аналогично получается с выживанием организмов и видов.
Проще всего убедиться в этом на примере колонии бактерий,
хотя интересующее нас обстоятельство типично для всех организ-
мов.. Пусть каждая бактерия делится ежечасно, причем общий
размер колонии постоянен. Потомство одной особи, полученное
бесполым путем, называется клоном. Если следить за десятком
тысяч клонов, то окажется, что через 100 поколений сохранилось
всего около 200 клонов, тогда как остальные 9800 за это время
вымерли. Зато оставшиеся в живых 200 клонов окажутся весьма
различными по численности; в них будет в среднем по 50 бактерий
на клон, что и даст необходимые 10 000 особей, т. е. неизменный
баланс колонии — одну особь на исходный клон [Чайковский,
1977а].
Как видим (рис. 3), распределение живых клонов по числен-
ности в каждый заданный момент тоже оказывается «однохво-
стым»; при этом клонов, представленных одной-единственной
особью, т. е. средним числом, будет всего около трех (из 10 тысяч!),
следовательно, термин «среднее» не несет здесь того смысла «наи-
более вероятного», что в гауссовой статистике. Наоборот — наи-
более вероятная численность здесь всегда далека от средней:
она или гораздо ниже (в нашем случае — нулевая численность
вымерших клонов), или гораздо выше. Тем самым, к «однохво-
стым» плотностям бесполезно подходить с обычной статистической
меркой: среднее значение здесь не имеет сколько-нибудь интерес-
ного смысла, зато термин «самое вероятное» имеет сразу два
противоположных смысла. Если в обычной гауссовой статистике
можно хоть в каком-то смысле говорить о норме как о среднем,
то здесь норму одним числом выразить нельзя, а приходится вво-
дить новый язык, язык кластеров.
Кластер — это компактное множество элементов, имеющих
близкие свойства, причем каждый кластер имеет свои характери-
стики, свою норму. Так, разумно сравнивать размеры «пустых»
кластеров (доли непечатающихся, нецитированных, вымерших
и т. д.) или размеры «ведущих» кластеров (доли авторов, пишущих
половину всех статей, доли клонов, составляющих треть всей
колонии и т. д.).
3.4. Мажорирующая модель отбора
Итак, дарвинское «уравновешенное число форм» скрывает в себе
никак не колебания вокруг среднего, а бурю, губящую почти всех
и дающую право на колоссальное потомство немногим. Это не имеет
пока никакого отношения к отбору, так как плодовитость всех
особей предполагалась в наших расчетах одинаковой. Что же будет,
если в этой буре примут удары не равные, а несколько различные
по плодовитости организмы?
Как мы видели, при отсутствии отбора уже через несколько
десятков поколений клоны стационарной колонии почти целиком
распределяются между двумя кластерами — вымерших и много-
79
кратно размножившихся. Существенно, что никакого сглаживания
между этими двумя процессами (вымиранием и бурным размно-
жением) со временем не происходит', в каждом поколении около
половины особей гибнет (т. е. отправляет свое потомство в кластер
вымерших), а другая половина обеспечивает сохранение общей
численности. Введение отбора лишь несколько изменяет эти доли,
не меняя существо процесса, который оказывается слишком хаоти-
чен, чтобы чувствовать малые вариации плодовитостей. В настоя-
щее время этот факт хорошо описан [Кимура, 1985] и никем из
анализировавших математические выкладки не оспаривается. Од-
нако на общебиологическую мысль они до сих пор оказали мало
влияния. Причин этому можно указать несколько.
Во-первых, неоправданно сложный аппарат диффузионных
уравнений, используемый в теории отбора, биологам непонятен;
они принимают или не принимают его. лишь на основе убеждений,
сложившихся у них независимо от математической теории. Столет-
ний опыт математического моделирования эволюции показал: тот,
чьи убеждения модель подтверждает, склонен видеть в ней строгую
теорию, обоснование подлинно научных воззрений; а тот, чьим
убеждениям модель противоречит, склонен говорить о невозмож-
ности адекватного моделирования «столь сложного процесса». Во-
вторых, ориентация статистической генетики на финальные вели-
чины (характеризующие объект через бесконечное время или же
в среднем по бесконечному числу независимых испытаний) не так
уж интересна для теории эволюции. В-третьих, статистическая
генетика, как и почти вся математическая статистика, до сих пор
игнорирует «однохвостые» распределения. Интерпретация матема-
тических результатов обычно дается в терминах средних величин
и потому оказывается беднее самих результатов. И, в-четвертых,
эволюционные модели статистической генетики традиционно носят
имитационный характер, т. е. пытаются максимально (насколько
позволяет аппарат) приблизиться к реальной ситуации, которую
хочется описать. Это — тупиковый путь, поскольку никогда нет
гарантии, что неучтенные обстоятельства играют меньшую роль,
чем учтенные. Такие модели имеют смысл тогда, когда ставится
цель проверить определенное свойство объекта (меняя это свойство
в модели, посмотреть, как меняется ее поведение в целом, чтобы
оценить возможную реакцию объекта на воздействие), но они
бессмысленны, если ждать от них прогноза поведения реального
объекта на значительных временах.
Сказанное приводит к следующему выводу. Поскольку, с одной
стороны, не представляется возможным сколько-нибудь строго
описать даже простейший процесс эволюции, а с другой стороны,
произвольные упрощения лишают работу биологического смысла,
необходимо применить метод, известный в математике как мажо-
рирование. Именно, желая доказать некоторый факт, будем делать
только те упрощения, которые заведомо не благоприятствуют дока-
зательству, т. е. те упрощения, которые, будучи реализованы в при-
роде, заведомо не облегчили бы осуществление данного факта.
80
Если факт удастся доказать в этих допущениях, то он заведомо
верен и без них, т. е. в реальной природе. Такие допущения
назовем мажорирующими. Подробно мажорирующая модель есте-
ственного отбора малых вариаций рассмотрена ранее [Чайковский,
1977а], там же можно найти нужную литературу, а здесь
ограничимся упрощенным изложением основных допущений и по-
лученных выводов.
Пусть мы следим за клоном, каждая особь которого оставляет
в среднем 5=1,01 потомка, т. е. с вероятностью 0,495 гибнет, а
с вероятностью 0,505 делится на две. Легко вычислить, что такой
клон с вполне ощутимой вероятностью 1/50 доживает до бесконеч-
ности, тогда как любой клон, оставляющий в среднем на особь
одного потомка или менее, наверняка рано или поздно вымрет.
Еще в 1922 г. Роналд Фишер, основоположник эволюционной
статистики, получив похожий результат, счел его решающим аргу-
ментом в пользу эффективности процесса отбора малых вариаций
(хотя, в отличие от своих последователей, признавал, что речь
идет лишь о модели, а не о теории реального процесса). По
Фишеру, если полезная мутация возникнет хотя бы 100 раз,
то ее обладатели, вернее всего, сохранятся и в конце концов
вытеснят всех остальных. Он, по-видимому, просто не подумал
о самых очевидных трудностях — ведь отбору подвергаются не
мутации, а их полезные комбинации (их не то что сто раз,
а и дважды не всегда дождешься), причем отбор каждого полезного
клона должен закончиться за ограниченное время. В течение этого
времени не только может сохраняться любой клон-конкурент,
но и будут неизбежно меняться сами условия конкуренции. Стоит
немного приблизить расчет к реальности, чтобы получить противо-
положные выводы.
Задача о выживании мутантного клона поставлена Фишером
как задача вычисления вероятности выживания единичного мутан-
та в зависимости от селективной ценности (называемой также
коэффициентом отбора) s мутантного генотипа. Фишер рассматри-
вал однородный ветвящийся процесс, в котором математическое
ожидание численности клона начавшего размножение при t=0,
равно
Et (s) = (l-(-s)' (1), причем 1+s = 2 гф, t = 0, 1, 2..., (2),
1 = 0
где t — номер поколения, a qr — вероятность оставления одной
особью числа i потомков в одном поколении. Кроме Et (s) и s для
исследования процесса полезно знать условную среднюю числен-
ность Mt мутанта, не утерянного до момента t:
Mt (s)=Et (s)/Pi (s),
где pt — вероятность неутери клона до t. Поскольку числа ф(. опи-
сывают процесс размножения точно, постольку формулы (1) — (3)
дают точную вероятностную динамику клона, и в этом преимущест-
во подхода Фишера. Однако, если ф не постоянны, то процесс
является неоднородным, и основной аппарат теории ветвящихся
81
процессов (аппарат производящих функций) обычно нельзя ис-
пользовать. Поэтому в математической генетике более популярна
предложенная Вильямом Феллером в 1951 г. диффузионная мо-
дель, использующая аппарат уравнений в частных производных,
в принципе допускающая широкий класс переменных параметров,
характеризующих клон. Хотя сам Феллер указал на ограничен-
ность модели, имеющей смысл только при больших численностях
исследуемых клонов и большом числе поколений, а также на
необходимость аккуратного задания переменных и осторожного
толкования результатов, в последующие годы эту модель исполь-
зовали для самых смелых выводов. Вначале «оправданием» служи-
ло правдоподобие получаемых результатов (т. е. решались биоло-
гически тривиальные задачи, ответ которых предсказуем качест-
венно) , однако теперь положение радикально изменилось. Как
показал анализ [Чайковский, 1977а], целый ряд утверждений,
обсуждавшихся в качестве реальных эволюционных закономер-
ностей на страницах эволюционно-статистических публикаций,
является лишь следствием некорректного перехода от ветвящегося
процесса к его диффузионной модели. Чтобы не повторять таких
ошибок, лучше всего предельно упростить аппарат, но так, чтобы
все упрощения носили мажорирующий характер.
С тех пор как Фишер показал для пуассоновского клона, что
селективной ценности мутанта s = 0,01 достаточно для его выжива-
ния с вероятностью p(s) ~0,02, в математической генетике популя-
ций (а затем и в биологической литературе) стала господствующей
следующая модель эволюции клонов под действием естественного
отбора: единственным первичным источником изменений являются
случайные мутации (случайные как в смысле момента возникно-
вения, так и в смысле связи с потребностями адаптации), которые
все можно разделить на селективно полезные (з>0), селективно
нейтральные (з=0) и селективно вредные (s<0); даже малые
з>0 достаточны, согласно этой модели, для вытеснения их облада-
телями тех, которые имеют з^О, чем и определяется фактический
ход эволюции. Установилось молчаливое согласие, что эта модель
мажорирует реальную эволюцию в том смысле, что в реальной
природе регулярно реализуются мутанты с з>0,01. Однако в дейст-
вительности даже в простейшем случае (конкуренции двух клонов)
не всегда можно приписать мутанту определенное s =£0 [Чайков-
ский, 1977а, с. 1476], т. е. упрощения, сделанные Фишером,
противоречат идее мажорирования. Зато его результаты вполне
применимы практически, если ставить противоположную цель —
показать те пределы, в которых естественный отбор заведомо
не действует. Для этого надо ввести в модель только мажорирую-
щие допущения, облегчающие естественный отбор полезных
свойств.
Таких допущений сделано три. 1) Поскольку моделировалось
распространение полезного свойства (а не его возникновение),
то это свойство описывалось как постоянный объект действия
отбора, не меняющийся с течением поколений за счет скрещивания.
82
Это достигалось рассмотрением клона (а не популяции). 2) Малое
s>0 допускалось в течение всего процесса, хотя среда, уступающая
клону одну и ту же малую долю в каждом поколении, нереальна;
допущение малого s>0 мажорирует утверждение об эволюционной
роли малых селективно полезных изменений. 3) Величина s>0
считалась постоянной. Если бы допустить, что s зависит от времени
и лишь в среднем положительна, то роль отбора оказалась бы еще
более скромной. Допуская постоянное s, мы постулируем постоян-
ное перемешивание всех условий существования, что осуществимо,
и то с трудом, лишь в микробиологическом эксперименте.
Сделав эти допущения и учтя, что реальный интерес представ-
ляет не формула (1), выражающая полную среднюю численность,
а формула (3). характеризующая кластер выживших, легко полу-
чить основной вывод: за приемлемое время выгодный мутант
не может, как правило, не только завладеть колонией, но и просто
выделиться на фоне невыгодных. В частности, даже численность
клона мутантов, имеющих s=0,04 (такую мутацию уже нельзя
назвать малой) при £=128 превосходит численность клона мутан-
тов, имеющих s=—0,04, всего в 132 раза, тогда как самих этих
невыгодных клонов за это время возникает в сотни раз больше,
чем выгодных.
Нерегулярность клонального роста формально выражается в
том, что дисперсия численности клона неограниченно растет по
сравнению с самой численностью (это и делает средние величины
неинформативными). Поэтому, в частности, оказывается, что судь-
ба любого мунтанта, как вредного, так и полезного, фактически
решается в ходе первых поколений его существования: почти все
они в это время исчезают. Разумеется, для судьбы мутанта,
пережившего несколько поколений, важно, полезен он или вреден
(в предположении, что постоянное s>0 возможно), но сам факт
неутраты в течение этих поколений слабо зависит от селективной
ценности: из каждой тысячи мутантных клонов за восемь поко-
лений при s=—0,04 утрачивается 868 клонов, при s=0 утрачи-
вается 836, а при s=0,04 утрачивается 801 клон. Отбор не столько
выбирает полезных мутантов, сколько распространяет тех, у кото-
рых s не слишком мало и которые случайно (безотносительно
к отбору) пережили первые несколько поколений; уже поэтому
роль отбора здесь довольно скромна.
Итак, малое различие в постоянных селективных ценностях
серьезным фактором эволюции не является. Не приходится ожи-
дать значительной роли и от малых изменений s, но вот роль резко-
го их изменения отрицать априори нельзя. Поэтому пришлось
исследовать и такую модель: в условиях катастрофы, т. е. монотон-
ного сокращения численностей клонов за время порядка 100 поко-
лений, исследовано выживание клонов с различными близкими
st (естественно, все они были отрицательны) и значительными
численностями в начале катастрофы. Получены мажорирующие
оценки вероятностей выживания, показавшие, что судьба клона
может вообще не зависеть от st: легко наблюдать преимуществен-
83
ное выживание клона с меньшей s [Маленков, Чайковский,
1979].
Фишер был прав, утверждая, что одна полезная мутация может
вытеснить когда-нибудь одну вредную, но оказывается, что редкие
слабовыгодные мутации тонут в потоке слабовредных. Отбор,
как проверено и в природе, и в лаборатории, и в практике
селекционеров, эффективно действует тогда, когда разница в плодо-
витости участников велика — когда один тип организмов раз-
множается свободно, а его конкуренты почти не размножаются.
Наоборот, слабые различия размножаемостей отбор использовать
эффективно не может.
Зато в ходе расчетов обнаружились другие процессы, быть
может, влияющие всерьез на судьбу клона: рост мутабильности
при плохих условиях среды, падение размножаемости в тех же
условиях, быстрый выход колонии из катастрофических условий
при резком изменении ее генотипического состава — все они могут
в принципе играть не меньшую роль в элементарных актах
эволюции, нежели селективная ценность. Однако надо отметить,
что мажорирующий характер моделей, позволивший сделать опре-
деленные выводы о невозможности определенных процессов, ли-
шает нас права делать определенные выводы о возможности какого-
либо определенного изменения — для этого надо бы делать иные
допущения. Указание на какие-то параметры, более существенные,
чем селективная ценность, можно расценивать лишь как повод
к размышлению, к построению иных моделей.
3.5 Гиперболические распределения Виллиса
В 1900 г. ботаник Гуго Де Фриз предложил свою знаменитую
«мутационную теорию» — модель эволюции, в которой новый вид
появляется скачкообразно, за,счет «видовой мутации», которую
Де Фриз считал более важной формой изменчивости, чем медленное
накопление мелких преимуществ. Он писал: «Как известно, Дар-
вин принимал обе формы происхождения новых видов, но его
последователи всегда отдавали больше предпочтения медленному
происхождению» [Де Фриз, 1932, с. 55]. Естественно, они не при-
няли идею Де Фриза, но она стимулировала первую эволюционно-
статистическую работу, где разнообразие выступало как феномен
гиперболической статистики.
Джон Кристофер Виллис, как почти все английские ботаники,
был в юности очарован учением Дарвина; однако, видя его слабую
фактическую обоснованность, он решил собрать недостающую ар-
гументацию сам. В своем «Словаре цветковых растений и папорот-
ников» он насчитал 12 561 род цветковых, из которых 4853 рода
содержали по одному виду. Поразительно: ведь понятие рода для
того и введено, чтобы объединять сходные виды, на одновидовой
род принято смотреть как на исключение из правила — то ли
следствие плохой изученности, то ли плохой работы систематика,
то ли вымирания видов. Но вот выясняется, что таких родов —
84
более трети, зато несколько родов — просто гиганты: астрагал (из
бобовых) — 1600 видов, крестовник (из сложноцветных) — 1450 ви-
дов. Виллис построил графики: по оси абсцисс число видов в роде,
а по оси ординат количество соответствующих родов — и получил
хорошие гиперболы. Приятель Виллиса, математик Гаролд Юл,
посоветовал откладывать по осям графика не сами величины, а их
логарифмы, и почти все точки аккуратно легли на прямую, причем
прямые для разных семейств легли почти параллельно. Для
сравнения Виллис просчитал некоторые семейства низших расте-
ний, а также жуков, змей, ящериц и нашел ту же закономер-
ность [Yule, 1924, с. 85 — 87]. Число родов в семействе также
выразилось гиперболой, только более пологой — доля однородовых
семейств составила 18% [Willis, 1922, с. 188]. Этот феномен ока-
зался всеобщим [Численко, 1977]. Разные исследователи, разная
изученность и разный возраст таксонов, а результат один и тот же.
Если бы разнообразие видов зависело просто от разнообразия
условий обитания, как учил Дарвин, то оно не могло бы так четко
следовать закону, игнорирующему всякую экологию. Гиперболы
подтверждали, как счел Виллис, правоту де Фриза: структура
рода отражает еще не понятый закон «мутаций» [Willis, 1922,
с. 221].
В 1924 г. Юл взялся объяснить открытый Виллисом закон
математически и воспользовался тем же аппаратом, что и Фишер,—
теорией ветвящихся случайных процессов. Юл положил, что вид
делится на два скачкообразно (т. е. в результате «видовой мута-
ции» ), и получил однохвостое распределение видов по родам. Далее
Юл допустил, что роды образуются подобно видам — за счет
«родовых мутаций», и после этого его модельные кривые распре-
деления видов по родам стали почти не отличимы от гипербол
Виллиса, описывающих реальность [Yule, 1924, с. 54, табл].
Совпадение оказалось так точно, что удивило самого Юла (ведь
в группировке таксонов некоторый произвол систематиков все-таки
неизбежен). Каждый вид в модели Юла мог порождать новые
независимо от своего возраста и численности, что было против
всех представлений об изменчивости — ведь мутация мыслится
как нечто, происходящее в организме, т. е. тем более вероятное,
чем больше имеется самих организмов. Это было уже серьезным
вызовом господствовавшим эволюционным представлениям: ведь,
по Дарвину, новый род образуется за счет многократных видообра-
зований, а не за счет однократной «родовой мутации», почти столь
же частой, как и «видовая мутация».
Именно тогда, в двадцатые годы, с расцветом генетики, в био-
логию вошел и утвердился тезис (заимствованный из физики
и химии), что истина проверяется только экспериментом. Четкие
повторяющиеся результаты — вот истина, а отвлеченные рассуж-
дения — вообще не наука. Однако крайности нередко смыкаются,
и судьба идеи Виллиса—Юла тому пример: они предъявили
четкий, многократно проверенный результат, но ученый мир прене-
брег и самими наблюдениями, и фактом многократного точного
85
совпадения этих наблюдений с простой и ясной теорией. Кривые
Виллиса не представляют, говорили тогда и потом, интереса для
биологии, поскольку таким же кривым подчиняется распределение
частот букв в языке, химических элементов в земной коре, ученых
по их публикациям и многое другое. (С тем же правом можно
заявить, что волновое уравнение бесполезно для радиотехники,
поскольку ему же подчиняются и круги на воде.) Сходство кривых
чисто случайно, поскольку эти явления несопоставимы, они опре-
деляются разными законами природы — таков был приговор тех,
кто считал себя специалистами в этой, еще не родившейся, дис-
циплине. (В действительности феномен Виллиса — типичный реф-
рен и в этом смысле неслучаен. Подробнее см. [Диалектика раз-
вития..., 1978, с. 156].)
Отчасти была виновата математика, которой пользовался Юл,—
теория ветвящихся процессов только зарождалась, Юлу пришлось
почти все доказывать самому, и он не мог не ввести ряда упроще-
ний, ставших вызовом биологии. Сейчас изучены ветвящиеся про-
цессы разных типов, и можно добиться совпадения с виллисовыми
кривыми, не шокируя биологов. Например, теперь известно, что
распределение числа п видов крупного таксона, существовавших за
все время жизни таксона, выражается кривой n~s/2 [Золотарев,
1983, с. 37 — 38]. Основная же беда была в неподготовленности поч-
вы: во все времена новые законы природы воспринимались с тру-
дом, а Виллис и Юл указали на один из самых общих законов при-
роды — «однохвостую» статистику.
Строго говоря, она была известна и до них итальянским стати-
стикам: еще в 1830 г. статистик Адриано Бальби обнаружил, что
так распределены люди по их доходам, причем было отмечено, что
более 75% лиц имеют доход ниже среднего по стране [Степанов,
1844, с. 278]. Однако лишь через 70 лет этот факт вошел в обиход
науки — благодаря трудам Вильфредо Парето, с чьим именем
обычно и связывается. В 1927 г. Дж. Ципф установил гиперболиче-
ское распределение слов по частоте их употребления в языке [Ара-
пов, 1981].
Феномен Виллиса интересен в нескольких отношениях. Во-пер-
вых, следует усомниться в прежнем взгляде на эволюцию как на
сдвиг средних. Скорее речь может идти о смене ведущих кластеров.
Во-вторых, возможно, что однохвосто всякое распределение, несу-
щее в себе свою историю, а изображенные на рис. 2 кривые, с кото-
рыми традиционно работали эволюционисты, как раз и не содержат
исторического элемента. В-третьих, система организмов оказалась
не беспорядочным нагромождением таксонов, а именно системой,
т. е. чем-то единым, имеющим свою четкую структуру. Биологи
энергично пытаются избавиться и от массы мелких родов, и от не-
удобных гигантских родов, но ничего не выходит, (В частности, те
же роды астрагал и крестовник насчитывают теперь более чем
по 2 тыс. видов.)
Виллис обнаружил гиперболы не только в распределении таксо-
нов, но и в распределении индивидов по видам, явившись тем са-
86
мым одним из основателей учения об относительном обилии ви-
дов (подробнее см. [Engen, 1979]). Именно это обилие может быть
сопоставлено с частотой встречаемости слова по Ципфу, так что
понятие статуса слова [Арапов, 1981, с. 137 — 138], означающее
его значимость в речи, можно сопоставить со статусом вида (его
значимости в экосистеме).
Очевидно, что для самых разных наук важно понять, почему
одни случайные величины распределены симметрично (по Гауссу,
Пуассону и т. и.), а другие — несимметрично (по Виллису, Ципфу
или Парето).
3.6. Симметрия и экстремальность случайного
Случайность привыкли сопоставлять с отсутствием всякого поряд-
ка, в том числе — и симметрии. Однако если игральную кость ки-
дать много раз, то разнообразие исходов окажется довольно симмет-
ричным: частота выпадения каждой грани будет близка к ‘/б. Еще
300 лет назад математик Якоб Бернулли заметил, что симметрия
случайного явления здесь гораздо более глубока, нежели симмет-
рия самой кости. Бернулли предлагал рассматривать всякое слу-
чайное событие как набор равновозможных. Это можно пояснить
так: если кость несимметрична и потому падает на грань № 1 с ча-
стотой p/q в среднем, то можно вместо этой кости взять другую,
симметричную д-гранную кость, у которой р граней имеют один
и тот же номер, № 1; эта кость будет падать на грань № 1 с частотой
p/q. (Заметим, что сам Бернулли понятием кости на пользовался.
Кстати, симметричную д-гранную кость проще всего представить
себе как длинную призму с правильным д-гранником в основании.)
В этих предположениях Бернулли доказал свой знаменитый закон
больших чисел, гласящий, что при большом числе повторений слу-
чайного испытания частота каждого исхода близится к определен-
ному пределу, именуемому вероятностью этого исхода (анализ см.:
[Чайковский, 1990]).
Ни одна теорема не вызывала столько споров, сколько эта, хотя
ее формулировка и доказательство вполне ясны. Неясно было, ка-
кое она имеет отношение к случайности, поскольку никаких
свойств случайности в доказательстве не используется. Бернулли
в действительности доказал лишь следующее. Вместо бросания
одной д-гранной кости N раз, можно бросить N костей 1 раз, причем
число различных исходов не зависит от того, брошены они или раз-
ложены по определенному правилу: их будет qN. Если у каждой
кости закрашено р граней, то можно сосчитать, сколько костей ле-
жит на закрашенных гранях. Доля их равна p/q, это очевидно,
но Бернулли доказал большее: эта доля близка к p/q не только в
полной совокупности раскладок, но и почти в каждой раскладке
(если все они различны); а это значит, что если взять раскладок
много, то доля близка к p/q практически всегда.
Учтя каждую возможную раскладку ровно 1 раз, Бернулли тем
самым признал все раскладки равновозможными, т. е. вместо са-
87
мой случайности рассматривал ее симметрию. Этот-то ход мысли ’
и вызвал споры, но нам здесь достаточно того, что фактическое вы- I
полнение закона больших чисел в природных явлениях говорит
о действительном наличии этой симметрии в реализации случай-
ных явлений.
Изучая случайное явление, мы обычно интересуемся какой-то
случайной величиной — например, числом п выпадений грани № 1,
если кость брошена N раз. Величину p/q можно назвать центром
распределения случайной величины n/N (для обычной игральной
кости p/q={ /в). Частоты n/N, которые намного больше или намно-
го меньше, чем p/q, реализуются очень редко, что и выражается
колоколообразной кривой плотности распределения, похожей на
гауссову (рис. 1). Такие распределения, равно как и порождающие
их случайные процедуры, назовем центральными. Они очень удоб- ,
ны тем, что вместо самих случайных величин можно, как правило,
рассматривать их средние значения и делать, если нужно, поправ-
ку на дисперсию (разброс случайных значений). Так, если дорога
на работу занимает у вас в среднем час, то вы можете выходить из
дому за час 20 минут до начала работы и не думать о случайности.
Возникает соблазн считать саму природу неким царством средних
величин, а цель науки — в выявлении и взаимоувязке этих сред-
них. Идеология Кетле к этому и призывала. Однако в природе
взаимодействуют не средние значения, а сами объекты, и если их
взаимодействие достаточно сложно, то апелляция к средним может
оказаться бессмысленной, как мы уже видели в 3.3.
Однако бывают случайные величины совсем других видов. Если,
например, следить не за данной гранью, а за числом выпадающих
очков, то при бросании 6-гранной кости мы будем с равной часто-
той получать значения 1, 2, 3, 4, 5, 6. Это — равномерно распреде-
ленная случайная величина.-Так же равномерно распределено зна-
чение угла <р между начальным и конечным положением стрелки, ,
свободно вращающейся на оси, если ее ^раскрутить и ждать, пока |
она остановится. А вот tg ф распределен сложнее, с плотностью (
g(х) 77, 2\ ’ где
л(1+г )
Она тоже имеет колоколообразную форму, но затухает (при
х—>-оо) гораздо медленнее, чем кривая Гаусса, а потому случайная
величина х не имеет ни среднего значения, ни дисперсии. Ее рас- ,
пределение называется распределением Коши.
Замечательно, что если взять очень много независимых случай-
ных величин (не обязательно центральных) и складывать одно-
временно реализующиеся их значения, то суммарная случайная ве-
личина почти всегда распределена по Гауссу — в этом состоит цен- ;
тральная предельная теорема теории вероятностей. У нее есть два
веских основания называться центральной: во-первых, она —
центр теории и приложений, объяснение того, почему мы столь
часто встречаемся с гауссовым распределением; а, во-вторых, это
название подчеркивает главную роль центральной части [Феллер,
88
1964, с. 189], т. е. пиков плотности распределений, имеющих коло-
колообразную форму и затухающих быстро.
Теорема утверждает, в наших терминах, что сложение незави-
симых случайных величин — тоже центральная процедура (более
сложная, чем схема Бернулли, т. е. чем серия независимых броса-
ний кости). Центральными являются и многие случайные процес-
сы (процедуры, в которых случайное событие зависит от исхо-
да предыдущих) — они часто приводят к гауссову распреде-
лению.
Кроме биномиального и нормального, к центральным относятся
пуассоново, логнормальное и другие подобные распределения, но
не относится, например, распределение Коши, хотя и похоже на
гауссово по форме кривой: из-за отсутствия конечной дисперсии
здесь нельзя «сделать поправку на дисперсию», т. е. нельзя ни
в каком приближении пренебречь нерегулярностью случайной ве-
личины.
Нормальный закон встречается столь часто (и в этом смысле
нормален) потому, что образуется при сложении почти всего, что
угодно. Именно поэтому издавна общепринят прием: чтобы опреде-
лить какую-то величину поточнее, надо измерить ее п раз, сложить
результаты и сумму разделить на п. С осмысления этого приема
и началась история нормального закона [Adams, 1974, с. 33].
Сходная картина — при перемножении случайных величин:
при этом складываются их логарифмы, и сумма сходится к лог-
нормальному распределению. Однако при более сложном взаимо-
действии случайностей картина меняется принципиально. Так, ес-
ли складываются квадраты случайных величин, то получается рас-
пределение «хи-квадрат», затухающее (при .т—>-оо) как е~х [Вар-
ден, 1960, с. 118]. Еще сложнее дело в ветвящемся процессе —
здесь перемножаются только средние численности Et, а не сами
случайные величины: если в момент t каждый индивид оставляет в
среднем (l-f-sj потомков, то Е t— (l-|-si) (1+«2) •••(1+«1), что в
частном случае постоянного s дает формулу (1); но реальная чис-
ленность большинства клонов, как мы видели в 3.3, вовсе не приб-
лижается со временем к Et. Здесь отсутствие центральности ра-
спределения вызвано тем, что случайная численность в момент t
зависит от численности в момент t—1. Бывает, что даже сумма
независимых случайных величин совсем не центральна. Так, если
каждое слагаемое распределено по Коши, то и сумма будет распре-
делена тоже по Коши [там же, с. 125]. Когда же справедлива цент-
ральная предельная теорема?
Она справедлива, грубо говоря, тогда, когда случайности обла-
дают тройной симметрией — равновозможностью, аддитивностью
и равномерностью (подробнее см.: [Чайковский, 19886]). Равно-
возможность означает, что случайность представима в виде броска
или серии бросков одной и той же симметричной кости, аддитив-
ность — что случайности складываются (или перемножаются),
а равномерность — что ни одна из случайностей не имеет слиш-
ком большой дисперсии (разброса значений). Тройная симметрия
89
перемалывает, словно мельница, любую совокупность обладающих
ею случайностей в одну случайность — гауссову (см. рис. 1в).
Долгое время молчаливо считалось, что такой симметрией обла-
дают все случайности, и до сих пор мало кто задумывается над тем,
что, например, слова «мутация случайна» подразумевают совсем не
ту случайность, которую мы видим, бросая кость.
Кость, сколько ее ни бросай, всегда упадет на одну из шести
граней — вот весь спектр ее элементарных возможностей, так что
можно говорить об определенном соотношении частот разных гра-
ней. Если же вернуться к примеру с клоном бактерий, то оказывает-
ся, что элементарная возможность здесь — численность клона в мо-
мент Т; а она может оказаться любой — от 0 до 2 т; другими слова-
ми, здесь можно ввести только очень странную «игральную кость» :
перед каждым бросанием у нее случайным образом изменяется
число граней, причем пределы этих изменений (спектр возмож-
ностей) очень быстро растут. Так же в принципе, хотя и слож-
нее, растет со временем спектр возможных мутаций, поскольку
каждая тоже может зависеть от любой из предыдущих. Эти случай-
ности тройной симметрией не обладают и не приводят к централь-
ным процессам.
Мы видели, что спектр исходов ветвящегося процесса распада-
ется на два основных кластера. При описании на традиционном
языке это выражается в неограниченном росте дисперсии, т. е. для
оценки вероятностей событий по их наблюдаемым частотам следует
брать нереально большое число независимых опытов. Другими сло-
вами, закон больших чисел фактически не выполняется, и о вероят-
ностях (предельных частотах) говорить нельзя. Привычная стати-
стическая методология рушится, требуется прилагать другую —
методологию системности.
Случайное выживание порождает возможность следующей слу-
чайности (выживания в следующем поколении), тогда как случай-
ная гибель никакой новой случайности породить не может — вот,
наглядно говоря, источник асимметрии, порождающий однохво-
стость распределения клонов по численности. Со временем левый
конец плотности распределения (доля вымерших клонов) все более
поднимается, чтобы стать высшей точкой плотности распределе-
ния. Левый хвост, так сказать, редуцируется, зато правый стано-
вится, столь же образно говоря, все толще и толще — он выражает-
ся не кривой вида е~х‘ (как у гауссоиды), а кривой вида е~х или
даже х~1~у, где х — число особей в клоне 0<у<2 (о роли пара-
метра у в сходимости распределения см. [Золотарев, 1983]). Вот
почему на кривой 1 рис. 3 мы не видим левого склона — кривая ста-
ла монотонно падающей уже в третьем поколении.
Самое частое нарушение симметрии случайного процесса —
взаимодействие случайностей, зависимость случайных компонент
друг от друга. Ведь в реальной колонии, где особи конкурируют
за «место под солнцем», размножение одной способствует гибели
другой. Поэтому можно сказать, что наблюдаемые случайности
принадлежат единой системе, в которой существенные параметры
90
связаны нежестко; при этом Ожидать гауссовости нельзя 3. Когда
симметрия нарушена, в системе обычно возникает краевой процесс,
т. е. процесс, описываемый однохвостым распределением, у кото-
рого, максимум плотности — с левого края кривой.
Естественно встает вопрос о краевой предельной теореме, кото-
рая указала бы простой способ обходиться с краевыми распределе-
ниями й определила бы те условия, в которых эти распределения
естественно ожидать. Для нее предложено уже много частных фор-
мулировок и еще более частных их доказательств. Для бактерий на
чашках «теорема» звучит примерно так: распределение клонов по
их численности принимает со временем краевой характер, вслед-
ствие чего большинство клонов имеет нулевую численность, но
большинство живых особей сосредоточено в немногих крупных
клонах. То же можно сказать о распределении видов организмов по
родам и о многом другом. Лингвистический вариант теоремы см.
у Арапова [1981, с. 139]. Сама общность феномена краевых про-
цессов заставляет пересмотреть отношение к ним при построении
теорий упорядочения конкретных разнообразий: представляется
излишним построение множества частных моделей, призванных
объяснить, откуда в данном конкретном разнообразии возникает
краевое распределение,— ведь не поясняем же мы, откуда в каждом
конкретном случае берется нормальное распределение.
Как взаимодействие зарядов искривляет силовые поля, нару-
шая симметрию силовых линий, так и взаимодействие случайно-
стей, выражаясь фигурально, искривляет их распределение, нару-
шая симметрию случайных событий (подробнее см. [Чайковский,
19886]). Отсюда простое правило: встретив краевое распределение,
искать систему, объединяющую случайности, а встретив нежест-
кую систему — искать краевое распределение. И наоборот: встре-
тив гауссово распределение, можно, хотя и с некоторым риском,
допустить, что соответствующие параметры независимы 4 — таким
правилом давно и с успехом пользуется зоолог Л. Л. Численко
[1977] в своих поисках органической системности.
Нарушение тройной симметрии сопровождается возникнове-
нием новой, более сложной симметрии (нового инварианта): в са-
мых разных системах мы видим один и тот же тип распределений —
гиперболический. (Это, как и законы листорасположения,— до-
вольно редкий для биологии пример рефрена, выражаемого форму-
лой.) Недаром системолог Э. Янч считал усложнение симметрии
одним из главных свойств эволюции, о чем пойдет речь в 9.4.
3 Пониманием этого обстоятельства я обязан беседе с техноэволюционистом
Б. И. Кудриным. Насколько мне известно, нет пока четкого критерия, позволя-
ющего различать системы с гауссовой и негауссовой случайностью.
4 Риск в том, что наблюдаемая статистическая независимость не означает
подлинного отсутствия причинной связи (см. пример с числом л в 3.2), которая
обычно и интересует исследователя. Самый наглядный пример, когда статисти-
ческая независимость и гауссовость возникают в детерминированной системе,—
так называемые стохастические динамические системы [Заславский, 1984].
Хотя в них система (задаваемая, например, дифференциальными уравнениями)
и налицо, но случайность их не носит системного характера, а обладает тройной
симметрией.
91
Почему тройная симметрия приводит именно к нормальному
распределению, а ее нарушение чаще всего — к гиперболическим?
Вопросы столь высокой общности могут решаться лишь с позиций
самых общих принципов естествознания; в качестве таковых со
времен Мопертюи выступают экстремальные принципы. Этот круг
идей мы обсуждали ранее [Чайковский, 19886] и здесь отметим
только, что симметрия нормального распределения более глубока,
чем других, и носит экстремальный характер [Уиттл, 1982, с. 199—
202, 270]. В частности, нормальное распределение можно вывести
из требования максимума энтропии 5 распределения. С этой пози-
ции нормально распределено то, что обладает максимальной неупо-
рядоченностью, какая еще допускает устойчивость частот (наличие
вероятностей), а при снятии этого допущения распределение ста-
новится краевым. Однако познавательный статус подобных экстре-
мумов пока неясен.
Глава 4. РАЗНООБРАЗИЕ, ПОИСК
И ПРИСПОСОБЛЕННОСТЬ
Предыдущие результаты могли относиться и к живым, и к нежи-
вым разнообразиям, например, ветвящиеся процессы и отбор ис-
следуются в химии автокатализа. Специфика же разнообразия ор-
ганизмов в том, что оно создается путем наследования изменений
при смене поколений. В ходе этого процесса заполняется таблица
архетипа (по Мейену), т. е. идет эволюция. Мы будем различать
заполнение прежней таблицы (идиогенез) и появление принци-
пиально новых строк в таблице (арогенез), хотя эти процессы
в природе нерасчленимы. В этой главе мы в основном займемся
адаптивным идиогенезом, т. е. идиоадаптацией по А. Н. Северцову,
чтобы выявить элементарные механизмы надорганизменного обуче-
ния.
4.1. Дискомфорт и генетический поиск
С появлением генетики (1900-е годы) приспособление вида стали
понимать как распространение полезных мутаций и их комбина-
ций, происходящее при смене поколений. В 1960-х годах на языке
генетики стали описывать и прижизненное приспособление особи,
толкуя его как изменение активности генов (точнее, оперонов).
Обе схемы следовало увязать в одну, тем более, что появились дан-
ные о наследовании клеткой не только ее генов, но и ее процессов;
исследователи стали допускать наличие «состояния, при котором
одни гены активны, а другие подавлены и которое передается во
время клеточного деления» [Женермон, 1970, с. 669]. В 1973 г. бы-
ла предложена математическая модель двухоперонного триггера,
демонстрировавшая, как дочерняя клетка может получать от мате-
ринской индукторы и репрессоры, чтобы у дочерней были активи-
рованы, в основном,те же гены, что и у материнской [Ратнер, 1975,
92
с. 156]. Если внешнее воздействие переключило такой триггер, то
оно создало наследственное (пусть и недолговечное) изменение без
изменения в генах. Позже один из авторов модели, Р. Н. Чураев,
развил на этой основе целую концепцию «эпигеномного» наследо-
вания, обходящегося без изменений в текстах ДНК [Молекулярные
механизмы..., 1985, с. 154].
Триггер (логическая схема с двумя действиями, чередующими-
ся в зависимости от внешнего сигнала) обладает некоторой разум-
ностью по сравнению с чисто случайным «поведением» бросаемой
монеты, что видно из следующего. Пусть среда может действовать
на объект двумя способами — штрафом и поощрением, причем
первое действие объекта штрафуется с вероятностью р\, а второе —
с вероятностью р2^=р|. Если объект — триггер, меняющий дейст-
вие при штрафе и не меняющий при поощрении, то, как легко вы-
числить [Цетлин, 1969, с. 25], его средний выигрыш W больше,
чем выигрыш ИД объекта, меняющего действие чисто случайно.
Улучшить его поведение (получить W>Wr) можно, сделав триггер
инерционным, т. е. меняющим действие не при первом штрафе.
Для этого он должен уметь считать время, т. е. иметь цепочку
внутренних состояний, которые меняются одно за другим без изме-
нения действия (невидимо для внешней среды). Таков автомат
с линейной тактикой (АЛТ), предложенный М. Л. Цетлиным
в 1961 г.
Вообще, конечным автоматом называется объект, имеющий п
состояний и заданное правило перехода из одного состояния в дру-
гое. Состояния объединяются в к групп, именуемых действиями
автомата; автомат, выполняющий действие г, штрафуется внеш-
ней средой с вероятностью р (поощряется с вероятностью 1—р ),
не зависящей от номера состояний внутри группы. Правило смены
состояний для АЛТ таково: если в момент t было поощрение, то
в момент t-[-l номер состояния увеличивается на 1 или (если он
уже был равен максимальному числу d — глубине памяти АЛТ)
не меняется; если же в момент t был штраф, то номер состояния
уменьшается на 1 или (если он уже был равен 1) происходит смена
действия на следующее (действия образуют кольцо), и в нем авто-
мат занимает тоже номер 1. Автомат задается двумя графами:
один — поведение при поощрении, другой при штрафе. На рис. 4а
изображен АЛТ для случая к=2, d=5. При d=l АЛТ становится
триггером, а при d—>-оо он оптимален, т. е. реализует практически
только то действие, штраф за которое минимален. Для приемлемой
разумности d должно заметно превышать к—1, ар — достаточно
различаться.
АЛТ можно по-разному модифицировать. На рис. 46 изображен
автомат, после поощрения сразу занимающий устойчивое (самое
глубокое) состояние, а при штрафе ведущий себя аналогично АЛТ,
с той разницей, что номер действия выбирается случайно, с вероят-
ностью 1/к—1 (пунктир). Он также оптимален при d—>-оо, но инте-
ресен тем, что при fc=4 моделирует точковый мутагенез.
Механизм репликации ДНК (как и механизм транскрипции)
93
Граф поведения £ случае
поощрения
Граф поведения £ случае
штрафа
7$
ОЧ-НХ—О< о< о
РИС. 4. Разумноинерционные автоматы: А) автомат с линейной тактикой (к—2)
d=5); Б) автомат, моделирующий инерционный точковый мутагенез (k=4, d=4);
В) автомат со сравнивающей тактикой (детерминированный автомат Милютина,
к=2, d=8), демонстрирующий антиинтуитивное оптимальное поведение. Действия
автоматов обозначены через число возможных действий — через к, число со-
стояний в каждом действии — через d.
воспроизводит в данной точке новой полинуклеотидной цепи тот из
четырех нуклеотидов, какой нужно (согласно прежней цепи), но
механизм может ошибиться и вставить в цепь не тот нуклеотид —
это и есть точковая мутация; в норме ошибки происходят с ничтож-
ной частотой 10“ —10“ на нуклеотид, и мы ими пренебрежем.
(Иногда по традиции толковой мутацией называют любое измене-
ние в рамках одного гена — тогда ее частота в норме близка
к 10“ —10“ на особь.) Зато, как давно известно (литературу см.
[Чайковский, 1976]), действие любого фактора, резко ненормаль-
ного для жизни, заметно повышает частоту мутаций. При. этом
большинство факторов вызывают не сразу саму мутацию, а пред-
мутационное повреждение, которое чаще всего исправляется одним
из механизмов репарации ДНК; только если ни один из них не
94
сработал, происходит мутация. Поврежденный участок ДНК может
находиться в неустойчивом состоянии несколько поколений, и су-
ществует особый тип репарации (пострепликативная), работаю-
щий-в этих случаях [Общая генетика, 1978; Ауэрбах, 1978].
Задержка акта мутации, вне зависимости от ее молекулярного
механизма, играет ключевую роль: если клетка не повреждена, то
система репарации редко доводит локус до мутации; если же функ-
ционирование клетки нарушено, то это так или иначе помешает
правильной работе системы репарации, а значит — повысит веро-
ятность мутации для всех нестабильных локусов. Каждый локус,
подвергшийся действию мутагена, в течение некоторого времени
как бы выбирает, мутировать ему или нет, причем «выбор» его бы-
вает положительным тем реже, чем лучше адаптирована клетка.
Такой процесс назовем инерционным мутагенезом. Чем дольше за-
держка, тем более мутагенез зависит от интегральных, а не от мгно-
венных характеристик среды, т. е. тем более он целесообразен (со-
действует выживанию).
Нормальное считывание ДНК соответствует на рис. 46 устой-
чивому состоянию автомата, действие мутагена, а также срыв
каждого отдельного акта репарации являются штрафом и обозна-
чаются сдвигом на 1 шаг в сторону мутации, а пунктир обозначает
саму мутацию. В отсутствие «штрафов» мутабильность обычно
низка настолько, что нельзя ожидать появления даже одной пары
определенных нуклеотидных замен даже в огромной колонии бак-
терий. Чтобы гарантировать такую пару хотя бы для колонии мас-
сой в 100 т, надо поднять точковую мутабильность в тысячу раз.
Грубая оценка, произведенная мной в 1972 г., показала для од-
ного частного случая, что инерционный мутагенез в принципе
может обеспечить нахождение адаптивной пары мутаций, вредных
по отдельности [Чайковский, 1976, с. 158]. В 1976 г. был получен
первый экспериментальный результат такого рода: бактерии, обыч-
но растущие на рибитоле, почти (но не совсем!) лишенные способ-
ности жить на ксилитоле (эти пятиуглеродные спирты различают-
ся положением одной ОН-группы), приобретали способность роста
на ксилитоле за счет пары точковых мутаций, обеспечивающих,
как решили авторы, замену двух аминокислот в ферменте рибитол-
дегидрогеназе [Hartley е. а., 1976]. Тогда же появились данные
и о том, что рост мутабильности в сотни раз может быть вызван
чрезмерной интенсивностью работы гена, и Л. Н. Дроздов-Тихоми-
ров высказал (в беседе со мной) важную мысль, что таким путем
могут изменяться те самые гены, которые являют собой «узкое мес-
то» метаболизма. (Это похоже на правило Ле Шателье в химии.)
Были открыты и химические способы повышения мутабильности
в 105 раз, а также — избирательного ее повышения [Сох, Gibson,
1974]. Все это вместе взятое дало право уверенно говорить о прин-
ципиальной возможности нахождения адаптивной тройки мутаций,
вредных по отдельности, путем инерционного точкового мутагене-
за. Много это или мало? Увы, очень мало.
Комментируя первые данные по «экспериментальной эволюции
95
бактерий», Тай Тэ By [Wu, 1978] писал: таким путем можно объяс-
нить слабое изменение активного центра фермента, но уже чуть
более сложные изменения ферментов требуют других механизмов.
Так, он сравнил три дегидрогеназы, у которых активные центры
сходны, но находятся в разных участках белковых цепей — этого
толковый мутагенез сделать не мог. Добавим, что он вообще бес-
силен сделать что-либо у многоклеточных организмов, поскольку
те не исчисляются триллионами.
И все же толковый мутагенез и его автоматная модель представ-
ляют теоретический интерес, так как указывают на реальность
феномена генетического поиска. Нормальная функция реплика-
ции — копирование текстов, а генетическим поиском мы будем
называть те чрезвычайные режимы работы генетической системы,
когда в ней изготовляются новые тексты ДНК. Рассмотренный
выше инерционный мутагенез — самый маломощный вид генетиче-
ского поиска, при котором в тексте заменяются отдельные буквы.
О более мощных механизмах поиска речь пойдет в 4.2 и в части II.
Если сам по себе рост мутабильности не может помочь адапта-
ции (так как на одну полезную мутацию приходятся тысячи вред-
ных — см. 3.4), то инерционный мутагенез может. Объектом есте-
ственного отбора выступают при этом не сами мутации, а генетиче-
ская система, способная их порождать. Она здесь — квант селек-
ции [Чайковский, 1976, с. 162], т. е. несомненно полезное при-
обретение, неразложимое отбором на части (части этого качества
не могут в реальных условиях отбираться по отдельности), но
способное быть отобранным как целое. Напомним, что еще Богда-
нов [1913, с. 44, 91] полагал «основою подбора» не признаки орга-
низмов, а их жизнеспособность.
Если методологическая роль клональной модели отбора виде-
лась в мажорировании реальных процессов, то роль автоматных
моделей — ориентационная: они призваны показать новые логиче-
ские возможности, открывшиеся перед теоретиками эволюционной
генетики. С этой точки зрения интересен также автомат со сравни-
вающей тактикой — ACT [Цетлин, 1969, с. 32 — 36]. Разумность
этого автомата достигается не столько за счет инерционности (хотя
и она необходима), сколько за счет регулярной смены действий.
Если АЛТ считает штрафы и поощрения, сохраняя одно и то же
действие, то ACT — меняя их. Наиболее интересный вариант ACT
нашел в 1965 г. А. А. Милютин: его конструкция вообще наилуч-
шая, какая возможна при данной глубине памяти. Автомат ока-
зался вероятностным: в каждом действии есть одно состояние, по-
пав в которое, он никогда не меняет действие при поощрении и ред-
ко — при штрафе. Милютин предложил и детерминированный
автомат, ведущий себя очень сходно и, следовательно, получаю-
щий почти тот же средний выигрыш.
На рис. 4в этот ACT изображен для к=2, (1=8. Замечателен
он тем, что ведет себя антиинтуитивно: он состоит из двух под-
систем, каждая из которых почти или вовсе лишена целесообраз-
ности: в верхних четверках состояний смена действия обязательна,
96
независимо от штрафа, а в нижней четверке он при штрафе вовсе
не считает время, так что сама по себе нижняя четверка не лучше
триггера. Переход от нее к верхней четверке возможен только после
трех поощрений, а затем штрафа. И все же легко видеть, что если
действие fa чаще штрафуется, а /2 — чаще поощряется, то автомат
быстро попадает в нижнюю четверку справа, откуда выходит редко.
Пусть ACT моделирует какую-то клональную генетическую
систему, т. е. надо, наблюдая фенотипы /1 и /2, судить о генотипах
(состояниях). Пусть такт времени — поколение, а штраф — ненор-
мальное для особи состояние среды. Угадать закон перехода от
устойчивого проявления/1, к устойчивому проявлению /2 практиче-
ски невозможно, и генетик вернее всего определит здесь 3 генотипа:
два — с устойчивыми фенотипами fa и /2 и один — обеспечиваю-
щий чередование поколений. В большой неоднородной колонии он
будет то и дело наблюдать переходы (между этими тремя «гено-
типами»), которые, разумеется, идентифицирует как случай-
ные мутации (это — случайности типа А из 3.1). Если часть особей
гибнет, то постепенное накопление фенотипов /2 соблазнительно
приписать естественному отбору, а чередование поколений описать
как специальную адаптацию к изменчивости среды.
В действительности генотипов здесь 8, а случайных переходов,
отбора и адаптации к изменчивости нет вовсе. Автоматы реагируют
на состояние среды однозначно и могут все, без гибели (см. 4.4),
перейти к /г- Гибель фенотипов fa лишь ускоряет накопление /2 —
вот в чем состоит здесь роль отбора. ACT выступает тоже как квант
селекции. Такой переход генетической системы из одного устой-
чивого состояния в другое (через неустойчивое) называют генети-
ческой стабилизацией. Описанная выше абстрактная модель во
многом сходна с реальной генетической стабилизацией: возможна
ситуация, когда «фиксация еще не ставшего наследственным изме-
нения будет происходить в момент устранения фактора внешней
среды, вызывающего данную модификацию» [Оленов, 1946, с. 257,
сноска 4].
4.2. Организующая роль случайности
Как мы уже говорили, наилучшим автоматом является тот вариант
ACT, в саму структуру которого включен механизм случайного
выбора действия с определенной вероятностью. Детерминирован-
ный ACT (рис. 4в) не может работать без случайности среды: он не
сможет покинуть то невыгодное действие, за которое только штра-
фуют или если невозможны 3 поощрения подряд, а затем штраф
(в случайной среде это возможно всегда). Такая ситуация доста-
точно типична: существует множество объектов, которые без слу-
чайностного компонента распались бы на отдельные бессмыслен-
ные состояния или движения. Недаром с древности неотъемлемой
частью многих социальных действий является жеребьевка, а в наше
время в науке и технике распространены методы случайного поис-
ка. Покажем на примере коллектива автоматов, как введение слу-
4 Ю. В. Чайковский
97
чайности в акт поведения радикально упрощает поиск и повышает
надежность работы коллектива.
Что позволяет клеткам (одного организма или одной колонии)
делиться синхронно? Попытки моделировать этот процесс приве-
ли кибернетиков к задачам о синхронизации большого коллектива
N автоматов. Основной интерес представляла задача с одинаковыми
автоматами, имеющими небольшое число g‘ состояний, при котором
автомат неспособен считать ни время, ни число членов коллектива
[Варшавский, Поспелов, 1984]. Решения были остроумны, но все
они, пока рассматривались детерминированные задачи, обладали
общим дефектом: сбой одного автомата хотя бы в одном такте или
даже малое перемещение автомата среди соседей разрушали весь
процесс синхронизации. Биологического значения такие ненадеж-
ные схемы не имели. Однако в 1969 г. В. И. Варшавский изменил
постановку задачи: он предложил говорить об е-синхронизации.
т. е. о таком поведении, когда в синхронное состояние g=m прихо-
дят одновременно не менее (1 —е) автоматов, где 0<е<С1, при-
чем до этого в состоянии g =т побывало не более e.N автоматов.
Этот статистический подход позволил построить очень надежную
систему и сильно упростить сами автоматы.
Вот одна из схем Варшавского. Каждый автомат имеет (т-(-1)
состояний, в каждый такт t образуется N/2 случайных пар авто-
матов, и состояние каждого в момент t-1-1 есть следующая функция
от состояний обоих в момент t. В начальный момент все находятся
в нулевом состоянии: g (0)=0, а при t=l один из них (первый)
переводится внешним сигналом в состояние g, (1)=1. Остальные
действуют по правилу:
g(£+l)=gj (« + 1) =
'О, если gi(0=g/(f)=O
ш, если g (t) =g;. (t) =m
,min{gj ( t), (£)}-|- 1 в остальных случаях.
Таким образом, автомат остается в нулевом состоянии, пока не
встретит партнера с другим состоянием; пара, попавшая в конеч-
ное состояние, также остается в нем; остальные пары «выравнива-
ются на отстающего» и делают шаг вперед. Доказано, что почти все
автоматы достигают состояния m в один момент. Суть процесса
в том, что выравнивание создает «единый движущийся фронт».
При 2V=1O6 и т=12 синхронизация наступает одновременно через
35—40 тактов для 95% автоматов или более, т. е. е не превышает
5%.
Эта модель не описывает (так же, как АЛТ, ACT и т. и.) никако-
го реального феномена, но раскрывает неожиданные информацион-
ные достоинства случайных пар. Случайные пары, объединяющие
свою информацию, достаточно обычны в биологии: таковы пчелы,
обменивающиеся содержимым желудков по возвращении в улей,
или бактерии, осуществляющие половой процесс (который у них
отделен от процесса размножения).
Как и в других автоматных моделях, процесс здесь может идти
без гибели участников, т. е. без отбора. Хотя отбор и ускоряет про-
цесс, но.отсутствие жесткой связи с ним дает то огромное преиму-
98
щество, что в коллективе могут одновременно распространяться
многие новые признаки — например, он может одновременно при-
ближаться к синхронизациям по различным параметрам и в то же
время максимизировать какой-то иной параметр. Именно это дает
надежду на понимание эволюции — ведь если бы она управлялась
только через размножаемость, то даже два независимых признака
одновременно распространяться не могли бы: отбор в принципе
не различает обладателей равных размножаемостей, как бы ни раз-
личались наборы их признаков.
Изящество и поучительность автоматных моделей порождают
иногда надежду, что коллективы одинаковых объектов могут все,
что поэтому разнообразие объектов ненужно, что достаточно, если
каждый из одинаковых индивидов обладает некоторым разно-
образием возможностей: «Никто еще не доказал, что однородные
коллективы и структуры могут эффективно решать все задачи, воз-
никающие перед техническими системами. Но никто не доказал
и противоположного!» [Варшавский, Поспелов, 1984, с. 173].
Недоказанность может служить базой лишь для осторожности, тем
более, что надежды авторов выходят далеко за рамки технических
систем — тут и биология, и психология, и искусство. Кстати, един-
ственный пример действительно сложной задачи заимствован ими
из искусства: оркестр без дирижера, известный некогда Персим-
фанс. «Взаимодействуя непосредственно друг с другом, прекрас-
ные музыканты, входившие в Персимфанс, легко обходились без
дирижера» [там же, с. 3], поскольку «каждый слушал каждого
и всех».
Что и говорить, опыт интересный, но свидетельствует никак не
в пользу цитированных авторов. В ансамблях без дирижеров всегда
есть инициатор, ведущий репетиции («первая скрипка»), и, как
рассказывали, был он и в Персимфансе. Еще важнее для нашей
темы, что ансамбль не был однородным'. 140 оркестрантов дели-
лись на 18 инструментальных групп, на несколько групп по основ-
ному месту работы (Персимфанс не был самостоятельным оркест-
ром) , а около трети ансамбля составляли пары, тройки и т. д.
родственников (последнее ясно из того, что 17% его составляли
однофамильцы) [Персимфанс, 1932]. Естественно, если группа,
освоившая данное произведение в обычном оркестре, лидировала.
Главное однако, заключалось в том, что Персимфанс, несмотря на
поддержку сверху, встретил свое десятилетие при последнем изды-
хании, а другие подобные ансамбли (их возникло около 10, у нас и
за рубежом) быстро исчезли. Только Укрсимфанс в Киеве протя-
нул 3 года, но и он не был самостоятельным оркестром. Причина
видится в том, что ансамбль без дирижера демонстрировал только
образец самоорганизации, но не творчества. Это признала даже во-
сторженная юбилейная брошюра: «И если крепкая художествен-
ная дисциплина, прекрасная сыгранность и сплоченность Ансамб-
ля с успехом заменила дирижера как организатора и этим дала
Персимфансу его первую и основную победу, то в другой творче-
ской части — в части замены дирижера как интерпретатора
4*
99
музыкальных произведений, как творческого толкователя партиту-
ры, Персимфанс сделал лишь первые и робкие шаги» [там же, с. 6].
Самоорганизация, демонстрируемая при е-синхронизации,
очень красива, но за нее надо дорого платить — случайное парное
взаимодействие означает непрерывное перемешивание коллектива
(как перемешиваются молекулы в сосуде с газом), а это означает
невозможность образования не только структуры из индивидов, но
и существенного разнообразия самих индивидов. В реальных кол-
лективах пары образуются (например, при половом процессе выс-
ших организмов) если и с элементом случайности, то далеко не
равновероятно. В качестве основной случайности, организующей
элементы в целое, чаще выступает другой феномен — мероно-
таксономическое несоответствие (3.1). Этим термином Мейен обо-
значил «отсутствие соответствия между классами частей и класса-
ми самих индивидов» [Мейен, 1984, с. 18], т. е. феномен, в принци-
пе отсутствующий в однородных коллективах. Чем значительнее
это несоответствие, тем выше и организующая роль случайности.
4.3. Поиск как игра
До сих пор мы рассуждали об организме, находящемся в некоторой
среде, характеристики которой в среднем постоянны, но из главы
3 знаем, что усреднение не всегда имеет смысл. Более того, постоян-
ных сред вообще не бывает, поскольку среда тоже состоит из эволю-
ционирующих объектов, а потому стационарные модели годны
лишь для грубого описания отдельных приспособительных реак-
ций, но не приспособления в целом. Можно, например, считать
стационарными (в вероятностном смысле) многие климатические
факторы, но следует помнить, что вид, приспосабливаясь к ним,
в то же время испытывает множество влияний со стороны менее
устойчивых факторов среды. Те процессы, когда среда сама реаги-
рует на действие нашего вида приблизительно с той же скоростью,
с какой вид — на действия среды, удобно моделировать с помощью
игры (об играх см. простой учебник [Льюс, Райфа, 1961]).
Простейший вид игры — матричная игра', в ней участвуют два
игрока, причем выигрыш одного является проигрышем другого
(антагонизм). Таков, например, процесс конкуренции двух попу-
ляций за общий ресурс. Каждый игрок может совершать конечное
число действий; если I игрок выбрал определенное действие г,
а II игрок — определенное действие /, то считается заданным чис-
ло а. — выигрыш I игрока (сумма, которую ему платит II игрок;
если она отрицательна, то победил II). Игра полностью задается
таблицей (матрицей) чисел а,, где l^y^n, которую обо-
значают как Л=||а ||. Бывают таблицы, в которых прямо можно
указать такой номеф I (строку), который для I игрока выгоднее
других независимо от того, какой номер j (столбец) выбрал II иг-
рок; и, соответственно, этот номер / для II игрока выгоднее других,
независимо от действия I игрока. Тогда говорят, что матрица А име-
ет седловую точку. В общем же случае выгодность любого действия
100
i зависит от выбора /, и решение игры существует лишь в вероят-
ностном смысле. Еще Джон фон Нейман в 1928 г. доказал основ-
ную теорему: для любой матрицы А существует решение игры —
пара оптимальных стратегий х° и у0 игроков и цена игры v{A),
т. е. выигрыш I игрока при этих стратегиях. Стратегия — это набор
частот (вероятностей), с которыми игрок применяет свои способы
действия, а оптимальна та стратегия, применяя которую, игрок
гарантирует себе в среднем максимальный возможный при дан-
ных atj выигрыш (равный и). При и>0 игра выгодна для I, а при
у<0 — для II игрока. Если обозначить стратегию I игрока векто-
ром х=(х1,..., хп), а стратегию II игрока — вектором y=(yi, ..., уп),
то средний (по независимым актам игры) выигрыш I игрока равен
М(х, у)=хАу=Х ха. у. причем решение игры находится из
функционального уравнения
п=шах min М(х, у)=х°Ау°.
Тот факт, что решение игры в вероятностном смысле существует
всегда,— еще одно свидетельство организующей роли случайности.
Когда оба игрока отказываются от случайных выборов действий,
то процесс повторения игры состоит в чередовании выигрышей
и проигрышей каждого, но, как правило, без приближения среднего
(по времени) выигрыша к цене игры.
Игровые модели интересны как в ориентационном, так и в ма-
жорирующем планах, и в этом смысле как бы противостоят тради-
ционным диффузионным моделям конкуренции, носящим имита-
ционный характер, т. е. пытающимся попросту отобразить какие-то
желаемые свойства объекта. Ранее [Чайковский, 1977а] было пока-
зано, как качественный анализ игровой матрицы может легко
сориентировать исследователя среди разнобоя частных результа-
тов, даваемых диффузионными моделями конкуренции. «В рам-
ках этой модели выяснено, в частности, что ген-мутатор (ген, кото-
рый сам не влияет на фенотип, но увеличивает мутабильность
других генов) если и может быть зафиксирован отбором, то лишь
при наличии в генотипе механизма, управляющего активностью
мутатора (в зависимости от условий существования особей клона) »
[там же, с. 1475—1476]. Это — обобщение идеи генетического
поиска на конкурентную ситуацию, т. е. ориентационная модель.
Далее, именно игровая модель наводит на мысль, что биологи-
чески бессмысленно рассматривать постоянные селективные цен-
ности s>0, поскольку даже в простейшем случае конкуренции
двух объектов мы видим не число, а матрицу. Для этой матрицы
есть единственное число v(A), которое характеризует игру как
таковую, а не какой-то способ играть в нее, но, как мы узнаем ниже,
этой характеристикой могут воспользоваться лишь очень хорошо
организованные объекты, а в отношении других приходится при-
знать, что превосходство числом не выразить.
Впрочем, в этом нет необходимости, так как для выживания
клону вовсе не нужно иметь высокого значения какой-либо сред-
101
ней величины, характеризующей скорость размножения,— ему
необходимо и достаточно сохранить при всех t ненулевую числен-
ность (подробнее см. [Маленков, Чайковский, 1979]). То же можно
сказать о других, более сложных числовых характеристиках орга-
низмов, экосистем или таксонов— «адаптивной высоте», «эволю-
ционном потенциале», «отрицательной энтропии» и т. д. Ранее
[Чайковский, 19886] уже говорилось, что ни на чем не основано,
например, допущение об экстремизации энтропии в биологических
объектах.
Наиболее важен игровой подход в мажорирующем отношении,
точнее — при определении границ, далее которых тот или другой
тип поиска бессилен. Именно этот подход дает решающий аргу-
мент для отказа от самых примитивных селекционных толкова-
ний биологической целесообразности. Первые игровые модели по-
ведения простых объектов рассмотрел в 1963 г. Цетлин. Исследо-
ванные им автоматы (АЛТ со случайной сменой действия) играли
хоть и целесообразно, но очень грубо: если в игровой матрице
не было ни целиком неотрицательной строки, ни целиком неполо-
жительного столбца, то автоматы играли вничью (М(х, у)->0 с ро-
стом времени), как бы сильно v ни отличалось от нуля. Столь гру-
бое поведение никак не может быть типичным для биологических
систем, поскольку те, приспосабливаясь к среде, часто используют
очень тонкие и мало влияющие на выживаемость обстоятельства
(см. 4.4).
Встает вопрос: каковы должны быть минимальные требования
к автомату, чтобы он мог участвовать в поиске решения матрич-
ной игры — точного или приближенного? Для ответа надо иссле-
довать структуру решения. Решить игру — значит найти такое рас-
пределение частот xt собственных действий, чтобы получать вы-
игрыш не менее цены v, как бы ни вел себя противник, причем цену
v тоже надо найти. То есть надо, чтобы при любом действии j про-
тивника выполнялось нестрогое неравенство М\х) =2 а. х ~^v.
Если противник решил аналогичную задачу, т. е. нашел ¥акие у,
чтобы при любом i выполнялось: М. (у) =2 а. у^.v, то фактически
могут выполняться лишь равенства (один и тот же выигрыш v при
любом действии), а номера г, /, для которых неравенства строги,
в решение войти не смогут (т. е. соответствующие действия в опти-
мальную стратегию войдут с частотой, равной нулю). Поскольку
в окрестности решения все величины Д, Mj почти равны друг другу
(или не участвуют в формировании решения), то нахождение его
путем поиска — тонкая вычислительная задача даже при точном
знании нужных величин.
Нас же интересует случай, когда поиск решения игры происхо-
дит путем сравнения несложных оценок, доступных автоматам
с небольшой памятью. Оценивать величины типа Mf(y) автомат не
может (для этого надо иметь неограниченную память), но на это
может быть способен большой коллектив одинаковых автоматов,
рассматриваемый как единый игрок. Разумность I игрока как цело-
го достигается сравнением между выигрышами Д групп, приме-
102
няющих в данный момент одно и то же (i-oe) действие. Здесь для
разумности тоже нужна инерционность, но она может достигаться
и без собственной памяти автомата — за счет малости частот пере-
хода. При этом роль коллективной памяти играет распределение
автоматов по действиям.
Сама по себе приближенная оценка средних в коллективе не-
сложна: достаточно между актами игры несколько раз обменяться
частью своего выигрыша с ближайшими соседями, чтобы в коллек-
тиве быстро выровнялась величина, близкая к М(х, у). Так, напри-
мер, поступают пчелы в улье — они регулярно обмениваются друг
с другом содержимым желудков, так что каждая, прилетев со взят-
ком, может грубо оценить удачность своего вылета. Сложнее, но
ненамного, оценить среднее Мк(у) по «своим» автоматам.
Опуская детали, отметим, что если противник коллектива при-
меняет заданную (Стратегию у, то коллектив, члены которого умеют
оценивать выгодность Sk=Mk (у)—М(х, у) собственного действия,
может находить действие i, гарантирующее максимальный выиг-
рыш. Однако если противник сам является аналогичным коллек-
тивом (сравнивающим разности М(х, у) — М (г)}. и матрица А
не имеет седловой точки, то знания лишь собственной ,S’/C недостаточ-
но, а надо знать хотя бы один из номеров г действий, к которым вы-
годно переходить. Коллективы автоматов, способных оценивать
лишь собственные выгодности, бесконечно блуждают вокруг точки
решения игры, даже не приближаясь к ней,— это показано уже на
примере игры с тремя действиями (т=п=3) [Чайковский, 1971].
Другими словами, покидая невыгодное действие, необходимо знать,
к которому из остальных действий переходить,— иначе будет хаос.
При этом достаточно, чтобы каждый автомат мог узнавать номер
одного из них,— если разные автоматы, выполняющие к-ое дейст-
вие в разные моменты t случайно узнают разные номера i
(S. (i)>0; i=/=k). Само значение величины 5 (i) знать необязатель-
но. Здесь снова налицо организующая роль случайности.
Тем самым, информационная граница видна четко, и, обнару-
жив оптимальность, мы обязаны искать информационные каналы,
ее обеспечившие. Объяснять оптимальность «отбором ненаправлен-
ных вариаций» оснований нет. Далее, как бы ни были сложно и раз-
лично устроены игроки (они могут различаться умом, памятью,
вовлеченностью в другие конфликты и т. п.), это может повлиять
только на достижение ими оптимальных стратегий, но не на свой-
ства самих этих стратегий, поэтому оптимальность следует описы-
вать и объяснять, не отговариваясь чрезмерной сложностью объек-
та. (С характерным синдромом оптимальности — равенством вы-
игрышей независимо от действия — мы встретимся в 5.3.) В этом
и состоит мажорирующий аспект матричноигровой модели.
Однако в подавляющем большинстве реальных ситуаций нельзя
даже приближенно полагать выигрыш одного равным проигрышу
другого (антагонизм), и наша игровая модель не работает. О пове-
дении автоматов в неантагонистических играх тоже есть работы
(например, указанные в книге [Варшавский, Поспелов, 1984]),
103
мы же рассмотрим другое усложнение игры — многоходовость.
Обычно игра состоит из многих последовательных ходов, и на
каждом своем ходу игрок должен делать выбор — определенный
или вероятностный, а выигрыш определяется всей последователь-
ностью ходов всех участников. Для антагонистической игры двух
игроков в 1953 г. Говард Кун доказал теорему: игрок может га-
рантировать себе выигрыш, равный цене игры v, только в том слу-
чае, когда помнит все прошлые свои ходы [Льюс, Райфа, 1961,
с. 211]. Это свойство игроков выполнено, например, в шахматах,
но не в тех карточных играх, где один игрок представлен двумя
партнерами: каждый из них хотя знает, какими картами ходил
партнер, но не знает, из каких карт тот делал выбор. Теорема
Куна показывает, какой поток информации во времени так же
эффективен, как поток ее в коллективе одновременно действую-
щих. В частности, теорема выражает одно из свойств прогрессивной
эволюции — расширение множества запомненного.
Игровые ситуации демонстрируют важный источник разнооб-
разия: если в стационарной случайной среде, штрафующей каждый
автомат независимо, все автоматы переходят в наименее штрафуе-
мое действие, т. е. ведут себя однообразно ', то в игре они вынуж-
дены, как правило, вести себя по-разному. На примере антагони-
стической игры мы видели, что высоко организованный игрок
(коллектив) распределяется в оптимальных пропорциях по своим
возможным способам действия, а низко организованный бесцельно
блуждает (меняет действия), но в обоих случаях налицо разнооб-
разие. Налицо оно и в более сложных конфликтах.
4.4. Смена функционирования и наследование
В моделях предыдущих параграфов индивид мог реагировать на
Дискомфорт только сменой генотипа или фенотипа, хотя в дейст-
вительности первой реакцией организма бывает смена поведения
или физиологических реакций. Животные достаточно автономны
в своем поведении: так, бабочка, имеющая меланистическую
(черную) мутацию, не садится на светлый фон, хотя инстинкт
не мутировал. Растения тоже обладают поведением: изгибаются
в поисках света и опоры, принимают стоячую или плавающую
модификации, даже обучаются (например: закаливание теплолю-
бивых растений), и еще Дарвин видел аналогию в поведении
животных и растений. Описано даже активное поведение бактерий
[Никитин, Никитина, 1978, с. 150]. Правда, некоторые паразиты
почти не обладают поведением, но они и не дают, как принято
считать, материала для эволюции. Словом, поведение особи следу-
1 Стационарная среда, в которой все члены коллектива (автоматы) штрафуются
или поощряются одновременно (с определенной вероятностью), также не допу-
скает однообразия (таковы климатические факторы, обработка ядохимикатами и
т. д.). Если автоматы способны к случайной смене действия, то при этом устанав-
ливаются устойчивые частоты распределения их по действиям, хотя каждый то
и дело меняет действие (эргодический случайный процесс).
104
ет отнести (что не раз делалось) к элементарным актам, постав-
ляющим материал для эволюции.
Организм, в том числе и одноклеточный, стремится сохранить
неизменной свою внутреннюю среду, используя для этого все
доступные ему реакции — поведенческие, физиологические и гене-
тические. Как было выяснено в течение 1970-х годов, наследствен-
ный материал не только случайно мутирует, но и закономерно
преобразуется, что собственно наследственные реакции — лишь
более глубокий уровень целостной реакции организмов. Этот уро-
вень обнаруживается только в последовательности поколений.
В силу скрытости и сложности своих связей с физиологическим, он
часто воспринимается как совокупность «случайных мутаций».
Появилось несколько моделей, толковавших этот круг явлений
с единой точки зрения, например, «молекулярная машина»
Вайнцвайга—Либермана (о ней см.: [Чайковский, 1976]). Была
предложена и игровая модель [Чайковский, 1972; 1977а], на кото-
рой легко понять, насколько для эволюции важен феномен смены
поведения.
Еще Дарвин обращал внимание на то, что в ходе приспособле-
ния орган часто меняет свою функцию, а в 1876 г. зоолог Антон
Дорн сформулировал это положение в виде принципа смены
функций. (Пример: богатый набор наружных желез млекопитаю-
щих — молочные, сальные, потовые, пахучие и др.— берет начало
от слизистых желез их предков.) Принцип легко позволял понять
простые случаи эволюции: рука, например, смогла развиться пото-
му, что у предков передняя конечность служила для более простых
функций — плавания, ползания и т. д.,— но требовал важной ого-
ворки: смена функции невозможна без смены поведения (а часто
требуется и смена физиологии), причем организм предполагается
уже способным к пользованию еще только возникающей функ-
цией. В этом допущении и состоит по существу принцип смены
функционирования.
Принцип проявляется по крайней мере в шести формах: этоло-
гической (смена йоведения), экологической (уход в более приемле-
мую нишу, или эврибионтность), физиологической, онтогенетиче-
ской (изменение зародыша и незрелых особей), генетической
(генетический поиск) и эволюционной (смена функций). Эти 6
форм образуют множественный параллелизм, следовательно и сама
смена функционирования имеет общесистемную природу. В тер-
минах главы 2 это можно выразить так: любая смена функциони-
рования состоит в движении вдоль какого-то рефрена архетипиче-
ской таблицы, которая для данного таксона едина, с какой бы из
6 точек зрения его ни рассматривать.
Рассмотрим важный частный случай — параллелизм между
модификационной (возникающей в ходе онтогенеза) и наследст-
венной изменчивостью, обсуждаемый уже 90 лет (литературу см.
[Юдин, 1976; Хесин, 1984, с. 219; Экология..., 1984, с. 196]).
Так, растение может сменить стоячую форму на плавающую
(и наоборот) в зависимости от влажности в момент прорастания,
105
а может не иметь этой способности, т. е. жестко наследовать
форму. Генетик Рихард Гольдшмидт в 1930-е годы пришел к выво-
ду, что любая наследственная аномалия может быть наблюдаема
и в качестве модификационной. Смена второй на первую (частный
случай генетической стабилизации — см. 4.1) обычно объясняется
с помощью некой гипотетической формы (совпадающей, гармони-
ческой, косвенной, стабилизирующей и т. п.) отбора, но эпоха
подобных объяснений 2 уходит, да есть и веские данные в пользу
того, что такая замена просто мешает действию отбора. Во-первых,
давно замечено чисто логическое противоречие: отбор в принципе
не может отделить наследственную адаптацию от ненаследствен-
ной, поэтому способность организмов к модификации лишь затруд-
няет выявление параллельных ей мутаций [Чайковский, 1976,
с. 151]. Во-вторых, эта помеха действительно оказалась решающей
в машинном эксперименте [Чайковский, 1972]. И, в-третьих,
преимущественное выживание именно обладателей модификаций
по сравнению с обладателями параллельных им мутаций (геноко-
пий) еще в 1939—1941 гг. прямо показано в работах группы
Г. Ф. Гаузе: в случае приспособления инфузорий к естественному
фактору среды это преимущество сказывалось сразу, а в случае
искусственного яда (хинина) — с задержкой порядка 100 поколе-
ний [Экология..., 1984]. Последнее, заметим, ясно указывает на
длительный генетический поиск. Позже аналогичные результаты
получены группой Б. П. Ушакова на лягушках: лучшая генети-
ческая приспособленность к высокой температуре среды означает
меньшую способность приспособляться физиологически и, как
результат, худшую выживаемость [Ушаков, 1982].
Но если основу адаптивного процесса составляют прижизнен-
ные приспособления, а эволюция состоит в наследственных изме-
нениях, то должен существовать механизм наследования, т. е.
механизм, превращающий первые во вторые. Здесь прежде всего
надо, вслед за М. А. Шишкиным, отметить, что «деление феноти-
пических признаков на наследственные и ненаследственные»
ошибочно подменяет собою «единственно оправданное их деле-
ние — по степени устойчивости их выражения» [Экология..., 1984,
с. 199]. Другими словами, всякий признак возникает прижизнен-
но — как результат изменения пути онтогенеза, а наследование
означает лишь фиксацию этого пути в качестве единственного,
т. е. отсечение всех альтернативных вариантов. Наглядный пример
изменения онтогенеза привел И. А. Аршавский [1982, с. 195]:
обычно кролики отличаются от зайцев малой подвижностью, но
если кролик взрослеет в условиях регулярных физических нагру-
2 Для них характерна подача гипотезы как установленной истины, при полном
отсутствии ссылок на фактические исследования предлагаемой формы отбора.
Характерна работа Ю. М. Оленова [1946], задавшая тон последующим. Автор
прямо заявил, что его цель — «рациональное объяснение с позиций дарвинизма»,
в качестве которого привел цитату из Дарвина о том, что привычка и упражне-
ние «подкреплялись» отбором. Наоборот, Г. Ф. Гаузе еще в 1941 г. отметил, что
стабилизирующий отбор — постулат [Экология..., 1984, с. 51].
106
зок, то его физические характеристики и внешняя форма прибли-
жаются к заячьим — настолько, что он теряет способность спа-
риваться с единоплеменниками. Фиксации (наследования) «зая-
чьих» черт здесь не было (для фиксации, как мы увидим ниже,
нужны более жесткие условия), но опыты и не ставили этой цели.
Их цель была — показать, что межродовые различия достижимы
путем изменения физиологического режима позднего онтогенеза.
Фиксацию часто называют эффектом Болдуина — по имени
биолога, описавшего ее (1896 г.), в качестве фактора эволюции.
И. И. Шмальгаузен [1968, с. 410] полагал ее побочным эффектом
стабилизации, т. е. всегда выгодной. Наоборот, Любищев [1982,
с. 171—172] рассматривал ее как рутинизацию (см. 2.5), подчер-
кивая этим ее невыгодность и даже гибельность. Очевидно, что
само понятие фиксации нуждается в более аккуратном анализе.
Когда фиксация идет в течение ряда поколений, должны
существовать какие-то промежуточные ее стадии. Они давно
известны как длительные модификации (ДМ) — так именуют выз-
ванные внешними условиями модификации, сохраняющиеся
в нескольких поколениях. Например, карликовая высокогорная
форма дерева после пересадки на равнину обретает нормальный
рост примерно в пятом поколении. ДМ типичны при адаптации
одноклеточных к экстремальным условиям [Женермон,1970].
Особенно интересны ДМ, адаптирующие организмы к условиям,
в прежней их эволюции не встречавшимся (например, к синте-
тическим ядам). Неоднократно ДМ предлагались в качестве гипо-
тетической стадии эволюции отдельных признаков (например
[Martin, 1956]), но подобные гипотезы не имели успеха, поскольку
ДМ почти не были известны у многоклеточных. В настоящее
время можно уверенно сказать, что ДМ имеют всеобщее распрост-
ранение, но у многоклеточных выражаются не столько во внешних
признаках организмов, сколько в способах работы генетических
систем. Первые теоретические модели такого рода появились в
1970-е годы.
Сначала модели ДМ были эпигенетическими, т. е. описывали
процессы, не изменяющие текстов ДНК,— таков, например, двух-
оперонный триггер (4.1). С 1960-х годов Л. Б. Меклер разрабатыва-
ет эпигенетические схемы наследования, основанные как на общих
законах метаболизма, так и на особенностях высших животных
(вынос в кровоток соответствующих генетических фрагментов и их
встраивание в геном клеток, формирующих зародыш) [Меклер,
1980а, с. 337 — 338]. Некоторые авторы традиционно приписывали
ДМ, как и всякое наследуемое приспособление, отбору [Юдин,
1976, с. 25], но такой «отбор» (как справедливо отмечал протисто-
лог Жан Женермон [1970]) может идти вообще без смертности
адаптирующихся особей. Если добавить к этому, что ДМ легче
всего наблюдаются при бесполом размножении и захватывают сра-
зу (в отличие от мутаций) значительный процент исследуемых
особей, то станет очевидной аналогия между ДМ и поведением
генетических автоматов, описанных в 4.1—4.3. В самом деле,
107
реплицирующая нестабильность является ДМ по определению, но
она сама по себе не изменяет фенотип, а обеспечивает перестройки
генов, которые его меняют. При этом сама нестабильность насле-
дуется ненадолго, но перестроенные с ее помощью признаки
наследуются стойко.
Давно известно, что малые дозы радиации способствуют акти-
вации биологических процессов (вспомним хотя бы радоновые ван-
ны), ускоряют рост и увеличивают урожайность [Кузин, 1977].
Механизм этого удивительного явления состоит, прежде всего,
в производстве облученными клетками химических ядов (радио-
токсинов), которые в малых дозах полезны, причем здесь тоже
говорят о триггерах, точнее, о «триггер-эффекторах» [там же,
с. 81, 90]. Химический мутагенез хорошо известен и сам по себе
(вне связи с облучением), причем стимулирующий эффект малых
доз часто сказывается в течение нескольких поколений после об-
работки [Химический мутагенез..., 1987, с. 239, 247]. Этот круг
примеров методически важен и потому, что впервые показал, как
физиологическое изменение может через несколько поколений
(т. е. путем ДМ) закрепляться генетически. Сейчас о такой возмож-
ности говорят уже многие [Молекулярные механизмы..., 1985,
с. 158]. Хорошо известен стимулирующий эффект и обычных
физиологических воздействий (тепло, холод, растворы), применен-
ных в малых дозах. Можно даже сказать, что лекарство — это яд
или мутаген, примененный в стимулирующих дозах.
Известны и прямые наблюдения превращения ДМ в стойкий
наследственный признак. Так, мутантная дрозофила, страдающая
нехваткой 75% рибосомных генов, увеличивает их число модифи-
кационным путем, причем эта модификация сперва наследуется
нестойко, но через 5 — 7 поколений становится обычным наслед-
ственным признаком, кодируемым в хромосоме,— если только
организмы в течение этих поколений находились в состоянии
стресса [там же, с. 159].
О стрессе следует сказать особо. В норме вся физиология ор-
ганизма направлена на сохранение внутренней среды и, в частнос-
ти, на безошибочное наследование родительстких свойств. Гене-
тическая система изолирована от внешних влияний (в том числе
и от влияний самого организма) сложной системой физиологиче-
ских барьеров. Когда же внешние условия нельзя считать нормаль-
ными, ситуация может меняться в разные стороны. В 1936 г.
Ганс Селье предложил концепцию стресса — наиболее общей со-
вокупности реакций организма на ненормальное воздействие (на
стрессор). Он прилагал концепцию к высшим животным и челове-
ку, видя в стрессе три стадии — тревогу, резистентность и истоще-
ние, из которых важнейшей является вторая, в сущности и обес-
печивающая адаптацию. (Позже она описана и у отдельных кле-
ток — литературу см. [Чайковский, 1976; Браун, Моженок, 1987].
О стрессе и иммунитете см. [Здродовский, Гурвич, 1972].) Если
стрессор достаточно силен, то вместо тревоги приходится рас-
сматривать два процесса — шок и послешоковый поиск [там же].
108
Аналогичное явление также можно видеть на отдельных клетках —
оно описано как биохимический шок [Bacq, Goutier, 1968]. Если
же происходит адаптация к слабому или умеренному воздействию,
то она протекает более мягко. Обычно она начинается со стадии
ориентировки [Гаркави и др., 1979, с. 101]. В случае слабых
раздражителей адаптация ограничивается реакцией тренировки,
а при средних — реакцией активации. Для стадии ориентировки
характерна пассивная резистентность, а для реакций тренировки
и активации — резистентность активная [там же, с. 103—105].
Различие их — в активности разных физиологических механизмов
и в том, что для тренировки характерно превышение уровня
анаболизма (процессов синтеза) над катаболизмом (процессами
распада), а при реакции активации они уравновешены; реакция
активации обычна в молодом здоровом организме [там же, с. 64].
Только слишком сильный или слишком частый раздражитель
вызывает собственно стресс; Селье в конце жизни тоже пришел
к различию между целительным эустрессом и патологическим
дистрессом [Селье, 1979].
Выявление мягких форм адаптации открыло блестящие перс-
пективы для взаимодействия физиологии и медицины с эволюци-
онной теорией. Так, классик возрастной физиологии И. А. Ар-
шавский разработал эффективный метод закаливать детей с
помощью небольших доз резкого охлаждения, основанный на па-
раллели между индивидуальным и историческим развитием
человека [Чайковский, 1988г].
Если мягкие способы адаптации неэффективны, то начинается
собственно стресс, характеризуемый снижением внешней активно-
сти, прорывом физиологических барьеров и, как следствие, стойки-
ми генетическими перестройками. Этот феномен хорошо известен
как «экспериментальная эволюция» [Агаев, 1978; Wu, 1978]:
под влиянием резко ненормальных условий жизни за несколько
поколений может образовываться нечто вроде нового вида у насе-
комых (опыты Г. X. Шапошникова), злаков (опыты М. Г. Агаева)
и бактерий (см. 4.1). Налицо общая закономерность: если физио-
логическая система не смогла адаптироваться одним способом, то
включается другой, более глубокий и, следовательно, теснее свя-
занный с генетической системой. «Экспериментальная эволюция»
хорошо описывается в терминах генетического поиска, тогда как
отбор ненаправленных вариаций заведомо бессилен на столь малых
выборках.
Разным типам внешней смены функционирования грубо соот-
ветствуют разные ведущие формы внутреннего поиска: смене
поведения — физиологическая норма и ориентировка, уходу
в другую нишу — ориентировка и следующая за нею тренировка,
смена физиологических (включая иммунные) режимов принимает
формы тренировки и активации, нестойкие изменения в онтогенезе
(модификации) тоже означают активацию, а стойкие генетические
перестройки характерны для стресса — прежде всего для стадий
шока и истощения. Их длительное продолжение вызывает массо-
109
вую катастрофическую гибель и необратимый эволюционный
эффект у немногих выживших 3. Если не в целом, то по отдель-
ности элементы этой схемы приспособления уже хорошо иссле-
дованы. В частности, генетик М. Д. Голубовский аргументированно
пишет о трех формах наследственной изменчивости — эпигенети-
ческой (без изменения текстов ДНК), вариационной (модифика-
ции, основанные на изменении нехромосомных ДНК) и мутацион-
ной [Молекулярные механизмы..., 1985, с. 155]. По-видимому,
следует в качестве четвертой формы выделить перестройки целых
блоков генетической памяти — «транспозиционные взрывы» [там
же, с. 13, 143], трансформацию [там же, с. 84], гипермутации
[там же, с. 163] и многие другие. Все четыре формы вместе ха-
рактеризуют диапазон путей генетического поиска, т. е. генетиче-
ской формы смены функционирования.
До сих пор наследственность и разнообразие в основном изу-
чались порознь, но, как показывает генетик Л. И. Корочкин
[там же, с. 132], в организации генома тоже есть архетип (по
Оуэну) и параллелизмы. Остается добавить, во-первых, что смена
функционирования — это, как правило, движение вдоль рефренов,
т. е. развертывание потенций архетипа по Мейену, а наследова-
ние — фиксация одной или немногих позиций этого архетипа, и,
во-вторых, что одна структура может выполнять разные функции,
и наоборот. Поэтому Мейен [1984, с. 25] видел в смене функциони-
рования временной аспект мероно-таксономического несоответст-
вия. Отсюда (а вовсе не из хаотичности механизмов изменчивости)
вытекает известный феномен — разнообразие изменений организ-
мов воспринимается как случайное.
4.5. Что такое приспособленность?
Под приспособленностью организма понимают соответствие его
свойств требованиям жизни. Неживые объекты также проявляют
соответствие своих свойств свойствам среды (именно поэтому Лейб-
ниц говорил о мировой гармонии), но биологи издавна стремились
сформулировать сущность специфики живого. Отправным пунктом
им служило учение о целесообразности. Бытует мнение, что проб-
лема биологической целесообразности решена Дарвином, но это —
досадное смешение понятий: у Дарвина вообще нет речи о том, по-
чему организм растет, действует и размножается. Эти вопросы
3 Принятию этого вывода, уже почти очевидного, во многом мешает устаревшее ста-
тистическое мировоззрение (см. 1.2). Так, палеонтолог А. П. Расницын недоуме-
вает: «Если даже в нормальных условиях в потомстве каждой самки из всех ее
дочерей в среднем только одной удается оставить свое потомство... если шансы на
оставление потомства для каждой особи ничтожны в норме, то неясно, какие же
условия могут быть катастрофическими?» [Эволюция..., 1987, с. 57]. Дело здесь в
незаконном употреблении выделенных мною терминов: в 3.4 мы видели, что сле-
дует говорить не просто о среднем, но о среднем по выжившим; в норме для
каждого выжившего шансы оставить потомство велики, что диктуется его полно-
ценной физиологией, и он оставляет много потомков, в основном полноценных.
Катастрофа же — это состояние, при котором в популяции таких особей (почти)
нет, когда родителей и их немногочисленное потомство поражает дистресс.
110
обычно не затрагивались и у других эволюционистов, внимание же
уделялось чаще всего лишь причинам наследственных изменений.
Естественно, что эти причины мыслились как внешние организму,
а самая жизнь — как цепь реакций на них. Так, Ламарк утверждал,
что растения и низшие животные «живут только раздражениями,
получаемыми извне» [Ламарк, 1935, с. 13], а позже постулировал
(без дальнейшего анализа) такие основные свойства организмов,
как развитие, потребности и способности [Ламарк, 1959, с. 422].
Дарвин еще более сузил круг рассмотрения, ограничившись фено-
меном унаследования (inheritance) 4 изменений и полностью обхо-
дя вопрос о наследственности (heredity) как таковой. (Этот термин
стал основным после рождения генетики.) Он еще больше вынес
целесообразность за рамки анализа организмов, поместив во внеш-
нюю среду свой главный двигатель — отбор.
Вопросов собственной (внутренней) целесообразности организ-
мов Дарвин касался лишь вскользь (например: «Способность
к восстановлению частей... не более удивительна, чем рост и разви-
тие» [Дарвин, 1951, с. 676]). Для него эволюция была не становле-
нием организации, а лишь процессом накопления различий, повы-
шающих выживаемость, и в 1864 г. Герберт Спенсер дал этому
узкому (внешнему) пониманию целесообразности натурфилософ-
ское выражение в форме принципа «переживания приспособлен-
нейшего». Критики не раз указывали на тавтологичность этой
формулировки: «приспособленнейший» определяется лишь тем,
что переживает прочих. Естественно, что и само понятие целесооб-
разности живого при внешнем его толковании крайне упрощалось,
например: «Целесообразно все то, что увеличивает долговечность,
нецелесообразно все, что ее сокращает» [Оствальд, 1903, с. 241].
Не многим лучше нынешнее понимание приспособленности как
устойчивости индивидуального развития [Эволюция..., 1987, с. 77],
при котором эволюция как развитие тоже не рассматривается,
а ее цели сводятся к отслеживанию нормы, заданной от среды.
Иной была традиция Иммануила Канта. В «Критике способно-
сти суждения» (1790) он отметил, что ссылка на «причину,
действующую согласно целям», не дает ничего, кроме тавтологии,
но все же призвал «не упускать из виду в продуктах природы
принцип целей, т. к. этот принцип хотя и не делает способ возник-
новения более понятным для нас, все же [он] хороший эвристи-
ческий принцип для исследования частных законов природы»
[Кант, 1966, с. 440—441]. Полагая, что с помощью понятия цели
можно описывать природу, а с помощью механического 5 принци-
па — объяснять ее, Кант предлагал объединить оба подхода в одном
принципе. Ведь механический способ в сущности ограничен: «Как
бы мы ни преуспели в нем, он всегда будет недостаточен для
вещей, которые мы признаем целями природы», поэтому следует
4 В русских переводах это слово передают то как «наследование», то как «наследст-
венность», то как «наследуемость», смотря по контексту.
5 В наше время говорят не о механическом, а о физико-химическом способе объяс-
нения природы, но суть подхода от этого не изменилась.
111
«все продукты и события природы, даже самые целесообразные,
объяснять механически настолько, насколько это в нашей воз-
можности», но «те продукты, которые мы можем поставить для
изыскания только под понятие... разума, мы в конце концов
должны, несмотря на эти механические причины, подчинять целе-
вой каузальности» [там же, с. 446]. Берг присоединялся к мнению,
что этот двойственный принцип — «мысль самая глубокая, какую
можно найти у Канта» [Берг, 1977, с. 70]. Ныне этому «двойст-
венному принципу» Канта вполне можно придать иной познава-
тельный статус: причинно-целевое описание образует взаимодо-
полнительную пару по Бору, так же, как мероно-таксономическое
(2.2) и номотетико-идиографическое (3.2) описания.
Кант, дав не менее 16 пониманий целесообразности, пользовал-
ся единым ее обозначением — Zweckmassigkeit. Однако кроме
основного термина Zweck (цель), ему понадобились в роли сино-
нимов Absicht (намерение), Mittel (средство), Zweckahnlich (не-
что, подобное цели), scopus (мишень) и др. Этот разнобой пробовал
обойти Бэр, разделив понятия сознательной цели (Zweck) и окон-
чательного состояния (Ziel), завершающего направленный про-
цесс. Согласно Бэру, стрела летит в Ziel, ничего не зная про
Zweck стрельца. (Анализ понятий см. у Любищева [1982] и
Сутта [1977]). Отрицая Zweck за организмами, Бэр все-таки
признавал ее за природой как целым, поскольку без этого отдель-
ные цели (Ziele) развития были бы друг для друга случайны,
т. е. нецелесообразны (мы говорили об этом в 3.1). Тем самым
понятие цели как соответствия Бэром не разъяснено, хотя словар-
ное значение слова Zweck как раз это прежде всего и означает.
Поэтому полезно уточнить классификацию Бэра. Воспользовав-
шись двухсотлетним опытом анализа проблемы — в основном
немецкими авторами, будем понимать целесообразность организ-
мов в следующих смыслах:
(1) прочность, добротность, долговечность (Dauerhaftigkeit) —
против такого понимания возражал, кстати, Бэр [Берг, 1977,
с. 133];
(2) соразмерность и соответствие цели, пригодность, удобность
(Zweckmassigkeit), что близко к technica speciosa Канта [1966,
с. 417] и его Zutraglichkeit [Kant, 1790, с. 392];
(3) целестремительность, устремленность (Zielstrebigkeit), что
близко к technica naturalis Канта [1966, с. 417];
(4) преднамеренность, предположенность (Absichtlichkeit), что
близко к technica intentionalis [там же] и absichtiliche Zweckmas-
sigkeit [Kant, 1790, с. 318];
(о) самосознание желаемой и известной цели (Zielbewusstheit).
Это последнее скорее относится к человеческой деятельности
и потому далее не рассматривается, но именно ему принято
противопоставлять (4) как специфику биологии.
Типы (1) —(3) возможны в живой и неживой природе: вы-
живают виды достаточной экологической прочности и достаточно
стабильные изотопы; гриб и водоросль соответствуют друг другу
112
в лишайнике, а два электрона с разными спинами — на одном
уровне в атоме; зародыш стремится к состоянию взрослого орга-
низма, а пересыщенный раствор — к состоянию кристалла. На-
оборот, тип (4) специфичен для живого: так, морские организмы,
эволюционируя в наземных, занимали новые зоны обитания,
дотоле им неизвестные, причем реализовывались их потенции,
заложенные задолго до этого,— например, многолучевые конеч-
ности позвоночных (2.5). Знание конечной цели здесь вовсе не
требуется, о чем пойдет речь в части II. Наоборот, при сознательном
человеческом планировании (например, в планировании работы
завода) лежит в основе знание цели — в этом специфика, выражае-
мая типом (5).
Живое адаптируется преднамеренно в том смысле, что любой
дискомфорт понуждает к поиску приемлемых условий жизни,
даже если о них ничего не известно. Это — так называемый
стохастический поиск, который не следует путать со случайным
блужданием (даже контролируемым смертностью). Поиск реали-
зует ту внутреннюю целесообразность, на которую указывали до
Дарвина многие философы. В частности, Гегель писал в «Филосо-
фии природы» (1817): «Основательное определение, которое дал
живому Аристотель, учивший, что его следует понимать как
действующее согласно его (собственным.— Ю. Ч.) целям, было
почти утрачено в новое время, пока Кант не восстановил это
понятие в виде принципа внутренней целесообразности, согласно
которой живое должно быть понимаемо как самоцель... Инстинкт
есть целевая деятельность, действующая бессознательно» [Гегель,
1975, с. 507]. При этом инстинкт он понимал широко, как один
из основных способов жизнедеятельности, равный по значению
«стремлению к формообразованию» [с. 499]. Так понятому
инстинкту примерно соответствует «гедонический подбор» Богда-
нова [1913, с. 75]. С той же целью Ганс Дриш [1915, с. 182] и
Л. С. Берг [1977, с. 107] ввели понятие изначальной целесообраз-
ности живого, имея в виду не какую-то мистическую догму (как
часто их толкуют), а общеизвестный феномен, основной для физио-
логии. (Его частным случаем является сформулированный в 4.4
принцип смены функционирования.) Впоследствии об этом фунда-
ментальном свойстве живого писали многие — упомянем «телеоно-
мию» К. Питтендрая, «опережающее отражение действительнос-
ти» П. К. Анохина, «потенциальную направленность» Т. Сутта
(обзор см.: [Сутт, 1977, с. 100—103]), однако суть целесообраз-
ности типа (4) прояснилась лишь с пересмотром смысла категории
времени, о чем мы будем говорить в 9.4. Пока же подчеркнем
только, что многочисленные наблюдения целесообразных ответов
организма на ситуации, в его эволюционном прошлом не встречав-
шиеся, вполне достаточны для признания типа (4) реальностью.
Именно тип (4) отражает специфику биологической приспо-
собленности и, как принято считать, является следствием эволю-
ции. Однако «как раз наиболее важные для жизни... физиологиче-
ские процессы эволюционируют очень мало, они отличаются пора-
113
зительным консерватизмом, почти такой же однотипностью, как
и основной состав живых существ», тогда как компоненты живого,
«которые, возможно, никакого полезного значения для организма
не имеют... необычно разнообразны» [Рыжков, 1974, с. 95]. Этот
хорошо известный факт долго оставался в тени, так как противоре-
чил обычному взгляду на разнообразие как на совокупность
частных приспособлений (которые предстоит выявить) к локаль-
ным условиям. Здесь снова уместно сравнение с языком: если он
как целое безусловно приспособлен для передачи мыслей, то искать
какую-то полезность в каждом свойстве каждого слова нет смысла.
Замена старого короткого слова довлеть на новое громоздкое
удовлетворять нельзя назвать приспособлением к чему бы то ни
было, но, оставшись без употребления, слово довлеть было приспо-
соблено — к новой функции (получило значение «подавлять, го-
сподствовать» по созвучию со словом давление). «Концепция
эволюции как процесса, направляемого все большим и большим
приспособлением, сталкивается с целым рядом противоречий,
которые, в сущности, она может преодолеть только словесно»,
путем «хитроумия» толкователей [там же, с. 95, 97]. Мысль эта
(как и все аргументы в данной дискуссии) очень стара: еще Бэр,
иронизируя, вспоминал анатома XVII в., который объяснял нали-
чие ягодиц у человека тем, что таковые дают ему возможность
сидя размышлять о величии божием. Н. Я. Данилевский [1885,
ч. 1, с. 15] привел это как не нуждающийся в комментарии
курьез, но и через сто лет после иронии Бэра было выдвинуто
почти того же качества объяснение: ягодицы, оказывается, закреп-
лены отбором потому, что их обладатели могли, усевшись удобно
у костра, выточить лучший каменный топор, чем те, кто сидел на
корточках, и потому оставить большее потомство. (Морфологиче-
ски, в традиции Везалия —Монро—Бэра, развитие седалищных
и икроножных мышц объясняется вертикальной ориентацией
тела.)
Конечно, это курьез, но намного ли он хуже привычных нам
с детства объяснений? Воробей сер — это покровительственная
окраска, снегирь ярок — он привлекает самку, синицы и многие
другие (где ярки оба пола) опознают свой вид по расцветке,
а изобретательные турухтаны (у этих куликов голова каждого
самца раскрашена на свой лад) преодолевают, оказывается,
безразличие пресыщенных самок (анализ см.: [Любищев, 1973,
с. 40]). Если мы позволим себе перебирать любые доводы, пока
не найдем подходящий к данному случаю, то лишимся всяких
критериев истинности. Как заметил еще Кант, «разум умеет
давать вещам соответствие с его произвольными выдумками»
[1966, с. 393].
Разнообразие не сводится к приспособлению (по Гёбелю, «фор-
мы жизни несоизмеримо разнообразнее, чем условия жизни»
[Рыжков, 1974, с. 96]), и продолжать бесплодную традицию
«естественного богословия» мы не имеем морального права, но
столь же несомненно, что приспособленность существует: орга-
114
низмам полезны не только их общие (мало разнообразные)
свойства, но и многие из их частных особенностей. Для упорядоче-
ния разнообразия оказывается, поэтому, необходимым понять, что
такое сама приспособленность. Исследовав этот вопрос, Мейен
[1984] пришел к неутешительному выводу: вопрос всерьез даже не
поставлен, оппоненты противопоставляют друг другу только свою
убежденность — за отсутствием «методологического инструмента-
рия». Сказанное до сих пор позволяет предложить 4 таких
«инструмента», хотя общая теория адаптации — дело будущего.
Во-первых, надо четко различать общее приспособление (ту
самую внутреннюю целесообразность, которая делает живое само-
целью) от частного, связанного с разнообразием. Так, хлорофилл,
служащий для поглощения света при фотосинтезе, — общее
приспособление, а зеленый пигмент (иногда — тот же хлорофилл),
делающий гусеницу сходной по цвету с листом, — частное. Идиоге-
нез, рассматриваемый в данной главе, производит лишь частные
приспособления (об общих см. часть II). Хотя четкой границы
между общими и частными приспособлениями нет, но это, соглас-
но 2.3, не должно мешать четкому разграничению соответствующих
понятий.
Во-вторых, выясняя адаптивность данного признака, надо объя-
снять не отдельный этот факт, а соответствующий ряд фактов
(1.2). «Рассматривать природу теоретически следует на основании
учета большого числа явлений, ибо согласное с природой прояв-
ляется здесь или всегда, или по большей части» [Аристотель,
1937, с. 96]. До сих пор, к сожалению, распространен противо-
положный тезис, сформулированный зоологом Хью Коттом: «Каж-
дый случай нужно рассматривать в его специфике» [Котт, 1950,
с. 180]. Если поступать так, то не удается заметить, что отдельные
объяснения противоречат друг другу и потому ничего не дают
в целом. Почему одни виды обладают данным свойством, а другие,
экологически и морфлогически сходные — нет? Почему один сер,
другой зелен, третий ярок, а четвертый наделен сложнейшим
узором? Почему то же наблюдается, пусть и очень редко, у видов,
живущих в темноте? До тех пор, пока теоретик видел свою задачу
лишь в идеологически приемлемом объяснении (см. сноску 2 в 4.4),
тезис Котта был не хуже других, но сейчас, когда нужно действие
с целью сохранения разнообразия, он неприемлем.
В-третьих, «не следует смешивать механизм с функцией»,
т. е. средство с целью [Котт, 1950, с. 174]. Как мы видели, этот
призыв восходит к Канту. В частности, следует четко различать
процессы приспособления и отбора (см. 4.1). Пусть неселективное
приспособление звучит парадоксально, но оно идет всюду, где есть
генетический поиск, а последний запускается не избирательной
смертностью, а общим дискомфортом. Разумеется, покровитель-
ственная окраска полезна в том смысле, что маскирует от врагов,
но это вовсе не значит, что она могла появиться за счет накопления
случайных изменений: еще энтомолог Сэмюэл Скаддер [Scudder,
1870] предлагал выяснить в опыте зависимость судьбы насекомого
115
от его расцветки (он удивлялся: почему гусеница гротескной
формы и угрожающей окраски буквально кишит пожирающими
ее паразитами, тогда как близкий вид гусениц для них несъедо-
бен? Почему отбор растрачивал силы на такие детали, как рисунок
гусеницы или взрослой бабочки, а не на борьбу с паразитами,
от которых гибнет 99,9% гусениц?). Когда опыт был наконец-то
(через 76 лет) поставлен, выяснилось, что даже «крайний случай
наиболее утонченной маскировки» гусениц почти ничего им в це-
лом не дает: птицы тратили от 7 до 40 минут на поиск первой
гусеницы, но «после обнаружения первой гусеницы остальные
почти всегда отыскивались очень быстро», за секунды [Тинберген,
1970, с. 146, 149]. Если учесть, что сам рисунок гусеницы не может
сформироваться путем отбора быстрее, чем за десятки миллионов
лет (а за это время много раз изменятся и фон и вся экологическая
обстановка), то ясно, что механизм формирования приспособи-
тельной окраски еще предстоит найти, а ссылка на отбор случай-
ностей — наивный самообман.
В-четвертых, факты надо уметь не только получать, но и ви-
деть. Достаточно сказать, что приведенная выше демонстрация
селективной ничтожности покровительственной окраски была
воспринята как доказательство силы отбора: «Нас поразило, на-
сколько важной для выживания оказалась эта полная гармония
между насекомым и растением» [там же, с. 150]. Ситуация
вполне обычная: опыт ставился, чтобы подтвердить заданную схе-
му, и потому исход мыслился только как подтверждение. Если
у методологов уже давно признано, что факт не имеет значения
вне объясняющей схемы, то у многих биологов еще сильна пози-
тивистская уверенность, что науку движут именно факты. И вот
мимикрия, которую Любищев (ведущий практик-энтомолог)
аттестовал как наивный маскарад, который хищники легко разга-
дывают, экспериментаторам предстала гармонией выживания.
Таких примеров — когда блестяще задуманный опыт погублен
априорной уверенностью в справедливости того самого, что вроде
бы проверяется,— можно привести множество.
Попробуем использовать все сказанное выше для ответа на воп-
рос «что такое приспособленность?» на том же примере мимикрии.
Прежде всего, выстроив множество покровительственных окрасок
в ряды, легко убедиться, что они распадаются на два множества —
собственно мимикрию (подражание фону) и миметизм (подража-
ние экологически близких видов друг другу). Энтомолог Эндрю
Мёррей в 1859 г. заявил, что эти два феномена имеют различную
природу [Murray, 1862], почти то же объявил в 1935 г. Вавилов,
когда охарактеризовал миметизм (этого термина у него еще не бы-
ло) как «общее явление повторности форм, характерное для
всего органического мира», а собственно мимикрию — как «не
имеющую никакого отношения к... параллельной изменчивости»
[Вавилов, 1987, с. 94]. В последнем он был неправ: виды насеко-
мых отряда привиденьевых (палочников) независимо мимикриру-
ют под самые разные фоны, и то же мы видим у клопов-хищнецов
116
(отряд полужесткокрылых) — налицо параллелизм. В других от-
рядах мимикрия более редка, что, как мы знаем, обычно для
рефренной структуры разнообразий. Однако само различение
мимикрии и миметизма очень важно, поскольку механизм второго
достаточно ясен.
Известно, что культурное бобовое растение чечевица часто
засоряется сорняком викой, и с этим трудно бороться, так как
семена вики имитируют семена чечевицы. Считалось, что это
сходство — результат бессознательного отбора со стороны челове-
ка, но вот было исследовано 14 сортов чечевицы и для всех обнару-
жены соответствующие сорта вики с похожими семенами; из них
только 2 сорта засоряли посевы сходной чечевицы, а остальные
12 росли в других, нежели соответствующий сорт чечевицы,
регионах. Отбор мог повышать численность тех сортов вики,
попавшей в посевную культуру, которые обладали сходством с че-
чевицей, но исходный спектр возможностей был задан без всякой
связи с отбором. (Этот результат опубликовала в 1920 г. Е. И. Ба-
рулина, и на него не раз ссылался Вавилов.) На этом примере
видно, что миметизм сам по себе не связан с приспособлением,
хоть и может быть использован как таковое.
Гораздо сложнее с мимикрией. Факт копирования рисунка
листа на крыльях или надкрыльях насекомого очевиден, равно как
и его маскирующий эффект. Фантастическая гипотеза — что
каждая жилка рисунка возникла случайно и сохранилась потому,
что позволяла оставить больше потомства — была обычна, как и
всякая ссылка на случайность, пока о механизме сказать было
нечего. Каждая жилка отбору подвергаться не могла (никто и не
пробовал защищать этот тезис с цифрами в руках), но почему
надкрылье похоже на лист именно каждой жилкой? Сейчас мы уже
можем вчерне обрисовать черты механизма этого чуда.
Отбору вообще подвергается не признак, а целостная генети-
ческая система, способная жить (см. 4.1), т. е. обладающая при-
емлемым спектром общих приспособлений. Что касается какого-то
частного приспособления (например, расцветки), то она, как пра-
вило, быть отобрана не может — в силу своего слишком малого
влияния на выживаемость, слишком большой сложности и слиш-
ком малого времени, даваемого изменчивой средой на приспособ-
ление. Более того, приспособлением служит, по-видимому, не кон-
кретная расцветка, а физиологическая система, способная произ-
водить тот или другой тип расцветок. Она должна копировать не
отдельные жилки или пятна, а ту целостную картину, которая
создает общее впечатление сходства (например, крыла с листом).
Такие неселективные физиологические приспособления всем хоро-
шо известны.
Что удивительнее — бабочка, получившая рисунок от рожде-
ния, или камбала, умеющая воспроизводить тот фон, на котором
лежит сейчас? Положенная на разноцветные камешки, камбала
постепенно почти исчезает из виду, это всем известно, но почти
никто не видит здесь сходства с бабочкой, «исчезающей» среди
117
сухих листьев. Такова установка науки, господствовавшая до
недавнего времени и родившаяся в начале XX в. вместе с генети-
кой: для эволюции важно только то, что наследственно. Однако
недавно зоолог и палеонтолог М. А. Шишкин, анализируя об-
ширную литературу, обнаружил, что никакого различия между
«наследственным» и «ненаследственным» провести вообще нельзя
[Экология..., 1984; Эволюция..., 1987]. Если организм может что-то
реализовать, значит для этого у него есть подходящий механизм;
если же это «что-то» реализуется наследственно, значит генети-
ческая программа запрещает другие варианты — вот и все (4.4).
Никакой «проблемы наследования приобретенных признаков» про-
сто не существует при таком взгляде на факты. То, что обычно
называют мутацией, — не что-то новое, а переключение с одного
старого на другое. Новое же появляется совсем иначе.
Если так, то мимикрия должна создаваться двумя механизма-
ми: первый создает прижизненный рисунок, а второй отсекает
иные, кроме данного рисунка, варианты и этим делает его наслед-
ственным. Проблема, как видим, в том, чтобы понять механизм
маскировки, например, камбалы: каким образом нижняя сторона
тела передает информацию для рисунка на верхней его стороне.
Эта чисто физиологическая проблема выходит за рамки возмож-
ностей автора, но не входит и в круг задач данной книги о раз-
нообразии. Остается напомнить только, что и у насекомых по-
добный механизм имеется. Еще 100 лет назад его начал изучать
энтомолог Эдвард Паултон, который обнаружил, что гусеницы
некоторых бабочек могут менять свою раскраску: одни, сидя на
коре, покрытой лишайниками, становятся через несколько часов
бурыми в серых пятнах, другие, оказавшись на пожарище, по-
крытом. ростками свежей зелени — черными в зеленую полоску.
Это Котт [1950, с. 34] описал, но не счел интересным добавить
кое-какие детали, к примеру — удавалось получить имитацию фо-
на, на котором гусеницы в природе не живут, причем для этого
гусенице даже не требовались глаза — их густо замазывали. Ни
Паултон, ни Котт не придали значения тому, что насекомые
могут (хотя бы иногда) копировать на своем теле рисунок фона.
Запомним это и снова взглянем на разнообразие мимикрий.
Самки разных видов семейства листовидок (листотелок) из
отряда привиденьевых имитируют разные листья, следовательно,
от общего предка могли унаследовать не само сходство, а только
тенденцию — способность к копированию. Разные самки одного ви-
да могут имитировать листья разных расцветок. Уже это свиде-
тельствует против гипотезы длительного приспособления к конк-
ретному фону, но еще важнее то, что различные виды совершают
один и тот же «промах»: копируют своей верхней стороной
нижнюю сторону листа. Бесспорно, это — приспособление, но
воспользоваться им мудрено — надо перевернуться; Котт [1950,
с. 395] писал, что листотелка «иногда висит под ветвью или череш-
ком листа». В остальное время ей остается только полагаться на
неразборчивость птиц, не замечающих, что лист торчит не туда.
118
Может быть, ее враги нападают снизу, из листвы? Но у нее нет
способности оставаться спиной вниз, да и зачем бы это? Если
хищник приблизится настолько, что сможет разглядеть листопо-
добные прожилки, то заведомо увидит торчащие из-под «листа»
голову, лапки и заднюю часть насекомого. Такие приспособления
Любищев [1982] называл гипертелией (т. е. более чем целесооб-
разностью — признак развился дальше, чем было полезно) и объяс-
нять не брался, но в свете данных Паултона они теряют значитель-
ную долю загадочности: если вид листа насекомым скопирован, то
естественно, что копировалось то, что оно «видело» вблизи и снизу,
а не то, что может видеть птица издали и сверху.
Что и говорить, до полного понимания еще далеко; трудно
представить себе, как рисунок листа копируется прижизненно,
поскольку крыло или надкрылье насекогого — безжизненная
сухая пленка, в отличие от кожи камбалы. Однако, как отмечает
Шишкин [Экология..., 1984, с. 208], «у насекомых подсыхание
крыльев или формирование рисунков надкрыльев может длиться
многие часы после выхода из куколки и, очевидно, в какой-то
степени модифицируется средой» (имеются в виду результаты
Н. В. Тимофеева-Ресовского с сотрудниками, 1965 г.). Это
значит, что поведенческий поиск рисунка все-таки идти может.
Если он может много поколений чередоваться с рутинизацией
(о ней было сказано в 2.5 и 4.4), то оба процесса могут носить
характер ДМ. Следовательно, для понимания мимикрии надо экс-
перементировать всерьез (менять кормовое растение, исследовать
куколки и т. п.). Опыты перед их началом надо основательно
продумывать, чтобы сама их постановка не предопределяла ответ:
не раз случалось, что объекту подсказывали единственный желае-
мый ответ: например, когда насекомых прикалывали к разным
фонам и подсчитывали число съеденных птицами. Ясно ведь, что
на контрастном фоне несчастных быстро склюют и что, следова-
тельно, это не столько опыт, сколько полемический прием. Такой
опыт методологи называют «искусственно привязанным к теории»
[Чудинов, 1977, с. 124].
Правильнее, пожалуй, говорить здесь не о теории (ибо таковой
не может быть без полноценного эксперимента, если сама об-
ласть исследования — экспериментальная), а о натурфилософской
схеме, на что особо указывал Любищев. Суть ее он видел в псевдо-
телизме — направлении мысли, «признающем важное значение
проблемы целесообразности, но пытающемся решить возникно-
вение целесообразного простым взаимодействием сил, не заклю-
чающих в себе ничего целеполагающего»; причем «псевдотелизм
имеет наиболее полное выражение в натурфилософии Ч. Дарвина»
[Любищев, 1982, с. 150, 152]. Возражая против псевдотелизма как
самообмана, Любищев предлагал, вслед за Кантом, различать
утилитарную и эстетическую целесообразность. В 1943 г. он писал:
«Приспособление есть частный случай гармонического строения»,
и полагал целесообразность «частным случаем мировой гармонии,
все более теряющей утилитарный характер» [там же, с. 66];
119
а в 1946 г. сделал уточнение, допустив, что понятия «целесообраз-
ное» и «приспособленное» только частично перекрываются
[Любищев, 1978, с. 415]. Тем самым он указал адаптивному прин-
ципу границы. Посмотрим, как обстоит дело в природе.
4.6. Границы приспособленности
Итак, понятие «приспособленность» шире, чем понятие «резуль-
тат естественного отбора», но и в этих широких рамках попытки
объяснить каждый признак его полезностью часто наталкиваются
на непреодолимые трудности. Смысл многих признаков раскрыва-
ется только тогда, когда описаны и упорядочены те разнообразия,
членами которых эти признаки являются, и этот смысл не всегда
оказывается связанным с пользой. Так, почти все виды хищных
окрашены покровительственно, и это можно связать с приспособ-
ленностью, но само наличие немногих ярких видов указывает
этому границы: рыжая лиса ничего не извлекает из своей окраски
(редкие тусклые особи живут не хуже обычной «огнёвки»), она
сохраняется вопреки своей рыжей шерсти, сохраняется благодаря
некоторому запасу экологической прочности лисы как вида. Виль-
ям Бэтсон, теоретик ранней генетики, резюмировал подобные
факты в парадоксальной форме: животные существуют не благода-
ря своим признакам, но вопреки им [Берг, 1977, с. 179]. Ясно, что
речь здесь может идти только о частных признаках, ответственных
за разнообразие как таковое (см. 4.5). Там же отмечалось, что
прочность — простейший случай целесообразности, а сейчас можно
добавить, что и она не связана жестко с отбором.
Чем выше у вида экологическая прочность, тем меньше может
повлиять на него отбор, поскольку вид реагирует на изменение
условий изменением не смертности своих индивидов, а лишь
режима их жизни [Чайковский, 1977а, с. 1486]. Лишь виды, не
имеющие запаса прочности, подвержены избирательной элимина-
ции, но они и не дают материала для эволюции, на что указывал
еще Данилевский [1885, ч. II, гл. 13]. Чем слабее давление обора,
тем богаче разнообразие организмов вообще и обладателей невы-
годных черт, в частности. Для тропических видов это давно сформу-
лировано [Steenis, 1969], но у отдельных видов экологическая
прочность допускает и в умеренной зоне огромные отклонения
от принципа адаптивности. Они хорошо известны, но, пока к ним
не было теоретического подхода, говорить о них не было принято,
а если изредка кто-то их касался, то ему или просто не отвечали,
или напоминали, что вопрос не нов, и только. Однако некоторые
из них выглядят не столь уж неприступно, если взглянуть на них
с позиций разнообразия. Приведем три примера, а затем обсудим
их возможные механизмы.
(1) У самки пятнистной гиены наружные половые органы
неотличимы от мужских, и «все это устройство поражает своей
нецелесообразностью» [Шмальгаузен, 1968, с. 136]. (Заметим, что
изредка в литературе мелькают курьезные доводы в пользу «селек-
120
тивной ценности» этого дефекта строения тела, но они рушатся,
если вспомнить хотя бы то, что полосатая гиена дефектом не обла-
дает.) Приведя несколько примеров такого «переноса половых
признаков с одного пола на другой» (гамогетеротопии), Шмаль-
гаузен признал их полную неадаптивность, поэтому дальнейшая
его попытка объяснить их с позиции стабилизирующего отбора
[там же, с. 138 — 143] оказалась внутренне противоречивой, и обыч-
но исследователи ее просто не упоминают. Однако в принципе
Шмальгаузен скорее прав: позже выяснилось, что гамогетерото-
пия — обычный феномен эволюции, но проявляется не благодаря,
а вопреки отбору и, в частности, была главным актом при образова-
нии цветковых (см. 7.3). Отсутствие внешних различий является
редким для животных значением мерона «половые органы», и оно
возможно только у экологически прочного вида: пятнистая гиена
продолжает существовать несмотря на то, что копуляция и дето-
рождение у нее «чрезвычайно затруднены» [там же, с. 136].
(2) Всем известно, что обыкновенная кукушка — гнездовой
паразит, т. е. откладывает яйца в чужие гнезда. Поскольку она —
одна из самых обычных птиц, ее отличительные черты принято
считать полезными, т. е. результатом приспособления. Но к чему
она приспособилась? По сравнению с обычными птицами ее су-
ществование выглядит резким дискомфортом: кукушонок в чужом
гнезде до предела напрягает силы, выбрасывая хозяйских птен-
цов, отчего нередко сам гибнет; кукушка вынуждена откладывать
в 2 — 3 раза больше яиц, чем другие, имеет неограниченный
пищевой ресурс (волосатые гусеницы, которых избегает большин-
ство птиц), и «тем не менее она никогда не распложается
сильно: кажется, этому препятствует ее буйная, беспокойная
жизнь» [Брем, Россмесслер, 1867, с. 633]. «Постройка гнезда
и воспитание собственного потомства занимают подчас меньше
времени... Однако природа распорядилась так, что другого пути
у кукушки уже нет» [Мальчевский, 1987, с. 252]. Разумеется, это
возможно только в силу большой экологической прочности данного
вида — кукушка неприхотлива к пище и климату, не имеет серь-
езных врагов и селится повсюду. Главное же — в отряде кукуш-
ковых 52 вида (35%) — гнездовые паразиты, тогда как среди
остальных 27 отрядов птиц это редкость: 26 видов, т. е. 0,3%
видов этих отрядов [там же, с. 218, 229]. Налицо тенденция.
Изучавший кукушку всю жизнь А. С. Мальчевский отверг
бездумную ссылку на естественный отбор как «трафаретное
объяснение» и пришел к выводу, что «вся проблема эволюции
гнездового паразитизма еще далека от решения». Все же он решил
один важный частный вопрос — показал, что миметизм кукушечьих
яиц является сам по себе не приспособлением к паразитизму,
а рядом, параллельным ряду расцветок других птиц. Главное
отличие от миметизма семян (какой описала Барулина) состоит
здесь в том, что кукушка сознательно выбирает гнездо с яйцами,
похожими на ее собственные, и эта поведенческая мобильность
резко усиливает эффект миметизма.
121
Ценные наблюдения Мальчевского наводят на мысль о выявле-
нии рефренной структуры гнездового паразитизма в целом. Если
выписать в ряд все формы репродуктивного поведения птиц,
то окажется, что практически все они реализованы в рамках
семейства кукушек — от стойкой пары в аккуратном гнезде до
полного отсутствия и пар, и гнезд. В поведении кукушек нет
ничего, что было бы характерно только для них. Так, одна из
американских кукушек известна коллективным гнездом — то же
известно для многих колониальных птиц [Панов, 1983, с. 249]
и для «гнезд» некоторых сорных кур; другая выводит птенцов
в отнятом у других птиц гнезде — то же делают многие хищные
птицы; третья вьет гнездо хуже вороньего, а иногда и подбрасывает
свои яйца в гнёзда других видов — то же делает одна из воловьих
птиц (отряд воробьиных); у четвертой птенец-паразит буквально
загрызает птенцов хозяев — то же делают птицы медоуказчиков
(отряд дятловых); пятая паразитирует лишь на одном виде (что
экологически абсурдно) — то же делают некоторые вдовушки
(отряд воробьиных). Далее, у многих птиц (совы, сорные куры)
известны утеря инстинкта гнездостроения и непомерно растянутый
период кладки яиц, что однако не связано у них с гнездовым
паразитизмом; именно эти качества легли в основу стабильной
яйценоскости домашних кур. Словом, не продолжая перечня,
можно заключить, что кукушкам свойственно воспроизводить
черты самых разных птиц. Наконец, гнездовой паразитизм хоро-
шо известен у насекомых, будучи (как и у птиц) преимущест-
венной чертой одного отряда — перепончатокрылых, причем его
формы (особенно так называемый социальный паразитизм) прояв-
ляет хороший параллелизм с птичьими. Однако разнообразие
форм здесь гораздо выше, в соответствии с гораздо большим
обилием самих видов насекомых (их более миллиона).
Известны виды наездников (подотряд стебельчатобрюхих), ли-
чинки которых обращают себе на пользу поведение и физиологию
своих «хозяев» (точнее — жертв), например — заставляя их перед
смертью превращаться в прочное жилище для паразита (мумию).
Однако и здесь есть параллель с кукушкой, которой приемные
родители безропотно служат и даже отчасти помогают при уничто-
жении их собственного потомства. Хотя некоторые из видов
воробьиных выбрасывают кукушечьи яйца, но ни один не пресле-
дует кукушат и не мешает им выбрасывать из гнезда хозяйских
птенцов. Очень похож в этом отношении и гнездовой паразитизм,
описанный недавно у рыбы зубатки пятнистой [Sato, 1986].
(3) Наиболее удивительное порабощение хозяина паразитом
описано у одного плоского червя — ланцетовидного сосальщика
[Анохин, 1966] (на эту работу мне любезно указала Э. А. Зелик-
ман). Яйцо сосальщика из экскрементов овцы попадает в легкое
наземного моллюска (слизня) и превращается в личинку. Комочек
таких личинок, «выплюнутый» слизнем, поедается муравьем, и тут
происходит нечто невообразимое. Одна из личинок мигрирует из
кишечника муравья в его подглоточный ганглий (где и гибнет),
122
после чего муравей лезет на былинку, вцепляется в нее челюстями
и там остается. Когда овца съест его вместе с былинкой и перева-
рит, то остальные личинки (из его брюшной полости) мигрируют в
печень овцы, где и развиваются. До раскрытия этого процесса счи-
талось, что овцы поедают зараженных муравьев случайно. Заметим,
кстати, что сама по себе внутриорганизменная миграция давно и
хорошо известна: например, плоские черви могут включать в свои
наружные покровы и успешно использовать водоросли (т. е. эколо-
гически становиться растениями) и стрекательные капсулы съе-
денных ими кишечнополостных [Любищев, 1982, с. 195]. Это уди-
вительно, но не более, чем передача камбалой расцветки своего фо-
на с «брюшной» стороны на «спинную», о чем шла речь в 4.5.
Приведенные примеры, при всей их фрагментарности и неза-
вершенности самих исследований, позволяют сформулировать ог-
раничения принципа адаптивности: далеко не все наблюдаемое
адаптивно, а сами адаптации формируются из блоков, поставля-
емых возможностями архетипа (по Мейену) и потому достаточно
универсальных; приспособления, даже важные для выживания,
не имеют жесткой связи с отбором, тогда как многие качества орга-
низмов, явно приспособительные, не имеют заметной связи и с
выживанием (4.5). Все это вынуждает очертить, хотя бы грубо,
процесс приспособления, дабы указать его место в более общем
процессе идиогенеза.
Сперва отметим самое простое: во многих случаях ведущим
процессом можно полагать рутинизацию. Почему ланцетовидный
сосальщик живет в печени млекопитающего, а его личинка — в ки-
шечнике муравья в легком слизня? Ответ довольно прост: мы не
знаем, живет ли он только там, просто там он живет достаточно
часто, чтобы его обнаружили; если данный путь развития оказался
намного надежнее других, то сама собой пошла рутинизация —
«забывание» генетической системой тех возможностей организма,
которые использовались недостаточно часто — подобное описыва-
лось генетиками не раз.
Это не значит, разумеется, что в эволюции паразита все понят-
но,— наоборот, понять предстоит главное. Как получилось, хотя
бы, что его личинка порабощает муравья, заставляя его ждать на
былинке овцу, которая его съест? Механизм этого явления неизвес-
тен, но, чтобы искать его, надо заранее понять, что он не выработан
исключительно для данного случая, а взят из арсенала средств при-
роды, использующей его довольно часто. Словом, сперва следует
поместить его в соответствующий ряд. Данный прием полезен
и в других случаях. Мы уже видели, как синдром гнездового пара-
зита утратил значительную долю загадочности, когда был разложен
на элементы, обычные у самых различных животных. Важно, что
такой анализ допускают, в силу общесистемных законов, не только
материальные формы, но и социальная организация.
А как быть с примерами, стоящими особняком? Так, Котт, буду-
чи уверен в полезности всякой формы и окраски для селективного
выживания, честно привел несколько совсем непонятных ему при-
123
меров и объединил их рубрикой «Примеры случайного сходства».
Логика была простая: все полезное объединяется самою своею по-
лезностью в единый ряд, что и служит общим объяснением, а непо-
лезное тем самым остается без причины, беспричинное же равно-
сильно случайному. Действительно, его примеры на редкость
разнородны и ряда не образуют, но это не значит, что каждый из
них не принадлежит какому-то своему ряду, Коттом не замечен-
ному. Разве не наводит на размышление хотя бы то, что «портрет
самурая» (на панцире краба) и «герб клана Токугава» (на голове
рыбы) отмечены в японских водах, а рыба, у которой «рисунок
хвоста напоминает древние арабские письмена», причем можно
прочесть заклинание «Нет бога кроме Аллаха»,— в мусульман-
ских? Сам Котт смог написать по этому поводу лишь следующее:
«Если учесть невероятное разнообразие форм, цветов и рисунков
у бесчисленных видов животных, обитающих в различной среде,
то, пожалуй, было бы затруднительнее объяснить отсутствие
случаев такого сходства, чем их наличие» [Котт, 1950, с. 172]. Что
и говорить, на самом хвосте эта мысль выражена проще и яснее.
Как ни относиться к примерам (среди них возможны и анекдоты),
налицо капитуляция перед проблемой разнообразия.
Попробуем быть чуть рациональнее Котта: разве подлинный
герб не может принадлежать к тому же географическому стилю
(см. 2.5), что и орнаменты местных животных? Разве не могли
изобретатели письменности черпать материал для алфавитов из
природных орнаментов? Конечно, было бы абсурдно думать, что
знаменитое заклинание списано с хвоста рыбы (которая и не водит-
ся на родине Магомета), но ведь точное совпадение букв не утвер-
ждается, а совпадение стиля — вещь хоть и не всегда понятная,
но довольно обычная. Словом, ряды вроде географического стиля
могут упорядочивать знания и служить для поиска общих причин
не хуже, чем ряды вроде «приспособительной окраски» или «род-
ственных форм». Все они в какой-то мере необходимы для того,
чтобы перейти от наивного «рассмотрения каждого случая в его
специфике» к анализу разнообразия.
Здесь приходится говорить о стиле в более широком смысле.
Развивая идеи Любищева и Мейена, ботаники М. С. Игнатов
и С. В. Чебанов рассматривают стиль как устойчивый комплекс
периферических меронов данного архетипа. [Системность и эволю-
ция, 1984, с. 87]. В рамках биологии они различают географиче-
ский, исторический и экологический стили, хотя можно говорить о
морфологическом, физиологическом, этологическом и других сти-
лях. Общим здесь всюду явится понимание стиля как некоторой
надорганизменной целостности, не выводимой ни из общего про-
исхождения обладателей данных черт, ни из приспособления их
к общим условиям жизни. Зато возникнув, стиль может выполнять
определенные функции и потому даже служить объектом отбора.
Такова, например, поведенческая изоляция, характерная для
видов-двойников, так как «подбирается партнер сходного стиля»
[там же, с. 88]. Рассмотренные выше примеры миметизма оче-
124
видным образом подпадают под разные формы стиля, но когда
мы обратимся к редким явлениям, например, к переносу признаков
с одного пола на другой у зверей, то говорить о стиле вряд ли
уместно; и тем не менее видно нечто общее: во всех случаях пара-
доксальные варианты не образуют хаоса, но лишь выявляют спектр
возможностей архетипа (по Мейену). Замечательно, что та целост-
ность, которую призвано в последнем случае формализовать
понятие архетипа, оказывается не таксоном, не экосистемой и
даже не ландшафтом, а половым процессом (точнее — принципом
полового воспроизведения). Именно на его периферии наблю-
дается перенос признаков с пола на пол у зверей.
Понятие морфологического стиля тесно смыкается с понятием
общей гомологии по Оуэну (1.4), но не покрывается им. Таков
феномен меторизиса, когда, например, «гомологичные плавники
у крайних форм могут не содержать ни одной гомологической
части: форма сохранилась, содержание полностью сменилось»
[Любищев, 1982, с. 236]. О том же свидетельствуют галлы (обра-
зуемые насекомыми вздутия) хвойных, похожие на их шишки
[Берг, 1977, с. 177 — 179], и т. д.
Могут возразить, что понятие стиля — лишь новый способ обоз-
начать непонятное, но это будет неверно: понятие стиля позволяет
находить ряды там, где иначе их не видят (как не увидел их Котт),
а это — первый шаг в поисках механизма. Более того, если механи-
ческое и целевое описания взаимодополнительны, то уже этот пер-
вый шаг может в будущем вполне удовлетворять биологов, как
сейчас ссылка на стиль удовлетворяет многих гуманитариев.
Когда ряд построен, есть смысл искать единый механизм его
порождения. Так, все случаи гнездового паразитизма являются в
той или другой мере проявлением дисбаланса репродуктивной си-
стемы, и если здесь и можно говорить о приспособлении, то только
в том смысле, что наблюдаемые виды сумёли, отягченные дисбалан-
сом, выжить. Сам дисбаланс выступает как тенденция таксона,
и только к ней (а не к внешней среде) здесь можно искать приспо-
собление. Эта опасная для таксонов тенденция ясно проявляется
и у отряда сов, но, как всякие хищники, они не столь прочны эколо-
гически, чтобы выдержать те ее крайние формы, которые именуют
гнездовым паразитизмом, и можно полагать, что «кукушечьи»
варианты сов попросту вымерли. Вот где речь об отборе есте-
ственна.
Столь же естественно вспомнить, что всякий дисбаланс вызы-
вает дискомфорт, а с ним и различные формы уклонения от физио-
логической нормы, вплоть до дистресса (4. 4). Известно [Мальчев-
ский, 1987, с. 229], что кукушки гнездящихся видов подбрасывают
лишние яйца в гнезда других видов тогда, когда имеют пищу в из-
бытке. Эта скрытая форма инфантицида могла при длительном
избытке пищи рутинизироваться, т. е. привести к гнездовому
паразитизму. Стресс (в котором оказываются самки, регулярно не
находящие подходящих гнезд для откладки яйца, и кукушата,
растущие в неподходящих условиях) вполне мог быть фактором,
125
обеспечивавшим быстрые генетические перестройки. (То же можно
сказать и обо всех формах паразитизма.) Кстати, для некоторых
видов кукушек известно [Котт, 1950, с. 474], что имитирующий
рисунок бывает нанесен на скорлупу поверх ее основного (голу-
бого) цвета. Сам же этот рисунок не зависит от самца, т. е. видимо
подчиняется нехромосомной наследственности. Это, а также ог-
ромная изменчивость (обыкновенная кукушка паразитирует
на 150 видах воробьиных) говорят о незакрепленности генетиче-
ских программ. Генетический поиск здесь, вероятно, весьма интен-
сивен и сейчас, причем выбирает он варианты не из хаоса, а из
«клеток» архетипической «таблицы», откуда и сходство расцветок
яиц.
Главный методический вывод, какой можно сделать сейчас,
когда исследования едва начинаются, состоит в том, что следует
говорить не только о приспособлении к среде, но и о приспособ-
лении к наличной тенденции таксона. Спектр возможных при-
способительных реакций организма очень широк — от простой
смены поведения до дистресса, и мы почти еще не знаем, как свя-
зано это разнообразие реакций с разнообразием генетических
перестроек. Многие наблюдаемые свойства и действия могут быть
не приспособлениями, а просто остатком от прежнего разнообразия
морфо-физиологических реакций, результатом рутинизации;
они удивляют нас только потому, что мы смотрим на них, как на
специальные приспособления, невесть каким образом преду-
смотренные и достигнутые. Точно так же нас поражают изобрета-
тельность и проницательность персонажей детектива, пока мы не
узнаем, как это сделано: автор вычленял заране известную раз-
вязку, запрещая героям все, что не ведет к ней.
Глава 5. РАЗНООБРАЗИЕ, НОРМА И ЦЕЛОСТНОСТЬ
5.1. Что такое норма
В обыденном отношении к разнообразию норма является, пожалуй,
самым главным. Не будучи в силах охватить мыслью разнообразие,
неискушенный наблюдатель ищет в нем чего-то простого, устойчи-
вого и в то же время — достаточно существенного, словом, чего-то
нормального. Понимания нормы могут быть настолько различны-
ми, что между ними трудно указать связь. Это хорошо показал пи-
сатель Андрей Битов [1976, с. 536], приведя широкий спектр пони-
маний — от чего-то среднего и обычного, через некий идеал к «бо-
жественной норме» в искусстве и «норме творения» в природе.
В предыдущих главах мы уже встречались с разными понима-
ниями нормы, а сейчас надо выяснить связь между ними. Самое
простое — извлечь из разнообразия один, чем-то приглянувшийся,
элемент и назвать его нормой. Так поступает, например, система-
126
тик, давая отряду воробьиных (воробьинообразных) имя по име-
ни одного из родов этого отряда; само название «воробей» будет
всегда сохраняться за тем родом, к которому будет отнесен вид
«воробей домовый», точнее те экземпляры, которые описал Линней.
Таков общепринятый норматив систематики.
Понимание нормы как норматива (или соглашения) очень ста-
ро и проще всех усваивается, оно легко смыкается с житейским
«делай, как положено», но ведь и сам норматив должен быть выб-
ран разумно, чтобы им можно было руководствоваться. Музейный
экземпляр должен (если есть возможность выбора) быть в каком-то
смысле типичным для своего вида, чтобы его можно было счесть
нормальным, и вот, как мы уже говорили, со времен Кетле стали
склоняться к пониманию нормы как среднего. Оно-то и вошло в
дарвинизм. Против этого возражал зоолог С. А. Усов, говоря: дарви-
низм обычно трактует уклонения от среднего как случайные ано-
малии, но «во-первых, нельзя назвать аномалией то, что не укло-
няется от нормы, а в изменяющемся беспредельно и постоянно
норма не устанавливается, во-вторых, случайность здесь также не
у места, ибо уклонения происходят вовсе не случайно, а по закону
причинности и необходимости», пусть еще и не понятому [Усов,
1867, с. 86].
Мысль эта, не привлекшая тогда внимания, очень глубока:
пусть среднее и можно вычислить, но из этого еще не следует, что
им всегда можно осмысленно воспользоваться. Так, температура
тела человека ниже 36° и выше 37° считается ненормальной,
но не на основании сплошного усреднения, а в результате сравне-
ния больных со здоровыми: те, у кого все системы работают нор-
мально, почти всегда (к сожалению, без этого «почти» при анализе
разнообразий не обойтись) имеют температуру 36°—37° С. Только
в этом кластере норму можно ввести как среднее (36,5±0,5° С),
после чего положить (в качестве норматива) температуру выше
37° признаком заболевания, т. е. признаком принадлежности к дру-
гому кластеру (больных). Другими словами, нормальная темпера-
тура вводится как составная часть физиологической нормы —
понятия целостного, такого же целостного, как таксон, красота,
разумность и многие другие (в философии их называют категория-
ми) , которые мы понимаем, хотя и не можем дать им краткого
определения. Если врач, прибыв в чужую страну, обнаружит у
жителей среднюю температуру 38°, то он сочтет ее местной нормой
только при условии, что их можно будет всех счесть физиологиче-
ски здоровыми. В противном случае он признает, что в стране
эпидемия.
Часто физиологической норме все-таки пытаются дать краткое
определение, обычно в терминах кибернетики. Самое простое пони-
мание при этом состоит в том, что каждый параметр удерживается
в определенных рамках за счет работы некоторой следящей систе-
мы, подобной автоматическому регулятору в технике (в 1.5 мы ви-
дели, что традиция усматривать в природе подобие наших машин
восходит к Парацельсу и у потомков обычно вызывает только улыб-
127
ку). В технике норма, т. е. контрольное значение параметра (ус-
тавка) задается человеком, но кто задает ее в организме? Неужели
распределение температуры (ноги — холоднее тела, но разогре-
ваются при ходьбе; к вечеру температура чуть возрастает, при
инфекции - устанавливается на высоком уровне и т. п.) во всех
деталях предписано этими таинственными уставками? Подобные
вопросы породили более сложные понимания нормы, например,
иногда ее понимают не как вездесущее слежение, а как состояние,
в котором поисковая система не ведет поиска [Фицнер, 1977].
Система терморегуляции включается только тогда, когда тело не
может удовлетворительно работать из-за перегрева или переохлаж-
дения; болевая реакция означает, что следует искать утоляющее
боль средство — вот идеология этого подхода. Однако остается
неясным, почему температура так стабильна, почему иногда боль
утихает от согревания, почему боль не всегда связана с какой-то
патологией (болят мышцы после нагрузки) или даже препятствует
поиску (болевой шок) и т. д.
Эколог Юджин Одум [1986, т. 1, с. 61, 64] дополняет кибернети-
ческую схему в двух отношениях: во-первых, в биологических
системах нет особых «управляющих элементов» (а следовательно,
добавим, и уставок) — роль управляющей играет одна подсистема
относительно другой, а во-вторых, высокую стабильность пара-
метров может обеспечить избыточность функциональных подси-
систем, работающих параллельно. Он однако ничего не говорит
о природе нормы («постоянной точки», как он ее называет).
Часто норму трактуют как функциональную устойчивость
(ненормально то, что нарушает устойчивую работу органа или
организма) или функциональный оптимум (ненормально вообще
всякое ухудшение работы), но это — тоже лишь кибернетический
пересказ того, что норма является характеристикой целостной.
Более того, в самых простых случаях эти определения не работают:
летаргический сон так же функционально устойчив, как спячка,
но все же ненормален; Ньютон и Дарвин были слабы здоровьем
и в этом смысле далеки от нормы, но оба дожили до старости,
и трудно назвать их жизнь неоптимальной (хотя временами она
теряла устойчивость). Наоборот, при самой нормальной (опти-
мальной) беременности многие органы работают с большой пере-
грузкой (ненормально в физиологическом смысле) и все же не
обеспечивают устойчивой работы организма (он просто не успевает
разрушиться за время беременности). Вопрос о том, свойственна
ли вообще организмам оптимальность, далек от решения [Чайков-
ский, 19886].
Словом, идея рассматривать норму как кибернетический фено-
мен не более убедительна, чем идея объяснить все признаки их по-
лезностью. Обе способны объяснять специально подобранные при-
меры, но не выдерживают проверки разнообразием. И все-таки
кибернетическое понимание нормы, полезное для наших целей,
существует. Хоть и далекое от универсальности, оно тем не менее
способно разъяснить кое-что в многогранной проблеме разно-
128
образия. Его предложил, развивая идею Т. Уотермана, биокибер-
нетик В. Н. Новосельцев [1973], на чью работу мне любезно указа-
ла А. В. Григорьева, открывшая мне тем самым весь круг
соответствующей проблематики.
Будем рассматривать биологическую систему (клетку, орган,
организм, экосистему и т. и.) не как набор разнородных следя-
щих систем, а как открытую термодинамическую систему (термин
«открытая» означает, что через нее текут потоки веществ, энергии
и, возможно, информации). Баланс открытой системы заключа-
ется в том, что стабилизируются потоки через систему, а стабиль-
ность внутренних параметров достигается при этом сама собой,
причем Новосельцев показал, как это получается.
Сам по себе баланс достижим без следящих систем: цыплят, на-
пример, выводили в инкубаторах задолго до появления термомет-
ров — слабый подогрев оказался способным заменять стабилизиро-
ванное тепло наседки. Однако лучше (надежнее), если подогрев ре-
гулируется, и проще всего это достигается, если нагреватель вклю-
чается и выключается автоматическим термометром. Существенно,
что таким путем можно стабилизировать температуру инкубатора,
но — не ток через нагреватель: именно скачки его и стабилизируют
температуру. Очевидна асимметрия процесса: один параметр ста-
билизируется, другой, его стабилизирующий, сам колеблется,
Если мы хотим стабилизировать температуру очень точно или
в очень разнообразных внешних условиях, то систему надо услож-
нить: ввести, кроме грубого регулятора подогрева, второй, тонкий;
ввести охладитель, регулируемую теплоизоляцию и т. п. И тут ока-
зывается, что стабилизируется не только температура, но и регули-
рующие воздействия (электрический ток через нагреватель, ток
воды через охладитель, положение теплоизоляции). Разнообразие
регуляторных связей симметризовало систему: параметры стали
равноправными, их теперь нельзя разделить на регулируемые и
регуляторы. Здесь естественно назвать нормой ситуацию, когда
исследуемые параметры симметричны, равноправны, в равной
мере стабилизированы. Так, мы говорим, что режим инкубатора
ненормален, если постоянная температура в нем поддерживается
скачками в работе нагревателя или охладителя. Если стабилизация
одной величины достигается большими перепадами другой, то,
следовательно, регуляцию ведет один контур обратной связи
(одна следящая система) вне области фактической работоспособ-
ности других контуров — в этом и состоит ненормальность. В част-
ности, и в экологии колебания численности организмов «усили-
ваются в том случае, если численность популяции хозяина регу-
лируется только одним паразитом» [Макфедьен, 1965, с. 284].
Такое понимание нормы разъясняет много чисто физиологиче-
ских вопросов. Нет надобности предполагать наличие особой систе-
мы стабилизации для каждого параметра, который мы видим стаби-
лизированным: если стабилизированы потоки через систему, то
внутренние параметры стабилизируются сами собой. Не представ-
ляет исключения и температура тела, являющаяся результатом ста-
5 Ю. В. Чайковский
129
билизированного метаболизма; и ноги при ходьбе не мерзнут пото-
му, что при сокращении мышц выделяется тепло (баланс устанав-
ливается при большей температуре), а не вследствие изменения за-
данной свыше уставки. Кстати, именно при смене режима работы
(или условий среды) выявляется различие между регулируемыми
и регуляторами: первые сохраняются, а вторые устанавливаются на
новых уровнях.
Однако не всякую норму можно понимать как физиологиче-
скую — например, необычное число пальцев может никак не
влиять на физиологию и все-таки это считается ненормальным,
причем без всяких соглашений или статистики. Здесь норма —
соответствие объекта его архетипу (по Оуэну).
5.2. Логика ценоза
Итак, понятие нормы осмысленно только для целостной системы,
и стабилизировать, например, саму по себе температуру тела столь
же бессмысленно, как подогревать до 37° труп. Далее, в области
нормы нет смысла рассматривать одни параметры как причины,
а другие — как следствия, поскольку они для наблюдения равно-
правны. Это обстоятельство относится равно и к клеткам, и к орга-
низмам, и к их сообществам, что позволило математику С. Г. Смир-
нову охарактеризовать его в 1979 г. (на семинаре в МОИП) как
«логику ценоза», сменяющую в теории систем привычную причин-
но-следственную логику. Эта новая логика еще совсем не разрабо-
тана, но очевидно, что разрабатывать ее имеет смысл только для
систем с достаточно богатым внутренним разнообразием — ведь
сама идея равноправия переменных возникает, при исследовании
систем, в которых каждый параметр имеет богатое множество
регуляторов (и сам, соответственно, служит регулятором для
многих).
Тем самым намечается некоторый просвет в казалось бы бес-
просветной проблеме, которую поставил Спенсер, когда рассматри-
вал всякое развитие как взаимодействие противоположных тенден-
ций — дифференциации и интеграции (см. 1.2). На ряде примеров
он показывал, что «однородное неустойчиво» (эту идею он почерп-
нул из эмбриологических работ Бэра 1820-х годов), а сто лет спустя
экологи признали, что слишком однородные экосистемы действи-
тельно неустойчивы — северные экосистемы, содержащие немного
видов, чаще подвергаются вспышкам численности отдельных видов
и легче разрушаются человеком, чем экосистемы теплых стран;
в жарких странах такие вспышки случаются в бедных видами полу-
пустынях (превращение саранчи из безвредной оседлой формы
во всепожирающие мигрирующие полчища), но не в богатых вида-
ми влажных тропиках; что же касается искусственных экосистем,
где огромные поля засеваются единственной культурой, то они во-
обще без управления человеком существовать не могут.
Все это так, но уточнить тезис, что разнообразие видов способ-
ствует устойчивости экосистемы, не удается ни конкретными иссле-
130
дованиями, ни математическими моделями [Левин, 1980, с. 50].
«... Между видовым разнообразием как таковым и стабильностью
не существует сколько-нибудь значительной корреляции» [Одум,
1986, т. 1, с. 65]. Дело в том, что эколог чаще всего ищет обоснова-
ние тезиса Спенсера («однородное неустойчиво») просто: сравни-
вает две экосистемы с различным видовым разнообразием и изме-
ряет в обеих колебания численности видов, а не обнаружив боль-
шей стабильности в более разнообразной экосистеме, приходит
к выводу, что тезис Спенсера сомнителен или даже ложен. Еще
проще идеология математических моделей — в них считается, что
каждый вид встречается в своей жизни с одной-единственной труд-
ностью (или с голодом, или с холодом, или с засухой и т. и.), а хоро-
шей (нормальной) считают ту экосистему, в которой параметры
стабильны.
Взглянем на проблему с позиции понимания нормы системы
как симметрии ее внутренних параметров при стабилизации пото-
ков через нее. Прежде всего, если действительно стабилизируются
потоки (веществ и энергии), то измерять надо их, а не числен-
ности, которые могут быть, в лучшем случае, регуляторами. Далее,
стабильность можно считать нормальным явлением только тогда,
когда стабильны внешние воздействия на систему — в противном
случае нормальная система просто обязана изменяться, чтобы под-
страиваться к переменным условиям. Недаром ведущий эколог
Роберт Уиттекер склонялся к выводу, «что не сложность сообщест-
ва приводит к стабильности сообщества, а что стабильность среды
позволяет развиваться сложным сообществам» [1980, с. 68].
Наконец, тезис Спенсера (как и рассматриваемое нами понима-
ние нормы) ясно говорит, что для устойчивости нужна некоторая
неоднородность, но никак не безграничный рост разнообразия.
В самом деле, обычно пять слоев регуляции достаточны (в модели
Новосельцева) для симметризации регулируемых и регуляторов,
а вместе с тем и для стабилизации. Вспомним, что согласно канонам
экологии число звеньев трофической цепи (А поедает В, В поедает
Сит. д.) обычно не превышает шести. Например: зеленое растение,
травоядное животное, хищник, его паразит, мертвоядное живот-
ное, разлагающая бактерия (или гриб) — причем обычно цепь ко-
роче, так как большая часть разлагающих организмов питается
непосредственно отмершими растениями. Иногда цепь бывает
длиннее — за счет каннибализма, вторичного хищничества или
вторичного паразитизма, когда существенны семь звеньев. (Более
длинные цепи редки и не носят обязательного характера — напри-
мер, паразиты третьего и четвертого порядков известны, но у них
«наблюдается малая разборчивость по отношению к объекту, их
нападений» [Догель, 1941, с. 156], т. е. они могут поражать и
свободноживущих, и их паразитов.) Если бы разнообразие видов
распределялось по трофической цепи равномерно, то уже 5X7=35
видов было бы с избытком достаточно для стабилизации любой
экосистемы. Учтя неравномерность, можно ожидать, что для ста-
бильности достаточно 100 видов, что большее обилие ничего не
5*
131
добавит. Так оно и есть в действительности. Более того, С. В. Мейен
высказал в 1978 г. (на семинаре в МОИП) догадку, что рост разно-
образия системы выше некоторого — необходимое условие для ее
эволюции, а вовсе не для стабильности.
В известном руководстве [Риклефс, 1979] приводится такой
пример в пользу тезиса Спенсера. Сравнивали насекомых на ка-
пусте, засеянной среди сорняков и на чистом поле, и выяснили,
что сорняки способствуют сохранению капусты от вредителей. За-
мечательно, что на обоих полях хищников было достаточно, но на
чистом поле они не могли сдержать аппетиты вредителей, посколь-
ку размножались тем же темпом, что и вредители, но с запазды-
ванием. Наоборот, среди сорняков всегда находилась готовая орава
хищников, питавшихся до всхода капусты другими видами насе-
комых [там же, с. 392]. Здесь роль разнообразия очевидна. Столь
же ясна она и в других случаях — там, где автор руководства ее
не увидел. Наиболее показателен опыт по внесению удобрений на
луга с различным разнообразием животных: более разнообразная
система, разумеется, откликнулась на рост продуктивности расте-
ний более эффективно; но эколог ожидал почему-то, что она должна
«лучше противостоять вмешательству» [там же, с. 393]. Никаких
оснований сомневаться в тезисе Спенсера здесь нет.
В последние годы в связи с общесистемными концепциями
(«гипотеза Геи» Джеймса Лавлока [Заварзин Г. А., 1984; Одум,
1986], «новый функционализм» [Уголев, 1985] и др.) вновь обре-
ли популярность старые параллели между организмом и экосисте-
мой. С этой точки зрения естественно спросить, не следует ли ви-
деть взаимную подстройку и симметрию отношений там, где до сих
пор говорили только о хищнике и жертве? Все «цепи питания» имеют
кольцеобразную форму (вспомним древний афоризм: курица ест чер-
вя, человек — курицу, червь — человека), и в этом смысле звенья
их симметричны — по отношению не друг к другу, а ко всей
системе.
В 1932 г. зоолог У. Мак-Ати опубликовал удивительный подсчет:
анализ содержимого 80 тысяч птичьих желудков (500 видов птиц)
убедил его в том, что «число поимок пропорционально встречаемо-
сти данного вида» насекомых, т. е.— что класс птиц как целое охо-
тится за классом насекомых как целым, невзирая на окраску, ядо-
витость и прочие защитные качества конкретных видов насекомых.
Мак-Ати пришел к выводу о полной неэффективности всех средств
защиты [Котт, 1950, с. 196]. Котт резко осудил этот подход, шед-
ший вразрез со всей тогдашней идеологией (требовавшей «иссле-
довать каждый случай в его специфике»), но признал, что почти
каждая крупная группа животных дала бы сходные результаты,
и сделал неожиданный вывод: «Если ... гибель от хищников про-
порциональна численности жертв, то это лишь показывает, что
в природе существует известное равновесие между различными
типами жертв и что разные типы защитных приспособлений при-
мерно одинаково охраняют их обладателей» [там же, с. 198].
Следовательно, Котт признал данные Мак-Ати (ни одна защита
132
не дает преимуществ), но сделал из них противоположный вы-
вод — защита необходима. Мимикрирующих насекомых, пояснял
он, едят, в основном, те виды птиц, которые специализированы
в их- поиске; если бы не мимикрия, эти виды насекомых стали бы
общей добычей и были бы съедены целиком. Обе стороны были
в какой-то мере правы: Мак-Ати отметил феномен глобальной
экологии — баланс между крупными группами, а Котт указал,
что этот баланс достигается приблизительно равной эффектив-
ностью различных средств защиты.
Среди работ, обсуждавших этот вопрос, следует особо отметить
ту, где было введено понятие адаптивного компромисса [Norris,
Lowe, 1964]. Там оно касалось частного аспекта—приспособ-
ления к противоречивым условиям среды (например, когда живот-
ное обитает на нескольких фонах, имеющих различные окраски),
однако суть его гораздо значительнее. О ней говорил еще Дарвин
[1939, с. 420]: «Если подвести итог (в подлиннике: balance —
10. Ч.) добру и злу, причиняемому каждой частью организации,
то в целом каждая данная часть окажется скорее полезною». Одна-
ко такая мысль, если ее продумать глубже, опрокидывает всю идею
естественного отбора, поскольку отбор компромисса означает
отсутствие отбора отдельных свойств. Это отметил в 1894 г. Бэтсон.
Он писал: нет оснований утверждать полезность каждого призна-
ка, каждый инстинкт или орган может быть полезен, но нельзя
говорить что его полезность доказана и тем более, что она — причи-
на его происхождения. «Мы, животные, живем не только в силу,
но также и вопреки тому, чем обладаем... Помимо первоначального
знания о том, что организмы более или менее приспособлены к
своим обстоятельствам, мы можем достигать лишь расплывчатых
знаний об адаптации»; Бэтсон заключил: «Исследуя причины
вариаций, я не считаю необходимым делать допущение об их полез-
ности или гармоничности; такое допущение бесплодно и безуспеш-
но» [Steenis, 1969 с. 100]. Другими словами, если признать адап-
тивный компромисс, то придется отказаться от адаптивного толко-
вания отдельных (пусть и очень важных) свойств — основного
приема в традиции Ламарка—Дарвина; потребуется какой-то но-
могенетический принцип, дополнительный к адаптивному.
В последнее время в связи с идеями системности адаптивный
компромисс трактуют очень широко, и он грозит выродиться в пус-
той звук (как это произошло со «стабилизирующим отбором»
Шмальгаузена, первоначально содержательным понятием, вскоре,
однако, ставшим одним из обозначений любого адаптивного про-
цесса) . Так, А. П. Расницын относит к адаптивному компромиссу
не только все формы оптимизации (что само сомнительно), но и
прямо противоречащие компромиссу идеи конкурентного исклю-
чения [Эволюция..., 1987, с. 54].
Наиболее глубоким представляется мне тот смысл адаптивного
компромисса, на который указали Мак-Ати и Котт. Прекрасный
пример равной силы разных стратегий защиты представил гнездо-
вой паразитизм воловьей птицы, исследованный Н. Смитом
133
в 1968 г.: успешный вылет птенца-паразита происходил с частотой
0,73—0,76 (а хозяйского птенца — с частотой 0,39—0,43), т. е.
колебался очень мало, хотя использовались 4 вида хозяев, 2 типа
паразитизма (с мимикрией яиц и без нее) и 3 стратегии гнездо-
строения (рядом с гнездами ос или пчел и вдалеке от них) [Пианка,
1981, с 256 — 258]. Сходна (хоть и ие столь определенна) ситуация
для ядовитых растений: различные типы ядовитости «представляют
для травоядных мощный экологический барьер, но в эволюцион-
ном масштабе они являются слабой помехой» [там же, с. 246].
В этом аспекте адаптивный компромисс указывает еще на одну
симметрию нормы — на неразличимость оптимальных действий по
их выгодности. Рассмотрим ее подробнее.
5.3. Норма прибыли и норма приспособленности
Вывод Котта перекликается с известным тезисом политэкономии,
который обосновал в свое время Карл Маркс: норма прибыли во
всех отраслях производства приблизительно одна и та же. Норма
прибыли — единая для всей экономики величина, обусловленная
единством экономической системы, возможностью свободного пере-
мещения капиталов и тем, что каждый предприниматель знает
всю конъюнктуру.
Ту же, что в экономике, внешнюю картину мы видим в экологии,
и приблизительное равенство эффектов защиты естественно на-
звать нормой приспособленности. Однако как сопоставить внутрен-
ние механизмы? Разве класс насекомых может перераспределять
средства защиты между своими видами, как перераспределяет
финансы класс предпринимателей? Разве виды обладают какой-то
информацией, которую можно было бы сопоставить с информацией
рынка? Чтобы отвечать на подобные вопросы, надо сначала выяс-
нить, какая информация в принципе необходима для обеспечения
нормы прибыли и нормы приспособленности.
В 4.3 мы видели, что равенство результатов при различных спо-
собах действия характеризует оптимальную стратегию в матричной
игре. Точнее, если выигрыш и гарантирован игроку при любом
способе действия противника, значит игрок применяет оптималь-
ную стратегию. Эта простая теорема теории игр и является модель-
ной формулировкой вывода Котта, поскольку пищевое взаимо-
отношение насекомых с насекомоядными птицами приблизительно
является матричным: выигрыш птиц есть проигрыш насекомых,
а число способов действия можно считать конечным. Если считать
первым игроком совокупность насекомоядных птиц, то способом
его действия явится способ охоты; способом действия второго иг-
рока (класс насекомых) явится любое средство защиты — цвет, яд,
угроза и т. п., а также — любая их комбинация у одного вида. Стра-
тегия — распределение этих способов по отдельным видам данного
игрока. Если каждое средство защиты (способ действия) насеко-
мых приводит к передаче птицам примерно одной и той же живой
массы (выигрыша), то можно допустить, что стратегия птиц опти-
134
мальна. Можно, хотя и сложнее, вывести отсюда оптимальность по-
ведения и для класса насекомых. Мажорирующий аспект этой игры
состоит в следующем: как бы ни была сложна реальная жизнь, как
бы ни упрощала ее линейная игровая модель, все равно сам факт
согласованности поведения видов, будучи осознан как оптималь-
ный, требует объяснения. Из теории игр мы знаем, что такая сог-
ласованность сама собой (статистически) не устанавливается,
что для оптимальности в названном смысле игрок должен действо-
вать как информационное целое. Вспомним, что наш игрок — сово-
купность видов (и подвидов), т. е. представлен множеством парт-
неров. Для таких игр справедлива упомянутая в 4.3 теорема Куна:
игрок может гарантировать выигрыш и только в том случае, когда
он как целое обладает полной памятью о действиях своих партне-
ров. Результат Куна изящен и в сущности вполне достаточен, чтобы
понять — оптимальность целого класса не могла сложиться за счет
независимого приспособления каждого вида к своим частным усло-
виям существования. Остается искать зависимости, т. е. потоки
информации между видами. Общие соображения о подобных пото-
ках будут изложены в части II, здесь же отметим одно частное сооб-
ражение, высказанное П. Шеппардом.
Он обнаружил в 1951 г., что певчий дрозд выедает преимуще-
ственно тех садовых улиток, цвет раковин которых плохо их маски-
рует [Пианка, 1981, с. 239]. Затем, в 1956 г. Шеппард выяснил, что
когда один вид улиток становится менее уязвимым (за счет сезон-
ного изменения фона), то другой, хуже замаскированный, вид под-
вергается усиленному выеданию, и сделал из этого вывод: хищник
как бы выравнивает жертв в отношении селекционного пресса,
побуждая их всех к мимикрии [Макфедьен, 1965, с. 26]. Что каса-
ется конкретных путей мимикрирования, то мы можем указать на
следующее. Во-первых, оно идет не произвольными путями, а пос-
редством заполнения «клеток» соответствующих рефренов, причем
плотности заполнения этих «клеток» характерны для данных раз-
нообразий в такой же степени, как и для влияний среды. Во-вто-
рых, сохранение или несохранение «клетки» определяется адап-
тивным компромиссом, а вовсе не полным приспособлением к ка-
кому-то определенному фону (или другому типу защиты). И, в-
третьих, удивляющее нас подчас соответствие друг другу хищника
и жертвы может быть попросту результатом рутинизации (4.6).
Каждую из реализованных возможностей (заполненных «кле-
ток» разнообразия) можно условно назвать адаптивной нормой,
и в этом смысле эволюцию мимикрии действительно можно пони-
мать как «смену адаптивных норм» по Шмальгаузену, но отнюдь
не как «сдвиг» какой-либо нормы. Вообще, уверенность морфоло-
гов-эволюционистов в том, что «любое направленное эволюционное
изменение может быть охарактеризовано как сдвиг фенотипиче-
ской нормы» [Экология ..., 1984, с. 205] просто ошибочна — она
не учитывает дискретности архетипической таблицы по многим
(если не по большинству) параметров. К этому вопросу мы вер-
немся в части II, при обсуждении дилеммы «дискретность — не-
прерывность» в эволюции.
135
5.4. Многомерность нормы
Понимание нормы как оптимального поведения в игре позволяет
сформулировать ряд понятий, имеющих более общее значение, чем
сам игровой подход. Первое такое понятие — адаптивный компро-
мисс — заведомо шире, чем та антагонистическая схема, которой
мы его иллюстрировали. Оно заставляет отказаться от одномерно-
го понимания нормы: приспособление обычно не состоит в подгон-
ке какого-то одного параметра к требованиям среды — приемлемая
область его значений зависит от всей системы организм —среда.
Эта область редко включает экстремальное значение параметра —
в этом и состоит адаптивный компромисс.
Но если так, то приходится отказаться от многих традиционных
представлений. Например, нельзя считать нормой стремление ор-
ганизмов к неограниченному размножению. Такое стремление под-
час наблюдается, но не как норма, а как резкая аномалия, когда
размножаемость выходит из-под эко-физиологического контроля.
Таковы нашествия саранчи, эпидемии чумы и т. д., а в рамках одно-
го организма — злокачественные опухоли. Дарвин признавал толь-
ко экологическую форму контроля ’, да и ее — лишь в форме «борь-
бы за существование», мы же сейчас знаем множество поведенче-
ских и физиологических форм ограничения численности, как пря-
мых (рассасывание беременности у зверей, прекращение гнездо-
строения у птиц, каннибализм у самых различных групп и т. д.
[Панов, 1983]), так и косвенных (адаптивный компромисс, сим-
биоз, а заодно и все другие формы сосуществования — от индиви-
дуальных участков высших животных до «известной спаренности
некоторых свободноживущих и паразитических групп» [Догель,
1941, с. 39]).
Второе понятие — норма как устойчивость — также невозмож-
но при одномерном понимании нормы, но здесь надо отказаться
и от двумерных иллюстраций. Именно засилье двумерных (плос-
ких) графиков, должных иллюстрировать общие (многомерные)
процессы, долго не давало натуралистам увидеть главное — кон-
фликт и неустойчивость. Мы говорили в 4.3 о том, что выбор игрока
между тремя (и более) возможностями порождает бесконечные
блуждания, если его информированность неполна, а в 5.1 — что
одной пары переменных (регулируемое и регулятор) недостаточно
для стабилизации параметров. Следует сказать и про детермини-
рованные системы, в которых нормой является хаотическое дви-
жение, наблюдаемое при выполнении некоторого условия неустой-
чивости. В динамической (задаваемой дифференциальными урав-
нениями) системе это условие — критерий стохастичности — хо-
рошо изучено [Заславский, 1984, с. 42]. Здесь плоский случай тоже
вырожден — в том смысле, что лишь при размерности пространства
ЛС^З фазовая «траектория может уходить сколь угодно далеко от
1 В отличие от Мальтуса, у которого мальтузианцы заимствовали лишь самый при-
митивный тезис — «геометрическую прогрессию размножения». В действитель-
ности демографическая концепция Мальтуса достаточно сложна.
136
1
области невозмущенного движения» [там же, с. 26]. Уже при Л’=3
в таких системах могут возникать странные аттракторы [там же,
с. 250] — устойчивые области притяжения траекторий, между ко-
торыми траектория как бы блуждает, имитируя последовательность
независимых испытаний. Если не требовать непрерывности траек-
торий, то стохастичность детерминированной системы можно наб-
людать и при N=2 [там же, с. 31]. Вопрос о том, в какой мере та-
кая случайность является нормой для биологических явлений, еще
предстоит выяснить.
Сказанное позволяет, наконец, через 120 лет ответить на воз-
ражение Усова (см. 5.1): вводить случайность в рассмотрение де-
терминированного процесса вполне уместно, но вот называть одно
состояние нормой, а другое аномалией, исходя из одних лишь час-
тот наблюдения, в самом деле нельзя. Функциональная норма оп-
ределяется не статистически, а эко-физиологически, причем обыч-
но приходится говорить о нормальности или аномальности не от-
дельных объектов (элементов), а их разнообразий. Жизнеспособ-
ное разнообразие можно считать нормальным, равно как нормален
и всякий его жизнеспособный элемент. Отсекая ненормальные
(нежизнеспособные) элементы, создать новое разнообразие нель-
зя — по крайней мере, примеров тому неизвестно (подробнее об
этом см. в части II, при анализе транзитивного полиморфизма).
Если норма целостна, то, перефразируя афоризм Линнея из 1.3,
можно сказать: не параметры задают норму, но норма задает пара-
метры. Этот тезис (как и сам афоризм Линнея) имеет отношение
к систематике, поскольку норма задает ядро таксона, т. е. нечто ти-
пичное. Это нечто не всегда выразимо на языке параметров и при-
знаков, и все-таки классификация как-то производится: «Я мог бы
узнать англичанина где угодно,— подумал Майкл,— а общих физи-
ческих признаков нет» [Голсуорси, 1956, с. 97].
Глава 6. РАЗНООБРАЗИЕ, СИММЕТРИЯ
И МАКРОСИСТЕМА
Чтобы перейти от описания свойств разнообразий к описанию их
эволюции, надо хотя бы на одном примере увидеть, как какое-то
1 разнообразие устроено в целом. Сделаем это на примере макросис-
тем^ (системы царств) живой природы. Поскольку чисто биологиче-
ский анализ этой темы проведен ранее [Чайковский, 1986] и снаб-
жен ссылками на необходимую литературу, здесь мы можем
сосредоточиться в основном на историко-методологическом анализе
• проблем макросистематики.
Можно сказать, что до работ американского эколога Роберта
Уиттекера макросистематики не существовало: хотя царства и упо-
минались как высшие таксоны, но в систему увязаны не были. Под-
разумевалось, что царство — такой же таксон, как класс или род,
т. е. может быть определено филогенетически, в терминах родства
, 137
и приспособления. Теория Уиттекера и методологический анализ
Мейена убеждают, что это далеко не так: царство является прежде
всего эко-физиологическим единством, а таксономическое значение
всех форм родства и приспособления еще нуждается в разработке.
Достаточно заметить, что систематики часто говорят о приспособи-
тельном характере таксономических различий, однако на практике
поступают по-другому — признают наиболее ценными признаки,
не имеющие адаптивного значения, поскольку именно их принято
считать унаследованными от общего предка наверняка. Так, одно-
клеточные динофлагеллаты (панцырные жгутиконосцы) объедине-
ны в таксон прежде всего из-за наличия общего признака — коль-
цевого желобка и в нем жгутика. Признак явно бесполезен и со-
храняется во всех средах, в том числе при паразитизме, а потому
и считается предковым. Но если так, то почему он наблюдается
у клеток самого различного внутреннего строения? Не есть ли это
обычный пример параллелизма? Словом «существует несоответст-
вие между учением об адаптации как основной силе, направляю-
щей эволюцию, и анализом, который проводит филогенетическая
систематика» [Рыжков, 1974, с. 97]. Нельзя забывать, что «филоге-
нетическая система может передать лишь ретроспективные генеа-
логические отношения, а для отношений других типов нужны
другие системы» [Красилов, 1975, с. 119].
6.1. Система и макросистема
С древности организмы делили на растения и животных, и в этом
видели разделение их функций: растения производят, а животные
потребляют. За последние сто лет выяснилось, что животные отхо-
ды становятся пищей для растений в основном после их минерали-
зации микробами и грибами, которых, тем самым, нельзя относить
к растениям, коль скоро растения определены экологически.
В 1959 г. Уиттекер свел новые знания в формальную схему: царст-
во растений — продуценты, т. е. производят органическое вещество
путем фотосинтеза; царство грибов — редуценты, т. е. разлагают
тела и отходы растений и животных; а царство животных — про-
межуточное звено, консументы (потребители). Подобно трем ле-
песткам, эти три царства вырастали в схеме Уиттекера из общего
основания — царства протист (простейших), в котором тенденции
всех трех высших царств выражены одновременно, но в менее яс-
ной форме: одноклеточные водоросли и фотобактерии примыкают
к растениям, а гнилостные бактерии и актиномицеты — к грибам.
Красота и глубина этой простой схемы состояла в том, что Уит-
текером были выявлены три ведущие экологические тенденции:
растения — фототрофы, грибы — осмотрофы (питаются за счет
осмосорбционного транспорта растворов из среды), и животные —
фаготрофы (заглатывают пищу). Соответствие двух триад:
продуценты — редуценты — консументы, (1)
фототрофы — осмотрофы — фаготрофы (2)
— выполняется в целом, но не на всех объектах. Так, растение
138
росянка может ловить и переваривать насекомых. Все подобные
примеры свидетельствуют, что речь идет только о тенденциях, но
не о точных абсолютных законах и не о четких границах.
Главное же, чем важны для макросистематики триады, это
иной, нежели в остальной систематике, принцип классификации:
они рассекают мыслимое множество организмов по некоторым
априорным правилам, тогда как остальная систематика построена
по принципу конгрегации, т. е. объединения объектов с близкими
свойствами. Как секционный, так и конгрегационный метод может,
будучи применен от начала до конца процедуры классификации,
дать иерархическую систему, но первый не дает того, что обычно
называют видом, а второй — того, что называют царством, поэтому
их приходится комбинировать, и это одна из причин того, почему
иерархии не получается.
Как мы видели в 2.1, секционным путем нельзя дойти до по-
нятия вида потому, что вид состоит не из признаков, а из цельных
организмов; низшие таксоны разумно классифицировать конгре-
гационно. Наоборот, конгрегационные системы царств неизменно
оказываются лишь перечнями разрозненных названий царств, сос-
тав которых ни для кого не убедителен, и ни одна система не полу-
чает признания. Отсутствие единой системы влечет неустойчивость
названий, а это — бич систематики. Далее, конгрегацию тради-
ционно понимают как общность происхождения, а она (даже если
признать этот подход в принципе приемлемым) на уровне царств
бессмысленна: ведь древнейшие организмы тоже образовывали
экосистему, поэтому нет оснований выводить все растения (или
всех животных) из единого предка. Макросистему лучше строить
секционным путем. Желательно, чтобы она опускалась до типов,
поскольку именно до них поднимается обычная сегрегационная
систематика.
Тип — высший таксон, для которого существует какое-то еди-
ное морфологическое понимание. Морфолог В. Н. Беклемишев
[1964, с. 13] писал: «В основу разделения на типы следует класть
не столько органологические различия, сколько архитектониче-
ские». Другими словами, тип определяется планом строения орга-
низмов, а отдельные свойства органов играют в его определении
подчиненную роль. Напомним, что Агассиц рассматривал таксоны
от класса и ниже как последовательную детализацию и конкрети-
зацию (в соответствии с требованиями среды обитания) того плана
строения, который задан типом [Agassiz, 1857, с. 170]. Правда,
само понятие плана строения (а, следовательно, и типа) зависит от
позиции исследователя. Так, для ботаника динофлагеллаты — осо-
бый тип водорослей, поскольку обладают редким в ботанике дор-
зовентральным строением (можно выявить спинную и брюшную
стороны). Наоборот, для зоолога это строение обычно, а фотосин-
тез он к плану строения не относит, и в зоологической системе ди-
нофлагеллаты — всего лишь отряд. Поэтому только логика самой
макросистемы определит, считать ли данную группу типом или
нет. А там, где о плане строения говорить не приходится, нет смыс-
139
ла говорить и о типах. Это относится прежде всего к бактериям:
самое авторитетное руководство [Bergey, 1984] во вводной статье
предлагает разделить все бактерии на четыре типа, но не пользует-
ся этим делением при описании конкретного материала и, более
того, утверждает, что «официальной классификации бактерий не
существует» [там же, с. 3].
Как говорилось в 1.3 и 2.3, бесконечная смена названий связана
с тем, что со времен Жюссье на одну систему возложены три
несовместимые функции — номенклатура, адресация и естествен-
ность. Еще Линней понимал, что естественность — нечто более глу-
бокое, чем удобство адресации, а Бэр уточнил естественность как
сходство строения, а не отдельных (пусть и многих) частей или
свойств; он полагал, что естественную систему следует то и дело
пересматривать в связи с новыми фактами [Бэр, 1959, с. 300, 372].
Это мнение бытует до сих пор [Тахтаджян, 1987, с. 23], хотя есть
более конструктивная традиция Уэвелла—Любищева, разработан-
ная недавно Мейеном. Для Уэвелла естественность состояла
в устойчивости — чтобы систему не приходилось пересматривать
при появлении новых классификационных признаков. Любищев
соединил Линнея и Уэвелла, признав естественной ту систему,
«где все признаки объекта определяются положением его в си-
стеме». Для Дарвина и Геккеля естественность системы состояла
в общности предка каждой группы, для Вавилова и Кренке —
в прогностичности (возможности предсказывать новые формы),
для Е. С. Смирнова — в общепризнанности, окончательности и
единственности, «не допускающей вариантов» [Смирнов, 1969,
с. 6] и т. д. Вообще, как отмечает В. В. Корона [1987, с. 25], пони-
мание естественности определяется философскими представле-
ниями систематиков и неминуемо изменяется со сменой философ-
ских норм эпохи. Это куда разумнее, чем считать неизбежным
непрерывный пересмотр или, наоборот, видеть в своих взглядах
окончательную истину. Большинство же систематиков до сих пор
полагает, что сходство, как правило, свидетельствует о родстве
(если не доказано противное), не замечая, что само понятие
родства чаще всего основано на сходстве.
Налицо три методологические установки — бесконечное описа-
ние, выбор какой-то позиции в качестве окончательной и порочный
круг. Методолог В. Ю. Милитарев объединил их термином «трилем-
ма Мюнхаузена», имея в виду барона, вытащившего себя за волосы
из болота [Теория и методология..., 1983, с. 115]. Уход от трилеммы
он видит в том, чтобы основывать классификацию на предметных
принципах, отражающих свойства классифицируемого разнообра-
зия, а не способ классификации. Воспользуемся этим советом, для
чего необходимы два замечания.
Во-первых, как иерархия, так и комбинативность — не пред-
метные принципы, а, только способы классификации. Распростра-
ненное мнение, что «чистая комбинаторика всех признаков несов-
местима с иерархическим строением системы» [Смирнов, 1969,
с. 92], верно только тогда, когда признаки неупорядочены. Если же
140
какой-то признак комбинативной системы назван первым, то, про-
ведя членение по этому признаку, «мы можем вернуться к исход-
ному таксону и провести членение по другому признаку, соответ-
ственно выбрав другой мерой», и последовательность таких актов
«приводит к появлению иерархии таксонов и признаков» [Мейен,
Шрейдер, 1976, с. 76].
Во-вторых, номенклатура, (система названий) сама по себе
не связана ни с предметными принципами, ни со способом клас-
сификации. Ее назначение — обладать «мнемонической цен-
ностью» и «облегчать общение» [Джеффри, 1980, с. 34, 53],
ее «главная причина затруднений... в том, что наши системы
классификации непрерывно изменяются» [там же, с. 31]. Кодексы
запрещают произвольное переименование, но оно непрерывно идет.
Например, наглядный термин «цветковые» предложено заменить
на «магнолиофиты», потому что магнолиевые признаны как самый
архаичный порядок среди цветковых. (Это примерно то же, что
переименовать млекопитающих в утконосовых.) Однако самым
архаичным внутри порядка признана не магнолия, а дегенерия
[Тахтаджян, 1987, с. 24, 29], так что новый термин по сути неверен
и лишь затрудняет общение. Плох он и мнемонически, так как
связывает выявление исходной формы цветковых с именем Маньо-
ля — ботаника, который был вполне чужд данной проблеме.
Мы будем пользоваться более понятной широкому кругу ученых
прежней номенклатурой, основанной на существенных свойствах
высших таксонов.
6.2. Соотношение сети и древа
Иерархия, представимая в форме древа, и комбинативная система,
представимая сетью или таблицей, логически равноправны,
но если на практике реализуется большая часть логически мысли-
мых комбинаций, то удобнее сеть, а если меньшая, то — древо.
Систему организмов в форме сети еще в 1750 г. предлагал Ви-
талиано Донати, а затем и другие [Райков Б. Е., 1952, с. 74], но об-
щепринятой до 1960-х годов оставалась иерархическая система.
В настоящее время от нее отказались бактериологи и, отчасти,
палеоботаники. О последних мы будем говорить позже, а у бак-
териологов причина была именно в почти полной свободе комби-
наций свойств бактерий, почему их удобно классифицировать не по
планам строения, а по наборам физиологических свойств.
Бактерии являются прокариотами (не имеют клеточного ядра),
тогда как у всех прочих организмов (эвкариот) план строения
клетки совсем другой — она имеет ядро, в котором расположены
хромосомы. Одноклеточные эвкариоты принято классифицировать
по строению, иерархически, но число и состав типов здесь то и
дело меняются, таксоны чрезмерно дробятся — вместо обычного
распределения Виллиса (3.5), для которого характерно 40% и ме-
нее вырожденных таксонов (классов, имеющих по одному отряду
и т. д.), здесь мы видим их более половины. Причина этого в почти
141
столь же высокой, как и у бактерий, комбинативности свойств.
Однако границы между царствами в значительной части проходят
среди одноклеточных, поэтому удобно искать макросистему не в
форме древа, а в форме «парка», в котором «рассажены» частные
системы, каждая из которых имеет свою форму, например, форму
древа. В середине этого «парка» должно расположиться «озеро» —
система бактерий, в форме древ вообще не представимая. Пла-
нировка «парка» должна учесть как сходства, так и уровни орга-
низации, что на двумерной поверхности сделать трудно. Ясно,
что надо выбрать минимальный возможный набор сходств и мак-
симально использовать наличные системы групп (каждую из них
мы будем понимать как тип).
Иерархическую систему организмов часто сравнивают с биб-
лиотечным каталогом и с адресной книгой. Действительно, система
организмов для адресации используется, но вряд ли найдется
библиограф, который по мере получения новых книг переписывал
бы карточки старых, тасовал бы карточки в ящиках и переставлял
бы ящики. А ведь биологи-систематики заняты как раз чем-то
подобным. Аналогично, адрес жителя не меняется от того, что
в соседней квартире кто-то поселился. Каталог и адрес — искус-
ственные системы, от которых требуют только удобства поиска,
как от номенклатуры требуют постоянства. Другое дело — описа-
ние библиотеки (из каких фондов сложилась, чем комплектуется
и где в ней какие разделы), план города или макросистема. Каждое
из таких описаний можно во многих отношениях считать естествен-
ной системой, но нельзя требовать от них исчерпывающей пол-
ноты каталога. Каталог иерархичен (шкафы состоят из ящиков,
главы — из параграфов и т. д.) и потому допускает любую степень
детальности, зато крайне ограничен в отображении сходств,
потому что они выражаются здесь только фактом включения объек-
тов в общую группу. Наоборот, план (карта) имеет еще одно сред-
ство сближения — общую границу объектов (в систематике расте-
ний этим успешно пользовался Мейен [1982]), зато детальность
здесь ограничивается единым масштабом и двумерностью.
Не следует думать, что когда-нибудь таксономические трактаты
(сводки или определители) примут форму таблиц или карт: если
пользоваться аналогией с языком, то трактат подобен словарю,
а таблица или карта — грамматике; они взалмодополнительны,
поэтому естественная и искусственная системы друг друга заменить
не могут [Красилов, 1975], и понимал это еще Линней (см. 1.3).
По Линнею, «все растения показывают взаимное сродство
подобно территории на географической карте» [Linne, 1751, § 77,
с. 27]. Замечательно, что такой «географический» принцип вовсе
не ушел в прошлое. Так, в наши дни, когда химические элементы
естественно и наглядно упорядочены периодической системой
(таблицей), все-таки иногда приходится пользоваться картой:
только на рельефной карте элементов удается наглядно охаракте-
ризовать их изотопы по стабильности. Примеры «географических»
приемов в систематике показывают, что общий абрис системы в
142
виде «карты» вполне может сосуществовать с детальной системой
в форме древа или таблицы. Чтобы составить о городе общее впе-
чатление, нужна не пудовая адресная книга, а краткий путеводи-
тель с планом.
Однако иерархический принцип сам по себе обладает для боль-
шинства исследователей безусловной наглядностью и очевидно-
стью, которая воспринимается ими как естественность. (Подробнее
см.: [Корона, 1987, с. 29].) В явном виде это обычно формулируют
в эволюционных терминах (естественная система — это родослов-
ное древо), но, по-моему, дело не в эволюции. Во-первых, как мы
видели в 1.3, иерархия пришла в систематику отнюдь не из эволю-
ционизма, даже наоборот: именно система-древо послужила для
Дарвина аргументом в пользу его учения [Любищев, 1972; Краси-
лов, 1975]. Во-вторых, древо возможно только там, где' у каждого
таксона признается ровно один предок, однако иерархией продол-
жают пользоваться и те, кто признал симбиогенез, т. е. эволюцию
путем объединения предковых организмов в единый организм-
потомок [Воронцов, 1987]. Подлинным стимулом к построению
иерархий является, по-видимому, стремление избежать неодноз-
начности адресации, но оно оборачивается недолговечностью,
а потому и низкой полезностью. Как заметил Н. А. Заренков,
систематики «пытаются преодолеть неопределенность, не зада-
ваясь вопросом о природе неопределенности» [Теория и методо-
логия..., 1983, с. 40]. Если сам объект классификации нечеток
(например, обладает свойствами животного и растения), то обще-
принятой и долговечной может быть только та система, которая
честно определит его место как нечеткое. Нечеткость означает при
этом одновременное отнесение объекта к двум (или более) группам,
т. е. нарушение иерархии.
Положение нечеткого объекта удобно изображать, помещая его
между четко классифицируемыми объектами, например — на гра-
нице двух областей. При этом адрес его может быть приписан одной
из областей произвольно. Если таких объектов много, то сама
граница теряет четкость, и удобнее говорить не о ней, а об общей
периферии двух областей, причем каждая область должна зада-
ваться своим ядром. В 2.3 уже говорилось, что объединять в особый
таксон периферические виды бесперспективно. Напомним мнение,
высказанное в 1860 г. А. С. Фаминцыным: отсутствие четкой гра-
ницы царств растений и животных само по себе не дает оснований
вводить новое царство, поскольку здесь можно, в зависимости от
вкуса, получить сколько угодно царств. В новейшей литературе мы
это и видим.
Если нечеткие признаки принимают непрерывные значения,
то удобной формой изображения системы будет (для двумерного
случая) карта. Если же значения признаков дискретны, то удобна
таблица. Табличный метод не только экономит место, но нагляден
и потому удобен для знакомства с разнообразием. Так, справочник-
определитель «Деревья и кустарники СССР» (1966), рассчитанный
на начинающих, использовал таблицы, являющиеся как бы фраг-
143
ментами архетипической таблицы Мейена (2.2): строка здесь
обозначает вид растений, а столбец — мерой; на пересечении стро-
ки и столбца стоит цифра, означающая значение мерона у данного
вида. Например, у ели обыкновенной в столбце «хвоя» стоит
цифра 3; в перечне форм хвои под этой цифрой читаем: «Одиноч-
ная, торчащая, 4-гранная, жесткая». Если мерой может иметь у
данного вида несколько значений, то в клетке стоит несколько
цифр. К неудобствам такой формы надо отнести невозможность
отразить все нюансы и варианты. Поскольку для наших целей
это несущественно, мы будем карто-табличным методом пользо-
ваться. Тем самым на первое место у нас выйдут пары «таксон —
мерой» и «ядро — периферия», играющие пока в обычной система-
тике малую роль.
6.3. Диатропика клеток
Есть один случай, когда граница обнаруживается точно, без пери-
ферий — это граница между прокариотами и эвкариотами. К про-
кариотам относятся бактерии, цианеи (синезеленые водоросли)
и актиномицеты (лучистые грибки); в последние годы часто всех
прокариот именуют бактериями (различая эубактерии, цианобак-
терии и актинобактерии). В 1968 г. прокариоты получили офи-
циально статус царства, а в 1969 г. Уиттекер дополнил свою эко-
логическую схему понятием надцарства, т. е. чисто цитологическим
разграничением: прокариоты составили у него одно надцарство,
состоящее из единственного царства, а все эвкариоты — другое
надцарство, состоящее из четырех , царств — растения, грибы,
животные и протисты. Уиттекер, в сущности, отказался от фило-
генетического принципа, и его схема, несмотря на обширную
критику, была принята.
Ядерная организация либо есть в клетке целиком, либо ее нет у
нее вовсе. У прокариотной клетки всего одна оболочка, к внутрен-
ней стороне которой прикреплена одною своей точкой кольцевая
двуспиральная молекула ДНК, и самовоспроизведение наслед-
ственной информации сводится к удвоению этого кольца. К началу
деления клетки оно уже удвоено, и оба новых кольца прикреплены
к внутренней стороне клеточной оболочки. В эвкариотной клетке
все устроено гораздо сложнее: внутри клеточной оболочки есть
другая (двойная) оболочка — ядро, и ДНК расположена в
ядре, причем представлена она не одной молекулой, а несколькими
очень сложными конструкциями — хромосомами, где ДНК пере-
плетена с белковыми нитями — гистонами. Каждая хромосома до
начала деления клетки удваивается, т. е. клетка изготовляет вместо
нее две точно таких же хромосомы. Это само по себе чудо (ведь
хромосома — не просто молекула, ее надо расплести, скопировать
все петли ДНК, синтезировать новые гистоны и вновь заплести обе
конструкции) и происходит оно внешне очень эффектно: в покоя-
щейся клетке хромосомы не видны, и перед тем, как удваиваться,
они как бы заново рождаются внутри ядра; но затем начинают сме-
нять друг друга еще более эффектные картины.
144
Ядерная оболочка растворяется, но хромосомы, оказавшись
на свободе, не бегут кто куда, а как бы выстраиваются в две шерен-
ги, причем пару соседок образуют всегда две сестры, происшедшие
от одной хромосомы. Затем шеренги начинают четко расходиться к
противоположным полюсам клетки, так что сестры навсегда поки-
дают друг друга. Управляет этим движением, как в театре марионе-
ток, пара пучков тончайших ниточек (точнее — микротрубочек),
сходящихся к полюсам,— веретено деления. Когда хромосомы
покинут середину клетки, она начинает сужаться, ее как бы стяги-
вает тугой пояс (это, впрочем, происходит здесь так же, как у бак-
терии) , и она делится на две. Наконец, в каждой новой клетке
вокруг хромосом вырастает новая ядерная оболочка, и хромосомы
вновь становятся невидимыми, как бы растворяются. Таков ти-
пичный механизм клеточного деления — митоз.
Кроме ядра и хромосом эвкариотная клетка обязательно имеет
внутренний опорно-двигательный аппарат, тогда как другие ее
органеллы могут и отсутствовать [Серавин, 1986, с. 779]. Тем не
менее, для характеристики эвкариотной организации существенно,
что эти органеллы, когда они есть, всегда несут эвкариотную
специфику: так, окислительное фосфорилирование (основной
источник энергии) всегда локализовано у эвкариот в митохонд-
риях, фотосинтез — в хлоропластах, а эвкариотный жгутик устро-
ен совершенно иначе, чем прокариотный (содержит иные структур-
ные белки и в основании имеет микротрубочки — такие же, какими
образовано митотическое веретено).
Сто лет назад, когда митоз был открыт и детально изучен, уче-
ные были убеждены, что он в общем протекает одинаково во всех
организмах — у животных, у растений и в одноклеточных орга-
низмах, например в инфузориях. Однако в начале XX в. стало
выясняться, что картины митоза, типичные для многоклеточных,
далеко не так обычны среди одноклеточных и что инфузории скорее
стоят здесь особняком.
Прежде всего оказалось, что у многих одноклеточных ядерная
оболочка не растворяется в ходе деления, а делится пополам, как
и сама клетка, причем веретено деления располагается внутри
ядра. Затем были обнаружены одноклеточные (в основном — водо-
росли), у которых митоз организован еще удивительнее: ядро
делится, не растворяясь, но веретено деления странным образом
располагается вне ядра, пронизывая своими нитями ядерную обо-
лочку; или, что уже совсем непонятно, митоз, в котором хромосомы,
казалось, расходятся вообще без помощи веретена. Среди них наш-
лись и такие одноклеточные, у которых и хромосомы как бы не
вполне настоящие — между митозами не растворяются, а видны
всегда. Ими оказались динофлагеллаты, сочетающие растительный
(фототрофия) и животный (фаготрофия) типы питания. Есте-
ственно, возникло желание увидеть в динофлагеллатах промежу-
точную стадию эволюции от прокариот к эвкариотам. Такова дав-
няя традиция — использовать данные о нынешнем разнообразии
для мысленного воссоздания хода эволюции (сравнительный
145
метод -- см. 1.1). Как мы увидим в части II, пользоваться таким
методом надо с большой осторожностью, а сейчас ограничимся
классификационной стороной дела. Состоит она в том, что динофла-
геллаты были объявлены царством, а затем и надцарством (подим-
перией), о чем см.: [Воронцов, 1987].
Слов нет, отличия их велики и касаются фундаментальных
свойств клетки — строения хромосом и митоза. Вопрос в том, мож-
но ли выявить такое ядро таксона динофлагеллят, которое могло бы
быть сопоставлено с другими царствами. В 1965 г. цитолог
Дж. Додж назвал организацию динофлагеллат мезокариотной,
т. е. промежуточной между про- и эвкариотами, но таксономическо-
го анализа за этим термином не стояло. Вскоре же выяснилось, что
своеобразие динофлагеллат отнюдь не примитивно — наоборот,
они во многом сложнее других эвкариот, особенно их хромосомы,
образующие сложные пространственные структуры, скрепленные
не столько белками, сколько структурными РНК; прокариотные же
черты динофлагеллат носят характер отдельных признаков, а не
комплексов [Fritz, Triemer, 1983; Bhaud, Soyer-Gobillard, 1986;
Gavrila, Michaesku, 1986; Costas, Goyanes, 1987]. Как уже отме-
чалось [Чайковский, 19776], динофлагеллаты наиболее характер-
ны не примитивностью, а разнообразием — в их пределах можно
встретить едва ли не любое уклонение от обычной организации
клетки. Но на этом свойстве ядро таксона построить нельзя, ядро
строится всегда на сходствах, а не на различиях. Различия же
позволяют говорить здесь не об особой организации клетки,
а о концентрации изменчивости — тенденции, которую мы отмеча-
ли (см. гл. 4): если у кукушек реализованы почти все формы репро-
дуктивного поведения птиц, а у привиденьевых — почти все формы
мимикрии насекомых, то у динофлагеллат — почти все формы
митоза и хромосом. Единая тенденция говорит в пользу выделения
динофлагеллат в таксон, но определить этот таксон можно только
на основе того, что у них общего, что образует ядро таксона. А это
общее соответствует рангу не выше типа (см. 6.1). Если же пы-
таться дать «диагноз подимперии мезокариот» [Воронцов, 1987,
с. 1770], то он окажется просто диагнозом эвкариот плюс несколько
признаков, свойственных некоторым (далеко не всем) динофлагел-
латам.
Кстати, динофлагеллаты довольно молоды (известны лишь с
юрского периода), так что и с филогенетической позиции являются
только ответвлением. Не являют они и той эко-физиологической
особенности, какую обычно приписывают царствам. Словом, обо-
собление их в отдельное царство не может быть ни долговечным,
ни полезным для выявления основных черт живого. То же можно
было бы сказать и об остальных эфемерных царствах. Вообще,
стремление возвести свой любимый объект в ранг царства стало
в последние годы досадной модой, лишающей макросистематику
роли ключа к познанию разнообразия (как это уже давно происхо-
дит с систематикой родов — см. 2.3). Иногда на роль царства пре-
тендует такая совокупность видов, какую трудно назвать вообще
146
каким-либо таксоном. Рассмотрим только два примера, чаще дру-
гих мелькающие в литературе,— архебактерии и протисты.
Слово «архебактерии» родилось осенью 1977 г. в обстановке
журнальной шумихи и сразу — для обозначения нового, «первич-
ного» царства. В него были отобраны роды из самых разных в
физиологическом и морфологическом отношении групп бактерий,
сходные по составу клеточных оболочек и особенностям некоторых
структур, ответственных за синтез белка. Разумеется, эти отличия
были сочтены главными и самыми древними, хотя общая морфо-
функциональная организация здесь не отличается от большинства
бактерий [Дуда, 1984]. В отношении общей организации гораздо
больше отличаются микоплазмы: они вообще не имеют клеточной
стенки, их геном — самый короткий из всех изученных организмов
и приближается по размеру к геному крупного вируса, их рибосом-
ные РНК столь же уникальны, как и у архебактерий, а многие
биохимические черты сближают их с эвкариотами; наконец, неко-
торые микоплазмы имеют нечто вроде внутреннего скелета, что
прокариотам несвойственно [Борхсениус, Чернова, 1987а, б]. Есте-
ственно, что высказывались гипотезы об эволюционной первично-
сти микоплазм, и с помощью журналистов нетрудно бы й их
объявить царством.
Можно, с неменьшим правом, выделить «первичное царство»
совсем иначе, положив, что самое важное свойство -- наличие
у бактерии фотосинтеза [Lake е. а., 1985]. В такое царство Photocy-
ta вошли бы и некоторые архебактерии. Кстати, руководство Берги
[Bergey, 1984] называет архебактерии во вводной статье типом,
но не указывает его состава, а конкретные их семейства описаны
порознь, среди других групп. Одна из архебактерий, Thermoplasma,
попала в труппу микоплазм. Строить макросистемы можно на
любой вкус, если не заботиться о единой для всех групп методоло-
гической основе классификации.
Что касается протист (простейших), т. е. одноклеточных эвка-
риот, то основание выделять их в особое царство совсем не то, что
в случае бактерий, и носит чисто отрицательный характер: их счи-
тают царством только потому, что многие из них не проявляют
четко свойств других царств — грибов, растений или животных.
Выявить у протист ядро таксона нельзя, а таксон, собранный из
периферий (в данном случае — периферий царств растений, гри-
бов и животных) всегда ущербен. Однако это царство вот уже вто-
рой век то исчезает в одних системах, то появляется в других (см.,
напр.: [Журавлева, Мягкова, 1987]). Очевидно, что в нем есть
потребность, но нет достаточного основания для его признания.
6.4. Необходимый минимум высших таксонов
Недостаточность двухцарственной системы организмов (растения
и животные) была ясна давно. В 1546 г. Франсуа Рабле применил
термин «зоофиты» для обозначения организмов неопределенной
природы, а Баугин [Bauhin, 1623, с. 368] прямо назвал зоофиты
147
«третьей формой жизни», отличной и от животных, и от растений.
Наоборот, Линней, чтобы не портить красоты своей трехцарствен-
ной системы (камни, растения, животные), отнес зоофиты к низ-
шим животным [Linne, 1766, с. 1069]. Состав группы, сперва очень
широкий (у Баугина среди зоофитов числится даже один моллюск)
неуклонно сужался, так что к 1858 г. (в системе Бронна) в ней
остались только одноклеточные и губки (прочие были отнесены
к более высоко организованным животным). Наконец, в 1860 г.
Джон Хогг, развивая мысль Оуэна, предложил объединить все
низшие организмы в царство Protoctista (Protozoa-]-Protophy ta).
Поизошло это на том знаменитом съезде ученых в Оксфорде, где
впервые спорили об учении Дарвина. Геккель предпочел сузить
понятие низших: он ввел царство Protista, включающее только
одноклеточных [Haeckel, 1866]. (Характерно, что со времен Гекке-
ля системы одноклеточных являются иерархическими лишь номи-
нально, поскольку почти не содержат ветвлений, т. е. напоминают
не древо, а куст. Единственный признак древа — общий корень —
недавно тоже отвергнут в системах, изображаемых «бескорневым
древом» [Lake е. а., 1985].)
Параллельно формировалась тенденция к обособлению грибов.
У Баугина мы находим среди грибов группу «каменные грибы»
(Fungi lapidei), в XVIII в. к отдельным «идам грибов изредка
применялся термин Lithophyta (камнерастения), а в 1783 г. немец-
кий ботаник Наталь Йозеф Неккер предлагал выделить грибы в
особое царство [Ainsworth, 1976]. Однако лишь Уиттекер сочетал
царства грибов (Fungi) и простейших (Protista) в одной системе.
Система Уиттекера оставила без ответа массу вопросов. Почему
одноклеточные и многоклеточные водоросли попали в разные
царства? Куда отнести насекомоядные растения? К какому типу
питания отнести пиноцитоз *, т. е. захват одноклеточным капель-
ки жидкой или слизистой пищи? Уиттекеровы принципы следовало
дополнить какими-то еще, в том или другом смысле естественными.
Многие искали естественную систему в форме симметричного
рисунка.
Систему в форме рисунка предложил в 1792 г. П. Гизеке, один
из учеников Линнея, пытаясь графически изобразить «естествен-
ный метод» классификации цветковых, у Линнея едва намеченный
[Козо-Полянский, 1937]. Гизеке изображал семейства цветковых в
форме соседствующих кругов, причем размер каждого круга
соответствовал числу видов в семействе. Подобным методом поль-
зовались Фрис, Бэр и другие, причем зоологи добавили идею сим-
метрии рисунка (некоторые из них воспроизведены в статье
С. Р. Микулинского [1959].) Против такого увлечения возражал
в 1820 г. старый слепой Ламарк: «...ошибочен тот взгляд, будто
разветвления общего ряда... могут быть представлены различными
кругами, сетями или [точками] — как на географической карте,
так как подобное расположение противоречило бы порядку нара-
* Этот вопрос задал мне в 1984 г. С. В. Чебанов.
148
стающего усложнения» [Ламарк, 1959, с. 428], т. е. прогрессивной
эволюции. Действительно, все подобные системы были статичны,
неэволюционны.
Впоследствии А. А. Гроссгейм [1945] предложил рассматри-
вать систему кругов, аналогичную схеме Гизеке, в качестве кроны
эволюционного древа, причем круги у него упорядочены с помощью
трех концентрических колец, соответствующих трем уровням
эволюционного развития групп. Однако поиски симметричной
системы были безуспешны, пока их вели ботаники и зоологи по-
рознь и пока они искали красивую таблицу (прямоугольную, сово-
купность кругов, концентрических колец, комбинацию эллипсов
и т. д.) для уже известных таксонов примерно одного ранга. Если
система действительно симметрична, то и форма таблицы, и состав
таксонов одного ранга, и самое понятие таксона — все это должно
определяться, исходя из этой симметрии, а не браться извне. Сама
же симметрия должна отражать не нынешний этап эволюции (как
у Гроссгейма), а инварианты, которые сохраняются в ходе истори-
ческого развития. Такие инварианты изобразил своими триадами
Уиттекер, но затем были выявлены и другие инварианты. Важное
развитие схемы Уиттекера предпринял в 1973 г. Г. А. Заварзин.
В центре он поместил собственно бактерии, к которым слева
примкнули прокариоты-продуценты (цианеи, фотобактерии и др.),
а еще левее следуют эвкариоты-продуценты, т. е. растения (рис. 5).
РИС. 5. Схема царств по Уиттекеру — Заварзину
[Заварзин Г. А., 1979], изображенная в симметри-
зованном виде.
С другой стороны к собственно бактериям примыкают прокариоты-
редуценты (актиномицеты, гнилостные бактерии и др.), а к ним —
грибы. Естественным образом при этом собственно бактерии сопо-
ставляются с консументами, хотя и не являются (за исключением
миксобактериальных колоний) фаготрофами. Среди эквариот За-
варзин выделил два инварианта — уровня организации: низшие,
или нетканевые (водоросли, грибы и простейшие животные) и выс-
149
шие, или тканевые. Это — большой шаг вперед, хотя понятие тка-
невого уровня и требует уточнения (см. 6.6).
Заварзинская схема формализовала экологические парал-
лелизмы между прокариотами и эвкариотами. Водоросли, с одной
стороны,— аналоги цианей, а с другой — высших растений; про-
стейшие животные, с одной стороны,— аналоги собственно бакте-
рий, а с другой — высших животных; грибы — аналоги актиноми-
цетов. (Если не считать отсутствия «тканевых» грибов, то схема
симметрична, и это удобно для восприятия.) Царства изображены
в виде «карты», где соседство означает сходство. Тем самым род-
ство отображается столь же наглядно, как и параллелизм. Если ни
одна территория не входит одновременно в две «области», то иерар-
хический принцип не нарушен, и вполне можно сочетать удобства
иерархии с наглядностью карты, что с успехом проделал С. В. Мей-
ен [1982]. Иерархия означает последовательное включение групп
друг в друга (вид не входит в два рода и т. д.), но метаболическое
разнообразие одноклеточных (и близких к ним многоклеточных)
таково, что приходится включать некоторые «территории» сразу в
2—3 «области».
Хотя, как видим, на рис. 5 нет ущербных царств вроде архе-
бактерий, мезокариот или протист, но вопрос о числе царств не
решен: если, например, параллелизм между прокариотами и эвка-
риотами так существен, то почему бы не разделить прокариот тоже
на три царства или хотя бы — подцарства? Чтобы понять структуру
надцарства прокариот, пришлось снова обратиться к их экологии
и физиологии, но обратить внимание не на способ поглощения,
а на еще одну группу инвариантов — характер метаболизма.
6.5. Типы метаболизма и макросистема
В современных руководствах принято классифицировать типы
метаболизма следующим образом: (а) по источнику внешней энер-
гии — фото-и хемотрофия; (б) по источнику внешнего углерода —
неорганический (авто-) и органический (гетеротрофия); (в)
по донору электрона основной окислительно-восстановительной
реакции — неорганический (лито-) и органический (органотро-
фия). Всего эти три пары образуют 23=8 типов питания. У эвка-
риот электроны берутся из того же источника, что и углерод: грубо
говоря, в растениях расщепляется СОг, углерод которого усваи-
вается, а кислород в форме О2 (т. е. отдавший электроны) уходит
в атмосферу; у животных и грибов, наоборот, углерод берется в
форме сахаров, которые в ходе метаболизма окисляются (ли-
шаются соответствующих электронов). Тем самым, у эвкариот
реализуется либо фотолитоавтотрофия (растения), либо хемоорга-
ногетеротрофия, тогда как у прокариот возможны все 8 вариантов
метаболизма [Шлегель, 1987]. Однако по этим типам разделить
прокариоты на группы невозможно, в силу явления миксотроф-
ности — одна и та же бактерия может не только переключаться с
одного типа на другой, но и одновременно осуществлять процессы
150
разных типов — например, усваивать СО2 и органику. У эвкариот
миксотрофность — большая редкость, тогда как у прокариот она
обычна. Именно поэтому невозможно выделить в надцарстве прока-
риот различные царства, подобные уиттекеровым эвкариотным
царствам. Нужен иной тип классификации.
Тройку дихотомий (а), (б), (в) пришлось дополнить еще одной
дихотомией. Основное множество метаболизмов (все те метаболиз-
мы, которые реализуются у эвкариот) характерно, во-первых,
наличием двух типов (растительного и животно-грибного) усвое-
ния энергии и углерода и, во-вторых, общим для всех эвкариот
потоком минеральных (прежде всего — растворенных в воде со-
лей) и органических веществ. Все эти типы усвоения и потоки
имеются и у прокариот, но в них есть еще 6 других типов усвое-
ния и, сверх того, прокариоты способны включать и другие элемен-
ты (молекулярный азот из воздуха, элементарную серу и т. д.).
Все организмы, способные к другим типам метаболизма, не входя-
щим в основное множество,— идиотрофы (от греческого ’iSiog —
особенный). Они включают что-то из неорганических потоков
веществ в органические и в этом смысле являются продуцентами
по Уиттекеру. Само понятие продуцента, совпадающее в надцар-
стве эвкариот с растениями, получает в прокариотном мире более
широкий смысл, выражаемый формулой:
продуценты=фотолитоавтрофы-|-идиотрофы (3).
Идиотрофы — не таксон в линнеевом смысле, поэтому им нельзя
придавать определенный ранг (например, подцарства или типа).
У них нет ядра таксона, так как они определены в целом только
через отрицания — как обладатели свойств, отсутствующих у
эвкариот. Кроме того, их характерные свойства распределены
среди большей части прокариот: например, фиксируют из воздуха
азот самые разные микробы, в том числе многие цианеи. К идио-
трофам относятся и почти все (но не все!) архебактерии. Надцар-
ство прокариот не допускает разбиения на таксоны равного ранга.
Это, однако, не значит, что идиотрофы нельзя упорядочить.
Наоборт, именно их упорядочение удается сделать прогностичным.
Как показал опыт Менделеева и Вавилова, это — общее свойство
комбинативных систем. В частности, описан случай успешного
поиска бактерии, литотрофно восстанавливающей трехвалентное
железо водородом: зная свойства бактерий, обладающих поодиноч-
ке требуемыми свойствами, удалось верно предсказать свойства
искомой их комбинации и найти ее [Заварзин Г. А., 1979, с. 19].
Экологический принцип сам по себе не позволяет расчленить
царство прокариот на типы, зато он раскрывает суть этого царства
как экологического феномена. Заварзин прав, утверждая, как мы
отмечали выше, эко-физиологическую универсальность прокариот
и зависимость эквариот. Эта универсальность — одна из главных
тенденций прокариот, и с этой точки зрения они — более чем цар-
ство не только в цитологическом, но и в экологическом смысле.
Универсальность не надо понимать как экологическую самостоя-
151
тельность или, тем более, как экологическое господство; наоборот,
для прокариот в настоящее время характерна экологическая мар-
гинальность, т. е. занятие всевозможных граничных зон.
Сказанное порождает новый вопрос: в чем состоит экологиче-
ское значение другого надцарства — эвкариот? Обладают ли они
своей единой ведущей тенденцией? Здесь вновь существенны типы
метаболизма.
Метаболизм принято условно делить на два класса процессов:
анаболизм, при котором характерно удлинение ковалентных цепо-
чек молекул и поглощение энергии; и катаболизм, при котором
цепочки укорачиваются, а энергия выделяется. Оба класса тесно
сопряжены в любом организме, но в продуцентах, естественно,
преобладает анаболизм. Казалось бы, в других организмах должен
безусловно преобладать катаболизм (ведь они живут за счет раз-
рушения молекул, синтезированных продуцентами), но здесь дело
далеко не так просто. Осмотрофы, которыми является большинство
редуцентов, действительно разрушают гораздо больше органики,
чем создают,— это ясно уже из того, что они выделяют лизирующие
ферменты во внешнюю среду и поглощают мелкие молекулы,
образовавшиеся при лизисе крупных; эффективность такого про-
цесса всегда низка: органики разрушается гораздо больше, чем
используется для нужд собственного синтеза. Потому они и ре-
дуценты.
Фаготрофы питаются более эффективно: поглощенная пища
лизируется внутри организма, они могут, взяв, к примеру, 3 г пи-
щи, один из них лизировать и с помощью полученной при этом
энергии включить 2 г в состав своего тела. В отношении судьбы
всей взятой пищи это редукция (взято 3 г, а усвоено только 2),
но в отношении эффективности самого метаболизма животного
это — превосходство анаболической фазы над катаболической
вдвое. Аршавский [1982, 1986] считает превышение анаболизма
над катаболизмом основным свойством организмов, которое позво-
ляет им расти и активно действовать. Однако степень активности в
разных царствах различна.
И тут оказывается, что роль эвкариот в биосфере достаточно
очевидна — экологическое разнообразие. У них гораздо больше
возможностей как в отношении вовлечения биомассы (99% массы
живого вещества — высшие растения) и подвижности (основная
часть целенаправленно перемещающейся биомассы — животные),
так и в отношении создания различных структур (более 99% ви-
дов — эвкариоты). По-видимому, это можно связать с компартмен-
тализацией (пространственным обособлением разных механизмов в
различных органеллах), радикально повышающей эффективность
анаболических процессов. У продуцентов и фаготрофов это,
вероятно, оказалось определяющим. Наоборот, грибы не дали,
несмотря на компартментализацию, ни тканевых структур, подоб-
ных растительным (этот факт отражен на рис. 5, где снизу нет
внешнего «лепестка»), ни подвижности, подобной животной
(у высших грибов подвижна лишь цитоплазма). Это естественно
152
связать с низкой эффективностью осмотрофии: основная часть
биосинтеза обеспечивает не собственный анаболизм (рост, услож-
нение), а пул ферментов, выделяемых во внешнюю среду. Преиму-
щества эвкариотности выразились у грибов прежде всего в раз-
нообразии (100 тыс. видов, тогда как актиномицетов — менее 2 тыс.
видов), особенно — высших форм. Разнообразие растений еще
выше — около 300 тыс. видов, а животных прямо-таки огромно —
около 2 млн. Именно эвкариоты определяют ныне экологический
облик планеты, оттеснив большинство прокариот в маргинальные
зоны.
Зачем вообще нужны, с экологической точки зрения, животные?
Ведь круговороты веществ могут быть замкнуты, казалось бы, и без
них. Более того, неясна роль и других высших: «Бактерии суще-
ствуют... безотносительно к приспособленности других организмов,
и это доказано всей эпохой протерозоя, когда бактерии были един-
ственными обитателями Земли» [Заварзин Г. А., 1979, с. 19]. Од-
нако не будем забывать, что ныне большинство бактерий живет
за счет высших и должно погибнуть в случае их гибели. Кроме
синтеза и распада биосфере нужны многие другие свойства —
устойчивость, мобильность, способность к развитию. Сам же Завар-
зин согласен, что нынешние бактерии доминируют лишь в мар-
гинальных условиях, но, в основном, заменены высшими в «нор-
мальных условиях». Связи в нынешней биосфере характерны
своей специализацией, одна из черт которой состоит в том, что
наиболее сложные в своем поведении организмы не играют опреде-
ляющей роли ни в создании, ни в разрушении органики. Они-то и
называются животными.
Вот пример конкретной экологической роли животных: в био-
сфере нет круговорота фосфора, подобного круговороту углерода
или серы, соли фосфора необратимо вымываются в океан, и только
1% его возвращают обратно на сушу наземные животные,
поедающие продукты моря: в основном — птицы и человек. Напом-
ним, что, согласно Линнею, животные «хищением сдерживают
растительное население, чем и поддерживается общая соразмер-
ность» [Linne, 1766, с. 12].
6.6. Симметрийная основа макросистемы
Сложность организации, учтенная Г. А. Заварзиным в форме трех
уровней (рис. 5), еще явно недостаточна. Ясно ведь, что плесень
чем-то существенно ниже шляпочного гриба, а червь — ниже насе-
комого. Неясно пока и другое — как обходиться с промежуточны-
ми группами. Однако главная трудность — с параллелизмами: ведь
существенные сходства наблюдаются между любыми группами
организмов и могут запутать любую систему. Если прежде их
попросту игнорировали, то теперь, когда схема (рис. 5) построена
на межцарственных экологических параллелях, следует учесть
хотя бы другие параллели того же ранга, т. е. межцарственные.
Например, во всех царствах есть клетки со жгутиками, а есть и без
жгутиков.
153
Для наглядности и цельности надо иметь какой-то четкий прин- ;
цип расположения, подобный столбцам и строкам менделеевской
таблицы. Обычный иерархический принцип, применяемый в си-
стематике, выявляет, в качестве высших таксонов, только крупные
группы, а не какие-то уровни организации (например, как губки,
так и хордовые образуют тип). Группировка типов не входит в
задачи традиционной систематики: «...подразделения, стоящие над
типом, не следует рассматривать как систематические (таксоно-
мические) категории. Высшей таксономической категорией оста-
ется тип... Группировки, стоящие выше типа, позволяют подчерк-
нуть, с одной стороны, уровень морфофизиологической диф-
ференцировки, а с другой — родственные (филогенетические) свя-
зи между типами» [Догель, 1981, с. 19]. Для животных эти уровни
таковы: одноклеточные (Protozoa), пластинчатые (Placozoa), про-
стые тканевые, т. е. губки (Parazoa — Spongia), двуслойные,
т. е. лучистые (Radiata), простые трехслойные, т. е. бесполостные
(Scolecida), первичнополостные (Pseudocoelomata), вторичнопо- ;
лостные (Coelomata). Они общепризнаны, хотя отдельные системы
несколько различаются (ср.: [Догель, 1981] и [Маргелис, 1983]).
Для цельности макросистемы нужно провести подобные уровни
через все царства единым образом.
Приблизительно ту же роль, какую при определении уровня ор-
ганизации животных играет полость (coeloma), у растений играет
побег (cormus); а ту роль, которую у животных и растений играет
ткань (textus), у грибов выполняет плетеная из гиф «ложная
ткань» (plectenchyma) *. Это сопоставление позволило наметить
что-то вроде координатной сетки, дать некоторый принцип, упоря-
дочивающий параллелизмы и типы организации.
«Координатная сетка» должна быть симметричной, но о какой
симметрии можно говорить в систематике? Разнообразие таксонов
очень несимметрично, зато очевидна симметрия баланса биосферы,
существующей за счет почти замкнутых круговоротов вещества.
И хотя именно неполная замкнутость движет ее эволюцию (мы
увидим это в части II), единство макросистемы легче разглядеть,
если считать биосферу сбалансированной. Съедается примерно
столько же, сколько производится, и для каждого вещества сущест-
вует как механизм синтеза, так и механизм разложения. Да, 99%
земной биомассы — в растениях, но ведь 95% их массы — медлен-
но растущая древесина (медленно она потом и гниет), а активная
зеленая масса оборачивается в среднем дважды в год и тем обеспе-
чивает всех редуцентов и консументов пищей. Все комбинации
реализоваться не могут, так как их больше, чем атомов во Вселен-
ной; а поскольку систематику принято строить именно на ком-
бинациях свойств (систематику прокариот, в основном, на комби-
нациях функций, а эвкариот — в основном, на комбинациях струк-
тур), то возникает асимметрия. Следовательно, наша макросистема
будет тем симметричнее, чем меньше будет учитывать комбинато-
* На эту параллель мне указал С. В. Чебанов.
154
рику. Мы увидим, что симметрия еще видна, а крупные таксоны
уже можно расставить, если в макросистеме рассматриваются соче-
тания основных способов питания с основными структурными
принципами (но не с их комбинациями).
Биосистемолог С. В. Петухов [1981], предлагая выявлять в жи-
вой природе общие, но малозаметные симметрии, обратил внима-
ние на круговую симметрию (где осью симметрии служит окруж-
ность). Воспользуемся этим советом на чисто качественном уровне.
Кольцевые макросистемы давно известны, но носят характер лишь
графического приема, не ставящего цели отобразить какой-то
специфический, именно кольцами выражаемый, закон природы
(см., например, «кольцо Кандлера» [Воронцов, 1987]). У нас же
речь пойдет о криволинейной симметрии концентрических колец.
В качестве основной оси (окружности) симметрии естественно
взять границу прокариот с эвкариотами, поскольку это — един-
ственная четкая граница между таксонами, не осложненная пере-
ходными формами. На рис. 6 она изображена жирной линией.
В верхней части этой окружности располагаются жгутиковые одно-
клеточные организмы, обладающие всеми тремя способами пита-
ния, согласно триаде (2) из 6.1. К ним относятся некоторые эвгле-
ны. Обходя эту окружность по часовой стрелке, мы увидим посте-
пенное исчезновение этих свойств, а затем возвращение к ним —
эта объективно существующая цикличность и потребовала для
своего отображения кольцо. Правее эвглен расположатся жгути-
ковые без фотосинтеза (зоофлагеллаты), затем — жгутиковые с
чисто осмотрофным питанием (осмофлагеллаты), еще далее —
осмотрофы без жгутиков; в нижней части окружности — без-
жгутиковые с фотосинтезом, а в левой ее части — снова фотосин-
тетики со жгутиками (фитофлагеллаты), которые в верхней части
соседствуют со жгутиковыми смешанного растительно-животного
питания. Круг замкнулся.
Внутри окружности расположится кольцо одноклеточных про-
кариот. С них, по-видимому, началась эволюция организмов, и это
кольцо будет в нашей системе основным. Снаружи от окружности
поместим кольцо соответствующих одноклеточных эвкариот.
Параллельно с цианеями (прокариотными водорослями) располо-
жатся одноклеточные эвкариотные водоросли, параллельно со
жгутиковыми бактериями (псевдомонадами) — жгутиковые эвка-
риоты (их прежде называли монадами), параллельно с непод-
вижными бактериями-консументами — споровики и т. д. Пло-
щадь внутреннего (прокариотного) кольца меньше, чем внешнего,
и этим удачно отражается факт меньшего морфологического и
видового разнообразия прокариот: среди одноклеточных видов
прокариотные составляют едва 1/10.
Проведем внутри от кольца одноклеточных прокариот кольцо
поменьше и расположим на нем многоклеточные формы прока-
риот,— видоспецифические скопления клеток: колониальные,
мицелиальные, нитчатые, псевдоплазмодии и т. д. Всем им най-
дутся аналогичные эвкариотные формы, которые расположатся
155
ПРЕОБЛАДАЕНИЕ
I' ЖГУТИКОВЫХ
ФОРМ
ФАГОТРОФЫ
рвотные
моллюски
С ЗЕЛЕНЫМИ
СИМБИОНТАМИ
ЗООФИТЫ
б V
моллюски с
ПРЕОБЛАДАНИЕ
БЕЗЖГУТИКОВЫХ
Актчкоми-
иеты
ц^ТЧАТЫё
Ннфузори
е > IS
•X S ’Л
< чЛ
5 । t
*
ХЕМОТРОФНЫМИ
СИМБИОНТАМИ
РИС. 6. Логическая схема диатропической четырехцарственной макросистемы
организмов. Исходный уровень организации: одноклеточные прокариоты (1п).
Уровни интеграции: одноклеточные эвкариоты (1э); равноклеточные (2); разнокле-
точные, т. е. тканевые (3); разнотканевые, т. е. тканеорганные (4); централизован-
ные (5).
Таксоны обычной систематики написаны с большой буквы (Животные и т. д.),
прочие группы — заглавными буквами (ЗООФИТЫ и т. д.), а прямыми заглавными
буквами (НИТЧАТЫЕ и т. д.) обозначены уровни организации. Некоторые груп-
пы — Водоросли (Algae), Шлнпочные грибы (Boleti-Pileati) и др,— не являются
таксонами нынешних филогенетических систем, но в диатропической макросистеме
их естественно считать таксонами (подцарствами, надтипами, типами). Схему с ла-
тинскими названиями и более детальное изображение царства животных см.: [Чай-
ковский, 1986].
ВТОРЙЧНОПОЛОСТНЬЧ
. черви,
^мгевыв, Д»Ус?
плаэЙЗ
^гутиконой^
М О З'о и
*в Ч)
Ьт
as
IlX
ЦВЕТКОВЫЕ
ПАРАЗИТЫ
ХИЩНЫЕ
ГРИБЫ
*Е
Е а
вне кольца одноклеточных эвкариот и образуют второе эвкариотное
кольцо; там же следует поместить и типы организации, несвой-
ственные прокариотам: многоядерные структуры без клеточных
перегородок (ценоциты, плазмодии, эвкариотный мицелий). Таков
уровень простой многоклеточности.
156
Бывают, в качестве исключения из правила, и более сложные
прокариоты: цианеи в виде сантиметровых кустиков и плодовые
тела миксобактерий в виде ярких милиметровых грибочков — они
естественно займут небольшой кружок в центре схемы. Их эколо-
гическое значение невелико. Компактный слой или полость клеток,
специализированных для общей функции, называют тканью. Мож-
но сказать, что упомядутые исключения являют собой тканевый
уровень прокариот: функция питания исполняется здесь одной
компактной полостью клеток, а размножение — другой; у цианей
бывают и азотфиксирующие «ткани», отдельную «ткань» может
представлять и оболочка органа размножения. Зато у эвкариот
соответствующий (тканевый) уровень очень обширен: это все то,
что мы в обиходе называем водорослями и грибами, а также —
губки, кишечнополостные (медузы, гидры и т. п.) и низшие черви.
Этим симметрия между прокариотами и эвкариотами исчерпы-
вается, так как более сложных форм у прокариот нет. Нет их,
кстати, и у грибов.
Симметрия относительно границы надцарств означает, что
прокариотные -типы клеточной организации повторены низшими
эвкариотами: и тут и там мы видим клетки круглые, вытянутые
и спиральные, с жесткой стенкой и без, жгутиковые, скользящие и
неподвижные; повторены и прокариотные типы многоклеточности:
и тут и там мы видим нитчатые, мицелиальные (ветвящиеся),
кустистые, слизевые, корочковые и т. п. Однако ткани — то, что,
в сущности, создает все богатство многоклеточных,— у прокариот
почти отсутствуют. Тем самым, эвкариотная организация клетки
обеспечила реверсию разнообразия: если у прокариот разнообразие
падает по мере усложнения организации, то у эвкариот — растет.
Четвертым уровнем (кольцом) эвкариот будет тканеорганный:
здесь орган строится из различных тканей. Ствол дерева строится
из древесины, луба и коры; пищеварительный тракт даже у коль-
чатого червя включает железистые, соединительные и мускульные
ткани и т. д. У высших растений есть много различных разнотка-
невых органов, и в этом они проявляют параллелизм с животными
средней сложности (приблизительно — до уровня кольчатых чер-
вей), образуя тем самым четыре уровня эвкариотной интеграции.
Приблизительно те же уровни выявила недавно М. Шультер (ана-
лиз см.: [Журавлева, Мягкова, 1987, с. 154]). Этим организация
растений и завершилась, тогда как у животных есть еще и пятый
уровень, наиболее богатый видами — централизованный.
Интеграция никогда не доходит у растений до централизации:
из корня часто может вырасти новый стебель, отрезанный стебель
или ветка часто может давать новый корень, а разведение культур-
ных пород прививкой общеизвестно. В то же время животные выше
червей не переносят даже разрезания пополам — настолько их
органы интегрированы в единую систему. Поэтому среди них
естественно выделить пятый уровень, куда войдут все позвоночные,
членистоногие и моллюски, имеющие головной мозг и сердце.
Процедура образования колец тоже обладает своеобразной сим-
157
метрией: каждый уровень, считая от одноклеточных прокариот,
образует следующий путем интеграции: эвкариотная клетка есть
объединение прокариотных механизмов [Чайковский, 19776],
многоклеточный организм или ткань — объединение клеток и т. д.
На эту симметрию накладывается третья, вполне наглядная,
симметрия — уиттекерова триада, которая здесь лишь немного
изменена. Уиттекер сопоставлял экологическую триаду (1) и фи-
зиологическую, близкую с ней по видовому составу, триаду (2),
но проще оказалось взять за основу такую, синтетическую, триаду:
продуценты — осмотрофы — фаготрофы (4).
Триада (4) образована путем двух разнокачественных диад
и потому делит систему колец не просто на три сектора, а еще и на
три пограничные области, в которых нашли себе место все те формы
«неясного систематического положения», из-за которых возникло
добавочное царство протист. Выяснилось, что никакого царства
здесь нет, так как нет соответствующего ядра таксона.
6.7. Царства и межцарства. Логика построения
Вглядевшись в рис. 6, мы видим, что всю середину его занял стран-
ный «росток» из «стебля» и двух «листьев». Это — все те орга-
низмы, которые питаются каким-либо нестандартным образом: или
идиотрофы, или организмы, сочетающие разные способы питания
триады (4). Для «ростка» уместно название Myxobiota, отражаю-
щее факт смешанного (животно-растительного, фото-хемотрофного
и т. п.) или промежуточного (осмо-продуцентного) типа питания.
Миксобиоты — не таксон (как и входящие в них идиотрофы и как
пересекающие их фототрофы), но именно их надо мысленно изъять
из системы, чтобы увидеть эвкариотные царства, которые есте-
ственно задаются триадой (2), определяющей растения, грибы и
животных. Все организмы живут на органике, поставляемой авто-
трофами, частично разлагают ее. чтобы извлечь свободную энергию,
однако видны следующие три тенденции.
Грибы более проявляют себя (в глобальном масштабе) как
редуценты — они вместе с гнилостными (и другими подобными)
бактериями образуют так называемое детритное звено (разложение
мертвечины и экскрементов на неорганические компоненты); в то
же время животные более заметны создаваемым ими морфологи-
ческим разнообразием, тогда как выделяемая ими неорганика
(разлагая углеводы, они выделяют воду и углекислый газ) не столь
существенна. Фактически грибы выявляются тоже морфологиче-
ски — по ветвящемуся мицелию, внутри которого свободно перете-
кает цитоплазма (клеточных перегородок гораздо меньше, чем кле-
точных ядер). Ядро царства грибов образуют мицелиальные фор-
мы — это чисто морфологический, а не экологический критерий.
Наоборот, ядро царства растений определяется экологически — как
автотрофные эвкариоты.
Других эвкариотных царств при таком подходе не оказалось,
поскольку налицо лишь три ядра (фотосинтез, захват пищи и ми-
158
целий), зато периферии трех царств перекрываются. Это перекры-
тие присуще природе как таковой, а вовсе не связано с ее недоста-
точной изученностью. Нельзя, как еще часто делают, писать про эти
промежуточные формы: «Их положение в системе неясно» —
в действительности их промежуточный статус абсолютно ясен.
Каждое царство (Regnum) задается своим ядром, тогда как пери-
ферии их, пересекаясь, образуют межцарства. Каждое межцарство
(Interregnum) образуется сочетанием свойств двух царств, а не
собственным ядром. Каждое из трех межцарств — зоофиты,
зоомицеты и фитомицеты — имеет максимум разнообразия среди
одноклеточных (нет реверсии разнообразия, что резко отличает
межцарство от эвкариотного царства), но в каждом есть и высоко
развитые формы. В подцарстве зоофитов это — моллюски с недо-
развитым пищеварительным трактом, ставшие плантациями для
живущих в них одноклеточных водорослей, и хищные цветковые;
в подцарстве зоомицетов это — плеченогие, перешедшие к осмотро-
фии, миксомицеты и хищные грибы; в подцарстве фитомицетов —
паразитические цветковые, а также лишайники. Прокариоты об-
разуют одно царство и не образуют межцарств с эвкариотами,
поскольку параллелизмы их с эвкариотами не носят характера
переходных форм.
Итак, диатропическая система — четырехцарственная, но не
такая, как у Уиттекера: у него протисты были объединены с про-
кариотами (позже — выделены в пятое царство), а здесь — рас-
пределены вдоль всей границы прокариот с эвкариотами в форме
трех межцарств, не являющихся таксонами. Среди прокариот не
является таксоном группа идиотрофов, но ее статус не тот, что у
межцарств: идиотрофы тоже не образуют собственного ядра
(типичных форм), но не является и периферией каких-либо так-
сонов, а составляют, как и все миксобиоты, совокупность пери-
ферий.
Каждое межцарство входит сразу в два царства и тем явно
нарушает линнеев принцип иерархии. Это непривычно, не очень
удобно при адресации и поисках по данному адресу (для этих целей
лучше иерархическая систематика), но таков сам материал. Прихо-
дится отделить задачу адресации от описания системы как целого.
Зато появляются новые удобства, о которых раньше система-
тик и не мечтал: живое предстает как целое, в частности — видны
параллели развития, главные тенденции и симметрия. Указание
места группы в системе — расстояния (от центра) и угла (от вер-
тикали) — определяет многие свойства группы. Именно такую
систему Любищев называл естественной. Положение каждой
группы в этой системе не зависит от того, что известно о других
организмах, и это очень существенно: новые сведения об организ-
мах здесь, как и следует, только детализируют схему — позволяют
заполнить пустые места и провести новые параллели, но не менять
саму систему.
На детальном рисунке [Чайковский, 1986, с. 30] видно: некото-
рые традиционные типы оказались по нескольку на одном и том
159
же месте (например, некоторые водоросли), и естествен вопрос об
их слиянии. Нам, однако, важнее отметить методическое обстоя-
тельство: каждое царство описывается не только общими свойства-
ми, обычно включаемыми в диагноз, но и тенденциями организации
разнообразия в пределах царства, что видно из следующего описа-
ния макросистемы, уточненного по недавней работе [Воронцов,
1987].
I. 1) Regnum Procaryota (бактерии). Царство определяется ци-
тологически: нет клеточного ядра, а основной геном представлен
единственной молекулой ДНК, замкнутой в кольцо. Признаки
комбинируются почти свободно, поэтому иерархическая система-
тика неэффективна. Отсутствие переходных форм к ядерной орга-
низации клетки и эко-физиологическая универсальность позво-
ляют считать прокариоты надцарством. Разнообразие видов падает
с ростом числа клеток в организме, так что самые примитивные
«ткани» наблюдаются лишь как исключение. Характерна экологи-
ческая маргинальность.
II. Остальные организмы образуют надцарство Eucaryota, где
клетки содержат ядра. Каждое ядро содержит несколько хромо-
сом. Удвоение ядра и каждой хромосомы в нем происходит путем
митоза. Ряд функций эвкариотная клетка выполняет, в отличие от
прокариотной, посредством особых органелл, из которых важней-
шие — митохондрии, осуществляющие окислительное фосфорили-
рование. Митохондрии (а у растений еще и хлоропласты) содер-
жат собственную ДНК, сходную отчасти с прокариотной, но корот-
кую и кодирующую лишь фрагменты функциональных структур.
Жгутики, если имеются, являют значительную структурную общ-
ность, но совершенно отличны от прокариотных. Надцарство де-
лится на три царства.
2) Regnum Fungi (грибы) — эвкариотные осмотрофы. Клеточ-
ная оболочка из хитина. Ядро царства образуют безжгутиковые
формы, т. е. неподвижность характерна грибам более, чем любому
царству. Для высших характерно мицелиальное строение (с актив-
ным перетеканием цитоплазмы), для низших — видоспецифиче-
ская форма колоний (у растений это свойство редко). Многоклеточ-
ных зародышей не образуют. Характерны различные сочетания
гаплоидности, диплоидности и полиплоидности (за счет неиден-
тичности ядер) и симметрия плодовых тел.
3) Regnum Animalia (животные) — эвкариотные фаготрофы
(голозои). Клетки не имеют наружной оболочки. Характерна под-
вижность на всех стадиях, диплоидная раздельнополость и заро-
дышевое развитие многоклеточных. Характерна дифференциация
органов или органелл (что связано с превышением синтеза над
деструкцией и с подвижностью), редукция бесполого размножения,
регенерации и чередования поколений. У высших — билатераль-
ная симметрия и централизация.
4) Regnum Plantae (растения) — эвкариотные фототрофы.
Клеточная оболочка — из целлюлозы или пектина, иногда —
с кремнеземом. Фотосинтез происходит в хлоропластах. Характер-
160
ны: чередование поколений, диплоидная слитнополость спорофи-
тов и способность к бесполому размножению. Высшие формы (Ет-
bryophyta) образуют многоклеточные зародыши. Потенциальная
возможность развития организма из любой клетки связана с более
высокой комбинативностью свойств (меньшей долей запретов):
такие параллелизмы, как сходный характер расчленения листа
(или таллома), пронизывают все царство. Характерна криволиней-
ная симметрия тела и органов.
Остается много форм, не подпадающих под данные описания.
Почти все они легко описываются, если ввести понятие межцар-
ства (Interregnum). Именно:
(a) Interregnum Zoophyta — эвкариоты со смешанным фото-
трофно-голозойным питанием: жгутиковые (Phytoflagellata),
фототрофные амебы и плазмодии; животные, постоянно связанные
с фототрофными симбионтами. Завершают межцарство хищные
цветковые растения (насекомоядные и грибоядные).
(б) Interregnum Mycozoa (Mycetozoa) — эвкариоты со смешан-
ным осмотрофно-голозойным питанием, включая пиноцитоз:
некоторые из амеб, динофлагеллят, эвглен и т. п. (при наличии
фотосинтеза они входят сразу в два межцарства); плазмодиевые
и т. п. формы (миксомицеты и др.), чередующие осмотрофное и
фаготрофное питание. Завершают межцарство хищные грибы
(Zoopagales) и те плеченогие, которые перешли к осмотрофии.
(в) Interregnum Mycophyta — эвкариоты со смешанным осмо-
трофно-фототрофным питанием. Наиболее известны здесь лишай-
ники; к этому же межцарству относятся все фототрофные орга-
низмы, которые или не могут развиться без поступления извне
органических веществ или способны развиваться в темноте (многие
эвглены), и растения, лишившиеся фотосинтеза (Leucophyta).
Кроме того, к микофитам следует отнести фауну гидротерм —
недавно обнаруженных подводных экосистем, населенных хемо-
автотрофными прокариотами и живущими в симбиозе с ними
животными, лишенными пищеварительной системы. Завершают
межцарство цветковые-паразиты и моллюски гидротерм.
Три межцарства совместно с прокариотной группой Idiotropha
образуют эко-физиологическую группу Myxobiota, компактно за-
полняющую середину схемы. Вершины межцарств (высшие мик-
собиоты) проявляют сходство: тканеорганные животные перешли
к осмотрофии и симбиозу. Это наводит на мысль, что предложенная
плоская схема — лишь развертка на плоскость схемы с более
сложной топологией.
У миксобиот нет: реверсии разнообразия, ядер таксонов, четких
границ с другими (легче классифицируемыми) группами. Для
миксобиот обычны: малое разнообразие видов, большое разно-
образие и комбинативность цитологических структур, экологиче-
ская маргинальность (например, лишайники на голых камнях).
Этот квазидиагноз построен на одних тенденциях, что полезно бу-
дет иметь в виду при будущем методологическом анализе проблем
систематики.
6 Ю. В. Чайковский
Часть II. ЭВОЛЮЦИЯ КАК ПРЕОБРАЗОВАНИЕ
РАЗНООБРАЗИЯ
Для наблюдателя эволюция, особенно биологическая, предстает
как изменение разнообразия в течение исторического времени.
Несомненно, что разнообразие организмов в ходе эволюции жизни
росло: появление новых форм видно из данных палеонтологии,
а в первичной (архейской) жизни, по всей видимости, отсут-
ствовали основные носители нынешнего разнообразия — эвкарио-
ты. О первичном разнообразии живого мы почти ничего не знаем,
кроме того, что жизнь сразу предстает нам как сбалансированный
геохимический процесс, а это значит, что первые организмы (их
обычно представляют как очень простых бактерий) были уже
достаточно разнообразны — выполняли все • многочисленные
функции продуцентов и редуцентов (возвращавших в геохимиче-
ское лоно почти столько же, сколько брали из него продуценты).
Как удачно выразился палеонтолог-мыслитель Пьер Тейяр де
Шарден, «едва народившись... жизнь уже кишит» [Тейяр, 1987,
с. 82]. Тем самым, теория абиогенеза (происхождения жизни)
должна быть, в частности, теорией происхождения разнообразия
(но не одного первичного организма, как иногда пишут), а теория
биологической эволюции — теорией преобразования разнообразия.
Казалось бы, это самоочевидно, однако до недавнего времени эво-
люционисты предпочитали обсуждать возникновение и преобразо-
вание отдельных качеств, свойств и признаков, но не разнообразий.
Наблюдаемая сейчас переориентация эволюционной мысли
вряд ли может быть успешна без тщательного методологического
анализа, который и является целью дальнейших глав. При этом
конкретная эволюционная проблематика излагается постольку,
поскольку она необходима для выявления статуса и оснований
эволюционной диатропики.
Глава 7. ЗАКОНОМЕРНОСТИ БИОЛОГИЧЕСКОЙ
ЭВОЛЮЦИИ
7.1. Эволюция как физиологический процесс
Пожалуй, главным достижением эволюционной мысли последних
30 лет явилось понимание того, что эволюция — процесс преобразо-
вания систем, а наблюдаемый поток меняющихся признаков —
162
лишь эпифеномен (сопутствующее явление) этого процесса. Когда
в качестве системы выступает организм, то можно говорить о
морфологическом и физиологическом преобразовании. Если первое
удобно описывать как преобразование планов строения (особенно
см.: [Беклемишев. 1964; Корона, 1987]), то для второго этот язык
неудобен. Хотя Мейен [1978] и полагал, что физиологическое
разнообразие можно описать на том же архетипическом языке,
что и морфологическое, но приходится согласиться с Аршавским,
что при отсутствии собственного понятийного аппарата «на долю
эволюционной физиологии выпадает задача лишь регистрировать
эволюционные преобразования, но не раскрывать природу их
возникновения» [Аршавский, 1985, с. 165]. Понимание эволюции
как физиологического процесса было несомненным у Эразма Дар-
вина (1790-е годы), достаточно заметно у Ламарка и Этьена Жоф-
фруа Сент-Илера, но затем уступило место статистическому пони-
манию ее как процесса смены признаков. Целостность стали
описывать просто как «корреляцию признаков», в чем Дарвин
следовал за Кювье. Только в наши дни, когда статистическое миро-
воззрение стало уступать системному, на первый план понемногу
выходят собственно физиологические законы эволюции. Физиоло-
гия (как и этология) делает акцент на роль собственной активности
организма в эволюционном процессе, подчеркивая, что «жить —
значит реагировать, а отнюдь — не быть жертвой» [Grasse, 1973,
с. 351].
Исходным понятием является для Аршавского метаболиче-
ская спираль: «... Каждая очередная двигательная активность...
является фактором индукции избыточного анаболизма ’... что и
обусловливает спиралеобразный перевод системы на новый струк-
турноупорядоченный негэнтропийный уровень организации, уве-
личение степени неравновесности ее и позволяет осуществиться
процессам роста и развития», причем данный принцип «представ-
лен не только в клетках животных, но и у одноклеточных и расти-
тельных организмов» [Аршавский, 1985, с. 166]. С этой позиции
эволюция характеризуется степенью превышения анаболизма над
катаболизмом (усвоения над разрушением): там, где превышение
велико, мы наблюдаем эврибионтностъ (приспособленность к ши-
рокому спектру сред) и морфофизиологический прогресс по Север-
цову [там же, с. 167].
Новый язык — новый взгляд на вещи. Прежде эволюционные
схемы были привилегией морфологов и популяционных генетиков.
Они всегда признавали необходимость функционального взгляда
на эволюцию, но попытки их рассуждать за экологов и физиологов
часто порождали фантастические схемы, подчас заимствованные из
памфлетов. (Так, в экономическом памфлете Томаса Мальтуса
«Очерк о населении» (1798 г.) было введено понятие борьбы за
существование, перешедшее отсюда в биологию без специального
1 Об избыточном анаболизме мы говорили в 6.5. Добавим, что внутри эвкариот-
ной клетки «все движется» [Меклер, 19806, с. 462].
6*
163
анализа и усвоенное Дарвином [Schwartz, 1974].) В этих схемах
экология сводилась к «борьбе за существование», а физиология —
к всевозможной «оптимизации» (считалось, что организм стре-
мится добыть максимум пищи, оставить максимум потомков, жить
в наиболее комфортных условиях, с минимальной активностью
и т. д.). Теперь, когда собственно эволюционные физиологические
понятия даны, вырисовывается совсем другая схема: комфорт,
инертность и сытость в равной мере гибельны с позиций эво-
люционной перспективы, стремление к неограниченному размно-
жению наблюдается лишь как патология, а потому борьба вовсе
не является основным фактором взаимодействия (см. 5.4). Основ-
ным физиологическим фактором эволюции выступает дискомфорт,
как он описан в главе 4. Существенно, что организмы подвергаются
патологическому дискомфорту в тем большей степени, чем менее
они приспособлены к своим условиям обитания, и в этом смысле
прав был Э. С. Бауэр, писавший в 1935 г., что материал для эволю-
ции поставляют не победители (как у Дарвина), а побежденные.
В 1947 г. М. Е. Лобашев конкретизировал эту точку зрения, пред-
положив, что мутационный процесс носит «паранекротический»
(предсмертный) характер. (Развитие этой идеи см. в работах [Чай-
ковский, 1976; Ватти, Тихомирова, 1987].) Сюда же следует отне-
сти выводы М. Г. Агаева [1978, с. 240] о пусковых механизмах
мутагенеза.
При всей незавершенности такой схемы, она уже сейчас не
только объясняет феномен «экспериментальной эволюции» и со-
ответствующие данные об эволюции природной, но и раскрывает
причины долгого неуспеха попыток доказать или опровергнуть
«наследуемость приобретенных признаков» — до тех пор, пока они
игнорировали физиологию [Аршавский, 1985, с. 168].
Разумеется, эта схема эволюционного акта, которую уместно
назвать спираль-стрессовой, объясняет далеко не всё в эволюции —
хотя бы потому, что эволюция не состоит из одних приспособ-
лений. Более того, описанный физиологический процесс не создает
принципиально нового, он лишь реализует определенные возмож-
ности. заданные архетипом (по Мейену) таксона. Однако для
диатропического аспекта эволюционизма (не касающегося вопроса
о том, откуда берутся архетипы) спираль-стрессовая схема явля-
ется одной из главных. Она обобщает прежние представления,
в которых основными исходными понятиями были цикл и конку-
ренция.
Принято считать, что «на всех уровнях организации (субкле-
точном, клеточном, органном, организменном, популяционном,
экосистемном и планетарном) биологические системы полностью
или частично циклизированы» [Уголев, 1985, с. 458]. В действи-
тельности полной цикличности не существует («если бы она была
реальной в живых системах, самое живое не могло бы иметь места.
Возможность роста и развития была бы абсолютно исключена.
Биологическое время остановилось бы» [Аршавский, 1985,
с. 166]), а все «частичные» циклы можно рассматривать как витки
164
спиралей. Только в одних спиралях соседние витки почти совпа-
дают, и циклическая модель достаточно удобна (например, «круго-
ворот воды в природе», в котором почти вся масса воды действи-
тельно циркулирует, а лишь небольшая часть добавляется за счет
ювенильных вод из недр и безвозвратно уходит при фотолизе воды,
когда водород улетает в космос), в других же, наоборот, спираль
выглядит почти что прямой (например, «круговорот фосфора»,
упомянутый в конце 6.5), и упоминание цикла лишь затемняет
суть дела. Именно незамкнутость биологических «циклов» обеспе-
чивает эволюцию.
Вальтер Циммерман [Zimmermann, 1953] изобразил в виде спи-
рали историческое преобразование рыбы в птицу: каждый цикл
развития особи (половая клетка, зародыш, взрослый организм,
половая клетка), чуть отличаясь от предыдущего цикла, подыма-
ется незаметно в отношении высоты организации (рисунок воспро-
изведен в книге Канаева [1966, с. 151]). Здесь филогенез — след-
ствие преобразования онтогенеза, что прямо противоположно
«биогенетическому закону» Геккеля, где историческое преобра-
зование онтогенеза рассматривалось как следствие филогенеза
(литературу см.: [Чайковский, 1987а]). Аршавский [1985, с. 169]
видит основной механизм эволюции в том, что прижизненные
адаптивные модификации в цепи поколений сдвигаются на более
ранние этапы индивидуального развития, становясь из прижизнен-
ных наследственными (подробнее см.: [Чайковский, 1988г]).
Кроме принципа спиральности, физиологический эволюцио-
низм формулирует ряд других принципов столь же высокой общно-
сти. Некоторые из них конкретизируют принципы, сформулиро-
ванные в предыдущих главах: так, принципы множественности
и полиэссенциальности [Уголев, 1985, с. 459] продолжают тему
мероно-таксономического несоответствия, а принцип компромисса
[там же, с. 460] обсужден нами в главе 5 как принцип адаптивного
компромисса. Зато защищаемая А. М. Уголевым концепция эво-
люции путем комбинации функциональных блоков (ФБ) требует
отдельного анализа.
Издавна говорят о «блочности конструкции как общем принци-
пе» морфологического эволюционизма [Мужчинкин, 1978, с. 781].
Представление об организме как о наборе подсистем (ФБ), в ко-
тором различные ФБ преобразуются в какой-то мере независимо
при явно зависимых изменениях параметров в рамках каждого ФБ.
позволяет сформулировать важные обобщения. Например, «мини-
мальные перестройки могут иметь максимальный эффект: мак-
симальная скорость (плавания тюленей,— Ю. Ч.) достигается у
минимально специализированных форм», поскольку «перестрой-
ка одного блока решает сразу несколько функциональных проблем
(ласт служит и движителем и терморегуляционным органом)»
[там же, с. 780]. Блочность засвидетельствована и в морфологии
высших растений [Корона, 1987, с. 228], и в строении отдельной
клетки [Серавин, 1986, с. 781], она требует теоретического осмы-
сления.
165
Именно концепция относительно независимых ФБ позволяет
говорить о естественном отборе: все рассуждения о преимуществен-
ном выживании носителей каких-то качеств неявно основаны на
допущении, что остальные качества в данной популяции в данное
время совсем или почти не изменяются, т. е. — на идее поочеред-
ности изменений. Но тогда сама блочность выступает, как фунда-
ментальная номотетика, как закон изменчивости. Хотя сами при-
чины блочности не вполне понятны, но ясно, что ФБ реализуют
смену функционирования (4.4): именно тот блок, который наибо-
лее в данный момент уязвим и потому нуждается в изменении,
обычно работает наиболее интенсивно. На макроблоках (органах)
такую нагрузку выполняет смена поведения, на микроблоках —
избирательная экспрессия генов. К роли поочередности в про-
цедуре отбора мы вернемся в конце 7.2.
Суть концепции ФБ в том, что различные функции складыва-
ются из элементарных функций, реализуемых комбинациями
небольшого числа ФБ, причем эволюция однотипных структур
связана с перераспределением ФБ, которые близки или идентичны
у организмов, стоящих на разных эволюционных уровнях; «основ-
ные физиологические функции (рост, дыхание, пищеварение, дви-
жение, размножение и т. д.) в конечном итоге могут быть разложе-
ны на некоторые простые элементы» [Уголев, 1985, с. 212]. В эво-
люции блоки, в том числе ФБ, поразительно стабильны: «В сущно-
сти вся... теория эволюции живых систем в целом базируется на
представлениях о стабильности блоков» [там же, с. 364]. Именно
поэтому эволюция почти целиком комбинаторна. Более того, те ред-
кие эволюционные акты, которые не принято, в силу появления
принципиального новшества, называть комбинаторными, все-таки
обнаруживают комбинаторику — только на более глубинном уров-
не. В 8.3 мы убедимся в этом на примере самого крупного так-
сономического приобретения — становления эвкариотности.
Блочность не мешает организму действовать и эволюциониро-
вать как целому. Указать на то, что не обладавшие целостностью
не выжили, мало — надо разглядеть механизм, обеспечивающий
целостность. Поскольку мероно-таксономическое несоответствие
соединяет все со всем (в частности, всякая структура может быть,
вообще говоря, использована для всякой функции), то активность
особи должна обладать некоторым исходным свойством, позво-
ляющим ей находить структуру (орган) для исполнения данной
функции,— целесообразностью. О ней мы говорили в 4.5, а сейчас
обратим внимание на физиологический механизм ее действия —
доминанту. Понятие ввел в физиологию в 1890 г. Рихард Авена-
риус, для которого доминанта, выражаясь словами А. А. Ухтом-
ского,— «это преобразователь текущей реакции, фактор, направ-
ляющий поведение животного в данных условиях» [Ухтомский,
1950, с. 301]. Ухтомский разработал учение о доминанте сперва как
о способности центральной нервной системы поддерживать воз-
бужденным один нервный центр при торможении других, но вскоре
его стали трактовать чисто поведенчески и прилагать даже к орга-
166
низмам, не имеющим нервов [там же, с. 300]. В таком широком
понимании доминанта — это установка особи на реализацию опре-
деленной жизненной функции, ради которой остальные функции в
это время оставляются без внимания или даже подавляются. Так,
даже у одноклеточных можно говорить о пищевой, половой и за-
щитной доминантах, причем последняя выражается в повышении
порогов повреждающих клетку воздействий.
Замечательно, что доминанта может сохраняться вплоть до
крайнего истощения, причем «она гаснет последнею» [там же,
с. 165]. Сопоставив этот факт с тем, что в 4.4 говорилось об истоще-
нии и о стрессе вообще, легко видеть, что доминанта — то звено,
которое завершает спираль-стрессовую схему, уменьшая размер-
ность поискового пространства до одного или нескольких изме-
рений, т. е. делая поиск реально осуществимым. ФБ, много поколе-
ний подряд находящийся в сфере действия одной и той же доми-
нанты, имеет наибольший шанс быть преобразованным в' нечто
новое, о чем мы еще будем говорить. Сам же по себе стресс неспе-
цифичен и потому без доминанты не смог бы играть заметной
эволюционной роли.
Среди физиологов-эволюционистов никогда не гасла симпатия
к Ламарку, чью позицию в данном вопросе Карпов резюмировал
так: «Внешняя среда и ее изменения... вызывают особое состояние
напряжения в организме», которое «действует организующим
образом. Новые органы возникают в бесформенной „клеточной”
ткани, в которой движение жидкостей и флюидов прокладывает
каналы разного рода и отлагает различные вещества» [Ламарк,
1935, с. CXXXIV], Остается привести соответствующее мнение
нынешнего физиолога: «... В доминирующей системе образуется
структурный след, который увеличивает мощность системы и ста-
новится материальной основой долговременной адаптации»
[Меерсон, 1988, с. 11],—чтобы констатировать преемственность.
Когда-то Кювье записал шутливо об идее Ламарка: «Привычка
жевать привела через несколько столетий к появлению зубов;
привычка гулять — к появлению ног» [Burkhardt, 1977, с. 199],
и 150 лет ирония казалась уместной. Однако сейчас мы знаем, что
надо говорить не о «привычке гулять», а о необходимости пол-
зать — она действительно преобразует плавники в ласты и лапы,
если есть в наличии соответствующая морфологическая тенденция.
Возможна тенденция бйз необходимости в ней (кистеперые рыбы
имеют плавники-ласты с суставчатым скелетом, но плавают в
толще вод) и необходимость без достаточно явной тенденции (та-
ковы многие виды рыб, ползающие, ходящие и даже лазящие по-
средством обычных лучевых плавников), а их сочетание обуслов-
ливает эволюционный скачок. Недаром со времен Дарвина пишут о
необходимости синтеза морфологического и физиологического
подходов к эволюции [Bowler, 1977].
167
7.2. О факторах эволюции организмов
Вопрос о факторах довольно подробно рассмотрен в сборнике
[Диалектика развития..., 1978], где фактором названа «элементар-
ная (в смысле неразложимости в биологических понятиях) при-
чина или элементарное условие исторической трансформации
организмов» [там же, с. 90]. Там рассмотрено пять групп факто-
ров — изменчивости, наследования, взаимодействия, целостности
(системности) и прогресса. Сказанное выше позволяет сделать
уточнение: факторы изменчивости сами образуют две группы —
диатропические факторы (определяющие возможные варианты
изменений) и факторы активности (определяющие пути и способы
достижения этих вариантов). К первым относятся прежде всего
рефрены и другие тенденции, ко вторым — дискомфорт (т. е. внеш-
ние причины), смена функционирования и генетический поиск
(т. е. средства достижения эволюционного акта). Факторы актив-
ности и наследования — основные в обеспечении спираль-стрес-
совой схемы.
Говоря о шести группах факторов, надо иметь в виду, что
группировка эта достаточно условна - каждый акт эволюции
связан с несколькими факторами. Напомним, что Спенсер проводил
ту точку зрения, что процесс эволюции един и нерасчленим. Если
так, то фактор эволюции — феномен не природы, а познания.
В 4.4. мы уже говорили о тех взглядах, согласно которым
наследование является всего лишь стабилизацией одного из путей
онтогенеза, т. е. сокращает онтогенетическое разнообразие. При
этом либо сокращается полиморфизм вида (падает внутривидовая
изменчивость), либо каждый организм сокращает круг своих
возможностей (специализируется). В качестве возможного меха-
низма стабилизации Аршавский указал (ссылаясь на довоенные
работы А. А. Захваткина) на эмбрио низа.цию, т. е. на сдвиг (в ходе
эволюции таксона) развития органа или свойства на более ранние
стадии развития особи; при этом эмбрион потомка может иметь
признак, свойственный взрослому предку [Аршавский, 1982; Чай-
ковский, 1988 г]. Как приобретение, так и утрата первоначально
реализуются, с этой точки зрения, путем прижизненной модифи-
кации, которая завершает онтогенез. Это хорошо видно на примере,
когда-то поразившем Бэра: у одного слепого вида рыб мальки
имеют нормальные глаза, которые затем редуцируются, но не у всех
особей полностью [Ваег, 1861]. С нынешней точки зрения, столь
поздняя (на последней стадии онтогенеза) и не у всех особей пол-
ная редукция свидетельствует об эволюционной молодости данного
свойства, что подтверждается и малым возрастом соответствующего
биотопа (мутного озера). Редукцию неиспользуемого органа Бэр
истолковал там как физиологическую экономию, чем и заложил
основы неоламаркизма.
Наследованию, в таком его понимании, должны сопутствовать
противоположные явления — дестабилизация онтогенеза и неоте-
ния (когда взрослый потомок имеет признаки, бывшие у предка
эмбриональными). Без них историческое развитие то и дело захо-
168
дило бы в тупик. Чередование этих противоположностей в ходе
эволюции обычно. Еще в 1793 г. зоолог и натурфилософ К.-Ф. Киль-
мейер говорил, что «законы, по которым силы распределяются
между организмами, это те же законы, по которым силы распре-
деляются в особях одного и того же вида, и даже у одного и того же
индивида в разные периоды его развития», так что высшие формы
похожи, в начале развития, на низшие [Канаев, 1963, с. 159]. Воз-
ражая против универсальности такого вывода, Бэр в 1828 г. указы-
вал на противоположное явление, когда у эмбрионов «ниже стоя-
щих» животных наблюдаются свойства, исчезающие у них во
взрослом состоянии, но когда у «выше стоящих» эти свойства
сохраняются во взрослом состоянии. Например, у головастика мож-
но наблюдать клюв, подобный птичьему [Берг, 1977, с. 157]. Эти
феномены Агассиц позже назвал «пророческими типами», а геолог
А. П. Павлов в 1900 г. дал им эволюционное толкование и назвал
пророческими стадиями эволюции [Любищев, 1982]. Тем самым
онтогенез может как повторять в чем-то ход эволюции, так и пред-
варять его. Вместо рекапитуляции (повторения одним процессом
другого) теперь чаще говорят о гетерохронии (термин Геккеля),
т. е. об «изменении относительных сроков процессов развития»
[Рэфф, Кофмен, 1986, с. 185]. С диатропической точки зрения
это — следствие изоморфизма индивидуального и исторического
развития (онтогенеза и филогенеза). Точнее, разнообразие фило-
генезов заполняет ту же архетипическую таблицу, что и разнообра-
зие онтогенезов, и разнообразие ныне существующих видов, но как
перечень заполненных позиций, так и порядок их заполнения могут
быть различны. И в онто-, и в филогенезе процессы могут идти
как в прямом, так и в обратном направлении.
Соболев [1924, с. 92] отмечал, что широкое распространение в
эволюции феномена специализации возможно только потому, что
есть противоположный процесс, постоянно обеспечивающий приро-
ду неспециализированными формами. Этот процесс изучен много
хуже, чем специализация, поскольку происходит геологически
почти мгновенно и оставляет мало следов в палеонтологической
летописи. По-видимому, он всегда резко неадаптивен, сопровож-
дается массовым вымиранием деспециализирующихся таксонов и
потому не может быть сколько-то длительным. Тем не менее мас-
совый ископаемый материал, даваемый раковинами многих групп
моллюсков, позволил сделать ряд важных выводов. В 1930-х годах
палеонтолог Отто Шиндевольф разработал (развивая идею Павло-
ва) понятие протерогенеза, обозначающее исторический процесс,
во многом противоположный эмбрионизации. В ходе протерогенеза
«комплекс новых молодых признаков появляется на ранней стадии
индивидуального развития, а в дальнейшем онтогенезе исчезает;
в следующих звеньях генетического ряда он постепенно смещается
на всё более поздние онтогенетические стадии до тех пор, пока не
будут полностью вытеснены унаследованные древние признаки»
[Ивановский, 1976, с. 14]. В 1950 г. Шиндевольф сформулировал
следующую трехчастную схему развития филума (исторической
169
последовательности таксонов): «типогенез — первая фаза, на кото-
рой путем взрыва или количественной эволюции происходит вне-
запное возникновение крупных таксонов путем протерогенеза;
гипостаз — длительная стадия параллельного направленного
(ортогенетического) развития; типолиз — дисгармония, эволюци-
онный регресс, сверхспециализация, предшествующие вымира-
нию» [Ивановский, 1976, с. 17].
Сказанное здесь и в 7.1 можно свести в схему. Эволюция
крупной группы начинается с дестабилизации онтогенеза всех ее
особей, что легко представить себе как следствие массового дист-
ресса под влиянием внешних условий (в том числе и массовой
инфекции). В условиях дестабилизации, когда зародышу «нечего
терять», когда «сколько-нибудь закономерная организация еще не
возникла или почему-либо разрушена», возможны «весьма вариа-
бельные результаты» онтогенеза [Белоусов, 1987, с. 50]. Новое
создается при этом в ненаследуемой форме. Взрывной рост разно-
образия ведет к катастрофической гибели большинства, а остав-
шиеся демонстрируют во взрослом состоянии новые, в том числе
и зародышевые признаки. Затем следует длительное адаптивное
развитие группы, когда ищутся полезные прижизненные модифи-
кации, которые понемногу наследуются путем эмбрионизации.
Когда же процесс сокращения разнообразия берет верх, то история
группы завершается: таксоны чрезмерно специализируются и вы-
мирают. Хотя многое здесь еще предстоит выяснить в будущем, но
главный вывод диатропического характера виден уже сейчас: по-
скольку процесс эволюции состоит в заполнении архетипических
таблиц (объективно существующих в той же мере, что и таблица
химических элементов), постольку все факторы определяют лишь
характер заполнения позиций, заданных факторами первой груп-
пы. Пока, однако, степень заполнения данной таблицы низка, то все
факторы в равной мере можно рассматривать как творческие.
Особенно это ясно при заполнении ее первой клетки — при рож-
дении данного типа.
С этой точки зрения, генотип определяет собой не сам ход и
результат онтогенеза (их определяют законы природы как тако-
вые), а лишь его специфику — то, чем данный организм отли-
чается от других. Иными словами, изменение генов является не
причиной, а условием эволюции. Эмбриологи выражают эту мысль
по-разному. Они призывают «рассматривать генотип в качестве
параметризатора эпигенетических процессов» [Белоусов, 1987,
с. 206] или констатируют: «И мы до сих пор практически ничего не
знаем о том, связана ли реализация наследственной информации,
заключенной в ДНК и проявляющейся в последовательном синте-
зе различных органических веществ, со становлением специфиче-
ской формы или нет. Для разных организмов можно описать ста-
дии их химического развития и стадии их морфологического разви-
тия, можно однозначно сопоставить эти стадии... Но так называе-
мой каузальной (причинной) связи между этими двумя рядами
процессов не установлено и неизвестно, будет ли она установлена»
170
[Корочкин, 1986, с. 282]. Автономность молекулярного и морфо-
генетического «языков» Л. И. Корочкин называет «принципом
дополнительности в эмбриологии» [там же]. Напомним: Бор
считал •дополнительность общим законом познания.
Равноправие этих двух «языков» осознается с трудом, посколь-
ку гены принято считать высшим уровнем регуляции (против
этого всегда возражали ламаркисты [Wintrebert, 1962; Grasse,
1973]). Именно поэтому изменение самих генов трактуется как
случайное. В главе 3 мы выяснили, в каком смысле с этим можно
согласиться, а сейчас отметим, что вообще термин «случайность»
используется для обозначения сразу многих факторов эволюции,
относящихся сразу ко всем шести группам факторов и ко всем семи
типам случайности, названным в 3.1. Так, мероно-таксономиче-
ское несоответствие (диатропический фактор) выступает в наблю-
дении как случайность типов А, Б и Д, но при анализе механизмов
часто оказывается обычным следствием полифункциональности
органов и систем. Например, один и тот же молекулярный меха-
низм («соннеборновская наследственность» [Меклер, 1980а,
с. 353]), вероятно, ответствен за физиологическую и наследствен-
ную адаптацию клетки, за память и за репарацию двунитевых
разрывов ДНК. Пока все четыре функции рассматривали порознь,
они казались совершенно независимыми (взаимно случайными),
но выявление общего для них механизма позволяет не только све-
сти к минимуму ссылки на случайность в наследственной адапта-
ции, но и говорить о едином факторе целостности, определяющем
эволюцию, онтогенез и жизнь особи. То же можно сказать и о
«прыгающих генах», перемещения которых описаны в литературе
как случайные, но которые, как мне представляется, ответственны
за согласование двух «языков», о которых пишет Корочкин.
Совсем другой пример феномена, в котором посредством слу-
чайности взаимосвязаны различные факторы онтогенеза и эволю-
ции, являет собой миграция первичных половых клеток. Эти
клетки обычно образуются в самом раннем онтогенезе животных,
раньше, чем появятся зачатки гонад (половых желез), куда они
затем встраиваются и там служат источником спермиев или
яйцеклеток. Такой ход дел понятен: для размножения нужны
полноценные неспециализированные клетки («зародышевый
путь», о котором писал еще М. Нуссбаум в 1880 г.), тогда как соб-
ственные ткани гонад все специализированы. Непонятно другое:
почему-то у многих групп животных первичные половые клетки
сперва распределяются по всему зародышу, а затем только миг-
рируют в гонады. Один из первых исследователей миграции Луи
Бунур заметил мимоходом, что здесь проявляется двоякая природа
наследственности: «С одной стороны, это — механизм, обеспечи-
вающий передачу признаков в цепи поколений, т. е. относительное
постоянство видов, а с другой — иные механизмы, обусловливаю-
щие внутривидовую изменчивость, т. е. наличие в популяции осо-
бей, не вполне тождественных друг с другом» [Происхождение...,
1968, с. 186]. Клетки, прибывшие в гонаду из различных органов
171
зародыша, могут в принципе иметь различную информацию о том,
как протекает онтогенез, и это как раз та информация, которой
компетентная половая клетка должна отличаться от специализи-
рованных клеток, имеющих тот же набор генов. В настоящее время
известно, что половая клетка действительно может получать от за-
родыша генетическую информацию — например, в форме вирусов,
о чем можно прочесть в 6.2.2 книги Р. Хесина [1984] 2. Поэтому
организмы, развившиеся из клеток, пришедших в гонаду разными
путями, могут быть генетически различны и давать различный
материал для наследования.
Тем самым разнообразие в пределах популяции, которое обычно
описывается как случайное, в действительности может нести ту
информацию о строении и онтогенезе организмов, которая в одной
особи содержаться не может. Популяция выступает как целост-
ность, как минимальная самодовлеющая величина — даже в том
случае, когда особи генетически тождественны. Когда же геномы
особей различны, то каждая клетка зародыша — гибрид, и потому
информация о результате взаимной адаптации в зародыше двух
геномов тоже полезна. У многих низших животных (гидры, асци-
дии) миграции нет, как нет ее. возможно, у грибов и растений,
и встает вопрос: как отсутствие этого фактора сказалось на ходе
и результатах их эволюции? Он еще ждет ответа.
Как отмечал Мейен, хотя многие отрицают самодовлеющую
роль целостности, фактически ее признают все — хотя бы в том,
что принимают как нечто данное «готовые таксоны и архетипы»
[Диалектика развития.... 1978, с. 147]. Последовательный отказ
от факторов целостности ведет к абсурду [там же. с. 149—150].
поэтому лучше ввести их в рассмотрение сознательно. Этот вопрос
слабо разработан, но все-таки известно, что в результате эволюции
появляются таксоны, характеризуемые распределением Виллиса,
т. е. системной случайностью. Это наводит на мысль о наличии
факторов целостности, обладающих системной случайностью,
в механизме таксонообразования (Юл называл их видовыми и ро-
довыми мутациями).
Другой пример фактора целостности, связанного со случайно-
стью, являет собой невыводимость по Гёделю (подробнее о ней
см.: [Чайковский, 1985]). В 1931 г. логик Курт Гёдель доказал
знаменитую теорему о неполноте: в рамках достаточно сложной
логической системы можно сформулировать истинное утвержде-
ние, которое, однако, нельзя вывести из аксиом этой логики посред-
ством правил вывода этой логики. Существенно, что истинность
такого утверждения доказать можно, но это будет доказательство-
проверка. а не доказательство-вывод: другими словами, возможно
утверждение, в истинности которого можно убедиться средствами
данной логики, если утверждение уже сформулировано, но самый
вывод этого утверждения (нахождение формулировки) требует
2 Передача фрагментов ДНК между организмами посредством вирусов и т. и. пред-
ложена в качестве особого фактора эволюции [Тихоненко, 1979]. Это — один из
факторов взаимодействия организмов.
172
более общего аппарата, нежели данная логическая система. Все
формальные системы делятся на две группы в отношении разно-
образия их свойств — на примитивные и гёделевы. Примитивная
система полностью определяет свои свойства в том смысле, что
любой факт, формулируемый на ее языке, в ней формально выво-
дится: вся эволюция в рамках такой системы сводится к удлинению
текстов, записываемых по раз навсегда заданным правилам.
Наоборот, гёделева система способна к качественной эволюции:
в ее рамках можно записать осмысленный текст, смысл которого не
вытекает из правил его построения (на основе более коротких
текстов), хотя и согласован с этими правилами. (Именно это об-
стоятельство отметил и Корочкин своим «принципом дополни-
тельности в эмбриологии».) Получение такого текста в рамках
системы неформально и может быть формализовано только путем
добавления новой аксиомы (выхода на новый логический уро-
вень). Добавление аксиомы выглядит произвольным {случайным)
актом с позиций прежней логики, но закономерно с позиций
новой (расширенной) логики: добавляется именно аксиома, кото-
рая нужна для формального вывода заданного утверждения. Все
интересные нам системы — гёделевы.
Именно наличие у систем целостности позволяет прилагать к
ним такие понятия, как введенные в 1.2 causae fornialis. finalis.
exeniplaris.
В рамках гёделевой системы исходное разнообразие порождает
почти необозримое множество формально выводимых текстов
(редукционизм), но существуют тексты, задаваемые только ир-
редукционно. Расширение системы (введение новой аксиомы.)
формализует и эти тексты, но само расширение всегда неформаль-
но. Следовательно, эволюция состоит здесь в плавном заполнении
одного уровня, изредка сопровождаемом быстрым выходом на но-
вые, более широкие уровни (ароморфоз по Северцову). Такие акты
все авторы связывают с понятием прогресса, но обычно в этот
термин вкладывают еще и более широкий смысл, связывая с ним
всякое усложнение формы организмов, о чем мы говорили в 1.4.
Если понимать прогресс как всякое усложнение, то в качестве
факторов прогресса можно указать генетическую интеграцию
[Чайковский. 19776] и топологические преобразования онтогенеза
[Преснов, Исаева. 1985]. Они же содействуют ароморфозам,
однако ароморфозу можно указать и свою собственную причину —
градацию уровней, о которой мы узнаем в главе 9.
Другой тип факторов целостности составляют факторы, связан-
ные с позитивным взаимодействием организмов (взаимопомощь по
П. А. Кропоткину, спаривание, симбиоз и т. д.) и их частей («срод-
ство частиц» но Мопертюи, «корреляция частей» по Кювье, «при-
рода организма» по Дарвину, «творчество» по Бергсону, «двига-
тельная активность» по Аршавскому и т. д.). Наоборот, негативное
взаимодействие (конкуренция, каннибализм, инфекционные бо-
лезни и т. д.) в некотором смысле противостоит целостности,
хотя провести здесь четкую границу и не удается.
173
Только перечислив все группы факторов, можно обратиться к
естественному отбору, чтобы понять, что собственно является
объектом его действия. Многие еще считают, что таким объектом
являются признаки организма, которые действием отбора делятся
на позитивные (сохранившиеся) и негативные (утраченные).
Наиболее последовательная концепция — фенетика [Яблоков,
1980] — рассматривает и сам организм фактически как набор
сохранившихся признаков, В этом можно видеть логическое за-
вершение статистического мировоззрения, о котором шла речь
в 1.2. В противоположность ему. системное мировоззрение считает
конкретные признаки лишь эпифеноменом целостной организа-
ции — см. конец 5.4 и начало 7.1. Отбор отдельных признаков
малоэффективен (см. 3.4), отбор может быть эффективен только
тогда, когда объектом его действия являются цельные конструкции,
а не отдельные признаки. Это, в общем, соответствует взглядам
молодого Дарвина, который затем склонился к статистическим
взглядам. Однако с позиции последних нельзя ответить на вопрос,
почему новое качество сохраняется при скрещивании его носителя
с прочими членами популяции (этот вопрос, приведший в смятение
Дарвина, часто именуют «кошмаром Дженкина»), И в наши дни
приходится признать, что «генные комбинации разрушаются поло-
вым процессом так же быстро, как порождаются» [Kimura, Ohta.
1971, с. 160]. Те немногие примеры эффективного действия отбора,
какими располагает наука, демонстрируют, что он эффективен
тогда, когда близок к искусственному, т. е. когда обладатели
ненужных качеств почти или вовсе не участвуют в размножении,
а само размножение никак не зависит от конкуренции между
особями за дефицитный ресурс. Тем самым отбор как фактор
эволюции нет оснований толковать «по Мальтусу», и встает вопрос
об ином — системном — понимании отбора. Попыток в этом
направлении было много (см.: [Бабков, 1980]), но мне представ-
ляется наиболее конструктивным то понимание, которое в 1969 г.
дал химик А. П. Руденко: в открытой системе данный субстрат
в основном перерабатывается той подсистемой, которая делает
это наиболее активно. Здесь отбираются не единичные акты измен-
чивости. а массовые процессы, целостности [Диалектика разви-
тия..., 1978. с. 106].
При таком понимании отбора особенно ясно, что он — вовсе не
отрицательная обратная связь (как часто мимоходом говорят),
а положительная, что он дестабилизирует экосистему. О том, что
избыточность размножения (основа отбора по Дарвину) служит
положительной обратной связью, пишет и Одум [1986, т. 1, с. 61].
Для стабилизации же служат факторы, касающиеся целостности.
Это — никак не умаление роли отбора, но конкретизация того суб-
страта («основы подбора» по Богданову), на который он действует.
Об этом писал Любищев [1973, с. 43]: «Как известно, всякое учение
об эволюции заключает три момента: изменчивость, наследствен-
ность и естественный отбор», однако «все дело в том, как понимать
наследственность и изменчивость». В части I было показано, что
174
изменчивость сложно и многоуровнево упорядочена, а наследст-
венность мобильна, что отбор достаточно груб (малые различия
выгодности не влияют на вероятность выживания), что существен
только катастрофический отбор, т. е. чье-то преимущественное вы-
живание в условиях почти полной гибели группы. Все это позво-
ляет очертить следующую схему действия факторов, насколько она
видна в настоящее время.
Умеренный дискомфорт вызывает физиологический стресс (по
Аршавскому), активацию и тренировку (по Гаркави и др.), эуст-
ресс (по Селье), что ведет к адаптивным модификациям; а патоло-
гический дискомфорт влечет за собой дистресс, при котором (в ос-
новном, по-видимому, при шоке и истощении) прорываются защит-
ные барьеры генетической системы и тем открывается путь к актам
(пусть и редким) наследования, выражающимся в фиксации новых
путей окончания онтогенеза. Для популяции такой процесс высту-
пает как катастрофа (в этом — роль отбора), а сам эволюционный
акт выглядит исторически мгновенным.
Когда специализация или внешние обстоятельства подводят
таксон к катастрофе, то все его особи подвергаются одному и тому
же испытанию, и у всех апробируется одна и та же система (ор-
ган) — та, которая наиболее уязвима в данных условиях, посколь-
ку от нее более всего зависит смертность. Она функционирует в
предельном напряжении (как пищеварение тлей в опытах Шапош-
никова), находится в сфере доминанты по Ухтомскому, а организ-
мы в целом испытывают, в силу длительного дискомфорта, дистресс
по Селье. Дистресс вызывает прорыв барьеров генетической систе-
мы, т. е. синтез модифицированных ДНК, массовую дестабилиза-
цию онтогенезов и наследственности. Это возможно, если в основе
каждого онтогенеза лежит неспециализированная клетка, тогда как
конечная стадия развития (взрослая особь) может быть очень спе-
циализирована. Наличие такой клетки позволяет даже очень спе-
циализированному организму стать предком новых форм. Выдви-
нутый сто лет назад Копом «закон неспециализированного пред-
ка», по-видимому, места не имеет [Меуеп, 1987, с. 365], так как
предок мог быть специализирован и деспециализироваться в ходе
катастрофы. Немногие выжившие в ней дают материал для типоге-
неза по Шиндевольфу (или ароморфоза по Северцову). Группы, на-
шедшие физиологически и экологически приемлемый вариант, вы-
ходят из дистресса и стабилизируются, причем существен быст-
рый (горизонтальный) обмен генетической информацией. В сле-
дующей катастрофе объектом апробации могут послужить другие
системы (органы), т. е. отбор может действовать на них поочеред-
но. Это делает его реальным фактором.
Понятие «катастрофический отбор» не ново (см.: [Raven,
1964]), но до сих пор рассматривалось лишь как одна из многих
форм отбора, а не как общий принцип действия этого фактора эво-
люции. Встает вопрос: если отбор эффективен только в режиме
катастрофического сокращения численности, то насколько он во-
обще существен для эволюции? Здесь следует указать на следую-
175
щее соображение Мейена: во влажных тропиках, где не бывает
катастрофических перепадов температуры и влажности, эво-
люция высших растений течет гораздо интенсивнее и продуктив-
нее, следовательно «изрядная доля... макроэволюционных актов
происходила... при „попустительстве” (а не под давлением) отбо-
ра» [Мейен, 19876, с. 304], точнее — абиотического отбора. Пара-
докс в том, что специфическим фактором макроэволюции здесь ока-
зывается «всего лишь торможение одного из факторов — естест-
венного отбора» [там же, с. 308]. О торможении как факторе писа-
ли еще в 1979 г. палеозоологи В. В. Жерихин и А. П. Расницын
[Меуеп, 1987, с. 370]. Наоборот, движущая роль отбора в макроэво-
люции была Мейену сомнительна: «Мы знаем, что эффективность...
репродуктивного цикла у покрытосеменных несравненно выше,
чем у голосеменных. Тем не менее покрытосеменные не смогли
вытеснить голосеменных из лесного древостоя в течение многих
миллионов лет. Коэффициент селективности (эффективность от-
бора.— Ю. Ч.) оказался явно недостаточным. Почему же мы долж-
ны считать его достаточным, когда речь идет о гораздо меньших
различиях...?» [там же, с. 368 — 369]. Как мы уже говорили (см.
4.5 и 4.6), акты приспособления могут быть не связаны с отбо-
ром, а акты эволюции — с приспособлением. Согласно Мейену,
неселективные макроэволюционные акты происходят случайно, но
без определенной вероятности [Мейен, 19876, с. 303], что вполне
понятно, если трактовать их как произвольный выбор (по Гёделю,
а также в смысле типа Ж из 3.2) между различными целостными
конструкциями.
Концептуальная ценность схемы факторов, изложенной здесь,
видится в том, что каждый ее блок имеет обоснование — либо в
наблюдении, либо в эксперименте, либо в мажорирующей модели.
Хотя в целом схема пока еще гипотетична, но обоснованность бло-
ков подымает ее статус выше статуса прежних эволюционных мо-
делей, где гипотетичны и сами блоки. Сторонники традиционной
системы факторов (где отбор поставлен выше других факторов)
тоже понемногу меняют позицию. Например: «Становится все бо-
лее очевидным, что в определении направленности эволюционных
процессов существенную роль играет не только отбор, но и некото-
рые организменные факторы, такие, как механизмы морфогенеза»
[Татаринов, 1987, с. 5]. Однако ценность схемы факторов должна
определяться не столько сравнением ее с другими, сколько ее соб-
ственной работоспособностью, о которой и пойдет речь в сле-
дующих главах. Наиболее важна возможность давать конкретные
рекомендации прикладным научным дисциплинам (см. часть III).
7.3. Преобразование разнообразия
и происхождение таксонов
Эволюцию удобно описывать как два сопряженных процесса —
происхождение организмов и происхождение экосистем. Данное в
7.2 беглое описание хорошо бы, в принципе, допол-
176
нить описанием факторов эволюции экосистем, но на сегодня это
вряд ли возможно: хотя некоторые из таких факторов известны
(например, упомянутая в 7.1 незамкнутость круговоротов и за-
тронутая в 7.2 неустойчивость селективных процедур), но перечня
факторов, по-видимому, до сих пор нет [Татаринов, 1987, с. 5],
не говоря уж о какой-либо их системе. Поэтому приходится сопря-
гать достаточно разработанный анализ эволюции организмов с
весьма схематичными представлениями об эволюции экосистем,
образуемых ими. Основной сопрягающей процедурой пока что
служит анализ преобразования таксономического разнообразия
[Красилов, 1987].
Еще в 1858 г., до обнародования идей Дарвина, зоолог и мето-
долог Страхов писал, что «дело не в том, как произошли формы
и могут ли они переходить одна в другую, а в том, как они сущест-
вуют в своем разнообразии». И пояснял это химической аналогией:
если когда-то научатся превращать серу в кислород и обратно,
то «и тогда, как и теперь, это будут два различные вещества»;
поэтому ясно, что переход одного вида в другой должен происхо-
дить «быстро, скачком», подобно превращению льда в воду [Стра-
хов, 1858]. В сущности, этим было выражено то, что через 120 лет
Мейен назвал номотетической концепцией эволюции. Суть ее — в
указании на «пространство логических возможностей» (термин
Г. А. Заварзина), структура которого определяет спектр (архетип
по Мейену) возможных для данного таксона вариантов (целостных
структур). Тем самым акцент делается не на способ, как в ламар-
кизме, не на путь, как в дарвинизме, и не на результат, как в
номогенезе, а на разнообразие (способов, путей и результатов). Мы
уже говорили, что рефренная структура разнообразий свидетельст-
вует о структурированности этого «пространства». Когда степень
заполнения пространства высока, рефрены видны отчетливо, но су-
ществуют они всегда. Приспособительный характер эволюции вы-
ражен в рамках одного рефрена тем, что плохо приспособленные ва-
рианты редки, а совсем не приспособленных мы вообще не видим.
Рефренная структура, по-видимому, существует всюду, но не
всегда ее легко разглядеть. В 4.6 мы видели, например, что неудач-
ная конструкция гениталий пятнистой гиены, кажущаяся просто
курьезом, в действительности является реализацией редкого ар-
хетипического варианта, снижающего жизнеспособность, но все
же допустимого. Механизм, реализовавший этот вариант в эволю-
ции, достаточно очевиден: «имитация мужских гениталий у
самки пятнистой гиены не могла произойти помимо гамогетерото-
пии. В противном случае нужно идти на нелепое допущение, что
предки гиены были гермафродитными» [Мейен, 1986а, с. 304].
(Напомню, что гермафродитные виды не встречаются среди поз-
воночных, кроме некоторых рыб.)
У растений гамогетеротопия не связана со снижением жизне-
способности и потому смогла быть более существенна в их эволю-
ции: ее не только успешно привлекали для объяснения происхож-
дения нового рода (кукурузы) и некоторых семейств (голосемен-
177
ных), но она дала единственную на сегодня убедительную гипоте-
зу происхождения целого типа (отдела) — цветковых [Мейен,
1986а]. В этой гипотезе главный пункт — переход одного органа,
а именно, плодолистика (образовавшего первое подобие листовки,
т. е. примитивного стручка) с мужской фруктификации голосе-
менного предка на женскую фруктификацию (плод) покрытосе-
менного потомка. Этот переход (гамогетеротопия) мог произойти
только мгновенно, в одно поколение, хотя распространялся в
исходной популяции, вероятно, долго.
Другие гетеротопии (перемещения) могли возникать и сразу,
и постепенно. Например, перемещение брюшных плавников вперед
(югулярная тенденция — см. 2.5) возможно обоими способами.
Вообще, при гетеротопии «уже отработанная онтогенетическая
программа включается в новом месте и производит здесь тот же
орган» [Меуеп, 1987, с. 356], поэтому постепенность, которую тре-
бовали традиционные концепции эволюции, необязательна. Изме-
нение программы, осуществляемое переключением регуляторных
генов, происходит само по себе сразу, так что демонстрируемые
часто (например, в югулярной тенденции) ряды «переходных
форм» вовсе не обязательно отражают постепенность эволюцион-
ного процесса — о ней можно говорить только тогда, когда она чет-
ко прослеживается вверх по геологическому разрезу. На это указы-
вал, возражая Дарвину, еще палеонтолог Герман Траутшольд
[Trautschold, 1860, с. 528]. Чаще же «непрерывные» ряды отража-
ют лишь рефрен, характерный для данного таксона, а не эволюцию
таксона. Это ясно, в частности, из того факта, что «морфологиче-
ские тенденции часто бывают встречными. У одних растений идет
[эволюционный] процесс от А к В, а у других от В к А»
[Меуеп, 1987, с. 368].
От предков к потомкам переходят не столько отдельные призна-
ки, сколько целые рефрены. В силу феномена (Мейен [1978] наз-
вал его транзитивным полиморфизмом) разнообразие восстанав-
ливает себя почти независимо от того, какая его часть берется для
размножения. Яркий пример приводил Дарвин (см. 1.5): известно
огромное разнообразие бархатистых персиков — по вкусу, форме и
цвету плодов, по форме косточки и т. д.; среди них возникла
вариация (мутант) с гладкой кожицей, и вот она повторила своими
(вторичными) вариациями все формы бархатистых персиков. Гля-
дя на это вторичное разнообразие, нельзя понять, из какой формы
оно получено, т. е. какая форма послужила для транзита. Рефрен-
ная структура вида была воспроизведена заново. Подробнее см.:
[Чайковский, 19896].
Каждый признак, возможный в рамках данного рефрена, когда-
то реализуется — пусть и очень редко (см. 2.4). Это существенно
для эволюции, состоящей в значительной мере из исчезающе
редких актов: макроэволюционная новация у цветковых происхо-
дила «один раз за несколько сот тысяч лет на всей Земле. У голосе-
менных такие новации происходили в 5—7 раз реже», а вне тропи-
ков — вообще 1—2 раза за 100 млн лет [Мейен, 19876, с. 303]. Про
178
каждый из этих актов нельзя было бы сказать ничего (поскольку
они случайны, но не обладают вероятностями), если бы за
700 млн лет эволюции 3 они не наметили бы собою контуры соответ-
ствующих рефренов. Некоторые позиции столь редки, что для оцен-
ки принадлежности их к определенному рефрену приходится выхо-
дить даже за пределы царства. Так, известный с силура (около
420 млн лет назад) класс усоногих раков содержит в одном из своих
отрядов два семейства, некоторые виды которых гермафродиты, но
имеют карликовых дополнительных самцов. Дарвину, описавшему
этот феномен в 1854 г., мы обязаны и замечательным сопоставле-
нием: ничего похожего среди животных нет, но есть среди цветко-
вых [Дарвин, 1936, с. 86]. Дать эволюционное объяснение феноме-
ну (т. е., в его понятиях,— построить ряд постепенных улучшений
функции) Дарвин не смог (хоть и пытался) и ограничился в книге
1877 г. восклицанием: «Поразительна редкость таких случаев»
[Дарвин, 1948, с. 49],— которое, как ни странно, и заключало в се-
бе подход к объяснению: если вариант крайне редок и его носители
живут в различных средах, то бессмысленно пытаться включать
его в адаптивный ряд, зато есть смысл строить диатропическую
таблицу. И Дарвин ее мысленно построил и даже дал по ней прог-
ноз!
Он выписал все логически возможные сочетания полов и кон-
статировал, что один из вариантов — на одних особях только гер-
мафродитные цветки, а на других только мужские — неизвестен в
норме, хотя описан в качестве индивидуальной вариации у калуж-
ницы болотной. На тот случай, «если существуют» виды с таким
свойством, Дарвин дал ему название — андродиэция [там же].
(Заметим, что еще у Линнея названо четыре таких рода, в том
числе хурма [Linne, 1751, §149].) Замечательно, что обладающие
ею виды двух семейств принадлежат к разным подотрядам
(Lepadomorpha и Balanomorpha), возникшим с интервалом более
70 млн лет [Vermeij, 1987, с. 331], т. е. дополнительные самцы —
тоже, вероятно, элемент транзитивного полиморфизма.
Итак, номотетика налицо даже в столь редких явлениях, как га-
могетеротопия и андродиэция. Главным же объектом номотетиче-
ского эволюционизма являются массовые закономерности. Напри-
мер, Агассиц писал: «Почти в каждом классе имеются тропические
семейства, и они-то, как правило,— высшие в своих классах, а если
и бывает наоборот, когда они явно стоят на низком уровне, то име-
ются явственные отношения их с превалирующими типами прош-
лых эпох» [Agassiz, 1857, с. 121]. Пояснял он это такими, напри-
мер, доводами: обезьяны ближе всего к человеку и живут в тропи-
ках, а бегемоты (которые хоть и стоят среди зверей низко, но тоже
живут в тропиках) были ведущей группой в прошлом. Это, как те-
перь видно, было гениальной догадкой Агассица — ведущая роль
3 Если полагать начало фанерозоя (сколько-то надежной палеонтологической
летописи) с венда. Сама же жизнь возникла по меньшей мере на 3 млрд лет ранее.
179
тропических форм в глобальной эволюции. (Напомним, что самый
факт эволюции Агассиц отрицал.)
И у Агассица, и у Дарвина и, пожалуй, у всех, кто писал за и
против эволюции следующие 125 лет, всегда наблюдался разрыв
между разными корпусами фактов, разрыв, заполняемый полетом
фантазии, которую можно принять или отвергнуть в зависимости от
вкуса. Мейен поступил иначе: скрупулезно исследовал данные обо
всех семействах ископаемых высших растений (от мхов до цвет-
ковых) по всем эпохам (от девона до неогена, причем наиболее пол-
но это удалось для мезозоя) и всем континентам, чтобы спроециро-
вать геологические данные на палеогеографическую карту (Агас-
сиц называл это параллелью геологии с географией). Вот его выво-
ды. Все растения, насколько это сейчас известно, произошли мо-
нофилетично — от одного предкового класса, и все таксоны от се-
мейства и выше монофилетичны. С конца девона (~300 млн лет
назад, когда Пангея распалась на Гондвану и Лавразию) однород-
ная флора распалась на три области — экваториальную, северную
и южную. Все порядки, кроме одного, и почти все семейства прои-
зошли в безморозном климате, большинство — в тропическом. Гло-
бальный флорогенез представлял собою растекание таксонов (фи-
тоспрединг) от экватора к высоким широтам [Мейен, 19866;
1987а, б].
Вряд ли надо объяснять, насколько эта простая изящная схема,
если она будет принята, упростит и организует палеоботанику —
от самых высоких ее академических вопросов до чисто практиче-
ских задач поиска ископаемых. Поэтому обращу внимание лишь
на малозаметные слова «от семейства и выше». Ведь до сих пор
считалось прямо наоборот: низшим таксонам (родам и, где возмож-
но,— видам) приписывали монофилию, а вот о высших спорили и
самым высшим (классы, отделы) обычно находили по нескольку
предков. Мейен же убедился, что при образовании семейств часто
господствует транзитивный полиморфизм: роды данного семейства
складываются из родов предкового семейства так, что каждый бе-
рет признаки разных предковых родов вперемешку, синтетически.
«Синтетические типы» нередки среди хвойных и цветковых [Мей-
ен, 1987а, с. 217 и далее, с. 270; Меуеп, 1987], причем для хвойных
Мейен утверждал, что указать такому таксону определенный так-
сон-предок часто невозможно в силу определяющей роли транзи-
тивного полиморфизма [Меуеп, 1987, с. 362]. О других группах он
столь определенно не писал, но в одной из последних бесед со мной
характеризовал транзитивный полиморфизм родов как основное
препятствие для филогенетического подхода. В этом свете понятна
и фраза: «Вполне возможно, что само распределение признаков
между синтетическими типами разных уровней окажется препят-
ствием на пути выявления» предков, «особенно между таксонами
более низкого ранга» [там же].
Разумеется, не надо распространять эти выводы, касающиеся
высших растений, на все группы организмов, особенно — океани-
ческих, но все-таки их общетеоретическое значение огромно. В кон-
180
цепции глобального флорогенеза, по-моему, наука получила первое
серьезное доказательство феномена эволюции как естественного
процесса, тогда как прежде речь шла о толкованиях. На Западе
сейчас вновь усилились разговоры о том, что «доказательства»
эволюции суть не более, чем свидетельства творения [Morris,
1981], но пусть их сторонники попробуют теперь апеллировать не
к Дарвину, а к Мейену. Если Дарвин, следуя Ламарку и другим,
лишь проводил параллель между распределением нынешних орга-
низмов в пространстве и их гипотетической эволюцией во времени,
то Мейен первым показал, как время фактически преобразовывало
и расселяло разнообразие высших растений, начиная с девона;
и пусть сторонники творения объясняют именно это. Бога, созна-
тельно творившего такое преобразование, представить себе можно,
но он будет уже по существу неотличим от естественного процесса
эволюции.
Закономерности, открытые Мейеном в палеоботанике, увидеть
на зоологическом материале труднее, по крайней мере по двум
причинам: высшие животные более целостны (интегрированы) и
легче перемещаются, чем высшие растения. Поэтому блочность
(мозаичность) проявляется у них не столь ясно (гомологические
ряды обычно не столько полны), а приуроченность определенных
таксонов определенным зонам обитания не столь отчетлива. Однако
при хорошей изученности таксона аналогии с ботаникой могут быть
различимы. Так, еще в 1863 г. Карл Фогт, один из первых немец-
ких дарвинистов, горячо возражал против идеи общего предка (ее,
считал он, приписывают Дарвину противники, чтобы скомпромети-
ровать новое учение). По Фогту, если производить строение чело-
века от обезьяньего, то надо взять голову ревуна, руку шимпанзе,
скелет туловища гиббона, ногу гориллы и мозг оранга; в этом он ви-
дел свидетельство того, что эволюция идет не путем расхождения от
общего предка, а путем скрещиваний [Фогт, 1865]. Дальнейший
прогресс наук, в том числе находки ископаемых, не улучшил по-
ложения, и в 1977 г. ведущие английские антропологи отказались
указать филогению (последовательность предков) человека: хотя
для некоторых костей есть «непрерывная временная последова-
тельность в одной и той же географической области», но она не сог-
ласуется с их представлениями о ходе эволюции человека в целом
[Харрисон и др., 1979, с. 105]. Причина этой и подобных трудно-
стей гораздо более обща, чем феномен скрещивания, она носит
общедиатропический характер: в силу рефренной структуры разно-
образий, сходные мероны бывают у очень различных таксонов, а в
силу транзитивного полиморфизма, сходство не так уж много го-
ворит о родстве.
Фактически, всякая филогения, даже при наличии ископаемого
материала, строится путем группировки таксонов по сходным меро-
нам, и таксоны могут оказаться близки по одним меронам, но да-
леким по другим. Приходится говорить о семофилогениях, т. е. от-
делённых филогениях признаков, причем каждый мерой образует
свою семофилогению. Например, говорят об эволюции плавника в
181
лапу. Филогения достоверно строится только тогда, когда все семо-
филогении сходятся к единой предковой форме, но этого-то часто и
не бывает.
В частности, филогению высших растений удалось построить,
а филогению человека не удалось, потому что для растений основ-
ные семофилогении сошлись воедино, а для человека — нет. И дело
здесь не в различной изученности объектов («Нет другого сущест-
ва, так хорошо изученного по форме тела, как человек» [Яблоков,
1980, с. 97]), а в их таксономическом ранге: филогения существует
не столько для видов и родов, сколько для более обширных раз-
нообразий. Причина этого видится сегодня в том, что на уровне
рода и ниже степень заполнения архетипической таблицы до-
статочно высока, чтобы выстраивать гомологические ряды по боль-
шинству признаков, в том числе и диагностических. В такой ситу-
ации вид попросту не несет достаточной информации о своем пред-
ке, а разнообразие видов большого семейства высших организ-
мов — несет, если считать предком семейства как целого более
древнее семейство как целое. Для животных это еще далеко не так
выяснено, как для растений, но уже очевидно, что дальнейшие на-
ходки к построению филогении человека не приведут, если, ра-
зумеется, не пренебрегать известными на сегодня семофилоге-
ниями.
У низших организмов, бывших объектом классификации в гла-
ве 6, комбинативнось столь велика на всех таксономических уров-
нях, что филогении не выявляются (да их, возможно, и нет объек-
тивно — в той мере, в какой их эволюция протекала путем генети-
ческой интеграции). Поэтому здесь имеет смысл ставить только за-
дачи об эволюции механизмов и структур (но не о выявлении пред-
ков), зато сами эти задачи решаются легче и полнее именно при от-
казе от филогенетического метода. В самом деле, такой отказ дает
возможность следить прямо за логикой развития механизма:
такие феномены, как транзитивный полиморфизм, гетеротопия и
генетическая интеграция, позволяют развиться даже тому, что не-
прерывно (постепенно) развиться не может.
В частности, приходится отказаться от схемы эволюции геноти-
па путем постепенных добавок, изъятий и замен нуклеотидов —
хотя формально таким путем и можно получить из любого ге-
нотипа любой, однако среди промежуточных форм неминуемо ока-
жутся логически запрещенные (по Гёделю, т. е. сами себе не соот-
ветствующие) . Это неизбежно, в частности, когда конечный гено-
тип принадлежит более высокому уровню, чем начальный. В связи
с этим укажем недавнюю работу, в которой систематика крупной
группы растений построена на четком признании: а) исходного
разнообразия, б) неполной фиксированности логических правил и
в) отсутствия некоторых промежуточных форм [Мейен, 1982].
В следующей главе будет показано, как эти методологические
установки помогают прояснить ряд вопросов становления клеточ-
ной организации живого.
182
Глава 8. ДИАТРОПИЧЕСКИЙ АНАЛИЗ
НЕКОТОРЫХ ПРЕДСТАВЛЕНИЙ
ОБ ЭВОЛЮЦИИ КЛЕТОК
Следующий краткий анализ основан, если не оговорено иное, на
данных, которые можно найти в работах: [Чайковский, 19776,
1980; Шлегель, 1987; Эволюция..., 1987].
8Л. Связь концепций эволюции биосферы и клетки
Многие считают, что проблема возникновения жизни состоит,
прежде всего, в загадке появления аппарата наследственности
(пресловутый вопрос: что было раньше — курица или яйцо?). Ес-
ли, однако, всерьез задуматься над происхождением жизни, то
встает другой вопрос: что было раньше — организм или условия
для его существования, организм или экосистема? Оба эти аспекта
проблемы происхождения жизни (генетический и экологический)
сходны в том, что не удается говорить о происхождении элемента
отдельно от происхождения всей системы как таковой. Для обсуж-
дения подобных вопросов наука еще только начинает вырабатывать
язык.
В 1924 г. появилась книжка А. И. Опарина «Происхождение
жизни», в которой проблеме был придан научный статус: было по-
казано, что в условиях бескислородной атмосферы могло идти на-
копление мелких органических молекул (аминокислот, сахаров,
нуклеотидов и т. п.), которые, как предполагается, были нужны для
образования первых организмов, а также небольших цепочек из
таких молекул. Однако главные трудности оставались еще впереди,
так как здесь ставилась только проблема «начала начала» жизни.
Даже если мы поместим в сосуд все макромолекулы, образующие
клетку, это не даст нам клетки; а если мы поместим в океан даже
целую клетку, но только одну, это еще не даст океану жизни: если
клетка и будет делиться, то только пока будут в достатке подхо-
дящие молекулы, а потом погибнет. Мы знаем жизнь только в одной
форме — в той, которая окружает нас на Земле. Эта жизнь умеет
существовать только целиком, и мы не в силах всерьез пред-
ставить себе, как мог бы существовать один-единственный вид,
а не то что единственная особь.
В наш век экологического кризиса сказанное воспринимается
само собой и даже трудно представить, что в течение всей своей
истории человечество в целом находилось далеко от понимания эле-
ментарных экологических истин. В частности, не экологична и
концепция происхождения жизни в некоем «первичном бульоне»,
который содержит все, что требуется, и в котором (что не менее
таинственно) безболезненно исчезает все, что не требуется. Таким
«бульоном» для каждого организма служит окружающая его при-
рода, но она сама состоит из организмов, умеющих делать то, чего
не умеет данный.
183
Еще Антуан Лавуазье незадолго до казни (1794 г.) писал в
одной из рукописей: «Растения черпают материалы, необходимые
для их организации, в воздухе, который их окружает, в воде и во-
обще в минеральном царстве.
Животные питаются или растениями, или другими животными,
которые, в свою очередь, питались растениями, так что вещества,
из которых они состоят, в конце концов всегда почерпнуты из
воздуха или из минерального царства.
Наконец, брожение, гниение и горение постоянно возвращают
атмосфере и минеральному царству те элементы, которые растения
и животные из них заимствовали» [Лункевич, 1960, т. 2, с. 425].
Ламарк пошел дальше: в 1797 г. он изложил свою концепцию,
в которой, говоря словами Карпова, «считал земную кору продук-
том организмов, их распадом» [Ламарк, 1935, с. СХХШ].
Например, он полагал всякий известняк продуктом распада орга-
низмов, а кварц — продуктом трансформации известняка. В 1799 г.
он опубликовал рассуждение, близкое к приведенному отрывку из
Лавуазье, и добавил, что растения преобразуют материю «с по-
мощью своей жизненной деятельности и импульсов света» [там же,
с. СХ1Х]. Учтя уровень тогдашних знаний (считалось, например,
что растение питается из почвы, беря от солнца только тепло, хотя
формально фотосинтез был открыт в 1779 г. Джозефом Пристли),
можно лишь изумиться глубине мысли Ламарка, увидевшего то,
что мы называем биосферой, то, о чем едва заговорили только с
1860-х годов, когда врач Генри Фрик предпослал своей книге
[Freke, 1861] схему глобального круговорота веществ, когда в
1875 г. геолог Эдуард Зюсс ввел понятие биосферы. А вошло оно
в науку лишь с трудами В. И. Вернадского.
Еще 70 лет назад, когда Вернадский стал размышлять о проис-
хождении жизни, представление о геологической роли земной жиз-
ни было чуждо науке: «Изучая организмы — живую материю,—
оставляют без внимания, как неважное для ее понимания явление,
изменения, совершаемые ими в окружающей их внешней среде»
[Вернадский, 1978, с. 140]. Именно это — изменение организмами
их среды — Вернадский положил в основу своих размышлений о
начале жизни и пришел к парадоксальному выводу: земная жизнь
всегда была в геохимическом отношении такой же, какой мы ее
видим сейчас (живое вещество все время имело приблизительно
одну и ту же массу 1018 г, а каждый химический элемент совершал
в ней тот же тип круговорота, что и сейчас). Это обстоятельство он
назвал «геологической вечностью жизни»; жизнь, приняв свою
современную геохимическую форму, вообще, по Вернадскому, не
эволюционирует в течение известной нам истории — в том смысле,
что круговорот веществ в природе не изменяется при смене кон-
кретных видов организмов.
Казалось бы, что остается делать науке о происхождении жиз-
ни после таких пессимистических выводов? Однако именно после
них здесь начинается интересная работа: загадка происхождения
жизни предстает как загадка становления круговоротов. Так,
184
Г. А. Заварзин считает, что простейшее разнообразие, которое
могло бы выполнять основные геохимические функции жизни
(поддерживать баланс газов в воде и воздухе, отлагать известные
типы осадков и т. д.), в принципе могло бы состоять из реально из-
вестных бактерий. Развитие геологии архея подтверждает: все то,
чем для геохимика обитаемая планета отличается от необитаемой,
сделали уже бактерии.
Поскольку организмы размножаются несравненно быстрее, чем
эволюционируют, то нет смысла говорить о первых организмах как
о нахлебниках, исчерпывавших какой-то первичный бульон,
а следует искать возможный путь рождения жизни в форме ста-
бильных потоков. До появления жизни имелись потоки трех типов:
1) излучения из космоса (прежде всего — солнечное) и в космос (в
основном — тепло), 2) потоки тепла и веществ из недр земли (го-
рообразование, вулканы, источники и т. п.) ив недра (задвига-
ние океанической коры под континенты и т. и.), 3) минеральные
круговороты (например, круговорот воды). Жизнь должна была
включаться в эти потоки — либо в качестве добавочных звеньев
прежних цепей, либо создавая новые цепи (каналы). Так, свобод-
ный водород мог прежде только одно — уходить в космос, тогда как
водородные бактерии открыли ему путь в органический кругово-
рот и, через него, в захоронения (каменный уголь и т. п.). Свобод-
ный кислород возникал прежде только за счет разложения воды
солнечным светом (фотолиза) и расходовался только на окисление
минералов и газов, тогда как фотосинтез обеспечил новый мощный
источник свободного кислорода, создавший нынешнюю атмосферу,
а в ней живут аэробные организмы, в том числе — грибы и
животные.
В 1927 г. почвовед В. Р. Вильямс писал, что биологические кру-
говороты являются малой частью абиотических, что жизнь разви-
лась как «расширяющаяся спираль» на «безжизненном фоне гео-
логических процессов» [Камшилов, 1974, с. 39]. В 1940-х годах
ряд западных ученых (А. Львов, Н. Горовиц, Дж. Бернал и др.) не-
зависимо сформулировали некоторые положения, касающиеся
биосферных аспектов происхождения жизни: задолго до первых
организмов (клеток) возникали «су б витальные области», а затем
«су б витальные единицы», в которых шли соответствующие про-
цессы синтеза и распада, причем прогресс заключался в поэтапном
замещении в них геохимических процессов биогеохимическими;
обособление субвитальных систем на отдельные предорганизмы
(эобионты) и затем на организмы сопровождалось сокращением
возможностей каждого: «современные организмы происходят от
одного или нескольких сверхорганизмов с более широкими возмож-
ностями» [Возникновение жизни..., 1959, с. 51, 56, 84]. Сперва
возник метаболизм, а затем клетки и их размножение [Камшилов,
1974, с. 34].
Тем самым эволюцию жизни естественно делить на два главных
этапа: первый — становление биосферы и потоков (круговоротов),
второй — развитие жизни в пределах уже установившихся био-
185
сферных рамок. По мнению Заварзина, первый этап провели не
эобионты, а прокариоты, второй — эвкариоты. Это мнение согласу-
ется с наблюдаемым сейчас свойством прокариот занимать мар-
гинальные зоны (см. гл. 6); прокариоты как бы предназначены от-
воевывать прежде безжизненную зону, тогда как эвкариоты позже
замещают их или уживаются с ними. Время смены этапов неиз-
вестно, но условно можно отнести его к границе архей—протеро-
зой (2,7 млрд лет назад).
Заварзин считает правдоподобной гипотезу, согласно которой
колыбель жизни — в вулканах, извергающих много органических
молекул, игравших тогда роль неисчерпаемого «первичного буль-
она». Вовлечение органики в круговорот преджизни было возмож-
но только в окисляющей среде, а это отсекает все гипотезы, ос-
нованные на отсутствии окислителя в первичной атмосфере [За-
варзин, 1984]. Если для первой жизни нужен окислитель (вернее
всего — молекулярный кислород), то нужен постоянный меха-
низм, восстанавливающий его запас. Сперва этим механизмом слу-
жил фотолиз воды, затем фотосинтез. Откуда он впервые взялся,
никто объяснить не может, но вероятно, что он, как и другие ос-
новные биологические процессы, послужил органическим вариан-
том реакции, до этого шедшей неорганическим путем [Камши-
лов, 1974, с. 38].
Поскольку такие функции живого, как фотосинтез, дыхание и
т. п., прекрасно выполняют прокариоты, а многие функции живого
(например, усвоение азота из воздуха) до сих пор осуществляют
только они, то жизнь действительно могла долго состоять из про-
кариот, если, разумеется, они были достаточно разнообразны.
Не имея возможности дать здесь цельный рассказ о возникновении
клетки, проанализируем становление только одного механизма —
клеточного деления. На этом примере попробуем увидеть, как зна-
ние законов разнообразия помогает ставить и решать общие проб-
лемы эволюции. (О становлении энергетики, кроме работ, упомя-
нутых в начале главы, см. также: [Брода, 1978].)
8.2. Жизнь до клеток
Все живое создано из клеток, а клетки размножаются делением —
это выглядит банальной истиной, но все же не вполне верно. Труд-
но назвать клеткой такие многоядерные образования, как мицелий,
о котором шла речь в главе 6; бывают и другие не вполне клеточ-
ные организмы, но главное для нас даже не в этом. Деление клет-
ки — цепь невообразимо сложных процессов, и само рождение жиз-
ни стало бы намного понятнее, если бы не надо было воображать
самый первый организм уже делящимся. Самое простое, что вооб-
ще размножается,— вирус, но он не делится.
Всякий вирус состоит из нуклеиновой кислоты, компактной
спиралью уложенной в его сердцевине, и белковой оболочки. В нем
нет ни большинства ферментов, которые бы делали эти его вещест-
ва, ни системы транспорта, которая совершала бы обмен с внеш-
186
ней средой, — словом, всего того, что делает клетку клеткой,
нет в нем, разумеется, и механизма деления. Вирус — не организм,
и чтобы размножиться, должен попасть в клетку, где новые вирусы
синтезирует она сама, считывая информацию с нуклеиновой кисло-
ты вируса. Жизнь явно началась не с вируса, но он полезен при
понимании рождения жизни в другом — демонстрирует нам, на-
сколько различно может быть устроено размножение. Если бы не
вирусы, мы считали бы незыблемой истиной тот факт, что наслед-
ственную информацию может нести только двуспиральная ДНК и
что всякое размножение начинается с ее удвоения. Однако в виру-
сах информация хранится в четырех формах: в ДНК или РНК,
в двуспиральной или одиночной.
Копирование двойной спирали — самая сложная из всех форм
копирования, требующая многих ферментов. Ведь двойную спи-
раль надо сначала расплести, затем сделать копию с каждой поло-
винки (а направления записи и считывания на двух цепях ДНК
противоложны) и снова закрутить, теперь уже две двуспиральные
цепи. Появление такой системы сразу — фантастика. А вот с обыч-
ной одноцепочечной РНК, какая бывает в некоторых вирусах, ко-
пию может делать один-единственный фермент. Причем, а это не
менее важно, по РНК можно и строить основные «кирпичики» —
белки, тогда как с ДНК надо сперва переписать текст на РНК.
Наконец, РНК может служить ферментом. Поэтому для начала на-
до отыскать правдоподобный путь возникновения РНК, что еще
науке не вполне ясно.
Впрочем, даже РНК — не первое, что надо понять. Ведь чтобы
записывать и считывать информацию, надо ее иметь, надо иметь
какой-то жизненный опыт, который имеет смысл хранить и переда-
вать. У современного организма такой опыт всегда связан с белка-
ми (хотя никогда не сводится к одним белкам). Общеизвестное
утверждение «белки — длинные цепи из 20 различных аминоки-
слот, порядок которых считывается с цепи РНК, где закодирован
последовательностью нуклеотидов»,— говорит не все. Кроме 20
канонических аминокислот, кодируемых в РНК, в организмах
встречается еще около 160 аминокислот, иногда образующих це-
почки (таковы многие антибиотики). Эти цепочки (пептиды) син-
тезируются нематричным путем (т. е. не считываются с РНК), они
просто соединяются друг с другом при посредстве ферментов
[Диксон, Уэбб, 1982, т. 3, с. 882]. Многие склонны считать, что
так же синтезировались и предшественники первых белков.
Если так, то во множестве аминокислот естественно выявляется
ядро (канонические формы), бедное разнообразием, но подавляю-
щее количеством, и периферия (остальные 160 форм), до сих пор
не охваченная матричным синтезом.
Первые катализаторы могли быть неорганическими (глины),
затем их дополнили пептиды, а потом их заменили комплексы
пептидов. Затем уже могла произойти «встреча» (см. главу 9) их с
РНК, причем главную роль здесь играла самосборка (напомним,
что и сейчас ее роль огромна: в гене кодируется только последова-
187
тельность аминокислот белка, так сказать — ниточка, а в работаю-
щую машину эта ниточка складывается путем самосборки). Один
из пра-белков должен был хотя бы немного катализировать син-
тез полинуклеотидной цепочки. Другие пра-белки катализировали
распад лишних молекул, который должен быть пространственно от-
делен от зоны синтеза.
Получается, что первое живое образование (эобионт), еще не
обладавшее системой клеточного деления, проще всего представить
себе в виде: РНК плюс примитивное приспособление (пра-фер-
мент) для ее считывания. На этой РНК должна быть записана ка-
кая-то информация, полезная для «выживания» и распростране-
ния этой РНК. Что это за информация, мы можем только гадать,
так как еще не понимаем законов взаимодействия РНК и белков
современных клеток. Хотя синтез белка по РНК-овой матрице дав-
но умеют проводить в пробирке, но никто не знает, каким образом
участники этого процесса (и подобных) находят друг друга в реаль-
ных условиях клетки. В 1970 г. цитолог В. Я. Александров охарак-
теризовал эту ситуацию как «бездну нашего незнания», но только
сейчас на этот пробел знаний начинают обращать внимание. Мож-
но надеяться, что, поняв эти законы, мы сможем говорить и о мини-
мальной сложности систем, способных к их реализации.
От эобионта до самой примитивной клетки еще невообразимо
далеко: у клетки всегда есть оболочка, отделяющая внутреннюю
среду (цитоплазму) от внешней, причем оболочка умеет
избирательно пропускать внутрь все, что требуется цитоплазме для
жизни, и наружу — все, что требуется изъять; в цитоплазме и
оболочке содержится все, что нужно для выработки энергии, син-
теза всех нужных веществ, а наследственная ДНК содержит коды
всех этих веществ и еще — информацию о том, какие синтезы и ра-
спады за какими должны включаться и выключаться. Все это уст-
роено так, что в какой-то момент клетка делится на две (размножа-
ется), и каждая дочерняя клетка получает весь рабочий комплект,
да еще и замыкается собственной оболочкой.
Все без исключения организмы размножаются быстрее, чем
эволюционируют, но эобионту надо было начать не с роста своей
численности (размножения), а с роста разнообразия своих свойств.
Ни один организм не способен на это, зато именно так ведет себя
земная жизнь в целом — она с самого начала присутствует в
единственном экземпляре и все время увеличивает свое разнообра-
зие. Если допустить, что «предок» возник в одном экземпляре,—
это все-таки следует признать меньшим чудом, чем одновременное
появление сбалансированной совокупности организмов,— то его
надо сопоставлять не с организмом, а со всей биосферой. Этот пред-
теча жизни не был клеткой — микроскопической капелькой с набо-
ром всего необходимого внутри и оболочкой снаружи; гораздо
проще все понять, если считать эобионт просто некоторой системой
химических реакций; эта система не была обособлена пространст-
венно, зато была включена в общую систему тогдашней геохимии.
Например, ей нужны аминокислоты, а их, как мы знаем, в массе
188
поставляет неорганическая природа — вулканы, электрические
разряды и т. д.; она же, природа, и изымает аминокислоты, вклю-
чая их в последующие химические реакции, разлагая солнечным
ультрафиолетом, и т. д. Вероятно, сперва эобионт синтезировал
только полимеры-катализаторы, строительные белки, затем стал
собирать блоки памяти — из готовых «кирпичиков» геосферы, а
позже стал делать «кирпичики» сам. Первой могла быть синтезиро-
вана глутаминовая кислота (из нее до сих пор делаются остальные
19 аминокислот), причем сахар и источник азота брались до этого
из геосферы, пока не возникли системы и для их синтеза. Отслу-
жившие молекулы возвращались через системы распада в геохими-
ческое лоно. Не связывая эти системы с капельками, мы не встре-
тим и трудностей в понимании того, как они сопрягались в прост-
ранстве,— они могли переплетаться повсюду, хоть по всему Миро-
вому океану. Еще не разделенный на отдельные объекты, а потому
и не имеющий потребности размножаться, эобионт разрастался
и завоевывал доступное ему пространство, т. е. в некотором смысле
жил. Эту концепцию — эволюцию живого путем поэтапного вклю-
чения реакций, протекавших до этого вне рамок жизни, биохимик
Горовиц назвал в 1945 г. ретроградной эволюцией, т. е. эволюцией,
движущейся вспять. Термин прижился в науке [Брода, 1978, с. 60],
хотя не очень удачен: если что и отступало, то не жизнь, а неорга-
ническая химия Земли, а органическая эволюция все время насту-
пала. Разъединив понятия жизни, размножения и клеточного деле-
ния, мы избавились сразу от нескольких трудностей, так как про-
исхождение жизни перестало казаться просто фантасти-
ческим совпадением счастливых случайностей.
Все гипотезы происхождения жизни затрудняются объяснить,
как возникла клеточная оболочка. Пишут, например, что оболочка
возникла из веществ «первичного бульона», сама плавала в нем и
оказалась выгодной для случайно попавшего внутрь нее эобионта.
Это, конечно, замечательно, но не следует забывать, что там ему и
конец, поскольку он, не будучи скоординирован с синтезом оболоч-
ки химически, не сможет ни расти внутри нее, ни выбраться из
нее, ни управлять ее делением. Чтобы избежать тупика, придумали
допущение, что пузырьки, пленившие эобионтов, могли делиться
под ударами волн прибоя. Это еще остроумнее: деление оболочек
якобы регулярно совпадало с делением их содержимого до тех пор,
пока случайно не был найден способ сопрячь эти деления. Такое
нагромождение случайностей нельзя признать научным. Надо по-
нять химическое сопряжение каждого процесса со всей системой.
По-видимому, эобионт собирал оболочку из внешнего материа-
ла, включая в свою систему новые реакции, до этого протекавшие
в рамках геохимии, и разбирал ее, отдавая этот материал обрат-
но. Позже он, естественно, включил в свою систему и изготовление
упомянутого материала. Эобионт, не будучи изолирован прост-
ранственно, мог, вместо возвращения вещества в геохимическую
систему, передавать его от одной своей части к другой. Например,
он мог использовать материал отслужившей мембраны как источ-
189
ник энергии, т. е. как бы частично съедал сам себя — позже эти
функции отошли к разным организмам. Когда эобионту пришло
время разделиться на разные организмы, среди них были уже про-
образы и растений (продуцентов), и преобразователей (консумен-
тов), и гнилостных бактерий (редуцентов).
8.3. Первые клетки
До клеточного деления надо было научиться размножать генетиче-
ский материал — сперва в виде РНК, а затем ДНК. Зачем вообще
понадобилась ДНК? Ответ (разумеется, гипотетический, как и все,
о чем мы здесь говорим) позволяет дать та же концепция прибли-
зительного баланса первичной жизни с тогдашней геохимией:
включая в состав своей РНК все новый и новый нуклеотидный
материал, эобионт неизбежно был и сам включен в те пути геохи-
мического круговорота, которые разлагали нуклеотидные цепи,-
иначе эобионту скоро не из чего было бы строить новые РНК.
Это означало, что наряду с массовой, избыточной РНК подлежала
уничтожению и включению в геохимический баланс уникальная
РНК, кодировавшая только что найденные эобионтом «изобрете-
ния». От этого и спасло появление ДНК: как ни мало отличие ДНК
от РНК, его достаточно, чтобы ДНК могла быть свернута в более
тугую двойную спираль, чем РНК,— в так называемую В-форму
ДНК. Тем самым ДНК может находиться как в форме, похожей на
РНК (A-форма), и служить для считывания информации с по-
мощью тех же механизмов, какие читают РНК, так и сворачивать-
ся в нечитабельную В-форму. В ней она становилась недоступной
для катализаторов, разлагавших РНК. Первые ДНК, вероятно,
копировались с РНК, создавая тем самым генетическую память,
а репликация ДНК появилась позже. Замечательно, что один и тот
же фермент может вести разные типы матричного синтеза (транск-
рипции) РНК и ДНК [Диксон, Уэбб, 1982, т. 3, с. 825]. Возможно,
это самая древняя из функций ферментов.
Репликация ДНК открыла путь к размножению, но ее первона-
чальная функция, вероятно, была иной: для устойчивости и разви-
тия всякая генетическая система нуждается в «механизме РРР»
(рекомбинации, репарации, репликации); этот механизм нерас-
членим, и репликация вычленяется из него лишь мысленно. Это
мысленное вычленение, подобное вычленению отдельных факто-
ров, является по существу методическим наследием прежнего, ста-
тистического подхода к эволюции, которого мы не раз касались.
При всех его прежних заслугах, в настоящее время он в основном
исчерпан, и приходится признать, что описать эволюцию клетки
как последовательное накопление полезных признаков и свойств не
удалось. В частности, ни одно из трех «Р» механизма РРР нельзя
считать первичным, поскольку перед нами — три функции единого
ферментативного аппарата. Столь же нерасчленимой выглядит
описанная в 6.3 эвкариотность. Стало очевидным, что надо го-
ворить не о накоплении свойств, а о самоорганизации. Ей посвя-
190
щена следующая глава, а сейчас надо завершить эскиз картины ста-
новления клеточного деления.
Деление клетки тоже выглядит нерасчленимым: из простого де-
ления прокариот нельзя изъять ничего. Чтобы начать делиться,
эобионт должен был научиться растить клеточную оболочку, разре-
зать и сшивать ее края, замыкать ее в полость и при этом обеспечи-
вать каждую новую полость копией ДНК. Он мог решить эту проб-
лему, используя созданные им прежде механизмы, собирая их в но-
вые комплексы. Так, механизм поверхностного катализа мог по-
ставлять примитивные мембраны, а механизм РРР — копии ДНК.
Замкнутая полость впервые могла быть собрана не как клетка
(самодостаточная единица, способная к делению), а как органелла,
обеспечившая стабильность внутренней концентрации веществ.
Такие полости возникали в ходе эволюции и позже (митохондрии,
хлоропласты, липосомы и т. д.), демонстрируя реализацию одной и
той же архитектонической возможности в разных условиях. О том,
что липосома могла послужить протоклеткой, пишет Меклер
[19806].
При делении клетки (и про- и эвкариотной) оболочка делит-
ся, словно перетягивая сама себя тугим поясом. Перетяжка сопро-
вождается ростом оболочки — примерно так растет, закрывая от-
верстие, диафрагма фотоаппарата. Когда оба пузырька готовы, они
сами собой «зашиваются» (диафрагма схлопывается) и уплывают
в разные стороны. К началу деления прокариотной клетки ДНК
уже удвоена, обе двуспиральные копии прикреплены к одной точке
«экватора» клетки, и каждая копия увозится соответствующим
участком растущей в обе стороны оболочки. Как она растет, еще не
вполне известно, но это тоже самосборка.
Прежде, когда эволюционные теории строились на материале
геометрических форм крупных животных, а клетку (позже —
клеточную органеллу) рассматривали как бесструктурный «ко-
мочек протоплазмы», эволюцию удавалось описывать как постепен-
ное улучшение формы и (это считалось самоочевидным следстви-
ем) заданной функции. Казалось, что так же можно объяснить
и эволюцию клетки, но теперь мы знаем, что субмикроструктуры не
проще, а зачастую сложнее других структур. Поэтому характер эво-
люционных объяснений понемногу изменился: вместо сдвигов па-
раметров и деформаций органов чаще говорят о заменах и вставках
целых органелл и реакций (подробнее см.: [Чайковский, 19776,
с. 824, 826]). Фактически речь идет при этом всегда о самосборке.
Ее простейшим для понимания вариантом является генетическая
интеграция: за объединением двух генетических текстов, прежде
разрозненных, следует объединение произведенных ими элементов
в единый механизм.
Генетический материал, подвергнутый интеграции чужерод-
ным геномом, всегда используется не совсем так, как в прежнем
геноме, т. е. происходит смена его функционирования; но эта
смена не может сводиться к одной лишь модификации синтезируе-
мых молекул — главное в том, что новые молекулы должны быть
191
собраны вместе со старыми в новый целесообразный механизм.
Интегрируемый генетический материал должен «уметь изменить
свое поведение» вследствие контакта с новым геномом. Следова-
тельно, интегрироваться могут только целые авторегуляционные
системы. Само возникновение феномена авторегуляции (как и са-
моорганизации вообще) не объясняется в терминах интеграции и
будет обсуждаться в следующих главах. Здесь же отметим только,
что для понимания становления эвкариотной организации прихо-
дится привлекать те же понятия (самосборки, интеграции и т. и.),
что и для объяснения происхождения клетки вообще. Дело в том,
что основные эвкариотные механизмы можно мысленно сложить из
прокариотных элементов.
Сразу вслед за открытием (1882 г.) у низшей водоросли явле-
ния деления хлоропластов ряд крупных фитофизиологов (в том чи-
сле и Фаминцын) стали проводить параллель между хлоропласта-
ми растительных клеток и прокариотными водорослями (цианея-
ми), что дало толчок для развития теории симбиогенеза. Теория
утверждала, что безъядерные клетки (в нынешних терминах —
прокариоты) следует рассматривать, вместе с такими органеллами
ядерных клеток, как хлоропласты, в качестве элементарных орга-
низмов — биобластов. (Вскоре к ним отнесли митохондрии и
другие органеллы, включая даже ядро клетки.) Тем самым эвка-
риотная клетка трактовалась как система интегрированных био-
бластов. Затем идеи интеграции клеточных структур были призна-
ны не соответствующими данным цитологии и были надолго забыты,
но породившие их вопросы остались без ответа — это вопрос о при-
чинах сходств органелл друг с другом и с прокариотами и вопрос
о том, как объяснить генезис основных механизмов клетки.
В 1960-е годы интерес к интеграции снова возрос, но уже на основе
данных генетики микроорганизмов и электронной микроскопии.
Кроме симбиогенеза (признающего вторжение в клетку и сох-
ранение навсегда в ней целых прокариотных организмов) рассмат-
риваются различные другие гипотезы интеграции. По-видимому,
органеллы — результат последовательности разнородных актов
интеграции, а не только симбиогенеза. Очевидно одно: эвкариот-
ность связана с появлением нового принципа организации. Броса-
ется в глаза полирепликонностъ эвкариотной клетки: если у про-
кариот ДНК представлена единым кольцом, в котором репликация
осуществляется единым актом (начинается в точке крепления ДНК
к оболочке клетки и в ней же кончается), то в эвкариотной клетке
всегда несколько хромосом, каждая из которых состоит из многих
единиц репликации {репликонов). Поэтому у прокариот генетиче-
ская интеграция состоит либо во включении генома вирусов (и
подобных им частиц) в кольцо ДНК, либо просто в сохранении
их внутри клетки. Для эвкариот же характерна интеграция путем
образования новых репликонов (в том числе новых хромосом и
органелл, содержащих ДНК). При этом эвкариотные репликоны
короче прокариотных и (за исключением некоторых динофла-
геллат) сходны с репликонами вирусов, а часть генетической ин-
192
формации содержится в органеллах — митохондриях и хлоропла-
стах, хотя ДНК органелл не кодирует никакой или почти никакой
самостоятельной информации. Тем самым принцип эвкариотной
организации выступает как принцип прогрессивной децентрали-
зации генетического управления.
Очевидно, что этот принцип открывает и новые, чисто интегра-
ционные возможности: не случайно эвкариоты (в отличие от про-
кариот) имеют эндосимбионтов — эвкариот умеет координировать
свою деятельность с деятельностью эндосимбионта, что открывает
принципиальный путь и для симбиогенеза. В прокариоте, напро-
тив, по-видимому, может жить только вирус, т. е. объект, не явля-
ющийся генетической системой. Теперь можно переосмыслить
старую концепцию биобластов как концепцию полуавтономных
репликонов, способных к перегруппировке. Таксоны, которые
«изобрели» децентрализацию, не смогли остановиться на полпути
и все приобрели комплекс эвкариотности целиком, как нечто нерас-
членимое (6.3), зато утратили идиотрофность, о которой шла речь в
6.5. Первый шаг к эвкариотности легче всего представить как появ-
ление у клетки многих репликонов. Нечто сходное мы видим иног-
да в нынешних микоплазмах, где, кроме плазмид и вирусов, иног-
да наблюдается мультигеномность [Борхсениус, Чернова, 1987а,
с. 380].
Как говорилось в 6.3, обнаружение некоторых примитивных
черт митоза у динофлагеллат повлекло идею считать их переход-
ной группой, «мезокариотами». Идея не выдержала критики с
позиций диатропики, однако сами по себе примитивные черты
любопытны: так, разъединение сестринских хромосом динофлагел-
лат связано с раздвижением оболочки ядра [Bhaud, Soyer-Gobil-
lard, 1986, с. 24], словно бы это не ядро эвкариота, а прокариот-
ная клетка. Другой этап эволюции наблюдается у некоторых эв-
глен: митоз похож на динофлагеллатный, но хромосомы — обыч-
ные для всех эвкариот (не рыхлые). Третий этап — появление
дыр в ядерной оболочке, а затем и полное растворение ее в ходе
митоза (кариолиз). Таким образом можно поэтапно «собрать»
весь типичный митоз, но беда в том, что, во-первых, обладатели
примитивных свойств очень далеки таксономически (относятся к
разным типам, а иногда и царствам), а, во-вторых, они никак не
выстраиваются в ряд от простой формы к сложной. Например, один
из самых примитивных митозов (водоросль Chroomonas) обладает
кариолизом [Oakley, Dodge, 1973], тогда как многие группы одно-
клеточных обладают митозом, типичным во всех отношениях за
исключением отсутствия кариолиза; багрянки (красные водорос-
ли), обладая почти типичным митозом, сохранили ряд прокариот-
ных пигментов и других примитивных черт.
Зная рефренную структуру разнообразий, этому удивляться не
приходится. Лучше интересоваться не рядом последовательных
организмов с разными митозами, а прямо — преобразованием са-
мих рассматриваемых нами стуктур и функций (т. е. в терминах
7.3 — не филогениями таксонов, а семофилогениями меронов).
7 Ю. В. Чайковский
193
У эволюционистов принято считать, что происхождение понято, ес-
ли указано, кто от кого произошел, хотя от такого указания может
быть далеко до понимания сути процессов. Так, давно все согласны,
что ядовитая железа змей произошла от слюнной, но откуда взялся
яд, как он синтезируется и действует, яснее не стало; этот вопрос
надо решать отдельно, в других терминах. Мы же, наоборот, знаем,
из каких элементов складывался митоз, но не можем сказать, в ка-
кой последовательности и у каких организмов эти элементы поя-
вились. А может быть, это и не нужно? Ведь наблюдаемые орга-
низмы все — нынешние; а у тех подлинных, что впервые отрабаты-
вали варианты митоза два миллиарда лет назад, комбинации
свойств все равно были другими и комбинировались, надо полагать,
так же свободно, как у нынешних. Зато, зная разнообразие нынеш-
них форм, можно выявить независимые элементы и мысленно соб-
рать их в нынешний механизм.
В примитивном митозе ядро делится подобно бактерии, следова-
тельно, здесь мог работать уже известный бактериальный меха-
низм; примитивные хромосомы сходны с пучком бактериальных
ДНК, следовательно, и их могли поставить бактерии; от них же —
бактериальный принцип разъединения хромосом в ходе примитив-
ного митоза. От вирусов могла быть заимствована плотная упаков-
ка ДНК в хромосомах. Даже при самом примитивном митозе уда-
ется выявить несколько ниточек — по одной от полюса деления к
хромосоме; они же, по-видимому, служат для ориентации деляще-
гося ядра в делящейся клетке. В электронном микроскопе видно,
что каждая ниточка — полая внутри, т. е. является микротрубоч-
кой. Микротрубочек в мире бактерий не найдено, но более простые
и рыхлые структуры, похожие на трубочки, описаны в 1975 г.
у цианей и т. д.
Таким путем можно мысленно «собрать» деление ядерных кле-
ток из доядерных структур так, что не придется предполагать
таинственного одновременного появления сразу многих новых
структурных «кирпичиков». Этот чисто биологический результат
достигнут путем смены методологической установки — отказом от
филогенетического метода в пользу семофилогенетического.
8.4. Конец «центральной догмы»
и новая познавательная модель
Известные химики, обсуждая происхождение жизни, задали воп-
рос: «Почему существуют миллионы видов животных и растений,
несмотря на то что молекулярный аппарат клетки в своей основе
одинаков...?» [Эйген, Шустер, 1982, с. 15]. Сам вопрос — диатро-
пический, но поставлен неверно: разнообразие есть и в «молеку-
лярном аппарате клетки», только плохо пока изучено. Достаточно
отметить, что тимин — одно из четырех нуклеотидных основа-
ний — иногда заменен в ДНК на урацил, а иногда — на оксиме-
тилурацил [Кордюм, 1982, с. 61]. Последний обычен у динофла-
геллат, которые вариабельны во всем, хотя и не старше, чем
194
другие одноклеточные, и это наводит на мысль, что рефрен генети-
ческого кода пока целиком не выявлен. Еще примечательнее
данный химиками ответ: дело якобы в различии процедур отбора —
если у организмов он мягок, то у эобионтов был жестким, действо-
вал «раз и навсегда», и закрепившийся эобионт «не может быть
заменен» [Эйген, Шустер, 1982, с. 177]. Откуда это следует? По-
чему здесь так, а там иначе? Подобные вопросы интересуют авторов
не более, чем древних мифотворцев. Больше того, как мягкий,
так и жесткий отбор «выведены» ими из одной и той же, по сути,
формулы, только для эобионтов она истолкована буквально, а для
организмов — расплывчато. Те же авторы заявляют: принцип ес-
тественного отбора «является единственным понятным нам спосо-
бом создания информации» [с. 28], хотя далее справедливо при-
знают, что отбор лишь сохраняет информацию. Написано это через
год после получения Нобелевской премии Ильей Пригожиным
за описание принципа самоорганизации в диссипативных структу-
рах (альтернативного прежнему принципу естественного отбора),
причем эти работы нашим авторам формально известны, ибо на них
есть ссылка. Почему же принцип отбора — «единственный»?
А вот совсем другой пример: блестящий знаток разнообразия
митозов пишет, что у нынешних одноклеточных эвкариот «мы
имеем дело с отражением «периода проб и ошибок» в эволюции,
когда шли поиски наиболее эффективного митотического механиз-
ма», каковой мы видим у многоклеточных в форме закрепленного
отбором «наиболее эффективного и энергетически выгодного ва-
рианта митоза» [Райков И. Б., 1986, с. 53]. Разумеется, даже
не поставлен вопрос о том, что такое эффективность митоза,
чем она отлична от энергетической выгодности и как это можно бы
измерить, однако вывод для автора бесспорен, ибо он соответст-
вует той же умственной моде «проб и ошибок». Эффективность
примитивных форм митоза в действительности ничуть не низка:
их обладатели процветают среди водорослей и паразитов, а дино-
флагеллаты, появившись в мезозое, даже потеснили более древние
формы, имевшие, надо полагать, более сложный митоз.
Эти несообразности не стоили бы внимания, если бы не их
типичность для нынешней литературы по эволюции. Ни один
натуралист при анализе конкретных результатов (теоретических
или опытных) не позволит себе подобной небрежности в ссылках,
понятиях или логике, но едва речь заходит об эволюции, обычные
требования вдруг отпадают. Те, кто «за Дарвина», обычно считают
возможным ссылаться на любой аргумент, если и он «за Дарвина»,
как на истину, не заботясь даже о смысле используемых слов.
Не многим лучше строятся и тексты «против Дарвина». Оппоненты
попросту иллюстрируют свою априорную уверенность (например,
что всякое развитие целиком идет «путем проб и ошибок» [Эйген,
Шустер, 1982, с. 17]), а вовсе не выясняют, что происходит
в природе. Аналогична ситуация при обсуждении эволюции Сол-
нечной системы [Чайковский, 19876].
Очевидно, что эволюционизм пользуется не теми познаватель-
7*
195
ними установками, что остальное естествознание, и установки эти
естественно, по-моему, связать с самой старой познавательной
моделью (см. 1.2), полагавшей авторитет текста выше, чем автори-
тет фактов, отмеченных в природе. Еще полнее представлена
в эволюционизме вторая (механическая) модель, которую лучше
всего выразила «центральная догма молекулярной биологии», гос-
подствовавшая еще совсем недавно.
Согласно ей, ДНК служит матрицей для себя и для РНК,
а РНК — матрицей для белка; хотя с самого начала было известно,
что синтез матриц и их работу осуществляют белки, влияние
их на текст матриц отрицалось, и «догму» записывали формулой:
ДНК^ДНК-*РНК->белок.
В такой схеме эволюция могла быть связана только с самопроиз-
вольным изменением ДНК, откуда и отмеченная выше мода на
«пробы и ошибки» — то новое, что привнесла в механическое пони-
мание эволюции третья (статистическая) модель. Наоборот, авто-
ры, стоявшие на системных позициях, видели в ДНК равноправ-
ный элемент генетической системы. Так, ламаркист Поль Вентре-
бер, отметив полную «иммутабильность» ДНК в схеме молекуляр-
ных биологов, протестовал против идеи первичности генов и вто-
ричности всего прочего. Он писал: «Ферменты в клетке сами себя
синтезируют под контролем генов», тогда как сам ген — всего лишь
«продукт, сотворенный живой материей, ее делегат в хромосомах...
сохраняемый и используемый, где и когда понадобится» [Wintre-
bert, 1962, с. 154, 156, 183, 138]. Если так, то все стрелки формулы
должны быть обратимы. Это и было впоследствии доказано.
Здесь главную роль сыграл вирусолог Дейвид Балтимор: в
1970 г. он был одним из троих, открывших процесс РНК—>-ДНК
(его назвали обратной транскрипцией, хотя в 8.3 мы видели, что
он вероятно возник до процесса ДНК—>-РНК), а в 1982 г. ему же
посчастливилось быть в паре открывших нематричный синтез
ДНК. Оказывается, в ходе синтеза гена, кодирующего иммуногло-
булин (белок, атакующий чужеродное тело и тем, в частности,
обеспечивающий позвоночным устойчивость к инфекции), в текст
ДНК встраивается небольшой (8 нуклеотидных пар) фрагмент,
никакой матрицей не кодируемый [Alt, Baltimore, 1982]. (На эту
замечательную работу мое внимание любезно обратила
Т. И. Ульянкина, историк иммунологии.)
Открытие позволило вскоре решить давнюю загадку — каким
образом иммунная система мгновенно находит ответ на воздействия,
никогда ранее в истории вида не встречавшиеся. За открытие
«генетического принципа обеспечения разнообразия антител»
иммунолог Сусуму Тонегава получил Нобелевскую премию [То-
negawa, 1988], а сам нематричный синтез оказался свойством,
общим для самых различных организмов [Clark, 1988]. Если
учесть, что принцип молекулярного узнавания, открытый на имму-
ноглобулинах позвоночных, обеспечивает также межклеточные
взаимодействия при формировании зародышей [Эдельман, 1989],
196
то станет ясно: процесс белок—>-ДНК — один из главных в природе.
И увидеть его долго мешала «центральная догма».
Почему так произошло? Думаю, что виной были устаревшие
познавательные модели. Процесс белок—«-ДПК отрицали потому,
что стрелку хотели понимать только как считывание текста с мат-
рицы, а белок матрицей не является. Верный ответ был получен
в системных понятиях, но хочу отметить, что постановка задачи
здесь — диатропическая: разнообразие иммунных ответов должно
соответствовать разнообразию чужеродных воздействий. Рассмот-
рим пример, когда системный подход тоже завел исследователей
в тупик, поскольку требовался подход диатропический.
Цитолог Линн Маргелис, возродившая в 1969 г. идею симбио-
генеза эвкариотной клетки, связывала ее со становлением митоза:
веретеном деления якобы управляет унаследованный клеткой про-
кариот. Опыты опровергли эту гипотезу, Маргелис признала ее
несостоятельность [Margulis, 1973], а затем выступила с тради-
ционной (без симбиогенеза) концепцией эволюции митоза
[Margulis, 1974]. Одновременно симбиогенез был раскритикован
цитологами и по остальным позициям (литературу см.: [Чай-
ковский, 19776; 1986]). Стало ясно, что суть эвкариотной органи-
зации так объяснить нельзя, да в сущности этой цели никто и не
ставил. Симбиогенезом остроумно объясняется ряд сходств прока-
риот и эвкариот, однако даже не ставится вопроса о причине
различий, не говоря уже о рассмотрении эволюции клетки как
целого. Тем не менее, саму идею эволюции путем слияния малых
систем в большую надо отнести к системной познавательной мо-
дели.
Эволюционная парадигма, утверждающая, что всякое сходство
либо унаследовано от общего предка, либо приобретено независи-
мым приспособлением к сходным условиям, не нашла иного
объяснения факту сложности эвкариотных структур, кроме как
через симбиогенез. Он вошел в моду помймо воли Маргелис, и вот
она, никак не объяснив своего прежнего пессимизма, назвала
симбиогенез' «точной и проверяемой гипотезой» [Margulis, 1975,
с. 267], а затем вновь положила его в основу концепции эволюции
клетки [Маргелис, 1983], хотя теперь это — лишь набор очень
частных гипотез, объясняемых через другие гипотезы, а отнюдь
не «проверяемых».
По Лакатосу, всякая исследовательская программа со време-
нем вырождается: она «дает только запоздалые объяснения либо
случайных открытий, либо фактов, предвосхищаемых и откры-
ваемых конкурирующей программой»; тем не менее опровержения
«не могут одним ударом уничтожить исследовательскую програм-
му» [Структура..., 1978, с. 220, 222]. Продолжая существовать на
правах парадигмы, вырожденная программа долго еще может
казаться обществу серьезной за счет решения искусственных задач,
подбираемых под рутинные методы. Вырождение дарвинизма по-
родило целую дисциплину — математическую генетику популя-
ций, о которой ведущий специалист пишет: «В бункеры тео-
197
рии-машины в изобилии посыпались факты. Но на выходе не поя-
вилось... ничего!» [Левонтин, 1978, с. 194]. На явные черты вырож-
дения традиционной программы эволюционизма постоянно ука-
зывал Любищев, говоря, в частности, что в ее пользу собран
«Монблан фактов», но против нее говорят «Гималаи фактов».
Вместо цельной научной картины эволюции налицо множество
разрозненных объяснений разрозненных фактов, в отношении ко-
торых «есть возможность найти какие-то фразы, которые будут
имитировать функциональное объяснение» [Меуеп, 1987, с. 367].
Потребность в общей концепции чувствовалась давно, и многие
по-разному выражали ее (энтелехия Дриша, жизненный порыв
Бергсона, тектология Богданова, общая теория систем Людвига
Берталанфи и др.), но к нашей теме ближе всего (кроме рассмот-
ренных выше взглядов) новя эволюционная концепция, связанная
с нелинейной термодинамикой, оформившаяся в 1970-е годы и
привлекшая внимание методологов в 1980-е годы.
Суть эвкариотной организации не может быть раскрыта в чисто
системных рамках, поскольку, как это показывает материал гла-
вы 6, здесь требуется прежде всего уяснить рефренную структуру
клеточных разнообразий. Сходства и различия внутриклеточных
структур настолько перемешаны, что никакая физическая интегра-
ция предков ничего здесь не объяснит; однако рефрены видны
совершенно четко: хлоропласты образуют параллельный ряд
свойств с цианеями и т. д. Новая система взглядов неминуемо
будет связана с диатропикой, но, надеюсь, диатропика никогда
не станет парадигмой, ибо парадигмой Томас Кун назвал господст-
вующую точку зрения, претендующую на единственность и объек-
тивность. Сама идея разнообразия противоречит идее парадигмы,
о чем уже не раз писалось [Фейерабенд, 1986, с. 110].
Вообще же познавательная модель по Огурцову, насколько я
ее понимаю, отличается от парадигмы по Куну и темы по Холтону
своей всепроникающей междисциплинарностью. Ее можно считать
общенаучной макротемой; если в одной физике Холтон [1981,
с. 27] насчитывает десятки тем, то моделей я вижу только пять,
и вряд ли их можно увидеть намного больше. В отличие от пара-
дигмы моделью можно назвать систему взглядов, не претендующую
на единственность и даже не осознаваемую научным сообществом.
Проникая в дисциплину, модель может порождать в ней исследо-
вательскую программу по Лакатосу, причем одна программа может
использовать две модели: так, теория газов Максвелла — Больцмана
была сразу и механической, и статистической и дала физике
программу, получившую название статистической механики. Та-
кой же программой в биологии был дарвинизм, позволивший
ставить и решать новые задачи (например, эволюционное переос-
мысление палеонтологии дало ей мощный импульс к развитию)
[Чайковский, 1990].
198
Глава 9. СВЯЗЬ ИДЕЙ ЭВОЛЮЦИОННОЙ ТЕРМОДИНАМИКИ
И ДИАТРОПИКИ
9.1. Статус эволюционной термодинамики
Поразительно, что сто лет физика и биология радикально противо-
речили друг другу, и это мало кого волновало. Согласно класси-
ческой термодинамике (ее теперь чаще называют термостатикой).
устойчиво только состояние хаоса, тогда как какое-либо упорядо-
чение может идти исключительно с помощью внешнего воздейст-
вия. В качестве внешнего управления биологи указывали естест-
венный отбор, посредством которого живая природа непрерывно
сама собой упорядочивается и усложняется, все более удаляясь
от того молекулярного хаоса, в котором, по их представлениям,
она родилась. Мы уже говорили не раз о том, что отбор — фактор
маломощный (в отношении мелких редких изменений). Однако,
даже если верить в его силу, он все-таки должен оставаться
внешним фактором, т. е. может только действовать на вид со
стороны заданной среды.. А какой же тогда фактор усложняет
биосферу как целое? Ведь она одна-единственная и извне не управ-
ляется. Извне она получает только потоки веществ и энергии,
в общем одни и те же во все эпохи, а вовсе не информацию
о нынешних путях усложнения.
В свое время Джеймс Максвелл предложил мысленный экспе-
римент: пусть крохотный «демон» производит «отбор» быстрых
молекул, открывая перед ними дверцу сосуда и закрывая ее перед
остальными молекулами — тогда газ в сосуде будет нагреваться
без затраты энергии извне. Позже было доказано, что «демон
Максвелла» — самообман: на распознавание быстрых молекул
надо затратить не меньше энергии, чем на нагревание газа.
В 1964 г. Айзек Азимов остроумно назвал естественный отбор
«демоном Дарвина» — за (столь же подозрительную) способность
повышать сложность организации без притока внешней информа-
ции. В частности, творцов новой термодинамики озадачивал тот
факт, что «сам Дарвин объяснял необратимую эволюцию к более
организованным структурам в живых системах через естественный
отбор», который «благоприятствует исключительным ситуациям,...
т. е. организованные структуры становятся правилом в биологии.
Однако эти общие аргументы не могут помочь в решении проблемы.
Хотелось бы быть более точным и объяснить явления, исходя
из определенных физических законов» [Пригожин, Николис, 1973,
с. 519].
На этом пути были достигнуты блестящие результаты. Оказа-
лось, что хотя все выводы термостатики были корректны, все же
произошло досадное смещение акцентов, неверно ориентировавшее
ученый мир и общество. Сто лет считалось азбучной истиной,
что миром правит закон возрастания энтропии (меры беспорядка),
тогда как он верен лишь при довольно жестких условиях, а именно
199
вблизи термостатического равновесия и (что уж совсем нереально)
при отсутствии силовых полей (К. Э. Циолковский называл
гравитацию главным антиэнтропийным фактором). Если система
далека от равновесия, (например, находится в потоке) и притом
описывается существенно нелинейными уравнениями, то в ней
могут сами собой образовываться упорядоченные структуры. Их
называют диссипативными структурами (ДС), поскольку они
существуют за счет рассеяния (диссипации) внешней энергии в
окружающей данную систему среде.
Простейшая ДС возникает в ванне с водой, когда вынута
пробка. Хотя самопроизвольное движение воды по кругу невероят-
но, но при достаточно мощном стоке вода закручивается, причем
водоворот имеет свои простые онтогенез (воронка формируется
постепенно) и память (закрутившись влево, не изменит направле-
ния) . ДС немыслима без движения, она существует как побочный
результат потока. Она вбирает в себя элементы до этого друг другу
чуждые, тем самым в ней реализуется сродство частей к целому.
Заслугой школы Пригожина была демонстрация общности феноме-
на ДС (механические, тепловые, химические процессы, а также их
аналогия с биологическими).
Суммарная энтропия всякой системы растет (второй принцип
термостатики сохраняет силу и в термодинамике). Человек, напри-
мер, для поддержания своих 70 кг тела разрушает за жизнь
около 20 тонн органики, а саму органику создают на Земле расте-
ния, рассеивающие для этого в тепло солнечную энергию столь же
расточительно (при фотоситезе в химическую энергию переходит
менее 3% солнечной энергии). Тем самым усложнение (градация)
структур — не отбор «исключительных ситуаций», а побочный
эффект термодинамического (но не термостатического) процесса
деградации солнечной энергии и (в меньшей мере) веществ
Земли.
Коль скоро Земля пребывает в потоке солнечной энергии, а
уравнения взаимодействия частиц существенно нелинейны, то обя-
зательно будет то тут, то там происходить самосборка — сперва
молекул, затем их комплексов, затем более сложных структур,
вплоть до живых — вот идеология новой термодинамики в вопросе
начала жизни. Тем самым жизнь возникла вовсе не из «молекуляр-
ного хаоса», поскольку такового в чистом виде никогда не было —
среди «хаоса» всегда были ДС. Коль скоро законы самосборки
достаточно общи, то нечего удивляться параллелизму, так как
сходные процессы должны порождать и сходные структуры — вот
первый вклад термодинамики в проблематику разнообразия. Одна-
ко важно понять не только то, почему организмы сходны, но и
почему они различны более, чем условия их существования. Есть
соображения, по которым рост разнообразия видов вполне естест-
вен — как следствие общего роста энтропии в системе из многих
ДС; видоообразование в подобных построениях рассматривается
как нечто вроде фазового перехода (замерзания воды и т. п.),
а такие переходы возможны только скачком [Lewin, 1982].
200
Сразу отметим, что все это — вольные аналогии, а не доказа-
тельства: новая термодинамика разработана для узкого круга физи-
ко-химических систем, где самоорганизация выражается в очень
простых явлениях — устойчивых колебаниях, ячеистой структуре
и т. п. Многие даже протестуют против самой идеи сопостав-
лять ДС с организмами — например, потому, что суть живого не
в диссипации, а «в избыточно-анаболическом сопротивлении дисси-
пации» [Аршавский, 1986, с. 103]. Однако мы только что видели,
что это сопротивление диссипации внутренней идет за счет мощной
диссипации внешней. Тем самым, даже простые .аналогии очень
полезны, поскольку впервые показали, каким путем возможна
самоорганизация без «демонов». При этом возможны интересные
обобщения.
Неизбежно не только само возникновение структур, но и их
градация (обычно ее неудачно называют иерархией): по мере нако-
пления в сложной системе многих ДС одного уровня сложности,
они сами становятся элементами новой, более сложной системы,
образуются более крупные ДС. Однако здесь встает одна принци-
пиальная трудность: чем выше уровень ДС, тем сложнее должна
быть та первичная «затравка», которая сможет породить переход
к новому уровню, начать макроскопический процесс структуро-
образования. Этот «надкритический зародыш флуктуации» [Ни-
колис, Пригожин, 1979, с. 477] реализуется все труднее, так что
процесс усложнения все менее может быть назван самопроизволь-
ным. Парадоксальным образом то, что обычно именуется самоорга-
низацией, требует внешнего запуска или инструктирования. Не
меньшее затруднение возникло и при попытке указать хотя бы
гипотетический путь, каким усложняющиеся системы могут стано-
виться приспособленными к своей среде обитания.
Вероятно, что именно ввиду этих трудностей эволюционная
термодинамика в ее первом, пригожинском, варианте (1972) не
привлекла биологов. Если говорить языком методологов, она не
оказалась исследовательской программой. К тому же она страдала
явно излишним редукционизмом — пыталась свести биологиче-
скую проблему не просто к физико-химическим задачам, но к зада-
чам квазиравновесным. Дело в том, что до сих пор сами термоди-
намические процессы описываются, в основном, терминами термо-
статики, в силу чего процесс выступает как отклонение от
равновесия. В частности, центральное понятие — энтропия —
сформулировано для газа, находящегося в термостатическом рав-
новесии. Даже основной результат новой термодинамики — «уни-
версальный критерий эволюции», из которого следует существова-
ние ДС, — основан на достаточно туманном постулате о
«локальном равновесии», которое должно иметь место внутри каж-
дой макроскопической подсистемы исследуемой системы [Глен-
сдорф, Пригожин, 1973, с. 109]. И хотя трудно согласиться с тем
мнением, что «редукционизм Пригожина—Эйгена» является
«ошибкой века» [Trincher, 1984], поскольку подход группы При-
гожина представляется много более реалистичным, чем группы
201
Эйгена, все же следует признать, что настоящая эволюция — это
движение как таковое (не выразимое в равновесных терминах),
а вовсе не отклонение от равновесия. Здесь была нужна более об-
щая точка зрения, и с ней выступил в 1975 г. системолог Эрих
Янч.
Обычно эволюцию системы изучают после того, как сама систе-
ма описана, а Янч начал с предложения описывать изучаемую
систему через ее развитие. Западная наука, писал он, привыкла
исходить из неподвижных схем как чего-то исконного, строить
всякое движение как мысленную последовательность состояний,
тогда как естественнее считать движение первичным, а всякое
состояние — как мысленно остановленное движение. Традиционная
динамика, строящая движение как последовательность кинокадров
(аналогия принадлежит Бергсону), бессильна понять, почему
движение может быть направлено к некоторой цели, и поэтому
вынуждена ссылаться на внешние влияния: либо в форме отбора,
либо — управления, либо — замысла. Все эти ссылки Янч уподоб-
лял попытке Мюнхаузена вытащить самого себя за волосы из
болота (ср. с трилеммой Мюнхаузена из 6.1): если у барона не
было механической опоры, то здесь — системной. Наоборот, си-
стемная динамика, считая понятие движения первичным, легче
приводит к пониманию работы и цели систем, как велосипед легче
удержать в равновесии, когда он едет. Направленность движения
удивительна только для стороннего наблюдателя, но вполне есте-
ственна для наблюдателя, движущегося вместе с системой [Jantsch,
1975]. На данном пути, как мы увидим в 9.4, эволюционизм
впервые получает надежду подойти к проблеме приспособления,
не пользуясь услугами «демонов».
Эту философскую схему и призвана иллюстрировать термоди-
намика. Если школе Пригожина свойственно надеяться, что ее
уравнения когда-то окажутся достаточными для описания сути
биологии, то Янчу эта иллюзия чужда. Он ясно понимал, что
квазиравновесный метод термодинамики принадлежит «кинемато-
графической» традиции, что феномен ДС — всего лишь частная
иллюстрация феномена самоорганизации, пусть и самая на сегодня
понятная. По-моему, роль ДС в понимании процессов жизни такова
же, какова роль вычислительных машин в понимании мышления:
пусть машины и не мыслят, но аналогия мозга с ЭВМ на порядок
выше, чем его аналогия с телефонной станцией (а та была выше
аналогии с часами). Аналогия с ЭВМ выявила роль информацион-
ных процессов, а аналогия жизни с ДС выявляет в самоорганизации
роль диссипации. Это на порядок выше, чем прежняя (термоста-
тическая) аналогия мира и организма с тепловой машиной. Она
была глубоко пессимистична (прогнозировала тепловую смерть),
а свою аналогию Янч считал оптимистичной, ибо она обосновывает
не распад, а самоорганизацию.
Сто лет назад дарвинизм соответствовал статистическому взгля-
ду ученых и общества на мир, давал эволюционизму познаватель-
ную модель статистического характера, а теперь термодинамика
202
дает эволюционизму новую познавательную модель системного
характера. В 8.4 говорилось, что дарвинизм дал эволюционизму
и исследовательскую программу, ныне исчерпанную. Дает ли
новую программу эволюционная термодинамика? Для ответа доста-
точно привести примеры новых содержательных задач, которые без
термодинамики эволюционизм ставить не мог.
Вот первый из них: если биосфера — ДС, то может ли она
развиваться без деградации планеты? Нынешняя термодинамика
полагает, что ДС могут возникать только в открытых системах
(системах, которые обмениваются с окружающей средой как
энергией, так и веществом); наоборот, образование структур в
закрытых системах (лишь энергетический обмен со средой) отри-
цается [Николис, Пригожин, 1979, с. 13]. Насколько это отрицание
обосновано, нам не так важно, поскольку биосфера — существенно
открытая система: непрерывно расходует одни ископаемые и соз-
дает другие. Зато очень существен вопрос о необходимом соотно-
шении потоков вещества и энергии через ДС. Вызывающее тревогу
разрушение биосферы человеком — лишь ускоренная модель необ-
ратимого воздействия самой биосферы на планету, и следует
выяснить, есть ли принципиальная возможность понизить дисси-
пацию веществ за счет более эффективного использования дисси-
пации солнечной энергии. Вопрос о минимальном потоке веществ
через систему, обеспечивающий существование ДС, аналогичен
вопросу о максимальной эффективности тепловой машины, кото-
рый поставил и решил Сади Карно на заре термостатики. Если
диссипацию веществ можно довести до уровня, обеспечивающего
подстройку к геологическим ритмам, то жизнь на Земле сможет
существовать столько, сколько будет продолжаться активность
недр и светить Солнце. Если же нельзя, то мечтам Вернадского
о «геологической вечности жизни», основанной на якобы замкну-
тых циклах, и, возможно, о ноосфере (сфере жизни, основанной
на разумном управлении биосферой) не суждено сбыться, но
остается задача о максимальном продлении жизни на Земле. Таким
'образом, вопрос о ноосфере является вопросом эволюционной
термодинамики. Ее другие задачи рассматриваются далее.
9.2. Статистика и системность в эволюционизме
С развитием общества роль статистики не падает, а растет,
причем система балансов (например, в экономической статистике)
усложняется и совершенствуется. Поэтому естественно и в систем-
ном эволюционизме ожидать возрастания роли статистики. Надо
только не забывать афоризм: «Статистике часто принадлежит пер-
вое слово, но последнее — никогда», ибо статистика призвана опи-
сывать феномен, а не раскрывать его механизм. Статистика необ-
ходима, например, для выявления глобальных круговоротов ве-
ществ. Концепция замкнутых круговоротов по каждому элементу,
вошедшая в большую науку с Вернадским, лишь недавно подверг-
лась серьезному анализу, и выяснилось, что «циклы» разомкнуты,
203
т. е. являются не циклами, а спиралями. Жизнь непрерывно
расходует одни ископаемые и создает другие, что ясно показала
в 1970-х годах Н. И. Базилевич: «Несбалансированность между
созиданием живого и деструкцией мертвого органического вещест-
ва является движущей силой эволюции почв, природных экосистем
и биосферы в целом» [Моделирование..., 1981, с. 83].
Формально говоря, этот факт был известен и Вернадскому,
но фактических балансов тогда не было, и он мог ограничиться
таким замечанием о накоплении ископаемых: «Количество живой
материи, выходящей этим путем из круговорота,., составляет не-
большую ее часть. Даже огромные толщи углей или нефтей...
едва соизмеримы с углеродом организмов» [Вернадский, 1978,
с. 303—304]. Но по данным Базилевич, углерод организмов состав-
ляет 0,1% осадочного. Многие элементы поступают в биосферу, в
основном, при необратимом распаде горных пород. Таков фосфор,
который «быстро циркулирует между водой и планктоном, более
медленно проникает в прибрежные растения и постепенно накап-
ливается в донных отложениях» [Уиттекер, 1980, с. 313]. Оттуда
он может быть извлечен только геологическим катаклизмом (на-
пример, поднятием океанического шельфа) или человеком, следо-
вательно, биологического круговорота фосфора почти нет. Ни ка-
таклизмы, ни человек также не обеспечивают круговорота, посколь-
ку возвращают в оборот лишь малую долю фосфора, причем
человек создает его локальный избыток, а следовательно — и новые
залежи.
Жизнь зависит от глобальных факторов в большей мере, чем
это допускалось ранее. Тот же фосфор: его поток через биосферу
определяется глубинной активностью планеты и мог в определен-
ные моменты становиться главным ограничителем жизни. Доста-
точно исчерпаться его наличным источникам, и жизнь должна
резко сузиться в ожидании нового горообразования. Это выясни-
лось уже в рамках балансового, статистического, подхода, почти
не рассматривающего структуру биосферы.
Есть вопросы, которые в статистических рамках вообще нельзя
выразить. Каким образом возникает новое, например — новый
орган или новый трофический уровень в экосистеме? Как возник
хотя бы главный из промежуточных уровней — животные? В главе
6 мы говорили о том, зачем он биосфере, и в прежней парадигме
этого было бы достаточно, поскольку в ней вопросы «как» и «за-
чем» практически неразличимы, ибо статистика не различает ме-
ханизма и его назначения, ограничиваясь выявлением тенденции.
Системный эволюционизм с выявления тенденций начинает (для
этого ему служит диатропика), но здесь важно вербальное обоб-
щение, а не формульное усреднение, как это принято при
статистическом подходе.
Вопрос о роли царства животных (о которой мы говорили
в конце 6.5) допускает в настоящее время и такой ответ: сложным
системам свойственна специализация частей, и более сложная
система (надцарство) эвкариот сравнительно четко делится на три
204
экологических царства (чего у прокариот вообще нет), причем
царство животных специализировано на обеспечении сложных ти-
пов потоков (материальных, энергетических и информационных),
которые недоступны растениям и грибам в силу их малой подвиж-
ности. Малая же подвижность связана с расходом энергии и ве-
ществ на синтез и разложение органики. Речь при этом идет именно
о тенденциях, свойственных царствам как целым, а не о свойствах
отдельных видов: некоторые растения (например, насекомоядные)
подвижнее некоторых животных (например, кишечных паразитов)
и т. д.
Такой ответ на вопрос «зачем?» подводит и к ориентировочному
ответу на вопрос «как?»: в потоках возникают ДС, а в них —
сродство частей к целому (как водоворот собирает в единую
горсть все, что до этого плавало порознь). Множество разнородных
ДС демонстрирует как градацию уровней, так и рефренную струк-
туру, а потому здесь возможна классификация. Пусть эта картина
слишком туманна и предварительна, все-таки она лучше годится
для организации знаний и для поиска нового, чем прежняя идея,
видевшая главное в том, что у всех животных был абсолютно
неспециализированный (т. е. ни на что не годный) общий предок,
остальное же считавшая не столь важным. О сродстве будет
сказано в 9.3, а сейчас следует отметить, что перемена отношения
к тенденции (обобщение вместо усреднения) приводит и к измене-
нию отношения к случайности.
О системной случайности мы говорили в главах 3 и 4. В част-
ности, было отмечено, что случайность служит цементом, скреп-
ляющим в целостную систему как реальные процессы, так и теоре-
тические представления о них, хотя случайности при этом фигу-
рируют разные: в природе чаще всего играют роль скрещение
несогласованных процессов и неустойчивость движения, а в пред-
ставлениях о ней — непонятая закономерность и относительность
знания. При возникновении нового чаще всего случайность высту-
пает как уникальность и (при его толковании) — как произволь-
ный выбор или как имманентная случайность.
Наряду с этими типами случайности статистический эволю-
ционизм эксплуатировал и другие, но они не имеют серьезного
обоснования — ни онтологического, ни гносеологического. Так,
часто абсолютную однородность рассматривают в качестве основ-
ной исходной характеристики мира и объектов в нем. Для объясне-
ния становления реальных структур (неоднородностей) приходи-
лось вводить ad hoc диссимметризующие факторы, т. е. громоздить
случайность на случайность. Системное мировоззрение, наоборот,
исходит из наличия начальной структуры как данной (подроб-
нее см.: [Чайковский, 1985; 19886]).
Отказываясь от необоснованной случайности, системный эволю-
ционизм об остальных типах случайности может сказать больше,
чем статистический. Ведь далеко не все случайные явления допу-
скают статистический подход: для баланса требуется наличие сред-
ней величины, а для существования вероятности — наличие устой-
205
чивой частоты при неограниченном повторении испытаний. В эво-
люционных задачах эти требования обычно невыполнимы, поэтому-
то статистика и не дала здесь работоспособной теории. Наоборот,
качественные обобщения здесь вполне возможны (даже уникаль-
ное событие обычно допускает аналогии, образующие номотетиче-
ский ряд), поэтому теоретическое описание эволюции, обещающее
практические выходы, наконец-то становится чем-то достижимым,
реальным. При этом, как уже сейчас видно, теория окажется
не той, что в физике: вместо процедур формального вывода из
постулатов, системный эволюционизм будет в основном, по-видимо-
му, строиться на качественных номотетиках.
9.3. Понятие сродства в системном эволюционизме
Итак, начало жизни можно понимать как возникновение биологи-
ческих форм сродства, но в самом ли деле мы поняли что-то новое
или, как часто бывает, просто придумали для непонятного новый
термин? Ведь и раньше многие усматривали в феномене жизни
нечто специфическое и давали этому специфическому фактору
красивое название (в частности, «энтелехия» Дриша, «взаимопо-
мощь» Кропоткина, «гея-фактор» Лавлока), но оппоненты видели
в этом только уход от решения проблемы. Не является ли и срод-
ство новым именем непонятного? Дать уверенный ответ пока нель-
зя, но понемногу выясняется, что сродство — не просто новый
термин, а скорее общее имя для группы явлений природы, в кото-
рых прежде не видели ничего общего. Еще при Дарвине взгляд
на эволюцию с позиций баланса и круговорота привел ряд авторов
независимо к одной и той же идее — сродства частей к целому.
В частности, небольшая книга Фрика [Freke, 1861] так и называ-
лась: «Происхождение видов посредством органического срод-
ства». О сродстве до него говорил Меррей [Murray, 1862], назы-
вавший этим термином то свойство организмов, благодаря кото-
рому в ходе онтогенеза образуются независимые сходства, именуе-
мые ныне миметизмом. За сто лет до этого Мопертюи [Maupertuis,
1756, с. 120 и далее] объяснял сродством самый факт онтогенеза,
который он понимал как самосборку частиц наследственности.
Термин «сродство» (affinity), заимствованный ими из химии,
означал то свойство системы (атомов, клеток, органов или организ-
мов — безразлично), которое заставляет ее компоненты объеди-
няться друг с другом для совместного существования. Другими
словами, сродством была названа способность той или иной систе-
мы к самосборке. Если элементы таковы, что сами собой собирают-
ся в систему, то мы говорим, что между ними имеется сродство.
Феномен сродства буквально пронизывает природу на всех
уровнях. Элементарные частицы соединяются как в другие эле-
ментарные частицы, так и в атомы: атомы — как в кристаллы, так
и в молекулы; молекулы — как в кристаллы, так и в более слож-
ные комплексы; из молекулярных комплексов образуются как ви-
русы и клеточные органеллы, так и клетки; клетки могут как обра-
206
зовывать организм, так и сами становиться организмами, и чем
сложнее организм, тем богаче в нем формы сродства частей (о чем
фактически уже шла речь в 6.6). Наконец, организмы соединяют-
ся как в семьи, стаи и популяции, так и в экосистемы. При этом
на многих уровнях можно наблюдать один и тот же эффект: пара
частиц, стремящихся в одной ситуации избавиться друг от друга,
в другой ситуации соединяется в рамках устойчивой системы.
Так, два протона в свободном состоянии взаимно отталкивают-
ся, но они же устойчиво сосуществуют в атомном ядре. Аналогично
два самца, встретившись, могут начать драку, но они же могут вхо-
дить в устойчивую стаю; наоборот, самец и самка, влекомые друг
к другу, словно протон и электрон, нередко не способны жить вме-
сте (зубр может в условиях заповедника забить самку насмерть,
если люди не разведут их по разным загонам).
Итак, при взаимодействии частей в целом причудливо перепле-
таются притяжение и отталкивание. В трудах Дарвина тоже есть
отдельные примеры сродства — так, он указывал, что многие расте-
ния могут противостоять истреблению только в том случае, когда
растут достаточно тесными группами; на подобных примерах неко-
торые биологи пытались даже обосновать такое понимание «борь-
бы за существование», которое включало в себя и все формы срод-
ства. Однако суть теории определяется не тем, как понимать в ней
то или другое слово, а ее логикой (или, если угодно, сродством по-
нятий), логика же дарвинизма в этом пункте проста и достаточно
известна: в основе эволюции лежит естественный отбор, который
возникает вследствие избыточности размножения и (вытекающего
из этого) вытеснения одними особями других [Дарвин, 1939,
с. 213]. Только вытеснение, т. е. взаимное отталкивание, есть пред-
посылка естественного отбора, тогда как факты сродства, даже если
собрать их много, остаются внешними по отношению к этой теории.
Для включения этих фактов в теорию отбора было сделано допуще-
ние, что любые организмы, демонстрирующие сродство друг к дру-
гу, когда-то в прошлых поколениях вытеснили своих собратьев, не
обладавших сродством (так объясняют появление заботы о потом-
стве, симбиоз, охоту стаями и т. п.). Словом, реально наблюдае-
мое сродство толкуют как результат гипотетического прошлого вы-
теснения.
О подобных эволюционных увлечениях Гегель шутил в «Фило-
софии природы», что они трактуют развитие «как продукт внеш-
них причин, которые, для того чтобы сделать их более наглядными,
помещаются в темную глубину веков» (§ 249). Особенно наглядна
их несостоятельность при обсуждении возникновения жизни, когда
загадок больше всего, а прежних поколений просто нет. Как, напри-
мер, возникло взаимное сродство белков и нуклеиновых кислот?
Ничего кроме нагромождения невероятных случайностей идеоло-
гия отбора пока не предложила, а другие концепции все так
или иначе базируются на идее сродства. Разработав одну из таких
концепций рождения жизни, Меклер [1980а, б] справедливо на-
звал своим предшественником Ламарка. Однако, стоя на позиции
207
крайнего физико-химического редукционизма, Меклер решил вы-
вести биологическое сродство из химического, что представляется
мне столь же безнадежным, как вывести законы стихосложения из
законов электрофизиологии мозга. «Вывод» при этом всегда на по-
верку оказывается цепью недомолвок и произвольных допущений,
очевидных лишь автору. Представляется более перспективной по-
зиция Мейена (являющаяся в сущности следствием теоремы Гёде-
ля) : редукционизм правомерен лишь в рамках одного уровня
организации [Диалектика развития..., 1978, с. 163J. Всякий пере-
ход познания на качественно новый уровень требует новой «аксио-
мы». В частности, переход на новый уровень организованности
знаменуется новой формой сродства частей к целому.
С непривычки это кажется странным — разве мы объяснили
процесс тем, что сослались на сродство? Ведь большинство, ученых
твердо уверено: объяснить работу механизма эволюции — значит
описать механику молекулярных превращений от поколения к по-
колению, а вовсе не ввести какую-то новую аксиому. Заметим, одна-
ко, что опыт других наук говорит иное: понять новое как механику
превращений уже известных элементов можно только тогда, когда
по-настоящему нового не ожидается; наоборот, абсолютно новое по-
знается не иначе, как введением новых аксиом. Механическую кар-
тину атома удалось дать только тогда, когда ученые отважились
изменить само значение слова «механика» и ввести аксиомы кван-
товой механики, когда вместо «планетарной модели атома» возник
язык квантовых уровней. Аксиом не надо вводить тем, кто ищет не
вполне новое, тогда как рождение жизни — пожалуй, самое новое,
что случалось на Земле. Не исключено, что именно введение аксиом
сродства, которых так настойчиво требует проблема рождения жиз-
ни, позволит лучше понять и обычную эволюцию. Ведь так уже бы-
вало: теория относительности, разработанная сперва для больших
скоростей, затем — для больших масс, оказалась полезной и для
земных масштабов (при этом загадочное равенство гравитационной
и инерционной масс тела было осознано как новая аксиома физи-
ки); а принцип неопределенности, рожденный для объяснения
атомных явлений, привел Бора к общему принципу дополнитель-
ности, необходимому, как мы не раз видели, и в биологии: ко вся-
кой достаточно общей точке зрения на данный объект существует
дополнительная точка зрения, раскрывающая новые свойства
объекта. В частности, взгляд на разнообразие как на результат
приспособления дополнителен к взгляду на разнообразие как на
тенденцию. Если квантовая механика оказалась необходимым язы-
ком при изучении новых масштабов длин и энергий, то, встречаясь
с новой, по сравнению с обычной химией, мерой организованности
материи, мы тоже должны, по-видимому, ввести новые понятия
[Чайковский, 1980].
9.4. Прогресс и приспособление
Итак, эволюционной термодинамикой предложена схема, согласно
которой прогресс неизбежен в силу феномена градации уровней
208
ДС, а приспособление — в силу всевозможных явлений сродства.
Оба тезиса обоснованы, в основном, аналогиями (с химией) и в этом
их слабость, но зато оба включены в единую, предложенную Янчем,
натурфилософскую схему, для которой термодинамика выступает
системной познавательной моделью. Для этой схемы существен
системный релятивизм: вводится наблюдатель, движущийся вместе
с системой, причем под движением имеется в виду не столько пере-
мещение, сколько развитие, а это значит, что пространство рас-
сматривается не физическое трех- или четырехмерное, а систем-
ное (фазовое) многомерное. Янч нигде об этом прямо не сказал,
но этого требует логика его схемы.
Процесс, который для наблюдателя в обычном трехмерном про-
странстве выглядит как движение к заранее выбранной цели, в си-
стемном континууме окажется, по Янчу, обычным причинно обу-
словленным движением, поскольку выбор конечного состояния
здесь неотличим от обычного задания граничного условия. Осями
в фазовом пространстве служат не только расстояния, но и все-
возможные системные параметры (скорости, температуры, концен-
трации, корреляции и т. д.). Поэтому и время здесь вряд ли можно
задать одним скаляром. Именно в таком многомерном континууме
и можно представить себе, как это делал Янч, приспособление
в виде искривления траектории подсистемы в силу системных от-
ношений; оно подобно искривлению луча света в силу гравитации.
Гравитация искривляет геодезическую (кратчайшую линию, свя-
зывающую данные две точки) или, другими словами, экстремаль,
находимую вариационным методом. Естественно ожидать, что
и приспособление — экстремаль в системном континууме, который
еще предстоит аккуратно определить в будущих исследованиях.
В своей последней книге Янч заявил, что само понятие приспо-
собления надо переосмыслить; направление адаптации радикально
зависит от мгновенных нерегулярных причин и не может иметь
постоянного значения (даже, добавим мы. в среднем). Как Берг-
сон и Любищев, он видел в приспособлении не самоцель эволюции,
а ее средство, аспект целесообразности. Последнюю он описывал
в терминах, близких если не к аппарату, то к духу точных наук —
теоретической физики и теории систем: «Биологическая эволюция
совершает эксперимент с целым филумом, начиная с образования
первых биомолекул, эффективный сейчас» [Jantsch. 1980, с. 16].
Требуется лишь оговорка: вместо эффективности «сейчас» надо го-
ворить об эффективности в течение всей истории «филума»,
причем будущий конец экстремали (если позволительна столь
прямая аналогия с вариационным исчислением) рассматривается
как подвижный. Самая возможность таких интегральных решений
виделась ему примерно так же, как она видится сторонникам
самодовлеющего смысла экстремальных принципов, т. е. в соответ-
ствующем устройстве наблюдаемого мира (о сторонниках такого
устройства мира см.: [Чайковский, 19886]).
С этой позиции Янч сумел увидеть абстрактное решение пробле-
мы «затравок», о которой мы говорили в 9.1. Откуда берется «за-
8 Ю. В. Чайковский
209
травка» для каждого нового уровня ДС? Всякая ДС обладает дина-
мической памятью (например, направление водоворота), которая
громоздка и неэффективна, зато порождается самой ДС и ей же слу-
жит остовом. Возможна и другая, консервативная, память — она
привносится в ДС извне (таков, например, вирус, попавший
в клетку). Консервативной памятью являются ДНК, коралловые
рифы, книги и т. п. структуры долговечные и эффективные, но не
участвующие сами по себе в процессах обмена, поддерживающих
соответствующую ДС [Jantsch, 1980, с. 198]. Янч даже назвал кон-
сервативную память «мертвой неравновесной структурой», кото-
рую вызывают к жизни лишь динамические феномены («метаболи-
ческий и нервный контекст»). Тем самым она может порождаться
недолговечным информационным процессом, но порождать процесс
долговечный, т. е. служить ему затравкой. В этом состоит эпигенеа-
логический принцип Янча [там же, с. 201].
Конкретизируя данный принцип, можно сказать, что жизнь на-
чалась с момента, когда в первой субвитальной области (см. 8.1)
синтезировалась цепочка РНК, послужившая затравкой для син-
теза пра-белка (пептидной цепочки). Это вполне возможно, если
учесть, что РНК способны к ферментативной активности [Ceder-
gen, Grosejean, 1987]. Появление полипептида, катализирующего
синтез РНК, которая сама катализирует пептидный синтез, было,
по Янчу. первой «встречей», породившей жизнь. Подобные «встре-
чи» (к ним он относил, например, соединение партнеров симбио-
за) Янч рассматривал не как случайные удачи, а как следствие не-
симметрии (неоднородности и неизотропности) системного про-
странства-времени, в котором неизбежно «самопорождение» новых
и новых систем. Здесь-то и есть главное расхождение системного
видения мира с прежним, механическим и статистическим, господ-
ствовавшим в науке более трехсот лет. Если прежде в науке цари-
ла презумпция симметрии, выражавшаяся в том, что все направле-
ния движения считались априори (при отсутствии возмущений)
равновозможными, а события — независимыми, то теперь постули-
руется наличие априори избранных направлений, отношений и вза-
имодействий, т. е. речь идет о презумпции системности. Фактиче-
ски эта точка зрения давно признана космологией в форме антроп-
ного принципа, согласно которому основные мировые константы
априори согласованы так, чтобы реализовался тот самый мир, кото-
рый мы наблюдаем (см.: [Балашов, 1988]). Янч скептически отно-
сился к трактовке антропного принципа как результата отбора дан-
ного мира из множества возможных, называя это «поспешным
объяснением в духе Дарвина», основанным на неведении законов
«самопорождения» [Jantsch, 1980. с. 96].
Самопорождение Янч тесно связывал с нарушением (breaking)
симметрии (термин Пригожина), верно отметив, что при переходе
от одного уровня исследования к другому возникает несво-
димость, связанная с тем, что при переходе природы от одного
уровня развития к другому нарушается какая-то фундамен-
тальная симметрия. Так, при переходе от механики к термодина-
210
мике нарушается симметрия времени и тем порождается причин-
ность [там же, с. 27 — 28]. По Янчу, ДС податливы (malleable)
[там же, с. 41], т. е. способны, в силу их самоорганизации, зани-
мать доступный фазовый объем и в то же время сохранять свои
существенные характеристики, тогда как статические структуры
жестки (либо противостоят воздействию, либо пружинят, либо не-
обратимо теряют свои качества). Это их свойство (характерное
и для всего живого) позволяет достигать упорядочения без строго
детерминированных траекторий, мягко. Здесь работает новый
принцип: «Чем больше свободы в самоорганизации, тем больше
порядка» — принцип, который Янч называл оптимистическим
[там же, с. 40]. Очевидно, что системодинамика (динамика ДС
в частности) несводима ни к традиционной динамике (механике),
ни к статистике. (Формально несводимость следует из того факта,
что всякая ДС, возникая, проходит через неустойчивость.) В каче-
стве примера самостоятельных уровней развития Янч рассматри-
вал «три микроскопических уровня самопорождения» — ДС, про-
кариоты, эвкариоты. В ДС нарушена только пространственная сим-
метрия (возникает структура), а в прокариотах — еще и времен-
ная (возникает наследование), но сохраняется «анархичность» —
все может комбинироваться со всем, и горизонтальная передача
генов (инфекция, трансдукция) не отделена от вертикальной (от
наследования). У эвкариот формы передачи информации резко раз-
граничены, и горизонтальная передача генов сведена к миниму-
му [там же, с. 132 — 133]. Тут, однако, мысль Янча требует уточ-
нения.
Он безусловно был прав в том, что сама идея искать согласован-
ность в случайном совпадении — лишь примитивное повторение
познавательных ошибок прошлого, что познавательные барьеры
несводимости существуют объективно и что, тем не менее, объект-
ная (затравки) и познавательная (нарушение симметрии) стыков-
ки уровней возможны. Однако свою положительную программу он
оставил лишь в форме грубого эскиза. Попробуем чуть прояснить
его.
Задав вопрос — в каком случае консервативная память, по-
рожденная динамической, становится затравкой для будущего
уровня развития,— мы неизбежно придем к необходимости понять,
что такое уровень. По-моему, здесь следует вспомнить, что эволю-
ционирующие системы — гёделевы (в смысле 7.2). Покуда разви-
тие системы выразимо в терминах логических операций и удлине-
ния текстов, можно говорить, что она развивается в пределах еди-
ного уровня. На каждом уровне ДС изготовляют консервативную
память для будущих ДС того же уровня (будущих поколений),
и ее вариации, каким бы механизмом они ни порождались, почти
всегда принадлежат тому же уровню — логическому и эволюцион-
ному. Лишь исчезающе редко (и в этом смысле — случайно) из-
готовляется текст (консервативная память), невыводимый (по Гё-
делю) из прежних, но тем не менее оказывающийся затравкой для
новых ДС — тогда следует говорить о новом уровне эволюции.
8*
211
В процессе происхождения жизни в качестве примера такого ново-
го уровня можно назвать упомянутый в 8.3 синтез первой ДНК по
РНК-овой матрице: ДНК оказалась консервативнее, чем РНК, чем
и обеспечено становление настоящей наследственности.
Пусть отдельные эволюционные претензии термодинамики
и необоснованны, все же нельзя отрицать, что она дает эволюцио-
низму новую исследовательскую программу и наконец-то указыва-
ет связь приспособления с прогрессом. Оба процесса — аспекты
самоорганизации, но прогресс связан с выходом на новый логиче-
ский уровень. Как и всякая консервативная структура, ДНК сама
по себе эволюцию вести не может, она не локомотив, а инструкция.
В нее записываются найденные удачные варианты самоорганиза-
ции. как изобретатель записывает удачные мысли в блокнот.
Эти записи можно подчас назвать случайными, но не в том смыс-
ле, какой вкладывает в этот эпитет теория вероятностей, а, напри-
мер, в смысле невыводимости.
Понемногу новая программа вычленяется и становится попу-
лярной [Brooks, 1983; .Nagi, 1986; Glaser. 1987; и др.], сменяя гос-
подствовавшую 120 лет адаптационную программу. Характерно,
что в философском сборнике [О современном статусе..., 1986] из
17 авторов 10 сослались на Янча. Пусть включение В. А. Кутыре-
вым [1988] книги Янча в пятерку самых модных научных книг
последних 20 лет — преувеличение, но ее организующая роль
налицо.
Янч почти не касался проблемы «происхождения видов», едва
ли не главной в привычном эволюционизме. Достоинство такого
подхода в том, что он взглянул на дело с другой стороны и ввел
в оборот понятия, для которых и языка не было и которые, поэтому,
были едва видны на периферии научной мысли; недостаток же —
в том, что огромный потенциал прежнего эволюционизма остался
почти не использованным.
Глава 10. ЯЗЫК И СТРУКТУРА ЭВОЛЮЦИОНИЗМА
10.1. Сквозные закономерности эволюции
Коль скоро феномен самоорганизации всеобщ и на всех уровнях
демонстрирует системные сходства (прежде всего, в наших терми-
нах,— рефренную структуру), то нет смысла выделять какой-то
уровень в качестве особого и описывать его исключительно в своих
специфических понятиях, как это долго делалось, например, в дар-
винизме. Естественно выявить наряду со специфическим то общее,
что объединяет развитие организма и вида с развитием других объ-
ектов, как меньших (орган, клетка и мельче), так и больших (эко-
система, государство, биосфера, космос).
Наличие таких сквозных закономерностей было, между прочим,
очевидно статистикам прошлого. Так. в 1830—1840-х годах
212
Т. Ф. Степанов, харьковский профессор политэкономии, развивал
взгляд на земной шар как на единый организм, описать законы
развития которого должна статистика, а объяснить — политэконо-
мия' и другие науки. Для нашей темы важно, что он исходил из
убеждения в необходимости развития природы и общества для са-
мого их существования. Изменение как климата и организмов, так
и общественных отношений было для него непременным условием
сохранения жизни на Земле [Степанов, 1844, с. 204]. Необходимым
атрибутом развития он полагал разнообразие индивидов, считая
должным заботиться как о бедных (в чем резко расходился с попу-
лярным тогда Мальтусом), так и об уродах, поскольку «все мы, как
достоинства свои, так и самые недостатки, должны почитать
в некотором смысле общими» [там же, с. 207]. К сожалению, столь
глубокий подход (близкий к взглядам тогдашней итальянской шко-
лы статистиков) не получил развития и вскоре был забыт.
Лишь через сто лет появилась возможность выяснить, в чем
здесь было дело. Как отметил системолог Уоррен Уивер, всякая
дисциплина проходит в своем развитии три стадии: изучение «орга-
низованной простоты» (детерминизм, механицизм), изучение
«беспорядочной сложности» (статистический, в нынешнем смысле
слова, подход) и изучение «организованной сложности» (систем-
ный, или интегративный, подход) [Weaver, 1948]. Степанов, писав-
ший с позиций механистического взгляда на природу, пытался пе-
рейти к системному ее пониманию, минуя собственно статистиче-
ский этап развития естествознания, этап Максвелла —Дарвина,
и потому не был понят коллегами.
Являясь отрицанием отрицания механистического подхода,
системный подход часто воспринимается натуралистами как воз-
врат к примитивному механицизму прошлого, и эта элементарная
философская оплошность приводит к множеству бесплодных дис-
куссий, на что неоднократно указывали Любищев, Мейен и другие.
В частности, критика в адрес неодарвинизма до сих пор часто тре-
тируется как возврат к додарвинским учениям. В действительности
имеет место возврат к их «организованности» по Уиверу, но на сов-
сем иной (сложной) основе. Например, феномен рефрена не мог
быть понят в додарвинской биологии, поскольку прежние рассуж-
дения о рядах (см. 1.5) носили либо фрагментарный (Фрис,
Максимович), либо расплывчатый (Агассиц, да и сам Дарвин)
характер. Потребовалась скрупулезная обработка (описательная
статистика) параллелизмов (например, работы Кренке в ботани-
ке), чтобы выявить статистическую (в вероятностном значении
этого слова) природу параллелизмов и тогда уже сформулировать
системную суть рефрена: он существует всюду, но его члены вы-
глядят очень по-разному в зависимости от того, относятся ли они
к ядру (точнее, радикалу по Вавилову) или к периферии исследуе-
мого разнообразия, так что рефрен не всегда легко виден.
То же можно сказать про общие свойства организма, общества
и биосферы. Эту древнюю параллель, к которой позже была добав-
лена аналогия с клеткой [Александров Д. А., 1986], детально раз-
213
вил Янч, который воспользовался и здесь идеей градации уровней
ДС в качестве исходной. По Янчу, биосферный «организм» в неко-
торой мере способен к целесообразному поведению и даже к его
планированию. (Профессионально Янч занимался научным прог-
нозированием [Янч, 1974].) Онтогенез этого «организма» мы на-
блюдаем через эволюцию его «клеток» (видов) и «органов» (эко-
систем), а в онтогенезе, как известно, ненаправленные вариации
играют лишь подчиненную роль, отбор же их — и того меньше.
Естественно, что отбор как фактор эволюции состоял для Янча не
столько в гибели плохого мутанта, сколько в выборе нового текста
для записи в генотип, производимом соответствующей ДС: «Осо-
бую важность здесь имеет селективное и синхронизованное исправ-
ление консервативно записанной (т. е. генетической) информации
с помощью диссипативного процесса». Традиционному отбору он
отводил следующую роль: «Давление отбора, действующее через
среду, гораздо более эффективно в незрелых системах — как уско-
ритель эволюции. В зрелых же системах эволюцию ускоряют
флуктуации как таковые», т. е. процесс образования ДС. (Это до-
статочно близко к пониманию отбора, данному в 7.2.) Янч не спо-
рил с Дарвином, признавал его заслуги, но считал их «фрагмен-
тарными», как и заслуги пунктуализма ’, входившего в то время
в моду. Причину «фрагментарности» он видел в том, что обе докт-
рины исходят из равновесия как чего-то данного [Jantsch, 1980,
с. 16, 140. 239].
Но и одного лишь динамического подхода недостаточно для
преодоления фрагментарности: она неизбежно возникает, если
свойства «своего» объекта экстраполировать на разнообразие
объектов, как это прокламировал, например, бактериолог Жак Мо-
но тезисом: «То, что верно для бактерии, должно быть верно и для
слона». Зоолог Альбер Вандель отверг этот тезис [Vandel, 1968,
с. 166], напомнив, что закономерности эволюции бактерий и живот-
ных различны: вопреки частым мутациям и огромной числен-
ности, бактерии почти не эволюционировали за миллиарды лет, для
них типична адаптация, тогда как для животных она вторична,
а первичен типогенез по Шиндевольфу [там же, с. 161 —162].
Добавим, что вся эволюция слонов прошла за 3 млн лет, причем
десятки их видов появились и исчезли, хотя общее количество их
особей не достигло даже того числа, какое обычно требуется у бак-
терий для получения одной полезной мутации. Ясно, что язык
мутаций — не тот, на котором можно формулировать сквозные
закономерности эволюции.
1 Пунктуализм, или теория прерывистого равновесия,— представление об эволю-
ции как о чередовании быстрых актов таксонообразования, перемежаемых дли-
тельными стадиями относительного постоянства таксонов. Подробнее см.: [Ка-
тастрофы..., 1986; Меуеп, 1987]. В сущности, коцепция очень стара: еще Ост-
вальд Геер [Heer, 1859] на палеоботаническом материале описывал быструю
«перечеканку» (Umpragung) флор, перемежаемую длительным стазисом. Споры
о пунктуализме еще не завершились, но уже четко установлено, что периоды
резкого возрастания скорости преобразования таксономического разнообразия
действительно имели место.
214
Эти закономерности Вандель видел в другом — в изоморфизме
процессов эволюции, идущих на разных уровнях. Он перечислил
семь главных типов эволюции, которые удобно расположить в по-
рядке их гипотетического начала, указав для каждого типа основной
макрообъект и характерный для него «кирпичик» (элемент), т. е.
говорить об эволюции (1) галактик и элементарных частиц, (2)
звезд и химических элементов, (3) планет и химических соедине-
ний, (4) биоценозов и организмов, (5) общественных структур
и сигнальных языков, (6) мышления и концептуальных языков,
(7) научно-технических знаний и изделий. Подробнее см.: [Диа-
лектика развития..., 1978, с. 124].
Изоморфизм свойств эволюции материальных и идеальных (на-
пример, языков) объектов говорит о более глубоком сходстве, чем
то, какого можно ожидать, исходя из сходств формирования ДС.
Старая проблема — существовал ли единый предковый язык или
их сразу было несколько [Трубецкой, 1987] — вряд ли продвинет-
ся к решению, даже если аналогичная проблема в биологии будет
успешно моделирована термодинамически. Как уже говорилось,
общесистемные сходства имеют более глубокую природу. Здесь
следует отметить следующее: общим для всей семерки является то,
что любое разнообразие эволюционирует путем все более полной
реализации своих рефренов, путем заполнения клеток одной и той
же таблицы. Это приводит к трем общим выводам.
Во-первых, новизна сводится преимущественно к новым комби-
нациям прежних свойств, что отмечали не только многие биологи,
но и, например, науковед М. А. Розов, который назвал этот фено-
мен, выявленный им в истории знаний, пересечением традиций.
Во-вторых, поскольку таблица нынешнего разнообразия и преж-
него развития в основном одна и та же, постольку анализ первого
может дать для понимания второго гораздо больше, чем принято
считать. Так, легко разрешается противоречие между дарвинизмом,
видящим всюду непрерывное перетекание одного таксона в другой,
и пунктуализмом. Ныне живущие организмы делятся на резко обо-
собленные — по одному виду в роде или даже в более высоком
таксоне — и необособленные (см. 3.5). Исследуя ископаемые,
оставшиеся от обширного рода, где виды имели по многу подвидов,
палеонтолог легко увидит перетекание, а исследуя семейство, со-
стоявшее, в основном, из одновидовых родов — перечеканку. Даже
в одном организме могут сосуществовать органы, возникшие как
скачком, так и постепенно, поэтому «мы должны не выбирать меж-
ду моделями пунктуализма (сальтаций) и градуализма, а думать
о том, как эти модели сочетаются в реальном процессе эволюции»
[Меуеп, 1987, с. 357]. Узнать, совершались ли переходы за десятки
поколений или за десятки тысяч, по данным палеонтологии нельзя,
но она полезна в этой дискуссии для другого: как и в опытах
Шапошникова, здесь часто бывает видна ограниченность разно-
образия возможных форм — в ходе истории снова и снова обра-
зуются сходные конструкции организмов, о чем писал еще Коп.
В-третьих, новые мероны (строки таблицы) появляются редко
215
(в основном, в моменты типогенеза по Шиндевольфу), но все-таки
появляются. Экстраполируя назад, легко прийти к ошибочному за-
ключению, что какая-то клетка заполнена первой, что разнообра-
зие родилось из однообразия. Гипотетический первый представи-
тель типа и явится тем общим корнем, о котором мы говорили
в 6.4. Вера в общего предка оказывается частным случаем идеи
общего корня разнообразия.
Эта идея настолько распространена в науке, что можно гово-
рить о принципе общего корня как основном постулате эволюцио-
низма (сама Вселенная согласно теории «Большого взрыва» мы-
слится как первично однородная): разнообразие рождается из
однообразия. Только не следует думать, что этот принцип принят
потому, что хорошо обоснован,— основанием ему прежде всего слу-
жит его наглядность: новое возникает по единому механизму,
и притом простейшему. В условиях, когда акт новации не имеет
формального объяснения, его приходится трактовать как случай-
ность, притом исчезающе редкую, а это правдоподобнее, если допу-
щена максимальная мыслимая простота возникшего. Отсюда —
уникальность новации и однообразие нового. Итак, в основе эволю-
ционизма лежат (в обозначениях 3.1) случайность-уникальность
(тип В) и случайность-спонтанность (ее чаще всего поставляют
процессы типов А, Б, Г), а принцип общего корня основан на допу-
щении максимальной простоты механизма новации.
Однако это допущение оборачивается трудностями при
объяснении становления разнообразия: мы обычно не видим даже
следов исходного однообразия, не удается аккуратно описать
гипотетический механизм, порождающий разнообразие из одно-
образия, да и дальнейшую историю разнообразия приходится
описывать посредством многих добавочных предположений, требуе-
мых не логикой предмета, а возникающими по ходу исследования
трудностями (анализ см.: [Чайковский, 1985, с. 158—159]). Ес-
тественно рассмотреть альтернативную точку зрения, согласно ко-
торой новое возникает из старого разными путями и в разных
формах, а разнообразие всегда порождается каким-то прежним
разнообразием, но не рождается из однообразия, причем законо-
мерность разнообразия выражается в его внутренней упорядочен-
ности (номотетике), а не в возможности вывести его свойства из
единого принципа или закона. Выразим это короче двумя тезисами:
(а) разнообразие из разнообразия;
(б) упорядоченность неформальна.
Тезис (а) не означает никакого закона сохранения — в ходе
эволюции разнообразие меняется и количественно, и качественно.
Так, древнейшая жизнь, по-видимому, состояла из одних бактерий,
но они-то были достаточно разнообразны, чтобы образовать биосфе-
ру [Заварзин Г. А., 1979; 1984]. Без исходного разнообразия
нечему эволюционировать, и это относится даже к рождению
Вселенной; хотя теория ДС и формулирует условия, при которых
однородное становится неустойчивым, но лишь начальная неодно-
родность (затравка) решает, куда пойдет дальнейшее развитие,
216
которое породит остальное разнообразие. Напрашивается во-
прос: каково минимальное разнообразие, достаточное для начала
эволюции данной системы? Обсуждение его было дано ранее
[Чайковский, 1985, с. 165 168], и здесь достаточно привести
только вывод: система должна быть, образно говоря, гёделевой в
смысле 7.2 и 9.4 («образно говоря» добавлено в силу тезиса (б):
гёделева система может служить лишь формальной моделью
реальной системы). Такая система как бы определяет саму себя,
включая и собственное развитие, как это мы видели в отношении
естественного языка (3.2), и именно поэтому аналогии между раз-
витием языка и природы оказываются продуктивными.
Однако и образная речь требует важной оговорки: всякая
реальная система развивается лишь в определенной среде. Абсо-
лютно определяет саму себя только Вселенная как целое, а уже
биосфера мыслима лишь в потоке веществ и энергии. Называя
какую-то систему (государство, общественный институт, язык,
биоценоз, организм, клетку и т. д.) целостной, надо всегда иметь
в виду среду, в которой та может развиваться, и не забывать, что
среда тоже развивается. Тем самым всякая эволюция на поверку
оказывается коэволюцией системы и ее среды. Другой вопрос —
можно ли счесть среду тоже целостностью, обладает ли она для
этого необходимой связностью.
Как отметил Янч, для коэволюции характерно сопряжение
макро- и микроструктур, причем с течением времени в целом
растет размер основных объектов микроуровня (от фотонов до
людей), и падает размер систем макроуровня (от протогалактик до
семей). Эту мысль Янч иллюстрировал двумя схемами (космиче-
ской и земной эволюции), которые соединены мною на рис. 7
в одну, уточненную. В основу положены следующие соображения,
рассмотренные нами в главах 6—8: (1) метаболизм предшествовал
наследованию и возник на базе геохимических круговоротов; (2)
первичный метаболизм был гетеротрофным (потреблял небиологи-
ческие потоки органики), но автотрофность (в том числе, вероятно,
фотосинтез) возникла до начала экспансии живого, т. е. до появле-
ния самовоспроизведения; (3) генетический материал возник ранее
самовоспроизведения, и его первичным свойством была реком-
бинация, обеспечившая преобразование разнообразия еще до появ-
ления прочих функций живого; (4) самовоспроизведение (размно-
жение генетического материала) появилось до сегрегации живого
на отдельные организмы; (5) значение ядра клетки сказалось
прежде всего в симбиозе (прокариоты к симбиозу друг с другом
неспособны), но затем оказалось главным в порождении многокле-
точных (реверсия разнообразия).
Вся эволюция эвкариот прошла между двумя системными рубе-
жами. Момент (в геологическом смысле этого слова) ts характери-
зовался рождением симбиоза, а всякий симбиотический организм
является микробиоценозом, поскольку между организмами, его
составляющими, существуют круговороты. Следовательно, в мо-
мент ts объект микроуровня (организм) стал элементом макроуров-
217
Характерные процессы
i макроуровень
макроуровень
Стабилизация размеров небесных тел
Синтез тяжелых ядер
Планетная хемодинамика
Геодинамика
Анаболизм (5) + рекомбинация
Фотосинтез
Самовоспроизведение
Размножение
механизмов
3S
Обособление
внутри клетки
разнообразия
Половое размножение
морфологический прогресс
Обособление полов
Язык сигналов
3
о.® »
и
Автономизация
от среды
Реверсия
(1}
Охлаждение Вселенной
s
1
€
1
§
л
*
Язык
fs Я понятий
Е-е-
S S Й
s
5 3
218
ня (биоценозом); после этого переплетение макро- и микроуровня
нарастало (вспомним хотя бы муравейник — одновременно и ор-
ганизм и биоценоз со своим «домашним скотом» — тлями и пр.);
с появлением в момент t человека деление эволюции на эти
уровни просто потеряло смысл. Итак, эволюция эвкариот, в основ-
ном законченная, отличалась от всех предыдущих переплетением
уровней эволюционных процессов, а будущая эволюция (неизбеж-
ная, пока существует сама жизнь) должна описываться (если
не исчезнет человечество) другим языком, о котором можно будет
говорить только после выявления языка прежней эволюции.
10.2. Язык и понимание эволюции
Великие эволюционисты очень различно видели будущее. Если
не касаться спора космологов о том, сожмется ли Вселенная в точ-
ку, рассеется ли в пыль или будет бесконечно пульсировать,
а ограничиться близким будущим нашей жизни, то надо вспомнить,
что Ламарк [1959, с. 442] полагал технический прогресс причиной
будущей гибели человечества, а Тейяр и Вернадский верили, на-
оборот, что настанет эра разумной эволюции, которая породит
ноосферу — сферу разума. В последние годы возрождается доверие
к мрачному прогнозу Ламарка. В такой ситуации понять эволю-
цию — значит не столько объяснить происшедшее (что до сих пор
было у эволюционистов главным) или предсказать ее возможный ход
(о чем пойдет речь в части III), сколько осознать необходимость
немедленной реакции на процессы, которые идут сейчас. Однако
нельзя договориться о какой-либо реакции, если нет согласия
даже в том, что из происходящего относится к эволюции. А это
невозможно выяснить без ответа на вопрос — что такое эволюция,
что именуется этим словом.
В предыдущих главах мы видели, сколь неоднозначно понима-
РИС. 7 Ориентировочная схема коэволюции систем макро- и микроуровня от мо-
мента разделения вещества и излучения t0 (полагают, что это произошло через
100 тыс. лет после Большого взрыва [Девис, 1985, с. 46]) до появления человека
в момент tm. Под каждой системой указан ее характерный элемент. Масштабы
вдоль разных кривых различны (молекула больше атома и т. д.), а рост (умень-
шение) характерных размеров вдоль одной кривой следует рассматривать лишь
как общую тенденцию, допускающую отдельные отклонения. Интервал t0, th (около
15 млрд, лет) — неорганическая эволюция, интервал th, ts (около 2 млрд, лет) —
становление жизни (биохимическая и клеточная эволюция), интервал ts, tp (около
1 млрд лет) — становление тканевой организации, tp, tm (670- 700 млн лет) -
морфологическая и поведенческая эволюция (фанерозой).
Примечания: (1) звезда 1-го поколения (синтез гелия); (2) звезда 2-го поколения
(то же-|-синтез тяжелых элементов); (3) время выхода жизни в океан и станов-
ления биологических «циклов»; (4) биохория — территория распространения
определенной группы организмов (то же в биогеографии, что таксон в система-
тике) . Биохории наземных растений (фитохории) удается выявить с карбона
(350 млн. лет назад), тогда как массовый выход жизни на сушу происходил в девоне
(400 млн. лет назад) [Мейен, 19866; 19876]; (5) анаболизм — конструктивная
часть метаболизма (см. § 6.5). Деструктивная его часть (катаболизм) первоначаль-
но могла осуществляться неорганическим путем; (6) л и м е с — граница трех
фаз — твердой, жидкой и воздушной. Служила активной зоной протобиологиче-
ских реакций; (7) видоспецифические колонии, ценоциты, мицелии, плазмодии.
219
ние таких расхожих слов, как отбор или случайность, а ведь
понятие эволюции сложнее. До сих пор многие руководства утвер-
ждают, что эволюция всегда идет очень медленно, т. е. не относят
к ней идущие у нас на глазах быстрые изменения — приобретение
устойчивости бактерий к лекарствам и насекомых к ядам, приспо-
собление птиц к жизни в городах, морфологические (акселерация)
и физиологические (незрелость, аллергия) изменения людей и мно-
гое другое. Ключевую роль понятийного анализа подчеркивали,
например, Любищев, Янч и Мейен.
Смысл основных понятий Любищев анализировал посредством
апорий (так он называл пары антонимов). Для термина «эволю-
ция» он рассмотрел пять апорий: эволюция — постоянство, эволю-
ция (развертывание наследственных потенций) — эпигенез (но-
вообразование), эволюция — революция, эволюция (градация) —
эманация (деградация), эволюция (достижение в первый раз) —
инволюция (возврат к прежнему). Почти все споры эволюционис-
тов связаны с тем, что каждый отстаивает не только «свой»
механизм на «своем» круге фактов, но и свое понимание самого
феномена эволюции. Так, победа Дарвина состояла в том, что его
современники признали эволюцию в первом и третьем смыслах,
тогда как многие отрицали победу Дарвина на том основании,
что тот не рассмотрел эволюцию в смысле второй апории. Она,
кстати, относится к диатропике, поскольку заставляет задуматься,
создается ли новое разнообразие процессом ненаправленных му-
таций и рекомбинаций, или же этот процесс только преобразовыва-
ет прежнее разнообразие само в себя. Подробнее см.: [Системность
и эволюция, 1984, с. 35]. Для решения актуальных задач экологиче-
ского кризиса важно четко различать четвертую и пятую апории,
поскольку движение вперед может явиться деградацией.
К пятой апории Любищева близок анализ пары «новизна —
подтверждение» (novelty — confirmation) у Янча: информация по
Шеннону (как и энтропия по Больцману) — чисто статическое
понятие, не связанное с развитием (новизной). Она учитывает
лишь «синтаксический уровень», расположение знаков, тогда как
собственно развитие идет на «семантическом уровне» [Jantsch,
1980, с. 41, 51]. Выход он видел в разработке понятия прагматиче-
ской информации Христины Вайцзеккер (1974 г.); оно во многом
повторяет понятие семантически полезной информации
Ю. А. Шрейдера (см. 3.2). Однако главным для Янча была
(см. гл. 9) первичность движения — отсюда у него и такие
специфические понятия, как «встреча» (т. е. динамика сродства,
понятого в самом широком смысле), динамическая память и
нарушение симметрии.
С каждым типом и этапом эволюции Янч связывал свою
особую форму нарушения симметрии. По-моему, лучше было бы
говорить об усложнении симметрии, поскольку она в ходе эволю-
ции хотя и не сохраняется, но, как правило, и не исчезает целиком:
асимметризующее изменение обычно порождает разнообразие
частных симметрий. Так, по Пригожину [1985], время возникло
как результат необратимости, создав этим фундаментальную
220
асимметрию «прошлое—будущее», но при этом возникло множе-
ство периодических явлений (одно из них — смена поколений),
являющихся разновидностью трансляционной симметрии.
Впрочем, сама идея вводить динамическое понятие как наруше-
ние или усложнение статического инварианта вряд ли перспек-
тивна. Симметрия — один из таких инвариантов (сохранений,
балансов, констант), сам поиск которых — наследие прежнего под-
хода, который Янч пытался преодолеть. Здесь лучше прямо гово-
рить, что в ходе развития неизбежны невыводимые из прежней
логики акты, гёделевы ситуации. Картина мира, в которой первич-
но движение, требует нового языка, новых исходных понятий.
Вместо статических инвариантов уместен чисто динамический ин-
вариант — когерентность (согласованность фаз движения), необ-
ходимая в физике при рассмотрении кооперативных эффектов
(например, сверхпроводимости), для описания которых разраба-
тывается когерентная логика. В качестве примера логической
когерентности И. А. Акчурин [1985, с. 254] приводит то свойство
целостного объекта, которое позволяет дать ему логический диаг-
ноз (отнести его к определенному таксону). В этом аспекте
когерентная логика смыкается со статичной логикой ценоза (см.
5.2). Если так, то именно разработка когерентной логики должна
дать тот «синтаксис» биологической грамматики, о котором гово-
рилось в 2.2. Другой пример когерентности в биологии (одновре-
менно и статической, и динамической) дает взаимная приспособ-
ленность организмов, которую Янч (см. 9.4) рассматривал,
следуя физикам, как неотъемлемое свойство системного простран-
ства, но не как результат накопления независимых случайностей.
Наконец, «принцип дополнительности в эмбриологии» (7.2) Коро-
чкина — тоже пример когерентности, только динамической и на
меньшем масштабе времени.
Однако успехи нового языка пока невелики. В основном мы
наблюдаем парадоксальную ситуацию: для характеристики
процесссов общего (типичного) вида мы и слов-то не имеем,
характеризуя их только через отрицания, например, как
нелинейные. С тех пор, как 300 лет назад Ньютон предложил
описывать механику материальной точки линейным дифференци-
альным уравнением, этот математический аппарат стал в естест-
вознании основным. (Уравнения в частных производных, ставшие
основой физики сред, тоже линейны.) Это позволило создать
работоспособный аппарат, приведший к огромному прогрессу
наук о природе, но и издержки, как мы теперь видим, тоже были
огромны: всякую нелинейность стали трактовать как усложнение
соответствующей линейной схемы, как экзотику. Сложился некий
язык линейности, т. е. традиция описывать процессы в форме
приближения к линейным, посредством малых добавок в уравне-
ниях, отчего и сама линейность выступала в качестве познава-
тельной установки (общей для механической и статистической
моделей), часто мешая видеть суть дела. Так, считалось, что
процесс диффузии способен лишь разрушать структуры (отсюда
221
и вера в то, что «миром правит закон роста энтропии»). Действи-
тельно, линейное уравнение диффузии ut=Duxx имеет лишь реше-
ние и(х, t) стремящееся в каждой точке х к нулю с ростом t.
Однако, например, в горящей спичке диффузия тепла приводит не
к распаду, а к перемещению источника тепла, и по спичке бежит
компактная структура — огонек. Еще в 1937 г. Колмогоров
с соавторами показали, что если к правой части уравнения
добавить нелинейность F(u), описывающую условие загорания при
температуре uo(F=O при и<лщ, F=1 при и^ио), то возможно
решение в форме бегущей волны. Результат нагляден, но он про-
тиворечил тогдашним понятиям и убеждениям, а потому стал до-
стоянием большой науки лишь в наши дни [Свирежев, 1987].
И до сих пор термодинамика не избавилась от ограничений,
связанных с господством термостатического языка.
Стремление вводить для новых фактов и идей новые понятия
встречает противодействие со стороны укоренившейся традиции
обходиться раз навсегда установленным набором понятий. (При этом
ссылаются на «бритву Оккама», хотя сам Вильям Оккам говорил
вполне определенно: «Сущности не умножаются без необходимос-
ти»,— т. е. все дело в том, чтобы вовремя усмотреть необходи-
мость.) На практике это оборачивается необходимостью игнори-
ровать некоторые факты в надежде, что им найдется объяснение
в будущем. «Эти факты отмечаются как курьезы, но если их по-
добрать, то получится весьма импозантная картина» [Любищев,
1982, с. 161]. Еще чаще мы видим безудержное расширение
смысла понятий с целью уместить в них новые факты и идеи,
т. е. неконтролируемую понятийную экстраполяцию, которую надо
сменить на контролируемую.
Мейену принадлежит понятие типологической экстраполя-
ции — суждения обо всех представителях данного класса объектов
по результатам выборочного изучения его представителей. Все
суждения о природе являются такими экстраполяциями. Открыв
новый вид растений или животных, любой признает, не глядя
в микроскоп, что этот вид — эвкариот. Без таких экстраполяций
наука невозможна, так что вопрос лишь в том, как выбирать пре-
делы экстраполирования; «все ошибки в науке (если исключить
прямые ошибки в наблюдении) — это неверные типологические
экстраполяции» [Мейен, 1981, с. 154]. Например, утверждение
«в эволюции нет ничего, кроме отбора случайных мутаций» есть
экстраполяция круга собственных представлений («я не знаю ни-
чего, кроме...») на многообразие природных явлений. Причин
ошибок, по Мейену, всего две — «плохая типология» (неудачный
набор классов) и неправильное отнесение объекта к данному клас-
су. (Заметим, что набор непересекающихся классов почти всегда
неудачен в силу наличия периферии вокруг почти любого ядра.)
По Мейену, самое изощренное исследование «не стоит и ломаного
гроша, если не знать того круга объектов, в пределах которого
можно смело экстраполировать полученные сведения» [там же,
с. 155], и в этом отношении диатропика оказывается полезна.
222
Ведь экстраполяция — рассуждение по аналогии, а аналогия —
causa exemplaris Суареса.
Как отмечено в 3.2, почти каждое слово разговорного языка
может служить для определения других слов, что делает язык
основным инструментом познания. Сами явления природы теперь
трактуются рядом ученых как языковые, как некоторая символи-
ческая активность. Эта «онтологизация символических структур»
[Диалектика развития..., 1978, с. 17] не является гносеологиче-
ской новинкой: как уже отмечено в 1.5, ученым свойственно на-
ходить в природе то, что стало модным в их обиходе, и вот теперь
в природе нашли формальную логику. В действительности,
обнаруженное — не более чем параллелизм двух рефренных струк-
тур, что не умаляет важности обнаруженного, но что надо
помнить при контролируемой экстраполяции. Если рассматривать
пять причин (см. 1.2) как рефрен, повторяющийся во всех задачах
познания, то кроме целеполагания, формальной логики и прочего,
в природе естественно искать и логику уподобления. Ведь и Суарес
писал, что хотя подлинное суждение по аналогии требует работы
сознания, но метафорически оно присутствует и у «низших соз-
даний природы» [Suarez, 1600, с. 617].
По-моему, именно с онтологическим метафорическим уно- .
доблением связан тот вывод биофизика и методолога Говарда
Патти, что простота формулировок законов природы связана со
сложностью языка науки, что при упрощении языка теории ниже
некоторого можно обнаружить лишь физические взаимодействия,
а не существо системы с ее символической активностью [Диалек-
тика развития..., с. 32].
Одним из главных таких упрощений языка является использо-
вание упомянутого выше языка линейности, проникшего в научный
обиход глубже, чем обычно думают. Например, линейность
лежит в основе теории вероятностей: результаты случайных
испытаний для получения общего результата складываются.
В 3.6, перейдя от центральных случайных явлений к явлениям
краевым, мы отказались от одной, самой заметной линейности,
но главная, глубинная линейность осталась. Состоит она в том, что
сама вероятность исходного события (например, гибели бактерии)
определяется как сумма всех однотипных случаев (суммарное
число всех погибших), отнесенная ко всем случаям (ко всем бак-
териям данного опыта). Теперь вместо линейности главным ин-
струментом экстраполяции должна стать аккуратная аналогия
в рамках рефренов.
Как пишет Акчурин [1985, с. 256], наиболее глубоким понятием
при исследовании оснований физики является на сегодня плоский
морфизм — такое преобразование объекта (не обязательно мате-
риального), при котором, образно говоря, целое не становится
частью, а внешнее - внутренним, и обратно. Плоский морфизм —
наиболее широкое обобщение понятия линейности (или, если
угодно, наиболее продвинутая аналогия с линейностью). Не явля-
ется, например, плоским морфизмом преобразование объема жид-
223
кости, движущейся турбулентно (бурлящей), процесс, на ко-
тором, по Акчурину, впервые «споткнулась» классическая механи-
ка [там же, с. 260]. Неплоский морфизм становится основным
аппаратом физики элементарных частиц, причем дело не в самой
их малости, а в особенностях топологии их преобразований при
взаимодействии. Акчурин приводит примеры неплоских морфиз-
мов в явлениях мышления и общественной жизни, но тогда уж
следует отметить, что самые обычные биологические явления —
метаболизм и половой процесс — тоже перемешивают внутреннее
с внешним и целое с частью. Это наводит на мысль, что язык для
описания преобразований органических разнообразий еще пред-
стоит создать. Если статистика описывает средние, а биологическая
грамматика — спектр возможных состояний, то от будущей теории
естественно ожидать указания спектра возможных путей развития.
Пока же можно только заметить, что речь идет даже не об одном
будущем языке, а о нескольких.
В самом деле, новое приспособление — это новая целостная
система, а вовсе не набор мутаций; эту фразу часто произносят, но
не всегда задумываются о ее следствиях. Прежде всего, искать
следует не механизм удачных мутаций как таковой, а механизмы,
участвующие во взаимоувязке разноречивых требований и возмож-
ностей среды и организма. Речь идет, как до недавнего времени
считали, об увязке трех разнообразий — условий среды (экотипа),
действий организма (фенотипа) и возможностей наследственного
материала (генотипа). Однако цитолог В. Я. Александров [1970]
обратил внимание на четвертое разнообразие — свойств и воз-
можностей внутриклеточных механизмов. Этим механизмам Мар-
челло Барбиери [Barbieri, 1981] предложил название — риботип,
указывая на основную роль в них рибонуклеиновых кислот —
матричных, рибосомных и транспортных. По Барбиери, именно
в форме риботипа жизнь возникла, да и сейчас всякий организм
существует как взаимодействие генотипа, риботипа и фенотипа.
В этом взаимодействии существенна роль транспортных РНК
(тРНК), которые организуют взаимодействие биохимических сис-
тем клетки (а не только включают аминокислоты в белковый
синтез) и информируют полинуклеотидную систему о внешних
воздействиях на клетку; тРНК обладают пространственной
структурой, напоминающей белковую, что и делает их удобными
посредниками. (На данные свойства тРНК и их роль мое внимание
обратил медико-биолог А. М. Гусейнов в 1983 г. на семинаре в
МОИП.)
В действительности надо говорить даже о взаимоувязке еще
большего числа разнообразий. В эвкариотной клетке есть много
разнообразных органелл, так что вряд ли здесь можно ограничить-
ся одним понятием риботипа. В многоклеточном организме
следует выявить цитотип (совокупность разнообразных клеток),
а высшие организмы обладают разнообразием тканей и органов.
Наконец, как считает философ В. А. Шуков [1987], живое тоже
взаимодействует со средой в форме диалога. Каждое из этих раз-
224
нообразий имеет свой «язык» (свою когерентность), так что
требование одного уровня может иметь смысл для других уровней
только при наличии «переводчиков». Пусть тРНК и годятся на
роль «переводчиков» между рибо- и цитотипом, а молекулы
клеточной адгезии [Эдельман, 1989] — между клетками и тканя-
ми, но как организуется общение между другими уровнями? И
каковы правила таких переводов? Как видим, ситуация сходна с
той, какую Корочкин отметил для эмбриологии, и в биологии, так
же как в лингвистике, в будущем встанет вопрос о сравнительной
грамматике разных языков.
10.3. К концептуальной схеме биоэволюционизма
Если для классического эволюционизма Ламарка — Дарвина был
характерен акцент на приспособление к среде обитания, то теперь
на первый план выходит идея самоорганизации. Вместо приспо-
собления вида к заданной среде как чему-то внешнему говорят об
усложнении и взаимоприспособлении живого как едином процессе
преобразования разнообразия. Разумеется, при этом нельзя отказы-
ваться от достижений прежнего эволюционизма; даже если приз-
нать их фрагментарными, их следует включить в единую схему.
Традиционный эволюционизм включает не менее трех основных
течений. Ламаркизм основан на идеях градации (усложнения,
прогресса) и приспособления, дарвинизм - на идеях приспособле-
ния и разнообразия (вариаций), а номогенез — разнообразия
и прогресса. Хотя и Ламарк упоминал о разнообразии, и Дар-
вин — о прогрессе, и Берг — об изначальной целесообразности как
«основном свойстве живого», но не в этом был пафос (сохра-
нившийся поныне) их учений. Удивительно, как постоянно отме-
чал Мейен, отсутствие не только в номогенезе, но и в ламаркизме,
и в дарвинизме теории процесса приспособления. Вместо нее гово-
рят лишь, что феномен приспособления неудивителен: для
ламаркиста — потому, что организмы стремятся приспособиться,
а для дарвиниста — потому, что неприспособленные вымирают.
Отсутствие серьезного анализа было неизбежно, покуда само
понятие пользы трактовалось как нечто внешнее организму (см.
4.5). Как мы видели выше, теперь ситуация изменилась: прогресс,
приспособление и разнообразие выступают как три аспекта са-
моорганизации, точнее — коэволюции, как ее понимал Янч
(см. 10.1).
Если условно разделить биологическое знание на морфологию
(учение о структурах организмов), физиологию (учение об их
функциях) и взаимодействие (экология, этология), то собственно
эволюционная проблематика соответственно разделится на проис-
хождение форм и соответствующих им (в целом) таксонов (видов,
родов и т. д.), эволюцию функций и эволюцию взаимодействий.
Трем направлениям эволюции соответствуют (нежестко) три
главных эволюционных проблемы — разнообразия, прогресса
и приспособления. . Если поместить эти проблемы в вершины
225
треугольника, то его стороны грубо представят ламаркизм, дар-
винизм и номогенез. (Науки о становлении — генетика, эмбриоло-
гия и т. и,— распределятся между тремя вершинами, смотря по
тому, становление чего изучается.) Таков познавательный
треугольник эволюционизма, центр методологической структуры
системного эволюционизма [Chaikovsky, 1988]. Треугольник
показывает, что спор между ламаркизмом, дарвинизмом и номоге-
незом беспредметен, поскольку они рассматривают разные стороны
эволюции организмов. А эволюцию сообществ вообще фактически
рассматривают вне этих частных концепций.
Естествен вопрос (поставленный мне Огурцовым в 1983 г.)
о познавательной установке, обеспечивающей цельность данной
структуры. Она видится мне в самоорганизации, понятой на основе
идей экстремальности, системной случайности и невыводимости.
Еще Мопертюи в 1744 г. показал, что в ходе механического
движения сама собой минимизируется важная величина (дей-
ствие). Впоследствии эти соображения привели к общему принци-
пу, согласно которому физическую теорию можно строить, выводя
уравнения движения из априорного экстремального принципа.
Тот же путь возможен и вне физики: в 4.1 мы рассмотрели одну
модель генетического поиска, в основе которой была конструкция
(автомат Милютина), найденная из экстремального принципа
(максимума выигрыша), а в 3.6 мы коснулись экстремальности
случайного. Аналогично, у Янча феномен приспособления рас-
сматривался как оптимизация целого процесса исторического пре-
образования биологической системы (организма, таксона или эко-
системы). Причину такого хода событий он видел в структуре
фазового пространства-времени, которое неоднородно и неизотроп-
но. Для физика это теперь естественно, а для биолога — еще нет.
Как показывает номотетико-идиографический анализ (3.2),
взаимодействие объектов непрерывно порождает случайность,
а анализ случайности (3.6) позволяет утверждать, что она при
этом будет, вообще говоря, несимметричной. При историческом раз-
ворачивании разнообразия пространство элементарных событий
(как бы ни понимать его) неограниченно расширяется, так что
набор альтернатив, из которых возможен выбор (сознательный
или самопроизвольный, здесь это неважно), можно трактовать как
симметричный в каждый данный момент, но в каждый следующий
момент тип этой симметрии усложняется. Неуклонное расширение
набора альтернатив Янч именовал «открытым обучением»
[Jantsch, 1980, с. 202], поясняя термин таким образом: если обу-
чение в школе носит закрытый характер (все цели заранее
поставлены учителем), то в творческой работе (а значит, по Берг-
сону, и в эволюции) научиться — значит достичь цели, поставлен-
ной в ходе учения, т. е. открытой цели. Иметь намерение -
совсем не то же, что знать путь к его достижению, хотя и то и другое
именуют словами «иметь цель» (ср. 4.5). Так, древний математик,
изобретая абак (подобие канцелярских счетов), хотел автоматизи-
ровать счет, эта цель достигнута в компьютерах, о которых он
226
никакого понятия не имел. Открытая цель тесно связана с пробле-
мой возникновения нового.
Дарвин ее игнорировал и лишь замечал мимоходом, что она,
как и само рождение жизни, «вряд ли касается нас», что его
учение посвящено лишь анализу постепенных изменений того, что
уже появилось. На деффект такого понимания эволюции Дарвину
указал еще в 1860 г. Брони. (Подробнее см.: [Чайковский, 1988а,
с. 96, 112].) Философ и биолог Джордж Луэс (Льюис) писал тогда
же: польза «Происхождения видов» уменьшается «от чрезвычай-
ной недостаточности философского развития» [Льюис, 1860,
с. 285], и в частности, промежуточные формы показывают лишь
нечеткость понятия «вид» и необоснованность догмы о его неиз-
менности, но еще не доказывают изменяемости. В своей последней
работе он уточнил: суть развития не столько в самой постепеннос-
ти, сколько в новизне полученного. Все новации он разделяет
на результанты (продукты действия ранее существовавших факто-
ров) и эмердженты. Эмерджент «вырастает из комбинации аген-
тов» и потому «является результантом в его компонентах», но «мы
не можем проследить шаги этого процесса так, чтобы увидеть
в продукте характер действия каждого фактора» [Lewes, 1875,
с. 412]. Так соединение водорода с кислородом дает воду.
На этой основе в XX в. вырос эмерджентный эволюционизм,
основной упрек которому (А. С. Богомолов) состоял в том, что
материальное спутано с идеальным: наше неумение проследить
и предсказать перенесено на объективный материальный процесс,
в котором идет переход количества в качество. (Анализ старой
литературы см.: [Елфимов, 1983], более новой — [Jantsch, 1980;
Пригожин, 1985].) Упрек серьезный, но игнорировавший реально
существующую параллель между возникновением новой мысли, и
нового материального объекта.
Еще одну трудность являет параллель между развитием
зародыша и преобразованием видов. Оба процесса включают
элемент эмерджентности, но первое непрерывно, а во втором мы
наблюдаем только последовательность устойчивых видов, т. е. на-
лицо системный изоморфизм, а не повторение одним процессом
другого. (Мое внимание обратила на это в 1978 г. Е. Ш. Ярмо-
линская.) В чем же тогда состоит механизм эмерджентности?
Мысль способна преодолевать невыводимость — это видно хотя
бы потому, что удается доказывать теоремы вроде теоремы Гёделя.
Если, следуя Патти (10.2), видеть в реализации биологических
процессов такую же «символическую активность», как в мыш-
лении, то несколько проясняется (я не говорю — решается)
вопрос об эмерджентности: она становится в один ряд со способ-
ностью человека доказывать теоремы. Более того, три рассмотрен-
ные выше аспекта самоорганизации (экстремальность, системная
случайность и невыводимость) выступают в качестве конкретных
атрибутов эмерджентности.
Во-первых, акт самопроизвольного возникновения (эмерджен-
ции) имеет теперь свою модель — возникновение ДС, а это значит,
что логические конструкции могут (в понятиях Патти) порождать-
227
ся не только путем логического вывода, но и некоей самосборкой,
преодолевая гёделеву невыводимость. Во-вторых, доказательство
невыводимого утверждения направляется своим результатом, чис-
той causa finalis Аристотеля, это — открытое целенаправленное
движение; но точно таким же является и движение вдоль
вариационной экстремали с подвижным концом. В-третьих, то же
можно сказать о рассуждении по аналогии, о causa exemplaris
Суареса и о реализации рефренов. В-четвертых, когда к системе
аксиом добавляется новая, чтобы сделать невыводимые утвержде-
ния выводимыми ', она выступает как произвольный выбор, т. е.
как случайность, не имеющая вероятности, вообще говоря,
несимметричная. И, в-пятых, поскольку всякий процесс течет в
своем времени, резкое замедление выступает для стороннего на-
блюдателя как стабилизация, а резкое убыстрение — как скачок.
Учтя все это, эволюцию, в том числе и эмерджентную, можно
понимать как открытое обучение, как преобразование разнооб-
разия, детерминированное открытой целью.
Янч простодушно полагал, что эволюционное время подобно
четвертой координате эйнштейнова континуума, но ведь там нет
движений, а есть лишь следы их — траектории. «В таком четырех-
мерном мире всё, что должно быть в соответствии с законами
Природы, уже существует реально... Однако такая картина фаталь-
ного детерминизма противоречит свободе выбора и всему опыту
нашей жизни. Скорее всего это означает, что чисто геометрическое
представление времени является недопустимо упрощенным»
[Козырев, 1982, с. 59—60]. Мейен [1984, с. 11] признал недоста-
точным и физическое определение времени, предложил понимать
время как изменчивость индивида. Не как меру изменчивости,
а как саму многомерную изменчивость. Подробнее о гео- и биовре-
мени см/. [Мейен, 1989], а мы заметим лишь, что эволюционное
время многомерно. Так, «Гея» Лавлока обладает целостностью
и эмерджентностью [Lovelock, 1989], которая может проявляться
в разных астрономических временах — как в очень медленных
биосферных ритмах (см. 11.2), так и в относительно быстрых актах
подстройки класса насекомых и класса птиц (5.2, 5.3). Такое пони-
мание времени представляется более содержательным, чем
обычное [Пригожин, 1985, с. 246] понимание внутреннего времени
как среднего.
Таков эскиз концептуальной схемы биологического эволюцио-
низма, схемы, которая сможет претендовать на охват (с единой
позиции) основной эволюционной тематики. Без такой или по-
добной схемы эволюционизм будет вынужден, как и прежде,
служить для «закрытого обучения» уже известному, но не инст-
рументом выживания в быстро и угрожающе эволюционирующем
мире. Хотя до такой теории-инструмента еще далеко, но и сейчас
некоторые результаты налицо, что мы увидим в части III.
1 Таковы пятый постулат Эвклида, постулат Эйнштейна о равенстве инерционной
и гравитационной масс и т. д.; естественно полагать, что и эвкариотная органи-
зация не выводится из прокариотной.
Часть III. ПРИМЕНЕНИЕ ПРИНЦИПОВ
ДИАТРОПИКИ
Глава 11. ПЕРЕОРИЕНТАЦИЯ ИССЛЕДОВАНИЙ
ЭВОЛЮЦИИ
Все наземные растения принято считать потомками зеленых во-
дорослей, а их — потомками каких-то прокариот. Прокариотные
«водоросли» (цианеи) хорошо известны, но сходны не с зелеными,
а с красными водорослями (багрянками). Сходство состоит
в общности некоторых пигментов и в отсутствии жгутиков (так
что одиночные клетки не могут направленно плавать), тогда как
у зеленых водорослей жгутики есть. Багрянки устроены очень
своеобразно (их часто выделяют даже в особое подцарство), но
ничуть не проще других водорослей. Однако открытие в 1971 г.
«жгутика» у багрянки стало сенсацией, ибо поставлен был вопрос:
не багрянки ли предки всех водорослей, а с тем - и наземных
растений? «Жгутик багрянки» стал популярен в литературе, хотя
на последней и самой четкой микрофотографии ясно видно, что
речь может идти вовсе не о жгутике, а о невиданной структуре:
два изогнутых тяжа действительно похожи на сложенные жгутики,
но расположены внутри клетки и вроде бы прикреплены к ядерной
(а не клеточной) оболочке [Simon-Bichard-Breaud, 1972]. Разу-
меется, когда были открыты более «приемлемые» предки зеленых
водорослей — прокариоты, сходные пигментами и с цианеями, и с
зелеными водорослями [Withers е. а., 1978], о необычной багрянке
быстро забыли, как постоянно забывают и о том, что пропасть
между прокариотами и эвкариотами заполнить нечем.
Как это, увы, часто бывает, замечательные приборы фактически
сработали вхолостую: сама по себе невиданная структура мало
кого заинтересовала — «жгутик» был интересен только как свиде-
тельство о родстве. Его искали у близких видов багрянок и не
нашли — это окончательно охладило пыл «искателей предка».
Даже открытие у багрянок других семейств еще каких-то нео-
бычных внутриклеточных структур, трактуемых как «рудименты
жгутиков» [Tripodi, Masi, 1983], не привлекло внимания. Подоб-
ные структуры были истолкованы как полезные при разделении
мембран клетки и при переработке ее содержимого [Broadwater,
Scott, 1983], на чем дискуссия и выдохлась.
А ведь сам факт куда интереснее, чем свидетельство в пользу
гипотетического предка (наверняка непохожего на реального пред-
ка водорослей, жившего 2 млрд лет назад) или в пользу приспособ-
ления (без которого обходятся все прочие клетки). Что, если
229
зафиксирован редчайший акт эволюционного эксперимента?
Этот вопрос в головы не пришел, ибо эволюционисты привыкли
искать либо родство, либо приспособление. Исследовательская
программа, в дни Дарвина звавшая вперед, теперь помогла
закрыть глаза на новое. В точности по Лакатосу: вырожденная
программа вместо новых вопросов предлагает только вереницы
однотипных ответов на вопросы рутинные. Чем может здесь помочь
диатропика?
11.1. Выявление тенденций
Дело не в данном эксперименте (он может оказаться просто ошиб-
кой), а в установке теоретиков принимать на веру любой удобный
результат (мы говорили об этом в связи с работами Тинбергена
и Эйгена) и отвергать результаты, чем-то неудобные, не ложа-
щиеся в схему. Вопреки общепринятому мнению, что аккуратный
факт не стареет, мы то и дело видим, как факты стареют и умирают
по мере смены их интерпретаций. Если 50 лет назад почти все
считали, что «для своего времени Дарвин учел весь точно установ-
ленный материал биологии и геологии и дал поэтому исчерпываю-
щую теорию» [Комаров, 1937, с. 10], то теперь многие науковеды
вообще не находят у Дарвина фактов в ключевых местах теории,
а находят метафоры [Young, 1971], игнорирование морфологии
и эмбриологии [Goodwin, Webster, 1981], бездоказательные рас-
суждения [Gale, 1982] и т. п. Сами по себе упреки не новы —- их
высказали еще первые критики Дарвина (литературу см.:
[Чайковский, 1987а, с. 100]),— но тогда критика противоречила
основным симпатиям общества и была им быстро забыта. Впос-
ледствии не раз критика фактической обоснованности дарвинизма
возобновлялась. Так, книга Данилевского [1885] была финалом
уже второй волны такой критики. Об этой и следующей волне
можно прочесть у Филипченко [1977]. Подобные волны возникали
и позже (вспомним успех работ Любищева, недавний всплеск
креационизма в США, нынешние споры вокруг пунктуализма и
многое другое), поэтому можно говорить о своего рода тенденции,
проявляющейся в регулярных сменах научной моды. Однако
развитие науки состоит не в этих колебаниях и не в упорном
следовании одному из течений, а в некоторой равнодействующей,
выявить которую должна методология. В западной литературе
сейчас о сменах моды на дарвинизм пишут многие науковеды, но
на саму биологию это пока почти не повлияло. Как отмечал Мейен
[1987в], впечатление динамичности теоретической биологии об-
манчиво, в ней давно — застой, господство стремления доказать
свою правоту над стремлением решить фундаментальную пробле-
му. В том же сборнике палеоботаник В. А. Красилов выразился
еще резче: по его мнению, общебиологические проблемы стоят
там же, где их оставили натурфилософы прошлого.
Эти мысли следует уточнить. Да, в теоретической биологии
«равнодействующая» за последние 60 лет сдвинулась поразительно
230
мало (по сравнению со многими другими науками), й некоторые
методологи (например, В. В. Налимов) склонны даже считать,
что теоретической биологии просто нет. Да, после смерти Дарвина
мы почти не видим в биологии того, что прежде именовалось
натурфилософией, а ведь именно натурфилософы ставили (пусть
и в наивной для нынешней науки форме) основные теоретические
вопросы. Однако все это верно лишь по отношению к ядру био-
логии, как оно выглядит в учебниках и руководствах, тогда как на
ее периферии натурфилософская мысль никогда не затухала и,
в частности, видными ее представителями можно считать как раз
Мейена и Красилова. (Под натурфилософией зде.сь понимается
построение широких, обобщающих картин природы, не противо-
речащих фактам и даже базирующихся на фактах, но рисуемых
не «от факта к факту», а исходя из априорных установок, новых
для науки. Факты при этом играют, в основном, роль иллюстра-
ций. Так, Мейен, будучи вообще-то скрупулезным фактографом,
выступил как натурфилософ, когда предложил [Мейен, 1978]
описывать всю биологию в терминах мероно-таксономического
анализа.) Как и прежде, разнообразие биологических теорий вы-
растает из натурфилософских схем (обычно — конкурирующих),
как и прежде, срок роста часто измеряется столетиями (так,
господствующая сейчас в науке идея микроэволюции путем из-
менения дискретных единиц наследственности довольно ясно
сформулирована натурфилософами еще в XVII в. [Hale, 1677]),
но только срок этот непомерно велик для нынешних темпов раз-
бухания научной литературы. Другими словами, разнообразная
натурфилософская периферия стала непомерно малой для однооб-
разного респектабельного ядра науки, и потому обычный
защитный аргумент ретроградов: «Подавляющее большинство спе-
циалистов в этом уверенно»— приобрел непомерно большую силу.
Аргумент апеллирует к тенденции, но кроме тенденций существу-
ют рефрены, не связанные с массовостью, однако столь же
важные для процесса познания. Если какая-то позиция рефрена
стала массовой, это может стать главным для соответствующей
эпохи, но быть маловажным для исторического процесса в целом.
Примеров не счесть — как в самой эволюции, так и в науке о ней.
Проводя всевозможные параллели и строя сквозные ряды,
диатропйка постоянно сравнивает большое с малым (ядро с пери-
ферией) . Это позволяет, в частности, преодолеть традиционное для
прежнего эволюционизма препятствие, на которое указывал Тейяр.
По его словам, принято закономерности, отмеченные «в среднем
масштабе наших организмов и наших сооружений», считать основ-
ными, а все остальное — исключениями. Самый поразительный
его пример — пренебрежение феноменом человеческой мысли как
чем-то чуждым жизни в целом: «Причудливое исключение, стран-
ная функция, эпифеномен — мысль характеризовалась как-нибудь
так, чтобы от нее избавиться» [Тейяр, 1987, с. 54]. В то же время
истинная задача научного анализа, по Тейяру, в том, чтобы
«открыть в исключительном всеобщее», чтобы понять фундамен-
231
тальное: «В глубине нас самих, бесспорно, обнаруживается
внутреннее... Этого достаточно, чтобы в той или иной степени это
внутреннее существовало везде и всегда в природе. Раз в одной
точке самой себя ткань универсума имеет внутреннюю сторону,
то она неизбежно двустороння по самой своей структуре» [там
же, с. 55]. В сущности, именно эту мысль приняла теперь физика
(и биофизика) неплоских морфизмов (см. 10.2), а диатропика
пользуется (при построении рефренов) подобными приемами
обнаружения «в исключительном всеобщего» постоянно.
В связи с этим следует сказать об изменении понимания
термина «закон эволюции». По классификации историка эволюцио-
низма С. А. Орлова законы эволюции делятся на три группы:
«законы, определяющие организацию субстрата эволюции; законы,
детерминирующие процесс развития; и законы организации самого
этого процесса», причем отличие третьей группы в том, что ее
законы «проявляются не в каждом отдельном элементарном
эволюционном явлении, а лишь в определенной последовательнос-
ти их совокупности», что «это — законы-тенденции, реализующие-
ся в зависимости от условий с различной полнотой» [Орлов,
1979, с. 101]. Все сказанное ранее позволяет добавить: многие
законы диатропики тоже выявляются как тенденции, хотя, по
системе Орлова, в основном относятся к первой группе. Это приво-
дит к выводу, что выявление тенденций становится теперь одним
из главных направлений эволюционизма, тогда как прежде
тенденции считались лишь «эмпирическими обобщениями», чем-то
низшим по сравнению с законами, с теорией. Орлов справедливо
отметил, что «само деление обобщений на теоретические и эмпи-
рические весьма шатко» [там же], и теперь мы можем уточнить:
оба вида обобщений являются типологическими экстраполяциями
по Мейену, других обобщений в описательных науках, по-видимо-
му, просто нет.
Наиболее известны тенденции таксонов развиваться в опреде-
ленном направлении — зоолог Вильгельм Гакке объединил их
в 1893 г. термином «ортогенез». «К несчастью, этот термин приме-
няется не только как название определенного явления, но и для
обозначения теорий, имеющих целью его объяснение» [Симпсон,
1948, с. 232]. Если говорить об ортогенезе только как о явлении, то
его ярким примером явится гнездовой паразитизм кукушек (4.6)
и другие тенденции, о которых стараются не вспоминать, напри-
мер — метаморфоз камбал.
В 4.5 мы говорили о том, что камбалы поразительны умением
быстро мимикрировать, но у них (как и у кукушек) поразительно
сразу многое. Молодая камбалка нормальна формой и нормально
плавает в толще вод, но взрослая претерпевает метаморфоз: ее
высота (от спины до брюха) становится больше длины (от рта до
хвоста), и она буквально падает на один бок, который становится
«брюхом» и светлеет. Это посветление рассматривалось как дли-
тельная модификация, что-то вроде утраты загара [Martin, 1956],
подвергались анализу и другие черты метаморфоза порознь. Падение
232
на бок традиционно трактуется как «приспособление к донному
образу жизни», хотя другие донные рыбы приспосабливаются куда
проще. Глаз, оказавшийся на «брюшной» стороне, перемещается
на «спинную», и еще Дарвин трактовал это чисто ламаркистски —
глаз якобы стремится смотреть вбок и оттого сползает [Дарвин,
1939, с. 442]. Все это ни на шаг не приближало к пониманию
своеобразия камбал. Наоборот, ихтиолог Данилевский подробно
рассмотрел вопрос в целом. Отвергнув приспособительное объясне-
ние метаморфоза как не соответствующее фактам (в частности,
неуклюжие камбалы с трудом охотятся, и в этом им мешают как
раз глаза, «напрасно смотрящие наверх; где уже камбалам ничего
не добыть»), он воскликнул: «Не очевидно ли, что это произошло
не вследствие применения к внешним условиям жизни, а по
типу, сообразуясь с которым, как с чем-то данным, эти рыбы
живут, как могут» [Данилевский, 1885, ч. 2, с. 48, 229]. Объясним,
что значит «по типу».
Молодая камбалка внешне зеркально симметрична. От этой
симметрии затем мало что остается, зато появляется нечто вроде
новой симметрии — бывшие спина и брюхо (теперешние бока)
становятся похожи друг на друга. Вся рыба выглядит, как отражен-
ная в кривом зеркале. Наложение новой симметрии на прежнюю
(как бы скрещение двух онтогенетических программ) никак не
назовешь приспособлением к среде, это — тенденция, охватившая
отряд камбалообразных. Как и кукушки, камбалы выработали
много приспособлений к тенденции, перекосившей их тела: мимик-
рию, плавание «плашмя» (глазной стороной кверху) и т. д. Для
приспособлений камбалы характерна высокая обратимость: она не
только меняет свою расцветку, но и может в случае опасности
плыть «стоймя» (спинным плавником кверху) и очень быстро.
Вообще метаморфоз у рыб нередок и иногда несет явные следы
приспособления к тенденции: такова, например, луна-рыба, в ходе
метаморфоза компенсирующая (за счет изгиба назад непарных
плавников) утрату хвостового плавника. Необычно у камбал нару-
шение зеркальной симметрии, но, как отмечает Петухов [1981,
с. 36—37], видимое нарушение симметрии вообще часто связано
с переходом к симметрии высшего типа, выявляемой лишь
достаточно изощренной математикой. Смысл же поиска таких
симметрий в том, что ищутся универсальные принципы морфогене-
за, которых, вероятно, немного, и комбинацией которых
реализуется разнообразие форм.
Наиболее очевиден комбинаторный характер онтогенеза как раз
у растений, поскольку у них нет ни централизации (6.6), ни (веро-
ятно) первичных половых клеток (7.2), а это порождает частые
ситуации, когда «уже отработанная онтогенетическая программа
включается на новом месте и производит здесь новый орган»
[Меуеп, 1987, с. 356]. Онтогенез нельзя описывать в терминах
последовательных делений клеток, он прежде всего — последова-
тельность преобразований симметрий, топологии и физиологии
[Петухов, 1981; Аршавский, 1982; Преснов, Исаева, 1985]. Не
233
является онтогенез и реализацией какой-то готовой программы —
генетическая «программа», как теперь выясняется (см. 7.2), лишь
ограничивает варианты самосборки. Если так, то «жгутик» баг-
рянки мог появиться в результате переключения онтогенеза клетки
за счет мутации или случайной интеграции фрагмента чужеродной
хромосомы. Переключений естественно ждать там, где работа
репродуктивной системы разбалансирована (см. 4.6), и поэтому
необычные формы необычны сразу во многом.
Еще выше, чем у растений, комбинативность прокариот — это
естественно связать с тем, что консерватизм их наследственности
(основное свойство живого по Янчу — см. 9.4) вообще невысок.
Возрастание консерватизма наследственности в ходе эволюции —
важнейшая тенденция, выявление которой проливает некоторый
свет и на происхождение жизни.
Как говорилось в 9.4, первой субвитальной единицей можно
считать ту ДС, в которой синтезировались РНК, служившие для
синтеза пра-белков. Дальнейшая тенденция состояла в том, что на
каждом уровне развития изготовлялась консервативная структура,
служившая затравкой для следующего уровня. Во всех организ-
мах для синтеза белков служит рибосома, и в субвитальной еди-
нице должен был существовать ее пептидо-нуклеотидный предше-
ственник. Он может считаться той затравкой, которая изготовлена
первым уровнем преджизни и служила для порождения второго,
заведомо более разнообразного молекулами. Допустим, что уже на
втором уровне ДС порождали двуспиральную РНК (хотя могли
быть и промежуточные уровни). Спирализация РНК породила но-
вый (третий) уровень — тот, где появилась примитивная наслед-
ственность. Этот уровень ДС можно отождествить с эобионтом.
Четвертым и пятым уровнями могли стать появление ДНК и мемб-
ран (что раньше — сказать трудно). ДНК обеспечила эобионту ту
устойчивость информации, при которой одного экземпляра ее до-
статочно для сохранения и передачи свойств — появился ген.
А мембрана послужила затравкой для синхронизации и локализа-
ции биохимических процессов, без чего нельзя говорить о размно-
жении. Обладателя этих уровней уместно назвать термином проге-
нот, входящим сейчас в научный лексикон [Воронцов, 1987]. Нако-
нец, замыкание мембраны' в изолированную полость превратило
прогенота в прокариота (шестой уровень).
В течение следующего миллиарда лет прогрессивная эволюция
состояла в совершенствовании биохимических механизмов и преж-
де всего — энергетики. Какова была энергетика эобионта и проге-
нота, мы сейчас не знаем, а у нынешних прокариот находим ее уже
вполне совершенной. Простейшие из них — микоплазмы — полу-
чают энергию путем гликолиза, процесса все-таки очень сложного.
Остается лишь строить догадки о его происхождении, и до сих пор
единственным приемом служил поиск отдельных более примитив-
ных экзэргических реакций — некоторые бактерии,-в общем, гораз-
до более сложные, чем микоплазмы, добывают энергию путями,
которые в 2 — 3 раза короче гликолиза [Панцхава, 1973]. Однако
234
и здесь участвуют сложные молекулы-посредники, которых в
начале жизни не было; возможно, что удастся найти им неоргани-
ческие аналоги или указать возможность протекания подобных
реакций с более простыми посредниками, какие могли быть синте-
зированы уже эобионтом или прогенотом. То же самое можно
сказать о попытках понять рождение фотосинтеза. По-моему, стои-
ло бы попробовать выстроить цепочку уровней реакций, так, чтобы
каждый уровень порождал (при содействии пептидо-нуклеотидной
памяти) затравку для следующего уровня.
Зато достаточно очевидны следующие (седьмой и восьмой)
уровни генетической эволюции: появление ядерной оболочки и
объединение ДНК с белком (у динофлагеллат — с РНК) в ком-
пактную хромосому повысили консервативность генов еще больше,
что послужило к образованию эвкариот, описанному в главе 8.
Все ядро служит затравкой для клеточных процессов. Существен-
но, что прокариотный тип организации генов не был при этом
отброшен, и до сих пор мы видим, как работают затравки, иногда
именуемые якорями (anchors) [Bogorad, 1975]: митохондрия и
хлоропласт имеют свою ДНК прокариотного типа, которая коди-
рует не сами компоненты этих органелл, но лишь их фрагменты
(якоря), которые притягивают к себе (сродство!) все, что нужно
для сборки органелл и что синтезировано с помощью ядерной ДНК.
Кстати, этот феномен гипотеза симбиогенеза вынуждена игнориро-
вать, а схема Янча вполне объясняет, хотя сам Янч о «якорях»
не подозревал.
11.2. О становлении систем
Коэволюционная концепция рассматривает эволюцию данного объ-
екта (например, вида) как ячейку в общем процессе, и это, пожа-
луй, главное отличие от прежнего эволюционизма Ламарка-
Дарвина—Берга, сосредоточенного на эволюции отдельных видов,
якобы изменяющихся в довольно безликой среде. Экологи гово-
рят о коэволюции давно, а сам термин предложен в 1964 г. [Одум,
1986; Vermeij, 1987], но у них это понятие означает лишь взаимное
приспособление видов и не повлекло концептуальных изменений,
тогда как расширение смысла понятия Янчем отражает новое пони-
мание эволюции.
Об одностороннем «приспособлении к среде» можно говорить
только в отношении грубых физических факторов (географическая
широта, высокогорье, пустыня, рассол и т. п.), причем сказать здесь
можно не так уж много, поскольку почти в каждой крупной физи-
ческой зоне представлено все разнообразие крупных таксонов —
от бактерий до цветковых и зверей. Даже в таких специфических
и небольших зонах, как подводные горячие источники, мы видим
огромное разнообразие — от бактерий до моллюсков. Говорить о
том, что какое-то разнообразие порождено приспособлением к сре-
де, можно лишь в отношении низших таксонов, да и то с массой
оговорок. В остальном разнообразие порождается коэволюцией,
т. е. речь идет о становлении систем как целого. Даже вода не
235
является абиотическим параметром: грибы находят нужную им
влажность в тени деревьев, а сами деревья часто не могут жить без
грибов; как мы знаем хотя бы на примере катастрофического
наводнения в Бангладеш (сентябрь 1988 г.), замена лесов травами
ведет к трагическим изменениям климата, к бесконтрольному стоку
вод.
Необходима, как мы не раз говорили выше, переориентация
эволюционной мысли со статистического подхода на системный,
что дается с большим трудом. Даже такие, казалось бы, антиподы
эволюционной мысли, как Дарвин и Вернадский, оба выражали ста-
тистический взгляд на эволюцию, поскольку оба мыслили ее в рам-
ках баланса. В 1844 г. Дарвин писал: «Природу можно сравнить
с поверхностью, в которую вбиты десятки тысяч клиньев, располо-
женных вплотную и вгоняемых внутрь беспрестанными ударами»
[Дарвин, 1939, с. 136]. Аналогичное место есть в 1-м издании «Про-
исхождения видов», а из 2-го издания вычеркнуто — Дарвин, види-
мо, понял, что равновесие такой «поверхности» было бы неустойчи-
во; но он ничего не предложил взамен, Вернадский, наоборот,
видел в природе полную устойчивость, поскольку (если пользо-
ваться терминами Дарвина) считал «поверхность» неизменной,
сохраняющей «форму» независимо от последовательной смены
«клиньев». Уточним сказанное.
В 8.1 уже говорилось, что основой эволюции биосферы Вернад-
ский считал замкнутые круговороты, не меняющиеся при смене
одних видов организмов на другие. В этом он следовал Бэру [Вер-
надский, 1980, с. 76], сформулировавшему «закон бережливости»,
согласно которому элементы в течение миллионов лет не покидают
биогенных циклов [Вальт, 1978, с. 118]. Выход органики из круго-
ворота Вернадский считал незначительным, а прочие отложения,
по-видимому, полагал пренебрежимо малыми. Теперь мы знаем,
что это не так (9.2) и что в отдельные периоды истории Земли
скорости накопления осадков резко возрастали при одновременном
сокращении видового разнообразия [Неручев, 1988]. «Бережли-
вость» безусловно имеется, но в форме спиралей, а не балансов, да
и спираль может быть иногда смята катастрофой.
Ни у Дарвина, ни у Вернадского эволюция не понималась как
возникновение — статистический подход давал им возможность
обсуждать только переход одного живого в другое, без качествен-
ных новшеств. И само рождение жизни они обсуждать отказыва-
лись, а последующее развитие подчиняли идее баланса. Поэтому
полезно вспомнить Бэра, который посвятил свой первый эволю-
ционный доклад (1834 г.) вопросу — что можно считать постоян-
ным в процессе развития жизни на Земле. Ответ его был прост:
постоянной является не какая-то величина, а тенденция разви-
тия — «все время идущая вперед победа духа над материей». Ана-
лиз доклада см. у Ю. А. Филипченко [1977]. Сейчас, после
«эмерджентной парадигмы» Янча, мы можем сказать большее:
очевидна тенденция материи образовывать все новые и новые уров-
ни самоорганизации.
236
Не то чтобы о тенденциях вообще после Бэра не думали —
о них писали Брони, Бергсон, Вернадский и другие, но подобные
мысли были часто как бы подчинены господствовавшим идеям
выживания. Например: «Под влиянием концентрации или недо-
статка химических элементов создаются новые расы, виды и под-
виды» или: «При эволюции видов выживают те организмы, которые
своею жизнью увеличивают биогенную геохимическую энергию»
[Вернадский, 1980, с. 123, 260]. Разнообразие видов не представ-
лялось ему самодовлеющей сущностью. Сейчас, наоборот, ученые
понемногу склоняются к мысли, что без разнообразия большие
системы вообще быть поняты не могут. Всякое разнообразие —
срез (временной, функциональный, аспектный и т. д.) какой-то
системы.
Именно на этих идеях строится понемногу и современный эво-
люционизм, но возникают большие трудности, касающиеся пони-
мания одними эволюционистами других. Это прежде всего отно-
сится к эволюционной термодинамике. Некоторые ведущие эволю-
ционисты (например, Аршавский) полагают неприемлемым под-
ход группы Пригожина, а первую книгу, трактующую биологи-
ческую эволюцию термодинамически [Brooks, Wiley, 1986]. рецен-
зент назвал бессмыслицей [Morowitz, 1986]. Можно согласиться,
что нет большого смысла в таких, например, фразах: оплодотво-
рение «задает новую точку равновесия, к которой развивающийся
зародыш спонтанно движется в согласии со вторым законом» тер-
мостатики; или: «Статистическая энтропия сообщества может быть
выражена как функция относительного числа организмов каждого
вида и их экологических взаимодействий» [Brooks, Wiley. 1986,
с. 82, 264]. Ведь движение к термостатическому равновесию озна-
чает распад структур, а не их развитие; и «энтропия сообщества»
вряд ли имеет смысл, если вводить ее по аналогии с энтропией
газа: вспомним хотя бы, что последняя определена для хаотическо-
го движения молекул сосуда при отсутствии потоков через сосуд,
а экосистема существует в потоке веществ и энергии, существует
за счет целесообразной деятельности организмов. Отождествление
разнообразия с энтропией [там же, с. 287] вообще несерьез-
но. Однако не в этом суть термодинамического подхода к био-
логии.
Начиная с первой работы [Prigogine, Wiame, 1946] и вплоть
до работ Янча (которого Д. Брукс и Э. Вили едва упомянули в
общем перечне) данный подход всегда использовал термодинамику
для выявления параллелей развития между живыми и неживыми
системами, т. е. для выявления системных свойств, а не для описа-
ния биологии в физических терминах (что, по-моему, вообще смыс-
ла не имеет). В той мере, в какой наблюдается градация уровней
ДС (см. 9.1), неизбежно и развитие, поскольку каждая ДС данного
уровня существует лишь характерное для нее время, а затем
переходит в дочернюю ДС или распадается. Это обстоятельство по-
могает понять генезис систем вообще, в том числе и тех, где
теоретики обходятся без понятия энтропии. В частности, в любом
237
системогенезе следует выявлять характерные времена, и одно это
уже может сильно прояснять суть дела.
Это, кстати, хорошо понимают Брукс и Вили. Прежней точке
зрения на «баланс природы», якобы достигаемый противостоянием
разнонаправленных сил, они противополагают другую точку зре-
ния, согласно которой силы, сравнимые по порядку величины и
скорости изменения, не конкурентны, а комплементарны (в этом,
добавим, и состоит еще одно понимание коэволюции, соответствую-
щее «встрече» Янча): однако они определяют лишь общее направ-
ление развития, тогда как второстепенные феномены могут про-
текать и в иных направлениях [Brooks, Wiley, 1986, с. 299—300].
Поясняют это авторы таким примером: по отношению к более быст-
рым процессам мутирования более медленные процессы измене-
ния скрещиваемости (дивергенция одного вида на два) могут
выглядеть противоположно направленными. С этой позиции они
критикуют «синэргетику» Г. Хакена, справедливо упрекая его в
попытке истолковать новые, системные, термины в рамках идеоло-
гии неодарвинизма и подчеркивая, что требуется новая (систем-
ная) идеология, а не смена терминов.
Поясним значение характерных времен на примере проблемы
становления биосферы. Как уже говорилось в 8.1, жизнь должна
была возникнуть в ходе коэволюции биосферы и клетки. В запад-
ной литературе эту коэволюцию часто трактуют в рамках «гипоте-
зы Геи», которую в 1970-х гг. предложил физик Джеймс Лавлок:
он полагает, что условия для жизни, в том числе океан, атмосфера
и почва, сделаны жизнью самой для себя, поскольку вся Земля
является единой кибернетической системой. Лавлоку возражает
эколог М. И. Будыко, считающий, что влияние жизни на Землю
как целое ничем не подтверждено, что появление и дальнейшее
существование жизни есть следствие совпадения большого числа
редких случайностей, а потому, вероятно, жизнь уникальна во Все-
ленной [Будыко, 1984, с. 437—452]. Так, появление на Земле
жидкой воды 4 млрд лет назад он связывает не с жизнью, а с нагре-
ванием атмосферы за счет «парникового эффекта», вызванного
избытком в ней углекислого газа; позже, когда эффект исчез,
«как раз» усилилось солнечное излучение, а затем, когда Земля
была готова вновь замерзнуть, «как раз» усиливалась активность
вулканов, вновь добавлявших углекислый газ в атмосферу, и т. д.
Биосфера кажется ему малоустойчивой. Это — типичное рассужде-
ние в духе статистического мировоззрения.
Подобными «как раз» до недавнего времени научные гипотезы
изобиловали. Вспомним антропный принцип в космологии (9.4)
или объяснение феномена хиральности живого (преобладания
левых аминокислот и правых сахаров) как случайности. Однако
теперь надеются, что хиральность удастся понять термодинами-
чески [Кизель, 1985], а антропный принцип охарактеризован Ян-
чем как «поспешное объяснение в духе Дарвина», игнорирующее
системность Вселенной.
Сам процесс рождения жизни должен иметь характерное время.
238
По данным геологии оно не превышает полумиллиарда лет, и те,
кто считают этот срок малым, склонны приписывать земной жизни
космическое происхождение. При этом забывают, что 4 млрд лет
назад высокоразвитой (готовой к космической экспансии) жизни
просто неоткуда было взяться, поскольку нынешняя космогония
видит возможность появления планет с достаточно разнообразной
химией лишь у звезд второго поколения, подобных Солнцу. С дру-
гой стороны, полумиллиарда лет «мало» только с позиций прежне-
го, статистического, эволюционизма и то — в предположении, что
на молодой Земле все процессы текли так же, как на нынешней.
Это — в терминах Мейена,— неудачная типологическая экстрапо-
ляция.
Статистический подход отрицал связь явлений всюду, где не
видел ее, тогда как системный подход ищет ее всюду, где налицо
согласованность. Да, мы не знаем механизма влияния биосферы на
активность вулканов, но это не значит, что его нет. Ритмы биосферы
измеряются миллионами и миллиардами лет, они действительно
не могут поспевать за быстрыми (десятки тысяч лет) климатиче-
скими факторами, но вполне могут согласовываться с ритмами
горообразования и дрейфа континентов. Усиление фотосинтеза
снижает парниковый эффект, отчего у полюсов скапливаются гро-
мадные массы льда (до 5% объема Мирового океана), т. е. проги-
бается земная кора и ускоряется вращение Земли. Все это не может
не влиять на активность недр. Другими словами, медленные прог
цессы (например, оледенение в конце палеозоя и потепление в
мезозое) в принципе могли определяться самой биосферой. Вопрос
еще требует изучения. Отметим в связи с этим, что, по Одуму
[1986, т. 1, с. 38], «гипотеза Геи указывает на важность изуче-
ния и сохранения регулирующих механизмов» биосферы, причем
Одум советует для понимания этой проблемы прочесть также книгу
Янча.
Микробиолог Г. А. Заварзин [1984], наборот, не видит в гипо-
тезе Лавлока ничего нового, кроме названия, полагая, что основные
ее идеи можно найти у Вернадского. По Заварзину, жизнь могла
возникнуть вовсе не в океане, а в горячих потоках, исходивших из
недр Земли. До сих пор в горячих источниках наблюдаются своеоб-
разные примитивные экосистемы, использующие, в отличие от
всех других систем, не продукты разложения других организмов и
не солнечную энергию, а потоки из недр. Возможно, что они
способны существовать сами по себе, образуя как бы собственные
небольшие биосферы, понемногу снабжая большую биосферу орга-
никой.
Если так, то схема коэволюции (по Янчу) должна быть дополне-
на: кроме макро- и микроуровней существует еще мезоуровень,
играющий роль реактора, в котором элементы микроуровня рожда-
ются и преобразуются; реактор снабжает новыми элементами си-
стему макроуровня. В первичной (а возможно и в последующей)
биосфере мезоуровень обеспечен системой горячего источника,
в звезде — конвективной зоной ядерного синтеза и т. д.
239
По Г. А. Заварзину [1976], первая экосистема могла сложиться
из прокариот, потреблявших восстановленные вещества недр и
окислители атмосферы (например, кислород, возникший при раз-
ложении паров воды солнечным светом). Через миллиард лет она
могла погибнуть вследствие образования озонового экрана, сделав-
шего невозможным фотолиз воды у земной поверхности, но обеспе-
чить рождение биосферы и, в частности, фотосинтеза (который
мог сперва возникнуть в виде простых реакций защиты от облу-
чения). Еще через миллиард лет случилась вторая катастрофа —
повышение концентрации кислорода в атмосфере, позволившая
выжить на открытом воздухе и в толще вод лишь аэробам. Наобо-
рот, анаэробы, т. е. не переносящие кислорода организмы (в основ-
ном они представлены бактериями), существуют лишь огражден-
ные от кислорода химическими и геохимическими барьерами.
В связи с этим Заварзин поставил важный вопрос о создании
искусственных геохимических барьеров — например, ограждаю-
щих разнообразие дикой природы от однообразия окультуренной.
Это — яркий пример переориентации эволюционного мышления,
полезной для практики.
Глава 12. РАЗНООБРАЗИЕ И ПРОГНОЗ
12.1. Экологический диктат и замыкание затрат
Нынешняя охрана природы необходима, но скорее демонстрирует
озабоченность общества, чем служит преодолению кризиса. Исто-
рия знает ряд примеров, когда озабоченность не спасала. Так,
Италия еще в древности и в средние века знала законы, ограждав-
шие леса от чрезмерной вырубки, однако беспокойство законода-
теля ничего не спасло: равнинные леса в Италии почти исчезли.
В своем нашумевшем «Закате Европы» (1918 —1922) Освальд
Шпенглер связал даже само крушение Римской империи с тем,
что сейчас называют экологическим кризисом.
Надежную программу может дать только хорошо обоснованная
теория. Чаще всего в этой связи называют экологию, но правильнее
будет сказать, что спасение природы (как и повреждение ее) —
не отрасль знания или хозяйства, а аспект человеческой деятель-
ности вообще. Аспектом (познания) является и диатропика, и нас
интересует пересечение этих двух аспектов. Разнообразию явлений
надо сопоставить (но не противопоставить) разнообразие средств.
Общий пересмотр отношения к природе требует выработки та-
ких оснований каждой науки, какие будут объединять все науки
для единого дела. Пока что мы этого не видим. Так, для биолога
охрана природной территории означает снижение давления
эксплуатации до той величины, какая требуется, чтобы территория
сохраняла свой естественный набор растений и животных: для эко-
240
номиста же охрана этой территории означает набор мер, необхо-
димых для сохранения ее продуктивности. Могут ли они найти
общий язык?
Могут, но только при условии, что будут решать одну общую
задачу, а не каждый свою. Эту общую задачу и должна сформули-
ровать будущая теория. Представление о способе объединения двух
задач в одну может дать двойственная пара задач оптимизации ',
в каждой из которых оптимизируется своя функция, ограничения
на которую берутся из сопряженной с ней задачи. Так, биолог (если
он не фанатик какого-то одного любимого им вида организмов —
увы, такие тоже бывают) требует, чтобы численность каждого вида
на территории не падала ниже некоторой заданной величины,
а экономист — чтобы территория давала урожай (добычу) каждого
продукта не ниже требуемого. Роль математика здесь прежде
всего в том, чтобы определить, совместимы ли эти ограничения,
существует ли хоть одна стратегия, всем им удовлетворяющая.
Если нет, то какие-то ограничения надо ослабить — либо ослабить
эксплуатацию, либо смириться с исчезновением какого-то вида.
Только тогда, когда приемлемых стратегий оказывается больше
одной, встает задача оптимизации. Другими словами, в норме
оптимизируется нечто второстепенное (с данной точки зрения),
а главное задается граничными условиями (абсолютными запрета-
ми). Если же каждый готов оптимизировать «свое», значит систе-
ма распадается. Тогда говорят о хищнической добыче, тогда же
рождается и тактика вытеснения нежелательного предприятия на
«чужую» территорию. Этот безнадежный режим и соответствует,
в общем, нынешней тактике охраны природы, тогда как для спасе-
ния природы нужна новая стратегия, основанная на новой этике,
этике двойственной оптимизации, когда каждый участник задает
«свое» в качестве ограничений для других, а сам оптимизирует
в «чужих» ограничениях. Естественно, что для этого необходим
еще и властный посредник, задающий участникам приемлемые
законы, но не вмешивающийся в конкретные решения, если они
законны. Подробнее см.: [Чайковский, 1989в].
Возможна ли такая гармония? Вспомним описанную в 5.2, 5.3
симметрию нормы: оказывается, и природа и экономика способны
делать приемлемые стратегии различных своих подсистем слабо-
различимыми по их внешнему эффекту. Но там речь шла о произ-
водствах. Природоохранные работы, наоборот, резко отличаются
от производств тем, что обычно являются убыточными. Поэтому
спасение природы требует выработки новой стратегии экономи-
ки — такой, когда работа по сохранению и возобновлению природы
будет давать примерно те же прибыли, что и другие виды деятель-
ности, станет равноправным элементом разнообразия отраслей на-
родного хозяйства. В сущности, речь идет о резком усилении обыч-
ной функции государства — отчуждении прибыли производств для
обеспечения общественных нужд, дающих прибыль опосредован-
1 Простейший пример двойственной пары (задач линейного программирования),
легко сводимой к матричной игре, см.: [Льюс, Райфа, 1961, с. 520].
9 Ю. В. Чайковский
241
ную и не всегда скорую. Возникают непривычные трудности.
Рассмотрим пример.
Вырубка тропических лесов (около 1,5% в год) уже через 20 лет
должна привести к заметному ухудшению качества атмосферы пла-
неты. Экспорт леса — необходимая статья дохода для тропических
стран, и если развитые страны даже сумеют отказаться от тропи-
ческой древисины (для этого им надо научиться жить не только
без тары разового пользования, но и без большой части химии,
газет, плакатов, громоздких документов), то до решения проблемы
будет далеко. Во-первых, возникнет глобальная безработица:
в джунглях остановится добыча леса, в северных странах — его
переработка и потребление. Во-вторых, 2/3 тропического лесопова-
ла будет продолжаться, поскольку традиция местного населе-
ния — дровяная энергетика и подсечно-огневое земледелие [Био-
логия ..., 1983, с. 343 — 345]. Очевидно, что и обычные призывы к
развитым странам финансировать создание в джунглях огромных
заповедников бессмысленны, ибо ничего реально этим не решится,
а лишь усилится давление экономики на неохраняемые земли.
Чтобы обеспечить рекультивацию джунглей, надо увеличить тот
поток финансов, который течет из северных стран в тропические,
при резком сокращении (вот парадокс) встречного потока леса —
это должна быть плата за воздух и почву как общие всем природ-
ные ресурсы (об этом экономисты уже говорят [Олдак, 1983,
с. 26]). В этом и состоит задача нынешних наук: найти межгосу-
дарственные и внутригосударственные экономические рычаги, со-
циальные механизмы и юридические формы сознательного пере-
ключения людской активности с уничтожения лесов на их восста-
новление. Подробнее см.: [Реймерс, 1990].
Выход состоит в изменении принципа экономической деятель-
ности: «Политическая экономия со времен Адама Смита трактовала
общественное производство как преобразовательную деятельность,
направленную на превращение продуктов природы в хозяйствен-
ные блага... Сегодня недостаточность данной характеристики оче-
видна... Отсюда изменение общей постановки экономических задач:
от управления народнохозяйственным комплексом мы переходим к
управлению биосоциальными системами» [там же, с. 125—126].
Пришло время отказаться от попыток все мерить в рублях, т. е. от
попыток свести все ресурсы природы, общества и хозяйства к
одному — затратам труда. Труд — ресурс возобновимый, и только
возобновимое мы вправе им мерить. В частности, доход от древе-
сины можно всерьез измерить в рублях только тогда, когда потреб-
ление леса ограничено его воспроизводством. Если лес надо вос-
создавать, то соответствующий труд должен быть включен в стои-
мость лесопродуктов, чего лесозаготовители и потребители леса
не хотят. Надо указать способы радикального улучшения дел,
необходим «экологический диктат» [Воронцов, 1989], и тут мы
подходим к диатропике.
Если образно выражаться приведенными выше терминами, то
должна быть срочно сформулирована двойственная пара задач
242
оптимизации, где одну сторону (задачу) будут представлять нату-
ралисты (биологи, медики, географы, геологи и т. д.), а другую —
все «эксплуатационники» (экономисты, демографы, технологи и
т. д.). Как известно, доказать существование решения удается для
гораздо более широкого класса задач нежели те, которые удается
конкретно решить. Пусть задача и не может быть фактически
решена — цель этой работы другая: выяснить те значения
параметров экономического развития, какие окажутся необходи-
мыми для принципиального существования решения задачи, т. е.
существования устойчивой биосферы. Можно не сомневаться, что
для наличия решения придется уменьшить давление на природу,
т. е. сократить промышленное производство, ограничив его не-
обходимым для жизни людей. Откуда же тогда брать огромные
средства на рекультивацию земель и все прочее? Ответ неожиданно
прост: они есть в достатке, если не мерить их рублями, а оглядеть
их разнообразие.
Если до сих пор экономика носила механический (прямое
воздействие на природу), статистический (план, рынок) и систем-
ный (оптимизация) характер, то теперь встает диатропическая
задача — сопрячь разнообразие хозяйств, людей и природных
условий.
Разнообразие хозяйств отнюдь не сводится к сочетанию плана
и рынка, ибо и тот и другой способны губить природу. Экологиче-
ский диктат означает отказ мерить все в рублях, понимаемых как
прежние и нынешние затраты труда. Труд — товар, а многое в
хозяйстве товаром не является (например, чистый воздух или
чистая океанская вода), однако становится дефицитом. Пора пе-
рестать рассматривать слова «экономичный» и «дешевый» (или
«дающий прибыль») как синонимы. .Экономично в действитель-
ности то, что годно для экономики, для народного хозяйства;
в частности — все то, что сохраняет приемлемый быт людей и
природу. И если для такого сохранения надо непрерывно затра-
чивать труд, то это вовсе не надо называть убыточностью. Стремле-
ние минимизировать затраты труда унаследовано нами от XVII в.,
когда рождалась политэкономия. Тогда труд был главным дефици-
том (процветала работорговля) и был для очень многих каторж-
ным, зато ресурсы природы казались неисчерпаемыми. С тех пор
многое изменилось, и минимизировать теперь надо ущерб, наноси-
мый природе (в том числе и людям); но труд — не ущерб, а не-
отъемлемое свойство человека, без которого он деградирует. Одна-
ко, как сбалансировать хозяйство, экономичное в указанном смысле?
Ясно, что рынок тут не спасет. Что и говорить, рынок полезен,
и надо им всюду, где можно, пользоваться, но давайте не будем
пытаться торговать всем; будем торговать только товарами и при-
том бездефицитными, помня, что рынок возникает только тогда,
когда предложение превышает спрос. Если товар дефицитен, но
необходим, то его, увы, надо распределять, пока не исчезнет его
дефицитность, а дефицитные нетоварные ресурсы придется распре-
делять (и охранять от использования) всегда. Запад тоже распре-
9;
243
деляет дефицит отнюдь не рыночными средствами. Там хорошо
знают, что рынок лишь регулирует экономику, а не создает ее.
Но хотя у нас царит наивная вера в рынок-творец (см.: [Бога-
чев, 1990]), на самом деле мы все привыкли к распределению,
и нам принять экологический диктат будет легче. На Западе яд-
ро общества образуют прежде всего люди экономического типа
мотивации (см. 2.5), а у нас — политического и социального,
поэтому рынок у нас возможен пока лишь на периферии экономи-
ки. И частную собственность наше общество готово признать лишь
в смысле мелких предприятий [Заславская, 1990]. Однако и это
может тоже стать нашим преимуществом, поскольку экологический
диктат включает в себя отказ от индустриальных гигантов в пользу
меньших предприятий, легче стыкуемых друг с другом.
Отстав от западного мира, хорошо освоившего системную позна-
вательную модель (таково западное «постиндустриальное обще-
ство»), наше общество пока что стремится перейти от одной ста-
тистической идеи (спланированные балансы) к другой (рыночные
балансы). Этот наш недостаток может стать нашим преимущест-
вом, поскольку статистическая модель кое в чем близка диатропи-
чес.коп (см. 1.2). Другими словами, то, с чем Запад пока не справ-
ляется, нам может даться легче, если мы будем ориентироваться не
на пройденный этап истории, а на нечто новое. Это новое видится
мне в согласовании разнообразий.
Всякую потребность общество будет впредь вынуждено удовлет-
ворять не саму по себе, а вместе со всеми другими, поскольку все
виды деятельности понемногу приобретут характер перераспреде-
ления * дефицитных ресурсов. Для этого необходим, по-моему,
метод замыкаемых затрат, о котором я коротко писал [Чайков-
ский. 1988в, с. 83] в связи с таким примером..
На экологические нужды и на молодежный досуг тратятся ог-
ромные, но совершенно недостаточные средства, и опыт показал,
что бывает полезно замкнуть эти затраты друг на друга: молодежь
легко откликается на призывы к очистке загаженной природы, при-
нимая это как полезное (не показное) дело и как интересный
досуг; при этом финансировать надо, грубо говоря, не сумму,
а разность тех затрат, какие требуются при раздельном проведе-
нии досуга и очистки. То же справедливо буквально для всех
сторон жизни. Так, почти любой отход для чего-то полезен, пока не
перемешан с другими, не утерян. Пусть его утилизация и убыточна,
но при утилизации субсидировать надо только разность (убыток),
а не весь процесс очистки или уничтожения. Это приводит к важ-
ной аналогии.
В биосфере всякое вещество включено в замкнутые циклы мета-
болизма, так что, являясь «выходом» какой-то органической реак-
* Его не надо путать с уравниловкой. В кризисных ситуациях необходимо прибегать
и к уравнительному распределению, но распределять надо минимум жизненно не-
обходимых наименований (например, хлеб, молоко) и ниже среднедушевой доли
на каждого. Остальное же должно быть предметом либо торговли, либо распре-
деления, но должны соблюдаться естественные законы распределения величин
в больших системах, обычно носящие гиперболический характер (см. гл. 3).
244
ции, оно обязательно является и «входом» какой-то другой. Очень
существенно, что для многих реакций количественного сопряжения
между «входом» и «выходом» нет, так что ряд веществ накапли-
вается в органических осадках.
Биосфера выступает как огромный организм, в течение 4 млрд
лет медленно съедающий планету путем потребления одних иско-
паемых и создания других. Вопреки учебникам, для поддержания
жизни биосферы необходим не только солнечный свет, но и горооб-
разование. Когда оно через 2 — 3 млрд лет прекратится, то ряд хими-
ческих элементов, и прежде всего фосфор, скопятся понемногу в
океанических осадках, которые биологическими средствами почти
не возвращаются в биосферный круговорот. Однако пока горооб-
разование идет, продолжается и жизнь, поскольку ритмы ее под-
строены к ритмам геологии. Это важно иметь в виду: разнообра-
зие органических реакций согласовано не само с собою (как часто
думают), а с разнообразием геологических процессов — качествен-
но и количественно.
Экономическая сфера расходует ископаемые приблизительно в
миллион раз быстрее, чем экологическая, и поэтому биосфера вы-
ступает для экономики примерно тем же, чем для биосферы —
геосфера. Налицо пара рядов, наводящая на ту мысль, что замыка-
ние затрат должно означать вовсе не создание замкнутых эконо-
мических циклов (это почти нигде невозможно), а подстройку
экономических ритмов к биосферным. Это значит, что антропо-
генные'выбросы в биосферу должны быть ограничены не медицин-
скими и хозяйственными соображениями (чем обычно ограничи-
ваются сейчас), а прежде всего — экологической емкостью среды
[Воронцов, 1989], т. е. способностью экосистем их ассимилировать.
Такая стратегия и означает экологический диктат; варианты эко-
номического развития следует выбирать только из числа экологи-
чески приемлемых, что заставит отказаться: Западу — от потреби-
тельских излишеств, нам — от стремления к ним. Ясно, что наш
путь будет в этом смысле психологически легче.
Западное общество раньше нас осознало, что подоплекой поли-
тических и межнациональных конфликтов обычно бывает пере-
населенность и распад экосистем [Наше общее будущее, 1989;
гл. 4, 11]. Эколог Стэнли Роу пишет: «Как можно добиться гармо-
нии между миром природы и сообществом народов, у каждого
из которых своя национальная экономика?.. Человечество, люди
существуют и живут в мире природы... Поэтому, когда мы оптими-
стически заявляем, что экономическое развитие и сохранение окру-
жающей среды могут сочетаться друг с другом, к этому необходимо
немедленно добавить: только в случае, если сохранению биосферы
будет придаваться первостепенное значение. Экономическое разви-
тие должно занимать второе место и определяться строгими эко-
логическими нормами. Далеко не все согласны с этими основопо-
лагающими идеями» [там же, с. 263]. В нашей стране положение
еще хуже, поскольку среди идеологов перестройки популярна
прямо противоположная идея — экономический диктат, диктат
245
рынка. Экологическое движение иногда называют даже «зеленым
чудовищем», чем-то вроде нового фашизма, предлагая считать
нынешний образ жизни выше идеи выживания человечества [Соко-
лов, 1989]. При этом верно отмечается некоторая параллель между
обещаниями ранних фашистов и нынешних «зеленых», но ведь
параллель найдется между любыми системами (тезис Урманцева),
и искать надо не ее, а тенденцию. Увы, тенденция к зеленому
тоталитаризму — реальность (ее не изменят никакие статьи, и
надо не бороться с неизбежным, а направлять опасную тенденцию
в мирное русло), но она во многом противоположна той, что была
в годы рождения фашизма и сталинизма. Сейчас, в отличие от
1920-х годов, растет роль малых предприятий и децентрализации,
а это создает иной климат. Философ Джанна Томпсон выражает
надежду, что экологическое сознание придет на смену потребитель-
скому * мирно, как новая мода [Thompson, 1983, с. 96]. Чтобы это
могло случиться в нашей стране, где демократии не было никогда
и нет, зато самоуправление было (община, земство) и вновь наме-
чается, необходимо, по-моему, вовлечение всех активных людей в
замыкание затрат путем планирования снизу, т. е. следующее.
Если при обычном (сверху) планировании инициатива подав-
ляется, а сопряжение объекта со средой путем «согласований»
и «платы за ресурсы» не спасает природу, а подчас быстрее губит,
то планирование снизу означает подстройку объекта, заявленного
теми, кому он нужен, к локальным процессам. Каждому ресурсу
надо указать источник, продукту (отходу) — потребителя. Участок
отводится только волею местного Совета, т. е. объекты будут, в
основном, строиться на месте ликвидируемых. Если сопряжение
не удается, Совет либо отвергает проект, либо содействует его
сопряжению путем контакта с другими Советами. Низшие Сове-
ты — объекты соопряжения для высшего. Далее, налог с предприя-
тия можно взять деньгами, продукцией (ферма снабжает школу
ит. п.) или трудом — по договору. Трудом же могут платить налог
и другие (полк, райком, вуз, семья), и реален тот замысел, который
люди поддержат трудом. Тем самым, замыкание затрат пойдет не
столько в рублях, сколько в физических потоках; товаром станет
лишь то, что надо продать или купить «на стороне», на рынке.
Это снизит потери и, смею надеяться, улучшит моральный климат
за счет снятия отчуждения труда от продукта. Принимая к рас-
смотрению и замыкая друг на друга любые проекты (производст-
венные, природоспасающие, культурные), Совет может соединить
достоинства хозяйственного и благотворительного фондов с права-
ми местной власти, реализовать активность людей разных соци-
альных интересов и личных мотиваций. Именно разнообразие
активностей делает государдство самоорганизующейся системой,
а пренебрежение этим фактом уводит к идее рынка как чудесного
спасителя.
* Его часто считают общечеловеческой ценностью, хотя это скорее новый за-
падный идеал, сменивший марксизм, о чем подробнее см. [Солженицын, 1990].
246
Намеченная здесь модель близка к замыслам Римского клуба
[Одум, 1986, т. 2, с. 237], но контролирует более физические
потоки, нежели капитал. Ее можно назвать утопией, но вряд ли
это упрек: при смене тенденций реализуются как раз утопии
«Увы, либеральная доктрина всегда была слаба тем, что она не
могла создать идеологии и не имела утопий» [Чаянов, 1989, с. 70].
Более детальный анализ выходит за рамки нашей темы, и оста-
ется заметить только, что симпатии нашей научной общественности
быстро переходят от рыночного варианта экономики к различным
вариантам сопряжения потоков. Достаточно прочитать програм-
мный сборник «Постижение» (1989 г.), чтобы заметить этот сдвиг.
Главный мотив переориентации — осознание невозможности даль-
нейшей экономической экспансии, истощающей ресурсы природы
и общества, в том числе — нравственные ресурсы. Рыночная мо-
дель ориентирована на неограниченный рост производства, а можно
ли сделать экономику замыкаемых затрат стационарной?
Деятельность людей так или иначе включается в глобальные
циклы, но надо, чтобы это делалось сознательно, без отравления
природы. Можно ли замкнуть все круговороты, оставив прямоли-
нейным лишь один поток — диссипацию солнечной энергии? Разу-
меется, этот поток надо использовать гораздо интенсивнее, чем
сейчас — например, путем переработки света в электричество
[Goldsmith, 1981]. Вопрос о максимальном приближении спиралей
к циклам аналогичен вопросу о максимальном КПД тепловой ма-
шины, решенному Карно в 1824 г. (см. 9.1). Возможно, что удастся
вычислить ту степень замкнутости системы, ту границу, далее ко-
торой образование подсистем невозможно. В таком случае роль
новой термодинамики будет методологически эквивалентна преж-
ней роли термостатики.
12.2. Эволюционный прогноз. Заключение
Об однозначном прогнозе эволюции биосферы никто не говорит,
но, по-моему, нет смысла говорить и о вероятностном прогнозе:
любая альтернатива, коль скоро она осознана как в принципе
возможная, заслуживает полного внимания. Варианты настолько
многочисленны, что детальный анализ невозможен, но диатропика
обращает внимание на то, что разнообразие всегда создается
комбинаторикой обозримого числа элементов, обнаруживающих к
тому же рефренную структуру. Потепление или похолодание, осу-
шение или увлажнение, смещение небольшого числа климатиче-
ских зон к северу или югу — подобных альтернатив немного.
Пусть мы не можем вычислить, какая из них реализуется, но в
этом и нет надобности, поскольку любое такое вычисление может
быть неправильным из-за неучета параметра, казавшегося несу-
щественным, но оказавшимся главным. Реальный прогноз видится
в другом — в указании наиболее гибельных альтернатив.
Так, мы твердо знаем, что чрезмерное сжигание органических
топлив влечет рост концентрации СОг, а он в свою очередь создает
247
парниковый эффект. Последствия этого неисчислимы, и таяние
Антарктиды — далеко не самое вероятное из них, но это — одна
из самых гибельных альтернатив. Разрушение 1/3 антарктического
щита (именно такую его долю оценивают как неустойчивую) не
только уничтожит Голландию (что всем очевидно), но соединит
Черное море с Каспийским и сделает Тобольск морским портом,
а большинство речных долин — болотами. Убыль плодородных
территорий будет такова, что возможна полная деградация челове-
чества. Согласны ли мы на такой вариант (пусть и маловероятный)
или лучше все-таки резко сократить сжигание топлив? Таков один
из вопросов, какие ставит прогностика.
Сейчас мы вновь, как до Ламарка, начинаем рассматривать
эволюцию живого в качестве компонента эволюции Земли и. отча-
сти. Космоса. Это меняет всю логику эволюционных построений.
Когда в центре внимания было взаимодействие данного организма
с заданной средой, главным событием естественно оказывалось
преобразование признака, откуда, путем экстраполяции, выводи-
лось видообразование, и вся дальнейшая эволюция трактовалась
как таксонообразование (см. 7.3). Красилов [1987] назвал этот тип
рассуждений восходящей причинностью. Наоборот, когда в центре
внимания оказывается экосистема, определяющая как целое основ-
ные свойства входящих в нее видов и безразличная к их конкрет-
ным различиям, то главным эволюционным событием оказывается
преобразование не признака и даже не таксона, а разнообразия.
Например, периоды высокой геологической и космической актив-
ности, наиболее существенные для смены таксонов, характеризуют-
ся сокращением видового разнообразия [Красилов, 1987; Неручев,
1988]. Это — логика нисходящей причинности. Она близка к «ло-
гике ценоза» (5.2.).
Прежняя логика восходящей причинности оказалась неспособ-
ной к эволюционному прогнозу — «не только для отдаленного, но
и для ближайшего будущего» [Татаринов, 1987, с. 6]. Немногочис-
ленные попытки прогноза являют собой примитивные экстраполя-
ции и плохо продуманные комбинации известных форм организ-
мов [там же, с. 48, 188]. Элементарный диатропический анализ
обычно обнаруживает неправдоподобность таких прогнозов. Так,
гипотетические звери будущего, похожие на огромных бескрылых
птиц или ходящие на передних лапах [Dixon, 1981], ни в какие
ряды не становятся: у млекопитающих нет ни редукции передних
конечностей, ни переноса на них тяжести тела.
Тот же диатропический анализ сам приводит иногда к успешно-
му мероно-таксономическому прогнозу: вспомним андродиэцию
Дарвина (7.3), злаки Вавилова (1.5) или бактерии Заварзина
(6.5). Логику таких прогнозов можно бы назвать (продолжая
мысль Красилова) горизонтальной. Ей легко придать и эволю-
ционную внешность, утверждая, что пустующая сейчас клетка
какого-то ряда будет когда-нибудь заполнена — так, можно «про-
гнозировать» что-то вроде млекопитающего угря или утверждать
невозможность «ангела» (см. 2.4). Однако это не так уж интересно.
248
Гораздо важнее оказывается прогностическая роль нисходящей
причинности.
Разумеется, при этом прогнозируется не внешность отдельных
будущих видов, а общее направление преобразования того или дру-
гого разнообразия. Прогноз в принципе не может быть однознач-
ным — в той же мере, в какой неоднозначна филогения (см. 7.3),
что, однако, не умаляет его ценности; наоборот, полезен именно тот
прогноз, который разворачивает перед нами весь спектр возмож-
ностей. Для примера коснемся эволюционного аспекта той же проб-
лемы СОг в атмосфере.
Накопление СОг может изменить всю обстановку на плланете и
в частности может как увеличить продукцию кислорода (за счет
усиления фотосинтеза и потепления), так и уменьшить ее (за счет
сокращения лесов и расширения пустынь). Накопление кислорода
в атмосфере может как укрепить позиции животного царства (за
счет добавочной растительной пищи), так и ослабить их (если рас-
тения станут быстрее гнить, т. е. отдавать биомассу редуцентам,
минуя консументов). Если животная жизнь усилится, то поведет
ли это к прогрессу (усложнению организмов), просто к накопле-
нию биомассы или к регрессу (вытеснению высших низшими)?
С одной стороны, усиление активности вроде бы должно вести к
прогрессу, но с другой — Аршавский [1982, с. 197] показал, что
для лучшего развития желателен умеренный недостаток кислоро-
да, и склонен поэтому связывать происхождение человека с высо-
когорьем. Спектр вариантов можно продолжить, но и сказанного
достаточно, чтобы увидеть главное: они образуют не древо, а сеть,
поскольку к одному выводу можно прийти разными путями. Это
радикально упрощает задачу прогнозирования: вместо шп вершин
древа (где ш — число выборов, и — число вариантов в каждом из
них) мы имеет таблицу из шп клеток, некоторые из которых
пусты.
Одно ясно — на любое изменение атмосферы организмы ответят
быстрой и существенной эволюцией, в чем нас убеждают «экспе-
риментальная эволюция» (4.4), адаптация к ядам и т. п. Вернее
всего, параллельно реализуется большинство альтернатив: в одних
эонах растения повысят фотосинтез, в других понизят; в одних
местах будут наступать болота, в других пустыни; одни животные
будут прогрессировать, другие просто нарастят биомассу и т. д.
В такой ситуации самой подходящей методологией должен явиться
принцип множественных рабочих гипотез [Chamberlin, 1931], при-
зывающий не выбирать «лучшую» возможность, а оперировать со
всем их разнообразием как «единой семьей».
Если мы хотим прогнозировать более конкретно, то следует
привлечь морфологические тенденции, например, террипетную
тенденцию по Бронну (2.5). Для имеющих голову животных из-
вестна цефализация — тенденция увеличения головы от предков к
потомкам. Позвоночные, на которых геолог Джеймс Дана в 1852 г.
описал эту тенденцию, демонстрируют тот факт, что сочетание вы-
сокой степени террипетности и цефализации дает наиболее сложно
249
устроенные организмы. У насекомых цефализация выражена сла-
бее, но зато гораздо сильнее проявляется комбинативность (свобода
в комбинировании свойств, наиболее развитая у низших форм —
см. главу 6), тенденция, наиболее важная в приспособительной
эволюции. Это, а также прямые опыты Шапошникова, позволяют
предположить, что именно насекомые наиболее радикально отве-
тят морфологически на рост СОг. Группы, обладающие лишь ком-
бинативностью, скорее ответят на рост СОг изменением физиоло-
гии, но не формы.
Конкретнее можно говорить о заполнении определенных эко-
логических ниш, и здесь естественная эволюция смыкается с искус-
ственной. Можно ли, например, заполнить нишу, 200 лет пустую-
щую после исчезновения стеллеровой коровы? Разведение мясного
скота на подводных пастбищах морской капусты весьма жела-
тельно, но близкие «родственники» стеллеровой коровы — дюгонь
и ламантин — для целей селекции явно не годятся: сами находятся
на грани вымирания. К тому же весь отряд (сирены) никогда не
был богат видами, не проявлял ни террипетности, ни цефализации
(в отличие от богатого видами и цефализованного отряда китов),
а потому вряд ли здесь найдется подходящий спектр изменчивости.
Как же занять пустующие подводные поля? Очерченный в ча-
сти II экосистемный взгляд на эволюцию позволяет наметить
спектр подходов, быть может годных для того, чтобы совместными
усилиями решить эту или подобную задачу.
Во-первых, нам нужен вовсе не определенный вид, а приемле-
мая экосистема (на морской капусте удобнее выращивать не огром-
ную стеллерову корову, но совокупность некрупных, а потому эко-
логически более устойчивых, видов). Во-вторых, исходить надо не
из «родственников», так как таксономическая близость не говорит
об экологической (достаточно вспомнить, что ближайшим живым
«родственником» дюгоня и ламантина является слон), а из эколо-
гически удобных видов. В-третьих, селекцию нельзя вести только
на очевидно полезные признаки, надо управлять всей траекторией
преобразования желаемого вида, т. е., в частности,— становлением
зародыша. Для этого, в-четвертых, надо уметь разбалансировать
генетическую систему вида, чтобы на всех уровнях могли созда-
ваться новые объекты будущего отбора (4.4). В-пятых, чтобы обес-
печить приемлемую скорость преобразования, надо уяснить ход
индивидуального времени преобразуемого вида — этим отчасти за-
нимается так называемая хронобиология. В-шестых, для опытов
надо выбирать виды, проявляющие нужные тенденции, но не про-
являющие тенденции к вымиранию. Уже из этих наметок видно,
что попытки формировать экосистемы надо начинать не с крупных
зверей, а с мелких насекомых. Напомним, что Шапошников (как до
него генетики) строил эволюционный эксперимент именно так.
Далее говорить пока не о чем, если не вдаваться в чистые фан-
тазии.
Однако сами тенденции, по которым следует выбирать виды для
опытов, вполне можно уяснить уже сейчас. Прежде всего, речь
250
должна пойти о тенденции к вымиранию (по-видимому, она харак-
терна, например, для всего отряда сирен). Данилевский [1885,
ч. 2, с. 390 — 393] полагал, что стеллерова корова сама была на грани
вымирания, когда человек «доконал» ее, что немногочисленные
моряки просто не сумели бы столь быстро истребить столь обшир-
ное стадо, и указал ряд признаков его неблагополучия уже во вре-
мя, когда его обнаружили. Объективная тенденция к вымиранию
(«дряхлость таксона» по неоламаркистам, типолиз по Шиндеволь-
фу и т. д.) — феномен хорошо известный, но, к сожалению, редко
принимаемый во внимание при практической работе ввиду господ-
ства уверенности, что каждый вид произошел исключительно путем
приспособления к своей конкретной среде обитания. В то же время
неадаптивность иногда бывает прямо-таки очевидна, о чем шла
речь в 4.6. Неадаптивные легко вымирают — таковы многие неле-
тающие и плохо летающие птицы. Неадаптивность наиболее ярко
выступает как тенденция к вымиранию тогда, когда прямо снижает
способность к размножению. Таким путем вымирает стерх (белый
журавль), неуклонно снижающий (несмотря на заботу людей)
зону гнездования и число птенцов в выводке. Аналогичным обра-
зом уже вымер как вид зубр, причем это событие еще в 1865 г.
прогнозировано в книжке Усова «Зубр». Прогноз был основан
на проницательном анализе тенденции зубра к вымиранию, кото-
рую, по мнению Усова, могло предотвратить только скрещивание с
бизоном, подобной тенденции не проявляющим. Прогноз оправдал-
ся: немногие «чистокровные» зубры еле живут иждивением чело-
века, тогда как гибриды процветают.
Странно слышать призывы «полностью исключить спаривание»
даже зубров разных подвидов, «удалить все другие виды копыт-
ных» (не говоря уж о волках) с территории, предназначенной
«чистокровным» и, более того, к «полной элиминации гибридного
поголовья» [Биологические аспекты..., 1981, с. 12 — 13]. 80 лет ис-
кусственного размножения и подкормки зубров названо успешным
«возвращением к дикому образу жизни» [там же, с. 14], образцом
для восстановления других видов. Этим отрицается не только борь-
ба за существование (даже в той скромной стабилизирующей роли,
какую признавали [Schwartz, 1974] еще до Дарвина), но и сама
возможность эволюции. Досадно, что все это подается как торжест-
во дарвинизма, но еще хуже, что игнорирован оправдавшийся
эволюционный прогноз. Скромные природоохранные ресурсы рас-
ходуются при этом фактически на прежнюю «борьбу с природой»,
на борьбу с эволюцией.
Необходимо ясно различать сохранение разнообразия и сохра-
нение конкретного вида. Замена же одних видов другими — про-
цесс не только неизбежный, но подчас и желательный. Приоритет
инвалидным (нуждающимся в поддержке) стадам может быть
необходим экономически (скотоводство), но в остальных случаях
неприемлем, ибо спасению дикой природы лишь препятствует.
Развыв между эволюционными девизами и антиэволюционной
практикой говорит о методологическом тупике. Особенно тупик
251
виден при анализе попыток дать новое (экологическое) толкование
эволюции, понимаемой в старой (конкурентно-адаптационной)
парадигме. Так, зоолог Гирэт Вермей привлекает для этого толко-
вания старое соображение Дарвина, согласно которому прогресс
живого вызван усложнением среды, окружающей каждый данный
вид, и называет такое понимание прогресса «гипотезой эскалации»
[Vermeij, 1987, с. 4]. Изложив в этом ключе историю фанерозоя
(последние 670 млн лет), он приходит к неожиданной трудности:
«Тот факт, что эволюционные изменения идут гораздо легче в од-
них направлениях, чем в других, также означает, что возможны
многие тенденции, не согласующиеся с эскалацией» [там же,
с. 426], и что предсказательная сила идеи отбора в отношении
многих малораспространенных форм организмов мала. Не имея
других методологических средств, Вермей ограничился, как обыч-
но, надеждой, что будущие исследования разрешат все трудности
с позиции отбора. Эта уловка известна как asylum ignorantiae
(убежище невежества) [Любищев, 1982; Меуеп. 1987, с. 355] и
является одним из самых ходовых приемов эволюционизма. Сама
по себе надежда на будущие результаты вполне естественна, но
незаконно другое — использовать ее как аргумент, отводящий кон-
кретные возражения.
Методологический тупик возник, по-моему, оттого, что бурное
накопление фактов необычайно обогнало их теоретическое (не
говоря уж о философском) осмысление. В первой половине XX в.
в естествознании преобладала гипотетико-дедуктивная методоло-
гия, давшая тогда блестящие результаты в физике, но еще до этого
заявившая о себе в дарвинизме [Ghiselin, 1969]. Для нее характер-
ны попытки строить конкретные теории внеисторическим образом,
путем простого выведения суждений из гипотез. Этот подход ока-
зался ограниченным даже в физике [Чудинов, 1977], а в биологии
вряд ли вообще имеет смысл. Истины описательных наук склады-
ваются в ходе исторического осмысления, здесь гораздо богаче раз-
нообразие объектов. Поэтому, если физик может хотя бы на время
забывать, что все его теории суть модели, соответствующие нынеш-
нему состоянию развития знания, то для биолога «позиция наивного
реализма, полного отождествления модели... и самого объекта»
[Карпинская. 1984, с. 159] просто непродуктивна.
Последние 20 лет кристаллизуется собственный метод описа-
тельных наук (намеченный еще Страховым) — двойственно-ите-
ративный анализ, основанный на попеременном обращении к взаи-
модополнительным (по Бору) точкам зрения на данный объект.
В этой связи мы говорим о мероно-таксономическом и номотетико-
идиографическом методах, о двойственной оптимизации, о восхо-
дящей и нисходящей причинностях и т. п. В более сложных ситуа-
циях взаимодополнительностью обладает не пара, а тройка поня-
тий — таковы разнообразие, приспособление и прогресс (10.3),
а само разнообразие — результат дифференциации, "интеграции и
приспособления (1.2). На познавательную роль триад постоянно
указывают системологи Р. Г. Баранцев и П. А. Харченко. Возможны
252
познавательные тетрады, пентады и т. д., и это позволяет вновь с
почтением взглянуть на учение самого древнего из названных в
книге мыслителей -- Пифагора.
Развитие науки возвращает нас вновь и вновь к небольшому
числу сквозных тем, проходящих через всю историю науки [Хол-
тон, 1981]. В те периоды, когда данная тема оказывается связан-
ной с актуальной исследовательской программой (по Лакатосу),
она привлекает всеобщее внимание и облачается в покровы из
новых фактов, но ее концептуальный базис никогда не бывает
абсолютно нов, а потому анализ новой теории всегда требует зна-
ния истории.
Как мне представляется, данные тезисы в основном обоснованы
материалом настоящей книги. Разнообразие и эволюция — две
давние темы, на скрещении которых сейчас формируется диатро-
пика. Как сложится ее судьба, сейчас говорить рано, но если диат-
ропика позволяет (как я пытался показать) ставить новые задачи,
имеющие практический смысл, то, следовательно, она имеет свою
исследовательскую программу, а с тем и право на внимание.
Главного внимания, по-моему, заслуживают не столько резуль-
таты (хотя, например, глобальный флорогенез важен теоретиче-
ски и практически — см. 7.3) и даже программы, сколько смена
монистического подхода к природе и обществу на плюралистиче-
ский. Как писал в 1920 г. Вернадский [1988, с. 49], его учение о
живом веществе — не отрицание учения Дарвина, но «представ-
ляет другую сторону — другой аспект — эволюционного учения».
Сейчас существуют и многоаспектные подходы. Как считал Мейен,
априорная фиксация какого бы то ни было числа уровней или
аспектов умаляет возможности познания, а если так, то по исчерпа-
нии триады Уивера (см. 10.1) следует ожидать нового этапа раз-
вития науки, основанного на анализе разнообразия как такового.
Плюрализм моден и сейчас, но у нас он узко утилитарен: дадим
конкурировать идеям (товарам) и выберем лучшее. Эта давняя
идея отбора, идея рынка легко оборачивается монизмом. Плюра-
лизм рынка обманчив: товары здесь — только средства, и если
выгода требует, на рынке царит дефицит [Богачев, 1990]. От
огораживаний Англии до нынешней трагедии джунглей всегда
виден жестокий монизм самовластного рынка. Изобилие его легко
уживается с голодом. Достоинства Запада (защита природы, куль-
туры, слабых) — результат недавнего преодоления власти рынка
у себя за счет чисто рыночной эксплуатации чужих ресурсов —
наших и Юга. Этот недальновидный плюрализм обречен. Невоз-
можность дальнейшего роста экономики и численности населения
обращают взоры многих к равновесному природопользованию,
к отказу от главенства мотива выгоды, т. е. к более реальному плю-
рализму. Рынку надо оставить его полезную функцию регулятора.
Как дарвинская идея победителя в мире-рынке привела, помимо
воли ее авторов, к худшим идеологиям XX в., так, смею надеяться,
идея единства мира-сада (Лейбниц) поможет хоть немного смяг-
чить идеологии XXI в., с такой тревогой ожидаемого.
ЛИТЕРАТУРА
Агаев М. Г. Экспериментальная эволюция. Л., 1978.
Акчурин И. А. Новые теоретико-категорные и типологические методы в осно-
ваниях физики // Методы научного познания и физика. М., 1985.
Александров В. Я. Проблема поведения на клеточном уровне: (Цитоэтология') //
Успехи соврем, биологии. 1970. Т. 69, № 2.
Александров Д. А. Об организации живого вещества // Журн. общ. биологии. 1986.
№ 2.
Алексеев И. С. Концепция дополнительности. М., 1978.
Анохин И. А. Суточный ритм муравьев, инвазированных метацеркариями // Докл.
АН СССР. 1966. Т. 166, № 3.
Арапов М. В. Теория систем и изучение естественного языка // Системные иссле-
дования: Ежегодник, 1980. М., 1981.
Аристотель. О частях животных. М., 1937.
Аршавский И. А. Физиологические механизмы и закономерности индивидуаль-
ного развития. М., 1982.
Аршавский И. А. К обоснованию принципов и задач эволюционной физиологии //
Журн. эволюц. биохимии и физиологии. 1985. № 2.
Аршавский И. А. Некоторые методологические и теоретические аспекты анализа
индивидуального развития организмов // Вопр. философии. 1986. № 11.
Ауэрбах Ш. Проблемы мутагенеза. М., 1978.
Ахутин А. В. Понятие «природа» в античности и в Новое время. М., 1988.
Бабков В. В. Системный стиль в изучении естественного отбора // Системные ис-
следования: Ежегодник, 1980. М., 1981.
Балашов Ю. В. «Антропные аргументы» в современной космологии // Вопр. фило-
софии. 1988. № 7.
Беклемишев В. Я. Морфологические проблемы животных структур. Пермь, 1925.
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных. М., 1964.
Т. 1: Проморфология.
Белоусов Л. В. Биологический морфогенез. М., 1987.
Берг Л. С. Труды по теории эволюции. Л., 1977.
Бергсон А. Творческая эволюция. М., 1909.
Биологические аспекты охраны редких животных. М., 1981.
Биология охраны природы. М., 1983.
Битов А. Г. Семь путешествий. М., 1976.
Бляхер Л. Я. Проблемы морфологии животных. М., 1976.
Бобрецкий Н. В. Основания зоологии. Киев, 1891. Вып. 3.
Бобров Е. Г. Описательный метод Линнея // Карл Линней. М., 1958.
Богачев В. Я. Монополия // Знание — сила. 1990. № 3.
Богданов (Малиновский) А. А. Всеобщая организационная наука: (Тектология).
СПб., 1913. Ч. 1.
Богородицкий В. А. Очерки по языковедению и русскому языку. Казань, 1910.
Борхсениус С. Я., Чернова, О. А. Микоплазмы // Цитология. 1987а. № 4.
Борхсениус С. Я., Чернова. О. А. Геном микоплазм // Там же. 19876. № 6.
Браун А. Д., Моженок Т. Л. Неспецифический адаптационный синдром клеточ-
ной системы. Л., 1987.
Брем А. Д., Россмесслер Е. Лесные животные. СПб., 1867.
Брода Э. Эволюция биоэнергетических процессов. М., 1978.
Бруно Дж. Диалоги. М., 1949.
Будыко М. И. Эволюция биосферы. Л., 1984.
Бэр К. М. Неизданные рукописи // Анналы биологии. М., 1959. Т. 1.
254
Вавилов Н. И. Закон гомологических рядов в наследственной изменчивости. М.,
1987.
Валы М. X. Экологические исследования Бэра и концепция борьбы за существова-
ние// Петербургская академия наук и Эстония. Таллин, 1978.
Варден Б. Л. ван дер. Математическая статистика. М., 1960.
Варшавский В. И., Поспелов Д. А. Оркестр играет без дирижера. М., 1984.
Ватти К. В., Тихомирова М. М. Адаптация и мутагенез // Радиационный мутагенез
и его роль в эволюции и селекции. М., 1987.
Вернадский В. И. Живое вещество. М., 1978.
Вернадский В. И. Проблемы биогеохимии. М., 1980.
Вернадский В. И. Из дневников 1919—1920 гг.//Наука и жизнь. 1988. № 3.
Взаимодействие лингвистических ареалов. Л., 1980.
Вигнер Е. Этюды о симметрии. М., 1971.
Виндельбанд В. История новой философии. СПб., 1913. Т. 2.
Возникновение жизни на Земле. М., 1959.
Волк/Д. И. Бибиков. М., 1985.
Воронцов Н. Н. Системы органического мира и положение животных в них //
Зоол. журн. 1987. № И, 12.
Воронцов Н. Н. Экологический диктат в эпоху демократизации //Сов. культура.
1989. 2 сент.
Гайденко П. П. Эволюция понятия науки. М., 1980.
Гаркави Л. X., Квакина Е. Б., Уколова М. А. Адаптационные реакции и резистент-
ность организма. Ростов н/Д, 1979.
Гегель Г. Энциклопедия философских наук. М., 1975. Т. 2.
Гельмгольц Г. Популярные научные статьи. СПб., 1866. Вып. 1.
Гиппократ. Избранные книги. М., 1936.
Гленсдорф П., Пригожин И. Термодинамическая теория структуры, устойчивости
и флуктуаций. М., 1973.
Голсуорси Дж. Сага о Форсайтах. М., 1956. Т. 2.
Грамматика литовского языка/В. Амбразас. Вильнюс, 1985.
Гроссгейм А. А. О графическом изображении системы цветковых // Сов. ботаника.
1945. № 3.
Гумбольдт А. Путешествие в равноденственные области Нового света. М., 1963. Т. 1.
Данилевский Н. Я. Дарвинизм: Крит, исслед. СПб., 1885. Т. 1, ч. 1—2.
Дарвин Ч. Сочинения. М„ Л.: Изд-во АН СССР. Т. 2. 1936; Т. 3. 1939; Т. 7. 1948;
Т. 4. 1951; Т. 5. 1953; Т. 6. 1956; Т. 9. 1959.
Девис П. Случайная Вселенная. М., 1985.
Де Фриз Г. Избранные произведения. М., 1932.
Джеффри Ч. Биологическая номенклатура. М., 1980.
Диалектика развития в природе и научном познании: Сб. обзоров ИНИОН. М., 1978.
Диксон М., Уэбб Э. Ферменты. М., 1982. Т. 3.
Догель В. А. Курс общей паразитологии. Л., 1941.
Догель В. А. Зоология беспозвоночных. М., 1981.
Дриш Г. Витализм: Его история и система. М., 1915.
Дуда В. И. Архебактерии — новое царство живых организмов // Природа. 1984.
№ 2.
Елфимов Г. М. Возникновение нового. М., 1983.
Женермон Ж. Проблема длительных модификаций у простейших // Журн. общ.
биологии. 1970. № 6.
Журавлева И. Т., Мягкова Е. И. Низшие многоклеточные фанерозоя. М., 1987.
Заварзин А. А. Труды по теории параллелизма и эволюционной динамике тканей.
Л., 1986.
Заварзин Г. А. Экстенсивная микробиология // Изв. АН СССР. Сер. биол. 1976. № 1.
Заварзин Г. А. Пространство логических возможностей в многообразии бактерий
и их филогения // Природа. 1979. № 6.
Заварзин Г. А. Бактерии и состав атмосферы. М., Наука, 1984.
Заславская Т. И. Частная собственность в стране перестройки? (Социологическое
обследование) // Курьер латвийской ассоциации инженеров. 1990. № 5.
Заславский Г. М. Стохастичность динамических систем. М., 1984.
Здродовский П. Ф., Гурвич Г. А. Физиологические основы иммунитета и его регуля-
ции. М., 1972.
255
Золотарев В. М. Одномерные устойчивые распределения. М., 1983. ц
Иванов В. В. Историческая грамматика русского языка. М., 1983.
Ивановский А. Б. Палеонтология и теория эволюции. Новосибирск, 1976.
Камшилов М. М. Биотический круговорот. М., 1970.
Камшилов М. М. Эволюция биосферы. М., 1974.
Канаев И. И. Очерки из истории сравнительной анатомии до Дарвина. М.; Л., 1963.
Канаев И. И. Очерки из истории проблемы морфологического типа от Дарвина
до наших дней. Л., 1966.
Кант И. Сочинения: В 6 т. М., 1966. Т. 5.
Карпинская Р. С. Теория и эксперимент в биологии. М., 1984.
Карпов Вл. Пор. Основные черты органического понимания природы. М., 1912.
Катастрофы и история Земли: Новый униформизм. М., 1986.
Кизелъ В. А. Физические причины диссимметрии живых систем. М., 1985.
Кимура М. Молекулярная эволюция: Теория нейтральности. М., 1985.
Козо-Полянский Б. М. А. Жюссье и Линней // Бюл. МОИП. Отд. биол. 1937. № 5.
Козырев И. А. Время как физическое явление // Моделирование и прогнозирование
в биоэкологии. Рига, 1982.
Комаров В. Л. Введение // Дарвин Ч. Происхождение видов. М.; Л., 1937.
Кордюм В. А. Эволюция и биосфера. Киев, 1982.
Корона В. В. Основы структурного анализа в морфологии растений. Свердловск,
1987.
Корочкин Л. И. Молекулярно-генетические аспекты онтогенеза // Биология разви-
тия и управление наследственностью. М., 1986.
Котт X. Приспособительная окраска животных. М., 1950.
Красилов В. А. Современные проблемы соотношения филогении и систематики //
Итоги науки и техники. Зоология позвоночных. М., 1975. Т. 7.
Красилов В. А. Периодичность развития органического мира // Палеонтол. журн.
1987. № 3.
Кузин А. М. Стимулирующее действие ионизирующего излучения на биологиче-
ские процессы. М., 1977.
Кутырев В. А. Универсальный эволюционизм или коэволюция? // Природа. 1988.
№ 8.
Ламарк Ж. Б. Философия зоологии. М.; Л., 1935. Т. 1.
Ламарк Ж. Б. Избранные произведения. М., 1959. Т. 2.
Левич А. И. Структура экологических сообществ. М., 1980.
Левонтин Р. Генетические основы эволюции. М., 1978.
Лейбниц Г. В. Сочинения: В 4 т. М., 1982. Т. 1.
Лункевич В. В. От Гераклита до Дарвина. М., 1960. Т. 1 — 2.
Льюис Г. Г. Очерки животной жизни // Рус. вести. 1860. Т. 29, сент.
Льюс В. Д., Райфа X. Игры и решения. М., 1961.
Любищев А. А. О природе наследственных факторов: Крит, исслед. Пермь, 1925.
Любищев А. А. Проблемы систематики // Проблемы эволюции. Новосибирск, 1968.
Т. 1.
Любищев А. А. К логике систематики // Там же. Новосибирск, 1972. Т. 2.
Любищев А. А. О постулатах современного селектогенеза // Там же. Новосибирск,
1973. Т. 3.
Любищев А. А. Из переписи // Пути в незнаемое. М., 1978. Вып. 14.
Любищев А. А. Проблемы формы, систематики и эволюции организмов. М., 1982.
Магометов А. А. Табасаранский язык. Тбилиси. 1965.
Максимович М. А. О системах растительного царства. М., 1827.
Макфедъен Э. Экология животных. М., 1965.
Маленков Г. Г., Чайковский Ю. В. Выживание мутантного клона: Катастрофиче-
ский отбор // Генетика. 1979. № 10.
Малкей М. Наука и социология знания. М., 1983.
Мальчевский А. С. Кукушка и ее воспитатели. Л., 1987.
Маргелис Л. Роль симбиоза в эволюции клетки. М., 1983.
Меерсон Ф. 3. Алгебра и гармония адаптации // Знание — сила. 1988. А» 3.
Мейен С. В. О соотношении номогенетического и тихогенетического аспектов эво-
люции // Журн. общ. биологии. 1974. Т. 35, № 3.
Мейен С. В. Таксономия и мерономия // Вопросы методологии в геологических нау-
ках. Киев. 1977.
256,
Мейен С. В. Основные аспекты типологии организмов // Журн. общ. биологии
1978. № 4.
Мейен С. В. Следы трав индейских. М., 1981.
Мейен _С. В. Органы размножения голосеменных и их эволюция //Журн. общ.
биологии. 1982. № 3.
Мейен С. В. Принципы исторических реконструкций в биологии. // Системность
и эволюция. М„ 1984.
Мейен С. В. Гипотеза происхождения покрытосеменных от беннетитов путем гамо-
гетеротопии // Журн. общ. биологии. 1986а, № 3.
Мейен С. В. Флорогенез и эволюция растений // Природа. 19866. № 11.
Мейен С. В. Основы палеоботаники. М., 1987а.
Мейен С. В. География макроэволюции у высших растений // Журн. общ биоло-
гии. 19876. № 3.
Мейен С. В. Логико-методологические и теоретические стереотипы в биологии //
О специфике биологического познания. М., 1987в.
Мейен С. В., Соколов Б. С., Шрейдер Ю. А. Классическая и неклассическая био-
логия: Феномен Любищева // Вести. АН СССР. 1977. № 10.
Мейен С. В., Шрейдер Ю. А. Методологические аспекты теории классификации //
Вопр. философии. 1976. № 12.
Мейен С. В. Введение в теорию стратиграфии. М., 1989.
Меклер Л. Б. Общая теория биологической эволюции // ЖВХО. 1980а. № 3.
Меклер Л. Б. О происхождении живых клеток // Там же. 19806. № 4.
Микулинский С. Р. Взгляды К. М. Бэра на эволюцию в додарвиновский период //
Анналы биологии. М., 1959. Т. 1.
Моделирование биогеоценотических процессов. М., 1981.
Молекулярные механизмы генетических процессов: Молекуляр. генетика, эволю-
ция и основы селекции. М., 1985.
Мопертюи П.-Л. Законы движения и покоя, выведенные из метафизического прин-
ципа // Вариационные принципы механики. М., 1959.
Мужчинкин В. Ф. Блочность конструкции у млекопитающих на примере семей-
ства ушастых тюленей // Журн. общ. биологии. 1978. № 5.
Наше общее будущее. Доклад международной комиссии по окружающей среде
и развитию. М„ 1989.
Неручев С. Г. Глобальные геохимические аномалии и биосферные кризисы //
Природа. 1988. № 1.
Никитин Д. И., Никитина Э. С. Процессы самоочищения окружающей среды и пара-
зиты бактерий. М., 1978.
Николис Г., Пригожин И. Самоорганизация в неравновесных системах. М., 1979.
Новосельцев В. Н. Гомеостаз организма как система управления. М., 1973.
Общая генетика. Итоги науки и техники. М., 1978. Т. 3—4.
Одум Ю. Экология. М., 1986. Т. 1—2.
Олдак П. Г. Равновесное природопользование: Взгляд экономиста. Новосибирск,
1983.
О ленов Ю. М. Наследственная изменчивость и эволюция // Журн. общ. биологии.
1946. № 4.
Оленов Ю. М. Проблемы молекулярной генетики. Л., 1977.
Орлов С. А. Необратимость эволюции: некоторые итоги обсуждения вопроса // Воп-
росы развития эволюционной теории в XX в. Л., 1979.
О современном статусе идеи глобального эволюционизма. М., 1986.
Оствальд В. Философия природы. СПб., 1903.
Павлинов И. Я., Россолимо О. Л. Систематика млекопитающих СССР. М., 1987.
Панов Е. Н. Поведение животных и этологическая структура популяций. М., 1983.
Па.нцхава Е. С. Некоторые данные о возможности догликолитического пути в эволю-
ции // Проблемы возникновения и сущности жизни. М., 1973.
Персимфанс. X лет, 1922 — 1932: Программа концерта. М., 1932.
Петухов С. В. Биомеханика, бионика и симметрия. М., 1981.
Пианка. 3. Эволюционная экология. М., 1981.
Преснов Е. В., Исаева В. В. Перестройки топологии при морфогенезе. М., 1985.
Пригожин И. От существующего к возникающему. М., 1985.
Пригожин И., Николис Ж. Биологический порядок, структура и неустойчивости //
Успехи физ. наук. 1973. Т. 109, № 3.
257
Происхождение и развитие половых клеток в онтогенезе. Л., 1968.
Лялль Э„ Тотсель Э., Тукумцев Г. Сопоставительная грамматика эстонского и рус-
ского языков. Таллинн, 1962.
Райков Б. Е. Русские биологи-эволюционисты до Дарвина. М., 1952. Т. 1.
Райков И. Б. Пути эволюции митотического аппарата у низших эукариот// Тр.
Зоол. ин-та. 1986. Т. 144.
Ратнер В. А. Молекулярно-генетические системы управления. Новосибирск, 1975.
Раутиан А. С. Уникальное перо птицы из отложений юрского озера // Палеонтол.
журн. 1978. № 4.
Реймерс Н. Ф. Об экологии - и не только о ней // СССР: Демографический
диагноз. М., 1990.
Риклефс Р. Основы общей экологии. М., 1979.
Рыжков В. Л. Адаптация и эволюция: (Некоторые парадоксы неодарвинизма) //
Философия и теория эволюции. М., 1974.
Рэфф Р., Кофмен Т. Эмбрионы, гены и эволюция. М., 1986.
Свирежев Ю. М. Нелинейные волны, диссипативные структуры и катастрофы в эко-
логии. М., 1987.
Северцов А. Л. Собр. сочинений. М.; Л. Т. 3. 1945; Т. 5. 1949.
Селье Г. Стресс без дистресса. М., 1979.
Серавин Л. Н. Происхождение эукариот // Цитология. 1986. № 6—9.-
Симпсон Дж. Г. Темпы и формы эволюции. М., 1948.
Система, симметрия, гармония. М., 1988.
Системность и эволюция. М., 1984.
Смирнов Е. С. Таксономический анализ. М., 1969.
Соболев Д. Л. Начала исторической биогенетики. Симферополь, 1924.
Соколов Максим. Возможен ли «зеленый» тоталитаризм? // Век XX и мир. 1989.
№ 10.
Солженицын А. И. Письмо вождям Советского Союза (1973 г.) // Диалог Журнал
ЦК КПСС. 1990. № 4.
Спенсер Г. Основания биологии. СПб., 1870. Т. 1 — 2.
Степанов Т. Ф. Записки о политической экономии. СПб., 1844. Ч. 1.
Страхов Л. В. О методе наук наблюдательных / / Журн. М-ва нар. просвещения.
1858. № 1.
Структура и развитие науки. М., 1978.
Сутт Т. Проблема направленности органической эволюции. Таллин, 1977.
Татаринов Л. Л. Очерки по теории эволюции. М., 1987.
Тахтаджян А. Л. Система магнолиофитов. Л., 1987.
Тейяр де Шарден П. Феномен человека. М., 1987.
Теория и методология биологических классификаций. М., 1983.
Тинберген Л. Осы, птицы, люди. М., 1970.
Тихоненко Т. И. О возможности межвидового обмена генетической информацией
в биосфере // Вести. АН СССР. 1979. № 8.
Томилин А. Г. Снова в воду. М., 1977.
Трубецкой Л. С. Избранные труды по филологии. М., 1987.
Уголев А. М. Эволюция пищеварения и принципы эволюции функций. Л., 1985.
Уиттекер Р. Сообщества и экосистемы. М., 1980.
Уиттл Л. Вероятность. М., 1982.
Усов С. А. Таксономические единицы и группы. М., 1867.
Ухтомский А. А. Сочинения. М., 1950. Т. 1.
Ушаков Б. П. Эволюционное значение температурных адаптаций животных //
Успехи соврем, биологии. 1982. Т. 93, № 2.
Фейерабенд П. Избранные труды по методологии науки. М., 1986.
Феллер В. Введение в теорию вероятностей и ее приложения. М., 1964.
Филиппенко ТО. А. О параллелизме в живой природе // Успехи эксперим. биологии.
1924. Вып. 4.
Филиппенко Ю. А. Эволюционная идея в биологии. М., 1977.
Фицнер Л. Л. Биологические поисковые системы. М., 1977.
Фогт (Фохт) К. Человек: Место его в мироздании. СПб., 1865.
Харрисон Дж., Уайнер Дж., Тэннер Дж. Биология человека. М., 1979.
Хесин Р. Б. Непостоянство генома. М., 1984.
Химический мутагенез в селекционном процессе. М., 1987.
258
Холтон Дж. Тематический анализ науки. М., 1981.
Целищев В. В. Философские проблемы семантики возможных миров. Новосибирск,
1977.
Цетлин-М. Л. Исследования по теории автоматов и моделированию биологических
систем. М., 1969.
Чайковский Ю. В. О непрерывном процессе решения матричной игры // Докл. АН
СССР. 1971. Т. 199, № 5.
Чайковский Ю. В. Некоторые проблемы дарвинизма и возможности машинного
моделирования // Журн. общ. биологии. 1972. № 3.
Чайковский Ю. В. Проблема наследования и генетический поиск // Теоретическая
и экспериментальная биофизика. Калининград. 1976. Вып. 6.
Чайковский Ю. В. Выживание мутантного клона // Генетика. 1977а. № 8.
Чайковский Ю. В. Генетическая интеграция клеточных структур как фактор эво-
люции // Журн. общ. биологии. 19776. № 6.
Чайковский Ю. В. Где начало того конца? // Знание — сила. 1980. № 1.
Чайковский Ю. В. Рождение дарвинизма // Теоретические проблемы современной
биологии. Пущино, 1983.
Чайковский Ю. В. «Происхождение видов»: Загадки первого перевода // Природа.
1984. № 7.
Чайковский Ю. В. Разнообразие и случайность // Методы научного познания
и физика. М., 1985.
Чайковский Ю. В. Опыт эко-физиологической макросистемы // Методы исследова-
ния в экологии и этологии. Пущино, 1986.
Чайковский Ю. В. История науки и обучение науке: (На примере дарвинизма) //
Вопр. истории естествознания и техники. 1987а. № 2.
Чайковский Ю. В. Нечеткие закономерности в планетной астрономии // Историко-
астрономические исследования. М., 19876. Вып. 19.
Чайковский Ю. В. О формировании концепции Ч. Дарвина // Науки в их взаимо-
связи. М„ 1988а.
Чайковский Ю. В. Экстремальность как междисциплинарная эвристика // Взаимо-
действие наук как фактор их развития. Новосибирск, 19886.
Чайковский Ю. В. Молодежь в разнообразном мире // Социол. исслед. 1988в. № 1.
Чайковский Ю. В. Человек эволюционирует // Химия и жизнь. 1988г. № 12.
Чайковский Ю. В. История науки и обучение науке: (На примере понятий «слу-
чайности» и «вероятности») //Вопр. истории естествознания и техники. 1989а.
№ 4.
Чайковский Ю. В. Наука о разнообразии // Химия и жизнь. 19896. № 1.
Чайковский Ю. В. Охрана природы и интеграция знаний // Естественнонаучное
мышление и современность. Киев, 1989в.
Чайковский Ю. В. Идея равновозможности в физике и биологии // Физическое
знание: его генезис и развитие. М., 1990.
Чаянов А. В. Путешествие моего брата Алексея в страну крестьянской утопии
(1924 г.) // Чаянов А. В. Избранные повести. М., 1989.
Численко Л. Л. О структуре таксонов и таксономическом разнообразии. // Журн.
общ. биологии. 1977. № 3.
Чудинов Э. М. Природа научной истины. М., 1977.
Шлегель Г. Общая микробиология. М., 1987.
Шмальгаузен И. И. Основы сравнительной анатомии позвоночных. М., 1938.
Шмальгаузен И. И. Факторы эволюции. М., 1968.
Шрейдер Ю. А. Об одной модели семантической теории информации // Проблемы
кибернетики. М., 1965. Вып. 13.
Шуков В. А. Образ биологической реальности // О специфике биологического по-
знания. М., 1987.
Эволюция и биоценотические кризисы. М., 1987.
Эгертон Ф. Развитие концепции баланса природы // Историко-биологические ис-
следования. М„ 1978. Вып. 6.
Эдельман Дж. М. Топобиология //В мире науки. 1989. № 7.
Эйген М., Шустер П. Гиперцикл. М., 1982.
Экология и эволюционная теория. Л., 1984.
259
Юдин Л. Л. О возможностях генетического исследования агамно размножающихся
простейших // Кариология и генетика простейших. Л., 1976.
Яблоков А. В. Фенетика: эволюция, популяция, признак. М., 1980.
Янч Э. Прогнозирование научно-технического прогресса. М., 1974.
Adams W. J. Life and times of the central limit theorem. N. Y., 1974.
Agassiz L. Contributions to the natural history of USA: First monograph: Essay on
classification. Boston, 1857. Pt 1.
Ainsworth G. C. Introduction to the history of mycology. Cambridge, 1976.
Alt F. W., Baltimore D. Joining of immunoglobulin heavy chain gene segments //
Proc. Nat. Acad. Sci. US. 1982. Vol. 79, N 13.
Bacq Z. Al., Goutier R. Mechanisms of action of sulfur-containing radioprotectors //
Brookhaven Symp. Biol. 1968. N 20.
Baer К. E. Ein Wort fiber einen blinden Fisch — als Bildungs-Hemmung//Bull.
Acad. Sci. St. Petersbourgh. 1861. Vol. 4, N 1.
Barbieri M. The ribotype theory of the origin of the life //J. Theor. Biol. 1982. Vol.
91, N 4.
Bauhin C. Pinax theatri botanici. Basel, 1623.
Bergey’s manuel of systematic bacteriology. Baltimore, 1984. Vol. 1.
Bhaud Y., Soyer-Gobillard M.-O. DNA synthesis and cell cycle of a primitive Dino-
flagellate // Protistologica. 1986. T. 22, N 1.
Bogorad L. Evolution of organelles and eucariotic genoms //Science. 1975. Vol. 188,
N 4191.
Bowler P. J. Darwinism and the argument from design: Suggestion for reevaluations //
J. Hist. Biol. 1977. Vol. 10, N 1.
Broadwater S., Scott J. Fibrous vacuole associated organelles in the Florideophiceae:
A new interpretation of the «appareil cinetique» // Phycologia. 1983. Vol. 22, N 3.
Brooks D. R. What’s going on in evolution? A brief guide to some new ideas of evolutio-
nary theory //Canad. J. Zool. 1983. Vol. 61, N 12.
Brooks D. R., Wiley E. O. Evolution as entropy. Chicago; L., 1986.
Burkhardt R. W. The spirit of system: Lamarck and evolutionary biology. Cambridge
(Mass.), 1977.
Cedergren R., Grosejean H. On the primacy of primordial RNA // Biosystems. 1987.
Vol. 20, N 2.
Chaikovsky Yu. V. To the evolutionary thermodynamics//Lectures in theoretical
biology. Tallinn, 1988.
Chamberlin T. C. The method of multiple working hypotheses // J. Geol. 1931. Vol. 39,
N 2.
Clark J. M. Novel non-templated nucleotide addition-reactions catalized by procaryotic
and eucaryotic DNA polymerases // Nucl. Acids Res. 1988. Vol. 16, N 20.
Cluiton~Brock J., Corbet G. B., Hills M. A review of the family Canidae // Bull. Brit.
Mus. (Natur. Hist.). Zool. 1976. Vol. 29, N 3.
Costas E., Goyanes V. J. Ultrastructure and division behaviour of dinoflagellate
chromosomes // Chromosoma. 1987. Vol. 95, N 6.
Cox E. C., Gibson T. C. Selection for high mutation rates in chemostats // Genetics.
1974. Vol. 77, N 2.
Dixon D. After the man: A zoology of future. L., 1981.
Engen S. Stochastic abundance models in ecology // Biometrics. 1979. Vol. 35, N 1.
Freke H. On the origin of species through organic affinity. L., 1861.
Fritz L., Triemer R. E. An ultrastructural study of mitosis in a marine Dinoflagellate //
J. Protozool. 1983. Vol. 30, N 2.
Gale B. G. Evolution without evidence. Brighton, 1982.
Gavrila L., Michaesku G. The fine structure of mesokaryotic genome: Multiple nucleo-
lar-organizing chromosomes in Prorocentrum micans // Caryologia. 1986. Vol.
39. N 1.
Chiselin M. T. The triumph of the Darwinian method. Berkeley, 1969.
Closer H.-J. Allgemeine und biologische Evolutionstheorie // Biol. Zentr.-Bl. 1987.
Bd. 106, H. 6.
Goldsmith E. Thermodynamics of ecodynamics? // Ecologist. 1981. Vol. 11, N 4.
Goodwin В. C., Webster G. C. Rethinking of the origin of species by natural selection //
Rev. Biol. 1981. Vol. 74, N 1/2.
Grasse P~P. L’evolution du vivant. P., 1973.
260
Haeckel E. Generelle Morphologie der Organismen. B., 1866. Bd. 2: Allgemeine Ent-
wicklungsgeschichte der Organismen.
Hale M. The primitive origination of mankind considered and examined according
to the light of nature. L., 1677.
Hartley B. S., Altosaar I., Dothie J. M., Neuberger M. S. Experimental evolution of
a xylitol dehydrogenase // Structure-function relationships proteins. Amsterdam,
1976.
Heer O. Flora tertiaria Helvetiae. Winterthur, 1859. Bd. 3.
Hunde // Brockhaus’ Konversations-Lexicon. Leipzig, 1908. Bd. 9.
Jantsch E. Design for evolution. N. Y., 1975.
Jantsch E. The self-organizing universe: Scientific and human implications of emer-
ging paradigm of evolution. Oxford; N. Y., 1980
Jussieu A. L. Exposition d’un nouvel ordre de plantes adopte dans les demostrations du
jardin royal // Hist. Acad. roy. sci. annee 1774. Avec mem. math. phys. P., 1778.
Pag. II.
Kant I. Kritik der Urteilskraft. B., 1790.
Kepler J. Gesammte Werke. Mfinchen, 1938. Bd. 1.
Kimura M., Ohta T. Theoretical aspects of population genetics. Princeton, 1971.
Lake J. A., Clark M. И7., Henderson E. Eubacteria, halobacteria and the origin of
photosynthesis: the photocytes // Proc. Nat. Acad. Sci. US. 1985. Vol. 82, N 11.
Lewes G. H. Problems of life and mind. L., 1875. Vol. 2.
Lewin H. A downward stope to greater diversity // Science. 1982. Vol. 217, N 4566.
Linne (Linnaeus) C. Philosophia botanica. Stockholm, 1751.
Linne C. Species plantarum. Stockholm, 1753. T. 1 — 2.
Linne C. Systema naturae per regna tria naturae. 12 ed., reform. Stockholm, 1766.
T. 1.
Lovelock J. E. Geophysiology // Transactions Roy. Sci. Soc. Edinburgh: Earth Sci.
1989. Vol. 80. P. 170.
Margulis L. Colchicine-sensitive microtubules // Intern. Rev. Cvfol 1973. Vol. 34.
Margulis L. On the evolutionary origin and possible mechanism of colchicine-sensitive
mitotic movements // Biosystems. 1974. Vol. 6, N 1.
Margulis L. The microbes’ contribution to evolution // Ibid. 1975. Vol. 7, N 2.
Martin С. P. Psychology, evolution and sex. Springfield, 1956.
Maupertuis P. L. Oeuvres. Lyon, 1756. T. 2.
Меуеп S. V. Plant morphology in its nomothetical aspects // Bot. Rev. 1973. Vol. 39,
N 3.
Меуеп S. V. Fundamentals of palaeobotany. L., 1987.
Morowitz H. J. Entropy and nonsense // Biol, and Philos. 1986. Vol. 1, N 4.
Morris H. The scientific case for creation. San Diego, 1981.
Murray A. On Mr Darwin’s theory of the origin of species // Proc. Roy. Soc. Edinburgh.
1857/1862. Vol. 4, N 52.
Nagi W. Molecular phylogeny // Patterns and process. B. etc., 1986.
Norris K. S., Lowe С. H. An analysis of background color-matching in amphibians and
reptils // Ecology. 1964. Vol. 45, N 3.
Oakley В. R., Dodge J. D. Mitosis in cryptophyceae // Nature. 1973. Vol. 244, N 5417.
Ostwald W. Vorlesungen fiber Naturphilosophie. Leipzig, 1902.
Owen R. Principes d’osteologie comparee ou recherches sur [’archetype et les homo-
logies du squelette vertebre. P., 1855.
Prigogine I., Wiame J. M. Biologie et thermodynamique des phenomenes irreversibles //.
Experientia. 1946. Vol. 2, N 11.
Raven P. H. Catastrophic selection and edaphic endemism // Evolution. 1964. Vol.
18, N 2.
Sato T. A broad parasitic catfish of mouthbroading cichlid fishes in lake Tanganyika //
Nature. 1986. Vol. 323, N 6083.
Schwartz J. B. Charles Darwin’s debt to Malthus and Edward Blyth // J. Hist. Biol.
1974. Vol. 7, N 2.
Scudder S. H. Is mimicry advantageous? Nature. 1870. Vol. 3, Dec. 22.
Simon- Bichar-Breaud J. Formation de la crypte flagellaire et evolution de son contenu
au cours de la gametogenese male chez Bonnemaisonia hamifera (Rodophycee) //
C. r. Acad. sci. D. 1972. Vol. 274, N 12.
[Spencer H.] The social organism // Westminster Rev. 1860. Vol. 17, N 1.
Spranger E. Lebensformen. Halle, 1914.
261
Steenis G. G. G. J. van. Plant speciation in Malesia, with special reference to the theory
of non-adaptive saltatory evolution // Biol. J. Linn. Soc. 1969. Vol. 1, N 1/2.
Stevens P. F. Metaphors and typology in the development of botanical systematics
1690—1960, or the art of putting new wine in old bottles // Taxon. 1984. Vol. 33,
N 2.
Suarez F. Disputationes metaphysicae. Mainz, 1600. T. 1.
Thompson J. L. Preservation of wilderness and the good life // Environmental philoso-
phy / R. Elliot, A. Gare. St. Lucia (Queensland); University Park (Pennsylvania);
London, 1983.
1 uiiegawa S. Somatic generation of immune diversity (Nobel lecture) //Biosci. Rep
1988. Vol. 8, N 1.
Trautschold H. Uberimnge und Zwischenvarietaten // Bull. Soc. Imp. Natur. Moscow.
1860. Vol. 33, N 4.
Trincher K. Der Irrtum des Jahrhundert: Manfred Eigen’s Umwalzung in der Biole-
gie // Cytobiol. Rev. 1984. Vol. 8, N 1.
Tripodi G., Masi F. Unusual structures in spermatial vesicles of the red algae // Plant
Syst. and Evol. 1983. Vol. 143, N 3.
Vandel A. La genese du vivant. P., 1968.
Van Gelder R. G. A review of canid classification//Amer. Mus. Novitates. 1978.
N 2646
Vermeij G. T. Evolution and escalation: An ecological history of life. Princeton, 1987.
Weaver W. Science and complexity // Amer. Sci. 1948. Vol. 36.
Willis J. C. Age and area. Cambridge, 1922.
Wintrebert P. Le vivant createur de son evolution. P., 1962.
Withers A. W., Albert R. C., Lewin R. A. et al. Photosynthetic init size, carotenoids and
chlorophyll-protein composition of Prochloron sp., a prokaryotic green algae //
Proc. Nat. Acad. Sci. US. 1978. Vol. 75. N 5.
Wu T.-T. Experimental evolution in Bacteria // CRC CriL Rev. Microbiol. 1978. Vol.
6, № 1.
Young R. M. Darwin’s metaphor: does nature select? // Monist. 1971. Vol. 55, № 3.
Yule G. U. A mathematical theory of evolution based on the conclusion of Dr J. C. Wil-
lis, FRS // Philos. Trans. Roy. Soc. London B. 1924. Vol. 213.
Zimmermann W. Evolution: Die Geschichte ihrer Probleme und Erkenntnisse. Frei-
burg; Munchen, 1953.
ИМЕННОЙ УКАЗАТЕЛЬ *
Абрамсон Н. И. 4
Авенариус Р. (Avenarius) 166
Агаев М. Г. 109, 164
Агассиц Л. (Agassiz) 55, 139, 169, 179.
180, 213
Адансон М. (Adanson) 21
Азимов A. (Asimov I.) 199
Акчурин И. А. 221, 223, 224
Александров В. Я. 188, 224
Александров Д. А. 213
Анохин И. А. 122
Арапов М. В. 66, 86, 87, 91
Аристотель 9, 10, 11, 14, 16, 21, 34, 113,
115, 228
Аршавский И. А. 4
— адаптивные модификации, сдвиг
в онтогенезе и эволюции 165
— анаболизм и катаболизм 152, 163
— время биологическое 164
— гипоксия, фактор эволюции 249
— двигательная активность как фактор
эволюции 173
— закаливание детей холодом 109
— и Бэр 168
— и Пригожин 201, 237
— изменение онтогенеза при нагрузке
106
— индивидуальное и историческое раз
витие 109
— метаболическая спираль 163, 164
— наследуемость приобретенных приз-
наков — ложная проблема 164
— негэнтропийный уровень организа-
ции 163
— онтогенез как преобразование фи-
зиологии 233
— основной механизм эволюции 165
— физиологический стресс 178
— эволюционная физиология 163
— экспериментальная эволюция 164
— эмбрионизация как фактор эволюции
168
Ауэрбах Ш. (Auerbach) 95
Ахутин А. В. 12
Бабков В. В. 4, 32, 174
Базилевич Н. И. 204
Балашов Ю. В. 7, 210
Балтимор Д. (Baltimore) 196
Бальби A. (Balbi) 86
Баранцев Р. Г. 252
Барбиери М. (Barbieri) 224
Барулина Е. И. 117, 121
Баугин К. (Bauhin) 18, 19, 30, 47, 147,
148
Бауэр Э. С. (Bauer) 164
Беклемишев В. Н. 43, 72, 139, 163
Белоусов Л. В. 4, 170
Берг Л. С. 32, 64, 69, 112, ИЗ, 120, 125,
169, 225, 235
Берги Д. (Bergey) 140, 147
Бергсон A. (Bergson Н.) 60, 173, 198,
202, 209, 226, 237
Бернал Дж. (Bernal J.) 12, 185
Бернгарди И. (Bernhardi J.) 30
Бернулли Я. (Bernoulli J.) 87, 89
Берталанфи Л. (Bertalanffy) 198
Битов А. Г. 126
Бляхер Л. Я. 17, 39, 64
Бобрецкий Н. В. 59
Богачев В. Н. 244, 253
Богданов А. А. [Малиновский] 12, 13.
96, ИЗ, 174, 198
Богомолов А. С. 227
Богородицкий В. А. 49, 56
Бок И. (Bock/Tragus) 17, 18
Болдуин Дж. (Baldwin J.) 107
Больцман Л. [Boltzmann] 198, 220
Бор Н. (Bohr) 16, 31, 65, 68, 69, 112,
208, 252
Борхсениус С. Н. 147, 193
Браун А. Д. 108
Брем A. (Brehm) 52, 121
Брода Э. (Broda) 186, 189
Брони Г. (Bronn Н.) 24, 26, 27, 57, 58,
148, 227, 237, 249
Брукс Д. (Brooks) 212, 237, 238
Бруно Дж. (Bruno) И
Брунфельс О. (Brunfels) 17
Будыко М. И. 238
Бунур Л. (Bounoure) 171
Бэр К. (Baer) 21, 237
— естественная система 140, 148
— «закон бережливости» 236
— и Агассиц, и Кильмейер 169
— и Брони 24
— и Геккель 27
— и естественное богословие 114
— и Кант 112
— и Спенсер 130
— и Янч 236
— критерий Мильн-Эдвардса — Бэра
24
* Ссылки на вторых соавторов указаны
лишь при необходимости
263
— неоламаркизм 168
— переходные формы 44
— «победа духа над материей» 236
— прогресс морфологический 24
— рекапитуляция 169
— сходство и родство 27
— типы Кювье и онтогенез 24
— традиция Везалия — Монро — Бэра
114
— целесообразность и случайность 69
— эмбрионизация у слепой рыбы 168
— ядро и периферия 43
— ядро класса млекопитающих 44
— Ziel und Zweck 112
Бэтсон У. (Bateson W.) 123, 133
Бюффон Ж.-Л. (Buffon) 23, 29, 30, 44
Вавилов Н. И. 8, 16, 27, 31 -33, 35, 37,
51, 52, 63-65, 116, 117, 140, 151, 213,
248
Вайнцвайг М. Н. 105
Вайцзеккер X. (Waizsacker) 220
Вальт — см. Реммель
Ван Гельдер Р. (Van Gelder) 45, 48, 65
Вандель A. (Vandel) 214, 215
Варден Б. (Waerden) 76—78, 89
Варшавский В. И. 98, 99, 103
Ватти К. В. 164
Везалий A. (Vesalius) 21
Вентребер П. (Wintrebert) 171, 196
Вермей Г. (Vermeij) 179, 235, 252
Вернадский В. И. 13, 24, 184, 203, 204,
219, 236, 237, 239, 253
Вигнер Е. (Wigner) 7, 67
Вили Э. (Wiley) 237, 238
Виллис Дж. (Willis) 84—87, 141, 172
Виллис Т. (Willis) 22
Вильямс В. Р. 185
Виндельбанд В. (Windelband) 15, 71
Вольтерра В. (Volterra) 4
Воронцов Н. Н. 143, 146, 155, 160, 234.
242, 245
By Т.-Т. (Wu) 96, 109
Гайденко П. П. 9
Гакке (Нааске) 232
Галилей (Galilei) 12, 13
Гаркави Л. X. 109, 175
Гартман Н. (Hartmann) 73
Гаузе Г. Ф. 106
Гегель Г. (Hegel) 113, 207
Гегенбаур К. (Gegenbaur С.) 63
Геер О. (Heer) 214
Геккель Э. (Haeckel) 21 27, 55, 63, 64
140, 148, 165, 169
Гексли Т. (Huxley) 27, 45
Гельмгольц Г. (Helmholtz) 43
Герцшпрунг Э. (Hertzsprung) 70
Геснер К. (Gesner) 18, 20, 21
Гёбель К. (Goebel) 63, 114
Гёдель К. (Godel) 172, 173, 176, 182, 208.
211, 227
Гёте И. (Goethe) 10, 23
Гизеке П. (Giseke) 148, 149
Гиппократ 9
Гленсдорф П. (Glansdorf) 201
Голсуорси Дж. (Galsworthy) 137
Голубовский М. Д. 110
Гольдшмидт Р. (Goldschmidt) 106
Горовиц Н. (Horowitz) 185, 189
Григорьева А. В. 4, 129
Гроссгейм А. А. 149
Гумбольдт A. (Humboldt) 35
Гурвич А. Г. 108
Гусейнов А. М. 224
Дана Дж. (Dana) 249
Данилевский Н. Я. 114, 120, 230, 233,
251
Дарвин Ч. (Darwin) 15, 17, 23, 24, 58,
68, 84, 85, 113, 128, 148, 167, 177, 178,
180, 181, 206, 213, 227, 231
— адаптивный компромисс 133
— аналогия и гомология изменчивости
63
— андродиэция 179
— борьба за жизнь 136, 164, 207
— гомология и родство 66
— «демон Дарвина» 199
— диатропический прогноз 179, 248
— естественная система 140
— естественное богословие 15
— за и против него 195, 230
- и Берг 225, 235
— и Вавилов 63
— и Вернадский 236
— и Геккель 27, 140
— и Кетле 76—78
— и Кювье 163
- и Ламарк 27, 133, 181, 225, 233
— и Максвелл 12, 213
— и Мальтус 136, 164, 174
— и Мопертюи 30
— и эмерджентность 227, 236
— и Янч 214
— идеология 253
— избыточность размножения и отбор
174, 207
— «кошмар Дженкина» 174
— метаморфоз камбал 233
— .методология 252
— морфология 27
— наименьшее действие творения 30
— наследственность и унаследование
111
— натурфилософ 119
— отбор, смена взглядов 174
— пангенез 29
— параллелизм изменчивости 31—33
— победители и побежденные 164
— причина победы дарвинизма 220
— прогресс 252
— разнообразие и эволюция 15, 181
264
— родословное древо 143
— случайность и сложность 75
— сродство и родство 21, 207
— статистическое мировоззрение 77
— целесообразность 110, 111, 119
— эволюция и поведение 104
— — и смена функций 105
— — и статистика 12, 77, 236
— — как накопление различий 111
— экология борьбы 136, 164, 207, 251
Дарвин Э. (Darwin) 29, 163
Девис П. (Davies) 10, 219
Декарт Р. (Descartes) 12, 28, 29
Де Фриз Г. (De Vries) 29, 84, 85
Дженкин Ф. (Jenkin) 174
Джеффри Ч. (Jeffrey) 47, 141
Диксон М. (Dixon) 187, 190
Догель В. А. 58, 131, 136, 154
Додж Дж. (Dodge) 146, 193
Донати В. (Donati) 141
Дорн A. (Dohrn) 105
Дроздов—Тихомиров Л. Н. 95
Дриш Г. (Driesch) 113, 198, 206
Дуда В. И. 147
Еленкин А. А. 37
Елфимов Г. М. 227
Женермон Ж. (Genermont) 92, 107
Жерихин В. В. 176
Жоффруа Сент-Илер Э. (Geoffroy
Saint-Hilaire) 23, 25—27, 64, 163
Журавлева И. Т. 147, 157
Жюссье A. (Jussieu) 17, 20, 21, 33, 40
Жюсье Б. (Jussieu) 20
Заварзин А. А. 32, 63, 64, 66
Заварзин Г. А. 4, 132
— биосфера прокариотная 153, 185, 186, ,
216
— вулканы как колыбель жизни 186.
239
— геохимические катастрофы архея и
протерозоя 240
— геохимические функции жизни 185
— диатропический прогноз у бактерий
151, 248
— и Вернадский 239
— и Лавлок 239
— и Мейен 177
— и Уиттекер 149
— искусственные геохимические барье-
ры 240
межцарственные параллелизмы 150
— окисляющая среда первичная 186
— первая экосистема 240
— пространство логических возможнос-
тей 177
— разнообразие первичное 216
— система царств 149, 153
— эвкариоты и прокариоты, их роль
151, 153, 186
— экологические параллелизмы 150
— эко-физиологическая универсаль-
ность прокариот 151
Заренков Н. А. 143
Заславская Т. И. 244
Заславский Г. М. 68, 91, 136
Захваткин А. А. 168
Здродовский П. Ф. 108
Зевс 10
Зеликман Э. А. 122
Золотарев В. М..86, 90
Зюсс Э. (Suss) 184
Иванов В. В. 49, 56
Ивановский А. Б. 169, 170
Игнатов М. С. 124
Йорений М. (Johrenius) 34
Камшилов М. М. 185, 186
Канаев И. И. 17, 58, 63, 165, 169
Канцлер (Kandler) 155
Кант И. (Kant) 111-115, 119
Карно С. (Carnot) 203, 247
Карпинская Р. С. 13, 152
Карпов В. П. 9, 167, 184
Кеплер И. (Kepler) 13, 68, 74
Кетле A. (Quetelet) 76, 77, 88
Кизель В. А. 238
Кильмейер К.-Ф. (Kielmeyer) 169
Кимура М. (Kimura) 80, 174
Кириллова Н. П. 4, 5
Козополянский Б. М. 19, 148
Козырев Н. А. 228
Колмогоров А. Н. 70, 71, 222
Комаров В. Л. 230
Коп Э. (Соре) 31, 175, 215
Коперник Н. (Copernicus) 74
Кордюм В. А. 194
Корогодин В. И. 4
Корона В. В. 40, 140, 143, 163, 165
Корочкин Л. И. 4, 110, 171, 173, 221
КоттХ. (Cott) 115, 118, 123-126, 132—
134
Коши О. (Cauchy А.) 88, 89
Красилов В. А. 138, 142, 143, 177, 231,
248
Кренке Н. П. 40, 51, 57, 140, 213
Кропоткин П. А. 173, 206
Кудрин Б. И. 4, 90
Кузин А. М. 108
Кун Г. (Kuhn) 104, 135
Кун Т. (Kuhn) 198
Кутырев В. А. 212
Кювье Ж. (Cuvier) 22—24, 26, 27, 163.
173
Лавлок Дж. (Lovelock) 13, 132, 206,
228, 238, 239
Лавуазье A. (Lavoisier) 184
Лакатос/Лакатош И. (Lakatos) 67, 197,
198 230 253
Ламарк Ж.-Б. (Lamarck) 27, 58, 133,
207, 248
265
— биосфера 184
— дихотомический ключ 34, 35
— и Аршавский 163, 164
— и Э. Дарвин 163
— и Карпов, и Кювье 167
— место в классическом эволюционизме
225, 235
— низшие живут за счет внешних разд-
ражений 111
— органическая природа земной коры
184
— сравнительный метод 181
— прогресс техники влечет гибель при-
роды 219
— формы систем 148
— усложнение организмов и система-
тика 149
— эволюция как физиологический про-
цесс 163, 167
Лаплас П.-С. (Laplace) 12, 13
Левич А. П. 71, 131
Леви-Строс К. (Levi-Straus) 35
Левонтин Р. (Lewontin) 198
Ле Шателье A. (Le Chatelier) 95
Лейбниц Г.-В. (Leibniz) 3, 11-14, 22,
28, 29, 110, 253
Леритье Ф. (L’Heritier) 7
Либерман Е. А. 105
Линней К. (Linne) 17, 21, 24, 46
— адрес и систематика 20, 46
— андродиэция 179
— бинарная и биномиальная номенкла-
туры 19
— бытовые названия 19
— возврат систематиков к Л. 45
— «географический» принцип класси-
фикации 142
— дихотомия 20, 34, 35
— естественная и искусственная систе-
мы 19, 20, 140
— «естественный метод» 148
— животные, роль в природе 153
— зоофиты 148
— и Вюффон 30
— и Уэвелл 140
— иерархический принцип 18, 20
— «не признаки задают род...» 20, 137
— нормальные экземпляры (голотипы)
127
— параллелизмы и систематика 30
— разнообразие, единый взгляд 20
— расхожее мнение о Л. 19
— семь семейств по Л. 20
— «систематика по Линнею» 33
— сродство 142
— царство человека 19
— целостность объекта 137
— человек в отряде приматов 19
— югулярные плавники рыб 59
Лобашев М. Е. 164
Лункевич В. В. 17, 184
Луэс/Льюис Дж. (Lewes) 227
Львов A. (Lwoff) 185
Льюс В. (Luce) 100, 104, 241
Любищев А. А. 17, 124, 140, 143, 213,
222, 230, 252
— анализ системы Линнея 20
— — целесообразности 112
— апории 220
— географический стиль 62
— гипертелия (сверхцелесообразность)
119
— и Янч 209
— изменчивость, наследственность
и отбор 174
— кладистический метод 46
— клептогенез (присвоение органов)
123
— меторизис 125
— «Монблан фактов» 198
— морозные узоры 16
— морфология историческая и конст-
руктивная 27
— непонятное игнорируют 67
— общность против точности 68—69
— параллелизмы и Дарвин 32
— польза отлична от приспособленнос-
ти 120
— понятийный анализ 220
— повторность в систематике 8
— «приспособительная» окраска 114,
116
— пробабилизм 68
— пророческие стадии эволюции 169
— псевдотелизм 119
— рутинизация 59, 107
— унаследование (фиксация) 107
— четыре причины по Аристотелю 14
Магомет 124
Магометов А. А. 41
Майварт Дж. (Mivart) 63
Мак-Ати У. (McAtee) 132, 133
Максвелл Дж. Клерк (Maxwell) 12, 13,
198, 199, 213
Максимович М. А. 21, 31, 34, 44, 213
Макфедьен Э. (MacFadyen) 129, 135
Маленков Г. Г. 4, 8, 84, 102
Малкей М. (Mulkay) 15
Мальбранш Н. (Malebranche) 29
Мальтус Т. (Malthus) 136, 163, 174,
213
Мальчевский А. С. 121, 122, 125
Маньоль П. (Magnol) 18—20, 141
Маргелис Л. (Margulis) 154, 197
Маркс К. (Marx) 134
Мейен С. В. 5, 6, 12, 17, 138, 198, 213,
220
— активная методология 3
— архетип 39, 41, 49, 92, 132 — 135, 144.
164, 177
— бытовая классификация 34
266
— время индивида 228
— гамогетеротопия 177
— — и происхождение цветковых 178
— глобальный флорогенез 180
— гомологизация 65
— гомологические ряды, природа их 66
— грамматика системы 36
— граница (в системе) 142, 150
— диатропическая концепция 3, 32
— — — и структурализм 4, 35
— доказательство эволюции 181
— «закон» неспециализированного
предка 175
— законы образования форм 53
— застой в теории биологии 230
— и Агассиц 179
— и Вавилов, и Кренке 51
— и Оуэн 38
— иерархия системы 141
— историко-методологический анализ
3, 38
— итеративный метод 38
— логика системы голосеменных 182
— мерой 38, 124
— мероно-таксономическое несоответ-
ствие 100, 110
— механизм эволюции 4, 177, 180
— морфология и систематика 33
— натурфилософ 231
— номотетическая эволюция 177
— обобщение научное 232
— обратимость тенденции 178
— онтогенез, переключение 233
— параллель уродств и таксонов 40
— понимание типологии 33, 38
— «попустительство отбора» 176
— приспособленность, методология ис-
следования 115, 225
— проблема разнообразия 15
— пунктуализм, концепция 214, 215
— расчленение листа 52, 53
— редукционизм и холизм 208, 253
— рефрен 42, 51, 213
— синтетические типы 180
— случайность 67, 176
— стиль, понятие 124
— типологическая упорядоченность 44,
48, 54 (см. также Акчурин)
— - экстраполяция 222, 232
— типологический прогноз 43
— транзитивный полиморфизм 178, 180.
181
— «убежище невежества» 252
— устойчивость таксономии 140
— физиологическое разнообразие 163
— фиксации уровней 253
— фитоспрединг 180
— фитохории 219
— форма и функция 4, 163, 232
— целостность используют все 172
— эволюционный метод 180
— эволюция путем отбора мутаций 222
Меклер Л. Б. 107, 163, 171, 191, 207, 208
Менделеев Д. И. 151
Мёррей Э. (Murray А.) 116, 206
Микулинский С. Р. 148
Милитарев В. Ю. 140
Мильн-Эдвардс A. (Milne Edwards Н.)
24, 25, 27, 30
Милютин А. А. 94, 96, 226
Моно Ж. (Monod) 214
Монро A. (Monro) 22, 114
Мопертюи П.-Л. Моро (Maupertuis) 29.
30, 40, 54, 92, 173, 206, 226
Мужчинкип В. Ф. 165
Мюнхаузен К.-Ф. И. (Miinchhausen)
140, 202
Мюнтцинг A. (Miintzing) И
Налимов В. В. 231
Нейман Дж. фон (Neumann) 101
Неккер Н. (Necker) 148
Неручев С. Г. 236, 248
Никитин Д. И. 104
Николис Г. (Nicolis) 109, 201, 203
Новиков М. (Nowikoff) 64
Новосельцев В. Н. 129, 131
Нуссбаум М. (Nussbaum) 171
Ньютон И. (Newton) 12, 13, 29, 74.
128, 221
Огурцов А. П. 12, 198, 226
Одум Ю. (Odum Е.) 128, 131, 132, 174,
235, 239, 247
Окен Л. (Oken) 25
Оккам В. (Ockham, Occam) 222
Олдак Г. П. 242
Оленов Ю. М. 97, 106
Опарин А. И. 183
Орлов С. А. 232
Оствальд В. (Ostwald) 7, 15—17, 111
Оуэн Р. (Owen) 27
— аналогия 25, 63, 64
— архетип 25, 39, 41, 130
— гемапофиз 26
— гомология 25, 38, 49, 63, 64, 125
— — конвергентное понимание 64
— — общая 25, 26
— — серийная 25, 26, 49, 65
— — частная 25, 26, 65
— — ядро и периферия 49
— гомотип 26
— и Вавилов 65
— и Гегенбаур 63
— и Геккель, и Новиков 64
— и Жоффруа 25, 26, 64
— и Любищев 125
— и Мейен 38, 39, 41
— коракоид 26
— позвонок идеальный 25
— — естественный типичный 25
— царство простейших 148
Павлинов И. Я. 46
267
Павлов А. П. 169
Панов Е. Н. 37, 122, 136
Панцхава Е. С. 234
Парацельс (Paracelsus) 27, 28, 127
Парето В. (Pareto) 86, 87
Патти Г. (Pattee) 223, 227, 228
Паултон Э. (Poulton) 118, 119
Петухов С. В. 50, 51, 155, 233
Пианка Э. (Pianka) 134, 135
Питтендрай К. (Pittendrigh) ИЗ
Пифагор 9, 11, 253
Платон 10, 11
Поп A. (Pope) 29
Преснов Е. В. 64, 173, 233
Пригожин И. (Prigogine) 195, 199,
201-203, 210, 220, 227, 228, 237
Пристли Дж. (Priestley) 184
Пуассон С. (Poisson) 87
Пэйли У. (Paley) 15
Пялль Э. (Pall) 49, 56
Рабле Ф. (Rabelais) 147
Райков Б. Е. 141
Райков И. Б. 195
Расницын А. П. 110, 133, 176
Ратнер В. А. 92
Раутиан А. С. 4, 40, 43
Реймерс Н. Ф. 242
Ремане A. (Remane) 63
Реммель М. (Remmel/Valt) 4, 236
Риклефс Р. (Ricklefs) 132
Робинэ Ж.-Б. (Robinet) 23
Розов М. А. 215
Розова С. С. 47
Роу С. (Rowe) 245
Руденко А. П. 174
Рыжков В. Л. 114, 138
Рэй Дж. (Ray) 18, 21
Рэфф Р. (Raff) 169
Свирежев Ю. М. 222
Северино A. (Severino) 22
Северцов А. Н. 25, 58, 92, 163, 173, 175
Селье Г. (Selye) 108, 109, 175
Серавин Л. Н. 145, 165
Сибатани A. (Sibatani) 4
Симпсон Дж. (Simpson) 232
Скаддер С. (Scudder) 115
Смирнов Е. С. 21, 33, 140
Смирнов С. Г. 130
Смит Адам (Smith) 242
Смит Н. (Smith) 133
Соболев Д. Н. 32, 40, 54, 169
Соколов Максим 246
Солженицын А. И. 246
Спенсер Г. (Spenser) 15, 111, 130—132,
168
Степанов Т. Ф. 86, 213,
Страхов Н. И. 38, 177, 252
Суарес Ф. (Suarez) 14, 223, 228
Сутт Т. (Sutt) 112—113
Татаринов Л. П. 176, 177, 248
Тахтаджян А. Л. 21, 140, 141
Тейяр де Шарден П. (Telliard de
Chardin) 162, 219, 231
Тимофеев-Ресовский Hi В. 119
Тинберген Н. (Tinbergen) 116, 230
Тихоненко Т. И. 172
Тициус И. (Titius) 49
Томилин А. Г. 61
Томпсон Дж. (Thompson) 246
Тонегава С. (Tonegawa) 196
Траутшольд Г. (Trautschold) 178
Тролль В. (Troll) 63
Трубецкой Н. С. 49, 56, 215
Уголев А. М. 132, 164-166
Уивер У. (Weaver) 213, 253
Уиттекер Р. (Whittaker) 131, 137, 138.
144, 148, 149, 158, 159, 204
Ульянкина Т. И. 196
Уотермэн Т. (Waterman) 129
Урманцев Ю. А. 8, 51, 67, 246
Усов С. А. 127, 137, 251
Ухтомский А. А. 166, 175
Ушаков Б. П. 4, 106
Уэвелл /Юэлл/ (Whewell) 44, 48, 140
Фаминцын А. С. 143, 192
Фейерабенд П. (Feyerabend) 198
Феллер У. (Feller) 82, 88
Филипченко Ю. А. 31, 32, 230, 236
фицнер Л. Н. 128
Фишер Р. (Fisher) 4, 81, 82, 85
фогт К. (Vogt) 181
Фрешкоп С. (Frechkop) 65
Фрик Г. (Freke) 184, 206
Фрис Э. (Fries) 31, 148, 213
Хакен Г. (Haken) 238
Хантер Дж. (Hunter) 22, 23
Харрисон Дж. (Harrison) 181
Харченко П. А. 252
Хесин Р. Б. 105, 172
Хогг Дж. (Hogg) 148
Холтон Дж. (Holton) 9, 198, 253
Хэйл М. (Hale) 28, 231
Цезальпин A. (Cesalpinus) 18, 21, 24
Целищев В. В. 39
Цетлин М. Л. 93, 96, 102
Циммерман В. (Zimmermann) 165
Циолковский К. Э. 200
Ципф Дж. (Zypf) 86, 87
Чайковский Ю. В. 6, 12, 13, 16, 25, 49,
57, 60, 61, 63, 66, 67, 74, 79, 81, 82, 84,
87, 89, 91, 94-96, 101-103, 105, 106,
108, 109, 120, 128, 137, 146, 156, 158,
159, 164, 165, 168, 172, 173, 178, 183,
191, 195, 197, 198, 205, 208, 209, 216,
217, 226, 227, 230, 241, 244
268
Чаянов А. В. 247
Чебанов С. В. 4, 124, 148
Численно Л. Л. 85, 91, 92
Чудинов Э. М. 119, 252
Чураев Р. Н. 93
Шапошников Г. X. 109, 175, 215, 250
Шеннон К. (Shannon) 220
Шеппард П. (Sheppard) 135
Шимкевич В. М. 37
Шиндевольф О. (Schindewolf) 169, 175,
214, 216, 251
Шишкин М. А. 106, 118, 119
Шлегель Г. (Schlegel) 150, 183
Шмальгаузен И. И. 58, 77, 107, 120, 121,
133, 135
Шпенглер О. (Spengler) 240
Шпрангер Э, (Spranger) 60
Шрейдер Ю. А. 72, 141, 220
Шуков В. А. 5, 224
Шультер М. (Schulter) 157
Шустер П. (Schuster) 194, 195
Эвклид 228
Эгертон Ф. (Egerton) 11
Эдельман Дж. (Edelman) 196, 225
Эйген М. (Eigen) 194, 195
Эйнштейн A. (Einstein) 228
Эмпедокл 9
Эразм /Роттердамский/ 43
Юдин А. Л. 105, 107
Adams W. I. 89
Ainsworth G. С. 19, 148
Alt F. W. 196
Bacq Z. M. 109
Bhaud Y. 146, 193
Bogorad L. 235
Bowler P. J. 167
Broadwater S. 229
Burkhardt R. W. 167
Cedergren R. 210
Chamberlin T. C. 249
Clark J. M. 196
Clutton-Brock J. 45, 48, 65
Cox E. C. 95; Costas E. 146
Dixon D. 248
Engen S. 87
Fritz L. 146
Gale B. G. 230
Gavrila L. 146
Ghiselin M. T. 252
Glaser H.-J. 211
Goldsmith E.. 247
Юл Г. (Yule) 85, 86, 172
Юнг И. (Jung/Jungius) 22, 33
Яблоков А. В. 174, 182
Янч Э. (Jantsch) 221, 227, 237, 239
— антропный принцип 210, 238
— биосфера как организм 13, 214
— время системы 228
— «встреча» 210, 220, 238
— динамический взгляд на мир 202, 211
— «затравка» развития 209, 211
— и Дарвин 214
— консервативная и динамическая па-
мять 210
— консервативность как фактор разви-
тия 212, 234
— коэволюция 217, 221, 225, 235
— — уровни 239
— мягкое самоупорндочение 211
— наблюдатель сторонний и включен-
ный в систему 202
-- отбор и макрофлуктуации 214
— открытая цель 226
— понятийный анализ 220
— приспособление, экстремаль 209
— самопорождение, три уровня 211
— симметрия 91, 210, 211
- фазовое пространство 209, 221
— эмерджентная парадигма 236
— эпигенеалогический принцип 210
Ярмолинская Е. Ш. 4, 227
Goodwin В. С. 230
Grasse Р.-Р. 163, 171
Hartley В. S. 95
Lake J. А. 147, 148
Lewin R. 200
Martin С. Р. 107, 232
Morowitz Н. J. 237
Morris Н. 181
Nagi W. 211
Norris К. S. 133
Oakley 193
Raven Р. Н. 175
Sato Т. 122
Schwartz J. В. 164, 251
Simon-Bichar-Breaud J. 229
Soyer-Gobillard M.-O. 146, 193
Steenis G. G. G. J. 120, 133
Stevens P. F. 18
Trincher K. 201
Tripodi G. 229
Withers A. W. 229
Young R. M. 230
ОГЛАВЛЕНИЕ
Предисловие ........................................ 3
Часть 1. УПОРЯДОЧЕННОСТЬ РАЗНООБРАЗИЯ............... 6
Глава 1. Введение............................................ 6
1.1. Феномен разнообразия............................... 6
1.2. Проблема разнообразия. Познавательные модели .... 9
1.3. Систематика. От первых реестров до Линнея и Жюссье 17
1.4. Морфология. От Везалия до Бэра и Геккеля.......... 21
1.5. Комбинативность. От Парацельса до школы Вавилова . . 27
Глава 2. Разнообразие, ряды и тенденции..................... 32
2.1. О биологической грамматике........................ 33
2.2. Мероно-таксономический анализ (типология по Мейену) 38
2.3. Ядро и периферия.................................. 43
2.4. Параллельные ряды................................. 50
2.5. Другие тенденции.......................... ... . 57
2.6. Гомология и аналогия взаимодополнительны.......... 63
Глава 3. Разнообразие, случайность и отбор.................. 66
3.1. Случайность и закономерность .......... 66
3.2. Номотетико-идиографическое чередование............ 72
3.3. Статистика гауссова и гиперболическая . .............. 76
3.4. Мажорирующая модель отбора............................ 79
3.5. Гиперболические распределения Виллиса................. 84
3.6. Симметрия и экстремальность случайного................ 87
Глава 4. Разнообразие, поиск и приспособленность................ 92
4.1. Дискомфорт и генетический поиск....................... 92
4.2. Организующая роль случайности.................... . 97
4.3. Поиск как игра......................................100
4.4. Смена функционирования и наследование...............104
4.5. Что такое приспособленность?........................110
4.6. Границы приспособленности........................... 120
Глава 5. Разнообразие, норма и целостность....................126
5.1. Что такое норма.....................................126
5.2. Логика ценоза.......................................130
5.3. Норма прибыли и норма приспособленности.............134
5.4. Многомерность нормы.................................136
Глава 6. Разнообразие, симметрия и макросистема..............137
6.1. Система и макросистема..............................138
6.2. Соотношение сети и древа............................ 141
270
6.3. Диатропика клеток...................................... 144
6.4. Необходимый минимум высших таксонов.....................147
6.5. Типы метаболизма и макросистема.........................150
6.6. Симметрийная основа макросистемы........................153
6.7. Царства и межцарства. Логика построения.................158
Часть II. ЭВОЛЮЦИЯ КАК ПРЕОБРАЗОВАНИЕ РАЗНООБРАЗИЯ 162
Глава 7. Закономерности биологической эволюции.................. 162
7.1. Эволюция как физиологический процесс....................162
7.2. О факторах эволюции организмов..........................168
7.3. Преобразование разнообразия и происхождение таксонов 176
Глава 8. Диатропический анализ некоторых представлений об эво-
люции клеток................................................ 183
8.1. Связь концепций эволюции биосферы и клетки . . . 183
8.2. Жизнь до клеток.........................................186
8.3. Первые клетки.......................................... 190
8.4. Конец «центральной догмы» и новая познавательная модель 194
Глава 9. Связь идей эволюционной термодинамики и диатропики 199
9.1. Статус эволюционной термодинамики.......................199
9.2. Статистика и системность в эволюционизме................203
9.3. Понятие сродства в системном эволюционизме .... 206
9.4. Прогресс и приспособление...............................208
Глава 10. Язык и структура эволюционизма.........................212
10.1. Сквозные закономерности эволюции.......................212
10.2. Язык и понимание эволюции..............................219
10.3. К концептуальной схеме биоэволюционизма................225
Часть III. ПРИМЕНЕНИЕ ПРИНЦИПОВ ДИАТРОПИКИ 229
Глава 11, Переориентация исследовании эволюции ...... 229
11.1. Выявление тенденций..................................230
11.2. О становлении систем.................................235
Глава 12. Разнообразие и прогноз...............................240
12.1. Экологический диктат и замыкание затрат..............240
12.2. Эволюционный прогноз. Заключение.....................247
Литература.....................................................254
Именной указатель..............................................263
Научное издание
Чайковский Юрий Викторович
ЭЛЕМЕНТЫ
ЭВОЛЮЦИОННОЙ
ЦИАТРОПИКИ
Утверждено к печати
Институтом истории
естествознания и техники
Редактор В. А. Чудинов
Художник В. Н. Тикунов
Художественный редактор М. Л. Храмцов
Технический редактор Т. С. Жарикова
Корректоры Н. Б. Габасова, Р. 3. Землянская
И Б № 46448
Сдано в набор 27.02.90.
Подписано к печати 21.05.90.
Формат 60X90 '/16
Бумага офсетная № 1
Гарнитура обыкновенная
Печать офсетная
Усл. печ. л. 17. Усл. кр.отт. 17. Уч.-изд. л. 20,7
Тираж 1000 экз. Тип. зак. 4288
Цена 2 р. 80 к.
Ордена Трудового Красного Знамени
издательство «Наука»
117864 ГСП-7, Москва В-485
Профсоюзная ул., 90
2-я типография издательства «Наука»
121099, Москва, Г-99, Шубинский пер., 6.