



































































































































































































































































































































































































































































































































Похожие




Текст
НЛ.БОБРИНСКИЙ, Б.А.КУЗНЕЦОВ, А.П.КУЗЯКИН
Определитель
МЛЕКОПИТАЮЩИХ
Н.А. БОБРИНСКИМ, Б.А.КУЗНЕЦОВ, А.П.КУЗЯКИН
Определитель
МЛЕКОПИТАЮЩИХ
СССР
ПОСОБИЕ ДЛЯ СТУДЕНТОВ
ПЕДАГОГИЧЕСКИХ ИНСТИТУТОВ
И УЧИТЕЛЕЙ
ИЗДАНИЕ ВТОРОЕ,
ИСПРАВЛЕННОЕ И ДОПОЛНЕННОЕ
Под редакцией
проф. А. П. КУЗЯКИНА
екае
москва 1965
Рецензенты:
Доктор биологических наук В. Ф. Натали, доктор биологических наук
И. И. Соколов, доктор биологических наук К. К. Чапский, кандидат биоло-
гических наук И, М. Громов.
ПРЕДИСЛОВИЕ
Первое издание полного однотомного «Определителя мле-
копитающих СССР» (1944) было подготовлено авторами
в 1940 г., а в атласе карт ареалов использованы материалы,
опубликованные до 1938 г. За истекший период накопилось
большое количество новых фактов по видовому составу, си-
стематике, номенклатуре, изменчивости, распространению,
образу жизни, практическому значению млекопитающих и по
активному воздействию человека на их ареалы. При подго-
товке книги к новому изданию авторы использовали более
1500 книг и статей, опубликованных после 1938 г., не считая
мелких заметок в различных журналах и большого количе-
ства экспонатов в коллекциях как советских, так и некото-
рых зарубежных музеев.
Кроме дополнений, в новом издании введены следующие
изменения в план книги и в ее отдельные разделы.
Из оглавления выделен полный список видов млекопитаю-
щих фауны СССР и помещен в начале книги. В списке видов,
как и в тексте, уточнена латинская номенклатура и предпри-
нята первая попытка упорядочения русских названий. За
каждым видом авторы предлагают закрепить одно русское
название, состоящее не более чем из двух слов. Расположе-
ние видов и групп в списке, в основном тексте, в определи-
тельных таблицах и атласах унифицировано и посильно упо-
рядочено в филогенетическом отношении.
За полным списком видов помещен полный атлас цветных
тотальных изображений всех видов млекопитающих фауны
СССР, выполненный художниками Н. Н. Кондаковым и
О. Ф. Хлудовой.
Основная литература приведена в одном списке, помещен-
ном перед атласом ареалов. Список литературы разделен на
три части: 1) общие сводки и периодические издания по си-
стематике, распространению, териогеографии и биологии мле-
копитающих фауны СССР, 2) региональные териологические
сводки и статьи с материалами не менее чем по двум отря-
дам млекопитающих и 3) основная литература по отдельным
отрядам и видам.
1*
3
Атлас ареалов, приведенный в конце книги, расширен до
110 карт; в него включены ареалы всех наземных видов мле-
копитающих, а также части ареалов некоторых ластоногих и
китообразных, связанные с побережьями и полосой террито-
риальных вод СССР.
Разделы между авторами распределены следующим об-
разом:
Н. А. Бобринский—автор текста по отрядам Хищные,
Ластоногие, Китообразные, Копытные.
Б. А. Кузнецов переработал свой обширный раздел по
отряду Грызуны (вместе с зайцеобразными) и принял неко-
торое участие в переработке текста по отрядам Хищные, Ла-
стоногие и Копытные.
А. П. Кузякин составил полный список млекопитающих
фауны СССР, главу «Основные сведения по систематике»,
тексты по отрядам Насекомоядные и Рукокрылые, принял не-
которое участие в переработке текста по отряду Китообраз-
ные, составил зональную основу для атласа ареалов.
При подготовке определителя к новому изданию авторы
пользовались любезным содействием и помощью многих своих
коллег и учеников. Директор Зоологического музея МГУ
проф. С. Г. Соин разрешил пользоваться обширными коллек-
циями, а сотрудники музея О. Л. Россолимо, М. В. Васильева
и Е. Б. Климик неизменно оказывали большую помощь при
работе с коллекциями. В обработке материалов рукописи
наибольшее участие принимала В. И. Комкова. Много неопуб-
ликованных данных авторам представили с разрешением ис-
пользовать их А. А. Слудский, А. Г. Томилин,. В. И. Цалкин,
Б. М. Касаткин, Д. И. Бибиков, А. С. Бурделов, Р. П. Зи-
мина, Н. В. Щепотьев, Н. И. Ларина, А. П. Шведов, А. Н. Ко-
ротких, Е. И. Страутман, Г. К. Корсаков, Р. Р. Гюсян и
другие.
Выписками по распространению и составлением чернови-
ков ареалов на картах были заняты более 70 студентов трех
институтов Москвы, особенно Московского областного педа-
гогического института. Наибольшее старание проявили Э. Ко-
ренберг, А. Дмитриев, И. Соколова и другие.
Всем, любезно оказавшим содействие в подготовке книги
к новому изданию, авторы выражают глубокую благодар-
ность.
МЛЕКОПИТАЮЩИЕ ФАУНЫ СССР
(СПИСОК ВИДОВ)
А. П. Кузякин
КЛАСС МЛЕКОПИТАЮЩИЕ. CLASS MAMMALIA
ОТРЯД НАСЕКОМОЯДНЫЕ. ORDO INSECTIVORA
Семейство Ежовые. Familia
Erinaceidae
Род Ежи. Genus Erinaceus Linnaeus,
1758 ............................. 38
1. Еж обыкновенный. Erinaceus
europaeus Linnaeus, 1758 ... 40
E. e. dauricus Sundevall,
1842 >.
2. Ушастый еж. Erinaceus auritus
Gmelin, 1770 .................... 42
3. Лысый еж. Erinaceus hypome-
las Brandt, 1836 ................ 43
Семейство Выхухолевые. Familia
Desmanidae
Род Выхухоли. Genus Desmana
Giildenstaedt, 1777 ............ 44
4. Выхухоль. Desmana moschata
Linnaeus, 1758 ............... 44
Семейство Кротовые. Familia Talpidae
Подсемейство настоящих
кротовых. Subfamilia
T a 1 p i n a e
Род Кроты. Genus Talpa Linnaeus,
1758 ............................. 49
5. Малый крот. Talpa herzegovien-
sis Bolcay, 1925 ............. 50
T. h. minima Deparma, 1959.
6. Крот обыкновенный. Talpa
europaea Linnaeus, 1758 ... 50
T. e. levantis Thomas, 1906.
T. e. orientalis Ognev, 1926.
T. e. caucasica Satunin, 1908.
1 Виды, систематический ранг кото-
рых пока не вполне установлен или вы-
зывает сомнения, приведены в списке
без порядковых номеров. Из подвидов
включены в список лишь обозначав-
шиеся ранее (и в других сводках и ста-
тьях) видовыми названиями.
7. Алтайский крот. Talpa altaica
Nikolsky, 1883 ................. 52
Род Могеры. Genus Mogera Pomel,
1848 ........................... 53
8. Средняя могера. Mogera wo-
gura Temminck, 1842 .... 54
9. Большая могера. Mogera robu-
sta Nehring, 1891................54
Семейство Землеройковые. Familia
Soricidae
Род Бурозубки. Genus Sorex Lin-
naeus, 1758 ...................... 58
10. Крошечная бурозубка. Sorex
minutissimus Zimmermann,
1780 ............................ 61
11. Малая бурозубка. Sorex minu-
tus Linnaeus, 1766 .............. 62
S. m. cinereus Kerr, 1792.
12. Бухарская бурозубка. Sorex
buchariensis Ognev, 1921 . . 62
13. Темнозубая бурозубка. Sorex
daphaenodon Thomas, 1907 . 63
14. Средняя бурозубка. Sorex cae-
cutiens Laxmann, 1788 ... 63
15. Бурозубка обыкновенная. So-
rex araneus Linnaeus, 1758 . 64
S. a. asper Thomas, 1914.
S. a. vir G. Allen, 1914.
16. Когтистая бурозубка. Sorex
unguiculatus Dobson, 1890 . . 66
17. Альпийская бурозубка. Sorex
alpinus Schinz, 1837 .... 66
18. Гигантская бурозубка. Sorex
pacificus Coues, 1877 .... 66
S. p. mirabilis Ognev, 1937.
Род Куторы. Genus Neomys Каир,
1829 ............................... 67
19. Малая кутора. Neomys anoma-
lus Cabrera, 1907 .............. 67
20. Водяная кутора. Neomys fo-
diens Pennant, 1771 .... 68
Род Многозубые белозубки. Genus
Suncus Ehrenberg, 1833 .... 69
5
21. Карликовая белозубка. Suncus
etruscus Savi, 1822 .... 69
Род Белозубки. Genus Crocidura
Wagler, 1832 ...................... 69
22. Малая белозубка. Crocidura
suaveolens Pallas, 1811 . . . 73
Белохвостая белозубка. Crcci-
dura pergrisea Miller, 1913 . 74
23. Бурая белозубка. Crocidura
russula Hermann, 1780 ... 74
24. Большая белозубка. Crocidura
lasiura Dobson, 1890 .... 75
Серая белозубка. Crocidura?
lasia Thom?:s, 1906 ........... 76
25. Белобрюхая белозубка. Croci-
dura leucodon Hermann, 1780
Сибирская белозубка. Croci-
dura 1. sibirica Dukelsky, 1930 76
Род Пуюраки. Genus Diplomeso-
don Brandt, 1852 .................. 78
26. Пегий путорак. Diplomesodon
pulchellum Lichtenstein, 1823 78
ОТРЯД РУКОКРЫЛЫЕ. ORDO CHIROPTERA
ПОДОТРЯД ЛЕТУЧИХ МЫШЕИ.
SUBORDO MICROCHIROPTERA
Семейство Подковоносые. Familia
Rhinolophidae
Род Подковоносы. Genus Rhinolo-
phus Lacepede, 1799 .............. 82
1. Малый подковонос. Rhinolo-
phus hipposideros Bechstein,
1800 .......................... 85
2. Средиземный подковонос. RhL
nolophus blasii Peters, 1866 . 85
3. Южный подковонос. Rhinolo-
phus euryale Blasius, 1853 . . 86
4. Подковонос Мегели. Rhinolo-
phus mehelyi Matschie, 1901 87
5. Большой подковонос. Rhino-
lophus ferrumequinum Schre-
ber, 1774 ..................... 87
6. Бухарский подковонос. Rhino-
lophus bocharicus Kastschenko
et Akimow, 1917...............88
Семейство Гладконосые. Familia
Vespertilionidae
Род Ночницы. Genus Myotis Каир,
1829 ....................... 90
7. Длинноухая ночница. Myotis
bechsteini Kuhl, 1818 .... 92
8. Остроухая ночница. Myotis
blythi Tomes, 1857 .... 92
9. Большая ночница. Myotis
myotis Borkhausen, 1797 . . 93
10. Прудовая ночница Myotis
dasycneme Boie, 1825 ... 93
11. Водяная ночница. Myotis dau-
bentoni Kuhl, 1819...............94
12. Длиннопалая ночница. Myotis
capaccinri Bonaparte, 1837 . 94
13. Ночница Наттерера. Myotis
nattereri Kuhl, 1818 . . . . 94
14. Трехцветная ночница. Myotis
emarginatus Geoffrey, 1806 . 95
15. Малая ночница. Myotis muri-
cola Gray, 1846 ................ 95
16. Усатая ночница. Myotis mysta-
cinus Kuhl, 1819.................96
17. Длиннохвостая ночница. Myo-
tis frater G. Allen, 1923 ... 97
Род Длиннокрылы. Genus Miniop-
terus Bonaparte, 1837 .......... 97
18. Длиннокрыл обыкновенный.
Miniopterus schreibersi Kuhl,
1819...........................98
Род Ушаны. Genus Plecotus Geoff-
rey, 1818..........................98
19. Ушан. Plecotus auritus Lin-
naeus, 1758 ...................... 98
Род Широкоушки. Genus Barbastella
Gray, 1821........................100
20. Европейская широкоушка.
Barbastella barbastellus Schre-
ber, 1774 . ................. 100
21. Азиатская широкоушка. Bar-
bastella leucomelas Cretzsch-
mar, 1826 ....................... 100
Род Вечерницы. Genus Nyctalus
Bowdich, 1825 ................... 101
22. Гигантская вечерница. Nycta-
lus lasiopterus Schreber, 1780
Восточная вечерница. Nycta-
lus aviator Thomas, 1911 . . 102
23. Рыжая вечерница. Nyctalus
noctula Schreber, 1774 . . .102
24. Малая вечерница. Nyctalus
leisleri Kuhl, 1818 . . . . .103
Род Кожаны и Нетопыри. Genus
Vespertilio Linnaeus, 1758 . . .103
25. Нетопырь-карлик. Vespertilio
pipistrellus Schreber, 1775 . .107
26. Лесной нетопырь. Vespertilio
nathusii Keys et Blasius, 1839 108
27. Восточный нетопырь. Vesperti-
lio abramus Temminck, 1840 . 108
28. Средиземный нетопырь. Ves-
pertine kuhli Kuhl, 1819 . . .108
29. Кожановидный нетопырь. Ves-
pertilio savii Bonaparte, 1837 109
30. Северный кожанок. Vespertilio
nilssoni Keys, et Blasius, 1839 109
31. Кожанок Бобринского. Vesper-
tilio bobrinskii Kuzyakin, 1935 110
32. Восточный кожан. Vespertilio
superans Thomas, 1899 . . .Ill
33. Двухцветный кожан. Vesper-
tilio murinus Linnaeus, 1758 . Ill
34. Кожан Огнева. Vespertilio og-
nevi Bobrinskiy, 1918 . . .112
35. Поздний кожан. Vespertilio se-
rotinus Schreber, 1774 . . .112
Род Стрелоухи. Genus Otonycteris
Peters, 1859 ...................... 113
36. Белобрюхий стрелоух. Otonyc-
teris hemprichi Peters, 1859 . 113
Род Трубконосы. Genus Murina
Gray, 1942 ........................ 113
6
37. Большой трубконос. Murina
leucogaster Milne-Edwards,
1872 .................. .... 114
38. Малый трубконос. Murina
aurata Milne-Edwards, 1872 . 114
Семейство Бульдоговые. Familfa
Molossidae
Род Складчатогубы. Genus Tadarida
Rafinesque, 1814....................115
39. Широкоухий складчатогуб.
Tadarida teniotis Rafinesque,
1814..........................115
ОТРЯД ХИЩНЫЕ. ORDO CARNIVORA
Семейство Собачьи. Familia Canide
Семейство Кошачьи. Familia Felidae
Род Собаки. Genus Canis Linnaeus,
1758 ............................ 121
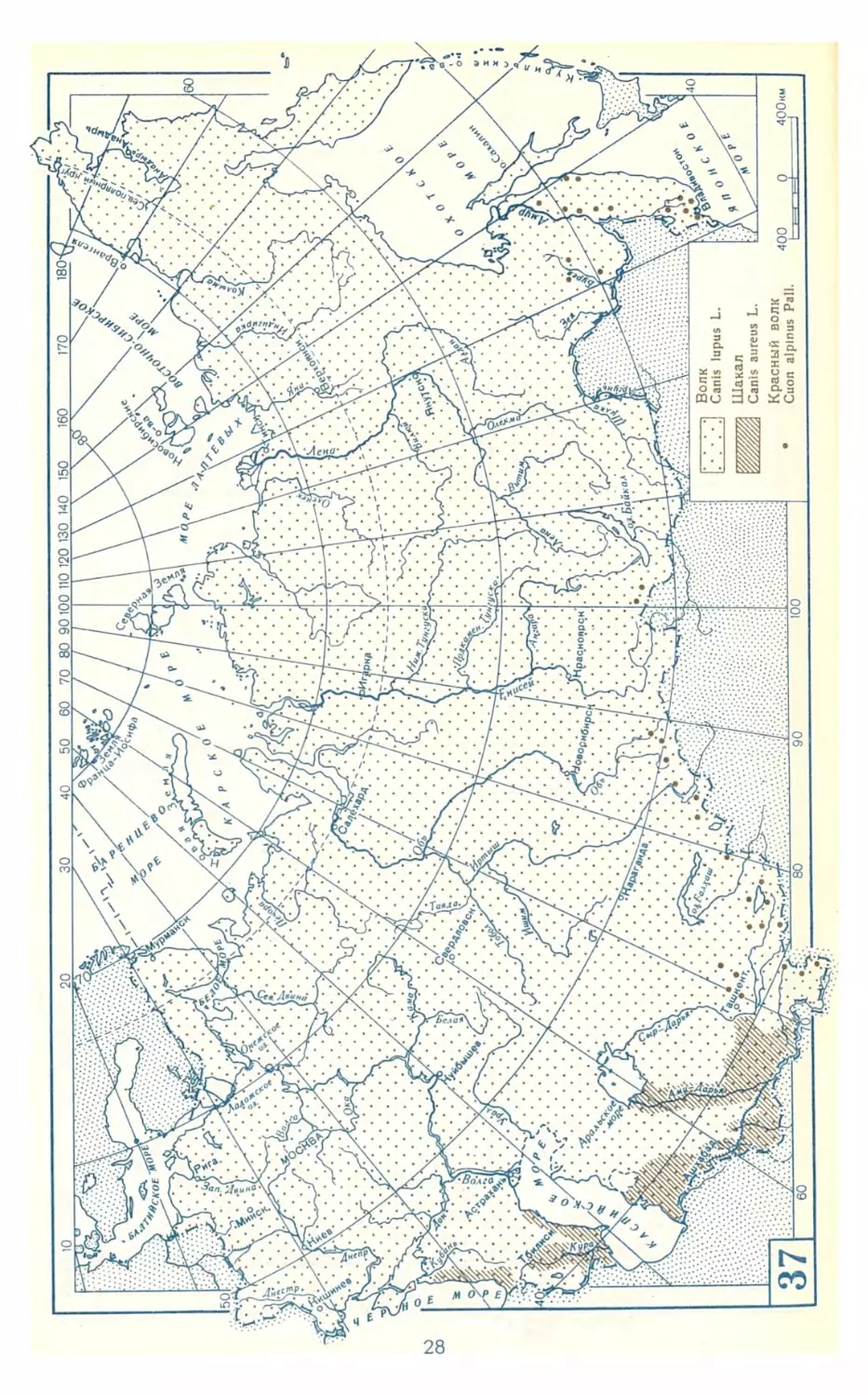
1. Волк. Canis lupus Linnaeus,
1758 ........................ 121
2. Шакал. Canis aureus Linnaeus,
1758 ........................ 122
Род Красные волки. Genus Cupn
Hodgson, 1838 ................... 123
3. Красный волк. Cuon alpinus
Pallas, 1811..................123
Род Лисицы. Genus Vulpes Oken,
1816..............................124
Подрод настоящих лисиц. Subge-
nus Vulpes Oken, 1816.............125
4. Лисица. Vulpes vulpes Linnae-
us, 1758 .....................125
5. Корсак. Vulpes corsac Linnae-
us, 1768 .....................127
6. Афганская лисица. Vulpes сапа
Blanford, 1877 ............ 127
Подрод песцов. Subgenus Alopex
Каир, 1829 .................... 127
7. Песец. Vulpes lagopus Linnae-
us, 1758 .................... 127
Род Енотовидные собаки. Genus
Nyctereutes Temminck, 1839 . .128
8. Енотовидная собака. Nyctereu-
tes procyonoides Gray, 1834 . 128
Семейство Енотовые. Familia
Procyonidae
Род Еноты. Genus Procyon Storr,
1780 ............................ 129
9. Енот-полоскун. Procyon lotor
Linnaeus, 1758 .............. 129
Семейство Медвежьи. Familia Ursidae
Род Черные медведи. Genus Sele-
narctos Heude, 1901.................130
10. Черный медведь. Selenarctos
tibetanus G. Cuvier, 1823 . . 130
Род Бурые медведи. Genus Ursus
Linnaeus, 1758 .................... 131
11. Бурый медведь. Ursus arctos
Linnaeus, 1758 ............. 131
Род Белые медведи. Genus Thalas-
sarctos Gray, 1825 ................ 132
12. Белый медведь. Thalassarctos
maritimus Phipps, 1774 . . . 132
Семейство Гиеновые. Familia Hyaenidae
Род Гиены. Genus Hyaena Brisson,
1762 ............................ 133
13. Полосатая гиена. Hyaena hya-
ena Linnaeus, 1758 ...........133
Род Гепарды. Genus Acinonyx Broo-
kes, 1828 ...................... 134
14. Гепард. Acinonyx jubatus
Schreber, 1776 .............. 135
Род Пантеры. Genus Panthera Oken,
1816.............................135
Подрод барсов. Subgenus Uncia
Gray, 1854 ...................'. . 136
15. Барс. Panthera uncia Schreber,
1776 ........................... 136
Подрод леопардов. Subgenus Pan-
thera Oken, 1816.................136
16. Леопард. Panthera pardus
Linnaeus, 1758 ................. 136
Подрод тигров. Subgenus Tigris
Oken. 1816.......................136
17. Тигр. Panthera tigris Linnaeus,
1758 ........................... 136
Род Кошки. Genus Felis Linnaeus,
1758 ...............•............137
Подрод настоящих кошек. Subgenus
Felis Linnaeus, 1758 ........... 139
18. Лесная кошка. Felis silvestris
Schreber, 1777 .............. 139
। 19. Степная кошка. Felis libyca
Forster, 1780 ............... 139
20. Бенгальская кошка. Felis ben-
galensis Kerr, 1792 .......... 140
* 21. Барханная кошка. Felis mar-
garita Lache, 1858 .............. 140
22. Манул. Felis manul Pallas,
1776 .............................. 140
23. Xayc. Felis chaus Giildenstaedt,
1776 ........................... 141
Подрод каракалов. Subgenus Caracal
Gray, 1843 ...................... 141
24. Каракал. Felis caracal Schre-
ber, 1776 ...................... 141
Подрод рысей. Subgenus Lynx Kerr,
1792 ............................. 141
25. Рысь. Felis lynx Linnaeus, 1758 141
Семейство Куньи. Familia Mustelidae
Подсемейство барсуков.
Subfamilia Melinae
Род Барсуки. Genus Meles Brisson,
1762 .......................... 146
26. Барсук. Meles meles Linnaeus,
1758 . ...................... 146
Подсемейство медоедов.
Subfamilia Mellivorinae
Род Медоеды. Genus Mellivora
Storr, 1780 ............ 147
27. Медоед. Mellivora indica Kerr,
1776 .................... 147
7
Подсемейство куньих.
Subfamilia Mustelinae
Род Росомахи. Genus Gulo Storr,
1780 . ....................... . 147
28. Росомаха. Gulo gulo Linnae-
us, 1758 ...................... . 148
Род Куницы. Genus Martes Pinel,
1792 ...............................148
Подрод харзы. Subgenus Charronia
Gray, 1865 ........................ 150
29. Харза. Martes flavigula Bod-
daert, 1785 .................... 150
Подрод куниц. Subgenus Martes Pi-
nel, 1792 .................... 150
30. Каменная куница. Martes
foina Erxleben, 1777 . . . .150
31. Лесная куница. Martes martes
Linnaeus, 1758 .............. 150
32. Соболь. Martes zibellina Lin-
naeus, 1758 .................... 151
Род Перевязки. Genus Vormela Bla-
sius, 1884 ........................ 153
33. Перевязка. Vormela peregusna
Giildenstaedt, 1770 ......... 153
Род Ласки и Хорьки. Genus Mustela
Linnaeus, 1758 . .............. 153
Подрод ласок. Subgenus Mustela
Linnaeus, 1758 ............... 156
34. Ласка. Mustela nivalis Linnae- ’
us, 1766 ....................... 156
35. Горностай. Mustela erminea
Linnaeus, 1758 .................157
36. Солонгой. ’Mustela altaica Pal-
las, 1811.......................158
37. Колонок. Mustela sibirica Pal-
las, 1773 ......................158
Подрод хорьков. Subgenus Putorius
Cuvier, 1817....................159
38. Светлый хорек. Mustela evers-
manni Lesson, 1827 .... 159
39. Черный хорек. Mustela putori-
us Linnaeus, 1758 ............. 160
Подрод норок. Subgenus Lutreola
Wagner, 1761....................160
40. Европейская норка. Mustela
lutreola Linnaeus, 1761 . . .160
41. Американская норка. Mustela
vison Brisson, 1756 ... 161
Подсемейство выдр.
Subfamilia Lutrinae
Род Выдры. Genus Lutra Brisson,
1762 .......................... 162
42. Выдра. Lutra lutra Linnaeus,
1758 .......................... 162
Род Каланы. Genus Enhydra Fle-
ming, 1822 .................... 163
43. Калан. Enhydra lutris Linnae-
us, 1758 .......................163
ОТРЯД ЛАСТОНОГИЕ. ORDO PINNIPEDIA
Семейство Котиковые. Familia Otariidae
Род Морские котики. Genus Callo-
rhinus Gray, 1859 ............... 167
1. Морской котик. Callorhinus ur-
sinus Linnaeus, 1758 .... 167
Род Сивучи. Genus Eumetopias Gill,
1866 ............................ 168
2. Сивуч. Eumetopias jubata
Schreber, 1776 ............... 168
Семейство Моржовые. Familia
Odobenidae
Род Моржи. Genus Odobenus Bris-
son, 1762 ....................... 169
3. Морж. Odobenus rosmarus
Linnaeus, 1758 ...... 169
Семейство Тюленевые. Familia Phocidae
Подсемейство тюленевых.
Subfamilia Phocinae
Род Тюлени. Genus Phoca Linnaeus,
1758 .......................... 171
Подрод гренландских тюленей. Sub-
genus Pagophoca Trouessart, 1904. 173
4. Лысун. Phoca groenlandica
Erxleben, 1777 ............... 173
Подрод крылаток. Subgenus Histrio-
phoca Gill, 1873 ................ 174
5. Крылатка. Phoca fasciata Zim-
mermann, 1783 ................ 174
Подрод нерп. Subgenus Phoca Lin-
naeus, 1758 .................... 174
6. Обыкновенный тюлень. Phoca
vitulina Linnaeus, 1758 . . . 174
7. Кольчатая нерпа. Phoca his-
pida Schreber, 1775 . ... 175
8. Каспийская нерпа. Phoca cas-
pica Gmelin, 1788 ..... 175
9. Байкальская нерпа. Phoca si-
birica Gmelin, 1788 . . . . 177
Род Морские зайцы. Genus Erigna-
thus Gill, 1866 ................ 177
10. Морской заяц. Erignathus bar-
batus Erxleben, 1777 . . . .177
Род ДлинноМордые тюлени. Genus
Halichoerus Nilsson, 1820 . . . 178
11. Длинномордый тюлень. Hali-
choerus gripus Fabricius, 1791 178
Подсемейство тюленей-
монахов. Subfamilia
Monachinae
Род Тюлени-монахи. Genus Mona-
chus Fleming, 1822 .......... 178
12. Тюлень-монах. Monachus mo-
nachus Hermann, 1779 . . .178
Подсемейство хохлачей.
Subfamilia С у s t о p h о r i n a e
Род Хохлачи. Genus Cystophora
Nilsson, 1820 ............... 179
13. Хохлач. Cystophora cristata
Erxleben, 1777 .......... 179
8
ОТРЯД КИТООБРАЗНЫЕ. ORDO CETACEA
ПОДОТРЯД ЗУБАТЫХ
КИТОВ. SUBORDO
ODONTOCETI
Семейство Дельфиновые. Familia
delphinidae
Подсемейство дельфиновых.
Subfamilia Delphininae
Род Дельфины-белобочки. Genus
Delphinus Linnaeus, 1758 .... 185
I. Дельфин-белобочка. Delphinus
delphis Linnaeus, 1-758 . . .185
Род Продельфины. Genus Stenella
Gray, 1866 ..................... 186
Полосатый продельфин. Ste-
nella caeruleoalbus Mayen, 1833 186
Род Афалины. Genus Tursiops Ger-
vais, 1855 . ..................... 186
2. Афалина. Tursiops truncatus
Montagu, 1821..................186
Род Китовидные дельфины. Genus
Lissodelphis Gloger, 1841 . . 187
Китовидный дельфин. Lisso-
delphis borealis Peale, 1848 . 187
Род Короткоголовые дельфины. Ge-
nus Lagenorhynchus Gray, 1846 . 187
Белобокий дельфин. Lageno-
rhynchus acutus Gray, 1828 . 188
3. Полосатый дельфин. Lageno-
rhynchus obliquidens Gill, 1865 188
4. Беломордый дельфин. Lageno-
rhynchus albirostris Gray, 1846 188
Род Косатки. Genus Orcinus Fitzin-
ger, I860 ................. 189
5. Косатка. Orcinus orca Linnae-
us, 1758..................... 189
Род Черные косатки. Genus Pseu-
dorca Reinchardt, 1862 ....... 189
Черная косатка. Pseudorca
crassidens Owen, 1846 . . .189
Род Гринды. Genus Globicephala
Lesson, 1828 ................. 190
6. Гринда. Globicephala melaena
Traill, 1809 ................ 190
Род Серые дельфины. Genus Gram-
pus Gray, 1828 ............... 190
Серый дельфин. Grampus gri-
seus Cuvier, 1812...........190
Род Морские свиньи. Genus Phoca-
ena G. Cuvier, 1817............191
7. Морская свинья. Phocaena
phocaena Linnaeus, 1758 . . 191
Род Белокрылые свиньи. Genus
Phocaenoides Andrews, 1911 . . . 192
8. Белокрылая свинья. Phocae-
noides dalli True, 1885 . . .192
Род Бесперые свиньи. Genus Neo-
meris Gray, 1846 .............. 192
Бесперая свинья. Neomeris
phocaenoides G. Cuvier, 1829 . 192
Подсемейство белуховых.
Subfamilia Delphinapterinae.
Род Белухи. Genus Delphinapterus
Lacepede, 1804 ............... 192
9. Белуха. Delphinapterus leucas
Pallas, 1776 .................. 192
Род Единозубы. Genus Monodon
Linnaeus, 1758 ................. 193
10. Нарвал. Monodon monoceros
Linnaeus, 1758 .............. 193
Семейство Клюворылые. Familia
Ziphiidae
Род Бутылконосы. Genus Hyperoo-
don Lacepede, 1804 ............. 194
II. Высоколобый бутылконос. Hy-
peroodon ampullatus Forster,
1770 ........................... 195
Род Плавуны. Genus Berardius Du-
vernoy, 1851.................? . . 195
12. Северный плавун. Berardius
bairdi Steineger, 1883 . . . .195
Род Клюворылы. Genus Ziphius Cu-
vier, 1823 ...................... 195
13. Клюворыл. Ziphius cavirostris
G. Cuvier, 1823 ................ 195
Род Ремнезубы. Genus Mesoplodon
Gervais, 1850 .................. 196
14. Командорский ремнезуб. Me-
soplodon steinegeri True, 1885 196
Атлантический ремнезуб. Me-
soplodon bidens Sowerby, 1804 197
Семейство Кашалотовые. Familia
Physeteridae
Род Карликовые кашалоты. Genus
Kogia Gray, 1846 ................. 198
Карликовый кашалот. Kogia
breviceps Blainville, 1838 . .198
Род Кашалоты. Genus Physeter Lin-
naeus, 1758 ........................ 198
15. Кашалот. Physeter catodon
Linnaeus, 1758 ............... 198
ПОДОТРЯД БЕЗЗУБЫХ КИТОВ.
SUBORDO MYSTACOCETI
Семейство Серокитовые.
Familia Eschrichtiidae
Род Серые киты. Genus Eschrichtius
Gray, 1864 ....................... 199
16. Серый кит. Eschrichtius gibbo-
sus Erxleben, 1777 .............. 199
Семейство Полосатиковые. Familia
Balaenopteridae
Род Полосатики. Genus Balaenop-
tera Lacepede, 1804 .............. 200
17. Малый полосатик. Balaenop-
tera acutirostrata Lacepede,
1804 ............................ 201
18. Сейвал. Balaenoptera borealis
Lesson, 1828 .................. . 202
19. Финвал. Balaenoptera physa-
lis Linnaeus, 1758 .............. 202
20. Синий кит. Balaenoptera mus-
culus Linnaeus, 1758 .... 203
Род Горбатые киты. Genus Megap-
tera Gray, 1846 .................. 203
9
21. Горбач. Megaptera novaeang-
liae Borowski, 1781 .... . . 203
Семейство Гладкокитовые.
Familia Balaenidae
Род Южные киты. Genus Euba-
laena Gray, 1864 .............. 204
22. Южный кит. Eubalaena glacia-
lis Borowski, 1781 ............. 204
Род Гренландские киты. Genus Ba-
laena Linnaeus, 1758 . . . . . 205
23. Гренландский кит. Balaena
mysticetus Linnaeus, 1758 . . 205
ОТРЯД НЕПАРНОКОПЫТНЫЕ. ORDO PERISSODACTYLA
Семейство Лошадиные. Familia Equidae
Род Лошади. Genus Equus Linnae-
us, 1758 ....................206
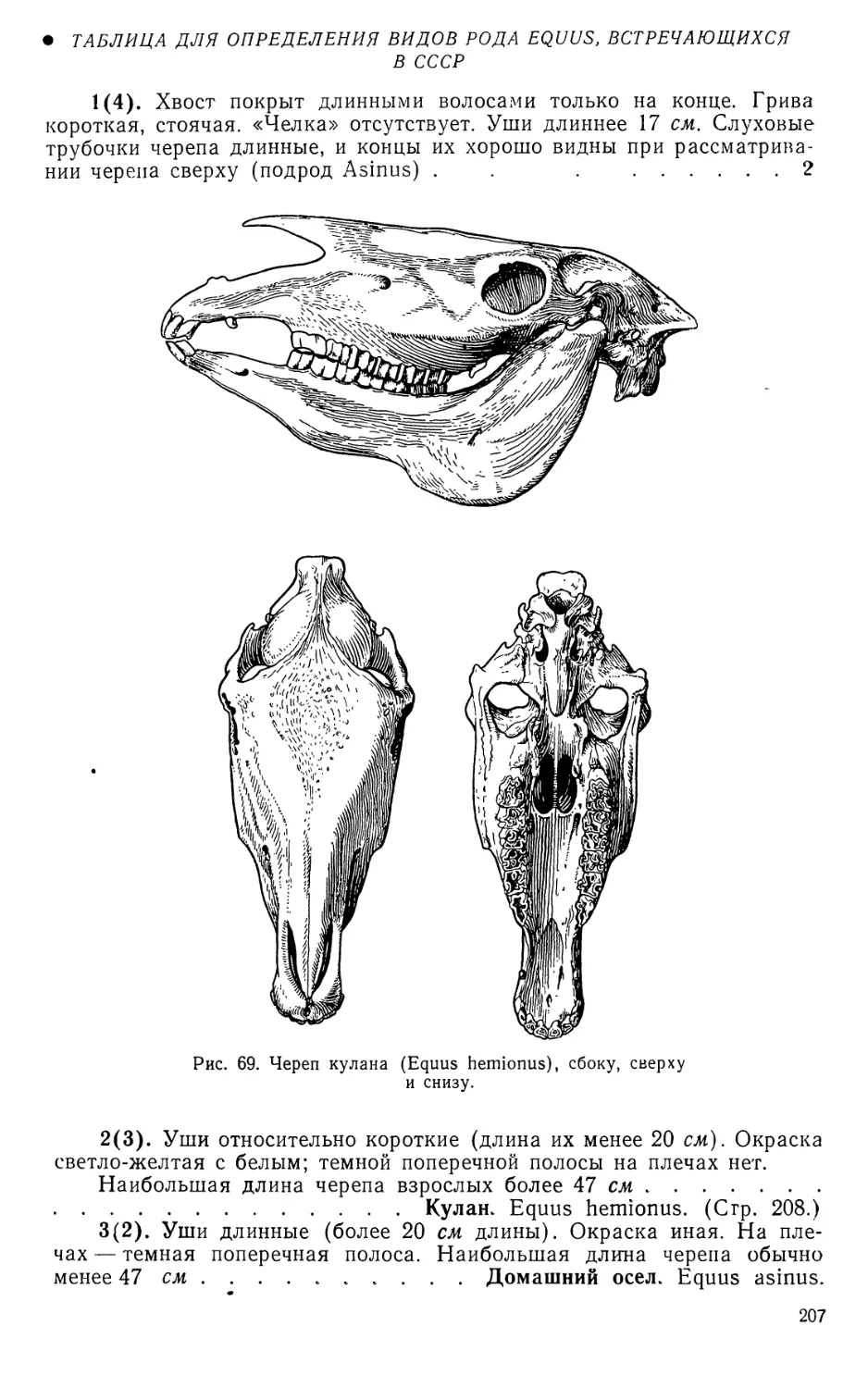
1. Кулан. Equus hemionus Pallas,
1775 ..................... 208
Домашний осел. Equus asinus
Linnaeus, 1758 ............ 208
Домашняя лошадь. Equus ca-
ballus Linnaeus, 1758 .... 208
ОТРЯД ПАРНОКОПЫТНЫЕ. ORDO ARTIODACTYLA
ПОДОТРЯД НЕЖВАЧНЫХ.
SUBORDO NONRUMINANTIA
Семейство Свиные. Familia Suidae
Род Кабаны. Genus Sus Linnaeus,
1718...............................210
1. Кабан. Sus scrofa Linnaeus,
1758 ........................... 210
ПОДОТРЯД МОЗОЛЕНОГИХ.
SUBORDO TYLOPODA
Семейство Верблюдовые. Familia
Camelidae
Род Верблюды. Genus Camelus Lin-
naeus, 1758 .............. 212
Двугорбый верблюд. Camelus
bactrianus Linnaeus, 1758 . . 212
Одногорбый верблюд. Camelus
dromedarius Linnaeus, 1758 . . 212
ПОДОТРЯД ЖВАЧНЫХ. SUBORDO
RUMINANTIA
Семейство Оленьи. Familia Cervidae
Подсемейство кабарговых.
Subfamilia Moschinae
Род Кабарги. Genus Moschus Lin-
naeus, 1758 ................ 213
2. Кабарга. Moschus moschiferus
Linnaeus, 1758 .......... 213
Подсемейство оленьих.
Subfamilia Cervinae
Род Настоящие олени. Genus Сег-
vus Linnaeus, 1758 ......... 214
Подрод ланей. Subgenus Dama
Frisch, 1775 ................ 216
3. Лань. Cervus dama Linnaeus,
1758 ..................... 216
Подрод пятнистых оленей. Subgenus
Sica Sclater, 1870 .......... 216
4. Пятнистый олень. Cervus nip-
pon Temminck, 1838 .... 216
Подрод настоящих оленей. Subgenus
Cervus Linnaeus, 1758 .... 216
5. Благородный олень. Cervus ela-
phus Linnaeus, 1758 ........... 216
Род Косули. Genus Capreolus Gray,
1821..............................218
6. Косуля. Capreolus capreolus
Linnaeus, 1758 ............. 218
Род Лоси. Genus Alces Gray, 1821
7. Лось. Alces alces Linnaeus,
1758 . ........................ 219
Род Северные олени. Genus Rangi-
fer H. Smith, 1827 .............. 220
8. Северный олень. Rangifer taran-
dus Linnaeus, 1758 ........ 220
Семейство Полорогие. Familia Bovidae
Подсемейство газелевых.
Subfamilia Gazellinae
Род Газели. Genus Gazella Blain-
ville, 1816......................222
9. Джейран. Gazella subgutturosa
Giildenstaedt, 1780 ........ 222
Род Дзерены. Genus Procapra
Hodgson, 1846 .................. 223
10. Дзерен. Gazella gutturosa
Gmelin, 1760 . ........... 223
Подсемейство козлиных
Subfamilia Caprinae
Род Сайги. Genus Saiga Gray, 1843 225
11. Сайга. Saiga tatarica Linnaeus,
1766 ........................ 225
Род Горалы. Genus Nemorhaedus H.
Smith, 1827 ................... 225
12. Горал. Nemorhaedus goral
Hardwicke, 1825 .......... 225
Род Серны. Genus Rupicapra Blain-
ville, 1816......................226
13. Серна. Rupicapra rupicapra
Linnaeus, 1758 ........... 226
Род Козлы. Genus Capra Linnaeus,
1758 ........................... 226
14. Винторогий козел. Capra falco-
neri Wagner, 1839 ........ 228
15. Безоаровый козел. Capra
aegagrus Erxleben, 1777 . . . 229
16. Центральноазиатский козел.
Capra sibirica Pallas, 1776 . 229
10
17. Кавказский тур. Capra cauca-
sica Giildenstaedt et Pallas,
1783 . ......................... 229
Дагестанский тур. Capra cylin-
dricornis Blyth, 1841 .... 230
Род Бараны. Genus Ovis Linnaeus,
1758 . ......................... 230
Подрод настоящих баранов. Subge-
nus Ovis Linnaeus, 1758 .... 230
18. Архар. Ovis ammon Linnaeus,
1758 . ...................... 230
Подрод толсторогов. Subgenus Pa-
chyceros Gromova, 1936 .... 233
19. Толсторогий баран. Ovis cana-
densis Shaw, 1804 ............ 233
Подсемейство бычьих.
Subfamilia Bovinae
Род Быки. Genus Bos Linnaeus, 1758 234
Подрод бизонов. Subgenus Bison H.
Smith, 1827 .................... 235
20. Зубр. Bos bonasus Linnaeus,
1758 ........................ 235
Род Яки. Genus Phoephagus Gray,
1843 .......................... 235
Як. Phoephagus mutus Przewal-
ski, 1883 .................... 235
Род Буйволы. Genus Bubalus H.
Smith, 1827 .................... 235
Буйвол. Bubalus bubalus Lin-
naeus, 1758 .................. 235
ОТРЯД ГРЫЗУНЫ. ORDO RODENTIA
ПОДОТРЯД ДВУПАРНОРЕЗЦОВЫХ.
SUBORDO DUPLICIDENTATA
Семейство Пищуховые. Familia
Ochotonidae
Род Пищухи. Genus Ochotona Link,
1795 .......................... 239
1. Северная пищуха. Ochotona
hyperborea Pallas, 1811 . . . 241
2. Алтайская пищуха. Ochotona
alpina Pallas, 1773 ........... 241
3. Палласова пищуха. Ochotona
pallasi Gray, 1867 ............ 242
4. Красная пищуха. Ochotona ru-
tila Severtzov, 1873 .... 242
5. Степная (малая) пищуха. Ocho-
tona pusilia Pallas, 1769 . . .242
6. Даурская пищуха. -Ochotona
daurica Pallas, 1776 .... 242
7. Рыжеватая пищуха. Ochotona
rufescens Gray, 1842 .... 243
8. Большеухая пищуха. Ochotona
roylei Ogilby, 1839 (-O. macrotis
Gunther, 1875)..................243
Семейство Заячьи. Familia Leporidae
Род Кролики. Genus Oryctolagus
Lilljeborg, 1874 .............. 244
9. Дикий кролик. Oryctolagus cu-
niculus Linnaeus, 1758 . . . 244
Род Зайцы. Genus Lepus Linnaeus,
1758 . ........................ 244
10. Дальневосточный заяц. Lepus
brachyurus Temminck, 1845 . 245
11. Заяц-беляк. Lepus timidus Lin-
naeus, 1758 .................... 246
12. Заяц-толай. Lepus tolai Pallas,
1778 ........................... 247
Заяц-песчаник. L. c. tibetanus
Waterhouse, 1841 ........... 247
13. Заяц-русак. Lepus europaeus
Pallas, 1778 ................... 247
ПОДОТРЯД ОДНОПАРНОРЕЗЦОВЫХ.
SUBORDO SIMPLICIDENTATA
Семейство Дикобразовые. Familia
Hystricidae
Род Дикобразы. Genus Hystrix Lin-
naeus, 1758 ............. 253
14. Дикобраз. Hystrix cristata
Linnaeus, 1758 .................. 253
Семейство Бобровые. Familia
Castoridae
Род Бобры. Genus Castor Linnaeus,
1758 ............. 254
15. Бобр. Castor fiber Linnaeus,
1758 ............................ 254
Семейство Нутриевые. Familia
Myocastoridae
Род Нутрии. Genus Myocastor Kerr,
1792 ........................ 255
16. Нутрия. Myocastor coypus Mo-
lina, 1782 ...................... 255
Семейство Беличьи. Familia
Sciuridae
Род Сурки. Genus Marmota Blu-
menbach, 1779 .................... 257
17. Длиннохвостый сурок. Mar-
mota caudata Jacquemont, 1844 258
18. Сурок обыкновенный. Mar-
mota bobak Muller, 1776 . . 259
Сурок Мензбира. Marmota
menzbieri Kaschkarov, 1925 . 260
Род Суслики. Genus Citellus Oken,
1816...............................260
19. Желтый суслик. Citellus maxi-
mus Pallas, 1778 ............ 261
20. Большой суслик. Citellus ma-
jor Pallas, 1779 ................ 262
21. Крапчатый суслик. Citellus sus-
licus Giildenstaedt, 1770 . . . 262
22. Малый суслик. Citellus pyg-
maeus Pallas, 1779 .............. 263
23. Серый суслик. Citellus citellus
Linnaeus, 1766 .................. 264
24. Длиннохвостый суслик. Citel-
lus undulatus Pallas, 1779 . . 264
Род Тонкопалые суслики. Genus
Spermophilopsis Blasius, 1884 . . 265
25. Тонкопалый суслик. Spermo-
philopsis leptodactylus Lichten-
stein, 1823 ..................... 265
Род Бурундуки. Genus Tamias lili-
ger, 1811...........................266
11
26. Бурундук. Tamias sibiricus
Laxmann, 1769 . . .... 266
Род Белки. Genus Sciurus Linnaeus,
1758 .............................. 266
27. Кавказская белка. Sciurus ano-
malus Giildenstaedt, 1785 . . 267
28. Белка обыкновенная. Sciurus
vulgaris Linnaeus, 1758 . . . 267
Семейство Летяжьи. Familia Pteromyidae
Род Летяги. Genus Pteromys Cuvier,
1800 . ......................... 270
29. Летяга. Pteromys volans Lin-
naeus, 1758 .................. 270
Семейство Соневые. Familia
Muscardinidae
Род Сони-полчки. Genus Glis Bris-
son, 1762 ...................... 272
30. Соня-полчок. Glis glis Linnae-
us, 1766 ......................272
Род Лесные сони. Genus Dryomys
Thomas, 1906 . ................ 273
31. Лесная соня. Dryomys nitedula
Pallas, 1779 .............. 273
Род Садовые сони. Genus Eliomys
Wagner, 1840 ................... 274
32. Садовая соня. Eliomys querci-
nus Linnaeus, 1766 ........... 274
Род Орешниковые сони. Genus
Muscardinus Каир, 1829 .... 274
33. Орешниковая соня. Miiscardi-
nus avellanarius Linnaeus, 1758 274
Род Мышевидные сони. Genus Myo-
mimus Ognev, 1924 .............. 274
34. Мышевидная соня. Myomimus
personatus Ognev, 1924 . . . 275
Семейство Селевиниевые. Familia
Seleviniidae
Род Селевинии. Genus Selevinia
Belosliidov et Bazhanov, 1938 . . 275
35. Селевиния. Selevinia betpakda-
lensis Belosl. et Bazhanov, 1938 275
Семейство Тушканчпковые. Familia
Dipodidae
Подсемейство
пятипалых тушканчиков.
Subfamilia Allactaginae
Род Пятипалые тушканчики. Genus
Allactaga Cuvier, 1836 ......... 277
36. Большой тушканчик. Allactaga
major Kerr, 1792 ..... 279
37. Тушканчик Северцова. Allac-
taga severtzovi Vinogradov,
1925 ........................... 279
38. Тушканчик-прыгун. Allactaga
sibirica Forster, 1778 .... 279
39. Горный тушканчик. Allactaga
williamsi Thomas, 1897 . . . 280
40. Малый тушканчик. Allactaga
elater Lichtenstein, 1825 . . . 280
41. Тушканчик Бобринского. Al-
lactaga bobrinskii Kolesnikov,
1937 ........................... 281
Род Земляные зайчики. Genus Alac-
tagulus Nehring, 1897 ........... 281
42. Земляной зайчик. Alactagulus
pygmaeus Pallas, 1779 . . . 281
Род Толстохвостые тушканчики. Ge-
nus Pygeretmus Gloger, 1841 . .281
43. Тушканчик Житкова. Pygeret-
mus zhitkovi Kuznetzov, 1930 282
44. Толстохвостый тушканчик. Py-
geretmus platyurus Lichten-
stein, 1823 ................... 282
Тушканчик Виноградова. Py-
geretmus vinogradovi Voront-
zov, 1958 .................. 282
Подсемейство
трехпалых тушканчиков.
Subfamilia Dipodinae
Род Емуранчики. Genus Stylodipus
G. Allen, 1925 ................ 283
45. Емуранчик обыкновенный. Sty-
lodipus telum Lichtenstein, 1823 283
Род Мохноногие тушканчики. Ge-
nus Dipus Zimmermann, 1780 . . 284
46. Мохноногий тушканчик. Dipus
sagitta Pallas, 1773 .... 284
Род Гребнепалые тушканчики. Ge-
nus Paradipus Vinogradov, 1930 . 284
47. Гребнепалый тушканчик. Para-
dipus ctenodactylus Vinogra-
dov, 1929 ...................... 284
P о.д Песчаные тушканчики. Genus
Jaculus Erxleben, 1777 ........ 285
48. Туркменский тушканчик. Jacu-
lus turkmenicus Vinogradov et
Bondar, 1949 ............... 285
49- Тушканчик Лихтенштейна. Ja-
culus lichtensteini Vinogradov,
1927 ........................... 285
Подсемейство
карликовых тушканчиков
Subfamilia С a г d i о с г a n i i n a e
Род Пятипалые карликовые туш-
канчики. Genus Cardiocranius Sa-
tunin, 1902 ...................... 286
50. Пятипалый карликовый туш-
канчик. Cardiocranius parado-
xus Satunin, 1903 ................ 286
Род Трехпалые карликовые туш-
канчики. Genus Salpingotus Vino-
gradov, 1924 ..................... 286
51. Трехпалый карликовый туш-
канчик. Salpingotus crassi-
cauda Vinogradov, 1924 . . . 286
Семейство Мышовковые. Familia
Zapodidae
Род Мышовки. Genus Sicista Gray,
1827 ........................ 287
52. Степная мышовка. Sicista sub-
tilis Pallas, 1773 ....... 288
53. Лесная мышовка. Sicista betu-
lina Pallas, 1779 ............ 288
54. Одноцветная мышовка. Sicista
concolor Buchner, 1892 . . . 289
12
Кавказская мышовка. Sicista
caucasica Vinogradov, 1925 . 289
Рыжая алтайская мышовка.
Sicista napaea Hollister, 1912 289
Серая алтайская мышовка. Si-
cista pseudonapaea Е. Straut-
man, 1949 289
Семейство Мышиные. Familia
Muridae
Подсемейство мышей.
Subfamilia Murinae
род Домовые мыши. Genus Mus
Linnaeus, 1758 ................. 292
55. Домовая мышь. Mus musculus
Linnaeus, 1758 ............ 292
Род Мыши-малютки. Genus Micro-
mys Dehne, 1841 ................ 293
56. Мышь-малютка. Micromys mi-
nutus Pallas, 1771 ........ 293
Род Лесные и Полевые мыши. Ge-
nus Apodemus Каир, 1829 . . . 294
57. Полевая мышь. Apodemus ag-
rarius Pallas, 1771 ....... 295
58. Восточноазиатская мышь. Apo-
demus speciosus Temminck,
1845 .......................... 295
59. Лесная мышь. Apodemus syl-
vaticus Linnaeus, 1758 . . . 296
60. Желтогорлая мышь. Apodemus
flavicollis Melchior, 1834 . . 296
61. Горная мышь. Apodemus mys-
tacinus Danford et Alston, 1877 297
Род Крысы. Genus Rattus Fischer,
1803 . ......................... 297
62. Черная крыса. Rattus rattus
Linnaeus, 1758 ............ 298
63. Туркестанская крыса. Rattus
rattoides Hodgson, 1845 . . 299
64. Пасюк. Rattus norvegicus Ber-
kenhout, 1769 ................. 299
Род Пластинчатозубые крысы. Ge-
nus Nesokia Gray, 1842 .... 299
65. Пластинчатозубая крыса. Ne-
sokia indica Gray et Hardwicke,
1832 .......................... 300
Подсемейство песчанок.
Subfamilia Gerbillinae
Род Малые песчанки. Genus Merio-
nes liliger, 1811................300
66. Персидская песчанка. Merio-
nes persicus Blanford, 1875 . 303
67. Когтистая песчанка. Meriones
unguiculatus Milne-Edwards,
1867 .......................... 303
68. Полуденная песчанка. Merio-
nes meridianus Pallas, 1773 . 303
69. Краснохвостая песчанка. Me-
riones libycus Lichtenstein, 1823 304
70. Песчанка Виноградова. Merio-
nes vinogradovi Heptner, 1931 304
71. Песчанка Зарудного. Meriones
zarudnii Heptner, 1937 . . . 305
72. Малоазиатская песчанка. Me-
riones blackleri Thomas, 1903 305
73. Гребенчуковая песчанка. Me-
riones tamariscinus Pallas, 1773 305
Род Большие песчанки. Genus
Rhombomys Wagner, 1841 . . . 306
74. Большая песчанка. Rhombo-
mys opimus Lichtenstein, 1823 306
Подсемейство хомяков.
Subfamilia Cricetinae
Род Мышевидные хомячки. Genus
Calomyscus Thomas, 1905 . . . 307
75. Мышевидный хомячок. Calo-
myscus bailwardi Thomas, 1905 307
Род Пестрые хомяки. Genus Crice-
tus Leske, 1779 ................ 307
Подрод обыкновенных хомяков.
Subgenus Cricetus Leske, 1779 . . 308
76. Хомяк обыкновенный. Cricetus
cricetus Linnaeus, 1758 . . 308
Подрод средних хомяков. Subgenus
Mesocricetus Nehring, 1898 . . . 309
77. Предкавказский хомяк. Crice-
tus raddei Nehring, 1894 . . 309
78. Переднеазиатский хомяк. Cri-
cetus auratus Waterhouse, 1839 309
Род Мохноногие хомячки. Genus
Phodopus Miller, 1910.........309
79. Джунгарский хомячок. Pho-
dopus sungorus Pallas, 1773 . 310
80. Хомячок Роборовского. Phodo-
pus roborovskii Satunin, 1903 310
Род Хомячки. Genus Cricetulus
Milne-Edwards, 1867 ............. 310
Подрод короткохвостых хомячков.
Subgenus Allocricetulus Argyro-
pulo, 1933 .................... 311
81. Хомячок Эверсманна. Cricetu-
lus eversmanni Brandt, 1859 . 311
Подрод серых хомячков. Subgenus
Cricetulus Milne-Edwards, 1867 . 311
82. Даурский хомячок. Cricetulus
barabensis Pallas, 1773 . . .311
83. Длиннохвостый хомячок. Cri-
cetulus longicaudatus Milne-Ed-
wards, 1867 ................ 312
84. Серый хомячок. Cricetulus
migratorius Pallas, 1773 . . . 312
Подрод крысовидных хомяков. Sub-
genus Tscherskia Ognev, 1914 . . 313
85. Крысовидный хомяк. Cricetu-
lus triton Winton, 1899 . . . 313
Подсемейство полевок.
Subfamilia Microtinae
Род Ондатры. Genus Ondatra Link,
1795 ........................... 316
86. Ондатра. Ondatra zibethicus
Linnaeus, 1766 .............. 316
Род Водяные крысы. Genus Arvi-
cola Lacepede, 1799 ............. 316
87. Водяная крыса. Arvicola ter-
restris Linnaeus, 1758 . . . 317
Род Серые полевки. Genus Micro-
tus Schrenk, 1798 ............... 318
Подрод снежных полевок. Subgenus
Chionomys Miller, 1908 .... 320
13
88. Полевка Роберта. Microtus
roberti Thomas, 1906 .... 320
89. Снежная полевка. Microtus ni-
valis Martins, 1842 .... 321
Подрод серых полевок. Subgenus
Microtus Schrenk, 1798 .... 321
90. Полевка-экономка. Microtus
oeconomus Pallas, 1776 . . 323
91. Темная полевка. Microtus ag-
restis Linnaeus, 1761 .... 325
92. Обыкновенная полевка. Micro-
tus arvalis Pallas, 1779 . . . 325
93. Полевка Миддендорфа. Micro-
tus middendorfi Poliakov, 1881 326
94. Северосибирская полевка. Mic-
rotus hyperboreus Vinogradov,
1934 . ................ .327
95. Общественная полевка. Micro-
tus socialis Pallas, 1773 . . . 327
Подрод афганских полевок. Subge-
nus Blanfordimys Argyropulo, 1933 327
96. Афганская полевка. Microtus
afghanus Thomas, 1912 . . . 328
Подрод монгольских полевок.
Subgenus Lasiopodomys Lataste,
1887 ..... .................... 328
97. Полевка Брандта. Microtus
brandtii Radde, 1861 .... 328
98. Китайская полевка. Microtus
mandarinus Milne-Edwards,
1871........................... 329
Подрод арчевых полевок. Subgenus
Phaiomys Blyth, 1863 ............ 329
99. Арчевая полевка. Microtus
juldaschi Severtzov, 1879 . . 329
Подрод узкочерепных полевок. Sub-
genus Stenocranius Kastschenko,
1901............................. 329
100. Узкочерепная полевка. Micro-
tus gregalis Pallas, 1779 . . 330
Подрод кустарниковых полевок.
Subgenus Micrurus Major, 1877 . 330
101. Кустарниковая полевка. Mic-
rotus subterraneus de Selys
Longchamps, 1835 .... 331
Род Лесные полевки. Genus Cleth-
rionomys Tilesius, 1850 .... 331
102. Красно-серая полевка. Cleth-
rionomys rufocanus Sunde-
vall, 1846 ...................... 332
103. Рыжая полевка. Clethriono-
mys glareolus Schreber, 1780
Cl. g. frater Thomas, 1908 . 333
104. Красная полевка. Clethriono-
mys rutilus Pallas, 1779 . . 334
Род Каменные полевки. Genus Alti-
cola Blanford, 1881............ 335
Подрод плоскочерепных полевок.
Subgenus Platycranius Kastschen-
ko, 1901 .......................335
105. Плоскочерепная полевка. Al-
ticola strelzovi Kastschenko,
1900 .......................... 335
Подрод каменных полевок. Subge-
nus Alticola Blanford, 1881 . . . 336
106. Серебристая полевка. Alticola
roylei Gray, 1842 ......... 336
107. Гоби — Алтайская полевка.
Alticola barakshin Bannikov,
1954 ...................... 337
108. Большеухая полевка. Altico-
la macrotis Radde, 1862 . . 337
Род Лесные лемминги. Genus Myo-
pus Miller, 1910.................337
109. Лесной лемминг. Myopus П
schisticolor Lilljeborg, 1844 . 337
Род Настоящие лемминги. Genus
Lemmus Link, 1795 .............. 338
ПО. Амурский лемминг. Lemmus
amurensis Vinogradov, 1924 . 338
111. Сибирский лемминг. Lemmus
sibiricus Kerr, 1792 .... 338
112. Норвежский лемминг. Lem-
mus lemmus Linnaeus, 1758 339
Род Копытные лемминги. Genus
Dicrostonyx Gloger, 1841 . . . . 339
113. Копытный лемминг. Dicrosto-
nyx torquatus Pallas, 1779 . 339
Род Пеструшки. Genus Lagurus
Gloger, 1841...................... 340
114. Степная пеструшка. Lagurus
lagurus Pallas, 1773 .... 340
Желтая пеструшка. Lagurus
luteus Eversmann, 1840 . . 341
Род Прометеевы полевки. Genus
Prometheomys Satunin, 1901 . . 341
115. Прометеева полевка. Prome-
tlieomys schaposchnikovi Satu-
nin, 1901.................. 341
Род Слепушонки. Genus Ellobius
Fischer, 1814......................341
116. Слепушонка обыкновенная.
Ellobius talpinus Pallas, 1770 342
117. Афганская слепушонка. Ello-
bius fuscocapillus Blyth, 1843 343
Подсемейство ц о к о р о в.
Subfamilia М у о s р а 1 а с i п а е
Род Цокоры. Genus Myospalax Lax-
mann, 1769 .............. 344
118. Цокор обыкновенный. Myos-
palax myospalax Laxmann,
1773 ..................... 344
Семейство Слепышовые. Familia
Spalacidae
Род Слепыши. Genus Spalax Gtil-
denstaedt, 1770 ................. 345
119. Малый слепыш. Spalax leuco-
d-on Nordmann, 1840 . . . 345
120. Слепыш обыкновенный. Spa-
lax microphthalmus Giilden-
staedt, 1770 ................. 346
121. Гигантский слепыш. Spalax
giganteus Nehring, 1898 . . 346
ЦВЕТНЫЕ
ТАБЛИЦЫ
ТАБЛИЦА 1
Ежи: 1-3—обыкновенный (Erinaceus europaeus: Е. е. europaeus, Е. е. roumanicus,
Е. е. dauricus), 4-5— ушастый (Е. auritus: Е. a. auritus, Е. a. albulus), 6-7 — лысый
(Е. hypomelas: светлая и темная морфы).
1
ТАБЛИЦА II
1 — выхухоль (Desmana moschata). Кроты: 2— малый (Talpa Hercegoviensis),
3 — обыкновенный (T. europaea), 4 — алтайский (Т. altaica). Могеры. 5 — средняя
(Mogera wogura); 6 — большая (М. robusta).
1
2
ТАБЛИЦА III
Бурозубки: 1 — крошечная (Sorex minutissimus), 2— малая (S. minutus), 3— тем-
нозубая (S. daphaenodon), 4-5 — средняя (S. caecutiens), 6-8— обыкновенная (S. ara-
neus), 9 — когтистая (S. unguiculatus), 10 — альпийская (S. alpinus), 11— гигантская
(S. pacificus). Куторы: 12 — малая (Neomys anomalus), 13-14 — водяная (N. fodiens).
1
2
ТАБЛИЦА IV
1 — карликовая белозубка (Suncus etruscus). Белозубки: 2-3— малая (Crocidura
suaveolens), 4 — белохвостая (Сг. pergrisea), 5 — бурая (Сг. russula), 6 — большая
(Сг. lasiura), 7-9— белобрюхая (Сг. leucodon), 10— сибирская (Сг. 1. sibirica), 11—
пегий путорак (Diplomesodon pulchellum).
ТАБЛИЦА V
Подковоносы: 1-3 — малый (Rhinolophus hipposideros), 4 — средиземный (Rh.
blasii), 5— южный (Rh. euryale), 6 — Мегели (Rh. mehelyi), 7-9 — большой (Rh. fer-
rumequinum), 10 — бухарский (Rh. bocharicus).
ТАБЛИЦА VI
Ночницы: 1—длинноухая (Myotis bechsteini), 2 — остроухая (М. blythi), 3—боль-
шая (М. myotis), 4-5 — прудовая (М. dasycneme), 6 — водяная (М. daubentoni), 7 —
длиннопалая (М. capaccinii), 8 — Наттерера (М. nattereri), 9 — трехцветная (М. етаг-
ginatus), 10 — малая (М. muricola), 11-12— усатая (М. mystacinus), 13 — длиннохво-
стая (М. frater), 14 — длиннокрыл обыкновенный (Miniopterus schreibersi).
ТАБЛИЦА VII
1-2 — ушан (Plecotus auritus). Широкоушки: 3— европейская (Barbastella bar-
bastellus), 4 — азиатская (В. leucomelas). Вечерницы: 5— гигантская (Nyctalus
lasiopterus), 6— рыжая (N. noctula), 7 — малая (N. leisleri). Нетопыри: 8-9 —
карлик (Vespertilio pipistrellus), 10 — лесной (V. nathusii), 11 — средиземный (V. kuhli),
12-13 — кожановидный (V. savii). Кожанки: 14 — северный (V. nilssoni), 15— Боб-
ринского (V. bobrinskii),
1
3
ТАБЛИЦА VIII
Кожаны: 1-2— двухцветный (Vespertilio murinus), 3 — восточный (V. superans),
4 — Огнева (V. cgnevi), 5-6 — поздний (V. serotinus), 7 — белобрюхий стрелоух (Oto-
nycteris hemprichi). Трубконосы: 8-9— большой (Murina leucogaster), 10 — малый
(М. aurata), // — широкоухий складчатогуб (Tadarida teniotis).
ТАБЛИЦА IX
/ — волк (Canis lupus): 2 — шакал (С. aureus); 5 —красный волк (Cuon alpinus);
4-6— лисица (Vulpes vulpes); 7 — корсак (V. corsac); 8 — афганская лисица (V. сапа);
9-10 — песец (V. lagopus).
ТАБЛИЦА X
/—енотовидная собака (Nyctereutes procyonoides), 2 — енот-полоскун (Procyon lotor).
Медведи: 3— черный (Selenarctos tibetanus), 4 — бурый (Ursus arctos), 5 — белый
(Thalassarctos maritimus); 6 — полосатая гиена (Hyaena hyaena).
ТАБЛИЦА XI
1 — гепард (Acinonyx jubatus), 2— тигр (Panthera tigris), 3 — барс (P. uncia), 4 —
Леопард (P. pardus). Кошки: 5 — лесная (Felis silvestris), 6 — степная (F. libyca),
7 — бенгальская (F. bengalensis), 8— барханная (F. margarita),
ТАБЛИЦА ХП
1 — манул (Felis manul); 2—хаус (F. chaus); 3 — каракал (F. caracal); 4 — рысь (F.
lynx); 5-7 — барсук (Meles meles); 8— медоед (Mellivora indica); 9—росомаха (Gulo
gulo).
1— харза (Martes flavigula). Куницы: 2—каменная (М. foina), 3— лесная (М. mar-
tes); 4-5— соболь (М. zibellina; уральский и баргузинский); 6 — перевязка (Vormela
peregusna); 7-10— ласка (Mustela nivalis); 11-12 — горностай (М. erminea).
ТАБЛИЦА XIV
1 — солонгой (Mustela altaica); 2— колонок (М. sibirica). Хорьки: 3— светлый
(М. eversmanni), 4 — черный (М. putorius). Норки: 5 — европейская (М. lutreola),
6 — американская (М. vison); 7 —выдра (Lutra lutra); 8 — калан (Enhydra lutris).
ТАБЛИЦА XV
1-3— морской котик (Callorhinus ursinus); 4 — сивуч (Eumetopias jubata); 5-6— морж
(Odobenus rosmarus).
ТАБЛИЦА XVI
1-4 — лысун (Phoca groenlandica); 5 — крылатка ((Ph. fasciata); 6-7— тюлень обыкно-
венный (Ph. vitulina). Нерпы: 8 — кольчатая (Ph. hispida), 9 — каспийская (Ph. cas-
pica), 10 — байкальская (Ph. sibirica); 11 — морской заяц (Erignathus barbatus); 12 —
длинномордый тюлень (Halichoerus gripus), 13 — тюлень-монах (Monachus monachus),
14 — хохлач (Cystophora cristata).
2
ТАБЛИЦА XVII
/ — дельфин-белобочка (Delphinus delphis); 2— афалина (Tursiops truncatus). Дель-
фины: 3— полосатый (Lagenorhynchus obliquidens), 4 — беломордый (L. albirostris);
5 —косатка (Orcinus orca); 6 — гринда (Globicephala melaena). Свиньи: 7 — морская
(Phocaena phocaena), 8 — белокрылая (Phocaenoides dalli); 9 — белуха (Delphinapterus
leucas); 10 — нарвал (Monodon monoceros); 11— высоколобый бутылконос (Hyperoodon
•unpiillaius); 12— северный плавун (Berardius bairdi); 13— клюворыл (Ziphius cavi
rostris); 14 — командорский ремнезуб (Mesoplodon steinegeri).
1
ТАБЛИЦА XVIII
1 — кашалот (Physeter catodon); 2 — серый кит (Eschrichtius gibbosus). Полоса-
тики: 3 — малый (Balaenoptera acutirostrata), 4 — сейвал (В. borealis), 5 — финвал
(В. physalis), 6 — синий (В. musculus), 7 — горбач (Megaptera novaeangliae). Киты:
8— южный (Eubalaena glacialis), 9— гренландский (Balaena mysticetus).
ТАБЛИЦА XIX
/ — кулан (Equus hemionus); 2-4— кабан (Sus scrofa). Верблюды: 5 — двугорбый
(Camelus bactrianus), 6 — одногорбый (C. dromedarius); 7-8— кабарга (Moschus mo-
schiferus).
ТАБЛИЦА XX
Олени: 1 — лань (Cervus dama), 2-3— пятнистый (С. nippon), 4-5— благородный
(С. elaphus); 6-7— косуля (Capreolus capreolus); 8-9 — лось (Alces alces); 10 — север-
ный олень (Rangifer tarandus).
1-2— джейран (Gazella subgutturosa); 3— дзерен (P. gutturosa); 4-5— сайга (Saiga
tatarica); 6 — горал (Nemorhaedus goral); 7 — серна (Rupicapra rupicapra). Козлы:
8 — винторогий (Capra falconeri), 9 — безоаровый (C. aegagrus), 10 — центральноази-
атский (C. sibirica). Туры: 11 — кавказский (C. caucasica), 12 — дагестанский (С. су-
I indricornis).
ТАБЛИЦА XXII
1-2 — архар (Ovis ammon). Муфлоны: 3— азиатский (О. orientalis), 4 — европей-
ский (О. musimon); 5 — толсторогий баран (О. canadensis); 6— зубр (Bos bonasus);
7 — домашний як (Р. grunniens); 8 — домашний буйвол (Buballus buballus).
4
5
ТАБЛИЦА XXIII
Пищухи: 1-3— северная (Ochotona hyperborea), 4-5—алтайская (О. alpina), 6 —
Палласова (О. pallasi), 7 — красная (О. rutila), 8— степная, или малая (О. pusilia),
9 — даурская (О. daurica), 10 — рыжеватая (О. rufescens), 11 — большеухая
(О. macrotis).
2
ТАБЛИЦА XXIV
1 — дикий кролик (Oryctolagus cuniculus). Зайцы: 2 — дальневосточный (Lepus
brachyurus), 3-4 — беляк (L. timidus), 5-6— толай (L. tolai, L. t. bucharensis), 7-9— ру-
сак (L. europaeus).
2
ТАБЛИЦА XXV
Сурки: 1—длиннохвостый (Marmota caudata), 2-5— обыкновенный (М. bobak),
6 — Мензбира (М.? menzbieri); 7 — нутрия (Myocastor coypus); 8— дикобраз (Hystrix
cristata); 9 — бобр (Castor fiber).
ТАБЛИЦА XXVI
Суслики: 1 — желтый (Citellus maximus), 2 — большой (С. major), 3-4 — малый
(С. pygmaeus), 5-6— крапчатый (С. suslicus), 7-10 — серый (С. citellus: С. с. citellus,
С. с. xanthoprymnus, С. с. dauricus, С. с. relictus), 11-12 — длиннохвостый (С. undulatus-
С. u. undulatus, С. u. leucostictus).
ТАБЛИЦА XXVII
1-2— тонкопалый суслик (Spermophilopsis leptodactylus); 3— бурундук (Tamias sibiri-
cus). Белки: 4 — кавказская (Sciurus anomalus), 5-9—обыкновенная (S. vulgaris).
ТАБЛИЦА XXVIII
1-2 — летяга (Pteromys volans). Сони: 3-4— полчок (Glis glis), 5-7 — лесная (Dryo-
mys nitedula), 8 — садовая (Eliomys quercinus), 9— орешниковая (Muscardinus avella-
narius), 10 — мышевидная (Myomimus personatus): 11 —селевиния (Selevinia betpakda-
lensis).
ТАБЛИЦА XXIX
Пятипалые тушканчики: /—большой (Allactaga major), 2— Северцова (А.
severtzovi), 3—прыгун (A. sibirica), 4 — горный (A. williamsi), 5 — малый (A. elater),
6 — Бобринского (A. bobrinskii); 7 — земляной зайчик (Alactagulus pygmaeus); 8 —
тушканчик Житкова (Pygeretmus zhitkovi), 9 — толстохвостый (Р. platyurus).
ТАБЛИЦА XXX
Трехпалые тушканчики: 1 — емуранчик (Stylodipus telum), 2— мохноногий
(Dipus sagitta), 3— гребнепалый (Paradipus ctenodactylus), 4—туркменский (Jaculus
turkmenicus), 5 — Лихтенштейна (J. lichtensteini). Карликовые тушканчики:
6 — карликовый (Cardiocranius paradoxus), 7—трубкоухнй (Salpingotus crassicauda).
Мышовкн: 8 — степная (Sicista subtilis), 9— лесная (S. betulina), 10-12 — одноцвет-
ные (S. concolor).
ТАБЛИЦА XXXI
Мыши: 1 — малютка (Micromys minutus), 2-3— домовая (Mus musculus; 3 — М. m.
bactrianus), 4 — полевая (Apodemus agrarius), 5 — восточноазиатская (A. speciosus),
6 — лесная (A. sylvaticus), 7 — желтогорлая (A. flavicollis), 8 — горная (A. mystacinus).
Крысы: 9-10— черная (Rattus rattus), // — туркестанская (R. rattoides), 12-13 — па-
сюк (R. norvegicus). 14 — пластинчатозубая (Nesokia indica).
ТАБЛИЦА ХХХ11
Песчанки: /—персидская (Meriones persicus), 2-3 — полуденная (М. meridianus),
4 — когтистая (М. unguiculatus), 5 — краснохвостая (М. libycus), 6—Виноградова
(М. vinogradovi), 7 —Зарудною (М. zarudnyi), 8 — малоазиатская (М. blackleri), 9 —
гребенчуковая (М. tamaiiscinus), 10-11— большая (Rhombomys opinus).
ТАБЛИЦА XXXIII
1-2— мышевидный хомячок (Calomyscus baiwardi). Хомяки: 3-4— обыкновенный
(Cricetus cricetus), 5—предкавказский (Cr. radclei), 6— переднеазиатский (Сг. auratus).
Хомячки: 7— джунгарский (Phodopus sungorus), 8 — Роборовского (Rh. roborovskii).
ТАБЛИЦА XXXIV
Хомячки: 1 — Эверсманна (Cricetulus eversmanni), 2— даурский (Сг. barabensis),
3 — длиннохвостый (Сг. longicaudatus), 4-6— серый (Сг. migratorius), 7—крысовидный
(Сг. triton).
ТАБЛИЦА XXXV
1-2— ондатра (Ondatra zibethicus); 3-5— водяная крыса (Arvicola terrestris).
Полевки (серые): 1 — Роберта (Microtus roberti), 2—снежная (М. nivalis), 3—эко-
номка (М. oeconomus), 4 — большая (М. maximowiczi), 5 — темная (М. agrestis), 6 —
обыкновенная (М. arvalis), 7 — закаспийская (М. a. transcaspicus), 8 — Миддендорфа
(М. middendorfi), 9 — северосибирская (М. hyperboreus).
ТАБЛИЦА XXXVII
Полевки (серые): 1-3— общественная (Microtus socialis), 4 — афганская (М. afgha-
nus), 5 — Брандта (М. brandti), 6 — китайская (М. mandarinus), 7-8 — арчевая (М. jul-
daschi; 8— М. j. carruthersi), 9-10 — узкочерепная (М. gregalis), 11-12 — кустарниковая
(М. subterraneus; 12 — М. s. majori).
ТАБЛИЦА XXXVIII
Полевки (лесные): 1-2— шикотанская (Clethrionomys sicotanensis), 3— красно-се-
рая (CL rufocanus), 4-6— рыжая (Cl. glareolus; 6 — Cl. g. frater), 7 — красная (Cl. ru-
tilus). Полевки (каменные): 8 — плоскочерепная (Alticola strelzovi), 9-10 — серебри*
стая (A. roylei), 11—большеухая (A. macrotis), 12—лемминговидная (A. m. lemminus).
ТАБЛИЦА XXXIX
Лемминги: 1 — лесной (Myopus schisticolor), 2— амурский (Lemmus amurensis),
3-4 — сибирский (L. sibiricus), 5 — норвежский (L. lemmus), 6-8— копытный (Dicrosto-
nyx torquatus, зимний, летний и молодой). Пеструшки: 9 — степная (Lagurus
lagurus), 10 — желтая (L. luteus).
1
2
1-2— прометеева полевка (Prometheomys schaposchnikovi). Слепушонки: 3-5 —
обыкновенная (Ellobius talpinus), 6 — афганская (Е. fuseocapillus), 7-9— цокор (Myo-
spalax myospalax; 8 — M. m. aspalax; 9— M. m. psilurus). Слепыши: 10 — малый
(Spalax leucodon), 11 — обыкновенный (S. microphthalmus), 12— гигантский (S. gigan-
teus).
А. П. Кузякин
ОСНОВНЫЕ СВЕДЕНИЯ ПО СИСТЕМАТИКЕ
Систематика — наука о видах животных и растений, о формах
внутривидовой изменчивости и объединениях видов в систематические
группы, или категории, разного ранга.
ВИДЫ, СИСТЕМАТИЧЕСКИЕ ГРУППЫ ВИДОВ
И РАЗНОВИДНОСТИ
Вид (species) — систематическая единица. Никаких других единиц
в биологической систематике нет. Части вида — разновидности (разные
формы внутривидовой изменчивости) или объединения видов в группы
разного ранга называть единицами неправильно. Разновидности и
группы видов суть систематические категории разного ранга, но не
систематические единицы.
Виды группируются в следующие основные категории (по степени
возрастания ранга): род (genus), семейство (familia), отряд
(ordo, в ботанической систематике отряд называют порядком), класс
(class), тип (philum), мир, или царство (regnum). Каждый вид
животного или растения обязательно относится к определенному роду,
семейству, отряду, классу и типу. В целях уточнения сходств и различий
между видами, кроме основных категорий, выделяют и промежуточные:
подроды и реже надроды (или трибы), подсемейства и (редко) надсе-
мейства, подотряды и надотряды, подклассы, надклассы и подтипы.
Группы видов любого ранга (роды, семейства, отряды...) составля-
ются (систематиками) на основе больших или меньших сходств и раз-
личий между ними. Степень морфологических сходств и различий в свою
очередь указывает на большую или меньшую филогенетическую бли-
зость (родственность) разных видов между собой. Например, виды, от-
носящиеся к разным семействам, определенно отличаются друг от друга
больше, чем виды, относящиеся к разным родам одного и того же се-
мейства. Но все эти категории условны. Никаких определенных кри-
териев для объединения видов в род или семейство нет. Это объедине-
ние отражает большую или меньшую эквивалентность различий и за-
висит в значительной степени от опытности, такта, личных взглядов и
склонностей систематика.
Многие виды по морфологическим (реже, физиологическим или эко-
логическим) признакам бывают неоднородными. В таких случаях ви-
довую популяцию делят на разновидности.
Легко различающиеся разновидности, обособленные географически
(имеющие особый ареал), называют подвидами (subcpecies), г е о -
Н. А. Бобринский и др.
графическими расами или просто расами. Подвиды с боль-
шими ареалами или широкой морфологической пластичностью в свою
очередь могут быть неоднородными. В отдельных частях ареала под-
вида могут быть отмечены (обычно на большом, серийном коллекцион-
ном материале) незначительные различия. Такие географические раз-
новидности второго порядка называют племенами. Племя (natio)
слабее отличается от других племен того же подвида, чем подвиды од-
ного вида различаются между собой. Племя тоже имеет свой ареал,
составляющий часть ареала подвида.
Кроме географических (ареальных) разновидностей (подвидов и
племен), бывают разновидности, географически не обособленные. На-
пример, в любой части ареала песца наряду с особями, которые меняют
темно окрашенный летний мех на чисто белый зимний, встречаются «го-
лубые» песцы, круглый год покрытые мехом буровато-серой окраски
разных оттенков. Местами среди обычных хомяков, окраска меха ко-
торых имеет сложный рисунок (охристо-сероватую спину, черный низ,
белые и красноватые пятна на боках тела и головы), встречаются (и
даже в большом количестве) хомяки, мех которых сплошь черный или
черный с белыми пятнами. Среди рыжевато-бурых альпийских пищух
тоже нередки особи с черной окраской всего меха. Такие безареальные
разновидности называют морфами (morpha).
Морфы иного порядка образуются в результате сезонных измене-
ний окраски, густоты и высоты волосяного покрова, а бывают и внеш-
ние половые различия между особями (половой диморфизм). Так, об-
щеизвестны различия между самцами и самками большинства видов
оленей, у которых самцы имеют ветвистые рога, а самки — безрогие.
У многих, например у морских котиков, сивучей, кашалотов, самцы от-
личаются от самок значительно большими размерами и весом. Общеиз-
вестны и возрастные различия у животных любого вида и пола.
Между особями любого вида, подвида, племени или морфы всегда
имеются, хотя бы и совеем небольшие, индивидуальные различия, диа-
пазон которых отражает степень относительного постоянства или, на-
оборот, морфологической пластичности вида.
Индивидуальные и географические различия между особями одного
вида называют изменчивостью. Безареальные же морфы, воз-
растные, половые и сезонные различия правильнее называть не измен-
чивостью, а морфизмами.
Разновидности любого вида домашних животных принято назы-
вать породами. Общеизвестны, например, породы домашних собак:
лайки, немецкие овчарки, таксы, доги и др. В отличие от пород домаш-
них животных, разновидности культурных растенйй принято называть
сортами.
Разновидность — термин, применявшийся пока в неопределенном
значении. Одни разновидностями называли подвиды, другие писали
«подвиды и разновидности», считая их разными понятиями. Во избежа-
ние терминологического разнобоя мы предлагаем называть разновид-
ностями любые формы групповой внутривидовой изменчивости и мор-
физмы: географические расы (подвиды), племена, морфы, породы
домашних животных (и сорта культурных растений). Называть
разновидностями возрастные, сезонные и половые морфизмы, ви-
димо, не следует, и в этом не бывает нужды, хотя не будет и боль-
шой ошибки в выражении: «сезонная разновидность» или «возрастная
разновидность».
Любая разновидность коренным образом, принципиально, качест-
венно отличается от вида.
18
Виды, относящиеся к разным родам или семействам, обычно опре-
деляются без затруднений. Однако определение морфологически близ-
ких видов в пределах одного рода часто вызывает большие затруднения.
Иногда бывает трудно решить, что это: вид или разновидность (подвид,
морфа и т. д.). Как виды, так и разновидности могут различаться
между собой большим количеством признаков: морфологических, физио-
логических, генетических, экологических, биохимических, серологиче-
ских, цитологических и т. д. Для характеристики любого вида или раз-
новидности все эти признаки имеют большое значение, и, чем точнее они
изучены, чем полнее дана характеристика вида, тем лучше. Но в прак-
тике систематической работы используют только признаки, имеющие
диагностическое (определяющее или распознавательное) значение.
Этими признаками и конкретизируются критерии вида.
Критериев вида в систематике три: морфологический, географиче-
ский (основные) и генетический (определяющий или решающий). Из
этих трех критериев в практической систематике используют чаще всего
два первых: морфологический и географический, а к генетическому при-
бегают лишь в крайних (наиболее затруднительных, спорных) случаях.
Использование морфологического и географического критериев как ос-
новных получило название морфолого-географического метода в си-
стематике.
Морфологический критерий вида сводится к тому, что между лю-
быми двумя, даже самыми близкими видами обязательно должны быть
ясные и устойчивые морфологические различия. Степень устойчивости
морфологических признаков выясняется путем детального изучения из-
менчивости: индивидуальной, географической и разного рода морфиз-
мов (возрастного, группового, полового, сезонного). Без знания измен-
чивости у животных изучаемой группы невозможно определить степени
устойчивости морфологических признаков и, следовательно, их диагно-
стической ценности (значимости). Обязательно также условие, что
между двумя видами не может быть особей с промежуточными при-
знаками, т. е. таких особей, о которых по морфологическим признакам
трудно сказать, к какому виду они относятся. Если две определяемые
формы животных (или растений) связаны между собой особями с про-
межуточными признаками, значит, эти формы — не виды, а разновид-
ности. Одного морфологического критерия для определения вида бывает
иногда недостаточно, так как некоторые морфы, подвиды, породы,
сорта и т. д. могут иметь резко выраженные и устойчивые морфологи-
ческие отличия, но их все-таки нельзя называть видами. Тогда исполь-
зуют другие критерии.
Географический критерий сводится к тому, что каждый неодомаш-
ненный вид имеет свой ареал (область распространения). Ареалы двух
близких видов могут совпадать (взаимно перекрываться) или могут
быть в разных местах, но не может быть таких случаев, чтобы ареалы
двух видов примыкали друг к другу, а в месте их соединения (смыка-
ния) встречались бы особи с переходными (промежуточными) призна-
ками. Если между двумя формами бывает такой случай, то это не
виды, а географические формы, подвиды. В ботанической систематике
этим важным критерием обычно не пользуются и географические формы
(подвиды) зачастую называют видовыми названиями (считают их раз-
ными «видами»).
Может быть такой случай, что ареалы двух близких форм примы-
кают один к другому. «Переходных» особей между этими формами нет,
но различия между ними не очень резкие. Изменчивость признаков^
изучена недостаточно. В таких случаях решению вопроса, что это: виды
2*
19
или подвиды, помогает знание общих закономерностей географической
изменчивости. Еще Дарвин указывал, что в определенных географиче-
ских условиях животные (или растения) разных систематических групп
приобретают определенные признаки. Например, чем дальше на север
распространено животное, тем гуще, выше и мягче становится у него
зимний мех, увеличиваются размеры тела, но уменьшается относитель-
ная поверхность тела за счет относительного укорочения конечностей,
хвоста и ушных раковин.
В песчаных пустынях почти у всех животных сильно светлеет ок-
раска (меха, оперения, чешуй); в окраске появляются желтоватые,
или «песчаные», тона и т. д.
Определенность проявления географической изменчивости называют
географическим изоморфизмом. Зная характер географического изомор-
физма, можно не только оценить существо отличий между близкими
формами со смежными ареалами, но можно даже предсказать, какими
признаками должна обладать географическая форма изучаемого вида
в том или ином географическом районе. В закономерностях географи-
ческого изоморфизма оба основных критерия (морфологический и гео-
графический) выступают и неразрывном единстве.
В подавляющем большинстве случаев морфолого-географический
метод в систематике бывает вполне достаточным. Но в некоторых (ред-
ких и особо трудных) случаях для решения вопроса: виды это или раз-
новидности — двух основных критериев бывает недостаточно. Тогда ис-
пользуют третий — генетический критерий.
Генетический критерий заключается в том, что два вида не дают
между собой помесей (гибридов), а если помеси получаются, то они
оказываются бесплодными или почти бесплодными. Например, мул —
помесь между ослом и лошадью. Мулы в основной массе бесплодны. Их
бесплодие указывает на то, что лошадь и осел — разные виды. Отличия
между многими породами собак (например, между овчаркой и бульдо-
гом или между борзой и таксой) гораздо больше, чем между лошадью
и ослом, но помеси между этими породами всегда бывают нормаль-
ными, плодовитыми. Значит, разные породы собак, несмотря на резкие
морфологические различия между ними, относятся к одному виду.
Еще пример: до сих пор все систематики считали, что европейско-
кавказский зубр и североамериканский бизон — разные виды. Однако
они легко дают между собой помеси, вполне жизнеспособные и одина-
ково плодовитые в любых комбинациях и в любом числе поколений.
Биологически это явные разновидности одного вида. Но вместе с этим
не остается оснований и для того, чтобы в систематике зубра и бизона
считать разными видами. Это типичные подвиды одного вида.
Учет физиологического критерия не только в теории, но и в прак-
тике систематики может привести к существенным изменениям в трак-
товке многих форм, разделяемых в разные виды или даже роды, а фак-
тически представляющих собой лишь разновидности.
Генетический критерий настолько важный, что мы назвали его оп-
ределяющим или отвечающим на вопрос, с какой формой имеем дело:
с видом или разновидностью. Это признается и другими зоологами-си-
стематиками. Генетический критерий не только включают в определе-
ние понятия «вид», но считают возможным даже в самых кратких опре-
делениях вида ограничиваться указанием только на этот критерий. На-
пример, Н. А. Бобринский (1944) 1 писал, что вид можно рассматривать
как «свободно скрещивающееся сообщество» (стр. 20).
1 В скобках указан год издания статьи или книги данного автора, приведенной
в списке литературы.
:20
Соглашаясь с правильностью высокой оценки генетического (или
физиолого-генетического) критерия, нельзя согласиться, что только
посредством этого критерия можно выяснить, с какой формой вида
имеем дело.
Среди более миллиона видов и нескольких миллионов разновид-
ностей описанных в настоящее время диких животных только в отно-
шении ничтожного числа форм известны условия и результаты их скре-
щиваемости (или нескрещиваемссти).
Систематики, как известно, имеют дело с коллекционным материа-
лом. Сравнивая целые серии тушек и черепов млекопитающих, собран-
ных зоологами в разные годы и в разных местах, трудно определить,
могут эти (определяемые или сравниваемые) формы скрещиваться
между собой или не могут.
В литературе, касающейся теории вида, имеются правильные
указания на нередкое несоответствие между «биологическими» и «так-
сономическими» видами. Это особенно касается ботанической систе-
матики.
Представляется неправомерным сохранять два различных понятия:
1) «биологический вид» и 2) «таксономический», или «систематиче-
ский», вид.
Таксономия (систематика) не может иметь какие-то свои, особые
«виды», отличные от «биологических» видов. Вид должен быть од-
ним и тем же как в природе, так и в таксономии. Выяснение видов
(видового состава) в природе составляет одну из первых и главных за-
дач систематики. А если в систематике некоторые разновидности пока
еще называются разными «видами», так это указывает лишь на недо-
статочное совершенство систематики, подлежащее по мере накопления
материалов быстрым уточнениям и исправлениям.
Понимание и трактовка вида неразрывно связаны с пониманием
и трактовкой видообразования.
Проблема видообразования — одна из самых острых и сложных
проблем современной биологии. Возникла она после трудов Ламарка
(1809) и особенно Ч. Дарвина (11859).
Бессмертная заслуга Дарвина заключается в доказательстве того,
что виды животных и растений происходят одни из других, преемст-
венно связаны между собой, что все современные виды животных и
растений есть продукты длительного исторического (развития. Эти ос-
новные положения Дарвина все современные биологи признают, раз-
деляют, считают бесспорными.
Теперь речь идет не о том, что естественным путем произошли раз-
ные виды животных и растений или они были кем-то созданы. Теперь
биологи стремятся понять, как происходили одни виды животных и
растений из других.
Вид—совокупность особей животных (или растений), морфологи-
чески относительно одинаковых или представленных несколькими раз-
новидностями— географически обособленными расами (подвидами),
племенами, или безареальными морфами (диких животных), породами
(одомашненных животных) или сортами (культурных растений). Для
каждой разновидности характерны одинаковые возрастные, половые и
сезонные морфизмы. Диапазон морфологической изменчивости у диких
материковых (и океанических) видов наименьший, у обитателей мелких
островов и замкнутых водоемов значительно шире, а у одомашненных
(окультуренных) видов практически неограниченный. В любой части
сплошного или расчлененного ареала любого неодомашненного вида не
должно быть особей с признаками, переходными к любому другому
21
виду. Кроме ясных и устойчивых морфологических различий, для любой
пары близких видов характерна физиологическая несовместимость, т. е.
неспособность давать нормально плодовитые помеси. Взаимоотношения
между близкими (преемственно связанными) видами при их совмест-
ном распространении всегда антагонистические.
ОСНОВНЫЕ ПРАВИЛА СИСТЕМАТИЧЕСКОЙ
НОМЕНКЛАТУРЫ
Основателем современной научной номенклатуры считается все-
мирно известный шведский систематик Карл Линней (К. Linnaeus).
В своем труде «Sistema naturae» («Система природы») он дал краткие
характеристики всем известным в то время видам, родам, отрядам и
классам животных и растений. Книга Линнея выдержала 12 изданий
при жизни автора и одно, 13-е, посмертное издание. В настоящее время
исходным (основным) считают 10-е издание, вышедшее в 1758 г., в ко-
тором Линней впервые со всей строгостью применил для каждого вида
латинское название, составленное из двух слов — из родового и собст-
венно видового. Например, зайца-беляка он назвал Lepus timidus.
Слово Lepus (заяц) означает название рода. Второе слово в названии —
собственно видовое: timidus—в буквальном переводе означает «трус-
ливый».
Латинское название из двух слов дается каждому виду животного
и растения, распространенному в любой части земного шара, в стране,
населенной народом, говорящим на любом языке. За исходные при-
нимают названия, данные видам в 10-м издании книги Линнея (1758).
Все названия, в том числе и латинские, данные различными авторами
до 1758 г., в расчет не принимаются.
Название рода всегда и обязательно пишется с прописной (заглав-
ной) буквы, а название вида (в зоологической систематике) пишут
всегда со строчной (маленькой) буквы, если даже оно означает имя
собственное. Например, в названии кожана Огнева — Vespertilio og-
nevi — слово ognevi означает фамилию русского зоолога-систематика
С. И. Огнева, в честь которого дано это название, но пишется оно
с маленькой буквы. (В ботанической систематике этого правила не при-
держиваются.) Слово, означающее название вида, писать без родового
или первой буквы родового названия не принято, но сокращение родо-
вого названия до одной, двух или трех букв допускается. Например,
timidus писать нельзя, a L. timidus — можно.
Обозначение вида двумя латинскими словами называют бинарной
(двухсловной) номенклатурой.
Если вид в пределах своего ареала образует две или несколько хо-
рошо отличимых географических рас (подвидов), для их научного на-
звания применяют уже тройную (трехсловную) номенклатуру, причем
за подвидом из первого места описания (за «номинальной формой»)
закрепляют видовое же название, но написанное второй раз, а для всех
вновь описываемых подвидов дают новые, отличные от видового назва-
ния. Например, Линнеем был описан заяц-беляк из Швеции. Но позднее
описали несколько подвидов беляка из других мест его ареала. Тогда
беляков из Швеции (номинальную форму) стали называть: Lepus timi-
dus timidus, а белякам из Амуро-Уссурийского края Goodwin в 1933 г.
дал название: Lepus timidus. mordeni. Третье слово означает название
подвида.
Если в ареале подвида имеются географически обособленные, но
очень слабо отличающиеся формы — племена (natio), то им тоже
22
иногда давали особые латинские названия в виде четвертого слова. На-
пример, описано светло окрашенное племя обыкновенного ежа (из
окрестностей г. Тобольска), и название его получилось таким: Erinaceus
europaeus centralrossicus natio pallidus Ognev, 1928.
Хотя четырехсловные названия слабо различающихся natio есть,
но их можно и лучше не употреблять. Даже в систематических рабо-
тах лучше ограничиваться тройной (трехсловной) номенклатурой,
а в экологических работах вполне достаточно видового (бинарного)
названия.
В систематических списках, монографиях и определителях, кроме
полного латинского названия, принято писать (тоже латинскими бук-
вами) фамилию автора, впервые описавшего данный вид или подвид,
и год этого (первого) описания. Например, L. timidus Linnaeus, 1758;
или (если определяется до подвида): Lepus timidus timidus Linnaeus,
1758; Lepus timidus mordeni Goodwin, 1933.
Если две формы, описанные вначале в качестве самостоятельных
видов, при дальнейшем изучении окажутся только подвидами одного
вида, то за ними закрепляется одно видовое название, и обязательно
то, какое было дано раньше. Это называется правилом приоритета.
Год описания обычно ставят лишь в подзаголовках и в синонимике
систематических сводок и определителей или в специальных системати-
ческих списках. В статьях же фаунистического или экологического со-
держания, а также в тексте и определительных таблицах систематиче-
ских работ год описания вида или подвида можно не ставить.
В отношении авторов описаний видов и подвидов придерживаются
того же режима.
Название, закрепленное за видом (или подвидом) по правилу прио-
ритета, изменять нельзя, если даже оно по своему значению стало не-
точным. Например, пасюка (серую крысу) раньше всех описал Вег-
kenhout (в 1769 г.) по экземплярам, добытым в Англии, но завезенным,
по мнению автора, из Норвегии, и назвал его A^us norvegicus (норвеж-
ская мышь). В дальнейшем оказалось, что пасюк живет не только
в Норвегии, но и во многих других странах мира и Норвегия не яв-
ляется местом возникновения этого вида, однако заменить слово nor-
vegicus в названии пасюка другим словом уже нельзя.
Правило приоритета относится только к названиям родов и под-
родов, видов и подвидов. За всеми видами (и подвидами), описанными
впервые Линнеем и другими авторами после 1758 г., закрепляются на-
звания, данные раньше других, но при условии, что автор придержи-
вался латинской бинарной (для вида) или тройной (для подвида)
номенклатуры и опубликовал хотя бы краткое описание данного вида
(или подвида). Название, не снабженное описанием, считается за no-
men nudum — «голое название» и в расчет не принимается. Все назва-
ния, данные видам (подвидам, подродам -и родам), уже названным
ранее, отходят в синонимы.
Если форма, первоначально описанная в качестве подвида, в даль-
нейшем оказывается самостоятельным видом, то ее подвидовое назва-
ние становится видовым. Например, О. Томас в 1899 г. описал из Ичана
в пров. Хубэй кожана Vespertilio murinus superans. Как видно по на-
званию, Томас считал его подвидом двухцветного кожана (V. muri-
nus L.). Однако дальнейшее изучение показало, что описанная Томасом
форма не подвид двухцветного кожана, а самостоятельный вид. Под-
видовое название стало теперь видовым — V. superans Thomas, 1899.
И наоборот, если форма вначале была описана в качестве самостоя-
тельного вида, а более детальное изучение показало, что она представ-
23
ляет собой лишь географическую расу (подвид) другого, ранее описан-
ного вида, то видовое название становится подвидовым с указанием
автора и года первоописания.
Изменения в названиях родов (или подродов) на видовые (и под-
видовые) названия влияния не оказывают. Например, К. Линней,
II. Паллас и многие другие систематики всех мышей, крыс и даже по-
левок относили к одному роду Mus. Теперь этот род разделили па под-
роды, роды и даже подсемейства. Однако изменения родовых (и под-
родовых) названий не оказали влияния на собственно видовые назва-
ния. Черная крыса — Mus rattus Linnaeus, 1758 — относится теперь
к особому роду Rattus и пишется: Rattus rattus Linnaeus, 1758, или
обыкновенная полевка — Mus arvalis Pallas, 1799 — относится теперь
к роду серых полевок (и особому от мышей подсемейству полевок) и
называется Microtus arvalis Pallas, 1799.
Некоторое время существовало правило, что если вид (или подвид)
при первом описании относился не к тому роду, к какому он относится
теперь, то фамилию автора и гсд первоописания заключали в скобки.
Например, в двух выше приведенных случаях писали: Rattus rattus
(Linnaeus, 1758) и Microtus arvalis (Pallas, 1779). Теперь этого пра-
вила не придерживаются.
По правилам научной номенклатуры в течение некоторого срока
было обязательным отражать в названиях видов (и подвидов) принад-
лежность не только к роду, но и к подроду. При этом подродовое на-
звание ставилось после родового в скобках. Например, обыкновенная и
узкочерепная полевки относятся к роду серых полевок — Microtus, но
к двум различным подродам — собственно Microtus и Stenocranius. Так,
их подродовую принадлежность обязательным было отражать в видо-
вых названиях и писать так: Microtus (Microtus) arvalis Pallas, 1779;
M. (Microtus) arvalis Pallas; M. (M). arvalis Pallas, Microtus (s. str.)
arvalis Pall.; Microtus (Stenocranius) gregalis Pallas, 1779; M. (Steno-
cranius) gregalis Pallas; M. (St.) gregalis Pall., M. (Stenocranius) gre-
galis major, Ognev, 1923. Хотя выделение подродов имеет частный,
узкоспециальный систематический (точнее — филогенетический) смысл,
этого правила придерживались даже в статьях и книгах по морфоло-
гии, экологии, фаунистике и т. д.
Теперь даже в специальных систематических сводках не отвергают
подродовые группировки видов, указывая в подзаголовках названия
подродов, но в видовые названия их не включают. Узкочерепную по-
левку теперь законно и правильно называть: Microtus gregalis Pallas,
1779, без включения в это название подродового слова Stenocranius. Та-
кое изменение упрощает номенклатуру, делает ее менее громоздкой и
не причиняет ущерба решению узкоспециальных задач по филогенети-
ческим взаимоотношениям между отдельными видами. Выделения под-
родов теперь, даже в солидных систематических сводках, ограничивают
лишь указанием их названий, автора и года описания без подробной
характеристики и даже без кратких диагнозов.
Названия семейств животных образуются путем прибавления
к основе типичного родового названия окончания idae. Например, род
кошки — Felis, а семейство кошачьи — Felidae; род обыкновенных тю-
лений — Phoca, семейство тюленевые — Phocidae. Названия подсе-
мейств всегда оканчиваются на inae. Например, подсемейство настоя-
щих кошек — Felinae; подсемейство полевок — Microtinae (от типичного
рода серых полевок — Microtus). Выделение подсемейств имеет тоже
узкоспециальный смысл, и в последнее время даже в солидных систе-
матических сводках подсемейства приводятся только в подзаголовках
24
без диагнозов и подробных характеристик. Правило приоритета на
названия подсемейств, семейств и вышестоящих систематических кате-
горий не применяется.
В названиях отрядов, подотрядов, надотрядов, классов, подклассов,
типов и подтипов общие правила пока не установлены. В систематике
рыб уже приняты и в систематике птиц частично приняты для латин-
ских названий всех отрядов одинаковые окончания formes (.. .образ-
ные) . Например, сельдеобразные — Clupeiformes; ракшеобразные —
Coraciiformes, аистообразные — Ciconiiformes и т. д. В систематике
млекопитающих такой унификации в названиях отрядов пока не при-
нято, хотя она имела бы смысл с точки зрения упорядочения номенкла-
туры крупных таксономических категорий.
Правила научной систематической номенклатуры периодически
пересматриваются, совершенствуются и утверждаются на специальнькх
международных конгрессах. Правила эти касаются только обязательных
для каждого вида международных латинских названий.
Местные, национальные названия животных правилам научной но-
менклатуры не подчиняются, их устанавливают в каждой стране по-раз-
ному, в зависимости от языка, национальных традиций и т. д.
В задачу систематиков должно бы входить не только выполнение
международных правил латинской номенклатуры, но и упорядочение
национальных названий.
Русские названия млекопитающих многими авторами даже в боль-
ших систематических сводках давались произвольно. Например, в свод-
ках Академии наук СССР (1952, 1963) Nesokia indica названа: «индий-
ская земляная, или пластинчатозубая, крыса»; Rattus norvegicus —
«крыса рыжая, или амбарная, пасюк»; «серая, или амбарная, крыса,
пасюк».
Для многих видов даны русские названия из трех слов без какой-
либо необходимости к этому. Например, красная полевка (Clethriono-
mys rutilus Pall.) названа Б. С. Виноградовым (1952) и И. М. Громо-
вым (1963) «сибирской красной полевкой», хотя она, кроме Сибири,
населяет весь север Европы, от Урала до Скандинавии включительно,
и весь север Америки, от Тихоокеанского до Атлантическою побережий;
живет она также в Казахстане, Монголии, на Дальнем Востоке. Дру-
гих полевок, в названиях которых было бы слово «красная», нет. Зна-
чит, слово «сибирская» не уточняет названия красной полевки, а де-
лает его только излишне громоздким и неправильным по существу.
То же можно сказать и о «европейской рыжей полевке», в названии ко-
торой слово «европейская» ничем не оправдано и лишне. Бывали на-
звания и еще более произвольные. Например, большеухая полевка
(Alticola macrotis) называлась Б. А. Кузнецовым (1944) «саянской
высокогорной полевкой», Б. С. Виноградовым (1952) —«забайкальской
высокогорной полевкой», А. Г. Банниковым (1954)—«забайкальской
горной полевкой» и т. д.
В новом издании «Определителя» для каждого вида млекопитаю-
щих фауны СССР оставлено по одному русскому названию, состоящему
не более чем из двух слов, так как произвольные изменения ни к чему,
кроме затруднений и путаницы, не приводят. Названия некоторых ви-
дов явно неудачны, но оставлены потому, что стали общепринятыми.
Таковы, например, краснохвостая песчанка, у которой хвост никогда не
бывает красным, или желтая пеструшка, которая никогда не бывает
пестрой.
По аналогии с латинскими, в русских названиях следовало бы ста-
вить на первом месте родовое название, а на втором собственно видо-
25
вое. Однако в русских названиях существительное обычно шире ро-
дового латинского названия. Например, слово «полевка» означает Mi-
crotus, Clethrionomys, Alticola, Prometheomys, «лемминг» фигурирует
в названиях грызунов из родов Lemmus, Myopus, Dicrostonyx; тушкан-
чиками называют грызунов из родов Allactaga, Pygere,tmus, Dipus Pa-
radipus, Jaculus, Salpingotus и др. Прилагательное в русских назва-
ниях точнее отражает специфику как вида, так и рода. Поэтому на
первом месте правильнее ставить прилагательное, что больше соот-
ветствует и специфике русской речи.
О ПОРЯДКЕ РАСПОЛОЖЕНИЯ ВИДОВ И ГРУПП
МЛЕКОПИТАЮЩИХ ФАУНЫ СССР
Трудно представить себе систему без обоснованного порядка в рас-
положении видов и видовых групп разного ранга. Сами слова «си-
стема» и «порядок» означают синонимы.
Обоснованный порядок приводит к унификации в расположении
материалов в фаунистических статьях, региональных сводках, учебни-
ках, справочниках, в музейных экспозициях, в картотеках и сильно
облегчает пользование ими. В порядке взаимного расположения дол-
жны отражаться филогенетические связи между отдельными видами
и группами разного ранга. Порядок — одна из главных целей систе-
матики.
В отрядах насекомоядных и рукокрылых эта идея была уже прак-
тически осуществлена в прежних изданиях. Ранее предложенный по-
рядок по этим отрядам уже принят большинством советских и зару-
бежных зоологов. Теперь хотелось бы упорядочить расположение ма-
териала и в других отрядах, особенно в наиболее трудном и обширном
отряде грызунов.
В порядке расположения видов неизбежны большие трудности,
преодолеть которые не всегда удается. Развитие органического мира
шло не в одном направлении, не по одной прямой линии от низших
к высшим, а в разных направлениях, обусловленных приспособлениями
к разным условиям существования, в виде целых пучков филогенети-
ческой дивергенции, в виде веток единого «древа». Пытаясь располо-
жить все ветви в одну линию (в один список), зачастую не удается из-
бежать перерывов, провалов между отдельными группами и разъеди-
нения сравнительно близких видов и групп другими, менее родствен-
ными. Эти трудности, однако, не освобождают систематиков от попы-
ток расположения видов и групп в более или менее стройном порядке.
Приведенный выше список млекопитающих фауны СССР содер-
жит первую в этом направлении попытку.
Отряд китообразных мы начали с зубатых китов. В расположении
семейств, родов и видов китообразных мы не нашли обоснованного по-
рядка.
КАК ПОЛЬЗОВАТЬСЯ ОПРЕДЕЛИТЕЛЕМ
Основу определителя составляют таблицы для определения, рас-
положенные последовательно от высших таксономических групп к низ-
шим. В каждой группе таблица расположена после краткой характери-
стики этой группы.
В определитель включены все виды млекопитающих фауны СССР.
Для определения надо иметь перед глазами тушку, шкурку (или
шкуру) с целыми конечностями и череп взрослого или полувзрослого
26
^животного с целыми зубами. В таблицах не отражены признаки моло-
дых особей.
Таблицы составлены по общепринятому дихотомическому прин-
ципу. Текст таблицы содержит перенумерованные тезы и антитезы.
В каждой тезе по возможности указаны один или несколько внешних
признаков и один или несколько признаков строения черепа, зубной
системы или отдельных зубов.
Этот принцип, к сожалению, не везде удалось выдержать, особенно
в таблицах по определению некоторых групп грызунов. Например,
в таблицах для определения видов сусликов указаны почти одни внеш-
ние признаки, а для определения серых полевок — почти одно строение
коренных зубов.
Если неизвестно, к какому отряду относится подлежащий опреде-
лению экспонат, то надо начинать с «Таблицы для определения отря-
дов млекопитающих фауны СССР», помещенной на странице 34.
Начинать определение надо с первой строки таблицы, в начале ко-
торой стоит цифра «1» (это номер тезы), а рядом с ней другая цифра,
заключенная в круглые скобки и означающая номер антитезы. Если
признаки, перечисленные в тезе, соответствуют признакам определяе-
мого экспоната, то надо читать следующую тезу, номер которой ука-
зан в конце последней строки первой тезы. Если же признаки не под-
ходят, не соответствуют признакам определяемого экспоната, то надо
читать антитезу, номер которой указан в круглых скобках. Если текст
антитезы подошел, но название группы или вида еще не указано, то
после этого надо читать текст тезы, номер которой указан в конце ан-
титезы. Такое сопоставление признаков, перечисленных в тезах и анти-
тезах, непременно приведет к названию группы (или вида), к которой
относится определяемый экспонат.
Когда станет известным, к какому отряду относится определяемый
вид, в «Таблице для определения отрядов...» после названия отряда,
к которому относится этот экспонат, указана страница, на которой дана
краткая характеристика отряда. После характеристики отряда следует
«Таблица для определения семейств...» этого отряда, которой надо
пользоваться так же, как предыдущей. Сопоставлением признаков оп-
ределяемого экспоната с признаками в тезах и антитезах будет опре-
делено название семейства, после которого указана страница книги,
где дана краткая характеристика этого семейства и «Таблица для
определения родов...».
После названия рода в таблице указана страница, где дана харак-
теристика этого рода, и если к этому роду относится не один, а два
или несколько видов, то следует «Таблица для определения видов» этого
рода. После русского и латинского названий вида, к которому отно-
сится определяемый экспонат, указана страница, на которой приведены
по этому виду краткие сведения по распространению, образу жизни,
практическому значению и по географической изменчивости его, т. е.
по географическим расам (подвидам), распространенным в пределах
СССР.
Сведения о подвидах набраны мелким шрифтом (петитом). Они
содержат только латинские названия животных, фамилии авторов, ко-
торыми описаны эти подвиды (в общепринятой латинской транскрип-
ции), годы описания подвидов, основные отличительные признаки каж-
дого подвида и очень краткие сведения об их ареалах. Таблиц для
определения подвидов в этой книге нет, так как географическая измен-
чивость подавляющего большинства видов изучена пока недостаточно
обстоятельно. Описано подвидов больше, чем их можно практически
27
различить. Отличия многих подвидов можно заметить лишь при непо-
средственном сличении определяемых экспонатов с экспонатами дру-
гих подвидов.
Подвидовая систематика, смысл которой сводится только к отраже-
нию географической изменчивости видов, имеет узкоспециальное зна-
чение, и доводить определение до подвида не обязательно. В статьях,
сводках, отчетах, дипломных и курсовых работах не специального си-
стематического содержания названия подвидов лучше вообще не ука-
зывать, можно ограничиться названием только единственной реаль-
ной качественно обособленной систематической единицы — вида.
Во избежание путаницы в определителе подвидам не даны осо-
бые русские названия.
Перед характеристикой или кратким описанием ареала каждого
наземного вида млекопитающих фауны СССР имеется ссылка на номер
цветного изображения данного вида; римская цифра означает поряд-
ковый номер таблицы, арабская — номер рисунка на этой таблице. За-
тем указан номер карты ареала (если она есть). Перенумерованные
карты ареалов всех наземных видов млекопитающих и части ластоно-
гих и китообразных сведены в единый атлас в конце книги.
ОБЪЯСНЕНИЕ ГЛАВНЕЙШИХ ПРОМЕРОВ
Длина тела (означает длину головы и туловища):
1. У мелких млекопитающих (величиной до зайца, лисицы и др.) —
расстояние от конца носа до заднего края таза, основания хвоста или
анального отверстия. Точки эти, как правило, совпадают. Если же не
совпадают, то задний конец таза — точка наиболее определенная и
устойчивая. Для измерения длины тела зверька кладут на спину, при
этом позвоночник его выпрямляют. Измерение производят мерной ли-
нейкой или штангенциркулем.
2. У крупных хищников, ластоногих и копытных — расстояние от
конца носа до основания хвоста; измеряется лентой по спине, следуя
ее изгибам.
3. У китообразных — расстояние от конца морды до вырезки между
хвостовыми ластами; измеряется лентой, следуя изгибам тела.
Длину хвоста, если не оговорено, измеряют без концевых
волос.
Длину ступни измеряют (у мелких животных — штангенцир-
кулем, у крупных — лентой) по прямой от задней стороны пятки до
конца самого длинного пальца без когтя. При описании вида часто
не пишется «длина задней ступни», так как ступня бывает только
задней, а вентральную сторону конца передней конечности называют
ладонью.
Длина (или высота) уха — расстояние от нижней вырезки,
или от места приращения внешнего края, до вершины ушной раковины
без концевых волос. Если вырезка (место соединения внешнего и внут-
реннего краев) ушной раковины не совпадает с ее основанием (как
у зайцев и копытных), то ухо измеряют от его основания до вершины
по тыльной стороне.
Ко иди л о б а з ал ь н а я длина черепа — расстояние от наи-
более выступающей вперед части межчелюстной (или предчелюстной)
кости (в которой сидят резцы) до задней стороны затылочных мыщел-
ков. Если написано просто «длина черепа», то речь идет только о кон-
дилобазальной длине.
28
Общая длина че-
репа— расстояние от на-
иболее выступающей вперед
точки межчелюстных костей
до наиболее выступающей
назад части черепа.
Наибольшая дли-
на черепа — расстояние
от наиболее выдающейся
вперед части верхних рез-
цов до наиболее выступаю-
щей назад части черепа.
Это один из наиболее точ-
ных промеров, но почему-то
редко производимый.
Скуловая шири-
на — наибольшее расстоя-
ние между внешними сто-
ронами скуловых дуг.
Ширина черепа —
наибольшее расстояние
между внешними сторо-
нами черепа в области слу-
ховых костей или выступаю-
щих в бока костных гребней
около них.
Ширина мозговой
капсулы обычно совпа-
дает с шириной черепа, но
если сильно развиты боко-
вые гребни, то измеряется
над ними.
Межглазничный
промежуток (или меж-
глазничная ширина чере^
па) —наименьшее расстоя-
ние между внутренними
стенками левой и правой
глазниц, измеряется сверху.
Ширина носового
отдела черепа — рас-
стояние между внешними
сторонами верхних клыков
(у ежей, кротов, хищников,
рукокрылых) или между
внешними сторонами пер-
вых промежуточных зубов
(у землероек).
Ширина между
верхними корен-
ными — наибольшее рас-
стояние между внешними
сторонами верхних корен-
ных зубов (очень показа-
тельный и точный промер).
Рис. 1. Череп ежа (Erinaceus euro-
paeus dauricus Sund.) сверху.
Измерения черепа: а — кондилоба-
зальная длина; б — наибольшая длина;
в — скуловая ширина; г—наибольшая ши-
рина черепа; д — ширина мозговой капсу-
лы; е — межглазиичный промежуток.
Места приложения ножек штангенциркуля при из-
мерениях черепа: ВРЗ — длина верхнего ряда зубов;
ДЧ — кондилобазальная длина черепа (ЗМ — заты-
лочный мыщелок); ЗШ — ширина между рядами
коренных зубов; МШ — межглазиичная ширина;
НД — наибольшая длина черепа; НШ — носовая ши-
рина; 770 — предглазиичпое отверстие; ПШ — пред-
глазничная ширина; ШЧ — ширина черепа.
29
Длина верхнего ряда зубов — очень важный промер, про-
изводится всегда до задней стороны последнего заднекоренного зуба;
у землероек (Soricidae) — от передней стороны крупных резцов сред-
ней пары; у других насекомоядных, летучих мышей и хищников — от
передней стороны верхнего клыка (такой промер точнее называть дли-
ной максиллярного ряда-зубов); у грызунов — от передней стороны пе-
реднего предкоренного или заднекоренного зуба. В последнем случае
точнее пишется: длина верхнего ряда коренных зубов.
Высота нижней челюсти — расстояние от прогиба у осно-
вания углового отростка до вершины венечного отростка • нижней
челюсти.
КЛАСС МЛЕКОПИТАЮЩИЕ.
CLASS MAMMALIA
А. П. Кузякин
ОБЩАЯ ХАРАКТЕРИСТИКА
Млекопитающие, или звери,— высшие позвоночные, выкармливаю-
щие своих детенышей молоком.
Покровы млекопитающих сложного строения. Эпидермис состоит
из двух слоев: нижний — мальпигиев слой, образованный жизнедея-
тельными кубическими или цилиндрическими клетками, и верхний —
роговой слой эпидермиса, клетки которого подвержены постепенному
перерождению (ороговению) и отмиранию. Под эпидермисом распо-
лагается собственно кожа, или кутис, образованная волокнистой
тканью. В нижнем, более рыхлом слое кожи может развиваться слой
подкожной жировой клетчатки, особенно характерной для водных жи-
вотных (китообразных, ластоногих) и для животных, впадающих в дли-
тельную зимнюю спячку (как многие грызуны, ежи, летучие мыши, не-
которые хищники). Сильного развития достигают производные кожи:
различные кожные железы, волосы и другие роговые образования.
Волосы для млекопитающих так же характерны, как перья для
птиц, роговые чешуи для рептилий или костные чешуи для рыб. Лишь
у немногих млекопитающих волосяной покров вторично утрачивается
(у китообразных, моржей и некоторых других). Волосы по их строению
и функции делят на несколько типов. У большинства видов основу во-
лосяного покрова составляет густой мягкий и низкий пух, подпушь или
подшерсток. Среди пуховых волос рассеяны более длинные и жесткие
остевые, или направляющие, волосы. Видоизменения остевых волос при-
водят к образованию щетины или даже иглистого панциря. На многих
частях тела располагаются вибриссы, выполняющие осязательную функ-
цию. Волосяной покров периодически меняется (линяет). У некоторых
видов бывает одна линька в году, у других — две и у некоторых (как
кроты) — три линьки, при одной из которых (летней) меняется только
часть покрова. Окраска волос имеет большое диагностическое (система-
тическое) значение. Для некоторых видов характерны возрастные и се-
зонные различия в окраске меха.
Из прочих роговых образований для многих млекопитающих харак-
терны когти (ногти или копыта), чехлы на рогах (у полорогих парно-
копытных), роговые пластинки или чешуйки на лапах, хвостах и т. д.
Особенно сильного развития покров из роговых пластин достигает
у ящеров и броненосцев.
Кожные железы многочисленны и разнообразны. Различают: по-
товые, сальные, пахучие железы, но особенно характерны молочные,
31
или млечные, железы, возникшие в результате видоизменения потовых
желез. У водных млекопитающих потовые железы не развиты; у назем-
ных видов они располагаются на разных участках кожи и выполняют
функцию органов теплообмена (терморегуляции) и дополнительных ор-
ганов выделения.
Череп массивный с большой мозговой коробкой. Нижняя челюсть
представлена только одной (парной или непарной) зубной костью по-
кровного (кожного) происхождения. Сочленованная и квадратная кости
челюстного аппарата преобразованы в слуховые косточки, которых
у 'млекопитающих, в отличие от всех прочих позвоночных животных, 3:
стремечко, молоточек и наковальня; располагаются они в полости сред-
него уха, возникшей в результате преобразования одной из жаберных
полостей — полости брызгальца. У большинства видов хорошо развиты
скуловые дуги, образованные особыми скуловыми костями и скуловыми
отростками верхнечелюстных и чешуйчатых костей. Характерны для
млекопитающих костные слуховые барабаны, отсутствующие у прочих
позвоночных. Затылочных мыщелков 2; расположены они по бокам
от большого затылочного отверстия.
Зубная система сложная. Все зубы располагаются в глубоких ям-
ках— альвеолах и имеют по одному, 2 или (у заднекоренных зубов) по
3 корня. У большинства зубная система содержит от 2 до 5 категорий,
различающихся по строению и функциям. Зубные системы в целом и
строение отдельных зубов имеют очень большое диагностическое зна-
чение. Для удобства зубную систему любого вида или группы можно
изобразить в виде зубной формулы. Формула содержит буквы и цифры.
Буквы означают сокращенные латинские названия отдельных катего-
рий зубов: I — incisivi (резцы); С — canini (клыки); Рш — praemolares
(предкоренные) и М— molares (заднекоренные). После буквы пи-
шется количество зубов в виде простой дроби, в числителе которой ука-
зывается количество зубов этой категории с одной стороны верхней
челюсти, а в знаменателе — то же в нижней челюсти. За исходную,
наиболее примитивную считают зубную систему, состоящую из 44 зубов.
Исходная зубная формула выглядит так:
I —С—Pm —М —-2 = 44.
3 1 4 3
Среди задних предкоренных и реже передних коренных имеются
зубы несколько отличного от других строения; их целесообразно вы-
делять в особую категорию крупных предкоренных зубов (praemolares
prominantes, Pmp). Среди плацентарных млекопитающих только
у части зубатых китообразных (Odontoceti) зубная система содержит
более 44 (до 260) зубов относительно однородного строения — одновер-
шинных, конических. У остальных современных плацентарных млеко-
питающих зубов 44 или (у большинства видов) меньше. Уменьшение
числа идет за счет редукции (исчезновения) зубов разных категорий.
Последними редуцируются самые устойчивые из всех зубов — крупные
предкоренные (у наземных хищников верхние из них называются «хищ-
ническими» зубами). Беззубые киты (Mystacoceti) во взрослом состоя-
нии зубов не имеют. Для зубной системы всех высших млекопитающих
характерны две возрастные стадии: молочная и постоянная. Молочных
предшественников имеют зубы всех категорий, кроме заднекореиных.
В осевом скелете характерно постоянное (за редкими исключе-
ниями) число шейных позвонков (7) с плоскоповерхностными (плати-
цельными) телами. Два первых шейных позвонка (атлас и эпистрофей)
по своему строению отличаются от последующих (на кольцевидном ат-
32
ласе спереди есть сочленовные ямки и нет тела, а у эпистрофея, кроме
своего тела, спереди есть зубовидный отросток, который является по
своему происхождению телом первого позвонка). От грудных позвонков
отходят ребра, часть которых достигает грудины (настоящие ребра),
другая часть оканчивается свободно (ложные ребра). В поясничном от-
деле — от 2 до 9 позвонков, в крестцовом — от 2 до 4 обычно сросшихся
позвонков и в хвостовом отделе — от 3 до 49.
В плечевом поясе коракоидная кость самостоятельна только у яй-
цекладущих; у остальных она срастается с лопаткой, образуя коракоид-
ный отросток лопатки. Ключицы у многих млекопитающих нет. Тазовый
пояс состоит из трех костей: седалищной, подвздошной и лобковой, ко-
торые у многих срастаются в одну, безымянную кость.
Скелет парных конечностей построен по схеме конечностей пяти-
палого типа. Концевые (дистальные) отделы конечностей имеют раз-
нообразное строение, в зависимости от функций, и широко используются
в диагностике отрядов, реже — семейств.
В мускульной системе млекопитающих особенно характерна плос-
кая куполообразно изогнутая мышца — диафрагма, разделяющая по-
лость тела на грудной и брюшной отделы. Сокращение диафрагмы
(изменение кривизны купола) ведет к увеличению объема грудной по-
лости и акту вдоха.
Центральная нервная система у млекопитающих достигает высо-
кого развития, особенно за счет коры больших полушарий головного
мозга — центра высшей нервной деятельности.
Органы чувств у видов разных групп имеют разный уровень раз-
вития.
Пищеварительная система сложная; кишечник сильнее дифферен-
цирован, чем у других позвоночных; разнообразны пищеварительные
железы.
Дыхательная система представлена легкими сложно-губчатого
строения; альвеолы, в стенках которых проходят капиллярные сосуды,
исчисляются у одной особи десятками и сотнями миллионов. Гортань
поддерживается 5 хрящами. Голосовые связки располагаются между
щитовидным и черпаловидными хрящами.
Сердце четырехкамерное с одной (левой) дугой аорты, отходящей
от левого же толстостенного желудочка. Воротной системы в почках
нет. Эритроциты мелкие, безъядерные.
Почки тазовые, метанефрические. Мочеточник впадает в мочевой
пузырь, из которого идет далее особый мочеиспускательный канал.
Семенники парные, яйцевидной формы, расположенные в полости
тела или в особом мешочке — мошонке. У всех имеется совокупитель-
ный орган втяжной или невтяжной; у многих внутри этого органа обра-
зуется кость (os penis), достигающая особенно больших размеров у ла-
стоногих. Яичники парные, всегда расположенные в полости тела. Верх-
ние отделы яйцеводов впадают в их расширенный отдел — матку. Матки
бывают двойные, двураздельные, двурогие или простые. Оплодотворе-
ние внутреннее.
Все млекопитающие, кроме однопроходных, родят живых детены-
шей. У большинства на месте прикрепления зародыша к стенке матки
образуется плацента, через которую осуществляется связь кровеносной
системы зародышей с кровеносной системой матери. Через плаценту
осуществляется питание развивающихся эмбрионов и вывод из них
неусвояемых организмом продуктов обмена.
В класс млекопитающих объединяется 3 подкласса: 1) яйцекладу-
3 Н. А. Бобринский и др. 33
щие (Prototheria), 2) сумчатые (Metatheria) и 3) высшие млекопитаю-
щие (Eutheria).
Высшие млекопитающие, к которым относятся все современные
виды этого класса, представленные в фауне СССР,— наиболее обшир-
ный из всех пёречисленных подклассов. От двух предыдущих они от-
личаются постоянным отсутствием в стенке брюшной полости сумчатых
костей, обычно сильным развитием плаценты и рождением детенышей,
способных по крайней мере сосать молоко матери.
Распространены высшие млекопитающие почти повсеместно, за
исключением Антарктиды и некоторых небольших высокоарктических и
океанических островов.
Все современные виды высших млекопитающих (общее число ко-
торых около 6 тысяч) группируют в 17—18 отрядов, из которых
в фауне СССР представлены 8 отрядов (около 290 видов), а именно:
Насекомоядных..........около 30
Рукокрылых ................39
Хищных ....................43
Ластоногих.................13
Китообразных................23
Непарнокопытных ............ 1
Парнокопытных ............. 20
Грызунов...............около 120
В это число не входит несколько видов домашних животных, не
встречающихся в диком состоянии (на территории СССР или вообще).
В некоторых отрядах (особенно в отрядах грызунов, насекомоядных и
отчасти парнокопытных) число видов еще не окончательно установлено.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ОТРЯДОВ МЛЕКОПИТАЮЩИХ
ФАУНЫ СССР
1(10). На теле имеются волосы (густые или хотя бы редко рассе-
янные). Задние конечности есть. Форма тела не рыбообразная, и хвост
не в виде двухлопастного плавника. Носовые отверстия открываются
на переднем конце черепа, а носовой проход располагается параллельно
главной оси черепа..........................................2.
2(3). Носовой отдел головы вытянут в виде подвижного хоботка.
Зубные ряды сплошные (без широких беззубых промежутков между
резцами и коренными), причем резцы средней пары (как в верхней, так
и в нижней челюсти) обычно сильно увеличены, но не долотообразной
формы; если же все верхние резцы мелкие, то внешние из них вплот-
ную примыкают к клыкам (мех обычно густой, ровный, бархатистый или
тело сверху покрыто короткими иглами: ежи, выхухоли, кроты, земле-
ройки) .....................Насекомоядные. Insectivora. (Стр. 36.)
3(2). Носовой отдел головы не вытянут в подвижный хоботок.
Зубные ряды с широким беззубым промежутком (между резцами
и коренными); если же ряды оплошные, то средние пары верхних и
нижних резцов не увеличены, а между внешним верхним резцом и
клыком имеется явственный промежуток . . ...................4.
4(5). Передние конечности превращены в крылья; II—V пальцы
передних конечностей сильно удлинены (третий палец длиннее туло-
вища), и на них, как на спицах зонтика, натянута тонкая, почти голая
летательная перепонка; от пальцев она тянется к бокам тела, охватывает
задние конечности и хотя бы часть хвоста или весь хвост. Межчелю-
стные кости черепа недоразвиты; твердое нёбо имеет спереди широкую
и глубокую вырезку; верхние резцы правой и левой стороны поэтому
разделены широким промежутком (средней пары резцов никогда не
бывает)........................Рукокрылые. Chiroptera. (Стр. 79.)
5(4). Передние конечности не похожи на крылья, и любой палец
передней конечности во много раз короче длины туловища. Межчелю-
стные кости нормально развиты без широкой и глубокой вырезки спе-
34
реди; верхние резцы левой и правой стороны (если они есть) сомкнуты
между собой...................................................6.
6(14). В нижней челюсти с каждой стороны по 2 или 3 резца.
Нижние клыки (иногда похожие на резцы) есть. Промежуток между
нижним клыком и предкоренными меньше длины ряда коренных зубов
(вместе с предкоренными). Средняя пара верхних резцов по сравнению
с другими зубами не сильно увеличена или верхних резцов вообще
нет (по внешнему виду животные непохожи на мышь, крысу, суслика,
белку, дикобраза, бобра или зайца)............................7.
7(11). Пальцы оканчиваются когтями, ногтями или совсем лишены
роговых образований. Верхних коренных зубов (вместе с предкорен-
ными) не более 6, причем жевательная поверхность их коническая
или многобугорчатая...........................................8.
8(9). Локтевое и коленное сочленения конечностей отчетливо вы-
ступают на некотором расстоянии от туловища или легко прощупыва-
ются. Передние конечности не в виде плавательных ластов. Заднеко-
ренные и предкоренные зубы резко различны по форме и размерам
.....................................Хищные. Carnivora. (Стр. 117.)
9(8) От туловища выступают только дистальные (концевые) от-
делы конечностей; локтевые и коленные сочленения не видны и не
прощупываются. Передние (как и задние) конечности в виде плава-
тельных ластов. Все коренные зубы (включая и предкоренные) сходны
между собой по форме и мало отличаются по размерам...............
..............................Ластоногие. Pinnipedia. (Стр. 164)
10(1). Тело покрыто голой кожей, лишенной волос. Задних конечно-
стей нет. Форма тела рыбообразная и хвост в виде двухраздельного
плавника. Носовые отверстия открываются наверху черепа, а носовой
проход располагается почти под прямым углом к продольной оси черепа.
...............................Китообразные. Cetacea. (Стр. 180.)
11(7). На концах пальцев копыта. Если верхние коренные зубы
(вместе с предкоренными) с бугорчатой жевательной поверхностью, то
их на одной стороне челюсти 7, а если же меньше 7, то жевательная
поверхность их не бугорчатая и не коническая, а плоская со сложным
рисунком из полосок эмали и заключенных между ними участков ден-
тина ........................................................12.
12(13). На ногах только по одному копыту. Верхние резцы долото-
образной формы (их по 3 с каждой стороны). Все коренные приблизи-
тельно одинаковых размеров ......................................
......................Непарнокопытные. Perissodactyla. (Стр. 206.)
13(12). На ногах по 4 или (реже) по 2 копыта (если 4, то 2 пе-
редних больших и 2 задних поменьше). Верхние резцы или отсутст-
вуют, или имеют не долотообразную форму. Задние коренные явно
крупнее передних .... Парнокопытные. Artiodactyla. (Стр. 209.)
14(6). В нижней челюсти по одному длинному долотообразному
резцу. Клыков (как нижних, так и верхних) нет. Между резцами и пе-
редними коренными беззубый промежуток (диастема), длина которого
больше длины ряда коренных зубов (по внешнему виду животные по-
хожи на мышь, крысу, белку, суслика, дикобраза, бобра или зайца).
....................................Грызуны. Rodentia. (Стр. 236.)
А. П. Кузякин
ОТРЯД НАСЕКОМОЯДНЫЕ. ORDO INSECTIVORA
Насекомоядные — разнообразные по внешнему виду и анатомиче-
скому строению млекопитающие. Размеры — от средних до самых мел-
ких из современных млекопитающих, как карликовая белозубка (Sun-
cus etruscus Savi), длина тела которой не превышает 45 мм и вес 1,2—
1,7 г (в среднем около 1,5 г), или крошечная бурозубка (Sorex minutis-
simus Zimmermann) с длиной тела не более 50 мм и весом от 1,6 до
2,9 г (в среднем около 2 г). Оба эти вида представлены в фауне СССР.
Самые (Крупные из насекомоядных нашей фауны — ежи. Вес лысого ежа
до 1,5—1,6 кг. Мех у большинства мягкий, густой, бархатистый; для
ежей характерен иглистый панцирь. Конечности в большинстве пятипа-
лые стопоходящие, редко полустопоходящие. Пальцы всегда вооружены
когтями. Большой, первый (внутренний) палец не противопоставляется
остальным. Носовой отдел у всех видов фауны СССР вытянут в хобо-
ток, у некоторых весьма подвижный. У многих имеются боковые, цри-
анальные или хвостовые пахучие (муксусные) железы. Сосков от 3
до 6 пар.
Череп с удлиненным носовым отделом и сравнительно небольшой
мозговой коробкой, без затлазничных отростков. Слуховые барабаны
обычно не развиты, представлены лишь плоскими кольцеобразными
пластинками.
Зубная система примитивная: число зубов от 44 до 26. Зубы слабо-
дифференцированы, и в каждом семействе по-разному. Резцы внутрен-
ней (передней) пары у многих (кроме части кротов) сильно увеличены,
но, в отличие от грызунов, имеют не широкую долотообразную режу-
щую поверхность, а сведенную на конус вершину (или 2 вершины) и не
отделены от прочих зубов широким беззубым промежутком. Отсут-
ствие широкого беззубого промежутка можно видеть у зверька, даже
не извлекая черепа, лишь приподняв губы (с боков). Клыки у многих
(кроме части кротов) не увеличены и мало отличаются или совсем
не отличаются от смежных с ними внешних резцов и малых предкорен-
ных зубов. Предкоренные зубы у насекомоядных, как у рукокрылых и
хищников, следует делить на 2 категории: на малые предкорен-
ные (praemolares, сокращенно — pm или при раздельном обозна-
чении Pm — для верхней челюсти и pm — для нижней) и на боль-
шие предкоренные (praemolares prominantes — Pmp и pmp). Большие
предкоренные по одному с каждой стороны в верхней и нижней челю-
сти имеются у всех насекомоядных; они примыкают к первым заднеко-
ренным зубам (molares — М.1 и mi); по размерам они всегда крупнее
смежных с ними 'малых предкоренных, имеют по 2 и по 3 корня, часто
несколько острых вершин, и этим мало отличаются от заднекоренных
36
зубов; по размерам и строению они часто бывают сложнее заднекорен-
ных зубов (М3 и Шз). Специализация зубной системы, сопровождаемая
уменьшением числа зубов (от исходных 44), идет за счет исчезновения
части малых предкоренных, средних и 'внешних резцов, клыков или
даже заднекоренных зубов, но никогда не исчезают большие предко-
ренные зубы. Заднекоренные зубы (molares) насекомоядных имеют до
5—7 острых вершин, часто соединенных режущими гребнями (комис-
сурами); зубы- такого строения называют зубами «насекомоядного
типа».
Насекомоядные — один из самых примитивных отрядов плацен-
тарных млекопитающих. Ископаемые остатки их известны начиная
с верхнемеловых отложений мезозоя. До появления многих из высших
плацентарных насекомоядные были, видимо, господствующим среди
млекопитающих отрядом, отдельные группы которого приспособились
к разным условиям существования. Лишь небольшая часть из них за
счет узкой специализации сохранилась до сих пор. Этим объясняется
крайнее разнообразие «жизненных форм» современных насекомоядных.
Одни из них приспособлены к наземному существованию (ежи, земле-
ройки), другие — к подземно-роющему (кроты), третьи — к наземно-
водному (куторы, выдровые землеройки, выхухоли), есть прыгающие
(наподобие тушканчиков) формы (прыгунчики) и полудревесные (ту-
пайи).
Распространены насекомоядные почти по всей суше земного шара,
кроме Антарктиды, Австралии, некоторых арктических островов и
Южной Америки, где они встречаются только на крайнем северо-
западе материка.
Систематика насекомоядных разработана пока недостаточно. До
настоящего времени описано около 400 видов. Многие из них, вероятно,
окажутся лишь разновидностями. Все они группируются в 10 семейств.
В фауне СССР представлено 4 семейства (около 30 видов).
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ НАСЕКОМОЯДНЫХ
ФАУНЫ СССР
1(2). Тело сверху покрыто твердыми иглами. Всех зубов 36. Верх-
ние резцы внутренней (передней) пары (левый и правый) разделены
широким промежутком (рис. 2, 3). Скуловые дуги хорошо развиты и
широко расставлены, так что скуловая ширина больше ширины черепа
в слуховой области.................Ежовые. Erinaceidae. (Стр. 38.)
2(1). Все тело покрыто мягким густым мехом. Всех зубов не 36,
а 42—44 или 26—32. Верхние резцы внутренней (передней) пары сомк-
нуты между собой. Скуловых дуг совсем нет или они очень тонкие и
узкорасставленные, так что скуловая ширина меньше ширины черепа
в слуховой области..........................................3.
3(6). Длина тела (с головой) более 10 см. Ушных раковин нет.
Череп со скуловыми дугами. Общее число зубов 42 или 44 .... 4.
4(5). Передние лапы (ладони) значительно меньше и уже задних
(ступней). Пальцы задних лап соединены широкими плавательными
перепонками, доходящими до основания когтей (полуводные живот-
ные). Хвост более чем в два раза превышает длину головы; он покрыт
роговыми чешуйками, среди которых рассеяны редкие волосы. Верх-
ние внутренние (передние) резцы во много раз больше расположен-
ных за ними резцов и клыка. Нижние резцы средней пары тоже зна-
чительно крупнее расположенных за ними зубов: внешних резцов, клы-
ков и малых предкоренных . . Выхухолевые. Desmanidae. (Стр. 43.)
37
5(4). Передние лапы (ладони) значительно больше и шире задних
(ступней). Задние лапы без широких плавательных перепонок (роющие,
подземные животные). Хвост не длиннее головы и густо покрыт воло-
сами. Резцы мелкие, и все почти одинаковых размеров. Верхний клык
значительно больше резцов............Кротовые. Talpidae. (Стр. 47.)
6(3). Длина тела (с головой) менее 10 см. Имеются ушные рако-
вины, хотя бы слаборазвитые, в виде кожных, покрытых волосами
лоскутков. Череп без скуловых дуг (рис. 8). Общее число зубов не
более 32 (от 26).............Землеройковые. Soricidae. (Стр. 54.)
СЕМЕЙСТВО ЕЖОВЫЕ. FAMILIA ERINACEIDAE
Тело сверху и с боков покрыто твердыми иглами. Ушные раковины
и глаза хорошо развиты. Скуловые дуги мощные, широко расставлен-
ные (шире мозговой коробки). Верхние передние резцы разделены
между собой широким промежутком. Всех зубов 36.
Распространены в Европе, Азии и Африке.
К семейству ежовых обычно относят ежеземлероек, или гимнур,
в качестве подсемейства Echinosoricinae. От ежей они отличаются
очень резко. Размеры их небольшие (величиной с обыкновенного
крота). По внешнему виду похожи на крупных землероек. Тело по-
крыто грубым мехом, но не иглами. Хвост длинный. Зубная система
примитивная—из 42—44 зубов. Все эти отличия так велики, что еже-
землеройковых правильнее выделять в особое семейство — Echinosori-
cidae.
Виды настоящих ежей обычно группируют в несколько родов и
подродов (Erinaceus, Hemiechinus, Paraechinus, Atelerix и др.). В ка-
честве отличительных признаков между ними приводят: отроение и раз-
меры послесочленового отростка, крыловидных костей-и крыловидной
ямки, строение поверхности игл, расположение игл на темени и т. д.
Изучение коллекционного материала показало, что все эти при-
знаки крайне неустойчивы и не могут быть использованы для разделе-
ния видов настоящих ежей на самостоятельные роды.
Всех ежей фауны СССР мы объединяем в один род и не видим
оснований для разделения его на подроды.
РОД ЕЖИ. GENUS ERINACEUS Linnaeus, 1758
В дополнение к характеристике семейства приведем лишь описа-
ние зубной системы.
Зубная формула: 1-|-Су Pm у Ртру Му-2 = 36.
Резцы внутренней (передней) пары как в верхней, так и в нижней
челюсти сильно увеличены; верхние разделены между собой широким
промежутком, и нижние не вполне сомкнуты друг с другом. Верхние
резцы средней пары самые мелкие. Клыки мелкие (гораздо меньше пе-
редних резцов), по форме похожи на малые предкоренные зубы. Боль-
шие предкоренные (praemolares prominantes) хорошо отличаются от
малых предкоренных своими размерами и более сложным строением.
Верхний Ртр имеет 3—4 вершины и 3 корня; нижний —2—3 вершины
и 2 корня. Из заднекоренных зубов самый крупный первый, а послед-
ние заднекоренные малы и очень простого строения (рис. 1, 3).
Распространены в Европе, Азии и Африке.
38
Населяют различные ландшафтные зоны, от тропических лесов
и пустынь к северу до лесолуговой зоны и речных долин южной и
средней тайги. Летом укрываются в норах и логовах, зимой — в норах,
видимо, всегда закупоренных земляной пробкой. В тропиках активны
круглый год, в умеренной полосе осенью накопляют большие запасы
жира и впадают в спячку. Пробуждаются ранней весной; истощенные
длительной спячкой бывают активными (питаются) почти круглые
сутки, а летом активны главным образом ночью и в сумерки.
Размножаются один раз в году (летом). В приплоде бывает от 2
до 8 (обычно от 4 до 6) детенышей. Родятся они слепыми и голыми;
на спине новорожденных выступают ряды бугорков, из которых уже
в первые сутки жизни вырастают мягкие
белые иглы длиной до 5—6 мм. Прозре-
вают на 15—18-й день. Нижняя сторона
тела покрывается мехом на 20-й день.
Рис. 3. Череп ежа (Е. europaeus dauricus Sund.).
А — сбоку, Б — снизу. Зубы: 1,1 — резцы (Z1 — /3 — верхние; ii — i2 — нижние); С, с—клыки (С — верх-
ний; с —нижний); Рт, рт — малые предкоренные (Ртх, рт2 — верхние; рт' — нижний); Ртр,
ртр — крупные предкоренные (Ртр — верхний; ртр — нижний); М, т — заднекоренные (ЛГ—Л43—
верхние; mi—т3 — нижние).
Питаются ежи преимущественно животной пищей: крупными бес-
позвоночными (дождевыми червями, насекомыми) и мелкими позво-
ночными, главным образом малоподвижными грызунами (полевками),
птенцами и яйцами гнездящихся на земле птиц. Высокоподвижных
грызунов они поймать не могут. Лягушек и змей едят редко или совсем
не едят. Врагов у ежей мало. За счет очень сильно развитой продольной
и поперечно-кольцевой подкожной мускулатуры они могут свертываться
в шар, со всех сторон одетый колючками (иглами), что и служит ос-
новным средством защиты. Из пернатых хищников ежей одолевает фи-
лин, из хищных млекопитающих — лисица. Характерно и важно обилие
паразитирующих на ежах и ксодовых клещей в разных стадиях их разви-
тия. Ежи обладают высокой живучестью, долго могут быть голодными,
слабо реагируют на многие сильные яды (например, на пары хлорпи-
крина), на них почти не действует яд гадюковых змей.
Всего было описано около 40 видов ежей. Фактически их вряд ли
более 10; в пределах Палеарктики — 8, в фауне СССР — 3 близких
вида, различимых лишь по комплексу малоустойчивых признаков. Воз-
можно нахождение четвертого вида, признаки которого включены
в определительную таблицу.
39
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ЕЖЕЙ ФАУНЫ СССР
1(2). Уши короткие: менее 35 мм (редко до 38 мм). Конец сосце-
видного отростка располагается примерно на одном уровне или ниже
конца послесочленовного отростка (рис. 4) .......................
...............Еж обыкновенный. Erinaceus europaeus. (Стр. 40.)
2(1). Уши большие: длина их у взрослых обычно более 35 мм
(до 57 мм). Конец сосцевидного отростка располагается выше конца
послесочленовного отростка (рис. 4) ......................... 5.
3(6). На темени нет большого голого участка («лысины»). Иглы
сравнительно короткие: длина их не более 30 мм. Верхний второй ма-
лый предкоренной зуб с отчетливо развитым внутренним талоном ко-
ронки ........................................................4.
4(5). Размеры не очень крупные: кондилобазальная длина черепа
не более 53 мм; скуловая ширина менее 33 мм. Длина уха менее
5 см; длина тела не более 22 см..................................
. . . . .........Ушастый еж. Erinaceus auritus. (Стр. 42.)
Рис. 4. Соотношение сосцевидных и послесочленовных отростков у ,ежей (вид
сбоку):
/1 — южная форма обыкновенного ежа (Е. europaeus roumanicus Barr. — Ham.); Б, В—
даурская форма обыкновенного ежа (Е. europaeus dauricus Sund.); Г — ушастый еж
(Е. auritus Gmel.).
5(4). Размеры крупнее: кондилобазальная длина черепа более
54 мм; скуловая ширина более 33 мм. Длина уха более 5 см; длина
тела взрослых более 23 (до 30) см..................................
....................Афганский еж. Erinaceus megalotis. (Стр. 43.)
6(3). На темени располагается большой участок голой кожи («лы-
сина»), резко отграниченная спереди мехом, а сбоков и сзади — иг-
лами. Иглы длинные: длина их у взрослых 32—42 мм. Верхний вто-
рой малый предкоренной зуб (примыкающий к большому предкорен-
ному) округлый, без внутреннего талона коронки.....................
....................Лысый еж. Erinaceus hypomelas. (Стр. 43.)
1. Еж обыкновенный. Erinaceus europaeus Linnaeus, 1758
Табл. I, 1—3. Карта 1
Ареал состоит из 2 частей: 1. Европа (от Ирландии и Великобри-
тании на север до Норвегии и Швеции, на юг до островов Средизем-
ного моря), Малая Азия (Палестина, вероятно, Турция и Северный
Иран), Кавказ (с Закавказьем), Северо-Западный Казахстан и За-
падная Сибирь к востоку до Омска, ir. Тары и верховьев р. Васюгана
(левого притока Оби), к северу (в СССР) до оз. Ладожское и Онеж-
ское, до ст. Пундоги (к северу от Вологды), окрестности Ки-
рова, Конды и Васюгана. 2. Восточная Азия, от Забайкалья и Се-
веро-Восточной Монголии до Амуро-Уссурийского края (долины
Среднего Амура, Уссури и их притоков) и южнее расположенных рай-
онов Восточной Азии.
40
Типичный обитатель лиственных и смешанных лесов, лугов, луго-
вых степей и речных долин, по которым проникает в южные окраины
тайги, в степи и в западные полупустыни (до верховьев р. Эмбы). Из-
любленные места обитания — опушки лиственных лесов, светлые рощи,
сады, кустарниковые заросли. В ставропольских степях, например,
более 90% ежей было добыто в лесных полосах и посадках шелковицы
и только около 9%—на целине и в посевах. В подходящих ландшаф-
тах численность их достигает 15—18 особей на 1 км2. Основную пищу
составляют насекомые: в Новгородской обл. (по Б. И. Померанцеву,
1935)—главным образом личинки комаров-долгоножек (Tipulidae),
в Татарии, Чувашии и Ставрополье — жуки, в том числе майские жуки
и волосатые жужелицы (Harpalus ocneus), вредящие проросткам хвой-
ных и злаков. Из чешуекрылых еж поедает гусениц монашенки и не-
парного шелкопряда. Остатки позвоночных встречены в 20% желуд-
ков; большую часть их составляли мышевидные грызуны (полевки).
Еж охотно поедает птенцов и яйца мелких насекомоядных и охот-
ничье-промысловых видов (рябчика, тетерева, фазана), гнездящихся
на земле. Во многих сотнях желудков ежей, добытых в природе, остат-
ков змей не было обнаружено. Прежние утверждения, что ежи пое-
дают ядовитых змей, фактами не подтверждаются. Большой вред ежи
причиняют как основные прокормители иксодовых клещей в очагах
клещевого энцефалита человека, пироплазмоза лошадей и бабезиеллеза
крупного рогатого скота. В очагах клещевого энцефалита в Приморье
автор собирал с одного ежа до 600—650 клещей, преимущественно
главного переносчика вируса — Ixodes persulcaitus — в разных фазах
его развития (личинок, нимф и половозрелых). Иглистый панцирь хо-
рошо защищает присосавшихся между иглами клещей от самого ежа.
Своим иглистым покровом он, как щеткой, собирает на себя клещей,
заползающих на стебли и листья травянистого покрова. Н. Г. Олсуфьев
использовал ежей даже для количественного учета клещей и предло-
жил стандартную единицу учета клещей — «ежечас».
Еж подвержен широкой индивидуальной и географической изменчивости. В пре-
делах СССР можно выделить 4 подвида, различающихся главным образом по окраске
иглистого и волосяного покрова:
а) Е. е. europaeus Linnaeus, 1758. (Описан по экземпляру из Южной Швеции.)
Все иглы по окраске относительно однородные: вершины их светлые (беловатые),
ниже — темно-бурый или черный участок, еще ниже — светлый желГговато-белесый
участок, у основания иглы — темно-бурый поясок. Окраска меха на нижней стороне
тела обычно относительно светлая, серовато-белесая; особенно светлые волосы
на горле, груди и брюхе. Ухо обычно короче 30 мм. Распространен от Балтийского
побережья на юг до Чехословакии, Южной Украины и Нижнего Поволжья, на восток
до побережья Иртыша и верховьев р. Васюгана. К этому же подвиду мы относим
очень слабо различающиеся племена (natio), выделяемые некоторыми систематиками
в ранг подвидов: Е. е. centralrossicus Ognev, 1926 (тип из бывшего Сычевского уезда
Смоленской губернии), и Е е. с. natio pallidus Ognev, 1928 (тип с р. Тавды Тюмен-
ского района и области).
b) Е. е. concolor Martin, 1838. (Тип из окрестностей Трабзона, Северо-Восточная
Турция.) Окраска игл, как у номинальной формы или с большим развитием темных
поясков. Окраска меха наверху головы, боках и на брюхе темно-бурая, резко кон-
трастирующая с более светлЬ окрашенными низом головы и груди. Отличия этой
формы от позднее описанной Е. е. roumanicus Barret-Hamilton, 1900 (по типу из
Румынии) остаются недостаточно выясненными и, вероятно, недостаточными для их
разделения как особых подвидов. В Чехословакии эта форма на большой территории
встречается совместно с номинальной, и чехословацкие зоологи (в частности, И. Кра-
тохвил) считают Е. roumanicus В.-Ham. отдельным видом. По материалам с террито-
рии СССР для разделения этих подвидов в особые виды нет достаточных оснований.
Е. г. transcaucasicus Satunin, 1905 (по типу из Ордубата, на Араксе) и Е. г. abasgicus
Satunin, 1907 (с верхнего и среднего течения р. Кодори, Абхазия) даже С. У. Строга-
новым (1957) сведены в синонимы Е. е. roumanicus В.-Ham. Возможность их подви-
дового отличия от Е. е. concolor Martin представляется нам исключенной. Чехослова-
41
кия, Румыния, Венгрия, Болгария, Южная Украина, Нижнее Поволжье, Западный
Казахстан, Кавказ, Закавказье, Турция.
с) Е. е. dauricus 1 Sundevall, 1842. (Тип из Даурии, Забайкалье.) Размеры и
окраска, как у номинальной формы. Уши немного больше, у взрослых особей обычно
от 30 до 38 мм. По деталям строения черепа некоторые особи похожи на ушастых
ежей, но эти отличия весьма неустойчивы и недостаточны для выделения даурских
ежей в ранг особого вида. Степи Южного и Юго-Восточного Забайкалья, северо-
восточных районов Монгольской Народной Республики, Внутренней Монголии и
смежных с ней районов.
d) Е. е. amurensis Schrenk, 1859. (Тип с Агуна на Амуре.) Среди нормально окра-
шенных пестрых игл со светлыми и буроватыми поясками около 30% игл одноцвет-
ных, желтовато-белесых. Окраска меха на нижней стороне тела у старых особей
сравнительно светлая, палево-сероватая, у средневозрастных землистая с расплывча-
тым светлым пятном на груди и брюхе. Долины рек Амура (в его среднем отрезке),
Уссури и Сунгари с их притоками.
2. Ушастый еж. Erinaceus auritus Gmelin, 1770
Табл, I, 4—5, Карта 2
Степи от берега Таганрогского залива Азовского моря и среднего
отрезка Северного Донца, Северного Кавказа, Поволжья (к северу
до г, Пугачева и с, Ферапонтовки; западносибирские степи от верхо-
вий р. Тобола до предгорий Алтая и к северу до г, Петропавловска и
Семипалатинска; полупустыни долины Аракса, Апшеронского полу-
острова и Прикаспия; полупустыни и пустыни всего Казахстана и
Средней Азии, предгорья и долины рек Тянь-Шаня, За пределами
СССР от ОАР, Сирии и Кипра через Малую Азию, Иран, Афганистан,
Северную Индию, полупустыни и пустыни Южной Монголии к востоку
до сухих степей Внутренней Монголии,
Оптимальные условия — в бугристых, всхолмленных и предгорных
пустынях и полупустынях; менее типичны степные ландшафты, В став-
ропольских степях придерживается зарослей кустарников и полезащит-
ных полос. Летом укрывается в норах, вырытых самим или крупными
грызунами и черепахами. Активен ночью и в сумерки. Очень подвиж-
ный: бегает быстрее обыкновенного ежа. Неохотно свертывается в клу-
бок. Питается преимущественно насекомыми, особенно жесткокрылыми.
Позвоночных (ящериц, жаб, мелких птичек) ест редко и в небольшом
количестве. Размножается поздно: в ставропольских степях в июле,
В приплоде бывает от 2 до 8 (в среднем около 6) детенышей. Прокор-
митель множества иксодовых клещей.
Подвержен очень широкой индивидуальной и географической из-
менчивости, что вызвало описание большого количества видов и подви-
дов, реальность которых (особенно видов) не подтверждается факти-
ческим материалом, Из 8 видов ушастых ежей, указывавшихся разными
авторами для фауны СССР, ни один не имеет отличий, достаточных
для обособления от Erinaceus auritus Gmelin, На сериях экспонатов
можно различить следующие подвиды:
а) Е. a, auritus Gmelin, 1770. (Описан из окрестностей г. Астрахани.) Окраска
темная: самые кончики иголок бурые, ниже — беловатый предвершинный участок,
затем — широкий темно-бурый или черный пояс и светлое желтовато-охристое осно-
вание. Мех на лбу, щеках и на боках тела темный, палево-бурый. Иглы относительно
короткие: 17—22,5 мм. Длина уха 29—47 мм. Синоним этой формы: Е. a. minor
Satunin, 1907. (Из окрестностей Барнаула.) Степи Восточной Украины, Северного
Кавказа, Северного Казахстана и Западной Сибири до предгорий Алтая.
b) Е. a. calligoni Satunin, =1901, (Тип из песков Аралых, у подножия горы Арарат,
Южная Армения.) Окраска игл и меха светлее; вершинки игл светлые; темные
1 В первоописании — Е. dauuricus; второе «и» в этом слове, видимо, неисправ-
ленная опечатка.
42
пояски уже и бледнее. Мех на лбу и щеках рыжевато-палевый. Размеры тела, ушей,
черепа, как у номинальной формы. Иглы тоже короткие: 16—23 мм. Синонимы этой
слабодифференцированной формы: Е. a. brachyotus Satunin, 1908 (тип из Ахсу
Шемахинского района, Азербайджан) и Е. a. turkestanicus Ognev, 1928 (тип со ст.
Кара-Узяк, к северу от Кызыл-Орды, Казахстан). Полупустыни Южной Армении,
Восточного Азербайджана, Прикаспия; полупустыни и северные пустыни Казахстана.
с) Е. a. albulus Stoliczka, 1872. (Описан из Яркенда, Кашгария.) От двух преды-
дущих форм отличается более длинными иглами (26—28 мм) и очень светлой
окраской густого мягкого меха (снизу и на ушах чисто белого). По размерам
близок к предыдущим (чуть крупнее). Широко распространен в полупустынях и
пустынях Южной Монголии. В СССР — в пустынях Средней Азии и в Тувинской
области у оз. Тере-Холь. К этому же подвиду следует отнести: Е. a. persicus
Satunin, 1907 (тип из Гуляндара, Иран), Е. a. major Ognev, 1928 (тип из Аннау, близ
Ашхабада, Южная Туркмения) и Е. a. insularis Timofejew, 1934 (тип с о-ва Барса-
Кельмес, на Аральском море).
Афганский еж. Erinaceus megalotis Blyth, 1845
Один экземпляр этого вида был добыт, и по нему был описан осо-
бый вид — Hemiechinus chorassanicus М. К. Лаптевым (1926). Экземп-
ляр этот не сохранился. Осталось только его описание, по которому
можно предполагать, что он относится к Е. megalotis Blyth, 1845, опи-
санному из окрестностей Кандагара (Афганистан). От ушастого ежа
этот вид отличается очень крупными размерами: длина тела 265—
300 мм, очень большими ушами: длина уха 52 мм; общая длина че-
репа 57,4 мм; скуловая ширина 34,4 мм; длина верхнего ряда зубов
27,8 мм. Длина иглы 34 мм. Каждая игла с тремя темными поясками.
С. У. Строганов (1957) пишет, что в окраске игл Н. megalotis бывает
и «две темные зоны».
Афганистан, Белуджистан, Юго-Западная Туркмения (долина
р. Чандыра, в Западном Копет-Даге).
3. Лысый еж. Erinaceus hypomelas Brandt, 1836
Табл. I, 6—7. Карта 3
Устюрт, Южный Узбекистан (к северу до Самаркандской обла-
сти), Афганистан, Аравия, Пакистан (Синд).
Живет в предгорных и горных пустынях, песчаных и глинистых,
в садах селений; в горы поднимается до пояса арчи. Днем укрывается
в норах. Во второй половине мая — первой половине июня самки ро-
дят по 3—4 детеныша. Образ жизни изучен очень слабо.
В СССР обитает номинальная форма: Е. h. hypomelas Brandt,
1936, описанная из Северного Ирана. Две морфы: одна — светло окра-
шенная с иглами желтовато-белесого цвета, другая — темная, черно-
ватая, необычная для пустынных условий.
СЕМЕЙСТВО ВЫХУХОЛЕВЫЕ. FAMILIA DESMANIDAE
Тело яйцевидной формы, заостренное в сторону головы и покры-
тое очень густым, ровным и мягким мехом, у здоровых зверьков нена-
мокающим. Задние лапы значительно больше и шире передних. Как
короткие пальцы передних, так и длинные изогнутые пальцы задних
лап соединены плавательными перепонками до основания когтей.
Длина тела от 12—15 до 18—22 см. Хвост длинный, почти равный
длине тела, покрытый роговыми чешуйками, среди которых разбросаны
короткие редкие волосы. По гребню хвоста тянется киль из относи-
тельно густых и длинных волос. У самого основания хвост «перетя-
нут» (имеет наименьший диаметр), в первой трети длины грушевидно
43
утолщен. В утолщении расположены мускусные железы, маслянистый,
сильно пахнущий мускус которых выдавливается через многочислен-
ные отверстия, расположенные на нижней стороне хвоста (им смазы-
вается меховой покров). Средняя и концевая части хвоста уплощены
с боков и служат локомоторным органом и рулем поворота при дви-
жении выхухоли в воде. Ушных раковин нет. Глаза очень маленькие
(с булавочную головку). Зрение почти атрофировано. На свет выхухоль
реагирует, но предметов, видимо, не различает. Хоботок, несколько рас-
ширенный на конце, выполняет, видимо, функцию осязания. Обоня-
ние развито слабо (Г. А. Скребицкий, 1945).
Череп длинный (до 50—58 мм), гребнистый. Тонкие скуловые дуги
узко расставлены (уже мозговой капсулы). Зубная система исходная
(из 44 зубов).
Распространены выхухолевые только в Европе, на Пиренейском
полуострове и в бассейнах южных рек Восточной Европы.
К 'семейству относятся два 'современных монотипических рода: Des-
mana и Galemys. В фауне СССР — один род.
РОД ВЫХУХОЛИ. GENUS DESMANA Giildenstaedt, 1777
Размеры относительно крупные: длина тела 18—22 см', хвоста
17—20 см. Общая длина черепа 53—58 мм. Вес 280—520 г.
Зубная формула: I у С у Рш у Ршр-рМ —2 =44.
Верхние резцы внутренней пары сильно увеличенные, треугольные,
с режущими боковыми краями, не разделенные между собой. Мелкие
резцы второй и третьей пар располагаются в одном ряду с небольшими
клыками и коренными. Нижние резцы внутренней пары малы, а резцы
второй пары крупные, направленные под углом вперед. Нижний клык
такой же -миленький, как резец внешней -пары и первый малый предко-
ренной. Большие предкоренные (как нижние, так и верхние) хорошо
отличаются от малых предкоренных крупными размерами и более
сложным строением. Заднекоренные многобугорчатые; задний из них
несильно уменьшен и тоже с многими бугорками.
Бассейны южных рек Восточной Европы.
Один вид.
4. Выхухоль. Desmana moschata Linnaeus, 1758
Табл. II, 1. Карта 4
Поймы малых рек всего бассейна Дона и среднего отрезка Волги
от Ярославля до Саратова (ниже Саратова крайне редка). В меньшем
количестве — в пойме нижней половины р. Урала и реакклиматизиро-
вана на разных отрезках Днепра.
Наиболее благоприятны для жизни выхухоли замкнутые поймен-
ные водоемы типа стариц с площадью водного зеркала более 0,1 —
0,5 га и средней глубиной от 1,3 до 5 м, с участками невысоких, но су-
хих обрывистых берегов и с водной растительностью. Очень благопри-
ятно расположение водоема близ коренного берега или пойменного
леса. Обилие выхухоли на отдельных отрезках пойм средних рек (Оки,
Цны, Клязьмы и др.) примерно пропорционально обилию замкнутых
пойменных водоемов при малых расстояниях между ними. В наиболее
густо заселенных участках поймы р. Хопра (по М. Г. Неемченко-Хит-
ровой, 1954) на 1 км береговой линии насчитывалось от 7 до 20 зверь-
44
ков. Большую часть года живет в ‘.норах, вырытых самостоятельно или
связанных с ворами бобра. Обычно нора имеет один выход, располо-
женный ниже поверхности воды, так что начальный отдел норы бывает
затоплен и несколько размыт. Основная часть хода, расположенного
над уровнем воды, тянется почти «горизонтально на 2,5—3 м и имеет
до 2—4 расширений (камер) с выстилкой (гнездом) из растительных
материалов, собранных на дне водоема. Выходов на сушу из обитаемых
нор никогда не бывает. В период половодья норы затопляются,
и зверьки в это время укрываются на полузатопленных деревьях, на-
носах или в неглубоких временных норах, вырытых в незатопленных
Рис. 5. Череп выхухоли (Desmana moschata L.).
А — сбоку, Б — сверху, В — снизу. Зубы: /, i — резцы; С, с —
клыки; Рт, рт — малые предкоренные; Ртр, ртр — крупные
предкоренные; М, т — заднекоренные (/’—/3, С, Рт'—Рт\
Ртр, М'—М3 — верхние; ij—i3< с, pmx—pmZt ртр, т}—т3 — ниж-
ние).
45
частях коренного берега. На дне водоема перед входом в жилую нору
всегда образуется хорошо выраженная дорожка, прорезывающая всю
толщу ила (до песчаной основы). Во время кормежки выхухоль при-
держивается этих придонных путей. Нормальная задержка ее под во-
дой обычно не превышает 1 «минуты (.максимальная до 4 минут). За
это время набранный в легкие воздух выхухоль постепенно выпускает
в виде небольших пузырьков. Мелкие пузырьки воздуха выходят так-
же из мехового покрова (под давлением воды). Зимой пузырьки воз-
духа скопляются над траншеей под «нижней поверхностью льда, посте-
пенно вмерзая в него в виде пустот разной величины; лед над тран-
шеей становится пористым и непрочным. При опасных для жизни
выхухоли ранних паводках лед ломается прежде всего по линиям
с высокой пористостью (над траншеями), где образуются трещины,
через которые зверьки уходят от затопления.
Летом выхухоли живут поодиночке, парами и семьями, а зимой
в одной норе «может жить до 12—13 зверьков разного пола и возраста.
Каждый зверек или их скопление имеет целую группу временно посе-
щаемых нор, расположенных на расстоянии 25—30 м одна от другой.
Такое расстояние выхухоль проплывает за нормальный срок ее пре-
бывания под водой — за 1 минуту (Л. В. Шапошников, 1936).
Питается выхухоль малоподвижными «и неподвижными формами
пресноводных беспозвоночных. Летом основное значение в питании
выхухоли имеют брюхоногие моллюски, личинки насекомых (особенно
ручейников и жука-радужницы) и пиявки. Зимой существенную при-
месь составляют мелкие рыбы и растительные корма: крахмалонос-
ные корневища кубышки, ежеголовки и клубни стрелолиста. За счет
пузырьков воздуха над донными траншеями выхухоли создаются усло-
вия лучшей аэрации, привлекающие сюда моллюсков, пиявок и мел-
ких рыб. Привлекающее действие на водных беспозвоночных оказывает,
видимо, и мускусный след вдоль траншеи. Выхухоль не рыщет по дну
всего водоема в поисках пищи, а плывет только по системе своих тран-
шей, к которым поедаемые животные сами активно стягиваются.
В период весеннего половодья зверьки соединяются парами; в тихие
пасмурные дни этого периода они (обычно молчаливые) издают свое-
образные звуки. Срок беременности 45—50 дней. Число детенышей
в весеннем приплоде от 3 до 5. В ноябре — декабре (до января) по-
является молодняк второго (в году) приплода от части взрослых са-
мок, родившихся весной, и, видимо, части самок весеннего приплода.
Число детенышей в зимнем приплоде от 1 до 5 (в среднем менее 3).
Слепой, голый, беспомощный детеныш весит около 2—3,3 г (почти
вдвое меньше новорожденного крысенка). В июне — июле , (в период
воспитания молодых весеннего выводка) взрослые выхухоли линяют.
Вторая линька взрослых и первая молодых (весенней генерации)
бывает в сентябре — октябре. Живут выхухоли в природе более 4 лет.
Мех выхухоли очень густой, плотный, шелковистый; шкурка проч-
ная и используется как ценное меховое сырье в натуральном виде или
для одной из лучших имитаций под котика. Однако ареал этого вида
невелик, благоприятных пойменных водоемов мало, численность и пло-
довитость невелики, поэтому и промысловое значение вида мало. На
снижение его 'численности действуют зимние подъемы воды и слишком
ранние (подледные) паводки, отсутствие древесной и кустарниковой
растительности по берегам, сильные засухи, но особенно рыболовные
снасти типа вентерей, в которые выхухоль легко попадает и гибнет.
В период засух при переселениях зверьков из высохших водоемов они
становятся жертвой пернатых хищников. Хищные звери (включая до-
46
машних собак) выхухоль не едят (из-за мускуса), но при встрече
душат ее.
В толще -плотного мехового покрова выхухоли образуется на-
столько постоянная .воздушная прослойка, что на выхухоли паразити-
рует маленький жук—Silphopsyllus desmanae, не имеющий приспособ-
лений для дыхания в воде.
Уровень развития высшей нервной деятельности можно пояснить
одним примером. Зверьков, добытых для пересадки в другие водоемы,
вначале не знали, как накормить: положенную в клетку рыбу они не
видели и не обнаруживали с помощью слабого обоняния. Выхухоль
брали за хвост, подносили к лежащей на дне клетки рыбе; когда она
касалась тела рыбы хоботком, то немедленно начинала ее поедать, не
обращая внимания на свое положение: ее держали за хвост навесу без
какой-либо опоры.
Таковы некоторые особенности образа жизни этого замечательного
«живого ископаемого» — эндемика русской природы.
СЕМЕЙСТВО КРОТОВЫЕ. FAMILIA TALPIDAE
Кротовые — относительно мелкие подземные (роющие), реже, на-
земные (бегающие) насекомоядные, по внешнему виду похожие на
широко известного -обыкновенного крота или на землероек. Ушных ра-
ковин у настоящих кротов нет. Глаза очень малы и у некоторых скрыты
под кожей. Кистевой отдел передней конечности широкий, плоский,
лопатообразный, окаймленный рядом жестких волос; когти длинные,
расположенные в плоскости вывернутых наружу ладоней. Ступни зна-
чительно уже ладоней. Разный по длине хвост не уплощен с боков и
обычно густо покрыт волосами. Мех мягкий, ровный, густой, бархати-
стый, однообразного черного, темно-серого или коричневатого цвета.
Мощная ключица непосредственно сочленяется с плечевой костью.
Таз длинный, узкий, без вентрального симфиза. Два крестцовых и три
хвостовых позвонка срослись между собой и с подвздошной костью;
сросшиеся остистые отростки этих позвонков образуют гребень. Череп
длинный, конический, со слаборазвитыми гребнями. Скуловые дуги
очень тонкие и узко расставленные, так что скуловая ширина меньше
ширины мозговой капсулы. Зубов от 44 до 34. Как число, так и строе-
ние зубов имеет большое значение в диагностике подсемейств и родов
кротовых.
Распространены в Европе, Азии и Северной Америке.
По многим признакам кротовые занимают промежуточное положе-
ние между выхухолевыми и землер-ойковыми, хотя обособились они
уже с начала третичного периода. Около 30 современных видов кро-
товых группируются в 4 подсемейства:
1) Talpinae (4 рода, около 13—15 видов); 2) Condilurinae (1 род,
1 вид); 3) Scalopinae (7 родов, 11 видов) и 4) Uropsilinae (3 рода,
3 вида). Наименее специализированные из них к подземному (рою-
щему) образу жизни имеют наиболее специализированную зубную си-
стему и большее сходство с землеройковыми (Soricidae). В фауне
СССР представлено -одно подсемейство: Talpinae.
Подсемейство НАСТОЯЩИХ КРОТОВЫХ. Subfamilia TALPINAE
Внешний вид типично кротовый. Ушных раковин нет. Ладони ло-
патообразные, значительно шире ступней. Звездообразного кожистого
нароста на носу нет. Хвост короткий (длина его не превышает длины
головы), густо покрытый волосами. Зубов 44—40. Верхние резцы мел-
47
кие и почти одинаковые по размерам и форме. Верхние клыки боль-
шие, конические, резко отличные по размерам и форме от смежных
с ними резцов и 'малых предкоренных. Нижние клыки -мало отличаются
по размерам и форме от мелких и ровных нижних резцов. Форму клыка
в нижней челюсти приобрел крупный по размерам первый малый пред-
коренной зуб. Большие предкоренные (Ртр и ртр) составляют особую
группу зубов.
Распространены в Европе и Азии.
Кроты — подземные животные. Они всю жизнь проводят в темных,
не связанных с поверхностью ходах, проложенных ими в разных гори-
зонтах почвы. По расположению ходы бывают разных типов. В рыхлой
и влажной лесной почве прокладываются так называемые «поверхност-
ные» ходы, расположенные на глубине 2—5 см\ при их прокладке крот
поднимает «потолок» хода в виде заметного снаружи земляного ва-
лика. Выброшенных на поверхность кучек земли (кротовин) не бы-
вает. В местах с часто и сильно подсыхающей почвой прокладываются
«глубинные» ходы, расположенные на глубине 10—50 см от поверхно-
сти. Слой земли такой мощности крот поднять не может и поэтому
избыток земли выбрасывает через временные отнорки на поверхность
в виде характерных кучек — кротовин. Отнорки потом закупориваются
земляной пробкой. Бывают еще системы кольцевых и радиальных хо-
дов вокруг гнездовой камеры^ расположенной на глубине до 1—2 м.
Кротовина над системой пригнездовых (кольцевых и радиальных хо-
дов) бывает особенно большой. Питаются кроты почвенными беспозво-
ночными, особенно дождевыми червями, и в меньшей степени — насе-
Рис. 6. Черепа кротов:
А, а — алтайский крот (Talpa altaica Nik.); Б — большая могера (Mogera robusta Nehr.). Зубы
/3. h—i'3 — резцы; С, с — клыки (у могеры нижнего клыка нет); РпР—Рт\ рт{—рт3—малые
предкоренные; Ртр, ртр — крупные предкоренные; ЛР—ЛР, т}—т3 — заднекоренные зубы.
48
комыми и их личинками. В зимнюю спячку не впадают. Размножаются
1—2 раза в году. В каждом (приплоде бывает от 3 до 10 детенышей,
голых, беспомощных. Растет .молодняк очень быстро.
Мех кротов ровный, густой, бархатистый, со слабо выраженным
ворсом. Шкурки средних и крупных видов кротов используют как
пушно-'меховое сырье. Качество их меха снижается в период линек.
У взрослых кротов бывает 3 линьки в году: весенняя, неполная (ком-
пенсаторная) летняя и полная осенняя.
В это подсемейство объединяют 4 рода (13—15 видов): Talpa (sin.
Euroscaptor, Eoscalops и Asioscalops; всего около 7—9 видов), Mogera
(3 вида), Parascaptor (2 вида) и Scaptochirus (1 вид). В фауне СССР
представлено 2 рода: Talpa и Mogera.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ КРОТОВЫХ ФАУНЫ СССР
1(2). Всего зубов 44; в нижней челюсти—11 + 11; есть нижние
клыки, примыкающие к резцам и похожие на них по размерам ....
........................................Кроты. Talpa. (Стр. 49.)
2(1). Всего зубов 42; в нижней челюсти всех зубов 10+10; ниж-
них клыков нет (крупные зубы, похожие на клыки, — первые предко-
ренные) ...............................Могеры. Mogera. (Стр. 53.)
РОД КРОТЫ. GENUS TALPA Linnaeus, 1758
Зубная формула: IyC -|-Pm-|- Pmp уМ-|--2 = 44.
Нижние клыки входят -в группу нижних резцов и мало отличаются
от последних чуть большей высотой и конической формой коронки.
Широко распространены в Евразии.
В роде Talpa (в принятом нами объеме) без родов Mogera, Paras-
captor и Scaptochirus можно различить 7 видов, из которых в фауне
СССР представлены 3; положение двух или трех форм остается не-
окончательно установленным.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ КРОТОВ РОДА TALPA
ФАУНЫ СССР
1(2). Размеры мелкие: наибольшая длина черепа 27—29,2 мм;
кондилобазальная длина 26,5—28,5 мм; ширина носового отдела че-
репа (в области верхних клыков) 2,9—3,8 juju; длина верхнего ряда трех
заднекоренных зубов 4,8—5,5 мм. Вес не более 30 г (в одном случае —
34,5 г, в другом — 39,2 г).......................................
......................Малый крот. Talpa hercegoviensis. (Стр. 50.)
2(1). Размеры крупнее: наибольшая длина черепа более 29 мм
(почти всегда более 29,4 мм); кондилобазальная длина черепа не
менее 28,8 мм; длина верхнего ряда трех заднекоренных зубов не ме-
нее 5,5 juju. Вес больше 32 г..................................3
3(4). Зубы относительно крупные: длина максиллярного ряда зу-
бов более 35% кондилобазальной длины черепа. Длина основания
коронки верхнего клыка менее чем в 2 раза превышает продольный
диаметр средней части того же зуба. Длина хвоста не менее чем на
8 мм превосходит длину ступни (без когтей).......................
......................Крот обыкновенный. Talpa europaea. (Стр. 50.)
4(3). Зубы относительно мелкие: длина максиллярного ряда зубов
менее 35% кондилобазальной длины черепа. Длина основания коронки
4 Н. А. Бобринский к др.
49
верхнего клыка более чем в 2 раза превышает продольный диаметр
средней части того же зуба. Длина хвоста менее чем на 8 мм превы-
шает длину ступни (без когтей)........................................
..........................Алтайский крот. Talpa altaica. (Стр. 52.)
5. Малый крот. Talpa hercegoviensis Bolkay, 1925
Табл. II, 2. Карта 5
Западный Кавказ: верховья р. Белой и ее притоков (Курджипс,
Киша), среднее течение р. Бзыч, южные склоны горы Дудугуш, север-
ные склоны горы Черный Шахан, перевал Псеашхо, 2100 мм над у. м.;
Западная Грузия, 24 км к юго-западу от Зугдиди (Н. К- Депарма,
1961); лагерь Холодный в Кавказском заповеднике; Зеленый мыс,
близ Батуми.
Живет в широколиственных лесах, на субальпийских и альпийских
лугах Западного Кавказа. Места его обитания вкраплены в места
обитания кавказской формы обыкновенного крота. Численность его
обычно ниже последнего, но местами (например, в высокогорье у исто-
ков р. Желобной и в 1963 г. у лагеря Холодного в Кавказском заповед-
нике) малый крот численно преобладал над обыкновенным (кавказ-
ским). Ходы малого крота отличаются малым диаметром (2,1—2,9 cjh);
ходы поверхностные, без кротовин. Обычно кроты бегают по своим хо-
дам, но в июне — июле (в период расселения молодняка) изредка попа-
дают и в ходы обыкновенного крота (диаметр которых 3,5X5,5ел). Все
ходы малого крота встречались только в рыхлой (лесной и луговой)
почве и обычно поблизости от воды. Биология размножения остается
совсем не изученной. Среди самок, добытых с мая до июля, не было
ни одной беременной. В мазках из придатков семенника самцов, добы-
тых за тот же период и даже у добытого 3 апреля, сперматозоиды не
обнаружены. В желудках малых кротов преобладали личинки и ку-
колки насекомых, особенно бабочек (в частности, совок), и много-
ножки; дождевых червей поедают в меньшем количестве, чем насеко-
мых. В желудках Уз—V* малых кротов встречены паразитические
черви (Spirura talpae и Soboliphime hirundiformes). Из эктопарази-
тов в небольшом количестве обнаружены блохи: Palaeopsylla caucasi-
са и Hystichopsylla satunini.
Известны 2 подвида:
а) Т. h. herzegoviensis Bolkay, 1925. (Тип из Герцеговины, Югославия.) Размеры
относительно крупные: кондилобазальная длина черепа 27,2—28,5 мм; наибольшая
ширина черепа 13,9—14,5 мм; ширина носового отдела 3,3—3,8 мм; максиллярный
ряд зубов 10,5—11,5 мм. Югославия. Один взрослый самец, добытый нами 27/IX
1958 г. на Зеленом мысе, близ Батуми, по размерам не отличается от этой формы.
b) Т. h. minima Deparma, 1959. (Тип из окрестностей пос. Хамышки, близ северной
границы Кавказского заповедника.) Размеры меньше: кондилобазальная длина че-
репа 26,5—27,5 мм; наибольшая ширина черепа 12,4—14,0 мм; ширина носового
отдела 2,9—3,4 мм; максиллярный ряд зубов 8,5—10,4 мм. Западный Кавказ (пос.
Хамышки, разные участки Кавказского заповедника); Западная Грузия, близ Зуг-
диди; ст. Приближная в Кабардино-Балкарии.
6. Крот обыкновенный. Talpa europaea Linnaeus, 1758
Табл. II, 3. Карта 6
Западная Европа от Британии, Южной Швеции и Южной Финлян-
дии к югу до Испании, Италии, Сицилии, Северной Греции. В СССР —
Европейская часть, Урал и Западная Сибирь к востоку до левых при-
токов Нижней Оби и правых притоков Иртыша. По долинам рек да-
50
леко заходит в зону тайги (до Архангельска, Мезени, устья Печоры
и Березова, на Оби). На юг ареал идет до широты Кишинева, в Мол-
давии, до Кировограда, Днепропетровска и Луганска, на Украине, до
Славянска, Воронежа, Саратова, Куйбышева, Оренбурга, Тюмени,
Ишима и Омска. Другая (изолированная) часть ареала расположена
от устья Дона и восточных берегов Азовского моря через все Западное
Предкавказье и северные склоны Большого Кавказа, а на юг по Чер-
номорскому побережью до Батуми и, вероятно, уходит в Северо-Во-
сточную Турцию.
Крот — типичный обитатель лесостепья, широколиственных и сме-
шанных лесов, а также долин рек с лугами и лиственными насажде-
ниями в них, по которым он к северу проникает до средней тайги, а
к югу — до типичных степей. В сухих степях, полупустынях, пустынях,
в водораздельных ландшафтах тайги, в лесотундрах и тундрах крот
не живет или встречается крайне редко (в тайге и степях). В лесных
ландшафтах с рыхлой и влажной почвой он прокладывает сложные
системы поверхностных ходов (без кротовин), а в осветленных лесах
и открытых ландшафтах преобладают глубинные ходы. Ходы отдель-
ных кротов и семей соединяются в сложнейшие системы. В лесных
ландшафтах, особенно густо заселенных кротами, сложные системы
поверхностных ходов соединяются нечастыми глубинными переходами,
проложенными в уплотненной земле под лесными тропами и дорогами.
Такими переходами пользуются (как подземными «мостами») кроты
многих семей и ремонтируют эти переходы в случае их разрушения.
Глубинные отрезки ходов под лесными тропами и дорогами служат
основными местами промысловой расстановки ловушек (кротоловок).
В одном переходе за неделю ловят до 10 и более кротов. В широко-
лиственных лесах (Тульских засеках) летом насчитывалось до 200—
225 кротов на 1 км2 старолесья.
Размножение изучено слабо. Спаривание бывает в марте—апреле.
Период беременности точно не установлен (по разным авторам от
30—32 до 50—60 дней). Детеныши в числе 3—9 (в среднем около 4)
родятся с конца апреля до первой половины июня. Изредка беремен-
ные самки встречались и летом. В месячном возрасте молодняк по
размерам уже не отличается от родителей (крупный молодняк отли-
чают от старых по землистой окраске ладоней). Половое созревание
самок наступает лишь к следующей весне (в возрасте примерно 10 ме-
сяцев).
Важный промысловый вид: ежегодно добывают около 20 млн. кро-
тов. По товарному выходу в общесоюзных заготовках крот занимал
6—7-е место, а в средней полосе Европейской части СССР и Урала —
1-е место. Качество большей части заготовляемых шкурок снижено из-
за линек (весенней, летней и осенней), протекающих почти во весь
промысловый сезон. Лучшее качество меха и мездры бывает поздней
осенью и зимой (после полной осенней линьки), когда промысел уже
прекращается. В морозные малоснежные зимы отмечалась массовая
гибель (вымерзание) кротов (в частности, на Урале).
Реально можно различить следующие подвиды:
а) Т. е. europaea Linnaeus, 1758. (Описан из окрестностей г. Кристианшада, Южная
Швеция.) У большинства особей имеется зрительное отверстие длиной около 0,5—1 мм.
(Настоящих подвижных век и ресниц у крота нет.) Описанные рядом авторов под-
виды: Т. е. brauneri Satunin, 1908 (тип из Чегульского поста Белецкого района Мол-
давии), Т. е. uralensis Ognev, 1925 (тип из Пермского района), Т. е. obensis Scalon
et Raewski, 1940 (тип из окрестностей пос. Шухтункурт, в среднем течении р. Малой
Сосьвы, притока Оби), Т. е. transuralensis Stroganov, 1956 (тип из окрестностей дер.
Соровой, в среднем течении р. Демьянки Уватского района Тюменской области) —
практически неразличимы.
4*
51
b) T. e. caucasica Satunin, 1908. (Тип из окрестностей г. Ставрополя Кавказского.)
Глаза у подавляющего числа особей прикрыты тонким слоем кожи (изредка бывает
небольшая прорезь). Других устойчивых отличий от европейско-сибирской формы
нет. Коренные зубы у кавказского подвида несколько крупнее, чем у европейских и
особенно западносибирских кротов. Закавказского «крупнозубого» крота, иногда
выделяемого в качестве особого вида (Т. romana Thomas, 1902), мы относим к этой
же форме. Размеры, как у номинальной формы: наибольшая длина черепа 32,5—
36,0 мм; кондилобазальная длина черепа 33,1—35,7 мм; ширина носового отдела 4,5—
5,0 мм} максиллярный ряд зубов 12,3—13,8 мм. Северный Кавказ, Главный Кавказский
хребет и Закавказье до Батуми включительно. Вероятно есть в Северной Турции.
с) Т. е. orientalis Ognev, 1926. (Тип из Хосты, Черноморский берег Кавказа.)
Размеры немного меньше, чем у предыдущей формы: наибольшая длина черепа
29,7—34 мм} кондилобазальная длина 29,2—32,8 мм; ширина носового отдела
4,0—4,5 (до 4,7) мм; максиллярный ряд зубов 11,6—12,8 мм. Эта форма была
описана С. И. Огневым (1926) в качестве подвида слепого крота (Talpa caeca Savi,
1822), описанного из Италии. В Италии слепые кроты как будто хорошо отличаются
по размерам от обыкновенных (см. A. Toschi, 1959). Однако в условиях Кавказа они
по размерам перекрываются мелкими особями кавказской формы обыкновенного
крота. Отличия в строении таза и относительных размерах верхних резцов для диагно-
стики видов и подвидов не пригодны по причине их широкой индивидуальной измен-
чивости. Черноморский берег Кавказа, от Туапсе до Сухуми; Красная Поляна; южный
участок Кавказского заповедника.
d) Т. е. levantis Thomas, 1906. (Тип из Скалита, близ Трабзона, Северо-Восточная
Турция.) Наибольшая длина черепа 29,4—31,8 мм; кондилобазальная длина 28,8—
31,0 мм} максиллярный ряд зубов 10,5—11,7 мм. Видимо, в синонимы этой формы
следует отнести Т. е. olimpica Chaworth-Musters, 1932 (тип с горы Олимп, в Греции),
и Т. е. transcaucasica Dahl, 1944 (тип из окрестностей с. Воскресеновка Кироваканского
района Армянской ССР).
е) Т. е. taiyschensis Verestschagin, 4945. (Тип из ущелья Виляж-чай, в Талышских
горах, Азербайджан.) По размерам, как мелкие особи предыдущей формы: наиболь-
шая длина черепа 29,4—30,1 мм; кондилобазальная длина 28,9—29,8 мм. Носовой отдел
черепа широкий: 3,8—4,3 мм. Коренные зубы относительно крупные. Желательно
уточнить отличия от мелких кротов из Армении. Тал^ш в Азербайджане.
7. Алтайский крот. Talpa altaica Nikolsky, 1883
Табл. II, 4. Карта 6
Западная и Средняя Сибирь от оз. Зайсан, с. Убинского, Семи-
палатинска и Тары, на Иртыше, и от Сургута, на Оби, к востоку до
поймы Селенги (в Забайкалье), Верхней Лены (до Усть-Кута) и верх-
ней части бассейна р. Вилюя. На севере почти по всему бассейну Ниж-
ней Тунгуски, а по Енисею вниз до г. Дудинки (69°30'). К югу насе-
ляет долины рек и лесные ландшафты Алтая, Саян, Тувинской области
и уходит за пределы нашей страны в северо-западные районы Монго-
лии. В горах Алтая и Саян поднимается до субальпийского пояса (до
2000—3500 м).
Наибольшая численность алтайского крота бывает в поймах рек
(облесенных), в лиственных и смешанных насаждениях с густым разно-
травьем и рыхлой, богатой гумусом почвой. Мало или совсем не бывает
в чистых хвойных лесах, в сильно заболоченных почвах и на быстро
просыхающих песках. В сырых лесах предпочитает опушки и вырубки.
Ходы в лесных ландшафтах преобладают поверхностные, а на открытых
местах — глубинные, с кротовинами. Питается, как и другие виды кро-
тов, главным образом дождевыми червями. Размножается с латентной
фазой в развитии эмбрионов. Спаривание происходит летом (с июня до
начала августа), эмбрионы обнаруживаются лишь в апреле, детеныши
родятся в мае. Период беременности (с латентной фазой) составляет
около 9 месяцев. В одном приплоде бывает около 5 детенышей. В июне
молодняк уже мало отличается от взрослых и попадает в ловушки. Ли-
нек 3. Шкурки алтайского крота больше и прочнее, чем обыкновен-
ного. В южных частях ареала (на Алтае, в Саянах) ведется интенсив-
52
ный промысел, а в средней и северной частях ареала промысел пока не
развит. В поймах больших рек в периоды высоких половодий отмеча-
лась массовая гибель кротов от затопления.
Географическая изменчивость алтайского крота выражена очень слабо, при нали-
чии широкой индивидуальной изменчивости и резко выраженного полового диморфизма
(самцы значительно крупнее самок; у европейского крота диморфизм, пожалуй, менее
резок, хотя тоже имеется). С. У. Строганов (1948, 1957) считал «легко различимыми»
по окраске и размерам 5 подвидов. Различия их по размерам ничтожно малы.
Для иллюстрации приводим частичную выписку из сводной таблицы С. У. Строга-
нова (1948, стр. 380):
Промеры Т. a. altaica Т. a. suschkini Т. a. salairica Т. a. tymensis
Длина тела с голоеой 152—203 163—232 150—179 150—162
Длина ступни (без когтей) . . . 20,2—24,0 21,4—24,6 19,3—23,0 19,0—22,0
Кондилобазальная длина черепа 36,3—39,6 37,2—41,0 36,7—39,7 35,5—38,4
Наибольшая ширина черепа . . 16,8—19,0 17,2—18,7 17,1—18,5 17,'3—18,5
Ширина рострума над клыками 5,2—6,0 5,4—6,3 5,2—6,2 5,2—6,0
Высота черепа 10,3—11,5 10,5—11,8 10,5—11,8 10,5—11,8
Длина максиллярного ряда зу-
бов 12,2—13,4 12,5—13,7 12,0—13,5 11,8—12,4
С. У. Строганов составил таблицу, чтобы показать «различия» в размерах
отдельных подвидов. Мы же привели выписку по важнейшим (в диагностике кротов)
размерам с противоположной целью: чтобы каждый мог убедиться, что реальных
различий между ними нет.
В крайнем случае можно выделить 2 формы, слабо различающиеся по тону
окраски меха:
а) Т. a. altaica Nikolsky, 1883. (Описана по экземпляру из окрестностей дер. Тоу-
рак, на Алтае.) Синоним ее: Т. a. var. suschkini Kastschenko, 1905. (Тип из окрестностей
дер. Бараксан, близ с. Ермаковского, Западный Саян.) В окраске меха есть коричне-
ватые оттенки. Алтай и Саяны к востоку до Иркутска.
b) Т. a. salairica Egorin, 1936. (Тип с Салаирского кряжа.) Синонимы ее: Т. а.
tymensis Egorin, 1937 (тип из окрестностей пос. Наунак, на р. Васюгане), и позднее
описанная А. С. Фетисовым вариация из Тувинской области и Южного Забайкалья.
В окраске меха коричневатых оттенков нет. Весь ареал вида, кроме Саян и Алтая.
РОД МОГЕРЫ. GENUS MOGERA Pomel, 1848
Зубная формула: I у С у Pm у Ртр у Му-2 = 42.
Нижних клыков нет; их функцию (как и у настоящих кротов) вы-
полняют увеличенные первые нижние предкоренные зубы. Отсутствие
нижних клыков — признак постоянный, вполне устойчивый, и нет осно-
ваний для объединения могер с родом Talpa или родами Parascaptor
и Scaptochirus. У последних нижние клыки есть, а отсутствуют малые
предкоренные зубы; У Parascaptor Pm2, а у Scaptochirus — Pm2 и pm2.
Южная половина Восточной Азии до Японских островов включительно.
К роду могер относятся 3 вида: из них 2 представлены в фауне
СССР.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ МОГЕР, ПРЕДСТАВЛЕННЫХ
В ФАУНЕ СССР
1(2). Размеры средние: длина тела 120—156 juju; длина ступни
17—21 juju. Наибольшая длина черепа 33,4—37,9 juju; кондилобазальная
длина 30—38,7 juju; наибольшая ширина черепа 14,3—18,3 juju; длина
максиллярного ряда зубов 10,6—14,9 juju. Вес 85—96 г...........
......................Средняя могера. Mogera wogura. (Стр. 54.)
53
2(1). Размеры крупные: длина тела 160—206 мм; длина ступни
21—26 мм. Наибольшая длина черепа 41,2—46,2 мм; кондилобазальная
длина 40,1—45,4 мм; наибольшая ширина черепа 19,2—20,2 мм; длина
максиллярного ряда зубов 15,4—17,2 мм. Вес НО—232 г...............
.........................Большая могера. Mogera robusta. (Стр. 54.)
8. Средняя могера. Mogera wogura Temminck, 1842
Табл. II, 5. Карта 6
Японские острова, Тайвань, Хайнань, Фуцзян, Ляонин, Корея, край-
ний юг Советского Приморья: в июле 1948 г. добыта Д. И. Бибиковым
у оз. Хасан (до 1960 г. была в коллекции Зоологического музея МГУ
под ошибочным названием М. robusta).
Биология не изучена.
В провинции Ляонин населяет горные широколиственные и сме-
шанные леса с выходами камней. Очень осторожная. В цилиндры, вры-
тые под ходы, и в кротоловки добыть не удалось. Ходы в лесу поверх-
ностные, по диаметру похожие на ходы обыкновенных кротов. На юге
Приморья живет в безлесной равнине и на увалах с редким кустарни-
ком (М. В. Охотина, 1963).
Кроме номинальной формы: М. w. wogura Temmink, 1842 (тип из окрестностей
Нагасаки, Кюсю, Япония), описаны еще 3 подвида, различия которых требуют де-
тальной проверки и уточнения. В СССР встречается, вероятно, М. w. coreana Thomas,
1907. (Тип из Ким-хоа, в 60 милях к северо-востоку от Сеула, Корея.)
9. Большая могера. Mogera robusta Nehring, 1891
Табл. II, 6. Карта 6
Южное Приморье (к северу до р.Иманпо Уссури идо бухты Свет-
лой по берегу Японского моря), горные леса по долинам Уссури и Сун-
гари, Корея.
Населяет широколиственные леса и долины рек Дальнего Востока
с рыхлой (местами сильно каменистой) почвой и густым разнотравьем.
В чистых хвойных лесах и на открытых местах встречается редко или
совсем не поселяется. Ходы преобладают поверхностные без выбросов.
Основную пищу составляют дождевые черви и личинки насекомых.
В году бывает 1—2 приплода, до 8—10 детенышей в каждом (в сред-
нем около 7). Беременные самки с мая до июля. Вес новорожденного
около 2—2,5 г; лактация около 30 дней; в 10-недельном возрасте они
достигают 3/4 величины взрослых (М. И. Зубаровский, 1939). Линяют
4 раза в году (весной, летом, осенью и зимой). Шкурки большой мо-
геры раза в два больше по площади, чем обыкновенного и алтайского
кротов, и гораздо прочнее; меховые качества шкурок значительно выше,
но промысел могеры не развит и не имеет перспектив для широкого
развития, потому что ареал вида небольшой, а площади, населенные
с промысловой плотностью, еще меньше. Вероятно, целесообразными
будут опыты по акклиматизации могеры в широколиственных лесах
западных областей нашей страны.
В СССР встречается номинальная форма: М. г. robusta Nehring, 1891. (Тип из
окрестностей г. Владивостока.)
СЕМЕЙСТВО ЗЕМЛЕРОЙКОВЫЕ. FAMILIA SORICIDAE
Землеройковые — мелкие и очень мелкие насекомоядные, внешне
похожие на мышей, но с очень маленькими глазами, с вытянутым
в виде подвижного хоботка носом и покрытые бархатистым мехом.
54
Длина тела от 4,5 до 10 (у видов фауны СССР) и до 13 см (у южно-
азиатских видов). Вес от 1,2 до 17—75 г. Конечности нормального строе-
ния: пятипалые, стопоходящие, бегающего типа. Ладони короче и уже
ступней. Плавательных перепонок между пальцами нет. Хвост обычно
длинный, равный 7з—V2 или всей длине тела, густо покрытый воло-
сами, а если волосы редкие, то среди них выступают кольцеобразные
роговые чешуйки. Ушные раковины у большинства имеются в виде
покрытых короткими волосками кожных лоскутов.
Череп относительно узкий и длинный, заостренный в носовом от-
деле (рис. 2). Скуловых дуг нет (отличие от всех прочих млекопи-
тающих нашей фауны). Барабанные кости имеют вид плоских колец,
не приращенных к черепу. Многие кости черепа рано срастаются.
Зубов от 32 до 26. Внутренние (передние) рез-
цы сильно увеличены, особенно нижние (рис. 8);
они значительно крупнее смежных с ними зубов.
Коронки левых и правых передних резцов сомкнуты.
Верхние внутренние резцы двухвершинные; перед-
ний край их серповидно изогнут. Нам удалось вы-
яснить, что верхний резец землероек представляет
собой продукт сращения между собой двух резцов
(внутренней и средней пары). У добытой нами очень
молодой белозубки (Сг. russula attenuata) коронки
этих зубов были еще раздельными. Корни этих зу-
бов остаются раздельными и у взрослых белозубок
(рис. 7). У бурозубок срастание резцов проис-
ходит раньше и полнее. Корни у них тоже сра-
стаются, но у основания корней сохраняется глубо-
кая, разделяющая их впадина, и стволы корней че-
рез общий футляр просвечивают раздельно. Ниж-
ние резцы особенно велики и вытянуты вперед: верх-
ний край их гладкий или снабженный рядом тупо-
вершинных возвышенностей. Впереди многобугор-
чатых коренных зубов (характерного для насеко-
моядных строения) хорошо выделяются большие
(praemolares prominantes, Pmp — верхние и pmp — нижние): четырех-
или пятивершинные в верхней челюсти, одно- или двухвершинные —
в нижней.
Между передними (внутренними) резцами и большими предкорен-
ными располагаются мелкие одновершинные (конические) промежу-
точные зубы (dentes intermedins, Din и din, или D — верхние и d — ниж-
ние). В нижней челюсти у всех землероек с каждой стороны только по
одному промежуточному зубу, расположенному над основанием коронки
резца. К какой категории этот зуб относится (резец, клык или малый
предкоренной), установить невозможно. Обычно его считают малым
предкоренным, и это наиболее вероятно. В верхней челюсти с каждой
стороны от 5 до 2 промежуточных зубов; в их состав входят внешние
(задние) резцы, клыки и малые предкоренные. Число верхних про-
межуточных зубов — один из важнейших признаков для разделения
землероек на роды.
Семенники располагаются в полости тела. Копулятивный орган
очень большой; у взрослых самцов длина его превышает 7з—2/з длины
туловища. Кости (os penis) нет.
Распространены в Евразии (включая Малайский архипелаг),
в Африке, Северной Америке и на крайнем северо-западе Южной
Америки.
Ч -
Рис. 7. Верхний ре-
зец бурой белозубки
(Crocidura russula
Негш.). Хорошо вид-
ны два обособленных
корня, соответствен-
ные двум вершинам.
предкоренные
55
Населяют они различные зоны и ландшафты, от равнинных и гор-
ных тундр до пустынь разного типа. В горы поднимаются до альпийских
лугов. Большинство видов ведут наземную жизнь, немногие — полу-
водную. Укрываются в своих норах или вырытых другими животными
(чаще всего мышевидными грызунами); в пустотах пней и поваленных
древесных стволов, под кучами валежника, реже, в постройках чело-
века. Питаются почти исключительно насекомыми в разных фазах раз-
вития и другими мелкими беспозвоночными, редко — семенами расте-
ний (в частности, хвойных). Свой корм они обычно разыскивают в слое
листовой подстилки и под всякими укрытиями. Характерна для земле-
Рис. 8. Череп и зубы бурозубки обыкновенной
(Sorex araneus L.), вид сбоку.
ВО — венечный отросток. СО — сочленовный отросток.
1—2
УО — угловой отросток нижней челюсти. Зубы I —
верхний резец (двойной); Dl—D5 — верхние промежуточ-
ные зубы (Dentes intermedins); D1 — третий резец (Z3);
D2 — клык (С); D3—D5 — верхние малые предкоренные
(Praemolares); Ртр— крупный предкоренной (Ргаето-
lares prominantes); At'— М3 — верхние заднекоренные
(Molares); А — нижний резец; d\ — первый нижний про-
межуточный зуб (неизвестной категории); ртр — вто-
рой нижний промежуточный зуб, или нижний крупный
предкоренной (praemolares prominantes); тх—т3 — ниж-
ние заднекоренные зубы (molares). Справа — отно-
сительные размеры верхних промежуточных зубов у
обыкновенной бурозубки.
роек очень высокая интенсивность обмена. Свой высококалорийный
(животный) корм они поедают в громадном количестве, и, чем меньше
размеры зверька, тем относительно больше корма он потребляет. Об-
щий вес, который они съедают за сутки, превосходит вес самого
зверька до 2—4,2 раза. Переваривание пищи протекает быстро, а по-
требность в ней очень высокая. Поэтому землеройки кормятся много
раз в сутки, а промежутки между кормежками проводят-во сне. Астро-
номические сутки у землероек бывают разделены на несколько «своих»
биологических суток, причем, чем меньше зверек, тем больше бывает
периодов сна и кормежки в течение суток. Крошечная бурозубка, на-
пример, спит до 78 раз в сутки, а кормится до 120 раз. Долго жить без
пищи они не могут. Мелкие виды, оставленные без корма, погибают за
срок от 7—9 до 28—57 часов. Потребность в питьевой воде у разных
видов различна. Размножаются землеройки 1—2, видимо, в редких слу-
чаях до трех раз в году. Указания А. К- Тарковского (1955) на 4—5
приплодов в сезон не доказаны. В одном приплоде бывает от 1—3 до
56
11 —14 детенышей, в среднем — от 4 до 8. Детеныши родятся голыми,
слепыми, с неразвитым хоботком (курносые). Развиваются они быстро
и через 3—4 недели становятся самостоятельными; в этом возрасте они
отличаются от родителей по весу, но размеры, особенно размеры черепа,
становятся такими же, как у взрослых. Осенью у молодых зверьков
наблюдается уменьшение размеров тела, уплощение черепа и даже
уменьшение объема и веса головного мозга (С. Боровский и А. Денель,
1952; К. Кабон, 1955). В зимнюю спячку не впадают. В средней полосе,
линька протекает с сентября по ноябрь (осенняя) и в апреле — начале
мая. С апреля по июнь происходит увеличение объема мозговой капсулы,
увеличение веса и объема головного мозга (К. Кабон, 1955; Виелак и
Пуцек, 1960). В средней Европе обыкновенные бурозубки живут до 14—
16, самое большое до 18 месяцев (А. Денель, 1949).
Землеройки — полезная группа млекопитающих. Главная польза их
заключается в уничтожении огромного количества насекомых во все се-
зоны года и в местах, недоступных для многих, а то и для любых видов
насекомоядных птиц (под снегом, под валежником, камнями, в толще
листовой подстилки, в глубине нор и т. д.). Численность землероек
местами бывает очень высокой.
В семейство землеройковых объединяют около 20 родов, хорошо
различающихся; из них в фауне СССР представлено 5 родов. А коли-
чество видов землероек до сих пор окончательно не установлено.
Систематика землероек — один из труднейших разделов современ-
ной териологической систематики. Главное затруднение заключается
в том, что пока остается не изученной степень устойчивости или диапа-
зоны изменчивости отдельных признаков. Систематики и морфологи,
имевшие возможность изучить изменчивость, не уделяли этому долж-
ного внимания, а без глубокого изучения изменчивости выяснить видо-
вой состав землероек невозможно. Односторонняя концентрация вни-
мания на отдельных, случайно избранных признаках с неизвестным диа-
пазоном их изменчивости приводит только к путанице.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ЗЕМЛЕРОЕК,
ПРЕДСТАВЛЕННЫХ В ФАУНЕ СССР
1(8). Окраска спины более или менее одноцветная. В верхней
челюсти с каждой стороны не менее 3 промежуточных зубов ... 2
2(5). Хвост сверху и с боков покрыт однообразными короткими
волосами. Ушные раковины не выступают над уровнем меха. Вер-
шины и жевательные поверхности зубов окрашены в красновато-бурый
цвет (разной интенсивности и оттенков). Крыша черепа в заглазнич-
ной области имеет пару небольших сближенных отверстий . . . . 3
3(4). В верхней челюсти с каждой стороны по 5 промежуточных
зубов (в очень редких случаях по 4). Поверхность переднего нижнего
резца снабжена рядом отчетливо выраженных туповершинных возвы-
шенностей (рис. 8). Края лапок покрыты обычными короткими
волосами, не образующими гребневидных оторочек. Хвост как сверху,
так и снизу покрыт однообразными короткими волосами (рис. 9, А) . .
....................................Бурозубки. Sorex. (Стр. 58.)
4(3). В верхней челюсти с каждой стороны по 4 промежуточных
зуба. Поверхность переднего нижнего резца гладкая или слегка волни-
стая. Края лапок (и внешние пальцы) у взрослых особей несут обычно
хорошо выраженную оторочку из относительно длинных жестких волос,
а на нижней стороне хвоста из удлиненных волос образуется явствен-
ный киль............................Куторы. Neomys. (Стр. 67.)
57
5(2). На хвосте сверху и с боков из густых и коротких волос вы-
ступают отдельные редкие длинные волосы (рис. 9, Б). Ушные рако-
вины всегда выступают над уровнем меха. Все зубы сплошь белые.
Крыша черепа в заглазничной области лишена отверстий .... 6
6(7). Размеры очень мелкие: длина тела (головы и туловища)
меньше 46 мм; задняя ступня без когтей не более 8,5 мм; общая длина
черепа меньше 13 мм. В верхней челюсти с каждой стороны по 4 про-
межуточных зуба (самый задний из них очень маленький и в редких
случаях может отсутствовать).......................................
. . . Многозубые белозубки. Suncus. (Стр. 69.)
роек:
А — бурозубки (Sorex) и
Б — белозубки (Croci-
dura).
7(6). Размеры крупнее: длина тела (головы
и туловища) больше 48 мм; задняя ступня без ког-
тей не менее 10 мм; общая длина черепа более
15 мм. В верхней челюсти с каждой стороны по
3 промежуточных зуба (очень редко по 2) . . .
...............Белозубки. Crocidura. (Стр. 69.)
8(1). На середине спины, окрашенной в пе-
пельно-серый цвет, расположено большое белое
пятно продолговатой или эллипсовидной формы.
В верхней челюсти с каждой стороны по 2 проме-
жуточных зуба.................................
............Путораки. Diplomesodon. (Стр. 78.)
РОД БУРОЗУБКИ. GENUS SOREX Linnaeus, 1758
Зубная формула: I -j-Din-j-Pmpy Му-2 = 32.
Группа «Din» означает мелкие одновершинные
конические промежуточные зубы (dentes interme-
dins), не разделенные на общепринятые категории
(резцы, клыки, предкоренные). Расшифрованная
по обычным категориям зубов формула должна
быть написана, видимо, так:
I — С —Pm — Pmp—М —-2 = 32.
10 1 1 3
Соотношение размеров (высоты, или уровня вершин) верхних про-
межуточных зубов используется в диагностике видов бурозубок, хотя
устойчивость этого признака ограниченная. Обычно размеры верхних
промежуточных зубов уменьшаются от первого (самого переднего,
Din1) до пятого (Din5). Уменьшение может быть как равномерным, так
и ступенчатым, что можно выразить формулами:
1>2>3>4>5; 1=2>3 = 4>5; 1 = 2 = 3> 4 > 5 и т. д.
Соотношение, показанное во второй формуле, называют двупар-
ной группировкой зубов. Такие варианты могут быть у особей боль-
шинства видов, что исключает их диагностическое значение. Однако
случаи, в которых последующий (по порядку) зуб больше преды-
дущего, бывают редко и свойственны немногим видам. Например,
у бухарской бурозубки (Sorex buchariensis Ognev) Din 1>2<3>4>5,
т. e. третий промежуточный зуб больше второго, и это характерно для
данного вида. Однако у малой бурозубки (S. minutus) соотношение
58
этих же зубов оказалось менее устойчивым. Обычно у нее бывает
1=2<3>4>5, но бывает также 2 = 3 и 2>3. У гигантской бурозубки
(S. pacificus) обычно Din 3<4>5, но бывает 3 = 4 и 3>4.
Относительная высота задней вершины верхнего резца и нижнего
промежуточного зуба, величина и расположение бугорков (мозолей) на
ступне не имеют существенного значения в диагностике в силу их ши-
рокой изменчивости и неточности самих измерений (нижнего промежу-
точного зуба из-за его сложной формы; мозолей из-за деформации на
шкурках и при разных условиях фиксации). Строение головки копуля-
тивного органа самцов нуждается в детальном изучении диапазонов ее
изменчивости: индивидуальной, географической, возрастной, сезонной и
зависимой даже от состояния в момент добычи самца и от условий фик-
сации (в спирте, формалине, при разных концентрациях фиксатора
и т. д.). В случае подтверждения некоторых различий в строении этого
органа включать его описание в характеристики видов желательно, од-
нако сводить различия видов только к этому признаку недопустимо.
Между видом и признаком, да еще свойственным только половине осо-
бей, нельзя ставить знак равенства.
Распространены в Евразии, к югу до Северо-Западного Памира и
Тибета включительно, Северной Америке и на северо-западе Южной
Америки.
Высокой численности бурозубки достигают в лиственных и сме-
шанных лесах, на лугах, особенно пойменных, облесенных; меньше их
бывает в луговых степях, хвойных насаждениях и тундрах и очень мало
или совсем не встречаются в сухих плакорных степях, полупустынях и
пустынях. В лесных ландшафтах они часто выходят в ранг доминантов
или согосподствующих видов. Это значит, что до половины или более
половины всех особей приходится на долю одного-двух видов бурозу-
бок, а другая половина — на всех остальных млекопитающих, вместе
взятых. При столь высокой численности и характерной для них прожор-
ливости бурозубки имеют, видимо, огромное значение в структуре з о о т
(биоценозов) лесных ландшафтов, и круглогодичным уничтожением
большого количества вредных насекомых они оказывают очень боль-
шую пользу в охране наших лесов.
Систематика бурозубок разработана слабо и некоторыми авто-
рами сильно запутана.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ БУРОЗУБОК ФАУНЫ СССР
1(2). Размеры очень мелкие: кондилобазальная длина черепа
11,7—14,1 мм; наибольшая ширина черепа 5,7—7 мм. Длина задней
ступни (без когтей) 6,2—8,9 мм. Вес 1,6—2,9 г....................
............Крошечная бурозубка. Sorex minutissimus. (Стр. 61.)
2(1). Размеры крупнее: кондилобазальная длина черепа не менее
14,4 мм; наибольшая ширина черепа (за редчайшими исключениями)
более 7 мм. Длина ступни (без когтей) не менее 9 мм. Вес взрослых
особей более 3 г...............................................3
3(4). Размеры относительно мелкие: ширина черепа не более
8 мм (в единичных случаях до 8,2 мм); длина верхнего ряда зубов
менее 7 мм (в редчайших случаях до 7,2 мм). Дополнительно: третий
верхний промежуточный зуб немного крупнее второго, реже они равны
друг другу, и очень редко третий зуб чуть меньше второго. Вес 3—5 г.
.........................Малая бурозубка. Sorex minutus. (Стр. 62.)
4(3). Размеры крупнее: ширина черепа больше 8—8,2 мм (в очень
редких случаях от 7,7 мм); длина верхнего ряда зубов не менее 7—
59
7,2 мм. Третий верхний промежуточный зуб заметно меньше второго
или, наоборот, значительно крупнее второго. Вес взрослых особей более
5 г (редко от 4—4,5 г).........................................5
5(6). Третий верхний промежуточный зуб значительно крупнее
второго. Размеры средние: длина ступни 11,3—12 мм; кондилобазальная
длина черепа 16—17,5 мм; ширина черепа 8—8,4 мм. Хребет Петра Вели-
кого, Северный Таджикистан . . •.................................
...............Бухарская бурозубка. Sorex buchariensis. (Стр. 62.)
6(5). Третий верхний промежуточный зуб заметно меньше второго
или они равны; если же третий крупнее второго, то размеры крупные:
ступня взрослых особей больше 12 мм; кондилобазальная длина черепа
не менее 18—19 мм; ширина черепа больше 8,5 мм (географически не
Северный Таджикистан)..........................................7
7(16). Размеры средние: длина ступни (без когтей) не более
15,5 мм; кондилобазальная длина черепа менее 21,7 мм; длина верхнего
ряда зубов не более 10 мм. Четвертый верхний промежуточный зуб
меньше третьего или равен третьему.............................8
8(9). Пигментация зубов очень сильная: у верхнего большого
предкоренного почти всегда пигментированы обе внутренние вершины
и даже впадина между основаниями наружного и внутреннего рядов
бугров; у первого и второго заднекоренных сильно пигментированы
тоже обе внутренние вершины. Верхние промежуточные зубы массив-
ные с широкими полого округленными и сильно пигментированными
вершинами. Мех на тыльной стороне задних лап такой же темный,
как на спине. Дополнительно: третий верхний промежуточный зуб по
размерам примерно равен второму. Размеры, как у средней бурозубки:
ступня 10—12,5 мм; кондилобазальная длина черепа 17—18,7 мм;
ширина черепа 8,7—9,5 мм; длина верхнего ряда зубов 7,8 —
8,6 мм . . . Темнозубая бурозубка. Sorex daphaenodon. (Стр. 63.)
9(8). У большого верхнего предкоренного зуба пигментирована
только передняя внутренняя вершина или обе вершины белые; задние
внутренние вершины первого и второго верхних заднекоренных в боль-
шинстве случаев белые, и лишь изредка на них имеется маленькое
бурое пятнышко. Верхние промежуточные зубы (если они не стерты)
с остроконечными вершинами. Мех на тыльной ‘ стороне лап значи-
тельно светлее меха спины. Дополнительно: размеры ступни и черепа
обычно крупнее, чем указано в тезе 8........................ 10
10(15). Длина хвоста менее 53 мм; длина ступни не более 14 мм.
Окраска меха (верха и низа) с коричневатыми, шоколадными, бурыми,
палевыми тонами, но не однообразная дымчато-серая. (Нижний проме-
жуточный зуб, сидящий над основанием резца, в большинстве случаев
одновершинный).......................•........................11
11(12). Размеры средние: кондилобазальная длина черепа менее
18 (18,8) мм. Длина ступни обычно не более 12 (12,8) мм. В скобках
указаны максимальные размеры зверьков с Алтая, Саян и прилегаю-
щих к ним территорий Сибири...................•..................
......................Средняя бурозубка. Sorex caecutiens. (Стр. 63.)
12(11). Размеры крупные: кондилобазальная длина черепа не ме-
нее 18(19,4) мм; длина верхнего ряда зубов обычно более 8(8,8) мм.
Длина ступни обычно не менее 12 мм. В скобках указаны наименьшие
размеры зверьков с Алтая, Саян и прилегающих к ним территорий
Сибири....................................................... 13
13(14). Когти на пальцах передней лапы короче 3 мм. Второй
верхний промежуточный зуб равен или немного меньше первого и
60
больше третьего (высота первого нижнего промежуточного зуба обычно
около 2/3 длины основания коронки этого зуба).......................
...............Бурозубка обыкновенная. Sorex araneus. (Стр. 64.)
14(13). Когти на пальцах передней лапы длиннее 3 мм (до 4,8 мм).
Второй верхний промежуточный зуб значительно (раза в полтора)
меньше первого и равен или меньше третьего. Высота первого
нижнего промежуточного зуба составляет около Уг его основания или
меньше; вершина его смещена вперед..................................
...............Когтистая бурозубка. Sorex unguiculatus. (Стр. 66.)
15(10). Длина хвоста 58—71 мм; длина ступни 14—15,5 мм.
Окраска меха однообразная темная дымчато-серая (на брюхе немного
светлее, чем на спине) без примеси коричневатых, шоколадных, или па-
левых тонов. (Нижний промежуточный зуб, сидящий над основанием
резца, всегда двухвершинный)........................................
..................Альпийская бурозубка. Sorex alpinus. (Стр. 66.)
16(7). Размеры крупные: длина ступни (без когтей) 16—17,5 мм;
кондилобазальная длина черепа 22,3—24,3 мм; длина верхнего ряда
зубов 10,0—10,5 мм. Четвертый верхний промежуточный зуб (за редким
исключением) больше третьего........................................
..................Гигантская бурозубка. Sorex pacificus. (Стр. 66.)
10. Крошечная бурозубка. Sorex minutissimus Zimmermann, 1780
Табл. Ill, 1. Карта 7
От Западной Карелии, оз. Селигер (Калининской области),Москов-
ской и Орловской областей к востоку до Чукотки, Камчатки, о. Куна-
шира и Японии (о. Хонсю). К северу до оз. Суоярви в Карелии, долины
р. Сосьвы, Нижней Тунгуски, Якутска, Нижне-Колымска и с. Марково,
на Анадыре; к югу до Брянского лесничества Орловской области, Чел-
кара, оз. Кургальджин (в Целинном крае), до Алтая, Тувы, Северной
Монголии и Южного Приморья.
Места обитания разнообразны, но преимущественно лиственные
леса, луга и поймы рек таежной, лесотундровой и лесолуговой зон. Ис-
ключение составляют две находки в Казахстане: в степи у оз. Кургаль-
джин и в северной пустыне близ Челкара. Убежища (норы) и гнезда ее
неизвестны. Суточный режим крошечной бурозубки заключается в ча-
стом чередовании коротких периодов активности и столь же коротких
периодов сна. По К. Н. Благосклонову (1957), бывшая под его наблю-
дением взрослая самка спала 78 раз в сутки, что составило в общей
сложности И часов 44 минуты. Кормилась за те же сутки она 121 раз
и съела более 10 г корма (муравьиных куколок), что в 4,2 раза превы-
шало ее собственный вес. У двух известных беременных самок было 4
и 5 эмбрионов. Молодые попадались с 10 июня до конца октября.
Из пределов громадного ареала описано несколько форм. Из них практически раз-
личить можно следующие:
a) S. m. minutissimus Zimmermann, 1780. (Описана по экземпляру из окрестно-
стей бывшего с. Кийского, теперь г. Мариинска Кемеровской области.) Размеры
относительно крупные: длина ступни (без когтей) 7,6—9 мм; кондилобазальная
длина черепа 13,2—14,1 мм; ширина черепа 6,2—7 мм. Окраска меха тусклая,
серовато-бурая; низ серовато-белесый; бока светлые с ясной границей между верхом
боков и темно окрашенной спиной. Хвост нерезко двухцветный. От номинальной формы
практически не отличимы: S. m. tscherskii Ognev, 1913 (тип с р. Одарки в бассейне
оз. Ханка); S. m. hawkerl Thomas, 1905 (о. Хондо, Япония); S. т. neglectus Ognev, 1921
(тип из Можайского района Московской области); последняя отличается от номиналь-
ной только более узким черепом, ширина которого 5,8—6,2 мм. Очень близка к номи-
нальной и вряд ли может быть выделена в качестве подвида S. m. karelicus Stroganov,
1949 (тип с оз. Суоярви, Карелия); автор отличал ее по более темной окраске меха.
61
b) S. m. tschuktschorum Stroganov, 1949. (Тип из окрестностей с. Марково, на
Анадыре.) Мех пышный, густой, шелковистый, окраска яркая; хвост резко двухцвет-
ный. Размеры несколько меньше номинальной формы: длина ступни (без когтей)
7,3—8,3 мм; кондилобазальная длина черепа 12,8—13,2 мм; ширина черепа 6—6,3 мм.
Поймы рек Колымы, Анадыря и Камчатка.
с) S. m. barabensis Stroganov, 1956. (Тип из окрестностей дер. Эгербаш Барабин-
ского района Новосибирской области.) Окраска меха близка к номинальной. Раз-
меры очень мелкие: длина ступни (без когтей) 6,2—8 мм; кондилобазальная длина
черепа 11,7—12,3 мм; ширина черепа 5,7—6 мм. Западносибирское лесостепье.
d) S. m. abnoimis Stroganov, 1949. (Тип из окрестностей г. Челкара, Казахстан.)
Окраска летнего меха бледная, буровато-коричневая; бока и низ серебристо-серо-
ватые. Размеры немного крупнее, чем у барабинской формы (сходные с чукотско-
камчатской). Пески Большие Барсуки, близ Челкара, и степь у оз. Кургальджин
Целинного края.
11. Малая бурозубка. Sorex minutus Linnaeus, 1766
Табл. Ill, 2. Карта 8
Лесолуговая зона и долины рек степной и таежной зон Европы,
Горный Крым, Кавказ и Закавказье, Тянь-Шань, лесолуговая зона За-
падной Сибири и долины рек к северу до кустарниковых тундр Ямала;
Алтай, Саяны, Прибайкалье, южный и Баргузинский берег Байкала,
Центральная Якутия, низовья р. Анадырь, Шантарские острова, ни-
зовья Амура, Приморский край, Сахалин, Курильские острова. За пре-
делами СССР — Кашмир, Палестина, Цайдам и др.
Населяют леса, преимущественно лиственные и смешанные, и луга.
Заболоченные участки предпочитает, видимо, сухим. Ежедневно съедает
насекомых, вес которых более чем в 2 раза превосходит вес самого
зверька. Без корма может прожить не более 9 часов (Н. В. Тупикова,
1949). В году бывает 1 или 2 выводка, по 4—12 детенышей в каждом
(обычно 6—8).
Географическая изменчивость изучена недостаточно. Можно выделить следую-
щие слабо различимые формы:
a) S. m. minutus Linnaeus, 1766. (Описана из окрестностей г. Барнаула, Западная
Сибирь.) Относительно мелкая форма: длина ступни 9,5—М мм; кондилобазальная
длина черепа 14,4—15,6 мм; ширина носового отдела 1,5—1,9 мм. Средняя и Северная
Европа, Западная и Средняя Сибирь до ближнего Забайкалья, на юг до Семиречья
включительно (синоним: S. m. heptapotamicus Stroganov, 1956; тип из Терскей-Алатау,
Тянь-Шань).
b) S. m. gracillimus Thomas, 1907. (Тип из окрестностей Дарине, к северо-западу
от Корсакова, на Сахалине.) От номинальной отличается более узкой носовой частью
черепа, ширина которой составляет лишь 1,3—1,5 мм. Дальневосточная часть ареала.
Недостаточно ясны отличия от этой формы S. leucogaster Kuroda, 1933, описанной по
типу с Северных Курил.
с) S. m. gmelini Pallas, 1811. (Описана из Крыма.) Размеры значительно крупнее
номинальной формы: длина ступни 11,9—12 мм; кондилобазальная длина черепа
16,6—17,3 мм. Горный Крым.
Положение кавказской малой бурозубки, описанной под названием S. m. volnu-
chini Ognev, 1921 (по типу с р. Киши, Кавказский заповедник), остается пока недо-
статочно ясным. По размерам она занимает промежуточное положение между номи-
нальной и крымской формами.
d) S. m. portenkoi Stroganov, 1956. (Тип с побережья Анадырского лимана, близ
пос. Анадырь.) Череп относительно широкий: 7,1—8,2 мм. Третий верхний промежуточ-
ный зуб меньше второго. Хвост короткий: 23—26 мм. Пойма р. Анадырь от Усть-
Белой и Снежной до лимана и в 40 км к юго-востоку от г. Якутска.
12. Бухарская бурозубка. Sorex buchariensis Ognev, 1921
Карта 8
До 1954 г. была известна по единственному экземпляру, найден-
ному А. К. Гольбеком 9/VIII 1911 г. в долине р. Диван-су (хребет
Петра Первого, Таджикистан). В 1954 г. сотрудниками Академии наук
62 *
Таджикской ССР В. И.Чернышевым, С. А. Саид-Алиевым и другими
добыто 18 бухарских бурозубок в районе того же хребта Петра Первого
в ряде мест Джиргательского и Товиль-Доринского районов Северного
Таджикистана.
Добывалась она в высокогорных березовых, тополевых и реже ар-
чевых лесах по склонам ущелий, в относительно влажных местах с пыш-
ным травянистым покровом. В июле — августе попадались молодые
зверьки. В тот же период была добыта кормящая самка и другая самка
с 7 эмбрионами.
13. Темнозубая бурозубка. Sorex daphaenodon Thomas, 1907
Табл. Ill, 3. Карта 9
Сибирь и Дальний Восток от Кондо-Сосьвинского заповедника (ле-
вобережье Нижней Оби) и верховьев р. Ишима до Охотского побе-
режья, Курильских островов и Сахалина; к северу до р. Ем-Еган,
в Нижней Оби, устья р. Мотльки (в бассейне Таза), верхнего отрезка
р. Вилюя, Джемана на р. Яне, дельты р. Индигирки, Нижне-Колымска
и Ола, близ Магадана, на юг до Аксуата Кустанайской области, Бара-
бинского лесостепья, Верхнеобского бора (между Барнаулом и Бий-
ском), южного берега Байкала, крайнего юга Приморья и Южного
Сахалина.
Обитатель лесных ландшафтов лесолуговой зоны Сибири и Даль-
него Востока, речных долин таежной зоны, лесотундры и типичных
тундр. Предпочитает буреломы и валежники старолесья. В Барабин-
ском лесостепье в июне — августе одновременно добывались яловые,
беременные и кормящие самки. Число эмбрионов 4—9 (в среднем 6).
Численность значительно ниже, чем обыкновенной, средней и малой
бурозубок.
Можно различить 3 подвида:
a) S. d. daphaenodon Thomas, 1907. (Тип из Дорине, близ Корсакова, Южный Са-
халин.) Хвост относительно длинный: 35—39 мм. Окраска меха темно-бурая с коричне-
ватым оттенком. Кондилобазальная .длина черепа 17,3—17,7 мм. Забайкалье, Амуро-
Уссурийский край, Сахалин, Курилы.
b) S. d. sanguinidens Gl. Allen, 1914. (Тип из окрестностей г. Нижне-Колымска.)
Хвост короткий, не превышает 33 мм. Окраска меха яркая, рыжевато-шоколадная.
Мех зимой густой, пышный, высокий: на спине до 11—12 мм. Размеры черепа, как
у номинальной. Северо-восток Сибири от Вилюя, Якутска и Яны до Колымы и
Охотского побережья.
с) S. d. scaloni Ognev, 1933. (Тип из устья р. Мотльки, бассейн р. Таза.) Крупная
форма: кондилобазальная длина черепа 17,8—18,7 мм; хвост 35—39,5 мм. Окраска
верха очень темная, черно-бурая. Западносибирская часть ареала.
Близка к этому виду (особенно по размерам и сильной пигментации зубов)
тянь-шаньская бурозубка Sorex araneus asper Thomas, 1914
14. Средняя бурозубка. Sorex caecutiens Laxmann, 1788
Табл. Ill, 4—5. Карта 10
В Евразии от Швеции и Финляндии до Чукотки, Камчатки и Саха-
лина; на север до берегов Ледовитого океана; на юг до Испании, Сред-
ней Украины, Горьковской области, верховий р. Урала, Алтая, Северной
Монголии, Кореи и Японских островов.
Места обитания разнообразны: леса разного типа, кустарниковые
заросли, луга, болота, поймы рек. Питается главным образом насеко-
мыми, почти не ест дождевых червей и растительные корма. В Томской
и Новосибирской областях беременные и кормящие самки попадаются
с мая до октября. Число эмбрионов от 2 до 11 (чаще 7—8). Местами
63
средняя бурозубка численно преобладает над любыми другими видами
землероек, например в лесах Приморского края.
Индивидуальная, групповая, географическая изменчивость и морфизмы (возраст-
ные и сезонные) в окраске, строении промежуточных зубов у средней бурозубки не
изучены. С. У. Строганов (1957) к S. caecutiens относил только особей с более или
менее парной группировкой верхних промежуточных зубов (хотя на точных рисунках
зубов этого вида, помещенных в его же книге на стр. 203, из 8 вариантов в 6 груп-
пировка недвухпарная) и часть взрослых самцов с расширенным концом головки
полового органа. Всех бурозубок, не подходивших под этот произвольно установ-
ленный стандарт, он относил к другому виду — S. arcticus. Последний, по С. У. Стро-
ганову, получился сборным видом, к которому он отнес основную массу средних
бурозубок (S. caecutiens) с явно недвухпарной группировкой верхних промежуточ-
ных зубов, часть самцов того же вида, не имевших расширения на конце головки
копулятивного органа, часть малых бурозубок (S. minutus) с чепрачной окраской меха
и одну форму обыкновенной бурозубки (S. araneus schnitnikovi), у одного или неболь-
шого числа взрослых самцов которой головка копулятивного органа в деталях оказа-
лась неотличимой от S. arcticus. Этим в систематику бурозубок внесена большая пу-
таница.
Уместно также обратить внимание, что искусственно скомбинированный вид под
названием S. arcticus не имеет ничего общего с американской S. arcticus Kerr и ее гео-
графической формой S. a. tundrensis Merriam. Две последние формы хорошо отличаются
от любых вариантов средних бурозубок нашей фауны и в дальнейшем, видимо,
будут признаны подвидами S. araneus, к которой они по всем признакам крайне
близки.
Для сравнения приводим размеры: I — ступни и II — кондилобазальной длины
черепа: 1) S. minutus; 2) S. caecutiens; 3) «S. arcticus», по С. У. Строганову, 1957;
4) S. arcticus и S. a. tundrensis по Jackson 1928, по Hall and Kelson, 1959, и 5) S. araneus:
I: 1) 9—11,2 мм', 2) 10—12,5 мм- 3) 9—12,8 мм; 4) 12,5—15 мм; 5) 12—44 мм.
II: 1) 14,4—17,3 мм; 2) 16,2—18,7 лш; 3) 15,7—18,3 мм; 4) 17,8—19,1 мм;
5) 18,0—21,7 мм.
При такой путанице оказалось пока невозможным дать полную характеристику
географической изменчивости средней бурозубки.
Реально, видимо, различаются следующие подвиды:
a) S. с. caecutiens Laxmann, 1788. (Описана по экземпляру с юго-западного берега
оз. Байкал.) Длина хвоста 34—44 мм; ступни 10,5—12,5 лш; кондилобазальная
длина черепа 17—18,7 мм. Прибайкалье, Саяны, Тувинская область, Алтай. Синоним
этой формы: S. daphaenodon sibiriensis Ognev, 1921 (тип со ст. Кольчугино Кемеров-
ской области). Очень близка к номинальной форме европейско-западносибирская
S. с. pleskei Ognev, 1921 (тип из окрестностей Гдрва Псковской области) и камчат-
ская S. с. macropygmaeus Miller, 1901 (тип из окрестностей Петропавловска-на-Кам-
чатке).
b) S. с. buxtoni J. Allen, 1903. (Тип из Гижиги.) По размерам, как европейско-
сибирские, но короткохвостые: длина хвоста 26—34 мм. Окраска часто бывает резко
контрастной чепрачного типа (темная спина, отделенная резкой границей от почти
белых боков), хотя у типа, по которому описан этот подвид, окраска была нечепрач-
ной. Равнинные и горные тундры от Северо-Восточной Европы до Чукотки и север-
ных берегов Охотского моря. Синонимы ее: S. с. borealis Kastschenko, 1905 (тип
с Таймыра); S. с. koreni Gl. Allen, 1914 (тип из Нижне-Колымска). Такая же
короткохвостая форма населяет Забайкалье (S. с. baicalensis Ognev, 1913, тип из Гор-
ного Зерентуя).
с) S. с. shinto Thomas, 1905. (Тип с о-ва Хонсю, Япония.) По размерам и
окраске похожа на номинальную форму, но длина хвоста 42—50 мм. Япония, Саха-
лин, Курильские острова.
15. Обыкновенная бурозубка. Sorex araneus Linnaeus, 1758
Табл. Ill, 6—8. Карта И
От Великобритании и Франции на восток до берегов Тихого океана
к северу до устьев северных рек Сибири, к югу до Палестины, Малой
Азии и Юго-Восточной Азии. Очень близкая форма под названием So-
rex arcticus Kerr, 1792, широко распространена на севере Северной
Америки от Тихоокеанского до Атлантического побережья.
В пределах столь огромного ареала живет в разных ландшафтах,
но особенно высокой численности достигает в лесолуговой зоне и в пой-
64
мах рек, пересекающих зоны тайги и степей. В сухих степях живет
только по речным долинам; в полупустынях и пустынях совсем не встре-
чается. Очень важный компонент биоценозов в лесных ландшафтах,
особенно в западной половине Евразии, где она численно преобладает
над прочими видами бурозубок, а иногда и над прочими видами млеко-
питающих вообще (типичный фоновый вид). Общий очерк экологии бу-
розубок составлен главным образом по этому фоновому виду.
Изменчивость обыкновенной бурозубки в размерах тела и черепа, в соотноше-
нии промежуточных зубов, в окраске меха очень широкая. Некоторые варианты
обособлены географически, другие проявляются в виде морф. С. У. Строгановым
(1957) изменчивость этого, как и других видов насекомоядных, не изучена, а систе-
матика запутана. Поэтому состав географических форм обыкновенной бурозубки
нуждается в основательном пересмотре на базе тщательного изучения морфизмов,
индивидуальной и- географической изменчивости. Здесь мы приводим лишь предвари-
тельное деление видовой популяции на подвиды.
a) S. a. araneus Linnaeus, 1758. (Описана по экземпляру из окрестностей г. Упсала,
Швеция.) Размеры некрупные: ступня 12—13 мм; кондилобазальная длина черепа
18—19,2 мм; длина верхнего ряда зубов 7,9—9 мм; высота черепа 5,4—6,2 мм.
Окраска меха разных типов, но нижняя сторона в любом возрасте и в любой сезон
светлая, беловатая, беловато-охристая или беловато-серая, но не кофейно-бурая
и не каштаново-бурая. Бока у молодых зверьков окрашены почти одинаково со спи-
ной. Летом преобладает коричневато-бурая окраска боков и верха. Для зимних
характерна трехцветная окраска: черно-бурая на спине, рыжевато-палевая на боках
тела и светлая — на нижней стороне. Ширина черепа больше половины кондилоба-
зальной1 длины. Европа, Урал, равнинная часть Западной Сибири, Южный Алтай,
Тарбагатай, Саур, Джунгарский Алатау.
Синонимы номинальной формы: S. a. uralensis Ognev, 1933 (тип с истоков
р. Няысь, Северный Урал); S. a. tomensis Ognev, 1921 (partim)—часть особей, ранее
относившихся к этой форме, из равнинных районов Западной Сибири; S. a. schnit-
nikovi Ognev, 1921 (тип из окрестностей Копала, в Семиречье). С. У. Строганов (1957,
стр. 153—154) считал- последнюю подвидом S. arcticus, от которой она действительно
отличается так же, как обыкновенная бурозубка отличается от средней; однако
отличий от номинальной формы S. araneus им не показано никаких, кроме деталей
строения головки гениталий части взрослых самцов.
b) S. a. raddei Satunin, 1895. (Тип из окрестностей Кутаиси, Грузия.) Размеры
относительно мелкие: кондилобазальная длина черепа 18—19 мм; длина верхнего ряда
зубов 7,6—8,1 мм; ступня 11,6—12,2 мм. Хвост относительно длинный: до 45 мм
(в среднем более 40 мм). Окраска двух резко различных типов: 1) темная, кофейно-
бурая, на нижней стороне чуть светлее, чем на спине; 2) светлая и яркая, рыжевато-
бурая сверху и беловатая снизу. Последняя описана в качестве особой формы
S. a. satunini Ognev 1921 (тип из Мюзарета, Закавказье), Кавказ и Закавказье.
с) S. a. roboratus Hollister, 1913. (Тип из Тануча, у оз. Телецкое, на Алтае.)
От европейской, формы отличается более крупными размерами черепа: кондилоба-
зальная длина его (по С. И. Огневу, 1928) 20—20.7 мм, длина верхнего ряда зубов
8,4—9,3 мм. Окраска меха, как у европейской формы или темнее (бывают особи
с кофейно-бурым низом). Среди перезимовавших преобладает трехцветный тип
с очень темным (черноватым) чепраком. Синоним этой формы: S. a. tomensis Ognev,
1921 (тип из окрестностей г. Барнаула). Алтай, Саяны. Вероятно, к этой же форме
относятся обыкновенные бурозубки из Забайкалья, Северной Монголии и Амуро-Уссу-
рийского края.
d) S. a. asper Thomas, 1914. (Тип из долины р. Текес, Тянь-Шань.) Близка к но-
минальной, но череп при одинаковой длине (18,7—19,4 мм) с более широкой мозговой
капсулой: 9—9,6 мм (вместо 7,9—9 мм). Верхние промежуточные зубы относительно
крупные п туповершинные; все зубы сильно пигментированы; у верхнего крупного
предкоренного бывает пигментирована и задняя внутренняя вершина зуба (гипокон),
как у S. daphaenodon.
е) S. a. vir Gl. Allen, 1914. (Тип из Нижне-Колымска, в низовьях р. Колымы.)
От прочих сибирских обыкновенных бурозубок отличается преобладанием особей
с уплощенным и узким черепом (высота его 4,8—5,6 мм); ширина черепа в боль-
• шпнстве случаев меньше половины кондилобазальной длины (иногда на 2,5—3 мм)
и относительно коротким хвостом: 30—37 мм (вместо 34—50 мм у других форм).
Окраска меха обычно двухцветная: бока окрашены почти одинаково с верхом тела,
а низ светлый беловатый или со слабым охристым или палевым налетом. С. У. Стро-
ганов (1957) выделял эту форму в качестве самостоятельного вида с многими подви-
дами, но характеристики их основывал на неустойчивых признаках. Горный Северо-Во-
сток Сибири, от Енисея до Колымы и Анадыря. В Приенисейской Сибири встречаются
особи, переходные между обыкновенной и плоскочерепной формами бурозубок.
5 Н. А. Бобринский и др.
65
16. Когтистая бурозубка. Sorex unguiculatus Dobson, 1890
Табл. Ill, 9. Карта 12
Япония, Курильские острова, Сахалин, Камчатка, Охотское побе-
режье, Советское Приморье. Северные и западные пределы ареала не
выяснены.
Экология не изучена, так как этот вид многими авторами не от-
делялся от обыкновенной бурозубки. На Южном Сахалине и на острове
Кунашир живет в долинах лесных рек и по численности в 1962 г. пре-
обладала там над всеми прочими видами бурозубок.
На Сахалине, Южных Курилах и в Приморье живет номинальная форма: S. п.
unguiculatus Dobson, 1890. (Тип не выделен; описание основано на сборах Л. И. Шренка
с о-ва Сахалин.) Характерны крупные размеры ладоней и частое отрастание у ста-
рых зверьков когтей на пальцах передних конечностей (до 4,8 лш). Размеры крупные:
кондилобазальная длина черепа 19,0—20,7 мм; ширина черепа 9,8—10,8 мм. Длина
хвоста составляет около */2 длины тела. Второй верхний промежуточный зуб значи-
тельно (раза в полтора) меньше первого и равен или меньше третьего.
17. Альпийская бурозубка. Sorex alpinus Schinz, 1837
Табл. Ill, 10. Карта 12
Юг Западной Европы: Франция до Пиренеев, Германия, Швейца-
рия, Италия, Югославия, Чехословакия, Польша, Карпаты (включая
советские).
Обитатель альпийского и субальпийского поясов гор Южной и За-
падной Европы, по долинам рек и заболоченным участкам местами
опускается до подножий. Сырые места с высокотравьем предпочитает
другим местам обитания. В Восточных Карпатах редка.
В СССР встречается, видимо, номинальная форма: S. a. alpinus Schinz, 1837 (опи-
сана из Швейцарии).
18. Гигантская бурозубка. Sorex pacificus Cones, 1877
Табл. Ill, 11. Карта 12
Восточная Азия и запад Северной Америки (Орегон, Калифорния).
В СССР найдена пока в трех точках Южного Приморья: 1) в долине
р. Кишинки (впадающей с юга в оз. Ханка), 2) в заповеднике Кедро-
вая Падь и 3) в Судзухинском заповеднике.
Экология не изучена.
Изменчивость обширна, а номенклатура запутана. По экземпляру с р. Кишинки
(сбор С. А. Надецкого) С. И. Огнев (1937) описал особый вид: S. mirabilis. Мы не
могли отличить этот вид от подробного описания и прекрасных фотографий черепа
и зубов по монографии Н. Jackson, 1928, от S. pacificus Coues, распространенной
в Орегоне и Калифорнии.
Американские систематики: Findley, 1955, Hall and Kelson, 1959, объединили
S. pacificus в один вид с S. vagrans (-S. obscurus). Такое объединение недостаточно
обосновано. S. pacificus ( + S. yaquinae) хорошо отличается от S. vagrans своими
очень крупными размерами, что можно видеть по прилагаемой сравнительной таблице.
Размеры двух видов американских бурозубок (по Н. Jackson, 1928):
Длина ступни Кондилобазальная длина черепа . . Ширина черепа Максиллярный ряд зубов S. vagrans + 4- S. obscurus S. pacificus + । + S. yaquinae
II—15 16,1 — 19,1 7,8—9,1 5,5—6,9 16—17,5 20,0—23,0 9,7—10,8 7,4—8,8 i 1
66
В группе vagrans-obscurus есть узколокализованные островные формы (напри-
мер, S. v. malitiosus, S v. elassodon, S. v. pravostensis и др.), у которых ступня дости-
гает 16 лш, а длина хвоста 52—62 мм. Однако череп их не выходит за пределы
приведенных в таблице показателей. Удлиненные хвост и ступня характерны для
островных форм бурозубок и в нашей фауне (см., например, курильско-японскую
S. caecutiens shinto). У материковых же форм S. vagrans длина хвоста изменяется
от 38 до 50 мм, а у S. pacificus — от 54 до-67 мм.
За крупными, длиннохвостыми, темно окрашенными бурозубками мы сохраняем
(по правилу приоритета) название: Sorex pacificus Coues, 1877. Азиатскую форму,
вероятно, можно выделить в качестве подвида: S. р. mirabilis Ognev, 1937 (тип с р. Ки-
шинки, бассейн оз. Ханка).
РОД КУТОРЫ. GENUS NEOMYS .Каир/1829
Зубная формула: I -у Din -у Ртр у-М -у • 2 = 30.
Средних размеров землеройки, в большей или меньшей степени
приспособленные к полуводному образу жизни. Окраска меха обычно
двухцветная: верх и бока черные или черноватые, брюхо белое или бе-
ловатое; лишь изредка встречаются (на Карпатах) особи с темно окра-
шенным низом. Хвост длинный с различно развитым килем из удлинен-
ных волос вдоль нижней стороны хвоста.
Череп с выпуклой мозговой капсулой. Вершины зубов пигментиро-
ваны (как у бурозубок). Размеры верхних промежуточных зубов посте-
пенно уменьшаются от первого к третьему, а четвертый промежуточный
зуб значительно меньше третьего и обычно оттеснен к внутренней сто-
роне зубного ряда. Верхний край нижнего резца обычно с одной пологой
возвышенностью над его серединой.
Два близких вида кутор населяют среднюю и северную Палеар-
ктику. Оба представлены в фауне СССР.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ КУТОР
1(2). Размеры сравнительно мелкие: кондилобазальная длина че-
репа 17,9—21 мм; ширина черепа 9,3—10,6 мм; ступня 13,5—17,5 мм
(в одном случае 18). Киль из удлиненных волос на нижней стороне
хвоста совсем не развит или слабо выражен лишь на протяжении ко-
нечной трети хвоста; плавательные оторочки из волос на лапах тоже
развиты слабо .... Малая кутора. Neomys anomalus. (Стр. 67.)
2(1). Размеры крупнее: кондилобазальная длина черепа 21 —
23,2 мм; ширина черепа 10,7—12,3 жж; ступня в подавляющем большин-
стве случаев более 18 жж. Киль из удлиненных волос на нижней стороне
хвоста у взрослых особей хорошо развит и занимает обычно не менее
двух конечных третей длины хвоста; плавательные оторочки из волос
окаймляют ступню и обычно сильно развиты на всех пальцах . . . .
.....................Водяная кутора. Neomys fodiens. (Стр. 68.)
19. Малая кутора. Neomys anomalus Cabrera, 1907
Табл. Ill, 12. Карта 13
Испания, Южная Франция (Пиренеи), Швейцария, Италия, Авст-
рия, Чехословакия, Юго-Восточная Польша, Венгрия, Румыния, Юго-
славия, Албания, Греция, Турция, Болгария. В СССР — Украина, пре-
имущественно Правобережная, в полесских и лесостепных районах
Левобережья (Броварский и Березанский районы Киевской области,
в бассейне р. Ворсклы Полтавской области), в плавнях Днепра и
67
южной по.ловине Крыма; в Молдавии, Южной Белоруссии (Беловежская
пуща, Василевичский и Речицкий районы Гомельской области), Воро-
нежский заповедник. Всюду, за исключением Испании, Крыма и Одес-
ской области, она встречается вместе с водяной куторой.
Живет по берегам небольших рек и ручьев в широколиственных
лесах, преимущественно горных (Крым, Карпаты), у незамерзающих
зимой источников. С водоемами биологически связана не менее чем
водяная кутора. Биология размножения не изучена. Нередко встре-
чается в погадках сов.
Подвиды:
a) N. a. anomalus Cabrera, 1907. (Тип из окрестностей- г. Мадрида, Испания.)
Относительно крупная и длиннохвостая: длина хвоста 51—61 мм; длина ступни
16,4—17,5 мм (18); кондилобазальная длина черепа 19,9—21 мм-, длина верхнего
ряда зубов 9,7—10,7 мм. Горный Крым и плавни Днепра. Крымских малых кутор
В. и Е. Мартино (1917) описали под названием N. fodiens mocrzecki. Мы эту форму
не выделяем потому, что не знаем ее отличий от номинальной испанской формы.
b) N. a. milleri Mottaz, 1907. (Тип из Швейцарских Альп.) Размеры меньше:
длина хвоста 41—49 мм; длина ступни 13,5—15 мм; кондилобазальная длина черепа
17,9—19,5 мм; длина верхнего ряда зубов 8,2—9,5 мм. Северная, преимущественно
равнинная часть ареала вида.
20. Водяная кутора. Neomys fodiens Pennant, 1771
Табл. Ill, 13—14, Карта 13
От Норвегии, Великобритании, Франции на восток до устья Амура
и Сахалина, на север почти до берегов Ледовитого океана в Сканди-
навии до Полярного круга на северо-востоке Европы, до 60—62° с. ш.
в Западной Сибири, до Баргузинского побережья Байкала и низовьев
Амура. На юг лишь по долинам рек вклинивается в степную зону
Украины, Поволжья и Казахстана. В Крыму этого вида нет. Широко
распространена на Кавказе и в Закавказье до Батуми, долины Аракса
и Южного Азербайджана (несомненно, должна быть найдена в север-
ных районах Ирана и Турции). В горах Восточного Казахстана п
Киргизии идет до берегов Иссык-Куля, встречается в Северной Мон-
голии. На Кавказе поднимается в горы до альпийского пояса.
Поселяется обычно по берегам водоемов. Охотно и с большим
мастерством плавает и ныряет. Питается мелкими позвоночными — ля-
гушками, головастиками, детенышами мелких грызунов, земляными
червями, насекомыми. Вес корма, съедаемого за сутки, несколько пре-
восходит вес самого зверька. Голодать может больше, чем бурозубки
(гибнет от голода лишь на третьи сутки). Беременные самки встреча-
ются весной и в середине лета. Число эмбрионов от 4 до 10. Числа
приплодов в году пока точно не установлено.
Морфизмы, индивидуальная и географическая изменчивость у ку-
торы выражены слабее, чем у бурозубок.
Подвиды:
a) N. f. fodiens Pennant, 1771. (Описан по экземпляру из Берлина, Германия.)
Длина уха 7,8—9,8 мм. Киль на хвосте и оторочки на лапах хорошо развиты.
Европа, Западная Сибирь к востоку — до Оби и приалтайских степей; Кавказ п
Закавказье (без Дагестана).
b) N. Г. orientis Thomas, 1914. (Тип с Каменной речки, близ Джаркента.) От но-
минальной формы отличается лишь несколько меньшими размерами ушной раковины:
длина уха 6,5—7,7 мм. От правобережья Оби и Алтая к востоку до Охотского
моря и Сахалина.
с) N. f. dagestanicus Heptner et Formozov, 1928. (Тип из Гуниба, Дагестан, Восточ-
ный Кавказ.) По недоразвитому килю на хвосте и оторочкам на лапах хорошо отли-
чается от номинальной формы п приближается к малой куторе. Известна пока
только из Дагестана.
68
РОД МНОГОЗУБЫЕ БЕЛОЗУБКИ. GENUS SUNCUS Ehrenberg, 1833
Зубная формула: I Din у Pmp -j-M -2 = 30.
По четырем промежуточным зубам с каждой стороны верхней че-
люсти представители этого рода отличаются от настоящих белозубой
(рода Crocidura), хотя задний (четвертый) промежуточный зуб развит
у них слабо, обычно оттеснен к внутренней стороне зубного ряда и
в редких случаях даже может отсутствовать. К этому роду относится
представленная в нашей фауне карликовая белозубка S. etruscus —
самое маленькое из современных млекопитающих:' длина тела ее не
превышает 4,5 см, вес от 1,2 до 1,7 г (в среднем около 1,5 г), и к этому
же роду относят самую крупную из современных землероек — Suncus
murinus, распространенную в Южной и Юго-Восточной Азии; длина
тела ее более 12 см, а вес взрослых зверьков более 70 г. Есть виды и
средние по размерам.
Распространены в Африке, на Мадагаскаре, на юге Европы,
в Средней, Южной и Юго-Восточной Азии и на островах Тернат, Джа-
лоло, Тимор, Банда-Непра.
Всего различают около двух десятков видов. В фауне СССР
только один вид.
21. Карликовая белозубка. Suncus etruscus Savi, 1822
Табл. IV, 1. Карта 14
Южная Европа (Испания, Франция, Италия, Сицилия, Сардиния,
Венгрия, Греция), Алжир и Северная Нигерия, Аравия, Иран, Ирак,
Индия, Цейлон и страны Индокитая. В СССР — Закавказье (Ленко-
рань, Муганская степь, окрестности Нухи и Тбилиси), Юго-Западная
Туркмения, г. Термез.
Образ жизни совсем не изучен.
Географическая изменчивость в пределах Южной Европы, Закавказья и Средней
Азии не изучена. Номинальная форма (S. е. etruscus Savi, 1822) описана из Пизы,
Италия.
РОД БЕЛОЗУБКИ. GENUS CROCIDURA Wagner, 1832
Зубная формула: I у Din Pmp уМ у -2 = 28, редко 26.
Правильно расшифрованной формулы по общепринятым катего-
риям зубов в литературе пока не было. Что передний верхний резец
представляет собой не один зуб (I1), а продукт слияния (спайки)
в области основания коронок двух резцов (11+2), хорошо видно именно
у белозубок. У совсем молодых особей коронки развиваются раздельно.
Затем они соединяются, но вершины и корни их сохраняют свою обо-
собленность. Первый промежуточный зуб (Din1), видимо, внешний
резец (I3); он всегда значительно (в 2—5 раз и более) превосходит
по размерам два следующих за ним промежуточных зуба, которые
в отлцчие от крупного первого можно называть малыми промежуточ-
ными зубами; соотношение между ними (по размерам) сильно варьи-
рует: обычно Din2 крупнее, чем Din3, часто они равны между собой,
реже Din2 меньше, чем Din3. Теперь известны случаи отсутствия одного
из малых промежуточных зубов (у Cr. leucodon с Кавказа и Алтая)
то с обеих сторон, то лишь с одной стороны. Редуцируется, вероятно,
69
Din3. Din2, видимо, следует считать клыком, а задний промежуточный
(если он есть)—первым малым предкоренным зубом (Рт1). За ним
следует крупный предкоренной зуб (Ртр) с хорошо выраженной на
переднем крае коронки вершинкой, имеющей обособленный центр
окостенения. К какой категории относится первый нижний промежу-
точный зуб (dini), установить невозможно; условно его считают ма-
лым предкоренным (рт). Следующий за ним более крупный одновер-
шинный зуб мы склонны считать крупным предкоренным зубом (ртр).
Если изложенные предположения верны, то формула по общеприня-
тым категориям зубов для белозубок будет выглядеть так:
I —С—Pm-^-^Pmp —М—-2 = 28 или 26.
10 1 1 3
Распространены белозубки по всей Африке, Южной и Средней
Евразии до Японии, Тимора и Цейлона на востоке и на юге. В СССР —
от Эстонии, юга Московской и Пермской областей, Свердловска, Вер-
хотурья, Томска, Минусинской степи, Забайкалья до Приморья вклю-
чительно.
Биологически более всего связаны с открытыми ландшафтами
степей, полупустынь, пустынь и гор с зарослями кустарников и травя-
ным покровом. Редко заходят в леса умеренной полосы. Иногда посе-
ляются в постройках человека.
Систематика белозубок очень трудная и разработана слабо. Все признаки,
используемые в диагностике видов: размеры черепа и промежуточных зубов, соотно-
шение длины хвоста и тела,’ окраска меха и др., подвержены широкой индивидуаль-
ной и географической изменчивости. Видимо, сильно выражены у них безареальные
морфы; возрастные и сезонные морфизмы пока совсем не изучены.
Разработка систематики рода затрудняется недостатком коллекционных матери-
алов, особенно по зарубежным странам. Разновременно и неполноценно подобран-
ные серии (как в Академии наук Грузинской ССР, где обрабатывали зверьков после
длительного хранения в формалине) не уменьшили затруднений.
Ненадежность различий по размерам и соотношению частей показана ниже
в кратких описаниях форм. Здесь уместно лишь обратить внимание на обнаружен-
ное нами быстрое изменение окраски меха на тушках (набитых шкурках), хранящихся
в коллекциях.
В сентябре и октябре 1953 г. Л. Петровская собрала целую серию бурых бело-
зубок (Сг. russula) в Хосте (Черноморский берег Кавказа). В конце августа и сен-
тябре 1961 г. там же (в Хосте) было добыто несколько белозубок того же вида, и
они оказались отличными по окраске меха от добытых в тот же сезон в 1953 г.
в такой степени, в какой, по М. В. Шидловскому (1953), Сг. russula gtildenstaedti
Pall отличается от Сг. г. monacha Thomas. У первой формы и у зверьков, добытых
в Хосте в 1961 г., окраска меха мышино-серо-бурая почти без примеси каштановых и
коричневатых тонов, а у Сг. г. monacha, как и у добытых в 1953 г., мех коричневато-или
каштаново-бурый.
В нашем эталонном наборе и в Зоомузее МГУ есть белозубки (Сг. leucodon),
добытые в августе и сентябре 1959 г. на Алтае, в долине р. Вии в окрестностях
с. Кебезень. В 1961 г. сотрудниками той же экспедиции в тот же сезон (в ав-
густе— начале сентября), в том же месте (в окрестностях с Кебезень) добыто не-
сколько особей того же вида, и все свеже добытые отличаются по окраске меха от
добытых ровно два года назад как «хорошо обособленные подвиды»: добытые
в 1959 г. по окраске сходны с Сг. 1. sibirica Dukelsky (коричневато-бурые), а добытые
там же и в тот же сезон 1961 г. сходны с Сг. 1. ognevi Stroganov: темные мы-шино-серые
без примеси коричневатых тонов.
Что при хранении шкурок окраска меха меняется, было давно известно. Однако
изменение за такой короткий срок (как 8 лет и даже 2 года) до уровня отчетливых
подвидовых различий отмечается нами впервые, и это должно учитываться в диа-
гностике видов и географических форм белозубок.
Использование строения наружных гениталий взрослых самцов белозубок как
решающего признака в .диагностике видов представляется неприемлемым по ряду
причин:
1) Изменчивость наружных гениталий самцов (индивидуальная, географическая,
популяционная) и морфизмы (возрастные и сезонные) пока совсем не изучены. Поэ-
тому возведение в ранг «особого вида» на основе деталей строения головки «одного
70
взрослого самца, пенис которого, однако, был сильно деформирован при фиксации»
(Б. С. Виноградов, 1958, стр. 1239), мы не можем считать обоснованным. Речь идет
при этом о «белозубке Огнева» (Сг. leucodon ognevi), описанной даже С. У. Строгано-
вым (1956) в качестве подвида белобрюхой белозубки, отличающегося от Сг. 1. sibirica
Dukelsky (1930) по окраске меха, изменяющейся в этом именно направлении за очень
короткий срок хранения в коллекции (за два года). Кстати сказать, тип описания Сг. 1.
ognevi, добытый в июле 1928 г. и хранящийся в Зоомузее МГУ, отличается теперь по
окраске, детально описанной С. И. Огневым (1935, стр. 623), от самого же себя
больше, чем от шкурок «белозубки Огнева» двухлетней давности. Строение гениталий
сибирской белозубки совсем не известно. Разве можно признать приемлемым, научно
обоснованным описание такого вида?
2) Осенью и зимой взрослые самцы землероек составляют не более 10—20%
от общего количества добываемых зверьков (любого вида). Остальные 80—90% бело-
зубой обрекаются на «неопределимость», а хранящиеся в виде шкурок (с черепами)
экспонаты всех музеев мира становятся неопределимыми до вида на 100%- В дан-
ном случае пытаются ставить знак равенства не только между видом и признаком,
Рис. 10. Копии рисунков половых органов самцов одних и тех же видов:
а, б, в, — по С. У. Строганову, 1957 и Д, Б, В — по Б. С. Виноградову, 1958: Д, а —
малая белозубка (Crocidura suaveolens); Б, б — белобрюхая белозубка (Сг. leucodon);
В, в — большая (дальневосточная) белозубка (Сг. lasiura).
но даже между видом и десятой частью одного признака. Это уже не наука, не
систематика.
3) На рисунке 10 представлены фотокопии рисунков гениталий взрослых самцов
одних и тех же видов белозубок по С. У. Строганову (1957) и Б. С. Виноградову
(1958). Различия между изображениями так велики, что по ним особей одного вида
следовало бы относить к разным родам или даже семействам. То же относится и к
описаниям гениталий разными авторами (даже единомышленниками). Б. С. Виноградов
особое внимание обращал на роговые шипики, а С. У. Строганов о шипиках не
писал ни слова, он их просто не видел.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ БЕЛОЗУБОК ФАУНЫ СССР
1(2). Размеры мелкие: наибольшая длина черепа (от переднего
края крупных верхних резцов) 15,9—17}9 мм; кондилобазальная длина
черепа 15,2—17,3 мм (в редких непроверенных случаях до 17,7 —
17,9 мм). Бока окрашены одинаково с верхней стороной тела; нижняя
сторона (тела и головы) значительно светлее боков: вершинки волос
низа белые или со 'слабым охристым налетом...........................
.............Малая белозубка. Crocidura suaveolens. (Стр. 73.)
2(1). Размеры крупнее: наибольшая длина черепа (от переднего
края крупных верхних резцов) более 18 мм; кондилобазальная длина
черепа более 17,7—17,9 мм (в редких случаях от 17,2 и даже от
16,7 мм)...........................................................3
3(4). Хвост белый или беловатый. Длина хвоста 39—47,5 мм
(58}2—78% от длины тела). Окраска меха на спине п боках очень свет-
71
лая с короткими цветными вершинками; низ белый. Кондилобазальная
длина черепа 17,9—18,8 мм. Горы Средней Азии......................
...............Белохвостая белозубка. Crocidura pergrisea. (Стр. 74.)
4(3). Хвост темно окрашенный или двухцветный: сверху темный
и только снизу светлый. Окраска меха и размеры или соотношение
длины хвоста и тела иные........................................5
5(8). Окраска меха, лап и хвоста темная коричневато-бурая,
серо- или черно-бурая. Бока такие же темные, как верх; низ окрашен
немногим светлее боков; переход в окраске от темных боков к не-
сколько более светлому низу постепенный. Тыльная сторона лап окра-
шена под цвет верхней стороны, тела.............................6
6(7). Размеры средние: наибольшая длина черепа (от переднего
края верхних резцов) 18—21,3 мм; кондилобазальная длина черепа
(16,7) 17,5—21,0 мм; длина верхнего ряда зубов 7,6—9,6 мм (редко
от 7,3 мм). Кавказ, Закавказье..................................;
....................Бурая белозубка. Crocidura russula. • (Стр. 74.)
7(6). Размеры крупнее: наибольшая длина черепа (от переднего
края верхних резцов) 21,1—25,8 мм; кондилобазальная длина черепа
Рис. 11. Караван: самка малой белозубки (Crocidura suaveolens) со своими детенышами.
По фотоснимку Яна Ганзака
20,3—25,0 мм; длина верхнего ряда зубов 9,5—11,2 мм. Уссурийский
край и другие районы Восточной Азии..............................
.................Большая белозубка. Crocidura lasiura. (Стр. 75.)
8(5). Окраска меха иная: сверху обычно темная, серая или корич-
невато-бурая; бока (тела и головы) двухцветные: темные сверху и бе-
ловатые снизу с резкой прямолинейной границей между ними; если
бока темные, то хотя бы короткие вершинки волос низа беловатые и
граница между окраской боков н низа отчетливая. Большая часть
тыльной поверхности лап окрашена под цвет нижней стороны тела (бе-
лые или белесые) или темно окрашен только наружный край лапы и
внешний палец (мизинец) до когтя...............................9
9(10). Размеры относительно крупные: кондилобазальная длина
черепа 20,1—21,8 мм; длина верхнего ряда зубов 9,6—10 мм. Западное
Закавказье...........Серая белозубка. Crocidura lasia. (Стр. 76.)
10(9). Размеры средние: кондилобазальная длина черепа (17,7)
17,9—20,2 мм; длина верхнего ряда зубов (7,5) 7,8—9,6 мм (очень
редко до 9,8 мм...............................................11
11(12). Верхние малые промежуточные зубы (второй п третий)
относительно крупные: по высоте они лишь в 2—3 раза уступают пер-
вому промежуточному, а суммарная длина их оснований (смотреть
снизу или с внутренней стороны) больше продольного диаметра пер-
вого промежуточного зуба. Бока тела и хвост резко двухцветные:
с прямолинейной границей между темным верхом и белым низом
............Белобрюхая белозубка. Crocidura leucodon. (Стр. 76.)
12(11). Верхние малые промежуточные зубы (второй и третий)
очень мелкие: по высоте они в 4—5 раз ниже первого промежуточного
зуба. У некоторых особей одного из малых промежуточных зубов во-
обще не бывает. Бока тела темно окрашенные (серо-бурые или корич-
певато-бурые); светлые вершинки волос очень короткие, поэтому низ
кажется тоже темно окрашенным; граница между окраской боков и
низа не очень отчетливая. Хвост темно окрашенный (серо- или корич-
невато-бурый), почти одинаковый сверху и снизу, но тыльная сторона
лап, 'кроме внешнего края и мизинца, белая или беловатая..............
Сибирская белобрюхая белозубка. Сг, leucodon sibirica. (Стр. 77,)
22. Малая белозубка. Crocidura suaveolens Pallas, 1811
Табл. IV, 2—3. Карта 14
Западная и Средняя Европа. В СССР—к северу до Эстонии, юж-
ных районов Московской, Горьковской, Пермской областей, Троицка,
Челябинской области; в Казахстане до Семиозерного района Куста-
найской области, до широты Целинограда и Караганды; Прибалхашье
и Джунгарский Алатау 1 и Приморский край (Супутинский заповедник,
Хасанский район, долина р, Сучан). Известна из Кореи, Монголии,
Ирана, Палестины, Алжира, Марокко.
Местами обычный (по численности) вид.
Живет в различных ландшафтах: в песчаных и глинистых пусты-
нях (Кара-Кумы, Аральское побережье), в оазисах пустынь, в степях
(Европы, Монголии), на обширных заболоченных лугах Приморья,
в кустарниковых зарослях, в разреженных лесах и садах, на посевах
зерновых культур. Нередко поселяется в жилье человека. Молодых
белозубок находили в разные части теплого периода года. В одном
приплоде бывает от 5 до 10 детенышей. Размножается в условиях не-
воли (например, несколько приплодов получено в лаборатории док-
тора Яна Ганзака в Национальном музее в Праге), Подрастающий
молодняк при передвижениях выстраивается в «караван»: детеныш
цепляется зубами за кожу самки, другой детеныш — за первого, тре-
тий— за второго и т, д. (см. рис, 11), Питается главным образом мел-
кими насекомыми. Вес корма, съедаемого за сутки, почти в полтора
раза (на 133%) превосходит вес самого зверька. Без пищи может быть
немногим более суток (до 28 часов).
Малая белозубка характеризуется относительно мелкими размерами черепа
и темно окрашенными боками тела с не очень резким переходом к светлой беловатой
или охристо-белесой окраске низа. Однако все эти отличия не абсолютны. По нашим
промерам (25 экземпляров из разных мест) и промерам А. К. Темботова (1957, 15 экземп-
ляров из Кабардино-Балкарии) кондилобазальная длина черепа 15,4—17,1 мм-. па
промерам С. У. Строганова (1957) у европейских и восточноазиатских особей этого
вида кондилобазальная длина черепа 15,2—17,0 лии, но по С. И. Огневу (1928) —
16,0—17,6—17,8 мм; у среднеазиатских малых белозубок, по С. У. Строганову
и С. И. Огневу, — до 17,8—17,9 мм. Если эти максимальные показатели правильны,,
то по размерам черепа становятся неотличимыми кавказские малые белозубки
(Сг. s. dinniki Ognev), кондилобазальная длина которых 17,6—17,9 мм. Последние отли-
чаются от малых белозубок из других мест только более длинным хвостом (по С. И. Ог-
неву — 37,3—44,4 мм). Через относительно крупную и длиннохвостую кавказскую
форму (Сг. s. dinniki) малая белозубка становится практически неотличимой от закав-
казских же мелких особей бурой белозубки (Сг. russula gtildenstaedti), трансгрессирует
по размерам черепа с мелкими особями белобрюхой белозубки (Сг. .leucodon); вплот-
ную примыкает по размерам черепа и хвоста к ирано-памирской белохвостой бело-
зубке (Сг. pergrisea). Еще менее отчетливы и менее устойчивы различия в окраске
меха, которая у малой белозубки варьирует от очень светлой, палево- или голубовато-
серой до темной, коричнево-бурой, что перекрывает различия в окраске между боль-
шинством форм белозубок фауны СССР. Широкая индивидуальная изменчивость,
1 Б, С. Виноградов (1958) указывал Алтай, Северную Монголию и южную часть
Бурятской АССР; нам сборы из этих мест не известны. Не проверены также прежние
указания Сабанеева о находках этого вида в окрестностях Свердловска, Верхотурья
п бывшего Богословского завода (на Урале).
73-
сезонные и возрастные морфизмы осложняют географическую изменчивость и диагно-
стику подвидов. Например, восточноазиатские малые белозубки по размерам и соотно-
шению частей варьируют в тех же пределах, как европейские, и по окраске меха там
встречаются особи, неотличимые от европейских (и от крымских, в частности). Поэтому
восточноазиатская форма (Сг. s. shantungensis Miller-1901 = Сг. s. orientis Ognev, 1921)
практически неотличима от номинальной.
В фауне СССР можно различить 3 подвида:
а) Сг. s. suaveolens Pallas, 1811. (Тип из Херсонеса Таврического, близ Севасто-
поля, Крым.) Мелкая, короткохвостая форма с темно окрашенным верхом и боками
тела: кондилобазальная длина черепа 15,2—17,3 мм; длина хвоста 24—37 мм. Крым,
большая часть европейского сектора ареала, Кабардино-Балкария, Приморье, Корея,
Внутренняя Монголия.
Ь) Сг. s. ilensis Miller, 1901. (Тип из Куктурука, на р. Или.) Размеры немного
крупнее номинальной формы, по С. И. Огневу. (1928) и С. У. Строганову (1957):
кондилобазальная длина черепа от 16,3—16,7 до 17,8—17,9 мм. Хвост короткий:
до 37,3 мм. Окраска меха очень светлая, наверху и боках голубовато- или палево-
серая; низ белый. Пустыни Казахстана, Средней и Центральной Азии. Неясны отличия
этой формы от ранее описанной Сг. s. lignicolor Miller, 1900. (Тип из Маралбаши,
на р. Яркенд.) Синоним: Сг. s. hyrcana Goodvin, 1940. (Тип с р. Горган, Северо-
Восточный Иран.)
с) Сг. s. dinniki Ognev, 1921. (Тип из Ставрополя, Северный Кавказ.) Относи-
тельно крупная и длиннохвостая: кондилобазальная длина черепа 17,3—17,9 мм;
длина хвоста 37,3—44,4 мм (по С. И. Огневу, 1928). Окраска меха яркая с корот-
кими цветными кончиками волос. Ставрополь, разные места Закавказья (Абастумани,
в Западной Грузии, Тбилиси; Кировабад, разные районы Армении), Северный Иран.
Неясными остаются отличия кавказских малых белозубой от ранее описанной
Сг. s. mimula Miller, 1901. (Тип из Швейцарии.)
Белохвостая белозубка. Crocidura pergrisea Miller, 1913
Табл. IV, 4.
Южный и Восточный Иран, Копет-Даг, Памир, Гиссарский хребет
и, вероятно, другие горы Таджикистана.
Биология совсем не изучена.
Сг. pergrisea описана из Южного Ирана (Белуджистана). В музеях СССР нет
экспонатов, соответственных номинальной форме. Здесь белохвостая белозубка выде-
лена условно (без порядкового номера), так как ее систематическое положение остается
недостаточно выясненным.
Размеры средние: кондилобазальная длина черепа 17,9—18,8 мм. Хвост относи-
тельно длинный: 39—47,5 мм, что составляет ат 58,2 до 78% от длины тела.
Окраска меха очень светлая с яркими вершинками волос, создающих характерную
цветистую рябь; низ белый; переход от ярко окрашенных боков к белому низу
постепенный. Хвост беловатый или чисто белый как снизу, так и сверху. По изло-
женному отличия могут показаться достаточными. Однако по размерам она вплотную
примыкает к малой белозубке (Сг. suaveoleus), особенно к ее кавказской форме
(смежной географически), от которой не отличается по длине хвоста (и ступни); по
окраске меха слабо отличается от западно-копетдагских малых белозубой. Трудно
отличить ее и от светло окрашенных белобрюхих белозубок (Сг. leucodon), кондило-
базальная длина черепа которых от 17,7—17,9 до 20,2 мм; длина хвоста до 43 мм,
что составляет до 63,3% от длины тела. По размерам и длине хвоста она неотли-
чима также от закавказских (в частности, восточногрузинских) Сг. russula giilden-
staedti; различия их выражены только в окраске (меха, хвоста); географически они
сменяют друг друга, и более светлая окраска обитателей гор пустынной зоны нор-
мальна и естественна для подвидов; имея в виду это, мы трактовали (в первом
издании определителя) памирских белозубок в качестве подвида Сг. russula.
23. Бурая белозубка. Crocidura russula Hermann, 1780
Табл. IV, 5. Карта 15
Западная и Средняя Европа (Польша, Бельгия, Голландия, Герма-
ния, Южная Болгария, Швейцария, Италия, Франция, Испания, Пор-
тугалия), острова Средиземного моря (Сардиния, Корсика, Крит), Се-
верная Африка (Марокко, Алжир, Тунис), Палестина, Малая Азия,
Иран, Афганистан и далее к востоку до Японии.
74
В СССР эта белозубка особенно характерна для Закавказья, где
она численно преобладает над всеми другими видами землероек. На
Черноморском побережье Кавказа и в предгорьях Талыша (в Ленко-
рани) она поселяется в зарослях ежевики вдоль берега моря, по бере-
гам водохранилищ (в Ленкорани) и горных речек. Заходит в широко-
лиственные леса нижнего пояса гор и местами поднимается до альпий-
ских лугов. Встречается в постройках человека (например, в Тбилиси).
По К- А. Сатунину, гнездо, построенное из травы, выстилает пухом и
перьями; самка приносит от 5 до 10 детенышей. Питается насекомыми,
их личинками и червями. Охотно идет на приманку из хлеба с расти-
тельным маслом.
Бурая белозубка от других средних (по размерам) белозубой отличается только
темной коричневато-бурой или серовато-бурой окраской меха наверху и боках тела
и головы; нижняя сторона тела (и головы) окрашена немногим светлее боков и верха
без отчетливой границы между окраской боков и низа. Хвост темно-бурый, почти
одинаковый сверху и снизу. Волосы на тыльной стороне лап окрашены под цвет
спины. По бурой окраске, как наиболее характерному признаку, мы и предлагаем
называть эту белозубку. Ранее данное русское название — «длиннохвостая» (по кав-
казской форме) оказалось менее характерным. Номинальная форма не длиннохво-
стая, а у кавказских особей этого вида длина хвоста варьирует от 35—38 до 60 мм,
что составляет от 39,3—45,5 до 88,3% от длины тела (с головой). Эти показатели
перекрывают абсолютную и относительную длину хвоста всех форм белозубок
фауны СССР.
При широкой индивидуальной изменчивости географическая изменчивость выра-
жена слабо, хотя описано более 30 подвидов. Номинальная форма — Сг. russula
Hermann, 1780 — описана из окрестностей Страсбурга, Восточная Франция. Длина
хвоста (по Г. С. Миллер, 1912) 33—45 мм (в среднем 35,6—41 лш); ступня 12,2—
13,9 мм (в среднем 12,8 льч). Из нескольких подвидов, описанных по экземплярам
из Северной Турции, Северного Ирана и Закавказья, по сериям и средним показа-
телям размеров можно признать две очень слабо различающиеся расы:
а) Сг. г. giildenstaedti Pallas, I8II. (Тип из Душети, к северу от Тбилиси,
Восточная Грузия.) Преобладают особи с относительно длинным хвостом: от 38 до
60 мм (в среднем около 47 льи), что составляет о г 49,5 до 88,3% (в среднем 66,4%)
о г длины тела. Размеры, как у номинальной формы: кондилобазальная длина черепа
(16,7) 17,5—20,0 мм; ширина черепа 7,7—9,7 мм (М8,9 мм); высота черепа 4,8—
5,6 мм (М 5,3 мм). Синонимы этой формы: Сг. longicaudata Tichomirov et Kortchagin,
1889 (Сухуми, Черноморский берег Кавказа); Сг. г. monacha Thomas, 1906 (тип из
окрестностей Трабзона, юго-восточный берег Черного моря, Турция). Попытка
М. В. Шидловского (1953) восстановить Сг. г. monacha в качестве особого подвида
осталась не подкрепленной фактами: по размерам эти формы неотличимы, а разница
в окраске меха не превосходит изменений, какие теперь точно зафиксированы за
короткий срок хранения в коллекции. Среднее Закавказье и Черноморское побережье
Кавказа и Турции.
Ь) Сг. г. caspica Thomas, 1906. (Тип с южного берега Каспийского моря, Север-
ный Иран.) Отличается преобладанием особей относительно короткохвостых и круп-
ных по размерам черепа. Длина хвоста 35—50 мм, что составляет от 39,3 до 65,3%
(в среднем 55,5%) от длины тела. Кондилобазальная длина черепа 18,6—20,8 мм
(М 19,8 мм); ' ширина черепа 8,9—10,2 мм (М 9,5 мм); высота черепа 5,4—6,4 мм
(М около 5,7 мм). Неясно отличие этой формы от Sorex (Crocidura) fumigatus
de Filippi, 1863, описанной по экземпляру из Тегерана (Северный Иран). Вероятно,
они тождественны, и по приоритету эта форма должна называться Сг. russula fumi-
gata de Filippi, 1863. Ленкоранская низменность. Талыш и прикаспийские районы
Северного Ирана.
24. Большая белозубка. Crocidura lasiura Dobson, 1890
Табл. IV, 6. Карта 15
Южные районы Приморского края (Ханкайская низменность, при-
хасанские луга и др.), Корея и южнее расположенные районы Восточ-
ной Азии.
Живет на сырых вейниковых лугах и болотах (в Приморье),
в зарослях кустарников по берегам речек (в бассейне р. Сунгари).
75
.Местами нередка. Как все землеройки, очень прожорлива. В неволе
деятельна почти круглые сутки: охотно поедает предложенных ей бабо-
чек, земляных червей, тушки грызунов и т. д. На очень короткое время,
но многократно в течение суток она засыпает.
Взрослые большие белозубки по окраске неотличимы от бурых белозубок; окра-
ска меха молодых темнее: черно-бурая. Хвост относительно короткий: 33—59 мм
(М 41 мм), что составляет 32—60,8% (М около 45%) от длины тела; у некоторых
особей хвост сильно утолщен. Размеры отдельных крупных взрослых зверьков бывают
значительными: длина тела до 124 мм; ступня до 18 мм; кондилобазальная длина
черепа до 25,0 льн, ширина черепа до П,5 мм; высота черепа до 7,0 мм. Однако
мелкие особи по размерам (и окраске) не отличаются от крупных особей бурой бело-
зубки, особенно ее прикаспийской формы (Сг. russula caspica), что правильно отме-
тили Эллерман и Моррисон-Скотт (1951).
В южных районах Восточной Азии землеройки (любые виды бурозубок и малая
белозубка) не образуют географических форм, отличающихся от европейских круп-
ными размерами. У европейских бурых белозубок (Сг. russula) не наблюдается
и резко выраженного полиморфизма в размерах тела и черепа, свойственного большой
белозубке. Эти обстоятельства могут быть основанием для видового обособления
Сг. lasiura. В Японии встречаются оба эти вида. Не исключена возможность, что оба
они будут отмечены и в материковых условиях Восточной Азии.
Серая белозубка. Crocidura lasia Thomas, 1906
Турция и Черноморский берег Закавказья (тип из Скалита, близ
Трабзона, юго-восточный берег Черного моря, Турция).
Места обитания и другие стороны биологии не изучены.
По размерам сходна с большой белозубкой, от которой отличается
только окраской меха (серая сверху, беловатая снизу и с резкой гра-
ницей на боках). От сходной по окраске белобрюхой белозубки отли-
чается только размерами черепа и то незначительно, особенно от круп-
ных особей смежной географически иранской формы (Сг. leucodon рег-
sica). Восстановление ее в качестве особого вида было бы недостаточно
обоснованным, а проверить пока невозможно из-за недостатка коллек-
ционных материалов.
25. Белобрюхая белозубка. Crocidura leucodon Hermann, 1780
Табл. IV, 7—10. Карта 16
Западная и Средняя Европа (Франция, Бельгия, Голландия, Гер-
мания, Польша, Швейцария, Италия, Чехословакия, Югославия, Бол-
гария), Северный Иран, юг Монголии, В СССР — Украина (включая
Крым), Курская область, Нижнее Поволжье, Кавказ и Закавказье,
Юго-Западная Туркмения, Южный Таджикистан, Южный' Казахстан
(Семиречье), Алтай и другие районы Западной Сибири.
Живет в различных ландшафтах: в степях (Европа), в оазисах
пустынной зоны (Туркмения), на высокотравных лугах и в пойменном
лесу (Кабардино-Балкария), в сосновых борах (на Оби), в черневой
тайге (на Алтае), в большом городе (Алма-Ата), Другие стороны об-
раза жизни не изучены. С. У. Строганов (1957) пишет, что молодые
«встречаются в течение всего весенне-летнего периода. Число детены-
шей в помете от 5 до 10». 7
Белобрюхая белозубка, средняя по размерам, относительно короткохвостая, свет-
лоногая, с отчетливой линией на боках тела и головы между темной окраской верха
п светлой низа. Но она с трудом отличается от большинства видов белозубок фауны
СССР. Мелкие (молодые) особи по размерам черепа, длине хвоста и деталям окра-
ски почти неотличимы от крупных особей малой белозубки (Сг. suaveolens). Часть
особей из Закавказья практически неотличима от светлобрюхих (длительно храня-
76
щихся в коллекциях) экземпляров бурой белозубки (Сг. russula). Некоторые особи
могут быть практически неотличимыми от белохвостой белозубки (Сг. pergrisea).
Очень близка по размерам и сходна по другим признакам с закавказско-турецкой
серой белозубкой (Сг. lasia); с последней она географически викарирует (замещает).
От всех средних по размерам белозубок она отличается фактически лишь деталями
окраски меха, диапазон изменчивости которой почти не изучен. У бурозубок, напри-
мер, особей с «чепрачной» окраской меха не выделяют в качестве особых видов,
а белобрюхая белозубка отличается от сходной с нею по другим признакам бурой
белозубки только широким «чепраком». Не исключена возможность, что белобрюхая
белозубка представляет собой лишь один из типов изменчивости окраски бурой
белозубки (Сг. russula).
Однако географическая изменчивость этой формы выражена относительно хорошо.
По имеющимся материалам, можно различить (в пределах СССР) 4 подвида:
а) Сг. 1. leucodon Hermann, 1780. (Тип из окрестностей Страсбурга, Восточная
Франция.) Окраска верха коричневато-бурая. Размеры средние: наибольшая длина
черепа 19,1—20,8 мм; кондилобазальная длина 18—20 мм; ширина черепа 8,6—10 мм;
длина верхнего ряда зубов 8,0—9,3 мм. Большая часть Европы, включая Европейскую
часть СССР, и Кавказ.
b) Сг. 1. persica Thomas, 1907. (Тип из системы Эльбурса, гора Демавенд, около
2000 м над у. м., Северный Иран.) От номинальной формы отличается несколько
более крупными размерами: длина верхнего ряда зубов 9,0—9,8 мм; наибольшая длина
черепа до 21,4 мм. По окраске сходна с предыдущей. Горы Северного Ирана, Запад-
ный Копет-Даг (долины рек Сумбара, Чандыра) и, вероятно, Южное Закавказье.
Б. С. Виноградов (1958) на основе деталей строения гениталий самцов из Запад-
ного Копет-Дага возводил эту форму в ранг особого вида, не зная, однако, строе-
ния гениталий самцов из Ирана и Западной Европы.
с) Сг. 1. heptapotamica Stroganov, 1956. (Тип из г. Алма-Аты.) Размеры относи-
тельно мелкие: кондилобазальная длина черепа (по С. У. Строганову, 1956) 17,2—
18,3 мм; окрашена «темнее, чем номинальная форма», однако окраска меха имеюще-
гося у нас экземпляра (пз Алма-Аты, т. е. из места описания типа, взрослая самка
из сборов Зернова, 28.XI.1940 г.) очень светлая и яркая, сверху золотистая, снизу
белая. Соотношение этой формы с Сг. myoides Blanford, 1875 (описанной из Ле,
Кашмир, Северная Индия) остается недостаточно ясным.
d) Сг. 1. sibirica Dukelsky, 1930. (Тип из окрестностей с. Означенное, на р. Енисее,
в 96 км к югу от Минусинска.) Размеры, как у предыдущей формы (относительно
мелкие): кондилобазальная длина черепа 17,7—19,3 мм. Окраска меха очень темная:
у свежедобытых зверьков темно- или черновато-серая, на длительно хранившихся
шкурках темная коричневато-бурая. Вершинки волос на нижней стороне тела и ниж-
ней части боков беловатые, но они так коротки, что едва покрывают темные основа-
ния волос, и общий фон низа кажется поэтому бледно-серым или охристо-серым.
По этой же причине уменьшается контраст между окраской верха и низа, хотя
граница между ними отчетлива. Хвост темный, почти одноцветный, но тыльные сто-
роны лап темные только по внешнему краю (с мизинцем), а на остальной части
белые. Характерны малые размеры второго и третьего верхних промежуточных зубов:
они до 4—5 раз уступают по высоте первому промежуточному зубу; иногда одного
пз малых промежуточных зубов вообще не бывает (с одной или обеих сторон). Тип
добыт на берегу Енисея среди камней. В пойме Оби живет в сосновых борах, а на
Алтае — в черневой (пихтово-осиновой) тайге. Синоним этой формы: Сг. leucodon
ognevi Stroganov, 1956 (тип из окрестностей с. Боровлянка Троицкого района Алтай-
ского края). Сибирская часть ареала вида. Новосибирская, Томская, Кемеровская
области, Алтай, Минусинский район Красноярского края и, вероятно, юго-восточный
берег оз. Байкал (Бурятия).
Сибирская форма белобрюхой белозубки отличается от других форм этого вида,
как и от других близких видов лучше, чем последние различаются между собой.
Особенно показательны и устойчивы для этой формы мелкие размеры малых верх-
них промежуточных зубов, необычное сочетание почти одноцветного темно окрашен-
ного хвоста с контрастной окраской (белой и черноватой) тыльной стороны задних
лап. Своеобразны и места обитания (боры, горные пихтово-осиновые леса), и поло-
жение ареала (в суровых условиях Сибири, где других видов белозубок нет). Накоп-
ляющийся материал, быть может, даст основание для выделения этой формы в каче-
стве особого вида.
В 1961 г. на Восточном Кавказе, в низовьях р. Самур, Г. Космиади добыл
замечательную белозубку. По размерам она похожа на мелких белобрюхих белозубок.
Окраска верха и боков серая; низ беловатый; граница красок на боках слабо выра-
жена. Хвост короткий, двухцветный. Тыльная сторона задних лап белая с темными
полосками у внешней стороны пяток. Череп относительно короткий (наибольшая
длина 18,8 мм; кондилобазальная длина 17,8 мм), широкий (8,9 мм), плоский (5,0 лш).
Пары малых промежуточных зубов нет; всего зубов 26 (формула, как у путорака).
Верхний задний коренной сильно упрощенного строения.
77
РОД ПУТОРАКИ. GENUS DIPLOMESODON Brandt, 1852
Зубная формула: I у Din у Pmp у М у • 2 = 26.
От всех землероек отличаются своеобразной окраской: вся нижняя
сторона тела и головы, бока, лапы, хвост чисто белые; верх головы и
спина серые; на середине спины большое белое пятно.
Всего 1 вид.
26. Пегий путорак. Diplomesodon pulchellum Lichtenstein, 1823
Табл. IV, 11. Карта 16
Известен только из песчаных пустынь Средней Азии и Казахстана.
Найден в песках между низовьями Волги, Урала и Эмбы, в песках
Большие Барсуки (к северу от Аральского моря), на северо-западном
берегу Арала, на полуострове Дарджа (юго-восточный берег Каспий-
ского моря), в Каракумах, Кызылкумах, в песках к югу от р. Чу и в пе-
счаной пустыне к югу от Балхаша, между реками Или и Аксу.
Живет среди сыпучих и слабозадерненных барханных и бугристых
песков. На поверхности наблюдался только в темную половину суток.
Бегает, в отличие от других землероек, медленно, становится легкой
добычей ночных хищников и, несмотря на относительную малочислен-
ность, в погадках сычей встречается в большом количестве. Хорошо
роет и может быстро зарываться в песок. Питается, по-видимому, на-
секомыми, мелкими ящерицами. Период размножения с апреля до ав-
густа. Среднее число детенышей 5. В один сезон размножения самка
приносит, вероятно, не один приплод.
Индивидуальная изменчивость окраски выражена главным образом в размерах
белого пятна на спине. Известны полные альбиносы. Географическая изменчивость
слабая. Для описания D. р. pallidus Heptner, 1938 (по экземпляру из Учаджи, Вос-
точные Каракумы) не было достаточных оснований.
А. П. Кузякин
ОТРЯД РУКОКРЫЛЫЕ. ORDO CHIROPTERA
Рукокрылые — мелкие и средних размеров млекопитающие, спо-
собные к настоящему длительному полету. Передние конечности их
видоизменены в крылья: предплечье, пястные (метакарпальные) кости
и фаланги всех пальцев (кроме первого) чрезмерно удлинены; между
плечом, предплечьем, пальцами, боками тела и задними конечностями
натянута тонкая эластичная летательная перепонка. Задние конечно-
сти вывернуты так, что колени обращены дорзально. Ушные раковины
относительно большие, у многих с хорошо развитым козелком. Хвост
у большинства видов длинный, полностью или частично заключенный
в межбедренную, перепонку; свободный край этой перепонки поддер-
живается парной хрящевой или костной шпорой, отходящей от пятки.
Вдоль основания шпоры у многих видов тяйется своеобразная кожная
лопасть — эпиблема (рис. 12, Ер). Межчелюстные кости всегда недо-
развиты или даже отсутствуют. В зубной системе есть все категории
зубов. Средняя пара верхних резцов всегда отсутствует. Нижние резцы
очень мелкие. Клыки (особенно верхние) большие, типичной для хищ-
Рис. 12. Правильно расправленная шкурка малой вечерницы (Nyctalus
leisleri):
I—V — первый — пятый пальцы передней конечности, Е — боковая перепонка (endo-
patagium); Ер — эпиблема (epiblema), IF3 — первая фаланга третьего пальца,
2F3 — вторая фаланга третьего пальца, Mt3 — метакарпальная (пястная) кость третьего
пальца, МП — та же кость четвертого пальца, R — предплечье, Тг — козелок, Up —
межбедренная перепонка, z — место прикрепления свободного края крыловой пере-
понки к ноге.
79
ников формы. Коренные зубы делятся на три естественные группы
(рис. 13): малые предкоренные — praemolares (Pm и pm) мелкие,
одновершинные, конические, каждый с единственным корнем; число
их варьирует и имеет большое значение в распознавании родов и ви-
дов. От многобугорчатых задних коренных зубов — molares (М и ш)
они отделены характерными для рукокрылых большими предкорен-
ными — praemolares prominantes (Ртр и*-ртр), вершины которых
почти достигают уровня вершин клыков; каждый снабжен двумя кор-
нями. Наиболее полная зубная формула выглядит так:
I Ас-Рт - Ртр -М —-2 = 38.
3 1 2 13
Молочные зубы резко отличаются от постоянных. Все виды нашей
фауны питаются насекомыми, которых схватывают и поедают на лету.
Рис. 13. Череп большой ночницы. (Myotis myotis Borkh). Вид сбоку.
Зубы Г—Р, i\—1'3 — резцы (incisivi); С, с — клыки (canini), Рт'—Рт2 и pmt—
рт2 — малые предкоренные (praemolares); Ртр, ртр — крупные предкоренные
(praemolares prominantes); М1— М3, т,—т3 — заднекоренные (molares).
В связи с характером пищи, содержащей твердые хитиновые образова-
ния, эпителий пищевода ороговевает. Кишечник необычайно короткий
(лишь в 1,5—4 раза превышающий длину тела). Характерна крайняя
бедность кишечной флоры. Ориентация в пространстве осущест-
вляется главным образом за счет восприятия отраженных ультразвуко-
вых сигналов (эхолокация). Издают они ультразвуковые сигналы не-
зависимо от слышимых звуков и независимо от акта дыхания (как при
выдохе, так и при вдохе). Диапазон слышимости очень широкий — от
12 до 100 000 гц (колебаний в секунду). Это указывает на исключи-
тельно высокую остроту слуха, тогда как зрение у большинства раз-
вито очень слабо, и летучие мыши видят плохо независимо от времени
суток. В странах с умеренным климатом рукокрылые совершают се-
зонные перелеты и в подходящих убежищах впадают в зимнюю спячку.
Температура тела летучей мыши может изменяться (в условиях экспе-
римента) в пределах 56° (от — 7,5 до +48,5°). Весьма характерно
продолжительное (с осени до весны, т. е. до 6—7 месяцев) «пережива-
ние» сперматозоидов в семенных канальцах самцов и в половых путях
80
самок. В широтах с умеренным климатом ежегодно бывает лишь одна
генерация: самка родит только одного или двух (в исключительных
случаях трех) детенышей.
Изменчивость и морфизмы кратко можно характеризовать сле-
дующим образом. Развитие молодняка протекает очень быстро. На
третью — шестую неделю жизни молодые уже достигают размеров
родителей, сохраняя отличие лишь в более темной и тусклой окраске
ювенального меха и в хрящевых образованиях на концах длинных
костей (пястных, фаланг). После первой (ювенальной) линьки, которая
заканчивается в возрасте одного-двух месяцев, молодой уже й по окра-
ске теряет отличие от взрослых. Индивидуальная изменчивость незначи-
тельна; большинство признаков показывает поразительную устойчи-
вость. Сезонные морфизмы проявляются лишь в характере (высоте,
шелковистости), меха и в тоне или (реже) цвете его окраски. Геогра-
фическая изменчивость (размеров, окраски и реже структурных при-
знаков) у многих-видов отчетлива. Половой диморфизм у видов нашей
фауны совсем не выражен или выражен очень слабо (у подковоносов,
например, самцы несколько мельче самок). Не редок полиморфизм
окраски.
Распространение отряда охватывает весь земной шар до полярных
границ древесной растительности. Рукокрылыми не заселены Крайний
Север, Антарктика и некоторые океанические острова. Наиболее мно-
гочисленны и разнообразны рукокрылые в тропических и субтропиче-
ских областях.
Отряд рукокрылых делится на два хорошо обособленных подот-
ряда: 1) Крыланы (Megachiroptera)—плодоядные формы с примитив-
ными чертами организации (например, на втором пальце крыла всегда
имеется концевая фаланта, обычно 'снабженная когтем). Распростра-
нены в тропиках и субтропиках только восточного полушария. Около
150 форм крыланов объединяются в одно семейство — Pteropidae. 2) Ле-
тучие мыши (A^icrochiroptera)—в основной массе насекомоядные,
реже плодоядные, хищные и кровососущие формы с более специализи-
рованной организацией (на втором пальце крыла концевой фаланги
нет, усложнен летательный аппарат и т. д.). Ареал подотряда совпа-
дает с ареалом всего отряда. Около 800 видов летучих мышей группи-
руются в 15 семейств, из которых в фауне СССР представлены только 3.-
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ ЛЕТУЧИХ МЫШЕИ,
ПРЕДСТАВЛЕННЫХ В ФАУНЕ СССР
• 1(2). На лицевой части головы имеются сложной формы голые
кожистые образования (рис. 15). Ухо в виде свернутого в воронку и
заостренного на конце листика. Козелка нет. Очень тонкий, целиком
заключенный в межбедренную перепонку хвост в ненапряженном со-
стоянии отгибается к верхней стороне тела. У самок в паховой области
имеется пара сосцевидных придатков. Очень мелкие верхние резцы
(всегда в числе одной пары) расположены на хрящевой пластинке.
Нижних резцов 2 пары . . . Подковоносые. Rhinolophidae. (Стр. 82.)
2(1). На лицевой части головы нет сложных голых кожистых об-
разований. Ухо с округленной вершиной. Козелок есть. Хвост в нена-
пряженном состоянии отгибается к нижней стороне тела. У самок
в паховой области сосцевидных придатков нет. Верхние резцы (в чис-
ле одной или двух пар) сидят в межчелюстных костях. Нижних резцов
3 пары.......................................................................3
6 Н. А. Бобринский и др. 81
3(4). Тонкий хвост весь заключен в межбедренную перепонку или
выступает из нее на длину 1—2 концевых позвонков. Верхние губы
не образуют мясистых нависающих складок. Голый, хорошо развитый
козелок с округленной или заостренной вершиной. Уши в большинстве
случаев разделены между собой. В верхней челюсти у огромного
большинства видов по 2 пары резцов. Костная крышка барабанной ка-
меры с внутренней стороны не вырезана.........................
.......................Гладконосые. Vespertilionidae. (Стр. 88.)
4(3). Утолщенный хвост на половину своей длины выступает из
межбедренной перепонки. Верхние губы собраны в нависающие мяси-
стые складки. Маленький козелок имеет форму неправильного квад-
рата, вершинная часть которого усажена рядом густых и сравнительно
длинных волос. Широкие толстокожие уши на лбу сращены между со-
бой. В верхней челюсти одна пара резцов. Костная крышка барабан-
ной камеры с внутренней стороны несет глубокую вырезку (рис. 30).
.............................Бульдоговые. Molossidae. (Стр. 114.)
СЕМЕЙСТВО ПОДКОВОНОСЫЕ. FAMILIA RHINOLOPHIDAE
Около 100 ныне известных форм подковоносов населяют тропиче-
ские и субтропические области восточного полушария; все они объеди-
няются в единственный род—Rhinolophus. В некоторых систематиче-
ских сводках подковоносов объединяют с полковниками (Hipposideri-
dae). По многим признакам полковники действительно близки к подко-
воносам, но по ряду признаков (строению позвоночника, скелета передней
конечности) они так сильно отличаются, что без новой ревизии этих
групп на большом фактическом материале мы пока предпочитаем
придерживаться мнения Г. С. Миллера, что эти группы надо рассмат-
ривать в качестве разных семейств.
РОД ПОДКОВОНОСЫ. GENUS RHINOLOPHUS Lacepede, 1799
Зубная формула: I — С —Pm —-^-Pmp —М — -2 = 32, реже 30
21 2 (*) 13 или 28.
На лицевой части головы расположены почти голые кожистые
образования (рис. 15). В составе их различают: подкову, ланцет и
седло. Подкова — широкая пластинка, огибающая ноздри спереди и
с боков. Ланцет — продолжение подковы с заостренной вершиной.
Седло — небольшой гребень, расположенный между подковой и лан-
цетом. На седле имеются два выступа: нижний (обращенный к концу
морды) и верхний (обращенный ближе к лобной области головы).
Форма верхнего выступа седла имеет большое значение в характери-
стике видов подковоносов.
Ареал рода совпадает с ареалом семейства. В СССР встречается
6 видов, и ни один из них не выходит к северу за пределы Средней
Азии, Кавказа и .Южной Украины.
Светлое время суток подковоносы проводят в пещерах, под купо-
лами древних гробниц и мечетей, в необитаемых темных постройках,
реже на чердаках церквей и домов и в виде редчайших исключений
в больших дуплах деревьев. В одном убежище обычно держатся они
большими сообществами, не избегая тесного сожительства с другими
видами летучих мышей. Острыми, круто изогнутыми когтями ног они
искусно прицепляются к неровностям потолка или свода убежища,
где висят вниз головой на некотором расстоянии один от другого. Из-
82
за особенностей строения задних конечностей они не могут бегать по
твердому субстрату, поэтому они или висят, или перелетают, как бы
мало расстояние ни было. Дневной сон их чуток. Потревоженные
зверьки быстро поворачивают из стороны в сторону большие головы
с листовидными ушами, издают низкие, похожие на скрип звуки, пере-
пархивают с места на место, прячутся в малодоступные части пещеры
или вылетают наружу независимо от времени суток. Полет их шумный
с частыми взмахами крыльев. По своеобразной конфигурации широких
крыльев и остроконечным ушам они хорошо отличаются на лету от
других летучих мышей. Питаются преимущественно бабочками и дву-
крылыми, летающими поздно вечером и ночью. В условиях нашей
страны размножаются один раз в год. Самка приносит только одного
детеныша.
Рис. 14. Череп подковоноса Мегели (Rhinolophus
mehelyi Matschie). А — снизу; Б — сбоку.
Зубы I, ti—t2 —резцы; С, с — клыки; Рт, рт}, рт^ —ма-
лые предкоренные; Ртр, ртр — крупные подкоренные;
М1—М3, mi—/п3 — заднекоренные.
Первые дни своей жизни детеныш висит
на теле матери, крепко прицепившись зу-
бами к сосцевидным придаткам в паховой
области, самка при этом окутывает его
своими широкими крыльями. Лишь время
от времени детеныш переползает к груд-
ному соску и, напитавшись молоком, сно-
ва прикрепляется к паховым придаткам,
не связанным с молочными железами.
Подросших, но еще неспособных летать
детенышей самки оставляют на потолке
пещеры. Зимуют частично в тех же пеще-
рах, где жили летом, а часть совершает
дальние перелеты в подходящие для зи-
мовок пещеры, где впадает в спячку.
Из нескольких десятков видов под-
ковоносов в фауне СССР представлено
6 видов.
Рис. 15. Голова подковоноса
(Rhinolophus).
6*
83
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ПОДКОВОНОСОВ ФАУНЫ СССР
1(2). Размеры мелкие: кондилобазальная длина черепа 13—14 мм;
длина максиллярного ряда зубов 5,2—5,8 мм. Предплечье 34,5—42 мм.
Верхний выступ седла прямоугольный (рис. 16,Л). Верхний малый пред-
коренной зуб расположен в середине линии зубного ряда и имеет
острую коническую • вершину......................................
.............Малый подковонос. Rhinolophus hipposideros. (Стр. 85.)
2(1). Размеры крупнее: кондилобазальная длина черепа более
15 мм; длина максиллярного ряда зубов более 6,2 мм. Предплечье не
менее 43 мм. Верхний выступ седла округленный или заостренный.
Верхний малый предкоренной зуб с плоской или зерновидной вершиной
и обычно оттеснен к внешней стороне зубного ряда..............3
Рис. 16. Форма верхнего вы-
ступа седла (S) у разных ви-
дов подковоносов фауны СССР;
вид сбоку (левая сторона):
А — малый (Rhinolophus hipposide-
ros); В — бухарский (Rh. bochari-
cus); С — средиземный (Rh. blasii);
D — южный (Rh. euryale); E—F —
подковонос мегели (Rh. mehelyi).
3(8). Верхний выступ седла удлиненный и заостренный в своей
вершинной части (рис. 16, С — F). Между верхним клыком и крупным
предкоренным зубом есть хотя бы узенький промежуток, в котором
полностью или частично расположен малый предкоренной зуб . . 4
4(5). Верхний выступ седла прямой, сильно суженный и направ-
ленный вверх (почти параллельно ланцету, рис. 16, С). В нижней че-
люсти между первым малым предкоренным и большим предкоренным
зубами имеется промежуток, в котором расположен второй малый пред-
коренной зуб . Средиземный подковонос. Rhinolophus blasii. (Стр. 86).
5(4). Верхний выступ седла сравнительно широкий в средней ча-
сти, а вершина его в разной степени отклонена вперед (рис. 16, D—F).
В нижней челюсти первый малый предкоренной и крупный предкорен-
ной сомкнуты; второй малый предкоренной зуб оттеснен к внешней сто-
роне зубного ряда.............................................6
6(7). Размеры относительно мелкие: кондилобазальная длина че-
репа 15,4—16 мм; скуловая ширина 9,2—9,8 мм. Предплечье 43—45 мм.
...............Южный подковонос. Rhinolophus euryale. (Стр. 86.)
7(6). Размеры крупнее: кондилобазальная длина черепа 16,6—
17,5 мм; скуловая ширина 10—11 мм. Предплечье 50—55 мм . . . .
...............Подковонос Мегели. Rhinolophus mehelyi. (Стр. 87.)
8(3). Верхний выступ седла укороченный с широкой округленной
вершиной (рис. 16, В). Верхний клык и крупный предкоренной зуб
84
Рис. 17. Малые подковоносы (Rh. hipposideros)
в состоянии зимней спячки.
сомкнуты, а очень мелкий верхний малый. предкоренной оттеснен к
внешней стороне зубного ряда или отсутствует.....................9
9(10)г Размеры крупные: кондилобазальная длина черепа 18,3—
22 мм; длина верхнего (максиллярного) ряда зубов 8—9,5 мм. Пред-
плечье 53—63 мм........................... , .....................
. . , Большой подковонос. Rhinolophus ferrumequinum, (Стр. 87,)
10(9), Размеры мельче; кондилобазальная длина черепа 16—
17,2 мм; длина верхнего (максиллярного) ряда зубов 6,8—7,2 мм;
предплечье 49,5—52,5 мм , . .......................................
, . , , . Бухарский подковонос, Rhinolophus bocharicus, (Стр, 88,)
1, Малый подковонос, Rhinolophus hipposideros Bechstein, 1800
Табл, V, 1—3. Карта 17
Западная Европа к северу до Ирландии, Великобритании, Немец-
кого и Балтийского побережий, Северная Африка, Малая и Передняя
Азия, Северо-Западные Гималаи, В СССР — Молдавия, Южная Укра-
ина, Кавказ (от северных склонов Главного Кавказского хребта) и
Закавказье, республики Средней Азии (Южная Туркмения, в Узбе-
кистане— хребет Нуратау и Фергана, в Таджикистане— в долине
Зеравшана, окрестности Душанбе и Гиссара, в Киргизии — у г, Ош),
85
Летом живет в небольших пещерах, в темных редко посещаемых
постройках (в древних мечетях, заброшенных сараях, конюшнях) и
на чердаках домов, даже покрытых железом, где другие виды подко-
воносов никогда не поселяются. Больших (многосотенных) колоний
не образует, встречаясь в летних убежищах поодиночке, парами или
скоплениями до 2—3 десятков особей. В Закавказье (в Куринской низ-
менности) детеныши родились в первых числах июля. Вес детеныша
составлял более 40% веса своей матери. Даже в первые дни жизни
самки оставляют детенышей на время кормежки в убежище (не носят
с собой). Но. подросший, уже свободно и хорошо летающий детеныш
днем забирается на тело матери, зубами держится за ее паховые со-
сцевидные придатки, а мать завертывает его в свои широкие крылья,
как новорожденного. Время от времени детеныш перевертывается вниз
головой и, энергично шевеля челюстями, лезет в рот самки. В при-
роде самки, вероятно, подкармливают своих детенышей принесенной
добычей. Зимовок в нашей стране пока не найдено. В Западной Европе
малые подковоносы зимуют в пещерах. Тело находящегося в спячке
зверька завернуто в крылья (рис. 17).
Размеры и окраска меха подвержены не совсем отчетливо выраженной географи-
ческой изменчивости, общую схему которой можно выразить так: по мере движения
(в пределах ареала вида) с северо-запада на юго-восток идет посветление окраски, а
по мере подъема над уровнем моря — увеличение размеров. Описано несколько подви-
дов, из которых в пределах СССР можно различить 2:
a) Rh. h. hipposideros Bechstein, '1800. (Тип из Франции.) Окраска меха взрослых
с примесью серых тонов. Молдавия, Южная Украина (включая Крым), Западный Кавказ.
b) Rh. h. midas Andersen, 1905'. (Тип из Джаска, Южный Иран.) Окраска взрослых
светлая, снизу почти белая, сверху с примесью охристых и желтоватых тонов. Восточ-
ное Закавказье, Средняя Азия, Иран.
2. Средиземный подковонос. Rhinolophus blasii Peters, 1866
Табл. V, 4. Карта 17
Южная Европа, Северная Африка, Палестина, Малая Азия, Афга-
нистан. В СССР — Южное Закавказье (долина Аракса у г. Ордубата
и Нахичевани) и Южная Туркмения (Бахарденская пещера и окрест-
ности Ашхабада).
В 1937 г. в Бахарденской пещере было более 200 особей. Днем
они висели на потолках куполовидных сводов группами от 2—3 до
15—20 особей отдельно от других видов. Один взрослый самец прихо-
дился примерно на 15—20 самок. В 1962 г. в той же пещере держалось
около 100 особей этого рёдкого вида. Рождение молодняка и линька
происходят значительно позднее, чем у всех остальных летучих мы-
шей нашей фауны. Вечерний вылет недружный, а ночной перерыв кор-
межки неотчетливо выраженный. Кормятся низко, среди редкой травя-
нистой растительности, покрывающей пологие склоны гор.
Географическая изменчивость не изучена. Окраска зверьков из Бахарденской пе-
щеры и Палестины заметно светлее, чем у западноевропейских. Номинальная форма
(Rh. blasii Peters, 1866) описана по экземпляру из Италии; тип не выделен.
3. Южный подковонос. Rhinolophus euryale Blasius, 1853
Табл. V, 5. Карта 18
Юг Западной Европы, Северная Африка, Малая Азия. В СССР —
Черноморское побережье Кавказа (к югу от Сочи), Нагорный Карабах
(Шуша), Бахарденская пещера в Юго-Западной Туркмении.
В 1937 г. в Бахарденской пещере несколько десятков особей дер-
жались в осенние дни в среднем отделе пещеры поодиночке в середи-
86
нах плотных куч многочисленных там длиннокрылов. В 1956 г. во
всех доступных для осмотра местах Бахарденской пещеры зверьков
этого редкого вида уже не было обнаружено. В 1962 г. обнаружено
3 зверька. В пещере около Шуши (в Нагорном Карабахе) в 1939 г.
один южный подковонос был в середине скопления из 25—30 особей
подковоноса Мегели. В Агурской пещере у Сочи южные подковоносы
висели вперемежку с длиннокрылами. Число самцов и самок в любых
возрастных группах примерно одинаково. Рождение молодняка про-
исходит, как у большинства других видов, в начале лета, а линька —
в конце лета (значительно раньше, чем у средиземного подковоноса).
Мех южных подковоносов из пещер Черноморского побережья Кавказа окрашен
темнее, чем у зверьков из Бахарденской пещеры. По экземплярам из пещеры близ
с, Павловского бывшего Сухумского округа К. А. Сатунин (1911) описал Rh. е. nord-
manni. Из-за недостатка доброкачественного сравнительного материала из Италии
(Милана — места описания номинальной формы) для сравнения с западнозакавказ-
скими и экземпляров из Алжира, Марокко и Палестины (откуда описаны 3 подвида)
для сравнения с бахарденскими вопрос о характере географической изменчивости и
подвидовой систематике остается пока открытым. Номинальная форма (Rh. е-'euryale
Blasius, 1853) описана из Милана, Италия.
4. Подковонос Мегели. Rhinolophus mehelyi Matschie, 1901
Табл. V, 6. Карта 18
Северная Африка (Алжир, Марокко), Южная Европа (Португа-
лия, Испания, Франция, Корсика, Сардиния, Сицилия, Италия, Юго-
славия, Греция, Венгрия, Румыния, Болгария). В СССР — Закав-
казье: в Грузии — Мцхета (близ Тбилиси), в Армении — Алучалу Мар-
тунинского района, в Азербайджане — Шуша (в Нагорном Карабахе)
и Шемахинский район.
В Шушинской пещере летом 1939 г. была небольшая колония
самцов (около 25—30 особей), а в пещере около Шемахи в колонии
было примерно 700—800 зверьков.
Закавказские подковоносы Мегели по размерам несколько крупнее южноевро-
пепских. Номинальная форма описана по экземпляру из Бухареста, Румыния.
5. Большой подковонос. Rhinolophus ferrumequinum Schreber, 1774
Табл. V, 7—9. Карта 19
От Великобритании, Португалии и Северной Африки ареал про-
стирается через горы Центральной Европы, Балканы и Малую Азию
до Вьетнама и Японии включительно. В СССР — Закарпатская Укра-
ина, Крым, Кавказ, горы и предгорные районы Средней Азии.
Поселяется в пещерах и пещерообразных сооружениях, под купо-
лами древних мечетей и мавзолеев, на чердаках, крытых черепицей.
Взрослые самцы и холостые самки поселяются летом поодиночке или
по 2—3 особи в одном месте, а размножающиеся самки и молодняк об-
разуют колонии до 400—500 особей. В состав таких колоний часто вклю-
чаются зверьки других видов, особенно неразлучной спутницы, большого
подковоноса — трехцветной ночницы. После продолжительной вечерней
кормежки следует полуночный перерыв, а перед рассветом зверьки вновь
вылетают. Несмотря на крупные размеры, питаются относительно мел-
кими насекомыми (моли, двукрылые), если же крупными, то с не очень
массивным хитиновым панцирем. Зимовки найдены В. И. Абеленцевым
(1950) в Закарпатье и О. П. Богдановым в окрестностях Ташкента и
в долине Зеравшана. Часть подковоносов на зиму улетает за пределы
нашей страны (в частности, в Иран и Афганистан).
87
Географическая изменчивость имеет примерно такой же характер, как у малого
подковоноса. Описано несколько форм, из которых в фауне СССР можно разли-
чить 2:
a) Rh. f. ferrumequinum Schreber, 1774. (Тип из Франции.) Окраска меха относи-
тельно темная, у взрослых с бурым или коричневатым налетом. Закарпатье, Молдавия,
Крым и Западный Кавказ.
b) Rh. f. proximus Andersen, 1905. (Тип из Гильгита, Кашмир.) Окраска меха
светлая: снизу чисто белая, сверху у взрослых с бледным охристым или палевым на-
летом. Восточное Закавказье и Средняя Азия.
6. Бухарский подковонос. Rhinolophus bocharicus Kastschenko et
Akimov, 1917
Табл. V, 10. Карта 19
Средняя Азия: Юго-Восточная Туркмения (долины рек Мургаба
и Катана), долины рек Среднего и Южного Узбекистана, включая
долину Сурхандарьи от Термеза до ст. Шурчи, в Таджикистане.
В 1957 г. добыт V. Aellen (18.Х) в пещере Зармаст, близ г. Меймене,
в Афганистане. Вероятно, будет обнаружен и в других местах Север-
ного Афганистана, особенно в зимние месяцы.
Летом населяет темные и влажные лессовые пещеры. Обычно
живет колониями до 200—300, а осенью даже до 600 особей. Самцы
и самки держатся вместе. Одиночки встречаются редко. Часть зверь-
ков на зиму остается в тех же пещерах, где жили летом (например,
в окрестностях Ташкента), другие (в Кашано-Мургабской долине) на
зиму улетают в сторону Афганистана. На юге Туркмении рождение
молодняка происходит в конце июня и самом начале июля. Молодняк
в возрасте 30—45 дней поодиночке, без сопровождения родителей, уле-
тает на юг, на зимовку (такие перелеты наблюдались в Кашано-Мур-
габской долине).
Подвидов нет. (Тип из Термеза, Южный Узбекистан.) V. Aellen (1959) объединил
бухарского подковоноса с Rh. clivosus Cretzschmar, 1828. Мы считаем это объединение
совершенно необоснованным.
СЕМЕЙСТВО ГЛАДКОНОСЫЕ. FAMILIA VESPERT1LIONIDAE
Представители этого семейства населяют весь земной шар до по-
лярных границ древесной растительности. Три подсемейства: Тошоре-
atinae (один вид, Перу), Kerivoulinae (два рода с 10 видами, Африка
к югу от Сахары, Индокитай, Индия, Малайский архипелаг, Новая
Гвинея) и Vespertilioninae (около 22 родов, объединяющих более
150 форм, распространенных в пределах всего ареала семейства).
В фауне СССР к этому семейству относится наибольшее число видов,
а фауна почти всей Восточной Европы, всей Сибири и Дальнего Во-
стока представлена исключительно им. 32 вида обыкновенных летучих
мышей, распространенных в пределах Советского Союза, группиру-
ются в 8 родов.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ГЛАДКОНОСЫХ ЛЕТУЧИХ МЫШЕИ,
ПРЕДСТАВЛЕННЫХ В ФАУНЕ СССР
1(2). В верхней челюсти с каждой стороны по два малых предко-
ренных зуба (рис. 13), в нижней тоже по два. Из дополнительных при-
знаков можно указать следующие. Эпиблемы (рис. 12, Ер) у большин-
ства видов нет. Тонкокожие, разделенные между собой почти всей ши-
риной головы уши на половину и более своей длины выступают над
уровнем меха. Относительно длинный козелок всегда сужен и часто за-
88
острен в своей вершинной части. В окраске меха спинной стороны пре-
обладают темно-бурые тона с примесью коричневых или палевых (у пу-
стынных форм).........................Ночницы. Myotis. (Стр. 90.)
2(1). В верхней челюсти с каждой стороны не более чем по одному
малому предкоренному зубу. Сочетание внешних признаков иное . 3
3(6). В нижней челюсти с каждой стороны по 2 малых предкорен-
ных зуба. Вторая фаланга третьего пальца крыла почти втрое длиннее
первой фаланги того же пальца или длина уха едва уступает длине
предплечья ..................................................... 4
4(5). Вторая' фаланга третьего пальца крыла почти в три раза
длиннее первой фаланги этого же пальца. Уши очень короткие, едва
выступающие над уровнем меха. Скуловая ширина больше ширины
черепа в слуховой области . . Длиннокрылы. Miniopterus. (Стр. 97.)
5(4). Вторая фаланга третьего пальца крыла не более чем в пол-
тора раза длиннее первой фаланги этого же пальца. Уши огромные:
длина уха едва уступает длине предплечья. Скуловая ширина меньше
ширины черепа в слуховой области . . Ушаны. Plecotus. (Стр. 98.)
6(3). В нижней челюсти с каждой стороны по одному малому
предкоренному зубу. Вторая фаланга третьего пальца не более чем
в 1,5—2 раза длиннее первой фаланги того же пальца, а длина уха ко-
роче половины длины предплечья ................................. 7
7(8). Уши соединены между собой кожистой складкой, и внутрен-
ние края их почти соприкасаются друг с другом. Пространство между
глазами и межноздревым промежутком голое. Скуловая ширина уже
ширины черепа в слуховой области...................................
...............................Широкоушки. Barbastella. (Стр. 100.)
8(7). Уши разделены между собой почти всей шириной головы и
не имеют соединяющей их кожистой складки. Пространство между гла-
зами и межноздревым промежутком покрыто волосами. Скуловая ши-
рина превышает ширину черепа в слуховой области..................9
9(14). Верхняя сторона межбедренной перепонки голая или только
у основания покрыта редкими волосами. Ноздри открываются на почти
гладкой поверхности носа. Череп иной конфигурации, чем показано на
рисунке 28......................................................10
10(13). В верхней челюсти с каждой стороны по 2 резца. Длина
уха менее 27 лии; длина козелка не более 10 мм..................11
11(12). Третья метакарпальная кость (рис. 12, Mt) на 8—12 мм
длиннее пятой (Mt 5). Подмышечная область крыловой перепонки (рис. 12
£) густо покрыта волосами; свободный край этой перепонки прикреп-
ляется выше основания внешнего пальца ступни. В окраске меха пре-
обладают рыже-коричневые тона . . Вечерницы. Nyctalus. (Стр. 101)
12(11). Третья метакарпальная кость не более чем на 3—4 мм
длиннее пятой. Подмышечная область крыловой перепонки не имеет
густого волосяного покрова; свободный край этой перепонки прикреп-
ляется к основанию внешнего пальца ступни .........................
.......................Кожаны и Нетопыри. Vespertilio. (Стр. 103.)
13(10). В верхней челюсти с каждой стороны по одному (двухвер-
шинному) резцу. Длина уха более 30 мм; длина козелка не менее
14 мм...........................Стрелоухи. Otonycteris. (Стр. 113.)
14(9). Верхняя сторона межбедренной перепонки от основания до
самого свободного края густо покрыта волосами. Ноздри открываются
на концах особых кожистых трубок, сращенных у основания и направ-
ленных в стороны. Череп своеобразной конфигурации (рис. 28) . . .
...................................Трубконосы. Murina. (Стр. 113.)
89
РОД НОЧНИЦЫ. GENUS MYOTIS Каир, 1829
9 1 9 14
Зубная формула: I у С у Pm — Pmp — № — • 2 = 38.
Число малых предкоренных зубов в верхней челюсти (по 2 с каж-
дой стороны)—важнейший отличительный признак рода Myotis; про-
чие приведенные в таблице признаки в большей или меньшей степени
переплетаются с признаками других родов, представленных в нашей
фауне.
Многочисленные (около 100) формы Myotis (количество настоящих
видов точно не установлено) распространены почти во всем ареале
отряда. В фауне СССР представлено 11 видов.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ НОЧНИЦ, ПРЕДСТАВЛЕННЫХ
В ФАУНЕ СССР
1(2). Длина уха более т/2 длины предплечья (22—25,5 мм); пред-
плечье 42—45 мм; кондилобазальная длина черепа 16,5—17,1 мм; меж-
глазничный промежуток 4—4,4 мм. Крыловая перепонка прикрепляется
к основанию внешнего пальца......................................
...............Длинноухая ночница. Myotis bechsteini. (Стр. 92.)
2(1). Длина уха меньше V2 длины предплечья. Сочетание дополни-
тельных признаков иное ................. ......... 3
3(6). Размеры крупные: предплечье более 53 мм; кондилобазаль-
ная длина черепа не менее 19 мм................................4
4(5). Кондилобазальная длина черепа 19—22 мм; длина верхнего
ряда зубов 8,2—9,7 мм; предплечье 53,5—60,5 мм; ухо 21—25 мм . . .
......................Остроухая ночница. Myotis blythi (Стр. 92.)
5(4). Кондилобазальная длина черепа 22,2—23,6 мм; длина верх-
него ряда зубов 9,8—10,6 мм; предплечье 61—64,5 мм; ухо 26—28 мм
. •.....................Большая ночница. Myotis myotis. (Стр. 93.)
6(3). Размеры значительно мельче: предплечье не более 49 мм;
кондилобазальная длина черепа менее 17,5 мм..............•. . 7
7(8). Нёбных складок 8, и все они цельные. Свободный край кры-
ловой перепонки прикрепляется к голеностопному сочленению. Меж-
глазничный промежуток очень широкий: 4,9—5,7 мм..................
....................Прудовая ночница. Myotis dasycneme. (Стр. 93.)
Рис. 18. Место прикрепления свободного края крыловой
видов ночниц:
А — Myotis dasycneme; Б — М. daubentonii; В — М. capaccini;
перепонки у разных
Г — М. mystacinus.
90
8(7). Нёбных складок 7, причем 4, 5 и 6-я всегда разделены (прер-
ваны) посередине. Свободный край крыловой перепонки прикрепляется
к другим точкам ноги (у основания внешнего пальца, к середине плю-
сны или выше голеностопного сочленения). Межглазничный промежу-
ток не шире 4,7 мм.................................................9
9(12). Свободный край крыловой перепонки прикрепляется к сере-
дине плюсны или выше голеностопного сочленения....................10
10(11). Крыловая перепонка прикрепляется к середине или основ-
ной трети плюсны. Участок верхней стороны крыловой перепонки, при-
лежащий к бедру и голени, голый. Межглазничный промежуток черепа
шире расстояния между внешними сторонами верхних клыков .
. . . . Водяная ночница.
Myotis daubentoni. (Стр. 94.)
11(10). Крыловая перепон-
ка прикрепляется на 3—5 мм
выше голеностопного сочлене-
ния. На участке верхней сторо-
ны этой перепонки вдоль бедра
п голени до свободного края
тянется полоса темно окрашен-
ных, довольно густых волос.
Межглазничный промежуток
черепа равен или уже расстоя-
ния между внешними сторона-
ми верхних клыков . . . .
. . . Длиннопалая ночница.
Myotis capaccinii. (Стр. 94.)
12(9). Свободный край
крыловой перепонки прикреп-
ляется к основанию внешнего
пальца ноги или в непосредст-
венной близости от него . 13
13(14). Свободный край
межбедренной перепонки меж-
Рис. 19. Свободный край межбедренной пере-
понки:
А — ночницы Наттереца (Myotis nattereri) — уса-
жен ресничками; Б — трехцветной ночницы (М.
emarginatus).
ду концами шпор и хвостом
резко утолщен, зазубрен и густо усажен жесткими ресничками, располо-
женными по самому краю в два параллельных ряда (рис. 19,Л). . .
....................Ночница Неттерера. Myotis nattereri. (Стр. 94.)
14(13). Свободный край межбедренной перепонки между концами
шпор и хвостом не имеет ни утолщения, ни зазубренности, ни ресничек,
которые резко отличались бы от редких и тонких волос, покрывающих
перепонку (рис. 19,5)..........................................15
15(16). Размеры средние: кондилобазальная длина черепа 14,8—
16 мм. Волосы, покрывающие верхнюю сторону тела, трехцветные . .
...............Трехцветная ночница. Myotis emarginatus. (Стр. 95.)
16(15). Размеры мельче: кондилобазальная длина черепа не
превышает 14,3 мм. Волосы, покрывающие верхнюю сторону тела,
двухцветные....................................................17
17(20). Предплечье не более 39 мм; длина голени не более 15—
18 мм..........................................................18
18(19). Размеры очень мелкие: общая длина черепа 13,2—15 мм;
кондилобазальная длина 12,2—12,4 мм. Вдоль основания шпоры тя-
нется ясно заметный эпиблемообразный выступ (рис. 20, Б)............
.......................Малая ночница. Myotis muricola. (Стр. 95.)
91
Рис. 20. Шпоры ночниц:
А — усатой (Myotis mystacinus); Б — ма-
лой (М. muricola ikonnikovi); В — длинно-
хвостой М. frater bucharensis; Г — М. fra-
ter longicaudatus.
19(18). Размеры крупнее: общая длина черепа 13,2—15 мм; конди-
лобазальная длина 12,4—14,3 мм; скуловая ширина 8,0—9,2 мм; пред-
плечье 32—39 мм; ухо 13—16 мм. У основания шпоры эпиблемообраз-
ного выступа нет (рис. 20, Л).......................................
.......................Усатая ночница. Myotis mystacinus. (Стр. 96.)
20(17). Длина предплечья 39—41 мм, длина голени около 20 мм.
Дополнительно: хвост длиннее тела. Первый малый предкоренной зуб
верхней челюсти вплотную примыкает к большому предкоренному зубу;
второй малый предкоренной оттес-
нен к внутренней стороне зубного
ряда и при рассматривании черепа
в профиль не виден. Вдоль основа-
ния шпоры тянется различно раз-
витая эпиблема (рис. 20, В, Г) . .
. . . . Длиннохвостая ночница.
Myotis frater. (Стр. 97.)
7. Длинноухая ночница. Myotis
bechsteini Kuhl, 1818
Табл. VI, 1. Карта 20
Западная Европа к северу до
Южной Англии, Норвегии и Сред-
ней Швеции, Литва, Ольгополь на
Украине, советские Карпаты, гора
Ачишхо в Кавказском заповеднике,
Ставрополь и с. Зугдиди, в Запад-
ном Закавказье.
Летом одиночных особей нахо-
дили в колониях других видов дуп-
логнездников (водяной ночницы и
ночницы Наттерера), реже встре-
чаются небольшие колонии в дуп-
лах деревьев, состоящие из особей только этого вида. Зимой нахо-
дили в пещерах, в подземных ходах и погребах (Закарпатская Украина,
Новая Александрия бывшей Люблинской губернии в Польше).
Географическая изменчивость не выражена. Номинальная форма описана по
экземпляру из Германии (Гессен, Генау).
8. Остроухая ночница. Myotis blythi Tomes, 1857
Табл. VI, 2. Карта 21
Южная Европа, Северная Африка, Палестина, Малая Азия, Иран,
Центральная Азия, Индия. В СССР населяет Молдавию, Крым, весь
Кавказ, Туркмению, Туранскую низменность, Западный Тянь-Шань,
Гиссаро-Алтай и Алтай.
Поселяется в больших пещерах, под куполами древних гробниц,
в мечетях, на чердаках больших домов и церквей, образуя колонии до
нескольких сот особей. На лето колонии не разделяются. Одиночки
встречаются сравнительно редко. Самцы обыкновенно круглый год жи-
вут вместе с самками. На вечернюю кормежку вылетают поздно, и
взрослые зверьки кормятся всю ночь без перерыва, а молодые, недавно
92
перешедшие к самостоятельной жизни, после вечерней кормежки отды-
хают в убежищах, вновь вылетая лишь перед рассветом. Поедает остро-
ухая ночница крупных жуков и ночных бабочек.
Географическая изменчивость (размеров и окраски) выражена очень слабо, и
большинство описанных форм практически различить невозможно. М. b. blythi Tomes,
1857, описана из Западной Индии (Насирабад, Раджастхан); ЛА. b. oxygnathus Monti-
celli, 1855 (из Италии, Матера, Базилиата); и ЛА. b. omari Thomas, 1906 (из Ирана,
Дербент, близ Исфахана), видимо, синонимы номинальной.
9. Большая ночница. Myotis myotis Borkhausen, 1797
Табл. VI, 3. Карта 21
Северная Африка (на восток до Абиссинии), Южная и Западная
Европа к северу до Великобритании, Южной Швеции и Средней Поль-
ши, а к востоку до советских Карпат включительно.
На зимовках образует скопления до 5000 и более особей в одной
пещере, а весной откочевывает в разных направлениях от места зи-
мовки на расстояния до 260 км. Самки летом образуют небольшие обо-
собленные колонии на чердаках домов, за обшивками стен и карни-
зов, а самцы в подобных же убежищах держатся отдельно от них,
большей частью поодиночке. В остальных чертах образ жизни сходен
с предыдущим видом. Подобно всем ночницам (кроме усатой), самки
родят по одному детенышу, эмбриональное развитие которого проходит
в правом роге матки.
В СССР, как и в других странах Европы, номинальная форма: М. m. myotis
Borkhausen, 1797. (Описана из Тюрингии, Германия.)
10. Прудовая ночница. Myotis dasycneme Boie, 1825
Табл. VI, 4—5. Карта 20
Своеобразный ареал прудовой ночницы в виде узкой полосы, за-
ключенной между 49—63° с. ш., тянется ог Северной Франции, Бельгии
и Голландии через всю Европейскую часть СССР и Западную Сибирь
до Енисея.
Колонии размножающихся самок прудовой ночницы найдены
только в четырех пунктах: на одном из островов оз. Селигер, в Вольске,
Орске и в Воронежском заповеднике. Скопления их, достигающие не-
скольких сотен особей, помещаются под куполами деревянных церквей
или над карнизами крытых железом построек. Самцы и неразмножаю-
щиеся (холостые) самки поодиночке или небольшими скоплениями дер-
жатся чаще всего в дуплах деревьев особняком или в колониях других
видов (двухцветного кожана, лесного нетопыря). Любое из населенных
этим видом убежищ всегда расположено близ стоячего или с тихим те-
чением водоема и непременно имеющего участок открытого зеркала.
Кормится прудовая ночница исключительно над поверхностью таких
водоемов и в других местах, подолгу никогда не летает. На вечернюю
кормежку вылетает относительно поздно и вскоре возвращается в днев-
ное убежище, а с наступлением рассвета начинается второй, еще более
дружный и кратковременный вылет. Несмотря на крупные размеры,
поедает, вероятно, множество мелких насекомых, особенно комаров,
летающих в приводных слоях воздуха. Зимовки обнаружены в пещерах
Урала; одиночки найдены на зимовке в пещерах Эстонии.
93
Известно 5 основных типов окраски, почти не соединенных между собой перехо-
дами и необособленных или неотчетливо разграниченных географически. М. dasycneme
описана из Виборга (Дания). М. d. major Ognev et Worobiev, 1923. (Тип из с. Хреновое
Бобровского района Воронежской области) — ее синоним.
11. Водяная ночница. Myotis daubentoni Kuhl, 1819
Табл. VI, 6. Карта 22
От Шотландии, Великобритании и Франции на восток до Камчатки,
Сахалина. Северная граница ареала идет близ 60° с. ш., а южная тя-
няется от Северной Италии через Румынию, Южную Украину, Нижнюю
Волгу, Северный Казахстан, Алтай, Северную Монголию до Южного
Приморья и Южного Сахалина.
Летом живет небольшими колониями (по нескольку десятков осо-
бей) в дуплах деревьев или на чердаках, преимущественно деревянных
построек. Самцы и самки держатся вместе. В остальном сходна с пру-
довой ночницей, хотя связь с приводными слоями воздуха выражена
у нее не так строго, и кормящихся водяных ночниц можно видеть лета-
ющими среди деревьев в лесах, парках, даже на озелененных улицах
поселков и городов.
М. d. daubentoni Kuhl, 1819, описана из Германии (Генау, Гессен). Восточную
Европу, Западную и Среднюю Сибирь населяют зверьки, несколько отличающиеся от
западноевропейских и восточносибирских относительно купными размерами. Кроме
того, по мере движения с запада на восток наблюдается некоторое изменение окраски
(уменьшение примеси палевых и коричневато-бурых тонов), но различия эти так
незначительны и географически так слабо отграничены, что описанные ранее формы
практически различить во многих случаях не удается.
12. Длиннопалая ночница. Myotis capaccinii Bonaparte, 1837
Табл. VI, 7. Карта 22
Средиземноморье, Нукус (Кара-Калпакия, низовье Аму-Дарьи), до-
лина рек Сучан и Шмаковка (Южное Приморье), Южные Курилы и
Япония.
Летом 1928 г. колония в несколько десятков особей была найдена
в небольшой каменной пещере (в долине р. Сучана). Летнее поселение
в пещерах отличает ее от большинства видов этого рода.
а) М. с. capaccinii Bonaparte, 1837. (Описан с о-ва Сицилия.) Второй нижний
предкоренной зуб равен или едва уступает в размерах первому. Волосы на верхней
стороне межбедренной перепонки заходят до коленных суставов и первой трети дли-
ны хвоста. Окраска верха относительно светлая палево-дымчато-бурая. Западная
часть ареала, в СССР — Нукус, в низовьях Аму-Дарьи.
b) М. с. macrodactylus Temminck, 1840. (Описан по экземпляру из Японии.)
Второй нижний предкоренной зуб в 1,5—2 раза меньше смежного с ним рпц. Волосы
на верхней стороне межбедренной перепонки заходят до средней части голеней и поло-
вины длины хвоста. Окраска верха темная, дымчато-бурая. Эллерман и Моррисон-Скот
(1951) считают эту форму особым видом. По нашему мнению, такие различия для
разных видов ночниц недостаточны. В СССР — Южные Курилы и Южное Приморье
(Перетинская пещера на р. Сучан и окрестности пос. Шмаковка); Япония.
13. Ночница Наттерера. Myotis nattereri Kuhl, 1818
Табл. VI, 8. Карта 23
Западная Европа, от Средиземного моря на север до Ирландии,
Шотландии и Южной Швеции, Лебяжье (близ Петергофа), Саблино
(в 40 км к югу от Ленинграда), г. Киров, Московская область: Подоль-
ский и Серпуховский районы, окрестности Симферополя, Дагестан, мно-
гие пункты Закавказья, Чули (в Северо-Западном Копет-Даге), Тункин-
94
ская крепость на р. Иркут (Восточный Саян), ст. Тунгир (в Южной
Якутии), долина Амура (без более точного обозначения), Владиво-
сток, о-ва Японии.
Данных об образе жизни нет. В Северном Азербайджане (Кусары)
одну колонию из 9 зверьков (6 самок и 3 самца) мы нашли в дупле
грецкого ореха. Зимует в пещерах, не образуя больших скоплений.
Описаны следующие подвиды:
а) М. n. nattereri Kuhl, 1818. (Описан по экземпляру из Генау, Гессен, Германия.)
Размеры относительно мелкие: предплечье 36,5—40,5 мм; максиллярный ряд зубов
5,8—6,3 мм; козелок 10—И мм, прямой или почти прямой. Вырезка на внешнем крае
уха едва заметная. Окраска верха темная, коричневато-бурая. Европа, Кавказ.
Ь) ДА. п. araxenus Dahl, 1947. (Тип из окрестностей с. Амгу, Вайкский хребет, Ар-
мения.) Размеры крупные: предплечье 42,3—47,8 мм; максиллярный ряд зубов 6,4—7,3 мм;
козелок 11,8—12,5 мм, вершина его слабо отклонена наружу. Вырезка на внешнем
крае уха слабая. Окраска верха светлая, серо-палевая. Долина р. Аракса, Армения.
с) М. п. tschuliensis Kuzyakin, 1935. (Тип из Чули, Копет-Даг.) Размеры средние:
предплечье 41,7—42 мм; максиллярный ряд зубов 6,3—6,6 мм; козелок 11—12,2 мм,
саблеобразно изогнутый. Вырезка на внешнем крае уха отчетливая. Окраска меха
очень бледная, серовато-палевая. Копет-Даг (Юго-Западная Туркмения):
d) М. n. amurensis Ognev, 1927. (Тип из г. Владивостока.) Размеры и форма ко-
зелка, как у европейской формы. Вырезка на внешнем крае уха хорошо выражена.
Окраска темная. Восточная Сибирь, Амуро-Уссурийский край.
е) М. n. bombinus Thomas, 1905. (Тип из Тако, Миясаки, Кен, о. Киушиу, Япония.)
Размеры, как у номинальной формы, но ухо больше, козелок уже и резко отклонен
к внешнему краю. Череп уплощен, верхняя линия профиля слабо изогнута в лобном
отделе. Окраска светлая. Япония.
Накопляющийся коллекционный материал показывает, что индивидуальная из-
менчивость у этого вида обширна и часть указанных признаков, может быть, утратит
свою диагностическую ценность.
14. Трехцветная ночница. Myotis emarginatus Geoffroy, 1806
Табл. VI, 9. Карта 23
Западная Европа (к северу до Голландии), Северная Африка,
Иран. В СССР — Закарпатская область, Крым, Закавказье, Южная
Туркмения, Узбекистан и Таджикистан.
Летом живет в пещерах и под куполами древних мавзолеев, церк-
вей и мечетей. В пределах СССР она во всех случаях находилась в ко-
лониях самок большого подковоноса, с которым эта ночница, оче^
видно, тесно биологически связана.
Географическая изменчивость выражена слабо. В фауне СССР 3 слабо разли-
чающиеся формы:
а) М. е. emarginatus Geoffroy, 1806. (Арденны, Северная Франция.) Размеры
относительно мелкие: предплечье 39—41 мм; кондилобазальная длина черепа 14,8—
15,2 мм; максиллярный ряд зубов 6,2—6,4 мм. Вершинки волос на спине относительно
темные, бурые или красновато-бурые. Западная Европа, в СССР — Закарпатье, Юж-
ный Крым и Среднее Закавказье (среднее течение р. Куры).
b) М. е. desertorum Dobson, 1875. (Джальк, Белуджистан, Южный Иран).
По размерам близка к Европейской, но окраска меха светлее и ярче- вершинки волос
на спине кирпично-красные. Южная Туркмения и пустынные районы Ирана.
с) М. е. saturatus Kuzyakin, 1934. (Тип из г. Ташкента, Узбекистан.) Окраска
близка к европейской форме, но размеры крупнее: предплечье 41,5—43,5 мм; кондило-
базальная длина черепа 15,2—16 мм; максиллярный ряд зубов 6,4—6,8 мм. Узбекистан
и Таджикистан.
15. Малая ночница. Myotis muricola Gray, 1846
Табл. VI, 10. Карта 24
От Южного Сахалина, Южной Якутии, окрестностей Красноярска,
Алтая (оз. Телецкое, Кебезень, Катон-Карагай) и Закарпатской обла-
сти на юг до Бирмы и Гималаев (Непал).
95
Обитатель горных лесов. По словам А. С. Фетисова, поселяется
в глубоких трещинах скал; из-за слабого полета избегает открытых
(ветреных) мест. Других данных об образе жизни нет.
Одиночных зимующих малых ночниц Н. Оводов находил в пеще-
рах около г. Красноярска.
В СССР — М. m. ikonnikovi Ognev, 1912. (Тип из долины р. Иман, приток Уссури.)
Окраска меха коричнево-бурая. Северная часть ареала.
16. Усатая ночница. Myotis mystacinus Kuhl, 1819
Табл. VI, 11—12. Карта 24
От Великобритании, Швейцарии и Южной Швеции на восток до
Камчатки и Сахалина. Северная граница ареала в Восточной Европе
и Западной Сибири достигает 62—63° с, ш., а далее на восток идет
несколько ниже 60°. .Южная граница от Швейцарии тянется через
Румынию, Черное море, Иран и Афганистан, далее огибает с юга всю
Гималайскую горную цепь и от нее — к южной части Приморья.
Поселяется в самых разнообразных убежищах: на чердаках до-
мов, за обшивками стен, наличниками окон, в дуплах деревьев, за
отставшей корой, в поленницах дров, в норах обрывистых берегов,
в щелях скал, погребах и пещерах. Больших колоний не образует,
встречаясь, как правило, поодиночке или группами не более 10—15
особей. Вечером вылетает поздно и кормится, видимо, всю ночь без
заметного перерыва. Р. Н. Мекленбурцев (1930) находил самок усатых
ночниц с двумя эмбрионами. Для других видов этого рода подобные
случаи неизвестны.
Географическая изменчивость усатой ночницы, осложнена широкой популяционной
и индивидуальной изменчивостью. Описано большое число подвидов, часть которых до
подбора лучших коллекционных материалов целесообразно сохранить в синонимике.
В СССР можно различить:
а) М. m. mystacinus Kuhl, 1819. (Германия.) Окраска- меха сильно варьирует, но
бледные желтоватые и песчано-палевые тона встречаются у небольшой части особей.
Череп удлиненный (особенно в носовом отделе); ширина мозговой капсулы на 1,2—
1,5 мм больше высоты черепа. Малые предкоренные зубы хорошо развиты и распола-
гаются вдоль средней линии зубного ряда. Penis крупный, расширенный
в предконцевой части; длина его 4—5 мм; диаметр 1,5—2,5 мм. Синонимы этой формы
М. ш. gracilis Ognev, 1927. (Тип из Николаевска-на-Амуре); М. m. brandti Eversmann,
1845. (Тип с р. Сакмары, в предгорьях Урала). Зверьки из Восточной Европы, Запад-
ной Сибири и с Камчатки немного крупнее, чем западноевропейские и амуро-уссурий-
ские, но различия можно выразить лишь в средних показателях. Вся европейско-си-
бирская часть ареала вида.
b) М. ш. przewalskii Bobrinskoy, 1926. (Тип из ущелья р. Мольджа, на северном
склоне хребта Русского, Хотан-Таг.) Окраска меха очень светлая: общий тон верха от
палево-желтоватой до буланой и песчано-палевой; низ белый. Череп укороченный
с округлой, сильно вздутой мозговой капсулой; ширина последней на 0,5—0,8 мм боль-
ше высоты. Малые предкоренные сильно уменьшены; Pm1 и Pmp часто сомкнуты;
очень мелкий Pm2 вытеснен к внутренней стороне зубного ряда. Penis маленький;
тонкий на всем протяжении; длина его 3—4 мм; диаметр около 1—1,5 мм. М. m. аи-
rascens Kuzyakim, 1935 (тип из с. Куркужин, близ Владикавказа), по строению зубов,
черепа и окраске занимает промежуточное положение между европейско-сибирской и
центральноазиатской формами. Пустыни Средней и Центральной Азин, Кавказ и За-
кавказье.
с) М. m. pamirensis Kuzyakin, 1935. (Тип из окрестностей оз. Яшилькуль, Восточ-
ный Памир.) Окраска меха взрослых относительно светлая, палево-желтоватая сверху
и беловатая на нижней стороне. Череп по форме, как у европейских, а зубная си-
стема, как у центральноазиатской формы. Недостаточно ясны отличия от гималайских:
М. m. caliginosus Tomes, 1859 (описана по экземпляру из Симла); М. т. nipalensis
Dobson, 1871 (тип из Катманду, Непал); М. т. meinertzhageni Thomas, 1926 (тип из
долины р. Шайок, у впадения в нее р. Нубра, Ладакх, Кашмир, Гималаи). Памир.
d) М. т. sogdianus Kuzyakin, 1934. (тип из г. Ташкента, Узбекистан.) Окраска
сверху относительно темная, свинцово-бурая. Размеры очень мелкие, близкие к М.
96
muricola. Место описания типа (Ташкент) выбрано неудачно, так как там такие ноч-
ницы встречены лишь осенью, а местные (ташкентские) близки к М. m. przewalskii.
Гиссаро-Алай, Западный Тянь-Шань.
17. Длиннохвостая ночница. Myotis frater G. Allen, 1923
Табл. VI, /5. Карта 23
Юго-Восточная Азия, Корея, Владивосток, Иманский район (При-
морского края), дер. Додонова Красноярского района; с. Кебезень,
на Алтае; Айвадж, в Юго-Западном Таджикистане, окрестности Са-
марканда.
Длиннохвостая ночница в пределах ее огромного ареала почти
всюду крайне редка. До 1957 г. в СССР было добыто лишь 14 особей.
8/VII—1957 г. О. П. Богдановым впервые было найдено большое скоп-
ление этих ночниц (около 500 особей) в подземных лабиринтах в ок-
рестностях Самарканда. Зверьки висели там плотной массой в узкой
(4 см) и длинной (150 см) щели потолка в 60 м от входа в пещеру.
30/VII—1957 г. колония оставалась там же. На другой год (1/VII—
1958 г.) в глубине той же пещеры было не менее 100 длиннохвостых
ночниц. Среди 100 зверьков, осмотренных 8/VII—1957 г., были только
взрослые самки с детенышами разных размеров; часть детенышей уже
почти достигла размеров взрослых, другие были меньше почти вдвое.
Это свидетельствовало о растянутости периода размножения. Из 27 мо-
лодых было 15 самок и 12 самцов. На вечернюю охоту (8/VII) зверьки
вылетели поздно, с наступлением густых сумерек, и, не задерживаясь
у пещеры, направились в пойму Зеравшана.
28/IX—1963 г. мы осмотрели группу искусственных пещер в окрест-
ностях Самарканда и в одной из них нашли скопление (около 100 осо-
бей) длиннохвостых ночниц. Они висели на неровности потолка плот-
ной кучей. Среди 28 взятых зверьков преобладали самцы. Все зверьки
(особенно самки) были очень жирными.
М. f. frater Gl. Allen описана из Юго-Восточной Азии. Морфологически она очень
близка (судя по описаниям) к западноамериканской М. volans Н. Allen, 1866.
В фауне СССР 2 формы:
а) М. f. longicaudatus Ognev, 1927. (Тип из г. Владивостока.) Размеры сравни-
тельно с номинальной формой немного крупнее: предплечье 39—41 мм; ухо 12 мм;
максиллярный ряд зубов 5,2—5,3 мм. Окраска верха темная, коричнево-бурая, низ
серо-белесый. Южное Приморье, вероятно, Корея, Красноярский район и с. Кебезень,
на Алтае.
b) М. f. bucharensis Kuzyakin, 1950. (Тип из Айваджа на Аму-Дарье, Юго-Запад-
ный Таджикистан.) Размеры крупнее: предплечье 40—43 мм; ухо 13—14,5 мм; мак-
силлярный ряд зубов 5,3—5,7 мм. Окраска меха светлая: сверху палево-буроватая;
низ беловатый. Таджикистан и Узбекистан.
РОД ДЛИННОКРЫЛЫ. GENUS MINIOPTERUS Bonaparte, 1837
Зубная формула: I — С-- PmTpmp-J-M — -2=36.
3 1 2 13
По многим признакам (зубная система, органы размножения
и др.) длиннокрылы близки к роду ночниц (Myotis), но, в отличие от
них, да и от большинства обыкновенных летучих мышей, они достигли
высокого совершенства в летательном аппарате. Это выражено в рез-
ком сужении концевого отдела крыльев за счет удлинения второй фа-
ланги третьего пальца, в наличии мощно развитых шпор, произвольно
отгибающихся внутрь межбедренной области и образующих (при на-
добности) из межбедренной перепонки мешок, выполняющий роль тор-
моза, и т. д.
7 Н. А. Бобринский и др.
97
Распространены длиннокрылы в тропических и субтропических
областях восточного полушария. Описано более 10 видов, из них в фа-
уне СССР один вид.
18. Длиннокрыл обыкновенный. Miniopterus schreibersi Kuhl, 1819
Табл. VI, 14. Карта 25
Огромный ареал вида тянется от Западной Африки и Пиреней-
ского полуострова до Японии, Филиппин и Северо-Западной Австра-
лии. В СССР представлен несколькими колониями, находящимися
в пещерах Закарпатской области, Южного Крыма, на Черноморском
побережье Кавказа, в долине Аракса (около Ордубата), в Нагорном
Карабахе (Шуша), Бахарденской пещере (в Северо-Западном Копет-
Даге); остатки когда-то большой колонии сохранились в Мцхетском
соборе (Грузия).
Полет длиннокрыла легкий, стремительный. С высоким мастер-
ством полета, связано образование огромных колоний: в Бахарденской
пещере, например, длиннокрылов жило около 40 000. На вечернюю кор-
межку более 3/4 этой колонии вылетало минут за 30. В момент наибо-
лее напряженного вылета через небольшое отверстие, соединяющее
пещеру с внешним миром, каждую секунду вылетало наружу более
20 зверьков; за 10 минут такого вылета в сплошном потоке вылетало
более 12 000 животных. Часа через два после начала кормежки начи-
налось возвращение, которое затягивалось до самого рассвета. Полу-
ночного перерыва и повторного (утреннего) вылета не бывает. Более
всего длиннокрыл поедает мелких чешуекрылых. В июле самки родят
по одному детенышу. Осенью длиннокрылы покидают свои летние убе-
жища, совершая миграции, вероятно, в тропические области (в Индию,
Африку), где они не только ведут активную жизнь (не впадают в спяч-
ку), но, может быть, даже приносят второе потомство. В СССР зимую-
щие найдены в пещере у г. Берегово Закарпатской области УССР и
в марте 1962 г. обнаружены в Бахарденской пещере (X. Бабаев).
Из разных мест обширного ареала описано несколько подвидов, отличающихся
только по размерам и окраске. В СССР встречаются 2 из них:
а) М. sch. schreibersi Kuhl. (Южный Баннат, Венгрия.) Относительно темно
окрашенный без примеси палевых тонов. Закарпатье, Крым, Черноморское побережье
Кавказа.
b) М. sch. pallidus Thomas, 1907. (Тип с южного берега Каспийского моря, Север-
ный Иран.) Окраска меха бледная, «пустынная», с преобладанием палевых тонов. Во-
сточное Закавказье, Иран, Туркмения (западная часть Копет-Дага).
РОД УШАНЫ. GENUS PLECOTUS Geoffroy, 1818
Зубная формула: I — С — Pm — Ртр —М — -2 = 36.
3 1 2 13
К этому роду относится один палеарктический вид — Plecotus auri-
tus L. и несколько видов в Северной Америке.
19. Ушан. Plecotus auritus Linnaeus, 1758
Табл. VII, 1—2. Карта 26
Палеарктика от Канарских островов и Португалии до Камчатки,
Сахалина и Японии; от севера Сахары, Палестины, Ирана, западных
и южных склонов Гималайских гор до Скандинавии, Финляндии и
60—62° с. ш. в пределах СССР.
98
Летом ушан поселяется в самых разнообразных убежищах: на чер-
даках домов, за обшивками стен, наличниками окон, в дуплах де-
ревьев, в дуплянках и скворечниках, за отставшими кусками коры,
в разного типа пещерах и т. д. Больших колоний не образует. Дальних
сезонных перекочевок не совершает. Зимует в естественных пещерах
и искусственных подземных сооружениях (погреба, подвалы). Осенью
часто залетает через открытые двери и форточки в жилые помещения.
Во время зимней спячки принимает своеобразную позу: огромные уши
он закладывает под крылья, а основаниями торчащих козелков при-
крывает слуховые отверстия. На кормежку вылетает поздно и кор-
мится почти всю ночь без длительных перерывов. Чаще других лету-
чих мышей ловит ползающих и спокойно сидящих насекомых и пауков.
Если они сидят на листьях или на концах тонких веток, то ушан,
подлетев, держится в воздухе некоторое время в одной точке
(подобно пустельге или зимородку). Самки родят по одному дете-
нышу, реже по два.
В фауне СССР представлены 4 географические формы:
а) Р. a. auritus Linnaeus, 1758. (Описан из Швеции.) Относительно мелкий:
предплечье 37—42,5 мм; кондилобазальная длина черепа 14,8—15,8 мм (редко до
16,2 мм); наибольший диаметр слуховых барабанов (bullae tympani) обычно в пре-
делах 3,8—4,2 мм (редко до 4,5 мм); максиллярный ряд зубов 5,2—5,8 мм. Окраска
меха темная: вершины волос на нижней стороне тела палево-серые. Европейско-си-
бирская часть ареала к востоку до Охотского и Японского морей; Кавказ (исключая
высокогорья).
b) Р. a. sacrimontis G. Allen, 1908. (Тип из Fuyi, Япония.) Размеры немного круп-
нее: кондилобазальная длина черепа 16—16,4 мм; наибольший диаметр слуховых ба-
рабанов 4—4,3 мм. Окраска меха темная (как у европейских или еще темнее).
Япония. В СССР — Шантарские острова и, вероятно, Сахалин,
с) Plecotus auritus macrobullaris Kuzyakin subsp. nova. Тип: взрослый самец,
шкурка и череп, IX, 1914 г., окрестности г. Орджоникидзе, Северо-Осетинская АССР,
Кавказ, Л. Б. Беме, № 1435, коллекция Зоологического института АН СССР. Котип:
сГ ad из того же места спиртовая коллекция Зоологического музея МГУ. Размеры, как
у номинальной формы: предплечье 41—42,5 мм; кондилобазальная длина черепа 15,6—
16,0 мм; максиллярный ряд зубов 5,6—5,8 мм. Слуховые барабаны (bullae tympani)
очень крупные: 4,5—4,9 мм. Окраска верха темная, как у европейской формы, но
срединные пояски слабее выражены. Вершины волос нижней стороны тела белые,
а основания всех волос черные или темно-бурые. Мы неоднократно (1935, 1950) от-
мечали своеобразие горно-кавказских ушанов, но не решались им дать особое под-
видовое название, потому что близкая (а может быть идентичная) форма встреча-
ется в высокогорьях Альп и Карпат, где они, вероятно, были описаны под другими
названиями, например, Р. a. austriacus Fischer, 1829 (Vienna, Austria), P. kirschbaumi
Koch, 1860 (Dillenburg, Oberhessen, Германия), P. auritus var. montanus Koch, 1863
(Westerwald, Nassau, Germany) или P. a. meridionalis Martino, 1940 (Словения, Юго-
славия). Типов большинства этих форм не сохранилось. Сравнить с ними не было
возможности. Остается неясным: которые из них синонимы обыкновенного ушана и ко-
торые близки к горнокавказским. К последним близки также (отличаются лишь более-
светлой окраской меха) ушаны из Южного Казахстана (Копальский район) и Узбеки-
стана (Самарканд). Ареал новой формы: высокогорья Главного Кавказского хребта,
в пределах Северной Осетии й Кабардино-Балкарии, и у оз. Севан в Армении. Морфо-
логически близкие формы — в Карпатах, Альпах и предгорных районах Южного Казах-
стана и Узбекистана. Форма эта интересна в том отношении, что занимает промежу-
точное положение между номинальной Р. a. auritus L., североафриканской Р. a. christiei
Gray, 1838, центральноазиатским Р. a. wardi Thomas и Р. a. ariel Thomas, 1911. Морфо-
логические различия между этими формами так сложно переплетаются, что разделить
их на два или несколько видов нет оснований.
d) Р. a. wardi Thomas, 1911. (Leh, Ладак, Кашмир.) Размеры относительно круп-
ные: предплечье 43—46,5 мм; кондилобазальная длина черепа 16—18,5 мм; максилляр-
ный ряд зубов 5,4—6,5 мм. Слуховые барабаны крупные: их наибольший диаметр
4,2—5,0 мм- Окраска меха очень светлая: сверху желтовато-палевая, снизу белая. Сино-
нимы этой формы: Р. a. kozlovi Bobrinskoy, 1929 (из Восточного Цайдама), и Р. mordax
Thomas, 1926 (из Кашгарии.) Горные пустыни Кашмира (Северная Индия), Южная
Кашгария, Восточный Цайдам, центральная Гоби, полуостров Мангышлак (Устюрт).
7*
99
РОД ШИРОКОУШКИ. GENUS BARBASTELLA Gray, 1821
Зубная формула: I С Pm у Ртр у М у-2 = 34.
Два вида, относящиеся к этому роду, распространены от Англии
и юга Скандинавии до Юго-Восточной Азии. Оба вида представлены
в фауне СССР.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ШИРОКОУШЕК
1(2). На второй трети длины внешнего края уха расположена хо-
рошо обособленная кожная лопасть (рис. 21). Размеры относительно
Рис. 21. Головы
двух видов ши-
рокоушек.
Вверху — евро -
пейской (В. barba-
stellus). и вни-
зу — азиатской (В.,
leucomelas).
мелкие: кондилобазальная длина черепа 13—13,6 мм;
предплечье 36—41 мм . . Европейская широкоушка.
Barbastella barbastellus. (Стр. 100.)
2(1). Кожной лопасти на внешнем крае уха нет
(рис. 21). Размеры крупнее: кондилобазальная
длина черепа 14,2—14,9 мм; предплечье 41,5—45 мм.
. . Азиатская широкоушка. Barbastella leucomelas.
....................................(Стр. 100.)
20. Европейская широкоушка. Barbastella
barbastellus Schreber, 1774
Табл. VII, 3, Карта 27
Англия, Испания, Франция, Швейцария, Герма-
ния, Голландия, Дания, юг Скандинавии, запад и юг
Польши. В СССР — Литва, Беловежская пуща (в Бе-
лоруссии), Украина, Крым, Кавказ, Закавказье.
Поселяется в пещерах и разного типа подземных
сооружениях, реже (летом) в дуплах деревьев. Боль-
ших колоний не образует. На зимовках можно встре-
тить по нескольку зверьков вместе. Самки родят по
2 детеныша.
Географических форм не образует. Номинальная форма опи-
сана из Бургундии, Восточная Франция.
21. Азиатская широкоушка. Barbastella leucomelas Cretzchmar, 1826
Табл. VII, 4. Карта 27
В СССР — Закавказье (Армения, Азербайджан), Восточный Кав-
каз (Дагестан) и Средняя Азия: западная часть Копет-Дага (у Айдере
в долине р. Сумбар), долина р. Мургаб, в Туркмении, Восточный Узбе-
кистан, Западный Таджикистан и Южная Киргизия (Ош). За преде-
лами СССР — Синай (Западная Аравия); Яркенд, в Кашгарии; Север-
ная Индия (Гильгит, Дарджилинг), Непал, Индокитай и Япония (Хон-
сю) . Всюду редка.
Только в одном случае найден в древнем монастыре (Кегарт, Ар-
мения). Остальные жили в обширдых, преимущественно лессовых пе-
щерах. В одной пещере никогда не встречали более трех животных.
Зимуют там же, где живут летом.
Окраска подвержена широкой индивидуальной и географической изменчивости,
но для точного описания форм не хватает доброкачественно подобранного коллекцион-
ного материала. Номинальная форма описана из Синая (Аравия).
100
РОД ВЕЧЕРНИЦЫ. GENUS NYCTALUS Bowdich, 1825
Зубная формула: I -|-С у Pm -j-Pmp = 34.
Вечерницы населяют широколиственные леса северной половины
восточного полушария. На огромной территории сибирской тайги и
безлесных пространств Центральной Азии они не встречаются. Из
8 описанных видов вечерниц в фауне СССР представлено 3 и четвер-
тый, может быть, будет найден в пределах Южного Приморья.
Рис. 22. Черепа вечерниц:
А — В гигантской (Nyctalus lasiop-
terus; Г — Е — малой (N. leisleri).
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ВЕЧЕРНИЦ ФАУНЫ СССР
1(4). Размеры крупные; предплечье 59—69 лии; длина тела 82—
104 мм; хвост 55—67 мм; кондилобазальная длина черепа более 20 мм;
наибольшая ширина черепа не менее 13 мм; максиллярный ряд зубов
не менее 8 мм..................................................2
2 (3). Размеры очень крупные: предплечье 65—69 мм (редко от
64 мм); кондилобазальная длина черепа 22,2—23,4 мм; наибольшая ши-
рина черепа 13,9—14,8 мм; скуловая ширина 15,5—16,2 мм; максилляр-
ный ряд зубов 8,5—9,1 мм. Ухо 21—26 мм...........................
..............Гигантская вечерница. Nyctalus lasiopterus. (Стр. 102.)
3(2). Размеры немного меньше: предплечье 59—63 мм (редко до
65 мм); кондилобазальная длина черепа 20,3—21,4 мм; наибольшая ши-
рина черепа 13,4—13,8 мм; скуловая ширина 14,2—15,1 мм; максил-
лярный ряд зубов 8,1—8,5 мм. Ухо 18—20,5 мм......................
.............................Восточная вечерница. Nyctalus aviator.
4(1). Размеры меньше: предплечье не более 57 мм; длина тела
менее 81 мм; хвост не более 54 мм; кондилобазальная длина черепа
101
менее 19,5 мм; наибольшая ширина черепа не более 12,5 мм; максил-
лярный ряд зубов менее 7,5 мм........................................5
5(6). Размеры средние: предплечье 51—57 мм; кондилобазальная
длина черепа 17,4—19,4 мм; наибольшая ширина черепа 10,8—12,3 мм;
максиллярный ряд зубов 6,5—7,5 мм. Коронки верхнего клыка и
крупного предкоренного зуба сомкнуты; малый предкоренной зуб сбоку
не виден. Основания всех волос почти такой же окраски, как и вер-
шинные части или немного светлее........................................
......................Рыжая вечерница. Nyctalus noctula. (Стр. 102.)
6(5). Размеры относительно мелкие: предплечье 42,5—46 мм; кон-
дилобазальная длина черепа 15,2—16,1 мм; наибольшая ширина че-
репа 9,5—10,1 мм; максиллярный ряд зубов 5,8—6,3 мм. Между ко-
ронками верхних клыков и крупных предкоренных имеется промежуток
(хотя бы в виде узкой щели), в который сбоку обычно виден (хотя
бы частично) малый предкоренной зуб. Все волосы резко двухцвет-
ные: их основания значительно темнее вершинных частей...................
......................Малая вечерница. Nyctalus leisleri. (Стр. 103.)
22. Гигантская вечерница. Nyctalus lasiopterus Schreber, 1780
Табл. VII, 5. Карта 28
Лиственные леса Европы, в пределах СССР к северу до Москов-
ской области (Новый Иерусалим) и Красных Баков (на р. Ветлуге),
на восток до Бузулукского бора, на юг до Западного Закавказья и
Южного Крыма включительно.
Летом поселяется в дуплах деревьев, чаще всего по 2—3 особи
в колониях рыжих вечерниц, реже колониями по 2—3 десятка особей
среди себе подобных.
На востоке Азии живет близкий вид: N. aviator Thomas, 1911.
23. Рыжая вечерница. Nyctalus noctula Schreber, 1775
Табл. VII, 6. Карта 29
Лиственные леса Европы к северу до Англии, Балтийского по-
бережья, Ленинграда, южной части Кировской области, Западная Си-
бирь до Алтая (оз. Телецкое), Тарбагатай, Кульджа, Гималаи, Семи-
речье, Средняя Азия (лиственные насаждения Узбекистана), Кавказ.
В отличие от прочих видов, рыжая вечерница поселяется не только
в дуплах деревьев (которые, однако, явно предпочитает другим типам
убежищ), но также на чердаках больших домов и церквей (в Закав-
казье) или в камышовых настилах крыш (зимой в Ташкенте). Летние
колонии содержат от 2—3 или нескольких десятков до нескольких со-
тен особей. В Западной Европе окольцованные рыжие вечерницы со-
вершили перелеты на расстояние до 750 км.
Географическая изменчивость (в размерах и окраске меха) выражена очень
слабо.
Номинальная форма — N. n. noctula Schreber, 1774 — описана по экземпляру из
Франции. В фауне СССР представлена двумя очень близкими к ней формами:
a) N. п. princeps Ognev et Worobiev, 1923. (Тип из с. Хреновое Бобровского района
Воронежской области.) От номинальной западноевропейской формы отличается чуть
более крупными размерами черепа. Восточная Европа, Западная Сибирь и Кавказ.
b) N. n. meklenburzevi Kuzyakin, 1934. (Тип из г. Ташкента, Узбекистан.) По раз-
мерам сходна с предыдущей, но окраска меха светлее (буланая). Древесные насаж-
дения Узбекистана и Южного Казахстана.
102
24. Малая вечерница. Nyctalus leisleri Kuhl, 1819
Табл. Vll, 7. Карта 30
Лиственные леса Европы и Кавказа на север до Ирландии, а
в СССР — до Московской (или Ярославской?) области и Среднего По-
волжья.
Летом поселяется исключительно в дуплах лиственных деревьев,
образуя колонии, состоящие обыкновенно из 10—40 особей. Подобно
другим видам вечерниц, на кормежку вылетает рано, вскоре после за-
ката солнца. Всю среднюю часть ночи проводит в убежищах, а перед
рассветом вновь вылетает. Насекомых ловит в высоких слоях воздуха
над лесными полянами или над сплошным пологом леса. В питании
даже этой (малой) вечерницы главную роль играют крупные жестко-
крылые (например, майские и июньские жуки). Самки родят обыкно-
венно по 2, реже по 1 детенышу. О зимовках сведений нет.
В фауне СССР — номинальная форма (описана из Генау, Гессен, Германия).
РОД НЕТОПЫРИ И КОЖАНЫ. GENUS VESPERTILIO Linnaeus, 1758
Зубная формула: 1-|-СуРт^^Ртр-j-M-|--2 = 34 или 32.
К роду Vespertilio мы относим все виды, которые раньше разде-
лялись на несколько родов: Vespertilio, Pipistrellus, Amblyotus, Eptesi-
cus. К такому решению мы пришли в результате детального изучения
большого коллекционного материала по двум малоизвестным видам —
по восточному (или большому двухцветному) кожану, Vespertilio su-
perans Thomas, и особенно по кожановидному нетопырю, Vespertilio
savii Bonaparte. Эти виды нивелировали все основные различия между
прежними, выше перечисленными, родами. Для зоологов, незнакомых
с морфологией упомянутых видов, кажется объединение нетопырей, ко-
жанков и кожанов в один род Vespertilio искусственным, недостаточно
обоснованным. Мы приводили обоснования в книге «Летучие мыши»,
1950, но многие систематики не обратили должного внимания на эти
аргументы и продолжают считать в качестве самостоятельных родов
Pipistrellus, Eptesicus, некоторые и Amblyotus'. Уместно поэтому за-
ново разъяснить причины объединения перечисленных «родов».
Главным отличием рода Pipistrellus от Vespertilio и Eptesicus считают наличие
у Pipistrellus верхнего малого предкоренного зуба (Рт!); у Vespertilio и Eptesicus
этого зуба нет. Однако у дальневосточной формы V. savii, в большом количестве
добытой нами на юге Приморского края, в любой колонии, у части особей, неотли-
чимых по всем признакам, малые верхние предкоренные зубы имелись (как у ти-
пичных Pipistrellus). у другой части особей этих зубов не было (как у Vespertilio и
Eptesicus), а у некоторых особей на одной стороне верхней челюсти был ничтожно
мелкий малый предкоренной зуб (Рт1), но на другой стороне верхней челюсти его не
было. У V. savii верхний малый предкоренной зуб (если он имеется) очень мелкий
(рудиментарный). Неустойчивость рудиментарных органов общеизвестна, и наличие
(в данном случае) или отсутствие такого (рудиментарного) органа у особей одной и
той же колонии, сходных между собой по всем другим признакам, вполне естественно.
Выдающийся специалист по систематике рукокрылых — Г. С. Миллер (G. S. Mil-
ler, 1907, 1912) в качестве основного отличия рода Vespertilio от рода Eptesicus
приводил ямки на крыше носового отдела черепа. Однако такие же ямки есть у всех
взрослых и старых особей. V. savii, а несколько менее выраженные ямки имеются
у взрослых V. abramus и V. kuhli. Значит, этот признак для различения родов тоже
негоден.
С. И. Огнев (1928) для отличия Vespertilio от других родов приводил наличие
у самок V. murinus двух пар сосков. У большинства самок V. murinus сосков действи-
103
тельнО' две пары. Но В. С. и Л. С. Лавровы добыли в Воронежском заповеднике не-
сколько самок V. murinus, у которых было по одной паре сосков. А у V. superans,
описанного О. Томасом (1898) в качестве подвида V..murinus (так близки они морфо-
логически), у всех самок бывает только по одной паре сосков. Значит, и этот признак
для обособления Vespertilio от Eptesicus и Pipistrellus негоден.
Г. Топал (G. Topal, 1958) аргументирует разделение Vespertilio, Pipistrellus и
Eptesicus в разные роды по размерам и конфигурации os penis. Однако если этому
признаку придавать такое значение; то V. nathusii и V. abramus надо отнести к разным
семействам, хотя самки этих видов почти неразличимы, a Rhinolophus и Myotis по
такому признаку следовало бы объединить в один род.
В. А. Попов (1960, стр. 149 и 165) принял наше понимание рода Vespertilio, но
назвал его «несколько искусственным». Сравнивая только виды средней полосы Ев-
ропы, он ссылается на наличие малого предкоренного зуба, общие мелкие размеры и
некоторые черты экологии, будто бы позволяющие обособить Pipistrellus от Vespertilio
в особый род. Положение с Pm 1 мы уже пояснили. С размерами дело обстоит так:
V. bobrinskii, с большим трудом отличимый от центральноазиатских форм. V. nilssoni,
по размерам трансгрессирует со средними Pipistrellus. Во Вьетнаме и Бирме есть не-
топырь (V. mimus) по размерам еще меньше нашего нетопыря-карлика (V. pipistrel-
lus), но Эллерман и Моррисон-Скот к роду Pipistrellus относят сравнительно крупных
V. annectus (подрод Megapipistrellus) и даже гигантских кожанов, выделенных О. То-
массом (1902) и Г. С. Миллером (1907) в особый род Ja с одним видом Ja io — са-
мый крупный вид из всего семейства Vespertionidae.
Экологические различия, которые указывали бы на родовые несходства Vesper-
tilio, Pipistrellus и Eptesicus, нам не известны, а экологические сходства между ними
(по местам обитания, убежищам, миграциям, биологии размножения и т. д.) нам
известны.
Нет ни одного признака, по которому можно было бы отделить Pipistrellus
Eptesicus и Amblyotus от Vespertilio в качестве не только самостоятельных родов, но
даже подродов. С этим согласится каждый, кто сможет внимательно ознакомиться
с морфологией азиатских форм V. savii и V. superans. Недаром V. savii назван кожа-
новидным нетопырем.
Многочисленные виды кожанов и нетопырей распространены как в восточном,
так и в западном полушарии, к северу до границ высокоствольного леса.
В фауне СССР 11 видов.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ КОЖАНОВ И НЕТОПЫРЕЙ,
ПРЕДСТАВЛЕННЫХ В ФАУНЕ СССР
1(8). Имеется верхний малый предкоренной зуб (Pm1). Межбед-
ренная перепонка охватывает весь хвост или из нее выступает лишь
самый кончик хвоста длиной не более чем 2—3 мм, Эпиблема хо-
рошо развита и снабжена хрящевой поперечной перегородкой. Раз-
меры мелкие: предплечье не более 37 мм\ кондилобазальная длина
черепа меньше 13,4 мм\ длина максиллярного ряда зубов не более
5 мм.........................................................2
2(7). Внутренний верхний резец двухвершинный. Свободный край
крыловой и межбедренной перепонок без резко выраженной белесой
каймы .......................................................3
3(6). Длина penis не более 6 мм, Длина ois penis менее 2 мм.
Промежуток между верхним клыком и большим предкоренным зубом
настолько широк, что через него виден почти весь малый предкорен-
ной зуб (при рассматривании черепа сбоку). Если череп не очень
мелкий (кондилобазальная длина его более 12 мм и длина максилляр-
ного ряда зубов более 4,5 мм), то длина уха не менее 12 мм и козелок
не менее 6 мм.............................................. 4
4(5). Размеры очень мелкие: предплечье 28—33 мм\ кондилоба-
зальная длина черепа 11 —11,8 мм\ длина максиллярного ряда зубов
4—4,3 мм , , , Нетопырь-карлик. Vespertilio pipistrellus. (Стр. 107.)
5(4). Размеры крупнее: предплечье 33,5—36,5 мм\ кондилобазаль-
ная длина черепа 12,4—13,3 мм\ длина максиллярного ряда зубов
4,7—5 мм .... Лесной нетопырь. Vespertilio nathusii. (Стр. 108.)
104
6(3). Длина penis 10—12 мм\ хорошо развитая os penis длиной
около 5 мм. Промежуток между верхним клыком и большим предко-
ренным зубом очень узок: через него можно увидеть (при рассматри-
вании черепа сбоку) только вершинную часть малого предкоренного
зуба, оттесненного к внутренней стороне зубного ряда. Череп относи-
тельно крупный (кондилобазальная длина его 12,1—12,6 мм; длина
максиллярного ряда зубов 4,6—4,8 мм), но уши и козелок короткие;
длина уха 10—И мм, козелка 5—5,5 мм................................
. . . . Восточный нетопырь. Vespertilio abramus. (Стр. 108.)
7(2). Внутренний верхний резец одновершинный. Свободный край
межбедренной и крыловой перепонки (от ноги до пятого пальца)
с резко выраженной белесой каймой..................................
..........Средиземный нетопырь. Vespertilio kuhli. (Стр. 108.)
8(1). Верхнего малого предкоренного зуба нет, а если есть (неко-
торые особи кожановидного нетопыря), то penis Г-образно изогнутый.
Конец хвоста на 4—5 мм выступает из межбедерной перепонки. Эпи-
блема развита слабо и не имеет хрящевой поперечной перегородки, а
если эпиблема хорошо развита (у двуцветных кожанов), то размеры
относительно крупные; предплечье более 41 мм; кондилобазальная
длина черепа не менее 13,9 мм; длина максиллярного ряда зубов более
5 мм..............................................................9
9(14). Внешний верхний резец относительно крупный: вершина его
заходит за половину длины внутреннего резца, почти достигая уровня
добавочной вершины последнего. Предплечье не более 43 мм (обычно
значительно меньше)............................................. 10
10(11). Penis Г-образно изогнутый (в отличие от всех Vespertilio).
У некоторых особей есть верхний малый предкоренной зуб. Череп мел-
Рис. 23. Наружные половые органы самцов нетопырей:
,4. а — лесной нетопырь (Vespertilio nathusii); Б — б — нетопырь карлик (V. pipistrcl-
lus); В — в — восточный нетопырь (V. abramus); Г —,г — кожановидный нетопырь
(V. savii)
105
кий: кондилобазальная длина не более 14,2 лш, обычно значительно
меньше. Предплечье 33—40 мм.....................................
...............Кожановидный нетопырь. Vespertilio savii. (Стр. 109.)
11(10). Penis без прямоугольного перегиба в основной трети. Верх-
ний малый предкоренной зуб всегда отсутствует. Череп крупнее: кон-
дилобазальная длина не менее 14,4 мм, обычно значительно больше.
12(13). Размеры средние: предплечье 38—43 мм. Череп слабо
уплощен; высота его в области слуховых барабанов 6,4—7 мм. Ши-
рина между внешними сторонами
верхних клыков на 0,4—0,8 мм
превышает ширину межглазнич-
ного промежутка................
. . . . Северный кожанок.
Vespertilio nilssoni. (Стр. 109.)
13(12). Размеры мельче:
предплечье 34,5—36 мм. Череп
резко уплощен; высота его в об-
ласти слуховых барабанов 5, 4—
6 мм. Ширина между внешними
сторонами верхних клыков равна
ширине межглазничного проме-
жутка или превышает последний
не более чем на 0,2 мм . . . .
. . . Кожанок Бобринского.
Vespertilio bobrinskii. (Стр. 110.)
очень мелкий, не достигающий
Рис. 24. Зубы кожановидного нетопыря
(Vespertilio savii); правые верхние ряды
трех особей, добытых в одном месте
(близ Амурского залива, 1940).
14(9). Внешний верхний pi
даже трети длины внутреннего резца. Предплечье не менее 41 мм
(обычно значительно больше).....................................15
15(18). Задний верхний коренной зуб слабо сжат в передне-зад-
нем направлении, снабжен двумя внутренними вершинами и третьей
комиссурой. Эпиблема хорошо развита и обычно имеет поперечную
хрящевую перегородку. Короткие белые или светло-палевые вершинки
спинных волос на коричневом или темно-буром фоне спины создают ха-
рактерную серебристость.........................................16
16(17). Размеры относительно мелкие: кондилобазальная длина
черепа 13,9—16,2 мм; скуловая ширина 9,1—10,9 мм; длина максилляр-
ного ряда зубов 5—6,1. Предплечье 41—48 мм. Нёбных складок 7.
У самок две пары сосков (отличие от всех Vespertilio нашей фауны)
................Двухцветный кожан. Vespertilio murinus. (Стр. 111.)
Рис. 25. Хвосты и шпоры:
Л — кожановидного нетопыря (Vespertilio savii); ^ — лесного нетопыря
(V. nathnsii).
106
0,5cm
0
A
В
Рис. 26. Зубы:
А — кожан Огнева (V. ognevi); В —
северный кожанок (V. nllssoni).
17(16). Размеры крупнее: кондилобазальная длина черепа 16,7—
17 мм] скуловая ширина 11,5—11,6 мм] длина максиллярного ряда зу-
бов 6,3—6,6 мм. Предплечье 48—54 мм. Нёбных складок 8—9. У са-
мок одна пара сосков ................................................
................Восточный кожан. Vespertilio superans. (Стр. 111.)
18(15). Задний верхний коренной зуб резко сжат в переднезаднем
направлении, лишен задней внутренней вершины и третьей комиссуры
(в отличие от прочих Vespertilio). Вдоль
основания шпоры вместо эпиблемы тя-
нется мясистое утолщение, лишенное по-
перечной перегородки. Спина более или
менее одноцветная, без серебристой
ряби................................19
19(20). Размеры средние: пред-
плечье 42—46 мм; кондилобазальная
длина черепа 14,7—17 мм; скуловая ши-
рина 10,4—11,7 мм................Кожан
Огнева. Vespertilio ognevi. (Стр. 112.)
10(19). Размеры крупные: предплечье
48,5—55,5 мм] кондилобазальная длина
черепа 19—21 мм; скуловая ширина 13—
15 мм..........................Поздний
кожан. Vespertilio serotinus. (Стр. 112.)
25. Нетопырь-карлик. Vespertilio pipistrellus Schreber, 1775
Табл. VII, 8—9. Карта 31
От Ирландии и Западной Испании через Южную и Среднюю Ев-
ропу, Южную Норвегию и Швецию, Кавказ и Среднюю Азию до Запад-
ного Китая, в пределах которого граница распространения точно не
выяснена. Есть сборы из экваториальной Африки (Букоба, на запад-
ном берегу оз. Виктория). Северные находки в европейской части аре-
ала: Упсала (Юго-Восточная Швеция), оз. Селигер, окрестности Мо-
сквы, Рязанские Мещеры, Кузнецк (Саратовская область). В Средней
Азии наиболее северные находки были сделаны в Кзыл-Орде и Алма-
Ате.
Поселяется карлик в разнообразных убежищах, но непременно
связанных с жильем человека или расположенных вблизи от него;
в дуплах деревьев, в пещерах и в трещинах скал поселяется редко.
В конце лета (в Средней Азии) можно находить до 400—500
зверьков в одном убежище. На вечернюю кормежку вылетают рано,
вскоре после заката солнца. При обилии корма уже минут через 15—
20 возвращаются в свои убежища и сидят там до наступления рас-
света, когда вылетают на еще более короткое время и при этом не
кормятся, летая, видимо, для моциона. Главную пищу составляют мел-
кие насекомые, преимущественно двукрылые. Каждая самка нормально
приносит по два детеныша, реже по одному. Родятся самцы и самки
в одинаковом числе, а среди взрослых самцы становятся большой ред-
костью. Это характерно для всех видов нетопырей и кожанов. В конце
лета у взрослых самцов семенники сильно увеличиваются. По ночам
в воздухе происходят своеобразные брачные игры. Как старые самки,
так, видимо, и родившиеся в текущем году (т. е. в возрасте 3—4 ме-
сяцев) спариваются осенью со старыми самцами, а молодые самцы
достигают половой зрелости лишь на следующий год. Перед осенним
107
брачным периодом происходит единственная в году линька. Основная
масса нетопырей-карликов дальних сезонных миграций не совершает^
проводя зиму в районах летнего обитания.
В фауне СССР представлены 2 подвида, хорошо отличающиеся по окраске:
а) V. р. pipistrellus Schreber, 1775. (Описан из Франции.) Окраска меха на спине
темная, интенсивно-коричневая. Европа, Кавказ.
b) V. р. bactrianus Satunin, 1905. (Тип из Теджена, Южная Туркмения.) Окраска
меха бледная («пустынная»), на спине палево-желтоватая (различной интенсивности)
и узкая белесая каемка на свободном крае крыловой перепонки между ступней и
концом пятого пальца крыла. Азиатская часть ареала.
В тропической Азии есть близкие формы под другими видовыми названиями,
например индийский V. coromandra Gray, 1838.
26. Лесной нетопырь. Vespertilio nathusii Keyserling et Blasius, 1839
Табл. VII, 10. Карта 31
От запада Франции на восток до Оренбурга. Северные пределы
распространения от Балтийского побережья примерно по линии: Ле-
нинград— низовья р. Вятки. На юг ареал простирается до Среди-
земного и Черного морей, а в Закавказье — до границы с Ираном и
Турцией; несомненно, будет найден в Северном Иране и, вероятно^
в Турции.
Не избегая селений, лесной нетопырь теснее связан с лиственными
лесами и парками. Поселяясь в убежищах, связанных с жильем че-
ловека, он все-таки предпочитает им дупла деревьев с летным отвер-
стием в виде длинной щели. Неразмножающиеся зверьки поселяются
в скворечниках и дуплянках с круглым летным отверстием. Весной
в одном дупле находили до 30—60 особей, а к концу лета за счет под-
росшего молодняка колонии увеличиваются до 100 зверьков и более.
Весной нетопыри кормятся час-полтора, а в середине лета летают
почти всю ночь. Опушки лесов, небольшие поляны, просеки и аллеи —
наиболее характерные места их кормежки. В конце августа — начале
сентября лесные нетопыри в средней полосе из районов летнего оби-
тания исчезают. Пролетных зверьков наблюдали осенью в разных ме-
стах Украины и Закавказья. О зимовках ничего не известно. В осталь-
ном сходен с предыдущим видом.
Географическая изменчивость не изучена. В тропической Азии есть очень близкие
формы под другими видовыми названиями, например V. pulveratus Peters, 1871.
27. Восточный нетопырь. Vespertilio abramus Temminck, 1840
Карта 31
От Южно-Уссурийского края и Японии до Индии, Цейлона, Ма-
лайского архипелага. Филиппинских островов и севера Австралии
включительно. В пределах СССР найден только у залива Америки
(в Южном Приморье).
По образу жизни и географической изменчивости в условиях на-
шей страны сведений нет.
28. Средиземный нетопырь. Vespertilio kuhli Kuhl, 1819
Табл. VII, 11. Карта 32
Южная Европа (к северу до ГДР и ФРГ), острова Средиземного'
моря, Африка, Аравия, Малая Азия, Кашмир. В пределах СССР най-
ден в Крыму (долина р. Салгира, Ай-Тодор), на Кавказе (Баку, Куба>.
Евлах, Барда; Аджи-Кабул Сальянского района; урочище Аралых
у Арарата, Ереван, долина Аракса) и в низовьях Аму-Дарьи (Хива).
108
В Закавказье наблюдались колонии самок численностью от 3—5
до 20 особей в щелях кирпичных стен и за наличниками окон. На ве-
чернюю кормежку вылетают несколько позднее других нетопырей. Мо-
лодые особи кормятся среди построек, а старые куда-то улетают за
пределы селений. По причине крайней осторожности стремительно-
сти движений добывать этого редкого нетопыря очень трудно. Дете-
ныши родятся в июне.
Географическая изменчивость изучена слабо. Номинальная форма описана из
Триеста. На Аму-Дарье и в Закавказье живет, вероятно, V. k. lepidus Blyth, 1845.
{Описан из Кандагара, Афганистан.)
29. Кожановидный нетопырь. Vespertilio savii Bonaparte, 1837
Табл. VII, 12, 13. Карта 32
Южные страны Западной Европы, Крым (Карадаг, Кекенеиз),
Кавказ (Тбилиси, Кавказский заповедник), Юго-Западная Туркмения
(Кара-Кала), Устюрт, Западный Тянь-Шань (ущелье Тамерлана в го-
рах Куги-Танг), многие пункты Центральной и Восточной Азии, Юж-
ное Приморье (Владивосток, Новокисловская, Славянка, Краскино).
Южная часть ареала уходит до-Индии.
На Дальнем Востоке в период размножения и роста молодняка
самки поселяются на чердаках домов, крытых железом, колониями от
15—20 до 50—70 особей. На кормежку в этот период вылетают одно-
временно с обитающими в тех же местах двухцветными кожанами,
вместе .с ними на большой высоте в определенном направлении тянут
за пределы населенного пункта. Направление таких вечерних полетов
на места кормежки время от времени изменяется. Самки родят по 2 де-
теныша. К концу лета как вполне самостоятельный молодняк, так и
взрослые зверьки накопляют основательные жировые запасы; правиль-
ные вечерние перелеты на большие расстояния прекращаются, на день
зверьки прячутся в это время уже не группами, а порознь; если на
одном чердаке удается найти 2—3 животных, то они сидят непременно
вдали один от другого. Колонии размножающихся самок распадаются,
и вид становится «редким». О зимовках ничего не известно.
Был описан ряд форм под разными видовыми и даже родовыми названиями
(Eptesicus alachanis, Vesperugo caucasicus, Amlyotus tauricus, A. velox и др.), но все они
оказались лишь типами индивидуальной и географической изменчивости одного вида.
По накопившемуся теперь коллекционному материалу можно выделить 3 хорошо
дифференцированные географические формы:
а) V. s. savii Bonaparte, 1837. (Описана из Пизы, Италия.) Мелкий: предплечье
31—33 мм; кондилобазальная длина черепа 13—14 мм; темно окрашенный, с корич-
нево-золотистым блеском спины и постоянным наличием верхнего малого предкорен-
ного зуба. Средиземноморье, Западная Европа.
b) V. s. caucasicus Satunin, 1901. (Тип из Тбилиси, Грузия.) Крупный: предплечье
33—37 мм; кондилобазальная длина черепа 12,5—13,7 мм; очень светло окрашенный:
общий тон спины варьирует от желтовато-белого до серовато-песчаного, а нижняя
сторона — от серо-белесого до чисто белого; верхнего малого предкоренного зуба
почти во всех случаях нет. Крым, Кавказ, Средняя и Центральная Азия.
с) V. s. alaschanicus Bobrinskoy, 1926. (Тип из Хатынгол, Алашань.) Крупный:
предплечье 33—40 лш; кондилобазальная длина черепа 14,2 мм, и очень темно окра-
шенный со слабо развитым золотистым блеском на верхней стороне тела; верхние
малые предкоренные зубы у половины особей есть, у другой половины их нет. Ала-
шань, Южное Приморье.
30. Северный кожанок. Vespertilio nilssoni Keyserling et Blasius, 1839
Табл. VII, 14. Карта. 33
От Норвегии и Франции до Тихоокеанского побережья, на север
в западной части ареала заходит за полярный круг (оз. Имандра и
Эноре на Кольском полуострове). В Сибири к северу идет примерно
109
до 60° с. ш. Южные пределы летнего обитания в Европейской части
СССР: севернее Московской и Горьковской областей: лишь осенью
находили его и в более южных районах (Средняя Волга, Смоленск,
Восточные Карпаты). В Западной Европе найден в Швейцарии, Се-
верной Италии, Чехословакии. Широко распространен по центрально-
азиатским пустыням, указан для Тибета и Кашмира.
Летом северных кожанков находили на чердаках домов и в тре-
щинах скал. Часть животных остается в районах летнего обитания и
зимует в пещерах (например, в саблинских песчаниковых галереях,
под Ленинградом), другие совершают дальние сезонные переселения
(например, из Норвегии в пещеры Швейцарии). Это одна из наиболее
холодоустойчивых форм. А. Н. Формозов (1929) наблюдал кожанков
Рис. 27. Контуры двух близких видов:
А — северного кожанка (Vespertilio nilssoni) и Б — кожанка Бобринско-
го (V. bobrinskii).
в одном из горных, ущелий Монголии кормящимися в полуденные часы,
так как ночью там было настолько холодно, что насекомые не летали.
Шренк наблюдал кожанка летающим над Аргунью 12 октября, когда
у берегов реки образовались уже ледяные припаи. Самки родят по
2 детеныша.
2 хорошо отличающихся подвида:
а) V. п. nilssonii Keyserling et Blasius, 1839. (Описан из Швеции.) Окраска меха
темная: общий тон верхней стороны тела темный, коричнево-золотистый, с ярким
металлическим блеском; нижняя сторона темная, палево-бурая; череп с высокой моз-
говой капсулой. Вся европейско-сибирская часть ареала.
b) V. п. gobiensis Bobrinskoy, 1926. (Тип из гор Бурхастей-тала, Гобийский Алтай.)
Окраска меха очень светлая, шелковисто-блестящая, песчано- или желтовато-бурая
(разных тонов); мозговая капсула заметно уплощена. Центральноазиатская часть
ареала. Синонимы этой формы: Е. n. centrasiaticus Bobrinskoy, 1926 (тип из Хоту,
близ оз. Русское, Северный Тибет) и Е. n. kashgaricus Bobrinskoy, 1926 (тип с хребта
Русского, Хотан, Южная Кашгария).
31. Кожанок Бобринского. Vespertilio bobrinskii Kuzyakin, 1935
Табл. VII, 15. Карта 33
Пустыни Казахстана и Средней Азии (у оз. Шонтыкуль Куста-
найской области, к северу от г. Иргиза, Казалинск, колодец Тюлек,
к востоку от г. Аральска; пески Малые Барсуки, Бет-Пак-Дала);
ПО
окрестности Фаскала в Северной Осетии (?) и Якутск (?). Экземпляры
из двух последних мест молодые и плохой сохранности.
Этот своеобразный короткокрылый и, очевидно, быстролетный ко-
жанок поселяется в глинобитных стенах разрушенных построек и ста-
рых казахских гробницах, разбросанных в пустыне. В Малых Бар-
суках 3 зверька были пойманы в жилых помещениях, куда они зале-
тали через открытые окна на свет. В колониях около Иргиза в конце
июня 1947 г. были только беременные самки.
Этот вид очень близок к северному кожанку (V. nilssonii), особенно к его светло
окрашенной и плоскочерепной центральноазиатской форме (V. n. gobiensis Bobr.), но
вряд ли он имеет сходство с аравийским V. nasutus Dobson, 1877, экспонатов которого
в музеях СССР нет.
32. Восточный кожан. Vespertilio superans Thomas, 1890
Табл. VIII, 3. Карта 34
От Ордоса до Кореи и Советского Приморья: Краскино, Уссурийск,
ст. Козловская, около 160 км к югу от Хабаровска, дер. Кугелево Ле-
нинского района Хабаровской области; Япония (Хоккайдо и Хондо).
Летом поселяется (в Южном Приморье) на чердаках домов, кры-
тых железом. В середине ясного дня температура в местах, где сидят
эти животные, поднимается до 47°, а вероятно, и выше. В Уссурийске
в июле—августе 1940 г. в колониях находили до 100 и более животных,
из которых около 2/з приходилось на долю, подросшего молодняка.
В одном районе города колонии были почти на каждом чердаке боль-
шого дома. С наступлением сумерек восточные кожаны вместе с оби-
тающими в тех же местах двухцветными кожанами дружно тянули на
большой высоте за город; в разгар такой тяги все небо покрывалось
множеством равномерно движущихся в одном направлении силуэтов.
В Краскино в потоки двухцветных кожанов включались кожановидные
нетопыри. Возвращение насытившихся зверьков растягивалось от се-
редины ночи до наступления рассвета. В конце августа большие коло-
нии кожанов разбились на мелкие группы, а в сентябре все куда-то
исчезли. Подобно другим кожанам и нетопырям, самки родят по
2 детеныша.
Характерна резко выраженная сезонная изменчивость окраски меха: свежий
(осенний) мех темно-бурый, почти черный (с характерной серебристостью), к сле-
дующему лету становится коричнево-рыжим и серебристость его (за счет снашивания
коротких светло окрашенных вершинок) в значительной мере утрачивается.
33. Двухцветный кожан. Vespertilio murinus Linnaeus, 1758
Табл. VIII, 7, 2, Карта 34
От Англии, Норвегии, Франции до Тихоокеанского побережья к се-
веру местами до 60° с. ш. Южные находки лежат в Ордосе, Северо-За-
падных Гималаях (Гильгит), Южной Туркмении, Иране и Швейцарии.
Летом поселяется на чердаках домов, за обшивками стен, в дуп-
лах деревьев, под отставшей корой и т. д. На вечернюю кормежку вы-
летает сравнительно поздно и при недостатке корма летает почти всю
ночь без длительных перерывов. Редкие самцы живут отдельно от са-
мок, образующих колонии чаще всего по нескольку десятков особей.
Самки родят по 2 детеныша, в редких случаях — по 3. О зимовках
двухцветных кожанов ничего не известно.
111
Есть два типа окраски меха: черная и коричнево-рыжая; однако географического
разделения их нет. На Дальнем Востоке преобладают черные, на Кавказе — яркие,
коричнево-рыжие; во многих местах ареала встречаются и те и другие. Поношенный
мех даже у черной вариации заметно рыжеет.
34. Кожан Огнева. Vespertilio ognevi Bobrinskoy, 1918
Табл. VIII, < Карта 35
Найден в Туркмении (Кара-Кала, колодец Кафаклы, Огри-Даг,
Репетек, Тахта-Базар, Рабат-Кашан), в горной Бухаре (Сохта-Чинар,
Киик-Даган, Фейзабад, ущелье Тамерлана), в Казахстане (заповедник
Аксу-Джебоглы, северный берег Аральского моря) и в Юго-Западной
Монголии (Эцзин).
Поселяется этот редкий кожан в щелях глинобитных и каменных
стен, в трещинах скал, в небольших пещерообразных укрытиях и т. д.
Больших колоний, видимо, не образует, встречаясь поодиночке или по
нескольку особей в одном месте. На кормежку вылетает одновременно
с поздним кожаном и, например, в Тахта-Базаре вместе с ним тянет на
большой высоте против сильного ветра от города к -правобережным
сопкам Мургаба. За насекомыми же гоняется в местах, наиболее за-
щищенных от ветра. В кормных участках кормежка длится около 1 —
2 часов. Утром появляются в воздухе лишь отдельные особи, а осталь-
ные до следующего вечера не покидают своих укрытий.
Этот вид не имеет никакого отношения к Eptesicus sodalis Barret-Hamilton, 1910,
описанному из Румынских Карпат; Е. sodalis не вид, а короткокрылая морфа V. sero-
tinus, встречающаяся в разных частях его ареала.
35. Поздний кожан. Vespertilio serotinus Schreber, 1774
Табл. VIII, 5—6. Карта 35
От Англии, Франции, Испании и Западной Африки через Цент-
ральную и Южную Европу, Кавказ, Малую, Среднюю и Центральную
Азию до Кореи. Северная граница ареала идет от Англии через Вар-
шаву, Харьков и Оренбург примерно на оз. Балхаш. Указан для Каш-
мира (Северная Индия).
Летом поздние кожаны поселяются в разных убежищах, но непре-
менно связанных с жильем человека (чердаки домов, щели за дощатой
обшивкой стен и т. д.). Число особей, составляющих одну колонию, ко-
леблется от нескольких десятков до 3—4 сотен. Вылетают поздно и
сплошным потоком тянут к месту кормежки — в горы, открытую степь
или пустыню, где в ущельях, защищенных от ветра, около деревьев или
над скудной растительностью песчаных барханов преследуют свою до-
бычу, главным образом крупных жуков. Самки родят нормально по
2 детеныша. О зимовках надежных сведений нет. Дальних перелетов
поздние кожаны, видимо, не совершают.
Хорошо отличающиеся по окраске 2 географические формы:
а) V. s. serotinus Schreber, 1774. (Описана из Франции.) Окраска меха темная,
коричневато-бурая. Европа, Кавказ.
b) V. s. turcomanus Eversmann, 1840. (Описана по экземпляру с Устюрта.) Окраска
верха очень бледная, желтовато-соломенная сверху и бурая или грязно-белесая на
нижней стороне тела. Азиатская часть ареала.
В разных частях ареала этого вида встречаются отдельные особи с укорочен-
ным предплечьем (до 46—47 мм), но ничем более не отличающиеся от нормальных
По такому короткокрылому экземпляру (из Румынских Карпат) описан фактически не
существующий вид: Eptesicus sodalis Barret-Hamilton, 1910.
112
РОД СТРЕЛОУХИ. GENUS OTONYCTER1S Peters, 1859
Зубная формула: 1уС уРгпу PmpyM-|-2=30.
Крупные летучие мыши (предплечье 57—67 лш; кондилобазальная
длина черепа 21,8—23 мм; длина уха 30—41 мм).
Обитают в Северной Африке, Передней и Средней Азии.
Из двух известных видов в фауне СССР представлен один вид.
36. Белобрюхий стрелоух. Otonycteris hemprichi Peters, 1859
Табл. VIII, 7. Карта 36
От Алжирской Сахары через ОАР, Малую, Переднюю Азию и
Туркмению до Гиссаро-Алая, Западного Тянь-Шаня, Памира и Северо-
Западных Гималаев включительно. Найден в следующих местах Сред-
ней Азии: Большие Балханы, Кзыл-Арват, Гермаб (в Копет-Даге)
Аннау близ Ашхабада, Тедженский и Мургабский оазисы, Айвадж
в низовьях Кафирнигана, па ст. Шурча,
в долине Сурхан-Дарьи, Душанбе, Хо-
рог на Западном Памире; в ущелье
Тамерлана (Средняя Бухара), на Зе-
равшане и в Джан-Булаке (между
Ташкентом и Ходжентом).
Образ жизни не изучен. О. П. Бог-
данов нашел колонию из 68 стрелоухов
в глинобитном заборе поселка ст.
Шурча.
РОД ТРУБКОНОСЫ.
GENUS MURINA Gray, 1842
Зубная формула:
I-С- Pm —Pmp —М —-2 = 34.
3 1 1 н 1 3
По густой опушенности верхней
стороны межбедренной перепонки, на-
личию кожистых трубок, на концах ко-
торых открываются ноздри, по харак-
терно всклокоченному густому меху
с выступающей из него блестяще окра-
шенной остью, по правильно эллипсо-
видной форме ушей с длинным заост-
ренным козелком и очень широким ту-
поконечным крыльям эти очень редкие
вымирающие зверьки легко отличают-
ся от любых летучих мышей фауны
СССР. Азия от Красноярска, Алтая и
Сахалина до Юго-Восточной Азии и
Цейлона.
Из 5 видов Murina в фауне
СССР — 2 вида.
8 Н. А. Бобринский и др.
Рис. 28. Череп малого трубконоса
(Murina aurata):
а — сбоку; б — снизу; в — сверху.
Рис. 29. Голова малого трубко-
носа (Murina aurata).
113
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ТРУБКОНОСОВ ФАУНЫ СССР
1(2). Размеры относительно крупные: предплечье 40—42,5 мм; кон-
дилобазальная длина черепа 17,7—17,8 мм; длина максиллярного ряда
зубов 6,3—6,6 мм. Вершина козелка отклонена наружу. Свободный
край крыловой перепонки прикрепляется близ основания внешнего
пальца ноги. Сагиттальный гребень на черепе хорошо выражен . . .
............... Большой трубконос. Murina leucogaster. (Стр. 114.)
2(1). Размеры значительно мельче: предплечье 29,5—31,5 мм; кон-
дилобазальная длина черепа 14,3—14,5 мм; длина максиллярного ряда
зубов 5—5,1 мм. Козелок прямой. Свободный край крыловой перепонки
прикрепляется к конечной половине внешнего пальца ноги, почти до-
стигая основания когтя последнего. Сагиттального гребня на черепе нет.
......................Малый трубконос. Murina aurata. (Стр. 114.)
37. Большой трубконос. Murina leucogaster Milne-Edwards, 1872
Табл. VIII, 8—9, Карта 35
Найден в следующих местах: близ г. Красноярска, у с. Б. Кемчуг
Красноярского края, в бассейне р. Казыр-Сук (верховья Енисея); улус
Кечино в Кузнецком Алатау; у оз. Телецкое на Алтае; в нескольких
местах Уссурийского края (Супутинский заповедник, Владивосток, Кед-
ровая падь); Сахалин (у Южно-Сахалинска, в частности); Иезо (в Япо-
нии); Дарджилинг (Гималаи).
На берегу Телецкого озера взрослый зверек на день забрался
в сверток березовой коры, где и был добыт К. Н. Благосклоновым. В пе-
щерах около г. Красноярска Н. Оводов и Н. Высоковский нашли в ян-
варе 1960 г. на зимовке 9 особей этого вида. Летом 1961 г. в лесу близ
с. Б. Кемчуг Э. Шейнфельд поймала 3 больших трубконосов в давилки,
расставленные (на земле) для вылова мелких грызунов и землероек.
Характерны две резко различные красочные вариации (морфы): однотонно серая
и яркая, кирпично-красная с белым брюхом. Географическая изменчивость не выра-
жена. Вид описан по экземпляру из Сычуани.
38. Малый трубконос. Murina aurata Milne-Edwards, 1872
Табл. VIII, Ю. Карта 35
На территории СССР известны только 6 экземпляров: 4 из них до-
быты в Уссурийских лесах (близ дер. Евсеевки Иманского района,
в долине р. Одарки, в Супутинском заповеднике) и 2 — на Южном Са-
халине. За пределами СССР — на о-ве Яку (в Южной Японии) и в ма-
териковых районах Юго-Восточной Азии.
В середине августа 1940 г. в Супутинском заповеднике один моло-
дой зверек кормился в глубине тайги, шумно летая между деревьями
и вдоль таежной тропинки. Появлялся он поздно, с наступлением гу-
стой темноты, а в середине ночи исчезал и не показывался до следую-
щего вечера. 16 августа этот зверек влетел через открытую дверь в лес-
ную сторожку на свет керосиновой лампы.
СЕМЕЙСТВО БУЛЬДОГОВЫЕ. FAMILIA MOLOSSIDAE
Многочисленные представители этого семейства распространены
в тропических и субтропических областях всего земного шара. Благо-
даря предельному для рукокрылых совершенству летательного аппа-
рата (весьма прочное сочленение плеча с лопаткой, узкие и длинные
114
крылья, совершенный рулёвой и тормозной аппараты), они господ-
ствуют над всеми насекомоядными летучими мышами тропической
фауны. Г. С. Миллер (1907) делил это семейство на 10 родов, из кото-
рых в фауне СССР представлен только один — Tadarida.
РОД СКЛАДЧАТОГУБЫ. GENUS TADARIDA Rafinesque, 1814
Зубная формула: I ^ууС уРт у Ртр у Му-2 = 32 или 30.
Рис. 30. Череп широкоухого складчатогуба (Tadarida teniotis):
а — сбоку; б — сверху; в — снизу.
Около 60 форм складчатогубов распространено в пределах всего
ареала семейства (т. е. в тропиках и субтропиках земного шара). Осо-
бенно большим числом форм они представлены в пределах Экватори-
альной Африки. В фауне СССР представлен только один вид — Tada-
rida teniotis.
39. Широкоухий складчатогуб. Tadarida teniotis Rafinesque, 1814
Табл. VIII, 11. Карта 36
Отдельные находки этого редкого вида разбросаны на огромной
территории субтропиков Африки, Европы и Азии (от Атлантического
до Тихоокеанского побережья). В СССР найден только в следующих
8*
115
Рис. 31. Голова широко-
ухого складчатогуба
(Tadarida teniotis):
Профиль (точное изображение)
и вид снизу (изображение не
вполне точное).
местах: в окрестностях ст. Кара-Куль (в
50 км от г. Бухары), в горах Бабатаг, на
границе между Узбекистаном и Таджики-
станом (О. П. Богданов), близ г. Ош
в Киргизии (А. А. Тагильцев), у Айдере,
в долине Сумбара (X. Бабаев), в Шушин-
ском ущелье (Нагорный Карабах, За-
кавказье), у г. Кисловодска и на юге Ка-
бардино-Балкарии (А. К. Темботов).
Взрослый самец широкоухого склад-
чатогуба, добытый в 1936 г. близ Кара-
Куля Кадыровым, в середине летнего дня
залетел в открытую дверь кибитки и при-
цепился к висевшему на стене халату. Ка-
дыров подошел к нему и взял в руки.
В Шушинском ущелье за 1938 и 1939 г.
было добыто 15 складчатогубов. На день
они забирались там в узкие щели отвесных
каменных стен каньона то поодиночке, то
группами от 3—4 до 10—12 особей. На кор-
межку вылетали поздно, уже в темноте,
а возвращение растягивалось до рассвета,
когда можно было свободно рассмотреть
их своеобразные 'огромные острокрылые
стремительно движущиеся силуэты с характерно выступающей вперед
головой, окаймленной широкими лопастями ушных раковин, которые,
очевидно, служат прекрасным рулем высоты.
Широкоухий складчатогуб— самый крупный из всех бульдоговых
летучих мышей: предплечье его достигает 60—63 мм\ кондилобазальная
длина черепа 23—24 мм. На лету он более всего похож на крупных бе-
лобрюхих стрижей. Любопытно, что эти животные из 4—5 различных
голосовых вариаций составляют своеобразную песенку; при содержа-
нии других летучих мышей в неволе ничего подобного слышать не
приходилось.
Вид представлен двумя резко различными вариациями окраски: рыжей и одно-
тонной пепельно-серой. На Кавказе и в Средней Азии добывались только особи с пе-
пельно-серой окраской меха.
Н. А. Бобринский
ОТРЯД ХИЩНЫЕ. ORDO CARNIVORA
Хищные—наземные, реже полуводные млекопитающие различных
размеров (от ласки до белого медведя) с хорошо обособленными от
туловища стопо-, полустопо- или пальцеходящими конечностями с че-
тырьмя или пятью пальцами, вооруженными хорошо развитыми ког-
тями. Большие полушария головного мозга с ясно выраженными изви-
линами. В основном плотоядные, хотя имеются всеядные и даже ра-
стительноядные виды. Зубы всех четырех типов. Резцы небольшие,
слабые. Клыки сильные и длинные. Число и форма коренных зубов из-
меняются в широких пределах, но передние из них (за редким исклю-
чением) сжатые с боков и остробугорчатые. Последний предкоренной
зуб верхней челюсти и первый коренной нижней челюсти обыкновенно
резко выделяются своей величиной и формой. Эти зубы называют
«хищническими». Зубы, расположенные за хищническими, имеют остро-
или тупобугорчатую жевательную поверхность.
Распространены в Евразии с прилегающими островами, в Африке
с Мадагаскаром, в Северной и Южной Америке. В Австралии един-
ственный вид — собака динго.
В СССР хищные имеют важное значение в пушном промысле, за-
нимая первое место среди всех прочих отрядов. Некоторые виды раз-
водят в звероводческих хозяйствах. Многие виды полезны истребле-
нием вредных грызунов. Волки сильно вредят уничтожением домаш-
него скота.
Отряд хищных делят на 7 семейств, из которых в фауне СССР
представлены 6 (отсутствует семейство виверровых).
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ ОТРЯДА ХИЩНЫХ
ФАУНЫ СССР
1(4). На задних лапах по 4 пальца. Костное нёбо сравнительно
короткое и широкое; оно не доходит до линии, соединяющей задние
края последних зубов (рис. 39), а если заходит за эту линию, то менее
чем на половину своей ширины между этими зубами...........2
2(9). Голова вытянутая (собачьего типа). Хвост опушен нерав-
номерно: волосы у основания короче, чем посредине, вследствие чего
хвост суживается у корня. Наружные края ноздрей окаймлены более
или менее широким пространством голой кожи. Когти невтяжные, при-
тупленные, слабо искривленные. В верхней челюсти с каждой стороны по
4 предкоренных зуба (1 хищнический большой и 3 малых) ..... 3
3(8). На передних лапах по 5 пальцев. Задние ноги равны
или длиннее передних, благодаря чему у стоящего зверя крестец
117
располагается не ниже лопаток. Окраска меха без поперечных полос.
В верхней челюсти с каждой стороны по два собственно коренных зуба
(рис. 33, А). В нижней челюсти с каждой стороны по 6 или 7 предко-
ренных и коренных зубов..............Собачьи. Canidae. (Стр. 120.)
4(1). На задних лапах по 5 пальцев. Костное нёбо сравнительно
длинное и узкое: оно продолжается за линию, соединяющую задние
рр
Рис. 32. Череп и зубы лисицы (Vulpes vulpes): А — сбоку;
Б — снизу.
Зубы: /*—/3, Z|—i3—резцы; С, с—клыки; Ртх— Ptn\ p/ni—p/n, — малые
предкоренные; Ртр, ртр — крупные предкоренные; ЛГ—М2, тх—т9 — задне-
коренные зубы. Ртр и т1 — хищнические зубы.
Череп: Ъо — слуховой барабан; fi — предглазничное отверстие; fin, — перед-
ненёбное отверстие; frt — крыловидная впадина; тр — задний край костного
нёба; рая —угловой отросток; par — сочленовный отросток; рс — венечный
отросток нижней челюсти; рр — надглазничный отросток; ppt — крыловидный
отросток.
Точки приложения ножек штангенциркуля при промерах черепа: а—b — кон-
дилобазальная длина; d — е — наибольшая длина черепа; f — g — скуловая
ширина.
края последних зубов, более чем на половину своей ширины между
этими зубами....................................................................5
5(7). Хвост длинный (длиннее задней ступни) и всегда явственно
выступает из меха туловища. Размеры средние или мелкие: длина тела
не более 1 ж; кондилобазальная длина черепа менее 170 мм. Нижний
край нижней челюсти не имеет направленного внутрь отростка. Слухо-
вые барабаны крупные и сильно вздутые...........................................6
118
6(10). На хвосте чередуются темные и светлые поперечные кольца.
Пальцы передних лап голые, задних покрыты редкими волосами.
В верхней челюсти по 2 собственно коренных зуба (рис. 33, В) . . .
..............................Енотовые. Procyonidae. (Стр. 129.)
7(5). Хвост очень короткий (короче задней ступни) и почти скрыт
в меху туловища. Размеры зверя очень крупные: длина тела взрослых
1,5—2 м и более; кондилобазальная длина черепа более 250 мм. На
нижнем крае челюсти
под угловым отростком
имеется небольшой за-
гнутый внутрь альвеоляр-
ный отросток (рис. 34).
Слуховые барабаны ма-
лы и уплощены ....
............. Медвежьи.
Ursidae. (Стр. 130.)_
Рис. 34. Нижняя челюсть
медведя (Ursus), основ-
ная часть правой сто-
роны, вид снизу:
ра—альвеолярный отросток.
Рис. 33. Верхние ряды зубов левой стороны:
А — волка (Canis lupus); Б — медведя (Ursus arctos);
В — енота (Ргосуоп lotor); Г—куницы (Martes martes);
Д — барсука (Meles meles); Е — полосатой гнены (Hyaena
hyaena); Ж — леопарда (Panihera pardus).
8(3). На передних лапах по 4 пальца. Задние ноги короче перед-
них, благодаря чему у стоящего зверя крестец располагается ниже ло-
паток. Окраска меха с неясными поперечными полосами. В верхней
челюсти с каждой стороны по 1 собственно коренному зубу (рис. 33).
В нижней челюсти с каждой стороны по 4 предкоренных и коренных
зуба............................Гиеновые. Hyaenidae. (Стр. 133.)
9(2). Голова округлая (кошачьего типа). Хвост опушен равно-
мерно и поэтому имеет одинаковую толщину на всем протяжении. Во-
лосяной покров достигает наружного края ноздрей. Когти втяжные,
очень острые, сильно искривленные (за исключением гепарда, имею-
щего полувтяжные притупленные когти). В верхней челюсти с каждой
стороны не более 3 (хищнический и 2 малых) предкоренных зубов
(рис. 33)..........................Кошачьи. Felidae. (Стр. 133.)
119
10(6). Хвост не имеет поперечных полос. Пальцы, по крайней мере
сверху, густо покрыты волосами. В верхней челюсти с каждой стороны
по 1 собственно коренному зубу (рис. 33)............................
......................................Куньи. Mustelidae. (Стр. 142.)
СЕМЕЙСТВО СОБАЧЬИ. FAMILIA CANIDAE
Средних размеров пальцеходящие хищники с удлиненной мордой,
длинным, обычно пушистым хвостом, относительно высокими ногами.
Передние конечности пятипалые, задние четырехпалые (у одного аф-
риканского рода обе пары конечностей четырехпалые). Когти тупые,
невтяжные. Слуховые барабаны черепа умеренно или сильно, взду-
тые. Хищнические зубы хорошо
Рис. 35. Череп енотовидной собаки (Nycte-
reutes procyonoides), вид сбоку. Виден вы-
ступ нижнего края нижней челюсти под
угловым отростком.
развиты.
Зубная формула:
т 3 r 1 п 4 ,, (3) 2 о
I — С — Pm — .2 = 42, реже
3 1 4 3 (2) г
44 или 40.
Образ жизни наземный.
Плотоядные, частично раститель-
ноядные.
Распространены по всем
материкам, кроме Антарктиды,
и по большинству материковых
островов.
Всего около 10 родов, из них
в СССР представлено 4.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ СЕМЕЙСТВА СОБАЧЬИХ
ФАУНЫ СССР
1(6). Щеки светлые (рыжеватые, серые, желтоватые, белые). Во-
лосы на боках головы не удлинены и не образуют пышных «бак». На
нижней челюсти под угловым отростком большого полукруглого выро-
ста нет. Вершины верхних клыков направлены вниз..................2
2(5). Хвост, измеренный с концевыми волосами, равен половине
длины головы и туловища. На подошвах задних ног пяточные мозоли
лишены волос. Зрачок круглый. Заглазничные отростки черепа сверху
выпуклые.........................................................3
3(4). Окраска спины серая, буровато-серая или рыжевато-серая.
На спине имеется примесь черных остевых волос. Хвост малопышный.
В нижней челюсти 7 предкоренных и коренных зубов...................
.....................................Собаки. Canis. (Стр. 121.)
4(3). Окраска спины изменяется от светлой палевой до насы-
щенной охристо-рыжей. На спине примеси черных волос нет. Хвост
очень пышный. В нижней челюсти только 6 предкоренных и коренны?;
зубов............................Красные волки. Cuon. (Стр. 123.)
5(2). Длина хвоста, измеренная с концевыми волосами, значи-
тельно превышает половину длины головы и туловища. На подошвах
задних лап пяточные мозоли покрыты волосами. Зрачок вытянутый
в вертикальном направлении, продолговатый. Заглазничные отростки
сверху плоские или даже вогнутые . . . Лисицы. Vulpes. (Стр. 124.)
6(1). Щеки черные или черноватые, окаймленные светлыми воло-
120
основания коронки пер-
Рис. 36. Верхние коренные
зубы:
А — волка (Canis lupus); Б —
шакала (Canis aureus).
сами. На боках головы волосы удлинены и образуют пышные «баки».
На нижней челюсти под угловым отростком имеется большой закруг-
ленный выступ (рис. 35). Вершины верхних клыков направлены вниз и
наружу...............Енотовидные собаки. Nyctereutes. (Стр. 128.)
РОД СОБАКИ. GENUS CANIS Linnaeus, 1758
Зубная формула: СуРшуМу-2 = 42.
Евразия, Африка (на Мадагаскаре нет), Северная Америка, Авст-
ралия (динго).
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CANIS ФАУНЫ СССР
1(2). Размеры крупные: длина тела взрослых более 90 см; общая
длина черепа более 20 см. Ободок (cingulum)
вого коренного зуба верхней челюсти (М1)
с наружной стороны зуба узкий и иногда пре-
рывается посредине (рис. 36, А)............
...............Волк. Cams lupus. (Стр. 121.)
2(1). Размеры мельче: длина тела менее
90 см; общая длина черепа меньше 19 см.
Ободок основания, коронки первого коренного
зуба (М1) верхней челюсти с наружной сто-
роны широкий и хорошо выраженный на всем
протяжении (рис. 36, Б)....................
. . . . Шакал. Canis aureus. (Стр. 122.)
1. Волк. Canis lupus Linnaeus, 1758
Табл. IX, 1. Карта 37
Прежде населял' всю Европу, большую
часть Азии и Северную Америку; ныне в ряде
стран Западной Европы (Англия, Голландия, Дания, большая часть
Германии) истреблен. В СССР встречается почти повсеместно, кроме
Крыма (где истреблен) и некоторых островов Арктики.
Живет в самых различных местностях: в тундре, в лесах, в степи
и пустыне, на равнинах и высоко в горах. Избегает только сплошной
тайги, где нет домашнего скота и мало диких копытных. Охотится в су-
мерки и ночью, но в глухих местах иногда рыщет и среди дня. Основ-
ная пища — домашний скот (особенно овцы) и дикие копытные, зайцы,
мелкие грызуны, различные птицы. Но вообще пища, в зависимости
о г места и времени года, весьма разнообразна. Волки поедают всякую
падаль, гнездящихся на земле птиц, их птенцов и яйца, лягушек, яще-
риц, даже насекомых, различные ягоды, плоды и т. д. Течка у старых
волчиц протекает в южных районах в конце декабря — начале января,
в средней полосе — в январе, на севере — в феврале. Течка у молодых
волчиц происходит на 2—3 недели позже. Самцы нередко сильно
дерутся из-за самок, особенно молодых. Спарившиеся волки выбирают
место будущего логова. Логово располагается в различных угодьях, но
всегда поблизости от воды, в сухом, укрытом месте. В средней полосе
оно устраивается обычно по лесистым грядам среди болот, в чаще лес-
ного молодняка, в тенистых участках строевых лесов, по оврагам, по-
росшим кустарником, в зарослях камыша по берегам озер и болот;
в горах логово располагается под скалой, в расселине или в пещере.
121
В степях волки обычно роют неглубокие норы или занимают норы бар-
сука, сурка или лисицы (расширив их), на склонах оврага или балки
либо устраивают логово в камышах по озерам. После 63—65-дневной
беременности волчица родит от 2 до 12 волчат (молодые обычно 3—
4 волчат, старые 6—8). На юге волчата родятся обычно в конце марта
и в апреле, в средней полосе — в апреле—мае, на севере — в июне и
даже в июле. Щенки прозревают на 10—12-й день. Волчица кормит
волчат молоком в течение 5—6 недель, но уже через месяц после их
рождения начинает подкармливать отрыжкой своей полупереваренной
пищи. В первое время после рождения молодняка самка не отходит от
гнезда и самец приносит ей пищу. Позднее оба родителя начинают
приучать волчат ловить и убивать добычу. Осенью к выводку волков
присоединяются «переярки» (молодые, родившиеся в прошлом году), и
вся семья начинает кочевать в поисках пищи. Такую кочевую жизнь
семья волков ведет всю зиму, постепенно расширяя район своей дея-
тельности. Когда начинается течка волчиц, эти семьи распадаются.
Половозрелыми молодые волки становятся в возрасте 21—22 ме-
сяцев.
Волки приносят огромный вред животноводству и охотничьему
хозяйству уничтожением скота и охотничьих животных. В очень ред-
ких случаях нападают на человека. Охота на волков разрешена круг-
лый год всеми способами, кроме общеопасных. За истребленных волков
выплачивают премию.
Географическая изменчивость волков очень велика, но еще слабо изучена. На
территории СССР можно различить следующие формы:
а) С. 1. lupus Linnaeus, 1758. (Место описания — Швеция.) Размеры крупные: вес
до 78 кг. Мех средней высоты, густоты и мягкости. Окраска довольно темная, обычно
буровато-серая или рыжевато-серая, реже чисто-серая. В степных районах преобла-
дают волки с рыжеватым мехом. Брюхо обычно беловатое. Европейская часть СССР.
Степных волков (С. 1. campestris Dwigubski, 1804) с более грубым и рыжеватым ме-
хом мы относим к этому же подвиду, в качестве особой natio.
b) С. 1. albus Kerr, 1792. (Описана с низовьев р. Енисея.) Размеры меньше: вес
до 60 кг. Волосяной покров очень пышный и мягкий. Окраска зимнего меха светлая,
серая иногда с легким рыжеватым оттенком. Черные волосы расположены обычно
по хребту. Тундровая зона СССР.
с) С. 1. altaicus Noack, 1911. (Тип с Алтая.) Размеры крупные: вес самцов до
72 кг. Волосяной покров высокий и мягкий, но все же значительно более низкий
и не столь шелковистый, как у тундровых волков. Окраска зимнего меха красивого
голубовато-серого цвета, в южных районах ареала иногда с рыжеватым оттенком.
Черные направляющие волосы не только на спине, но, и на боках. Таежная зона Си-
бири и Север Казахстана.
d) С. 1. cubanensis Ognev, 1923. (Тип из г. Майкопа.) Размеры средние: вес до
50 кг. Волосяной покров редкий, грубый. Зимний мех грязного серо-охристого или
буро-серого цвета. Брюхо сероватое или желтоватое. Кавказ.
е) С. 1. desertorum Bogdanov, 1882. (Описана из Хивинского оазиса и Кызылку-
мов). Размеры очень мелкие: вес до 40 кг. Мех грубый и редкий. Окраска зимнего
меха светлая, желтовато-серая с рыжим оттенком на спине. Пустыни Средней Азии
и Юго-Западного Казахстана.
f) С. 1. chanco Gray, 1863. (Описана из Гималаев.) Размеры мелкие: вес до 45 кг.
Волосяной покров зимой длинный и мягкий. Окраска светлая, чисто-серая или с при-
месью желтых оттенков и с темным «ремнем» по хребту. Горы Тянь-Шаня и Памира.
2. Шакал. Canis aureus Linnaeus, 1758
Табл. IX, 2. Карта 37
Вся Индия с Цейлоном, Пакистан, Афганистан, Средняя Азия,
Иран, Турция, Кавказ, Балканский полуостров, Ближний Восток и Се-
верная Африка. В СССР — Черноморское побережье Кавказа на север
до Новороссийска, Рионская низменность, равнины Восточного Закав-
казья, долины рек Аракса и Куры и побережье Каспийского моря до
122
устья Терека, Западная и Южная Туркмения, равнинная и предгорная
части Таджикистана, вся долина Аму-Дарьи, низовья р. Зеравшана и
район среднего течения р. Сыр-Дарьи.
Излюбленные местообитания — густые тростниковые и кустарнико-
вые заросли по берегам рек и озер, низменные леса (в Закавказье),
а также предгорья, богатые расселинами, скалами и пещерами. Охотно
держится близ человеческих 'поселений. В горах поднимается обычно
не выше 1000 м. Охотится парами или небольшими группами после за-
хода солнца, но иногда и днем. Продолжительность беременности
60 дней. Самка щенится обыкновенно в норе, вырытой ею в обрывах,
в скрытных местах. Иногда использует норы лис и барсуков. Молодые
родятся в апреле — начале мая. Число щенков в помете от 4 до 9
(обычно 6—7). Питается разнообразной животной и растительной пи-
щей: падалью, мелкими зверями и птицами, яйцами, ящерицами,
змеями, лягушками, рыбой, насекомыми, моллюсками, червями, раз-
личными плодами- и ягодами. Приносит существенный вред, уничтожая
мелких домашних животных и охотничьих зверей и птиц (особенно
фазанов). Поэтому добыча шакалов разрешена в любое время года.
В силу низкой ценности шкурки и малочисленности шакалов промысло-
вое значение их ничтожно.
В СССР распространены 2 формы:
а) С. a. aureus Linnaeus, 1758. (Описана из провинции Лар, Иран.) Мех относи-
тельно мягкий, сероватой окраски. Средняя Азия.
b) С. a. moreoticus Geoffrey, 1835. (Тип из Мореа, Греция.) Мех несколько грубее,
в окраске его развиты ржавые тона. Кавказ.
РОД КРАСНЫЕ ВОЛКИ. GENUS CUON Hodgson, 1838
Зубная формула: I у С -?-Ргп -^-М — 2 = 40.
Несколько мельче обыкновенного волка: длина тела около 1 м.
Хвост очень пушистый, длина его равна примерно половине длины тела.
Горы Центральной и Восточной Азии к югу до Малакки, острова
Индонезии, вся Индия.
3. Красный волк. Cuon alpinus Pallas, 1811
Табл. IX, 3. Карта 37
Широко распространен в горах Южной и Центральной Азии.
В СССР — Восточный Памир, система Тянь-Шаня с Джунгарским Ала-
тау, Тарбагатай, Алтай, Саяны, Южное Прибайкалье и Забайкалье,
среднее течение Амура и Уссурийский край. В СССР всюду крайне ре-
док, лишь в Уссурийском крае встречается немного чаще.
Горное животное. Питается дикими копытными (горными козлами,
косулями, маралами), сурками и другими зверями, а также птицами.
Зимой держится группами (семьями), а иногда стаями до 20 зверей.
Биология слабо изучена.
В СССР описано 2 подвида:
а) С. a. alpinus Pallas, 1811. (Тип с р. Уды, на Дальнем Востоке.) Размеры круп-
ные. Окраска зимнего меха интенсивная, ржаво-красная. Лобная часть черепа вы-
пуклая, носовые кости сравнительно короткие. Приморье, Приамурье, Южное Забай-
калье, Восточные Саяны.
b) С. a. hesperius Afanasiev et Zolotarev, 1935. (Тип с Аксая, в Семиречье.) Раз-
меры мельче. Ржаво-красные оттенки в окраске зимнего меха выражены слабо. Лоб-
ная часть черепа плоская. Носовые кости удлиненные. Алтай, Тянь-Шань, Памир.
123
РОД ЛИСИЦЫ. GENUS VULPES Oken, 1816
Зубная формула: I у Су PmyMy -2 = 42.
Средние и мелкие представители семейства с длинным телом и от-
носительно короткими ногами. Хвост пышно опушенный, длинный
(длина его с волосами более V2 длины тела). Ступни ног сплошь по-
крыты мехом, или кроме небольших мозолей на концах пальцев. Зра-
чок эллиптический.
Евразия, Африка (без Мадагаскара), Северная Америка.
Из 7—8 видов в СССР 4 вида (2 подрода).
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА ЛИСИЦ ФАУНЫ СССР
1(6). Уши заостренные и длинные: если их пригнуть к голове, они
доходят своими вершинами по крайней мере до глаз. Мозоли на кон-
цах пальцев лишены волос. Заглазничные отростки сверху ясновогну-
тые. При сомкнутых челюстях вершины нижних клыков далеко заходят
за альвеолы верхних клыков (рис. 37, £>, В)....................2
2(3). Тыльная сторона ушей черная или черноватая, резко отлич-
ная от цвета шеи и спины. Конец хвоста белый. На передней стороне
ног черные или темно-серые пятна. Общая длина черепа более 12 см.
.................Обыкновенная лисица. Vulpes vulpes. (Стр. 125.)
3(2). Тыльная сторона ушей светлая, сероватая, цвета шеи и
спины. Конец хвоста темный. На передней стороне лап черных или тем-
но-серых пятен нет. Череп мелкий: общая длина его менее 16,5 см . 4
Рис. 37. Зубы спереди:
А — песец (Vulpes lagopus); Б — корсак (Vulpes corsac); В — лисицы (V. vulpes).
4(5). Спина рыжеватая или ржаво-серая с сильно развитой се-
ребристостью, но без примеси черных остевых волос. Нижняя губа и
подбородок белые. Хвост лишь немного длиннее V2 длины тела. Длина
тела более 50 см. Скуловая ширина более 55 мм......................
..............................Корсак. Vulpes corsac. (Стр. 127.)
5(4). Спина серая с примесью черных остевых волос. Нижняя губа
и подбородок темно-бурые. Хвост значительно длиннее V2 длины тела.
Длина тела менее 50 см. Скуловая ширина менее 55 мм................
...............• . . . Афганская лисица. Vulpes сапа. (Стр. 127.)
6(1). Уши закругленные, короткие, будучи пригнутыми к голове,
они не доходят своими вершинами до глаз. Подошвы лап сплошь по-
крыты мехом. Заглазничные отростки сверху плоские или слабовогну-
124
тые. При сомкнутых челюстях вершины нижних клыков лишь немного
заходят за края лунок верхних клыков (рис. 37, Л)......................
........................Песец, Vulpes (Alopex) lagopus. (Стр. 127.)
Подрод настоящих лисиц. Subgenus Vulpes Oken, 1816
4, Обыкновенная лисица, Vulpes vulpes Linnaeus, 1758
Табл. IX, 4—6. Карта 38
Евразия (кроме крайнего Юга) и вся Северная Америка. В СССР
встречается почти повсеместно, кроме Крайнего Севера.
Живет в самых различных условиях: в лесотундре, тайге, степях и
пустынях, на равнинах и высоко в горах.
Питается разнообразной, как животной, так и растительной, пи-
щей, но в основном мелкими грызунами, птицами и насекомыми. Течка
протекает в средней полосе в марте или начале апреля, на севере —
в апреле, на Кавказе — в феврале. Самки в это время «подлаивают»
самцов, между которыми из-за самок происходят сильные драки. Бере-
менность длится 52—56 дней. Детеныши (от 3 до 12, обычно 5—6) ро-
дятся в норе. Самец держится около выводка. К началу осени молодые
переходят к самостоятельному образу жизни. Размножаться начинают
иа первом году жизни. Весенняя линька протекает в средней полосе
в феврале—апреле (иногда затягиваясь до конца мая), а развитие
зимнего меха заканчивается только в конце ноября — начале декабря.
Лисица — очень важный объект промысловой и спортивной охоты.
Ежегодно в СССР добывается 500—700 тыс. лисиц. Серебристо-черную
и платиновую разновидности лисицы разводят в большом’ количестве
в звероводческих хозяйствах. Местами причиняет вред уничтожением
дичи. Полезна истреблением вредных грызунов.
Размеры и окраска меха, подвержены широкой индивидуальной и географической
изменчивости. Описано много подвидов, которые можно объединить в несколько групп.
I. Группа северных подвидов. Распространена в тундровой, лесотундровой и та-
ежной зонах. Размеры этих подвидов относительно крупные: кондилобазальная длина
черепа более 13 см; мех пышный, с яркой, насыщенной, рыжей окраской спины, белой
расцветкой груди и большими черными пятнами на ногах.
a) V.‘ v. vulpes Linnaeus, 1758. (Описан из Швеции.) Размеры крупные: кондило-
базальная длина черепа 14—15 см. Мех пышный, мягкий. Спина ярко-рыжая с более
темным крестом на плечах: белые части на остевых волосах развиты сильно. Брюхо
белое, реже черноватое. Грудь белая. Черные пятна на ногах невелики. Таежная
зона Северной Европы.
К этому подвиду относится natio V. v. v. crucigera Bechstein, 1801 (тип из Герма-
нии), с грубоватым мехом и кирпично-рыжей расцветкой спины, населяющая Прибал-
тику и Белоруссию. В центральных и западных областях РСФСР водится особая natio
с менее пышным, чем у номинальной формы, и чуть более светлым и тусклым мехом.
b) V. v. tobolica Ognev, 1926. (Тип из окрестностей Салехарда.) Размеры очень
крупные: кондилобазальная длина черепа 15—46 см. Мех зимней шкурки особо пыш-
ный, гривистый, мягкий. Окраска его неяркая, ржаво-желтая с буроватым крестом
на плечах. Низ сероватый или белый. На лапах сильно развиты черные пятна. Тайга
и лесотундра Западно-Сибирской низменности. На севере Приенисейской Сибири жи-
вет пока неописанная, но хорошо выделяющаяся форма. Размеры крупные, но мельче,
чем у тобольской. Мех пышный, шелковистый, но не гривистый. Окраска зимнего во-
лосяного покрова яркая, чистая, обычно светло-рыжая; брюхо белое. Лапы с большим
черным пятном.
с) V. v. jakutensis Ognev, 4923. (Тип из тайги к югу от Якутска.) Размеры, как
у североевропейской лисицы: кондилобазальная длина черепа обычно 14—15 см. Зим-
ний мех пышный, но не гривистый, особо шелковистый. Окраска его у большинства
особей темная, но не яркая, буровато- или желтовато-ржавая с темным крестом на
плечах. Брюхо темно-серое. Якутия.
d) V. v. beringiana Middendorf, 1875. (Описана с побережья Берингова пролива.)
Размеры очень крупные: кондилобазальная длина черепа 14—16 см. Мех очень пыш-
ный, но менее мягкий, чем у предыдущего подвида. У большинства особей в зимнем
мехе спина интенсивного огненно-рыжего цвета. Белые части иа остевых волосах
125
развиты слабо. Брюхо красноватое или белое. Встречаются особи и с более светлой ок-
раской, обусловленной сильным развитием белых частей остевых волос. Камчатка,
Анадырский край и Чукотский полуостров.
f) V. V. dolychocrania Ognev, 1926. (Тип из Сидеми, Южное Приморье.) Размеры
средние. Мех пышный, но малошелковистый. Окраска зимнего меха довольно яр-
кая, кирпично-красная с хорошо развитыми белыми частями остевых волос. Брюхо
красноватое или черноватое. Лапы с небольшим темным пятном. Приморье.
g) V. v. splendidissima Kishida, 1924. (Описан с Северных Курил.) Крупная ли-
сица с пышным, но малошелковистым мехом. Окраска меха темная, буровато-рыжая;
много особей с признаками меланизма. Курильские острова.
II. Степные подвиды. Населяют степную и лесостепную зоны нашей страны. Раз-
меры этих лисиц средние: кондилобазальная длина черепа обычно колеблется от 12
до 14 см. Мех средней высоты, густоты и мягкости. Окраска заметно светлее, чем
у лесных форм. Спина обычно светло-рыжего или желто-рыжего цвета, бока желто-
ватые, брюхо беловатое, с серым оттенком от просвечивающей голубоватой подпуши.
Темные пятна на лапах невелики и не черные, а темно-серые или черноватые.
h) V. Nr diluta Ognev, 1924. (Тип из Каменной степи Воронежской области.) Раз-
меры сравнительно крупные. Мех среднего качества. Окраска его в зимнее время на
плечах ржаво-желтая, на огузке темнее. Бока светло-желтые. Брюхо белесое. Лесо-
степная зона Европейской части СССР и Западной Сибири и Северный Казахстан.
i) Nr Nr stepensis Brauner, 1914. (Тип из окрестностей г, Херсона.) Мельче преды-
дущей расы: кондилобазальная длина черепа 12—14 см. Мех низкий, грубый. Окраска
тусклая, грязно-желтая или ржаво-желтая с сильным развитием черных волос на
загривке. Брюхо сероватое. Степи Юга Европейской части СССР. К этому же подвиду
относятся лисы гор Крыма, отличающиеся более яркой окраской меха спины (V. v.
st crimeamontana Brauner, 1914).
j) V. v. daurica Ognev, 1931. (Тип из района г. Троицкосавска.) Сравнительно
крупный подвид: кондилобазальная длина черепа 13—16 см. Мех пышный, но грубова-
тый. Окраска бледная, тусклая: спина палево-желтовато-ржавого цвета со светло-
рыжим крестом на плечах. Бока очень светлые, желтоватые. Брюхо беловатое. На
лапах лишь узкая светлая сероватая полоска. Забайкалье, Верхний Амур.
111. Пустынные подвиды. Населяют пустыни и полупустыни Прикаспийской низ-
менности, Казахстана и Средней Азии. Размеры мелкие и средние: кондилобазальная
длина черепа 12—14 см. Мех низкий, редкий, грубый. Окраска зимнего меха обычно
светло-желтая, серая или желто-серая. Брюхо грязно-белое. Грудь сероватая. На
лапках небольшие сероватые полоски.
k) V. v. karagan Erxleben, 1777, (Описана из степей Казахстана.) Размеры сред-
ние: кондилобазальная длина черепа до 14 см. Мех не пышный, грубоватый. Окраска
светло-желтая (песчаного цвета) или светло-серая. Грудь и брюхо сероватые. Полупу-
стыни и пустыни Нижнего Поволжья и Казахстана.
1) V. v, flavescens Gray, 1843. (Описана из Северного Ирана), Вероятно, к этому
подвиду относятся лисицы пустынь Средней Азии. Размеры мелкие: кондилобазальная
длина черепа до 13 см. Мех низкий, малошелковистый. Преобладают особи с тусклой
серо-желтой окраской спины и буроватым крестом на плечах. Бока сероватые. Брюхо
серое. Передняя сторона лап серая.
m) V. v. alpherakyi Satunin, 1905. (Тип из Муганской степи, Азербайджан.) Раз-
меры средние: кондилобазальная длина черепа до 13,5 см. Мех малопышный, грубова-
тый, Окраска спины варьирует от бледного палево-желтовато-ржавого до рыжевато-
серого и светло-рыжего цвета. Низ сероватый. Равнины Восточного Закавказья.
IV. Подвиды гор Кавказа. Лисицы среднего размера; кондилобазальная длина
черепа от 13 до 15 см. Мех грубоватый, малопышный. Окраска очень изменчива
(обычно грязно-рыжая, серая с красным ремнем по хребту или серая средней темноты).
n) V. v. caucasica Dinnik, 1914. (Тип из окрестностей г. Орджоникидзе.) Раз-
меры крупные: кондилобазальная длина черепа до 15 см. Мех средней пышности, но
грубоватый. Окраска различная: преобладают особи серого цвета с кирпично-красной
полосой по хребту. Попадаются также рыжие и серые особи. Северный Кавказ.
о) V, v. alticola Ognev, 1926. (Тип из окрестностей оз. Севан, Армения.) Размеры
средние: кондилобазальная длина черепа 12—'14 см. Мех сравнительно пышный и
мягкий. Окраска зимнего' меха светлая, блеклая, палево-желтая или палево-серая.
Бока бледно-серые или желтоватые. Брюхо грязно-белое. Нередки особи с признаками
меланизма. Армянское нагорье. По-видимому, особую расу образуют лисы Грузии,
отличающиеся грубоватым, малопышным мехом довольно темной, желтовато-ры-
жей окраски.
V. Подвиды гор Средней Азии. Лисы среднего и мелкого размера с волосяным
покровом средней мягкости и пышности. Окраска меха различная, но всегда светлая.
р) V. v. ochioxanta Ognev, 1926. (Тип с Аксая, в Семиречье.) ^Размеры, средние:
кондилобазальная длина черепа 14—15 см. Волосяной покров мягкий, средней высоты.
Окраска светлая, охристо-желтая или рыжевато-желтая. Низ тела беловатый. Горы
Тянь-Шаня.
126
5. Корсак. Vulpes corsac Linnaeus, 1768
Табл. IX, 7. Карта 39
Калмыцкие и Волжско-Уральские полупустыни, Казахстан, приле-
гающие к нему районы Западно-Сибирской низменности, равнины
Средней Азии (кроме Ферганы). Водится также в Северном Афгани-
стане, Джунгарии, Монголии и в Южном Забайкалье.
Типичный житель сухих степей, полупустынь, пустынь и сухих
предгорьев. Ведет преимущественно ночной образ жизни. Пища раз-
нообразная, но большую ее часть составляют мелкие грызуны, степные
птицы, пресмыкающиеся и насекомые. Живет в норах, обычно лишь
с одним входом. Охотно занимает норы сурков. Течка самок, во время
которой происходят драки самцов, протекает в конце января — в фев-
рале. Беременность около 55 дней. Молодые, числом обычно 2—6, ро-
дятся в конце марта — в апреле. К зиме выводки распадаются. Чис-
ленность сильно колеблется по годам. Промысловое значение невелико.
Географическая изменчивость значительная. По направлению с севера на юг
размеры корсаков мельчают, их мех становится менее пышным, а окраска — более
рыжеватой. Различают следующие подвиды:
а) V. с. corsac Linnaeus, 1768. (Описан из степей между Уралом и Иртышем.)
Размеры крупные: общая длина черепа до 125 мм. Мех пышный. Окраска серебристо-
серая со слабой примесью рыжих тонов. Тыльная сторона ушей серая. Большая часть
Казахстана. Очень близкую к номинальной форме V. с. scorodumovi Dorogostajski,
1935 (тип из Южного Забайкалья), с чуть более желтой окраской меха вряд ли сле-
дует обособлять в качестве подвида.
b) V. с. kalmykorum Ognev, 1935. (Тип из Калмыкии.) Размеры немного меньше:
общая длина черепа до 117 мм. Мех средней пышности. В окраске ясно выступают
рыжеватые тона. Бока желтые. Тыльная сторона ушей охристая. Калмыцкие и волго-
уральские степи.
с) V. с. turkmenicus Ognev, 1935. (Тип из Туркмении.) Размеры мелкие: общая
длина черепа до ПО мм. Мех низкий и грубый. Окраска темная с преобладанием
рыжих оттенков. Пустыни Средней Азии.
6. Афганская лисица. Vulpes сапа Blanford, 1877
Табл. IX, 8. Карта 39
Афганистан, Пакистан и Северо-Восточный Иран. В СССР най-
дена в Южной Туркмении. Один из наиболее редких зверей нашей
фауны. Образ жизни в Туркмении не изучен.
Подрод песцов. Subgenus A lop ex Каир, 1829
7. Песец. Vulpes lagopus Linnaeus, 1758
Табл. IX, 9, 10. Карта 39
Кругополярное животное, населяющее все материковые и остров-
ные берега Северного Ледовитого океана, лесотундры и тундры Ев-
ропы, Азии и Северной Америки. Встречается на Командорских и Але-
утских островах. По льдам доходит на север до 85° с. ш. Зимой значи-
тельная часть -песцов откочевывает из тундры к югу в лесотундру и
в тайгу, в Европейской части СССР — до 65° с. ш., а в Сибири—еще
южнее. Отдельные особи иногда заходят в Калининскую область, окре-
стности г. Сургута, верховья р. Вилюя.
Характерный обитатель арктических побережий тундры и лесо-
тундры. Охотится в любые часы суток. Питается разнообразной пищей,
в состав которой входят различные грызуны (особенно лемминги, се-
рые и лесные полевки), птицы и их яйца, падаль и дохлая рыба, раз-
личные морские беспозвоночные, выброшенные на берег морем; насе-
комые, ягоды, травы, водоросли. Течка самок вызывает сильные драки
127
самцов. Она происходит в марте или апреле. Продолжительность бере-
менности около 52—53 дней. Молодые, числом от 3 до 12, иногда и до
21, родятся в мае или июне. Гнездовая нора роется обычно на возвы-
шенном месте, иногда бывает очень глубокой и сложно устроенной,
имея много выходов и отнорков. В воспитании молодых принимают
участие оба родителя; самец кормит лактирующую самку. К осени мо-
лодые звери постепенно начинают вести самостоятельную жизнь. Зим-
ний период наиболее тяжел для песцов в силу трудностей добывания
пищи, что и заставляет их откочевывать к югу или уходить на льды
океана, где они сопровождают белых медведей, подбирая остатки их
пищи. Высокая плодовитость и массовые падежи из-за зимних голо-
довок и эпизоотий вызывают сильное колебание численности по годам.
Весной 'пышный зимний мех сменяется на низкий летний, а осенью
происходит смена летнего волосяного покрова на зимний. В нашей
фауне песец представлен двумя разновидностями (морфами):
1) белой, особи которой зимой окрашены в чисто-белый цвет, а ле-
том имеют темную окраску — буровато-серую на спине, желтоватую
на боках и беловатую на брюхе;
2) голубой, особи которой круглый год носят темно окрашенный
мех; расцветка его колеблется от светло-серой до черноватой или от
кофейной до темно-бурой, иногда с ясно выраженной серебристостью.
Очень важный объект пушного промысла Севера; за последние
годы в общесоюзных заготовках шкурки песца занимают четвертое
место, в годы «неурожая» отходя на 5—6-е место. Голубых песцов раз-
водят в звероводческих хозяйствах.
Различают 2 подвида песцов, обитающих в нашей стране:
а) V. 1. lagopus Linnaeus, 1758. (Описан с севера Европы.) Размеры сравнительно
мелкие: кондилобазальная длина черепа самцов 12—13 см. Мех мягкий, шелковистый.
Голубые особи составляют ничтожный процент популяции. Тундры Северной Европы
и Азии и острова Ледовитого океана.
b) V. 1. beringensis Merriam, 1902. (Тип с о-ва Беринга.) Размеры крупные: кон-
дилобазальная длина черепа 13—14 см. Волосяной покров грубоватый. Популяция
почти полностью состоит из особей голубой окраски. Командорские острова.
Указание ряда авторов, что на Новой Земле водится мелкий подвид V. 1. spitz-
bergensis, ни на чем не обосновано; там песец очень крупный.
РОД ЕНОТОВИДНЫЕ СОБАКИ. GENUS NYCTEREUTES Temminck, 1839
Зубная формула: I у С -J- Pm М • 2 = 42.
Всего один вид.
8. Енотовидная собака. Nyctereutes procyonoides Gray, 1834
Табл. X, 1, Карта 40
Юго-Восточная Азия; в СССР — Амуро-Уссурийский край. Заве-
зена во многие районы Европейской части СССР и Кавказа, где ныне
встречается почти повсеместно, кроме районов Севера и некоторых
южных областей. Опыты акклиматизации в Сибири и Средней Азии пе-
дали положительных результатов.
Излюбленные места обитания — низменности, богатые водой и ча-
стично заболоченные, покрытые лесом или кустарником, поймы рек, за-
росли камыша по озерам и рекам, смешанные и лиственные леса.
Норы роет в сухих буграх, в склонах оврагов, под кустарником. Нора
редко превышает в длину 3 м и обычно имеет только один вход. Часто
использует брошенные норы барсуков. Иногда на болотах устраивает
логово под корнями деревьев или под валежником. Пища разнооб-
128
разна. Истребляет большое количество мелких грызунов, лягушек,
моллюсков, насекомых. Осенью ест ягоды и плоды. Поедает также яйца
и птенцов птиц, чем местами может приносить значительный вред охот-
ничьему хозяйству. Впадает в зимний сон, который длится в средней
полосе с ноября по март. Но в оттепели нередко выходит из норы в по-
исках пищи. Иногда в одной норе залегает целый выводок. Течка —
в марте. Беременность 60—64 дня. Число щенят от 3 до 15 (обычно 5—
6). Прозревают на 10—15-й день. Самец активно участвует в выкарм-
ливании молодых. Половозрелость наступает в конце первого года
жизни. В Советском Союзе добывается в настоящее время ежегодно
около 50 тысяч.
На Дальнем Востоке обитает подвид N. р. ussuriensis Matschie, 1907. (Тип пз
района впадения Уссури в Амур).
Под влиянием жизни в новых условиях признаки енотовидных собак, акклима-
тизированных в Европейской части СССР, заметно изменились, что дало возможность
даже выделить особый подвид: N. р. kalininensis Sorokin, 1956, отличающийся более
мягким мехом и более серой окраской.
СЕМЕЙСТВО ЕНОТОВЫЕ. FAMILIA PROCYONIDAE
Среднего размера стопоходящие и полустопоходящие хищные мле-
копитающие с пятипалыми конечностями, причем третий палец длин-
нее остальных. Хвост обычно длинный.
Зубные формулы: I С -j- Pm уМ -2 = 40
3 1 3 2
или (у зарубежных форм) I — С —Pm -М —-2 = 36.
3 1 3 2
Хищнические зубы слабо отличаются от смежных с ними коренных
и предкоренных. Жевательная поверхность коренных тупо- или остро-
бугорчатая.
Распространены в Южной, Центральной и южной части Северной
Америки. Два рода и вида в Азии. Один вид рода Procyon ввезен
в СССР из Северной Америки с целью акклиматизации.
РОД ЕНОТЫ. GENUS PROCYON Storr, 1780
Средней величины стопоходящие хищники с пышным, довольно
длинным хвостом, в окраске которого чередуются светлые и темно-бу-
рые кольца. Общая окраска меха буроватая или буровато-серая. На
щеках черные пятна.
Зубная формула: I-|-С у PmМ-^-2 = 40.
Родина — Америка. В СССР акклиматизирован 1 вид.
9. Енот-полоскун. Procyon lotor Linnaeus, 1758
Табл. X, 2. Карта 40
Центральная и Северная Америка до Южной Канады включи-
тельно. В СССР завезен в Азербайджан, на Северный Кавказ, в Бе-
лоруссию, в Молдавию, на Тянь-Шань и Дальний Восток. Особенно ус-
пешно прошла акклиматизация в Азербайджане, где енот заселил уже
значительную территорию. Здесь он живет в низинных лесах по доли-
нам рек и среди рисовых полей, укрываясь на день в дуплах деревьев.
Активен ночью. На зиму впадает в непродолжительный, сон. .Питается
разнообразной, как животной (лягушки, яйца птиц и т. п.), так
9 Н. А. Бобринский и др.
129
и растительной, пищей (фрукты, ягоды, орехи, зерна и др.). Обычно вся-
кую пищу перед поеданием полощет в воде (отсюда название). Спари-
вается в феврале. Беременность около 63 дней. Молодые (от 2—6 до 8)
родятся в конце апреля — в мае; остаются при родителях до зимы,
а иногда и до весны следующего года.
Ценный пушной зверь. Работа по его расселению в СССР про-
должается.
СЕМЕЙСТВО МЕДВЕЖЬИ. FAMILIA URSIDAE
Крупные, тяжелого склада стопоходящие звери с очень коротким,
спрятанным в меху хвостом. Пятипалые ’.передние и задние ноги воору-
жены большими, но тупыми, не втяжными когтями. Уши небольшие,
закругленные. Коренные зубы вытянуты вдоль черепа, йх жевательная
поверхность покрыта многими мелкими бугорками. Три первых предко-
ренных очень малы и у старых особей нередко выпадают. Последний
предкоренной зуб верхней челюсти и первый истинно коренной нижней
челюсти малы и не носят характера хищнического зуба.
Зубная формула: I у С у Pm у М -|- • 2 = 42.
Распространены в Северной Африке (Марокко), Евразии, Север-
ной Америке, откуда по Андам спускаются в пределы Южной Америки.
В фауне СССР — 3 вида, относящиеся к трем различным родам.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ МЕДВЕДЕЙ ФАУНЫ СССР
1(4). Мех черный, бурый, коричневый или песочного цвета. Под-
пушь темная. Подошвы голые. Ширина костного нёба между перед-
ними коренными зубами меньше общей длины обоих коренных зубов
верхней челюсти (М*+М2)........................................2
2(3). Мех блестяще-черный. Натруди белое или желтое полулунное
пятно. Большие уши расширены раструбом. Лицевой отдел черепа от-
носительно короткий: расстояние между концом межчелюстных костей
до переднего края глазницы меньше расстояния между концами над-
глазничных отростков . . Черные медведи. Selenarctos. (Стр. 130.
3(2). Мех различной темноты и разных оттенков бурого цвета —
ог буровато-палевого до черно-бурого. На груди белого или желтого
полулунного пятна нет. Уши небольшие, округлые. Лицевой отдел че-
репа сравнительно длинный: расстояние от переднего конца межче-
люстных костей до переднего края глазниц больше расстояния между
концами заглазничных отростков . Бурые медведи. Ursus. (Стр. 131.)
4(1). Мех белый или желтовато-белый. Подпушь чисто-белая. По-
дошвы ног покрыты густой шерстью. Ширина костного нёба между пе-
редними коренными зубами больше длины обоих коренных зубов
(МЦМ2) . . Белые медведи. Thalassarctos. (Стр. 132.)
РОД ЧЕРНЫЕ МЕДВЕДИ. GENUS SELENARCTOS Heude, 1901
Один вид.
10. Черный медведь. Selenarctos tibetanus G. Cuvier, 1823
Табл. X, 3. Карта 41
От Афганистана через Северо-Западный Пакистан, Северную Ин-
дию, Непал, Бирму до Японии и Амуро-Уссурийского края включи-
тельно.
130
В Амуро-Уссурийском крае держится преимущественно в листвен-
ных и хвойно-лиственных лесах маньчжурского типа. Превосходно ла-
зает по деревьям. На зиму залегает в зимний сон обычно в дупле боль-
шого дерева. Течка — в мае—июне. 1—2 медвежонка родятся в январе
или феврале. Питается преимущественно растительной пищей (ягоды,
плоды, орехи, желуди и даже листья). Поедает также насекомых, мол-
люсков, червей, мелких зверьков, птенцов и яйца птиц. На крупных
зверей нападает редко.
Промысловое значение ничтожно.
В СССР особый подвид: S. t. ussuricus Heude, 1901. (Тип из Уссурийского края,,
без более точного указания). От типичной (гималайской) формы отличается большими
размерами и более пышным мехом.
РОД БУРЫЕ МЕДВЕДИ. GENUS URSUS Linnaeus, 1758
В Евразии 1 вид.
11. Бурый медведь. Ursus arctos Linnaeus, 1758
Табл. X, 4. Карта 41
Европа, горы Атласские в Северной Африке, большая часть Азии
(к югу до Гималаев) и Северная Америка. В СССР водится от лесо-
тундры по всей лесной полосе, до некоторых районов лесостепи вклю-
чительно. Летом заходит далеко в тундры. Встречается в горах Кав-
каза и Средней Азии.
Типичный обитатель больших лесов, хотя в горах Средней Азии
иногда населяет и безлесные хребты и плоскогорья. Всеяден: пищу его
составляют различной величины звери (от мыши до лося), птенцы и
яйца птиц, пресмыкающиеся, земноводные, рыбы, насекомые, черви,
моллюски, различная падаль, мед шчел, самые различные растения,
плоды, ягоды, семена. На зиму впадает в длительный сон, который про-
водит в берлоге. Берлога расположена обычно в труднодоступных и
скрытых местах — на гривах среди болот, в буреломе, густолесье,
в ущельях гор, в пещерах и т. п. Сама берлога в большинстве случаев
представляет собой просторную, но неглубокую яму у корней повален-
ного дерева, под валежиной или большим камнем, выстланную мохом,
травой, листьями. Снег заносит залегающего медведя, так что в снегу
остается лишь небольшое отверстие — «чело» берлоги. В Восточной
Сибири медведи залегают в берлогу обычно еще в октябре, в цент-
ральных областях — во второй половине ноября, а в западных районах
СССР — даже в декабре. В Северо-Восточной Сибири медведи про-
буждаются от зимнего сна в первой половине мая, в средней полосе
Европейской части СССР — в апреле. В низменных местах Закавказья
в теплые зимы большинство медведей остается активными в течение
круглого года. Течка проходит в средней полосе в июле, на Кавказе —
в мае. Беременность около 7 месяцев. Медвежата, обычно. 2, реже 1,.
иногда 3 и даже 4, родятся в берлоге с конца декабря до начала фев-
раля. Родятся они слепыми, беспомощными; вес новорожденного лишь
около 500 г. Прозревают в месячном возрасте. Ко времени выхода мед-
ведицы из берлоги медвежата вырастают настолько, что уже могут
следовать за матерью. Сосут мать около 4 месяцев, но держатся при
ней до зимы и обычно ложатся вместе с ней в берлогу на зимний сон
и даже следующее лето все еще не покидают ее. По-видимому, иногда
молодые медведи (пестуны) держатся три матери и третье лето своей
жизни, хотя у медведицы уже имеются детеныши следующего помета.
9*
1311
Половозрелой медведица становится на третьем году жизни, приносит
первых детенышей в четырехлетием возрасте. Щенятся медведицы нор-
мально через год. Самец в выкармливании и воспитании медвежат уча-
стия не’принимает и всегда ложится в отдельной берлоге.
Промысловое значение невелико как в силу невысокой числен-
ности, так и невысокой цены шкуры. Но эти огромные хищники служат
важным объектом спортивной охоты. Местами медведи приносят ощу-
тительный вред животноводству и охотничьему хозяйству.
Признаки медведей, особенно их размеры и окраска меха, подвер-
жены широкой индивидуальной и географической изменчивости.
Из большого числа описанных подвидов медведей реально различить можно сле-
дующие:
a) U. a. arctos Linnaeus, 1758. (Описан из Швеции.) Размеры средние: кондило-
базальная длина черепа самцов 30—35 см. Наиболее обычны особи с темно-бурой
окраской меха; изредка встречаются черно-бурые или светло-коричневые особи. Мех
мягкий и пышный. Леса Севера Европейской! части СССР и Сибири.
b) U. a. beringianus Middendorf, 1851. (Описан по экземплярам с побережья Бе-
рингова моря.) Размеры очень крупные: кондилобазальная длина черепа самцов от
35 до 40 см. Окраска очень изменчива: от светло-коричневой до черно-бурой; наибо-
лее обычны особи бурого цвета. Мех пышный и мягкий. Огромные когти темные.
Анадырский край, Камчатка, Охотское побережье. Особую natio этого подвида состав-
ляет U. a. mandchuricus Heude, 1898. (Тип с р. Уссури.) Размеры тоже очень крупные:
кондилобазальная длина черепа самцов 36—42 см. Окраска варьирует со светло-бурой
до черной. Череп более вытянутый. Приморье.
с) U. a. isabellinus Horsfield, 1826. (Тип из Непала.) Размеры средние. Окраска
меха светлая: буровато-палевая или светло-бурая. Мех пышный, лохматый. Когти
светлые. Горы Средней Азии.
d) U. a. caucasicus Smirnov, 1916. (Тип с Пассанаур на Кавказе.) Размеры сред-
ние: кондилобазальная длина черепа самцов 29—35 см. Окраска светлая, варьирую-
щая от палево-серой до светло-рыжевато-бурой. Мех малопышный, грубоватый. Когти
темные. Кавказ и Закавказье, кроме юго-западных районов.
е) U. a. syriacus Hemprich et Ehrenberg, 1828. (Описан из Сирии.) Близок к пре-
дыдущей форме. Размеры мелкие. Окраска меха очень светлая: серо-палевая или
светло-серая с темным пятном на загривке. Мех малопышный. Когти светло-бурые
с темными концами. Юго-западные районы Закавказья.
РОД БЕЛЫЕ МЕДВЕДИ. GENUS THALASSARCTOS 1 Gray. 1825
Один вид.
12. Белый медведь. Thalassarctos maritimus Phipps, 1774
Табл. X, 5. Карта 41
Область плавучих льдов Северного Ледовитого океана, его острова,
азиатское и американское побережья Крайнего Севера. У европейских
берегов Арктики появляется только случайно. Иногда заносится на
.льдинах далеко к югу (до берегов Камчатки и побережья Охотского
моря). Основное место размножения — о. Врангеля.
Жизнь белого медведя связана с морем и плавучими льдами.
В глубину тундры редко заходит более чем на 1—2 км. Превосходно
плавает, иногда заплывая на 20 км и более от ближайшего берега или
.ледяных полей. Основная пища — тюлени; ловит также рыбу, поедает
разных морских животных, выброшенных волнами, охотится за моло-
дыми моржами. В берлогу на зиму ложатся только беременные самки.
Здесь они после 7-месячной беременности приносят в феврале —
марте 1, реже 2 медвежат. Берлога устраивается обычно на берегу, под
скалой или обрывом, изредка среди торосов. Ранней весной медведица
1 По Эллерману и Моррисон-Скотту (1951) —Thalarctos.
132
с медвежатами выходит из берлоги и ведет с ними бродячую жизнь,
кочуя по льдам и островам. Молодые держатся при матери обычно не
более года. Самки приносят детенышей через год. Самцы и холостые
самки весь год кочуют по льдам и побережьям Арктики.
Добывают медведей в небольшом количестве; их шкуры исполь-
зуют на ковры. Ценный объект зооэкспорта.
Описаны 2 слабо различающиеся формы:
а) Т. m. maritimus Phipps, 1774. (Описана с о. Шпицбергена.) Размеры более
мелкие: кондилобазальная длина черепа самцов 35—37 см. Лобная площадка черепа
слабо приподнята. Западная часть Северного Ледовитого океана.
b) Т. m. marinus Pallas, 1776. (Описана с севера Сибири.) Размеры немного круп-
нее: кондилобазальная длина черепа самцов 36—39 см. Череп с более приподнятой
лобной площадкой. Восточная часть Арктики.
СЕМЕЙСТВО ГИЕНОВЫЕ. FAMILIA HYAENIDAE
Наземные пальцеходящие хищники собачьего склада с высокими
четырехпалыми конечностями и притупленными невтяжными когтями.
Крестец располагается ниже уровня лопаток. Основная пища — трупы
крупных млекопитающих. Череп с мощными челюстями и большим са-
гиттальным гребнем. Верхний хищнический зуб очень большой, а рас-
положенный за ним единственный коренной зуб маленький.
Распространены по Африке, Передней и Южной Азии.
3 рода (с 4 видами). В СССР — 1 род.
РОД ГИЕНЫ. GENUS HYAENA Brisson, 1762
В области лопаток — грива из удлиненных волос. Окраска меха
на боках с темными поперечными полосами.
Зубная формула: I у С уРш уМу-2 = 34.-
13. Полосатая гиена. Hyaena hyaena Linnaeus, 1758
Табл. X, 6J. Карта 42
Африка, Аравия, страны Ближнего Востока, Иран, Афганистан,
Пакистан, Индия; в СССР — Закавказье и южные части Средней Азии.
В Закавказье в небольшом количестве населяет низменные районы и
предгорья Азербайджана и Восточной Грузии. В Средней Азии гиены
редки в южных частях Узбекистана, Таджикистане и Туркмении.
Излюбленные места обитания — пустынные предгорья с ущельями
и пещерами, глинистая пустыня с оврагами, тугаи по большим рекам.
Активны ночью. Основная пища — различная падаль, но иногда гиены
нападают и на животных, в том числе на мелкий домашний скот. Ло-
гово устраивают в пещерах под нависшими камнями, реже — в неглубо-
кой норе. Течка — в январе — феврале, рождение молодых — в ап-
реле - -- мае. Беременность около 3 месяцев. Число детенышей в по-
мете— 2—4. Редкий зверь. Промыслового значения не имеет. Ценный
объект зооэкспорта.
Полосатые гиены СССР относятся к номинальной форме: Н. h. hyaena L.
СЕМЕЙСТВО КОШАЧЬИ. FAMILIA FELIDAE
Средних и крупных размеров наземные пальцеходящие хищники
с пятипалыми передними и четырехпалыми задними конечностями.
Из всех хищных наиболее специализированы и приспособлены
133
К добыванию и питанию живой добычей — другими млекопитающими
и птицами. Когти (кроме одного вида — гепарда) втяжные, изогнутые,
-сжатые с боков, очень острые. С каждой стороны верхней и нижней
челюстей не более 3 предкоренных и 1 коренного зуба. Первые предко-
ренные обеих челюстей и задние коренные верхней челюсти развиты
очень слабо и нередко могут отсутствовать. Общее число зубов варьи-
рует от 26 до 32. Хищнические зубы развиты сильно.
Широко распространены по всем материкам (кроме Австралии п
Антарктики) и многим материковым островам.
В фауне СССР — 3 рода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ СЕМЕЙСТВА КОШАЧЬИХ
ФАУНЫ СССР
Рис. 38. Глазница (вид сверху):
А — леопард (Panthera pardus); Б — рысь (Felis lynx); pz —
передний отросток скуловой кости; fl — слезное отверстие;
РР — надглазничный отросток.
1(2). Конечности длинные и тонкие, «собачьего» типа, с притуплен-
ными невтяжными когтями. По верхней стороне шеи идет грива из
удлиненных грубых волос. Подглазничное отверстие черепа мало и вытя-
нуто в вертикальном на-
правлении: его диаметр
в 2 раза меньше рассто-
яния между верхним
краем этого отверстия н
глазницей. . . Гепарды.
Acinonyx. (Стр. 134).
2(1). Конечности срав-
нительно короткие и мощ-
ные, со вполне втяжными,
очень острыми когтями.
Гривы на верхней стороне
шеи нет. Подглазничное
отверстие черепа боль-
шое: его диаметр равен
или превышает расстоя-
ние между верхним кра-
ем этого отверстия п
глазницей............3
3(4). Размеры круп-
ные: длина тела взрослых
зверей более 120 см. Дли-
на хвоста значительно
превышает половину длины тела. Кондилобазальная длина черепа
взрослых более 16 см. Верхне-внутренний отросток скуловой кости не
доходит или только достигает внешнего края слезного отверстия
(рис. 38, А)........................Пантеры. Panthera. (Стр. 135.)
4(3). Размеры более мелкие: длина тела до 120 см. Длина хвоста
примерно равна или меньше половины длины тела. Кондилобазальная
длина черепа менее 16 см. Верхне-внутренний отросток скуловой кости
заходит за внешний край -слезного отверстия (рис. 38, Б).........
............... . . . . . Кошки. Felis. (Стр. 137.)
РОД ГЕПАРДЫ. GENUS ACINONYX Brookes, 1828
Своеобразные крупные кошачьи, со стройным легким телом, длин-
ными, тонкими ногами, пальцы которых несут невтяжные, туповатые
когти. Хвост длинный, тонкий. По верху шеи идет невысокая, но густая
134
стоячая грива. Пятна небольшие, резко выделяющиеся па рыжеватом
фоне.
Зубная формула: I у С -j- Pm у М у • 2 = 30.
Вся Африка и Юго-Западная Азия, от Сирии до Индии включи-
тельно.
14. Гепард. Acinonyx jubatus Schreber, 1776
Табл. XI, /. Карта 42
Широко распространен по степям и пустыням Африки и Юго-За-
падной Азии. В СССР найден только в Южной Туркмении по р. Ат-
реку, в Копет-Даге, в долинах Мургаба и Теджена, в районе г. Кушки.
Заходит на Устюрт и Мангышлак. Очень редкий зверь нашей фауны.
Обитает в равнинных или слегка всхолмленных глинистых и песча-
ных пустынях. Питается джейранами, зайцами, мелкими грызунами и
птицами. Беременность продолжается около 95 дней. Число детены-
шей — от 2 до 4. Деторождение происходит весной.
Гепарды Туркмении относятся к подвиду A. j. venaticus Griffith,
1821 (описан из Индии).
РОД ПАНТЕРЫ. GENUS PANTHERA Oken, 1816
Охватывает ряд наиболее крупных видов семейства кошачьих, от-
личающихся длинным хвостом и особенностями черепа, указанными
в определенной таблице. Обитатели преимущественно тропических
стран. В Советском Союзе водятся только в южных районах.
В СССР обитает 3 вида этого рода, относимые к разным подродам.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА ПАНТЕР ФАУНЫ СССР
1(4). Тело покрыто темными пятнами, иногда расплывающимися
в темный мраморный рисунок. Длина тела взрослых до 160 см. Общая
длина черепа до 26 см. Расстояние между передними наружными кон-
цами носовых костей всегда меньше ширины черепа позади заглазнич-
ных отростков................................................2
2(3). Пятна на спине большие (более 5 см в поперечнике), неясно
очерченные, расплывчатые, бледные. Хвост очень пушистый: его тол-
щина примерно равна толщине средней части предплечья. Общая
длина черепа обычно менее 19 см. Лобная площадка черепа резко вы-
пуклая с заметной впадиной посередине.............. ...
...............................Барс. Panthera uncia. (Стр. 136.)
3(2). Пятна на спине сравнительно мелкие (менее 5 см в попереч-
нике), обыкновенно резко очерченные, яркие. Хвост малопушистый, по-
крытый коротким мехом: его толщина значительно меньше толщины
средней части предплечья. Общая длина черепа взрослых обычно
больше 19 см. Лобная площадка черепа слабо и полого выпуклая, упло-
щенная, с едва заметной впадиной посередине ...................
...........................Леопард.. Panthera pardus. (Стр. 136.)
4(1). Тело покрыто черными или черноватыми поперечными поло-
сами. Длина тела взрослых животных более 160 см. Общая длина че-
репа более 26 см. Расстояние между передними наружными концами
носовых костей равно или превышает ширину черепа позади заглазнич-
ных отростков . ............Тигр. Panthera tigris. (Стр. 136.)
135
Подрод барсов. Subgenus Uncia. Gray, 1854
15. Барс. Panthera uncia Schreber, 1778
Табл. XI, 3. Карта 42
Горы Центральной Азии, от Алтая до Гималаев и от Тянь-Шаня
до Восточного Тибета. В Советском Союзе встречается на Памире, на
Гиссаро-Алае, на хребтах Тянь-Шаня, на Джунгарском Алатау, Тарба-
гатае, в Саурах и изредка на Алтае.
Высокогорное животное; лишь зимой спускается несколько ниже..
Активен преимущественно в ночное и сумеречное время. Основная
пища — дикие копытные (главным о'бразом горные козлы и бараны),
грызуны (особенно сурки) и птицы. Иногда нападает и на мелкий скот,
выгоняемый на горные пастбища. Течка — в конце зимы. Беременность
около 90 дней. Котята (обычно 2, реже 1 или 3) родятся во второй по-
ловине мая или в июне. Гнездо барс устраивает в пещере или под на-
висшей скалой.
Шкуры ценятся как пушнина, но промысловое значение барса не-
велико. Ценный объект зооэкспорта. Местами приносит вред животно-
водству.
Географические формы не описаны.
Подрод леопардов. Subgenus Panthera Oken, 1816
16. Леопард. Panthera pardus Linnaeus, 1758
Табл. XI, 4. Карта 42
Почти вся Африка, Южная и Восточная Азия. В СССР встре-
чается, с одной стороны, на Дальнем Востоке (к северу до 50° с. ш.)„
с другой — в горах Туркмении и на Кавказе.
Держится преимущественно в горных лесах, но иногда на безлес-
ных горных хребтах и в равнинных лесах. В горах поднимается до
3500 м. Хорошо лазает по деревьям. Предпочитает охотиться ночью.
Основная пища — дикие копытные, зайцы и другие грызуны, а также
птицы. Иногда нападает на мелкий скот. Во время размножения ведет
одиночную кочевую жизнь. Гнездо для рождения детенышей самка
устраивает в глухих местах, в пещере, под скалой или просто в чаще
леса. Течка — в конце зимы. Беременность — от 3 до 31/3 месяцев. Чи-
сло детенышей — 2—5.
В силу своей малочисленности добывается в количестве лишь не-
скольких десятков.
В СССР встречается 2 подвида:
а) Р. р. tulliana Valenciennes, 1856. (Описан из окрестности Смирны, Малая
Азия.) Основной фон окраски бледный, тускло-желтый, пятна буровато-черные. Кон-
дилобазальная длина черепа 20—22 см. Северный Кавказ и Закавказье. Р. р. ciscau-
casica Satunin, 1914,— синоним этого подвида.
b) Р. р. orientalis Schlegel, 1857. (Описан из Кореи.) Основной фон окраски яркий,
рыжеватый, пятна черные. Кондилобазальная длина черепа 18—20 см. Южная часть
Дальнего Востока.
Подрод тигров. Subgenus Tigris Oken, 1816
17. Тигр. Panthera tigris Linnaeus, 1758
Табл. XI, 2. Карта 43
Южная и Восточная Азия. В Советском Союзе встречается, с од-
ной стороны, на юге Дальнего Востока (на Сихоте-Алине и по сред-
нему течению Амура), с другой — местами в Средней Азии; иногда
136
заходит в Закавказье. В Средней Азии прежде тигры были распро-
странены широко, но ныне сохранились главным образом по долине
р. Аму-Дарьи и в низовьях Пянджа и Вахша. В Закавказье изредка
встречается только на Талыше, заходя из Ирана.
В Средней Азии тигры держатся главным образом в тугаях —
густых зарослях кустов и тростника по долинам рек. На Дальнем Во-
стоке они живут в горных лесах с ущельями и скалами. Вне периода
размножения тигры кочуют в поисках пищи, состоящей в основном из
диких копытных: кабанов, оленей, косуль и др. Иногда нападают на
домашний скот. Течка — в конце декабря, в январе. Продолжитель-
ность беременности — 98—НО дней. Гнездо, где родятся детеныши (2—
6 в помете), устраивается в глухом месте прямо на земле или под
скалой; оно иногда бывает выстлано травой и сухим листом. Молодые
сосут мать около полугода, но держатся около нее до двух лет и даже
дольше. Половозрелости достигают к пяти годам. Самка приносит де-
тенышей раз в три года.
В СССР ежегодно добывается по нескольку десятков тигров, пре-
имущественно живых (для зоопарков и зоосадов).
Географическая изменчивость тигров, выражающаяся главным образом в раз-
личиях интенсивности окраски и рисунка черных полос, довольно значительна. На
территории СССР различают 3 подвида:
а) Р. t. virgata liliger, 1815. (Описан из Северного Ирана.) Мех даже зимой
малопышный. Окраска зимнего меха интенсивная, с частыми черными полосами. Сред-
няя Азия, Закавказье.
b) Р. t. coreensis Brass, 1904. (Описан из Кореи.) Мех средней пышности. Окраска
его менее интенсивная. Полосы на теле уже и расположены реже, чем у предыду-
щего подвида. Размеры меньше, чем у амурских тигров. Приморье.
с) Р. t. longipilis Fitzinger, 1868. (Описан из Амурской области.) Мех- зимой
особо пышный. Окраска его еще более светлая, полосы шире, чем у предыдущего под-
вида. Размеры более крупные. Амурский край.
РОД КОШКИ. GENUS FELIS Linnaeus, 1758
Объединяет большое количество (более 60) видов, отличающихся
небольшими или средними размерами, относительно коротким хвостом
и отмеченными в таблице особенностями черепа.
Зубная формула: I у С-5- Рш|^М 5^1-2 = 30 или 28, реже 32 или 26.
Вся Евразия к северу до северного предела лесов., Африка, Се-
верная Америка к югу от тундры и вся Южная Америка.
В СССР обитает 8 видов этого рода, относящихся к 2 подродам.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА КОШЕК ФАУНЫ СССР
1(14). Размеры относительно небольшие: длина тела до 90 см.
Хвост длинный: его длина значительно больше длины головы; конец
его выдается за пальцы вытянутых назад ног. Только самый кон-
чик хвоста иногда бывает черным. Кондилобазальная длина черепа
менее 120 мм. Предглазничные отверстия вытянуты в .косом направ-
лении .............................................................2
2 (13). Тыльная сторона ушей не черная или черная только на вер-
шине. Над верхней губой по бокам головы черных пятен нет. Ушные
раковины без кисточки волос на концах или с небольшими кисточками,
длина волос которых не превышает 3 см. Надглазничные отростки че-
репа длинные, тонкие, заостренные; они настолько сближены с верх-
ними отростками скуловых костей, что расстояния между ними меньше
длины самих заглазничных отростков............................ . . 3
137
3(12). Длина хвоста (измеренная с концевыми волосами) при-
мерно равна У2 длины тела. На ушах кисточек из удлиненных волос
нет. Волосяной покров мягкий. Кондилобазальная длина черепа менее
104 мм..........................................................4
4(9). Тело покрыто ясным рисунком из темных пятен или полос
или вдоль хребта проходит двойная темная полоса. Барабанные ка-
меры черепа сравнительно невелики: их передние концы едва доходят
до заднего края сочленовной ямки................................5
5(8). Спина с многочисленными черноватыми пятнами или на бо-
ках выступают ясные черные полосы и вдоль хребта тянется двойная
темная полоса. По верху головы проходит 4 продольные темные по-
лоски. Резко выраженных светлых полос, идущих от носа вверх к те-
мени, нет. На груди поперечных полос нет. Костное нёбо короткое,
обычно не выступающее за задний край рядов зубов или продолжаю-
щееся назад не более чем на V2 ширины задненёбной выемки ... 6
6(7). Вдоль хребта проходит двойная черноватая полоса. На бо-
ках выступает 8—12 темных поперечных полос. Общий тон окраски
спины буровато-серый. Конец хвоста толстый, как бы обрубленный.
Наружный слуховой проход узкий: его наибольший диаметр меньше
длины коронки третьего предкоренного зуба верхней челюсти (лежа-
щего перед хищническим зубом). Концы носовых костей, как правило,
заходят назад за концы верхнечелюстных костей......................
.........................Лесная кошка. Felis silvestris. (Стр. 139.)
7(6). Спина и бока усеяны округлыми черноватыми пятнами и
небольшими полосками. Общий тон зимнего меха светлый, палево-
серый. Конец хвоста заостренный. Наружный слуховой проход более
широкий: его наибольший диаметр больше длины коронки третьего
предкоренного зуба верхней челюсти. Концы носовых костей не про-
стираются дальше назад, чем концы верхнечелюстных костей ....
.........................Степная кошка. Felis libyca. (Стр. 139.)
8(5). Все тело и хвост равномерно покрыты частыми ржаво-
бурыми пятнами. От носа вверх, к темени, тянутся две резко очерчен-
ные полоски. На груди выделяются темные поперечные полосы. Кост-
ное нёбо длинное, продолжающееся за концы зубных рядов на рас-
стояние, равное или даже превышающее !/2 ширины задненёбной
вырезки .... Бенгальская кошка. Felis bengalensus. (Стр. 140.)
9(4). Окраска спины и боков одноцветная (без рисунка из темных
пятен или полос), с легкой рябью или иногда с чуть заметными уз-
кими, темными, поперечными полосками в задней части тела. Барабан-
ные’ камеры черепа крупные: их передние концы заходят за передний
край сочленовных ямок................................ ... 10
10(11). На хвосте темных колец нет. Уши большие, заостренные,
выдающиеся из меха. Подошвы лап покрыты густым мехом. Баков по
бокам головы нет. Наименьшее расстояние между барабанными каме-
рами значительно меньше ширины задненёбной вырезки. Внутренняя
лопасть верхнего хищнического зуба развита хорошо..................
............. . . Барханная кошка. Felis margarita. (Стр. 140.)
11(10). На хвосте имеются ясные черные кольца. Уши небольшие,
округлые, слабо выдающиеся из меха. Подошвы лап голые. По бокам
головы удлиненные волосы образуют баки. Наименьшее расстояние
между барабанными камерами равно или лишь немного уступает ши-
рине задненёбной вырезки. Внутренняя лопасть верхнего хищнического
зуба едва заметна...................Манул. Felis manul. (Стр. 140.)
12(3). Хвост относительно короткий: длина его с концевыми во-
лосами приблизительно равна !/з длины тела. На ушах имеются не-
138
большие кисточки из удлиненных волос. Волосяной покров грубый.
Кондилобазальная длина черепа взрослых особей более 104 мм . . .
......................................Хаус. Felis chaus. (Стр. 141.)
13(2). Тыльная сторона ушей черная. По бокам головы над верх-
ней губой расположены черные пятна. На ушных раковинах большие
кисточки волос, длина которых превышает 3 см. Надглазничные от-
ростки относительно короткие и тупые: их длина меньше расстояния
от их концов до вершин отростков скуловых костей....................
....................................Каракал. Caracal. (Стр. 141.)
14(1). Размеры более крупные: длина тела взрослых особей более
90 см. Хвост короткий: его длина меньше длины головы, конец его не
достигает концов вытянутых назад задних ног. Последняя треть хво-
ста черная. Кондилобазальная длина черепа взрослых более 120 мм.
Предглазничные отверстия вытянуты почти вертикально.................
- . . . Рысь. Felis lynx. (Стр. 141.)
Подрод настоящих кошек. Subgenus Felis Linnaeus, 1758
18. Лесная кошка. Felis silvestris Schreber, 1777
Табл. XI, 5. Карта 43
Почти вся Западная Европа (во многих местах истреблена); Ма-
лая и Передняя Азия, Кавказ. В СССР — в Молдавии и западных об-
ластях Украины (прежде была в BGCP и Литве), в Закавказье и на
Северном Кавказе до нижнего течения Кубани, ставропольских остров-
ных лесов и долины Нижнего Терека.
Характерные места обитания — высокоствольные густые горные и
пойменные леса. По склонам гор поднимается до 2000 ж, но более
обычна в среднем поясе гор (900—1200 м над у. м.). Вне времени раз-
множения ведет одиночную кочевую жизнь, скрываясь на день в тре-
щинах скал, в дуплах, в заброшенных норах или в густой кроне де-
ревьев. Основная пйща— птицы и мелкие зверьки. Течка, сопровож-
даемая драками самцов, происходит на Кавказе в середине февраля —
начале марта. Беременность около 9 недель. Котята (2—6, обычно 4—5)
родятся в апреле или мае. Прозревают через 9—12 дней. Гнездо устра-
ивает в дуплах деревьев, в расселинах скал, в старых чужих норах.
Промысловое значение ничтожно. Приносит вред истреблением
пернатой дичи, зайцев и новорожденных косуль.
В фауне СССР 2 подвида:
a) F. s. silvestris Schreber, 1777. (Описан по экземпляру из Германии.) Карпаты,
Молдавия.
b) F. s. caucasica Satunin, 1905. (Боржоми.) Отличается от типичной формы не-
сколько более крупными размерами тела и черепа. Кавказ.
19. Степная кошка. Felis libyca Forster, 1780
Табл. XI, 6. Карта 43
Почти вся Африка, Аравия, Передняя Азия, Иран, Ирак, Афга-
нистан, Пакистан, некоторые северные области Индии. В СССР —
Восточное Закавказье (в Прикуринских полупустынях Азербайджана
и по долине р. Аракса — в Нахичевани и в Армении) и равнинные об-
ласти Средней Азии и Казахстана на север до волго-уральских полу-
пустынь, аральских Каракумов и котловин оз. Зайсан и оз. Иссык-
Куль.
139
Чаще всего держится в тугаях по долинам рек и в зарослях ку-
стов гребенчука среди пустыни. Поднимается по безлесным склонам
и долинам рек в горы до высоты 1500—2000 м. Основная пища — мелкие
и средних размеров звери и птицы, в частности уничтожает много фа-
занов. Течка — в феврале. Котята родятся обычно в конце апреля —
в мае; число их от 2 до 10. Гнездо помещается чаще всего в норе.
Ежегодно в нашей стране добывается около 10—12 тысяч степных
кошек. Ценность их шкурок невелика.
Описано несколько подвидов; большая часть из них описана по 1—2 шкуркам,
хотя индивидуальная изменчивость окраски особей этого вида огромна. Поэтому ре-
альность этих форм вызывает большие сомнения. Вероятно, в Советском Союзе встре-
чается только один подвид: F. libyca caudata Gray, 1874.
20. Бенгальская кошка. Felis bengalensis Kerr, 1792
Табл. XI, 7. Карта 44
Широко распространена по странам Южной и Юго-Восточной
Азии, в советском Приморье и по р. Амуру.
В пределах советского Дальнего Востока населяет смешанные и
широколиственные леса. Ночью охотится, а на день скрывается в рас-
селинах скал, под корнями деревьев, в дуплах, среди бурелома. Пи-
тается преимущественно мелкими грызунами, птицами, их птенцами
и яйцами. Ведет скрытную одиночную жизнь и только во время гона
держится парами. Течка происходит в марте. Беременность около
2 месяцев. Котята родятся- в мае. Гнезда устраивает среди камней,
в скалах, в дуплах.
Промысловое значение ничтожно.
В Советском Союзе встречается F. b. chinensis Gray, 1837 (Квантуй), которая
отличается от номинальной формы относительно тусклой окраской меха.
21. Барханная кошка. Felis margarita Loche, 1858
Табл. XI, 8. Карта 44
Северная Африка, Аравия и Средняя Азия. В пределах последней
обитает в пустынях Каракум и Кызылкум. Держится среди барханных
песков. Активна ночью. Днем скрывается в неглубокой норе, вырытой
в песке у корней саксаула ил'и других пустынных кустарников. Пита-
ется преимущественно пустынными грызунами (песчанками, тушканчи-
ками 'и др.). ААожет долго обходиться без воды. Котята (2—4) появля-
ются в начале апреля. К осени выводки распадаются.
Промысловое значение незначительное.
В пустынях Средней Азии обитает F. m. thinobius Ognev, 1926. (Тип из Репетека,
Восточные Каракумы.) От номинальной формы из Сахары отличается более крупными
размерами и некоторыми другими признаками.
22. Манул. Felis manul Pallas, 1776
Табл. XII, 1. Карта 44
Центральная и Средняя Азия, Иран. В СССР встречается в сте-
пях Забайкалья, в Туве, в горных степях Алтая, в Казахстане (кроме
его северных частей), в пустынях Средней Азии и в Армении (на
границе с Нахичеванью). Населяет степи и пустыни, предпочитая
держаться среди каменистых нагорий, пустынных горных гряд и мел-
140
косопочника. Живет в расселинах скал, среди камней, в норах, бро-
шенных другими животными. Охотится преимущественно ночью. Пита-
ется почти исключительно грызунами. Биология размножения изучена
плохо.
Промысловое значение невелико.
В СССР, видимо, 2 формы:
a) F. m. manul Pallas, '1776. (Тип с р. Джиды, Забайкалье.)
b) F. m. ferrugineus Ognev, 1928. (Описана по экземпляру из Туркмении.) Хроми-
стическая форма.
23. Хаус. Камышовый кот. Felis chaus Giildenstaedt, 1776
Табл. XII, 2, Карта 45
Вся Южная Азия, Афганистан, Иран, Средняя Азия, страны Ближ-
него Востока, большая часть Африки. В Советском Союзе обитает по
западному берегу Каспия на север до дельты Волги, в низменных ча-
стях Восточного Закавказья, в горах Копет-Дага, в Мур га беком базисе,
по Вахшу, Пянджу, Аму-Дарье и, по-видимому, по нижнему течению
Сыр-Дарьи и Чу.
Излюбленные места обитания — густые заросли кустарника, трост-
ника и камыша по берегам моря, рек и озер. Питается главным обра-
зом болотными и водными птицами, фазанами и другими обитателями
тугаев. Течка проходит в Закавказье в феврале. Котята (от 2 до 10)
родятся в мае или июне. Гнездо помещается или прямо на земле
в зарослях, или в брошенных норах дикобраза, барсука или других
зверей.
Значение как пушного зверя мало. Приносит вред охотничьему
хозяйству.
В СССР обитает номинальная форма: F. ch. chaus Giildenstaedt, 1776, описанная
по экземпляру из долины’ Терека.
Подрод каракалов. Subgenus Caracal Gray, 1843
24. Каракал. Felis caracal Schreber, 1776
Табл. XII, 3. Карта 45
Африка, Аравия, Сирия, Палестина, Ирак, Иран, Средняя Азия,
Афганистан, Пакистан и Индия. В нашей стране встречается только
в пустынях Туркмении, к северу до Аральского моря. Всюду редок.
Обитает в песчаных пустынях и в предгорьях. Питается зайцами,
сусликами, песчанками и другими грызунами. Биология не изучена.
Промыслового значения не имеет; ценность шкуры невелика.
В СССР — особый подвид: F. с. michaelis Heptner, 1945.
Подрод рысей. Subgenus Lynx Kerr, 1792
25. Рысь. Felis lynx Linnaeus, 1758
Табл. XII, 4. Карта 45
Лесные и горные области Европы, Азии (на юг до Гималаев) и
Северной Америки. В Советском Союзе распространена почти по всей
лесной зоне тайги, в смешанных и широколиственных лесах, откуда
заходит в лесотундру и лесостепь. На Камчатке нет. Южная граница
ареала идет от Карпат по южным окраинам Полесья через Брянские
леса, Рязанскую Мещеру, леса Мордовии, Татарию, Южный Урал,
141
боры и колки Барабинской лесостепи, Алтай, Северную Монголию,
Забайкалье, Приморье. Известны забеги далеко к югу от этой линии.
Кроме того, рыси встречаются в горах Кавказа, Закавказья и Средней
Азии.
Типичный обитатель глухих лесов, но в горах иногда встречается
и на безлесных склонах, среди скал и осыпей. Поднимается во многих
горных районах до 3000 м над у. м. Ведет обычно кочевой образ жизни
в пределах определенного охотничьего участка. Но иногда предприни-
мает далекие миграции, заходя даже в степь. Охотится обычно в су-
мерках и ночью, скрываясь на день в буреломе и валежнике, в скалах
и среди камней, в чаще пли в густой кроне дерева. Хорошо лазает по
деревьям. Вне периода размножения держится либо поодиночке, либо
стайками до 6 зверей (вероятно, семьями, не распавшимися осенью).
Основная пища — зайцы, лесные птицы, мышевидные грызуны. Реша-
ется нападать и на более крупных животных — оленей, косуль, моло-
дых лосей и др. Течка, сопровождаемая жестокими драками самцов,
на Кавказе бывает в феврале, в средней полосе — в марте и начале
апреля, в северной Сибири — в апреле. Беременность — 9—10 недель.
В выводке обычно 2—3, редко до 5 котят. Прозревают они на 12-й
день. В выкармливании и воспитании молодых принимают участие,
видимо, оба родителя. Выводки держатся до февраля, а то и дольше.
Половой зрелести достигают в двухлетнем возрасте.
Промысловое значение рыси невелико. Шкуры ее весьма ценятся
на мировом рынке. Приносит вред истреблением ценных охотничьих
животных.
Окраска рысей, как основной тон, так и рисунок пз пятен, сильно варьирует
в зависимости от района их обитания. Это позволяет выделить в пределах СССР
4 подвида:
a) F. 1. lynx Linnaeus, 1758. (Описан из Швеции.) Размеры средние: общая длина
черепа самцов 14—15 см. Мех средней пышности и мягкости. Окраска зимнего меха
разнообразна, но преобладают особи со слаборазвитым рисунком; спина обычно ры-
жеватая с сединой, бока ржаво-серые. Европа и Западная Сибирь.
b) F. 1. wrangeli Ognev, 1928. (Тип из района Верхоянска.) Размеры крупнее:
общая длина черепа самцов 15—17 см. Мех особо пышный и мягкий. Преобладают
одноцветные (без пятнистого рисунка) особи с серебристо-белесой или светло-серой
окраской спины и боков. Восточная Сибирь, Якутия.
с) F. 1. isabellina Blyth, 1874. (Описан с Тибета.) Размеры средние, как у евро-
пейской формы. Мех мягкий, пышный. Окраска зимнего меха светлая, одноцветная,
буровато-белесая или сероватая. Горы Средней Азии.
d) F. 1. orientals Satunin, 1905. (Описан из Закавказья.) Размеры более мелкие.
Мех грубый, низкий. В партиях шкур этой рыси резко преобладают особи с рыжей
окраской зимнего меха, обычно с очень ярким рисунком из черных пятен и полосок.
Хорошо дифференцированный подвид, отличимый даже по отдельным шкурам. На ми-
ровом рынке шкуры кавказской рыси ценятся почти в два раза ниже, чем северные
формы. Кавказ и Закавказье.
СЕМЕЙСТВО КУНЬИ. FAMILIA MUSTELIDAE
Мелкие или среднего размера стопо- или полу-стопоходящие хищ-
ники весьма различного роста и телосложения. Как передние, так и
задние ноги пятипалые, вооруженные острыми, но невтяжными ког-
тями. Зубная формула различна у разных видов, но число истинно
коренных обычно у , редко у . Последний верхний предкоренной п
первый нижний коренной почти всегда выделяются как «хищнические»
зубы.
Евразия, Африка (без Мадагаскара), Северная и Южная Америка.
Распадается на 4 подсемейства, представители которых встре-
чаются в СССР.
142
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ СЕМЕЙСТВА КУНЬИХ
ФАУНЫ СССР
1(14). Пальцы как передних, так и задних ног вполне обособлены,
хотя могут быть соединены плавательными перепонками. Задние конеч-
ности не в виде плавательных ластов и средние пальцы длиннее боко-
вых. В каждой половине нижней челюсти по 3 резца..............2
2(5). Мех на брюхе черный или черно-бурый; спина и бока серо-
ватые без примеси коричневатых тонов. От носа через глаз к уху тя-
нется черная полоса. Коронка верхнего заднего коренного зуба в 2—3
раза больше коронки хищнического зуба.........................3
3(4). Ушные раковины имеются. В нижней челюсти с каждой сто-
роны всех коренных зубов (включая предкоренные) не менее 5. Слухо-
вые барабаны далеко не доходят до сочленовных впадин для нижней
челюсти..............................Барсуки. Meles. (Стр. 146.)
4(3). Ушных раковин нет. В нижней челюсти с каждой стороны
по 4 коренных зуба (вместе с предкоренными). Передние концы слу-
ховых барабанов доходят до линии, соединяющей задние края сочле-
новных впадин для нижней челюсти (рис. 39, Л)....................
..................................Медоеды. Mellivora. (Стр. 147.)
5(2). Окраска иная, причем если верх тела светлее низа, то пер-
вый с примесью желтоватых или коричневатых тонов. От носа через
глаз к уху нет черной полосы. Коронка заднего верхнего коренного
зуба меньше, равна или чуть больше коронки хищнического зуба . . 6
6(13). Пальцы не соединены широкой, почти голой плавательной
перепонкой. На подошвах, хотя бы между пальцами, есть волосы.
Хвост не расширен у корня. Ушные раковины значительно шире го-
лого пространства носа. Передний верхний предкоренной зуб распола-
гается между клыком и следующим предкоренным зубом. Предкорен-
4 3
них - Или у ................................................. . 7
7(8). Размеры крупные: длина тела более 65 см. Кондилобазаль-
ная длина черепа взрослых 134—168 мм. Сагиттальный гребень черепа
образует сзади большой выступ, нависающий над затылком. Верхние
хищнические зубы располагаются параллельно друг другу (рис. 39, Г)
.................................... Росомахи. Gulo. (Стр. 147.)
8(7). Размеры значительно мельче: длина тела менее 65 см. Кон-
дилобазальная длина черепа менее НО мм. Сагиттальный гребень не
образует сзади большого выступа. Верхние хищнические зубы распо-
ложены под углом друг к другу.................................9
9(10). Верх и бока головы и по крайней мере передние две трети
верхней губы однотонно бурые или коричневые. Предкоренных и корен-
ных зубов с каждой стороны верхней челюсти по 5, в нижней — по 6.
(Передние предкоренные обеих челюстей иногда выпадают, но их лунки
почти всегда сохраняются.) Задний нижний предкоренной зуб с допол-
нительным зубцом позади главной вершины (рис. 40)................
..................................Куницы. Martes. (Стр. 148.)
10(9). Конец морды или по крайней мере вся верхняя губа белая
или белесая. Предкоренных и коренных зубов с каждой стороны верх-
ней челюсти по 4, в нижней — по 5. Задний нижний предкоренной зуб
без добавочного зубца........................................11
11(12). Окраска спины пестрая с резко очерченными беловато-
желтоватыми пятнами и полосками, густо рассеянными по коричневому
143
Рис. 39 Черепа куньих (снизу):
Л — медоеда (Mellivora indica); Б — барсука (Meles melcs); В — выдры (Lutra lutra);
Г — росомахи (Gulo gulo).
1-14
Рис. 40. Задний нижний
предкоренной зуб (рт4) со-
боля (Martes zibellina);
вид изнутри и несколько
спереди 4- добавочная вер-
шина.
фону. Отростки крыловидных костей до-
ходят до слуховых барабанов и сращены
с ними (рис. 41, Л). На внутренней стороне
средней вершины нижнего хищнического зуба
имеется добавочная вершина (рис. 42) . . .
............Перевязки. Vormela. (Стр. 153.)
12(11). Окраска спины иная, без резко
очерченных пятен и полосок. Отростки крыло-
видных костей далеко не доходят до слухо-
вых барабанов (рис. 41, Б), Нижний хищни-
ческий зуб без добавочной вершины на внутренней стороне...........
..............................Ласки и хорьки. Mustela. (Сгр. 153.)
13(6). Пальцы соединены широкой, почти голой плавательной пе-
репонкой, доходящей на задних лапах до когтей. Подошвы совершенно
Рис. 41. Черепа мелких куньих (снизу):
А — перевязки (Vormela peregusna); Б — светлого хорька (Mustela eversmanni).
Рис. 42. Нижний хищ-
нический зуб (/Hi) пе-
ревязки (Vormela pe-
regusna). Вид изну-
три.
голые. Хвост у корня сильно расширен и постепенно суживается к кон-
цу. Ушные раковины приблизительно одной ширины с голой частью
носа. Передний верхний предкоренной зуб сильно
сдвинут внутрь и располагается с внутренней сто-
4
роны клыка (рис. 39, В). Предкоренных зубов -у
...................Выдры. Lutra. (Стр. 162.)
14(1). Пальцы передних конечностей пол-
ностью срослись. Задние конечности имеют вид
ластов, равномерно покрытых мехом как сверху,
так и снизу; наружный (5-й) палец самый длин-
ный, остальные же постепенно укорачиваются.
В каждой половине нижней челюсти лишь по 2
резца..............Каланы. Enhydra. (Стр. 163.)
Ю Н. А. Бобринский и др.
145
Подсемейство БАРСУКОВ. Subfamilia MELINAE
РОД БАРСУКИ. GENUS MELES Brisson, 1762
Относительно крупные стопоходящие звери. Спина и бока тела се-
ребристо-серые или буровато-серые; брюхо черное или черноватое. Го-
лова беловатая или сероватая с черными полосами, идущими по бокам
головы через глаза к ушам.
Зубная формула: I у С уРт|^М-^--2 = 34 или 38.
Умеренный пояс Евразии.
Один вид.
26. Барсук. Meles meles Linnaeus, 1758
Табл. XII, 5—7. Карта 46
Почти вся Европа (кроме ее северных частей) и большая часть
Азии (за исключением северной полосы Сибири и стран, лежащих
к югу от Гималаев). В СССР встречается на всей территории, лежа-
щей к югу от линии: Средняя Карелия, низовья Северной Двины, вер-
ховья р. Печоры, район г. Ханты-Мансийска, среднее течение р. Под-
каменной Тунгуски, устье р. Витима и хр. Тукурингра. На Сахалине
не водится.
Места обитания весьма разнообразны: различные типы лесов (где
барсуки селятся главным образом по оврагам), степи и полупустыни
(где роют свои норы преимущественно по балкам и в зарослях кустов),
песчаные и глинистые пустыни, горы и горные плато. Норы, в которых
барсуки проводят всю светлую часть суток и залегают в зимний сон,
имеют очень сложное строение: с большим количеством отнорков и
входных отверстий. Эти «барсучьи городки» используются многими по-
колениями зверей. Питается разнообразной, как животной, так и ра-
стительной, пищей; мелкими зверьками, птицами и их яйцами, пресмы-
кающимися, лягушками, насекомыми, моллюсками, червями, падалью,
ягодами, плодами, орехами, желудями, корнями, корневищами, лукови-
цами, семенами, некоторыми травами, грибами. На зиму сильно жи-
реет. Зимний сон длится в средней полосе страны с октября по ап-
рель, а на юге — с ноября по февраль — март; на Кавказе и в Средней
Азии иногда бодрствует всю зиму. Весной или в начале лета самка
•приносит в норе от 2 до 6 (обычно 3—4) детенышей. Вскоре после ро-
дов происходит новое оплодотворение самки. Беременность обычно
длится около года (340—376 дней). Молодые прозревают в месячном
возрасте. Из нор начинают выходить в июне или июле, но остаются
при матери до осени, когда расходятся и начинают рыть себе отдель-
ные норы. Половозрелыми становятся на втором году жизни. Линька
происходит раз в году — летом.
Промысловое значение барсуков невелико: щетинистый остевой во-
лос используют на изготовление кисточек для бритья, а шкурки с бо-
лее мягким волосяным покровом идут на меховые изделия.
Географическая изменчивость различных признаков барсуков, особенно их
окраски, велика. Это позволило описать до 15 форм, обитающих в СССР, но реаль-
ность многих из них вызывает большие сомнения. Вслед за Г. А. Новиковым мы
считаем, что в пределах СССР водится лишь 4 хорошо выраженных подвида:
а) М. m. meles Linnaeus, 1758. (Описан из Швеции.) Размеры относительно круп-
ные: кондилобазальная длина черепа самцов 125—140 мм. Голова беловатая с широ-
кими черными полосами на боках, которые охватывают сверху и снизу
основание ушей. Первые предкоренные зубы обычно имеются. Подглазничное
146
отверстие обычно вытянуто в вертикальном направлении. Западная Европа, Европей-
ская часть СССР на восток до Волги, Крым, Северный Кавказ. (К этому подвиду
относятся: М. m. tauricus Ognev, 1926; М. m. caucasicus Ognev, 1926; M. m. heptneri
Ognev, 1931.)
b) M. m. canescens Blanford, 1878. (Описан из Ирана.) Размеры мельче: кондило-
базальная длина черепа самцов 116—123 мм. Окраска головы, как у предыдущего
подвида. Спина несколько светлее. Зубы, как у номинальной формы. Закавказье, Се-
верный Иран, Узбекистан. (К этому же подвиду относятся: М. m. minor Satunin, 1905,
Закавказье, и М. m. severtzovi Heptner, 1940, Узбекистан.)
с) М. m. leptorhynchus Milne-Edwards, 1867. (Описан из окрестностей г. Пекина.)
Размеры близки к предыдущей форме, кондилобазальная длина черепа самцов
115—131 мм. Голова белесая с желтоватым оттенком; темные полосы по бокам ее узкие
и проходят над основанием уха. Первые предкоренные зубы обычно отсутствуют.
Подглазничное отверстие черепа обычно вытянуто в горизонтальном направлении.
Заволжье, Казахстан, горы Тянь-Шаня, Центральная Азия, Южная Сибирь, Забай-
калье. (К этому подвиду в качестве отдельных natio относятся: М. m. 1. arenarius
Satunin, 1895; М. m. 1. sibiricus Kastschenko, 1900; M. m. 1. raddei Kastschenko, 1901;
M. m. 1. tianschanensis Hoyningen-Huene, 1910.)
d) M. m. amurensis Schrenck, 1858. (Тип из района устья р. Уссури.) Размеры
относительно мелкие: кондилобазальная длина черепа самцов 113—117 мм. Окраска
головы буроватая, темная; поэтому темные боковые полосы слабо заметны. Спина
тоже темйая. В СССР — южная часть Дальнего Востока.
Подсемейство МЕДОЕДОВ. Subfamilia MELLIVORINAE
РОД МЕДОЕДЫ. GENUS MELLIVORA Storr, 1780
По внешности несколько напоминают барсуков, но окраска меха
оригинальна: верх белесого цвета, низ черно-бурый. Размеры немного
мельче, чем у барсуков. Медоеды отличаются от всех остальных куньих
наличием в нижней челюсти только одного коренного зуба с каждой
стороны.
Один вид.
27. Медоед. Mellivora indica Kerr, 1776
Табл. XII, 8. Карта 46
Африка, Индия, Пакистан, Афганистан, Иран. В СССР — Туркме-
ния (по р. Атреку, на Копет-Даге, в бассейне рек Мургаба и Теджена,
Южный Чинк Устюрта). Один из наиболее редких видов нашей фауны.
В Туркмении живет преимущественно в горах и предгорьях, а также
по долинам рек и среди песков. Деятелен обычно ночью. Живет в глу-
боких норах парами. Питается как животной (мелкие позвоночные, на-
секомые), так и растительной пищей (плоды, луковицы). На зиму
в спячку, по-видимому, не впадает. Биология размножения не изучена.
В силу своей малочисленности никакого значения не имеет.
Подсемейство КУНЬИХ. Subfamilia MUSTELINAE
РОД РОСОМАХИ. GENUS GULO Frisch, 1775
Крупные куньи, полустопоходящие, с большими полувтяжными
когтями. Окраска меха варьирует от песчаной до черно-бурой; по бо-
кам тела от плеч к корню хвоста проходят полосы более светлого меха.
Череп массивный с большим сагиттальным гребнем.
Зубная формула: I-|-СyPniyMy-2 = 38.
Один вид.
ю*
147
28. Росомаха. Gulo gulo Linnaeus, 1758
Табл. XII, .9. Карта 46
Вся таежная и лесотундровая зоны Евразии и Северной Америки.
Летом заходит в тундру. В СССР встречается по всей таежной зоне,
от Карелии до берегов Тихого океана. Изолированная часть ареала —
в лесах Полесья.
Населяет как горную, так и равнинную тайгу, криволесье, голь-
цовый пояс гор. Вне времени размножения ведет кочевую жизнь. На
отдых укрывается среди бурелома, в камнях, в чужих норах или про-
сто в лесной чаще. Берлогу делает в таких же скрытных местах, а
иногда и в неглубоких норах. Спаривание — осенью. Детеныши (от 1
до 5) родятся весной, к осени выводки расходятся. Питается весьма
разнообразной пищей: мелкими копытными (молодыми оленями, кабар-
гами, косулями), различными грызунами, птицами, лягушками, рыбой,
не брезгует падалью. Нередко таскает из капканов охотников при-
манку и попавшую в них добычу. Этим, а также истреблением охот-
ничьих животных приносит большой вред охотничьему хозяйству. По-
чти по всему ареалу малочисленна; ценность шкуры невелика. Мало
и промысловое значение росомахи.
Индивидуальная изменчивость окраски меха очень широкая, а географическая
изменчивость слабо выражена.
РОД КУНИЦЫ. GENUS MARTES Pinel, 1792
Средней величины полустопоходящие звери, длина тела 35—80 см,
со стройным туловищем и короткими ногами.
Зубная формула: I у С у Pm у М у • 2 = 38.
Северное полушарие, от северной границы лесной полосы до Сре-
диземного моря, Индии, Индонезии и южных частей США.
Из 7—8 видов этого рода в фауне СССР — 4 вида (2 подрода).
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ КУНИЦ ФАУНЫ СССР
1(2). Окраска пестрая: голова сверху и с боков, концы передних
лап, задние ноги, задняя часть спины и хвост темно-бурые, нижняя
губа и подбородок белые, бока шеи, горло и грудь золотисто-желтые,
передняя часть спины буровато-желтая, брюхо грязно-желтое. Хвост
длинный (35—50 см), малопышный; длина его около 2/з длины тела.
Длина тела взрослых 75—80 см. Кондилобазальная длина черепа более
100 мм..........................Харза. Martes flavigula. (Стр. 150.)
2(1). Окраска всей шкурки почти одноцветная, изменяющаяся от
песочно-желтой до черно-бурой; лишь на груди обычно имеется белое
или желтое пятно. Хвост пушистый, относительно короткий, длина его
без концевых волос менее 30 см, она примерно равна половине длины
тела или еще короче. Длина тела менее 57 см. Кондилобазальная дли-
на черепа менее 100 мм..........................................3
3(6). Хвост относительно длинный: длина его с концевыми воло-
сами заметно превышает х/г длины тела; конец хвоста сильно выдается
за пальцы вытянутых назад задних ног. На груди расположено боль-
шое, резко ограниченное белое, желтое или оранжевое пятно. Верх го-
148
ловы того же цвета, что и спина. Слуховые барабаны относительно
широко расставлены: расстояние между ними в области отверстий
для сонных артерий больше V2 длины барабанов (рис. 43, 5, В) . . 4
4(5). Горловое белое пятно продолжается назад двумя отростками,
спускающимися на передние стороны передних ног. Хвост гораздо тем-
нее туловища, черно-бурый. Подпушь меха спины светлая, белесая.
Подошвы лап покрыты редкими волосами, из которых ясно выступают
голые подушечки. Расстояние от заднего края лунки клыков верхней
челюсти до заднего края подглазничного отверстия равно или лишь
немного больше V2 расстояния между концами заглазничных отрост-
ков. Третий предкоренной зуб верхней челюсти не имеет на внутренней
стороне ясного выступа . Каменная куница. Martes foina. (Стр. 150.)
Рис. 43. Черепа куниц и соболя
(вид снизу):
А — соболь (Martes zibellina); Б — лесная куница (Martes martes); В — каменная
куница (Martes foina); С—клык; Рпг3— третий предкоренной зуб; Ртр —
крупный (четвертый) предкоренной (или хищнический) зуб; М1 — первый
заднекоренной; рр — предзатылочный отросток; ppt — отросток крыловидной кости;
feu — отверстие евстахиевой трубы; fc — сонное отверстие; tau — слуховая трубка.
5(4). Горловое пятно продолжается назад одним клинообразным
выступом, уходящим между передними ногами. Окраска его оранже-
вая, желтая или кремовая (как исключение почти белая). Окраска
хвоста того же цвета, что и спины, или лишь слегка темнее ее. Под-
пушь голубовато-серая или желтовато-серая, относительно темная. По-
дошвы лап покрыты мехом. Расстояние от заднего края лунки клыка
верхней челюсти до заднего края подглазничного отверстия значи-
тельно больше V2 расстояния между концами заглазничных отростков.
Третий предкоренной зуб верхней челюсти несет на внутренней стороне
ясный выступ.............Лесная куница. Martes martes. (Стр. 150.)
6(3). Хвост относительно короткий: длина его с концевыми волос-
ками обычно менее V2 длины тела; конец хвоста едва выдается за
пальцы вытянутых назад задних ног. Светлого пятна на горле нет
совсем или оно имеет вид небольшой желтой «звездочки», либо боль-
шое, но неясно отграниченное по цвету от окружающего его меха.
Верх головы обычно седоватый, более светлый, чем спина. Слуховые
149
барабаны черепа вытянуты в продольном направлении и сближены друг
с другом; расстояние между ними в области отверстий для сонных
артерий меньше или реже равно х/г длины барабанов...................
...............................Соболь. Martes zibellina. (Стр. 151.)
Подрод харз. Subgenus Charronia Gray, 1785
29. Харза. Martes flavigula Boddaert, 1785
Табл. XIII, 1. Карта 47
Юго-Восточная Азия. В СССР — Амуро-Уссурийский край.
Живет в лесах разного типа, предпочитая лиственные перестойные
насаждения с большим количеством дуплистых деревьев. Превосходно
лазает по деревьям. Питается различными животными, решаясь напа-
дать даже на крупную добычу (косуля, кабарга). Спаривание — в июне
и июле. Молодые (2—4 в помете) родятся в мае. Приносят вред истреб-
лением ценных охотничьих животных, в том числе соболя. Поэтому
добыча ее разрешается круглый год. Ценность шкурки невелика.
В СССР ежегодно добывается всего лишь несколько десятков их.
Харзы, обитающие на Дальнем Востоке, относятся к подвиду: М. f. aterrima Pallas.
1811. (Описан по экземпляру с Амура.)
Подрод куниц. Subgenus Martes Pinel, 1792
30. Каменная куница. Martes foina Erxleben, 1777
Табл. XIII, 2. Карта 47
Западная Европа, Передняя, Средняя, Центральная и Восточная
Азия. В СССР — Украина, Белоруссия, Латвия, Литва, Кавказ, Закав-
казье, горы Средней Азии, Тарбагатай, Алтай. Поднимается в горы
до 3000 м.
Обитает главным образом в горах, где держится как в лесах, так
и среди скал и каменистых осыпей. В равнинных районах встречается
в поселках и даже крупных городах, где живет на чердаках, в забро-
шенных каменных постройках, в парках. Питается мелкими грызунами,
птицами, пресмыкающимися, лягушками, насекомыми, моллюсками.
Охотно поедает плоды и ягоды. Спаривается в июле. Беременность
длится 236—274 дня. Рождение детенышей (в числе от 2 до 8) — в ап-
реле— мае.
Ценный пушной вид. Но промысловое значение не очень большое,
так как добывается раз в десять меньше, чем лесных куниц.
В фауне СССР — 2 слабо различающиеся формы:
а) М. f. foina Erxleben, 1777. (Описана по экземпляру из Германии.) Окраска отно-
сительно темная. Подпушь сероватая. Западная Европа, Европейская часть СССР,
Кавказ.
b) М. f. intermedia Severtzov, 1873. (Описана по экземпляру из гор Средней Азии.)
Окраска светлее, подпушь беловатая. Средняя Азия, Алтай.
31. Лесная куница. Martes martes Linnaeus, 1758
Табл. XIII, 3. Карта 47
Леса таежной и лесостепной зоны Европейской части СССР и за-
падных районов Западной Сибири (на восток до Оби и Нижнего Ир-
тыша), лесные ландшафты Кавказа и Закавказья, Западная Европа.
Типично лесной зверек, населяющий леса различных типов, особенно
по. долинам рек. В горах поднимается до верхней границы леса. Ведет
150
ночной, преимущественно древесный образ жизни. Гнездо устраивает
в дупле, нередко пользуется беличьим тайном и другими укрытиями.
Вне периода размножения обычно ведет кочевую жизнь. Гон в июле.
Беременность — 230—270 дней. Латентная стадия развития зародыша
продолжается до 6—7 месяцев. Число детенышей в помете— от 2 до
8 (обычно 3—4). Родятся они в апреле — мае. К осени начинают вести
самостоятельную жизнь. Пища — мелкие зверьки (белки, лесные по-
левки и мыши), птицы, их яйца и птенцы, реже ящерицы, насекомые.
Охотно ест ягоды.
Имеет важное промысловое значение: шкура ценится высоко.
Географической изменчивости подвержены размеры и тон окраски меха. Разли-
чают следующие подвиды:
а) М. т. martes Linnaeus, 1758. (Описан из Швеции.) Размеры относительно круп-
ные: длина тела самцов 48—55 см, хвоста 20—24 см; кондилобазальная длина черепа
самцов 81—88 мм. Мех очень пышный, мягкий. Окраска меха сверху темно-кашта-
новая. Горловое пятно обычно оранжевое. Западная Европа, прибалтийские респуб-
лики, Карелия, Мурманская область, БССР, западные области РСФСР.
b) М. m. borealis Kuznetzov, 1941. (Тип из Шенкурского района.) Размеры мельче:
длина тела самцов 38—46 см, хвоста 17—24 см; кондилобазальная длина черепа сам-
цов менее 82 мм. Мех пышный, очень шелковистый. Окраска светлая, горловое пятно
светло-желтое. Северные области Европейской части СССР к востоку от Белого моря.
с) М. ш. uralensis Kuznetzov, 1941. (Тип из окрестностей г. Миасса.) Размеры, как
у номинальной формы: кондилобазальная длина черепа самцов 80—87 мм. Окраска
очень светлая, с характерным желтым оттенком на огузке. Горловое пятно белесое.
Урал, Башкирия.
d) М. ш. ruthena Ognev, 1926. (Тип из Московской области.) Размеры средние:
длина тела самцов 45—50 см, хвоста 16—20 см; кондилобазальная длина черепа
самцов 75—82 мм. Окраска разнообразная, обычно средней темноты с рыжеватыми
оттенками. Центральные области Европейской части РСФСР.
е) М. m. lorenzi Ognev, 1926. (Тип с Кубани.) Размеры крупные: длина тела сам-
цов 50—58 см, хвоста 23—25 см; кондилобазальная длина черепа самцов 75—86 мм.
Мех густой, но невысокий и грубоватый. Окраска темная с характерным рыжевато-
сливковым оттенком. Кавказ.
32. Соболь. Martes zibellina Linnaeus, 1758
Табл. XIII, 4—5. Карта 47
Прежде (до XVIII века) обитал по всей таежной области, от Ка-
релии до Тихого океана. Вследствие хищнического промысла в XVIII
и XIX веках в ряде частей своего ареала был истреблен и сохранился
только в отдельных районах Северного Урала, Сибири и Дальнего
Востока. В результате принятых Советской властью мер численность
соболя вновь стала высокой, а его ареал путем как естественного рас-
селения, так и специальных выпусков в места прежнего обитания в зна-
чительной степени восстановлен (см. карту). Кроме СССР, обитает
в некоторых районах Северной Монголии, в горах Большого и Ма-
лого Хингана и в Корее.
Типичный обитатель тайги; встречается также среди каменистых
россыпей, покрытых кедровым сланцем .Ведет преимущественно назем-
ную жизнь, хотя хорошо лазает по деревьям. Чаще охотится в сумер-
ках и ночью, но в глухих местах нередко бодрствует и днем. Основная
пища — мелкие птицы, их яйца и птенцы, мелкие грызуны, но напа-
дает и на более крупных животных. Охотно ест кедровые орешки и
ягоды. Гнездо устраивает в валежнике, среди камней, в расщелинах
скал, среди корней деревьев. Вне периода спаривания взрослые со-
боли живут поодиночке. Гон — в июле. Беременность — 253—297 дней,
из которых около 7 месяцев приходится на латентную стадию развития
эмбриона. Молодые, числом от 2 до 6 (обычно 3—4), родятся в ап-
реле. Появляются они на свет слепыми, покрытыми редкой короткой
151
беловатой шерстью. На 34—35-й день прозревают. В возрасте 6—8 не-
дель начинают выходить из гнезда. Сосут мать 1,5—2 месяца. К осени
достигают размеров взрослых. Весенняя линька начинается в марте
и заканчивается в мае. Осенняя смена волосяного покрова длится
с конца августа по ноябрь.
Ценнейший пушной вид. В последние годы шкурки соболя вышли
по ценности на первое место в общесоюзных заготовках пушниньт
В последние годы численность соболей резко возросла.
Для соболей характерна широкая индивидуальная изменчивость. Особенно из-
менчива окраска меха. Например, окраска прибайкальских соболей изменяется от
песчано-желтой до черно-бурой. Так же сильно выражена у соболей и географическая
изменчивость в размерах, пышности и мягкости волосяного покрова и особенно
в окраске меха.
На территории СССР можно выделить следующие подвиды, различающиеся глав-
ным образом по преобладающему (в больших партиях шкурок) типу окраски, пыш-
ности и мягкости меха:
а) М. z. zibellina Linnaeus, 1758. (Тип из окрестностей г. Тобольска.) Кондилоба-
зальная длина черепа самцов 84—89 мм, самок 77—79 мм. Окраска светлая: преобла-
дают особи песочно-желтой или светлой, ржаво-коричневой окраски. Шкурки свет-
лой окраски составляют около 60%, а средней темноты — до 40%. Темные шкурки
попадаются редко. Мех пышный, но грубоватый. Тайга от бассейна Печоры и Север-
ного Урала до Енисея.
На Северном Урале и в Печорской тайге встречается помесь соболя и ку-
ницы — кидас.
b) М. z. averini Bashanov, 1943. (Тип из Катон-Карагайского района, на Алтае.}
Размеры крупные. Окраска меха сравнительно темная: темных шкурок 26%, средней
темноты 66% и светлых 8%. Мех малопышный, с короткой остью. Тайга Алтая.
с) М. z. jeniseensis Ognev, 1925. (Тип из Красноярского района.) Окраска меха
средней темноты: шкурок темных около 10%, средней темноты около 40% и свет-
лых до 50%. Мех средней пышности и мягкости. Кузнецкий Алатау, равнинная тайга
по рекам Чулыму, Кети, Сыма и Касса и плоскогорье на правом берегу р. Енисея
на восток до р. Оки и на север до водораздела Ангары и Подкаменной Тунгуски.
(К этой же форме, по-видимому, относится М. z. tomensis Timofeev et Nadeev, 1955x
с Кузнецкого Алатау.)
d) M. z. sajanensis Ognev, 1925. (Тип с p. Орзыбай, в Саянах.) Размеры средние.
Окраска преобладает темная: в больших партиях шкурок темных 17%, средних по
темноте 71% и светлых 12%. Преобладают шкурки коричневато-бурой расцветки с се-
диной. Мех малопышный, редковатый, но мягкий. Саяны, горы Тувы.
е) М. z. tungussensis Kuznetzov, 1941. (Тип с р. Тунгуски.) Размеры крупные.
Окраска средней темноты. Преобладают шкурки коричневого тона с рыжеватыми бо-
ками. В партиях шкурок 10—16% темных, около 50—53% средней темноты и около*
31—40% светлых. Мех пышный, но грубоватый. Бассейн рек Подкаменной и Нижней
Тунгуски, Хатанги и Пясины. (К этому же подвиду в качестве natio относится М. z.
ilimpiensis Timofeev et Nadeev, 1955.)
f) M. z. princeps Birula, 1916. (Тип с Баргузинского хребта). Окраска очень тем-
ная. В партиях шкурок преобладают темные, а светлые редки. Мех очень шелкови-
стый и пышный. Распространен этот подвид в горах Забайкалья. Степень темноты шку-
рок соболей из разных мест Забайкалья неодинакова. В. В. Тимофеевым и В. Н. Наде-
евым (1955) выделено несколько рас забайкальских соболей, которые мы считаем не-
более чем за natio данного подвида (М. z. р. vitimensis, Правобережье Киренги и Лены„
верховья Витима и В. Ангары; М. z. р. obscura, юго-восточная часть Яблонового
хребта; М. z. р. jacutensis Kuznetzov, верховья рек Алдана и Зеи).
g) М. z. sachalinensis Ognev, 1925. (Тип с о-ва Сахалин.) Размеры сравни-
тельно мелкие: длина тела самцов 39—45 см, самок 36—39 см; кондилобазальная
длина черепа самцов 76—82 мм, самок 70—76 мм. Окраска грубоватого и малопышного
меха светлая; темных шкурок около 10%, средних по темноте около 65% и светлых до
25%. (К этому же подвиду относятся: М. z. s. arsenjevi Kuznetzov, 1941; M. z. s.
schantaricus Kuznetzov, 1945; Нижний Амур, Шантарские острова.)
h) М. z. kamtschadalica Birula, 1918. (Тип с Камчатки.) Наиболее крупная форма:
длина тела самцов 45—58 см, самок 42—52 см: кондилобазальная длина черепа сам-
цов 86—95 мм, самок 79—85 мм. Мех очень пышный, но грубоватый. Окраска очень
разнообразная, но преобладают шкурки бурой окраски с сединой. Темных шкурок
16%, средних по темноте 76°/о, светлых 8%.
152
РОД ПЕРЕВЯЗКИ. GENUS VORMELA Blasius, 1884
По размерам и форме тела сходны с хорьками, но резко отли-
чаются от них своеобразной окраской и некоторыми особенностями
строения зубов. Спина коричневого или бурого цвета, испещренная
многочисленными мелкими и желтыми пятнами и полосками. Нижняя
сторона тела и конечности темно-бурые. Голова бурая с широкими бе-
лыми полосами над глазами, вокруг рта и по краям ушей. Пышный
хвост в своей средней части светлый, в конечной темно-бурый.
Зубная формула: I — С— Pm — М — -2 = 34.
3 1 3 2
Нижний хищнический зуб с хорошо развитой дополнительной вер-
шиной с внутренней стороны главного зубца.
Степи и пустыни Евразии.
Один вид.
33. Перевязка. Vormela peregusna Giildenstaedt, 1770
Табл. XIII, 6. Карта 48
Степи Болгарии, Румынии, южных областей Украины, Северного
Кавказа, полупустыни Закавказья, Нижнего Поволжья, Казахстана,
пустыни Средней Азии, Ирана, Центральная Азия. Почти повсеместно
редка. Ночной зверек, проводящий день в норе, которую роет сам или
отнимает у других зверьков. Питается мелкими грызунами, неболь-
шими птицами, ящерицами, насекомыми. Биология размножения не
изучена. Течка происходит, по-видимому, в конце зимы. Беременность,
вероятно, около 2 месяцев. Самка ранней весной приносит 3—8 дете-
нышей.
В силу малочисленности и малой ценности шкурки перевязка
в пушном промысле СССР играет ничтожную роль.
В пределах Советского Союза можно выделить 3 подвида, различия между ко-
торыми недостаточно четки в силу большой индивидуальной изменчивости:
а) V. р. peregusna Giildenstaedt, 1770. (Тип из долины р. Дона.) Основной фон
спины бурый, пятна буровато-желтые. Надглазничная белая полоса на лбу прервана.
Европа, Передняя Азия, Закавказье, Западная Туркмения.
b) V. р. alpherakii Birula, 1910. (Тип из окрестностей Ашхабада.) Спина черновато-
бурая с очень светлыми бледно-желтыми или беловатыми пятнышками. Надглазнич-
ная полоса обычно сплошная. Средняя и Восточная Туркмения.
с) V. р. negans Miller, 1912. (Тип из Ордоса.) Спина палево-желтая с очень мел-
кими ржаво-бурыми пятнышками. Пятна имеются и на брюхе. Лобная перевязь под
глазами соединяется с белым горлом. От Юго-Западного Казахстана до Внутренней
Монголии. (К той же форме относятся слабо выраженные уклонения, описанные
С. У. Строгановым (1948) в качестве особых подвидов: V. р. pallidior из Семиречья,
и V. р. obscura из Южного Таджикистана.)
РОД ЛАСКИ И ХОРЬКИ. GENUS MUSTELA Linnaeus, 1758
Окраска спины без рисунка из пятен и полос.
3 1 3 1
Зубная формула: I — С — Pm — М — • 2 = 34.
3 1 3 2
Нижний хищнический зуб без добавочного зубца внутренней сто-
роны главной вершины.
Северное полушарие на юг до Северной Африки, Северной Индии,
Индонезии и северных районов Южной Америки.
Из 12—13 видов в фауне СССР — 8 видов (в том числе один, заве-
зенный из Северной Америки).
153
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА MUSTELA
ФАУНЫ СССР
1(4). Зимний мех обычно белый. Летний мех (а у южных форм и
зимний) на спине коричневый, а на брюхе белый или желтоватый; гра-
ница между темной окраской спины и светлым брюхом резкая. Пред-
глазничные отверстия черепа велики: их поперечный диаметр равен или
больше продольного поперечника альвеолы верхнего клыка .... 2
2(3). Хвост на всем протяжении коричневый или белый (лишь
иногда на самом конце хвоста имеется небольшое количество черных
волос). Длина хвоста заметно меньше V2 длины тела. Ширина черепа
над клыками приблизительно равна ширине межглазничного проме-
жутка.........................Ласка. Mustela nivalis. (Стр. 156.)
Рис. 44. Черепа хорьков сверху:
А — светлый хорек (Mustela eversmanni); Б — черный хорек (Mustela
putorius).
3(2). Конечная половина или !/3 хвоста черная, а основная часть
его белая или коричневая. Длина хвоста с концевыми волосами равна
или превышает V2 длины тела. Ширина черепа над клыками заметно
меньше ширины межглазничного промежутка............................
...........................Горностай. Mustela erminea. (Стр. 157.)
4(1). Окраска как летнего, так и зимнего меха на спине и на
брюхе одинаковая или окраска брюха темнее спины. Граница между
окраской спины и брюха неясная, с постепенным переходом. Предглаз-
ничные отверстия малы: их поперечный диаметр меньше продольного
поперечника альвеолы верхнего клыка..............................5
5(8). Окраска спинной и брюшной стороны тела, лап и хвоста ры-
жая, темно-желтая или серовато-желтая. Слуховые барабаны бобовид-
ной или удлиненно-четырехугольной формы. Их внутренние края тя-
нутся более или менее параллельно друг другу.....................6
154
барабаны
б
Верхние
зу-
45.
а
Рис.
предкоренные
бы норок:
а — европейская нор-
ка (Mustela lutreola);
б — американская
норка (Mustela vi-
son).
бр). Размеры мелкие: длина тела до 26 см. Губы и подбородок
белесые; их окраска постепенно переходит в окраску смежных частей
головы. Верх головы, от конца морды до ушей, буровато-серый. Конди-
лобазальная длина черепа самцов 47—53 мм, самок 40—47 мм . . .
.............................Солонгой. Mustela altaica. (Стр. 158.)
7(6). Размеры крупнее: длина тела взрослых более 26 см. Губы и
подбородок белые; их окраска резко отграничена от окраски смежных
частей головы. Конец морды бурый, лоб рыжеватый. Кондилобазаль-
ная длина черепа самцов 55—68 мм, самок 50—55 мм..................
..............................Колонок. Mustela sibirica. (Стр. 158.)
8(5). Окраска всего тела темно-бурая или коричневая, либо темно-
бурым мехом одеты конечности, грудь и паха. Слуховые барабаны
треугольной формы. Их внутренние края сближены
спереди и расходятся кзади ................. 9
9(12). Окраска спины заметно отличается от
расцветки брюха. Ноги, грудь и пахи покрыты
черно-бурым или бурым мехом. Края ушей свет-
лые. Ширина черепа в области слуховых бараба-
нов значительно больше половины его кондилоба-
зальной длины. Лобная площадка черепа выпу-
клая .......................................10
10(11). Основная часть хвоста светлая, а ко-
нечная черно-бурая. На спине светлый подшерсток
ясно просвечивает через короткие черноватые
окончания остевых волос. Брюхо светлое с тем-
ными пятнами между передними и задними но-
гами. Область черепа сзади надглазничных отрост-
ков с ясным сужением посередине................
. Светлый хорек. Mustela eversmanni. (Стр. 159.)
11(10). Хвост черный или черно-бурый на всем
Брюхо черноватое. На спине черная ость скрывает или почти скрывает
светлый подшерсток. Область черепа позади надглазничных отростков
без резкого сужения посередине, с почти параллельными боковыми
краями....................Черный хорек. Mustela putorius. (Стр. 160.)
12(9). Окраска всего тела одноцветная темно-бурая, коричневая
или ржаво-коричневая; лишь на губах, иногда на груди и брюхе, име-
ются белые пятна. Края ушей темные. Ширина черепа в области слу-
ховых барабанов приблизительно равна V2 его кондилобазальной
длины. Лобная площадка уплощенная..............................13
13(14). Как нижняя, так и верхняя губа белая. Наименьшая ши-
рина черепа позади заглазничных отростков равна или превышает ши-
рину межглазничного промежутка. Второй предкоренной зуб верхней
челюсти своим задним концом соприкасается с передним краем внеш-
ней лопасти хищнического зуба (рис. 45, а)........................
..................Европейская норка. Mustela lutreola. (Стр. 160.)
14(13). Нижняя губа покрыта белым, а верхняя коричневым ме-
хом. Наименьшая ширина черепа позади заглазничных отростков
меньше ширины межглазничного промежутка. Второй предкоренной
зуб верхней челюсти своим задним концом заходит в выемку между
наружной и внутренней лопастями переднего конца хищнического зуба
(рис. 45, б) . . . . Американская норка. Mustela vison. (Стр. 161.)
своем протяжении.
155
Подрод ласок. Subgenus Mustela Linnaeus, 1758
34. Ласка. Mustela nivalis Linnaeus, 1766
Табл. XIII, 7—10. Карта 48
Европа, Северная Африка, Азия (кроме ее южных стран), Север-
ная Америка. Почти вся территория СССР, кроме некоторых островов
Арктики.
Встречается в самых разнообразных ландшафтах. В горах подни-
мается до высоты 4 тыс. м. Более обычна в южных частях ареала.
Не избегает близости человека и поселяется иногда даже в городских
постройках. Биология изучена слабо. Молодых находили в разное
время теплого периода года. Число их колеблется от 3 до 10. Основная
пища — мелкие грызуны, истребляя которых на полях и в скирдах,
ласка приносит пользу сельскому хозяйству.
Промысловое значение ничтожно, так как шкурка ласки ценится
весьма низко.
У ласки резко выражен половой диморфизм: самцы могут быть вдвое крупнее
самок (по весу). У северных форм отчетливо выражен сезонный диморфизм (зимой
белые, летом сверху коричневые).
Индивидуальная изменчивость сложно переплетается с географической изменчи-
востью. Общую схему последней можно выразить так: «Ласки из северных частей
ареала отличаются мелкими размерами тела, очень коротким хвостом и чисто белой
окраской меха зимой. К югу размеры увеличиваются, появляется тенденция к сохра-
нению бурой окраски в течение всего года» (Г. А. Новиков, 1956). Добавим к этому,
что в горах юга Палеарктики имеет место уменьшение размеров по мере подъема над
уровнем моря. В пределах СССР можно различить следующие основные подвиды;
а) М. п. nivalis Linnaeus, 1766. (Тип из окрестностей Вестерботтена, Северная’
Швеция.) Размеры средние: длина тела самцов 131—208 мм\ хвост короткий —
21—40 мм\ кондилобазальная длина черепа самцов 32—37 мм. Окраска спины
летом интенсивная буро-коричневая. Скандинавия, Европейская часть СССР, Урал
и Зауралье от берегов Белого, Баренцева и Карского морей до лесостепи вклю-
чительно.
b) М. п. pygmaeus J. Allen, 1903. (Тип из Гижиги, северо-восточный берег Охот-
ского моря.) Размеры мелкие: длина тела самцов 142—162 мм\ хвост очень ко-
роткий— 16—20 мм (самая короткохвостая форма!); кондилобазальная длина черепа
самцов 30—34 мм. Окраска летнего меха спины темно-шоколадная. Почти вся Си-
бирь к востоку от Иртыша и Оби, лесостепь Северного Казахстана.
с) М. п. vulgaris Erxleben, 1777. (Тип из окрестностей г. Лейпцига, Германия.)
Размеры крупные: длина тела самцов 200—220 мм; хвост относительно длинный —
45—61 мм; кондилобазальная длина черепа самцов 36—44 мм. Окраска летнего меха
на спине интенсивно-коричневая. Часть зверьков на зиму не белеет или белеет непол-
ностью. Значительная часть Западной Европы, степи и полупустыни Европейской
части СССР и Западного Казахстана. К этому же подвиду мы относим в качестве
natio крымскую М. п. v. nikolskii Semenov, 1899, отличающуюся чуть более светлой
окраской летнего меха.
d) М. п. caucasica Barret-Hamilton, 1990. (Тип с горы Гочал, Кавказ.) Размеры
мелкие: длина тела самцов 107—185 мм; хвост относительно длинный—43—56 мм;
кондилобазальная длина черепа 31—34 мм. Окраска спины летом бледная, рыжевато-
охристо-коричневая; зимой белая. Кавказ и Закавказье (преимущественно горные
районы).
е) М. п. dinniki Satunin, 1907. (Тип из окрестностей Ставрополя Кавказского.)
Размеры очень крупные: длина тела самцов 176—260 мм; хвост длинный —55—78 мм;
кондилобазальная длина черепа самцов 38—44 мм. Летняя окраска меха, как
у М. п. vulgaris. На зиму часть особей белеет, у других побеление наблюдается лишь
па боках. Кавказ и Закавказье (преимущественно равнинные, предгорные и низко-
горные ландшафты). Местами встречается вместе с мелкой кавказской формой
(М. п. caucasicus). Очень близка к средиземноморской М. п. boccamela Bechstein, 1800,
описанной по экземпляру с острова Сардиния.
f) М. п. stoliczkana Blanford, 1877. (Тип из Яркенда.) Размеры крупные: длина
тела самцов около 240—250 мм; хвост длинный — 60—77 мм; кондилобазальная длина
черепа 42—46 мм. Окраска меха бледная, буровато-песчаная; на зиму обычно не бе-
леет. Кашгария, пустыни Казахстана и Средней Азии. К этому подвиду мы относим
natio М. п. s. heptners Morozova-Turova, 1953, описанную из Южной Туркмении.
156
g) M. n. pallida Barret-Hamilton, 1900. (Тип из окрестностей г. Коканда, Фергана.)
Размеры средние: длина тела самцов 180—213 мм; хвост 37—60 мм; кондилобазальная
длина черепа самцов 32—41 мм. Окраска летнего меха неотличима от предыдущей
формы; зимний мех белый. Горы Средней Азии.
35. Горностай. Mustela erminea Linnaeus, 1758
Табл. XIII, 11—12. Карта 49.
Европа, кроме стран Средиземноморья, Азия к югу до Гималаев,
Северная Америка. В Советском Союзе почти повсеместно, кроме
Крыма, Закавказья, пустынь Средней Азии, Южного Казахстана и не-
которых островов Арктики.
Места обитания весьма разнообразны. В тундровой зоне больше
по долинам рек, хотя водится и в болотистой, и в каменистой тундре.
В таежной и лесолуговой зонах встречается в самых различных ланд-
шафтах. В степи и полупустыне держится главным образом по доли-
нам рек, по берегам озер, в колках и зарослях кустов, близ поселений
человека. В горных районах предпочитает лесные ландшафты, берега
рек и осыпи камней. Основная пища — мелкие грызуны: пищухи, водя-
ные крысы, полевки, мыши. Свое убежище устраивает в кучах камней,
в трещинах скал, в валежнике, под корнями деревьев, в низко распо-
ложенных дуплах; нередко роет неглубокую нору или занимает норы
грызунов. Активен преимущественно в ночное время. Беременность —
9—10 месяцев. Латентная стадия развития зародыша более 7 меся-
цев. Гон — в начале лета. Молодые, в количестве от 3 до 18 (обычно
7—9), родятся в средней полосе в конце апреля — в начале мая. Про-
зревают на 9—12-й день, а в возрасте двух месяцев покидают нору.
К осени, почти достигнув размеров взрослых, начинают вести самосто-
ятельную жизнь. Почти во всех районах нашей страны на зиму оде-
вает белый мех. Весенняя линька в центральных районах длится с на-
чала марта по май, осенняя — с сентября по ноябрь.
Принадлежит к числу важных объектов пушного промысла. В по-
следние годы в СССР ежегодно заготовляется 200—300 тыс. шкурок.
Особенно много горностаев, притом дающих шкурки высокого качества,
добывается в Западной Сибири и в Северном Казахстане.
Половой и сезонный диморфизмы горностая выражены очень резко, а географи-
ческая изменчивость размеров тела, пропорции отдельных частей черепа, размеров
зубов и отчасти окраски летнего меха — слабо. В пределах СССР различалось 13
подвидов, однако большинство их практически неразличимы. По очень слабым откло-
нениям в сериях можно реально различить лишь следующие из описанных форм:
а) М. е. erminea Linnaeus, 1758. (Описана из Швеции.) Размеры средние: кондило-
базальная длина черепа самцов 43—4'9 мм. Носовой отдел черепа относительно ши-
рокий. Скандинавия; в СССР — Кольский полуостров, Карелия.
b) М. е. aestiva Кегг, 1792. (Описана из Германии.) Размеры чуть крупнее: кон-
дилобазальная длина черепа самцов 45—52 мм. Носовой отдел черепа несколько уже.
Мех средней высоты и мягкости. В СССР — Европейская часть страны, Урал, Север-
ный Казахстан, Алтай. (Форма очень близкая к номинальной и не отличается от
М. е. birulai Martino, 1931; М. е. martinoi Ellerman et Morrison-Scott, 1951, из окрестно-
стей г. Актюбинска, и от М. е. lymani Hollister, 1912, с Алтая.)
с) М. е. tobolica Ognev, 1923. (Тип из окрестностей г. Тары, Омская область.)
Размеры очень крупные: длина до 30 см, кондилобазальная длина черепа самцов
в среднем около 49 мм. Мех пышный и мягкий. Равнинная Западная Сибирь.
d) М. е. kaneii Baird, 1857. Размеры средние: кондилобазальная длина черепа
самцов 44—49 мм. Окраска летнего меха светлая. Слуховые барабаны несколько уко-
рочены. Северная часть Восточной Сибири, от Енисея до Камчатки. Синонимами этой
формы следует признать: М. е. orientalis Ognev, 1928 (тип из окрестностей сел. Поход-
ского на Колыме); М. е. naumovi Jurgenson, 1936 (тип с р. Хатанги), и М. е. arctica
Merriam, 1896 (тип из Аляски).
е) М. е. transbaicalica Ognev, 1928. (Тип из Баргузинского заповедника.) Размеры
мелкие: кондилобазальная длина черепа самцов 42—46 мм. Забайкалье, Южная
157
Якутия, Амуро-Уссурийский край, Охотское побережье. (К этой же форме мы относим
мелкую островную форму горностаев: М. е. baturini Ognev, 1929, описанную с о-ва
Большой Шантар.)
f) М е. ferghanae Thomas, 1895. (Тип из Кара-Карык, Фергана.) Размеры отно-
сительно мелкие: кондилобазальная длина черепа самцов 41—47 мм. Мех редкий,
низкий. Окраска светлая.
Особи, живущие на равнинах и в предгорьях, на зиму не белеют. Средняя Азия.
Вероятно, в особую форму будут выделены горностаи с Кавказа.
36. Солонгой. Mustela altaica Pallas, 1811
Табл. XIV, /. Карта 50
Восточная и Центральная Азия, В СССР — Приморье, по Амуру,
Забайкалье, Прибайкалье, Саяны, Алтай, Тарбагатай у оз. Балхаш,
Джунгарский Алатау, восточные районы Киргизии и Памир.
Держится главным образом в горах, но встречается и в низменных
местах, даже в камышах по берегам оз. Балхаш. Иногда живет в по-
селениях человека. Питается преимущественно мелкими грызунами, а
также небольшими птицами, пресмыкающимися, лягушками, насеко-
мыми, ягодами. Гон — в феврале — марте. Число детенышей 3—8. Про-
мысловое значение невелико.
Различают следующие подвиды, встречающиеся в СССР.
а) М. a. altaica Pallas, 1811. (Описан с Алтая.) Размеры относительно крупные:
кондилобазальная длина черепа самцов 51—53 мм. Окраска зимнего меха бледная,
сероватая. Алтай, Саяны, Прибайкалье, по-видимому, Тянь-Шань.
b) М. a. raddei Ognev, 1928. (Тип из окрестностей оз. Торей-Нур, Забайкалье.}
Размеры мельче: кондилобазальная длина черепа 48—51 мм, Окраска зимнего меха
заметно более яркая. Забайкалье, Амурский край, Приморье.
с) М. a. birulai Ognev, 1928, (Тип из Лянгара, Западный Памир.) Размеры средние.
Окраска зимнего меха очень бледная; хвост резко двухцветный (снизу почти белый).
Не вполне ясны отличия этой формы от М. a. temon Hodgson, 1857, описанного из
Сиккима (Гималаи).
37. Колонок. Mustela sibirica Pallas, 1773
Табл. XIV, 2. Карта 50
Лесные районы Дальнего Востока (кроме Камчатки), таежная и ле-
солуговая зоны Сибири и восточных областей Европейской части СССР
на запад до Кировской области, Татарии и Башкирии. Продолжает
расселяться на запад: отдельные особи стали появляться уже в Горь-
ковской области. Широко распространен в Восточной и Центральной
Азии. Успешно акклиматизировался в котловине оз. Иссык-Куль.
Обитает в лесах различного типа, по долинам рек, по кочковатым
болотам, среди каменистых осыпей в горах, в колках лесостепи. Не-
редко живет в поселениях человека. Питается преимущественно мел-
кими грызунами: полевками, мышами, водяными крысами, бурунду-
ками, пищухами и даже зайцами. Поедает также птиц, ящериц, ля-
гушек, насекомых, ягоды и кедровые орешки. Иногда нападает на
домашнюю птицу. Норы устраивает под пнями, валежником, камнями,
нередко селится в низко расположенных дуплах. Охотится преимущест-
венно на земле, выходя на охоту с наступлением сумерек. Течка проис-
ходит в конце зимы — в начале весны. Беременность длится 28—30'
дней. Число детенышей в помете — 2—10, редко больше. К концу лета
выводки распадаются.
Ценный пушной зверек.
Географическая изменчивость выражена слабо.
Реально можно различить лишь 3 подвида:
158
a) M. s. sibirica Pallas, 1773. (Описан с Алтая.) Размеры более мелкие. Окраска
как зимнего, так и летнего меха светлая, неяркая. Мех густой, мягкий. Большая часть
ареала.
b) М. s. manchurica Brass, 1911, (Описан из Северо-Восточного Китая.) Размеры
крупные. Окраска яркая, интенсивная: зимой ярко-рыжая, летом буровато-рыжая. Мех
пышный, но грубоватый. Приморье, Амурский край.
с) М. s. itatsi Temminck, 1844. (Описан из Японии.) Размеры очень крупные.
Брюхо светлее спины, лапы темные. Акклиматизирован на о-ве Сахалин.
Подрод хорьков. Subgenus Р u tori us Cuvier, 1817
38. Светлый хорек. Mustela eversmanni Lesson, 1827
Табл. XIV, 3, Карта 51
Южные зоны СССР, Западная Европа и Центральная Азия, Се-
верная граница ареала в СССР тянется примерно через Киев, Чернигов
к Калуге, а оттуда по Оке, Волге, Каме и Белой к южной оконечности
Урала; далее проходит через район г, Свердловска, южнее Тобольска,
по Иртышу к г. Таре и далее через Томск к северному концу Байкала
и по Северному Забайкалью к низовьям р, Бурей,
Наиболее характерные места обитания — открытые ландшафты
с плотными почвами. По травянистым склонам и долинам рек подни-
мается высоко в горы, где обычен по сыртам и степным плато. Живет
в норах, которые лишь изредка роет сам, обычно же приспосабливает
для своего жилья норы сусликов, хомяков и других животных, Охотит-
ся как ночью, так и днем. Питается главным образом мелкими грызу-
нами, в основном сусликами, которых ловит на поверхности земли или
чаще душит в глубине норы. Поедает также мелких птиц, их яйца и
птенцов, пресмыкающихся, насекомых, Течка протекает в феврале —
марте. Продолжительность беременности около 7 недель. Молодые,
в числе обычно 8—11, иногда и до 18, родятся в норе в конце апреля—
начале мая. Прозревают в двухнедельном возрасте. К осени выводки
распадаются,
Важный объект пушного промысла, Приносит пользу истреблением
вредных грызунов, в частности сусликов,
Географическая изменчивость выражена сравнительно резко, что позволяет вы-
делить ряд подвидов:
а) М. е. satunini Migulin, 1928. (Тип из Ногайской степи.) Мех низкйй, редкова-
тый. Окраска пуха зимнего меха на спине желтоватая, концов ости буроватая. Пе-
редняя часть спины серовато-жёлтая. Хвост наполовину темный, наполовину светлый.
Степи Европейской части СССР.
b) М. е. eversmanni Lesson, 1827. (Описан из степей между Оренбургом и Буха-
рой). Зимний мех средней пышности. Пух на спине белый или беловатый, а концы
ости черные или черно-бурые. Передняя часть спины беловатая. Черная ость на
хребте заметно скрывает светлый подшерсток. Конечная темная часть хвоста состав-
ляет примерно 7з его длины. Казахстан и юг Западной Сибири (М. е. nobilis Stroganov).
с) М. е. talassicus Ognev, 1928. (Тип с Таласского Алатау.) Мех рослый. Пух
зимнего меха на спине беловатый. Черноватые концы остевых волос невелики и лишь
слегка затемняют светлую подпушь. Передняя часть спины беловатая. Вся спина
очень светлая. Конечная темная часть хвоста составляет не более х/з его длины.
Средняя Азия.
d) М. е. michnoi Kastschenko, 1910. (Тип из окрестностей г. Троицкосавска.)
Волосяной покров зимой особо рослый, лохматый, с редкой подпушью. Остевые во-
лосы зимнего меха на спине в основной части белесые, посередине ржаво-коричневые,
на концах темно-бурые. Пух беловатый. Передняя часть спины белесая с ржавым
налетом. Хвост светлый с небольшим темно-бурым концом. Степи Забайкалья и гор-
ные степи Алтая. Очень близки М. е. lineiventer Holl. (Чуйская степь) и М. е. tuvini-
cus Stroganov (Тува).
е) М. е. amurensis Ognev, 1930. (Описан по шкуркам из района г. Благовещенска.)
Мех низкий, но плотный. Окраска хребта и боков буровато-рыжая. Пух желтоватый.
Хвост рыжеватый с небольшой черно-бурой кистью на конце. Долина среднего те-
чения Амура.
159
39. Черный хорек. Mustela putorius Linnaeus, 1758
Табл. XIV, 4. Карта 51
Почти'вся Европа, кроме ее северных районов. В Советском Союзе
распространен на север до Средней Карелии, южного берега Белого
моря, г. Сыктывкара и г. Ирбита. На востоке граница ареала тянется
по восточному склону Уральского хребта. Южная граница идет до
г. Орска, чуть севернее г. Чкалова, южнее г. Куйбышева, мимо г. Вол-
гограда к низовьям Дона и далее по южным районам Украины.
В Крыму черного хоря нет.
Основные места обитания: разреженные леса, поймы рек, заросшие
кустарником, рощи, чередующиеся с лугами и полями, лесные острова
и посадки среди степи; часто селится в селах и даже больших городах.
Активен преимущественно ночью.
Питается мелкими грызунами (главным образом полевками и мы-
шами), птицами, пресмыкающимися, лягушками, рыбой. Нередко напа-
дает на домашнюю птицу. Время течки — март — апрель. Беременность
около 40 дней. Количество детенышей в помете — от 2 до 12, обычно 4—6.
Растут молодые сравнительно медленно, достигая полного роста и по-
ловозрелости только на втором году жизни. Весенняя линька начи-
нается в марте и заканчивается в мае, а осенняя тянется с сентября
по ноябрь.
Местами наносят ущерб птицеводству, поедая домашних птиц.
Шкурки черных хорей ценятся как пушное сырье.
Географическая изменчивость выражена слабо.
Можно различить 2 подвида:
а) М. р. putorius Linnaeus, 1758. (Описан из Швеции.) Размеры более крупные.
Окраска более темная. Западные области Европейской части СССР.
b) М. р. aurea Pocock, 1936. (Тип из Татарии.) Размеры мельче, а окраска свет-
лее. Восточные области Европейской части СССР.
Нередко попадаются помеси черного хоря с норкой (так называемый хорь-тумак)
и со светлым хорем. Кроме того, в западных областях иногда попадаются хори-хро-
мисты ярко-рыжей окраски.
Подрод норок. Subgenus Lutreola Wagner, 1761
40. Европейская норка. Mustela lutreola Linnaeus, 1766
Табл. XIV, 5. Карта 52
Большая часть Европы, кроме стран Средиземноморья, Англии
и Скандинавии. В СССР встречается на севере до средних частей
Карелии, южного побережья Белого моря, южных пределов лесо-
тундры, в Архангельской области и Коми АССР. Продолжает рассе-
ляться на восток, перевалив через Уральский хребет и достигнув
р. Иртыша. На юг спускается до среднего течения р. Урала, ни-
зовьев Волги, Кавказа. В Закавказье (кроме Абхазии) и в Крыму
отсутствует.
Ведет полуводный образ жизни. Селится обычно по заросшим ку-
стами и деревьями берегам лесных речек или около прудов и озер.
Особенно часто держится на перекатах, у заброшенных плотин, близ
ключей, где вода не замерзает в зимнее время. Питается в основном
водяными крысами, полевками, мышами, лягушками, рыбой, раками,
в меньшей степени — пресмыкающимися, птицами, их яйцами и птенца-
ми, моллюсками, насекомыми. Нору устраивает в непосредственной
близости от воды; иногда ее роет сама норка, но нередко завладевает
норой водяной крысы. Известны случаи, когда гнездо устраивалось
160
в низко расположенном дупле, в трещине склона берега, под корягой.
Кроме постоянной норы, имеет обычно несколько временных убежищ.
Гон происходит во время вскрытия водоемов. Срок беременности сильно
изменчив. Детеныши (от 2 до 7) родятся в апреле — мае. В августе вы-
водки уже распадаются. Зимой нередко ведет кочевой образ жизни,
иногда передвигаясь под нависшим над берегом льдом.
Шкурки имеют значение в пушной торговле СССР. Ежегодно
в стране заготовляется до 15—20 тыс. шкурок.
По направлению с севера на юг размер норок увеличивается, пышность и мяг-
кость волосяного покрова падает, а окраска меха становится более светлой и рыже-
ватой. Можно различить 4 подвида:
а) М. 1. lutreola Linnaeus, 1766. (Описан из Финляндии.) Размеры относительно
мелкие: кондилобазальная длина черепа самцов 56—64 мм. Окраска темно-бурая
с почти черным размытым ремнем по хребту. Мех высокий, густой и мягкий. Северные
области СССР, к югу до Вологды и Горького.
b) М. 1. cylipena Matschie, 1912. (Тип из Калининградской области.) Размеры
крупнее: .кондилобазальная длина черепа самцов 59—70 мм. Окраска относительно
темная, каштановая, без рыжины. Прибалтика, Белоруссия.
с) М. 1. novikovi Ellerman et Morrison-Scott, 1951. (Тип из Московской области.)
Размеры, как у предыдущей: кондилобазальная длина черепа самцов 59—67 мм.
Окраска спины однотонная, темно-коричневая, обычно с рыжеватым оттенком. Мех
средней пышности, и мягкости. Центральные области Европейской части РСФСР.
d) М. 1. turovi Kuznetzov et Novikov, 1939. (Тип из окрестностей ст. Прохладной.)
Размеры крупные: кондилобазальная длина черепа самцов 61—68 мм. Окраска свет-
ло-коричневая или коричневая с ясным рыжим оттенком. Мех редкий и грубый. Кав-
каз, Нижнее Поволжье, Нижний Дон.
41. Американская норка. Mustela vison Brisson, 1756
Табл. XIV, 6. Карта 52
Естественный ареал охватывает Северную Америку, от северной
границы леса до Мексики. В Советский Союз завезена с целью раз-
ведения в звероводческих хозяйствах и расселения в природных усло-
виях. Успешно прижилась в ряде районов Приморья, Хабаровского
края, Читинской области,-Бурятии, Иркутской области, Красноярского
и Алтайского краев, Новосибирской, Кемеровской, Омской и Сверд-
ловской областей, Татарской АССР, Карелии, Киргизии, Грузии, Азер-
байджана и некоторых других частей СССР. В ряде этих районов чис-
ленность американской норки стала столь значительной, что ее уже до-
бывают в довольно большом количестве.
Селится в Сибири главным образом по лесным и горным речкам,
особенно близ мест, не замерзающих на зиму. Норы устраивает обычно
под нависшим берегом; нередко делает гнездо под колодиной, в валеж-
нике, в невысоком дупле, среди камней. Основная пища — мелкие гры-
зуны, рыбы, лягушки, насекомые. Гон — в феврале — марте. Беремен-
ность— от 36 до 75 дней. Количество детенышей в помете — от 1 до 12,
как исключение до 17. К концу лета молодые достигают полного роста
и выводки распадаются.
Шкурки американской норки ценятся выше, чем шкурки европей-
ского вида.
Завезенные в СССР норки приобрели некоторые отличия от исходных, и на
основе этого уже выделены новые формы:
а) М. v. tatarica Popov, 1940. (Из Татарии.)
b) М. v. altaica Ternovskii, 1958. (Тип из окрестностей Яже-ящ Горно-Алтайской
автономной области.)
П Н. А. Бобринский и др.
161
Подсемейство ВЫДР. Subfamilia LUTRINAE
РОД ВЫДРЫ. GENUS LUTRA В risson, 1762
Относительно крупные хищники (длина тела до 80 см), ведущие
полуводный образ жизни. Тело длинное, слегка уплощенное. Ноги очень
короткие, пятипалые, с короткими тупыми когтями; пальцы связаны
плавательной перепонкой. Хвост длинный, конический, сужающийся
к концу, сплюснутый сверху вниз. Голова плоская с небольшими ушны-
ми раковинами. Мех низкий, прилегающий, с очень густым, шелкови-
стым пухом и частой грубой остью. Череп уплощенный, с укороченным
лицевым отделом.
Зубная формула: I С -j- Pm у М у -2 = 36.
В СССР— один вид.
42. Выдра. Lutra lutra Linnaeus, 1758
Табл. XIV, 7. Карта 53
Евразия (кроме Крайнего Севера, Индокитая и Аравии), Марокко
и Алжир. Близкие формы водятся в Северной Америке. В Советском
Союзе выдра встречается почти всюду, но отсутствует в тундрах край-
него северо-востока Европейской части страны и северных окраин Си-
бири, а также в Крыму, в большей части Казахстана (кроме Южного
Алтая и гор Семиречья) и в пустынях Средней Азии. Почти повсеместно
редка.
Ведет ночной, скрытный образ жизни. Места обитания — берега
различных пресных водоемов, но в районе Мурманска живет и на по-
бережье моря. Превосходно плавает и ныряет. Питается в основном
рыбой, в меньшей степени — раками, лягушками, моллюсками, водными
насекомыми, а также водоплавающими птицами, их яйцами и птен-
цами, водяными крысами и другими живущими по берегам водоемов
грызунами и некоторыми прибрежными растениями. Нора устроена
обычно в откосе берега под кустом или в корнях прибрежного дерева;
вход в нее открывается под водой. Гнездо выстлано травой, листьями,
мхом. Кроме того, в пределах своего охотничьего участка выдра всегда
имеет ряд временных убежищ в виде неглубоких нор, ямок под навис-
шим берегом, под нанесенными водой стволами деревьев. Гон чаще
всего в феврале — марте — апреле. Продолжительность беременности
не установлена. Молодые, обычно 2—5, появляются чаще всего весной,
но иногда новорожденных зверьков находили в середине лета и зимой.
Выводки не распадаются до осени, а иногда и до середины зимы.
Весьма ценный пушной зверь, добыча которого ограничивается вы-
дачей особых разрешений — лицензий.
Географическая изменчивость незначительна. В СССР различают 3 подвида:
a) L. 1. lutra Linnaeus, 1758. (Описан из Швеции.) Окраска спины темно-коричне-
вая или бурая. Пух серебристо-белесый с темно-каштановыми концами волос. Брюхо
серебристое. Хвост цвета спины. Вся Европейская часть СССР, Сибирь, Дальний
Восток.
b) L. 1. meridionalis Ognev, 1931. (Тип из окрестностей г. Тегерана.) Окраска спины
светло-коричневая (цвета кофе с молоком). Пух серебристо-белесый с желтоватым от-
тенком и светло-коричневыми концами волос. Хвост цвета спины. Закавказье, Запад-
ный Иран.
с) L. 1. seistanicus Birula, 1912. (Тип из Сейстана, Иран.) Окраска светлая, серо-
вато-коричневая, брюхо желтоватое. Пух белесый с желтоватыми концами. Хвост
сверху коричневый, снизу белесый. Горы Средней Азии, Афганистана, Восточный Иран.
162
РОД КАЛАНЫ. GENUS ENHYDRA Fleming, 1822
Крупные куницеобразные хищники, наиболее специализированные
к полуводному образу жизни. Пальцы передних конечностей срослись
между собой. Задние конечности имеют вид ластов, равномерно покры-
тых мехом.
Зубная формула: 1-|- С у Рт у М. — • 2 = 32.
Один вид.
43. Калан. Enhydra lutris Linnaeus, 1758
Табл. XIV, 8. Карта 53
Когда-то был широко распространен в прибрежных водах Куриль-
ской гряды, юго-западного побережья Камчатки, Командорских остро-
вов, всей Алеутской гряды и западных берегов Северной Америки на
юг до Калифорнии. В настоящее время сохранился у некоторых остро-
вов Курильского архипелага, Командорских островов и кое-где у Але-
утских островов и побережья Северной Америки.
Характерное местообитание — прибрежная зона моря с зарослями
морской капусты и других водорослей у каменистых берегов с боль-
шими надводными и подводными камнями. Живут обычно группами
разной величины. Ночью спят на выдающихся из воды камнях, днем же
плавают и ныряют, временами отдыхая лежа на воде или вылезая на
камни. Во время бурь укрываются на берегу или на прибрежных кам-
нях. Основная пища — морские ежи, моллюски, крабы, рыба, а также,
по-видимому, морская капуста. Определенного срока спаривания нет,
но наблюдается весной. Продолжительность беременности — 8—9 меся-
цев. Самка приносит одного детеныша на берегу или на прибрежных
скалах. Но сразу после родов самка с новорожденным уходит в море.
Рост молодых животных идет медленно, полного роста они достигают
только на третьем году жизни. Самки спариваются впервые в трех-
летием возрасте.
Шкурки каланов на мировом рынке ценятся дороже шкурок всех
остальных пушных зверей. В Советском Союзе добыча калана запре-
щена законом.
И*
Н. А. Бобринский
ОТРЯД ЛАСТОНОГИЕ. ORDO PINNIPEDIA
Ластоногие — средних и крупных размеров водные хищные звери,
нуждающиеся, однако, для размножения и линьки в твердом субстрате
и длительном пребывании в воздушной среде. Тело вальковатое с тол-
стой, относительно слабо выраженной шеей. Передние и задние конеч-
ности видоизменены в плавательные ласты. На задних ластах первый
и пятый пальцы длиннее средних. Хвост короткий или недоразвитый.
Мех у большинства взрослых из грубых остевых волос и обычно очень
редкого подшерстка; лишь у морского котика подшерсток хорошо раз-
вит. У взрослых животных некоторых видов волосяной покров почти
редуцирован. Ушные раковины очень малы или отсутствуют. Глаза
обычно большие с несколько уплощенной роговицей. Головной мозг
сравнительно велик, и поверхность его полушарий изборождена много-
численными извилинами. Зубная система из резцов, клыков и коренных.
В общем она сходна с таковой наземных хищных, но зубы слабее диф-
ференцированы: коренные (включая и предкоренные) почти однород-
ны (рис. 46). Пища — рыба, моллюски, ракообразные.
Ластоногие весьма близки к наземным хищникам и, вероятно, про-
изошли от них. Многие зоологи рассматривают ластоногих лишь как
подотряд отряда хищных.
Распространены преимущественно в холодных и умеренных морях
северного и южного полушарий. В морях тропической зоны малочис-
ленны. Живут и в некоторых замкнутых водоемах (Каспий, озера
Байкал, Ладожское и Др.). Отдельные виды регулярно заплывают
в устья рек.
Некоторые виды служат важным объектом морского зверобойного
промысла и дают нашему народному хозяйству много ценного техниче-
ского жира (ворвани), кожевенного и мехового сырья.
Отряд ластоногих подразделяется на 3 семейства, представители
которых встречаются и в водах Советского Союза.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ ОТРЯДА ЛАСТОНОГИХ
1(4). Задние ласты могут подгибаться под туловище и служить
опорой при передвижении зверя по суше. Передние конечности лишь
немного короче задних. Нижняя поверхность передних и задних ластов
голдя. Когти на передних ластах очень малы или отсутствуют, а на зад-
них ластах расположены далеко от их наружного края,- Слуховые ба-
рабаны малы и неправильной формы. Носовые кости своими задними
концами не вклиниваются между лобными костями (рис. 47) ... 2
164
2(3). Ушные раковины имеются. Клыки верхних челюстей лишь
в 2—3 раза длиннее крайних резцов и не выдаются из закрытого рта.
Все зубы остроконечные. В верхней челюсти с каждой стороны по
3 резца, в нижней — по 2. Лобные кости своими острыми передними
концами вклиниваются между носовыми костями (рис. 47, Б) . . . .
................................Котиковые. Otariidae. (Стр. 166.)
3(2). Ушных раковин нет. Верхние клыки в виде огромных бивней,
Рис. 46. Череп и зубы лысуна (Phoca groenlandtca): А — сбоку;
Б — сверху; В — снизу.
Точки приложения ножек штангенциркуля при измерении черепа: а —Ь — кондило-
базальная длина; а — d — наибольшая длина черепа; е — J — мастондная ширина;
g — h — ширина мозговой коробки; k — I — высота черепа; л — о — скуловая
ширина.
Череп: Ьо — слуховой барабан; ft — предглазничное отверстие; fin — передне-
нёбное (резцовое) отверстие; fr — лобная кость; тр — задний край костного нёба;
тх — верхнечелюстная кость; nas — носовая кость; ос — затылочная кость; par — те-
менная кость; ртх— предчелюстная кость; pt — крыловидный отросток; z — скуловая
кость.
Зубы: /3 — третий (внешний) верхний резец; i’, i2 — нижние резцы; С, с — клыки;
Рт\ Рт2, pmlt рт3 — предкоренные; М, т — заднекоренные зубы.
165
далеко выдающихся из закрытого рта. Жевательные поверхности рез-
цов и коренных зубов плоские. В верхней челюсти с каждой стороны
по 1 или по 2 резца, в нижней — резцы отсутствуют. Лобные кости не
вклиниваются между носовыми костями (рис. 47, А).....................
...............................Моржовые. Odobenidae. (Стр. 168.)
4(1). Задние ласты не могут подгибаться под туловище и всегда
направлены назад. Передние ласты значительно короче задних. Как
передние, так и задние ласты сверху и снизу покрыты мехом. Когти
передних ластов хорошо развиты (исключение — тюлень-монах). Слу-
ховые. барабаны развиты хорошо, грушевидной формы. Носовые кости
своими задними концами вклиниваются между лобными костями
(рис. 47, В).........................Тюленевые. Phocidae. (Стр. 169.)
Рис. 47. Передние части черепов ластоногих (вид сверху):
А — морж (Odobenus rosmarus); Б — сивуч (Eumetopias jubata): В — тюлень-монах (Monachus
monachus); fr — лобная кость; nas — носовая кость. Сильно уменьшены.
СЕМЕЙСТВО КОТИКОВЫЕ. FAMILIA OTARIIDAE
Крупные морские звери с торпедообразным телом, относительно,
длинной подвижной шеей, небольшой головой с маленькими ушными
раковинами. Ласты большие, голые снизу и частично сверху. Когти на
задних ластах малы и сидят далеко от их внешнего края. Волосяной
покров хорошо развит, обычно с наличием подшерстка (у сивучей очень
редкого) . Самцы намного крупнее самок.
Череп тонкостенный, вытянутый. Зубы остроконечные. Клыки не
превращены в выдающиеся изо рта бивни.
Зубная формула: I СРш у М-Ц^-2 =34 или 36.
В водах СССР — 2 рода.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ КОТИКОВЫХ ВОД СССР
1(2). На передних ластах вся кисть сверху голая. Подшерсток
очень густой. Межчелюстные кости в середине резко сужены, а перед
вершиной сильно расширены (рис. 48, Б). Расстояние между послед-
166
ним предкоренным и первым коренным зубами верхней челюсти мень-
ше длины первого из этих зубов.....................................
............................Морские котики. Callorhinus. (Стр. 167.)
2(1). На передних ластах шерсть покрывает сверху значительную
часть кистей. Подшерстка почти нет. Межчелюстные кости постепенно
суживаются кверху (рис. 48, Л). Последний предкоренной зуб верхней
челюсти отделен от первого коренного промежутком, величина которого
больше длины первого из этих зубов. Сивучи. Eumetopias. (Стр. 168.)
черепов
Рис. 48. Носовые отделы
котиковых:
Л — сивуч (Eumetopias jubata); Б —
морской котик (Callorhinus ursinus);
ртх — межчелюстная кость.
РОД морские котики.
GENUS CALLORHINUS Gray, 1859
Длина тела взрослых самцов до 2 ж,
самок до 1,5 м. Кондилобазальная дли-
на черепа самцов 22—24 см. Мех из гру-
бой ости и очень густого, хотя и невысо-
кого подшерстка: у взрослых самцов во-
лосы на шее удлинены в своеобразную
«гриву». Кисти передних ластов сверху
голые. Окраска взрослых самцов серо-
вато-бурая, самок коричневатая, моло-
дых (сеголеток) сероватая, новорожден-
ных черная (эту черную окраску моло-
дые сохраняют около трех месяцев).
Один вид.
1. Морской котик. Callorhinus
ursinus Linnaeus, 1758
Табл. XV, 1—3. Карта 54
Северная часть Тихого океана и его
моря: Берингово, Охотское (у Курил) и
Японское. Основные лежбища в настоящее время расположены на
Командорских островах, на островах средней части Курильской гряды,
на о-ве Тюленьем, в заливе Терпения, о-ве Сахалин и на Прибыловых
островах (принадлежащих США). На зиму совершают далекие мигра-
ции к югу: котики Командорских и Курильских островов проводят
зиму в водах Тихого океана, к востоку от Японии и в Японском море.
Пища — различная рыба, отчасти головоногие. В конце весны
взрослые самцы-секачи выходят на лежбища и занимают отдельные
участки, из-за которых дерутся с другими самцами, оттесняя слабых и
не достигших полного развития самцов (полусекачей) на края детного
лежбища. Через 10—20 дней на лежбище выходят самки и неполово-
зрелые самцы (так называемые холостяки); последние залегают от-
дельно, образуя особые, «холостяковые» лежбища. Самки немедленно
захватываются секачами в «гаремы». Спустя 1—3 дня самки родят по
одному детенышу, а через несколько дней вновь оплодотворяются сека-
чами. Продолжительность беременности маток — около года. Во время
«гаремного» периода, который длится до августа, секачи ничего не едят
и почти не спускаются в воду. В начале августа «гаремы» начинают
распадаться, а спустя некоторое время секачи уходят в море. В сен-
тябре— октябре лежбища покидают и самки с молодыми, которые до
того питались молоком матери. С середины августа до середины сен-
тября волосяной покров котиков сменяется, причем черные котики
превращаются в серых. Поздней осенью лежбища пустеют. Самки
167
становятся половозрелыми к двум годам, самцы — к трем, но обычно
самки оплодотворяются лишь на третьем году жизни, а самцы обзаво-
дятся «гаремами» только в возрасте 5—6 лет, так как более молодые
отгоняются от самок мощными секачами.
Шкуры, особенно молодых самцов-холостяков, высоко ценятся как
меховое сырье; при их выделке грубые остевые волосы удаляют, а ос-
тавшийся очень густой, бархатистый пуховой покров обычно красят
в черный цвет.
До последних лет считалось, что котики Командорских островов
отличаются от животных с Курил и о-ва Тюлений окраской подшерстка
и некоторыми другими признаками. Но ныне доказано, что сколько-ни-
будь заметных отличий между котиками обоих указанных стад нет.
Все они относятся к номинальной форме: С, u. ursinus Linnaeus, 1758
(описан по экземпляру с о-ва Беринга) .
РОД СИВУЧИ, GENUS EUMETOPIAS Gill, 1866
Огромные морские звери, самцы которых достигают длины 3,5 м
п веса 1 т, длина самки 2—2,6 м и вес до 350 кг. Кондилобазальная
длина черепа самцов 37—40 см. Тело взрослых особей покрыто жест-
ким, прилегающим к коже волосяным покровом, почти без подшерстка.
Волосами покрыта и верхняя сторона передних ластов, кроме их внеш-
него края. Окраска сивучей варьирует от соломенно-желтой до темно-
шоколадной.
Один вид,-
2. Сивуч, Eumetopias jubata Schreber, 1776
Табл, XV, 4, Карта 54
Распространен в северной части Тихого океана и его морей: Бе-
рингова, Охотского и Японского, В наших водах лежбища расположены
на Командорских островах, местами на побережье Камчатки, на ряде
островов Курильской гряды, кое-где по берегам Сахалина и на распо-
ложенном близ него о-ве Тюленьем, на о-ве Ионы в Охотском море,, на
Ямских островах, на скалах побережья некоторых частей Приморья.
Питается рыбой, крупными головоногими моллюсками и другими
морскими животными, В погоне за ними сивучи ныряют на большую
глубину. Заглатывают по нескольку килограммов камней. Зиму прово-
дят в море, кочуя поодиночке или стаями. Весной, в мае, взрослые
самцы-секачи выходят на лежбища, расположенные на каменистых бе-
регах или на выступающих из воды скалах. Через 2—3 недели к леж-
бищам подходят самки. Секачи собирают около себя «гаремы» из 10—
20 самок, из-за которых между самцами происходят жестокие драки.
Вскоре после .выхода на сушу самки родят по одному детенышу, а че-
рез несколько дней вновь оплодотворяются. Следовательно, беремен-
ность самки продолжается около года. Самцы становятся половозре-
лыми, по-видимому, только с пятилетнего возраста, а самки способны
к оплодотворению лишь в возрасте трех лет.
Промышляются в небольшом 'количестве местным населением ради
шкуры и сала,
СЕМЕЙСТВО МОРЖОВЫЕ. FAMILIA ODOBENIDAE
Огромные морские звери, обитатели Арктики, Длина тела самцов
до 4,5 X 'Самок до 3,5 м. Вес самцов до 1,5 т. Кожа очень толстая, об-
разующая складки. Молодые покрыты низким, редким, буроватым ме-
хом, у взрослых волосяной покров становится совсем редким, а у ста-
168
Рис. 49. Череп мор-
жа (Odobenus ros-
marus) сбоку.
рых особей почти исчезает. Усы очень толстые, густые, свисающие
с верхней губы. Ушных раковин нет. У взрослых моржей верхние клыки
далеко выдаются из закрытого рта. Коронки резцов и 'коренных зубов
имеют плоскую жевательную поверхность. Череп очень массивный:
кондилобазальная длина его у самцов равна 36—
43 см, у самок — 30—36 см. Передний отдел черепа
очень широк в силу огромных альвеол клыков.
Один род.
РОД МОРЖИ. GENUS ODOBENUS Brisson, 1762
Один ВИД.
3. Морж. Odobenus rosmarus Linnaeus, 1758
Табл. XV, 5, 6. Карта 54
Северный Ледовитый океан. Вдоль Атлантиче-
ского побережья Америки спускается к югу до
Лабрадора, изредка льдами заносится к Ислан-
дии; прежде был нередок близ Шпицбергена и
у Земли Франца-Иосифа, но теперь там стал ма-
лочисленным. Встречается у Новой Земли, в Крас-
ном мо-ре, у о-ва Северная Земля, в море Лапте-
вых. Всюду относительно редок. Численность его
выше в Чукотском море и северной части Берин-
гова моря; как исключение встречается здесь
к югу от 60°.
Моржи — обитатели прибрежных и мелковод-
ных участков моря, изобилующих донными живот-
ными: двустворчатыми и брюхоногими моллюска-
ми, червями, ракообразными, составляющими их
основную пищу. Держатся обычно стадами. Охотно
выходят на берег или на льдины, где ложатся отдыхать (плотными
группами по нескольку десятков голов. На зиму моржи откочевывают
в более южные части Арктики, не столь сильно закрытые льдами. Де-
торождение обычно в мае — июне, в некоторых районах — в апреле.
Роды протекают на отмелях или на льдинах. Самка '.кормит молоком
своего единственного детеныша два года. Обычно молодой держится
при матери три года.
На Чукотке моржей добывают ради мяса, сала и кож. Бивни ис-
пользуют в косторезном деле.
Различают 2 подвида:
а) О. г. rosinarum Linnaeus, 1758. (По экземпляру из Северной Атлантики.) Кон-
дилобазальная длина черепа самцов 36—38 см. Бивни относительно короткие и тонкие;
у взрослых ширина носового отдела черепа у клыков равна 60—64% ширины черепной
коробки у слуховых проходов. Западный сектор Арктики, от Канадского архипелага
до Новосибирских островов.
Ь) О. г. divergens liliger, 1815. (По экземпляру близ мыса Айс, Северная Аляска.)
Размеры крупнее: кондилобазальная длина черепа самцов 38—43 см. Бивни взрослых
очень длинны и толсты; у взрослых ширина носового отдела черепа у клыков равна
65—89% ширины черепной коробки у слуховых проходов. Восточный сектор Арктики,
от Новосибирских островов до мыса Барро.
СЕМЕЙСТВО ТЮЛЕНЕВЫЕ. FAMILIA PHOCIDAE
Мелкие и среднего размера ластоногие с вальковатым телом и зад-
ними ластами, неспособными подгибаться под тело. Передние конеч-
ности значительно короче задних. Когти на задних ластах хорошо
169
развиты и расположены на их концах. Волосяной покров взрослых
и полувзрослых низкий, прилегающий, состоящий из остевых волос,
почти без подшерстка. Детеныши покрыты густым и пышным мехом.
Общая длина черепа до 30 см. Носовые кости вклиниваются задними
концами между лобными (рис. 47, В).
Распространены почти по всем морям и океанам, хотя в тропиче-
ских морях численность их невысокая. Живут и в некоторых внутрен-
них бассейнах (Каспийское море, озера Байкал, Ладожское и др.).
Всего около 10 родов; в водах СССР — 5 родов. Они группируются
в 3 подсемейства: 1) настоящих тюленей, 2) тюленей-монахов и 3) хо-
хлачей.
предчелюстных
Рис. 50. Носовой
отдел черепа хох-
лача (Cystophora
cristat а):
nas — носовая кость;
ртх, — предчелюст-
ная кость.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ТЮЛЕНЕВЫХ ФАУНЫ СССР
1(8). Верхняя сторона морды гладкая, без растяжимого кожистого
мешка. В нижней челюсти с каждой стороны по 2 резца. Отростки
эстей достигают носовых костей................................2
2(7). Конец носа голый или почти голый. Когти
как на передних, так и на задних ластах хорошо раз-
виты. В верхней челюсти с каждой стороны по 3
резца..........................................3
3(6). Морда относительно короткая: расстояние
между концом носа и глазом значительно меньше уд-
военного расстояния от глаза до слухового отверстия.
Высота носового отдела черепа значительно ниже моз-
гового отдела................................. 4
4(5). Третий палец передних ластов короче пер-
вого и второго. «Усы» редкие; волосы их неровные:
с расширениями и перехватами (рис. 51, А). Длина
тела не более 2,2 м. У самки только 2 соска. Скуловая
кость длинная и узкая, так что высота ее в средней
части укладывается в ее длине более двух раз
(рис. 52, Б). Зубы сохраняются в течение всей жизни.
............Собственно тюлени. Phoca. (Стр. 171.)
5(4). Третий палец передних ластов самый длин-
ный. «Усы» густые; волосы их гладкие, равномерно
утончающиеся к концу (рис. 51, Б). Длина взрослых
более 2 м. У самки 4 соска. Скуловая кость короткая
и широкая, так что высота ее в средней части укла-
дывается в ее длине не более двух раз (рис. 52, А),
часто не сохраняются...........................
...........................Морские зайцы. Erygnathus. (Стр. 177.)
6(3). Морда очень длинная: расстояние между концом носа и глазом
почти в 2 раза больше расстояния между глазом и слуховым отверстием.
Носовой и мозговой отделы черепа одинаковой высоты.............
.....................Длинномордые тюлени. Halichoerus. (Стр. 178.)
7(2). Конец морды до самых ноздрей покрыт волосами. Когти раз-
виты слабо, на задних ластах нередко отсутствуют. В верхней челюсти
с каждой стороны только по 2 резца.............................
.............................Тюлени-монахи. Monachus. (Стр. 178.)
8 (1). На верхней стороне морды большой растяжимый мешок,
который может раздуваться, достигая у самцов огромных размеров.
В нижней челюсти с каждой стороны лишь по 1 резцу. Отростки меж-
челюстных костей далеко не достигают носовых костей (рис. 50) . . .
.................................Хохлачи. Cystophora. (Стр. 179.)
Зубы у взрослых
170
Рис. 51. Вибриссы тюленей:
А—нерпы (Phoca); Б—морского зайца (Erignathus barbatus).
РОД ТЮЛЕНИ. GENUS PHOCA Linnaeus, 1758
Зубная формула:
I — С —Pm —М—-2 = 34.
2 1 4 1
Зубы сохраняются в течение всей жиз-
ни. Коренные, пока не стерты, с хорошо за-
метными добавочными вершинами.
Арктические и умеренные моря север-
ного полушария (в Средиземном и Черном
морях отсутствуют).
Всего 6 видов, группируемых в 4 под-
рода (Pagophoca, Histriophoca, Phoca и
Pusa); все 6 видов представлены в фауне
СССР.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ
СОБСТВЕННО ТЮЛЕНЕЙ
1(4). Окраска взрослых светлая с боль-
шими черными или бурыми полями на бо-
ках. Костная носовая перегородка доходит
или почти доходит до заднего края костного
нёба. Задний край костного нёба образует
почти прямую линию или пологую двойную
дугу (рис. 53, Б)......................2
2(3). Передние ласты и прилегающие
части тела светлые; тело иногда покрыто
мелкими темными пятнами. Кондилобазаль-
ная длина черепа взрослых более 200 мм.
Костная носовая перегородка доходит до
заднего края костного нёба. Зубной край
верхней челюсти не изогнут...............
Лысун. Phoca groenlandica. (Стр. 173.)
Рис. 52. Скуловая дуга:
А — морского зайца (Erigna-
thus barbatus): Б — обыкновен-
ного тюленя (Phoca vitulina);
tnx — скуловой отросток верхне-
челюстной кости: t — скуловой
отросток теменной кости; z —
скуловая кость.
А
Рис. 53. Задняя часть
костного нёба снизу:
Д — обыкновенный тюлень
(Phoca vitulina); Б — лысун
(Phoca groenlandica).
171
3(2). Передние ласты и прилегающие части тела темные; мелких
темных пятен на теле не бывает. Кондилобазальная длина черепа ме-
нее 200 мм. Костная носовая перегородка немного не доходит до зад-
него края костного нёба. Зубной край верхней челюсти изогнут в гори-
зонтальной и вертикальной плоскостях.............................
...........................Крылатка. Phoca fasciata. (Стр. 174.)
4(1). Окраска обычно мелкопятнистая, реже однотонно темная,
сверху постепенно светлеющая к брюшной стороне.
Костная носовая перегородка далеко не доходит до заднего края
костного нёба. Задний край костного нёба имеет вид круглой дуги,
обычно с заостренной вершиной (рис. 53, Л).....................5
Рис. 54. Черепа тюленей:
Д — обыкновенный тюлень (Phoca vitulina); Б—кольчатая нерпа (Phoca hispida).
Вид сверху.
5(6). Скуловая ширина взрослых более 120 мм. Ширина межглаз-
ничного промежутка в 2—2,5 раза превосходит наибольший поперечник
предглазничного отверстия. У взрослых на верхней поверхности мозго-
вой коробки хорошо развит продольный гребень. Окраска взрослых
обычно желтовато-оливковая с рисунком из мелких (некольчатых) тем-
ных пятен .... Обыкновенный тюлень. Phoca vitulina. (Стр. 174.)
6(5). Скуловая ширина черепа менее 120 мм. Ширина межглазнич-
ного промежутка меньше или равна наибольшему поперечнику под-
глазничного отверстия. На верхней поверхности мозговой коробки про-
дольного гребня нет. Окраска взрослых иная: крупнопятнистая, коль-
чато-пятнистая или одноцветная.................................7
7(10). Окраска взрослых (за редким исключением) пятнистая или
со светлыми прожилками. Ширина когтей передних ластов значительно
меньше промежутков между ними. Верхняя поверхность их не образует
ясно выраженного гребня. Скуловые дуги расставлены узко, так, что
при взгляде на череп сзади они не видны, будучи скрыты черепной ко-
робкой ...................................................... 8
8(9). По основному темному фону, цвет которого сильно варьирует,
узор из кольчатых пятен или светлых прожилок. Подглазничное отвер-
172
а б
Рис. 55. Передние ласты
нерп:
Д — байкальской (Phoca
sibirica); Б — каспийской
(Phoca caspica);, а и б —
когти в поперечном разрезе.
стие равно альвеоле клыка. Длина слуховых барабанов значительно
больше расстояния между их внутренними краями...................
......................Кольчатая нерпа. Phoca hispida. (Стр. 175.)
9(8). По светлому, обычно желтому фону узор из больших рас-
плывчатых темных пятен, образующих мраморный рисунок. Иногда
попадаются одноцветные особи соломенно-жел-
того цвета. Подглазничное отверстие шире аль-
веолы клыка. Длина слуховых барабанов значи-
тельно меньше расстояния между их внутрен-
ними краями. Когти без ребра на верхней сто-
роне (рис. 55, Б)..........................
Каспийский тюлень. Phoca caspica. (Стр. 176.)
10(7). Окраска взрослых одноцветная, без
пятен, обычно серо-стального цвета, светлеющая
на брюхе. Ширина когтей передних ластов по-
чти не уступает ширине промежутков между
ними. Верхняя поверхность их с ясно выражен-
ными продольными гребнями (рис. 55, Л). Ску-
ловые дуги расставлены широко, так, что при
взгляде на череп сзади они хорошо видны . . .
Байкальская нерпа. Phoca sibirica. (Стр. 177.)
Подрод гренландских тюленей. Subgenus Pagophoca
Trouessart, 1904
4. Лысун. Phoca groenlandica Erxleben, 1777
Табл. XVI, 1—4. Карта 56
Самый крупный представитель рода. Длина тела взрослых самцов
1,7—2 ju, самок 1,5—1,8 м. Вес 100—160 кг. Кондилобазальная длина
черепа 20—24 см; скуловая ширина 10—13 см. Взрослый самец (в воз-
расте более четырех лет) имеет беловатую окраску с черным пятном на
конце морды и большими полулунными черноватыми пятнами на боках
тела, сходящимися своими передними концами на плечах («крылан»).
Окраска половозрелых самок в зависимости от возраста трех типов:
1) пепельно-серая с темными неясными пятнами; спина темнее брюха;
2) светло-серая, более темная снизу; на боках расплывчатые пятна,
образующие темно-серые* поля полулунной формы; 3) белесая с темно-
коричневыми или черноватыми полулунными полями на боках. Неполо-
возрелые осо'би (в возрасте 2—4 лет) пепельно-серые с неясным рисун-
ком из более темных пятен. Новорожденные (бельки) покрыты густой
пышной шерстью белого или желтоватого цвета.
Населяют арктические воды между Канадским архипелагом и Се-
верной Землей. Образуют три стада: 1) ньюфаундлендское, населяющее
воды между Канадским архипелагом и Гренландией; в период размно-
жения и линьки концентрируется в районе о-ва Ньюфаундленд; 2) ян-
майенское — в области между Гренландией, Исландией и Шпицберге-
ном; размножается на льдах у о. Ян-Майен; 3) беломорское — между
Шпицбергеном и Северной Землей; для размножения собирается
в Горле Белого моря и отчасти в Чешской губе.
Основная пища — пелагические ракообразные, пелагические рыбы
(сайка, мойва, сельдь и др.) и моллюски. Летом держатся рассеянными
стадами, преимущественно у кромки льда. К зиме скапливаются в боль-
шом количестве в местах щенки. В водах СССР основные лежбища
лысуна расположены в Горле Белого моря, у побережья Канина и
173
в Чешслой губе, на льду, у трещим и по краям полыней. Молодые
(1—2) родятся во второй половине февраля — начале марта. В течение
примерно недели они сохраняют свой белый пышный мех, не сходят
в воду и питаются лишь молоком матери. Сами 'матки в это время ни-
чего не едят и держатся около детенышей. По окончании кормления
молоком они вновь оплодотворяются; следовательно, беременность их
длится около одиннадцати с половиной месяцев. С марта массы зверей
разного пола и возраста скопляются на линных залежках (тоже на
льду). Перелиняв, тюлени откочевывают на север.
Важный промысловый вид, добыча которого дает ценные техниче-
ские жиры, кожевенные и 'меховые шкуры; мясо частично используется
в корм зверей, разводимых в звероводческих хозяйствах. Особенно це-
нятся шкурки новорожденных детенышей ('бельков), обладающих пыш-
ным и мягким белым мехом, который можно красить в самые различ-
ные цвета.
В водах СССР водится наиболее крупная форма лысуна: Ph. gr. oceanica Lepechin,
1778. (Описана по экземпляру с Белого моря.)
Подрод крылаток. Subgenus Histriphoca Gill, 1873
5. Крылатка. Phoca fasciata Zimmermann, 1783
Табл. XVI, 5. Карта 56
Размеры средние: длина тела 1,5—1,7 ж; кондилобазальная длина
черепа 18—20 см. Размеры самцов и самок почти одинаковы. Окраска
взрослого самца черная с широкими беловатыми полосами вокруг зад-
ней части головы, передних ластов и крестца. Взрослые самки окра-
шены сходно, но основной тон их более светлый, буровато-серый. Гра-
ницы светлых полос не столь резкие. Неполовозрелые особи почти од-
ноцветные, буровато-серые, новорожденные белые.
Северная часть Тихого океана; через Берингов пролив заходит
в Северный Ледовитый океан. В водах СССР встречается в Татарском
проливе, Охотском, Беринговом и Чукотском морях, проникает в Во-
сточно-Сибирское море; водится и у Курильских островов. Повсеместно
весьма редка. Совершает большие миграции. Основная пища — рыба и
головоногие моллюски. Щенение происходит на льду в марте и
апреле.
Промысловое значение невелико.
Подрод нерп. Subgenus Phoca Linnaeus, 1758
6. Обыкновенный тюлень. Phoca vitulina Linnaeus, 1758
(на Дальнем Востоке — ларга)
Табл. XVI, 6, 7. Карта 55
Средних размеров: длина тела 112—205 см. Самцы несколько круп-
нее самок. По окраске самцы, самки и молодые (после смены белого
первичного меха) различаются сравнительно мало, но индивидуальная
изменчивость расцветки меха очень велика. Обычно основной фон жел-
товато-палевый; по нему разбросаны многочисленные мелкие сплош-
ные черноватые пятна, особенно четкие на спине. Иногда этих пятен
бывает так много, что спина делается почти черной со светлым сетча-
тым узором. Чаще темные пятна весьма малочисленны и расположены
174
только на спине; иногда наряду с темными ’оплошными пятнами
имеются светлые кольчатые.
Встречаются в северных частях Атлантического и Тихого океанов,
заходят в Северный Ледовитый океан. В Северной Атлантике — при-
брежные воды американского побережья от Северной Каролины до
Канадского архипелага, вокруг Гренландии, у берегов Западной Ев-
ропы, заходя в Балтийское и Баренцево моря. В северной части Тихого
океана на азиатском побережье встречается от Японии и Приморья до
Чукотки, а на американском берегу — от Калифорнии до Аляски.
Прибрежное животное, избегающее открытого' моря. Охотно вы-
ходит на берег, подолгу лежит на отмелях и прибрежных камнях.
Ведет относительно оседлый образ жизни, совершая недалекие переко-
чевки, связанные с миграциями косяков рыб, которыми главным обра-
разом питается. В погоне за стаями рыб нередко заходит в низовья
рек. Держится поодиночке, парами или небольшими табунками, не об-
разуя массовых детных и линных залежей. Только на Дальнем Востоке
осенью иногда скопляется в некоторых местах большими стаями. Био-
логия размножения тюленей, обитающих у берегов Европы и Дальнего
Востока, различна. Европейские особи спариваются в начале осени.
Самки приносят детеныша в первой половине лета на берегу. Детеныши
покрыты прилегающим серым мехом. Дальневосточные (ларга) ще-
нятся в конце зимы или в начале весны на льду. Детеныш родится
покрытым пышным белым мехом, который быстро сменяется на серый
низкий вторичный покров.
В настоящее время добывается на Дальнем Востоке в сравни-
тельно небольшом количестве, хотя запасы его там велики.
Образует 2 резко обособленных подвида:
a) Ph. v. vitulina Linnaeus, 1758. (Описан с берегов Западной Европы.) Размеры
относительно мелкие: длина тела до 180 см; кондилобазальная длина черепа до 22 см.
Зубы сближены друг с другом. Окраска тусклая, обычно мелкопятнистая, особи
с кольчатыми пятнами встречаются очень редко. Северная Атлантика. В СССР — Бал-
тийское и Баренцево моря.
b) Ph. v. largha Pallas, 1811. (Описан с побережья Камчатки.) Размеры крупные:
длина тела до 220 см; кондилобазальная длина черепа до 25 см. Зубы расставлены
более широко. Окраска яркая, с преобладанием желтого или желто-серого цвета. Чер-
ные пятна более часты и крупны; иногда они кольчатые. Побережье Дальнего Во-
стока.
7. Кольчатая нерпа. Phoca hispida Schreber, 1775
(на Дальнем Востоке—акиба)
Табл. XVI, 8. Карта 55
Размеры мелкие: длина тела взрослых 1 —1,5 м; кондилобазальная
длина черепа 15—19 см. Окраска сильно варьирует. Она обычно тем-
ная с узором из более светлых кольчатых пятен или прожилок. Попа-
даются очень темные особи, почти черные сверху и темно-серые снизу,
пятна плохо заметны. Но бывают особи и светлой желтоватой окраски
с неясными пятнами на спине.
Северный Ледовитый океан, северные части Атлантического и Ти-
хого океанов. В СССР — Балтийское море (заходит в р. Неву), оз. Ла-
дожское, по всему северному и дальневосточному побережью.
Населяет преимущественно прибрежные области морей, но с дрей-
фующими льдами нередко заносится далеко в открытое 'море. Особенно
охотно придерживается бухт, заливов, проливов, устьев рек. Длитель-
ных и массовых перемещений не совершает, и кочевки носят ограни-
ченные размеры: они связаны с передвижением льдов и наличием пищи.
175
Держатся обычно поодиночке или небольшими группами и распреде-
ляются по побережью относительно равномерно, без скоплений. Пища
разнообразна: мелкая рыба (в северных морях в основном сайка),
планктонные ракообразные, некоторые донные морские животные. Спа-
ривание на льду, по-видимому, в апреле—июне. Щенение — в начале
полярной весны (в Баренцевом море — в начале апреля). Детеныши ро-
дятся на льду, нередко в небольшой снежной норе среди торосов. Но-
ворожденный покрыт пышным белым мехом.
Важный промысловый зверь Арктики и Дальнего Востока. Дает
хорошую ворвань, кожевенные и меховые шкуры. Мясо идет в корм
собакам и пушным зверям на зверофермах, а также для подкормки ди-
ких песцов.
Описано значительное количество подвидов, обитающих в водах СССР, но ре-
ально различаются лишь 3:
a) Ph. h. botnica Gmelin, 1788. (Описан из Ботнического залива Балтийского моря.)
Размеры средние: кондилобазальная длина черепа 16—19 см. Преобладают особи
с темной, черновато-бурой окраской спины и оливково-бурым брюхом. Балтийское
море, Ладожское озеро (Ph. h. b. ladogensis Nordguist, 1899).
b) Ph. h. pomororum Smirnov, 1929. (Тип с западного берега Новой Земли.) Раз-
меры немного крупнее: кондилобазальная длина черепа 17—20 см. Окраска обычно
светлая: преобладают особи оливково-буроватые сверху и желтые снизу, с узором
из пятен, образованных буроватым серединным ядром и желтоватой каймой. Север-
ное побережье СССР, от Мурмана до Чукотки.
с) Ph. h. ochotensis Pallas, 1811, акиба. (Описан из северной части Охотского
моря.) Размеры, как у балтийской формы: кондилобазальная длина черепа 16—18 см.
Окраска взрослых обычно светлая, сероватая, со светлой белесой кольчатой пятни-
стостью, степень развития которой различна. Дальневосточное побережье СССР.
8^ Каспийский тюлень. Phoca caspica Gmelin, 1778
Табл. XVI, 9. Карта 55
Близок к кольчатой нерпе. Длина тела взрослых самцов 125—
148 см, самок 120—140 см. Кондилобазальная длина черепа самцов
18—19 см, самок 17—18 см. Окраска особей обоих полов одинакова,
но сильно варьирует индивидуально и изменяется с возрастом. Взрос-
лые обычно имеют соломенно-желтый или оливково-желтый основной
фон, по которому разбросаны сплошные расплывчатые пятна. По-
падаются одноцветные (желтяки) и особи, у которых между частыми
и большими пятнами основной светлый фон выступает в виде мрамор-
ного кольчатого узора.
Распространен по всему Каспийскому морю, но в разное время
года концентрируется в различных его частях. Летом держится неболь-
шими косяками, преимущественно в средней и южной частях Каспия.
С осени начинает массовое переселение в северную и особенно северо-
восточную часть моря, где на льдах образует детные залежки. Здесь
в январе (частично в декабре или в феврале) самка приносит одного
одетого в пышный белый 'мех детеныша (белька). Кормление молоком
длится около месяца. К этому времени заканчивается смена белого
меха детеныша на низкий серый вторичный покров и белек превра-
щается в «сиваря». Новое спаривание происходит в конце февраля или
в первой половине марта. После того как молодняк сойдет в воду,
взрослые и полувзрослые особи переходят на линные лежбища. После
линьки (весной) основная масса зверей откочевывает к югу. Питается
преимущественно мелкой рыбой, ракообразными и моллюсками.
Промысел имеет большое хозяйственное значение. Добыча зверя
ведется главным образом на лежбищах.
176
9. Байкальская нерпа. Phoca sibirica Gmelin, 1788
Табл. XVI, 10. Карта 55
Близка к кольчатой нерпе. Длина тела взрослых от 125—140 до
160 см. Кондилобазальная длина черепа 16—20 см. Спина обычно од-
ноцветная, оливково-серая или буровато-серебристо-серая, бока и
брюхо светлее и желтее, пятна, как правило, отсутствуют. Молодые
серебристо-серые, новорожденные желтовато-белые.
Распространена по всему Байкалу, но более многочисленна в его
северной части, особенно зимой.
Питается рыбой, .преимущественно бычками и голомянкой. Взрос-
лые самцы зимуют рассеянно по всему Байкалу, неполовозрелые особи
обоих полов обычно держатся зимой у западного побережья, а взрос-
лые самки — у восточного берега. Всю зиму (кроме 'беременных самок)
проводят в воде, пользуясь для дыхания продушинами, которые де-
лают еще в тонком льду. Беременные самки выходят на лед и устраи-
вают под снегом с продушиной логово, в котором приносят в феврале—
марте детеныша. Массовых детных лежек не образует. Кормление мо-
локом— 1,5—3 месяца. Весной, >с апреля, все звери, кроме кормящих
самок, выходят на лед, образуя скопления до 200 голов. Течка и линька
протекают в мае и июне. В начале лета, после распада ледяного по-
крова, сосредоточиваются главным образом в северной части Байкала,
куда ветер наносит плавающие льдины. С конца июня образуют летние
залежки близ берегов на выступающих из воды камнях. С образова-
нием ледяного покрова залежки -распадаются.
Служит объектом промысла, но количество добываемых зверей
невелико.
РОД МОРСКИЕ ЗАЙЦЫ. GENUS ER1GNATHUS Gill, 1866
Один вид.
10. Морской заяц. Erignathus barbatus Erxleben, 1777
Табл. XVI, 11. Карта 56
Крупный тюлень: длина тела взрослых самцов 2,5—3 м, самок
2—2,5 м. Вес первых до 350 кг. Кондилобазальная длина черепа сам-
цов 21—23 см, самок 19—22 см. Окраска взрослых одноцветная, буро-
вато-серая, более темная на спине, у самцов и самок одинаковая. У мо-
лодых на спине иногда выступают едва заметные темные округлые
пятна. Новорожденные покрыты коричневым или бурым мехом.
Зубная формула: I -|-С -у Pm М у • 2 = 34.
Распространен круглополярно. В Атлантическом океане — к югу до
Ньюфаундленда и Англии. В водах Тихого океана — к югу до Японии
и Канады. В водах СССР обитает по всему северному побережью
страны и у островов Берингова и Охотского морей на юг до Татарского
пролива у берегов Сахалина.
Придерживается главным образом мелководных областей и при-
брежной зоны. Иногда заходит в реки на несколько километров. Ведет
одиночную жизнь, но на Дальнем Востоке иногда осенью собирается
стаями на определенных лежбищах. Миграции выражены слабо и но-
сят местный характер. Питается в основном донными беспозвоноч-
12 Н. А. Бобринский и др.
177
ними: ракообразными, моллюсками, иглокожими, иногда рыбой. Ще-
нится на льду весной. Линяет летом.
В связи с малочисленностью и разрозненностью распределения
промысловое значение невелико.
Выделяют 2 слабо различающиеся географические формы:
а) Е. b. barbatus Erxleben, 1777. (Описана по экземпляру из Южной Гренландии.)
Распространена к западу от Новосибирских островов.
b) Е. b. nauticus Pallas, 1811. (Описана по экземпляру из Охотского моря.) Оби-
тает к востоку от Новосибирских островов и в Тихом океане. Череп этой формы от-
личается более массивной и укороченной носовой частью.
РОД ДЛИННОМОРДЫЕ ТЮЛЕНИ. GENUS HALICHOERUS Nilsson, 1820
Один вид.
11. Длинномордый тюлень (тевяк). Halichoerus grypus Fabricius, 1791
Табл. XVI, 12. Карта 56
Размеры крупные: длина тела самцов до 3,3 м. Бес до 300 кг.
Самки несколько мельче. Общая длина черепа 24—32 см. Бросается в
глаза удлиненная морда. Окраска очень изменчива. У самцов верх го-
ловы и тела обычно светло-серый, низ тела белый; по всему туловищу
разбросаны круглые черные пятна, на голове и ластах расположены
мелкие редкие пятнышки. Значительно реже попадаются самцы-почти
черного цвета или, напротив, светлой окраски со светло-серым и корич-
неватым брюхом; по плечам через шею проходят черноватые попереч-
ные полосы. Самки обычно похожи на самцов первого типа окраски,
но с более слаборазвитой пятнистостью. Новорожденный покрыт жел-
товато-белым мехом.
Зубная формула: I у C-J- Pm у М 1 у2--2 = от 34 до 36.
Северная часть Атлантического океана, Балтийское и Баренцево
моря; заходит в Горло Белого моря. Численность в водах СССР мала.
Питается исключительно рыбой (лосось, сельдь, треска, мойва
и др.). Совершает только небольшие перекочевки. На детные лежбища
собирается до 500—600 особей; один самец собирает «гарем» из
2—5 самок. В Балтийском море щенение происходит в конце зимы на
льду. По берегам Атлантического океана деторождение приурочено
чаще к началу зимы; самки родят на прибрежных камнях или на льди-
нах. Детеныш питается молоком матери около трех недель.
В силу малочисленности у берегов СССР почти не промышляется,
но советские зверобойные суда ведут промысел в Северной Атлантике.
Географическая изменчивость не изучена.
Подсемейство ТЮЛЕНЕЙ-МОНАХОВ. Subfamilia MONACHINAE
РОД ТЮЛЕНИ-МОНАХИ. GENUS MONACHUS Fleming, 1822
•В водах СССР водится один вид.,
12. Тюлень-монах. Monachus monachus Hermann, 1779
Табл. XVI, 13
Длина тела от 2,0 до 3,5 м (иногда до 3,8 м). Кондилобазальная
длина черепа около 30 см. Спина и бока тела взрослых темно-бурые;
178
морда, ласты и низ тела грязно-белые; на боках иногда заметны мел-
кие беловатые пятна и полоски.
Зубная формула: I -| Су Рт у Му -2 = 32.
Западная часть Атлантики, Средиземное, Адриатическое, Эгейское
и Черное моря. В СССР — очень редко у Кавказского и Крымского
побережий.
Биология не изучена.
Подсемейство ХОХЛАЧЕЙ. Subfamilia CYSTOPHORINAE
РОД ХОХЛАЧИ. GENUS CYSTOPHORA Nilsson, 1820
Один вид.
13. Хохлач. Cystophora cristata Erxleben, 1777
Табл. XVI, 14. Карта 56
Длина самцов до 2,5 ж, самок до 2 м. Вес самцов до 350 кг. Кон-
дилобазальная длина черепа взрослых 21—29 см. Основной фон всего
тела светло-серый (у самки почти белый); по всему туловищу разбро-
саны крупные, неправильной формы темно-бурые пятна; морда и лоб
бурые. Молодые (до года) сверху буровато-серые с бёловатыми кон-
цами волос, книзу белые.
Зубная формула: I у С у Рт у М у • 2 = 30.
Северная часть Атлантического океана и прилегающие части
Арктического бассейна. Из морей, омывающих СССР, изредка встре-
чается только в Баренцевом море; отдельные особи заплывают в Кар-
ское море.
Обитатель открытых морских пространств. Совершает далекие
миграции, заплывая иногда в реки. Держится или поодиночке, или не-
большими стаями. Для деторождения собирается в районах островов
Ньюфаундленд и Ян-Мейена (между Гренландией и Исландией), где
образует разреженные детные лежбища на льдах. Молодые (1—2) ро-
дятся в марте — мае. Самки кормят их лишь 2—3 недели, после чего
спариваются. По-видимому, моногам. Из линных лежбищ самая мощ-
ная в Датском проливе (между Гренландией и Исландией). Питаются
разнообразной рыбой и головоногими моллюсками, за которыми спо-
собны нырять на большую глубину.
Важный промысловый вид, но в водах СССР ввиду своей малочи-
сленности добывается редко.
Географическая изменчивость не изучена.
12*
Н. А. Бобринский
ОТРЯД КИТООБРАЗНЫЕ. ORDO CETACEA
Рис. 56. Череп дельфина-белобочки (Delphinus
delphis) сверху и снизу.
Китообразные — крупные, вполне водные млекопитающие рыбо-
образного облика: тело веретеновидное, легкообтекаемое, голова обыч-
но большая, шейного 'перехвата нет или он лишь намечается; передние
конечности видоизменены в плавники, задние отсутствуют (сохраня-
ются лишь рудименты скелета, лежащие в толще мускулатуры);
имеется горизонтально расположенный двухлопастной хвостовой плав-
ник. Наружные носовые от-
верстия (дыхала) отодвину-
ты далеко назад, располо-
жены на наивысшей точке
головы (единственное ис-
ключение кашалот, дыхало
которого открывается на
конце морды) и снабжены
пассивно закрывающимися
клапанами. Кожа голая. По-
движных губ нет- Наружное
ухо представлено лишь не-
большим отверстием. Глаза
очень малы, с шаровидным
хрусталиком и уплощенной
роговицей. Семенники ле-
жат в полости тела. Penis
помещается в особом кар-
мане. У самок имеется па-
ра сосков, которые распо-
лагаются по бокам половой
щели и тоже скрыты в особых кожных карманах.
Верхнечелюстные и межчелюстные кости вытянуты в рострум
(клюв), обычно длинный и заостренный. Носовые, лобные и теменные
кости сильно укорочены. Ноздри открываются сверху черепа, и носо-
вой проход направлен почти вертикально вниз.
Зубы либо очень многочисленные, одновершинные и сходны между
собой, либо сильно уменьшены в числе: до одной верхней пары и двух
или одной нижней пары, либо совсем утрачены.
Распространены по всем морям земного шара (в Каспии и Арале
отсутствуют), а также в низовьях крупных рек тропической Азии, Аф-
рики и Южной Америки.
Отряд разделяется на два резко обособленных подотряда. Оба
представлены в фауне СССР.
180
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДОТРЯДОВ КИТООБРАЗНЫХ
1(2). Зубы имеются, хотя в старости могут выпадать. Дыхало не-
парное. На верхней челюсти роговых пластин нет. Череп в носовой об-
ласти асимметричен. Носовые кости участвуют в образовании передней
стенки черепной коробки. Обе половины нижней челюсти сочленены
неподвижно. К грудине причленяются не менее 3 пар ребер.......
..........................Зубатые киты. Odontoceti. (Стр. 181.)
2(1). Зубов нет. Дыхало парное. На верхних челюстях по краям
нёба расположены многочисленные роговые пластины «китового уса».
Череп вполне симметричный. Носовые кости образуют крышу над зад-
ней частью носового отверстия. Обе половины нижней челюсти под-
вижно соединены друг с другом лишь при помощи связок. К грудине
причленяется только одна пара ребер ..........................
. . .........Беззубые киты. Mystacoceti. (Стр. 199.)
Подотряд ЗУБАТЫХ КИТОВ. Subordo ODONTOCETI
См. таблицу для определения подотрядов. Кроме того, ротовая
щель относительно небольшая. Нижние челюсти либо прямые, либо
вогнуты внутрь, хватают пищу поштучно. Питаются рыбой, головоно-
гими моллюсками, другими китообразными и тюленями.
Космополиты.
4 семейства; в водах СССР — 3 семейства.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ ЗУБАТЫХ КИТОВ,
ПРЕДСТАВЛЕННЫХ В ВОДАХ СССР
1(4). Рот на переднем конце рыла. Щель дыхала обращена выпук-
лостью назад. Зубы есть в верхней и нижней челюстях, а если только
в нижней, то в числе не более 7 пар.........................2
2(3). Под горлом нет борозд. Хвостовой плавник с глубокой сере-
динной выемкой на заднем крае (разделен на правую и левую лопа-
сти). Спинного плавника нет, а если есть, то расположен примерно на
середине спины. В нижней челюсти много зубов (исключение составля-
ют только нарвал, у которого в нижней челюсти зубов нет, а в верхней —
по 1 или 1 непарный, и серый дельфин, у которого в нижней челюсти
от 2—4 до 7—14 зубов, а верхняя челюсть беззубая).............
..........................Дельфиновые. Delphinidae. (Стр. 181.)
3(2). Под горлом 2—4 сходящиеся кпереди борозды. Хвостовой
плавник на заднем крае без серединной выемки. Спинной плавник рас-
положен позади середины тела (ближе к хвостовому плавнику, чем
к грудным). В верхней челюсти зубов нет, а в нижней не более двух
зубов с каждой стороны .... Клюворылые. Ziphiidae. (Стр. 193.)
4(1). Рот целиком на нижней стороне головы. Щель дыхала вы-
пуклостью повернута в сторону (не назад). Зубы есть на нижних че-
люстях в числе 7—30 пар; на верхних челюстях зубов нет или не более
1—2 пар ... .... Кашалотовые. Physeteridae. (Стр. 197.)
СЕМЕЙСТВО ДЕЛЬФИНОВЫЕ. FAMILIA DELPHINIDAE
См. определительную таблицу на странице 181. Кроме того, раз-
меры обычно мелкие или средние; жировая подушка на голове развита
слабее, чем у других зубатых китов.
181
Распространены по всем морям (кроме замкнутых: в Арале и
Каспии нет).
Из 13 родов в морях СССР — 7 родов.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ДЕЛЬФИНОВЫХ ФАУНЫ СССР
И СМЕЖНЫХ МОРЕН
1(24). Голова некруглая. Имеется клюв хотя бы в виде узкой ото-
рочки спереди рыла. Спинной плавник хорошо развит, если же его нет
(роды бесперых свиней и китовидных дельфинов), то всех зубов бо-
лее 70.
2(9). Клюв хорошо выраженный, длинный, ясно отделенный от жи-
ровой подушки V-образной бороздой. Рострум значительно длиннее
собственно черепа, измеренного от переднего края
скулового отростка верхней челюсти до задней сто-
роны затылочного мыщелка.........................................3
3(8). Клюв длинный: расстояние от конца
верхней челюсти до переднего края жировой по-
душки явно более 7з расстояния от конца верхней
челюсти до глаза. Длина тела не более 2,5 м. Всех
зубов от 120 и более.........................4
4(7). Спинной плавник хорошо развит. Рост-
рум очень длинный и узкий: ширина рострума у
основания укладывается в его длине значительно
более двух раз...............................5
5(6). От грудных плавников тянется по тем-
ной полосе к подбородку. На костном нёбе имеет-
ся пара глубоких и широких продольных борозд.
Дельфины-белобочки. Delphinus. (Стр. 185.)
6(5). От грудных плавников тянется по тем-
ной полосе к глазу. Костное нёбо без продольных
борозд . . . Продельфины. Stenella. (Стр. 186.)
7(4). Спинного плавника нет. Рострум относительно короткий и
широкий: ширина его у основания укладывается в его длине лишь
около двух раз. . . Китовидные дельфины. Lissodelphis. (Стр. 187.)
8(3). Клюв сравнительно короткий: расстояние от конца верхней
челюсти до переднего края жировой подушки равно приблизительно
!/з расстояния от конца верхней челюсти до глаза. Длина тела взрос-
лых более 2,5 м. Всех зубов не более 104..........................
...................................Афалины. Tursiops. (Стр. 186.)
9(2). Клюв в виде узкой оторочки, слабо отграниченной от жиро-
вой подушки. Рострум равен или короче собственно черепа, измерен-
ного от переднего края скуловой дуги до задней стороны затылочного
мыщелка.........................................................10
10(19). Размеры относительно крупные: длина тела взрослых нс
менее 2 м. Зубы конические без перехвата между коронкой и корнем.
Межчелюстные кости впереди носового отверстия не вздуты бугром . И
11(12). Длина тела не более 3 м. Всех зубов не менее 100 . . . .
. . . . Короткоголовые дельфины. Lagenorhynchus. (Стр. 187.)
12(11). Всех зубов не более 52..............................13
13(18). Окраска одноцветно черная, иногда с узкой белой полосой
вдоль брюха либо пестрая из резко очерченных черных и белых уча-
стков. В верхней и нижней челюстях с каждой стороны не менее
8 зубов ... ... ... . . . . .14
182
14(17). Грудные плавники короткие и широкие. Спинной плавник
лежит приблизительно на середине длины тела или даже на задней
половине его. Расщеп рта прямой, горизонтальный. Лоб не вздут, по-
катый. Зубы мощные и расположены нормально. Один зуб приходится
не менее как на 25 мм зубной линии челюсти......................15
15(16). Окраска пестрая из резко очерченных черных и белых уча-
стков. Грудные плавники очень широкие и закругленные. Зубы мощ-
ные: их поперечный диаметр 2,5—5 см . . Косатки. Orca. (Стр. 189.)
16(15). Окраска сплошь черная. Грудные плавники сравнительно
узкие и к вершине заостряются. Зубы сравнительно слабые, их попе-
речный диаметр около 1,9 см . . .
Рис. 58. Череп косатки
(Orcinus orca).
................Малые, или черные,
косатки. Pseudorca. Стр. 189).
17(14). Грудные плавники очень
длинные и узкие. Спинной плав-
Рис. 59. Череп гринды
(Globicephal a melaena).
ник на передней половине тела. Расщеп рта косой, направленный вниз.
Лоб сильно вздутый, округлый. Зубы слабые, с возрастом часто выпа-
дают и расположены только в передних отделах челюстей. Один зуб
приходится на каждые 15—18 мм зубной линии челюсти.................
...................................Гринды. Globicephala. (Стр. 190.)
18(13). Верх серый, иногда черноватый, брюхо белое, причем на
боках эти цвета постепенно переходят один в другой. В верхней челю-
сти зубов нет, в нижней по 2—7 с каждой стороны....................
.........................Серые дельфины. Grampus. (Стр, 190,)
19(10). Размеры мелкие: общая длина тела не более 1,8 м. Зубы
сдавлены с боков, долотообразной формы, с явной шейкой между ко-
ронкой и корнем. Межчелюстные кости впереди носового отверстия
имеют по большому вздутию в виде бугра..........................20
20(23). Спинной плавник хорошо развит. Всех зубов более 80 . 21
21(22). Черный цвет верха на боках постепенно светлеет к брюху.
Спинной плавник сплошь черный. Задняя часть тела имеет сверху и
снизу лишь по притупленному килю. Зубов в верхних челюстях (16—
30 пар) столько же или больше, чем в нижних (16—25 пар,) . , . ,
Морские свиньи. Phocaena, (Стр. 191,)
183
22(21). Черный цвет спины и белый цвет боков резко разграни-
чены. Спинной плавник с большим или меньшим развитием белого
цвета. В задней части тела сверху и снизу проходит по узкому гребню,
из них верхний тянется от спинного плавника до хвоста, высок в своей
Рис. 60. Череп серого дель-
фина (Grampus griseus).
Рис. 61. Череп морской
свиньи (Phocaena pho-
caena). -
Рис. 63. Череп нарвала
(Monodon monoceros).
средней части, где имеет характер плавника. Зубов в верхних челюстях
(15—24 пары) меньше, чем в нижних (22—28 пар).......................
. . .............Белокрылые свиньи. Phocaenoides. (Стр. 192.)
23(20). Спинного плавника нет. Зубов обычно не более 80 (редко
до 84).....................Бесперые свиньи. Neomeris. (Стр. 192.)
184
24(1). Голова круглая или лишь со слабым намеком на клюв.
Спинного плавника нет. Всех зубов не более 40.....................25
25(26). Окраска однотонная: взрослые особи белые, молодые се-
рые. Зубы имеются как в верхней, так и в нижней челюсти..............
...............................Белухи. Delphinapterus. (Стр. 192.)
26(25). Окраска взрослых пятнистая. Зубы, если имеются, то толь-
ко в верхней челюсти и в количестве не более 2, причем они направ-
лены прямо вперед . ... Единозубы. Monodon. (Стр. 193.)
Подсемейство ДЕЛЬФИНОВЫХ. Subfamilia DELPHININAE
РОД ДЕЛЬФИНЫ-БЕЛОБОЧКИ. GENUS DELPHINUS Linnaeus, 1758
См. определительную таблицу на странице 182. Кроме того, клюв
длинный, узкий, резко отделенный V-образной бороздой от пологого
лба; спинной
Зубы мелкие
и грудные плавники умеренных размеров, серповидные.
40—60 .
<----. Рострум приблизительно в полтора раза длин-
ней черепной коробки. С каждой стороны костного нёба проходит по
продольной глубокой и широкой борозде (отличие от всех прочих ро-
дов дельфинов).
По-видимому, 1 вид.
1. Дельфин-белобочка. Delphinus delphis Linnaeus, 1758
Табл. XVII, 1, Карта 57
Длина тела взрослых самцов 0,9—2,5 ж, самок 0,8—2,3 ж. Об-
щая длина черепа от 36—45 см до 48,5 см. Окраска сильно варьирует,
обычно спина, верх головы, челюсти, грудные плавники, хвостовой сте-
бель и хвост черные, брюхо и низ хвостового стебля серого или белого
цвета с темными продольными полосами.
Основная пища — пелагическая рыба, добываемая чаще вдали от
берегов в верхней толще воды или на незначительной глубине. Как
второстепенный корм нечерноморских особей составляют головоногие
моллюски и ракообразные. Держатся небольшими стаями. Численно
преобладают самцы, составляющие 53%. Спаривание растянуто с июля
по декабрь. Беременность длится около 10 месяцев. Основной срок
спаривания — август — октябрь, максимум в сентябре, щенки — в
июне—августе, максимум в июле. В Черном море два цикла: весенний
и летне-осенний, главный в июле. Детенышей обычно 1, очень редко 2.
Детеныши рождаются обычно под водой. Кормление молоком длится
около 4 месяцев. Половозрелые самки щенятся, no-видимому, еже-
годно.
Космополит, в большом количестве в теплых и умеренных областях
Атлантического и Тихого океанов.
Промышляется в большом количестве в Черном 'море ради сала,
мяса и кожи.
Подвиды изучены слабо. В водах СССР — 3 подвида:
a) D. d. delphis Linnaeus, 1758. Европейские моря.
b) D. d. ponticus Barabasch, 1935. Отличается от номинального меньшей длиной
тела и меньшим черепом, возможно деталями окраски. Черное море.
с) D. d. bairdi Dall, 1873. (Описан из Калифорнии, мыс Аргуэлло.) Отличается от
предыдущих подвидов отсутствием боковых полос и тем, что темный и светлый
цвета на боках разграничены резче. Северная половина Тихого океана.
185
РОД ПРОДЕЛЬФИНЫ. GENUS STENELIA Gray, 1866
См. определительную таблицу на странице 182. По морфологиче-
ским признакам занимает промежуточное положение между родами
дельфинов-белобочек и афалин.
Теплые и умеренные воды Тихого, Индийского и Атлантического
океанов.
Различают 6 видов,' из которых в водах СССР возможно встретить
один.
Полосатый продельфин. Stenella caeruleoalbus Meyen, 1833
Размеры взрослых около 2—2,4 м. Окраска верха темно-голубая,
низ белый; на боках — косые полосы. Число зубов от 160 до 200; обыч-
но в верхней челюсти от 48 до 50 пар, в нижней — от 44 до 46 пар.
Теплый и умеренный пояса Тихого и Атлантического океанов. Воз-
можна встреча у южных островов Курильской гряды.
РОД АФАЛИНЫ. GENUS TURSIOPS Gervalis, 1855
См. определительную таблицу на странице 182. Кроме того, клюв
четко отделен бороздой от выпуклого лба, зубы до 7 мм в диаметре.
По-видимому, 1 вид, возможно 2.
2. Афалина. Tursiops truncatus Montagu, 1821
Табл. XVII, 2
Длина тела 2,3 ж, вес до 400 кг. Верхняя челюсть, верх головы,
спина, хвостовой стебель, спинной и грудные плавники и хвостовые
лопасти буровато-черные; нижняя челюсть, горло, грудь, бока и осно-
вание хвостового стебля белые.
Прибрежные области умеренных и теплых вод северного и южного
полушарий; есть ли в Баренцевом море, не установлено; в Черном море
держится в шрибрежной полосе повсеместно, от Одессы до Батуми,
сезонные и кормовые передвижения в Черном море не изучены, в наи-
большем количестве встречается у крымских берегов, в меньшем —
у кавказских.
Питается рыбой, преимущественно придонной. Ест также креветок,
морских тараканов, моллюсков. Придерживается прибрежной полосы
и избегает открытого моря. Иногда заходит в реки на значительные
расстояния. Размножение изучено слабо. Сроки размножения, по-види-
мому, растянуты, но совпадают преимущественно с теплым временем
года. Держится сравнительно небольшими косяками до нескольких де-
сятков голов. Иногда дает брызговые фонтаны до 1 —1,5 м высоты.
Успешно содержится в неволе в нью-йоркском и флоридском океана-
риях.
Промышляется у атлантических берегов США и у нас в Черном
море.
В водах СССР, по-видимому, 3 подвида:
а) Т. t. truncatus Montagu, 1821. (Описан из Англии.) Размеры относительно
крупные: длина тела до 360—390 см; длина черепа до 55—58 см. Зубов вверху обычно
21—26 пар, внизу 20—25 пар. Предчелюстные кости в средней части сильно вздуты.
Атлантический сектор ареала.
b) Т. t. ponticus Barabasch, 1940. (Тип из Черного моря.) Размеры меньше: длина
тела до 310 см; длина черепа до 50,3 см. Зубов меньше: верхних 19—22 пары; ниж-
них 23—24 пары. Предчелюстные кости со слабо выраженным вздутием. Черное море.
186
с) Т. t. gilli Dahl, 1873. (Описан по экземпляру из Японии.) Очень слабо изу-
ченная форма. По размерам близка к номинальной. Отличается некоторыми дета-
лями в строении черепа. Моря Дальнего Востока.
РОД КИТОВИДНЫЕ ДЕЛЬФИНЫ. GENUS LISSODELPHIS Gloger, 1841
См. определительную таблицу на странице 182. Кроме того, клюв
умеренной длины, узкий, четко отграниченный от сравнительно низ-
кого лба; грудные плавники серпообразные; зубы мелкие: в диаметре
около 3 мм.
Тихий океан.
2 вида; в водах СССР наблюдали (но не добывали) 1 вид.
Китовидный дельфин. Lissodelphis borealis Peale, 1848
Размеры взрослых около 2,4—2,5 м. Тело относительно стройное.
Нижняя челюсть несколько выступает за верхнюю. Окраска спины и
боков бархатно-черная, верхняя челюсть, кончик нижней челюсти и
большое пятно, идущее от горла, расширенное между грудными плав-
никами, суженное в узкую полосу вдоль средней линии брюха и раз-
двоенное на нижней поверхности хвостового плавника, белые.
Северная половина Тихого океана, от Калифорнии и Японии до
Берингова моря; в водах СССР наблюдали у островов Парамушир.
Итуруп и Шикотан.
Пищу, по-видимому, составляют рыба и головоногие моллюски.
Поведение известно плохо. Держится в одиночку и группами по
2—3 особи. Пуглив, исключительно быстрый, подвижный. Встречается
обычно вдали.от берегов.
РОД КОРОТКОГОЛОВЫЕ ДЕЛЬФИНЫ. GENUS LAGENORHYNCHUS Gray,1846
См. определительную таблицу на странице 182. Кроме того, хво-
стовой стержень сжат с боков и снабжен ясным верхним и нижним ки-
лями, спинной плавник высокий, серповидный; над верхней челюстью
нависает лобный выступ.
Космополит.
По-видимому, 5 видов, из них в водах СССР — 3 вида.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ КОРОТКОГОЛОВЫХ ДЕЛЬФИНОВ
ФАУНЫ СССР
1(4). Конец морды темный. На боках выше грудного плавника
есть белый участок. Зубов в верхней челюсти обычно 29—40 пар. Ди-
аметр зубов не больше 5 мм. Позвонков 74—82 ...................2
2(3). Темные грудные плавники прикрепляются к белым бокам.
Зубов в верхней челюсти 35—40 пар. Позвонков 77—82. Ребер
14—15 пар. . Белобокий дельфин. Lagenorhynchus acutus. (Стр. 188.)
3(2). Темные грудные плавники прикрепляются к темной полосе,
идущей вдоль боков тела. Зубов в верхней челюсти обычно 30—32 пары.
Позвонков 74—78. Ребер 12—14 пар.................................
. . Полосатый дельфин. Lagenorhynchus obliquidens. (Стр. 188.)
4(1). Конец клюва белый. Выше грудного плавника нет белого
участка. Количество зубов в верхней и нижней челюстях обычно
22—28 пар. Диаметр зубов в середине ряда у взрослых 6—7 мм. По-
звонков 88—94 ............................................. . .
Беломордый дельфин. Lagenorhynchus albirostris. (Стр. 188.)
187
Белобокий дельфин. Lagenorhynchus acutus Gray, 1828
Обычные размеры самцов 2,3—2,75 ж, до 3 ж, самки обычно
2—2,4 ж. Относительно короткий клюв нечетко отграничен от слабо
закругленного покатого лба. На хвостовом стебле сильно развиты верх-
ний и нижний гребни. Верх всего тела от конца морды до конца хво-
ста, грудные плавники и клюв черные. На боках тела широкая белая
полоса, заходящая на хвостовой стебель. Сверху она резко ограничена
черным цветом спины, снизу — желто-сероватой окраской нижней части
боков. Брюхо чисто белое. Грудные плавники темные. От их оснований
вперед к глазу тянется по белому фону узкая черная полоса. Другая
темная полоска идет вдоль боков тела, от заднего конца хвостового
стебля до вертикали, опущенной от переднего края спинного плавника.
Нижняя сторона хвостовых лопастей светлая. Глаза окружены тем-
ным кругом.
Прибрежные воды Северной Атлантики к югу до Балтийского и
Баренцева морей, к северу до Шпицбергена и Гренландии. В водах
СССР не найден, но возможен у берегов Мурмана и в советских водах
Балтийского моря.
Биология известна крайне слабо. На основании скудных данных
пищу составляет пелагическая и придонная рыба, а также ракообраз-
ные и моллюски. Беременность длится около 10 месяцев, роды, по-ви-
димому, в середине лета. Образ жизни стадный, обычно держится
группами в 10—50 голов, но иногда скопляется до 1 —1,5 тыс. голов.
Промысел ведется только в Норвегии.
3. Полосатый дельфин. Lagenorhynchus obliquidens Gill, 1865
Табл. XVII, 3
Несколько меньше белобокого дельфина: самец до 2,3 ж, самка до
2,2 ж. Края обеих челюстей, самый конец мордьц верх головы и спина
черные, бока белые, от глаза до вертикали переднего края спинного
плавника тянется черная полоса, брюхо белое, задний край спинного
и грудных плавников и хвостовых лопастей беловатый; однако рас-
цветка значительно варьирует индивидуально. Зубов верхних 27—34,
нижних 28—32 пары. Северная половина Тихого океана у берегов Азии
и Северной Америки.
Биология изучена слабо. Пища, судя по отрывочным данным, пре-
имущественно мелкая: стайная рыба и головоногие моллюски. Спари-
вание, по-видимому, происходит летом. Активны в любые часы суток.
Держатся группами в 2—3 головы, иногда стаями до тысячи (часто
вместе с белобочками). Неволю переносят хорошо (в Японии).
Не промышляется, добывается лишь случайно, между тем содер-
жит много жира, мясо вкусное, напоминает телятину; шкуру можно
использовать для выделки тонких и мягких кож.
4. Беломордый дельфин. Lagenorhynchus albirostris Gray, 1846
Табл. XVII, 4
Длина тела самцов обычно 2,7—3 ж; вес самца до 275 кг; самки
2,5 ж. Низ белый, верх черный, бока также темные, клюв и передняя
часть жировой подушки белые, хвостовые лопасти сверху темные,
снизу сероватые.
Умеренные воды Северной Атлантики, от Баренцева моря до бе-
регов Франции и от пролива Девиса до Массачусетса. Наиболее обы-
188
чен в Северном море, особенно у побережья Норвегии, восточных бе-
регов Великобритании и Фарерских островов, где встречается круглый
год, но преимущественно летом. Для Баренцева моря характерен не
менее чем пыхтун. В Варангер-фиорде наблюдались тысячами. Вполне
допустимо, что встречается у полуострова Рыбачий и Мурманского
побережья. В Балтийское море проникает до Финского залива. По-ви-
димому, на зиму уходит в южные части Северного моря.
Биология известна слабо. Прибрежный вид. В основном питается
придонной рыбой, а также ест ракообразных и моллюсков, в частности
головоногих. Роды происходят главным образом в середине лета.
В июле, августе и сентябре чаще держится парами, но иногда стадами
в несколько сот голов.
Промысел развит лишь у берегов Норвегии.
РОД КОСАТКИ. GENUS ORCINUS Fitzinger, 1860
См. определительную таблицу на странице 183. Кроме того, го-
лова с широкой мордой, без ясно выраженного клюва; грудные плав-
ники широкие, овальные, костный клюв широкий, несколько вогнутый
впереди ноздрей.
Один вид.
5. Косатка. Orcinus orca Linnaeus, 1758
Табл. XVII, 5
Длина тела самцов 6—8 м, реже 9—10 м, самок не превышает 7 м,
новорожденные около 2,5 м. Спинной плавник треугольный, очень вы-
сокий, особенно у самцов; с возрастом он увеличивается. Верх головы,
спина и хвостовой стебель черные или черно-серые, низ белый, позади
грудного плавника белое пятно широким языком поднимается на бока
тела, грудные и хвостовой плавники обычно сверху и снизу темные,
реже хвостовой плавник снизу целиком или частично белый; за спин-
ным плавником находится большое полулунное пятно с вершиной,
обращенной к хвосту, над глазом часто имеется овальное белое пятно.
Космополит, но в Черном и Азовском морях, в Арктике и Антарк-
тике отсутствует и не заходит в плотные льды; наиболее обычна в за-
падной части Баренцева моря, особенно у берегов Мурманска и Ры-
бачьего полуострова, где даже многочисленна; нередко заплывает
в Белое море, зимой посещает даже’Балтийское море. Миграции не
изучены. Основная пища — стайная рыба, морские млекопитающие.
РОД ЧЕРНЫЕ КОСАТКИ. GENUS PSEUDORCA Reinchardt, 1862
См. определительную таблицу на странице 183. Кроме того, жиро-
вая подушка хорошо развита, клюв тупой и очень короткий, спинной
плавник умеренно развит и глубоко вырезан сзади, грудные плавники
заострены и сужены.
Один вид.
Черная косатка. Pseudorca crassidens Owen, 1846
Длина 4—6 ж; вес до 1,5 т; самцы на 60 см—1м крупнее самок.
Клюв очень короткий, едва заметный. Верхняя челюсть слегка вы-
дается над нижней. Окраска всего тела черная. Зубы —~~8 ; ветре-
8—11 1 ~10
чается ------.
8—12
189
Умеренные воды северного и южного полушарий. Миграции совер-
шенно не изучены. Ведет стадную жизнь, объединяясь в косяки из
100—200 голов. Питается в основном головоногими моллюсками,
а также рыбой.
Добывается только случайно, хотя ее жир и мясо не хуже, чем
у других дельфинов.
РОД ГРИНДЫ. GENUS GLOBICEPHALA Lesson, 1828
См.'определительную таблицу на странице 183. Кроме того, череп
массивный с широким тупым рылом, межчелюстные кости очень ши-
рокие.
Один вид.
6. Гринда. Globicephala melaena Traill, 1809
Табл. XVII, 6
Длина тела обычно не превышает 6 м, наибольшие размеры самца
6,5 ж, самки до 5 ж. Окраска черная, вдоль брюха тянется узкая свет-
лая полоска, резко расширяющаяся в пятно якоревидной формы между
грудными плавниками и на горле. Зубов 7—13 пар верхних и 7—15 пар
нижних. С возрастом зубы частично выпадают, частично снашиваются
в вершинах.
Космополит.
Основная пища — головоногие моллюски и стайная рыба. Полово-
зрелость самок наступает в возрасте 6 лет. Щениться они могут, по-
видимому, до 18 лет, принося за это время 5—6 детенышей.
Размножение растянуто с весны до зимы включительно, бере-
менность и лактационный период длится почти по году. Полигам,
холостые самцы которого иногда образуют самостоятельные стада.
Стадный инстинкт сильно развит, животные держатся группами от
нескольких особей до нескольких тысяч. Хорошо переносит неволю
в Флоридском океанарии.
Регулярно добывается в летнее время у Фарерских островов (от
нескольких сот до нескольких тысяч в год), у берегов о-ва Ньюфаунд-
ленд, у берегов Виргинии и Северной Каролины. В СССР промысла
нет. Мясо несколько напоминает говядину, используется в пищу.
В водах СССР, по-видимому, 2 подвида:.
a) G. гл. melaena Traill, 1809. (Описан из района Оркнейских островов.) Имеет
сравнительно длинные грудные плавники, равные около Vs длины тела, белую полосу
на брюхе, расширяющуюся под горлом, и до 13 пар зубов наверху и внизу. Барен-
цево море.
b) G. m. macrorhyncha Gray, 1846. (Из южных морей.) Длина грудных плавни-
ков около Vs длины тела, сплошь черной окраски; имеет 7—8 пар зубов наверху и
внизу. Моря Дальнего Востока.
РОД СЕРЫЕ ДЕЛЬФИНЫ GENUS GRAMPUS Gray, 1828
См. определительную таблицу на странице 183. Кроме того, вслед-
ствие большой жировой подушки клюв внешне едва выражен, нижняя
челюсть немного не доходит до конца морды’; рот косой; клюв корот-
кий; длина его составляет V2 общей длины черепа, зубы очень часто
выпадают, грудные плавники сужены, заостренные, серповидные.
Один вид.
Серый дельфин. Grampus griseus Cuvier, 1812
Размеры взрослых особей 2,5—4 ж, самок до 3,4 ж, общая длина
черепа около 487—540 мм. Окраска весьма своеобразная: голова и пе-
редняя часть туловища серые, спина и плавники черноватые до чер-
190
ного, брюхо светло-серое или белое, все эти цвета постепенно перехо-
дят друг в друга, встречаются самцы светло-голубые с темно-коричне-
выми пятнами и самки одноцветно коричневатые. На теле 'многочис-
ленные неправильные светлые полоски, по-видимому, следы различного
рода повреждений. Питается головоногими моллюсками. Держится
большей частью одиночно либо парами, редко группами. Размножение
почти не изучено.
Космополит.
Подсемейство МОРСКИХ СВИНЕЙ. Subfamilia PHOCAENINAE
Имеются рудименты роговых бугорков на коже. Часть зубов с до-
лотообразной коронкой.
РОД МОРСКИЕ СВИНЬИ. GENUS PHOCAENA G. Cuvier, 1817
См. определительную таблицу на странице 183. Кроме того, череп-
ная коробка и клюв одинаковой или почти одинаковой длины, -задние
края межчелюстных костей образуют предносовое вздутие.
Мировой океан, исключая арктические и антарктические воды.
По-видимому, 4 вида, из которых один относится к особому под-
роду. В водах СССР — 1 вид.
7. Морская свинья. Phocaena phocaena Linnaeus, 1758
Табл. XVII, 7
Наибольшая длина тела 186 см (обычно около 1,5 ж), самки не-
сколько крупнее самцов. Голова, спина, хвостовой стебель, грудные,
хвостовой и спинной плавники черные, бока сероватые, брюхо белое; от
угла рта к грудному плавнику проходит узкая темная полоска; оттенки
изменяются ог темно-серого до черного и от ярко-белого до серого.
Верхних зубов 22—30 пар, нижних 22—23 пары. У взрослых осо-
бей диаметр зубов (расположенных в середине ряда) близ десен 2 мм,
ширина коронки в вершине 2,5—3 мм; у старых особей зубы вы-
падают.
Прибрежные воды северных частей Атлантического и Тихого океа-
нов, в морях Баренцевом, Белом, Балтийском, Северном, Черном, Азов-
ском, Японском, Охотском, Беринговом и Чукотском. В Средиземном
море, по-видимому, не водится и лишь изредка проникает в него через
проливы Гибралтарский и Босфор.
Разгар спаривания — в конце июня — августе. Беременность —
9—10 месяцев. Роды со второй половины апреля по июль. Новорож-
денный очень велик: до !/2 длины тела 'матери, вес его 2,8—7,9 кг. Лак-
тационный период длится, вероятно, около 4 месяцев. Обыкновенно
держится небольшими группами в 2—10 голов, нередко стайками
в 15—20 особей. Брызговых фонтанов не дает. Регулярный промысел
кошельковым неводом в СССР ведется только в Черном море с ноября
по апрель включительно. Питается мелкой рыбой, головоногими мол-
люсками, креветками, придонными беспозвоночными.
Географическая изменчивость слабо выражена. В водах СССР различают 3 слабо
обособленных подвида:
a) Ph. ph. phocaena Linnaeus, 1758. Размеры относительно крупные. Рострум уко-
роченный. На передних краях плавников есть бугорки. Северная Атлантика.
b) Ph. ph. relicta Abel, 1905. Размеры немного меньше, чем у атлантической формы.
Рострум менее укорочен. Бугорки на передних краях плавников редуцированы. Чер-
ное и Азовское моря.
с) Ph. ph. vomerina Gill , 1865. (Описан из американских вод.) Отличается дета-
лями строения черепа. Северотихоокеанская часть ареала.
191
РОД БЕЛОКРЫЛЫЕ СВИНЬИ. GENUS PHOCAENOIDES Andrews, 1911
См. определительную таблицу на странице 184.
Один вид.
8. Белокрылая свинья. Phocaenoides dalli True, 1885
Табл. XVII, 8
Самцы достигают 2 м длины и 114 кг веса, самки до 1,8 м. Зубов
15—24 ~
-----. Они очень малы, долотовидные, едва заметные, сжатые
22—28
в шейке, чуть выступают над деснами. На хвостовом стебле сильно
развиты спинной и брюшной гребни. На боках тела большие светлые
пятна. Задние края спинного и хвостового плавников тоже светлые,
беловатые. Окраска остальной поверхности тела черная.
Северная половина Тихого океана от Японии (о. Ниппон) и Ка-
лифорнии до Охотского и Берингова морей включительно.
Биология изучена слабо. Питается головоногими моллюсками и
рыбой. О размножении данных нет. Придерживается прибрежных вод,
где плавает стайками в 3—15 голов, иногда скопляется в стада до
200 особей. Миграции не изучены.
2 подвида:
a) Ph. d. dalli True, 1885. (Описан из района Алеутских островов.) Белое поле
боков и тела доходит только до вертикали от переднего края спинного плавника, но
высоко поднимается к спине. Охотское и Берингово моря.
b) Ph. d. truei Andrews, 191'1. (Описан из вод близ о-ва Хонсю, Япония.) Белое
поле боков доходит почти до глаз, но поднимается к спине менее высоко. Япон-
ское море.
РОД БЕСПЕРЫЕ СВИНЬИ. GENUS NEOMERIS Gray, 1846
См. определительную таблицу на странице 184.
Один вид.
Бесперая свинья. Neomeris phocaenoides. G. Cuvier, 1829
Размеры не более 160 см. На спине узкая (3—6 см) полоса из
многочисленных роговых бугорков (бородавок).
Теплый пояс Тихого и Индийского океанов, от Японии и Кореи до
мыса Доброй Надежды. Возможны заплывы к островам Курильской
гряды и берегам Южного Приморья.
Подсемейство БЕЛУХОВЫХ. Subfamilia DELPHINAPTERINAE
РОД БЕЛУХИ. GENUS DELPHINAPTERUS Lacepede, 1804
См. определительную таблицу на странице 185.
Один вид.
9. Белуха. Delphinapterus leucas Pallas, 1776
Табл. XVII, 9. Карта 57
Длина до 6 м\ вес до 1 т. Окраска взрослых особей белая, моло-
дых тем»но-серая, полувзрослых синевато-серая.
Распространение кругополярное (есть во всех морях Ледовитого
океана). В Тихом океане к югу заходит до Татарского пролива и Япон-
ского моря. Далеко заходит в большие реки Северной Америки и Си-
192
бири, например в Амур, Анадырь, Обь, Енисей и др. (по Енисею под-
нималась более чем на 1000 км).
Основная пища — рыба, в частности мойва (в Белом море), сайка
(в Ледовитом океане), лососевые и сельди (в морях Дальнего Во-
стока); ест также ракообразных и головоногих моллюсков. Держится
косяками от нескольких десятков до сотен голов, иногда до нескольких
тысяч. Избегая участки сплошных ледяных полей, совершает большие
и регулярные сезонные миграции.
Деторождение приурочено в основном, по-видимому, к весеннему
времени. Самки приносят по одному детенышу раз в два года.
Половозрелость — па 2—3-м году.
Важный промысловый вид, добываемый ради кожи и жира.
В морях СССР — 3 формы, слабо различающиеся по средним показателям раз-
меров тела и черепа:
a) D. 1. leucas Pallas, 1776. (Описана из Обской губы.) Длина взрослых (белых
самцов и самок) 318—464 см (М 390); кондилобазальная длина черепа 502—635 мм
(М 566). От Баренцева моря до моря Лаптевых включительно («карское стадо»).
b) D. 1. freimani Klumow, 1935, (Описана из Белого моря.) Размеры немногим
меньше: длина тела взрослых (белых самцов и самок) 253—376 см (М 312); конди-
лобазальная длина черепа 424—585 мм (М 520). Летом — в Белом море («беломор-
ское стадо»), зимой (предположительно) —в Баренцевом море.
с) D. 1. dorofeevi Barabasch et Klumov, 1935. (Описана из Охотского моря.) Отно-
сительно крупная: длина тела взрослых 320—600 см (М 424); кондилобазальная
длина черепа 580—648 мм (М 591). Татарский пролив и моря; Охотское, Берингово,
Чукотское и Восточно-Сибирское (восточная половина) — «охотское стадо».
РОД ЕДИНОЗУБЫ. GENUS MONODON Linnaeus, 1758 *
См. определительную таблицу на странице 185.
Один вид.
10. Нарвал. Monodon monoceros Linnaeus, 1758
Табл. XVII, 10
Размеры взрослых 4—6 м (без бивня); вес около 1 т. У самцов,
как правило, развивается только левый бивень, достигающий длины
2—3 м. У са'мок зубы обычно остаются в деснах, реже прорезаются;
иногда один или оба зуба достигают больших размеров. Окраска взрос-
лых обычно желтоватая с многочисленными серыми и темпо-бурыми
пятнами; на боках пятна реже и светлее, на брюхе их нет. Молодые
голубовато-серые с более светлым низом и без пятен.
Коренной обитатель Арктики, распространен кругополярно к се-
веру до линии плавучих льдов. В северных морях СССР редок, осо-
бенно в Чукотском море. Небольшие косяки их в последнее десятиле-
тие наблюдались в Карском и Баренцевом морях, около Новой Земли
и между островами Земли Франца-Иосифа. Раньше в зимние месяцы
отмечался у Мурмана, Мезенской губы, устья Печоры.
Питается головоногими моллюсками и, видимо, в меньшей степени
рыбами и ракообразными. Беременные самки и детеныши встречались
в разное время года. Нарвал — стадный вид. В прошлом столетии его
встречали косяками до нескольких тысяч голов. Теперь встречаются
стада лишь до 15—20 голов.
СЕМЕЙСТВО КЛЮВОРЫЛЫЕ. FAMILIA ZIPHIIDAE
См. определительную таблицу на странице 181, Кроме того, раз-
меры для китообразных средние. Челюсти удлинены в «клюв». Дыхало
в виде полумесяца, обращенного рогами к рылу. Грудные плавники
13 Н. А. Бобринский и др.
193
сравнительно малы, ланцетовидные. Череп сзади носового отверстия
резко и сильно приподнят.
Тихий, Атлантический и Индийский океаны. 5 родов, в водах
СССР — 4 рода.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ КЛЮВОРЫЛЫХ, ОБИТАЮЩИХ
В ВОДАХ СССР
1(6). Зубы более или менее конические; сидят они на самом конце
нижней челюсти. Клюв относительно широкий........................2
2(5). Лоб высокий, выпуклый. С каждой стороны нижней челюсти
по 2 зуба. На черепе глубокая
предглазничная выемка.............5
3(4). Клюв не уплощен. Длина
тела менее 10 м. Передние зубы до
5 см высоты; у самок они иногда
скрыты в деснах; задние зубы у
самок всегда, у самцов иногда
скрыты в деснах...............
. . . Бутылконосы. Hyperoodon.
(Стр. 194.)
Рис. 64. Черепа китов: слева — бутылконоса (Hyperoodon ampullatus);
справа — северного плавуна (Berardius bairdi).
4(3). Клюв уплощен. Длина тела взрослых около 12 м. Передние
зубы около 8 см высоты; задние около 5—7 см, но у самок обычно не
прорезаются.....................Плавуны. Berardius. (Стр. 195.)
5(2). Лоб низкий, плоский. С каждой стороны нижней челюсти по
1 зубу. На черепе предглазничная выемка слабо намечена ....
...................................Клюворылы. Ziphius. (Стр. 195.)
6(1). Зубы в виде удлиненных пластинок. Расположены они зна-
чительно отступя от конца нижней челюсти. Клюв длинный, тонкий . .
..............................Ремнезубы. Mesoplodon. (Стр. 196.)
РОД БУТЫЛКОНОСЫ. GENUS HYPEROODON Lacepede, 1804
См. определительную таблицу на странице 194.
Мировой океан.
2 вида, в водах СССР— 1 вид.
194
11. Высоколобый бутылконос. Hyperoodon ampullatus Forster, il[770
Табл. XVII, 11 ' - J
Длина тела 6—9 ju.
Северная половина Атлантического океана. Придерживаясь теп-
лого течения (Гольфстрима), доходит до Новой Земли, Шпицбергена,
Белого моря. Зимой заходит иногда в Балтийское море.
Питается в основном головоногими моллюсками, дополнительный
корм — рыба, голотурии, морские звезды. Беременность около года.
Роды весной и летом. Детеныш, по-видимому, сосет мать не более
5—7 месяцев. Держатся небольшими стай-
ками от 2—4 до 10—20 голов, хотя извест-
ны стада в несколько сот голов.
РОД ПЛАВУНЫ. GENUS BERARDIUS
Duvernoy, 1851
См. определительную таблицу на стра-
нице 194. Кроме того, зубы сжаты с боков:
2 вида, в водах СССР— 1 вид.
12. Северный плавун. Berardius bairdi
Steineger, 1883
Табл. XVII, 12
Северная половина Тихого океана (от
Охотского и Берингова морей до Камчатки
и южнояпонских вод).
Питается главным образом головоно-
гими моллюсками, частично рыбой. Бере-
менность длится 10 месяцев. Спаривание
сильно растянуто, преимущественно в фев-
рале. Деторождение — от декабря до июня.
Стадный инстинкт сильно развит. К берегу
подходит редко. Обычно держится за де-
Рис. 65. Череп клюворыла
(Ziphius cavirostris).
сятки миль от берега.
Охотиться с обычных китобойных судов
невыгодно, поэтому его
добывают в СССР пока случайно, однако запасы позволяют расширить
промысел. В пищу непригоден. Используют для выработки подошвен-
ной кожи сравнительно низкого качества.
РОД КЛЮВОРЫЛЫ. GENUS ZIPHIUS Cuvier, 1823
См. определительную таблицу на странице 194. Кроме того, клюв
короткий, треугольный.
Мировой океан.
Один вид.
13. Клюворыл. Ziphius cavirostris G. Cuvier, 1823
Табл. XVII, 13
Длина тела взрослых до 8 м; кондилобазальная длина 82—99 см.
Мировой океан, но всюду малочислен.
Экология изучена слабо;. По-видимому, питается головоногими
моллюсками. Время, родов растянуто, но большинство, по-видимому,
рожает осенью.
13*
195*
Промышляется в небольшом количестве в Японии. В СССР ие до-
бывается.
РОД РЕМНЕЗУБЫ. GENUS MESOPLODON Gervais, 1850
См. таблицу для определения родов на странице 194. Кроме того,
лоб низкий, зубов одна пара. При закрытом рте губы охватывают верх-
ние челюсти с боков, прочно фиксируя их. Длина тела меньше 7 м\
кондилобазальная длина около 75—83 см. Нижнечелюстной симфиз за-
1 1
нимает от — до — длины челюсти.
5 3
Тихий и Атлантический океаны с Северным, Балтийским и Среди-
земным морями.
8 видов, в водах СССР найден 1 и 1 возможен.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РЕМНЕЗУБОВ, КОТОРЫЕ МОГУТ
ВОДИТЬСЯ В ВОДАХ СССР
1(2). Отверстие для нерва в межчелюстной кости лежит позади
отверстия для нерва в верхнечелюстной кости (рис. 66, Б). Зубы четы-
рехугольные (рис. 67, 5)........................................
. . . Командорский ремнезуб. Mesoplodon steinegeri. (Стр. 196.)
2(1). Отверстие для
нерва в межчелюстной
кости лежит впереди от-
верстия для нерва в верх-
нечелюстной (рис. 66,А).
Зубы треугольные (рис.
67, А) . . . Атланти-
ческий ремнезуб. Meso-
plodon bidens. (Стр. 197.)
Рис. 67. Зубы ремнезубов:
А — атлантический ремнезуб (Me-
soplodon bidens); Б — командорский
ремнезуб (М. steinegeri).
Рис. 66. Черепа ремнезубов (сверху):
Л — атлантический ремнезуб (Mesoplodon bidens);
Б — командорский ремнезуб (М. steinegeri).
14. Командорский ремнезуб. Mesoplodon steinegeri True, 1885
Табл. XVII, 14
См. определительную таблицу на странице 196. Кроме того, длина
тела около 5 м. Предглазничная выемка на черепе выражена слабо.
Правая и левая нёбные кости раздвинуты. Затылочная кость уплощена.
196
Тихий океан. Всего известно 13 находок, из них 3 — в СССР, на
о-ве Беринга.
Образ жизни не изучен. По-видимому, питается головоногими
моллюсками.
Атлантический ремнезуб. Mesoplodon bidens Sowerby, 1804
См. определительную таблицу на странице 196. Кроме того, длина
тела взрослых 4,5—5,5 м. Предглазничная выемка на черепе развита
сравнительно хорошо. Правая и левая нёбные кости прилегают друг
к другу. Затылочная ко-сть резко выпуклая.
Атлантический океан и Средиземное море. В водах СССР не до-
быт, ближайшие к водам СССР места — берега Норвегии и Польши.
Как и все ремнезубы, редок, хотя обычнее других.
Образ жизни изучен крайне слабо. Близ берегов бывает редко.
Держится, по-видимому, в одиночку или парами. Не промышляется.
СЕМЕЙСТВО КАШАЛОТОВЫЕ. FAMILIA PHYSETERIDAE
См. определительную таблицу на странице 181. Кроме того, рото-
вое отверстие расположено на нижней стороне головы; на верхней че-
люсти 1—2 пары зубов или зубов нет, на нижней челюсти 7—30 пар
зубов.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ КАШАЛОТОВЫХ
1(2). Голова занимает около Vs общей длины тела. Дыхало на те-
мени. Длина тела не свыше 4 м. На нижней челюсти не более 15 пар
зубов. Спинной плавник серпообразный .....................
........... . . . Карликовые кашалоты. Kogia. (Стр. 198.)
Рис. 68. Черепа кашалотов:
А — карликовый кашалот (Kogia brevirostris); Б — кашалот (Physeter catodon);
А, Б — сбоку; а, б — сверху.
197
2(1). Голова занимает V2 общей длины тела. Дыхало лежит в пе-
реднем левом углу головы. Взрослые свыше 7 м длины. На нижней че-
люсти не менее 18 пар зубов. Спинной плавник низкий, толстый, в виде
горба..............................Кашалоты. Physeter. (Стр. 198.)
РОД КАРЛИКОВЫЕ КАШАЛОТЫ. GENUS KOGIA Gray, 1846
См. определительную таблицу на странице 197. Кроме того, голова
спереди закруглена.
Индийский, Атлантический и Тихий океаны, между 50° с. ш. и 45°
ю. ш. В водах СССР возможны заплывы в Японское море и к южной
части Курильской гряды.
Один вид.
Карликовый кашалот. Kogia breviceps Blainville, 1838
Длина тела самца 3—4 ж, самок 2—3 м. Грудные плавники заост-
ренные. Биология изучена крайне слабо. Пища в основном головоно-
гие моллюски, ест также крабов, креветок, рыб. Спаривание, по-види-
мому, поздним летом. Беременность — 9 месяцев. Роды весной. Моло-
дые держатся около матери почти год. Все эти даты весьма неточны.
Индийский, Атлантический и Тихий океаны, между 50° с. ш. и
45° ю. ш. В водах СССР крайне редок. Возможен в Японском море и
водах южной части Курильской гряды.
Географическая изменчивость не изучена.
РОД КАШАЛОТЫ. G’ENUS PHYSETER Linnaeus, 1758
См. определительную таблицу на странице 198. Кроме того, в че-
репе клюв заостренный, треугольной формы; если на верхней челюсти
есть зубы, то они недоразвиты и слабо сидят в десне, в нижней челюсти
с каждой стороны 20—27 зубов.
Мировой океан.
Один вид.
15. Кашалот. Physeter catodon Linnaeus, 1758
Табл. XVIII, А Карта 57
Самцы 14—20 ж (в настоящее время самцы свыше 18 м редки);
самки 10—15 м. Вес самцов до 100 т.
Основная пища — головоногие моллюски, второстепенная — рыба,
крабы. Спариваются, плодятся и выкармливают молодняк в теплых во-
дах. Держатся обычно группами («гаремами») из самца и 10—15 са-
мок. Иногда несколько «гаремов» объединяются в стадо из нескольких
сот голов. Спаривание, по-видимому, в июне—июле. Беременность
длится, по-видимому, -16 месяцев. Деторождение — с декабря по ап-
рель, максимум в феврале.
Самки, как правило, не выходят из тропического и подтропиче-
ского поясов, а холостые самцы в летнее время доходят в северном по-
лушарии до Баренцева и Берингова морей, в южном — до Антарктики
и плавучих льдов. Главная масса держится между 40° северной и юж-
ной широты.
В дальневосточных морях СССР — важнейший промысловый вид;
за 22 года (1933—1954) он дал половину всей продукции китобойного
промысла (51%), а за 1950—1954 гг.— 87%. С очень крупного самца
198
снимали до 25 т жира. Кроме жира, у кашалота берут от 2 до И т
спермацета, используемого для косметических изделий (крема, помады,
мыла). Высоко ценится также образующаяся в пищеварительном
тракте кашалота амбра — похожее на воск вещество, используемое
в качестве фиксатора ароматических веществ.
Подотряд БЕЗЗУБЫХ КИТОВ. Subordo MYSTACOCETI
См. определительную таблицу на странице 181. Кроме того, рот
очень велик, и разрез его простирается за глаза. Нижние челюсти вы-
гнуты, хотя зубы .и закладываются, но рассасываются еще у зародыша.
Пластины китового уса имеют треугольную форму, цельный наружный
край и бахромчатый внутренний. В целом эти пластины образуют це-
дильный аппарат, позволяющий этим огромным животным питаться
мелкими ракообразными, моллюсками и небольшой рыбой. Самки
в среднем, как правило, несколько крупнее самцов.
Космополиты.
3 семейства.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ БЕЗЗУБЫХ КИТОВ
1(4). На нижней стороне тела есть глубокие продольные борозды.
Есть спинной плавник, хотя бы зачаточный, в виде невысокого гребня.
Голова относительно небольшая и укладывается в общей длине тела
более четырех раз. Передние конечности четырехпалые.
2(3). На горле 2—4 глубокие продольные борозды. Зачаточный
спинной плавник в виде невысокого гребня. Голова небольшая. В каж-
дом ряду китового уса не более 180 пластин.....................
........................Серокитовые. Eschrichtiidae. (Стр. 199.)
3(2). На горле и груди свыше И глубоких продольных борозд.
Спинной плавник вполне развит. В каждом ряду китового уса более
220 пластин..........Полосатиковые. Balaenopteridae. (Стр. 200.)
4(1). Нижняя сторона тела гладкая, без глубоких продольных бо-
розд. Нет никакого намека на спинной плавник. Голова огромная и
укладывается в общей длине тела менее трех раз. Передние конеч-
ности пятипалые...........Гладкокитовые. Balaenidae. (Стр. 204.)
СЕМЕЙСТВО СЕРОКИТОВЫЕ. FAMILIA ESCHRICHTIIDAE
См. определительную таблицу на странице 199. Кроме того, голова
сравнительно короткая, сжата с боков и около пяти раз короче общей
длины тела. Горловые борозды кзади несколько расходятся. Пластины
китового уса низкие, толстые, числом 130—180, с очень грубой и тол-
стой бахромой с каждой стороны.
Один род.
РОД СЕРЫЕ КИТЫ. GENUS ESCHR1CHTHJS Gray, 1864
16. Серый кит. Eschrichtius gibbosus Erxleben, 1777
Табл. XVIII, 2. Карта 57
Наибольший размер 15,5 м; самки на 55—87 см больше самцов.
Новорожденные — 3,6—5,5 м. Пластин в одном ряду 138—180. Наи-
большая высота пластин (включая часть, скрытую в десне) 48 см.
199
Грудные плавники тупее и шире, чем у полосатиков, но уже и острее,
чем у гладких китов. Общая окраска серая; по всему телу разбросаны
светлые пятна — бывшие места прикрепления усоногих ракообраз-
ных, результаты ушибов, ссадин и т. д.
Прибрежный пояс северной половины Тихого океана и Чукотского
моря. Два стада: чукотско-калифорнийское мигрирует от мест зимовок
в прибрежных водах Калифорнии к летним местам нагула в Беринго-
вом проливе и Чукотском море; охотско-корейское стадо кочует
между Южной Кореей, где зимует, и Охотским морем — местом
нагула. Прежде (возможно даже до XVIII века) водился в Балтий-
ском море.
Питается исключительно придонными организмами — бокоплавами
и другими низшими раками, морской капустой. Спаривание — с де-
кабря по март, максимум в феврале. Беременность длится около года.
Щенение происходит в мелких лагунах Нижней Калифорнии и Кореи.
Разгар ее наступает в январе. Лактация длится, вероятно, около полу-
года. Половой цикл, как и у других усатых китов, двухгодичный. По-
ловозрелое™ достигают в двухлетнем возрасте, некоторые — немного
старше года. Сосунки растут быстро: по 1,6—2,7 см в сутки. Такой
быстрый рост связывают с исключительной питательностью молока, со-
держащего до 53% жира. Материнская привязанность исключительно
сильная, и мать защищает детеныша, рискуя жизнью, иногда даже пер-
вая нападает на лодки охотников. Обитатель прибрежных вод, где и
кормится, и плодится, и спасается от врагов на отмелях, и избавляется
от паразитов, скребясь о грунт.
СЕМЕЙСТВО ПОЛОСАТИКОВЫЕ. FAMILIA BALAENOPTERIDAE
См. определительную таблицу на странице 199. Кроме того, на
горле и груди 12—14 продольных борозд. Голова сравнительно неболь-
шая, укладывающаяся в общей длине тела от 3,5 до 5 раз. Сверху она
плоская. Разрез рта более или менее прямой. Пластин китового уса
с каждой стороны челюсти 270—430. Череп при взгляде сверху более
или менее треугольной формы. Верхнечелюстные кости шире межче-
люстных.
Питаются сравнительно крупным планктоном.
Космополиты.
2 рода; представители их есть в морях СССР.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ПОЛОСАТИКОВЫХ
1(2). Грудные плавники умеренных размеров, не более V? общей
длины тела, ланцетообразной формы с ровными краями. Продольных
полос более 40. Голова и нижние челюсти гладкие, без шишек . . . .
...........................Полосатики. Balaenoptera. (Стр. 200.)
2(1). Грудные плавники огромные, не менее */4 общей длины тела,
с фестончатыми краями. Продольных полос на нижней части тела не
более 36. На голове и нижних челюстях многочисленные шишки разме-
ром с куриное яйцо. . . Горбатые киты. Megaptera. (Стр. 203.)
РОД ПОЛОСАТИКИ. GENUS BALAENOPTERA Lacepede, 1804
См. определительную таблицу на странице 200.
Космополиты.
4 вида; все встречаются в морях СССР.
200
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ПОЛОСАТИКОВ
1(6). Нёбо светло-розовое или белое. Бахрома усов не черная.
Верх тела серовато-черный, низ белый или светло-серый. Рострум че-
репа при взгляде сверху благодаря прямым краям имеет треугольную
форму .............................................................2
2(3). Грудные плавники с белым пояском или наполовину белые.
Пластины китового уса или сплошь белые, или желтые с белой
бахромой . Малый полосатик. Balaenopterus acutirostrata. (Стр. 201.)
3(2). Грудные плавники сверху темные, иногда с белой оторочкой.
Бахрома пластин1 китового уса серая................................4
4(5). Спинной плавник расположен над заднепроходным отвер-
стием. Граница между темной спиной и светлым брюхом ровная. Ре-
бер 13—14 пар. Позвонков 55—57 ......................................
............................Сейвал. Balaenoptera borealis. (Стр. 202.)
5(4). Спинной плавник расположен впереди заднепроходного от-
верстия. Пластины китового уса черные с серой бахромой. Граница
между темной спиной и светлым брюхом крупнофестончатая. Ребер
15—16 пар. Позвонков 60—63 ..........................................
................. Финвал. Balaenoptera physalis. (Стр. 202.)
6(1). Нёбо черное. Ус сплошь черный. Все тело темно-серое с си-
ним отливом, а брюхо лишь немного светлее. Рострум черепа при рас-
смотрении сверху благодаря дугообразно выпуклым краям ложкооб-
разной формы . . . Синий кит. Balaenoptera musculus. (Стр. 203.)
17. Малый полосатик. Balaenoptera acutirostrata Lacepede, 1804
Табл. XVIII, 3
Длина самок 3,75—8,75 до 10 м, самки несколько больше самцов,
новорожденные 2—2,7 ж, в южном полушарии, по-видимому, крупнее.
Спинной плавник по своей форме, относительным размерам и положе-
нию больше сходен с плавником сейвала, чем финвала. На средней ли-
нии рыла хорошо развит продольный гребень, образованный слиянием
межчелюстных костей. Продольных полос на горле и брюхе 50—70.
В верхней челюсти с каждой стороны 231—325 белых или желтовато-
белых роговых пластин, наибольшая длина их 35 см и ширина при
основании 15 см. Верх головы, туловища, хвостового плавника и
весь спинной плавник черно-серые, бока светлее, низ тела белый,
грудные плавники светлые или черные с широкой белой поперечной
полосой.
От льдов Арктики и Антарктики до субтропических вод; в тропи-
ческом поясе редок; известны два случая (в 1880 и 1926 гг.) обсыха-
ния в Черном море.
Питается рыбой и пелагическими ракообразными, в северном по-
лушарии — прибрежной рыбой, в антарктических водах — планктон-
ными ракообразными (вдали от берегов).
Время спаривания и родов сильно растянуто. Беременность длится
10—11 месяцев. Охотнее других полосатиков держится у берегов, пре-
имущественно в одиночку, реже парами и совсем редко, при скоплении
пищи, группами.
Промышляется лишь в немногих местах, главным образом в Нор-
вегии. В СССР добывается случайно на Дальнем Востоке.
Географическая изменчивость выражена очень слабо. Особи с севера Тихого
океана отличаются от североатлантических чуть более коротким клювом.
201
18. Сейвал. Balaenoptera borealis Lesson, 1828
Табл. XVIII, 4. Карта 57
Длина тела самцов 18,8 ж, самок до 21 м. Окраска спины от сине-
вато-черной до черно-сероватой, бока светлей, хвостовые лопасти
сверху цвета спины, а снизу светлее или чаще серые; нижняя часть
головы и область грудных плавников обычно белые, позади грудных
плавников тянется широкая полоса цвета боков, за ней по брюху идет
белая, часто неровная полоса; на спине и боках светлые овальные
пятна; бока передней части нижней челюсти светлее верха головы,
а задняя часть челюсти темная. Продольных борозд на горле и брюхе
32—62. На каждой половине верхней челюсти 296—403 роговые пла-
стины китового уса, длина пластин 35—80 см, ширина их у оснований
от 4—5 до 25 см, цвет черный, бахрома светло-серая.
Воды от льдов Северной Атлантики и Чукотского моря до льдов
Антарктики. Теплолюбивее финвала, далеко к полюсам не заходит.
Питается мелкими пелагическими ракообразными, рыбой и голово-
ногими моллюсками. Сроки спаривания сильно растянуты, по-види-
^юму, способны к круглогодичной половой деятельности. Спаривание
обычно зимой в теплых водах, но иногда летом. Беременность длится,
надо думать, около года; роды бывают лишь в теплых широтах. Ново-
рожденный 4—5 м длины. Лактационный период длится около 5 меся-
цев. Половая зрелость достигается в возрасте не менее 18 месяцев.
Плодятся раз в два года. Чаще всего встречается поодиночке или по-
парного иногда, в годы изобилия пищи, группами до 50 и более особей.
Второстепенный объект промысла.
19. Финвал. Balaenoptera physalis Linnaeus, 1758
Табл. XVIII, 5. Карта 57
Длина тела половозрелых 17,7—27,5 м. Борозд 68—114. Верх тела
ют почти черного цвета до свинцового, бока светлее, низ белый, не-
редко окраска пятнистая, благодаря округлым, вытянутым или звезд-
чатым шрамам (отметинам) от эктопаразитов; расцветка челюстей
асимметричная; правая половина нижней челюсти и передняя часть
верхней светлые, левая половина нижней челюсти и передняя верхней
темные. Количество роговых пластин 262—433, чаще 356—365, длина
их от 10—15 до 85 см, ширина в основании от 4—5 до 25 см. Усы серо-
голубые, бахрома серая; усы первой трети правой половины усового
ряда верхней челюсти белые или желтоватые.
Все океаны, но в северном полушарии гораздо меньше, чем в юж-
ном, в северной половине Тихого океана многочисленней, чем в Север-
ной Атлантике, в наименьших количествах держится в низких широтах.
В морях СССР чаще всего встречается в Беринговом и Чукотском, не-
редко в Охотском и Японском, но отсутствует в Восточно-Сибирском,
Лаптевых и Черном.
Питается пелагическими ракообразными, рыбой, иногда поедает
головоногих моллюсков. Спаривание, по-видимому, возможно в те-
чение почти круглого года, но разгар его происходит в 2—3 зимних
месяца.
Придерживаются в основном открытых частей моря. В местах на-
гула держатся поодиночке, реже попарно и тройками.
Важнейший объект мирового китобойного промысла.
2 подвида:
202
a) В. ph. physalis Linnaeus, 1758. (Описан из района Шпицбергена.) Сравнительно
небольшой, средние размеры самцов около 19 м, самок около 20 м. Северное полу-
шарие.
Ь) В. ph. quoyi Fischer, 1830. (Описан из района Южно-Фолклендских островов.)
Сравнительно крупный, средние размеры самцов 20,4, самок 21 м. Южное полушарие.
20. Синий кит. Balaenoptera musculus Linnaeus, 1758
Табл. XVIII, 6
Длина самок до 33,3 м, самцов до 32,6 м. Параллельных полос
70—118. Роговых пластин на каждой половине верхней челюсти 270—
400; длина их до 130 см, цвет черный.
Космополит, придерживающийся открытых океанических про-
странств, предпочитает холодные воды и редко заходит в тропические.
Питается сравнительно крупными массовыми рачками. Половая
зрелость наступает не ранее 2—3 лет. Продолжительность беремен-
ности несколько меньше года. Лактационный период около 7 месяцев.
Обитает, как правило, вдали от берегов и встречается одиночками, па-
рами или тройками (редко большим числом особей). Значительных
скоплений не образует.
Важнейший вид, на котором до недавнего времени основывался
китобойный промысел в южном полушарии.
2 подвида:
а) В. ш. musculus Linnaeus, 1758. (Описан из Шотландии.) Северное полушарие.
Ь) В. гл. intermedia Burmeister, 1866. (Описан из Буэнос-Айреса.) Более крупный:
рекордная длина самки 33,27 м, самца 32,64 м. Южная часть Мирового океана.
РОД ГОРБАТЫЕ КИТЫ. GENUS MEGAPTERA Gray, 1846
См. определительную таблицу на странице 200. Кроме того, тело
короткое и толстое с огромными грудными плавниками: от !/4 до !/3
длины тела.
Один вид.
21. Горбач. Megaptera novaeangliae Borowski, 1781
Табл. XVIII, 7
Длина тела половозрелых самок 13,8—18 м, самцов 13,5—15 м.
Задний край хвостового плавника фестончатый. От нижней челюсти до
середины брюха тянутся 14—40 продольных полос. На каждой поло-
вине верхней челюсти от 270 до 400 пепельно-черных, черно-серых или
коричневатых роговых пластин, иногда 15 передних пластин белые.
Окраска крайне изменчивая (индивидуально). Верх головы, спина,
бока, спинной и хвостовой плавники сверху однообразно черные,
иногда с коричневатым оттенком; горло, грудь и брюхо от черно-серого
цвета до сплошь белого и белого с черноватыми пестринами самой раз-
нообразной величины; грудные плавники сверху бывают черными, пе-
стрыми, черно-белыми, иногда с мраморным рисунком или совсем бе-
лыми, снизу чаще белые, иногда с крапинами и пятнами, реже серые,
очень редко черные; хвостовые лопасти снизу светлые, пятнистые,
сплошь черные.
Почти все моря от Арктики до Антарктики.
Питается рыбой, придонными и пелагическими ракообразными,
иногда крылоногими и головоногими моллюсками. Спаривание растя-
нуто почти на круглый год с максимумом в зимне-весеннее время. Бе-
ременность длится 11 —11,5 месяцев. Самки, как правило, рожают
раз в два года. Лактационный период длится 5 месяцев. Детеныши
203
держатся при матери по крайней мере до года или, может быть, дольше-
Встречаются обыкновенно парами: самка и самец или мать и подрос-
ший сосунок. Держатся преимущественно в прибрежной полосе.
В XIX и особенно в начале XX в. промысел был широко распро-
странен. В результате сильно поредели стада у берегов Японии и Юж-
ной Африки.
Слабо различаются 2 подвида:
а) М. п. novaeangliae Borowski, 1781. (Описан по экземпляру из Новой Англии.)-
Северное полушарие.
b) М. п. lalandii Fischer, 1829. (Описан по экземпляру с мыса Доброй Надежды.)-
По размерам несколько крупнее номинальной формы. Южное полушарие.
СЕМЕЙСТВО ГЛАДКОКИТОВЫЕ. FAMILIA BALAENIDAE
См. определительную таблицу на странице 199. Кроме того, верх-
ние и нижние челюсти сильно изогнуты, поэтому разрез рта имеет дуго-
образную форму: череп при взгляде сверху Т-образной формы, роговые
пластины узкие, длиной до 4,5 м.
Космополиты.
3 рода; в водах СССР — 2 рода.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ГЛАДКИХ КИТОВ ФАУНЫ СССР
1(2). Голова составляет около 7з общей длины тела. Край нижней
челюсти ровный. Поверхность рыла гладкая, без нароста. Клюв черепа
очень длинный, так что скуловая ширина укладывается в длине черепа
от затылочного отверстия до конца рыла около двух раз ...........
........................Гренландские киты. Balaena. (Стр. 205.)
2(1). Голова составляет около 74 общей длины тела. Край нижней
челюсти фестончатый. Наверху рыла имеется толстый роговой нарост.
Клюв черепа короткий; скуловая ширина укладывается в длине черепа
от затылочного отверстия до конца рыла около полутора раз ....
.................................Южные киты. Eubalaena. (Стр. 204.)
РОД ЮЖНЫЕ КИТЫ. GENUS EUBALAENA Gray, 1864
См. определительную таблицу на странице 204.
Один вид.
22. Южный кит. Eubalaena glacialis Borowski, 1781
Табл. XVIII, 8
Длина самок 13,4—15,5, максимум 17 ж, самцов 13—15,5 м. Ново-
рожденные длиной 4,5—6 м. Туловище менее толстое, чем у полярного
кита, шейный перехват выражен слабее. Умеренные воды Северной
Атлантики к югу до Северо-Западной Африки и Флориды, к северу до
Шпицбергена, Исландии, Девнсова пролива. Избегает льдов, в тропи-
ческом поясе отсутствует. В настоящее время в результате перепро-
мысла крайне редок.
Основная пища — мелкие планктонные ракообразные. Плодятся
раз в два года. Спаривание, по-видимому, происходит главным образом
в конце зимы и весной. Беременность длится около года. Активнее и
подвижнее полярного кита. Держится в одиночку или небольшими
группами в 2—3 особи, но в местах скопления — стадами в сотню
зверей.
В настоящее время взят под защиту по международной конвенции.
204
Различают 3 подвида; из них 2 встречаются в водах СССР:
а) Е. g. glacialis Borowski, 1781. (Описан из вод между Норвегией и Исландией.)
Длина пластины уса не более 223 см. Умеренная Атлантика.
b) Е. g. sieboldi Gray, 1864. (Описан из Тихого океана у Японии.) Длина пла-
стины уса до 240—259 см. Умеренная полоса Тихого океана.
РОД ГРЕНЛАНДСКИЕ КИТЫ. GENUS BALAENA Linnaeus, 1758
См. определительную таблицу на странице 204.
Один вид.
13. Гренландский кит. Balaena mysticetus Linnaeus, 1758
Табл. XV1I1, 9
Размеры взрослого обычно 15—18 м, редко до 20 м. Новорожден-
ный 3—4,5 м.
Пластин китового уса 300—400 с каждой стороны, длина их до
4,5 ж, цвет черный. Верхняя часть головы, спина, грудные плавники и
хвостовой плавник сверху у взрослых обычно черные, иногда белый
цвет заходит на нижнюю часть шеи и переднюю часть брюха; неполо-
возрелые серовато или голубовато-черные. Некоторые особи (старые)
коричневатые. Очень редко встречаются альбиносы (частично или пол-
ностью).
Раньше был широко распространен в Северном Ледовитом океане,
откуда спускался до Гренландии и Лабрадора и был очень многочис-
лен в Беринговом и Охотском морях. В настоящее время в результате
перепромысла крайне редок и встречается в Гудзоновом заливе, в мо-
рях: Гренландском, Беринговом, Карском, Лаптевых, Восточно-Сибир-
ском, Чукотском, Охотском и Бофорта. Относительно более обычен
в водах, омывающих Чукотский полуостров, а наиболее редок в водах
от Баренцева моря до Восточно-Сибирского. Три основных стада:
1) шпицбергенское — мигрирует от Гренландии и Исландии к Шпиц-
бергену; 2) западногренландское — мигрирует из Девисова пролива
в Баффинов залив; 3) бсрингово-чукотское — мигрирует из моря Бо-
форта, вдоль Аляски в Чукотском море, далее в Берингово море и об-
ратно. Запасы этих стад были последовательно подорваны: первое —
в XVII в., второе — в XVI11 в., третье — в XIX в.
Питается в основном мелкими массовыми ракообразными и крыло-
видными моллюсками. Период спаривания растянут и точно не уста-
новлен. Одни считают, что он продолжается с июня по август, другие
указывают январь—февраль. Мнения о длительности беременности
расходятся: одни считают 9—10 месяцев, другие— 13—14 месяцев. Де-
теныши рождаются в холодных арктических водах. Время деторожде-
ния растянуто, но в общем приурочено к весне. Родительский инстинкт
развит очень сильно. Держится парами, в одиночку или группами по
3—5 особей.
Н. А. Бобринский
ОТРЯД НЕПАРНОКОПЫТНЫЕ. ORDO PERISSODACTYLA
Непарнокопытные — крупные наземные растительноядные копыт-
ные, млекопитающие. На передних конечностях 1, 3 или 5 пальцев, на
задних 1 или 3 пальца. Наиболее развит третий (средний) палец, че-
рез который проходит главная ось конечности. Боковые пальцы раз-
виты слабее или даже рудиментированы. Копыто среднего пальца сим-
метрично и больше других. У представителей отряда, встречающихся
в СССР, все четыре конечности имеют только один палец, одетый на
конце округлым копытом. Рогов нет.
Верхние челюсти несут долотообразные резцы. Коренные зубы
имеют плоскую жевательную поверхность с очень сложным рисунком
эмалевых петель.
Непарнокопытные населяют большую часть Африки, Переднюю,.
Среднюю, Центральную и Южную Азию (с Малайским архипелагом),.
Центральную Америку и северную половину Южной Америки. Совре-
менные непарнокопытные образуют 3 хорошо обособленных подотряда:
тапировых, носороговых и лошадиных.
В СССР встречаются представители только последнего подотряда —
семейства лошадиных.
СЕМЕЙСТВО ЛОШАДИНЫЕ. FAMILIA EQUIDAE
В отличие от других непарнокопытных, у лошадиных на всех четы-
рех конечностях хорошо развит и одет копытом только один, третий,,
палец. Боковые пальцы рудиментарны и имеют вид скрытых под ко-
жей тонких «грифельных» косточек. Волосы на верхней стороне шеи
удлинены, о'бразуя гриву. Глазница замкнутая.
Зубная формула: I у С -р Рш у М у • 2 = 42.
Первый предкоренной верхней челюсти очень мал и обычно с воз-
растом выпадает; клыки развиты только у самцов, да и то не всегда.
Единственный род.
РОД ЛОШАДИ. GENUS EQUUS Linnaeus, 1758
Охватывает около 10 видов, относящихся к 3 подродам: лошадей,
ослов и зебр. В СССР имеется только 1. дикий вид — кулан и 2 вида —
лошадь и осел разводятся как домашние животные.
206
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА EQUUS, ВСТРЕЧАЮЩИХСЯ
В СССР
1(4). Хвост покрыт длинными волосами только на конце. Грива
короткая, стоячая. «Челка» отсутствует. Уши длиннее 17 см. Слуховые
трубочки черепа длинные, и концы их хорошо видны при рассматрива-
нии черепа сверху (подрод Asinus) . . . ..............2
2(3). Уши относительно короткие (длина их менее 20 см). Окраска
светло-желтая с белым; темной поперечной полосы на плечах нет.
Наибольшая длина черепа взрослых более 47 см.................
.............................Кулан. Equus hemionus. (Сгр. 208.)
3(2). Уши длинные (более 20 см длины). Окраска иная. На пле-
чах — темная поперечная полоса. Наибольшая длина черепа обычно
менее 47 см...........................Домашний осел. Equus asinus.
207
4(1). Хвост на всем протяжении покрыт длинными волосами.
Длинные волосы гривы свешиваются на боковую сторону шеи. На лоб
свисает пучок длинных волос («челка»). Уши короче 17 см. Слуховые
трубочки черепа короткие и не видны при рассматривании черепа
сверху .... . . . .... Лошадь. Equus caballus.
1. Кулан. Equus hemionus Pallas, 1775
Табл. XIX, 1. Карта 58
Встречается в Центральной, Средней и Передней Азии. В нашей
стране в XVIII в. был широко распространен по низменностям Сред-
ней Азии и Казахстана, но позднее в большинстве районов был истреб-
лен. В настоящее время встречается в Бадхызском заповеднике, близ
г. Кушки (Южная Туркмения), где обитает до 600 голов этого интерес-
ного животного. Акклиматизирован на острове Барса-Кельмес, в Араль-
ском море. Возможны заходы в Прибалхашье из Китая.
Куланы обитают в равнинных и холмисто-увалистых пустынях и
полупустынях. Летом держатся небольшими табунами, главным обра-
зом у водопоев, зимой же сбиваются в более крупные стада, кочующие
по пустыне. Гон в Туркмении проходит в июле — августе. Молодые ро-
дятся преимущественно в мае и июне. Летом питается главным обра-
зом злаками, осенью — полынью и 'солянками, зимой — полынью, зла-
ками, ветками кустарниковых солянок.
Охота на куланов в СССР запрещена.
В Туркмении встречается особый подвид Е. h. onager B-oddaert, 1785, отличаю-
щийся от типичной формы из Монголии более мелкими размерами и очень светлой
окраской.
Н. А. Бобринский
ОТРЯД ПАРНОКОПЫТНЫЕ. ORDO ARTIODACTYLA
Крупные и средней величины наземные (кроме земноводного -беге-
мота) копытные млекопитающие. Главной опорой тела служат одина-
ково хорошо развитые третий и четвертый пальцы, между которыми
проходит главная ось конечности; копыта этих пальцев асимметричны.
Второй и пятый пальцы либо отсутствуют, либо более или менее недо-
развиты и обычно имеют вид копытец, лишенных костного скелета.
Многие виды имеют рога той или иной формы (обычно рога имеют
только самцы). Резцы верхней челюсти сильно редуцированы или от-
сутствуют. Коренные зубы либо многобугорчатые, либо имеют на своей
жевательной поверхности эмалевые лунки (рис. 72).
Парнокопытные населяют все материки, кроме Антарктиды и Ав-
стралии (в последнюю завезены человеком). Встречаются на многих
островах Северного полярного моря, на островах Индонезии, на Мада-
гаскаре, на Огненной Земле. Ряд форм завезен на острова Новой Зе-
ландии.
Отряд подразделяется на 3 подотряда: нежвачные, мозоленогие и
жвачные.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДОТРЯДОВ ПАРНОКОПЫТНЫХ
ФАУНЫ СССР
1(2). Морда сильно вытянутая, конусообразная, оканчивающаяся
голым, плоским хрящеватым кружком («пятачком»), на котором откры-
ваются ноздри. Тело покрыто жесткой щетиной. Клыки верхней че-
люсти, изгибаясь, направлены концами в стороны или вверх. Коренных
зубов в каждой челюсти 7. На их жевательной поверхности — ряд ту-
пых бугорков. Глазница незамкнутая ..........................
.........................Нежвачные. Nonruminantia. (Стр. 210.)
2(1). Морда не столь удлиненная, иной формы и не несет на конце
хрящеватого «пятачка». Тело покрыто более или менее мягкими воло-
сами. Клыки верхней челюсти направлены вершинами вниз. Коренных
зубов в каждой челюсти не более 6. На их жевательной поверхности —
эмалевые лунки........................................... 3
3(4). Как передние, так и задние конечности имеют только по
2 пальца, несущих спереди ногтевидные копыта. На спине один или два
жировых горба. На груди и ногах имеются большие роговые мозоли.
В верхней челюсти имеется одна пара резцов. Клыки нижних челюстей
отделены от резцов значительным промежутком..................
...........................Мозоленогие. Tylopoda. (Стр. 212.)
14 Н. А. Бобринский и др.
209
4(3). На конечностях, кроме двух хорошо развитых средних паль-
цев, имеются еще в той или иной степени недоразвитые боковые (вто-
рой и пятый) пальцы с копытцами. Жировых горбов на спине нет. Ро-
говые мозоли на груди и ногах отсутствуют. Резцов в верхней челюсти
нет. Клыки нижней челюсти непосредственно следуют за резцами . . .
................................Жвачные. Ruminantia. (Стр. 212.)
Подотряд НЕЖВАЧНЫХ. Subordo NONRUMINANTIA
К данному подотряду относятся семейства свиней, пекари и беге-
мотов. В Советском Союзе встречаются представители только первого
из них.
СЕМЕЙСТВО СВИНЫЕ. FAMILIA SUIDAE
Представители этого семейства населяют Африку (с Мадагаска-
ром), Европу и Азию (с Малайским архипелагом).
В СССР только один род.
РОД КАБАНЫ. GENUS SUS Linnaeus, 1758
5 или 6 видов этого рода населяют Европу, Азию и Северную Аф-
рику. В Советском Союзе только один вид.
1. Кабан. Sus scrota Linnaeus, 1758
Табл. XIX, 2—4. Карта 58
Встречается в Европе, Северной Африке и в Азии (на севере до
Южной Сибири). В Европейской части Советского Союза распростра-
нен к востоку до Ленинградской, Псковской, Смоленской, Московской,
Калужской, Брянской, Орловской, Воронежской, Харьковской, Кирово-
градской областей и Молдавской ССР. Известны заходы и в более во-
сточные районы; акклиматизирован в некоторых охотничьих хозяйствах
Московской, Рязанской и других областей центральной части страны.
Широко распространен на Кавказе (к северу до низовьев р. Кубани^
района г. Краснодара и г. Армавира, среднего течения р. Терека).
Встречается в низовьях рек Кумы, Волги и Урала. В Казахстане север-
ная граница тянется от среднего течения Малого и Большого Узеней,
через район г. Уральска, южнее Оренбурга, на Кустанай — Курган —
Петропавловск — Кокчетав — Караганду к оз. Зайсан. Но распро
странение его в Казахстане и в Средней Азии носит спорадический
характер, так как кабан ’избегает пусты-нных и полупустынных про-
странств. Обитает на Алтае, в Саянах, в Прибайкалье, в Южном За-
байкалье, на Амуре (вниз до Комсомольска), по всему Ус-сурий-.
скому краю.
Кабаны селятся в весьма разнообразных угодьях: низменных ле-
сах, в зарослях камыша и кустарника, по берегам рек и озер, по боло-
там (и в горах, где поднимаются местами выше зоны древесной расти-
тельности. Для существования кабана необходима близость какого-
либо водоема. Образ жизни в основном ночной. Всеядное животное,
пожирающее всевозможные растительные вещества (корневища, кор-
неплоды, плоды, желуди, орехи, ягоды, молодую зелень и т. п.),
а также насекомых, червей, моллюсков, мелких грызунов; не брезгует
и падалью. Местами сильно вредит посевам, огородам и бахчам. Ходят
табунами. Самки с поросятами (годовиками и сеголетками) держатся
210
обычно отдельно от взрослых самцов. Гон происходит в ноябре—ян-
варе. В это время секачи (взрослые самцы) разыскивают табуны са-
мок и яростно дерутся друг с другом. Беременность взрослых маток
длится 130—140 дней, молодых — несколько короче. Число поросят
в помете — 2—15 (обычно 4—6). Молодые родятся в марте — мае^
Самки достигают половой зрелости на втором году жизни.
В ряде районов юга нашей страны добывается значительное коли1
чество кабанов ради мяса, шкуры и щетины.
Рис. 70. Череп кабана (Sus acrofa). Вид сбоку.
Рис. 71. Череп двугорбого верблюда (Camelus bactria-
nus). Вид сбоку.
В СССР обитает 5 подвидов кабанов:
а) Западноевропейский кабан, S. s, scrofa Linnaeus, 1758, Размеры средние: наи-
большая д :нпз черепа взрослых 37—41 см. Окраска темная, подшерсток рыжевато-
бурый. Западные области Европейской части СССР, Западная Европа.
Ь) Кавказский кабан, S. s. attila Thomas, 1912. Размеры крупные: наиболь-
шая длина череча 41—47 см. Окраска немного светлее, Кавказ, Закавказье, низовье
Волги.
с) Среднеазиатский кабан, S. s, nigripes Bl., 1875. Размеры крупные. Окраска
туловища светлая, но ноги почти черные. Подшерсток светлый, песчаного цвета.
Средняя Азия, Казахстан,
d) Сибирский кабан, S. s. sibiricus Staffe, 1922, Самая мелкая форма: наибольшая
длина черепа 37—40 см. Окраска черно-бурая. По щекам тянется сероватая полоса,
Прибайкалье и Забайкалье,
е) Дальневосточный кабан, S, s, ussuricus Heude, 1888. Самый крупный подвид:
наибольшая длина черепа 49—51 см. Окраска изменчива, но преобладают темные
особи. На щеках светлая продольная полоса. Юг Дальнего Востока,
14* 211
Подотряд МОЗОЛЕНОГИХ. Subordo TYLOPODA
К этому подотряду относятся верблюды и ламы.
В Советском Союзе разводят 2 вида верблюдов: одногорбый вер-
блюд, Camelus dromedarius, Linnaeus, 1758 и двугорбый верблюд, Са-
melus bactrianus, Linnaeus, 1758. Дикие (одичавшие?) двугорбые вер-
блюды обитают в пустынях Центральной Азии.
Подотряд ЖВАЧНЫХ. Subordo RUMINANTIA
К подотряду жвачных относятся семейства: оленьковые, оленьи,
жирафовые, вилорогие и полорогие. В Советском Союзе встречаются
представители семейств оленьих и полорогих.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ ЖВАЧНЫХ ФАУНЫ СССР
1(2). Ноздри расставлены широко; промежуток между ними равен
или превышает расстояние от нижнего края ноздрей до края верхней
губы. Рога, если они имеются, представляют собой костные стержни
(у взрослых — ветвистые), которые ежегодно спадают и вырастают
вновь.
Молодые
отросшие же
Рис. 72. Верхние коренные зубы:
сверху — оленя (Cervus); снизу — козла
(Capra).
рога мягки и покрыты короткой бархатистой шерстью,
твердые и голые. Коренные зубы с низкими коронками
(бранхиодонтного типа); их наруж-
ные вертикальные грани не доходят
до края альвеолы, а соединяются
валиком, идущим вдоль основания
зуба. У ряда форм имеются клыки
верхней челюсти. Между лобной,
носовой, слезной и верхнечелюст-
ной костями имеется с каждой сто-
роны по большому отверстию не-
правильной треугольной формы . .
. . Оленьи. Cervidae. (Стр. 212.)
2(1). Ноздри расположены на-
столько близко друг к другу, что
промежуток между ними значи-
тельно меньше расстояния между
нижним краем ноздри и краем верхней губы (исключение составляют
зубры и быки яки). Рога, если они есть, имеют вид рогового футляра,
сидящего на костном стержне; они не ветвятся и не сменяются в тече-
ние всей жизни. Коренные зубы с высокими коронками (гипсодонтного
типа); наружные вертикальные грани их доходят до самых краев аль-
веол (по крайней мере на заднекоренных зубах). Верхних клыков нет.
Между носовой, лобной, слезной и верхнечелюстной костями отверстия
нет или оно имеет форму продольной щели...........................
................................Полорогие. Bovidae. (Стр. 221.)
СЕМЕЙСТВО ОЛЕНЬИ. FAMILIA CERVIDAE
Распространено в Европе, Северной Африке, в Азии, Южной Аме-
рике. Акклиматизировано в Австралии и в Новой Зеландии.
Распадается на 2 подсемейства: кабарожьих и собственно-оленьих.
212
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ
СЕМЕЙСТВА ОЛЕНЬИХ ФАУНЫ СССР
1(2). Рогов нет. Верхние клыки у самцов очень велики и далеко вы-
даются из-под верхней губы; у самок длина их превышает высоту коро-
нок коренных зубов. Длина головы и туловища менее 110 см. Задние
конечности почти на !/з длиннее передних. Слезных ямок нет. Общая
длина черепа меньше
17 см. Слезная кость без
вдавления . Кабарожьи.
Moschinae. (Стр. 213.)
2(1). У самцов, а у
некоторых видов и у са-
мок имеются рога. Клыки
верхних челюстей, если
они есть, у самцов при-
мерно равны по высоте
коренным зубам, а у са-
мок много ниже их. Дли-
Рис. 73. Череп самца кабарги
(Moschus moschiferus), вид сбоку.
на головы и туловища
более 110 см. Задние конечности примерно такой же длины, как и пе-
редние. Общая длина черепа взрослых более 18 см. Слезные кости
с вдавлением.........................Оленьи. Carvinae. (Стр. 214.)
Подсемейство КАБАРОЖЬИХ. Subfamilia MOSCHINAE
Представители подсемейства обитают в Центральной и Восточной
Азии. Подсемейство содержит только один род.
РОД КАБАРГИ. GENUS MOSCHUS Linnaeus, 1758
2. Кабарга. Moschus moschiferus Linnaeus, 1758
Табл. XIX, 7—8. Карта 59
Населяет в пределах СССР Алтай (с Саурами и Кузнецким Ала-
тау), Саяны, Восточную Сибирь, от Енисея до Охотского моря (на
Камчатке отсутствует), и Амуро-Уссурийский край. Водится также
в Корее, Северо-Восточном Китае и Северной Монголии.
Обитает преимущественно в горной тайге, богатой скалами и каме-
нистыми россыпями. Держится либо в одиночку, либо небольшими
группами, состоящими из самок и молодых. Основная пища — лишай-
ники. Гон протекает обычно в ноябре и декабре. Беременность длится
185—195 дней. Большая часть самок родит в июне 1—2 телят.
Добывается ради мяса, шкуры и мускуса, находящегося в железе
на брюхе самцов.
Из ряда описанных географических рас кабарги, обитающих в СССР, сколько-
нибудь хорошо обособленными можно считать следующие:
а) Сибирская кабарга, М. m. moschiferus Linnaeus, 1758. Размеры довольно круп-
ные: общая длина черепа до 164 мм. Окраска коричневая с заметной пятнистостью.
Носовые кости длинные. Алтай, Саяны, Енисейская тайга, Западная Якутия, Забайкалье.
Ь) Дальневосточная кабарга, М. m. parvipes Holl, 1911. Размеры мельче: общая
цлина черепа до 152 мм. Окраска темная, со слабой пятнистостью. Носовые кости
более короткие. Амуро-Уссурийский край.
К этой форме очень близка М. m. arcticus Fl. из Восточной Якутии.
с) Сахалинская кабарга, М. m. sachalinensis FL, 1929. Самая мелкая форма: об-
щая длина черепа до 151 мм. На черепе, в отличие от других форм, межглазничный
промежуток меньше (реже равен) ширины черепа за межглазничными отростками.
Сахалин.
213
Подсемейство ОЛЕНЬИХ. Subfamilia CERVINAE
Олени обитают в Европе, Северной Африке, Азии, Северной и Юж-
ной Америке.
Из ряда родов этого подсемейства в СССР встречаются
четыре.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ОЛЕНЬИХ ФАУНЫ СССР
1(6). Кожа на морде между ноздрями голая или покрыта воло-
сами, но с небольшим треугольным голым участком. Поверхность глав-
ного ствола, а обычно и ветвей рогов шероховатая, с костными рубчи-
ками и бугорками. Надглазничные отростки рогов, если они имеются,
не уплощены на концах. Задний край сошника не срастается с нёб-
ными костями и не делит хоаны на две .половины. Передний край глаз-
ницы расположен впереди уровня заднего конца зубного ряда ... 2
2(5). Вокруг или по бокам хвоста выделяется пятно светлого
меха — «зеркало». Все или почти все пространство между ноздрями
голое. Верхняя губа не увеличена и не нависает над нижней. Высота
в холке менее 170 см. Общая длина черепа менее 50 см. Межчелюст-
ные кости соприкасаются с носовыми............................3
3(4). Длина хвоста более 3 см\ он выступает из меха и потому
хорошо заметен. Рога имеют 1 или 2 надглазничных отростка. Длина
слезной кости не меньше передне-заднего диаметра глазницы ....
...........................Настоящие олени. Cervus. (Стр. 214.)
4(3). Длина хвоста менее 3 см\ он не выступает из окружающего
его меха. Рога без надглазничных отростков. Длина слезных костей
меньше передне-заднего диаметра глазницы.........................
..................................Косули. Capreolus. (Стр. 218.)
5(2). Задняя часть крупа того же цвета, что и остальные части
туловища; «зеркала» нет. Вся верхняя губа, за исключением неболь-
шого треугольного голого участка между ноздрями, покрыта воло-
сами. Верхняя губа очень большая и нависает над нижней. Высота
в холке взрослых особей более 170 см. Общая длина черепа более
50 см. Межчелюстные кости не соприкасаются с носовыми............
........................................Лоси. Alces. (Стр. 219.)
6(1). На морде пространство между ноздрями сплошь покрыто во-
лосами. Поверхность рогов гладкая. Надглазничные отростки рогов
уплощены на концах. Сошник срастается с нёбными костями и делит
хоаны на 2 половины. Передний край глазницы расположен позади
уровня заднего конца зубного ряда ...............................
...........................Северные олени. Rangifer. (Стр. 220.)
РОД НАСТОЯЩИЕ ОЛЕНИ. GENUS CERVUS Linnaeus, 1758
Ареал рода включает большую часть Европы, Северную Африку,
Азию (кроме ее северных частей), умеренную зону Северной Америки.
В Советском Союзе обитает 3 вида настоящих оленей, относя-
щихся к различным подродам (один из них завезен из Западной Ев-
ропы) .
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА НАСТОЯЩИХ ОЛЕНЕЙ
ФАУНЫ СССР
1(4). Размеры мелкие: высота в плечах самцов менее 115 см, са-
мок менее 100 см. Хвост более длинный: длина его с волосами равна
или превышает длину уха. Хвост сверху черный, с белой каймой по
214
бокам. «Зеркало» на крупе белое. Летом спина и бока в крупных белых
пятнах, зимой пятна слабо заметны или не выражены. Каждый рог не-
сет только один надглазничный отросток. Наибольшая длина черепа
менее 35 см.....................................................3
2(3). Вершины рогов самцов уплощены в виде лопаточек с не-
сколькими отростками. Голое пространство на конце морды невелико,
Рис. 74. Черепа оленей (сбоку):
А — олень обыкновенный (Cervus elaphus); Б — косуля (Capreolus
capreolus); В — лось (Alces alces); Г — северный олень (Rangifer
tarandus).
верхний край его едва поднимается выше уровня нижних краев нозд-
рей. Хвост значительно длиннее уха. Волосы, растущие у мочеполового
отверстия самца, удлинены и образуют кисть. Наибольшая длина че-
репа менее 30 см. Клыков в верхних челюстях нет....................
...........................Лань. Cervus (Dama) dama. (Стр. 216.)
3(2). Конечные части концов рогов самцов острые, неуплощенные.
Голое пространство на конце морды (носовое «зеркало») большое: его
верхний край поднимается почти до уровня верхних углов ноздрей.
215
Длина хвоста примерно равна длине уха. Волосы у мочеполового от-
верстия самца короткие и не образуют кисти. Наибольшая длина че-
репа больше 33 см. В верхней челюсти есть клыки................
.................Пятнистый олень. Cervus (Sica) nippon. (Стр. 216.)
4(1). Размеры крупнее: высота в плечах самцов свыше 120 см, са-
мок свыше 100 см. Хвост короткий. Длина его с волосами меньше
длины уха. Хвост сверху светлый. Светлое пятно на крупе («зеркало»)
ржавое или с ржавым оттенком. Спина и бока тела однотонные, без
пятен. Надглазничных отростков на каждом роге взрослых быков
обычно 2. Наибольшая длина черепа взрослых более 35 см.........
............Благородный олень. Cervus (Cervus) elaphus. (Стр. 216.)
Подрод ланей. Subgenus Dama Fischer, 1775
3. Лань. Cervus dama Linnaeus, 1758
Табл. XX, 1. Карта 59
В диком состоянии в настоящее время встречается только в Ма-
лой Азии и в Северо-Западной Африке. Полудомашние лани содер-
жатся в парках и охотничьих хозяйствах многих стран Западной Ев-
ропы.
В Советском Союзе небольшое количество ланей живет в лесах
Литвы, в Молодечненской области Белоруссии, в некоторых охотничьих
хозяйствах Украины и центральных районов, куда были завезены из
Западной Европы.
Лесные стадные животные питаются травой и ветками кустарников
и деревьев. Гон — в октябре. Беременность лани длится около 8 меся-
цев. В приплоде 1, редко 2 теленка.
Подрод.пятнистых оленей. Subgenus Sica Scleter, 1870
4. Пятнистый олень. Cervus nippon Temminck, 1838
Табл. XX, 2, 3. Карта 59
Населяет южную часть Восточной Азии. В СССР прежде обитал
только в Уссурийском крае. Ныне завезен и удачно акклиматизиро-
ван во многих заповедниках и охотничьих хозяйствах средней и южной
полосы Европейской части СССР. Разводится в оленесовхозах ради
ценных молодых рогов — пантов.
Обитает преимущественно в смешанных и лиственных лесах с под-
леском. В летнее время питается травой, листьями и побегами кустар-
ников и молодых деревьев, зимой использует, главным образом, дре-
весно-веточный корм. Гон — в сентябре — ноябре. Беременность самок
длится семь с половиной месяцев. В помете 1, очень редко 2 теленка.
Самцы сбрасывают рога в апреле —мае. Очищение рогов от кожного
покрова происходит в августе.
В СССР обитает особый подвид С. п. hortulorum Swinchoe, 1864.
Подрод настоящих оленей. Subgenus Cervus Linnaeus, 1758
5. Благородный олень. Cervus elaphus Linnaeus, 1758
Табл. XX, 4, 5. Карта 60
Распространен в Европе (на север и восток до Средней Сканди-
навии, Прибалтики, Белоруссии и Воронежской области), в Северной
Африке, в Азии (на север до Южной Сибири) и в Северной Америке.
216
В Европейской части СССР встречается в Прибалтике (главным
образом, в Литве), в Белоруссии (в Беловежской пуще, к югу от г. Мо-
лодечно, к западу от Минска и в некоторых других пунктах). На Укра-
ине (главным образом, на Карпатах и в горах Крыма), в Воронежском
заповеднике. Акклиматизирован в ряде охотничьих хозяйств и запо-
ведников западных, центральных и южных районов.
Значительное пространство занято оленями на Северном Кавказе
и в Закавказье. Они заходят в горы Копет-Дага. Обитают в тугаях
по рекам Пянджу и Аму-Дарье. В небольшом количестве сохранились
в горах Восточного Тянь-Шаня. Населяют горнотаежные районы Ал-
тая, Салаирского кряжа, Кузнецкого Алатау, Саян и Присаянья, При-
байкалья, Забайкалья, Южной Якутии, Амурского и Уссурийского
краев.
Обитает как в горных, так и равнинных лесах, предпочитая сме-
шанные и разреженные насаждения. В горах летом поднимается до
альпийских лугов, зимой же спускается в более низкие зоны. Обитает
также в тугаях и камышовых зарослях пойм больших рек. Питается
травянистыми и древесно-кустарниковыми растениями; последние име-
ют особое значение в питании оленя зимой. Поедают также ягоды,
плоды, грибы, лишайники. Охотно посещают солонцы. Гон происходит
осенью (обычно с конца августа до середины октября). Быки в это
время разыскивают самок и дерутся друг с другом. Сильные самцы
собирают целые «гаремы» самок. Зимой олени нередко собираются
в стада, но взрослые самцы держатся обычно отдельно. Беременность
оленух длится 34—35 недель. В мае или в начале, июня они родят
в укромном месте 1, редко 2 телят, которые вскоре могут следовать за
матерью. Во время течки самка отгоняет теленка, но потом он обычно
вновь возвращается к ней.
Во многих областях с целью восстановления запасов добыча оле-
ней запрещена. Но там, где численность их высока (например, в Кры-
му), производится планируемый отстрел этих животных. Молодые, еще
растущие рога (панты) сибирских оленей — маралов и изюбрей служат
сырьем для выработки лекарственного средства — пантокрина. В ряде
районов Южной Сибири имеются специальные совхозы для полуволь-
ного разведения маралов.
В Советском Союзе обитает ряд подвидов благородных оленей, которые могут
быть объединены в 3 основные группы, отличающиеся, главным образом, формой
рогов самцов.
А. Европейская группа. Рога взрослых самцов на концах несут не-
сколько отростков, расходящихся от одного места в различные стороны, образуя
«крону», или «чашу». Размеры животных средние.
К этой группе относятся следующие подвиды:
а) Среднеевропейский олень, С. е. elaphus Linnaeus, 1758. Размеры средние: вы-
сота в холке быков до 130 см. Окраска летнего меха интенсивная, рыжевато-коричне-
вая. «Зеркало» рыжеватое. Рога взрослых быков многоветвистые, обычно с хорошо
развитой кроной. Средняя Европа. В СССР — Прибалтика, Белоруссия, Украина (кроме
Крыма), Воронежская область.
Ь) Крымский олень, С. е. brauneri Charlemagne, 1920. Размеры примерно такие же,
как у предыдущей формы. Окраска менее интенсивная, с примесью желтоватых тонов.
Рога с меньшим числом отростков (не более 6) и часто без кроны. Горная часть Крыма.
с) Кавказский олень, С. е. maral Gray, 1850. Размеры более крупные: высота
в холке самцов до 135 см. Окраска менее насыщенная, желтовато-коричневая, обычно
с темными бедрами и плечами. Рога большие, но с малым числом отростков; они кон-
чаются либо кроной, либо развилкой двух концов. Кавказ.
Б. Равнинно-среднеазиатская группа. Включает только один
подвид:
d) Бухарский олень, С. е. bactrianus Lydekker, 1900. Размеры относительно мел-
кие: высота в холке самцов обычно до 120 см. Окраска летнего меха желтовато-серая
(песчаного цвета). Рога легкие, обычно не более чем с 5 концами; крона образуется
217
очень редко. Поймы рек Пянджа и Аму-Дарьи, раньше также Сыр-Дарьи. Близок
к североиндийской форме.
В. Сибирская группа. Подвиды этой группы характеризуются большими
рогами без кроны. На месте второго отростка ствол их делает резкий изгиб. Размеры
крупные. Группа объединяет следующие формы:
е) Семиречный олень, С. е. songaricus Severtzov, 1873. Размеры крупные: высота
в холке взрослых до 150 см. Окраска зимнего меха довольно темная. Горы Тянь-Шаня.
f) Марал, С. е. sibiricus Severtzov, 1873. Размеры такие же, как у предыдущей
формы, но окраска зимнего меха более светлая, серовато-буровато-желтоватая. Шея,
плечи и брюхо буроватые. Алтай, Саяны.
g) Изюбрь, С. е. xanthopygus Milne-Edwards, 1867. Размеры несколько мельче: вы-
сота в холке самцов до 145 см. Окраска зимнего меха более темная, а летнего
более красноватая, чем у марала. Рога заметно более легкие. Забайкалье, Якутия,
Дальний Восток.
РОД КОСУЛИ. GENUS CAPREOLUS Gray, 1821
Обитает в средней и южной полосе Европы, Передней и Средней
Азии, в Южной Сибири, Северной Монголии, Дальнем Востоке. Един-
ственный вид.
6. Косуля. Capreolus capreolus Linnaeus, 1758
Табл. XX, 6, 7. Карта 61
В Европейской части СССР распространена на восток до Ленин-
градской, Новгородской, Калининской, Московской, Брянской, Орлов-
ской, Харьковской, Воронежской, Днепропетровской областей и Молда-
вии. Обитает в горных лесах Крыма. Широко распространена в лесных
районах Северного Кавказа ’ и Закавказья. В Средней Азии встре-
чается в горах Тянь-Шаня и на северных склонах Алайского и Гиссар-
ского хребтов. Обитает в камышах рек Прибалхашья. В Сибири рас-
пространена от Урала до Тихого океана; северная граница ареала
идет примерно от Перми через верховья Тавды, к устью р. Иртыша
и далее вдоль правого берега этой реки к району г. Томска, к нижнему
течению р. Ангары, верховьям р. Лены, вдоль ее.до устья рек Витима
и Олекмы к Удскому заливу и далее через район г. Комсомольска-на-
Амуре к Советской Гавани. Южная граница распространения косули
на Урале и в Западной Сибири идет от г. Уральска к южным отрогам
Урала, через Курганскую, Омскую и Новосибирскую области к Алтаю.
Любимые места обитания косуль — разреженные, смешанные с
обильным подседом леса, перемежающиеся с полянами. В Средней
Азии встречаются в зарослях камыша и в тугаях. В горах поднима-
ются до пределов леса. Питаются травой, листьями и побегами кустар-
ников и деревьев, корой их, плодами, ягодами, мхами и лишайниками.
Гон продолжается с конца июля по середину сентября. Продолжитель-
ность беременности около 9 месяцев. Телята (1—2, но иногда 3—4)
родятся в апреле и мае. Осенью косули, сбившись в стайки, предпри-
нимают нередко дальние миграции. Рога у самцов сбрасываются в кон-
це осени, в декабре — январе начинается рост новых рогов, которые
полностью окостеневают в мае—июне.
Ценный промысловый вид, особенно в Сибири. В ряде областей
добыча косуль запрещена.
Подвиды фауны СССР:
а) Европейская косуля, С. с. capreolus Linnaeus, 1758. Размеры мелкие: высота
в плечах самцов 75—92 см, наибольшая длина черепа 19—22 см. Рога тонкие со
слаборазвитыми бугорками. Летний мех рыжий, «зеркало» белое, голова буроватая.
Западные области Европейской части СССР, Крым, Закавказье (в последней —
особая natio— С.с.с. armenius Blacler, 1916).
b) Сибирская косуля, С. с. pygargus Pallas, 1771. Размеры крупные: высота в плечах
самцов 85— 123 см, наибольшая длина черепа 22—25 см. Рога массивные с сильно
218
развитыми бугорками. Летний мех туловища и головы рыжий, «зеркало» белое.
Восточные окраины Европейской части СССР, Сибирь, Средняя Азия.
с) Дальневосточная косуля, С. с. bedfordi Thomas, 1908. Размеры средние: наи-
большая длина черепа 21—24 см. Окраска летнего меха насыщенно рыжая, более
яркая, чем у сибирской формы. «Зеркало» в зимнем мехе с рыжеватым оттенком.
Рога, как у предыдущего подвида. Южная часть Дальнего Востока.
d) Северокавказская косуля, С. с. caucasicus Dinnik, 1910. Размеры средние: наи-
большая длина черепа 22—24 см. Окраска и строение рогов, как у сибирского под-
вида. Северный Кавказ.
РОД ЛОСИ. GENUS ALCES Gray, 1821
Лоси распространены в лесной и лесостепной зонах Восточной Ев-
ропы, Азии и Северной Америки. Род включает только один вид.
7. Лось. Alces alces Linnaeus, 1758
Табл. XX, S, 9. Карта 62
В результате хищнического промысла в царской России числен-
ность лосей к началу настоящего столетия сильно сократилась и во
многих областях страны они исчезли. Но в результате охраны и ряда
биотехнических мероприятий численность лосей в последние десятиле-
тия в СССР снова стала весьма большой и прежний ареал этого жи-
вотного почти полностью восстановлен. В настоящее время северная
граница области распространения лосей в СССР совпадает с северным
пределом лесной области и лишь в бассейне р. Оби значительно спу-
скается к югу. Иногда заходят и в тундру. На востоке лоси распро-
странены до Тихого океана, но на Камчатке и на Сахалине их нет.
Южная граница сплошного ареала лосей в основном совпадает
в настоящее время с южным пределом лесной зоны, но они встре-
чаются и во многих лесных массивах лесостепной полосы и заходят
по долинам рек даже в степные районы.
Лось биологически связан с лесом. Обитает в лесах самых раз-
личных типов, но предпочитает сырые, заболоченные насаждения, в ко-
торых есть водоемы. Летом держится, главным образом, в долинах рек,
озер и по болотам, много времени проводит в воде, спасаясь от кро-
вососущих насекомых. В теплое время года питается преимущественно
травой, водными растениями, ветками кустов. Зимой поедает, главным
образом, побеги и кору различных деревьев, нередко причиняя большой
вред молодым лесным насаждениям. Гон проходит в сентябре—ок-
тябре. В большинстве случаев в период гона самца можно видеть с од-
ной самкой, но иногда самец собирает несколько лосих. Беременность
лосих длится 230—250 дней. Обычно взрослые самки приносят 2, а мо-
лодые 1 теленка.
Сбрасывание рогов самцами происходит в ноябре—декабре, а пол-
ное окостенение и очищение новых рогов — в августе.
Лоси нередко предпринимают (главным образом, осенью и весной)
далекие миграции.
Во многих областях, где охота на лосей разрешается (обычно по
лицензиям), добыча их дает значительное количество мяса и шкур.
Описано большое количество подвидов лосей фауны СССР, но, по-видимому,
достаточно ясно выделяются лишь 3:
а) Европейский лось, A. a. alces Linnaeus, 1758. Размеры средние: высота в холке
до 220 см. Окраска сравнительно однотонная, коричнево-бурая. Европа, Западная
Сибирь.
Ь) Восточносибирский лось, A. a. pfizenmayeri Zukovsky, 1910. Размеры круп-
ные: высота в холке до 235 см. Окраска разнотонная: верхняя часть туловища
219
ржаво-бурая или серо-бурая, а нижняя область его черная. Восточная Сибирь, север
Дальнего Востока.
с) Уссурийский лось, A. a. bedfordi Lydekker, 1902. Размеры мелкие: высота
в холке до 200 см. Окраска всего тела темная, насыщенная. Рога слабо развиты,
обычно без лопатообразного расширения. Дальний Восток, к югу от Амура.
РОД СЕВЕРНЫЕ ОЛЕНИ. GENUS RANGIFER Н. Smith, 1827
Полосы тундры и тайги Европы, Азии и Северной Америки. Опи-
сано много видов северных оленей (особенно из Америки), но, по-види-
мому, правильнее рассматривать их как географические расы одного
вида.
8. Северный олень. Rangifer tarandus Linnaeus, 1758
Табл. XX, 10. Карта 62
Распространен в северных областях Европейской части СССР, на
севере Западной Сибири, в тундре и тайге Восточной Сибири, на Сая-
нах и Восточном Алтае и в северной части Дальнего Востока. Южная
граница ареала северных оленей сильно меняется по годам из-за ми-
грации этих животных. В Европейской части Советского Союза ориен-
тировочно она может быть намечена так: от северного берега Онеж-
ского озера, через район г. Каргополя Вологодской области, по сред-
нему течению р. Онеги примерно вдоль р. Вычегды, через леса северной
части Пермской области к истокам р. Печоры. В тундровой зоне Во-
сточной Европы северные олени бывают только заходом. В Западной
Сибири эти олени водятся к югу до 58—59° с. ш., на Алтае — к во-
стоку от Телецкого озера и верхнего течения р. Оки; обитают в Куз-
нецком Алатау и в Горной Шории. Спорадично встречаются в Саянах.
Населяют как тундру, так и тайгу Восточной Сибири и Дальнего Во-
стока, спускаясь к югу до Северного Забайкалья и северных окраин
Амурского края.
Прежде (XIX в.) северные олени были распространены почти до
Новгорода, верховьев Волги, Казани, Южного Урала, Новосибирска.
Ведут стадный образ жизни, совершая большие миграции. Олени
северных районов весной продвигаются на север, в зону тундр, а
осенью откочевывают на юг — в лесотундру и тайгу. Зимой питаются
в основном лишайниками (главным образом, ягелем), а летом—тра-
вой, побегами кустарников, грибами. Гон происходит в сентябре—ок-
тябре. Сильные самцы кроют нескольких оленух (до 30). Беременность
самок до 8 месяцев. Телята (1, иногда 2, редко 3—4) родятся обычно
в мае, а на севере — в июне. Рога у самцов начинают спадать в де-
кабре, а в массе — в январе, а у самок весной. К началу гона рога бы-
ков окостеневают и очищаются от кожного покрова.
Дикие олени имеют большое промысловое значение. Во многих
районах Севера и в горах Южной Сибири северных оленей разводят
как транспортных животных, а также ради мяса, шкур и молока.
Основные подвиды северного оленя, встречающиеся в СССР:
а) Европейский северный олень, R. t. tarandus Linnaeus, >1758. Размеры средние:
наибольшая длина черепа 34—40 см. Зимний мех относительно темный, с ясно раз-
граниченными темными и светлыми участками. Северная Европа.
Ь) Сибирский тундровый северный олень, R. t. sibiricus Murray, 1866. Размеры бо-
лее крупные: наибольшая длина черепа 35—42 см. Окраска густого и высокого
зимнего меха очень светлая, но обычно с темноватой полосой на боках. Тундра,
лесотундровая зона Сибири и острова, лежащие к северу от нее
Очень близки к этой расе олени с островов Новой Земли (R. t. paersoni Lydekker,
1903), представляющие, вероятно, ее отдельную natio.
229
с) Сибирский лесной олень, R. t. valentinae Flerov, 1933. Размеры примерно такие
же, как и у предыдущей формы. Окраска зимнего меха более темная, рыжевато-
бурого или песчано-бурого цвета, без резко выраженного разграничения на темные
и светлые поля. Мех более короткий, чем у тундровой формы. Лесная зона Восточной
Сибири, горы Саян и Алтая.
d) Дальневосточный северный олень, Т. t. phylarchus Holl., 1912. Самая крупная
форма: наибольшая длина черепа 39—44 см. Окраска темная, светлые поля на боках
невелики. Ростральная часть черепа более выпуклая в профиль, чем у других форм.
Камчатка, Охотское побережье, северные части Амурского края, Сахалин.
СЕМЕЙСТВО ПОЛОРОГИЕ. FAMILIA BOVIDAE
Распространены в Африке, Европе, Азии (включая Малайский ар-
хипелаг) и в Северной Америке (с Гренландией).
Распадается на ряд подсемейств. В Советском Союзе обитают
представители подсемейств газелевых, козлиных и бычьих.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ ПОЛОРОГИХ
КОПЫТНЫХ ФАУНЫ СССР
1(4). Размеры более мелкие: длина тела (от морды до корня хво-
ста) менее 200 см, основная длина черепа не больше 35 см. Хвост ко-
роткий, его конец не достигает голеностопного скакательного сустава.
На его конце кисти длинных волос нет. Верхняя губа, кроме узкой
Рис. 75. Черепа (сбок>):
А — джейран (Gazella subgutturosa); Б — серна (Rupicarpa rupicarpa).
вертикальной голой полоски посередине, покрыта волосами. Промежу-
ток между ноздрями меньше расстояния их от края верхней губы . 2
2(3). Ягодицы, а иногда и круп покрыты белым мехом, участки
которого резко отграничены. Отверстия межпальцевых кожных желез
на передней стороне ног имеют вид вертикальных узких щелей. Наруж-
ный край коронок резцов первой пары примерно в два раза шире их
•основания, поэтому коронки их имеют вид сильно расширяющихся
к наружному краю асимметричных лопаточек. Пястные кости короче
лучевых.....................Газелевые. Gazellinae. (Стр. 222.)
3(2). Ягодицы и круп не выделяются заметно своей окраской. Во-
лосы, покрывающие их, не белые. В отдельных случаях, когда эти уча-
стки покрыты беловатым мехом, границы светлых участков размытые,
221
неясные. Отверстия межпальцевых кожных желез на передней стороне
ног (у тех видов, у которых они есть) имеют округлую форму. Наруж-
ный край коронок резцов первой пары менее чем в два раза превы-
шает ширину их основания, поэтому эти резцы имеют форму долота
или узких лопаток. Пястные кости длиннее лучевых...................
...................................Козлиные. Caprinae. (Стр. 223.)
4(1). Размеры крупные (с корову и более); длина тела (от морды
до корня хвоста) более 200 см, основная длина черепа взрослых более
35 см. Хвост длинный, спускающийся ниже скакательного сустава: на
конце его имеется кисть длинных волос. Конец морды (передняя часть
верхней губы и пространство между ноздрями) голый. Промежуток
между ноздрями больше расстояния между ними и краем верхней
губы..................................Бычьи. Bovinae. (Стр. 234.)
Подсемейство ГАЗЕЛЕВЫХ. Subfamilia GAZELLINAE
Подсемейство распространено по всей Африке, Передней, Средней
Центральной и Южной Азии.
Из ряда родов этого подсемейства в СССР водится 2: Gazella и
Ргосарга.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ГАЗЕЛЕВЫХ ФАУНЫ СССР
1(2). Основная часть хвоста желтая, конечная — черная. Белое
«зеркало» на ляжках ограничено с боков темной полоской. Рога чер-
ные. Передний край носовых костей с ясно выраженной выемкой. Ос-
нован длина черепа менее 21 см . . . . Газели. Gazella. (Стр. 222.)
2(1)..Хвост белый с буроватым концом. Белое «зеркало» на крупе
не ограничено темными полосками. Рога серые. Передний край носо-
вых костей без выемки. Основная длина черепа более 21 см...........
...................................Дзерены. Ргосарга. (Стр. 223.)
РОД ГАЗЕЛИ. GENUS GAZELLA Blainville, 1816
Ареал рода охватывает Северную и Восточную Африку, Перед-
нюю, Среднюю и Центральную Азию, а также большую часть Индии.
В СССР только один вид.
9. Джейран. Gazella subgutturosa Giildenstaedt, 1780
Табл. XXI, 1—2. Карта 63
Места обитания — равнины Восточного Закавказья, пустыни Ира-
ка, Ирана, Афганистана, Средней Азии и Южного Казахстана, пустын-
ные пространства Джунгарии, Кашгарии, Цайдама, Северного Тибета
и пустыни Гоби, на север до Южной Монголии, а на восток до Ала-
шаня и Ордоса. В Восточном Закавказье встречается спорадично.
В Казахстане и Средней Азии северная граница ареала, начинаясь
южнее устья р. Эмбы, тянется к северному берегу Аральского моря,
к среднему течению р. Сарысу и оз. Балхаш. В горах Средней Азии
отсутствует, кроме участка пустыни у западного берега оз. Иссык-Куль.
Типичный представитель фауны пустынь и полупустынь. Наиболее-
характерное местообитание джейрана — полынно-солянковые пустыни
222
и полынные полупустыни как равнинного, так и холмистого рельефа.
Совершают сезонные миграции. Весной и осенью взрослые самки с мо-
лодыми пасутся табунами по 2—10 животных. Зимой же джейраны
в тех местах, где их много, нередко собираются большими стадами.
Ведут преимущественно дневной образ жизни. Питаются различными
пустынными травянистыми и кустарниковыми растениями.
Гон происходит в ноябре и декабре, а деторождение обычно в ап-
реле— мае. В помете, как правило, 2 детеныша; изредка их бывает
1 или 3.
Джейран дает ценное мясо и шкуру, идущую на выделку замши.
Так как в ряде областей СССР численность джейранов в результате
перепромысла сократилась, добыча их в большинстве мест запрещена.
В СССР встречается типичная форма: G. s. subgutturosa Gulden-
staedt, 1780.
РОД ДЗЕРЕНЫ. GENUS PROCAPRA Hodgson, 1846
Распространение рода ограничивается Центральной Азией.
Единственный вид, встречающийся в Советском Союзе.
10. Дзерен. Ргосарга gutturosa Gmelin, 1760
Табл. XXI, 3. Карта 63
Распространен в Центральной Азии, от Ладака и Сиккима до Ал-
тая и Южного Забайкалья. В СССР встречается в Чуйской долине и
Чуйских Альпах на Алтае, южной части Тувы (котловина оз. Упса-
Нур) и в Даурской степи Южного Забайкалья. Предпочитает дер-
жаться в ковыльных, ковыльно-полынных и разнотравных сухих рав-
нинных и холмистых степях. Дзерены совершают далекие миграции.
Летом держатся табунами (взрослые самцы отдельно от самок с моло-
дыми), а зимой скопляются в большие стада. Питаются травянистыми
растениями, главным образом злаками. Спаривание происходит с конца
ноября до начала января, а рождение молодых (1—2) — в июне—июле.
Ввиду малочисленности и ограниченности распространения про-
мыслового значения дзерен в СССР не имеет.
Подсемейство КОЗЛИНЫХ. Subfamilia CAPRINAE
Следует отметить, что это подсемейство, вероятно, объединяет раз-
личные по своему происхождению группы копытных и в дальнейшем
будет разделено на ряд отдельных подсемейств (сайгачьих, серновых и
козлиных).
Представители подсемейства козлиных распространены в Европе,
Северной Африке, Азии и в Северной Америке.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ КОЗЛИНЫХ ФАУНЫ СССР
1(2). Морда сильно вздута, ее конец вытянут в нависающий над
ртом небольшой мягкий хобот. Ноздри округлые. Рога самцов (самки
безроги) воскового цвета. Носовое отверстие сильно вытянутое: длина
его (от переднего края межчелюстных костей до концов носовых ко-
стей) примерно в 3 раза превышает их ширину.....................
......................................Сайга. Saiga. (Стр. 225.)
2(1). Морда не вздута и не образует хобота. Ноздри щелевид-
ные. Рога бурого или серого цвета; самки обычно тоже имеют рога
223
Рис. 76. Черепа (сверху):
Л — серна (Rupicapra rupicapra); Б —горал
(Nemorhaedus goral).
Носовое отверстие черепа ме-
нее вытянуто; его длина не
более чем в 2 раза превышает
ширину .................. 3
3(6). Рога не несут про-
дольных гребней (килей) и
потому имеют округлое сече-
ние. У самок вымя имеет 4 хо-
рошо развитых соска. Верх-
ние отростки межчелюстных
костей не достигают носовых
костей. Резцы с ясно выра-
женной шейкой на границе
корня и коронки.
4(5). Рога прямые, ост-
рые, они направлены верши-
нами назад. Хвост доходит или
почти доходит до скакатель-
ного сустава, его длина значи-
тельно больше длины уха.
Верхнечелюстные кости не со-
прикасаются с носовыми . .
................... Горалы.
Nemorhaedus. (Стр. 225.)
5(4). Рога на концах за-
гнуты крючком. Они направ-
лены у живого зверя вверх.
Хвост более короткий, по длине равный уху или короче его. Верхне-
челюстные кости соприкасаются с носовыми.........................
....................................Серны. Rupicapra. (Стр. 226.)
6(3). Рога несут 1—2—3 продольных острых или тупых гребня
и потому в сечении имеют треугольную, эллиптическую или грушевид-
ную форму. У самок имеется только 2 хорошо развитых соска. Резцы
долотовидные, без шейки на границе коронки и корня ...... 7
7(8). Самцы, а часто и самки имеют на нижней стороне головы
пучок удлиненных волос (бороду). Хвост (с волосами) обычно длин-
нее уха. Рога серовато-черные, темные. Предглазничных желез нет.
Рис. 77. Затылочные отделы черепов сверху:
А — козы (Сарга); Б — овцы (Ovis).
224
На слезных костях вдавлений (слезных ямок) и продольных гре-
бешков нет............ ................Козлы. Сарга. (Стр. 226.)
8(7). Пучка удлиненных волос на нижней стороне головы (бо-
роды) нет. Хвост (с волосами) короче уха. Предглазничные железы
есть. На слезных костях имеются вдавления (слезные ямки) и про-
дольные гребешки..........................Бараны. Ovis. (Стр. 230.)
РОД САЙГИ. GENUS SAIGA Gray, 1843
Сайги водятся в степях и полупустынях Прикаспийской низмен-
ности, Казахстана, Джунгарии и Западной Монголии.
В роде только один вид.
11. Сайга. Saiga tatarica Linnaeus, 1758
Табл. XXI, 4—5. Карта 63
Еще несколько десятилетий назад сайга причислялась к числу ис-
чезающих животных. В результате охраны запасы и ареал сайги стали
быстро восстанавливаться. В настоящее время сайги распространены
в калмыцких, нижневолжских и казахстанских степях, откуда иногда
забегают и в сопредельные степные районы. Северная граница ареала,
начинаясь у устья Терека, тянется через Сальские степи к району
г. Волгограда и г. Уральска, огибает южные отроги Уральского хребта
и пересекает просторы Казахстана через районы гг. Атбасара и Кара-
ганды к хребту Чингиз-тау, к котловине оз. Зайсан. На юг сайгаки
распространены примерно до южного обрыва плато Усть-Урта, ни-
зовьев Аму-Дарьи, среднего течения Сыр-Дарьи и северных предгорьев
Тянь-Шаня.
Обитатель равнинных сухих степей и полупустынь. Питается тра-
вянистыми растениями, особенно различными злаками, солянками, по-
лынью и различными эфемерными видами. Дневные животные. Летом
пасутся обычно небольшими табунками, реже стадами, зимой иногда
образуют большие скопления. Нередко наблюдаются массовые мигра-
ции сайги, когда она заходит даже в районы, где обычно не бы-
вает. Полигамные животные. Гон бывает в ноябре и декабре. Беремен-
ность самок длится в среднем 145 дней. Молодые (обычно 2, реже 1
и совсем редко 3) родятся в мае.
В местах большого скопления сайги служат предметом промы-
сла, дающего ценное мясо и кожевенное сырье.
В СССР встречается номинальная форма.
РОД ГОРАЛЫ. GENUS NEMORHAEDUS1 Н. Smith, 1827
Представители рода обитают в Гималаях, в Юго-Восточном и
Северо-Восточном Китае, Корее и Амуро-Уссурийском крае.
Единственный вид рода.
12. Горал. Nemorhaedus goral Hardwicke, 1825
Табл. XXI, 6. Карта 63
В СССР ныне встречается только в горах Сихотэ-Алиня от их юж-
ной оконечности на север до р. Бикин (47° с. ш.). Численность горалов
в этом районе очень мала. Настоящее горное животное. Держится
1 По Эллерману и Моррисон-Скотту (1951) —Nemorhedus.
1^ Н. А. Бобринский и др.
225
небольшими группами среди скал и крутых обрывов с каменистыми
россыпями, поросших широколиственными деревьями и кедром. В сен-
тябре самки и самцы соединяются на два месяца в пары. Беременность
самок длится 250—260 дней. Молодые родятся в конце мая и в июне.
Добыча запрещена.
РОД СЕРНЫ. GENUS RUPICAPRA Blainville, 1816
Серны встречаются в горах Пиренеев, Аппенин, Альп, Карпат, Бал-
кап, Малой Азии и Кавказа.
Только один вид.
13. Серна. Rupicapra rupicapra Linnaeus, 1758
Табл. XXI, 7. Карта 63
Общее распространение указано выше. В СССР встречается только
на Кавказе, где обитает в районе Главного Кавказского хребта (от
Кавказского заповедника до Восточного Дагестана) и в некоторых
горных районах Закавказья.
Типичное горное животное, житель скал, утесов, каменных россы-
пей, альпийских лугов. На зиму серны спускаются обычно в более низ-
кие зоны. Пасутся преимущественно утром и вечером. Летом питаются
главным образом горной травяной растительностью, а зимой кормятся
также ветками кустарников, деревьев и лишайниками. Пасутся табун-
ками, иногда собираясь в большие стада. Гон — в конце октября и
в ноябре. Беременность самок длится 160—170 дней. Молодые (1—2)
появляются во второй половине апреля и в мае.
Добыча серн запрещена.
Кавказская серна выделяется в особый подвид: R. г. caucasica Lydekker, 1910.
РОД КОЗЛЫ. GENUS CAPRA Linnaeus, 1758
Дикие козлы населяют горы Южной Европы, Северо-Восточной
Африки, Передней, Средней и Центральной Азии (включая Северную
Индию). Домашние козы различных пород разводятся почти во всех
странах мира. Род насчитывает, по-видимому, 8 диких видов, из кото-
рых 5 обитает в СССР.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ДИКИХ ВИДОВ ГОРНЫХ КОЗЛОВ
ФАУНЫ СССР
1(2). Рога как самцов, так и самок уплощены с боков (образуя
передние и задние острые ребра), закручены в крутую спираль (што-
пором) и направлены вверх. У взрослых самцов на нижней части шеи
и груди имеется полоса удлиненных волос — «подвес» .......
....................Винторогий козел. Capra falconeri. (Стр. 228.)
2(1). Рога иной формы. Заднее ребро отсутствует или сглаженно.
«Подвес» из удлиненных волос снизу шеи отсутствует...............3
3(4). Передняя сторона рогов имеет вид острого ребра с выступа-
ющими на нем несколькими редко расположенными буграми. Слабо
выраженное заднее ребро рогов закругленное, тупое. Лобная часть че-
репа выпуклая, поэтому его поверхность в средней части расположена
выше верхних краев глазниц.......................................’
. . ............Безоаровый козел. Capra aegagrus. (Стр. 229.)
226
Рис. 78. Головы козлов:
А — винторогий козел (Capra falconeri); Б — безоаровый козел (С. aegagrus); В —
центральноазиатский козел (С. sibirica); Г— дагестанский тур (С. cylindricornis);
Д — кавказский тур (С. caucasica).
15*
227
Рис. 79. Рога кавказских туров:
4(3). Передняя поверхность ро-
гов не несет острого продольного
ребра, она плоская или выпуклая,
иногда с поперечными валиками,
поэтому поперечное сечение основ-
ной части рога треугольное или
округлое. Лобная часть черепа в
своей средней части лежит ниже
уровня верхних краев глазниц.
5(6). Передняя сторона рогов
широкая, уплощенная, несущая
многочисленные, часто расположен-
ные поперечные валики. Попереч-
ное сечение рога почти треугольное.
Рога расходятся в основной части
под углом меньше 45°. Вдоль хреб-
та тянется ясно выступающая по-
лоса . . . Центральноазиатский
козел. Capra sibirica. (Стр. 229.)
6(5). Передняя сторона рогов
выпуклая, без поперечных валиков
или лишь с неотчетливыми сгла-
женными складками. Поперечное
сечение рогов у их основания более
А — С. caucasica; Б —гибридная форма (С. cau-
casica XCz. cylindricornis); В — дагестанский тур
(С. cylindricornis).
или менее округлое. Рога расхо-
дятся в основной части более чем на 45°. Ясной темной полосы, идущей
вдоль хребта, обычно нет.....................................................7
7(8). Рога изогнуты дугообразно, их вершины направлены вниз
и внутрь. Борода длинная, отогнутая вперед, она заходит или доходит
до конца морды . . . Кавказский тур. Capra caucasica. (Стр. 229.)
8(7). Рога изогнуты растянутой спиралью. Концы их направлены
назад и вверх. Борода широкая, но короткая; отогнутая вперед, она не
доходит до конца морды.....................................................
- . . . . . Дагестанский тур. Capra cylindricornis. (Стр. 230.)
14. Винторогий козел. Capra falconeri Wagner, 1839
Табл. XXI, 8. Карта 64
Обитает в горах Таджикистана, Афганистана, Белуджистана, Се-
верной и Северо-Западной Индии. На территории СССР — в Средней
Азии водится в горном массиве Кугитанг-Тау, в начале р. Аму-Дарьи,
на хребте Баба-таг, между средними течениями Сурхан-Дарьи и Ка-
фирнаган, и в области гор, прилежащих к р. Пянджу, вниз от Дарваза
до линии Куляб-Чубек. Житель высокогорного пояса (1500—3500 м
над у. м.). Держится на альпийских лугах и горно-степных участках
с обилием скал и россыпей. Большую часть года самцы и самки с мо-
лодыми держатся отдельно группами по 3—5 голов; зимой иногда со-
бираются в небольшие стада. Основная пища — травянистые растения,
в меньшей степени (главным образом зимой) — побеги и кора кустов
и деревьев. Гон происходит в ноябре — декабре. Беременность коз
около 6 месяцев. В конце апреля — в мае самка приносит 1—2 козлят.
С целью сохранения этих редких животных добыча их запрещена.
В СССР особая форма: С. f. heptneri Zalkin, 1945.
228
15. Безоаровый, или бородатый, козел. Capra aegagrus Erxleben, 1777
Табл. XXI, 9. Карта 64
Обитает в горах островов Эгейского моря, Малой Азии, Ирана,
ряда районов Кавказа, Южной Туркмении, Белуджистана и Синда.
На Главном Кавказском хребте они водятся примерно от вер-
ховьев р. Аргуни до верховьев р. Самура. В Закавказье встречаются
спорадично в горах от окрестностей г. Батуми до Карабахского на-
горья. В Южной Туркмении встречаются в горах Копет-Дага и Боль-
ших Балхаиов. Держатся на скалистых склонах преимущественно
среднего пояса гор. Большую часть года бродят небольшими группами,
самцы отдельно от самок с козлятами. Зимой иногда соединяются
в стада. Питаются как травянистыми растениями, так и побегами кустов
и деревьев. Гон — в ноябре — декабре. Рождение козлят — в мае — июне.
В Советском Союзе водятся 2 формы безоаровых козлов:
а) Кавказский безоаровый козел, С. a. aegagrus Erxleben, 1777. Рога несколько
массивнее. Кавказ.
Ь) Туркменский безоаровый козел, С. a. turcmenica Zalkin, 1950. Рога более тон-
кие. Горы Южной Туркмении. Самостоятельность этих форм требует проверки.
16. Центральноазиатский козел. Capra sibirica Pallas, 1776
Табл. XXI, 10. Карта 64
Горы Афганистана, Северо-Западной Индии, Средней Азии, Юж-
ной Сибири (на восток до Байкала), Монголии, Северо-Западного Ки-
тая и Тибета.
В Советском Союзе широко распространен в горах Тянь-Шаня и
Памиро-Алая, обитает в горах Тарбагатая, Саур, Калбинского Алтая,
встречается на некоторых хребтах Алтая и Саян.
Обитает на склонах гор, начиная с предгорьев (около 500 м над
у. м.) до снежных вершин (местами до 5000 м над у. м.). Держится
на крутых, безлесных скалах и каменных осыпях, кормясь на травяни-
стых лужайках и альпийских и субальпийских лугах. Зимой спускается
в более низкие зоны гор. Летом питается преимущественно травой,
главным образом злаками, а зимой — ветками кустарников и деревьев.
Гон — в ноябре—декабре. Беременность длится 170—180 дней. Козлята
(1—2) родятся в мае—июне. Служит предметом спортивной и промыс-
ловой охоты.
Описано много географических форм центральноазиатских козлов, в том числе
и по экземплярам из Советского Союза. Но, вероятно, в пределах нашей страны
можно различить только 2 подвида этого козла:
а) Алтайский козел, С. s. sibirica Pallas, 1776. Размеры более мелкие: наибольшая
длина черепа самцов от 26 до 29 см, самок от 24 до 26 см. Рога самцов относи-
тельно короткие (длина по внешней стороне до 100 см). Окраска более светлая..
Алтай, Саяны, горы Монголии.
Ь) Среднеазиатский козел, С. s. alaiana Noack, 1902. Размеры более крупные:
наибольшая длина черепа самцов 28—31 см, самок 25—26 см. Рога очень мощные
длиной обычно более 100 см. Окраска более темная. Горы Тянь-Шаня и Памиро-Алая.
17. Кавказский тур. Capra caucasica Giildenstaedt et Pallas, 1783
Табл. XXI, 11. Карта 64
Распространен в западной части Главного Кавказского хребта, на
восток примерно до 43°30х в. д. Обитает по крутым скалистым склонам
гор, от 800 до 4200 м над у. м. Зимой обычно держится в более низких
229
зонах, чем летом. Летом пищей служат преимущественно различные
травянистые растения, зимой — высохшие пучки злаков, веточки
кустарников и деревьев, лишайники. В теплое время года самцы
держатся отдельно от самок, но осенью и зимой образуют смешанные
стада. Беременность длится 150—160 дней, с ноября—декабря до мая—
июня.
Там, где добыча туров разрешена, они служат предметом спортив-
ной и промысловой охоты.
Дагестанский тур. Capra cylindricornis Blyth, 1841
Табл. XXI, 12. Карта 64
Встречается в восточной части Главного Кавказского хребта, при-
мерно к востоку от 43° в. д.
Биология дагестанского тура сходна с образом жизни кавказ-
ского тура. В районе, где ареалы этих двух форм налегают друг на
друга, встречаются гибридные особи с признаками промежуточного ха-
рактера.
РОД БАРАНЫ. GENUS OVIS Linnaeus, 1758
Дикоживущие представители рода встречаются на некоторых ост-
ровах Средиземного моря, в Передней, Средней, Центральной и Се-
веро-Восточной Азии, а также в западных областях Северной Америки.
Одомашненные представители рода — домашние овцы разводятся
почти во всех странах мира. Еще недавно большинство зоологов на-
шей страны считали, что в Евразии обитает большое количество от-
дельных видов диких баранов. Но в настоящее время обычно признают,
что в СССР обитает только 2 вида баранов: горный баран — архар и
толсторогий баран.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ДИКИХ ВИДОВ БАРАНОВ ФАУНЫ СССР
1(2). Рога с резко выраженными несглаженными поперечными
складками. Лицевой отдел головы и черепа длинный; он составляет
обычно более 57% наибольшей длины черепа. Носовые кости узкие
и длинные: их ширина меньше 43% длины. Костные стержни рогов
самцов длинные; длина рогов больше их охвата у основания. Слезные
ямки на слезных костях выражены сильно...........................
.............................Архар. Ovis ammon. (Стр. 230.)
2(1). Рога со слабо выраженными сглаженными поперечными мор-
щинами. Лицевой отдел головы и черепа короткий; он составляет
обычно менее 57% наибольшей длины черепа. Носовые кости короткие
и широкие: их ширина обычно больше 43% длины. Костные стержни
рогов самцов короткие; длина рогов меньше их охвата у основания.
Слезные ямки на слезных костях выражены слабо....................
. . ..........Толсторогий баран. Ovis nivicola. (Стр. 233.)
Подрод настоящих баранов. Subgenus Ovis Linnaeus, 1758
18. Архар. Ovis ammon Linnaeus, 1758
Табл. XXII, 1—2. Карта 65
Сложный полиморфный вид, образующий большое число географи-
ческих рас. Вероятно, полиморфность вида находится в связи с боль-
шим разнообразием условий существования в различных районах об-
ширного ареала.
230
Область распространения данного вида охватывает острова Кор-
сика, Сардиния и Крит в Средиземном море, горы Передней, Средней
и Центральной Азии.
В Советском Союзе встречаются в некоторых районах Южного
Закавказья (лежащих между низовьями р. Азат и районами гг. Орду-
бада и Метри), в горах Копет-Дага, Малых и Больших Балханов, на
Южном чинке и в горах Западного Устюрта и Мангышлака, по ска-
лам восточного побережья Каспия, на останцовых возвышенностях
Рис. 80. Рога разных форм архаров и муфлонов:
/ — Ovis ammon gmelini; 2 — 0. a. urmiana; 3 — О. a. orientalis; 4, 5 — О. a. cycloceros;
6 — О. a. karelini: 7 — 0. a. polii; 8 — О. a. ammon.
Кызылкумов, на Памире, в горах Тянь-Шаня, в области мелкосопочника
Казахстана, на Тарбагатае, в Саурах, на некоторых хребтах горной
системы Алтая, на гряде Танну-Ола.
Хотя архары — горные животные, они все же предпочитают дер-
жаться на открытых пространствах со слабо пересеченным рельефом —
на сыртах, горных плато, травянистых склонах гор, на увалах пред-
горьев, среди мелкосопочника. Если на Памире архары живут на вы-
сокогорных сыртах, на высоте 3—5 тыс. м над у. м., то архары, обитаю-
щие на склонах восточного побережья Каспия, нередко спускаются
ниже уровня Мирового океана. Иногда архары живут на горных остан-
пах среди пустыни или степи. Иногда совершают значительные
231
миграции. Летом пасутся пре-
имущественно утром и вечером,
зимой — всю светлую часть су-
ток. Осенью и зимой архары
держатся стадами, весной —
небольшими группами. Летом
питаются травянистыми расте-
ниями, главным образом зла-
ками, зимой — сухой травой,
веточками кустарников, хвоей
арчи. Гон проходит обычно в
более северных районах в ок-
тябре— ноябре, на юге стра-
ны— в ноябре — декабре. Про-
должительность беременности
Рис. 81 ,Д. Голова архара (Ovis ammon polii). около 5 месяцев. Ягнение про-
исходит весной; в помете — 1,
реже 2, совсем редко 3 ягненка.
Архары — ценные охотничьи звери, но в ряде районов, где числен-
ность их сократилась, добыча архаров запрещена.
Из многочисленных описанных различными исследователями подвидов архара
наиболее четко выделяются следующие формы:
а) Алтайский горный баран (аргали), О. a. ammon Linnaeus, 1758. Самая крупная
из водящихся в СССР рас. Высота в холке взрослых самцов до 125 см, наиболь-
шая длина черепа самцов 35—39 см, самок 31—34 см. Рога очень мощные (длина
по наружной стороне до 125 txu). Подвес (удлиненные волосы спичу шеи)
мал. Окраска очень разнообразная. Алтай, Тува.
Ь) Памирский горный баран, О. a. polii Blyth, 1841. Размеры очень крупные: вы-
сота в холке самцов до 120 см, наибольшая длина черепа самцов 33—36 см. самок
28—31 см. Рога крупного размера и очень массивны: длина рогов до i90 см,
концы их сильно ютогнуты наружу. Подвес средней величины. Окраска темная,
серовато-красно-бурая или шоколадная. Памир, восточная часть Алтайского хребта,
сырты Аксая и Арпы на Тянь-Шане.
с) Тяньшаньский горный баран, О. a. karelini Severtzov, 1873. Размеры крупные,
но мельче, чем у предыдущего подвида: высота в холке самцов до 110—115 см,
наибольшая длина черепа самцов 31—34 см. Рога длинные, но менее массивные,
чем у О. a. ammon. Подвес отсутствует. Окраска несколько светлее. Тянь-Шань,
кроме сыртов Аксая и Арпы.
d) Казахстанский горный баран, О. a. coIlium Severtzov, 1873. Форма очень близ-
кая к предыдущей, но отличается несколько более темной окраской и некоторыми
краниологическими отличиями (удлиненным верхним отростком межчелюстных ко-
стей и др.). Мелкосопочник Центрального Казахстана, Сауры, Тарбагатай.
Рис. 81,5. Голова аргали (О. a. ammon).
232
Рис. 82. Географическая изменчивость архаров:
.4 — Ovis ammon gmelini (кавказский); Б — О. a. cycloceros (туркменский);
В — О. ammon (алтайский).
с) Каратауский горный баран, О. a. nigrimontana Severtzov, 1873. Размеры еще
мельче: наибольшая длина черепа самцов 29—32 см, самок 26—28 см. Рога крупные.
Подвес развит слабо. Окраска сравнительно темная, рыжевато- или коричневато-бурая.
Рога довольно большие: длина их до 105 см. Горы Каратау в Казахстане.
f) Кызылкумский горный баран, О. a. severtzovi asonov, 1914. Размеры средние:
наибольшая длина черепа самцов 26—26 см. Рога средних размеров и массивности:
длина их до 75 см. Подвес невелик и бороды не образует. Окраска средней тем-
ноты, серовато-коричневая или серовато-бурая. Останцовые возвышенности среди
пустыни Кызылкумов.
g) Туркменский горный баран, О. a. cycloceros Hutton, 1842. Размеры средние: вы-
сота в холке взрослых самцов до 95 см, наибольшая длина черепа самцов 26—30 см,
самок 23—26 см. Рога средней длины и мощности (длина до 92 см). Подвес
развит очень сильно, переходя снизу шеи в пышную бороду. Окраска довольно
светлая, рыжевато-желтоватая. Копет-Даг, Большие Балханы, Устюрт, Мангышлак.
h) Таджикский горный баран, О. a. bocharensis Nasonov, 1914. Размеры средние,
но мельче, чем у предыдущей формы: наибольшая длина черепа самцов 23—26 см,
самок 22—24 см. Длина рогов до 75 см, охват их невелик. Подвес развит сильно,
образуя на горле бороду. Окраска рыжевато-коричневая. Горы по верхнему течению
Аму-Дарьи и Нижнему Пянджу.
i) Закавказский горный баран, О. a. gmelini Blyth, 4840. Размеры довольно мел-
кие: высота в холке взрослых самцов до 95 см, наибольшая длина черепа самцов
25—26 см, самок 22—24 см. Рога небольшие, длиной обычно до 70 см. Подвес
небольшой, не доходящий до горла. Окраска от красновато-рыжей до рыжевато-
коричневой, у самцов — со светлыми пятнами на боках. Закавказье.
j) Муфлон, О. a. musimon Pallas, 1811. Размеры мелкие: высота в холке взрослых
самцов до 80 см, наибольшая длина черепа самцов 23—25 см, самок 21—22 см.
Рога небольшие, довольно тонкие. Окраска их темно-коричневая, значительно более
темная, чем у других форм. Подвес небольшой. Окраска темно-коричневая, у сам-
цов— со светлыми пятнами на боках. Родиной этой формы являются о-ва Корсика
и Сардиния. В СССР муфлоны акклиматизированы в горах Крыма.
Подрод толсторого в. Subgenus Pachiceros Gromova, 1936
19. Толсторогий баран. Ovis nivicola 1 Eschscholtz, 1829
Табл. XXII, 5. Карта 65
Распространен в горах Северо-Восточной Якутии и северной части
Дальнего Востока. Изолированные участки ареала охватывают горы
Путарона на Таймыре и горы Южной Якутии. Водятся также в горах
1 По Эллерману и Моррисон-Скотту — О. canadensis Shaw, 1804.
233
Аляски. Обитают в труднодоступных, скалистых, безлесных горных
районах, поднимаясь до высоты 3000 м над у. м. (обычно пояса от 1000
до 2000 м). Сезонные кочевки слабо выражены. Держатся обычно не-
большими группами. Ведут дневной образ жизни. Основная пища —
различные травы, в меньшей степени — ветки кустарников, зимой по-
едают также кору деревьев и лишайники. Гон в большинстве районов
ареала происходит в ноябре и декабре, а окот самок — в мае—июне.
Число детенышей в помете— 1, реже 2.
В ряде районов Севера толсторогий баран служит предметом про-
мысловой охоты, давая хорошее мясо. Ставится вопрос о скрещивании
толсторогов с домашними овцами для получения новых пород, приспо-
собленных к жизни в горных районах Арктики.
Толсторогие бараны образуют в СССР ряд географических форм:
а) Камчатский толсторогий баран, О. с. nivicola Eschscholtz, 1829. Окраска тем-
ная, более или менее однотонная. Рога очень массивные. Камчатка.
Ь) Охотский толсторогий баран, О. с. alleni Matschie, 1907. Окраска обычно тем-
ная. На боках выступают светлые пятна. Рога, как у предыдущей формы. Стано-
вой хребет, хребет Джугджур, южная часть Колымских гор.
с) Североякутский толсторогий баран, О. с. lydekkeri Kowarzin, 1913. Окраска
светлая, беловатая или желтоватая, с темным ремнем по хребту. Рога заметно более
короткие и тонкие. Горы Северо-Восточной Якутии и возможно Анадырского края.
d) Норильский толсторогий баран, О. с. borealis Severtzov, 1873. Признаки этой
расы изучены еще слабо. По окраске она близка к номинальной форме. Горы
в верховьях рек Пясины, Хеты и Котуя.
Подсемейство БЫЧЬИХ. Subfamilia BOVINAE
Представители этого подсемейства обитают в Европе, в Африке,
Передней и Южной Азии. Одомашненный крупный рогатый скот раз-
водится почти во всех странах мира.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ПОДСЕМЕЙСТВА БЫЧЬИХ
ФАУНЫ СССР
1(4). Тело покрыто хорошо развитым волосяным покровом, скры-
вающим кожу. Рога имеют в сечении округлую форму и не несут ясно
выраженных поперечных валиков в основной части. Сошник не сра-
стается с нёбными костями и не подразделяет заднехоанальное отвер-
стие на две половины.............................................: 2
2(3). Хвост покрыт короткими, прилегающими к коже волосами,
лишь на конце имеется кисть удлиненных волос. Длина волос на спине
и брюхе примерно одинакова. Верхние отростки межчелюстных костей
обычно достигают или почти достигают носовых костей..................
............................................Быки. Bos. (Стр. 234.)
3(2). Хвост покрыт густой, длинной, лохматой шерстью. Волосы на
брюхе сильно удлинены. Верхние отростки межчелюстных костей да-
леко не доходят до носовых костей . . Яки. Phoephagus. (Стр. 235.)
4(1). На теле разбросаны редкие, короткие, жесткие волосы, не
скрывающие поверхности кожи. Рога имеют в сечении треугольную
форму и несут в основной части крупные поперечные валики. Сошник
срастается с нёбными костями и делит заднехоанальное отверстие на
две половины..........................Буйволы. Bubalus. (Стр. 235.)
РОД БЫКИ. GENUS BOS Linnaeus, 1758
Представители этого рода встречаются в Европе, Южной Азии и
Северной Америке. Из диких видов быков в СССР обитает только один
234
вид — зубр. Кроме того, в нашей стране разводят различные породы
крупного рогатого скота, произошедшего от ныне вымершего дикого
быка —тура (Bos primigenus).
Подрод бизонов. Subgenus Bison Hodgson, 1837
20. Зубр. Bos bonasus Linnaeus, 1758
Табл. XXII, 6. Карта 65
Еще в историческое время ареал зубра охватывал большую часть
Европы. Но позднее в большинстве стран этого материка они были ис-
треблены. В СССР в настоящее время около 100 чистокровных зубров
содержится в заповеднике Бело-
вежская пуща, в Окском притер-
расном заповеднике и некоторых
других заповедниках. Кроме того,
там же имеется довольно большое
число помесей зубров с бизонами
или зубров с крупным рогатым ско-
том. Некоторое количество зубров
имеется в Польше, Швеции, Гер-
мании, Голландии и других стра-
нах.
Зубры — лесные стадные жи-
вотные, и только старые самцы ве-
дут иногда одиночную жизнь. Пи-
таются в основном травой, корень-
ями, корой и побегами лиственных
деревьев и кустарников. Течка
обычно протекает с августа по ок-
тябрь. Беременность зубрицы длит-
ся около 9 месяцев, отел происхо-
дит в мае и июне. Обычно родится 1
ление молоком продолжается до 6, г
зубрицы размножаются не каждый
Рис. 83. Череп зубра (Bos bonasus).
теленок, но иногда двойни. Корм-
1 иногда до 12 месяцев. Некоторые
год.
РОД ЯКИ. GENUS PHOEPHAGUS Gray, 1843
Единственный вид рода.
Як. Phoephagus mutus Przewalski, 1883
Табл. XXII, 7
В диком состоянии быки яки встречаются на нагорьях Тибета и
некоторых горных хребтах Центральной Азии. В Советском Союзе раз-
водятся как сельскохозяйственные животные на Алтае и на Памире.
РОД БУЙВОЛЫ. GENUS BUBALUS Н. Smith, 1827
Буйвол. Bubalus bubalus Linnaeus, 1758
Табл. XXII, 8
Одомашнен. В СССР разводится главным образом в Закавказье.
235
Б. А. Кузнецов
ОТРЯД ГРЫЗУНЫ. ORDO RODENTIA
Грызуны — мелкие или средних размеров сухопутные, иногда зем-
новодные млекопитающие, питающиеся преимущественно или исключи-
тельно растительной пищей. Внешний вид и строение грызунов разных
семейств и родов весьма различны в связи с разнообразием их образа
жизни и условий существования.
Наиболее характерной особенностью, объединяющей всех грызунов-
и отличающей их от представителей других отрядов млекопитающих,
является крайне своеобразное строение их зубной системы. Резцы, чи-
сло которых уменьшено до одной пары в нижних челюстях и до одной
или реже двух пар в верхних челюстях, очень велики, лишены корней
и обладают постоянным ростом. Их свободный конец долотообразно
заострен. Передняя стенка их покрыта толстым слоем очень твердой
эмали, тогда как боковые и задняя стороны этих зубов одеты тонким
эмалевым слоем либо совсем лишены эмалевого покрытия. Благодаря
этому резцы стачиваются неравномерно и всегда остаются острыми.
Клыки отсутствуют, и коренные зубы отделены от резцов широким без-
зубым промежутком — диастемой. Общее число коренных (рш + ш) ко-
леблется от — до — (у грызунов, обитающих в СССР, — от— до
6 2 6 3
Они приспособлены к перетиранию грубой растительной пищи и потому
имеют широкую жевательную поверхность, которая либо плоская, либо'
несет ряды тупых бугорков, либо снабжена невысокими гребнями..
У многих грызунов боковые эмалевые стенки коренных зубов глубо-
кими складками вдаются внутрь их коронок.
Для грызунов характерна также широкая суставная поверхность
головки нижней челюсти, позволяющая ей свободно двигаться не толь-
ко в бока, но также вперед и назад. Большинство грызунов (кроме
сонь) имеет очень большую слепую кишку. Мозг грызунов отличается
малым размером полушарий, которые не прикрывают мозжечка; их по-
верхность либо гладкая, либо несет небольшое количество слабо выра-
женных борозд. Матка самок двойная. Пальцы несут на концах когти;
только у некоторых леммингов зимой когти становятся широкими, не-
сколько напоминая своей формой копытца. Конец морды не вытянут
в хоботок, как у насекомоядных. Верхняя губа обычно глубоко рас-
щеплена, так что резцы верхней челюсти более или менее обнажены..
Грызуны населяют все материки (кроме Антарктики) и большин-
ство островов, на многие из которых они были завезены человеком.
В настоящее время известно более 2500 видов грызунов, что со-
ставляет около 7з всего числа видов млекопитающих, населяющих зем-
ной шар. В Советском Союзе обитает 125 видов грызунов, относящихся
к 2 подотрядам и 14 семействам этого отряда.
236
30
вч
Рис. 84. Череп грызуна (сурка, Marmota): А — сверху; Б—снизу; В — сбоку.
Пос — носовая кость; пч — предчелюстная кость; вч — верхнечелюетная коеть; л — лобная коеть; т — теменная кость; вк — впеоч'ная кость;
ск — екуловая коеть; зк — затылочная коеть; сб — слуховые барабаны; ок — основная клиновидная коеть; нк — нёбная кость; нч — нижняя че-
люсть; д'— хоаны; ро — резцовое (передненёбное) отверстие; зо—затылочное отверстие; во —венечный отросток нижней челюсти; со — сочле-
новып отроеток ее; уо — угловой отроеток ее; ко — крыловидный отроеток основной кости; 0 — диастема.
Зубы: / — верхние резцы, / — нижние резцы; Р/n1 — верхний малый предкоренной зуб; Ртр, ртр — крупные предкоренные зубы; А41 — АГ;
т\ — /??3 — заднекоренные зубы.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДОТРЯДОВ ГРЫЗУНОВ
1(2). В верхних челюстях позади мощно развитых передних рез-
цов расположены мелкие столбикообразные резцы второй пары. В каж-
дой ветви нижней челюсти по 5 коренных зубов....................
............Подотряд двупарнорезцовых. Duplicidentata. (Стр. 238.)
2(1). В верхних челюстях второй пары резцов нет. В каждой ветви
нижней челюсти только по 3—4 коренных зуба......................
Подотряд однопарнорезцовых. Simplicidentata. (Стр. 248.)
Подотряд ДВУПАРНОРЕЗЦОВЫХ ГРЫЗУНОВ.
Subordo DUPLICIDENTATA
них велики и
Рис. 85. Верхние
резцы зайца
(вид снизу).
Группа относительно примитивных грызунов, к которой относятся
пищухи, зайцы и кролики.
Верхние челюсти этих грызунов несут 2 пары резцов: передние из
отличаются долотообразной формой, а задние слабо раз-
виты и имеют вид небольших столбиков, расположен-
ных позади передних резцов. Коронки передних рез-
цов со всех сторон покрыты слоем эмали, но на перед-
ней поверхности он значительно толще. Коренных
зубов — или — ; они не имеют закрытых корней,
5 5
а на их коронках с боков глубокие складки эмали.
Костное нёбо в виде узкого поперечного костного мо-
стика, отделяющего нёбные отверстия от хоан. На
стенке слепой кишки имеется спиральная складка.
Половой член самцов направлен головкой назад.
Некоторые исследователи предлагают выделить
данный подотряд в особый, самостоятельный отряд
млекопитающих под названием Lagomorpha. Но сход-
ство ряда признаков (строения зубной системы, головного мозга, пище-
варительного тракта и др.) двупарнорезцовых и однопарнорезцовых
грызунов настолько велико, что разделение их на два отряда не может
быть, на наш взгляд, признано целесообразным и оправданным.
В фауне Советского Союза подотряд двупарнорезцовых грызунов
представлен двумя семействами: пищуховых и заячьих.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ
ПОДОТРЯДА ДВУПАРНОРЕЗЦОВЫХ ГРЫЗУНОВ ФАУНЫ СССР
1(2). Уши округлые; их длина примерно равна ширине. Хвост на-
столько короток, что не выдается из меха огузка. Задние ноги лишь
немного длиннее передних. В верхних челюстях по 5 коренных зубов.
Надглазничные отростки черепа отсутствуют.....................
............................. Пищуховые. Ochotonidae. (Стр. 239.)
2(1). Уши удлиненные: длина более чем в два раза превышает их
ширину. Хвост заметно выдается из меха огузка. Задние ноги примерно
в два раза длиннее передних. В верхних челюстях по 6 коренных зубов.
Череп несет хорошо развитые надглазничные отростки............
..................... . . . . Заячьи. Leporidae. (Стр. 243.)
238
СЕМЕЙСТВО ПИЩУХОВЫЕ. FAMILIA OCHOTONIDAE
Грызуны среднего размера: длина тела 12—25 см, с вальковатым
телом и уплощенной сверху головой, несущей большие округлые уши.
Хвост сильно редуцирован и не выдается из меха огузка. Задние ноги
лишь немного длиннее передних. Череп уплощен сверху. Надглаз-
ничные отростки отсутствуют. От скуловых дуг назад отходят тонкие
длинные отростки.
Зубная формула: I — Pm + М — • 2 = 26.
1 5
Один род — пищухи.
РОД ПИЩУХИ. GENUS OCHOTONA Link, 1795
Степи Юго-Восточной Европы, Западной Сибири и Северного Ка
захстана, горы Средней и Центральной Азии, Алтай, Саяны, горы Во
сточной Сибири, Дальнего Во-
стока и западной части Северной
Америки.
Степные виды живут на уча-
стках целины, в зарослях кустар-
ников и бурьяна, в оврагах и
балках, иногда на полях. Другие
встречаются в горах, по осыпям
камней, в скалах. Иногда оби-
тают в тайге и тундре среди ка-
менистых россыпей и бурелома.
Живут в неглубоких норках или
в щелях скал и в гнездах между
камнями. В год дают обычно два
помета из 3—12 детенышей. Пи-
таются травянистыми растения-
ми и веточками кустарников. На
зиму делают запасы высушенных
трав: степные виды складывают
их в стожки, а горные прячут
Рис. 86. Черепа:
А — пищухи; Б — зайца.
сено в трещины скал и между
камней. Активны преимущественно в светлое время суток. В неболь-
шом количестве добываются ради шкурок, ценность которых мала
вследствие тонины и слабости кожного покрова.
Легко заболевают чумой, чем способствуют распространению эпи-
зоотий в природных очагах этого заболевания.
Из II—12 видов пищух в фауне СССР — 8 видов.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ПИЩУХ ФАУНЫ СССР
1(6). Нёбное и резцовое отверстия черепа отделены друг от друга
сходящимися краями межчелюстных костей, которые полностью или
частично скрывают сошник (рис. 87, Л)........................2
2(3). Размеры мелкие: длина ступни менее 28 мм\ кондилобазаль-
ная длина черепа менее 40 мм...................................
................Северная пищуха. Ochotona hyperborea. (Стр. 241.)
3(2). Размеры крупнее: длина ступни взрослых более 28 мм\ коп-
дилобазальная длина черепа более 40 мм ... .......... 4
239
4(5). Подошвы лап покрыты темным буроватым мехом. Летом
спина коричнево-бурая, а брюшко желтовато-коричневое, зимой мех
несколько светлее и серее .......................................
......................Алтайская пищуха. Ochotona alpina. (Стр. 241.)
5(4). Подошвы лап покрыты светлым мехом. Окраска летнего
меха светлая, желтовато-серая, зимнего палево-серого цвета . . . .
..............Палласова пищуха.' Ochotona pallasi. (Стр. 242.)
6(1). Резцовое и нёбное отверстия черепа слиты в одно отверстие
или соединены узкой щелью. Сошник виден с нижней стороны черепа
на всем его протяжении.........................................7
7(10). Внутренние поверхности межчелюстных костей несут гори-
зонтальные, направленные друг к другу костные гребни. Но эти гребни
Рис. 87. Черепа различных видов пищух (снизу):
/1 — северная пищуха (Ochotona hyperborea); Б — степная пищуха (О. pusilia);
В — даурская пищуха (О. daurica).
не сходятся, оставляя между собой узкую продольную щель, через ко-
торую виден сошник. Эта щель соединяет резцовое и нёбное отверстия
(рис. 87, Б)..............................................: : 8
8(9). Размеры относительно крупные: длина тела более 20 см;
общая длина черепа более 43 мм. Летом спина ярко-рыжая, зимой
серая................Красная пищуха. Ochotona rutila. (Стр. 242.)
9(8). Размеры мельче: длина тела менее 20 см; общая длина че-
репа менее 43 мм. Окраска спины весь год темная, серовато-бурая . .
..............Степная (малая) пищуха. Ochotona pusilia. (Стр. 242.)
10(7). Резцовое и нёбное отверстия слиты в одно. Внутренние
края межчелюстных костей гладкие и не образуют гребней, нави-
сающих над сошником (при рассматривании черепа -снизу) (рис.
87, В).......................................................11
П(14). Межглазничный промежуток узкий: ширина его менее
4,5 мм. Вибриссы короткие (менее 6 см). Длина уха менее 25 мм . 12
12(13). Длина черепа менее 47 мм. Большинство вибрисс белые.
Ушные раковины с белой каймой. Окраска летнего меха на спине свет-
240
лая, песчано-желтая, на брюхе беловатая. Зимний мех того же цвета,
но светлее. Светлой полосы за ушами нет.................................
...................Даурская пищуха. Ochotona daurica. (Стр. 242.)
13(12). Длина черепа взрослых особей более 47 мм. Большинство
вибрисс темно-бурые, иногда со светлой вершиной. Ушные раковины
без белесой каймы. Летний мех на спине ржаво-буроватый, на брюхе
сероватый. Зимой мех на спине более серого оттенка. За ушами светлые
полосы .... Рыжеватая пищуха. Ochotona rufescens. (Стр. 243.)
14(11). Межглазничный промежуток широкий: ширина его более
4,5 мм. Вибриссы длиной 6—8 см. Длина уха взрослых более 25 мм . .
...................Большеухая пищуха. Ochotona roylei. (Стр. 243.)
1. Северная пищуха. Ochotona hyperborea Pallas, 1811
Табл. XXIII, 1—3. Карта 66
Основная часть ареала от р. Енисея до Камчатки и Сахалина
включительно; изолированно водится также на Северном Урале. Даль-
ше любых других палеарктических видов идет к северу: до устья Лены
и берегов Чукотского моря. Живет в тайге и каменистых тундрах. Се-
лится колониями или поодиночке в россыпях камней, в скалах, между
крупными камнями высохших русел рек, среди бурелома в тайге.
По направлению с севера на юг размеры северных пищух делаются все более
крупными, а окраска — более темной, но менее яркой. Можно различать следующие
подвиды фауны СССР:
а) О. h. hyperborea Pallas, 1811. (Местом описания типа принято считать Чукотку.)
Длина тела взрослых 119—160 мм. Общая длина черепа 35—41 мм. Окраска лет-
него меха спины светлая, серовато-бурая с рыжим налетом. Бока желтоватые. Чу-
котка, Анадырь, бассейн р. Колымы.
Ь) О. h. ferruginea Schrenk. 1858. ( = Och. h. turuchanensis Naum. Тип с Камчатки.)
Крупнее: длина тела взрослых до 176 мм; общая длина черепа 38—43 мм. Окраска
летнего меха спины яркая, ржаво-бурая или буровато-рыжая, бока охристые. Кам-
чатка, Якутия (кроме бассейна р. Колымы), северная часть бассейна р. Енисея.
с) О. h. cinereoflava Schrenk, 1858. (Тип из окрестностей Удского острога.) Раз-
меры, как у предыдущей расы. Окраска светлая. Спина летом желтовато-серая, бока
желтые. Охотское побережье.
d) О. h. mantchurica Thomas, d909. (Тип с Большого Хингана.) Размеры крупные:
длина тела 150—195 мм; общая длина черепа 39—44 мм. Окраска темная. Летний
мех на спине коричневато-бурый, на боках ржаво-буроватый. Приморье, Амурский
край, Забайкалье, Прибайкалье, Саяны.
е) О. h. uralensis Flerov, 1927. (Тип с р. Сыньи на Северном Урале.) Размеры
невелики: общая длина черепа 37—39 мм. Окраска летнего меха на спине относи-
тельно светлая, желтовато-ржавая. Бока охристые. Северный Урал.
Некоторые авторы объединяют О. hyperborea и О. alpina в один вид; мы считаем
такой взляд неверным, поскольку обе эти пищухи в ряде районов живут бок о бок
и, по-видимому, не скрещиваются.
Пищухи, распространенные в горах западных районов Северной Америки (О. colla-
ris Nelson, 1893, Аляска, и О. princeps Richardson, 1828), представляют собой, видимо,
подвиды О. hyperborea.
2. Алтайская пищуха. Ochotona alpina Pallas, 1773
Табл. XXIII, 4, 5. Карта 66
Встречается в горах Алтая, Саян, Тувы, Прибайкалья, Забайкалья
и области верхнего течения Амура. Распространена в Монголии и Се-
верном Китае.
Селится преимущественно среди скал и осыпей камней.
Размеры алтайских пищух постепенно уменьшаются по направлению от запад-
ных районов ареала к восточным. Кроме того, пищухи этого вида разных областей
16 Н. А. Бобринский и др.
241
отличаются интенсивностью, яркостью и оттенком окраски. Описаны следующие под-
виды алтайской пищухи:
а) О. a. alpina Pallas, 1773. (Тип с Тигерецкого хребта, Алтай.) Размеры крупные:
общая длина черепа до 53 мм. Летний мех на спине сравнительно светлый. Запад-
ные районы Алтая.
Ь) О. a. nitida Hollister, 1912. (Тип с р. Чуя, Алтай.) Размеры мельче: общая
длина черепа до 51 мм. Окраска летнего меха более насыщенная и сильнее затем-
ненная по хребту темными концами остевых волос, чем у номинальной расы.
Восточный Алтай, Саяны, Тувинская АССР.
с) О. a. svatoshi Turov, 1924. (Тип с Баргузинского хребта.) Размеры еще меньше:
общая длина черепа до 48 мм. Окраска летнего меха на спине яркая, ржаво-бурая,
слабо затемненная черноватой остью. Западное Забайкалье.
d) О. a. cinereofusca Schrenk, 1858. (Тип с Верхнего Амура.) Размеры, как у пре-
дыдущей формы. Окраска летнего меха более светлая и тусклая, чем у западнозабай-
кальских пищух, с менее выраженными рыжими оттенками (О. a scorodumovi Scalon
мы считаем синонимом О. a. cinereofusca Schr.).
Восточное Забайкалье, Верхний Амур.
3. Палласова пищуха. Ochotona pallasi Gray, 1867
Табл. XXIII, 6. Карта 66
Широко распространена в Монголии. В СССР найдена в Чуйской
степи на Алтае и в мелкосопочнике Восточного Казахстана.
Держится главным образом по пустынным сопкам, на горных ос-
танцах, среди скал. Селится обычно в щелях между каменными глы-
бами или в норах под ними.
Номинальная форма (О. р. pallasi Gray) живет в Северо-Западной Монголии.
В СССР водятся:
а) О. р. sushkini Thomas, 4924. (Тип из Чуйской степи.) Окраска летнего меха
относительно яркая, с ржаво-охристым оттенком на спине. Чуйская степь.
Ь) О. р. ораса Argyropulo, 1939. (Тип с р. Токрау.) Окраска тусклая, серо-бурова-
тая. Северное Прибалхашье.
4. Красная пищуха. Ochotona rutila Severtzov, 1872
Табл. XXIII, 7. Карта 66
Горы Тянь-Шаня, Гиссаро-Алая и Западного Памира.
Живет среди осыпей и скал. В СССР — типичная форма.
5. Степная (малая) пищуха. Ochotona pusilia Pallas, 1769
Табл. XXIII, 8. Карта 66
Степи Заволжья, Южного Урала и Северного Казахстана.
Живет в бурьянах, в зарослях степных кустарников, по оврагам.
По степным склонам поднимается в горы.
Подвиды:
а) О. р. pusilia Pallas, 1769. (Тип с Южного Урала.) Размеры мелкие. Окраска тем-
ная. Заволжье, Южный Урал.
Ь) О. р. angustifrons Argyropulo, 1932. Размеры крупные. Окраска светлая. Восточ-
ный Казахстан.
6. Даурская пищуха. Ochotona daurica Pallas, 1776
Табл. XXIII, 9. Карта 66
В СССР — степи Южного Забайкалья и Чуйской долины на Алтае.
Широко распространена в Монголии.
Поселяется колониями, чаще всего в зарослях ириса и карагана,
по обрывам, в западинах.
242
Подвиды-
а) О. d. daurica Pallas, 1776. (Описан из Забайкалья.) Окраска летнего меха
сравнительно яркая, охристая. Южное Забайкалье.
Ь) О. d. altaina Thomas, 4911. (Тип из Северо-Западной Монголии.) Окраска лет-
него меха более тусклая, с сероватыми оттенками. Чуйская степь.
7. Рыжеватая пищуха. Ochotona rufescens Gray, 1842
Табл. XXIII, 10. Карта 66
Иран и Афганистан. В СССР — в горах Копет-Дага и Больших
Балханов, в Туркмении.
Селится среди камней, осыпей, по склонам ущелий.
Номинальная форма (О. г, rufescens Gray) обитает в Афганистане. В СССР —
особый подвид: О. г. regina Thomas, 1911. (Тип с гор Копет-Дага.) Отличается от но-
минальной формы несколько большими размерами: общая длина черепа 44—53 мм.
8. Большеухая пищуха. Ochotona roylei Ogilby, 1839
(-0. macrotis Gunther, 1875)
Табл. XXIII, 11. Карта 66
В горах Центрального и Восточного Тянь-Шаня и Восточного Па-
мира. За пределами СССР — в Западном Китае, Ладакхе и Гималаях.
Живет в горах, преимущественно среди россыпей камней и в ска-
лах. Образ жизни изучен слабо.
Описанная Томасом из окрестностей Пржевальска О. sacana — синоним данного
вида пищух.
СЕМЕЙСТВО ЗАЯЧЬИ. FAMILIA LEPORIDAE
Сравнительно крупные грызуны с сжатым с боков телом, длин-
ными, сильными, четырехпалыми задними и более слаборазвитыми пя-
типалыми передними ногами, коротким хвостом и сжатой с боков го-
ловой, несущей длинные уши. Череп сдавлен с боков. Его надглазнич-
ные отростки хорошо выражены.
Зубная формула: I у Ргп + М~2 = 28.
Распространены в Европе, Азии, Африке, Америке, завезены в Ав-
стралию и Новую Зеландию. В СССР — 2 рода: зайцы и кролики.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ЗАЯЧЬИХ ФАУНЫ СССР
1(2). Длина задней ступни (без когтей) меньше 11 см. Длина уха,
измеренная от затылка, меньше длины черепа. Вырезка костного нёба
узкая; ширина ее значительно меньше длины костного нёба. Межте-
менная кость у взрослых особей остается обособленной...............
................................Кролики. Oryctolagus. (Стр. 244.)
2(1). Длина задней ступни (без когтей) взрослых особей обычно
11 см. Длина уха, измеренная от затылка, равна или превышает длину
черепа. Вырезка костного нёба широкая; ширина ее равна или больше
длины костного нёба. Межтеменная кость у взрослых животных сли-
вается с соседними костями черепа .... Зайцы. Lepus. (Стр. 244.)
16*
243
РОД КРОЛИКИ. GENUS ORYCTOLAGUS Lilljieborg, 1874
Коренной ареал — Южная Европа и Африка. Известно 2 вида:
один — средиземноморский, другой — южноафриканский. В СССР —
один вид.
9. Дикий кролик. Oryctolagus cuniculus Linnaeus, 1758
Табл. XXIV, 1. Карта 67
Западная Европа и Северная Африка. Завезен в Австралию, Но-
вую Зеландию, Америку. В нашу страну был завезен еще в конце про-
шлого столетия и выпущен в окрестностях Одессы и Херсона. В насто-
ящее время расселился по Черноморскому побережью Украины и в при-
легающих районах.
Живет колониями
в норах по обрывам
морского берега, по ов-
рагам, в лесных по-
садках. Ежегодно сам-
ка приносит до 4—5
пометов из 4—8 кроль-
чат. Летом питается
травой, зимой — корой
деревьев, . веточками
кустарников, сорняка-
ми, сеном. Служит
предметом охоты. Вре-
дит садам и лесным
посадкам.
Л — кролика
Рис. 88. Черепа сверху:
(Oryctolagus cunculus); Б — зайца-толая (Lepus
tolai).
РОД ЗАЙЦЫ. GENUS
LEPUS Linnaeus, 1758
Обитают в Европе,
Азии, Северной Аме-
рике и Африке.
летом травянистыми ра-
Места обитания разнообразны. Питаются
чтениями, зимой — побегами и корой деревьев и кустарников, сорня-
ками, сеном и т. п. За теплое время года приносят 2—4 приплода, по
2—8 детенышей в каждом. Детеныши родятся зрячими, покрыты гу-
стым мехом и способны сразу же бегать.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ЗАЙЦЕВ
ФАУНЫ СССР
1(2). Ухо сравнительно короткое: длина его (от затылка) равна
или даже меньше длины черепа. Тыльная сторона ушной раковины
сзади рыжеватая. Ширина вырезки костного нёба приблизительно
равна длине мостика костного нёба. Ширина носовых костей в их сред-
ней части более чем на !/з превышает ширину черепа позади заглазнич-
ных отростков . Дальневосточный заяц. Lepus brachyurus. (Стр. 245.)
2(1). Ухо длиннее: длина его, измеренная от затылка, больше
длины черепа. Тыльная сторона ушной раковины сзади не рыжая. Ши-
244
совсем, не белеет или белеет
рина вырезки костного нёба заметно больше длины мостика костного
нёба. Ширина носовых костей в их средней части не более чем на !/3
превышает ширину черепа позади заглазничных отростков .... 3
3(4). Верхняя сторона хвоста покрыта белым мехом зимой и серо-
ватым летом. На зиму зверек полностью белеет, кроме кончиков ушей,
которые остаются черными. Длина хвоста с концевыми волосами зна-
чительно короче длины задней ступни (без когтей). Передний предко-
ренной зуб нижней челюсти слабо наклонен назад.....................
............................Заяц-беляк. Lepus timidus. (Стр. 246.)
4(3). Верхняя сторона хвоста всегда с ясно выраженным чернььм
или черно-серым пятном. На зиму м(
только частично. Длина хвоста с кон-
цевыми волосами примерно равна
длине задней ступни (без когтей). Пе-
редний предкоренной зуб нижней че-
люсти сильно наклонен вперед . . 5
5(8). Размеры сравнительно мел-
кие: длина тела менее 56 см; вес
взрослых обычно не более 3,5 кг. На
спине концы пуховых волос сероватые
или желтоватые. Мех гладкий, не кур-
чавый. Общая длина черепа менее
95 мм.............................6
6(7). Ухо относительно короткое:
длина его (от затылка) превышает
длину черепа не более чем на !/3.
Концы ушей черные. Задняя пластин-
ка нижней челюсти велика (рис.
89, 4)..............................
Заяц-толай. Lepus tolai L (Стр. 247.)
7(6). Ухо относительно длинное:
длина его превышает длину черепа
более чем на !/3. Концы ушей лишь
немного темнее средней части. Задняя
пластинка нижней челюсти небольшая
.......................Заяц-песчаник. Lepus tibetanus !. (Стр. 247.)
8(5). Размеры крупные: длина тела взрослых обычно превышает
56 см; вес взрослых обычно от 3,5 до 7 кг. Мех обычно слегка курча-
вый. Общая длина черепа взрослых особей в большинстве случаев бо-
лее 95 мм.................Заяц-русак. Lepus europaeus. (Стр. 247.)
Рис. 89. Нижние челюсти:
А — зайца-толая (Lepus tolai); Б — зай-
ца-песчаника (L. tibetanus).
ОИС. 89. Б}............................
10. Дальневосточный заяц. Lepus brachyurus Temminck, 1861
Табл. XXIV, 2. Карта 67
Южные части советского Дальнего Востока, на север до Благове-
щенска, Малого Хингана, района г. Хабаровска, долины р. Самарги.
Водится в Северном и Северо-Восточном Китае, в Корее и в Японии.
Держится преимущественно в лесах с преобладанием лиственных
пород. Иногда залегает в низких дуплах деревьев и в полых повален-
ных стволах. Образ жизни изучен слабо. Промысловое значение ни*
чтожно.
1 Песчаник и толай так близки между собой, что их, вероятно, правильнее считать
географическими формами одного вида.
245
11. Заяц-беляк. Lepus timidus Linnaeus, 1758
Табл. XXIV, 3—4. Карта 67
Обитает в тундровой, лесной и в большей части лесостепной зонах
Восточной Европы, Азии и Северной Америки; водится в горах Альп
и в Ирландии, Шотландии и Скандинавии.
В СССР распространен на север до побережья Северного Ледови-
того океана, на восток — до берегов Тихого океана (встречаясь на о-ве
Сахалин), на запад — до пределов страны. Южная граница ареала тя-
нется примерно от Беловежа через Киев, Харьков, Воронеж, Камышин,
Оренбург, Актюбинск, Караганду, оз. Балхаш на Джунгарский Алатау.
Далее на Восток она проходит через Северную Монголию к оз.
Ханка, на широте которого упирается в Японское море.
Особенно многочислен в Якутии. В лесостепных районах сравни-
тельно редок. Держится преимущественно в лесных угодьях и по до-
линам рек, в тундре — в зарослях кустарника, в степях Северного Ка-
захстана и Западной Сибири — в камышах по озерам.
Самка ежегодно приносит 2—3 помета из 2—8 (чаще из 3—5)
зайчат. В центральных районах страны первый помет в большинстве лет
бывает в мае, второй — в конце июня и в июле, третий — осенью. Самец
кроет самку вскоре после родов. Беременность длится 49—51 день.
Через 9—10 дней после рождения зайчата начинают поедать траву.
Периодически происходящие массовые падежи беляков обычно вызы-
ваются поражением их глистами. Эти падежи приводят к большим ко-
лебаниям численности и изменениям размеров заготовки их шкурок (от
1 до 5 млн. в год).
Важный объект пушного промысла и спортивной охоты. Известны
случаи участия беляков в распространении туляремии.
Географическая изменчивость беляка выражена довольно сильно. В Европейской
части СССР и Западной Сибири обитают зайцы крупного размера, с довольно светлым
летним мехом, окраска которого светлеет по направлению с запада к востоку. В Восточ-
ной Сибири и на Дальнем Востоке (кроме Чукотки) обитают беляки мелкого размера:
окраска их летнего меха постепенно светлеет к югу. Описано большое число подвидов,
но многие из них в лучшем случае заслуживают выделения в качестве слабо обособ-
ленных natio, а некоторые должны быть сведены в синонимы. Реально различимые
подвиды могут быть объединены в три группы:
I. Европейско-западносибирские формы. Характеризуются крупными размерами:
вес до 5 и даже 6 кг\ общая длина черепа до 107 мм.
a) L. t. timidus Linnaeus, 1758. (Тип из Швеции.) Летний мех относительно тем-
ный, буровато-серый. Ухо спереди черно-бурое. Зимний мех с чисто белой подпушью.
Северные области Европейской части СССР и Западной Сибири (к югу до Вологды,
Кирова, Тобольска, Нарыма), Алтай.
b) L. t. kozhevnicovi Ognev, 1929. (Тип из Московской области.) Окраска лет-
него меха более светлая, ржаво-буроватая. Передний край уха ржаво-бурый. Зимний
мех с желтоватым пухом. Западные и центральные области Европейской части СССР.
с) L. t. sibiricorum Johansen, 1923. (Тип с р. Чулыма.) Летний мех светлый, зем-
листо-серый. Передний край уха сероватый. Зимний мех с белым пухом. Южные
части Западной Сибири и Северного Казахстана (кроме Алтая).
d) L. t. begitschevi Koljuschev, 1936. (Тип с западного берега Пясинского залива.)
Летний мех еще более светлый, сероватый. Передний край уха буроватый. Таймыр,
Гыданский полуостров.
II. Восточносибирские формы. Отличаются несколько меньшими размерами: вес
до 4 кг\ общая длина черепа до 100 мм.
е) L. t. kolymensis Ognev, 1922. (Тип из окрестностей Нижне-Колымска.) Летний
мех очень темный, серовато-бурый с чернотой. Передний край уха черный. Большая
часть Якутии, Забайкалье. (Зайцы Забайкалья отличаются чуть более светлой ок-
раской меха — natio L. t. k. transbaicalicus Ognev, 1929, тип из Сосновки на
Баргузине.)
f) L. t. gichiganus J. Allen, 1903. (Тип из Гижиги.) Окраска летнего меха более
светлая, передний край уха буровато-серый. Дальний Восток, кроме Чукотки. Зайцы
из Приморского края (L. t. g. mordeni Goodwin, 1933) чуть заметно отличаются более
246
светлой окраской с примесью ржавых тонов, а зайцы с Сахалина (L. t. g. orii Kuroda,
1928) — более серым тоном летнего меха.
III. Чукотский подвид заслуживает выделения в особую группу, в которую,
вероятно, входят также некоторые формы беляка из Северо-Западной Америки.
g) L. t. tschuktschorum Nordquist, 1883. (Тип с Чукотского полуострова.) Размеры
крупные: вес до 5,5 кг, общая длина черепа до 106 мм. Окраска летнего меха
тусклая, серо-буровато-палевая. Передний край уха темно-бурый с чернотой. Чукотка.
12. Заяц-толай. Lepus tolai Pallas, 1778
Табл. XXIV, 5, 6 Карта 67
Широко распространен в Монголии, В СССР обитает в степях За-
байкалья и в Чуйской степи на Алтае, Держится преимущественно по
зарослям кустарников, в падях и в кустарнике пойм рек. За год самка
приносит 2—3 помета. Иногда селится в норах сурков. Промысловое
значение невелико.
Подвиды:
a) L, t tolai Pallas, 1778. (Тип из долины р. Селенги.) Размеры относительно
крупные: общая длина черепа 87—96 мм. Окраска меха светлая. Забайкалье.
b) L. t. quercerus Hollister, 1912. (Тип из Чуйской степи.) Размеры немного
меньше: общая длина черепа 85—88 мм. Мех затемнен темными концами остевых
волос. Чуйская степь (на Алтае).
Заяц-песчаник. Lepus tibetanus Waterhouse, 1841
Карта 67
Центральная и Средняя Азия. В СССР обитает в Казахстане и
в среднеазиатских республиках на север до Устюрта, песков Большие
Барсуки, северного побережья оз, Балхаш и котловины оз, Зайсан,
Держится преимущественно в сухих глинистых и песчаных пусты-
нях, но в горах Средней Азии поднимается до альпийских лугов.
Охотно селится в тугаях и камышах по долинам рек и в зарослях чия.
Иногда занимает старые сурковые норы. В течение года самки прино-
сят по нескольку пометов (в южных районах размножаются почти
весь год), по 3—6 детенышей в каждом. Летом питается травянистыми
растениями (главным образом злаковыми, осокой, полынью), а зимой
грызет также побеги кустов. Промысловое значение невелико, но слу-
жит важным предметом спортивной охоты.
Подвиды выделены главным образом по слабым тоновым различиям окраски
меха:
a) L. t. desertorum Ognev et Heptner, 1928. (Тип из Аннау, близ Ашхабада.)
Окраска летом очень светлая, палево-песчаная. Пустыни Туркмении.
b) L. t. turcomanus Heptner, 1934. (Тип из района Джебеля.) Окраска летнего
меха палево-серая с сильным затемнением (темными концами) остевых волос.
Горы Копет-Дага.
с) L. t. buchariensis Ognev, 1922. (Тип из района Термеза.) Характерно наличие
красновато-ржавого оттенка. Таджикистан и Западный Узбекистан.
d) L. t. pamirensis Blanford, 1875. (Тип из окрестностей оз. Сары-Куль.) Отли-
чается очень пышным мехом и тусклой, бледной окраской. Памир.
е) L, t. lehmanni Severtzov, 1873. (Тип с низовьев Сыр-Дарьи.) Несколько круп-
нее предыдущих форм. Окраска немного темнее, более сероватого оттенка. Южный
Казахстан, Киргизия, Восточный Узбекистан.
13. Заяц-русак. Lepus europaeus Pallas, 1778
Табл. XXIV, 7, 9. Карта 67
Вся Европейская часть СССР (на север до Средней Карелии, рай-
она г. Архангельска, центральных частей Коми АССР) и Закавказье.
Постепенно распространяется по степям Зауралья и Казахстана.
247
Акклиматизирован в ряде районов Южной Сибири и Дальнего Востока.
Широко распространен в Западной Европе, завезен в Северную
Америку.
Держится преимущественно на открытых пространствах — в степи,
на полях и т. п. Самки приносят по 3—4 помета в год, из 2—5 (иногда
до 7) зайчат в каждом.
В СССР ежегодно добывается около 2 млн. (в отдельные годы до
4 млн.) русаков. Шкурки их используются как сырье для производства
фетра. Местами русаки зимой наносят серьезный вред садам и лесным
посадкам, повреждая кору и побеги деревьев и кустарников.
Географическая изменчивость выражена сравнительно резко. По направлению
с севера на юг размеры мельчают, мех становится более низким и редким,, степень
побеления зимнего волосяного покрова уменьшается. Ряд описанных форм, встре-
чающихся в СССР, можно объединить в 3 группы:
I. Северные формы. Размеры крупные: вес до 7 кг\ общая длина черепа 95—
ПО мм. На зиму мех заметно белеет.
a) L. е. hybridus Pallas, 1811. (Тип из бывшей Московской губернии.) Летний мех
на спине от буровато-палевой до глинисто-рыжевато-желтой окраски. На зиму белеют
загривок, бока и огузок. Северные, средние и западные области Европейской части
СССР, Татария, Башкирия, Зауралье, Оренбургская область.
b) L. е. tesquorum Ognev, 1923. (Тип из бывшего Бобровского уезда Воронеж-
ской области.) Окраска летнего меха на спине тусклая, буровато-желтовато-серая
без рыжеватых оттенков. На зиму слегка светлеют бока и огузок. Украина (кроме
южных районов), центрально-черноземные области.
II. Южные (степные) формы. Размеры средние: вес до 5 кг; общая длина черепа
90—105 мм. На зиму мех почти не белеет; лишь на огузке появляются седые волосы.
с) L. е. transsylvanicus Matschie, 1901. (Тип из Румынии.) Окраска летнего меха
более яркая, буровато-желтовато-серая с глинистым оттенком. Юг Украины: весьма
близкая форма (natio) обитает на Северном Кавказе (L. е. t. caucasicus Ognev, 1929).
d) L. e. caspicus Ehrenb, 1830. (Тип из окрестностей Астрахани.) Окраска лет-
него меха светлая, желтоватая. Уши короче, чем у других рас. Нижнее Поволжье,
Северо-Западный Казахстан.
III. Закавказская форма. Размеры мелкие: вес до 3,5 кг; общая длина черепа
90—100 мм. Окраска летнего и зимнего меха почти одинаковая.
е) L. е. cyrensis Satunin, 1905. (Тип из окрестностей Барды, в Азербайджане.)
Окраска летнего меха тусклая, буровато-палевая. Закавказье.
Подотряд ОДНОПАРНОРЕЗЦОВЫХ ГРЫЗУНОВ.
Subordo SIMPLICIDENTATA
В верхней челюсти только одна пара резцов. Долотообразные ко-
ронки этих зубов покрыты эмалью только с передней стороны. Число
коренных зубов у видов, встречающихся в СССР, колеблется в верх-
ней челюсти от 3 до 5, а в нижней — от 3 до 4. В слепой кишке спи-
ральной складки стенки нет.
В фауне СССР—11 семейств.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ ПОДОТРЯДА
ОДНОПАРНОРЕЗЦОВЫХ ГРЫЗУНОВ ФАУНЫ СССР
1(2). Верхняя сторона тела покрыта длинными толстыми иглами
(длиной до 30 см). Ширина межглазничного промежутка черепа равна
или больше ширины мозговой коробки, измеренной в области слухо-
вых проходов. Длина очень широких носовых костей составляет около
72 длины черепа.............Дикобразовые. Hystricidae. (Стр. 253.)
2(1). Верхняя сторона тела покрыта более или менее мягким ме-
хом; игл на спине нет. Ширина межглазничного промежутка черепа
меньше ширины мозговой коробки, измеренной в области слуховых
проходов. Длина носовых костей меньше */2 длины черепа ... 3
248
Рис. 90. Череп дикобраза (Hystrix cristata)
сбоку и сверху.
3(4). Хвост очень широкий, уплощенный сверху, лопатообразный.
Коготь второго (считая изнутри) пальца задней конечности раздвоен.
Наружные слуховые проходы охвачены выдающимися косо вверх и
в бока костными трубочками — частями слуховых барабанов черепа.
Перед затылочным отверстием в основании черепа имеется глубокая
ямка, окруженная костными валиками...............................
...............................Бобровые. Castoridae. (Стр. 254.)
4(3). Хвост округлый в сечении или сжат с боков. Коготь вто-
рого пальца задних ног не раздвоен. Костных трубочек, направленных
косо вверх и в бока, вокруг слуховых проходов нет, или они зачаточны.
Глубокой ямки, окруженной кост-
ными валиками, перед затылоч-
ным отверстием черепа нет . 5
5(6). Пальцы задних ног
связаны почти на всем протяже-
нии плавательной перепонкой.
Длина тела взрослых особей бо-
лее 40 см. Соски у самок распо-
ложены на боках тела. Сосце-
видные отростки черепа очень
длинные (длина их примерно
равна длине слуховых бараба-
нов) . Ряды коренных зубов резко
сходятся своими передними кон-
цами ..............Нутриевые.
Capromyidae. (Стр. 255.)
6(5). Пальцы задних ног не
соединены перепонкой (единст-
венное исключение — ондатра
имеет длину тела менее 30 см).
Соски у самок расположены по
бокам брюха. Сосцевидные от-
ростки черепа значительно ко-
роче слуховых барабанов. Ряды
коренных зубов верхней челюсти
идут более или менее параллель-
но друг другу...............7
7(10). Лобные кости несут хорошо развитые, направленные назад
и наружу, заостренные заглазничные отростки. Коренных зубов в ниж-
ней челюсти 4, а в верхней 5; первый коренной верхней челюсти иногда
очень мал или даже отсутствует (в последнем случае длина черепа
более 4 см)....................................................8
8(9). На боках тела между передними и задними конечностями
широкой складки кожи — летательной перепонки — нет. Нет также
длинной тонкой косточки, отходящей от запястья и поддерживающей
эту перепонку. Носовой отдел черепа у основания заметно шире, чем
в средней части....................Беличьи. Sciuridae. (Стр. 255.)
9(8). На боках тела между передними и задними конечностями
имеется широкая складка кожи — летательная перепонка, покрытая
мехом. Спереди она поддерживается тонкой длинной косточкой — шпо-
рой, отходящей от запястья. Носовой отдел черепа имеет примерно оди-
наковую ширину у основания и в средней части (рис. 92)...........
.............................Летяжьи. Pteromyidae. (Стр. 270.)
10(7). Заглазничных отростков на лобных костях черепа нет,
или они имеют вид небольших тупых, смотрящих в бока бугорков.
249
Коренных зубов в верхних челюстях не более 4. Общая длина черепа
менее 4 см.....................................................11
11(12). В нижней челюсти с каждой стороны по 4 коренных зуба.
Хвост у всех видов, кроме одного (водящегося только в Туркмении),
покрыт густым мехом, волосы которого на всем протяжении или только
на конечной половине хвоста расчесаны на бока. Длина волос, покры-
вающих бока основной половины хвоста, больше толщины хвостового
стебля........................ Соневые. Muscardinidae. (Стр. 271.)
12(11). В нижней челюсти с каждой стороны только по 3 корен-
ных зуба. Хвост либо покрыт кольцами кожных чешуек с торчащими
из-под них щетинками, либо одет короткими, прилегающими к стержню
хвоста волосами, лишь иногда образующими на его конце кисть удли-
ненных волос. Длина волос основной части хвоста меньше толщины
его стебля....................................................13
13(14). Длина ряда коренных зубов верхней челюсти менее 2 мм.
Их жевательная поверхность чашеобразно-вогнутая, без бугорков и
петель эмалевых стенок зуба (рис. 93). Передняя поверхность верхних
резцов несет глубокую продольную борозду, которая разделяет конец
зуба на два зубца. Длина тела менее 10 см. Хвост немного короче тела
и покрыт густым мехом из коротких волосков, среди которых кожных
роговых чешуек нет .... Селевиниевые. Seleviniidae. (Стр. 275.)
250
Рис. 93. Зубы селевинип
(Selevinia betpakdalensis):
Рис. 92. Черепа (сверху):
А — белки (Sciurus vulgaris); Б — летяги (Pteromys
volansh
/ — резцы; М — коренные зубы.
Рис. 94. Черепа:
А, а — малого тушканчика (Allactaga elater); Б, б — мышовки (Sicista); В, в — домовой мыши
(Mus musculus); А — В — вид сверху; а — в — вид снизу.
14(13). Длина ряда коренных зубов верхней челюсти более 2,5 мм.
Жевательная поверхность этих зубов плоская или выпуклая, с бугор-
ками или петлями эмалевых стенок коронок различной конфигурации.
Передняя поверхность резцов верхней челюсти гладкая или с неглу-
бокой продольной бороздой, не разделяющей концы зубов на два
зубца. Длина тела обычно более 10 см (если менее, то либо хвост
длиннее тела, либо он покрыт кожными роговыми чешуйками, между
которыми растут короткие редкие щетинки)...................... 15
Рис. 95. Черепа сбоку и сверху:
А, а — слепыша (Spalax); Б, б — цокора (Myospalax).
15(16). Задние конечности очень длинные: они в 3—4 раза длин-
нее передних. Пальцев на задних ногах 3 или 5, если 5, то когти край-
них из них не достигают оснований трех средних. Хвост обычно не
менее чем на длиннее тела, причем на конце хвоста имеется упло-
щенная кисточка волос («знамя»). У видов, хвост которых короче и
не несет «знамени», он сильно утолщен в своей основной части за счет
отложения жира под кожей. Предглазничные отверстия черепа очень
велики: их просвет в 5—10 раз больше площади носового отверстия
черепа.......................Тушканчиковые. Dipodidae. (Стр. 275.)
16(15). Задние конечности не более чем в два раза длиннее пе-
редних. Пальцев на задних ногах 4 или 5, причем коготь обоих или
одного из крайних пальцев выдается вперед за основание средних
пальцев. Хвост либо короче, либо длиннее тела; в последнем случае он
не имеет «знамени». Предглазничные отверстия черепа не более чем
в три раза превышают площадь носового отверстия...............17
17(18). С каждой стороны верхней челюсти по 4 коренных зуба.
Длина тела менее 9 см. Хвост длиннее тела (обычно значительно)
................................Мышевковые. Zapodidae. (Стр. 286.)
18(17). С каждой стороны верхней челюсти .только 3 коренных
зуба. Размеры обычно более крупные: длина тела обычно более 9 см,
а если менее, то хвост короче тела............................19
19(20). Глаза есть. Бока головы покрыты мягким мехом. Хвост
есть (иногда весьма короткий). Затылочные мыщелки не видны при
252
рассматривании черепа сверху. Поперечный (ламбдоидальный) шов
у молодых или поперечный гребень у взрослых лежит на черепе позади
вертикали основания скуловых отростков чешуйчатых костей ....
...................................Мышиные. Muridae. (Стр. 289.)
20(19). Глаза скрыты под кожей. На боках головы от носа через
глаза тянутся полоски жестких щетинок. Хвоста нет. Затылочные мы-
щелки хорошо видны при рассматривании черепа сверху. Ламбдои-
дальный шов или гребень расположен по вертикали, проведенной
от основания скуловых отростков чешуйчатых костей или даже впереди
нее (рис. 95) .... Семейство Слепышовые. Spalacidae. (Стр. 344.)
СЕМЕЙСТВО ДИКОБРАЗОВЫЕ. FAMILIA HYSTRICIDAE
Крупные наземные грызуны, тело которых на спине, боках и хвосте
покрыто длинными и толстыми иглами, а на других частях — жесткой
щетиной. В черепе очень сильно развиты лицевые кости. Длина очень
широких носовых костей почти равна V2 общей длины черепа. Коренных
. 4
зубов —, их жевательная поверхность плоская.
Африка, Средиземноморье, Передняя, Средняя и Южная Азия. Из
двух палеарктических родов в фауне СССР— 1 род.
РОД ДИКОБРАЗЫ. GENUS HYSTRIX Linnaeus, 1758
Основные признаки рода см. в определительной таблице и характе-
ристике семейства.
Ареал, как у всего семейства. Из трех палеарктических видов
в фауне СССР— 1 вид.
14. Дикобраз. Hystrix cristata Linnaeus, 1758
Табл. XXV, 8. Карта 68
Юг Западной Европы, Северная Африка, Передняя, Средняя и
Южная Азия. В Советском Союзе водится в Азербайджане (Ленко-
рань, Талыш), в большей части Средней Азии, к северу до Мангыш-
лака, Центральных Каракумов, Ташкентского оазиса, Киргизского
хребта и Заилийского Алатау; в горах Центральной и Восточной Кир-
гизии и на Восточном Памире отсутствует.
Поселяется преимущественно в предгорьях и нижних частях скло-
нов гор, по обрывам и оврагам, на обработанных землях. Живет ко-
лониями или поодиночке в глубоких норах. Ведет ночной образ жизни.
Зимой мало активен, хотя в спячку не впадает. Размножается, по-ви-
димому, один раз в год, весной; самка приносит 2—4 детенышей. Пи-
тается летом травянистыми растениями, корнеплодами, фруктами, по-
чатками кукурузы и т. п. Зимой грызет побеги кустарников и деревьев,
их кору, поедает сухие стебли сорняков и других трав. Местами сильно
вредит бахчевым и огородным растениям, садовым деревьям, посевам
кукурузы, люцерны и других сельскохозяйственных культур. Мясо съе-
добно.
Подвиды:
а) Н. с. hirsutirostris Brandt, 1835. (Тип из Талыша.) Азербайджан.
b) Н. с. satunini Muller, 1911. (Тип из Геок-Тепе, в Южной Туркмении.)
253
СЕМЕЙСТВО БОБРОВЫЕ. FAMILIA CASTORIDAE
Крупные полуводные грызуны. Тело массивное. Хвост уплощенный
сверху вниз, покрытый крупными роговыми щитками. Пальцы задних
ног связаны плавательной перепонкой, передние свободны. Коготь вто-
рого пальца задних конечностей раздвоен, образуя своеобразные
щипчики, которые, по-видимому, помогают бобру расчесывать волосы
и удалять эктопаразитов. Ушные раковины при погружении зверя
в воду складываются вдоль. Ноздри при этом замыкаются. Коренных
зубов — . Евразия и Северная Америка.
Один род.
РОД БОБРЫ. GENUS CASTOR Linnaeus, 1758
Признаки и ареал приведены в характеристике семейства. Из двух
близких видов, самостоятельность которых сомнительна, в фауне СССР
обитает 1 вид (другой — С. canadensis Kuhl, 1820 — распространен
в большей части Северной Америки).
15. Бобр. Castor fiber Linnaeus, 1758
Табл. XXV, 9. Карта 68
Прежде был широко распространен по всей лесной полосе Европы
и Азии, но хищнический промысел повлек исчезновение этого ценного
пушного зверя в большей части районов его прежнего ареала. В на-
шей стране в начале XX в. бобры в количестве 1000—1200 штук со-
хранились лишь кое-где в Белоруссии, в Северной Украине, Смолен-
ской, Воронежской и Тамбовской губерниях, в Северном Зауралье
и в верховьях Енисея. Ныне бобры тщательно охраняются и ведется
большая работа по их реакклиматизации в ряде районов их прежнего
распространения. Численность бобров возрастает.
Живут бобры колониями и поодиночке преимущественно по лесным
речкам в норах и в устроенных из кусков стволов деревьев куполооб-
разных «хатках» (последние строятся главным образом на топких ме-
стах). Иногда бобры делают поперек реки плотину, поднимая уровень
воды. Летом питаются травянистыми растениями, а зимой — корой
и побегами лиственных деревьев, которые валят, подгрызая у корня.
Спаривание происходит в январе — феврале. Беременность длится
105—107 дней. В помете обычно 2—4 детеныша. Половозрелыми
молодые становятся на третьем году жизни. Очень хорошо плавают
и ныряют, на суше редко отходят от берега. Мех бобров высоко
ценится.
В нашей стране различают следующие подвиды бобра:
а) С. f. fiber Linnaeus, 1758. (Тип из Швеции.) Окраска меха либо рыжевато-бу-
рая, либо почти черная, подпушь темная. Европейская часть СССР, завезен в ряд
районов Сибири.
b) С. f. pohlei Serebrennikov, 1929. (Тип с р. Сосьвы.) Окраска большинства осо-
бей светло-коричневая или светло-кофейная, подпушь светлая. Бассейны рек Конды
и Сосьвы, Тува; завезен в некоторые районы Сибири.
254
СЕМЕЙСТВО НУТРИЕВЫЕ. FAMILIA CAPROMYIDAE
Единственный род.
РОД НУТРИИ. GENUS MYOCASTOR Kerr, 1792
Крупные грызуны, ведущие земноводный образ жизни. Массивное
тело несколько сдавлено с боков. Длинный, округлый в сечении хвост
покрыт чешуйками и редкими короткими волосами. Передние лапы
малы, пальцы их не связаны перепонкой. Задние конечности более
длинные и сильные, четыре пальца их соединены плавательной пере-
понкой, пятый свободен. Крупная голова несет небольшие округлые
ушные раковины. Мех нутрии состоит из длинной жесткой ости и гу-
4
стого мягкого пуха. Коренных зубов — с каждой стороны челюсти. Ро-
4
дина этих животных — Южная Америка. В роде один вид, акклимати-
зированный в СССР.
16. Нутрия. Myocastor coypus Mollina, 1782
Табл. XXV, 7. Карта 68
Широко распространена в южной половине Южной Америки.
В СССР акклиматизирована в Закавказье, Таджикистане, Туркмении
и в некоторых районах Северного Кавказа. Разводится также в зверо-
водческих хозяйствах. В местах акклиматизации живет по берегам во-
доемов. Убежищами служат гнезда, лежки и неглубокие норы. Пита-
ется водяными и прибрежными растениями. Размножение происходит
в течение всего года. Беременность длится 128—133 дня, в помете — от
1 до 10 детенышей. Дает ценные шкурки.
СЕМЕЙСТВО БЕЛИЧЬИ. FAMILIA SCIURIDAE
Грызуны средних и крупных размеров (величиной от крысы до
кошки). Задние ноги не более чем в два раза превышают по длине
передние. Хвост разной длины, всегда покрыт волосами. Череп с широ-
ким межглазничным промежутком и хорошо развитыми, направлен-
ными назад и наружу заостренными заглазничными отростками.
1 2 (1) 3
Зубная формула: I — Pm——М, — -2 = 22(у одного вида 20 зубов).
1 13
У рода белок передний подкоренной верхней челюсти очень мал,
имеет вид небольшого тонкого столбика, прилегающего к передней сто-
роне следующего зуба; иногда этот зуб отсутствует.
Распространены беличьи по всем материкам, кроме Австралии
и Антарктиды. Из крупных островов они отсутствуют в Гренландии,
в Новой Зеландии, Новой Гвинее и на Мадагаскаре.
Это семейство, в состав которого входят сурки, суслики, бурундуки
и белки, — одно из наиболее богатых видами среди грызунов. В СССР
обитают представители 5 родов.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ БЕЛИЧЬИХ ФАУНЫ СССР
1(2). Размеры крупные: длина тела взрослых более 39 см; длина
ступни более 65 мм; кондилобазальная длина черепа не менее 75 мм;
длина максиллярного ряда коренных зубов более 18 мм..........
..................................Сурки. Marmota. (Стр. 257.)
255
2(1). Размеры мельче: длина тела менее 38 см\ кондилобазальная
длина черепа не более 63 мм\ длина максиллярного ряда коренных зу-
бов менее 17 мм.................................................3
3(6). Ушные раковины неразвиты: они имеют вид небольших кож-
ных валиков, окружающих слуховое отверстие. Длина хвоста (без кон-
цевых волос) составляет менее 60% длины тела. Первый верхний пред-
коренной зуб относительно крупный: он лишь в 2—3 раза уступает по
Рис. 96. Черепа грызунов семейства беличьих (изображены в разных масштабах):
А, а — крапчатый суслик (Cittellus suslicus); Б, б — тонкопалый суслик (Spermophilopsis leptodacty-
lus); В, в — бурундук (Tamias sibiricus); А — В — вид сверху; а — в — снизу.
площади жевательной поверхности второму предкоренному зубу. Если
же первый верхний предкоренной меньше, то концы надглазничных
отростков расположены значительно дальше кзади, чем задний конец
ряда верхних коренных зубов.......................................4
4(5). На пальцах задних ног нет пучков удлиненных волос, захо-
дящих за концы когтей. На пальцах передних ног длина самых длин-
ных когтей менее 10 мм. Первый предкоренной зуб верхней челюсти
относительно крупный: лишь в 2—3 раза уступающий (по площади же-
вательной поверхности) второму предкоренному зубу. Ширина меж-
глазничного промежутка составляет менее 28% (обычно менее 25%)
кондилобазальной длины черепа. У самок 5—7 пар сосков...............
......................................Суслики. Citellus. (Стр. 260.)
256
5(4). На крайних (внутренних и наружных) пальцах задних ног
есть пучки удлиненных волос, заходящих своими концами за концы
когтей. На пальцах передних ног длина самых длинных когтей более
10 мм. Первый предкоренной зуб верхней челюсти (если он есть)
мелкий: в 4—6 раз меньше второго предкоренного зуба. Ширина меж-
глазничного промежутка
составляет более 28%
(обычно более 30%) кон-
дилобазальной длины чере-
па. У самок 3—4 пары со-
сков ....................
Тонкопалые суслики.
Spermophilopsis. (Стр. 265.)
6(3). Ушные раковины
нормально развиты, иногда
с кисточками длинных во-
лос. Длина хвоста (без кон-
цевых волос) почти во всех
случаях составляет более
60% длины тела. Первый
верхний предкоренной зуб
очень мал: площадь его в
4—10 раз меньше площади
смежного с ним предкорен-
ного зуба. Иногда первого
предкоренного зуба вообще
нет. Концы надглазничных
отростков расположены над
(или впереди) задним кон-
цом ряда верхних коренных
зубов...................7
7(8). На спине имеет-
ся 5 черных или чернова-
тых продольных полос. Дли-
на тела менее 17 см. Кон-
дилобазальная длина чере-
па менее 39 см. Ширина
межглазничного промежут-
ка менее 28% кондилоба-
зальной длины черепа. Пер-
вый верхний предкоренной
зуб в 4—5 раз меньше вто-
рого .... Бурундуки.
Tamias. (Стр. 266.)
8(7). На спине нет чер-
ных продольных полос. Дли-
Рис. 97. Хвосты и ступни:
А, а — желтый суслик (Citellus maximus); Б, б — тон-
копалый суслик (Spermophilopsis leptodactylus). Видны
пучки удлиненных волос на крайних пальцах тонкопа-
лого суслика.
на тела взрослых более 18 см. Кондилобазальная длина черепа не ме-
нее 42 мм. Ширина межглазничного промежутка более 28% кондило-
базальной длины черепа. Первый верхний предкоренной зуб в 8—10 раз
меньше второго или совсем отсутствует . . Белки. Sciurus. (Стр. 266.)
РОД СУРКИ. GENUS MARMOTA Blumenbach, 1779
Крупные, весом до 10 кг, грызуны с массивным, мешкообразным
телом, короткими ногами, уплощенной сверху головой. Длина пышно-
17 Н. А. Бобринский и др.
257
опушенного хвоста составляет от V6 до V2 длины тела. Ушная раковина
редуцирована. Мех на спине и боках густой, на брюхе очень редкий.
Обитают среди целинной степи, по степным балкам, на сопках мел-
косопочника, по склонам гор, на горных сыртах. Живут в глубоких
норах обычно с многими входами. На зиму впадают в продолжитель-
ную (до 7 месяцев) спячку. Весной приносят помет из 2—8 детенышей.
Полного роста и развития достигают на третьем году жизни. Пита-
ются различными травянистыми растениями. Местами вредят пастби-
щам и особенно сенокосам (нарытые сурком холмики земли мешают
механизированному сенокошению). Служат объектом пушного про-
мысла. Являются разносчиками чумы.
Голарктика. Из 7—8 видов сурков в фауне СССР — 3 вида.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА СУРКОВ
ФАУНЫ СССР
1(2). Хвост с его концевыми волосами составляет около х/г длины
тела. Окраска всей шкурки ярко-рыжая или буровато-рыжая, более
или менее сильно затемненная на спине черной или бурой остью . . .
...............Длиннохвостый сурок. Marmota caudata. (Стр. 258.)
2(1). Длина хвоста составляет лишь около длины тела. Окраска
иная..........................................................: 3
3(4). Окраска щек и боков шеи постепенно переходит в более тем-
ную расцветку верха головы и шеи. Длина тела взрослых особей более
45 см. Общая длина черепа взрослых обычно более 87 мм...............
..................Сурок обыкновенный. Marmota bobak. (Стр. 259.)
4(3). Желтоватая или светло-коричневая окраска щек и боков шеи
резко граничит с темной расцветкой верха головы и шеи по линии,
идущей дугообразно от глаза над ухом к основанию плеча. Длина тела
обычно менее 45 см, а общая длина черепа менее 87 мм................
..................Сурок Мензбира. Marmota menzbieri. (Стр. 260.)
Примечание. До последнего времени большинство авторов считали, что
фауна СССР содержит значительно большее количество видов сурков, признавая за
сурком-байбаком, алтайским сурком, тарбаганом и камчатским сурком видовую
самостоятельность. Но исследования последних лет показали, что сурки мелкосопоч-
ника Казахстана носят промежуточные признаки между байбаком и алтайским сур-
ком, а сурки Хангая — между сурком алтайским и тарбаганом, и эти формы сле-
дует рассматривать лишь как подвиды единого вида — сурка обыкновенного.
17. Длиннохвостый сурок. Marmota caudata Jacquemont, 1844
Табл. XXV, 1. Карта 69
Горы Средней Азии, к югу — до Кашмира, к западу — до запад-
ных предгорьев Таласского и Туркестанского хребтов, к северу — до
предгорьев Западного Тянь-Шаня, к востоку — до линии: ущелье Аксу
на Киргизском хребте — долина р. Джумгала — район г. Нарына —
южная часть Ферганского хребта — восточные предгорья Заалайского
хребта — горы Кашгарского хребта.
Селится на высокогорных сыртах, на участках горной степи, альпий-
ских лугах, субальпийских лужайках лесного пояса и даже на увалах
предгорьев. Живет колониями и поодиночке.
Важный объект пушного промысла Киргизии.
В СССР М. с. aurea Blanford, 1875. (Тип из Яркенда, Кашгария.) Отличается от
номинальной формы, обитающей в Кашмире, более мелкими размерами.
258
18. Сурок обыкновенный. Marmota bobak Muller, 1776
Табл. XXV, 2—5. Карта 69
Еще в начале прошлого века этот сурок был широко распростра-
нен по степям и лесостепи Восточной Европы от Венгрии до Южного
Урала. Ныне в пределах Европы он сохранился отдельными колониями
в Восточной Украине, на Дону, в Среднем и Нижнем Поволжье, в За-
волжье и на Южном Урале. Более обычен среди целинных степей За-
уралья и Северного Казахстана; обитает среди сопок Казахского мел-
косопочника.
Водится в горах и местами в предгорьях Алтая, Тарбагатая и Во-
сточного Тянь-Шаня (к востоку от линии: ущелье Аксу на Киргизском
хребте — долина р. Джумгалы — район г. Нарына, урочище Арпа).
Широко распространен в горах и степях Монголии и Западного Китая,
заселяет степи Забайкалья, горы у оз. Байкал, горную часть Южной
и Восточной Якутии, Анадырского края, внутренней Чукотки.и Кам-
чатки. Акклиматизирован в Дагестане (на Гунибе).
Селится на участках целинной степи, по степным балкам, на соп-
ках мелкосопочника, на луговых пространствах предгорьев, на субаль-
пийских и альпийских лугах, среди горной степи, на сыртах. В обла-
стях распашки целины численность сурков сокращается.
Во многих местах служит предметом пушного промысла. Местами
опасен как источник чумы.
Образует ряд хорошо выраженных подвидов. Особи, обитающие в равнинной
степи, характеризуются низким, хотя и густым, мехом. Мех горных форм, наоборот,
отличается большой пышностью.
а) М. b. bobak Miiller, 1776. (Описан из Польши.) Окраска спины в осеннем мехе
рыжевато-песчано-желтая, брюхо рыжеватое. Мех низкий, малоплотный. Размеры
крупные: общая длина черепа 85—Q05 мм. Степи Европейской части СССР на восток
до Волги.
b) М. b. tschaganensis Bazhanov, 1930. (Тип с р. Чагана, приток Урала.) Окраска
осеннего меха несколько более светлая и тусклая, слабо затемненная буроватой
остью. Брюхо песчано-желтое. Степи Южного Урала и Северного Казахстана. Форма,
близкая к номинальной.
с) М. b. aphanasievi s. sp. nova. Форма, носящая промежуточные признаки между
двумя предыдущими и алтайским подвидами. Окраска осеннего меха буровато-желтая
с черноватой остыо. Мех средней пышности. Мелкосопочник Казахстана (горы Кызыл-
Рая, Нур-Тая, Кента, Каркары и других горных групп).
d) М. b. baibacina Brandt, 1843. (Тип из окрестностей Черги, на Алтае.) Окраска
осеннего меха спины буровато-желто-серая; подпушь песочно-желтая, ость чернова-
тая. Брюхо желто-ржаво-буроватое. Верх головы немного темнее спины. Мех пышный
и мягкий. Размеры крупные: общая длина черепа 85—105 мм. Алтай1, Тарбагатай,
Сауры. Отдельные natio: М. b. b. kastschenkoi Strog et Judin, 1956 (Томская и Кемеров-
ская области), и М. b. ognevi Scalon, 1950 (Юго-Западный Алтай).
е) М. b. centralis Thomas, 1909. (Тип с хребта Боро-Хото.) Сходен с предыдущим
подвидом, но окраска осеннего меха светлее в силу меньшего затемнения бурой
остью. Брюхо ржаво-буроватое. Горы Тянь-Шаня.
f) Al. b. caliginosus Bannikov et Scalon, 1949. (Тип с Шара-Усу, на Хангае.)
Этот подвид носит промежуточные признаки между М. b. baibacina и М. b. sibirica.
От алтайского сурка отличается более низким мехом, более светлой окраской,
на лбу и темени образуется темная шапочка. Хангай, Монгольский и Гобийский
Алтай.
g) М. b. sibirica Radde, 1862. (Тип с берегов оз. Торей-Нур, в Забайкалье.)
Окраска осеннего меха спины светлая, палево-буровато-серая; брюхо ржаво-бурова-
тое. Верх головы («шапочка») темный. Размеры чуть меньше предыдущих: общая
длина черепа 80—100 мм. Мех низкий, но плотный. Степи Забайкалья и Восточной
Монголии.
h) М. b. doppelmayeri Birula, 1922. (Тип с Баргузинского хребта.) Окраска осен-
него меха на спине светлая, палево-желтоватая, сравнительно слабо затемненная
черноватыми концами остевых волос. Темная «шапочка» на голове достигает только
ушной области. Брюхо тускло-ржавое. Мех средней пышности. Размеры мелкие:
общая длина черепа 78—85 мм. Горы Забайкалья.
17*
259*
i) M. b. bungei Kastschenko, 1901. (Тип с p. Омолоя, в Якутии.) Окраска осеннего
меха более темная, чем у предыдущей формы, сильно затемненная черноватыми
остевыми волосами. Брюхо интенсивно красно-рыжее. «Шапочка» на голове спускается
до глаз и охватывает междуушье. Мех очень пышный. Размеры немного крупнее:
общая длина черепа 82—98 мм. Горы Восточной и Северной Якутии.
j.) М, b. camtschatica Pallas, 1811. (Описан с Камчатки.) Окраска осеннего меха
спины еще более затемненная черноватой остью. Темная «шапочка» на голове сзади
незаметно переходит в темную расцветку загривка. Брюхо интенсивно рыжее. Мех
пышный. Размеры чуть крупнее предыдущей формы: общая длина черепа 88—101 мм.
Камчатка, Анадырь.
Сурок Мензбира. Marmota menzbieri Kaschkarov, 1925
Табл. XXV, 6, Карта 69
Найден в верховьях рек Угама, Пскема, Чаткала и Агрена, в За-
падном Тянь-Шане.
Селится на горных лугах, на травянистых склонах гор, близ каме-
нистых россыпей L
РОД СУСЛИКИ. GENUS CITELLUS Oken, 1816
Грызуны среднего размера (длина тела 20—38 см), с вальковатым
телом и относительно коротким хвостом, покрытым расчесанными на
бока волосами. Ушные раковины редуцированы и имеют вид кожного
валика вокруг слухового прохода. Мех на брюхе редкий, прилегающий.
Имеются защечные мешки.
Распространены на громадных пространствах Европы, Азии и Се-
верной Америки.
Обитают преимущественно в открытых ландшафтах от тундр Се-
веро-Восточной Сибири и якутских степей до пустынь и гор Средней
и Центральной Азии. Живут на лугах, пастбищах, выгонах, на межах
полей. Питаются преимущественно сочными травянистыми растениями,
луковицами, зернами (главным образом в ранних стадиях зрелости).
На зиму кормовых запасов не собирают. Размножаются один раз
в году — весной. В одном помете бывает от 2 до 15 слепых и го-
лых детенышей. Прозревший молодняк вскоре после выхода из нор
на поверхность расселяется; каждый зверек занимает отдельную нору.
Взрослые суслики летом, а молодые к началу осени сильно жиреют
и впадают в глубокую спячку до следующей весны.
Некоторые виды сусликов причиняют огромный вред сельскому
хозяйству уничтожением посевов различных сельскохозяйственных
культур и ухудшением пастбищ. Некоторые суслики, особенно малый,
служат источником болезней, опасных для человека и домашних жи-
вотных (чумы, бруцеллеза и др.). На обширных территориях, где оби-
тает большое количество сусликов, ежегодно проводят работы по ис-
треблению этих зверьков. В- одних случаях рассеивают с самолетов
отравленные зерновые приманки, в других — применяют «газовый ме-
тод»— затравливают норы цианоплавом и другими веществами.
Шкурки сусликов используют как пушное сырье. Вытопленный из
тушек жир может быть использован для технических целей.
Различают около 40 видов сусликов, из которых более 30 встреча-
ется в Северной Америке. В фауне СССР — 6 видов.
1 Систематическое положение сурка Мензбира пока остается неокончательно вы-
ясненным.
260
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА СУСЛИКОВ ФАУНЫ СССР
1(10). Хвост короткий: длина его без концевых волос не более
V2 длины тела.......................................................2
2(3). Окраска спины, боков тела и брюха песчано-желтая, затем-
ненная по хребту черноватыми окончаниями остевых волос................
.....................Желтый суслик. Citellus maximus. (Стр. 261.)
3(2). Окраска иная..............................................4
4(5). На щеках резко выделяются рыжие или коричневые пятна.
Размеры крупные: длина ступни обычно более 37 мм; кондилобазаль-
ная длина черепа в большинстве случаев более 40 мм....................
........................Большой суслик. Citellus major. (Стр. 262.)
5(4). На щеках ярких рыжеватых или коричневых пятен нет; если
намечаются очень нерезко выраженные пятна, то длина задней ступни
менее 35 мм, а кондилобазальная длина черепа менее 40 мм ... 6
6(7). На спине резко выступает рисунок из ясно очерченных бе-
ловатых крапин на коричневатом или буроватом фоне .......
...................Крапчатый суслик. Citellus suslicus. (Стр. .262.)
7(6). Окраска спины землисто-серая однотонная или со слабо
заметным рисунком из неясно очерченных чуть более светлых крапин . 8
8(9). На спине обычно заметны неясные более светлые крапины.
Подошвы задних лап голые. Хвост, как правило, короче V4 длины тела.
.....................Малый суслик. Citellus pygmaeus. (Стр. 263.)
9(8). Окраска спины однотонная, иногда с чуть заметной белесой
струйчатой рябью, но без крапин. Подошвы задних лап (кроме поду-
шечек) покрыты волосами (исключение — суслики этого вида с гор
Средней Азии). Хвост обычно более !/4 длины тела......................
..........................Серый суслик. Citellus citellus. (Стр. 264.)
10(1). Длина хвоста (без концевых волос) примерно равна поло-
вине длины тела.......................................................
.............Длиннохвостый суслик. Citellus undulatus. (Стр. 264.)
19. Желтый суслик. Citellus maximus Pallas, 1778
Табл. XXVI, 1. Карта 70
Обитает на равнинных и предгорных пространствах Средней Азии
(кроме песков Каракум и Кызылкум и Ферганской долины), Южного
Казахстана (на север до линии: район г. Чапаевска — верховья
р. Ишима — среднее течение р. Сырысу — низовья р. Чу) и в Южное
Заволжье. Встречается в Северном Иране и Афганистане.
Водится преимущественно в глинистых и песчанистых пустынях
и полупустынях, но голых песков избегает. Живет в норах, обычно
колониями. Спячка длится в южных пустынных районах ареала
с июня — июля по конец февраля — начало марта, а в северных его
частях — с конца июля по март. В помете от 4 до 11 (обычно 6—8)
детенышей. Питается пустынными травами и полукустарниками. Цен-
ный пушной зверек.
По окраске меха можно различить следующие подвиды:
а) С. m. maximus Pallas, 1778. (Описан с низовьев р. Урала.) Окраска серовато-
желтая с сильным затемнением черной остыо на спине. Актюбинская, Кзыл-Ордин-
ская и Карагандинская области.
b) С. m. orlovi Ognev, 1937. (Тип из района г. Вольска.) Окраска более яркая»
рыжевато-песчаная, менее сильно затемненная на спине темными окончаниями осте-
вых волос. Южное Заволжье.
с) С. m. oxianrs Thomas, 1915.-(Тип из района г. Бухары.) Окраска светлая,
тусклая, песчано-ж*лтая, слабо затемненная на спине темными концами остевых
волос. Южный Казахстан, Средняя Азия.
261
20. Большой суслик. Citellus major Pallas, 1779
Табл. XXVI, 2. Карта 70
Распространен в степных и лесостепных районах Заволжья, Юж-
ного Урала, Западной Сибири, в Центральном и в Восточном Казах-
стане. Водится в различных частях Центральной Азии. Селится на
целине, парах, близ пашен, на выгонах. Местами приносит вред на
полях.
Подвиды фауны СССР:
а) С. m. major Pallas, 1778. (Описан из района г. Куйбышева.) Наиболее круп-
ная форма: длина тела 25—38 см. Окраска спины серебристо-серая со струйчатой
рябью. Бока, брюхо и щеки рыжие. Заволжье.
b) С. m. argyropulo Bazhanov, 1947. (Тип с р. Уила.) Крупная форма: длина
тела до 30 см. Окраска немного светлее с примесью песчаных тонов. Брюхо и бока
охристые; мех ниже и реже. Бассейны Уила, Эмбы, Темира.
с) С. m. ungae Martino, 1923. (Тип из окрестностей Омска.) Размеры немного
меньше: длина тела 23—28 см. Окраска спины серая со светлой беловатой рябью.
Брюхо желтоватое. Под глазами резко очерченные ржаво-коричневые пятна. Степи
между Ишимом и Иртышом.
d) С. m. erythrogenys Brandt, 1841. (Описан из предгорьев Алтая.) Размеры, как
у предыдущей формы. Окраска землисто-серая с желтоватой рябью. Брюхо тускло-
желтое. Степи Западной Сибири к востоку от Иртыша.
е) С. m. brevicauda Brandt, 1843. (Описан из области «между Алтаем и Балха-
шем».) Размеры еще меньше: длина тела 22—25 см. Окраска светлее и желтее; крап-
чатый рисунок не выражен. Казахское нагорье, Зайсанская котловина, Бет-Пак-Дала.
Синонимы: С. m. intermedius Brandt, 1844, описанный из области к северу от Бал-
хаша, и С. m. iliensis Beljaev, 1945, описанный с левобережья р. Или, вряд ли можно
практически отличить от С. m. brevicauda.
f) С. m. carruthersi Thomas, 1912. (Тип с Джунгарского Алатау.) Близок к преды-
дущей форме: размеры такие же мелкие, но окраска тусклая с преобладанием
серых тонов. Джунгарский Алатау и Северный и Западный Китай.
В Центральной Азии широко распространен С. m. pallidicauda Satunin, 1903.
21. Крапчатый суслик. Citellus suslicus Giildenstaedt, 1770
Табл. XXVI, 5—6. Карта 70
Распространен к северу до линии: Ровно — Житомир—Киев —
Чернигов — Брянск—Калуга и далее на восток до р. Оки и Волги.
К востоку встречается до р. Волги на протяжении от Казани до Са-
ратова; к югу до линии: Саратов — Изюм — Хорол и далее до
Днепра и Черноморского побережья Украины; к западу до государст-
венной границы. Отдельные колонии имеются в южных районах Бело-
руссии.
Населяет степные участки лесостепи и разнотравные черноземные
степи; обитает на выгонах, по балкам, на межах полей, обочинах до-
рог. Живет колониями и отдельно. Зимняя спячка длится с августа —
начала сентября по конец марта — апрель. Раньше причинял огромный
вред посевам. После проведения ряда мер по его истреблению числен-
ность и вред крапчатого суслика сократились. Шкурка пригодна для
выделки меховых изделий.
В СССР различают 2 близкие формы:
а) С. s. suslica Giildenstaedt, 1770. (= С. s. averini и С. s. meridio occidentalis.
Описан из воронежских степей.) Окраска относительно светлая и тусклая. Крапины
выступают менее отчетливо. Южная часть ареала на север до Орловской и Тамбов-
ской областей.
b) С s guttatus Pallas, 1770. (Описан с рек Пьяны и Суры.) Окраска более тем-
ная, крапчатость более отчетливая. Северные части ареала.
262
22. Малый суслик. Citellus pygmaeus Pallas, 1779
Табл. XXVI, 3—4. Карта 71
Сухие степи, полупустыни, глинистые пустыни от Степного Крыма
и Перекопского перешейка через низовья Дона, Северный Кавказ,
Калмыкию, Нижнее Поволжье, Западный и Средний Казахстан к во-
стоку до Казахского нагорья, на юг до альпийского пояса Кабардино-
Балкарии, Дагестана, Среднего Устюрта, низовьев р. Сыр-Дарьи, к се-
веру почти до Полтавы и далее примерно до линии: Изюм — Мальчев-
ская — Калач — Урюпинская — Саратов — Кинель — Хайбуллино —
Троицк — Кустанай — Атбасар.
Обитает в глинистых (светло-каштановых и сероземных) полынно-
злаковых полупустынях, сухих степях, полынно-солянковых пустынях.
Где много полей, поселяется по их межам, по обочинам грунтовых до-
рог, на выгонах, пастбищах и т. д. Местами (например, на Ергенях)
весьма многочислен: весной до 50 и более взрослых сусликов на 1 га,
а летом, после расселения молодняка, до 120—150 зверьков на 1 га.
Живет в норах с наклонными и вертикальными выходами. Гнезда в вы-
водковых и зимовочных норах располагаются на глубине до 1,5—2 м.
На зиму все выходы из норы закупориваются земляными пробками.
Весной выходы снова открываются, очищаются и строятся новые норы.
Самый вредный из всех видов сусликов фауны СССР, совершает на-
беги на посевы сельскохозяйственных культур. На выбросах из нор
вырастают растения, не поедаемые домашним скотом, а выбросами бы-
вает покрыто до половины всей площади пастбищ. Миллионы сусликов
кормятся между выбросами, обедняя кормовую продуктивность паст-
бищ. Легко находят и поедают прорастающие желуди, высаженные при
создании полезащитных и противоэрозионных насаждений. Служат
источником бруцеллеза рогатого скота и чумы, опасной для человека.
Истребление малого суслика ежегодно проводится на громадных пло-
щадях. Самолетами проводят рассев зерна, обклеенного фосфидом
цинка на растительном масле. Около селений и птицеферм истребляют
затравкой нор цианплавом. В большом количестве их вылавливают
капканами. Шкурки используют как пушное сырье.
Описано большое количество подвидов, слабо различающихся по тону окраски
меха. Большую часть этих форм практически различить невозможно даже на боль-
ших сериях. Заметно различаются следующие подвиды:
а) С. р. pygmaeus Pallas, 1779. (Описан из полупустынь между Эмбой и Уралом.)
Окраска меха светлая, глинисто-серая с примесью желтых оттенков. Верх головы
сравнительно яркий, рыжевато-желтый. На конце хвоста темной каймы нет. Южные
районы Волго-Уральского и Урало-Эмбинского междуречий. От номинальной формы
практически не отличимы: С. р. herbicola Martino, 1916, описанный из актюбинских
степей; С. р. nikolskii Heptner, 1934, из Северного Приуралья (включая Барсуки) и
Устюрта; С. р. kasakstanicus Goodwin, описанный из Туэ-Булака, к северу от Кзыл-
Орды; С. р. arenicola Rail, 1935; С. р. ralli Heptner, 1948.
b) С. р. mugosaricus Lichtenstein, 1823. (Описан с Мугоджарских гор.) Близок
к номинальной форме. Окраска меха лишь немного темнее, насыщеннее.
с) С. р. septentrionalis Obolensky, 1927. (Тип из Ферапонтовки Куйбышевской
области.) По окраске близок к предыдущей форме: верх относительно темный; верх
головы яркий, рыжеватый; на конце хвоста иногда имеется темная кайма. Степи За-
волжья к югу, вероятно, до широты Уральска.
d) С. р. planicola Satunin, 1909. (Тип из карапогайских степей бывшего Кизляр-
ского округа.) В окраске верха преобладают сероватые тона; брюхо окрашено серее,
чем у номинальной формы; имеется темная кайма на хвосте. Низовья Терека и при-
лежащие полупустыни; предгорные районы Кабардино-Балкарии.
е) С. р. satunini Sviridenko, 1922. (Тип из Темир-Хана, Дагестан.) Отличается от
предыдущей формы примесью палево-желтого тона в окраске спины, более бледной
крапчатостью; в окраске хвоста проступает рыжеватый тон. Дагестан, ряд изолиро-
ванных колоний к югу до Сулака.
263
f) C. p. musicus Menetries, 1832. (Описан с подножия Эльбруса.) Окраска верха
темная; крапчатость плохо заметна; на конце хвоста широкая черноватая полоса.
Крупнее других форм. Горно-степной и альпийский пояс средней части Большого
Кавказа (верхние притоки Кубани и Терека, склоны Эльбруса).
g) С. р. boehrriii Krassovski, 1932. (Тип из Нижних Аталук, Ингушетия.) Форма
близкая к предыдущей, но несколько мельче и светлее; пятнистость выражена резче;
брюхо с примесью желтовато-ржавых тонов. Северная Ингушетия.
h) С. р. ellermani Harris, 1914. (Новое название вместо С. р. pallidus Orlov ct
Feniuk, 1927, и вместо С. р. orlovi Ellerman, 1940. Тип из урочища Улан-хол, в Калмы-
кии.) Окраска меха светлее и серее, чем у номинальной формы; верх головы одно-
тонный, сероватый; на конце хвоста темной полосы обычно нет. Калмыцкие степи и
полупустыни; пустыни приустьевой части Волги.
i) С. р. kalabuchovi Ognev, 1937. (Тип из Заветнинского района.) Окраска верха
бледная, палево-серая; крапчатость обычно ясная. Хвост насыщенно охристый без
черной каймы. Сальские и Манычские степи.
j) С. р. brauneri Martino, 1917. (Тип из бывшего Новомосковского уезда.)
Окраска верха относительно темная, серая с палевым оттенком; брюхо грязно-жел-
товатое. Крапчатость верха ясная. Предконцевая темная полоса на хвосте обычно
хорошо выражена. Крым, Левобережная Украина, приазовские и донские степи.
23. Серый суслик. Citellus citellus Linnaeus, 1766
Табл, XXVI, 7—10. Карта 71
Ареал этого вида тянется от Альп через юг средней Европы, Ма-
лую Азию, горы Средней Азии, степи и полупустыни Северного Китая
и Монголии на восток до степей Северо-Восточного Китая, В СССР
встречается в Западной Украине (в Черновицкой, Подольской, Винниц-
кой областях), в Молдавии, Западной Армении, горах Средней Азии и
в забайкальских степях. Обитает в самых различных степных угодьях:
на степных островках лесостепи, на участках горной степи, на полупу-
стынных увалах гор и предгорий, в сухой песчанистой степи. Живет
обычно колониями. В районах с развитым земледелием приносит вред
посевам. В Монголии и Китае этот вид сусликов включается в число
животных — распространителей чумных эпизоотий.
Подвиды фауны СССР:
а) С. с. citellus Linnaeus, 1776. (Описан из Австрии.) Размеры средние: длина
тела 18—24 см. Окраска спины буровато-охристая, обычно с заметной мелкой рябыо.
Брюхо желтоватое. Подошвы задних лап покрыты волосами. В СССР — Западная
Украина, Молдавия.
b) С. с. xanthoprymnus Bennett, 1835. (Описан из Эрзурума, в Восточной Турции.)
Размеры те же. Окраска с более выступающими рыжеватыми оттенками и слабо
выраженной рябыо. Подошвы лап покрыты волосами, но по средней линии их идет
голая полоска. Западная Армения, Малая Азия.
с) С. с. relictus Kaschkarov, 1923. (Тип с гор Таласский Алатау.) Размеры более
крупные: длина тела 23—28 см. Окраска спины насыщенная, землисто-серая, со слабо
заметными пестринами. Подошвы задних лап голые. Гиссарский, Ферганский, Чат-
кальский, Кавакский, Таласский хребты.
d) С. с. ralli Kuznetzov, 1948. (Тип из котловины оз. Иссык-Куль.) Крупнее. Окраска
спины более светлая с довольно- ясным рыжеватым оттенком, пестрины на спине вы-
ражены довольно ярко. Подошвы задних лап обычно голые. Восточная часть котловины
оз. Иссык-Куль.
е) С. с. dauricus Brandt, 1844. (Тип из окрестностей оз. Торей-Нор, Забайкалье.)
Размеры, как у европейской и малоазиатской форм. Окраска спины однотонная —
светлая, желтовато-серая. Подошвы задних лап частично покрыты волосами. Степи
Юго-Восточного Забайкалья и Восточной Монголии.
24. Длиннохвостый суслик. Citellus undulatus Pallas, 1779
Табл. XXVI, //, 12. Карта 72
Северная часть Северной Америки и Азии. В СССР водится в го-
рах Восточного Тянь-Шаня, Алтая, Саян, в присаянских районах,
в Прибайкалье, Забайкалье, по верхнему течению Амура, в Южной н
Восточной Якутии, в Анадырско-Чукотском крае и на Камчатке.
264
Обитает на горных лугах и травянистых склонах, по долинам рек,
на лужайках и степных участках среди тайги, по увалам предгорий.
Живет колониями. Питается луговыми травами. Местами вредит посе-
вам. Шкурки используют в меховой промышленности.
Подвиды фауны СССР:
а) С. u. eversmanni Brandt, 1841. (Тип с Алтая.) Размеры мелкие: длина тела
21—26 см. Окраска летнего меха яркая: брюхо рыжеватое, спина черноватая с бело-
ватой рябью. В окраске лба и темени много черноты. Зимний мех желтовато-серый
с белесой крапчатостью. Алтай, Саяны и их предгорья.
b) С. u. stramineus Obolensky, 1927. (Тип из окрестностей Ламан-Гегена, в Монго-
лии.) Размеры, как у предыдущей формы. Окраска летнего меха светлая, тусклая.
Брюхо тускло-желтоё. Спина менее интенсивно окрашена с более ясной белой рябью.
Лоб и темя ржаво-серые или желтовато-бурые. Зимний мех белесо-серый с желтова-
тым оттенком и ясной беловатой крапчатостью. Джунгарский Алатау, Монголия, Чуй-
ская степь на Алтае, южная часть Тувинской области.
с) С. u. undulatus Pallas, 1778. (Описан по экземплярам с р. Селенги.) Размеры,
как у предыдущих форм. Окраска летнего меха более светлая и тусклая, чем у алтай-
ской формы, но темнее и ярче, чем у особей из Монголии. Прибайкалье и Забайкалье
на восток до Читы.
d) С, u. intercedens Ognev, 1937. (Тип из Нерчинского района.) Размеры несколько
крупнее: длина тела 25—29 см. Окраска, как у предыдущего подвида. Восточное
Забайкалье.
е) С, u. menzbieri Ognev, 1937. (Тип из окрестностей г. Благовещенска-на-Амуре.)
Размеры еще крупнее: длина дела 30—33 см. Окраска, как у С. u. intercedens. Верхнее
течение Амура.
f) С. u. jacutensis Brandt, 1844. (Тип из окрестностей г. Якутска.) Размеры круп-
ные: длина тела 26—32 см. Окраска спины и брюха более тусклая, чем у сусликов из
Забайкалья. Южная и Восточная Якутия.
g) С. u. leucostictus Brandt, 1844. (Тип с р. Колымы.) Отличается от якутского
суслика более яркой, насыщенной окраской летнего меха и рыжеватой окраской лба
и темени. Крапины отчетливые. Бассейн р. Колымы, Чукотско-Анадырский край.
h) С. u. janensis Ognev, 1937. (Тип с р. Яны-) Форма, близкая к предыдущей, но
с еще более яркой окраской летнего меха. Бассейн р. Яны, Верхоянский хребет,
i) С. u. stejnegeri Allen, 1903. (Тип из окрестностей г. Петропавловска-Камчат-
ского.) Размеры средние. Окраска летнего меха относительно тусклая. Лоб и темя
рыжеватые. Камчатка.
РОД ТОНКОПАЛЫЕ СУСЛИКИ, GENUS SPERMORHILOPSIS Blasius, 1884
Своеобразные грызуны, приспособившиеся к обитанию среди пе-
сков пустыни. Тело вальковатое, несколько утолщенное кзади. Задние
ноги удлинены, их очень большая ступня несет длинные пальцы, бо-
ковые из которых снабжены пучками длинных волос, что еще более
увеличивает площадь опоры ноги. Более короткие передние конечно-
сти несут пальцы с очень длинными когтями. Хвост короткий с расче-
санными на бока волосами; с нижней стороны его конечная часть
черная со светлой каймой. Летний мех очень короткий, жесткий, приле-
гающий, зимний мягкий и рослый. Череп с широким межглазничным
промежутком и надглазничными отростками, отодвинутыми назад. Пер-
вый коренной зуб верхней челюсти примерно в 4 раза мельче второго.
Единственный вид,
25. Тонкопалый суслик. Spermophilopsis leptodactylus Lichtenstein, 1823
Табл. XXVII, 1—2. Карта 72
Обитает в песчаных пустынях Средней Азии и Южного Казах-
стана, Каракумы, Кызылкумы, Муюнкумы, Забалхашье и прилегающие
пески. Встречается в Северном Иране и Афганистане.
Живет в норах. Дневное животное. Питается растениями пустыни.
Дает в течение года один помет (в начале лета) из 3—7 детенышей.
Местами вредит пескозакрепляющим посадкам. В зимнюю спячку не
впадает.
265
Выделяют 3 слабо различающиеся формы:
a) S. 1. leptodactylus Lichtenstein, 1823. (Тип из Кара-Ата, к северо-западу от
Бухары.) Размеры относительно мелкие: длина тела 195—255 мм\ кондилобазальная
длина черепа 44—51 мм. Окраска зимнего меха светлая, охристая. Каракумы,
Кызылкумы.
b) S. 1. bactrianus Scully, 1887. (Тип из Северного Афганистана.) Размеры чуть
крупнее: длина тела 200—270 мм\ кондилобазальная длина черепа 48—54 мм. Окраска
зимнего меха более темная и тусклая. Бадхыз, Карабиль.
с) S. 1. heptopotamicus Heptner et Ismailov, 1952.-(Тип из Таукум, в Забалхашье.)
Размеры крупнее: длина тела 180—295 мм\ кондилобазальная длина черепа 48—55 мм.
Окраска зимнего меха темнее, чем у номинальной формы. Забалхашье, МуюнкуМ.
РОД БУРУНДУКИ. GENUS TAMIAS liliger, 1811
Небольшие зверьки с длинными хвостами. Волосы, покрывающие
хвост, расчесаны на две стороны. Окраска своеобразная: на спине
резко выделяется пять продольных черноватых полос.
В СССР — единственный вид (близкие формы обитают в Северной
Америке).
26. Бурундук. Tamias sibiricus Laxmann, 1769
Табл. XXVII, 3. Карта 72
В СССР распространен в лесных районах Дальнего Востока
(кроме Камчатки), Сибири, Северо-Восточного Казахстана и на севе-
ро-востоке Европейской части СССР (на запад до линии: Архан-
гельск— Верхняя Тойма — Вологда и на юг до Ветлуги, Казани, Уфы
и Южного Урала). Встречается .в Среднем, Северном и Северо-Восточ-
ном Китае, в Корее, на некоторых островах Японии и в Северной Мон-
голии.
Обитает в тайге, преимущественно в перелесках, по опушкам, в ку-
старниковых зарослях по долинам рек. Держится, главным образом,
на земле и среди валежника. Живет в неглубокой норе. Ведет днев-
ной образ жизни. Питается семенами различных древесных и травя-
нистых растений, ягодами, поедает также насекомых. На зиму делает
запасы, обычно из кедровых орешков. Зиму проводит в спячке. Раз-
множается весной, принося 4—10 детенышей. Местами вредит посевам.
Шкурки используют в меховом деле.
По окраске различают несколько подвидов:
а) Т. s. asiaticus Gmelin, 1788. (Описан с р. Северной Двины и р. Камы.) Окраска
светлая, серовато-желтая. Северо-восток Европейской части СССР, таежный Урал, За-
падная Сибирь (кроме Алтая).
b) Т. s. sibiricus Laxmann, 1769. (Тип из окрестностей г. Барнаула.) Окраска более
насыщенная, яркая. Южный Урал, лесостепь Западной Сибири, Алтай, Саяны,
Забайкалье.
с) Т. s. jacutensis Ognev, 1936. (Тип из окрестностей г. Якутска.) Окраска очень
бледная, белесо-желтоватая; темные полосы на спине узкие. Якутия.
d) Т. s. lineatus Siebold, 1826. (Тип с островов Иезо, Япония.) Окраска более яр-
кая, рыжеватая. Черные полосы на спине средней ширины. Амурский край, Шантар-
ские острова, Сахалин, Южно-Курильские острова.
е) Т. s. orientals Bonhote, 1899. (Тип с р. Сунгари, притока Уссури.) Окраска
наиболее насыщенная, яркая, рыжеватая. Черные полосы на спине широкие. Приморье.
РОД БЕЛКИ. GENUS SCIURUS Linnaeus, 1758
Среднего размера грызуны, ведущие древесный образ жизни.
Хвост длинный с расчесанными на бока длинными жесткими волосами.
Ушные раковины хорошо развиты и зимой обычно несут кисточку уд-
линенных волос. Защечные мешки отсутствуют. Череп с очень широ-
266
ким межглазничным промежутком; кости его значительно тоньше, чем
у черепов сусликов.
Представители рода широко распространены по лесам Европы,
Азии и Северной Америки. Из многих видов белок в СССР встреча-
ются 2 вида.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ БЕЛОК ФАУНЫ СССР
1(2). Мех на брюхе желтый или рыжеватый. Кисточек длинных
волос на ушах нет. На подошвах задних лап по 6 бугорков. В верхней
челюсти 'С каждой стороны обычно только по 4 коренных зуба ....
...................Кавказская белка. Sciurus anomalus. (Стр. 267.)
2(1). Мех на брюхе чисто белый. На концах ушей к зиме выра-
стают кисточки длинных волос. На подошвах задних лап по 4 бугорка.
В верхней челюсти с каждой стороны по 5 коренных зубов..........
...................Белка обыкновенная. Sciurus vulgaris. (Стр. 267.)
27. Кавказская белка. Sciurus anomalus Gmelin, 1778
Табл. XXVII, 4. Карта 73
Встречается в лесах Ирана, Восточной Турции, Сирии, Палестины,
Западного и Центрального Закавказья и южных склонов восточной
части Главного Кавказского хребта.
Обитает преимущественно в горных буковых лесах. Убежища —
в дуплах деревьев. Ведет дневной образ жизни. Питается буковыми
орешками, желудями, фруктами, ягодами и т. п. За год дает 2—3 по-
мета, по 3—7 детенышей в каждом. В силу малочисленности и невысо-
кой ценности шкурки промысловое значение ничтожно.
В Закавказье живет номинальная форма (S. a. anomalus Gmelin, 1778; тип из
Сабека близ Кутаиси).
28. Белка обыкновенная. Sciurus vulgaris Linnaeus, 1758
Табл. XXVII, 5—9. Карта 73
Вся лесная и лесостепная зона Европы и Азии. В СССР северная
граница распространения идет по Кольскому полуострову от г. Пе-
ченги, южнее Мурманска, севернее оз. Имандра к Терскому берегу и
далее, восточнее горла Белого моря, от устья р. Мезени по р. Суле,
северным притокам р. Усы к Северному Уралу. Реку Обь она пересе-
кает севернее г. Салехарда, р. Таз — в 250 км выше устья, р. Енисей —
у г. Дудинки, р. Хатангу — у ее устья, р. Лену — у с. Булуна, Яну и
Индигирку — у 70° с. ш., р. Колыму — у г. Нижне-Колымска, откуда
проходит к среднему течению р. Анадыря и далее к Пенжинской губе.
На восток белка распространена до Тихоокеанского побережья, встре-
чается в лесах Сахалина; в 20-х годах текущего столетия проникла и
заселила леса Камчатки. Южная граница распространения белки тя-
нется по Северо-Восточному и Сев-ерному Китаю, Северной Монголии
к Калбинскому Алтаю. Далее на запад она проходит по р. Иртышу
через окрестности г. Омска, г. Кургана и г. Магнитогорска к Южному
Уралу, а затем через Бузулук — Куйбышев — Саратов — Воронеж —
Купянск — Артемовск — Харьков — Полтаву — Днепропетровск — Ти-
располь к лесам Центральной Молдавии. Акклиматизирована в Крыму,
лесных районах Кавказа, горах Центрального Тянь-Шаня и в сосно-
вых борах Северного и Центрального Казахстана.
267
Лесной зверек, ведущий древесный образ жизни. Живет в гнездах
на деревьях или в дуплах. На зиму в спячку не впадает, но в метель
и сильные морозы подолгу не покидает гнезда. На севере приносит
1—2, в южных районах 2—3 помета в год по 4—10 (обычно 5—7)
бельчат в каждом. Питается главным образом семенами хвойных де-
ревьев, а также древесными почками, орехами, желудями, ягодами,
семенами некоторых трав, грибами и т. п.; поедает также насекомых,
яйца и птенцов мелких птиц. Численность 'белки по отдельным годам
сильно меняется, преимущественно в зависимости от урожая семян
хвойных. В некоторые годы белки мигрируют на большие расстояния.
Белка — основной объект пушного промысла лесных районов
СССР. Лишь в последние годы она уступила первое место в пушных
заготовках СССР соболю и ондатре.
Географическая изменчивость белки сильно осложнена резко выраженным сезон-
ным диморфизмом и очень широким полиморфизмом окраски меха в одних и тех же
местах и в одни и те же сезоны. По окраске волосяного покрова различают 5 сле-
дующих форм:
А. Краснохвост к а: окраска зимнего меха светло-серая, летнего ярко-рыжая;
хвост и кисточки на ушах рыжие; остевые волосы на хвосте у основания серые, на
конце рыжие.
В. Бурохвостка: окраска зимнего меха серая средней темноты или светло-
серая, летнего рыжая или буроватая; хвост и кисточки на ушах бурые; волосы на
хвосте у основания серые, посередине коричневые, на концах черные. Различают свет-
лых и темных бурохвосток.
С. Т е м н о х в о с т к а: окраска зимнего меха темно-серая, летнего бурая пли чер-
новатая; кисточки ушей черноватые; хвост черный, но с просвечивающими сероватыми
основаниями волос.
D. Чернохвостка: окраска зимнего меха темно-серая, летнего черная; хвост
и ушные кисточки черные.
Е. Серохвостка: окраска зимнего меха очень светлая, серебристо-серая;
хвост серый, седоватый; ушные кисточки обычно рыжие.
Описанные из СССР и смежных стран многочисленные подвиды белки могут
быть подразделены на 5 групп Е
I группа. Карпатские белки. В популяции в основном темно-бурохвостки и тем-
похвостки. Окраска летнего меха бурая или черно-бурая, зимнего темно серая. До-
вольно широкое белое поле на брюхе резко ограничено. Мех малопышный. К этой
группе относится только один подвид: S. v. carpathicus Pietruski, 1853.
II группа. Восточноевропейские белки. В популяциях бурохвостки и краснохво-
стки. Зимний мех серого цвета с развитыми в той или иной степени рыжеватыми
оттенками на спине, летний мех рыжий. Широкое белое поле на брюхе слабо огра-
ничено и тянется от нижней губы до основания хвоста. Мех малопышный.
a) S. v. ukrainicus Migulin, 1928. (Тип из Сумского района.) Размеры средние:
общая длина черепа 51—55 мм. Преимущественно краснохвостка. Окраска низкого и
редкого зимнего меха на спине рыжеватая или ржаво-коричневая, на боках рыжевато-
серая. Летний мех ярко-рыжий. Лесостепь Левобережной Украины, Курской и Воро-
нежской областей.
b) S. v. kessleri Migulin, 1928. (Тип из окрестностей г. Житомира.) Размеры, как
у предыдущей: общая длина черепа 51—55 мм. Мех низкий и редкий. Преимуще-
ственно бурохвостка. Окраска зимнего меха серая, обычно с сильной примесью на
спине рыжеватых и ржаво-коричневых оттенков. Летний мех ярко-рыжий. Лесостепь
Правобережной Украины, Северной Молдавии.
с) S. V. fedjuschini Ognev, 1935. (Туп из района г. Минска.) Размеры средние:
общая длина черепа 51—53 мм. Бурохвосток около 80%, краснохвосток около 20%.
Окраска зимнего меха бурохвосток грязновато-серая, средней темноты, обычно с сильно
рыжеватым налетом на спине. Окраска краснохвосток в зимнем мехе светло-серая
с рыжиной по хребту. Зимний мех малопышный. Летний мех рыжий. Украинское
Полесье, Белоруссия, западная часть Смоленской области, Литва, вероятно, Латвия.
d) S. v. varius Barret Hamilton, 1899. (Тнп из Северной Европы.) Размеры сред-
ние: общая длина черепа 49—54 мм. Бурохвосток около 70%, краснохвосток около
30%. Окраска довольно пышного зимнего меха относительно светлая (особенно у крас-
нохвосток), серая, обычно с рыжеватым оттенком на хребте. Летний мех рыжий.
1 Возможно, правильнее эти группы рассматривать как настоящие подвиды,
а большую часть подвидов рассматривать как natio.
268
Карельская АССР, Мурманская область, западные части Архангельской и Вологодской
областей. (Вероятно, неотличима от типичной формы.)
е) S. v. ognevi Migulin, 1928. (Тип из бывшего Боровского уезда Калужской гу-
бернии.) Размеры немного меньше, чем у предыдущих рас: общая длина черепа
47—53 мм. Бурохвосток около 75%, краснохвосток 25%. Окраска малопышного зимнего
меха белок обоих типов сходна с расцветкой белок предыдущего подвида, но тон
серого цвета более чистый с меньшей примесью рыжих оттенков' на спине (очень
близка к предыдущей). Летний волосяной покров рыжий.
Центральные районы Европейской части СССР.
III группа. Урало-западносибирские белки. В популяциях почти исключительно
бурохвостки, изредка краснохвостки и темнохвостки. Окраска довольно пышного зим-
него меха обычно светло-серая, без примеси рыжеватых оттенков или со слабым их
развитием. Летний мех рыжий. Широкая, нерезко ограниченная белая полоса на брюхе
тянется от нижней губы до корня хвоста.
f) S. v. formosovi Ognev, 1935. (Тип из ветлужских лесов.) Размеры средние:
общая длина черепа 49—53 мм. Зимний мех довольно чистого серого цвета средней
темноты, обычно со слабо выступающим коричневатым оттенком на спине. Летний
мех рыжий. Северо-восточные области Европейской части СССР.
g) S. v. baschkiricus Ognev, 1935. (Тип из Бузулукского бора.) Размеры довольно
крупные: общая длина черепа 50—55 мм. Окраска сравнительно пышного зимнего
меха светло-серая, иногда с охристым налетом по хребту. Летний мех светло-рыжий.
Заволжье, Средний и Южный Урал.
h) S. V. martensi Matschii, 1901; (Тип-с левого берега Нижнего Енисея.) Размеры
чуть крупнее предыдущей: общая длина черепа 51—56 мм. Окраска пышного зимнего
меха светлая, серебристо-серая, иногда с легким палевым оттенком на хребте. Лет-
ний мех светло-рыжий. Обская низменность.
IV группа. Белки-телеутки. Преимущественно серохвостки. Размеры крупные. Ок-
раска зимнего меха очень светлая, серебристо-серая, летнего светло-серая или ржаво-
сероватая. Широкая белая полоса на брюхе тянется от нижней губы до корня хвоста.
Ушные кисточки ярко-рыжие или черноватые (цвет их отличается от окраски хвоста).
i) S. v. exalbidus Pallas, 1778. (Описана из ленточных боров по рекам Оби и Ир-
тышу.) Размеры крупные: общая длина черепа 55—59 мм. Очень пышный зимний мех
светлого, белесо-серого цвета, иногда с палевым налетом на спине. Летний мех светло-
рыжий. Длинный хвост серого, желтовато-серого или рыжеватого цвета. Ушные ки-
сточки красные, лапки рыжеватые. Населяет ленточные боры по р. Иртышу (от
г. Усть-Каменогорска до г. Павлодара) и по р. Оби (от г. Горно-Алтайска до
г. Камня).
j) S. v. golzmajeri Smirnov, 1960. (Тип из Озернинского бора Звериноголовского
района Курганской области.) Размеры немного мельче, чем у предыдущего подвида.
Окраска зимнего меха светло-серебристая, летнего рыжевато-серая. Хвост серый или
ржаво-серый. Ушные кисточки серовато-черные. Лапки серые. Боры Курганской и юж-
ной части Тюменской областей.
V группа. Восточносибирские белки. В популяциях чернохвостки, темнохвостки,
бурохвостки и краснохвостки (у некоторых рас отдельные из этих типов выпадают).
Преобладают особи с темной окраской зимнего меха. Летний мех черный, бурый или
рыжий. Узкая, резко ограниченная белая полоса на брюхе тянется от горла до
передней границы пахов.
k) S. v. altaicus Serebrennikov, 1928. (Тип с р. Кок-су, на Алтае.) Размеры сред-
ние: общая длина черепа 49—53 мм. Преобладают темнохвостки и бурохвостки; есть
краснохвостки. Зимний мех темнохвосток темно-серый, обычно с буроватым оттенком,
бурохвосток серый, средней темноты, тоже с буроватым налетом, краснохвосток свет-
ло-серый с рыжиной. Летний мех от ярко-рыжего до черно-бурого. Горы и предгорья
Алтая и Саян.
1) S. v. kalbinensis Selevin, 1934. (Тип из Аюдинского бора.) Отличается от ал-
тайской белки более светлой окраской и большим числом особей с рыжей окраской
хвоста. Боры Калбинского Алтая.
m) S. v. jenissejensis Ognev, 1935. (Тип с р. Нижней Тунгуски.) Размеры средние:
общая длина черепа 51—54’лш. Больше всего бурохвосток, много темнохвосток, мало
краснохвосток. Окраска пышного зимнего меха, как у алтайской белки, но более
чистого голубоватого тона. Многие белки с рыжиной на хребте. Правобережье бас-
сейна р. Енисея.
n) S. v. jacutensis Ognev, 1929. (Тип из окрестностей г. Якутска.) Размеры, как
у енисейской белки. Около половины популяции составляют черно- и темнохвостки,
много темно-, бурохвосток; краснохвосток около 15%. Окраска очень пышного и шелко-
вистого зимнего меха отличается красивым чистым голубоватым оттенком. Рыжий
налет на спине обычно не выражен. Летний мех от ржаво-бурого до черного. Яку-
тия, Анадырский край.
о) S. v. fusconigricans Dvigubski, 1804. (Описан из Баргузина.) Размеры отно-
сительно крупные: общая длина черепа 52—56 мм. Чернохвостки, темнохвостки
269
и бурохвостки встречаются примерно одинаково часто; краснохвосток около 2%.
Окраска пышного, но грубоватого меха обычно темно-серая, с буроватым оттенком.
Летний мех от ржаво-бурого до черного. Забайкалье.
р) S. v. rupestris Thomas, 1907. (Тип из окрестностей г. Корсакова, на Сахалине.)
Размеры средние: общая длина черепа 51—55 мм. Чернохвостки, темнохвостки и буро-
хвостки встречаются примерно в равном количестве; краснохвосток немного. В ок-
раске пышного, но грубоватого зимнего меха ясно проступают буроватые оттенки.
Летний мех от ржаво-бурого до черного. Остров Сахалин, низовья Амура, Охотское
побережье.
q) S. v. mantchuricus Thomas, 1909. (Тип с Большого Хингана.) Размеры круп-
ные: общая длина черепа 53—58 мм. Окраска, как у предыдущего подвида. Амуро-
Уссурийский край, Северо-Восточный Китай.
СЕМЕЙСТВО ЛЕТЯЖЬИ. FAMILIA PTEROMYIDAE
Похожие на белок грызуны, но с покрытой мехом складкой кожи,
натянутой между передними и задними конечностями.
Населяют леса Южной и Восточной Азии, таежную зону Евразии
и Северной Америки.
Систематики Западной Европы и Америки (Эллерман и Моррисон-Скотт, 1951;
Холл и Келсон, 1959, и др.) относят летяг к семейству беличьих (Sciuridae).
Из 8 родов летяг в фауне СССР — 1 род.
РОД ЛЕТЯГИ. GENUS PTEROMYS Cuvier, 1800
Зверьки размерами несколько меньше белки и с менее длинным
хвостом. Зубная формула, как у беличьих.
Европа, Азия (к югу до Центрального Китая), остров Хоккайдо,
в Японии, и Северная Америка, к северу до южных границ лесотундры.
Из трех видов этого рода в фауне СССР — 1 вид.
29. Летяга. Pteromys volans Linnaeus, 1758
Табл. XXVIII, 1—2. Карта 73
Леса Северной Европы, Сибири, Дальнего Востока, Северной Мон-
голии, Центрального и Северо-Восточного Китая, остров Хоккайдо
в Японии. В СССР водится почти по всей лесной полосе, но и повсе-
местно довольно малочисленна. Северная граница ареала тянется по
Северной Карелии, по берегу Белого моря до г. Мезени и далее на во-
сток по северному пределу леса до Пенжинской губы. На востоке рас-
пространена до побережья Тихого океана, но на Камчатке отсутствует.
Южная граница тянется по Центральному Китаю, а в СССР — через
Калбинский Алтай, по сосновым борам правого берега р. Иртыша до
Омска, оттуда через Ялуторовск—Курган — Челябинск а Южному
Уралу и далее по рекам Белой и Каме до Казани, по р. Волге до
г. Горького, по р. Оке, до- г. Коломны, через Смоленскую область и
центральные части Белоруссии к западной границе страны.
Держится преимущественно в глухих лесах. Особенно охотно се-
лится в лесах по долинам рек. Живет обычно в дуплах деревьев.
Ведет ночной образ жизни. Планируя при помощи складки кожи на
боках тела, может «перелетать» с дерева на дерево на расстояние до
40—50 м. Приносит, по-видимому, один помет из 2—5 детенышей; воз-
можно, что иногда размножается два раза в год — весной и летом.
Питается, главным образом, семенами деревьев и трав, ягодами, гри-
бами, а зимой — почками хвойных деревьев. Промысловое значение
в связи с малочисленностью зверька и малой ценностью шкурки
невелико.
270
С территории СССР описано большое число форм, различия между которыми
нерезки и заметны лишь при сличении больших серий:
а) Р. v. volans Linnaeus, 1758. (Описан из Швеции.) Зимний мех спины пепельно-
серый с легким палевым оттенком, летний черно-серый со светлой серой рябыо. Хвост
темный. Северная Европа, север Западной Сибири.
b) Р. v. ognevi Stroganov, 1936. (Тип с оз. Пенс, в Калининской области.) Зим-
ний и летний мех на спине с ясно выступающими ржаво-палевыми оттенками. Хвост
палево-ржавый. Центральный район Европейской части СССР.
с) Р. v. gubari Ognev, 1934. (Тип из бывшего Бийского уезда.) Зимний мех спины
светлый, серебристо-серый, обычно без палевых оттенков. Летний покров бледного,
грязно-серого цвета с палевыми оттенками. Хвост светлый. Южная часть Западной
Сибири.
d) Р. v. betulinus Serebrennikov, 4929. (Тип из бывшего Павлодарского уезда.)
Зимний мех спины светлый, серебристо-серый с сильной примесью желтовато-палевых
оттенков. В летнем грязно-сером мехе резко выражены те же тона. Хвост светлый,
желтоватый. Сосновые боры Предалтая.
е) Р. v. turovi Ognev, 1929. (Тип с полуострова Большие Коты, на Байкале.) Зим-
ний мех спины темный, грязно-серый с палевым оттенком и черноватой рябью. Лет-
ний волосяной покров ржаво-серый, весьма темный. Хвост желтовато-серый. Алтай,
Саяны, Забайкалье, Верхний Амур.
f) Р. v. incanus Miller, 1918. (Тип из окрестностей Верхне-Колымска.) Окраска
очень пышного зимнего меха на спине светлая, серебристо-палево-серая. Летний мех
тускло-серый, со слаборазвитыми палевыми тонами. Хвост с палевым оттенком. Во-
сточная Сибирь, Якутия.
g) Р. vx anadyrensis Ognev, 1940. (Тип из окрестностей с. Ерополь, на Анадыре.)
Зимний мех голубовато-серебристо-серый без палевых оттенков. Хвост сероватый с па-
левым налетом. Анадырский край.
h) Р. v. athene Thomas, 1907. (Тип из окрестностей г. Корсакова, на Сахалине.)
Зимний мех спины довольно светлый, серебристо-серый с ясной примесью палевых
тонов. Летний волосяной покров с сильным развитием палево-ржавых оттенков. Хвост
палево-ржавый. Сахалин, Нижний Амур.
i) Р. v. arsenjevi Ognev, 1934. (Тип с р. Колумбе.) Зимний мех спины грязно-пале-
во-серый с интенсивным желтовато-коричневым оттенком. Летний мех серо-ржаво-
палевый. Хвост желтоватый или рыжеватый. Уссурийский край.
Реальность отличий некоторых из этих форм летяг требует проверки.
СЕМЕЙСТВО СОНЕВЫЕ. FAMILIA MUSCARDINIDAE
Мелкие или средней величины древесные или реже наземные гры-
зуны. По внешнему облику некоторые из них напоминают белку, дру-
гие— мышеобразных. Коренных зубов—; жевательная поверхность
их несет невысокие поперечные валики. В отличие от других грызунов,
у сонь отсутствует слепая кишка.
Из 8 родов в фауне СССР — 5 родов (по одному виду в каждом).
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ СОНЬ ФАУНЫ СССР
1(6). Размеры относительно крупные: ступня более 18 мм; конди-
лобазальная длина черепа взрослых более 23 мм ........ 2
2(3). На боках головы черных полос нет. Длина верхнего ряда
зубов более 6 мм........................Сони-полчки. Glis. (Стр. 272.)
3(2). На боках головы от носа через глаз к уху проходит черная
полоса. Длина верхнего ряда зубов менее 6 мм...........................4
4(5). Хвост на всем протяжении покрыт длинными волосами, «рас-
чесанными» на две стороны. Длина ступни (без когтей) 18—24 мм.
Кондилобазальная длина черепа 23—28 мм ...........
............. Лесные сони. Dryomys. (Стр. 273.)
5(4). Хвост в основной половине покрыт короткими волосами, по-
степенно удлиняющимися в концевой части. Длина ступни (без когтей)
25—31 мм. Кондилобазальная длина черепа взрослых 30—36 мм . . .
............. Садовые сони. Eliomys. (Стр. 274.)
271
Рис. 98. Хвосты различных видов
сонь:
А — полчок (Glis glis); Б — садовая соня
(Eliomys quercinus); В — лесная соня
(Dryomys nitedula); Г — орешниковая соня
(Muscardinus avellanarius); Д — мыше-
видная соня (Myomimus personatus).
6(1). Размеры мелкие: ступня менее 17 мм\ кондилобазальная
длина черепа менее 22 мм...................................... 7
7(8). Хвост густо покрыт рыжеватыми волосами, удлиненными по
бокам. Длина ступни 15—17 мм. Верхний предкоренной зуб раза в 4
короче смежного с ним коренного зуба.............................
.........................Орешниковые сони. Muscardinus. (Стр. 274.)
8(7). Хвост, как у мышей, покрыт кольцевыми чешуйками, между
которыми расположены редкие белые волоски. Длина ступни менее
15 мм. Верхний предкоренной зуб лишь раза в 2 короче смежного
с ним коренного зуба . . Мышевидные сони. Myomimus. (Стр. 274.)
РОД СОНИ-ПОЛЧКИ. GENUS GLIS Linnaeus, 1762
Всего один вид.
30. Соня-полчок. Glis glis. Linnaeus, 1766
Табл. XXVIII, 3—4. Карта 74
Распространен в Европе и Передней Азии. В СССР обитает по лес-
ным ландшафтам Кавказа, Закавказья, западных и центральных
районов Европейской части страны, к северу до Прибалтики, Тульской
и южных районов Горьковской области, на восток до р. Волги, на юг
до линии: Саратов — Воронеж — Чернигов — Киев — Кишинев. Вероят-
но, встречается в горах Копет-Дага.
272
Водится в лесах (главным образом, лиственных) и в садах. Жи-
вет в дуплах и гнездах. На зиму впадает в длительную спячку. Ведет
ночной образ жизни. Питается плодами, орехами, семенами, иногда
поедает насекомых и мелких птенцов. Дает 1 помет в году (в июне —
июле) из 3—6 детенышей. Причиняет вред садам и виноградникам.
Промышляется ради шкурки.
Подвиды:
a) G. g. glis Linnaeus, 1766. (Описан из Германии.) Размеры относительно мелкие.
Живет в лесах западных и центральных областей.
b) G. g. orientalis Nehring, 1903. (Описан из гор Алатаг, в Малой Азии.) Размеры
немного крупнее. В окраске спины сильнее выступают бурые оттенки. Кавказ и За-
кавказье, кроме Талыша и Ленкорани.
с) G. g. persicus Erxleben, 1777. (Описан по экземпляру из Ирана.) Отличается
крупными размерами, пышным хвостом, утолщенным носовым отделом черепа. Встре-
чается на Талыше и в Ленкорани.
РОД ЛЕСНЫЕ СОНИ. GENUS DRYOMYS Thomas, 1906
Только один вид.
31. Лесная соня. Dryomys nitedula Pallas, 1779
Табл. XXVIII, 5—7. Карта 75
Европа, Передняя и Средняя Азия. В СССР распространена в за-
падных, центральных и южных районах Европейской части страны
(к северу до Литвы, Калининской, Рязанской, Горьковской областей и
Татарии), на Кавказе, в Закавказье, горных лесах Средней Азии, Вос-
точного Казахстана и Южного Алтая.
Селится в лиственных и смешанных лесах, садах и зарослях ку-
старника. Поднимается высоко в горы. Живет обычно в построенных
самим зверьком гнездах, реже в дуплах и щелях в скалах. Ночной
зверек. Зиму проводит в спячке. Ловко лазает по деревьям и кустар-
никам, но нередко разыскивает пищу и на земле. Приносит ежегодно
1 (или возможно 2) помет из 3—6 детенышей. Питается семенами,
плодами, орехами, желудями, насекомыми. Местами вредит садовым
культурам.
Описано много подвидов, слабо различающихся по тонам окраски меха; реаль-
ность многих из них вызывает сомнение.
a) D. n. nitedula Pallas, 1778. (Описан из дубовых лесов бывшей Казанской губер-
нии.) Спина взрослых особей интенсивного серовато-желто-буроватого цвета с ясной
примесью коричневато-рыжеватых оттенков. Молодые сверху серые с легким бурова-
тым налетом. Хвост серый, почти без палевых тонов. Среднее и Верхнее Поволжье.
b) D. n. obolenskii Ognev et Worobiev, 1923. (Тип из Хреновскогс бора Воронеж-
ской области.) Окраска взрослых несколько более яркая с более выступающими жел-
товатыми оттенками. У молодых желтые оттенки на спине выступают более ясно.
Северная Украина, Курская, Воронежская, Орловская, Рязанская, Тамбовская обла-
сти. (D. n. tanaiticus Ognev et Turov, 1935, описанная по экземпляру из Тарасов-
ского района, на Дону, правильнее относить к синонимам этой формы.)
с) D. n. caucasicus Ognev et Turov, 1935. (Тип из окрестностей ст. Тарской, на
Северном Кавказе.) Окраска спины взрослых интенсивно серая с коричневым оттен-
ком, хвост серый с легким палевым оттенком. Центральное Предкавказье.
d) D. n. dagestanicus Ognev et Turov, 1935. (Тип из окрестностей г. Хасавюрта.)
Спина довольно светлая, серовато-желтая с охристым оттенком, бока охристые. Хвост
бледно окрашенный, палево-серый. Восточное Предкавказье.
е) D. n. ognevi Heptner et Formozov, 1928. (Тип из окрестностей с. Ахты, на р. Са-
муре.) Окраска спины очень тусклая, оливково-серая с легким палевым оттенком.
Хвост бледно-серый. Южный Дагестан.
f) D. n. tichomirovi Satunin, 1920. (Тип из окрестностей г. Тбилиси.) В окраске
спины сильно развиты охристые оттенки, особенно резко выступающие на боках тела.
Хвост светлый, палево-серый с палевым оттенком. Закавказье.
18 Н. А. Бобринский и др.
273
g) D. n. bilkjewiczi Ognev et Heptner, 1928. (Тип из окрестностей г. Гермаба,
в Южной Туркмении.) Окраска спины очень светлая, палево-серая. Хвост грязно-
серый с палевым оттенком. Копет-Даг.
h) D. n. pallidus Ognev et Turov, 1935. (Тип из долины р. Боскургай.) Отличается
от копет-дагского подвида большим развитием светлого, охристо-буроватого оттенка.
Узбекистан.
i) D. n. saxatilis Rosanov, 1932. (Тип из окрестностей поста Даршар, на Памире.)
Наиболее светлая форма. Западный Памир.
j) D. n. angelus Thomas, 1906. (Тип с Восточного Тянь-Шаня.) Окраска довольно
интенсивная с выступающими на спине красновато-буроватым оттенком. Хвост
светло-палево-серый. Горы Киргизии и Восточного Казахстана.
РОД САДОВЫЕ СОНИ. GENUS ELIOMYS Wagner, 1840
Только один вид.
32. Садовая соня. Eliomys quercinus. Linnaeus 1766
Табл. XXVIII, 8. Карта 76
Широко распространена в Западной Европе. В СССР обитает в
западных и центральных районах Европейской части страны к северу
до Южной Карелии, Калининской, Горьковской и Пермской областей,
на восток до Среднего и Южного Урала, на юг до Оренбургской, Куй-
бышевской, Ульяновской, Орловской областей, Украины и Молдавии
включительно.
Держится в садах, лиственных и смешанных лесах. Селится чаще
всего в дуплах и щелях, но иногда строит гнезда. Зимой спит. Пи-
тается, главным образом, желудями, орехами, ягодами, плодами,
а также насекомыми. Размножается, по-видимому, один раз в год,
принося по 3—6 детенышей. Местами приносит вред садам.
В СССР встречается особая форма: Е. q. superans Ognev et Stroganov, 1936. (Тип
из окрестностей г. Осташкова.) Отличается от типичной (описанной из Германии)
более крупными размерами.
РОД ОРЕШНИКОВЫЕ СОНИ. GENUS MUSCARDINUS Каир, 1829
Только один вид.
33. Орешниковая соня. Muscardinus avellanarius Linnaeus, 1758
Табл. XXVIII, 9. Карта 77
Европа и Малая Азия (на Кавказе отсутствует).
В СССР распространена в зоне широколиственных лесов и полосы
лесостепи на север до Северной Белоруссии, Калининской, Ивановской,
Горьковской областей и Татарии, на восток до Предуралья, на юг до
Куйбышевской, Харьковской, Полтавской, Днепропетровской и Одес-
ской областей.
Держится в подлеске лиственных и смешанных лесов, в переле-
сках, кустарнике, садах. Ведет ночной образ жизни. Живет обычно
в искусно построенных гнездах и в дуплах. Питается орехами, желу-
дями, семенами, ягодами; поедает насекомых. Дает ежегодно 1 или 2
помета по 3—5 детенышей.
В СССР встречается номинальная форма: М. a. avellanarius Linnaeus, 1758, опи-
санная из Средней Швеции.
РОД МЫШЕВИДНЫЕ СОНИ. GENUS MYOMIMUS Ognev, 1924
Только один вид.
274
34. Мышевидная соня. Myomimus personatus Ognev, 1924
Табл. XXVIII, 10. Карта 77
В СССР добыта только в районе поста Кайне-Кассыр и крепости
Кара-Кала в Туркмении. В последние годы обнаружена в Болгарии.
Вероятно, распространена в Иране и Турции. Возможно ее нахождение
в Закавказье.
Образ жизни наземный. Биология не изучена.
СЕМЕЙСТВО СЕЛЕВИНИЕВЫЕ. FAMILIA SELEVINIIDAE
Мелкие мышевидные грызуны с длинным, тонким, густо покры-
тым короткими волосками хвостом и короткими лапками. Особенности
строения черепа и отсутствие слепой кишки сближают их с семейством
сонь, но строение зубов в высокой степени своеобразно. Коренных зу-
бов обычно по 3 с каждой стороны как верхней, так и нижней челю-
стей, но иногда их бывает 4. Коренные зубы очень малы; их жева-
тельная поверхность чашеобразно вогнута. Огромные резцы верхней
челюсти несут спереди глубокие бороздки, отчего режущий край их
зигзагообразно изогнут.
Единственный род.
РОД СЕЛЕВИНИИ. GENUS SELEVINIA Belosludov et Bashanov, 1938
Только один вид.
35. Селевиния. Selevinia betpakdalensis Belosludov et Bashanov, 1938
Табл. XXVIII, 11. Карта 77
Обитает в пустыне Бет-Пак-Дала, в Северном Прибалхашье и
в котловине оз. Алакуль. Держится в глинистой пустыне, в зарослях
солянки-боялыча и белой полыни. Ведет сумеречный образ жизни. Пи-
тается преимущественно насекомыми. На зиму впадает в спячку. Раз-
множение, не изучено. Подвиды не описаны.
СЕМЕЙСТВО ТУШКАНЧИКОВЫЕ. FAMILIA DIPODIDAE
Своеобразные грызуны, приспособленные к передвижению по
земле большими прыжками. Передние конечности укорочены и слабы;
они служат для удержания пищй и для рытья нор. Задние ноги, на-
против, очень велики: их длина в 4 и более раз превышает длину
передних конечностей. Эти ноги несут 3 или 5 пальцев; в последнем
случае боковые пальцы заметно укорочены и концы их когтей не дости-
гают основания средних пальцев. Кости плюсны обычно слиты в еди-
ную кость — «цевку». Хвост длинный (почти всегда длиннее тела);
у большинства видов тушканчиков на конце хвоста имеется плоская
кисточка волос — «знамя».
Скуловые кости черепа с двумя ветвями: горизонтальной и верти-
кальной; последняя отделяет глазницу от большого предглазничного
TZ .43
отверстия. Коренных зубов —: или — .
3 3
Распространены в пустынях, полупустынях и степях Северной Аф-
рики, Европы и Азии.
В СССР встречается 3 подсемейства тушканчиков.
18*
275
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ
семейства тушканчиков ФАУНЫ СССР
1(4). Длина тела более 8 см. Внутренние края ушных раковин при
их основании не срослись. Сосцевидные кости черепа умеренной вели-
чины и не видны или почти не видны при рассматривании черепа
сверху. Межтеменная кость хорошо развита.....................2
2(3). Задние конечности пятипалые. Резцы верхних челюстей
сильно наклонены вперед .......................................
. . Подсемейство пятипалых тушканчиков. Allactaginae. (Стр. 277.)
Рис. 99. Черепа тушканчиков (сверху):
А — малого тушканчика (Allactaga elater); Б — мохноно-
гого тушканчика (Dipus sagitta); В — тушканчика Лихтен-
штейна (Jaculus lichtensteini); Г — карликового тушкан-
чика (Cardiocranius paradoxus).
3(2). Задние конечности трехпалые. Резцы верхних челюстей на-
правлены вершинами прямо вниз......................................
. . . Подсемейство трехпалых тушканчиков. Dipodinae. (Стр. 283.)
4(1). Длина тела менее 8 см. Внутренние края ушных раковин
при их основании срастаются в трубочку. Сосцевидные кости черепа
огромного размера: они почти сходятся своими краями на крыше че-
репа. Межтеменная кость слабо развита или отсутствует..............
. Подсемейство карликовых тушканчиков. Cardiocraniinae. (Стр. 285.)
276
Подсемейство ПЯТИПАЛЫХ ТУШКАНЧИКОВ.
Subfamilia ALLACTAGINAE
В СССР представлено тремя родами.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ПОДСЕМЕЙСТВА ПЯТИПАЛЫХ
ТУШКАНЧИКОВ ФАУНЫ СССР
1(2). Уши длинные: отогнутые вперед, они достигают конца морды
или заходят за него. На конце хвоста имеется хорошо развитое «зна-
мя»— кисточка «расчесанных» на бока волос. В верхней челюсти с каж-
дой стороны по 4 коренных зуба (передний из них иногда очень мал) .
....................Пятипалые тушканчики. Allactaga. (Стр. 277.)
2(1). Уши более короткие: отогнутые впе-
ред, они своими вершинами не достигают конца
морды. «Знамя» на конце хвоста слабо выра-
жено или отсутствует. В верхней челюсти с каж-
дой стороны только по 3 коренных зуба . . 3
3(4). Хвост длинный, слабо утолщенный
в средней части или тонкий. Длина его (без
концевых волос) 12—15 см. На концах хвоста
имеется слаборазвитое «знамя». Длина задней
ступни (без когтей) 45—50 мм. Первый (внут-
ренний) палец задней ноги немного длиннее
пятого. Межглазничный промежуток черепа
примерно в 3 раза меньше наибольшей длины
его............................................
Земляные зайчики. Alactagulus. (Стр. 281.)
4(3). Хвост более короткий (не более 11 см),
утолщенный (за счет отложения под кожей слоя
жира). «Знамя» на конце хвоста отсутствует.
Длина задней ступни (без когтей) менее 45 мм.
Первый палец задней ноги короче пятого. Меж-
глазничный промежуток черепа примерно в
4 раза меньше его наибольшей длины . . . .
...................Толстохвостые тушканчики.
Pygerethmus. (Стр. 281.)
Рис. 100. Хвосты различ-
ных тушканчиков:
А — малого тушканчика (Al-
lactaga elater); Б — толсто-
хвостого тушканчика (Pyge-
rethmus platyurus).
РОД ПЯТИПАЛЫЕ ТУШКАНЧИКИ. GENUS ALLACTAGA Cuvier, 1836
Крупные и среднего размера (реже мелкие) тушканчики с пятипа-
лыми задними ногами, длинным хвостом со «знаменем» на конце ’и
длинными ушами. Стержень хвоста на всем протяжении тонкий, без
заметных подкожных жировых отложений. Коренных зубов с каждой
стороны верхней челюсти по 4.
Обитатели степей, полупустынь и глинистых пустынь; сыпучих пес-
сков избегают. Живут в постоянных и временных норах. Ночные жи-
вотные. На зиму впадают в спячку. Размножаются весной.
В СССР встречается 6 видов этого рода.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА ПЯТИПАЛЫХ
ТУШКАНЧИКОВ ФАУНЫ СССР
1(10). На нижней стороне трех средних пальцев щетки длинных
волос нет. «Знамя» на хвосте либо двухцветное (черное у основания
и белое на конце), либо трехцветное: белое у основания и на конце
277
и черное в средней части. Вершины сосцевидных костей не сходятся
друг с другом..................................................2
2(9). Размеры крупные: длина взрослых животных более 12 см;
длина ступни более 6 см; кондилобазальная длина черепа более 30 мм
.............................................................. 3
3(7). На стержне хвоста перед черной частью «знамени» кольца
светлых волос нет. Верхний предкоренной зуб в 2—3 раза меньше по-
следнего коренного ........................................... 4
4(8). Ухо длинное: отогнутое вперед, оно своей вершиной заходит
за конец морды. Размеры крупные: длина ступни взрослых особей
более 7 см; кондилобазальная длина черепа более 32 мм .... 5
5(6). Основная половина «знамени» на хвосте снизу сплошь чер-
ная. Длина ступни взрослых обычно более 85 мм. Кондилобазальная
длина черепа более 41 мм ........................................
.................Большой тушканчик. Allactaga major. (Стр. 279.)
6(5). Через черную основную часть «знамени» по нижней стороне
стержня хвоста проходит узкая беловатая полоска. Длина ступни зад-
них ног обычно менее 85 мм. Кондилобазальная длина черепа менее
41 мм . . . Тушканчик Северцова. Allactaga severtzovi. (Стр. 279.)
7(3). Впереди от черной части «знамени» стержень хвоста покрыт
на протяжении 3—4 см беловатым мехом, образующим светлое кольцо.
Верхний предкоренной зуб почти такой же величины, как задний ко-
ренной ..........Тушканчик-прыгун. Allactaga sibirica. (Стр. 279.)
8(4). Ухо более короткое: отогнутое вперед, оно своей вершиной
едва достигает конца морды. Размеры мельче: длина ступни менее
7 см; кондилобазальная длина черепа менее 32 мм .................
...............Горный тушканчик. Allactaga williamsi. (Стр. 280.)
9(2). Размеры мелкие: длина тела менее 12 см; длина ступни
менее 6 см; кондилобазальная длина черепа 23—28 мм...............
.......................Малый тушканчик. Allactaga elater. (Стр. 280.)
10(1). Нижняя поверхность средних пальцев покрыта густой щет-
кой длинных волос. «Знамя» все дымчато-серое. Вершины сосцевидных
костей почти сходятся ...........................................
.............Тушканчик Бобринского. Allactaga bobrinskii. (Стр. 281.)
Рис. 101. Концевые части хвостов пятипалых тушканчиков:
А —. большого тушканчика (Allactaga major); Б — тушканчика Северцова (A. severtzovi); В —
тушканчика-прыгуна (A. sibirica); Г — тушканчика Бобринского (A. bobrinskii).
278
36. Большой тушканчик. Allactaga major Kerr, 1792
Табл. XXIX, /. Карта 78
Степи и пустыни Европейской части СССР, Западной Сибири и
Казахстана. Северная граница ареала тянется от Киева примерно че-
рез Чернигов—Брянск—Калугу по р. Оке до г. Горького, по Волге
до Казани, по нижней Каме и по Белой к Южному Уралу. Восточнее
она поднимается к северу почти до Свердловска и тянется по южным
районам Тюменской и по Омской области к Новосибирску, где сво-
рачивает на юг к Южному Алтаю. На восток этот вид в СССР рас-
пространен до Алтая и Джунгарского Алатау; водится в Джунгарии.
Южная граница ареала идет по северным предгорьям Тянь-Шаня, по
р. Сыр-Дарье, по юго-восточному берегу Арала, по Устюрту, побе-
режью Каспийского моря, подножию Кавказского хребта, берегам
Азовского моря, северным предгорьям Крымских гор, северному берегу
Черного моря.
Селится в пустынных, полупустынных и степных ландшафтах, на
выгонах, вдоль грунтовых дорог. Живет в норах, которые чаще всего
роет по обочинам дорог. Различают глубокие постоянные и небольшие
временные норы. Питается наземными и подземными частями травя-
нистых растений. Дает 1 (а возможно и 2) помет в год из 1—4 дете-
нышей.
Причиняет вред бахчевым культурам, выгрызая из арбузов и дынь
семена. Шкурки большого тушканчика используют как пушное сырье.
Описан ряд весьма слабо различающихся форм:
a) A. m. jaculus Pallas, 1778. (Описан из крымских степей.) Окраска спины туск-
лая, серовато-желтая; верх головы желтонпепельный. Степи юга Европейской части
страны, кроме Предкавказья.
b) A. m. fuscus Ognev, 1924. (Тип из Кизлярских степей.) Окраска спины более
насыщенная, рыжевато-охристая; в окраске головы выступают рыжеватые оттенки.
Предкавказье.
с) A. m. major Kerr, 1792. Описан из степей между Каспием и р. Иртышем. Спина
нежно-охристая с розоватым оттенком. Центральный Казахстан. (Sin.: A. m. intermedia
Ognev, 1948).
d) A. m. vexillarius Eversmann, 1840. (Описан с Устюрта.) Окраска спины бледная,
песчано-желтая. Верх головы светлый, пепельно-серый. Пустыни Казахстана, от
Устюрта до Прибалхашья включительно.
е) A. m. spiculum Lichtenstein, 1824. (Описан из окрестностей г. Барнаула.)
Окраска спины и верха головы тусклая, сероватая со слабым развитием желтых от-
тенков. Северный Казахстан и степи Западной Сибири. (К этой же форме относится
А. ш. decumanus Lichtenstein, 1825, описанный из окрестностей Златоуста.)
37. Тушканчик Северцова. Allactaga severtzovi Vinogradov, 1925
Табл. XXIX, 2. Карта 78
Равнинные пустыни Средней Азии и Южного Казахстана к северу
до Устюрта, Приаральских Каракумов, низовьев р. Чу, оз. Балхаш.
Обитает, главным образом, в глинистой пустыне, но попадается и
на участках щебенистой и солончаковой пустынь. По образу жизни, ко-
торый изучен еще очень слабо, по-видимому, сходен с большим туш-
канчиком.
Подвиды не описаны.
38. Тушканчик-прыгун. Allactaga sibirica Forster, 1778
Табл. XXIX, 5. Карта 79
От восточного берега Каспийского моря и низовьев р. Урала, че-
рез Казахстан и северные области Центральной Азии до Северного
и Северо-Восточного Китая. В СССР найден в Южном Казахстане,
279
в горах Восточной Киргизии, в Чуйской степи на Алтае, в степях За-
байкалья.
Встречается в сухих степях, среди глинистой и щебенистой пу-
стыни, на солончаках, на участках горной степи. Питается наземными
и подземными частями пустынных растений и насекомыми. Дает 1 по-
мет в год (в начале лета) из 2—6 детенышей.
Подвиды фауны СССР:
a) A. s. sibirica Forster, 4778. (Описан из Юго-Восточного Забайкалья.) Крупная
форма: общая длина черепа 39—42 мм. Окраска темная, спина охристо-бурая с олив-
ковым оттенком. Южное Забайкалье.
b) A. s. saltator Eversmann, 1848. (Тип с верховьев р. Чуи, на Алтае.) Размеры
мельче: общая длина черепа 33—37 мм. Окраска светлая: спина сероватая с палевым
оттенком. Долина р. Чуи, на Алтае, Северо-Восточная Монголия.
с) A. s. suschkini Satunin, 1900. (Тип из казахских степей.) Размеры средние:
общая длина черепа 36—39 мм. Окраска средней темноты: спина буровато-серая.
Равнинный Казахстан. К этой же форме относится: A. s. riickbeili Thomas, 1914, опи-
санный из окрестностей г. Панфилова.
d) A. s. altorum Ognev, 1946. (Тип с р. Арпы, в Киргизии.) Размеры средние: об-
щая длина черепа 36—40 мм. Уши и хвост длиннее, чем у других форм. Спина серо-
ватая с палевым оттенком. Горные степи сыртов Киргизии (Центрального Тань-Шаня).
е) A. s. dementievi Toktosunov, 1955. (Тип из западной части котловины оз. Иссык-
Куль.) Мелкая форма. Мех пышный. Спина со слабым развитием рыжих оттенков.
Бока серые. Котловина о<з. Иссык-Куль.
39. Горный тушканчик. Allactaga williamsi Thomas, 1897
Табл. XXIX, 4. Карта 79
Восточное и Южное Закавказье (Талыш, равнины Азербайджана,
нагорные степи Армении), Северная Турция.
Держится преимущественно по степи и увалам предгорий, покры-
тых ксерофитной растительностью, и в горных степях. Биология не
изучена.
Подвиды не описаны.
40. Малый тушканчик. Allactaga elater Lichtenstein, 1825
Табл. XXIX, 5. Карта 80
В пределах СССР встречается в полупустынях Азербайджана, гор-
ных степях Армении, на равнинах Восточного Предкавказья, в Калмы-
кии, Южном Заволжье, в Казахстане (на север до Калмыкова, Иргиза,
оз. Зайсан), в равнинной части Средней Азии (кроме Ферганы). Встре-
чается также в Джунгарии, в Северном Иране, Афганистане.
Обитает в глинистой и реже щебенистой пустыне. Приносит, по-
видимому, 1 помет в год из 2—6 детенышей. Питается пустынными
растениями, особенно их луковицами.
В пределах СССР описано несколько подвидов, слабо различающихся по тону-
окраски меха. На сериях можно различить следующие, очень близкие между собой
формы:
а) А. е. elater Lichtenstein, 1825. (Описан из Западного Казахстана.) Окраска
спины (летом) светлая, палево-сероватая с розоватым оттенком. Казахстан. (От этой
формы практически неотличимы: А. е. dzungariae Thomas, 1912, описанная из Джунга-
рии, и А. е. vinogradovi Argyropulo, 1941, описанная из Джамбульской области.)
b) А. е. indica Gray, 1842. (Описан из Афганистана.) Окраска меха несколько свет-
лее, чем у номинальной формы (=А. е. strandi Heptner, 1934, из окрестностей Мары
в Южной Туркмении).
с) А. е. caucasicus Nehring, 1900. (Тип из окрестностей Баку и Сальян.) От номи-
нальной формы отличается более интенсивной окраской летнего меха (что можно
заметить на сериях). Закавказье. (К этой же форме относится А. е. aralichensis Satu-
nin, 1901, описанный из урочища Аралых у подножия горы Арарат, в Армении.)
280
d) A. e. kizljaricus Satunin, 1907. (Тип из Ногайской степи, севернее г. Кизляра.)
Окраска спины еще интенсивнее, чем у предыдущей формы с явной примесью ржаво-
охристых оттенков, особенно развитых на боках. Предкавказье и Нижнее Поволжье-
41, Тушканчик Бобринского, Allactaga bobrinskii Kolesnikov, 1937
Табл. XXIX, 6. Карта 80
Пустыни Средней Азии.
Образ жизни его не изучен.
Надо выяснить отношение этого вида к A. hotsoni Thomas, 1920-
(Тип из Белуджистана.)
РОД ЗЕМЛЯНЫЕ ЗАЙЧИКИ. GENUS ALACTAGULUS Nehring, 1897
Пятипалые тушканчики, похожие на малого (A. elater). Главное
отличие заключается в отсутствие у земляных зайчиков верхнего пред-
коренного зуба. Уши немного короче; «знамя» на конце хвоста слабее
выражено, однако имеется; утолщения хвоста от жировых отложений
не бывает.
Один вид.
42, Земляной зайчик, Alactagulus pygmaeus Pallas, 1779
Табл. XXIX, 7. Карта 81
Пустыни и полупустыни Восточного Предкавказья, Нижнего По-
волжья, Казахстана (на север до Чапаевска, Актюбинска, Целинного
края и гор Тарбагатая) и Средней Азии (кроме гор и Ферганской
долины). Водится также в пустынях Центральной Азии к востоку до
Ордоса и Алашаня.
Держится, главным образом, на участках глинистой пустыни, осо-
бенно предпочитая солончаки и такыры. Дает ежегодно 1 или воз-
можно 2 помета из 2—6 детенышей. Питается пустынными расте-
ниями и насекомыми. Хозяйственного значения не имеет.
В фауне СССР следующие подвиды, отличающиеся, главным образом, окраской,
которая светлеет у более южных форм:
а) А. р. dinniki Satunin, 1920. (Тип из окрестностей с. Предтеча, в ставрополь-
ских степях.) Окраска спины темная, буровато-серая с примесью рыжих оттенков.
Предкавказье и Нижнее Поволжье; А. р. tanaiticus Ognev, 1948, описанный из окрестно-
стей с. Ремонтного, в Сальских степях, так слабо отличается от данной формы, что вря.ч
ли может считаться особым подвидом.
b) А. р. pygmaeus Pallas, 1779. (Описан из степей между Волгой и Уралом.)
Окраска относительно светлая, охристо-серая. Западный и Центральный Казахстан.
с) А. р. aralensis Ognev, 1948. (Тип с р. Сарысу.) Окраска спины еще более свет-
лая и с более выступающими серыми тонами. Южный Казахстан, Забалхашье.
d) А. р. turcomanus Heptner et Samorodov, 1939. (Тип из окрестностей г. Байрам-
Али, Туркмения.) Наиболее светлая форма. Темная часть «знамени» на хвосте раз-
вита слабо. Западный Узбекистан. Туркмения.
РОД ТОЛСТОХВОСТЫЕ ТУШКАНЧИКИ. GENUS PYGERETMUS Gloger, 1841
Мелкие пятипалые тушканчики с относительно короткими ушами,
укороченными цевками задних ног, с неразвитым «знаменем» и утол-
щающимся за счет жировых отложений основанием хвоста. Предко-
ренных зубов, как и у представителей предыдущего рода, нет.
Глинистые пустыни и полупустыни от низовий р. Урала до Юж-
ного Устюрта и Восточного Казахстана, 3 вида; все только в преде-
лах СССР.
281
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ТОЛСТОХВОСТЫХ ТУШКАНЧИКОВ
1(2). Белые концевые волосы хвоста имеют длину 13—15 мм. Пе-
ред кисточкой концевых волос на хвосте имеется черный поясок. Бо-
ковые стороны резцов нижней челюсти гладкие....................
...............Тушканчик Житкова. Pygeretmus zhitkovi. (Стр. 282.)
2(1). Темные концевые волосы хвоста не превышают в длину
10 мм. Черного пояска на концевой части хвоста нет. На боковых
сторонах резцов нижних челюстей имеются же-
а б в лобки..................................................3
Рис. 102. Хвосты толсто-
хвостых тушканчиков:
А — тушканчика Житкова
(Pygeretmus zhitkovi); Б--
толстохвостого тушканчика
(Р. platyurus): В —тушкан-
чика Виноградова (Р. vino-
gradovi).
3(4). На концевой части хвоста сверху по-
лоски из темных волос нет. Задненёбное отвер-
стие значительно короче длины первого корен-
ного зуба......................Толстохвостый
тушканчик. Pygeretmus platyurus. (Стр. 282.)
4(3). В концевой части хвоста сверху тянет-
ся полоска темных волос, переходящая в ки-
сточку черноватых конечных волос. Задненёбное
отверстие по длине примерно равно первому ко-
ренному зубу . . . Тушканчик Виноградова.
Pygeretmus vinogradovi. (Стр. 282.)
43. Тушканчик Житкова. Pygeretmus zhitkovi
Kuznetzov, 1930
Табл. XXIX, 8. Карта 81
Встречается в Восточном Казахстане — в
Ала-Кульской котловине, в Прибалхашье, в рай-
оне Чу-Илийских гор, в пустыне Бет-Пак-Дала,
в степях к северу от гор Таласского Алатау.
Живет в глинистой полынной полупустыне,
на участках солянковой пустыни и на пухлых
солончаках, в сравнительно неглубоких норках.
На поверхность земли выходит только с наступ-
лением сумерек. Размножение и питание слабо
изучены. Хозяйственного значения не имеет.
Подвиды не описаны.
44. Толстохвостый тушканчик. Pygeretmus platyurus Lichtenstein, 1823
Табл. XXIX, 9. Карта 81
Населяет пустыни по нижнему течению р. Урала и по р. Эмбе,
Устюрт, северное и северо-восточное побережье Аральского моря, степи
по рекам Иргизу и Тургаю.
Держится, главным образом, среди глинистой пустыни, полупу-
стыни и в сухих степях. Образ жизни изучен слабо.
Подвиды не описаны.
Тушканчик Виноградова. Pygeretmus vinogrdovi Vorontzov, 1958
Карта 81
Найден на южном берегу оз. Зайсан, в Восточно-Казахстанской
области. Видовая самостоятельность этой формы требует подтверж-
дения.
282
Подсемейство ТРЕХПАЛЫХ ТУШКАНЧИКОВ.
Subfamilia DIPODINAE
В фауне СССР представлено четырьмя родами.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ТРЕХПАЛЫХ ТУШКАНЧИКОВ
ФАУНЫ СССР
1(2). Конец хвоста темный. «Знамя» на конце хвоста не выра-
жено, так как волосы, покрывающие хвост, постепенно удлиняются от
его середины к концу. Нижняя поверхность пальцев задних конечно-
стей покрыта редкими изогнутыми волосками темной окраски ....
.............................Емуранчики. Stylodipus. (Стр. 283.)
2(1). Конец хвоста светлый. «Знамя» на конце хвоста выражено
ясно. Нижняя поверхность пальцев задних конечностей покрыта гу-
стой щеткой из прямых беловатых волос........................ . 3
3(4). Передняя поверхность резцов желтая. В верхней челюсти
с каждой стороны по 4 коренных зуба (передний более мелкий). Сосце-
видные кости не раздуты и при рассматривании черепа сверху не видны.
.........................Мохноногие тушканчики. Dipus. (Стр. 284.)
4(3). Передняя поверхность резцов белая. В верхней челюсти
с каждой стороны по 3 коренных зуба. Сосцевидные кости сильно раз-
дуты и видны при рассматривании черепа сверху ................ 5
5(6). Нижняя поверхность пальцев задних ног покрыта щеткой
сравнительно тонких волос; по краям концов этих пальцев располо-
жены ряды более коротких, но толстых и упругих щетинок. Длина уха
более 32 мм. Вершины барабанных камер черепа не соприкасаются
друг с другом . . . Гребнепалые тушканчики. Paradipus. (Стр. 284.)
6(3). Нижняя поверхность пальцев задних ног покрыта щеткой
однородных тонких и мягких волосков. Длина уха меньше 32 мм. Вер-
шины барабанных камер черепа тесно соприкасаются друг с другом.
.........................Песчаные тушканчики. Jaculus. (Стр. 285.)
РОД ЕМУРАНЧИКИ. GENUS STYLODIPUS G. Allen, 1925
Один вид.
45. Емуранчик обыкновенный. Stylodipus telum Lichtenstein, 1823
Табл. XXX, Карта 82
Распространен в пустынных и полупустынных пространствах Во-
сточного Предкавказья, Нижнего Поволжья, Казахстана (кроме его
северных частей). Изолированная часть ареала охватывает Алешкин-
ские пески, на левом берегу Нижнего Днепра; встречается также в пес-
ках по низовьям Дона.
Держится преимущественно среди глинистой и щебенистой пу-
стыни, но местами селится также на песчаных участках среди бахчей,
обнажениях песка, в степных борах, солончаках и на бахчах. Дает
ежегодно 1 или возможно 2 помета, из 3—6 детенышей каждый. Пи-
тается степными и пустынными растениями, особенно предпочитая их
луковицы. Хозяйственного значения не имеет.
Подвиды:
a) S. t. falzfeini Brauner, 1913. (Тип из Алешкинских песков.) Окраска верха свет-
лая, палево-охристая. Размеры относительно крупные. Низовья Днепра.
283
b) S. t. turovi Heptner, 1934. (Тип из окрестностей ст. Торговой.) Окраска спины
темная с примесью буровато-оливковых тонов, заметно затемненная черноватыми
окончаниями остевых волос. Восточное Предкавказье, Нижнее Поволжье.
с) S. t. amankaragai Selevin, 1934. (Тип из Наурзумского бора.) Окраска спины
относительно насыщенная с ясными желтоватыми оттенками и заметными бурова-
тыми крапинами. Южно-кустанайские, иргизские, акмолинские степи.
d) S. t. telum Lichtenstein, 1823. (Описан с северо-восточного побережья Араль-
ского моря.) Спина светлого, палево-желтого цвета, затемненного на огузке буровато-
серыми неясными крапинами. Низовья Урала и Эмбы, приаральские пустыни.
е) S. t. karelini Selevin, 1934. (Тип из сопок Семей-Tay, близ Семипалатинска.)
Окраска спины темная, сероватая с сильным развитием буроватой крапчатости. Во-
сточный и Центральный Казахстан.
РОД МОХНОНОГИЕ ТУШКАНЧИКИ. GENUS DI PUS Zimmermann, 1780
Один вид.
46. Мохноногий тушканчик. Dipus sagitta Pallas, 1773
Табл. XXX, 2. Карта 83
Пустыни и полупустыни Восточного Предкавказья и Нижнего По-
волжья, волго-уральские степи, Казахстан (кроме северных и горных
районов), равнинные области Средней Азии; широко распространен
в пустынях Центральной Азии к востоку до Внутренней Монголии.
Держится преимущественно на участках песчаной пустыни, встре-
чаясь как среди барханов, так и в песках, закрепленных раститель-
ностью. Живет также в ленточных борах на песках по р. Иртышу. В по-
мете 2—5 детенышей. Питается травянистыми растениями и веточками
кустарников. Поедает насекомых.
В фауне СССР представлен рядом подвидов:
a) D. s. nogai Satunin, 1907. (Тип из прикумских песков.) Размеры крупные: кон-
дилобазальная длина черепа 32—34 мм. Спина тусклого, буроватого цвета с легкой
примесью охристых оттенков. Восточное Предкавказье.
b) D. s. innae Ognev, 1930. (Тип из окрестностей Енотаевска.) Размеры мельче:
кондилобазальная длина черепа 30—32 мм. Окраска спины более яркая, с резко вы-
ступающими охристо-ржавыми оттенками. Нижнее Поволжье, Волго-Уральские пески.
с) D. s. lagopus Lichtenstein, 1823. (Описан с северо-восточного побережья Араль-
ского моря.) Размеры средние: кондилобазальная длина черепа 30—33 мм. Окраска
светлая, бледная, спина охристо-песчаного цвета. Южный Казахстан, Средняя Азия.
d) D. s. sagitta Pallas, 1773. (Тип с р. Иртыша, окрестности ст. Ямышевской.)
Размеры крупные: кондилобазальная длина черепа до 34 мм. Окраска спины темная,
охристо-коричневая с оливковым оттенком. Ленточные боры по р. Иртышу и в Пред-
алтайских степях.
е) D. s. zaissanensis Selevin, 1934. (Тип из Буконских песков.) Отличается от ти-
пичной формы более яркой и ржавой окраской спины. Зайсанская котловина.
РОД ГРЕБНЕПАЛЫЕ ТУШКАНЧИКИ. GENUS PARADIPUS Vinogradov, 1930
Только один вид.
47. Гребнепалый тушканчик. Paradipus ctenodactylus Vinogradov, 1929
Табл. XXX, 3. Карта 83
Найден в песчаных пустынях Каракумы и Кызылкумы.
Обитает преимущественно среди кустарниковой песчаной пустыни.
Норы роет по склонам барханов. Питается веточками пустынных ку-
старников и травянистыми растениями. Посередине зимы впадает
в кратковременную спячку. Размножение не изучено. Подвиды не
описаны.
284
РОД ПЕСЧАНЫЕ ТУШКАНЧИКИ. GENUS JACULUS Erxleben, 1777
Песчаные пустыни Северной Африки, Аравии, Передней и Сред-
ней Азии. Из 4 видов в фауне СССР — 2 вида.
« ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ПЕСЧАНЫХ ТУШКАНЧИКОВ
ФАУНЫ СССР
1 (2). Длина ступни более 60 мм. Длина уха более 25 мм. Отвер-
стие в чешуйчатой кости сзади от скулового отростка большое и имеет
форму равнобедренного треугольника ...........................
............Туркменский тушканчик. Jaculus turkmenicus. (Стр. 285.)
Рис. 103. Черепа песчаных тушкан-
чиков (сбоку):
.4 — туркменского тушканчика (Jaculus
turkmenicus); Б — тушканчика Лихтен-
штейна (J. lichtensteini); гтк — гребень
теменной кости; зтк — зубец теменной
кости.
ГТК
зтк
ь
2(1). Длина ступни менее 55 мм. Длина уха менее 20 мм, Отвер-
стие в чешуйчатой кости черепа сзади скулового отростка небольшое
и вытянутое...............................................................
. . . . Тушканчик Лихтенштейна. Jaculus lichtensteini. (Стр. 285.)
48. Туркменский тушканчик. Jaculus turkmenicus Vinogradov
et Bondar, 1949
Табл. XXX, 4, Карта 82
Известен по нескольким экземплярам из песчаных пустынь Турк-
мении. Обитает в песках, поросших пустынными кустарниками, и по
такырам.
Биология не изучена.
49. Тушканчик Лихтенштейна. Jaculus lichtensteini Vinogradov, 1927
Табл. XXX, 5. Карта 82
Распространен в пустынях Каракумы, Кызылкумы, Северного При-
аралья и Южного Прибалхашья. Живет по заросшим и развеянным
бугристым пескам. Образ жизни почти не известен.
Подсемейство КАРЛИКОВЫХ ТУШКАНЧИКОВ.
Subfamilia CARDIOCRANIINAE
Мелкие примитивные тушканчики с чертами высокой специализа-
ции к жизни в условиях пустыни. Размером не превышают домовую
мышь. Уши в основной части имеют вид трубочек, так как их внутренние
края у основания раковины срастаются. Задние конечности пяти- или
трехпалые. Кости плюсны не срастаются в цевку, как у других тушкан-
чиков. Череп с огромными сосцевидными костями, отчего приобретает
сердцевидную форму (рис. 99). Межтеменная кость не развита или
очень мала.
Распространены в пустынях Центральной Азии.
В подсемействе 2 рода; оба представлены в фауне СССР.
285
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ПОДСЕМЕЙСТВА
КАРЛИКОВЫХ ТУШКАНЧИКОВ
1(2). Задние ноги пятипалые. Длина хвоста менее 8 см. Скуловые
дуги черепа без отростков, отходящих от их середины вниз.........
. . . Пятипалые карликовые тушканчики. Cardiocranius. (Стр. 286.)
2(1). Задние ноги трехпалые. Длина хвоста более 8 см. Скуловые
дуги черепа с угловатым выступом или саблевидным отростком, на-
правленным от их средней части вниз..............................
. . . . Трехпалые карликовые тушканчики. Salpingotus. (Стр. 286.)
РОД ПЯТИПАЛЫЕ КАРЛИКОВЫЕ ТУШКАНЧИКИ.
GENUS CARDIOCRANIUS Satunin, 1902
Очень мелкие тушканчики: длина тела 5—6 см с большой головой
. и довольно коротким жирным хвостом. На задних ногах подошвы по-
крыты слаборазвитой щеткой жестких волос. Уши короткие.
Единственный вид.
50. Пятипалый карликовый тушканчик1. Cardiocranius paradoxus
Satunin, 1902
Табл. XXX, 6. Карта 81
Найден в Тувинской области. Житель глинистых и песчаных пу-
стынь. Ночное животное. Питается как растительной (в основном семе-
нами), так и животной (главным образом, насекомыми) пищей. Раз-
множение не изучено.
РОД ТРЕХПАЛЫЕ КАРЛИКОВЫЕ ТУШКАНЧИКИ.
GENUS SALPINGOTUS Vinogradov, 1922
Столь же мелкие тушканчики с более длинным тонким хвостом
(длина 8—И см). Задние ноги несут густую щетку длинных волосков.
Уши относительно длинные.
Три вида, из которых в СССР встречается только один.
51. Трехпалый карликовый тушканчик1. Salpingotus crassicauda
Vinogradov, 1924
Табл. XXX, 7. Карта 81
Распространен в степях Джунгарии. В СССР найден в Зайсан-
ской котловине. Обитает в песках. Питается насекомыми и семенами.
Очень редкий вид нашей фауны.
СЕМЕЙСТВО МЫШОВКОВЫЕ. FAMILIA ZAPODIDAE
Мелкие мышевидные грызуны с очень длинным хвостом, длина ко-
торого не менее чем на 15% длиннее тела, покрытым кольцами чешуек
с сидящими между ними отдельными редкими волосками. Задние ноги
1 Автором сохранены трехсловные видовые названия. Считая важным сохранить
в названии ссылку на число пальцев, предлагаем называть их: пятипалый карлик и
трехпалый карлик. (Ред.)
286
примерно в два раза длиннее передних. Укороченные первый и пятый
пальцы своими когтями не достигают основания средних пальцев. Уши
округлые. Череп внешне сходен с черепом мышей, но несет по 4 корен-
ных зуба с каждой стороны верхней челюсти.
Большинство авторов выделяет мышовок в особое подсемейство
семейства тушканчиков. Наличие родственных связей между мышов-
ками и тушканчиками бесспорно, но все же и различия между ними
настолько велики, что мышовки (вместе с американскими прыгунчи-
ками) могут быть выделены в отдельное семейство. Мы не видим осно-
ваний считать, что мышовки связаны с тушканчиками промежуточными
формами, как-то: Euchoreutes, Cardiocranius и Salpingotus. Нет сом-
нения, что все эти формы — высокоспециализированные представители
семейства тушканчиков и прямой филогенетической связи с мышов-
ками не имеют.
Семейство мышовок распространено в Европе, Азии и Северной
Америке.
В СССР оно представлено подсемейством Sicistinae, к которому
принадлежит род мышовок.
РОД МЫШОВКИ. GENUS SICISTA Gray, 1827
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ МЫШОВОК ФАУНЫ СССР
1(4). Вдоль спины тянется черная или черноватая полоска . . 2
2(3). Хвост не более чем на 73 длиннее тела. Длина задней ступни
менее 15,8 мм. По бокам черного спинного ремня расположены про-
долговатые участки сравнительно светлого
меха, которые на боках тела сменяются не-
сколько более темными полями................
Степная мышовка. Sicista subtilis. (Стр. 288.)
3(2). Хвост более чем на 7з длиннее те-
ла. Длина задней ступни взрослых особей бо-
лее 15,8 мм. Окраска спины и боков тела, за
исключением спинного черного ремня, одно-
тонная и равномерная, изменяющаяся от па-
лево-серой до охристо-рыжеватой.............
Лесная мышовка. Sicista betulina. (Стр. 288.)
4(1). Черной полоски на спине, проходя-
щей вдоль хребта, нет.....................5
5(8). Хвост обычно более чем в полтора
раза длиннее тела. Головка полового органа
самца покрыта большим количеством мел-
ких роговых шипиков; большие шипы от-
сутствуют ................................6
6(7). Окраска хвоста однотонная: верхняя
темнее нижней. Половой орган самца короткий и массивный; срединная
щель его доходит до половины его длины..............................
...............Одноцветная мышовка. Sicista concolor.* (Стр. 289.)
7(6). Окраска хвоста резко двухцветная: сверху — темная, снизу —
светлая. Половой орган самца вытянутый: срединная щель его доходит
только до 7з длины органа...........................................
...............Кавказская мышовка. Sicista caucasica Г (Стр. 289.)
Рис. 104. Половые ор-
ганы взрослых самцов
мышовок:
А—одноцветной, (Sicista con-
color); Б — алтайской (Si-
cista параеа).
сторона лишь немного
1 Возможно, все одноцветные мышовки относятся к одному виду. (Ред.)
287
8(5). Хвост обычно менее чем на */2 длиннее тела. На половом ор-
гане самца, помимо многих мелких роговых шипиков, имеется одна или
две пары больших шипов...............................................9
9(10). Спина рыжевато-бурая, брюхо серовато-ржавое. На головке
полового органа самца расположены две пары больших роговых ши-
пов, направленных назад ...............................................
..............Мышовка алтайская рыжая. Sicista параеа. (Стр. 289.)
10(9). Спина буровато-серая, брюхо сероватое. На головке поло-
вого органа, близ ее вершины, имеется лишь одна пара больших ши-
пов, направленных назад ...............................................
. . . . Мышовка алтайская серая. Sicista pseudonapaea. (Стр. 289.)
52. Степная мышовка. Sicista subtilis Pallas, 1773
Табл. XXX, 8. Карта 84
Населяет полосу степей от Венгрии до Байкала. В СССР северная
граница ареала проходит через Северную Украину, Татарию, Южный
Урал (Омскую, Новосибирскую, Иркутскую области), на юг — до се-
веро-западного берега Черного моря, северных предгорий Крыма, по-
бережья Азовского моря, гор Кавказского хребта, калмыцких и волго-
уральских степей и Северного Казахстана, где южная граница тянется
от р. Эмбы через Тургай, район Караганды, котловину Алакуля; оби-
тает также в кузнецких, минусинских и прибайкальских степях.
Держится преимущественно среди целинной степи, на выгонах,
сенокосах, по балкам. Живет в неглубоких норках. На зиму впадает
в спячку. Питается, главным образом, семенами и насекомыми. При-
носит 1 помет в году — весной.
Подвиды, водящиеся в СССР:
a) S. s. subtilis Pallas, 1773. (Описана из степей в верховьях Тобола.) Окраска
спины довольно светлая, бледно-серая с желтоватым оттенком, особенно заметным на
боках тела. Черный ремень вдоль хребта и светлые поля по бокам от него резко
выражены. Степи Южного Урала, Зауралья и Западной Сибири.
b) S. s. vaga Pallas, 1778. (Описана из полупустынь по нижнему течению Урала.)
Окраска спины очень бледная, светло-серая с желтоватым оттенком. Ремень на спине
узкий, черно-бурый. Темноватые поля на боках тела слабо развиты. Волго-уральские
степи, полупустыни Северного Казахстана.
с) S. s. nordmanni Keyserling et Blasius, 1840. (Тип из окрестностей Одессы.)
Близка к S. s. vaga, но спина серее, без желтоватых оттенков. Черный ремень на за-
гривке слабо выражен. Степи Юго-Западной Украины.
d) S. s. severtzovi Ognev, 1935. (Тип из Каменной степи, в Воронежской области.)
Окраска интенсивная, серовато-буроватая. Ремень на спине широкий, чисто черный.
Темные поля на боках тела выражены резко. Степи Европейской части СССР, кроме
Юго-Западной Украины и Южного Заволжья.
е) S. s. sibirica Ognev, 1935. (Тип с р. Котанды, на Алтае.) Сходна с S. s. severtzovi,
но с более рыжеватым оттенком меха, более узким черным ремнем, менее выражен-
ными темными полями на боках тела. Предалтайские, кузнецкие, минусинские и при-
байкальские степи.
53. Лесная мышовка. Sicista betulina Pallas, 1778
Табл. XXX, 9. Карта 84
Распространена в лесной и лесостепной зонах Европы и Азии, ме-
стами проникает в полосу степей. В Советском Союзе обитает к северу
до Карелии, Архангельской области, низовьев Печоры, среднего тече-
ния р. Таза. Южная граница ареала тянется по Северной Украине,
Северному Кавказу и Северному Казахстану; на восток — до Забай-
калья включительно.
Живет в лесах, кустарниковых зарослях, заросших кустами балках
в степи. Ведет скрытный ночной образ жизни. Зимой спит. Норку и
288
гнездо устраивает обычно в гнилых пнях и дуплах деревьев. Питается
насекомыми, семенами, ягодами. Размножение не изучено.
Подвидов, по-видимому, не образует.
54. Одноцветная мышовка. Sicista concolor Buchner, 1892
Табл. XXX, 10—12. Карта 84
Широко распространена в Азии, от Тихого океана до Афганистана.
В СССР водится на о-ве Сахалин, в Приморской области, в горах Се-
миреченского Тянь-Шаня.
Держится в лиственных и смешанных лесах, в кустарнике. В горах
поднимается до субальпийских лугов. На зиму впадает в спячку. Ве-
дет преимущественно ночной образ жизни. Пищей ей служат насеко-
мые, семена, ягоды.
Типичная форма встречается в Центральной Азии (тип из Гань-Су). В СССР —
2 подвида:
a) S. с. tianschanica Salensky, 1903. (Тип из Синьцзяня.) Окраска спины сероваго-
корпчневая, заметно затемненная черными концами остевых волос. Тянь-Шань.
b) S. с. caudata Thomas, 1907. (Тип с Южного Сахалина.) Окраска спины с более
выраженными рыжеватыми оттенками. Приморье, Южный Сахалин.
Кавказская мышовка. Sicista caiicasica Vinogradov, 1925
Карта 84
Найдена в различных частях Главного Кавказского хребта и в Ар-
мении. Образ жизни не изучен.
Рыжая алтайская мышовка. Sicista napaea Hollister, 1912
Карта 84
Встречается в горах и предгорьях Алтая. Держится, главным об-
разом, по лужайкам и на вырубках среди тайги.
Биология не изучена.
Серая алтайская мышовка. Sicista pseudonapaea Е. Strautman, 1949
Карта 84
Найдена в горах Южного Алтая. Обитает в тайге, по опушкам
леса, в кустарнике, на вырубках. Образ жизни известен слабо.
СЕМЕЙСТВО МЫШИНЫЕ. FAMILIA MURIDAE
Очень разнообразные по внешнему облику мелкие или средние по
размеру грызуны. Общим признаком всех представителей этого семей-
ства является строение их зубной системы, в которой отсутствуют пред-
коренные зубы. С каждой стороны как верхней, так и нижней челюсти
мышиных, водящихся в СССР, имеется по 3 коренных зуба с плоской,
несущей бугорки или поперечные гребни жевательной поверхностью
(у некоторых зарубежных форм число коренных зубов сокращено до
2 с каждой стороны); От семейства слепышей, у которых имеется такое
же количество коренных зубов, мышиные отличаются тем, что у них
глаза не скрыты под кожей, имеется хотя бы короткий хвост, по бокам
головы нет полоски из жестких щетинок.
Некоторые зоологи предлагают подразделить данное семейство на два:
1) Muridae, куда они относят мышей и крыс, и 2) Cricetidae, охватывающее все виды
19 Н. А. Бобринский и др.
289
Рис. 105. Передние лапы:
А— цодяной крысы (полевки)
(Arvicola terrestris); Б — цокора
(Myospalax myospalax).
хомяков, песчанок, полевок и цокоров. Первое пз ппх характеризуется тем, что у его
представителей верхние коренные зубы несут на жевательной поверхности три ряда
бугорков, тогда как виды семейства Cricetidae имеют двухрядное расположение бу-
горков на этих зубах. Нам кажется подразделение мышиных грызунов на два отдель-
ных семейства недостаточно обоснованным.
Распространены мышиные по всей суше, исключая Антарктиду и
некоторые океанические острова.
Описано около тысячи видов этого семейства, ведущих самый раз-
личный образ, жизни и населяющих самые различные места обитания.
Семейство мышиных делится на 12 подсемейств, В СССР обитают
представители 5 из них,
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ СЕМЕЙСТВА МЫШИНЫХ
ФАУНЫ СССР
1(2). Если верхние коренные зубы еще не стерты, их жевательная
поверхность несет три ряда небольших бугорков; если же они подвер-
гались стиранию, то их несколько сглаженная жевательная поверх-
ность имеет ряд прямых или изогнутых поперечных эмалевых петель,
дентиновые островки которых отделены друг от друга эмалевыми мо-
стиками. Хвост длинный, длина его обычно не менее 70% длины тела;
он покрыт кольцами чешуек, позади которых сидят одиночные редкие
упругие волоски. (Плавательной перепонки между пальцами задних
ног нет.)...............Подсемейство мышей. Murinae. (Стр. 291.)
2(1). Жевательная поверхность коренных зубов верхней челюсти
плоская или несет два ряда небольших бугорков; если зубы подверг-
лись стиранию, то на месте этих бугорков образуются поперечные эма-
левые петли, дентиновые средние части которых соединены друг с дру-
гом перемычками. Хвост либо короткий (длиной менее 70% длины
тела), либо длинный, но в последнем случае покрыт волосами, так что
кожных чешуек не видно (лишь у ондатры хвост длинный и чешуйча-
тый, но у нее пальцы задних ног связаны плавательной перепонкой) , 3
3(4). Передняя поверхность резцов верхней челюсти имеет одну
или две продольные бороздки. Хвост длиннее 70% длины тела, покрыт
волосами............Подсемейство песчанок. Gerbillinae. (Стр. 300.)
4(3). Передняя поверхность резцов верхней челюсти гладкая.
Хвост короче 60% длины тела, либо, если длиннее, покрыт кожными
чешуйками (лишь у водящегося в Туркмении и в Закавказье мыше-
видного хомячка он длинный и покрыт волосами)...................5
5(6). Жевательная поверхность верхних коренных зубов неровная,
несущая небольшие бугорки (если зубы еще не стерлись) или попереч-
ные эмалевые петли (у стертых зубов). Коронки этих зубов не подраз-
делены на ряд треугольных призм боковыми складками эмалевой
стенки (рис. 113) , , , Подсемейство хомяков. Cricetinae. (Стр. 306.)
6(5). Жевательная поверхность коренных зубов верхней челюсти
плоская. Наружные и внутренние эмалевые стенки этих зубов вдаются
вглубь острыми углами, разделяя коронку на ряд треугольных призм
(рис. 122)......................................................7
7(8). Когти передних лап примерно равны или короче длины паль-
цев. Ушные раковины обычно хорошо развиты.........................
..................Подсемейство полевок. Microtinae (Стр. 314.)
8(7). Когти передних лап огромны: длина их значительно превы-
шает длину самих пальцев. Ушные раковины зачаточны................
. . . Подсемейство цокоров. Myospalacinae. (Стр. 343.)
Подсемейство МЫШЕИ. Subfamilia MURINAE
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ПОДСЕМЕЙСТВА МЫШЕИ
ФАУНЫ СССР
1(6). Размеры мелкие; длина ступни менее 25 мм, кондилобазаль-
ная длина черепа менее 30 мм.................................2
2(3). Теменные кости образуют на передних наружных углах на-
правленные вперед узкие заостренные отростки (рис. 106, А). На внут-
ренней стороне верхних резцов имеется небольшой уступ, в который
упираются концы резцов нижней пары (рис. 107, А) .......
...............................Домовые мыши. Mus. (Стр. 292.)
3(2). Теменные кости не имеют направленных вперед узких от-
ростков (рис. 106, Б). Уступчика на внутренней поверхности верхних
резцов нет (рис. 107, Б).....................................4
4(5). Длина тела не более 70 мм; длина ступни задних ног менее
16 мм; кондилобазальная длина черепа не более 20 мм. Расстояние от
передней поверхности резцов до задней стороны последнего коренного
зуба верхней челюсти меньше расстояния от верхней челюсти до зад-
него края затылочного мыщелка..................................
. . . . . ..........Мыши-малютки. Micromys. (Стр. 293.)
5(4). Длина тела взрослых животных более 70 мм; длина ступни
задних ног свыше 16 мм; кондилобазальная длина черепа более 20 мм.
Рис. 106. Черепа мышей:
Л — домовая мышь (Mus musculus); Б — лесная
мышь (Apodemus sylvaticus).
Рис. 107. Верхние резцы мышеи
(вид сбоку):
А—домовая мышь (М. musculus);
Б — лесная мышь (Apodemus sylva-
ticus); У — уступ на задней поверх-
ности резца домовой мыши.
19*
291
Расстояние от передней поверхности резцов до задней стороны послед-
него коренного зуба верхней челюсти превышает расстояние от верх-
ней челюсти до заднего края затылочного мыщелка...................
......................Лесные и полевые мыши. Apodemus. (Стр. 294.)
6(1). Размеры крупные: длина ступни взрослых более 25 мм; кон-
.дилобазальная длина черепа свыше 30 мм.........................7
7(8). Хвост превышает 2/з длины тела. Скуловые дуги расставлены
узко: скуловая ширина менее 55% кондилобазальной длины черепа.
Расстояние между наружными сторонами резцов верхней челюсти у их
основания примерно равно ширине носового отверстия черепа (рис.
108, Л). Жевательная поверхность коренных зубов с бугорками или
с дугообразноизогнутыми трехлопастными эмалевыми петлями . . . .
.....................................Крысы. Rattus. (Стр. 297.)
8(7). Хвост равен '/2—2/з длины тела. Скуловые дуги расставлены
широко, так что скуловая ширина составляет более 55% кондилоба-
Рис. 108. Черепа крыс (спереди):
А — пасюк (Rattus norvegicus); Б — пластинчатозубая крыса
(Nesokia indica).
.зальной длины черепа. Резцы очень широкие: расстояние между внеш-
ними сторонами резцов у их основания значительно превышает ширину
носового отверстия черепа. У взрослых коренные зубы несут на жева-
тельной поверхности вытянутые в поперечном направлении овальные
эмалевые петли (на переднем — три, на втором и третьем — две) . . .
. . ..............Пластинчатозубые крысы. Nesokia. (Стр. 299.)
РОД ДОМОВЫЕ МЫШИ. GENUS MUS Linnaeus, 1758
Мыши средних размеров (длина тела до 11 см) с относительно ко-
ротким хвостом, длина которого составляет 65—100% длины тела.
Окраска спины буровато-серая, серая или песочного цвета. Брюшко
серое, беловатое или белое. Хвост сверху лишь немного темнее, чем
снизу. На передних боковых углах теменных костей черепа имеются
острые, направленные вперед отростки. На задней поверхности верх-
них резцов имеется небольшой уступ, в который упираются концы
нижних резцов. Распространены почти по всему земному шару.
В фауне СССР — один вид.
55. Домовая мышь. Mus musculus Linnaeus, 1758
Табл. XXXI, 2—3. Карта 86
Ныне с помощью человека расселилась по большой территории.
В СССР встречается почти повсеместно, кроме Крайнего Севера и не-
J292
которых глухих таежных районов. В северных частях страны живет
преимущественно в постройках человека или вблизи их. На юге-
обычна также в целинной степи, на пашнях. Размножается несколько-
раз в году, принося по 4—8 детенышей. Питается различной расти-
тельной и животной пищей. Приносит большой ущерб как амбарный и.
полевой вредитель.
Различными исследователями описано большое количество форм домовых мы-
шей из разных районов Советского Союза. Но большая часть их реально неразли-
чима. Можно наметить несколько наиболее обособленных географических рас:
а) М. m. musculus Linnaeus, 1758. (Описана из Швеции.) Крупная форма с длин-
ным хвостом: длина тела 70—105 мм; длина хвоста 60—100 мм; средняя длина зад-
ней ступни 17,0 мм. Окраска спины темная, буровато-серая, брюха грязно-серая,
иногда с желтым оттенком. Лесная и лесостепная полосы Европейской части СССР
и Сибири.
b) М. m. hortulanus Nordmann, 1840. (Тип из Одессы.) Размеры мельче: длина
тела 60—80 мм; хвост относительно длинный (от 65 до 90% длины тела); длина ступни
14,5—17,0 мм. Окраска спины светлая, сероватая или песчано-серая, брюшка беловатая
(основания волос серые). Степные области юга Европейской части СССР, Западной
Сибири, большей части Казахстана, равнины Азербайджана.
с) М. m. manchu Thomas, 1909. (Тип из провинции Гирин, в Северо-Восточном
Китае.) Размеры средние: длина тела 70—82 мм; хвост короткий — длина его 42—55 мм
(50—70% длины тела); ступня короткая — 13,5—16 мм. Окраска примерно, как у пре-
дыдущего подвида. Степи Забайкалья и Северо-Восточного Китая.
d) М. m. abbotti Waterhouse, 1839. (Тип из Малой Азии.) Размеры довольно
крупные: длина тела 65—95 мм; хвост относительно длинный — от 90 до 100% длины
тела; длина ступни 15,5—19 мм. Окраска спины средней темноты, серо-коричневая,,
брюшка сероватая. Горные районы Закавказья и Дагестана.
е) М. m. bactrianus Blyth, 1846. (Тип из Кандагара, в Афганистане.) Размерь?
сравнительно мелкие: хвост длинный (от 70 до 100% длины тела). Окраска очень,
светлая: спина светлая, песчано-желтая, брюшко чисто белое. Средняя Азия, Южный
Казахстан, пустыни Центральной Азии.
РОД МЫШИ-МАЛЮТКИ. GENUS MICROMYS Dehne, 1841
Мелкие мыши, длина тела которых не превышает 7 см. Длина!
хвоста 4—7 см. Окраска спины изменяется от ярко-рыжей до бурова-
той. Брюшко белое. Череп отличается короткой носовой частью и ок-
руглой мозговой коробкой.
Один вид.
56. Мышь-малютка. Micromys minutus Pallas, 1771
Табл. XXXI, 1, Карта 85
Распространена от Западной Европы до берегов Тихого океана,
В СССР обитает почти по всей Европейской части страны (кроме Се-
вера), в Северном Казахстане, в Южной Сибири и на Дальнем Во-
стоке. Северная граница ареала идет от Центральной Карелии через
южные районы Архангельской области, бассейн р. Конды, Нарымский
край и Центральную Якутию.
Чаще всего встречаются на полях, сенокосах, огородах, в зарос-
лях камыша по берегам рек. Летом живут обычно в шарообразных,
гнездах, которые укрепляют на стеблях хлебных злаков, трав или ка-
мыша. Размножаются несколько раз в году; число детенышей в поме-
те— от 4 до 8. На зиму значительная часть малюток укрывается в оме-
тах соломы и стогах сена. Питаются семенами и зелеными частями
травянистых растений. Местами вредят посевам зерновых.
293
Из пределов СССР описан ряд подвидов, различающихся преимущественно от-
тенками окраски; реальность существования большинства из них вызывает сомнение.
Вероятно, в нашей стране встречается только 3 подвида:
а) М. m. minutus Pallas, 1771. Окраска спины сравнительно светлая и яркая.
Европейская часть СССР, Западная Сибирь, Северный Казахстан.
b) М.-m. batarovi Kastschenko, 19L1. Окраска спины более темная и тусклая. При-
байкалье, Забайкалье, Южная Якутия..
с) М. ш. ussuricus Barret-Hamilton, 1889. Спина окрашена в насыщенный буро-
вато-ржавый цвет. Приморье, Уссурийский край.
РОД ЛЕСНЫЕ И ПОЛЕВЫЕ МЫШИ. GENUS APODEMUS Каир, 1829
Сравнительно крупные мыши: длина тела взрослых особей до
14 см. Окраска спины рыжеватая или буроватая, иногда с черной по-
лосой по хребту. Брюхо белое или сероватое. Хвост обычно сверху тем-
ный, снизу беловатый. Длина ступни 17—25 мм. Череп с удлиненным
носовым отделом. Его теменные кости имеют тупые передние наруж-
ные углы, без направленных вперед острых отростков.
Задняя поверхность верхних резцов гладкая, без уступа.
Широко распространены в Европе, Северной Африке и Азии.
5 видов; все представлены в фауне СССР.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ЛЕСНЫХ И ПОЛЕВЫХ МЫШЕИ
ФАУНЫ СССР
1(2). На спине вдоль хребта тянется резко выступающая черная
полоса. Наружный бугорок в первой петле второго коренного зуба
верхней челюсти отсутствует.....................................
......................Полевая мышь. Apodemus agrarius. (Стр. 295.)
2(1). На спине продольной черной полосы нет. Первая петля вто-
рого коренного зуба верхней челюсти образует два бугорка — наруж-
ный и внутренний ............................................. 3
3(8). Окраска спины светло-коричневая или серовато-коричневая,
обычно с рыжеватыми оттенками. С наружной стороны заднего корен-
ного зуба верхней челюсти в третьей петле только один бугорок ., . 4
4(7). Размеры относительно крупные: длина тела до 14 см\ длина
ступни взрослых более 22 мм\ кондилобазальная длина черепа более
22 мм..........................................................5
5(6). На груди между основаниями передних ног желтого пятна
нет. Края межглазничного промежутка черепа несут гребнеобразные
оторочки . Восточноазиатская мышь. Apodemus speciosus. (Стр. 295.)
6(5). На груди между основаниями передних ног имеется желтое
пятно. По краям межглазничного промежутка черепа гребневидных
оторочек нет . . Желтогорлая мышь. Apodemus flavicollis. (Стр. 296.)
7(4). Размеры более мелкие: длина тела, до 11 см\ длина ступни
задних ног менее 22 мм; кондилобазальная длина черепа обычно менее
22 мм.............Лесная мышь. Apodemus sylvaticus. (Стр. 296.)
8(3). Окраска спины серовато-бурая без заметной примеси корич-
неватых или рыжеватых оттенков. Задняя (третья) петля последнего
коренного зуба верхней челюсти несет на своем наружном крае два
бугорка...........Горная мышь. Apodemus mystacinus. (Стр. 297.)
Примечание. В ряде районов юго-востока Европейской части страны при-
знаки лесной и желтогорлой мышей настолько сближаются, что различать их трудно.
294
57. Полевая мышь. Apodemus agrarius Pallas, 1778
Табл. XXXI, 4. Карта 87
Распространена от Западной Европы до берегов Тихого океана.
В СССР обитает в Европейской части страны (кроме Крыма и север-
ных таежных и тундровых районов), в Северном и Восточном Казах-
стане, Северной Киргизии, южных районах Западной и Средней Си-
бири на восток до Байкала; в Забайкалье не найдена, но встречается
по Амуру и в Приморье. Северная граница ареала идет от Средней Ка-
релии, севернее Вологды и Кирова к г. Серову (на Урале), а оттуда
через Нарымский край, Енисейский и Братский районы к Байкалу. На
Дальнем Востоке она идет от Благовещенска, через Дуссе-Алинь, ни-
зовья р. Горюна к Советской гавани. На юг полевая мышь распростра-
нена до Черноморского побережья, перекопских степей, берегов Азов-
ского моря, северных склонов Кавказского хребта, северных окраин
пустынь Прикаспийской низменности, актюбинских, акмолинских и
карагандинских степей, Прибалхашья, Северной Киргизии.
Селится на пашнях, лугах, опушках леса, в кустарнике, по овра-
гам и берегам рек, на огородах. Живет в неглубоких норках. На зиму
обычно скопляется в ометах соломы, стогах сена и проникает в по-
стройки человека. Дает за год до 3 пометов из 3—9 детенышей. Пи-
тается семенами и зелеными частями травянистых растений, поедает
насекомых. Вредит посевам, особенно на Дальнем Востоке и в Запад-
ной Сибири.
Географическая изменчивость полевой мыши выражена слабо, но резко выра-
жены сезонные и возрастные морфизмы. На территории СССР реально можно раз-
личить (и то на сериях) только 2 подвида:
a) A. a. agrarius Pallas, 1778. (Описана из поймы Волги у г. Ульяновска.) Размеры
относительно мелкие: общая длина черепа 21'—24 мм. Окраска более тусклая. Запад-
ная часть ареала вида, на восток — до Иркутска.
b) A. a. mantchuricus Thomas, 1898. (Тип из Северо-Восточного Китая, близ корей-
ской границы.) Размеры крупные: общая длина черепа взрослых особей 26—29 мм.
Окраска более яркая. Дальневосточная часть ареала вида. Название подвида дано
условно, до выяснения различий с формами, описанными ранее: например, A. a. che-
vrieri Milhe-Edwards, 1868, или A. a. ningpoensis Swinhoe, 1870.
58. Восточноазиатская мышь. Apodemus speciosus Temminck, 1845
Табл. XXXI, 5. Карта 88
Япония, Китай, Монголия. В СССР распространена на Дальнем
Востоке, в Восточной Сибири. Прибайкалье, Саянах, Алтае. Западная
граница идет по предгорьям Алтая и Саян, через Северное Прибай-
калье к бассейну р. Вилюя. Есть указания на нахождения ее в ни,-
зовьях Лены. От Лены до Охотского побережья северная граница
ареала не выяснена.
Держится по опушкам леса,, в кустарниках, по оврагам, на полях.
Питается опавшими орехами и семенами деревьев, а также насе-
комыми.
В СССР встречаются 2 слабо различающиеся географические формы:
a) A. s. praetor Miller, 1914. (Тип из провинции Гирин, в Северо-Восточном Китае.)
Отличия от номинальной формы, описанной с о-ва Хондо, в Японии, точно не уста-
новлены. Размеры относительно крупные: длина тела 105—125 мм; длина хвоста
92—109 мм; кондилобазальная длина черепа 26—29 мм. Окраска меха относительно
светлая п яркая с развитием рыжеватых и кирпичных тонов. Уссурийский край,
низовья Амура, Сахалин.
b) A. s. major Radde, 1862. (Описана из Буреинских гор.) Размеры несколько
мельче: длина тела 100—118 мм-, длина хвоста 83—96 мм\ кондилобазальная длина
295
черепа 24—27 мм. Окраска меха относительно темная, рыжевато-бурая. От Забай-
калья на запад до Алтая включительно. (Синонимы этой формы: A. s. majusculus
Turov, 1924, с Баргузинского хребта, A. s. tscherga Kastschenko, 1898, с Алтая.)
59. Лесная мышь. Apodemus sylvaticus Linnaeus, 1758
Табл. XXXI, 6. Карта 90
Европа (кроме северных окраин), Северная Африка, Западная и
Южная Азия. В СССР встречается почти по всей Европейской части
страны ((на север — до Центральной Карелии, южных районов Архан-
гельской области и Коми АССР), на Кавказе, в большей части Сред-
ней Азии и Казахстана (исключая некоторые пустынные районы) и
в южной части Западной Сибири, на восток — до Алтая включительно.
Населяют леса, заросли кустарников, поля, долины рек, овраги,
склоны гор, сады, огороды, усадьбы человека. Живут в неглубоких но-
рах. Наиболее активны ночью. За год дают 2—4 помета, из 3—8 дете-
нышей в каждом. Питаются желудями, орехами, семенами деревьев и
трав и другими частями .растений; поедают также насекомых. Местами
(например, на юге Украины) вредят посевам зерновых и лесным и са-
довым посадкам.
Географическая изменчивость лесной мыши выражена слабо, а индивидуальная
изменчивость, возрастные и сезонные морфизмы значительны. Описано большое число
подвидов, однако различия между ними очень слабые, достаточные в лучшем слу-
чае для выделения natio. Мы перечислим лишь часть географических форм, которые
можно различить при сравнениях серийных коллекций.
a) A. s. sylvaticus Linnaeus, 1758. (Описана пз Упсалы, Швеция.) Размеры отно-
сительно мелкие: ступня 18—22 мм. Окраска меха на спине от темно-бурой до охристо-
рыжеватой. Небольшое желтое пятно на груди встречается редко. Европейская часть
СССР (без Кавказа, Крыма, Южной Украины п Южного Урала). Синонимы номиналь-
ной формы: A. s. mosquensis Ognev, 1913 (описанная из окрестностей Москвы); A. s.
charkovensis Migulin, 1936; A. s. vohlinensis Charleman, 1936, и др.
b) A. s. uralensis Pallas, 1811. (Описана с Южного Урала.) Размеры, как у но-
минальной формы. Окраска спины менее затемненная черными окончаниями остевых
волос. Южный Урал, Зауралье, Западная Сибирь, Северный и Восточный Казахстан,
Северная Киргизия.
с) A. s. ciscaucasicus Ognev, 1924. (Тип пз окрестностей с. Орджоникидзе, Север-
ный Кавказ.) Близка к предыдущей, но окраска меха светлее и ярче, с преобладанием
рыжеватых тонов. Часто бывает желтоватое или охристое пятно на груди. Кавказ,
Крым, Южная Украина, Нижнее Поволжье, Западный Казахстан. Лесные мыши За-
кавказья относятся, видимо, к этой же форме.
d) A. s. chorassanicus Ognev et Heptner, 1922. (Тип из окрестностей г. Ашхабада.)
Окраска спины светлая, песчано-желтая. Иран, Копет-Даг и его предгорья. Отличия
от иранской формы A. s. arianus Blanford не выяснены.
е) A. s. pallipes Barret — Hamilton, 1900. (Тип из Сурхад-Бакап.а, Памир.) Окраска
меха тусклая, охристо-палевая без желтоватых и песчаных оттенков. Таджикистан и
горы Южной Киргизии.
60. Желтогорлая мышь. Apodemus flavicollis Melchior, 1834
Табл. XXXI, 7. Карта 89
Широко распространена в Западной Европе. В СССР населяет
западные, центральные и южные районы Европейской части страны и
Закавказье. Северо-восточная граница ареала идет через Ленинград-
скую, Калининскую, Смоленскую, Тульскую, Рязанскую и Горьковскую
области, Татарию и Башкирию к Южному Уралу. Южный предел аре-
ала тянется по Оренбургской и Уральской областям, по западным
пределам прикаспийских степей к предгорьям Кавказа. В степях Азер-
байджана местами отсутствует.
Живет в лесах (преимущественно лиственных и смешанных), в ку-
старнике, по балкам, в степи и в садах. Нередко селится в дуплах.
296
Хорошо лазает по деревьям. Дает ежегодно 2—3 помета, по 4—8 дете-
нышей в каждом. Питается, главным образом, семенами деревьев и
трав, но. поедает также насекомых. Причиняет вред, повреждая лес-
ные и садовые посадки и уничтожая орехи и желуди.
В средней и северной частях ареала видовые отличия желтогорлой мыши от
близкой к ней A. sylvaticus выступают очень отчетливо. Даже полувзрослых желто-
горлых мышей по размерам ступни и черепа легко отличить от взрослых лесных мы-
шей. Однако на юге Украины, включая Крым, различия между этими видами стано-
вятся менее ясными. Еще сложнее диагностика лесных мышей Кавказа и Закавказья.
П. А. Свириденко (1936) и Н. К- Верещагин (1959) выделяют на Кавказе 3 вида:
A. sylvaticus, A. fulvipectus, A. flavicollis. Большинство советских териологов-система-
тиков считают, что A. fulvipectus не вид, а только одна из форм изменчивости желто-
горлой, или обычной, лесной мыши. Но одни авторы (А. И. Аргиропуло, 1940, 1941;
Б. С. Виноградов, 1952) считают ее подвидом A. flavicollis, другие (как С. И. Огнев,
1924) —подвидом A. sylvaticus, третьи (как В. Г. Гептнер, 1940) считают A. fulvipectus
гибридами между лесной и желтогорлой мышами. М. В. Шидловский (1941) и
А. К. Тембатов (1960) считают мелких лесных мышей с желтыми и охристыми пят-
нами на груди одним из типов изменчивости обыкновенной лесной мыши A. sylvaticus.
Как видно, вопрос.этот очень сложный и ожидает обстоятельного изучения на боль-
шом фактическом материале.
Желтогорлых мышей в пределах Восточной Европы (исключая Кавказ) делят на
2 трудно различимые формы:
a) A. f. flavicollis Melchior, 1834. (Описана из Дании.) Окраска верха относи-
тельно яркая, рыжеватая. Прибалтийские республики, Белорусская ССР, Ленинград-
ская, Калининская, Смоленская области, Молдавия и Западная Украина.
b) A. f. samariensis Ognev, 1922. (Тип из Бузулукского бора Куйбышевской обла-
сти.) Окраска верха немного тусклее. Воронежская и Тамбовская области, все По-
волжье и возможно Восточная Украина.
61. Горная мышь. Apodemus mystacinus Danford et Alston, 1877
Табл. XXXI, S. Карта 90
Распространена на Балканах и в Малой Азии, в СССР встречается
в горных 1И лесных районах Западного Закавказья.
Живет на горных склонах, покрытых лесом и кустарником. Образ
жизни в пределах нашей страны изучен слабо. В Советском Союзе, по-
видимому, обитает типичная форма, описанная из Малой Азии.
РОД КРЫСЫ. GENUS RATTUS Fischer, 1803
Грызуны среднего размера (длина тела взрослых от 15 до 25 см)
с длинным хвостом, покрытым кольцами кожных чешуек, позади кото-
рых сидят редкие щетинки. Длина хвоста либо немного меньше, либо
больше длины тела. Окраска спины буроватая, рыжеватая или черная,
брюшка белая, серая или черная. Голова несет большие, почти голые
уши. Череп с удлиненной носовой частью, узкой .расстановкой скуло-
вых дуг и большой мозговой коробкой.
Распространены по суше всего земного- шара, кроме Антарктиды
и некоторых океанических островов. Особенно богато представлены
в фауне Юго-Восточной Азии. В СССР встречается 3 вида.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА НАСТОЯЩИХ КРЫС
ФАУНЫ СССР
1(4). Хвост длиннее тела. На хвосте более 200 чешуйчатых колец.
Ухо, отогнутое вперед, заходит за глаз. Плавательной перепонки
между пальцами задних ног нет. Боковые гребни теменных костей вы-
гнуты дугой кнаружи (рис. 109, А) .... •...........................2
2(3). Верхняя и нижняя стороны хвоста одноцветные или первая
лишь немного темнее второй. Передний край вырезки костного нёба
297
лежит значительно позади линии, соединяющей задние поверхности
последних коренных зубов ..........................................
.........................Черная крыса. Rattus rattus. (Стр. 298.)
3(2). Хвост сверху темный, снизу беловатый. Передний край вы-
резки костного нёба лежит примерно на линии, соединяющей задние
поверхности последних коренных зубов ..............................
....................Туркестанская крыса. Rattus rattoides. (Стр. 299.)
4(1). Хвост короче тела. На хвосте не более 200 чешуйчатых колец.
Ухо, отогнутое вперед, не достигает глаза. Между пальцами задних
ног имеются зачаточные плавательные перепонки. Боковые гребни те-
Рис. 109. Черепа крыс (сверху):
Л — черная крыса (Rattus rattus); Б — туркестанская крыса (R. rattoides);В — пасюк
(R. norvegicus).
менных костей почти прямые, параллельные друг другу или слегка
расходящиеся кзади (рис. 109, В)...................................
. ............................Пасюк. Rattus norvegicus. (Стр. 299.)
62. Черная крыса. Rattus rattus Linnaeus, 1758
Табл. XXXI, 9—10. Карта 91
Спорадически распространена во многих странах Европы, Север-
ной Африки и Азии. В СССР найдена в ряде мест Европейской части
страны, в Закавказье и на Дальнем Востоке. Обитает в постройках
человека и вне их, чаще всего в широколиственных лесах. Хорошо ла-
зает по деревьям. За год дает по 2—3 помета, в среднем по 6 детены-
шей. В домах наносит вред, уничтожая и портя продукты, а в садах и
огородах поедая плоды и овощи.
Встречается в виде двух цветовых форм: 1) черной, мех которой
на спине окрашен в черный или черно-бурый, а на брюхе в темно-се-
рый цвет, и 2) серой, окрашенной сверху в серо-коричневый, а снизу
в беловатый цвет. Количественное соотношение этих двух цветовых
форм различно в разных районах.
298
Подвиды фауны СССР:
a) R. г. rattus Linnaeus, 1758. (Описана из Швеции.) В популяции преобладают
черные особи, в окраске которых коричневые оттенки не выражены. Размеры мелкие:
кондилобазальная длина черепа 36—40 мм. В СССР, по-видимому, в Ленинградской
области и в Прибалтике.
b) R. г. ruthenus Ognev et Stroganov, 1936. (Тип из Ельнинского района Смо-
ленской области.) Преобладают черные особи. В окраске меха выступают коричневые
•оттенки. Размеры средние: кондилобазальная длина черепа 37—43 мм. Центральные
районы Европейской части СССР (форма очень близкая к номинальной).
с) R. г. alexandrinus Geofr., 1803. (Тип из Александрии, ОАР.) Преобладают серые
•особи. Размеры крупные: кондилобазальная длина черепа 40—47 мм. В СССР встре-
чается в портах Черного моря.
63. Туркестанская крыса. Rattus rattoides Hodgson, 1845
Табл. XXXI, 11. Карта 91
Распространена в Индии, Индокитае, Афганистане, Средней Азии,
Южном и Юго-Восточном Китае. В СССР встречается в Таджикистане,
Узбекистане, Южной и Западной Киргизии и в Южном Казахстане.
Живет как в поселениях человека, так и вдали от них — в лесах, в гор-
ных ущельях, по берегам рек. В последнем случае питается орехами,
плодами, ягодами, семенами, насекомыми, яйцами и птенцами * птиц.
Размножается 2—3 раза в году. Местами повреждает огородные и са-
довые культуры, а в домах и на складах портит и поедает продукты.
Распространяет возбудителя клещевого возвратного тифа.
В СССР обитает особая форма: R. г. turkestanicus Satunin, 1903.
64. Пасюк. Rattus norvegicus Berkenhout, 1769
Табл. XXXI, 12—13. Карта 92
Распространена почти по всему земному шару. Населяет значи-
тельную часть территории СССР, кроме Крайнего Севера, таежных
районов Сибири и Дальнего Востока, пустынь и полупустынь Казах-
стана и Средней Азии. В 40-х годах завезена в Ташкент, где укорени-
лась и широко расселилась по городу и окрестным селениям.
Живет обычно в. постройках человека или вблизи .от них, но не-
редко поселяется по берегам водоема и вдали от человеческих посе-
лений. Размножается в природных биотопах только в теплые месяцы,
а в постройках — круглый год. В каждом помете в среднем по 8 дете-
нышей. ‘За год самка приносит 2, реже 3 приплода. Питается самой
разнообразной, преимущественно животной и в меньшей степени расти-
тельной, пищей. Приносит огромный вред истреблением и порчей про-
довольственных продуктов. Основной источник чумы и некоторых дру-
гих опасных для человека заболеваний: сыпнотифозных лихорадок
(риккетсиозов), бешенства, эризипелоида, листереллеза, лептоспироз-
ной желтухи и т. д. Самый опасный и вредный грызун фауны СССР.
В СССР 2 подвида:
a) R. n. norvegicus Berkenhout, 1769. (Описан из Англии.) Хвост составляет от
70 до 96% длины тела. Длина ступни взрослых 36—44 мм. Окраска брюха постепенно
шереходит в окраску боков. Европейская часть СССР и Сибирь на восток до Байкала.
b) R. n. саггасо Pallas, 1778. (Описан из Забайкалья.) Хвост короче: длина его
•составляет 61—77% длины тела. Длина ступни 31—36 мм. Окраска брюха и бокоз
резко отграничены. Забайкалье, Дальний Восток.
РОД ПЛАСТИНЧАТОЗУБЫЕ КРЫСЫ. GENUS NESOKIA Gray, 1842
По внешнему облику похожи на обыкновенных крыс, но отли-
чаются коротким хвостом, длина которого обычно составляет 60—70%
длины тела. Среди меха, особенно на огузке, резко выделяются особо
299
длинные направляющие волосы. Морда более тупая. Череп значительно
массивнее, чем у настоящих крыс. Скуловые дуги расставлены широко.
Носовой отдел черепа широкий. Резцы также очень широкие. Ко-
ренные зубы взрослых особей с уплощенной жевательной поверхно-
стью, образованной вытянутыми в поперечном направлении оваль-
ными эмалевыми петлями: тремя на первом и двумя на следующих
зубах.
Распространены в Южной Азии и Северо-Восточной Африке.
Один вид.
65. Пластинчатозубая крыса. Nesokia indica Gray et Hardwicke, 1832
Табл. XXXI, 14. Карта 92
От низовьев Нила до Северо-Восточной Индии. В СССР встречается
в Средней Азии — в бассейнах рек Зеравшана, Аму-Дарьи, Мургаба,
Теджена и в горах Копет-Дага. Селится по берегам рек и арыков, в ту-
гаях по речным долинам, на полях люцерны и риса, на огородах и в са-
дах, в жилищах человека. Живет обычно большими колониями в раз-
ветвленных сложных норах. На поверхность земли выходит редко, и
главным образом ночью. Приносит несколько пометов в год, размно-
жаясь даже зимой.
Питается, главным образом, подземными частями травянистых ра-
стений, но нередко грызет и их стебли, листья и плоды. Сильно вредит
посевам люцерны и огородным культурам. Нередко разрушает ирри-
гационные сооружения.
В фауне СССР 2 слабо различающихся подвида:
a) N. i. huttoni Blyth, 1846. (Тип из окрестностей г. Кандагара, Афганистан.)
Размеры относительно крупные. Окраска меха темная. Бассейны Аму-Дарьи
и Зеравшана.
b) N. i. satunini Nehring, 4899. (Тип из г. Мары, Туркмения.) Размеры немного
меньше. Окраска меха светлее. Северные предгорья Копет-Дага, долины рек Мур-
габа и Теджена.
Подсемейство ПЕСЧАНОК- Subfamilia GERBILLINAE
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ПОДСЕМЕЙСТВА ПЕСЧАНОК
ФАУНЫ СССР
1(2). На передней поверхности каждого из верхних резцов по од-
ной продольной бороздке. Коренные зубы у взрослых особей имеют
корни. Последний коренной зуб верхней челюсти мал; сверху он несет
одну петлю округлой формы . Малые песчанки. Meriones. (Стр. 300.)
2(1). На передней поверхности каждого из верхних резцов имеется
по две продольных бороздки. Коренные зубы особей всех возрастных
групп без корней. Последний коренной зуб верхней челюсти относи-
тельно велик; его верхняя эмалевая петля изогнута ................
.........................Большие песчанки. Rhombomys. (Стр. 306.)
РОД МАЛЫЕ ПЕСЧАНКИ. GENUS MERIONES liliger, 1811
Ряд видов мелкого и среднего размера (величиной с крупную»
мышь или небольшую крысу). Хвост либо немного превышает, либо»
несколько короче длины тела, густо покрыт прилегающими к стержню
волосами, обычно образующими на конце кисточку. Характерные при-
знаки в строении зубов отмечены выше.
300
Распространены по зонам пустынь и полупустынь Старого Света
от Сахары до Северо-Восточного Китая; местами проникают из полу-
пустынь в сухие степи.
В фауне СССР — 8 видов.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА МАЛЫХ ПЕСЧАНОК
ФАУНЫ СССР
1(2). Подошвы задних ног (за исключением краев) голые. Кисть
удлиненных волос на хвосте развита очень сильно: она занимает почти
всю конечную треть хвоста. Вибриссы длинные — до 90 мм........
..............Персидская песчанка. Meriones persicus. (Стр. 303.)
2(1). Подошвы задних ног покрыты волосами сплошь или кроме
узкой срединной полоски. Кисть удлиненных волос на конце хвоста
занимает не более т/4 его длины. Вибриссы длиной менее 70 мм . . 3
3(6). Размеры мелкие: длина
задней ступни менее 31 мм\ кондило-
базальная длина черепа менее 34 мм.
Подошвы задних ног сплошь покрыты
густой шерстью.....................4
4(5). Окраска спины глинисто-
серая. Брюхо беловатое; его волосы
сероватые в основной своей части.
Когти темные. Слуховые капсулы от-
носительно невелики и никогда не ка-
саются скуловых дуг..................
. . . . . Когтистая песчанка.
Meriones unguiculatus. (Стр. 303.)
5(4). Окраска спины песчано-
желтая. Брюхо чисто белое: его воло-
сы белого цвета на всем протяжении.
Когти светлые. Слуховые капсулы
Б А
Рис. НО. Верхние резцы песчанок:
А — гребенчуковая песчанка (Meriones ta-
mariscinus) из рода малых песчанок; Б —
большая песчанка (Rhombomys opimus).
очень большие и касаются скуловых дуг . . .....................
.............Полуденная песчанка. Meriones meridianus. (Стр. 303.)
6(3). Размеры крупнее: длина задней ступни более 31 мм; конди-
лобазальная длина черепа более 34 мм. Подошвы задних ног покрыты
шерстью, но с узкой полоской голой кожи, идущей от пятки до сере-
дины подошвы или основания пальцев..............................7
7(10). Брюхо сероватое; конечные части покрывающих его волос
белые, а основные — серые...................................8
8(9). Подошвы задних лап покрыты желто-серой шерстью. Слухо-
вые капсулы перед слуховым проходом образуют заметные вздутия
............Краснохвостая песчанка. Meriones libycus. (Стр. 304.)
9(8). Подошвы задних лап покрыты шерстью интенсивно охристого
цвета, сгущающегося у большинства особей позади мозолей в рыжее
пятно. Вздутий слуховой капсулы перед слуховым проходом нет . . .
...........Песчанка Виноградова. Meriones vinogradovi. (Стр. 304.)
10(7). Брюхо чисто белого цвета: его волосы на всем протяжении
белые......................................................11
11(14). Шерсть на подошвах задних ног светлая. Окраска хвоста
сверху и снизу различается слабо................................12
12(13). Слуховые капсулы велики; их наибольшая длина (без со-
сцевидной кости) значительно превышает длину шва между лобными
костями. Перед трубкой слухового прохода в черепе нет впадины
(рис. 112) . . . Песчанка Зарудного. Meriones zarudnyi. (Стр. 305.)
301
13(12). Слуховые капсулы невелики; их наибольшая длина (без.
сосцевидной кости) равна или лишь слегка превышает длину шва ме-
жду лобными костями. Перед трубкой слухового прохода имеется впа-
дина черепа (рис. 112)................•...........................
. . Малоазиатская песчанка. Meriones blackleri. (Стр. 305.)
Рис. 111. Черепа песчанок (вид снизу):
А — краснохвостая песчанка (Meriones libycus); Б — песчанка Виноградова
(М. vinogradcvi).
Рис. 112. Черепа песчанок (вид снизу):
.4 — песчанка Зарудного (Meriones zarudnyi); Б — малоазиатская
песчанка (М. blackleri).
14(11). Подошвы задних ног в средней своей части одеты темно-
бурой шерстью, а по бокам светлыми волосами. Хвост резко двухцвет-
ный: сверху буроватый, снизу почти белый. Кисточка волос на конце
хвоста зачаточна .................................................
. . . . Гребенчуковая песчанка. Meriones tamariscinus. (Стр. 305.)
332
66. Персидская песчанка. Meriones persicus Blanford, 1876
Табл. XXXII, 1. Карта 93
Иран и прилегающие части Турции, Афганистана и Белуджистана.
В СССР найдена в горах Копет-Дага и Больших Балханов, в Туркме-
нии и в Закавказье (Южная Армения, Нахичеванская АССР, Кара-
бах, Талыш и др.). Обитатель горных пустынь. Живет небольшими ко-
лониями или поодиночке. Ночной зверек. Дает 2 (возможно и 3) по-
мета в год. Питается горно-пустынными растениями.
В СССР встречается 2 очень слабо различающихся подвида:
а) М. р. suschkini Kaschkarov, 1925. (Тип с Больших Балханов.) От номинальной
формы (из Ирана) отличается менее яркой окраской и меньшими размерами черепа.
Горы Туркмении.
b) М. р. rossicus Heptner, 1931. (Тип из окрестностей г. Еревана.) Отличается от
туркменской и иранской форм желтоватой (а не белой) окраской нижней поверхности
хвоста. Южное Закавказье.
67. Когтистая песчанка. Meriones unguiculatus Milne-Edwards, 1867
Табл. XXXII, 4. Карта 93
Широко распространена в Монголии, Северном и Северо-Восточ-
ном Китае. В СССР водится ъ сухих степях Южного Забайкалья и на
юге Тувинской области. В Забайкалье найдена только в долине р. Се-
ленги, у монгольской границы, и в районе оз. Торей.
В этих местах О'на селится на участках сухой песчанистой степи.
Живет .колониями в просто устроенных норах. Летом питается, глав-
ным образом, зелеными частями травянистых растений, осенью и зи-
мой преимущественно их семенами, из которых делает запасы на зиму.
По-видимому, приносит 2 помета в год из 3—8 детенышей. В Монголии
относится к числу распространителей чумы.
В СССР представлена 2 слабо различающимися формами:
а) М. u. kozlovi Satunin, 1903. (Тип с низовьев р. Кобдо, в Монголии.) Длина тела
до 14 см. Окраска спины относительно светлая. Юг Тувинской области.
b) М. u. selenginus Heptner, 1949. (Тип из окрестностей г. Кяхты.) Размеры
немного крупнее: длина тела до 15 см. Окраска темная, глинистая. Южное Забайкалье.
68. Полуденная песчанка. Meriones meridianus Pallas, 1773
Табл. XXXII, 2—3. Карта 93
Распространена в Восточном Предкавказье, Нижнем Поволжье,
Казахстане (от низовьев Урала, иргизских степей и Аральских Кара-
кумов до Муюнкумов и прибалхашских песков включительно), в рав-
нинных частях Средней Аз-ии, Северном Афганистане, Северо-Восточ-
ном Иране, пустынях Центральной Азии, степях Тувинской области,
песчаных массивах Армении и Нахичевани.
Обитает преимущественно среди песков различной степени зара-
стания. Живет в одиночных постоянных -и временных норах. Активна
ночью и в сумерки. Питается пустынными травами и их семенами.
2—3 помета в году. Один из основных микробоносителей чумы.
Изредка вредит посевам, расположенным среди песков.
В пределах СССР встречаются следующие подвиды:
а) М. m. nogaiorum Heptner, 1927. (Тип из окрестностей ст. Терекли-Мектеб,
100—120 км к северо-западу от Кизляра.) Размеры относительно крупные: кондилоба-
зальная длина черепа в среднем около 30 мм. Окраска темная, желтовато-буроватая.
Полупустыни Восточного Предкавказья и Калмыкии.
b) М. ш. meridianus Pallas, 1773. (Описана из Рын-Песков между Волгой и Ура-
лом.) Размеры немного меньше: кондилобазальная длина черепа в среднем около
28 мм. Окраска тусклая, песчано-серая. Волжско-уральские пески.
303
с) М. m. karelini Kolossov, 1935. (Тип из района устья р. Эмбы.) Размеры, как
у номинальной формы. Окраска спины темная с преобладанием буровато-серых
тонов. Низовья Эмбы и Урала.
d) М. m. massagetes Heptner, 1933. (Тип из окрестностей г. Аральска.) Размеры
чуть крупнее, чем у типичной формы. Окраска спины очень светлая, песочная. Хвост
короткий без черной кисточки на конце. Пески Приаралья.
е) М. m. penicilliger Heptner, 1933. (Тип из окрестностей ст. Репетек, Восточные
Каракумы.) Размеры, как у предыдущей формы. Окраска спины светлая и бледная.
Хвост более длинный с черной кисточкой на конце. Каракумы, Кызылкумы, Муюн-
кумы, пески Прибалхашья.
f) М. m. zhitkovi Heptner, 1933. (Тип из окрестностей г. Коканда.) Размеры, как
у двух предыдущих форм. Окраска спины темная, желтовато-серая. Пески Централь-
ной Ферганы.
g) М. m. urianchaicus Vinogradov, 1927. (Тип из окрестностей Икиоттун, в Тувин-
ской АССР.) Наиболее крупная и темная форма. Тувинская область.
h) М. m. dahli Schidlovskyi, 1955. (Тип из Нахичеванской АССР.) Окраска тем-
ная, тусклая. Нахичевань и Армения.
69. Краснохвостая песчанка. Meriones libycus Lichtenstein, 1823
Табл. XXXII, 5. Карта 94
Пустыни Северной Африки, Передней и Средней Азии. В СССР
встречается спорадично по равнинам и предгорьям Средней Азии и
Южного Казахстана (к северу до низовьев р. Урала, Устюрта, Араль-
ских Каракумов, среднего течения р. Сарысу и Прибалхашья); в пес-
ках Каракумов и Кызылкумов, вдали от речных долин и поселений
человека отсутствует.
Поселяется в различных местах, особенно охотно, на культурных
землях и близ поселений человека. Больших массивов сыпучих песков
избегает. Живет колониями, причем норы отдельных особей нередко
сливаются в общую систему подземных ходов. Жизнедеятельна как
ночью, так и днем. Приносит несколько (обычно 2—3) пометов в год,
по 3—7 детенышей в каждом. Пища разнообразна. На зиму делает
большие запасы. Местами наносит сильный вред хлебным полям, по-
севам хлопчатника, бахчам и садам. Способствует распространению
чумы и является основным источником сезонного (кожного) лейшма-
ниоза. Типичная форма описана из Новой Александрии, в Египте.
В СССР различают следующие подвиды:
а) М. 1. eversmanni Bogdanov, 1873. (Описана из окрестностей порта Шевченко,
на восточном берегу Каспия.) Размеры относительно мелкие: кондилобазальная длина
черепа в среднем равна 34,6 мм. Окраска тусклая, серовато-песчаная. Западный и
Южный Казахстан, Устюрт.
b) М. 1. maxeratis Heptner, 1933. (Тип с р. Чандыр, в Копет-Даге.) Размеры не-
много крупнее. Окраска спины ярче. Копет-Даг, Большие Балханы. Форма очень близ-
кая к предыдущей. К ней относится описанная с Мургаба М. 1. marginiae Hept-
ner, 1933.
с) М. 1. oxianus Heptner, 1933. (Тип из Гузара, к югу от Карши.) Размеры те же.
Окраска спины светлее с преобладанием желтоватых оттенков. Южный Узбекистан.
d) М. 1. sogdianus Heptner, 1933. (Тип из района г. Коканда.) Размеры относи-
тельно крупные: кондилобазальная длина черепа в среднехм 37,1 мм. Окраска спины
темная, буровато-серая. Фергана.
е) М. 1. caucasicus Brandt, 1855. (Тип из окрестностей г. Сальян.) Размеры мелкие:
кондилобазальная длина черепа в среднем 34,6 мм. Окраска очень темная, сероватая.
Восточное Закавказье.
70. Песчанка Виноградова. Meriones vinogradovi Heptner, 1931
Табл. XXXII, 6, Карта 94
Форма, близкая к краснохвостой песчанке. В СССР найдена только
в Нахичеванской АССР и Юго-Восточной Армении. Известны места
нахождения ее в Северо-Западном Иране и Северо-Восточной Турции.
304
Обитатель солянковых полупустынь и прилегающих залежей. Живет
колониями. Образ жизни изучен слабо.
71. Песчанка Зарудного. Meriones zarudnyi Heptner, 1927
Табл. XXXII, 7. Карта 95
Пустыни Ирана и Афганистана. В СССР найдена только на юге
Туркмении.
Биология не изучена.
Требуется выяснение на коллекционном материале отличий этой формы от
Meriones crassus Sundevall, 1842, описанной из Синая и широко распространенной
в Северной Африке, Аравии, Иране, Ираке и Афганистане. Особенно неясны отличия
песчанки Зарудного от Meriones crassus swinhoei Scully, 1881, описанной из Афга-
нистана.
72. Малоазиатская песчанка. Meriones blackleri Thomas, 1903
Табл. XXXII, 8. Карта 95
Турция, Сирия, Иран и Закавказье. В пределах последнего най-
дена в Южной Армении, в Нахичеванской АССР, по р. Куре на. про-
тяжении от Тбилиси до Евлаха, в Карабахе, в Мильской и Муганской
степях, в западной части предгорьев Талыша.
Селится преимущественно в возвышенной глинистой полынной
степи и полупустыне на хрящевато-песчаных почвах. Норы обычно рас-
положены поодиночке на курганах, в развалинах, на кладбищах, по
насыпям .арыков и т. п. Деятельна днем и ночью. Самка дает несколько
пометов в год. Питается пустынными растениями и их семенами. Ме-
стами вредит посевам.
В СССР — М. b. bogdanovi Heptner, 1930. (Тип из Сальянского района, в Азер-
байджане.) Отличается от номинальной формы, описанной из Смирны и широко рас-
пространенной в Турции и Иране, меньшими размерами и более темной окраской.
73. Гребенчуковая песчанка. Meriones tamariscinus Pallas, 1773
Табл. XXXII, 9. Карта 95
Населяет полупустыни и пустыни Восточного Предкавказья, Кал-
мыкии, Нижнего Поволжья, Среднего и Южного Казахстана, ряд райо-
нов Средней Азии и Джунгарии. К северу распространена примерно
до Элисты, Енотаевска,-Калмыкова, Джамбейтинского района, г. Актю-
бинска, пос. Тургая, оз. Аккуль и Чубар-Тениз, области нижнего те-
чения р. Сарысу, центральной части пустыни Бет-Пак-Дала, Северного
Прибалхашья и котловины оз. Алаколь; найдена в долине оз. Зайсан;
на юг до песков Каракум, Самаркандского и Бухарского оазисов, Фер-
ганской долины; водится в котловине оз. Иссык-Куль, в некоторых до-
линах гор Киргизии.
Селится среди закрепленных или зарастающих-песков, на участках
глинистой пустыни, в зарослях гребенчука и саксаула; охотно дер-
жится на орошаемых участках посевов. Норы одиночные или реже
колониальные. Ведет преимущественно дневной образ жизни. Дает еже-
годно 2—3 помета из 4—7 детенышей. Питается различными пустын-
ными травами. Местами вредит посевам хлебов, проса, хлопчатника.
Различают следующие подвиды:
а) М. t. ciscaucasicus Satunin, 1907. (Тип из окрестностей ст. Червленной, на Те-
реке.) Окраска спины темная с преобладанием бурых оттенков. Предкавказье.
b) М. t. tamariscinus Pallas, 1773. (Описан из окрестностей ст. Сарайчиковской,
20 н. А. Бобринский и др. 205
на р. Урале.) Окраска светлее с примесью красноватых тонов. Волго-уральские и
урало-эмбинские полупустыни.
с) М. t. jaxartensis Ognev et Heptner, 1928. (Тип из окрестностей ст. Кара-Узяк,
близ г. Кзыл-Орды.) Окраска еще более светлая, красновато-песчаная, без примеси
коричневых оттенков. Пустыни Средней Азии и Южного Казахстана.
d) М. t. kokandicus Heptner, 1933. (Тип из окрестностей Коканда.) Окраска спины
темная, красновато-орехового цвета. Пески Ферганы.
РОД БОЛЬШИЕ ПЕСЧАНКИ. GENUS RHOMBOMYS Wagner, 1841
Крупные песчанки: длина тела их достигает 20 см, а хвоста — 18 см.
Летний мех редкий, грубый, ломкий; зимний густой, мягкий. Спина
песчано-желтая разной темноты, зависящей, главным образом, от сте-
пени затемнения черноватыми окончаниями остевых волос. Брюхо бе-
лое 'Или сероватое нередко с желтоватым оттенком. Отличия в строении
зубов указаны в таблице определения родов песчанок.
Один вид.
74. Большая песчанка. Rhombomys opimus Lichtenstein, 1823
Табл. XXXII, 10—11. Карта 95
Распространена от берегов Каспийского моря по равнинам Сред-
ней Азии и Южного Казахстана и пустыням Центральной Азии. Се-
верная граница ареала в нашей стране тянется от устьев рек Урала и
Эмбы, севернее г. Челкара, через окрестности г. Иргиза, по северной
окраине Приаральских Каракумов, низовье р. Сарысу, по Бет-Пак-
Дала, по южному берегу Балхаша. На юг распространена по Ирану и
Афганистану, на восток — до Северного Китая и Внутренней Монголии.
Типичный обитатель песчаных пустынь, где местами весьма много-
численна, но нередко селится и среди глинистой пустыни. Поднимается
в предгорья гор Средней Азии. Живет обычно колониями. Норы очень
сложного строения с большим числом (до 200—500) выходов. Ведет
дневной образ жизни. На зиму в спячку не впадает. Дает 2 или 3 по-
мета в год, из 2—8 (обычно 4—5) детенышей в каждом. Питается со-
лянками, песчаной осокой, вейником и другими пустынными растени-
ями. На зиму делает большие запасы, главным образом из веточек
саксаула. В некоторых районах вредит посевам, люцерны и других
культур. Местами наносит вред пескоукрепительным посадкам кустар-
ников. Портит насыпи дорог и стенки арыков. Известен в эпидемиоло-
гии и эпизоотологии как основной микробоноситель чумы в равнинно-
пустынных очагах и один из главных источников сезонного (кожного)
лейшманиоза.
В пределах СССР — 2 подвида:
a) Rh. op. opimus Lichtenstein, 1823. (Местом описания считают Приаральские
Каракумы.) Окраска светлая, песчано-желтая. Равнины Южного Казахстана, Турк-
мении и Узбекистана, кроме Ферганы.
b) Rh. op. fumicolor Heptner, 1933. (Тип из окрестностей г. Коканда.) Окраска тем-
ная, сероватая. Пески Ферганской долины.
Подсемейство ХОМЯКОВ. Subfamilia CRICETINAE
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ПОДСЕМЕЙСТВА ХОМЯКОВ
ФАУНЫ СССР
1(2). Хвост равен или длиннее тела. На конце хвоста имеется ки-
сточка удлиненных волос. Защечные мешки зачаточны. Сосков 3 пары.
Передняя петля первого верхнего коренного зуба не подразделена на
две части..................Мышевидные хомячки. Calomyscus. (Стр. 307.)
306
2(1). Хвост не длиннее 72 длины тела. Кисточки удлиненных волос
на конце хвоста нет. Защечные мешки развиты хорошо. Сосков не ме-
нее 4 пар. Передняя петля первого коренного зуба верхней челюсти
подразделена на две — левую и правую части (рис. 113)...........3
3(4). Вся нижняя сторона тела или только грудная область по-
крыта черной или черноватой шерстью. Сосков от 7 до 11 пар. На
верхней поверхности черепа взрослых зверьков хорошо выражены
гребни.....................Пестрые хомяки. Cricetus. (Стр. 307.)
4(3). Нижняя поверхность тела белая или светло-серая, иногда
с коричневым или желтым пятном на груди. Сосков 4 пары. На верх-
ней поверхности черепа гребней нет или
они выражены слабо....................5
5(6). Подошвы задних лап покрыты
густым мехом. Длина хвоста менее 14 мм.
Наружные углы коренных зубов верхней
челюсти лежат против соответствующих
внутренних углов .......................
Мохноногие хомячки. Phodopus. (Стр. 309.)
6(5). Подошвы задних лап полностью
или частично голые. Длина хвоста более
14 мм. Наружные углы коренных зубов
верхней челюсти лежат против выемок их
внутренней стороны ....................... .
. . . . Хомячки. Cricetulus. (Стр. 310.)
Рис. 113. Первый верхний ко-
ренной зуб хомячков:
Л — мышевидный хомячок (Calo-
myscus bailwardi); Б — серый хо-
мячок (Cricetulus migratorius).
РОД МЫШЕВИДНЫЕ ХОМЯЧКИ. GENUS CALOMYSCUS Thomas, 1905
Резко отличаются от других палеарктических хомяков хвостом,,
равным или превышающим длину тела и покрытым длинными воло-
сами, образующими на конце хвоста хорошо выраженную кисточку.
Ступни задних ног длинные и узкие. Уши велики. Бросаются в глаза
очень длинные вибриссы, доходящие своими концами до переднего*
края бедер. Сосков 3 пары. Защечные мешки рудиментарны. Череп
плоский с большой мозговой коробкой и узкой расстановкой скуло-
вых дуг.
Только один вид.
75. Мышевидный хомячок. Calomyscus bailwardi Thomas, 1905
Табл. XXXIII, /, 2. Карта 96
Водится в Белуджистане, Афганистане, Иране. В нашей стране
найден, с одной стороны, в Туркмении (в горах Копет-Дага и Больших
Балханов), а с другой — в Закавказье (Нахичеванская АССР).
Встречается по склонам гор (от их предгорий до значительных вы-
сот). Обитает среди скал, по осыпям крупных камней, в ущельях, пе-
щерах. Образ жизни изучен слабо.
В СССР встречается, видимо, номинальная форма: С. b. bailwardi Thomas, опи-
санная из Ирана; к ней относится и С. b. mystax Kaschkarov, 1925, с Больших Балханов.
РОД ПЕСТРЫЕ ХОМЯКИ. GENUS CRICETUS Leske, 1779
Грызуны среднего размера с массивным, несколько мешкообраз-
ным телом, коротким, конической формы хвостом и относительно корот-
кими конечностями. Голова несет большие тонкие округлые ушные ра-
ковины. Мех на спине и боках относительно высокий и густой, на брюхе
20* ЗОГ
редкий, прилегающий. Верх тела ржаво-буроватый или охристо-бурый,
брюхо черное или сероватое с черным пятном на груди.
Обитатели Европы и Западной Азии. В СССР — 3 вида, которые
относят к двум подродам (Cricetus и Mesocricetus).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ПЕСТРЫХ ХОМЯКОВ
ФАУНЫ СССР
1(2). Верхняя сторона тела ржаво-буроватая или ржаво-желтая,
нижняя черная или черно-бурая. Иногда попадаются особи черные и
черные с белыми пятнами. На боках головы и туловища с каждой сто-
роны выступают по 3 белых пятна. Нижняя часть скулового отростка
верхнечелюстной кости образует широкую пластинку, частично при-
крывающую подглазничное отверстие сбоку (рис. 114)...............
............ . . Хомяк обыкновенный. Cricetus cricetus. (Стр. 308.)
2(1). Окраска иная. Белых пятен по бокам головы и тела нет.
Нижняя ветвь скулового отростка верхнечелюстной кости не образует
пластинки, вдающейся в подглазничное отверстие ... . . . 3
Рис. 114. Скуловые дуги хомяков (вид сбоку)-:
А — хомяк обыкновенный (Cricetus cricetus); Б — предкавказский
хомяк (С. raddei); не — нижняя ветвь скулового отростка верхне-
челюстной кости.
3(4). Нижняя сторона тела на горле и на груди черная, а на брюхе
буроватая. Гребни на верхней стороне черепа взрослых особей сильно
развиты. Общая длина черепа обычно более 37 мм..................
.................Предкавказский хомяк. Cricetus raddei. (Стр. 309.)
4(3). Нижняя сторона тела сероватая с черным пятном или попе-
речной полосой на груди. Гребни на верхней стороне черепа взрослых
«особей выражены слабо. Общая длина черепа менее 37 мм..........
_.............Переднеазиатский хомяк. Cricetus auratus. (Стр. 309.)
Подрод хомяков обыкновенных. Subgenus Cricetus
Linnaeus, 1758
76. Хомяк обыкновенный. Cricetus cricetus Linnaeus, 1758
Табл.. XXXI1I,3,4. Карта 96
Европа, степи и лесолуговая зона Западной Сибири, Северного и
Восточного Казахстана. В Европейской части СССР идет к югу до
Крымских и Кавказских гор, а к северу — до Ярославля, Горького и
Кирова. В Западной Сибири — к северу до южной границы тайги, на
восток — до Енисея.
Живет в степи, на пашнях, бахчах, огородах, в садах, по берегам
рек, в степных балках, по опушкам леса. Живет поодиночке в глубоких
норах с многими выходами. Зиму проводит в спячке (хотя и не столь
глубокой, как суслики). Ведет преимущественно ночной образ жизни.
За лето дает обычно 2 помета, из 6—8 (до 18) детенышей в каждом.
Питается как растительной (зерно, корнеплоды, зеленые части расте-
ний и др.), так и животной пищей (насекомые, мелкие позвоночные).
308
На зиму делает в норе большие запасы зерна, семян и корнеплодов.
Сильно вредит растениеводству ряда степных районов страны. Шкурку
используют в качестве пушного сырья; ежегодно добывается от 2 до
11 млн. шкурок.
Различными авторами было описано много подвидов хомяков, обитающих в Со-
ветском Союзе. Но сравнение серий этих зверьков из разных районов страны убеж-
дает, что большая часть описанных форм реально не существует. Необходим крити-
ческий пересмотр всей внутривидовой систематики этого вида.
Подрод средних хомяков. Subgenus Mesocricetus
Nehring, 1898
77. Предкавказский хомяк. Cricetus raddei Nehring, 1894
Табл. XXXIII, 5. Карта 96
Обитает на северных и восточных склонах и предгорьях Главного
Кавказского хребта и в степях Предкавказья на север до низовьев
Дона и Маныча. Селится в равнинных степях, на травянистых увалах
предгорьев, среди участков горной степи, на пашнях, по бахчам, степ-
ным балкам. Живет -в глубоких и сложно устроенных норах, из кото-
рых выходит обычно только ночью. Зиму проводит в спячке. Размно-
жается два раза в год. Местами вредит полеводству и огородничеству.
Различают 2 подвида этого хомяка:
а) С. г. raddei Nehring, 1894. (Тип с р. Самур.) Размеры крупные: длина тела
взрослых 18—28 см. Дагестан. (К этой же форме относится С. г. avaricus Ognev et
Heptner, 1927.)
b) C. r. nigriculus Nehring, 1894. (Тип с p. Малки.) Размеры мельче: длина тела
15—20 см. Северные склоны Кавказа и степи Предкавказья.
78. Переднеазиатский хомяк. Cricetus auratus Waterhouse, 1839
Табл. XXXIII, 6. Карта 96
Широко распространен на Балканах, в Турции, Иране, Сирии, Ли-
ване. В СССР обитает в Закавказье (Армения, окрестности г. Тбилиси,
Ширванская степь, Нахичеванская АССР, Карабах, Талыш).
Селится, главным образом, по сухим безлесным нагорьям (до вы-
соты 2300 ;и), на пашнях, огородах. Глубокие норы его имеют обычно
1—3 входных отверстия. Ежегодно самка дает 2 помета, по 11 —15
детенышей в каждом. Летом поедает траву, семена, зерна, корни,
клубни. На зиму делает запасы. Приносит вред зерновым.
В Закавказье водится С. a brandti Nehring, 1894, описанная по экземпляру из
Центральной Грузии. (Ее синоним: С. a. koenigi Nehring, 1900. Описан из Армении.)
РОД МОХНОНОГИЕ ХОМЯЧКИ. GENUS PHODOPUS Miller, 1910
Мелкие зверьки (длина тела до 10 см), покрытые пышным мягким
мехом. Хвост едва выступает из меха. Подошвы задних ног сплошь
покрыты густой шерстью. Распространены в степях и пустынях Цент-
ральной Азии, в некоторых районах Южной Сибири и Восточного
Казахстана. 2 вида.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА МОХНОНОГИХ
хомячков
1(2). На спине вдоль хребта тянется ясно заметная черноватая
продольная полоска. Буровато-серая или желтоватая окраска меха
спины тремя выступами спускается вниз, вклиниваясь в белую рас-
цветку брюха . Джунгарский хомячок. Phodopus sungorus. (Стр. 310.)
309
2(1). На спине темной продольной полосы нет. Желтовато-розовая
или желтовато-серая расцветка спины граничит с белой окраской
брюха прямой линией ...........................................
...........Хомячок Роборовского. Phodopus roborovskii. (Стр. 310.)
79. Джунгарский хомячок. Phodopus sungorus Pallas, 1770
Табл. XXXIII, S. Карта 97
Широко распространен в степях и полупустынях Центральной
Азии. В СССР обитает в сухих степях Южного Забайкалья, ряда рай-
онов Южной Сибири и Северо-Восточного Казахстана.
Обитает по сухим целинным степям, на выгонах, пастбищах, паш-
нях. Живет в неглубоких норках. Размножается несколько раз в год,
в помете обычно 5—6 детенышей. В целинной степи питается, главным
образом, семенами различных трав. На пашнях вредит посевам.
В СССР встречается 2 формы:
а) Р. s. sungorus Pallas, 1770. (Описана из барабинских степей.) Окраска летнего
меха спины довольно темная, серовато-буроватая, сильно затемненная черными кон-
цами остевых волос, на плечах темные пятна. Низ тела чисто-белый. На зиму пол-
ностью белеет. Степи Южной Сибири и Северо-Восточного Казахстана.
b) Р. s. campbelli Thomas, 1905. (Тип из Монголии.) Окраска летнего меха на
спине светлая, сероватая без резко выраженных темных пятен на плечах. Брюшко
сероватое. На зиму мех белеет только частично. Монголия, Южное Забайкалье, Тува,
Чуйская степь (на Алтае).
80. Хомячок Роборовского. Phodopus roborovskii Satunin, 1903
Табл. XXXIII, 8. Карта 97
Распространен в пустынях Центральной Азии. В Советском Союзе
обнаружен только на юге Тувинской области. Обитает среди песков.
Норки неглубокие, просто устроенные. Питается как животной (насе-
комые), так и растительной пищей (ягоды селитрянки, семена различ-
ных трав и т. п.). Размножение не изучено.
РОД ХОМЯЧКИ. GENUS CRICETULUS Milne-Edwards, 1867
Мелкие и средних размеров хомяки с относительно длинным хво-
стом (длина которого колеблется от 7s до Vs длины тела), короткими
ступнями задних ног, подошвы которых полностью или частично голые.
Бугорки на подошве задних ног хорошо развиты. Межтеменная кость
черепа сильно вытянута поперек его оси.
Широко распространены в Европе и Азии. В СССР — 5 видов
(3 подрода).
« ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА ХОМЯЧКОВ ФАУНЫ СССР
1(2). На груди имеется буроватое или песчано-желтое пятно,
•остальные части нижней поверхности тела белые. Хвост короткий, рав-
ный лишь !/4—Vs длины тела. Межтеменная кость сильно вытянута по-
перек оси черепа; ее продольная длина равна д/4—Vs ее ширины . . .
...........Хомячок Эверсманна. Cricetulus eversmanni. (Стр. 311.)
2(1). На груди темного пятна нет; вся нижняя поверхность тела
белая. Хвост более длинный: длина его равна т/4—V2 длины тела. Меж-
теменная кость менее сильно вытянута в бока: ее продольная длина
равна примерно 7з ее ширины........................................3
3(8). Размеры относительно мелкие: длина тела менее 14 см\ кон-
дилобазальная длина черепа менее 35 жж; длина хвоста менее Vs длины
310
тела (менее 45 мм). Он двухцветный (сверху темный, снизу светлый)
или весь светлый. По бокам межглазничного промежутка черепа греб-
невидных оторочек нет.............................................4
4(5). На спине вдоль хребта тянется ясно выраженная черная по-
лоса ..........Даурский хомячок. Cricetulus barabensis. (Стр. 311.)
5(4). Черной продольной полосы на спине нет (иногда заметно не-
большое продолговатое темное пятно на загривке) ................. 6
6(7). Ухо темное с белой каймой. Длина хвоста равна или превы-
шает 7з длины тела. Хвост сверху темный, снизу светлый. Передне-
наружные углы теменных костей вытянуты в острые отростки ....
. . . Длиннохвостый хомячок. Cricetulus longicaudatus. (Стр. 312.)
7(6). Ухо без белой каймы. Длина хвоста менее 7з длины тела.
Хвост обычно весь светлый. Передне-наружные углы теменных костей
не вытянуты в острые отростки.......................................
..................Серый хомячок. Cricetulus migratorius. (Стр. 312.)
8(3). Размеры более крупные: длина тела взрослых более 15 см;
кондилобазальная длина черепа более 35 мм; длина хвоста примерно
равна V2 длины тела (более 50 мм). Он темный, обычно со светлым
концом. По бокам межглазничного промежутка черепа идут гребневид-
ные оторочки.......................‘................................
............. . Крысовидный хомяк. Cricetulus triton. (Стр. 313.)
Подрод короткохвостых хомячков. Subgenus
Allocricetulus Argyropulo, 1933
81. Хомячок Эверсманна. Cricetulus eversmanni Brandt, 1859
Табл. XXXIV, 1. Карта 97
Степи Заволжья, Южного Урала, Волго-Уральского междуречья.
Северного Казахстана и Монголии.
Обитает в целинной степи, в степных балках, на выгонах, пашнях
и бахчах, в поселениях человека. Живет в глубоких норках. Зиму про-
водит в спячке. Дает 2 помета в год из 3—7 детенышей. Питается
травянистыми растениями, их семенами, зернами хлебов и другой рас-
тительной пищей; поедает также насекомых и моллюсков. На зиму
делает большие -запасы. Вредитель зерновых и бахчевых культур.
По исследованиям И. П. Митиной, в пределах СССР встречается 2 подвида
хомячка Эверсманна:
а) С. е. eversmanni Brandt, 1859. (Тип из окрестностей г. Оренбурга.) Спина тем-
ная, буровато-коричневая, брюхо белое; граница этих двух цветовых зон резкая. На
груди широкая полоса бурого меха. Куйбышевская, Оренбургская области, Башкирия,
северные районы Западно-Казахстанской и Актюбинской областей, Челябинская,
Курганская, Кустанайская, Северо-Казахстанская области.
b) С. е. beljaevi Argyropulo, 1933. (Тип из котловины оз. Зайсан.) Спина более
светлая, песчано-палевая. Ее окраска постепенно переходит в белую расцветку брюха.
Пятно на груди желтоватое, бледное. Волжско-уральские степи, степи по р. Эмбе,
Актюбинская, Кзыл-Ординская, Карагандинская, Семипалатинская, Восточно-Казах-
станская области.
В /Монголии — особый подвид: С. е. curtatus.Allen.
Подрод серых хомячков. Subgenus Cricetulus
Milne-Edwards, 1867
82. Даурский хомячок. Cricetulus barabensis Pallas, 1773
Табл. XXXIV, 2. Карта 98
Степи и степные участки лесостепи южной полосы Сибири, За-
байкалья, южных районов Дальнего Востока, Монголии и Северного
311
Китая. В СССР распространен на юге Приморья, в долине среднего
течения р. Амура, в степях Забайкалья (к северу до Сретенска —
Читы — Улан-Удэ), в Прибайкалье к северу до верховьев р. Лены,
в Тувинской котловине, предалтайских степях, Барабинской лесо-
степи.
Обитает среди целинных степей, пашен, по опушкам леса, в доли-
нах степных рек, в поселениях человека. Сложные норки зверька имеют
по 1—5 входных отверстий. В спячку не впадает (южные части ареала),
или зимний сон* длится недолго (северные районы ареала). За лето
приносит несколько пометов из 4—11 детенышей. Питается семенами,
зелеными частями трав и насекомыми. Местами вредит посевам зер-
новых.
Образует несколько подвидов:
а) С. b. barabensis Pallas, 1773. (Тип из окрестностей г. Барнаула.) Наиболее тем-
ная форма. Спина ржаво-бурая. Ремень черный, резко выступающий. Степи Запад-
ной Сибири.
b) С. b. griseus Milne-Edwards, 1867. (Тип из окрестностей г. Пекина.) Окраска
более светлая и тусклая. Спина грязно-рыжеватая с узким, иногда слабо выражен-
ным ремнем по хребту. Южное Забайкалье.
с) с. b. fumatus Thomas, 1909. (Тип из провинции Гирин.) Окраска темная буро-
вато-ржавая с широким черноватым ремнем. Лесостепная полоса Забайкалья, долина
р. Амура.
d) С. b. ferrugineus Argyropulo, 1940. (Тип из Южно-Уссурийского края.) Окраска
яркая, насыщенная, хотя и не столь темная, с резко выступающими рыжими оттен-
ками. Южное Приморье.
83. Длиннохвостый хомячок. Cricetulus longicaudatus Milne-
Edwards, 1867
Табл. XXXIV, 3. Карта 98
Широко распространен в Китае и Монголии. В СССР найден в Ту-
винской области и на крайнем юге Красноярского края (Западный
Саян), а также по р. Джиде, в Забайкалье. Встречается по склонам
гор и в степи. Любит селиться среди скал, камней и в кустарнике. Пи-
тается как растениями, так и насекомыми. Образ жизни изучен слабо.
Как редкая форма, хозяйственного значения не имеет.
В Саянах обитает особая, темно окрашенная форма: С. 1. kozhantscikovi Vino-
gradov, 1927.
84. Серый хомячок. Cricetulus migratorius Pallas, 1770
Табл. XXXIV, 4—6. Карта 98
Распространен от Венгрии и Греции до Алтая и от северных пре-
делов лесостепи до Ирана и Афганистана. В нашей стране этот хомя-
чок встречается по всему югу Европейской части (к северу, примерно,
до Житомира, Киева, Чернигова, Калуги, Рязани, Горького, Казани,
Уфы), в Закавказье, по всей Средней Азии и Казахстану и в степях
юга Сибири (к северу до Тюмени, Барабинской степи, Новосибирска).
Встречается на степных участках, среди пашен, в горах, в посе-
лениях человека. Роет глубокие норки. Ведет преимущественно ночной
образ жизни. На зиму впадает в спячку. Дает 2, а возможно и 3 по-
мета, до 9 детенышей в каждом. Питается различными травянистыми
растениями и их семенами, поедает также насекомых и моллюсков. На
зиму делает большие запасы из семян различных растений. Вредит зер-
новым культурам.
312
В СССР встречаются следующие расы серого хомячка:
а) С. m. bellicosus Scharleman, 1915. (Тип из окрестностей Степанцы, Киевской
области.) Окраска спины темная, пепельно-серая с буроватым оттенком. Размеры
небольшие. Правобережная и север Левобережной части Украины, центрально-черно-
земные области.
b) С. ш. phaeus Pallas, 1778. (Тип из окрестностей г. Волгограда.) Окраска более
светлая с примесью рыжеватых тонов. Размеры крупнее. Слабо выраженными natio
этого подвида являются neglectus Ognev, 1916 (из мелитопольских степей), zviersombi
Pidoplitschko, 1928 (из окрестностей Ростова). Крым, Южная Украина, степи Предкав-
казья, Донецкий бассейн, Нижнее Поволжье.
с) С. ш. pulcher Ognev, 1924. (Тип из окрестностей Ларса, на Военно-Грузинской
дороге.) Окраска спины относительно светлая, мышино-серая с палевым оттенком.
Кавказ.
d) С. ш. vernula Thomas, 1917. (Тип из Турции.) Окраска весьма темная, серая.
Западное Закавказье.
е) С. m. migratorius Pallas, 1770. (Тип с низовьев р. Урала.) Окраска спины свет-
лая, бледная с песчаным оттенком. Западный Казахстан.
f) С. m. arenarius Pallas, 1770. (Тип из Барабинской степи.) Несколько более
темная форма с рыжеватыми оттенками. Прииртышье.
g) С. m. elisarjewi Afanasiev, 1953. (Тип из окрестностей с. Шемонаихн,. на Ал-
тае.) Окраска спины и боков очень темная, интенсивно серая с легким буроватым
оттенком; вдоль хребта тянется размытая черноватая полоса. Алтай.
h) С. m. fulvus Blanford, 1875. (Описан из Кашгара.) Окраска спины светлая,
песчано-серая. Восточные окраины- Казахстана.
i) С. m. caesius Kaschkarov, 1923. (Тип из окрестностей г. Фрунзе.) Спина кра-
сивого голубовато-серого цвета средней темноты; у старых особей с примесью бурых
оттенков. Горы Тянь-Шаня. Синонимы: С. m. griseus Kaschkarov, 1923, и С. fn. cinereus
Kaschkarov, 1926.
j) С. m. coerulescens Severtzov, 1879. (Тип с берегов оз. Кара-Куль, на Памире.)
Окраска очень пышного меха спины крайне бледного, розовато-палево-сероватого
цвета. Памир.
k) С. m. ognevi Argyropulo, 1936. (Тип из окрестностей г. Самарканда.) Очень
светлая, песчано-серая форма. Самаркандский и Бухарский оазисы.
I) С. m. isabellinus de Filippi, 1865. (Описан из Ирана.) Окраска также очень
бледная, палево-сероватая без примеси розоватых тонов. Копет-Даг.
Из этого обзора рас серого хомячка видно, что зону черноземных степей насе-
ляют особи темной, буровато-серой окраски. В зоне сухих степей и полупустынь во-
дятся хомячки более светлой, песчано-серой окраски с выступающими рыжеватыми
оттенками. В пустынях Средней Азии и в высокогорных пустынях Памира встре-
чаются очень светлые расы с бледной, палево-серой окраской спины. В горах Тянь-
Шаня и Кавказа обитают хомячки чисто серой окраски средней темноты.
По-видимому, эти группы форм серого хомячка соответствуют реально различи-
мым подвидам, а большинство из 12 перечисленных рас в лучшем случае могут быть
выделены в качестве natio.
Подрод крысовидных хомяков. Subgenus Tscherskia
Ognev, 1914
Единственный вид.
85. Крысовидный хомяк. Cricetulus triton Winton, 1899
Табл. XXXIV, 7. Карта 98
Юг советского Приморья, восточные провинции Китая и Корея.
Держится преимущественно по долинам рек. Роет сложные, со
многими входами, кладовыми и гнездами, норы, куда к зиме натаски-
вает большие запасы семян. Питается, главным образом, семенами ди-
корастущих растений, зернами хлебных злаков, насекомыми. Дает за
год несколько пометов, число детенышей в которых может достигать
20 (обычно 10—12).
В СССР, Северо-Восточном Китае и Корее обитает особая форма:
С. t. nestor Thomas, 1907. (Тип из окрестностей Сеула, в Корее.) Отли-
чия его от С. t. triton Wint. не выяснены.
313
Рис. 115. Костное нёбо полевок:
А — серой полевки (Microtus); Б — каменной полевки (Alticola).
Подсемейство ПОЛЕВОК- Subfamilia MICROTINAE
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ПОДСЕМЕЙСТВА ПОЛЕВОК
ФАУНЫ СССР
1(2). Хвост сильно сжат с боков. Размеры крупные: длина задней
ступни взрослых и полувзрослых особей более 30 мм; кондилобазаль-
ная длина черепа более 45 мм . . . . Ондатра. Ondatra. (Стр. 316.)
2(1). Хвост в сечении округлый. Размеры более мелкие: длина зад-
ней ступни (без когтей) менее 30 мм; кондилобазальная длина черепа
менее 45 мм...................................................3
3(4). Размеры относительно крупные (с домовую крысу): длина
ступни взрослых особей более 25 мм; кондилобазальная длина их че-
репа более 32 мм...........Водяные крысы. Arvicola. (Стр. 316.)
4(3). Размеры меньше: длина ступни не более 24 мм; кондилоба-
зальная длина черепа менее 32 мм..............................5
5(20). Верхние резцы направлены прямо вниз. Их передняя по-
верхность оранжевая или желтая. Эмалевые петли коренных зубов
образуют вполне замкнутые треугольники. Боковые углы их у взрос-
лых особей острые.............................................6
6(19). Когти пальцев передних лап короткие: длина самого длин-
ного из них менее 5 мм. Глаза развиты нормально. Задний коренной
зуб верхней челюсти имеет на внутренней стороне 3—4 выдающихся
зубца.........................................................7
7(12). Хвост (без концевых волос) хотя бы на 1—2 мм длиннее
ступни (без когтей)...........................................8
8(9). Задний край костного нёба черепа доходит до линии, соеди-
няющей средние части правого и левого задних коренных зубов верх-
ней челюсти. По бокам задней части костного нёба расположены ямки,
разделенные по средней линии нёба возвышением. Задний край кост-
ного нёба соединен с основаниями крыловидных отростков костными
перемычками (рис. 115). Окраска меха буроватая или серая . . . .
. . . .....................Серые полевки. Microtus. (Стр. 318.)
9(8). Задний край костного нёба доходит до линии, соединяющей
передние поверхности задних коренных зубов верхней челюсти. Ямок
на боках задней части костного нёба нет. Задний край костного нёба
обрывается над хоанами и не соединяется костными перемычками с ос-
нованиями крыловидных отростков (рис. 115)...................10
10(11). Коренные зубы у взрослых особей имеют корни, почему
ребра призм зубных коронок не доходят до краев альвеол или едва
достигают их. Длина вибрисс менее 35 мм. В окраске спины обычно
314
ясно выступают рыжие и красные оттен-
ки (кроме форм с Тянь-Шаня, Сахалина
и о. Шикотан) . . . Лесные полевки.
Clethrionomys. (Стр. 331.)
11(10). Коренные зубы как у моло-
дых, так и у старых особей корней не
Б
Рис. 116. Коготь первого
пальца передней лапы лем-
мингов: -
А — лесной лемминг (Myopus
schisticolor); Б — копытный лем-
минг (Dicrostonyx torquatus).
имеют, поэтому ребра призм их коронок
и отделяющие их борозды вдоль боков
зубов уходят в глубь альвеол. Длина ви-
брисс более 35 мм. Окраска спины обыч-
но серая с буроватым или желтоватым
оттенком .... Каменные полевки.
Alticola. (Стр. 335.)
12(7). Хвост (без концевых волос)
короче ступни (без когтей) .... 13
13(14). Подошва задней ноги в об-
ласти оснований пальцев голая; круп-
ные бугорки (мозоли) на подошве вы-
ступают отчетливо ....................
Лесные лемминги. Myopus. (Стр. 337.)
14(13). Вся подошва задних лап покрыта густой шерстью, скрыва-
ющей или почти скрывающей слаборазвитые мозоли..................15
15(16). Коготь первого (внутреннего) пальца передних лап круп-
ный, плоский, с параллельными боковыми краями и вырезкой на конце.
Заглазничные отростки черепа слабо выражены, закруглены . . . .
.......................Настоящие лемминги. Lemmus. (Стр. 338.)
16(15). Коготь первого (внутреннего) пальца передних лап мелкий,
конической формы, заостренный. Заглазничные отростки черепа хо-
рошо развиты, имеют вид тупых конусов (рис. 117)................17
17(18). Наружное ухо имеет вид небольшой каемки кожи вокруг
слухового прохода. Летом окраска сверху рыжевато-коричневая со
светлым ошейником, зимой белая или почти белая.....................
.......................Копытные лемминги. Dicrostonyx. (Стр. 339.)
18(17). Ушная раковина, хотя и мала, все же хорошо развита. Ок-
раска иная.................Степные пеструшки. Lagurus. (Стр. 340.)
Рис. 117. Черепа леммингов (сверху):
А — сибирский лемминг (Lemmus sibiricus); Б — копытный лемминг
(Dicrostonyx torquatus).
315
19(6). Когти передних лап очень длинные: обычно длина когтя
среднего пальца более 5 мм. Глаза редуцированы, малы. Задний ко-
ренной зуб верхней челюсти имеет на внутренней стороне коронки
только 2 выдающихся зубца........................................
....................Прометеевы полевки. Prometheomys. (Стр. 341.)
20(5). Верхние резцы направлены вниз и вперед. Их передняя по-
верхность белая. Эмалевые петли коренных зубов не образуют замкну-
тых треугольников. Боковые углы их у взрослых особей закругленные .
. . . . ....................Слепушонки. Ellobius. (Стр. 341.)
РОД ОНДАТРЫ. GENUS ONDATRA Link, 1795
Относительно крупные грызуны (длина тела взрослых 30—40 см)
с длинным, резко уплощенным с боков, покрытым кожными чешуй-
ками хвостом. Пальцы задних ног частично связаны плавательной пе-
репонкой. Мех пышный. Окраска от песочной до темно-бурой, брюхо
желто-серебристое. Длина черепа 60—65 мм. Коренные зубы с кор-
нями.
В СССР — один вид.
86. Ондатра. Ondatra zibethicus Linnaeus, 1766
Табл. XXXV, /, 2. Карта 99
Широко распространена по Северной Америке. С целью обогаще-
ния фауны пушных зверей завезена в ряд стран Европы (Германия,.
Чехословакия, Финляндия и др.). В СССР впервые ввезена в 1928 г.
Ныне заселяет водоемы почти всей территории севера Европейской ча-
сти СССР, многих районов средней и южной полос этой части страны,,
почти всей Западно-Сибирской низменности, большей части Восточной
Сибири, ряда районов Якутии, Забайкалья и Дальнего Востока.. В Ка-
захстане особенно успешно прошла акклиматизация ондатры в ни-
зовьях р. Или и по нижнему течению Сыр-Дарьи. Очень много ондатры
стало в дельте Аму-Дарьи. В настоящее время ареал ондатры охватил
уже примерно V2 площади СССР.
Живет по берегам озер, рек, стариц, на сплавинах и дамбах, в но-
рах или куполообразных «хатках». На зиму в спячку не впадает. За
год дает 2—3 помета из 2—16 (обычно 5—7) детенышей. Питается при-
брежной и водной растительностью. Дает ценный мех. Служит важным
объектом пушного промысла. Изредка бывает источником туляремии.
РОД ВОДЯНЫЕ КРЫСЫ. GENUS ARVICOLA Lacepede, 1799
Крупные полевки величиной с домовую крысу или несколько
мельче. Хвост длиной около V2—2/з длины тела имеет круглое сечение.
Окраска меха на спине варьирует от светло-коричневой или сероватой
до темно-бурой, брюха — от ржаво-белесой до бурой. Третий коренной
зуб верхней челюсти имеет упрощенное строение — только с двумя вхо-
дящими углами как с внешней, так и с внутренней стороны. Распро-
странены по всей Европе, в Сибири (на восток до р. Лены), в Казах-
стане и в Передней Азии.
Один вид.
316
87. Водяная крыса. Arvicola terrestris Linnaeus, 1758
Табл. XXXV, 3—5. Карта 100
От берегов Атлантического океана до Байкала и правобережья
р. Лены, от Малой Азии, Среднего Казахстана и Северо-Западной Мон-
голии до кустарниковых тундр к северу. На большей части громадного
ареала водяная крыса живет по поймам рек и берегам озер. В под-
таежной полосе заболоченных березовых лесов Западной Сибири, от
восточных склонов Урала до поймы Оби, она расселена равномерно и
повсеместно. На водораздельных участках тайги, лесотундры, степей
и полупустынь встречается только по берегам мелких рек и озер. В пу-
стынях и тундрах не встречается.
Летом живет в поймах, сырых заболоченных местах, на огородах,
•свекольных полях, в садах. Питается сочными частями растений, осо-
бенно прикорневыми частями крупных осок, рогоза, камыша, тростника
и других гидрофитов. Поселяется летом в норах, внутри осоковых ко-
чек, в дуплах пней и лежащих на земле стволов или в травянистых
гнездах на поверхности кочек. Хорошо, плавает и ныряет. Размно-
жается 2—4 раза в течение теплого времени года. В каждом приплоде
бывает от 2 до 10 (в среднем 6—7) детенышей. Часть'молодых (начи-
ная с двухмесячного возраста) приступает к размножению в том же
году. В конце лета переселяются на возвышенные участки, в том числе
на посевы зерновых культур, огороды и пр., где устраивают сложные
зимовочные норы, и питаются там овощами, • корнеплодами, картофе-
лем, поедают зеленые стебли зерновых культур (кукурузы, пшеницы,
овса) и семена этих культур. В условиях Западной Сибири, в заболо-
ченных районах Белоруссии и в поймах больших рек Европейской ча-
сти СССР водяная крыса — основной вредитель растениеводства.
В этих же районах она служит главным источником заражения туля-
ремией людей и ценных промысловых животных (особенно ондатры).
Истребляют ее, главным образом, авиационно-приманочным методом.
С самолетов рассеивают предварительно размоченные и оклеенные
фосфидом цинка на растительном масле зерна гороха или кукурузы.
С территории СССР описано более 20 подвидов водяной крысы. Но критиче-
ский анализ признаков этих форм показывает, что заметно отличаются друг
•от друга лишь сравнительно немногие из описанных рас. К числу таких мы относим
•следующие:
a) A. t. terrestris Linnaeus, 1758. (Описан из Швеции.) Размеры средние: конди-
лобазальная длина черепа 34—41 мм. Ширина скуловых дуг составляет около 60%
кондилобазальной длины черепа. Летний мех темный, серовато-бурый. Распростра-
нена в северных областях Европы на юг до южного предела лесной зоны. A. t. ferru-
ginous Ognev, 1933, отличающаяся немного более рыжеватым оттенком меха, относится
к этой же форме в качестве особой natio.
b) A. t. meridionalis Ognev, 1923. (Описан с Черенинского канала, в Заволжье.)
Размеры крупные: кондилобазальная длина черепа 36—44 мм; скуловая ширина со-
ставляет около 63% кондилобазальной длины черепа. Окраска летнего меха на спине
разнообразна, но обычно довольно светлая и тусклая. Встречается по всей степной-
и лесостепной полосе Европейской части СССР (до предгорий Б. Кавказа), в Зау-
ралье, Западном Казахстане.
с) A. t. pallasii Ognev, 1913. (Тип с р. Сосьвы.) Размеры крупные: кондилобазаль-
яая длина черепа 39—43 мм. Мех исключительно пышный. Окраска его темная, черно-
вато-бурая, но попадаются и светлые особи с буровато-серой расцветкой меха. Север
Сибири, от Урала до Якутии.
d) A. t. variabilis Ognev, 1938. (Тип из Барабинской степи.) Размеры средние:
кондилобазальная длина черепа 35—41 мм; скуловая ширина около 63% кондилоба-
зальной длины. Мех менее пышный, чем у северосибирской формы. Окраска летнего
меха обычно темно-бурая. Хвост довольно короткий. Южная Сибирь, Северо-Восточ-
ный Казахстан.
е) A. t. scithicus Thomas, 1914. (Тип из окрестностей г. Панфилова.) Размеры
крупные: кондилобазальная длина черепа 41—43 мм; скуловая ширина составляет
317
около 61% кондилобазальной длины черепа. Окраска летнего меха от темного, буро-
вато-серого до буро-черноватого. Семиречье.
f) A. t. persicus de Filippi, 1865. (Эльбрус, Северный Иран.) Череп крупный:
кондилобазальная длина его 37—42 мм- скуловая ширина составляет около 57% кон-
дилобазальной длины. Окраска преобладает яркая с примесью коричневато-рыжих
тонов. С Кавказа было описано различными авторами более 10 подвидов водяной
полевки, но нам кажется, что прав Н. К. Верещагин, который отмечает, что «геогра-
фический характер морфологических изменений зверька (кавказских водяных поле-
вок) уловить трудно». По-видимому, значительная часть описанных с Кавказа форм-
водяных полевок в действительности является синонимами или слабо выраженными
natio широко распространенного по Закавказью подвида A. t. persicus de Filippi. Си-
стема подвидов A. terrestris с Кавказа должна быть критически пересмотрена.
g) A. t. scherman Shaw, 1801. (Описан из окрестностей г. Страсбурга, Восточная
Франция.) Размеры относительно мелкие: кондилобазальная длина черепа 30—35 мм;
ширина скул составляет около 65% кондилобазальной длины черепа. Летний мех от
светло-коричневого до бурого. Эта форма рассматривается некоторыми териологами
как самостоятельный вид. Широко распространена в Центральной Европе; в СССР —
в Закарпатской, Драгобычской, Львовской, Тернопольской, Волынской и Ровенской
областях Украины и в юго-западных районах Белоруссии.
РОД СЕРЫЕ ПОЛЕВКИ. GENUS MICROTUS Schrank, 1798
Полевки средних и мелких размеров: длина тела до 15 см; длина
хвоста составляет обычно около !/3—ХД длины тела, реже до х/г и у по-
левки Роберта до 3Л длины тела. Окраска меха обычно буровато-серая
с примесью различных оттенков. Характерно строение костного нёба-
черепа. Его задний край доходит до линии, соединяющей задние концы
последних коренных зубов верхней челюсти. На задней части костного
нёба расположены два неглубоких углубления, отделенных средним
возвышением. Задне-боковые края нёба соединяются с крыловидными
отростками. Коренные зубы без корней. Широко распространены в Ев-
ропе, Азии, Северной Америке. К этому роду относят более 30 видов,
объединяемых в несколько подродов,
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДРОДОВ РОДА СЕРЫХ ПОЛЕВОК
ФАУНЫ СССР
1(2). Длина хвоста обычно больше х/2 длины тела; в случаях,
когда его длина близка к 50% длины тела, окраска хвоста как сверху,
так и снизу беловатая. Вибриссы значительно длиннее головы , . . .
..........................Снежные полевки. Chionomys, (Стр, 320.)
2(1), Длина хвоста обычно меньше ХА длины тела; в случаях,
когда его длина близка к 50% длины тела, окраска его ясно двух-
цветная: сверху темная, снизу светлая. Вибриссы короче длины го-
ловы ..............................................................3
3(8). Третий коренной зуб верхней челюсти имеет с внутренней
стороны 4 и более выдающихся углов.................................4
4(7). В первом коренном зубе нижней челюсти первый внутренний
и первый наружный треугольники разделены; эмалевые стенки третьего
наружного и третьего внутреннего (считая сзади) входящих углов
этого зуба соприкасаются друг с другом (рис. 118, А). На ступнях зад-
них лап почти всегда 6 мозолей.....................................5
5(6). Череп широкий: скуловая ширина превышает х/г общей
длины черепа; межглазничный промежуток почти всегда шире 3,3 мм .
...............................Серые полевки. Microtus. (Стр. 321.)
6(5). Череп узкий: скуловая ширина равна или меньше х/г общей
длины черепа; межглазничный промежуток почти всегда уже 3,3 мм . .
.....................Узкочерепные полевки. Stenocranius. (Стр. 329.)
7(4). В первом коренном зубе нижней челюсти первый внутрен-
ний и первый наружный треугольники слиты в один; эмалевые стенки
318
третьего наружного и третьего внутреннего
(считая сзади) входящих углов этого зуба
широко отодвинуты друг от друга (рис. 118,
Б). На ступне задней ноги только 5 мозолей.
Кустарниковые полевки. Micrurus. (Стр. 330.)
8(3). Третий коренной зуб верхней челю-
сти имеет с внутренней стороны только три
выдающихся угла..........................9
9(10). Первый наружный и первый внут-
ренний треугольники первого коренного зуба
нижней челюсти разделены соприкасающи-
мися эмалевыми стенками. Первый наружный
треугольник соответствует третьему выдаю-
щемуся углу внешней стенки (считая сзади),
а первый внутренний треугольник соответ-
ствует четвертому выдающемуся углу внут-
ренней стенки (тоже считая сзади) ....
....................Монгольские полевки.
Lasiopodomys. (Стр. 328.)
10(9). Первый наружный и первый внут-
ренний треугольники первого коренного зуба
нижней челюсти слиты друг с другом в своих
внутренних частях................... . 11
А Б
Рис. 118. Первый левый ко-
ренной зуб нижней челюсти
полевок:
А — обыкновенной полевки (Mi-
crotus arvalis); Б — кустарни-
ковой полевки (М. subterra-
neus); /в — первый внутренний
треугольник; 1н — первый на-
ружный треугольник.
11(12). Хвост светлый, однотонный 'сверху и снизу (или лишь сле!>
ка затемненный сверху). Длина его менее 35 мм. Межглазничное про-
странство широкое (более 4 мм),* плоское и гладкое (рис. 120, Л).
Затылочная ширина примерно равна V2 общей длины черепа . . . .
....................Афганские полевки. Blanfordimys. (Стр. 327.)
12(11). Хвост ясно двухцветный; длина его более 35 мм. Меж-
глазничное пространство узкое (менее 4 мм), у взрослых особей с яс-
ным сагиттальным гребнем или двумя менее выраженными дугооб-
разными гребнями, сближающимися в его средней части (рис. 120, Б).
Затылочная ширина значительно меньше У2 общей длины черепа . . .
. . . . . . Арчевые полевки. Phaiomys. (Стр. 329.)
Рис. 119. Черепа полевок (сверху):
А—обыкновенная полевка (Miceus arvalis);
узкочерепная полевка (М. gregalis).
Рис. 120. Черепа полевок:
А — афганская полевка (Microtus afghanus);
Б — арчевая полевка (М. juldaschi).
319
Подрод снежных полевок. Subgenus С h i о п о m у s Miller, 1908
Относительно крупные полевки (длина тела 10—15 см) с длинным
хвостом, длина которого равна 50—75% длины тела. Вибриссы длин-
нее головы. Окраска меха спины пепельно-серая или бурая. На ступ-
нях задних ног имеется по 6 мозолей. Ушные раковины большие, вы-
ступающие из меха.
Распространены в горах Европы и Передней Азии. В СССР—
2 вида.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ПОДРОДА СНЕЖНЫХ ПОЛЕВОК
ФАУНЫ СССР
1(2). Хвост всегда резко двухцветный. Длина его около 70—75%
длины тела. Спина темно-бурая с примесью рыжеватых оттенков. Ба-
рабанные камеры относительно мелкие: длина их примерно равна
длине ряда коренных зубов верхней челюсти (рис. 121, Л).............
.....................Полевка Роберга. Microtus roberti. (Стр. 320.)
Рис. 121. Черепа снежных полевок (вид снизу):
А — полевка Роберта (Microtus roberti); Б — снежная полевка
(М. nivalis).
2(1). Хвост обычно одноцветный, беловатый или реже двухцвет-
ный. Длина его около Чъ длины тела. В окраске спины преобладают
серые оттенки. Барабанные камеры крупные, длина их значительно
превышает длину ряда коренных зубов верхней челюсти................
....................Снежная полевка. Microtus nivalis. (Стр. 321.)
88. Полевка Роберта. Microtus roberti Thomas, 1906
Табл. XXXVI, /. Карта 100
Малая Азия, Западное Закавказье и западная часть Главного Кав-
казского хребта.
Живет в зарослях рододендрона, среди скал и камней, по окраи-
нам горных лесов. Образ жизни изучен слабо.
В СССР, по-видимому, номинальная форма.
320
89. Снежная полевка. Microtus nivalis Martins, 1842
Табл. XXXVI, 2, Карта 100
Горы Южной и Центральной Европы, Малая и Передняя Азия.
В Советском Союзе обитает на Карпатах, на Кавказе, на хребтах За-
кавказья и в горах Копет-Дага.
Живет преимущественно в альпийском и субальпийском поясах
гор, среди россыпей камней, в трещинах скал, на горных лугах, в ку-
старнике, по опушкам горных лесов. Дает в год до 3 пометов. Ле-
том питается травой, зимой также веточками рододендрона и других
кустарников. На зиму делает запасы сена. Хозяйственное значение не-
велико.
Подвиды (номинальная форма обитает в Альпах):
а) М. и. ulpius Miller, 1908. (Тип с гор Трансильвании.) Окраска темная, буро-
вато-серая. Хвост заметно двухцветный (сверху буроватый, снизу беловатый). Кон-
дилобазальная длина черепа 28—31 мм. Карпаты.
b) М. n. trialeticus Shidlcvskyi, 1919. (Тип из Бакуриани, в Грузии.) Окраска
спины светлая, сероватая с палевым оттенком. Хвост беловатый или слабо двухцвет-
ный. Кондилобазальная длина черепа 26—29 мм. Первый верхний коренной зуб имеет
обычно семь замкнутых пространств. Горы Закавказья.
с) М. n. loginovi Ognev, 1950. (Тип из Кавказского заповедника.) Окраска спины
довольно интенсивная, буровато-серая. Хвост двухцветный. Размеры мелкие: кондило-
базальная длина черепа 25—26 мм. Зубы, как у предыдущего подвида. Северо-Запад-
ный Кавказ.
d) М. n. gud Satunin, 1909. (Тип с Гудаурского перевала, в Грузии.) Окраска от
палево-серой до мышино-серой. Хвост или беловатый, или двухцветный. Кондило-
базальная длина черепа 27—29 мм. Строение третьего верхнего коренного зуба ослож-
нено: на внешней стороне его 3—5 входящих углов. Первый верхний коренной зуб,
напротив, несколько упрощен: в нем обычно только 6 замкнутых островков дентина.
Центральный Кавказ.
е) М. n. nenjukovi Formosov, 1931. (Тип из Карачая.) Окраска более интенсивная,
буроватая. Хвост двухцветный. Размеры крупнее, чем у предыдущей формы: кондило-
базальная длина черепа 30—32 мм. Зубы, как у М. n. gud. Северо-Западный Кавказ.
f) М. n. ighesicus Schidlovskyi, 1919. (Тип из Андийского округа Дагестана.)
Близка к М. n. gud, но заметно мельче: кондилобазальная длина черепа 26—28 мм.
Восточная часть Главного Кавказского хребта.
g) М. n. dementjevi Heptner, 1939. (Тип с массива Душак, в Копет-Даге.) Близок .
к М. n. trialeticus, но окраска светлая, палево-серая. Южная Туркмения.
Подрод серых полевок. Subgenus Microtus Schrank, 1798
Средней величины полевки с относительно коротким хвостом,
длина которого обычно равна или меньше половины длины тела. Уши
в большинстве случаев ясно выступают из меха. Вибриссы короче го-
ловы. Ступни задних ног с 5 или 6 мозолями. Окраска у большинства
видов буровато-серая, реже песчанисто-серая. Третий верхний корен-
ной зуб имеет с внутренней стороны 4 или 5 выдающихся углов.
Широко распространены в Европе, Азии, Северной Африке и Се-
верной Америке. В СССР обитает 9 видов.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ПОДРОДА СЕРЫХ ПОЛЕВОД
ФАУНЫ СССР
1(16). Хвост двухцветный (сверху темный, снизу светлый) или
весь темный. Окраска спины обычно сравнительно темная, буровато-
серая или мышино-серая. Сосцевидные части барабанных камер не
раздуты (рис. 125, А)................................................2
2(3). Первый коренной зуб нижней челюсти имеет на своей же-
вательной поверхности 6 замкнутых эмалевыми стенками островков
дентина. На наружной стороне этого зуба лишь три выдающихся угла
21 Н. А. Бобринский и др.
321
(рис. 122, А, Б). Длина хвоста около V2 длины тела. На ступнях задних
ног по 6 мозолей . Полевка-экономка. Microtus oeconomus. (Стр. 323.)
3(2). Первый коренной зуб нижней челюсти имеет на своей жева-
тельной поверхности 7 и более замкнутых эмалевыми стенками остров-
ков дентина. На наружной стороне этого зуба, как правило, 4 выдаю-
щихся угла (рис. 122, В). Длина хвоста обычно меньше V2 длины тела;
в тех случаях, когда она при-
мерно равна */2 длины тела,
на задних ступнях по 5 мозо-
Рис. 122. Коренные зубы нижней челюсти
полевок:
А, Б — полевки-экономки (Microtus oeconomus);
В — обыкновенной полевки (М. arvalis).
лей .......................4
4(5). Второй коренной
зуб верхней челюсти имеет
на своей жевательной поверх-
ности 4 большие и 1 (зад-
нюю) малую замкнутую пет-
лю. С внутренней стороны
этого зуба имеется 3 выдаю-
щихся угла (рис. 124, Б) . .
................................................Темная полевка.
Microtus agrestis. (Стр. 325.)
5(4). Второй коренной
зуб верхней челюсти имеет
на своей жевательной поверх-
ности только 4 большие замкнутые петли. С внутренней стороны этого
зуба только два выдающихся угла (рис. 124, Л) ........ 6
6(11). Длина хвоста более 7з длины тела. Ступни задних ног по-
крыты сверху темными буроватыми и буровато-серыми волосами.
У большинства особей на подошвах задних ног только по 5 мозолей . 7
7(8). Последний коренной зуб верхней челюсти усложненного
строения. Жевательная поверхность этого зуба имеет 6 изолированных
дентиновых островков. На его внешней стороне имеется 4 входящих
угла (рис. 123, В)................................................
.............Сахалинская полевка. Microtus sachalinensis. (Стр. 324.)
8(7). Последний коренной зуб верхней челюсти более простого
строения. Жевательная поверхность его имеет только 4—5 изолиро-
ванных дентиновых островков. На его внешней стороне имеется только
2—3 входящих угла (рис. 123, Б).................................9
9(10). Размеры крупные: длина тела взрослых особей обычно бо-
лее 12 см. На межглазничном пространстве крыши черепа ясно высту-
Рис. 123. Задний коренной зуб верхней челю- Рис. 124. Второй коренной зуб верх-
сти крупных серых полевок: пен челюсти серых полевок:
.4 — полевки-экономки (Microtus oeconomus); Б— А — обыкновенной полевки (Microtus arva-
большой полевки (М. maximowiczi): В — сахалинской lis); Б — темной полевки (М. agrestis); зп —
полевки (М. sachalinensis). задняя петля.
322
Рис. 125. Черепа полевок:
.4 — обыкновенной полевки (Microtus arvalis): Б — обще-
ственной полевки (М. socialis); бк— барабанная камера;
ск — сосцевидная кость.
пает сагиттальный гребень или два невысоких параллельных валика.
Межглазничный промежуток, как правило, менее 4,5 мм...........
..................Большая полевка. Microtus maximowiczi. (Стр. 324.)
10(9). Размеры более мелкие: длина тела почти всегда менее
12 см. На широком (шириной обычно более 4,5 мм) межглазничном
промежутке гребней или валиков нет: он почти гладкий............
.................Японская полевка. Microtus montebelli. (Стр. 325.)
11(6). Длина хвоста равна или менее 7з длины тела. Ступни зад-
них ног сверху светлые, сероватые. У большинства особей на подошвах
задних ног по 6 мозолей................................... 12
12(13). Темная окраска верхней поверхности хвоста постепенно,
без резкой границы, переходит в несколько более светлую расцветку
его нижней стороны. У большинства особей на внутренней стороне
третьего коренного зуба
верхней челюсти выступает
4 выдающихся угла, а на
его внешней поверхности—
3 угла....................
Обыкновенная полевка
Microtus arvalis. (Стр. 325.)
13(12). Темная окраска
верхней стороны хвоста
резко отграничена от бело-
ватой расцветки его ниж-
ней поверхности. На внут-
ренней и наружной сторо-
нах третьего коренного зу-
ба верхней челюсти имеется по 4 выдающихся угла..............14
14(15). Хвост равен примерно V3 длины тела. На межглазничном
пространстве черепа взрослых особей выступает продольный средин-
ный гребень. Длина слуховых барабанов менее 8,5 мм..............
. . . . Полевка Миддендорфа. Microtus middendorfi. (Стр. 326.)
15(14). Хвост равен примерно 74 длины тела. Межглазничное про-
странство черепа даже взрослых особей гладкое. Длина слуховых бара-
банов более 8,5 мм..............................................
. . Северосибирская полевка. Microtus hyperboreus. (Стр. 327.)
16(1). Хвост светлый как сверху, так и снизу. Окраска спины
обычно светлая, песчано-серая. Сосцевидные кости и костные барабаны
черепа сильно раздуты (рис. 125, Б).............................
.................Общественная полевка. Microtus socialis. (Стр. 327.)
90. Полевка-экономка. Microtus oeconomus Pallas, 1776
Табл. XXXVI, 3. Карта 101
Населяет тундры, лесотундры, всю таежную и часть лесостепной
зоны СССР к югу до Северной Украины, Воронежской области, Юж-
ного Урала, Северного Казахстана и Семиречья. Встречается также
в северной половине Западной Европы и в Монголии.
Селится в сырых местах, в зарослях кустов, на болотах, в поймах
рек, по оврагам. Живет в неглубоких норах с разветвленной системой
ходов. Летом питается, главным образом, зелеными частями травяни-
стых растений и их корнями и семенами, а зимой — корой и побегами
кустарников и молодых деревьев. Дает за год несколько пометов, по
6—12 детенышей в каждом. Вред, приносимый посевам, невелик. Во
многих местах средней полосы экономка служит основным источником
безжелтушного лептоспироза.
21*
323
Из различных районов СССР описано более 20 подвидов полевки-экономки.
Но многие из этих форм описаны на столь малом материале и столь поверхностно,
что реальность их признаков остается неясной. С большими затруднениями можно
различить (главным образом по тону окраски меха) следующие формы:
а) М. ое. oeconomus Pallas, 1776. (Описана по экземпляру из долины р. Ишима,
Западная Сибирь.) Размеры относительно крупные: кондилобазальная длина черепа
27—31 лш. Летний мех спины интенсивно-бурый. Западно-Сибирская низменность,
Урал и тайга Европы к югу до Минска, Калинина, Горького, Уфы. Европейские
особи от западносибирских слегка отличаются тоном окраски меха.
b) М. ое. petchorae Ognev, 1944. (Тип с низовьев р. Пеши.) Отличается от типич-
ной формы более светлой и рыжеватой окраской летнего меха спины. Тундровый
п лесотундровый районы Европейской части СССР, к востоку от Горла Белого моря.
с) М. ое. arenicola De S. Longchamps, 1841. (Тип из окрестностей Лейдена, в Гол-
ландии.) Относительно мелкая: кондилобазальная длина черепа 26—28 мм. Окраска
меха спины более светлая, рыжеватая, переходящая на боках в рыжевато-бурую.
Центральная Европа, Северная Украина и центрально-черноземные области.
d) М. ое. altaicus Ognev, 1944. (Тип с оз. Джулу-Куль, на Алтае.) Размеры сред-
ине: кондилобазальная длина черепа 27—30 мм. Окраска меха на спине тусклая,
сероватая. Алтай, Саяны, Тарбагатай.
е) М. ое. montiumcaelestinum Ognev, 1944. (Тип из долины р. Теректы, на Джун-
гарском Алатау.) Размеры немного меньше: кондилобазальная длина черепа
26—28 мм. Окраска большинства особей очень темная, черновато-бурая с рыжиной.
Семиречье.
f) М. ое. kjusjurensis Koljuschev, 1935. (Тип из пос. Кюсюр, на р. Лене.) Размеры
крупные: кондилобазальная длина черепа 29—31 мм. Мех спины тусклый, серый
с буроватым или желтоватым оттенком. Низовья рек Таза и Енисея, бассейн рек
Нижней и Подкаменной Тунгуски, низовья р. Лены.
g) М. ое. dauricus Kastschenko, 1910. (Тип из Южного Забайкалья.) Размеры сред-
ние: кондилобазальная длина черепа 27—30 мм. Окраска пышного меха темная,
черновато-серо-бурая с желтоватым оттенком. Прибайкалье и Забайкалье.
h) М. ое. suntaricus Dukelskaya, 1928. (Тип из дер. Сунтар, на Вилюе.) Размеры
невелики: кондилобазальная длина черепа 28—29 мм. Окраска летнего меха спины
насыщенного серо-бурого цвета с коричневым оттенком. Характерен длинный хвост.
Центральная и Южная Якутия.
i) М. ое. koreni Allen, 1914. (Тип из окрестностей Нижне-Колымска.) Размеры
относительно крупные: кондилобазальная длина черепа 30—32 мм. Окраска летнего
меха темная, коричиевато-бурая. Бассейн рек Колымы и Индигирки.
k) М. ое. tschuktschorum Miller, 1899. (Тип с побережья залива Креста.) Раз-
меры мелкие: кондилобазальная длина черепа 26—27 мм. Окраска летнего меха свет-
лая, палево-серая. Чукотский полуостров.
m) М. ое. shantaricus Ognev, 1929. (Тип с острова Большой Шантар.) Размеры
средние: кондилобазальная длина черепа 27—30 мм. Окраска буровато-серая с жел-
товатым оттенком. Камчатка, Анадырский край. (Видимо, к этой же форме относится
М. ое. uchida Kuroda, 1924, с острова Парамушир.)
m) М. ое. shantaricus Ognev, 1929. (Тип с острова Большой Шантар.) Размеры,
как у предыдущей: кондилобазальная длина черепа 26—30 мм. Хвост короткий (до
35 мм}. Окраска яркая, рыжеватая. Шантарские острова, Охотское побережье.
Сахалинская полевка. Microtus sachalinensis Vassin, 19551
Карта 101
Найдена в южной части о-ва Сахалин. Обитает на тундроподобных
болотистых местах. Живет колониями в сложно разветвленных норах.
Питается болотными и луговыми травами. В помете до 9 детенышей.
Хозяйственного значения не имеет. Систематическое положение тре-
бует уточнения.
Большая полевка. Microtus maximowiczi Schrenk, 1958 1
Табл. XXXVI, 4. Карта 101
Встречается в Забайкалье, по р. Амуру и в Приморье. Широко
распространена в Китае и Восточной Монголии.
Чаще всего селится по берегам рек, озер, по опушкам лесов, в ку-
1 Большую и сахалинскую полевки возможно следует считать не самостоятель-
ными видами, а только подвидами полевки-экономки. (Прим, ред.)
324
старнике. Нередко поселяется на пашнях и залежах. Роет сложные
разветвленные норы. Хорошо плавает. За год дает 3—5 (редко 6)
пометов, по 4—10 детенышей в каждом. Питается как дикорастущими,
так и культурными растениями. Местами причиняет вред.
Подвиды:
а) М. m. maximowiczi Schrenk, 1858. (Тип с Верхнего Амура у устья р. Омутной.)
Размеры не очень крупные: длина тела до 15 см; кондилобазальная длина черепа
27—33 мм. Забайкалье, верховье р. Амура.
b) М. m. pelliceus Thomas, 1911. (Тип с р. Уссури.) Размеры крупнее: длина тела
дс 17 см; кондилобазальная длина черепа 30—35 мм. Приморье.
Японская полевка. Microtus montebelli Milne-Edwards, 1872 1
Карта 101
Широко распространена на островах Японского архипелага. В пре-
делах СССР обнаружена Е. В. Соколовым на о-ве Шпанберга (Сико-
тан), в Курильской гряде. Селится в лиственных рощах по берегам
рек и в падях между сопками.
91. Темная полевка. Microtus agrestis Linnaeus, 1761
Табл. XXXVI, 5. Карта 102
Западная Европа, Европейская часть СССР (от зоны тундр на юг
до Северной Украины, Воронежской области и Южного Урала), За-
падная Сибирь (от тундр до Алтая и лесостепных районов), бассейн
р. Енисея, Прибайкалье и Якутия.
Держится преимущественно на лесных и болотистых участках,
в поймах рек, по оврагам. Самка за лето приносит несколько пометов
из 3—7 детенышей. Питание не изучено.
Подвиды:
а) М. a. agrestis Linnaeus, 1761. (Описан по экземпляру из Швеции.) Размеры
относительно крупные: кондилобазальная длина черепа 26—28 мм. Окраска темная,
серо-бурая. Почти у всех особей первый верхний коренной зуб имеет на задпе-внут-
реннем углу добавочный (4-й) зубец. Север Европы. Слабо выраженной natio
является М. a. a. ognevi Scalon из западносибирской тайги, отличающаяся еще более
темной окраской.
b) М. a. gregarius Linnaeus, 1766. (Описан по экземпляру из Германии.) Размеры
мельче: длина тела до 12,5 см; кондилобазальная длина черепа 25—27 мм. Окраска,
как у типичной формы. Добавочный зубец на задне-внутреннем углу первого верх-
него коренного зуба обычно отсутствует. Средняя полоса Европы.
с) М. a. argiropuloi Ognev, 1944. (Тип с р. Инзер, на Южном Урале.) Размеры
крупные: кондилобазальная длина черепа 27—29 мм. Отличается бледной, серова-
той окраской. Первый верхний коренной зуб, как у предыдущей формы. Южный
и Средний Урал, Башкирия.
d) М. a. mongol Thomas. 1912. (Тип с р. Кемчик, в Тувинской АССР.) Кондило-
базальная- длина черепа 25—29 мм. Окраска еще более тусклая и серая. Строение
первого верхнего коренного зуба сильно варьирует. Южная Сибирь, от Алтая до
Байкала.
92. Обыкновенная полевка. Microtus arvalis Pallas, 1779
Табл. XXXVI, 6. Карта 103
Европа, Южная Сибирь, Казахстан, Средняя Азия, Монголия и
Северо-Восточный Китай. В СССР водится в Европейской части (на
север до средней части Карелии, южных районов Архангельской
1 Полевки, добытые летом 1964 года на о. Шикотан, оказались не Microtus monte-
belli, a Clethrionomys (Neoaschisomys) sikotanensis Tokuda, 1935. Они же найдены на
Юго-Западном Сахалине. (Прим, ред.)
325
области и Коми АССР), на Кавказе, в. Закавказье, некоторых районах
Средней Азии и Казахстана, в степных и лесостепных районах Си-
бири, на восток до верховьев Амура.
Обитает на полях, залежах, бахчах, в целинной степи, по берегам
рек, на лугах горных склонов, по опушкам леса и в усадьбах чело-
века. На зиму нередко скапливается в стогах сена и ометах соломы.
Периодически наблюдаются массовые размножения. За лето прино-
сит до 4—6 пометов, обычно из 4—8 (иногда до 10) детенышей в каж-
дом. Основная пища — зеленые части травянистых растений, их семена
и корни. На пашнях сильно вредит посевам. Зимой нередко грызет
кору кустарников и фруктовых деревьев. Вредитель сельскохозяйствен-
ных культур и источник туляремии. Один из основных видов пищи на-
земных и пернатых хищников.
Описано большое число подвидов. Перечисляем их без критического пересмотра
коллекционных материалов:
а) М. аг. ruthenus Ognev, 1950. (Тип из окрестностей г. Кирова.) Размеры отно-
сительно мелкие: кондилобазальная длина черепа 22—25 мм. Окраска темная, насы-
щенная, буровато-серая. Кировская, Пермская области, Коми АССР, Марийская АССР.
b) М. аг. duplicatus Rorig et Borner, 1912. (Тип из Калининградской области.)
Размеры, как у предыдущей формы, но окраска светлее и тусклее с примесью
желтоватых оттенков. Средняя полоса Европейской части СССР.
с) М. аг. rossiaemeridionalis Ognev, 1924. (Тип из окрестностей г. Боброва Воро-
нежской области.) Размеры крупнее: .кондилобазальная длина черепа 24—28 мм.
Окраска сходна с окраской североевропейской формы — буровато-серая, без желтизны.
Южные области Европейской части СССР. На Северном Кавказе распространена
М. аг. г. macrocranius Ognev, 1924, отличающаяся несколько более массивным и
высоким черепом.
d) М. аг. iphigeniae Heptner, 1946. (Тип из Крымского заповедника.) Размеры,
как у предыдущей формы: кондилобазальная длина черепа 25—27 мм. Окраска лет-
него меха характеризуется развитием коричнево-ржавых тонов. Крым.
е) М. аг. transcaucasicus Ognev, 1924. (Тип из Борчалинского района Грузии.)
Размеры средние: кондилобазальная длина черепа 24—27 мм. Скулы расставлены
довольно широко. Окраска бледная, сероватая или песчано-желтоватая. Закавказье.
К этому подвиду весьма близка дагестанская М. аг. gudauricus Ognev, 1929.
f) М. аг. transcaspicus Satunin, 1905. (Тип из Чулийского ущелья, в Туркмении.)
Размеры очень крупные: кондилобазальная длина черепа 28—31 мм. Окраска спины
светлая, серо-палевая, буроватая или желтоватая. Туркмения.
g) М. аг. ilaeus Thomas, 1912. (Тип из окрестностей г. Панфилова.) По размеру
почти не уступает предыдущему подвиду: кондилобазальная длина черепа 26—30 мм.
Окраска значительно темнее: спина буровато-серая, иногда с желтизной. Семиречье.
h) М. аг. kirgisorum Ognev, 4950. (Тип с Киргизского хребта.) Размеры средние:
кондилобазальная длина черепа 24—26 мм. Окраска темная, буровато-серая. Горы
Киргизии.
i) М. аг. caspicus Ognev, 1950. (Тип из Астраханской степи.) Форма, близкая
к среднерусской полевке, но окраска светлее и желтее. Нижнее Поволжье, средняя
полоса Казахстана.
j) М. аг, obscurus Eversmann, 1841. (Описан по экземпляру с Алтая.) Размеры
средние: кондилобазальная длина черепа 23—27 мм. Окраска темная, буровато-серая.
Синонимом является М. a. transuralensis Ser и возможно отмеченная выше М. a. kir-
gisorum. Южный Урал, степи Западной Сибири, Алтай.
k) М. аг. mongolicus Radde, 1861. (Тип с верховьев Амура.) На ступне задней
ноги обычно 5 мозолей (редко 6); первый внешний зубец на М1 сглажен. Размеры,
как у алтайской формы: кондилобазальная длина черепа 23—26 мм. Окраска спины
темно-бурая, иногда с рыжеватым оттенком.
93. Полевка Миддендорфа. Microtus middendorfi Poljakov, 1881
Табл. XXXVI, 8, Карта 102
Тундры и лесотундры полярного Урала, низовьев рек Оби, Таза и
Енисея, Таймыр и Северная Якутия. Держится преимущественно на
болотистых участках, где роет норы в кочках и устраивает в покрываю-
щей их траве и ползучем кустарнике наземные гнезда. Реже селится
326
в норах на более сухих участках тундры. Питается тундровыми тра-
вами и побегами кустарников.
Подвиды:
а) М. m. middendorfi Poljakov, 1881. (Описана по экземпляру с Таймыра.) По-ви-
димому, синонимом ее следует считать описанную по экземпляру с р. Таза М. m. ta-
sensis Scalon, 1935.
b) M m. ryphaeus Heptner, 1948. (С Северного Урала.) Отличается несколько
более бледной окраской меха.
94. Северосибирская полевка. Microtus hyperboreus Vinogradov, 1933
Табл. XXXVI, 9. Карта 102
Найдена на Южном Таймыре, Верхоянском хребте, на Яне, Инди-
гирке и Колыме.
Обитает на склонах гор. Биология изучена слабо.
Систематически близка к М. middendorfi Poljakov.
95. Общественная полевка. Microtus socialis Pallas, 1773
Табл. XXXVII, 1—3. Карта 104
Встречается в степной части Крыма, на юге Левобережной Укра-
ины, в Приазовье, в степях Северного Кавказа, в Восточном Закав-
казье, в Калмыкии, в Казахстане и в равнинной части Средней Азии.
Селится, главным образом, среди сухой степи, в глинистой пустыне,
в плавнях, на парах, сенокосах. Живет обычно колониями в неглубо-
ких, но сложно устроенных и разветвленных норах. За теплое время
года самка родит 3—5 раз, принося по 4—11 детенышей. Питается степ-
ной растительностью и культурными растениями; местами вредит по-
севам.
Подвиды:
а) М. s. socialis Pallas, 1773. (Описан по экземпляру с низовьев р. Урала.)
Поскольку в настоящее время в низовьях Урала эта полевка не найдена, указать при-
знаки номинальной формы затруднительно.
b) М. s. parvus Satunin, 1901. (Тип с р. Калаус, в ставропольских степях.) Раз-
меры довольно крупные: кондилобазальная длина черепа 23—27 мм. Окраска лет-
него меха серо-палевая, с желтоватым оттенком. Степи Северного Кавказа.
с) М. s. nikolajevi Ognev, 1950. (Тип с Сиваша.) Размеры мельче: кондилобазаль-
ная длина черепа 23—25 мм. Окраска летнего меха несколько темнее и тусклее,
палево-буровато-серая. Юг Украины. (Форма, близкая к предыдущей.)
d) М. s. schidlovskii Argyropulo, 1933. (Тип добыт близ г. Ленинакана.) Размеры
средние: кондилобазальная длина черепа 23—26 мм. Окраска летнего меха темная,
землисто-серо-буроватая. Центральное Закавказье.
е) М. s. binominatus Ellerman, 1941. (Тип из окрестностей г. Тбилиси.) Размеры
чуть крупнее, чем у предыдущих форм: кондилобазальная длина черепа 24—28 мм.
Характерно большое вздутие барабанных камер. Закавказье — от г. Тбилиси до побе-
режья Каспия.
f) М. s. irani Thomas, 1921. (Описана по экземпляру из Северного Ирана.) Самая
крупная форма: кондилобазальная длина черепа 27—30 мм. Спина в летнем меху
серо-палевая с рыжиной. Характерный признак — наличие дополнительного, третьего
угла на внутренней стороне второго верхнего коренного зуба. Туркмения.
g) М. s. gravesi Goodvin. 1934. (Тип из Аральских Каракумов.) Размеры отно-
сительно мелкие: кондилобазальная длина черепа 23—25 мм. Окраска светлая, палево-
сероватая. Казахстан, Узбекистан.
Подрод афганских полевок. Subgenus Blanfordimys
Argyropulo, 1933
Внешне похожа на общественную полевку: хвост короткий, свет-
лый как сверху, так и снизу. Подошвы густо опушены. Сосков 4 пары.
Череп с очень большими раздутыми барабанными камерами. Третий
327
коренной зуб верхней челюсти имеет как с внутренней, так и с наруж-
ной стороны 3 выступающих и 2 входящих угла. Первый коренной зуб
нижней челюсти имеет 6 или реже 7 замкнутых пространств.
Один вид.
96. Афганская полевка. Microtus afghanus Thomas, 1912
Табл. XXXVII, 4. Карта 104
Распространена в Афганистане. В СССР найдена в горах Боль-
шого Балхана и на Копет-Даге, в долине р. Мургаб и его притоков,
от г. Мары до Кушки, в Серахском районе, на хребте Нуратау, на Гис-
сарском и Зеравшанском хребтах. Живет по предгорьям и склонам
гор, среди сухой степи и на участках богарных посевов. Роет очень
сложные системы подземных ходов на небольшой глубине. Нора имеет
от 30—50 до 200—300 выходов на поверхность. Вокруг каждого выхода
трава съедается почти начисто. Биология размножения не изучена.
Подвиды:
а) М. a. afghanus Thomas, 1912. (Тип из Афганистана.) Окраска светлая, палево-
охристая, хвост почти одноцветный. Юго-Восточная Туркмения.
b) М. a. bucharensis Vinogradov. 1930. (Тип из окрестностей Пенджпкента.)
Окраска темнее, сероватая. Хвост слабо двухцветный. Предгорья Зеравшанского и
Гиссарского хребтов.
с) М. a. balchanensis Heptner et Shukurov, 1950. (Тип с Большого Балхана.)
Окраска серая, тусклая. Западная Туркмения.
Подрод монгольских полевок. Subgenus Lasiopodomys
Lataste, 1887
Полевки средней величины. Передняя часть подошвы задних ног
голая с хорошо выступающими мозолями. Хвост едва превышает длину
ступни. Третий верхний коренной зуб упрощенного строения: на внеш-
ней стороне он образует только 2 или 3 выступающих угла.
Распространены в засушливых степях Монголии, восточной поло-
вины Китая и Забайкалья.
В фауне СССР — 2 близких вида.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ МОНГОЛЬСКИХ ПОЛЕВОК
ФАУНЫ СССР
1(2). Межглазничный промежуток черепа взрослых и старых осо-
бей узкий (менее 3,5 мм) с отчетливым гребнем вдоль середины меж-
глазничного пространства. На внешней стороне заднего верхнего ко-
ренного зуба 3 выступающих угла, причем задний угол хорошо выра-
жен ...............Полевка Брандта. Microtus brandti. (Стр. 328.)
2(1). Межглазничный промежуток черепа широкий (около 4 мм)
с отчетливым желобком (продольным углублением) вдоль его сере-
дины. На внешней стороне заднего верхнего коренного зуба 2 или 3
выступающих угла; третий зубец, если есть, то слабо выражен. Окраска
буроватая . . Китайская полевка. Microtus mandarinus. (Стр. 329.)
97. Полевка Брандта. Microtus brandti Radde, 1861
Табл. XXXVII, 5. Карта 104
Широко распространена в Монголии. В СССР — в степях Южного
Забайкалья. Селится обычно большими колониями на лугах западин
степных озер и долин рек, реже среди целинной степи. В годы мас-
328
сового размножения иногда предпринимает далекие миграции. Раз-
множается несколько раз в год, в помете 5—9 детенышей. Активна
в светлое время суток. Питается степными травами; на зиму делает
в норе большие запасы сухой травы и кореньев. В годы массового
размножения сильно вредит травостою сенокосов и пастбищ. Способ-
ствует распространению эпизоотий чумы в природных очагах.
В СССР номинальная форма: М. b. brandti Radde, описанная по экземпляру с бе-
регов оз. Торей в Забайкалье.
98. Китайская полевка. Microtus mandarinus Milne-Edwards, 1871
Табл. XXXVII, 6. Карта 104
Основная часть ареала этого вида лежит в пределах Китая.
В СССР найдена в южной части Бурятской АССР (Джидинский и
Закаменский районы).
Обитает на пашнях, лугах, на склонах сопок. Образ жизни в ус-
ловиях Советского Союза не изучен.
В СССР — особый подвид: М. m. vinogradovi Fetisov, 1936, отличающийся от осо-
бей из Внутренней Монголии более темной буроватой окраской меха на спине и боках.
Подрод арчевых полевок. Subgenus Phaiomys Blyth, 1863
Мелкие полевки. Длина хвоста около V3 длины тела. Подошвы
снизу густо опушены. Сосков 4 пары. Барабанные камеры небольшие.
На межглазничном пространстве черепа взрослых особей хорошо раз-
вит гребень или два дугообразных гребешка. Третий коренной зуб
верхней челюсти имеет как с наружной, так и с внутренней стороны по
3 выдающихся угла. Первый коренной зуб нижней челюсти обычно не-
сет на жевательной поверхности 5 замкнутых эмалевыми стенками
участков.
Распространены в горах Центральной и Средней Азии.
В СССР — один вид.
99. Арчевая полевка. Microtus juldaschi Severtzov, 1879
Табл. XXXVII, 7, 8. Карта 104
Встречается в горах Тянь-Шаня, Памира и Гиссаро-Алая.
Живет колониями на травянистых склонах, в зарослях арчи, на
лужайках среди горных лесов, по долинам горных рек, среди болот.
Норы роет под камнями или под корнями кустарников, в кочках. На
зиму собирает запасы сена. Местами уничтожает до 40% травостоя
пастбищ.
Подвиды:
а) М. j. juldaschi Severtzov, 1879. (Тип из окрестностей оз. Кара-Куль, на Па-
мире.) Окраска спины бледная, палево-серая; хвост почти одноцветный. Памир
и Алайская долина.
b) М. j. carruthersi Thomas, 1909. (Тип с Гиссарского хребта.) Окраска спины
темнее, буровато-серая. Хвост двухцветный. Горы хребтов Алайского, Гиссарского,
Зеравшанского и системы Тянь-Шаня до Таласского Алатау включительно.
Подрод узкочерепных полевок. Subgenus Stenocranius
Kastschenko, 1901
По внешнему виду сходны с настоящими серыми полевками, но
череп сильно вытянут, с узко расставленными скуловыми дугами и
узким межглазничным промежутком, ширина которого обычно менее
3,3 мм.
Единственный вид.
329
100. Узкочерепная полевка. Microtus gregalis Pallas, 1779
Табл. XXXVII, 9, 10. Карта 105
Распространена в горах Азии, степях Северного и Восточного
Казахстана, в степных районах Западной Сибири, на Алтае, в Саянах,
Тувинской АССР, Прибайкалье, Забайкалье, Якутии и по всей полосе
тундры и лесотундры от Берингова пролива до Горла Белого моря.
Встречается также в Монголии и Северо-Восточном Китае.
Места обитания разнообразны: высокогорные сырты, склоны гор,
целинные степи, пашни, луга по долинам рек, лужайки в тайге, тунд-
ровые пространства. Живет обычно колониями, в которых отдельные
норки соединены как подземными ходами, так и наземными тропин-
ками. Характерен очень малый диаметр ходов (около 1,5—2 см у мел-
ких форм). Размножается несколько раз в год, в помете 4—12 дете-
нышей. Питается травянистой растительностью. На зиму натаскивает
в норы большие запасы зерна, семян, луковиц, корней. В земледельче-
ских районах сильно вредит посевам.
Подвиды:
а) .М. g. gregalis Pallas, 1779. (Описан по экземпляру с р. Чулыма.) Размеры
средние: кондилобазальная длина черепа 24—28 лш, хвост длинный: 23—34 мм.
Окраска темная, буровато-серая. На затылке и передней части спины имеется раз-
мытая черная полоса. Степные районы Южной Сибири и Северного Казахстана.
Ь) Л4. gr. eversmanni Poljakov, 1881. (Тип с Уймона, на Алтае.) Размеры те же.
Окраска тусклая, буровато-серая без темной полоски на спине. Алтай и приалтай-
ские районы, Восточный Тянь-Шань. Очень близкие формы (natio?): М. g. dukel-
skiae Ognev, 1950 (тип с Саян) с более рыжеватым оттенком в окраске меха,
и М. g. castaneus Kaschkarov, 1923, с Западного Тянь-Шаня, отличающаяся немного
более темной окраской спины и более длинным хвостом.
с) М. g. montosus Argyropulo, 1932. (Тип из Сары-Таша, на Алае.) Размеры те
же. Окраска меха светлая, палево-сероватая. Алайская долина и прилегающие
хребты, Аксай, Арпа на Тянь-Шане.
d) М. g. dolguschini Afanasiev, 1939. (Тип из дельты р. Или.) Мелкая, светло
окрашенная форма. Низовья р. Или.
е) М. g. zachvatkini Heptner, 1945. (Тип из окрестностей г. Аральска.) Крупная
форма; кондилобазальная длина черепа 27—30 мм. Спина светлая, желтовато-серая,
иногда с рыжеватым оттенком. Центральный Казахстан.
f) М. g. raddei Poljakov, 1881. (Тип с оз. Торей.) Размеры средние: кондилоба-
зальная длина черепа 25—28 мм. Окраска спины светлая, тусклая, серо-палевая. Забай-
калье, Монголия, Чуйская долина, на Алтае.
g) М. g. brevicauda Kastschenko, 1901. (Тип с р. Алдана.) Кондилобазальная
длина черепа 25—29 мм. Спина буровато-серая, • средней темноты; по спине обычно
идет темная полоска. Длина хвоста около 30 мм. Центральная и Южная Якутия,
Северное Забайкалье.
h) М. g. buturlini Ognev, 1922. (Тип из дельты р. Индигирки.) Размеры крупные:
кондилобазальная длина черепа 26—30 мм. Хвост короткий: 21—26 мм. Окраска лет-
него меха темная, ржаво-коричневая. Тундровая зона Якутии и Дальнего Востока.
По-видимому, с ней идентична М. g. nordenskioldi Poliakov, 1881, с Таймыра.
i) М. g. major Ognev, 1923. (Тип с п-ова Ямал.) Размеры крупные: кондилоба-
зальная длина черепа 27—30 мм. Длина хвоста 24—35 мм. Окраска светлая, желтовато-
палево-серая. Ямал, Северный Урал, низовья р. Оби.
j) М. g. tundrae Ognev, 1944. (Тип из района г. Архангельска.) Размеры средние:
кондилобазальная длина черепа 26—28 мм. Хвост короткий: около 20 мм. Окраска
бледная, палево-сероватая. Тундры севера Европейской части СССР.
Подрод кустарниковых полевок. Subgenus Micrurus
F. Major, 1877 1
От других полевок рода Microtus отличаются относительно плоским
черепом, упрощенным строением первого нижнего коренного зуба, малым
числом сосков (2—3 пары), слаборазвитыми глазами. Распространены
в Европе, Малой Азии и на Кавказе. В фауне СССР — один вид.
1 Отнесение европейских кустарниковых полевок к американскому роду Pitymys
нам кажется необоснованным.
330
101, Кустарниковая полевка. Microtus subterraneus De Selys
Longchamps, 1835
Табл. XXXVII, 11, 12. Карта 105
Распространена в Европе, Передней Азии и на Кавказе. В СССР —
в Закавказье, на Северном Кавказе, почти по всей Украине, в Дон-
ской, Воронежской и Курской областях, некоторых частях Белоруссии,
Калининской и Вологодской областях.
Селится по лесостепным дубравам, в зарослях кустарника, в са-
дах, по краям полей; в горах поднимается до альпийских лугов. Живет
в норах с большим числом входов, камер и складов. В течение лета
у самок бывает 3—4 помета, число детенышей в помете обычно 3—4,
иногда до 7. Питается корнями и корневищами различных трав, их
листьями и побегами, семенами и желудями. На зиму делает большие
запасы, которые прячет в норах. Местами вредит молодым деревцам
.и кустарникам.
Подвиды:
а) М. s. ukrainicus Vinogradov, 1922. (Тип из окрестностей г. Змиева.) Длина
третьего коренного зуба верхней челюсти примерно равна длине второго. Украина.
b) М. s. transvolgensis Schanev et Schaposchnikov, 1958. (Тип из Максатихинского
.района Калининской области.) Длина третьего коренного зуба верхней челюсти при-
мерно на 10% больше длины второго. Калининская область.
с) М. s. atratus Stein, 1931. Отличается от двух предыдущих форм окраской меха,
в которой заметно выступают желтовато-бурые оттенки. Череп более узкий и длин-
ный. Белоруссия.
d) М. s. major! Thomas, 1906. (Тип из района г. Трабзона.) Размеры крупные:
длина тела до 11 см; кондилобазальная длина черепа 22—24 мм; длина хвоста
30—47 мм. Окраска довольно светлая — летом интенсивно палево-серо-бурая на спине
и белесая на брюхе. Малая Азия, Западный Кавказ, Черноморское побережье Кав-
каза. К этому же подвиду (в качестве особой natio) мы относим М. s. suramensis
Heptner, 1948, со средней части Главного Кавказского хребта и Малого Кавказа,
которая лишь на сериях слегка отличается более бледной окраской меха.
е) М. s. ciscaucasicus Ognev, 1924. (Тип из окрестностей г. Орджоникидзе.) Раз-
меры те же. Хвост короче: до 40 мм. Окраска более темная, буровато-серая без
ржавых оттенков. Северные склоны и предгорья Кавказа.
f) М. s. daghestanicus Schidlovskyi, 1919. (Тип из Гунибского района Дагестана.)
Размеры мельче: кондилобазальная длина черепа 21—22 мм; хвост до 40 мм. Окраска
•бледная, палево-серовато-бурая. Дагестан.
g) М. s. kaznakovi Satunin, 1903. (Тип с Талышских гор.) Хорошо обособленная
форма. Хвост короткий: около V4 длины тела. Окраска меха яркая, рыжевато-
коричневая на спине и боках и коричневато-серая на брюхе. Юго-Восточное
Закавказье.
Примечание. Остается неясным систематическое положение полевки Шел-
ковникова (М. schelkovnikovi Sat.), с Талыша, отличающейся, главным образом,
коротким хвостом.
РОД ЛЕСНЫЕ ПОЛЕВКИ. GENUS CLETHRIONOMYS Tilesius, 1850
Полевки мелкого или среднего размера. Длина хвоста составляет
1/4—7г длины тела. Глаза и ушные раковины развиты хорошо. Череп
без сагиттального гребня. Задняя часть нёба плоская, без боковых
ямок; задний край его резко обрывается над хоанами в виде полочки.
Он едва доходит до линии, соединяющей передние края последних ко-
ренных зубов в верхней челюсти. Коренные зубы взрослых с корнями,
В окраске меха спины большинства форм ясно выступают рыжие
цвета.
Распространены в Европе, Азии и Северной Америке.
В фауне СССР — 4 вида.
331
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ЛЕСНЫХ ПОЛЕВОК ФАУНЫ СССР
лишь низкие округлые валики, разделенные
Рис. 126. Черепа лесных полевок (сверху):
А — рыжая полевка (Clethrionomys glareolus); Б — красная
полевка (Cl. rutilus).
1(2). По бокам межглазничного промежутка черепа взрослых
особей идут два гребня, разделенные глубоким продольным желобком
между ними. Размеры относительно крупные: длина верхнего ряда
коренных зубов более 6 мм; кондилобазальная длина черепа взрослых
обычно более 25,5 мм. Верх головы и спина рыжеватые, а щеки и низ
боков тела серые...................................................
. . . . Красно-серая полевка. Clethrionomys rufocanus. (Стр. 332.)
2(1). Межглазничный промежуток гладкий или по бокам его идут
мелкой, слабо выражен-
ной ложбиной. Размеры
мельче: длина верхнего
ряда коренных зубов
менее 6 мм; кондилоба-
зальная длина черепа в
подавляющем большин-
стве случаев менее
25,5 мм. Щеки и бока
тела желтовато-охри-
стые, верх рыжий или
красноватый, или окра-
ска всего верха и боков
без рыжего тона (корич-
невато-бурая) . . . 3
3 (4). Длина хвоста
обычно более 40 мм
(очень редко от 35 мм).
Между относительно ред-
кими, плотно прилегаю-
щими волосами видна
чешуйчатая кожа хвоста.
Окраска хвоста резко
двухцветная: верх темно-бурый или черноватый, низ светлый; граница
между окраской верха и низа хвоста резкая .........................
...................Рыжая полевка. Clethrionomys glareolus. (Стр. 333.)
4(3), Длина хвоста не более 40 мм (обычно не более 35 мм).
Волосы на хвосте относительно длинные, густые, неплотно прилегаю-
щие, образующие пышную кисточку на конце; чешуйчатой кожи хво-
ста через волосяной покров не видно. Окраска хвоста обычно слабо
двухцветная: верх буровато-охристый (не черноватый), низ желтовато-
белесый; граница между окраской верха и низа хвоста обычно не
резкая .... Красная полевка, Clethrionomys rutilus. (Стр. 334.)1
102, Красно-серая полевка, Clethrionomys rufocanus Sundevall, 1846
Табл. XXXVIII, 3. Карта 106
Северная Скандинавия, северные районы Европейской части
СССР, почти вся Сибирь и Дальний Восток. Южная граница распро-
странения тянется от Средней Карелии, через Горьковскую область,
Южный Урал, Алтай, Северную Монголию и Северо-Восточный Китай.
Распространена преимущественно в таежных районах, но заходит
в лесотундру и даже в тундру. Летом питается зелеными частями
1 А. П. Кузякиным установлено, что на о. Шикотан живет Clethrionomys (Neo-
aschizomys) sikotanensis Tokuda, 1935 (табл. XXXVIII, /, 2).
332
трав, ягодами, грибами, кедровыми орешками, семенами. Зимой гры-
зет побеги и кору кустарников и деревцев. Самки дают несколько по-
метов в год, число детенышей в помете колеблется от 3 до 13. Один
из важных кормовых объектов ценных пушных зверей — соболя, ли-
сицы, колонка, горностая и др. Заметного вреда не причиняет.
Подвиды (наиболее резко обособленные):
а) С1. г. rufocanus Sundevall, 1846. (Тип из Северной Швеции.) Рыжая окраска на
спине тянется узкой полосой; бока и щеки чисто серые. Северные районы Европы.
b) С1. г. ircutensis Ognev, 1924. (Тип с хребта Хамар-Дабан.) Полоса рыжего
цвета довольно широкая. Бока и щеки с желтоватым оттенком. Сибирь, южная
часть Дальнего Востока.
с) С1. г. smithii Thomas, 1905. (Тип с острова Хондо, в Японии.) Отличается от
предыдущих более темной окраской меха. Длина хвоста до 63 мм (у номинальной
формы до 43 мм, у сибирских до 52 мм). Япония, Сахалин, Шантарские острова, побе-
режье Охотского моря.
Рис. 127. Задние коренные зубы лесных полевок (правый зубной ряд):
а, б, в — шикотанская полевка (Clethrionomys sikotanensis); г, д, е — красно-серая полевка (Cl. rufoca-
nus); ж, з, «—рыжая полевка (Cl. glareolus); к, л, м — красная полевка (Cl. rutilus).
d) Cl. г. wosnessenskii Poljakov, 1881. (Тип с Камчатки.) Окраска спины тусклая,
рыжие оттенки не яркие, полоса рыжего меха на спине узкая. Камчатка, бассейн
р. Колымы, Анадырский край, о. Парамушир.
103. Рыжая полевка. Clethrionomys glareolus Schreber, 1780
Табл. XXXVIII, 4—6. Карта 106
Населяет лесные ландшафты почти всей Западной Европы, таеж-
ной и лесотундровой зоны Европейской части СССР и Западной Си-
бири (до Алтая), гор Средней Азии, Малой Азии и Закавказья.
Держится в лесах, в кустарниках, в садах, по оврагам, долинам
рек. Живет в неглубоких норках. За лето приносит несколько пометов
(на севере 2—3, на юге 3—4), из 3—9 детенышей в каждом. Питается
летом травой, семенами деревьев, желудями, орехами, ягодами. Зимой
грызет побеги и кору кустарников и деревьев, чем причиняет некото-
рый вред садам и лесным насаждениям. Важный объект питания лес-
ной куницы, норки, черного хорька и других лесных хищников.
333
В СССР встречаются следующие подвиды:
a) Cl. g. glareolus Schreber, 1780. (Тип из Дании.) Размеры относительно мел-
кие: кондилобазальная длина черепа 22—25 мм. Окраска яркая, с заметным раз-
витием красновато-рыжих тонов. Западные и центральные области СССР.
b) Cl. g. istericus Miller, 1909. (Тип из Венгрии.) Размеры, как у предыдущей
формы. Окраска более светлая, ржаво-желтоватая. Молдавия, северные области
Украины, центрально-черноземные области, Среднее Поволжье.
с) Cl. g. suecicus Miller, 1900. (Описан по экземпляру из Швеции.) Размеры круп-
ные: длина тела до 12 см; кондилобазальная длина черепа 23—26 мм. Окраска тем-
ная. Спина в летнем мехе буровато-коричневая. Север Европейской части СССР
(к северу от линии: Эстония — Калининская, Ивановская и Горьковская области —
Южный Урал — Западная Сибирь). Синоним ее: Cl. g. devius Stroganov, 1948 (с ни-
зовьев Печоры).
d) Cl. g. saianicus Thomas, 1911. (Описана по экземпляру с Саянских гор.)
Кондилобазальная длина черепа 22—24 мм. Окраска еще темнее, чем у североевро-
пейской формы. Саяны и Алтай (синоним: Cl. g. sibiricus Egorin, 1936).
е) Cl. g. frater Thomas, 1908. (Тип с Тянь-Шаня.) Окраска меха еще темнее, чем
у алтайско-саянской формы; рыжеватый тон едва выступает.
f) Cl. g. ponticus Thomas, 1906. (Тип из Турции.) Окраска верха интенсивная,
серо-бурая с коричневато-ржавым оттенком по хребту. Турция; в СССР найдена
в Аджарии, к югу от г. Кутаиси.
104. Красная полевка. Clethrionomys rutilus Pallas, 1779
Табл. XXXVIII, 7. Карта 106
Север Европы, почти вся Сибирь, Дальний Восток и Северная Аме-
рика. Южная граница ареала идет от Южной Карелии, через Калинин-
скую, Горьковскую области, Татарию, Башкирию, Южный Урал, север-
ные области Казахстана, Северную Монголию и Северо-Восточный Ки-
тай. На севере заходит в лесотундровую зону и кустарниковые тундры.
Населяет леса (особенно таежные), лесные острова в лесостепи,,
кустарниковые заросли, кусты по долинам рек, лесные болота и дру-
гие подобные угодья. На севере иногда селится в домах и амбарах.
Свои гнезда устраивает в неглубоких норах, среди корней деревьев и
кустарников, под камнями, в кочках, в пустотах поваленных стволов и
в низко расположенных дуплах. Число детенышей в помете от 3 до 9
(обычно 6—7), число пометов 3—4 в год. Летом питается преимущест-
венно семенами хвойных и ягодами, в меньшей степени зелеными ча-
стями растений. Зимой грызет кору и веточки древесных растений.
Один из основных кормовых объектов ценцых пушных хищников в лес-
ных ландшафтах от лесотундры до лесостепи.
Многие из описанных форм в дальнейшем, вероятно, будут при-
знаны синонимами других подвидов:
a) Cl. г. rossicus Dukelsky, 1928. (Описана по экземпляру с Южного Урала.)
Длина хвоста 26—42 мм; кондилобазальная длина черепа 23—26 мм. Летний мех
на спине темный с преобладанием красно-коричневых токов. Европейская часть СССР.
b) Cl. г. rutilus Pallas, 1778. (Описана по экземпляру со среднего течения р. Оби.)
Размеры тела и черепа те же, что у предыдущей формы. Хвост длиннее: до 50 мм.
Окраска летнего меха на спине относительно светлая и чистая, ржаво-коричневая.
Брюшко белое. Западная Сибирь от Урала до Енисея. К синонимам относится: С1. г.
uralensis Koljuschev et Vinogradov, 1936, описана по экземпляру с р. Малой Сосьвы,
на Урале.
с) Cl. г. vinogradovi Naumov, 1938. (Тип с р. Нижней Тунгуски.) Длина хвоста
28—48 мм; кондилобазальная длина черепа 22—26 мм. Межглазничный промежуток
гладкий. Окраска летнего меха более цветистая, чем у номинальной расы: ржаво-
красновато-коричневая. Эта форма трудно отличима от номинальной. Бассейн сред-
него и нпжиего течения р. Енисея.
d) Cl. г. jacutensis Vinogradov, 1927. (Тип из г. Якутска.) Хвост короткий:
28—38 мм. Череп массивный, широкоскулый, по его межглазничному промежутку тя-
нется продольная ложбинка. Окраска яркая с насыщенными красноватыми оттенками,
бока огненно-желтые. Брюшко белое. Центральная Якутия, низовья р. Лены.
е) Cl. г. jochelsoni Allen, 1903. (Тип из окрестностей г. Верхне-Колымска.) Хвост
до 35 мм. Череп мельче, со слабо выраженной ложбиной посередине межглазнпчного
334
промежутка. Окраска спины в летнем мехе ржаво-охристая. Зимний мех очень
пышный* Бассейны рек Яйвы, Индигирки, Колымы, Анадыря; Камчатка.
f) Cl. г. amurensis Schrenk, 1858. (Тип из окрестностей г. Николаевска-на-Амуре.)
Размеры чуть крупнее: кондилобазальная длина черепа 23—27 мм. Хвост длинный:
35—47 леи. Окраска спины летом интенсивная, буро-красновато-коричневая. Брюшко
сероватое. Прибайкалье, Забайкалье, Амурский край, Приморье, Сахалин. И. М* Гро-
мов (1963) относит к этому же подвиду Cl. rjabovi Beljaeva, 1953, описанную по экзем-
пляру из окрестностей с. Зорино на Амуре.
g) CL г. lategriseus Argiropulo et Afanasiev, 1939. (Тип из окрестностей г. Карка-
ралинска.) Вдоль спины тянется узкая полоса коричнево-бурого цвета; серый цвет
боков поднимается кверху на спину. Казахстанское нагорье.
РОД КАМЕННЫЕ ПОЛЕВКИ1. GENUS ALTICOLA Blanford, 1881
Полевки мелкого и среднего размера с густым мехом, обычно пе-
пельно-серой или серебристо-серой окраски, часто с буроватым нале-
том* Хвост обычно густо покрыт волосами, образующими на конце
кисточку* Длина хвоста всегда превышает длину задней ступни и
у разных видов колеблется от 7? до V2 длины тела* Вибриссы очень
длинные: значительно длиннее головы* Первый палец передних конеч-
ностей зачаточный. Череп имеет тенденцию к уплощению* Задний край
твердого нёба почти доходит до линии, соединяющей передние края
задних коренных зубов верхней челюсти* Коренные зубы в течение
всей жизни не имеют корней*
От Тянь-Шаня до Чукотки и низовьев Лены и от гор Южной Си-
бири до Гималаев включительно.
Всего 5 видов; в фауне СССР — 4 вида, относящиеся к двум под-
родам*
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ КАМЕННЫХ ПОЛЕВОК ФАУНЫ СССР
1(2)* Череп очень сильно уплощен: ширина мозговой коробки
в 2 раза больше ее высоты..........................................
................Плоскочерепная полевка. Alticola strelzovi* (Стр 335.)
2(1)* Череп уплощен слабо или не уплощен: ширина мозговой ко-
робки лишь немного больше ее высоты..............................3
3(6). Хвост весь белый или лишь слегка двухцветный. Передняя
петля последнего коренного зуба верхней челюсти не замкнута и сое-
диняется с передним наружным треугольником.......................4
4(5). Хвост составляет 7з (или более) длины тела* Задний верх-
ний коренной зуб с внутренней стороны имеет 3 выдающихся угла*
....................Серебристая полевка. Alticola roylei. (Стр. 336.)
5(4). Хвост короче 7з длины тела* Задний верхний коренной зуб
с внутренней стороны имеет только 2 выдающихся угла................
. . * . Гоби-Алтайская полевка. Alticola barakshin* (Стр* 337*)
6(3)* Хвост резко двухцветный* Передняя петля последнего корен-
ного зуба верхней челюсти замкнутая................................
...............Большеухая полевка. Alticola macrotis* (Стр* 337.)
Подрод плоскочерепных полевок. Subgenus Platycranius
Kastschenko, 1901
105. Плоскочерепная полевка. Alticola strelzovi Kastschenko, 1900
Табл* XXXVIII, 8* Карта 107
Распространена в Казахском мелкосопочнике (Карагандинская,
и южная часть Павлодарской области), на хребте Тарбагатай,
в Зайсанской котловине, на Алтае к югу от Семинского перевала
1 Описание этого рода составлено по материалам, критически обработанным
А. П. Шведовым.
335
и в западной части Тувинской автономной области. За пределами
СССР — в северо-западной части Монгольского Алтая, к востоку до
меридиана Кобдо.
Заселяет скалы и каменные россыпи в степи и полупустыне, а
также горные каменистые степи; на Алтае была отмечена также в гор-
ной каменистой тундре. Устраивает своеобразные защитные сооруже-
ния из небольших камней, скрепляя их своим пометом. Питается тра-
вой, побегами и листьями кустарников, лишайниками и, возможно, неко-
торыми насекомыми. Как и другие виды этого рода, запасает себе корм
на зиму, высушив его предварительно на солнце. Запасы одной семьи
составляют несколько килограммов сена. Приносит 2—3 помета за
лето, от 3 до 12 (чаще 6—8) детенышей в каждом. Молодые первого
года в размножении участия, по-видимому, не принимают. В скотовод-
ческих районах в отдельные годы может наносить вред на пастбищах
и сенокосах.
Описано 3 подвида, незначительно различающихся по окраске меха:
a) A. s. strelzovi Kastschenko, 1900. (Тип с оз. Теньга, на Алтае.) Окраска спины
относительно темная, серая с буроватым оттенком. Центральный Алтай, Сауры,
Тарбагатай.
b) A. s. desertorum Kastschenko, 1901. (Описан, по-видимому, по экземпляру из
района г. Каркаралинска.) Окраска более светлая, палево-серая. Мелкосопочник
Казахстана.
с) A. s. depressus Ognev, 1944. (Тип из Чуйской нагорной степи.) Окраска светлая,
сероватая; брюхо чисто белое (у предыдущих форм с палевым налетом). Западная
Монголия; в СССР — Чуйская нагорная степь.
Подрод к а м е н н ы х п о л е в о к. Subgenus Alticola Blanford, 1881
106, Серебристая полевка. Alticola roylei Gray, 1842
Табл. XXXVIII, 9—10. Карта 107
Горы Памиро-Алая и Тянь-Шаня, Джунгарского Алатау, Тарба-
гатая и Саура. На юге Сибири обитает в Туве и проникает в Мину-
синскую котловину. За пределами СССР — в Северо-Западных Гима-
лаях и в Монголии.
Живет в альпийском, субальпийском и отчасти в лесном поясе гор,
но в любых условиях обязательно придерживается скал и россыпей
камней.
Активна в течение всего года: более деятельна в светлые часы
суток, чем ночью. Питается травянистыми растениями, веточками не-
которых кустарников и лишайниками. Запасает на зиму сено; отгры-
зенные части растений просушивает на солнце, а затем убирает в щели
между камнями. В течение лета дает 2—3 помета, по 3—10 детенышей
(обычно 4—5) в каждом. Служит кормом для некоторых пушных
зверей.
На сериях можно различить по тону окраски и размерам следующие формы:
а) А. г. argentatus Severtzov, 1879. (Тип с Аличура, на Памире.) 'Размеры отно-
сительно крупные. Окраска летнего меха спины серо-палевая, с преобладанием
серых, серебристых оттенков. Памир и Гиссар.
Ь) А. г. rosanovi Ognev, 1940. (Тип из Алайской долины.) Размеры немного
меньше. В окраске летнего меха спины ясно выступают ржаво-палевые оттенки.
Алайская долина и Заалайский хребет.
с) А. г. leucurus Severtzov, 1873. (Тип с горы Масат, близ Чимкента.) Размеры
те же. В окраске спины рыжевато-палевые оттенки выражены слабо. Горы
Тянь-Шаня.
d) А. г. saurica Afanasiev et Bachanov, 1948. (Тип с гор Саур.) От предыдущей
формы отличается более темной окраской меха. Сауры, Тарбагатай, Монрак.
е) А. г. tuvinicus Ognev, 1950. (Тип из окрестностей г. Кызыла.) Отличается ог
предыдущих форм более сложным строением верхнего заднего коренного зуба,
336
имеющего с наружной стороны 4 (а не 3) выдающихся угла. Горы юго-западной части
Тувинской автономной области.
f) А. г. olchonensis Litvinov, 1960. (Тип с острова Ольхон, на Байкале.) Окраска
песчаная. Хвост явно двухцветный. Остров Ольхон, па Байкале.
107. Гоби-Алтайская полевка. Alticola barakshin Bannikov, 1954
Карта 107
Распространена в горах Монголии. В СССР встречена только на
юге Тувинской области. Образ жизни в условиях Тувы не изучен.
108. Большеухая полевка. Alticola macrotis Radde, 1861
Табл, XXXVIII, 11—12, Карта 107
Распространена в горах Южной и Восточной Сибири,
Приурочена к каменистой горной тундре, россыпям камней среди
высокогорных лугов и кустарниковых зарослей. Взрослые самки успе-
вают принести за лето 1—2 помета на севере и 2—3 на юге, по 4—7
детенышей в каждом. Питается травой, молодыми побегами и листь-
ями кустарников, лишайниками. На зиму запасает сено. Имеет неко-
торое кормовое значение для соболя и горностая,
Можно выделить 3 подвида:
а) А. m. macrotis Radde, 1861. (Тип с Восточных Саян, Прибайкалье.) Хвост от-
носительно короткий: обычно менее 28 мм. Входящие углы наружной п внутренней
стенок заднего верхнего коренного зуба обычно не соприкасаются друг с другом,
окружая одно замкнутое пространство, Саяны, Прибайкалье.
b) A. m. vinogradovi Rasorenova, 1933. (Тип с Алтая.) Хвост немного длиннее:
обычно более 28 мм. Входящие углы наружной и внутренней стенок заднего корен-
ного зуба обычно соприкасаются, образуя 4—5 замкнутых пространств. Алтай.
с) A. m. lemminus Miller, 1898. (Описан по экземпляру с берегов залива Кре-
ста.) Своеобразная форма: с очень коротким хвостом, густо покрытым длинными
волосками, на зиму белеет. Задний верхний коренной зуб обычно с дополнитель-
ными зубцами на заднем конце. Горные тундры Северо-Восточной Сибири.
Примечание. До последнего времени эти три подвида большеухой полевки
считали самостоятельными видами. Но изучение индивидуальной изменчивости осо-
бей этих форм привело к заключению о их большой близости и нестойкости
отличительных признаков. Поэтому мы, как и И. М. Громов, рассматриваем их
лишь как подвиды одного вида.
РОД ЛЕСНЫЕ ЛЕММИНГИ. GENUS MYOPUS Miller, 1910
Небольшие грызуны (длина тела около 10—12 см) с коротким
хвостом и маленькими ступнями, подошвы которых (в противополож-
ность другим леммингам) не покрыты шерстью. Коготь первого (внут-
реннего) пальца передних лап крупный, уплощенный, с параллельными
боковыми краями и с вырезкой на конце (рис. 116). Ушные раковины
относительно велики и выдаются из шерсти, Окраска темно-аспидно-
серая с ржавым пятном на спине,
Только один вид,
109. Лесной лемминг. Myopus schisticolor Lilljeborg, 1884
Табл. XXXIX, /, Карта 108
От Северной Скандинавии по зоне тайги до Тихого океана.
В СССР найден в ряде пунктов севера Европейской части страны, Си-
бири и Дальнего Востока.
22 н. А. Бобринский и др.
337
Житель таежных лесов. Чаще всего держится на моховых боло-
тах. Повсеместно малочислен. Охотно поедает мхи, но ест и лесные
травы. За лето приносит несколько пометов. Хозяйственного значе-
ния не имеет.
РОД НАСТОЯЩИЕ ЛЕММИНГИ. GENUS LEMMUS Link, 1795
Полевки с очень коротким хвостом, длина которого меньше длины
задней ступни. Подошвы лап покрыты густой шерстью. Ушные рако-
вины малы и скрыты в мехе. Коготь первого пальца передних лап
велик, уплощен и имеет выемку на конце.
Тундры и лесотундры Европы, Азии и Северной Америки, а также
тайга Восточной Сибири.
Из 4 или 5 видов в фауне СССР — 3 вида.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ НАСТОЯЩИХ ЛЕММИНГОВ
ФАУНЫ СССР
1(4). Окраска меха без резко очерченного черного пятна на верх-
ней стороне головы и передней части спины. Иногда вдоль спины про-
ходит узкая темная полоска ................................... 2
2(3). Размеры относительно мелкие: длина задней ступни менее
15,5 мм; кондилобазальная длина черепа менее 28 мм...............
..................Амурский лемминг. Lemmus amurensis. (Стр. 338.)
3(2). Размеры крупнее: длина задней ступни более 15,5 мм; кон-
дилобазальная длина черепа взрослых особей более 28 мм...........
............Сибирский лемминг. Lemmus sibiricus. (Стр. 338.)
4(1). Окраска меха пестрая. От носа до лопаток простирается
резко ограниченное черное пятно, на фоне которого над глазами и
ушами расположены желтые пятна. Остальная часть спины желто-
вато-коричневая с темными пятнами или почти вся спина черная
...............Норвежский лемминг. Lemmus lemmus. (Стр. 339.)
ПО. Амурский лемминг. Lemmus amurensis Vinogradov, 1924
Табл. XXXIX, 2. Карта 108
Очень редкий вид, найден только в следующих четырех пунктах:
1) с. Пикан, на р. Зее Амурской области; 2) верховья р. Нельгехе,
в юго-восточной части Верхоянского хребта, у 63,5° с. ш.; 3) окрест-
ности пос. Александровский завод Читинской области; 4) голец Соп-
чикан, по р. Тершнаку, левому притоку р. Читы.
Найден в заболоченных участках тайги и гольцов.
111. Сибирский лемминг. Lemmus sibiricus Kerr, 1792
Табл. XXXIX, 5, 4, Карта 108
Тундра и лесотундра севера Европы (к востоку от Горла Белого
моря), Сибири и Дальнего Востока, а также на островах: Диксон,
Врангеля и Новосибирского архипелага.
Обитает в тундре и на тундровых участках лесотундровой зоны,
преимущественно по долинам рек, в зарослях ивы и карликовой бе-
резки. Летом живет в разветвленных норках, вырытых в земле или
в моховом покрове. Зимой роется под снегом. Селится обычно коло-
ниями. Дает за год несколько пометов. Иногда приносит детенышей
338
даже в зимнее время в гнездах из травы и мха под снегом. Числен-
ность сильно колеблется по годам. Служит пищей песцу и другим
тундровым хищникам.
Подвиды:
a) L. s. sibiricus Kerr, 1792. (Описан по экземпляру с Полярного Урала.) Летний
мех на спине ржаво-буроватый, обычно с темным ремнем по хребту. Темная окраска’
верха головы резко отграничена от белых щек. Зимний мех рыжеватый, без темной
полосы на спине. Тундра, от Горла Белого моря до дельты р. Лены включительно.
b) L. s. novosibiricus Vinogradov, 1924. (Тип с острова Котельного.) Летний мех,
как у типичной формы, но коричневая окраска верха головы не резко отграничена
от рыжеватой расцветки щек. Зимний мех очень светлый, беловатый. Новосибир-
ские острова.
с) L. s. chrysogaster Allen, 1903. (Тип из окрестностей пос. Гижиги.) Мельче пре-
дыдущих рас. Летний мех спины буровато-охристый, без темной продольной полоски.
Брюшко охристое. Тундры севера Дальнего Востока.
По-видимому, особая форма сибирского лемминга обитает на о-ве Врангеля.
112. Норвежский лемминг. Lemmus lemmus Linnaeus, 1758
Табл. XXXIX, 5. Карта 108
Распространен в лесотундре и горных тундрах Северной Сканди-
навии и Кольского полуострова.
Живет обычно колониями. Численность сильно колеблется по го-
дам. Иногда предпринимает далекие массовые миграции. Зимой про-
кладывает ходы под снегом. Размножается несколько раз в году, при-
нося по 4—10 детенышей. Питается мхами, травой и веточками кустар-
ников.
РОД КОПЫТНЫЕ ЛЕММИНГИ GENUS DICROSTONYX Gloger, 1841
В отличие от настоящих леммингов, ушные раковины не развиты:
они имеют вид небольшой складочки кожи вокруг слухового прохода;
коготь первого пальца передних ног конической формы; другие когти
пальцев передних конечностей летом имеют обычную для полевок
форму, а к зиме резко увеличиваются в своеобразные копытца. Для
черепа характерно наличие тупых конических заглазничных отростков.
Распространен в тундрах Евразии и Северной Америки.
Один вид.
113. Копытный лемминг. Dicrostonyx torquatus Pallas, 1779
Табл. XXXIX, 6—8. Карта 108
Распространен в тундрах Восточной Европы, Азии и Северной
Америки. В СССР населяет всю тундровую и лесотундровую зоны на
протяжении от Горла Белого моря до Берингова пролива.
Селится, главным образом, на участках каменистой и песчаной
тундр. Живет в норках или под камнями, в трещинах скал, в кочках.
Зимой роется под снегом. Приносит в течение года несколько пометов,
по 3—11 (обычно 4—6) детенышей в каждом. Питается побегами кар-
ликовой березки, ивы, веточками голубики и морошки, грибами, тра-
вой. Служит основной пищей песцу.
Подвиды:
a) D. t. torquatus Pallas, 1779. (Описан по экземпляру из устья р. Оби.) Окраска
спины в летнем мехе более или менее однотонная, буровато-рыжая, с ясно выра-
женной темной полоской по хребту. Плечи того же цвета, что и спина. Тундры
Восточной Европы и Западной Сибири.
22*
339
b) D. t. ungulatus Baer, 1841. (Тип с Новой Земли.) Окраска летнего меха на
спине более пестрая и с более выступающими серыми оттенками. Хребтовая темная
полоска не развита. Острова Новой Земли.
с) D. t. chionopaes Gl. Allen, 1914. (Тип из окрестностей г. Нижне-Колымска.)
Окраска летнего меха на спине более насыщенная и яркая, чем у типичной формы.
Темная полоска на спине обычно не выражена. Плечи ярко-рыжие. Тундры от Тай-
мыра до Чукотки.
d) D. t. vinogradovi Ognev, 1948. (Тип с о-ва Врангеля.) Окраска спины летом
пепельно-серая с каштаново-бурой крапчатостью. Ремня по хребту нет. Череп круп-
нее, чем у других рас. Остров Врангеля.
РОД ПЕСТРУШКИ. GENUS LAGURUS Gloger, 1842
Мелкие и средних размеров полевки с очень коротким хвостом и
небольшими лапками, подошвы которых покрыты густым мехом. Уш-
ные раковины нормального строения, но малы. Когти острые, кониче-
ские. Череп несет конические заднеглазничные отростки. Эмалевые
петли на жевательной поверхности коренных зубов имеют вид замкну-
тых треугольников.
Водятся в степях и пустынях Восточной Европы, Западной Си-
бири, Казахстана и Центральной Азии.
Два вида.
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ СТЕПНЫХ ПЕСТРУШЕК
ФАУНЫ СССР
1(2). Спина светло-серая или буровато-серая с черной полоской
по хребту. Кондилобазальная длина черепа менее 27 мм ................
..................Степная пеструшка. Lagurus lagurus. (Стр. 340.)
2(1). Спина песчано-желтого цвета; черной полоски вдоль хребта
нет. Кондилобазальная длина черепа более 27 мм ......................
Желтая пеструшка. Lagurus luteus. (Стр. 341.)
114. Степная пеструшка. Lagurus lagurus Pallas, 1773
Табл. XXXIX, 9. Карта 109
Распространена на юге Европейской части СССР (к востоку от
Днепра и к югу от Рязанской и Горьковской областей), в степных райо-
нах Западной Сибири (на восток до р. Енисея) и большей части терри-
тории Казахстана; водится также в Джунгарии и Западной Монголии.
Обитает на участках степей и глинистых пустынь различных ти-
пов, но преимущественно ковыльно-типчаковых и полынных. Селится
среди целинной степи, на пашнях, выгонах, толоках. Живет обычно
колониями. Норы зверька состоят из разветвленной сети ходов с рядом
гнездовых камер и многими входами. За лето дает до 5 пометов; число
детенышей в них колеблется от 3 до 7. В некоторые годы наблюдается
массовое размножение пеструшек и их далекие миграции. Питается
травянистой растительностью. Местами сильно вредит посевам.
Подвиды:
a) L. 1. lagurus Pallas, 1773. (Описан по экземпляру из степей по р. Уралу.)
Общий тон летнего меха па спине палево-серый, средней темноты, иногда с рыжим
оттенком. Спинной ремень узкий. Распространен в большей части Казахстана.
b) L. 1. agressus Serebrennikov, 1929. (Тип из Бузулукского района.) Окраска бо-
лее темная, серая; ремень на спине широкий. Юг Европейской части СССР.
с) L. 1. abacanicus Serebrennikov, 1929. (Тип из Минусинской степи.) Окраска свет-
лая, сероватая с палевым оттенком. Черный ремень узок, но отчетлив. Отличается
от предыдущих рас более крупным черепом и удлиненностью петель коренных зубов.
Степи по среднему и нижнему течению р. Абакана.
340
cl) L. 1. altorum Thomas, 1912. (Тип с хребта Барлык, в Джунгарии.) Окраска
спины очень светлая, палево-серая. Черноватая полоска на спине очень тонкая.
Широко распространена в Центральной Азии, в СССР — в Семиречье.
Желтая пеструшка. Lagurus luteus Eversmann, 1840
Табл. XXXIX, 10
Прежде была широко распространена по степям Казахстана,
Джунгарии и Монголии. Но в последние десятилетия в большинстве
областей Казахской ССР этот зверек по неизвестным причинам вымер.
Возможно, что в некоторых районах Центрального Казахстана он
в небольшом количестве все же сохранился; об этом свидетельствуют
найденные там относительно свежие черепа. В Джунгарии желтая
пеструшка обитает и в настоящее время.
Обитатель солянковых и полынно-солянковых пустынь, полупу-
стынь и сухих степей.
РОД ПРОМЕТЕЕВЫ ПОЛЕВКИ. GENUS PROMETHEOMYS Satunin, 1901
Средних размеров полевки (длина тела 13—16 см) с относительно
длинным хвостом (длина его обычно равна ’/3 длины тела). Глаза
очень малы. Когти пальцев передних лап настолько длинны, что пре-
вышают длину самих пальцев. Ушные раковины малы. Задний корен-
ной зуб верхней челюсти относительно простого строения: с наружной
стороны он имеет 2 выдающихся угла и 1 складку эмалевой стенки
между ними, а на наружной стороне 3 выдающихся угла и 2 впадины
между ними.
Эндемик Кавказа. Единственный вид.
115. Прометеева полевка. Prometheomys schaposchnikovi Satunin, 1901
Табл. XL, /, 2. Карта 109
Населяет альпийские и субальпийские луга и лужайки лесного
пояса Главного Кавказского хребта, от Черноморского побережья па
восток до Военно-Грузинской дороги.
Ведет подземный образ жизни; роет сложно устроенные норы,
землю из которых выбрасывает кучками на поверхность. За лето дает
1 или 2 помета. Кормится корнями и луковицами растений, иногда
затаскивает в норы и зеленые части трав. Местами портит горные се-
нокосы своей роящей деятельностью и поеданием травы.
РОД СЛЕПУШОНКИ, GENUS ELLOBIUS Fischer, 1814
Небольшие грызуны, приспособленные к подземному образу жизни.
Тело вальковатое, голова округлая, с торчащими изо рта большими
белыми резцами. Хвост очень короткий. Ушные раковины редуциро-
ваны, Глаза малы. Мех бархатистый. Резцы верхней челюсти (основ-
ные орудия рытья нор) очень велики и направлены вперед и вниз. Эма-
левые петли на жующей поверхности коренных зубов не образуют
замкнутых треугольников.
Обитает в степях Юго-Восточной Европы, Казахстане, Централь-
ной, Средней и Передней Азии.
Два вида.
341
Рис. 128. Черепа слепушонок (сверху):
4 — слепушонки обыкновенной (Ellobius talpinus); Б — афганской
слепушонки (Ellobius fuscocapillus).
• ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ СЛЕПУШОНОК ФАУНЫ СССР
1(2). Сагиттальный гребень отсутствует даже у взрослых особен.
Межтеменная кость всегда имеется, и границы ее сохраняются даже
у старых зверьков................................................
..........Обыкновенная слепушонка. Ellobius talpinus. (Стр. 342.)
2(1). Сагиттальный гребень у взрослых особей развит сильно.
Межтеменная кость на черепе взрослых особей сливается с соседними
костями черепа, и границы ее исчезают............................
. Афганская слепушонка. Ellobius fuscocapillus. (Стр. 343.)
116. Обыкновенная слепушонка. Ellobius talpinus Pallas, 1770
Табл. XL, 3—5. Карта 109
Распространена в степях Крыма, Южной Украины, Предкавказья,
Калмыкии, Нижнего Поволжья, Южного Урала, Зауралья, предалтай-
ских районов, почти всего Казахстана, а также в горах и равнинах
Средней Азии.
Селится в сухих степях, полупустынях и пустынях (преимущест-
венно глинистых), на травянистых склонах гор, на лугах, посевах трав,
в огородах и садах. Жизнь проходит под землей, в сложной системе
поверхностных и глубинных (обычно ведущих к гнезду) ходов. Роя
поверхностные ходы, слепушонка подрывает корни, луковицы и кор-
невища травянистых растений, которыми питается. Нарытую землю
выбрасывает через боковые отнорки наружу. Дает, по-видимому, 2
помета в год, из 1—7 (чаще 3—5) детенышей в каждом. В некоторых
районах приносит вред своей роющей деятельностью, повреждая куль-
турные растения.
Описано большое количество подвидов обыкновенной слепушонки,
отличающихся, главным образом, окраской меха. Зная огромную из-
менчивость окраски этих подземных грызунов, мы считаем, что боль-
шинство разновидностей носит индивидуальный, а не географический
характер. Впредь до глубокого и широкого анализа географической
изменчивости описываемого грызуна мы воздерживаемся от описания
его отдельных подвидов.
342
117. Афганская слепушонка. Ellobius fuscocapillus Blyth, 1841
Табл. XL, 6. Карта 109
Распространена в Афганистане, Иране, Белуджистане и Турции.
В СССР встречается в горах Копет-Дага и их предгорьях, в Бадхызе
и в Юго-Восточном Закавказье (Талыш, окрестности г. Еревана).
Селится на травянистых склонах гор. По образу жизни в основ-
ном сходна с обыкновенной слепушонкой.
Два резко выраженных подвида:
а) Е. f. fuscocapillus Blyth, 1841. (Описан по экземпляру из Белуджистана.) Сагит-
тальный гребень, не ветвясь, тянется от мсжглазннчного промежутка до затылка.
Афганистан, Восточный Иран, Южная Туркмения.
b) Е. f. lutescens Thomas, 1897. (Тип из Вана, в Турции.) Сагиттальный гребень
в области лобно-теменного шва делится на две ветви, расходящиеся кзади. Запад-
ный Иран, Турция, Юго-Восточное Закавказье.
Подсемейство ЦОКОРОВ. Subfamilia MYOSPALACINAE’
Сравнительно крупные грызуны (длина тела от 20 до 25 см), ве-
дущие подземный образ жизни. Тело цилиндрическое, шея короткая,
голова широкая, хвост небольшой, конический, покрытый короткими
волосами или почти голый. Лапы укороченные, пальцы передних лап
несут огромные когти, длина которых превышает длину пальцев. Глаза
развиты слабо. Ушные раковины отсутствуют. Коренные зубы без кор-
ней; жевательная поверхность их плоская, разделенная выдающимися
с боков углами эмалевой стенки на ряд треугольников.
Один род.
Рис. 129. Коренные зубы цокоров (варианты индивидуальной изменчивости):
верхний ряд —левый первый, (передний) верхний коренной зуб (Л1‘); средний р я д — ле-
вый третий (задний) верхний коренной зуб (Л13); нижний р я д — правый третий (задний)
коренной зуб нижней челюсти (ш3).
343
РОД ЦОКОРЫ. GENUS MYOSPALAX Laxmann, 1780
Встречается на Алтае и в приалтайских районах Западной Сибири,
в Забайкалье, Амуро-Уссурийском крае, в Северной Монголии, боль-
шей части Китая и в Корее.
До последнего времени считалось, что на территории СССР
водится три вида цокоров: алтайский — М. myospalax Laxmann. даур-
ский— М. aspalax Pallas и северокитайский — М. psilurus Milne-Ed-
wards. Но исследования А. П. Кузякина, Юй Вэнь-дао и Ли Жун-
гуана, располагавших большим материалом по цокорам как Совет-
ского Союза, так Китая и Монголии, показали, что отличия этих форм
весьма не стойки, что между ними существует немало особей с пере-
ходными признаками. По мнению этих исследователей, указанные
формы цокоров фауны СССР следует считать лишь за подвиды одного
вида.
118. Цокор обыкновенный. Myospalax myospalax Laxmann, 1773
Табл. XL, 7—9. Карта ПО
Распространен на Алтае и в прилегающих к нему районах Запад-
ной Сибири, на Тарбагатае, в Забайкалье, Северо-Восточной Монго-
лии, по р. Амуру, в Приморье, Корее и в Северо-Восточном Китае.
На Алтае и Тарбагатае селится, главным образом, на луговых и
горно-степных участках с мягким грунтом и богатой растительностью.
В Забайкалье и по Амуру он держится по опушкам лесных островов,
на лугах, выгонах, старых парах. Ведет подземный образ жизни.
Сложные норы цокора состоят из закрытых поверхностных ходов
с большими выбросами земли и глубинных лазов, ведущих к гнезду.
Весной приносит помет из 3—6 детенышей. На зиму в спячку не впа-
дает. Питается как подземными, так и наземными частями растений.
Приносит вред сенокосам как своей роющей деятельностью, так и
уничтожением травостоя. Местами повреждает огородные культуры.
Подвиды:
а) М. m. myospalax Laxmann, 1780. (Описан по экземпляру из окрестностей
г. Барнаула.) Окраска спины насыщенная, буровато-серая, обычно с охристо-ржавым
налетом. Бока охристые. Алтай и его предгорья, Тарбагатай, юг Томской области.
М. m. incertus Ognev, 1936 (из окрестностей Катон-Карагая), и М. m. tarbagataicus
Ognev, 1936 (из Тарбагатая), не имеют устойчивых различий для обособления
в качестве подвидов.
b) М. m. aspalax Pallas, 1778. (Описан по экземпляру из Южного Забайкалья.)
Спина обычно красивого, серебристо-серого цвета. Бока беловатые. На внутренней
стороне первого коренного зуба только один вдающийся внутрь угол (у номиналь-
ной формы их два, один или ни одного). Южное Забайкалье, Монгольская Народная
Республика и Внутренняя Монголия.
с) М. m. epsilanus Thomas, 1778. (Тип с Большого Хингана.) Спина песчано-се-
рая. Бока желтоватые. Хвост почти голый (у остальных форм он обычно покрыт
волосами). Амуро-Уссурийский край, Северо-Восточный Китай.
СЕМЕЙСТВО СЛЕПЫШОВЫЕ. FAMILIA SPALACIDAE
Своеобразные грызуны, ведущие постоянно подземный образ жиз-
ни. Тело вальковатое, шея очень короткая, голова широкая, уплощен-
ная сверху, лапы короткие, слабые с короткими небольшими когтями,
хвоста нет. Ушные раковины не развиты. Глаза скрыты под кожей. По
бокам головы от носа к ушам тянутся полоски жестких упругих ще-
тинок. Резцы торчат изо рта, так как губы сходятся за ними. Ко-
ренных зубов по три с каждой стороны как. верхней, так и нижней
челюсти.
344
Распространены в Юго-Восточной Европе, Передней Азии и Се-
веро-Восточной Африке.
Один род.
РОД СЛЕПЫШИ. GENUS SPALAX Guldenstaedt, 1770
Распространение то же, что у семейства.
В фауне СССР — 3 вида.
о ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ СЛЕПЫШЕЙ ФАУНЫ СССР
1(2). На затылочных костях над сочленовными мыщелками име-
ются маленькие и обычно глубокие ямки, как бы наколотые иглой;
обычно по одной ямке слева и справа, изредка только одна ямка
(с одной стороны); как исключение 3 ямки (одна с одной и две с дру-
гой стороны). Ширина основания носового отдела черепа не более
12 мм.................Малый слепыш. Spalax leucodon. (Стр. 345.)
2(1). На затылочных костях над сочленовными мыщелками ма-
леньких ямок нет. Ширина основания носового отдела черепа более
12 мм.............................................................3
3(4). Размеры средние: длина ступни (без когтей) менее 30 мм;
наибольшая длина черепа (от переднего края носовых костей до за-
тылочных мыщелков) не более 60 мм; кондилобазальная длина черепа
менее 58 мм; скуловая ширина не более 45 мм. Окраска меха сверху
относительно темная, охристо-бурая..................................
. . Обыкновенный слепыш. Spalax microphthalmus. (Стр. 346.)
4(3). Размеры крупнее: длина ступни (без когтей) у взрослых
особей более 30 мм; наибольшая длина черепа взрослых не менее
60 мм; кондилобазальная длина более 58 мм; скуловая ширина не
менее 45 мм. Окраска меха всегда, и даже у молодых, очень светлая,
серовато-охристая; у старых особей верх головы почти белый . . . .
................Гигантский слепыш. Spalax giganteus. (Стр. 346.)
119. Малый слепыш. Spalax leucodon Nordmann, 1840
Табл. XL, 10. Карта 110
Югославия, Венгрия, Румыния, Болгария, Греция, Турция.
В СССР — Молдавия и Южная Украина, к востоку до р. Буга, на се-
вер до Балты и Первомайска; кроме того, в Армении и Юго-Западной
Грузии.
В Закавказье обитает на пологих склонах гор, покрытых степной
и луговой растительностью. Отмечены их скопления в местах, изоби-
лующих луковичными растениями, составляющими важную часть их
рациона. В подземных ходах, кроме гнездовых камер, выстланных су-
хими растениями, имеются кладовые для пищевых запасов на зиму.
Кучи выброшенной на поверхность земли располагаются на расстоя-
нии от 0,25 до 7 ж одна от другой. Гнездовые камеры располагаются
под самыми большими кучами. Размножаются один раз в году и при-
носят не более двух детенышей.
В СССР — 2 хорошо различающихся подвида:
a) S. 1. leucodon Nordmann, 1840. (Тип из окрестностей г. Одессы.) Носовой отдел
черепа относительно широкий; длина черепа до 40 мм. Молдавия и Южная
Украина.
b) S. 1. nehringi Satunin. 1898. (Тип из окрестностей Казикопоран, в Армении.)
Носовой отдел черепа очень узкий; длина черепа 40—60 мм. Закавказье.
345
120. Обыкновенный слепыш. Spalax microphthalmus Giildenstaedt, 1770
Табл. XL, 11. Карта 110
Степи и лесостепь юга Европейской части СССР к северу до ли-
нии: Ровно—Киев—Чернигов—Тула—Пенза—Ульяновск, к востоку до
Волги, к югу до подножий Большого Кавказа и Перекопского пере-
шейка. На запад ареал идет в Румынию, Венгрию и Болгарию.
Селится по склонам балок, густо заросших травами, на выгонах,
толоках, опушках леса, реже на пахотных землях. Общая протяжен-
ность поверхностных ходов одного слепыша достигает 250 м. Гнездо-
вая камера располагается на глубине до 3,5 ж; кроме гнезда, в норе
имеется до 10 кладовых для запасов корма на зиму. Питается преиму-
щественно подземными частями растений, но втягивает в норы и зе-
леные (надземные) части (стебли и листья). Охотно поедает клубни
и луковицы. В году бывает, видимо, лишь один выводок из 1—3 дете-
нышей. Выбросами земли и поеданием растений, особенно клубней кар-
тофеля, причиняет существенный вред.
В пределах СССР различают 3 подвида:
a) S. m. microphthalmus Giildenstaedt, 1770. (Описан по экземпляру из окрестно-
стей Новохоперска, донские степи.) Лобно-носовой шов прямой или несколько выгнут
назад. От Волги до Днепра, за исключением нижнеприднепровских песков.
b) S. ш. zemni Erxleben, 1777. (Описан по экземпляру из Подолии.) Лобно-носо-
вой шов подковообразно выгнут вперед. Правобережная Украина, к северу от ареала
малого слепыша.
с) S. m. arenarius Reshetnik, 1938. (Тип из окрестностей Голой Пристани, Херсон-
ской области.) Отличается деталями конфигурации черепа и рисунком швов между
костями. Нижнеднепровские пески, от Каховки до Черного моря.
121. Гигантский слепыш. Spalax giganteus Nehring, 1898
Табл. XL, 12. Карта ПО
Полупустыни Северо-Восточного Предкавказья, между низовьями
Кумы и Терека, к югу до г. Махачкалы, к северу до южных районов
Астраханской области. Изолированные колонии имеются в полупусты-
нях к востоку от р. Урала (Кара-Агач, верховья и среднее течение
Эмбы).
Живет в глинистых и песчаных полупустынях, где, однако, предпо-
читает наиболее увлажненные участки (поймы рек, лесные насажде-
ния и пр.).
Гигантского слепыша некоторые считают лишь подвидом обыкно-
венного. Однако различия между ними (по окраске меха, размерам
тела и черепа) не меньше, чем между малым и обыкновенным слепы-
шами. Географически все три формы замещают друг друга. Раньше
малого слепыша по наличию отверстий на затылочной кости выделяли
даже в особый подрод (Microspalax). Но теперь выяснилось, что на
затылочной кости малого слепыша имеются не отверстия, а неглубокие
ямки диаметром со швейную иглу. Число ямок непостоянно (обычно 2,
но бывает 1 и 3). По одной ямке имеется и у отдельных экземпляров
гигантского слепыша. При такой неустойчивости этого признака разде-
ление слепышей на подроды оказалось совершенно несостоятельным.
А разделение их в качестве самостоятельных видов требует дальней-
шего уточнения. Если гигантского слепыша считать подвидом обыкно-
венного, то и малого слепыша тоже окажется необходимым свести на
положение подвида, так как различия между ними одинаково незначи-
тельны.
346
ЛИТЕРАТУРА
ПО МЛЕКОПИТАЮЩИМ ФАУНЫ СССР
В кратком списке приведены главным образом сводки и статьи, не отраженные
в библиографических списках сводок.
ОПРЕДЕЛИТЕЛИ И МОНОГРАФИИ ПО ВСЕМ ИЛИ НЕСКОЛЬКИМ
ОТРЯДАМ
И. И. Б а р а б а ш - Н и к и ф о р о в, А. Н. Формозов, Териология, изд.
«Высшая школа», М., 1963.
Н. А. Бобринский, Л. А. Зенкевич, Я. А. Б и р ш т е й н, География жи-
вотных, изд. «Советская наука», М., 1946.
Н. А. Бобринский, Б. А. Кузнецов и А. П. Кузякин, Определитель
млекопитающих СССР, изд. «Советская наука», М.. 1944.
И. АТ Громов, А. А. Гуреев, Г. А. Новиков, И. И. Соколов,
П. П. Стрелков, К. К. Ч а п с к и й, Млекопитающие фауны СССР, изд. АН СССР,
М.—Л., 1963.
В. И. Громова (ред.), Основы палеонтологии. Млекопитающие, изд. АН СССР,
М., 1962.
Д. Н. Данилов, Охотничье хозяйство СССР, М., 1963. Животный мир СССР,
тт. I—V, изд. АН СССР, М., 1936—1958.
А. М. Колосов, Н. П. Л а в р о в, С. П. Наумов, Биология промысловых
зверей СССР, изд. «Высшая школа», М., 1961.
Б. А. Кузнецов, Очерк зоогеографического районирования СССР, изд. МОИП,
М., 1950.
А. П. Кузякин, Зоогеография СССР, «Ученые записки МОПИ», т. 59. Биогео-
графия, вып. 1, М., 1962.
С. И. Огнев, Звери Восточной Европы и Северной Азии, т. I. Насекомоядные
н летучие мыши, '1928; т. II. Хищные млекопитающие, изд. АН СССР, М.— Л., 1931.
С. И. Огнев, Звери СССР и прилежащих стран, т. III. Хищные и ластоногие,
1935; т. IV—VII. Грызуны, 1940—1950; т. IX (А. Г. Томилин). Китообразные, изд.
АН СССР, М, 1957.
С. И. Огнев, Очерки экологии млекопитающих, изд. МОИП, М., 1951.
«Определители по фауне СССР», изд. ЗИН АН СССР, 48: Б. С. Виноградов
п И. М. Громов, Грызуны, М.— Л., 1952; 62: Г. А. Новиков, Хищные млекопи-
тающие, 1956; 79: А. Г. Томилин, Китообразные, 1962.
Эл лерман и Моррисон-Скотт, Ellerman J. R. and Morrison-Scott T.C.S.
Checklist of Palaearctic and Indian Mammals 1758 to 1946, London, 1951.
РЕГИОНАЛЬНЫЕ СВОДКИ И СТАТЬИ РСФСР
М. К. Виноградов, Морские млекопитающие Арктики, М.— Л., 1949.
М. Я. Марвин, Млекопитающие Карелии, Петрозаводск, 1959.
В. А. Савинов и А. Н. Лобанов, Звери Вологодской области, Во-
логда, 1958.
П. Белозеров, М. Торопова, В. Макридин, Растительность и живот-
ный мир Костромской области, Кострома, 1949.
А. Н. Формозов, Мелкие грызуны и насекомоядные Шарышского района
Костромской области в период 1930—194*0 гг. Материалы к познанию фауны и флоры
СССР. Новая серия II. Фауна и экология грызунов, вып. 3, М., 1948.
Т. С. Лукаш, А. И. Ш и р е н и н, П. В. П л е с с к и й, Животный мир Киров-
ской области. Сб. «Природа Кировской области», Киров, 1960.
Л. В. Шапошников, О. В. Головка, М. Г. Сорокин, А. Д. Тара-
канов, Животный мир Калининской области, Калинин, 1958.
347
Н. Кузнецов, Звери и птицы Ярославской области, Ярославль, 1947.
Т. Л. Граве, М. М. Емельянов, С. А. Плющепков, Е. А. Шмидт,
Животный мир Смоленской области, Смоленск, 1951.
Н. И. Пузанов, В. И. Козлов, Г. Л. Кипарисов, Животный мир
Горьковской области, г. Горький, 1955.
В. А. Попов, Млекопитающие Волжско-Камского края, Казань, 1960.
Г. А. Новиков, Экология зверей и птиц лесостепных дубрав, изд. АН СССР,
Л., 1959.
И. И. Барабаш-Никифоров, Звери юго-восточной части черноземного
центра, Воронеж, 1957.
А. К. Т е м б о т о в, Млекопитающие Кабардино-Балкарской АССР, Наль-
чик, 1960.
А. С. Строганова, Млекопитающие степного п полупустынного Заволжья,
«Труды Зоологического института АН СССР», т. XVI, М., 1948.
Я. Д а р к ш е в и ч, Птицы и звери Чкаловской области и охота на них, г. Чка-
лов, 1956.
С. В. Кирико в, Птицы п млекопитающие южной окраины Приуралья, «Труды
Института географии АН СССР», вып. 66. М., 1955.
С. С. Ш в а р ц, В. Н. Павлинин, Н. Н. Данилов, Животный мир Урала,
Свердловск, 1951.
В. Н. Павлинин, Очерк акклиматизации млекопитающих на Урале, «Труды
Института биологии УФ АН СССР», вып. 18, М., 1959.
С. В. Кирико в, Птицы и млекопитающие в условиям ландшафтов южной око-
нечности Урала, М., 1952.
И. П. Лаптев, Млекопитающие таежной зоны Западной Сибири. Томск, 1958.
Л. П. Никифоров, Опыт биосъемки населения млекопитающих Тоболо-Ишим-
ской лесостепи. География населения наземных животных и методы его изучения, изд.
АН СССР, М., 1959.
А. И. Я н у ш е в и ч, Фауна позвоночных Тувинской области, Новосибирск, 1952.
В. Е. Флинт, Материалы по фауне Тувы и перспективы ее изучения. Бюлле-
тень МОИП, отделение биологии, т. 64, вып. 2, М., '1959.
Э. В. Рогачева, Очерк фауны млекопитающих и птиц Енисейской тайги п
лесотундры. География населения наземных животных и методы его изучения, изд.
АН СССР, М, 1959.
А. С. Фетисов и В. П. X р у с ц е л е в с к и й, Млекопитающие Юго-Восточного
Забайкалья, «Труды Иркутского государственного университета», биологическая серия,
т. 111, вып. 8, Иркутск, 1948.
И. В. Измайлов, Зоологические исследования на Витимском плоскогорье
в 1959 году. Краеведческий сборник, вып. 5, Улан-Удэ, 1960.
В. И. Белык, Состав и распределение охотничье-промысловой фауны млекопи-
тающих Якутии. Сб. «Промысловая фауна и охотничье хозяйство Якутии», вып. 1,
Якутск, 1953.
Ю. В. Аверин, Наземные позвоночные Восточной Камчатки. «Труды Кропоц-
кого государственного заповедника», вып. 1, М., 1948.
Б. А. Кузнецов, Охотничье-промысловые звери Курильских островов. Сб.
«Пушные богатства СССР», М., 1949.
А. И. К у р е н ц о в, Животный мир Приморского края, Владивосток, 1951.
ПРИБАЛТИЙСКИЕ РЕСПУБЛИКИ И БЕЛОРУССИЯ
Ю. М. А у л ь, X. И. Лин г, К- Л. П а а в е р, Млекопитающие Эстонской ССР,
Таллин, 1957.
Б. А. Кузнецов, Материалы по фауне млекопитающих Литовской ССР.
«Бюллетень Московского отделения биологии», т. 59, вып. 4, М., 1954.
А. И. Калниньш, Охота п охотничье хозяйство в Латвийской ССР,
Рига, 1950.
И. Н. Сержанин, Млекопитающие Белорусской ССР, Минск, 1955.
УКРАИНА И МОЛДАВИЯ
О. П. Корнеев, Визначник 3BipiB УРСР, Ки!в, 1952.
В. 1. А б е л е и ц е в, 1. Г. П i д о п л i ч к о, Б. М. Попов, Фауна Украппк
т. 1, Ссавщ, вып. 1 (общий обзор млекопитающих, насекомоядные и рукокрылые),
Ки1в, 1956.
1. Т. Со кур, Ссавц! фауни Украши та i’x господарське значения, Ки!в, 1*960.
К. А. Татаринов, Зв1р1 Зах1дних областей Укра!ни, Кшв, 1956.
Б. А. Кузнецов, Фауна млекопитающих Молдавии, изд. Молдавского фили-
ала АН СССР, № 4—5, Кишинев, 1952.
348
КАВКАЗ И РЕСПУБЛИКИ ЗАКАВКАЗЬЯ
Н. К. Верещагин, Млекопитающие Кавказа, изд. АН СССР, М —Л., 1959.
С. К. Д а л ь, Животный мир Армянской ССР, т. I, Ереван, 1954.
Н. К. Верещагин, Класс млекопитающих. Сб. «Животный мир Азербайджана»,
Баку, 1951.
КАЗАХСТАН
А. В. Афанасьев, В. С. Бажанов, М. Н. К о р е л о в, А. А. С л у д с к и й,
Е. И. С т р а у т м а и, Звери Казахстана, Алма-Ата, 1953.
Б. А. Кузнецов, Млекопитающие Казахстана, изд. МОИП, М., 1948.
А. А. С л у д с к и й, Е. И. Страутман, Ю. В. Афанасьев, Пушные ре-
сурсы Казахстана и возможности развития охотничьего хозяйства в республике. «Труды
Института зоологии АН Казахской ССР», т. XVII, 1962.
А. Н. Формозов, Животный мир. Сб. «Казахстан», М.— Л., 1950.
А. Н. Формозов и В. И. Осмоловская, Численность некоторых ланд-
шафтных животных Центрального Казахстана. Материалы по биогеографии СССР,
вып. 54, «Труды Института географии АН Казахской ССР», М., 1953.
С. Н. Варшавский, М. Н. Шилов, Новые данные по распространению не-
которых млекопитающих, преимущественно грызунов, в Северном Приуралье и при-
лежащих районов, «Бюллетень МОИП», т. 60, вып. 5, 1955.
М. Н. Кор ел о в, Фауна позвоночных Бостандыкского района, «Труды АН Ка-
захской ССР», Алма-Ата, 1956.
РЕСПУБЛИКИ СРЕДНЕЙ АЗИИ
А. М. Андрушко, Млекопитающие высокогорной долины Алай (Памир), «Уче-
ные записки Ленинградского государственного университета», серия биологических
наук, вып. 38, Л., 1954.
В. Г. Г е п т н е р, Фауна позвоночных животных Бадхыза, «Труды Института
биологии АН Туркменской ССР», Ашхабад, 1956.
Г. П. Дементьев, К фауне наземных позвоночных Юго-Западной Туркме-
нии, «Ученые записки Московского государственного университета», вып. 83. Биология,
М., 1945.
Г. П. Дементьев, А. К. Рустамов, Е. П. Спангенберг, Материалы
по фауне наземных позвоночных Юго-Восточной Туркмении, «Труды Туркменского
сельскохозяйственного института», т. VII, Ашхабад, 1955.
Т. 3. 3 а х и д о в, Экологический обзор фауны позвоночных пустыни Кызылкум,
«Узбекский биологический журнал», № 2, Ташкент, 1958.
Г. И. И ш у н и н, Млекопитающие (хищные и копытные). Из серии «Фауна
Узбекской ССР», т. III, Ташкент, 1961.
В. П. Костин, Материалы по фауне млекопитающих левобережья низовьев
Аму-Дарьи и Устюрта и меры охраны их. Материалы по производительным силам
Узбекистана, вып. 2, Ташкент, 1956.
В. П. Костин, Некоторые данные о численности пушных зверей в Кара-
калпакии, «Труды Среднеазиатского университета». Зоология, изд. САГУ, Ташкент, 1959.
В. Т. Кривошеев, Эколого-географический очерк наземных позвоночных Се-
верных Кызылкумов, «Ученые записки МГПИ им. Ленина», т. 124, вып. 7, М., 1953.
В. И. Чернышев, Фауна и экология млекопитающих тугаев Таджикистана,
«Труды Института зоологии и паразитологии АН Таджикской ССР», т. 85, Душан-
бе, 1958.
В. И. Львова, Млекопитающие Гиссарской долины и прилежащих горных
районов, «Труды Таджикского филиала АН СССР». Зоология и паразитология, т. XIV,
Душанбе, 1945.
С. Ю. Мод а ми но в, Материалы по изучению фауны млекопитающих Север-
ного Таджикистана, «Ученые записки Ленинабадского педагогического института»,
вып. II, Ленинабад, 1953.
Р. Н. Мекленбурцев, Материалы по наземным позвоночным бассейна
р. Кашка-Дарья, Ташкент, 1958.
А. К. Рустамов и Е. С. П т у ш е н к о, Фаунистические материалы по на-
земным позвоночным Западных и Северо-Восточных Каракумов, Ашхабад, 1959.
X. С. Салихбаев, Позвоночные животные защитных лесных насаждений
Узбекистана, «Труды Института зоологии и паразитологии», вып. 8, Ташкент. 1956.
Б. К. Ф е н ю к и П. И. Камнев, Заметки о фауне млекопитающих Мангы-
шлака и Устюрта. Сб. «Грызуны и борьба с ними», вып. 5, 1957.
Б. А. Кузнецов, Звери Киргизии, изд. МОИП, М., 1948.
349
СВОДКИ И СТАТЬИ ПО ОТРЯДАМ И ВИДАМ
НАСЕКОМОЯДНЫЕ
Н. А. Бобринский, Б. А. Кузнецов, А. П. К у з я к и н, Определитель мле-
копитающих СССР. Насекомоядные, изд. «Советская наука», М., 1944.
А. А. Гуреев, Отряд насекомоядные, в кн. «Млекопитающие фауны СССР», под
ред. И. И. Соколова, изд. АН СССР, т. 1, М.— Л., 1963.
С. И. Огнев, Звери Восточной Европы и Северной Азии, т. I, Насекомоядные,
М., 1928.
С. У. Строганов, Звери Сибири. Насекомоядные, изд. АН СССР, М., 1957.
Д. И. А сп и сов, Выхухоль, М., 1952.
С. У. Строганов, Систематика кротовых (Talpidae), «Труды Зоологического
института АН СССР», том VIII, вып. 9, М.— Л., 1948.
Н. К. Д е п а р м а, Крот, изд. МОИП, М., 1951.
Н. К. Д е п а р м а, О малом кроте. Сообщение 2-е, «Бюллетень МОИП», т. 66,
вып. 6, 1961.
А. М. Колосов, Биология крота могеры. «Труды Московского пушно-мехового
института», т. 5, М., 1957.
К. Н. Благосклонов, О питании и характере суточной активности крошеч-
ной бурозубки, «Зоологический журнал», т. 36, вып. 3, 1957.
Б. С. Виноградов, О строении наружных гениталий у землероек-белозубок,
«Зоологический журнал», т. 37, вып. 8, 1958.
Е. В. Карасева и А. И. Ильенко, К изучению биологии и географиче-
ского распространения землероек в Северном Казахстане. Материалы по фауне и
экологии наземных позвоночных Казахстана АН Казахской ССР, т. XIII, «Труды Ин-
ститута зоологии», Алма-Ата, 1960.
В. И. Чернышев, К экологии бухарской бурозубки, «Известия Отделения
естественных наук АН Таджикской ССР», вып. 14, 1956.
С. С. III в а р ц, Биология землероек лесостепного Зауралья, «Зоологический жур-
нал», т. 34, вып. 4, 1956.
Б. С. Юдин, Экология бурозубок (род Sorex) Западной Сибири, «Труды
Института биологии Сибирского отделения АН СССР», вып. 8, Новосибирск, 1962.
РУКОКРЫЛЫЕ
А. П. Кузякин, Летучие мыши, изд. «Советская наука», М., 1950.
П. П. Стрелков, Отряд рукокрылые, в кн. «Млекопитающие фауны СССР»,
под ред. И. И. Соколова, изд. АН СССР, т. 1, М.— Л., 1963.
В. И. Абеленцев, О летучих мышах Закарпатской и других западных обла-
стей УССР, «Труды зоомузея Киевского университета», № 11, Киев, 1950.
О. П. Богданов, Рукокрылые. Из серии «Фауна Узбекской ССР», т. 3, вып. 2,
Ташкент, 1953.
О. П. Богданов, Летучие мыши Таджикистана, «Доклады АН Таджикской
ССР», вып. 15, 1956.
О. П. Богданов, Новые данные по распространению, местам обитания и
зимовкам рукокрылых в Узбекистане, «Доклады АН УзССР», № 3, Ташкент, 1956.
О. П. Богданов, К распространению и экологии длиннохвостой ночницы,
«Зоологический журнал», т. XXXIX, № 12, 1960.
Н. Н. Воронцов, Заметки о рукокрылых Восточного Казахстана, «Бюлле-
тень МОИП», отделение биологии, т. 63, вып., 1959.
М. Н. Корелов, Заметки по систематике летучих мышей Казахстана, «Вест-
ник АН КазССР», серия зоологии, вып. 8, Алма-Ата, 1948.
А. Н. Ку рек оу, Да распаусюджания i экалоги рукакрылых фауни Беларуская
ССР, «Висци АН БССР», № 2, Минск, 1960.
X. И. Ли и г, Материалы по фауне летучих мышей Эстонской ССР, Тал-
лин, 1953.
Г. Н. Лихачев, Использование летучими мышами птичьих искусственных гнез-
довий. «Труды Приокско-Террасного государственного заповедника», вып. 3, М., 1961.
Л. К. П о о т с, О зимовке летучих мышей в Эстонии, «Природа», № 10, 1953.
П. П. Стрелков, Материалы по зимовкам летучих мышей в Европейской
части СССР, «Труды Зоологического института АН СССР», т. XXV, 1958.
А. А. Т а г и л ь ц е в, К фауне летучих мышей Южной Киргизии, «Труды Инсти-
тута зоологии и паразитологии», Киргизский филиал АН СССР, вып. 2, Фрунзе, 1954.
ХИЩНЫЕ
Г. А. Новиков, Хищные млекопитающие фауны СССР, изд. АН СССР, М.— Л.,
1956; Отряд хищные, в кн. «Млекопитающие фауны СССР», под ред. И. И. Соколова,
изд. АН СССР, т. I, М.—Л., 1963.
350
А. А. С о к о л о в, Волк, M., 1957.
Ю. А. Герасимов, Лисица, М., 1950.
С. В. Шибанов, Песец, М., 1951.
В. Ф. Морозов, Уссурийский енот, М., 1951.
А. П. Корнеев, Енотовидная собака на Украине, 1954, стр. 13—72.
Ю. К. Попов, Материалы пб экологии енотовидной собаки в Татарской АССР,
«Биологический журнал», изд. Казанского филиала. АН СССР, № 5, 1956.
Н. Н. Руковский, Современный и возможный ареал епота-полоскуна (Рго-
cyon lotor) -в СССР. Проблемы зоогеографии суши, изд. Львовского государственного
университета, 1958.
А. Г. Панкратов, Акклиматизация енота-полоскуна в лесах Южного При-
морья, «Сообщения Дальневосточного филиала Сибирского отделения АН СССР»,
вып. II, 115, 1959.
Г. Ф. Бромлей, Гималайский медведь, «Зоологический журнал», т. XXXV,
вып. I, 1956.
Г. П. Дементьев, А. К. Рустамов, Заметки о некоторых видах кошачьих
Туркмении, «Известия АН Туркменской ССР», 1956.
Ех В. Ротшильд, Барханный кот, «Природа», № 9, 1956.
Н. А. П о л у ш и н а, Изменение распространения некоторых представителей се-
мейства куньих на западе Украины. Сб. «Проблемы зоогеографии суши», изд.. Львов-
ского государственного университета, 1958.
Г. Ф. Бромлей, Сборник материалов по результатам изучения млекопитаю-
щих в государственных заповедниках. Материалы по экологии соболя и харзы, рас-
пространённых в Приморском крае, М., 1956.
П. Б. Ю р г е н с о н, Очерки по сравнительному изучению соболя и куниц. К исто-
рии послеледниковых ареалов распространения соболя и лесной куницы в Европейской
части СССР, изд. Министерства сельского хозяйства, 1956.
В. Н. Над ее в, Соболь, М., 1955.
В. А. Тавров ск ий, Соболь Северо-Западной Якутии и пути восстановления
его промысла. Якутский филиал Сибирского отделения АН СССР, «Труды Института
биологии», вып. 4, 1958.
Ю. Г. Афанасьев, Темпы восстановления ареала алтайского соболя, «Труды
Института зоологии АН Казахской ССР», Алма-Ата, 1960.
Д. В. Т е р н о в с к и й, Биология и акклиматизация американской норки на
Алтае, 1958, стр. 32—40.
В. А. Попов, Материалы по экологии норки (Mustela vison) и результаты ак-
климатизации ее в Татарской АССР. «Труды Казанского филиала АН СССР», Ка-
зань, 1949.
«Калан». Сборник, М., 1947.
ЛАСТОНОГИЕ
К. К. Ч а п с к и й, Отряд ластоногие, в кн. «Млекопитающие фауны СССР», под
ред. И. И. Соколова, Отряд ластоногие, изд. АН СССР, ч. 2, М., 1963.
Н. А. Бобринский и др., Определитель млекопитающих СССР, Отряд ласто-
ногие, изд. «Советская наука», М., 1944.
С. П. Наумов, Тюлени СССР, М., 1933.
Н. А. Смирнов, Определитель ластоногих Европы и Средней Азии, «Изве-
стия Отделения промысловой ихтиологии», т. IX, вып. 3, Л., 1929.
КИТООБРАЗНЫЕ
А. Г. Томилин, Звери СССР и прилежащих стран, т. IX, Китообразные,
М., 1957.
А. Г. Томили н, Определитель китообразных по поведению и внешним при-
знакам, изд. АН СССР, М., 1951.
А. Г. Томилин, Китообразные фауны морей СССР, изд. АН СССР, М., 1962;
Отряд китообразные, в кн. «Млекопитающие фауны СССР», под ред. И. И. Соко-
лова, ч. 2, 1963.
Б. А. 3 е н к о в и ч, Киты и китобойный промысел, М., 1952.
С. Е. Клейненберг, Млекопитающие Черного и Азовского морей, М., 1956.
М. Н. Слепцов, Китообразные дальневосточных морей, Владивосток, 1955.
А. А. К и р п и ч н и к о в, О современном распространении кашалота в Мировом
океане по промысловым данным, «Бюллетень МОИП», т. 55, вып. 5,.М., 1950.
КОПЫТНЫЕ
И. И. Соколов, Копытные звери. «Фауна СССР. Млекопитающие», т. I, вып. 3,
изд. АН СССР, М.—Л., 1959.
И. И. Соколов, Отряд непарнопалые и отряд парнопалые, в кн. «Млекопи-
тающие фауны СССР», под ред. И. И. Соколова, изд. АН СССР, т. 2, М —Л., 1963.
351
A. A. H а си м о ви ч, А. Г. Банников, Млекопитающие Советского Союза,
т. I. Парнокопытные и непарнокопытные, изд. АН СССР, М., 1961.
В. В. Дмитриев, Копытные звери Алтайского заповедника и прилежащих
мест, «Труды Алтайского государственного заповедника», вып. I, М., 1938.
Т. В. Саблина, Копытные Беловежской пущи, «Труды института морфологии
животных им. А. Н. Северцова», изд. АН СССР, вып: 15.
А. А. Слудский, Кабан, Алма-Ата, 1956.
К. К. Флеров, Кабаны и олени. Из серии «Фауна СССР», т. I, вып. 2,
изд. АН СССР, М.—Л., 1952.
В. Г. Г е п т н е р, В. И. Ц а л к и н. Олени СССР, изд. МОИП, М., 1947.
Г. Ф. Б р о м л е й, Экология дикого пятнистого оленя в Приморском крае.
«Сборник материалов по результатам изучения млекопитающих в государственных
заповедниках», М., 1956.
Г. Ф. Бромлей, Горал, «Зоологический журнал», т. 35, вып. 9, М., 1956.
И. В. Друри, Дикий северный олень Советской Арктики и Субарктики, «Труды
Арктического института», т. 200, Л., 1949.
И. И. Соколов, Опыт естественной классификации полорогих, «Труды Зооло-
гического института АН СССР», М.— Л., 1953.
А. Г. Банников, Л. В. Жирнов, Л. С. Л е б е д е в а, А. А. Фа идее в,
Биология сайгака, М., 1961.
А. А. Наси мо вич, Новые данные по биологии серны из Западного Кавказа,
«Труды Кавказского государственного заповедника», вып. 3, М., 1949.
В. И. Ц а л к и н, Винторогий козел СССР, «Ученые записки Московского госу-
дарственного университета», вып. 83, М., 1945.
Г. С. Султанов, Винторогий козел в Узбекистане, «Труды института зооло-
гии и паразитологии», т. 2, «Зоологический сборник», 1953.
В. И. Ц а л к и н, Сибирский горный козел. Материалы к познанию фауны и
флоры СССР. Новая серия, отдел зоологии, вып. 21 (36), М., 1950.
О. В. Егоров, Экология сибирского горного козла (Capra sibirica), «Труды
Зоологического института АН СССР», т. XVIII, М.— Л., 1955.
В. И. Ц а л к и н, Изменчивость и систематика туров Западного Кавказа, «Бюл-
летень МОИП», отдел биологии, т. 60, вып. 4, М., 1955.
А. А. Н а с и м о в и ч, Очерк экологии западнокавказского тура, «Труды Кавказ-
ского государственного заповедника», вып. 3, М., 1949.
В. И. Цалкин, Горные бараны Европы и Азии. Материалы к познанию фауны
и флоры СССР. Новая серия, отдел зоологии, вып. 27 (42), М., 1951.
1. И. И ш у н и н, Баран Северцова — реликт Центральных Кызылкумов, «Бюл-
летень Окрсовета Всеармейского военно-охотничьего общества Туркестанского ВО»,
№ 1, 1960.
Р. Н. Мекленбурцев, Памирский архар, «Бюллетень МОИП», т 53, вып. 5,
М, 1948.
Ю. В. Аверин, О снежном баране на Камчатке, «Бюллетень МОИП», отдел
биологии, т. 56 (1), М., 1951.
Л. Н. Корочкина, Беловежский зубр, «Труды заповедника «Беловежская
пуща», вып. 1, Минск, 1958.
ГРЫЗУНЫ
Б. С. Виноградов, И. М. Громов, Грызуны фауны СССР, изд. АН СССР,
М.—Л, 1952.
Б. С. Виноградов, И. М. Громов, Краткий определитель грызунов фауны
СССР, изд. АН СССР, М.—Л., 1956.
И. М. Громов и др., Млекопитающие фауны СССР, под ред. И. И. Соколова.
Отряд грызуны, изд. АН СССР, М.— Л., 1963.
«Грызуны и борьба с ними», вып. 1, Алма-Ата, 1941; вып. 2, Саратов, 1946; вып. 3,
1950; вып. 4, 1954; вып. 5, 1957; вып. 6, Саратов, 1959.
Н. А. Бобринский и др., Определитель млекопитающих СССР. Отряд гры-
зуны, изд. «Советская наука», М., 1944.
А. М. Колосов, Грызуны — вредители сельского хозяйства, М., 1960.
А. П. Кузякин, К системе грызунов фауны СССР «Труды МОИП», т. X,
М., 1963.
«Материалы по грызунам» (под ред. А. Н. Формозова), изд. МОИП, вып. 1, 1946;
вып. 2, 1947; вып. 3, 1948; вып. 4, 1951; вып. 5, 1957.
Ю. М. Р а л л ь, Методика полевого изучения грызунов и борьбы с ними, изд.
Ростокинского книжного издательства, Ростов-на-Дону, 1947.
Н. В. Б а ш е н и н а, В. В. Груздев, Н. М. Д у к е л ь с к а я, И. А. Шилов,
Грызуны — вредители садов и огородов, М., 1954.
352
Т. В. Кошкина, Мышевидные грызуны Кольского полуострова и динамика
их численности, «Труды Кандалакшского государственного заповедника», вып. 1, Во-
логда, 1958.
В. В. Т у р ь е в а, Мышевидные грызуны лесной части Коми АССР, «Труды
Комифилиала АН СССР», № 1, Сыктывкар, 1953.
А. Г. Воронов, Материалы по экологии мелких грызунов Пермской области,.
«Ученые записки», изд. Пермского государственного педагогического института, вып.
11, 1948.
В. В. Кучерук, М. А. Рубина, Причины, определяющие видовой состав и
численность грызунов в скирдах, ометах и стогах южных районов Московской обла-
сти, «Зоологический журнал», т. 32, вып. 3, М., 1953.
И. Л. Кулик, Грызуны скирд и ометов. Отдел паразитологии и медицинской
зоологии Института эпидемиологии и микробиологии им. почетного акад. Н. Ф. Гама-
лея, изд. АМН СССР, М., 1951.
П. К. Смирнов, Фауна грызунов лесных полезащитных полос Курской обла-
сти, «Ученые записки ЛГУ им. Жданова». Зоология позвоночных, серия биологиче-
ских наук, вып. 38, № 181, 1955.
М. Я. Лаврова, Грызуны лесных полезащитных полос Ставропольских и Саль-
ских степей, «Труды Института географии АН СССР», вып. 66, 1955.
А. А. Л а в р о в с к и й,. Н. П. Миронов, Видовой состав, распространение и
численность грызунов Ергеней в местах закладки лесополос, «Зоологический журнал»,,
т. 20, вып. 1, М., 1951.
В. С. Елпатьевский, Н. И. Ларина, В. Л. Голикова, Видовой со-
став и экологическое размещение лесных и степных грызунов на трассе государствен-
ной лесной полосы Саратов — Камышин, «Ученые записки Саратовского универси-
тета», т. XXIX, вып. биолого-почвенного отделения, 1952.
Н. В. Щ е п о т ь е в, Мышевидные грызуны железнодорожных лесных полос Ниж-
него Поволжья, Сб. «Грызуны и борьба с ними», вып. 5, Саратов, 1957.
И. М. Громов и Н. М. Парфенова, Материалы по фауне грызунов Ин-
дерского Приуралья и ее истории, «Бюллетень МОИП», отдел биологии, т. 56, вып. 4,
М, 1951.
В. Н. Павлинин» С. С. Шварц, Мышевидные грызуны Урала, Сверд-
ловск, 1953.
С. У. Строганов, А. Ф. Потапкина, К характеристике фауны грызунов-
Томской области, «Ученые записки Томского государственного университета им. Куй-
бышева», № 14, 1950.
В. Н. Скалой, Н. Н. Скалой, К фауне Тувинской автономной области, «Из-'
вестия Иркутского государственного научно-исследовательского противочумного инсти-
тута Сибири и Дальнего Востока», т. VII, Иркутск, 1949.
Н. И. Фирстов, Грызуны района Братской ГЭС, «Известия Иркутского госу-
дарственного научно-исследовательского противочумного ин-та Сибири и Дальнего-
Востока», т. XXI11, Иркутск, 1960.
Е. П. Демин, Грызуны хребта Сайлюгем и южной части хребта Чихачева,
«Известия Иркутского государственного научно-исследовательского противочумного
института Сибири и Дальнего Востока», т. XXI11, Иркутск, 1960.
А. С. Фетисов, Грызуны Южного Забайкалья, «Известия Иркутского госу-
дарственного научно-исследовательского противочумного института Сибири и Дальнего
Востока», т. V, Иркутск, 1944.
П. Д. Ларионов, Мелкие грызуны Мегино-Кандаласского района и их ста-
циальное распределение, «Ученые записки Якутского педагогического института»,
вып. 3, 1954..
О. А. Козловская, И. Н. Аникеев, Грызуны и блохи Еврейской авто-
номной области, «Известия Иркутского государственного научно-исследовательского
противочумного института Сибири и Дальнего Востока», т. XXI11, Иркутск, 1960.
Н. С. Беляева, Мышевидные грызуны Амурской области, «Записки Амурского
областного музея краеведения и общества краеведения», Благовещенск, 1955.
Е. Н. Степин, Грызуны низовий Амура, «Известия Иркутского государствен-
ного научно-исследовательского противочумного института Сибири и Дальнего Востока»,
т. X, Иркутск, 1952.
Б. С. Виноградов, Краткий обзор грызунов южных областей Дальнего Во-
стока. Сб. «Паразитология Дальнего Востока», М., 1947.
И. И. Барабаш-Никифоров, О мышевидных грызунах Командорских
островов, «Зоологический журнал», изд. АН СССР, т. XXII, вып. 1, 1943.
А. А. Тимофеева, Некоторые наблюдения по стадиальному распределению
грызунов на юге о-ва Сахалина, «Известия Иркутского государственного научно-ис-
следовательского противочумного института Сибири и Дальнего Востока», т. 24, Ир-
кутск, 1962.
О. Н. Михалап, Грызуны Палесской низины.
23 н. А. Бобринский и др.
353
И. И. К а л юш ев, Краткий очерк фауны грызунов Закарпатской области, «Науч-
ные записки Ужгородского университета», т. VIII, Ужгород, 1953.
А. А. Лавровский, И. М. Колесников, Материалы к познанию грызунов
Дагестанской АССР, «Труды научно-исследовательского института Кавказа и Закав-
казья», вып. I, 1953.
М. В. Шидловский, Родентофауна Черноморского побережья Грузии.
(Эколого-фаунистический очерк.) «Труды Института зоологии АН Грузинской ССР»,
т. VII, 1947.
М. В. Шидловский, Грызуны Абхазии, «Труды Института зоологии АН
ГССР», т. IX, 1950.
М. В. Шидловский, Грызуны Аджарии, «Труды Института зоологии
АН ГССР», т. VIII, 1948.
А. Б. Алексанян, Материалы по изучению грызунов в Армянской ССР.
Сборник научных трудов Ереванского медицинского института, т. I, Ереван, 1940.
Е. И. Страутман, Влияние засухи на распределение и биологию некоторых
видов грызунов в Северном Казахстане, «Труды Института зоологии АН Казахстан-
ской ССР», т. VI, 1956.
Л. Б. Б е м е, К познанию грызунов Казахской складчатой страны, «Бюллетень
МОИП», отдел биологии, т. 55, вып. 2, М., 1952.
В. Е. Ф л и н т, К вопросу о структуре ареала и типах поселений некоторых
мышевидных грызунов в Северном Казахстане, «Бюллетень МОИП», отдел биоло-
гии, 1958.
В. Н. П а к и ж, К фауне грызунов Павлодарской области, «Труды Института
зоологии АН Казахской ССР», т. XIII, Алма-Ата, I960.
А. М. Андрушко, Грызуны восточной части Карагандинской области, «Уче-
ные записки ЛГУ», № 141, 1952.
А. В. Афанасьев, Грызуны Прибалхашья, «Известия Казахского филиала
АН СССР», серия биологии, вып. 5, Алма-Ата, 1945.
Н. В. Касаткина, Материалы по распространению грызунов в предгорьях
Заилийского Алатау, «Труды Государственного среднеазиатского противочумного
НИИ», вып. 4, 1958.
И. И. Колесников, Семейство беличьих, «Фауна Узбекской ССР», т. 3, вып. 5,
Ташкент, 1953.
В. П. П р и в а л о в, О стационарном распределении грызунов Центральных
Кызылкумов, «Труды Государственного среднеазиатского противочумного НИИ», вып. 3,
Алма-Ата, 1956.
В. П. Привалов, К вопросу о стадиальном размещении грызунов Ферганской
долины, «Труды Государственного среднеазиатского противочумного НИИ», вып. 3,
Алма-Ата, 1956.
В. П. Костин, Материалы по экологии грызунов южных склонов Каржан-
тау, «Труды Института зоологии и паразитологии», т. 2, изд. АН СССР, 1953.
В. А. Стальмакова, Пустыни СССР. Сб. «Грызуны Каракумов, их экология
и хозяйственное значение», вып. 2, М., 1956.
Б. К. Фен юк, П. И. Камнев, Г. Н. Скворцов, И. М. Гурьева, Эко-
логический очерк фауны грызунов западной части низменных Каракумов, «Грызуны
и борьба с ними», вып. 5, Саратов, 1957.
П. И. Камнев, Г. Н. Скворцов, И. М. Гурьева, Заметки по экологии
некоторых видов грызунов западной части Центральных Каракумов, «Грызуны и
борьба с ними», вып. 6, Саратов, 1959.
Л. Ф. Васильева, Л. А. Гувва, А. Н. Парамонова, Грызуны и их
эктопаразиты горного хребта Большого Балхана, «Тезисы докладов научной конфе-
ренции по природной очаговости», Саратов, 1957.
Н. В. Щепоть ев, Мышевидные грызуны населенных пунктов Нижнего По-
волжья, «Грызуны и борьба с ними», вып. 5, Саратов, 1957.
Е. П. Бондарь, Грызуны населенных пунктов Туркменистана, «Известия Турк-
менского филиала АН СССР», № I, 1946.
Н. В. Некипелов, Домовые грызуны в Юго-Восточном Забайкалье, «Изве-
стия Иркутского государственного научно-исследовательского противочумного инсти-
тута Сибири и Дальнего Востока», т. XIX (72—82), Иркутск, 1958.
В. В. Ш к и л е в, Материалы по динамике численности грызунов населенных
пунктов Приморского края, «Известия Иркутского государственного научно-исследо-
вательского противочумного института Сибири и Дальнего Востока», т. XII, Ир-
кутск, 1954.
'Р. П. Зимина, Грызуны — обитатели жилых построек в высокогорном Тянь-
Шане, «Зоологический журнал», изд. АН СССР, т. XXXI, вып. 1, 1952.
Б. С. Виноградов, А. И. Иванов, Грызуны Таджикистана, Душанбе, 1945.
А. Токтосунов, Грызуны Киргизии, Фрунзе, 1958.
Б. М. А й з и н, Определитель грызунов Киргизской ССР, изд. Киргизской респуб-
ликанской противочумной станции, Фрунзе, 1947.
354
сводки И СТАТЬИ ПО ГРУППАМ И ВИДАМ ГРЫЗУНОВ
Н. В. Некипелов, Изменение численности даурской пищухи в Юго-Восточ-
ном Забайкалье, «Известия Иркутского государственного противочумного института
Сибири и Дальнего Востока», т. XII (III—180). Данные по годовым изменениям сред-
него количества пар за 1941—1950 гг.
П. П. Т а р а с о в, К экологии монгольской пищухи, «Бюллетень МОИП», т. 55,
вып. 6, М., 1950.
Б. А. Ларин, Зайцы, М., 1956.
Т. Д. Д у л ь к е й т, Маньчжурский заяц в Уссурийском крае, «Зоологический
журнал», т. XXXV, вып. 6, 1956.
С. П. Наумов, Экология зайца-беляка, изд. МОИП, М., 1947.
С. П. Наумов, Общие особенности динамики численности зайца-беляка
в Якутии, «Ученые записки Московского государственного педагогического ин-та
им. В. И. Ленина», т. XCVI, вып. 6, М., 1956.
М. Н. Ко ре л о в, К экологии зайца-песчаника, «Вестник АН Казахской ССР»,
№ 5, Алма-Ата, 1947.
А. М. Колосов и Н. И. Бакеев, Биология зайца-русака, изд. МОИП,
М., 1947.
И. Н. Сержанин, Речной бобр и его биологические особенности, Минск, 1951.
И. М. Фомичева, Восстановление ареала речного бобра в СССР. В сб.'«Проб-
лемы зоогеографии суши», изд. Львовского государственного университета, 1958.
С. Н. Марин, Колония канадских бобров в СССР, «Зоологический журнал»,
т. XXV, вып. 7, 1956.
М. П. Павлов, Нутрия, М.. 1951.
Сурки. Экология, эктопаразиты, природная очаговость чумы, Алма-Ата,
Фрунзе, 1961.
А. А. Лавровский и Я. Ф. Шатас, Горноазиатский сурок в Дагестане,
«Зоологический журнал», т. XXXV, вып. 8, 1956.
В. И. Капитонов, Очерк биологии черношапочного сурка, «Зоологический
журнал», т. XXXIX, вып. 3, I960.
П. А. Ян ушко, Заметки по биологии сурка Мензбира, «Зоологический жур-
нал», т. XXX, вып. 6, 1951.
Е. А. Поляков, Суслики и меры борьбы с ними, М., 1956.
М. И. Исмагилов, Суслик-песчаник и способы его добывания, Алма-Ата, 1955.
М. И. Исмагилов, Половой и возрастной состав популяции и географиче-
ская изменчивость среднего суслика, «Зоологический журнал», т. XXXVIII, вып. 7,
изд. АН СССР, 1959.
А. П. Кузякин, Д. С. Резинко и Н. И. Макаров, Приманочный ме-
тод борьбы с малым сусликом и другими грызунами, вредными в сельском хозяйстве.
Сб. «Грызуны и борьба с ними», вып. 4, Саратов, 1955.
В. И. П а к и ж, Распространение европейского суслика в Молдавской ССР и
некоторые данные по его экологии, «Зоологический журнал», т. XXXVIII, вкп. I, 1958.
О. Р. Аветисян, Малоазиатский суслик и химические меры борьбы с ним
в Армянской ССР, 1951.
Ю. М. Р а л л ь, Очерк экологии иссык-кульского реликтового суслика, «Зооло-
гический журнал АН СССР», т. XXIV, вып. 6, М., 1945.
Д. И. Бибиков, К экологии .даурского суслика (сообщение 2-е), «Известия
Иркутского государственного противочумного института Сибири и Дальнего Во-
стока», т. X (3—21), Иркутск, 1952.
Н. С. Беляева, К биологии Амурского длиннохвостого суслика, «Записки
Амурского областного музея краеведения и общества краеведения», т. 3, Благове-
щенск, 1955.
В. Н. Скалой, Краткий обзор распространения и образа жизни длиннохво-
стого суслика в Якутии, «Бюллетень МОИП», отдел биологии, новая серия, вып.
4—5, М., 1946.
В. Г. Г е п т н е р, Внутривидовая изменчивость и распространение тонкопалого
суслика, «Зоологический журнал», т. XXXVIII, вып. 5, 1954.
С. А. Ларин, Белка, М., 1953.
В. С/ С ми рнов, Притобольская белка-телеутка и возможности ее акклимати-
зации в лесах Урала, изд. Уральского филиала АН СССР, «Труды Института био-
логии», вып. 18, 1959.
Г. Ф. Зворыкин, Биология некоторых видов сонь, «Природа», № 7, 1952.
М. Н. Л о з а н, Материалы по экологии сонь Молдавской ССР, изд. Молдав-
ского филиала АН СССР, № 8, 1959.
С. С. Д о н а у р о в, В. К. Попов, 3. П. К о н я к и н а, Соня-полчок в районе
Кавказского государственного заповедника, «Труды Кавказского государственного за-
поведника», вып. I, 1938.
А. Н. Данилович, К экологии лесной сони на Украине, «Природа», № 8, 1950.
23* 355
А. С. Бу рдел ов и О. Б. Российская. Об ареале селевинии (баялычной
сони) и некоторых особенностях ее экологии, «Зоологический журнал», т. XXXVIII,
вып. 6, 1959.
Б. С. Виноградов, Тушканчики, «Фауна СССР», т. III, вып. 4, изд. АН СССР,
М.—Л., 1937.
М. В. Андриевский и Л. Н. Елисеев, Материалы по экологии туш-
канчиков в районе государственной полезащитной лесной полосы Камышин — Волго-
град, «Бюллетень МОИП», т. 6, отдел биологии, изд. МГУ, 1962.
Ю. М. Ралль и Г. П. К а р а е р о в а, Новые данные о распространении и
вредной деятельности тушканчиков в Ростовской области, «Зоологический журнал»,
т. XXVIII, вып. 5, 1954.
А. Э. Аирапетьянц, И. М. Фокин, Тушканчик Бобринского в Каракумах,
«Природа», № 8, 1958.
А. М. К а р п у ш е в, Новые места нахождения тушканчика Бобринского. «Гры-
зуны и борьба с ними», вып. 6, Саратов, 1959.
Г. А. К о н д р а ш к и н, В. С. Е д ы к и н а, Очерк экологии земляного зайчика
дельты Волги. «Грызуны и борьба с ними», вып. 5, Саратов, 1957.
Н. Н. Воронцов, Новый вид толстохвостого тушканчика из Зайсайской кот-
ловины и заметки о тушканчиках рода Jaculus, «Зоологический журнал», т. XXXVII,
вып. I, 1958.
Н. Я. Мокроусов, Периодика жизнедеятельности и размножения емуранчика
в северо-западном Прикаспии. «Грызуны и борьба с ними», вып. 5, Саратов, 1957.
Б. С. Виноградов и Е. П. Бондарь, О нахождении в Туркмении нового
вида тушканчика из рода Jaculus, «Доклады АН СССР», новая серия, т. XV,
№ 4, 1949.
В. А. Стальмакова, О нахождении туркменского тушканчика (Jaculus
turkmenicus) в северных Каракумах и о некоторых его эколого-морфологических осо-
бенностях, «Зоологический журнал», т. XXXVI, вып. 2, 1957.
М. В. Елизарьева, Нахождение трубкоухого тушканчика в пределах СССР,
«Доклады АН СССР», т. XVI, № 3, 1949.
К. П. Пар а ск и в, Новые данные о карликовом тушканчике, «Труды института
зоологии АН Казахской ССР», т. XIII, I960.
Д. И. Берман, Карликовый пятипалый тушканчик — новый вид в фауне
СССР, «Бюллетень МОИП», отдел биологии, т. 67, вып. 5, 1962.
К. П. Благое к лоно в, К биологии лесной мышовки, «Бюллетень МОИП»,
отдел биологии, т. III, вып. 3, 1948.
С. К. Даль, Ю. Д. Чугунов, Лесная мышовка в Ставропольском крае
(эколого-географический очерк), «Труды Государственного научно-исследовательского
противочумного института Кавказа и Закавказья», вып. I, Ставропольское книжное
издательство, 1956.
В. Е. Флинт, Материалы по биологии степной мышовки, «Зоологический жур-
нал», т. XXXIX, вып. 6, I960.
Б. В. Образцов, Основные черты деятельности мышевидных грызунов в на-
горной дубраве, «Труды института леса», изд. АН СССР, т. VII, 1951.
А. И. Аргиропуло, Мыши. Из серии «Фауна СССР», т. III, вып. 5, изд. АН
СССР, М.—Л., 1940.
М. М. С л е п о ц о в, К биологии уссурийской мыши-малютки, «Фауна и эко-
логия грызунов». Материалы по грызунам, вып. 2, изд. МОИП, М., 1947.
Н. В. Тупикова, Экология домовой мыши, Материалы по грызунам, изд.
МОИП, вып. 2, М., 1947.
И. С. Солдаткин, Г. А. Асе но в, Ю. В. Р у д е н ч и к, Численность, раз-
множение и смертность домовых мышей оазиса низовьев Аму-Дарьи, «Грызуны и
борьба с ними», вып. 6, Саратов, 1959.
П. А. Свириденко, Экологические факторы, определяющие географическое
распространение и эйритопность полевой мыши, «Зоологический журнал», т. XXII,
вып. 5, М., 1943.
В. В. Ш к и л е в, Особенности изменений численности полевой мыши в Примор-
ском крае (Дальний Восток), изд. Иркутского государственного научно-исследова-
тельского противочумного института Сибири и Дальнего Востока, «Тезисы докладов
конференции», вып. 3, Улан-Удэ, 1958.
И. В. Жарков, Экология и значение лесных мышей в лесах Кавказского за-
поведника, «Труды Кавказского заповедника», вып. I, 1938.
П. А. Свириденко, Размножение и колебания численности желтогорлой
мыши, «Труды института зоологии», т. VI, М., 1949.
Н. И. Литвинов и Н. Г. Карнаухова, О видовом составе крыс, насе-
ляющих морские суда порта Владивостока, «Известия Иркутского государственного
научно-исследовательского института Сибири и Дальнего Востока», т. XIII, Ир-
кутск, 1954.
356
Л. Л. Гиренко, К вопросу о распространении черной крысы в УССР,
М., 1954.
В. П. Никитин, К распространению черной крысы на Дальнем Востоке, «Из-
вестия Иркутского государственного научно-исследовательского противочумного ин-
ститута Сибири и Дальнего Востока», т. X, Иркутск, 1952.
К. Бейшебаев, Туркестанская крыса, «Труды Института зоологии и парази-
тологии АН Киргизской ССР», вып. VI, 1957.
3. Козлова, К биологии туркестанской крысы. Сб. студенческих работ Сред-
неазиатского государственного университета им. В. И. Ленина, вып. XI, «Биоло-
гия», 1955.
А. П. Кузякин, К методике учета и вылова серых крыс в городских объек-
тах, «Грызуны и борьба с ними», вып. 3, Саратов, 1950.
А. П. Кузякин, История расселения, современное распространение и места
обитания пасюка в СССР. Материалы по грызунам, изд. МОИП, вып. IV, М., 1951.
И. С. Солдаткин, Краткий очерк экологии пластинчатозубой крысы и мер
борьбы с нею в дельте Аму-Дарьи, «Грызуны и борьба с ними», вып. IV, Саратов-
ское книжное издательство, 1955.
Т. 3. 3 а х и д о в, Песчанки и их значение в кормовом балансе пастбищ, «Из-
вестия АН Узбекской ССР», Ташкент, 1948.
А. С. Бур делов и М. Н. Леонтьева, Песчанки северного Прибалхашья,
вып. 3, 1956.
А. Н. Леонтьев, К экологии когтистой песчанки в Бурят-Монгольской
АССР, «Известия Иркутского государственного научно-исследовательского противочум-
ного института Сибири и Дальнего Востока», т. XII (137—348), Иркутск, 1957.
С. А. X а м г а н о в, К биологии когтистой песчанки в районе Торейских озер,
«Известия Иркутского государственного научно-исследовательского, противочумного
института Сибири и Дальнего Востока», т. XII, Иркутск, 1954.
Ю. М. Р а л л ь, Введение в экологию полуденных песчанок, «Вестник микробио-
логии и паразитологии», т. XIX, вып. I, Саратов, 1940.
Н. Н. Бакеев, Н. Ф. Д а р с к а я, Н. Г. К а д а ц к и й, Особенности некоторых
поселений краснохвостой песчанки по левобережью Куры и Азербайджанской ССР,
«География населения животных и методы его изучения», изд. АН СССР, 1959.
Шилова Е. С., О распространении и стационарном размещении краснохвостой
песчанки в Северном Приаралье, «Бюллетень МОИП», отдел биологии, т. VIII
(2), 1953.
Ю. М. Р а л л ь, Очерк экологии гребенщиковой песчанки, «Грызуны и борьба
с ними», изд. Алма-Атинской противочумной станции, вып. 1, 1941.
Т. А. К и м. Материалы по экологии тамарисковой песчанки пустыни Кызылкум,
«Зоологический журнал», т. XXXIX, вып. 5, М., 1960.
С. Н. Варшавский, М. Н. Шилов, Эколого-географические особенности рас-
пространения и территориального распределения большой песчанки в Северном При-
аралье, «Труды Государственного среднеазиатского научно-исследовательского противо-
чумного института», вып. 3, Казгосиздат, 1956.
А. С. Бурделов, В. С. Петров, В. П. Хрусцелевский, Большая пес-
чанка (Rhombomys opimus) как ландшафтный вид пустынь СССР, «Труды Государ-
ственного среднеазиатского противочумного института», изд. АН Казахской ССР, Алма-
Ата, 1960.
А. С. Бурделов, Е. П. Бондарь, Особенности экологии больших песча-
нок в южной и северной пустынях СССР, изд. АН Казахской ССР, Алма-Ата, 1960.
Б. М. Касаткин, Структура северной части ареала большой песчанки и
проблемы оздоровления этой части природного очага чумы, «Труды Государственного
среднеазиатского противочумного института», Алма-Ата, 1960.
Ю. М. С м и р и н, Распределение большой песчанки (Rhombomys opimus) на
правобережье среднего течения Сыр-Дарьи, «Зоологический журнал АН СССР»,
т. XXXIX, вып. 8, 1960.
Н. Н. Воронцов, Виды хомяков Палеарктики, «Доклады АН СССР», т. 132,
№ 6, 1960.
П. П. Гамбарян и Б. А. Мартиросян, Об экологии мышевидного хомячка
(Calomyscus bailwardi), «Зоологический журнал АН СССР», т. XXXIX, вып. 9, I960.
М. Г. Яковлев и М. М. Колесников, Некоторые новые данные по распро-
странению и экологии предкавказского хомяка в Ростовской области, «Зоологический
журнал», т. XXXIII, вып. 3, 1954.
И. П. Митина, Географическая изменчивость хомячка, «Зоологический жур-
нал», т. 38, вып. 12, 1959.
Н. В. Башен ин а, К экологии серого хомячка Европейской части СССР. Сб.
«Фауна и экология грызунов», вып. 3, изд. МОИП, М., 1951.
В. П. Никитин, К биологии крысовидного хомяка, «Известия Иркутского
государственного научно-исследовательского противочумного института Сибири и
Дальнего Востока», т. X, Иркутск, 1952.
357
Н. П. Лавров, Акклиматизация ондатры в СССР, М., 1957.
Е. И. Страутман, Ондатра в Казахстане, Алма-Ата, 1963.
«Водяная крыса и борьба с нею в Западной Сибири». Сб. под ред. А. П. Кузякина,
Новосибирск, 1959.
А. П. Кузякин, Авиационный и другие приманочные методы защиты урожая
от водяной крысы, Прага, 1960.
Т. Н. Дунаева, Сравнительный обзор экологии тундровых полевок, «Труды
Института'географии АН СССР», т. XXI, 1948.
П. Н. Степанов, К экологии гудаурской полевки. Сб. «Фауна и экология гры-
зунов», вып. 5, 1957.
Н. В. Карасева, А. И. Ильченко, Некоторые особенности биологии полев-
ки-экономки, изученные методом мечения зверьков. Сб. «Фауна и экология грызунов»,
вып. 5, 1957.
Г. И. Васильев, К экологии восточной полевки, «Известия Иркутского го-
сударственного научно-исследовательского противочумного института Сибири и Даль-
него Востока», т. XIX, Иркутск, 1958.
Ю. Г. Швецов, Систематические особенности восточной и уйгурской полевок,
«Тезисы докладов научной конференции по природной очаговости», Саратов, 1957.
Б. Н. Васин, Новый вид серой полевки с Сахалина, «Зоологический журнал»,
т. XXXIV, вып. 2, 1955.
Н. В. Б а ш е н и н а, Экология обыкновенной полевки, М., 1962.
С. Н. В а р ш а в с к и й, Г. А. Кондрашкин, У. С. Солдаткин, Заметки
о стадиальном размещении и экологии закаспийской полевки в Казахстане и Сред-
ней Азии, «Грызуны и борьба с ними», вып. 5, Саратов, 1957.
Л. П. С л и н к о, Динамика численности общественных полевок (Microtis socia-
lis) в засушливых районах Ростовской области, «Тезисы докладов научной конферен-
ции», 1960.
С. И. Мамедов, Некоторые данные по экологии общественной полевки в Азер-
байджане, «Тезисы докладов научной конференции по природной очаговости», Сара-
тов, 1957.
В. В. Кучерук и Т. Н. Дунаева, Материалы по динамике численности
полевки Брандта. Фауна и экология грызунов, под ред. А. Н Формозова. Материалы
по грызунам, вып. 3, изд. МОИП, М., 1948.
В. П. Хрусцелевский, Закономерности изменений численности полевки
Брандта в Юго-Восточном Забайкалье, «Известия Иркутского государственного науч-
но-исследовательского противочумного института Сибири и Дальнего Востока», т. XI11,
Иркутск, 1954.
А. Г. О б л е з н е в а, Некоторые наблюдения за полевками Виноградова, «Из-
вестия Иркутского государственного научно-исследовательского противочумного инсти-
тута Сибири и Дальнего Востока», т. XIX, Иркутск, 1958.
Г. А. Новиков, О. В. Петров, Экология подземной полевки в лесостепных
дубравах, «Зоологический журнал», т. XXXI1, вып. 1, 1953.
В. Д. Ш а н е в, Л. В. Шапошников, Подземная полевка в Калининской об-
ласти, «Бюллетень МОИП», Калининское отделение, вып. 1, 1958.
И. М. Громов и Б. Л. Воробьев, Кустарниковая полевка (Pytymys majori)
в высокогорье западной части Главного Кавказского хребта, «Труды Зоологического
ин-та АН СССР», т. XVII, изд. АН СССР, М.—Л., 1955.
Т. В. Кошкина, Сравнительная экология рыжих полевок в северной тайге.
Материалы по грызунам, вып. 5, изд. МОИП, М., 1957.
Н. Шубин, Материалы по экологии лесных полевок северной части Кузбасса,
«Тезисы 11 Всесоюзной научной конференции студентов-биологов», изд. МГУ, 1959.
В. П. Чернышев, К распространению и биологии высокогорной, или серебри-
стой, полевки в Таджикистане, вып. 16, изд. АН Таджикской ССР, 1956.
Н. И. Литвинов, Новые данные о распространении Саянской высокогорной
полевки, «Известия Иркутского государственного научно-исследовательского противо-
чумного института Сибири и Дальнего Востока», т. XIX, Иркутск, 1957.
В. И. Капитонов, О распространении и биологии лемминговидной полевки
в Хараулахских горах, «Зоологический журнал», т. XXXVI11, вып. 11. 1959.
Г. А. Новиков, К экологии лесного лемминга (Myopys schisticolor) на Коль-
ском полуострове, «Зоологический журнал», вып. 4—5, 1941.
А. Насимович, Г. Новиков, О. Семенов-Тяньшанский, Норвеж-
ский лемминг, «Фауна и экология грызунов», вып. 3, изд. МОИП, М., 1948.
В. М. Сдобников, Количественные соотношения популяции обского и копыт-
ного леммингов, «Зоологический журнал», т. XXXIV. вып. 6, М., 1955.
Е. М. Страутман, И. Г. Шубин, К биологии степной пеструшки и узко-
черепной полевки в Северном Казахстане, «Труды Института зоологии АН Казахской
ССР», т. ХШ, 1960.
АТЛАС АРЕАЛОВ
МЛЕКОПИТАЮЩИХ
СССР
60 70 80 90 100 110 120 130 140 150
400нм
0
Еж обыкновенный.
Erinaceus europaetis L.
Даурский ёж.
Erinaceus europacus dauricus Sundcv. 4QQ
/овосибирс/ Красноярск
2
3
3 d\0 И
50 60 70 80 90100 110 120 130 140 150
400км
/ов°сибИрс/ ^КРасноярск
Когтистая бурозубка.
Sorex unguiculatus Dobson.
Альпийская бурозубка.
Sorex alpinus Schinz.
Гигантская бурозубка.
Sorex paclficus Coues.
15
16
18(
40 50 60 70 80 90100 110 120 130 140 150
Ночница Наттерера. Myotis naitereri Kuhl.
+• Длиннохвостая ночница. Myotis frater G. Allen.
400км
Трехцветная ночница. Myotis emarginatus Geoffr.
'°в°сиб / х-4-^Расноярск
21
22
40 50 60 70 80 90100 110 120 130 140 150
&
400км
°вОс"би.
Красноярск
Нетопырь-карлик.
Vespertilio pipistrellus Schreb.
Лесной нетопырь.
Vespertilio nathusii Keys.et Blas.
® Места их общих нахождений
я Восточный нетопырь.
Vespertilio abramus Temm.
30
20
40 50 60 70 80 90 100 110 120 130 140 150
*^СКОЕ
400нм
->Красно"Рск
Корсак Vulpes corsac L.
Афганская лисица Vulpes сапа Blanf.
Песец Vulpes lagopus L.
Южная граница области летнего обитания
Южная граница области зимних кочевок
Отдельные заходы
180'
40 50 60 70 80 90100 110 120 130 140 150
Красноярск
400нм
Л
//
Ов°сибя
Енотовидная собака
Nyctereutes procyonoides Gray.
Места выпусков (основные)
Енот-полоскун Procyon lotor L.
60 70 80 90 100 110 120 130 140 150
42
400нм
Леопард Panihera pardus L.
Полосатая гиена Hyaena hyaena
Гепард Acinonyx jubatus Schreb.
^ChOF.
ибирс/ rJHpaCH0"PCH
50 60 70 80 90100 110 120 130 140 150
180’
400км
Тигр Panlhera tigris L.
Лесная кошка Felis Silvestris Schreb.
IIIIIIIIIIIIII Степная кошка Fells libyca Forster.
°*°c*ffwpc’ ^HpaCHO”PCH
50 60 70 80 90100 110 120 130 140 150 160
170
180-
Красноярск
400нм
Рысь Felis lynx L.
Хаус (Камышовый кот).
Felts chaus Gold.
Каракал Felis caracal Schreb
10
30
70 80 90 100 110 120 130 140
•рна
400
400нм
Европейская норка
Mustela lutreola L.
Американская норка.
Mustela vison Briss.
Основные места
акклиматизации.
60 70 80 90 100 110 120 130 140 150
Л
Его основные лежбища
1оа°СнбИр I л-^НРасноярсн
Морж Odobaenus rosmarus
Его основные береговые
сЭ лежбища
Морской котик Callorhinus
urslnus L.
(Основные лежбища)
Пути осенних миграций
00 Q 400нм
10
20
30
18(
40 50 60 70 80 90100 110 120 130 140 150 160
400мм
*-*САГО£'
ОвОс*бйрс/ х^Расноярск
Лысун
Phoca groenlandica Erxl.
Основные места детных
лежбищ
Крылатка.
Phoca fasciaia Zimm.
Основные места детных
лежбищ___________
Морской заяц.
Erignaihus barbatus Erxl.
Длинномордый
тюлень.(Тевяк)
Halichocrus grypus Fabr.
Хохлан Cysiopliora
cristaia Erxl.(Заплывы) 400
Карта составлена проф.А.Г. Томилиным
18(
400нм
Кабарга. Moschus moschiferus L.
Отдельные заходы
Пятнистый олень.Cervus nippon Temm
Места акклиматизации
JIaHb.Cervus dama L.
(Места акклиматизации)
70 80 90 100 110 120 130 14Q
Косуля. Capreolus capreolus L.
Места отдельных заходов
Места акклиматизации
400
400нм
10
20
30
18(
70 80 90100 110 120 130 140 150
Сайга. Saiga taiarica L.
- Отдельные заходы
Места акклиматизации
’Красноярск
[•.рСерна.Rupicapra rupicapra L.
И Джейран. GazelJa subguttu-
Orosa Giild.
Горал. Nemorhaedus Дзерен. Procapra gutturosa 4qq ~ 4C
g goral Hardwicke Gmel. | ।—। p—i ------1
30
20
40 50 60 70 80 90100 110 120 130 140 150
400 нм
Винторогий козел.
Capra falconeri Wagner.
Безоаровый козел.
»Capra aegagrus Erxl.
**С<О£
’**4x4 r^paCMO"PCH
Центрально - азиатский
козел.
° Cppra sibirica Pallas.
Кавказский тур.
Capra caucasica Gold, и
дагестанский тур.
Capra cylindricornis Blyth.
-у- <1. Iijubiiiiuu rail. aar^Jinma июацпп n
Места акклиматизации реакклиматизации. 400 0 400нм
30
20
GO
40 50 60 70 80 90100 110 120 130 140 150
400км
Расноярск
Северная пищуха.
Ochotona hyperborea Pall.
Алтайская пищуха.
Ochotona alpina Pall.
Палласова пищуха
Ochotona pallast Gray.
Красная пищуха.
Ochotona rutila Severtzov.
Малая пищуха
Ochotona pusilia Pall.
Даурская пищуха.
Ochotona daurica Pall.
Рыжеватая пищуха.
Ochotona rutesccns Gray.
Болылеухая пищуха.’
Ochotona macrotis Gunther.
60 70 80 90 100 110 120 130 140 150
400нм
песчаник.
Lepus tibetanus Wat.
1 Заяц-русак.
J Lepus europaeus Pali.
Места выпуска
зайца-русака.
Дикий кролик.
Oryctolagus cuniculus L.
Дальневосточный заяц.
Lepus brachyurus Temm.
Г] Заяц-беляк.
*1 Lepus timidus L.
Заяц-толай.
Lepus tolai Pall.
60 70 80 90 100 110 120 130 140 150
400нм
Длиннохвостый сурок. Marmota caudata Jacquemont.
Сурок обыкновенный. Marmota bobak Muller.
Места акклиматизации обыкновенного сурка.
Сурок Мензбира. Marmora menzbieri Kaschkarov.
**CKOt
Ираснояосн
61
64
65
60 70 80 90 100 110 120 130 140 150
90
• Тушканчик большой. Allactaga major Kerr.
д Тушканчик Северцова.
Allactaga severtzovi Vinogr
Оаос^рс
Красноярск
50 60 70 80 90100 110 120 130 140 150
Тушканчик малый. Allactaga elater Licht.
400км
100
90
Тушканчик Бобринского. Allactaga
bobrinskii Kolesn.
Zoe°c*»ffHpc' ^НрасноиРсн
60 70 80 90 100 110 120 130 140 150
//
400км
Толстохвостый тушканчик
Pygereimus platyurus Licht.
Земляной зайчик.
Alactagulus pygmaeus Pall.
Тушканчик Житкова.
Тушканчик Виноградова.
* Pygeretmus vinogradovi Vorontzov.
^шлапчпп mninuDa. ПяТИПаЛЫЙ КарЛИКОВЫЙ
Pygeretmus zhltkovi Kuznetzov. ♦ gX’craniui paradoxus Satunin.
A Трехпалый карликовый
А тушканчик.
Salpingotus crassicauda Vinogr.
•oc-e»pci
’Красноярск
70
Одноцветная мышовка
Sicista concoJor Blichner
Алтайская серая мышовка
Sicista pseudonapaea Str.
76
60 70
400км
Полуденная песчанка.
Meriones meridianus Pall.
Когтистая песчанка.
Meriones uhguiculatus Milne-Edw.
Персидская песчанка.
Meriones persicus Blanf.
'Красноярск
80
70 80 90 100 110 120 130 140 150
400нм
Переднеазиатский хомяк.
Cricetus auratus Waterh.
”1 Хомяк обыкновенный.
J Cricetus cricetus L.
Мышевидный хомячок. е Предкавказский хомяк.
Calomyscus bailwardi Thomas. Cricetus raddei Nehring.
90
18(
50 60 70 80 90 100 110 120 130 140 150
400км
100
Роборовского.
roborovskii Satunin.
Эверсманна.
eversmanni Brandt.
Джунгарский хомячок.
Phodopus sungorus Pall.
Хомячок
Phodopus
Хомячок
Cricetulus
) 80 90 100 110 120 130 140
Ондатра.
Ondatra zibethicus L. 400
400нм
Карта составлена Г.Корсаковым
10 20
00
Сп
м о
400нм
Северосибирская полевка. Microtus hyperboreus Vinogr.
Полевка Миддендорфа. Microtus middendorfi Poliakov.
Тёмная полевка. Microtus agrestis L.
'Красноярск
60 70 80 90 100 ПО 120 130 140 150
400км
Общественная полевка Microtus socialis Pall.
Афганская полевка Microtus afghanus Thomas
Арчевая полевка Microtus juldaschi Sevetz
Полевка Брандта Microtus brandti Radde
Полевка китайская Microtus mandarinus M.-Edw.
Г°в°сибИр / ^Красноярск
60 70 80 90 100 110 120 130 140 150
Красноярск
400нм
Узкочерепная полевка Microtus gregalis Pall
Кустарниковая полевка Microtus subterrancus de S. Long
Малоазиатская полевка Microtus majori Thom
4- Полевка Шелковникова Microtus schelkovnikovi Sat
10
30
60 70 80 90 100 110 120 130 140 150
400км
90
Серебристая полевка Alticola roylei Gray
«ф. Гоби-Алтайская полевка. Alticola barakschin Вапп.
А Большеухая полевка Alticola macrotis Radde
Плоскочерепная полевка Alticola strelzovi Kastsch.
**СКОЕ
ОвОС^ИРс/ z>HPaCMO«PCH
60 70 80 90 100 110 120 130 140 150
490км
Цокор обыкновенный Myospalax myospalax Laxsm
Слепыш обыкновенный Spalax microphthalmus Giild.
Гигантский слепыш Spalax giganteus Nehring
Малый слепыш Spalax leucodon Nordm.
Красноярск
М о Р Е
I
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
РУССКИХ НАЗВАНИЙ МЛЕКОПИТАЮЩИХ ФАУНЫ СССР
Обычными цифрами указаны страницы списка видов, определительной таблицы
и заглавия текста по данной форме или группе; римские цифры означают номер таб-
лицы в атласе цветных рисунков и цифры, выделенные жирным шрифтом, — номер
карты с ареалом данного вида.
Азиатская широкоушка 6, 100, VII, 27
Азиатский муфлон 11, 230, XXI1
Алтайская пищуха 11, 240, 241, XXI11, 66
Алтайская рыжая мышовка 13, 288, 289,
84
Алтайская серая мышовка 13, 288, 289,
84
Алтайский крот 5, 50, 52, 11, 6
Альпийская бурозубка 5, 61, 66, 111, 12
Американская норка 8, 155, 161, XIV, 52
Амурский лемминг 14, 338, XXXIX, 108
Архар 11, 230, XXII, 65
Арчевая полевка '14, 319, 329, XXXV11,
104
Атлантический ремнезуб 9, 196, 197
Афалина 9, 186, XVII
Афалины 9, 182, 186, XVII
Афганская лисица 7, 124, 127, IX, 39
Афганская полевка 14, 319, 328, XXXV11,
104
Афганская слепушонка 14, 342, 343, XL,
109
Афганский еж 40, 43
Баран толсторогий 11, 230, 233, ХХП, 65
Бараны 11, 225, 230, ХХП, 65
Байкальская нерпа 8, 173, 177, XVI, 55
Барс 7, 135, 136, XI, 42
Барсук 7, 146, XII, 46
Барсуки 7, 143, 146, XII, 46
Барханная кошка 7, 138, 140, XI, 44
Беззубых китов, подотряд 9, 181, 199,
XV111, 57
Безоаровый козел 10, 226, 229, XXI, 64
Беличьи 11, 249, 255, XXV—XXV11, 69—73
Белка кавказская 12, 267, XXVI1, 73
Белка обыкновенная 12, 267. XXVI1, 73
Белки 12, 257, 266, XXVI1, 73
Белобокий дельфин 9, 187, 188
Белобочка дельфин 9, 185, XVII, 57
Белобрюхая белозубка 6, 72, 76, IV, 16
Белобрюхий стрелоух 6, 113, V111, 36
Белозубка белобрюхая 6, 72. 76, IV, 16
— белохвостая 6, 72, 74, IV
— большая 6, 72, 75, IV, 15
— бурая 6, 72, 74, IV, 15
Белозубка карликовая 6, 69, IV, 14
— малая 6, 71, 73, IV, 14
— серая 6, 72, 76
— сибирская 6, 77, IV
Белозубки 6, 58, 69, IV, 14—16
Белозубки многозубые 6, 58, 69, IV
Белокрылая свинья 9, 192, XVII
Белокрылые свиньи 9, 184, 192, XVII
Беломордый дельфин 9, 187, 188, XVII
Белохвостая белозубка 6, 72, 74, IV
Белуха 9, 192, XVII, 57
Белухи 9, 185, 192, XVII, 57
Белые медведи 7, 130, 132, X, 41
Белый медведь 7, 132, X, 41
Бенгальская кошка 7, 138, 140, XI, 44
Бесперая свинья 9, 192
Бесперые свиньи 9, 184, 192
Благородный олень 10, 216, XX, 60
Бобр 11, 254, XXV, 68
Бобровые 11, 249, 254, XXV, 68
Бобры 11, 254, XXV, 68
Большая белозубка 6, 72, 75, IV, 15
Большая могера 5, 54, 11, 6
Большая ночница 6, 90, 93, VI, 21
Большая песчанка 13, 306, ХХХ11, 95
Большеухая пищуха 11, 241, 243, ХХШ,
66
Большеухая полевка 14, 335, 337,
XXXV111, 107
Большие песчанки 13, 300, 306, ХХХ11,95
Большой подковонос 6, 85, 87, V, 19
Большой суслик 11, 261, 262, XXVI, 70
Большой трубконос 7, Ц4, V111, 35
Большой тушканчик 12, 278, 279, XXIX,
78
Буйвол домашний 11, 235, ХХП
Буйволы 11, 235, ХХП
Бульдоговые 7, 82, 114, V111, 36
Бурая белозубка 6, 72, 74, IV, 15
Бурозубка альпийская 5, 61, 66, 111, 12
— бухарская 5, 60, 62, 8
— гигантская 5, 61, 66, 111, 12
— когтистая 5, 61, 66, 111, 12
— крошечная 5, 59, 61, 111, 7
— малая 5, 59, 62, 111, 8
— обыкновенная 5, 61, 64, III, 11
23а Бобринский
361
Бурозубка плоскочерепная 5, 11
— средняя 5, 60, 63, III, 10
— темнозубая 5, 60, 63, III, 9
Бурозубки 5, 57, 58, III, 7—12
Бурундук 12, 266, XXVII, 72
Бурундуки 12, 257, 266, XXVII, 72
Бурые медведи 7, 130, 131, X, 41
Бурый медведь 7, 131, X, 41
Бутылконос высоколобый 9, 195, XVII
Бутылконосы 9, 194, XVII
Бухарская бурозубка 5, 60, 62, 8
Бухарский подковонос 6, 85, 88, V, 19
Быки II, 234, 235, XXII, 65
Бычьих, подсемейство 234
Верблюд двугорбый 10, 212, XIX
— одногорбый 10, 212, XIX
Верблюдовые 10, XIX
Верблюды 10, XIX
Вечерница восточная 6, 101
— гигантская 6, 101, 102, VII, 28
— малая 6, 102, ЮЗ, VII, 30
— рыжая 6, 102, VII, 29
Вечерницы 6, 89, 101, VII, 28—30
Винторогий козел 10, 226, 228, XXI, 64
Водяная крыса 13, 317, XXXV, 100
Водяная кутора 5, 67, 68, III, 13
Водяная ночница 6, 91, 94, VI, 22
Водяные крысы 314, 316, XXXV, 100
Волк 7, 121, IX, 37
— красный 7, 123, IX, 37
Волки красные 7, 120, 123, IX, 37
Восточная вечерница 6, 101
Восточноазиатская мышь 13, 294, 295,
XXXI, 88
Восточный кожан 6, 107, III, VIII, 34
Восточный нетопырь 6, 105, 108, 31
Выдра 8, 162, XIV, 53
Выдры 8, 145, 162, XIV, 53
Высоколобый бутылконос 9, 195, XVII
Выхухолевые 5, 37, 43, II, 4
Выхухоли 5, 44, II, 4
Выхухоль 5, 44, II, 4
Газели 10, 222, XXI, 63
Газелевых, подсемейство 221, 222
Гепард 7, 135, XI, 42
Гепарды 7, 134, XI, 42
Гиена полосатая 7, 133, X, 42
Гиеновые 7, 119, 133, X, 42
Гиены 7, 133, X, 42
Гигантская бурозубка 5, 61, 66, III, 12
Гигантская вечерница 6, 101, 102, VII, 28
Гигантский слепыш 14, 345, 346, XL, ПО
Гладкокитовые 10, 199, 204, XVIII
Гладконосые 6, 82, 88, VI—VIII, 20—36
Гребенчуковая песчанка 13, 302, 305,
XXXII, 95
Гребнепалые тушканчики 12, 283, 284,
XXX, 83
Гребнепалый тушканчик 12, 284, XXX, 83
Гренландские киты 10, 204, 205, XVIII
Гренландский кит 10, 205, XVIII
Гринда 9, 190, XVII
Гринды 9, 183, 190, XVII
Гоби-алтанская полевка 14, 335, 337, 107
Горал 10, 225, XXI, 63
Горалы 10, 224, 225, XXI, 63
Горбатые киты 9, 200, 203, XVIII
Горбач 10, 203, XVIII
Горностай 8, 154, 157, XIII, 49
Горная мышь 13, 294, 297, XXXI, 90
Горный тушканчик 12, 278, 280, XXIX, 79
Грызуны II, 35, 236, XXIII—XL, 66—110
Дагестанский тур II, 228, 230, XXI, 64
Дальневосточный заяц II, 244, 245,
XXIV, 67
Даурская пищуха II, 241, 242, XXIII, 66
Даурский еж 1, I
Даурский хомячок 13, 311, XXXIV, 98
Двупарнорезцовых, подотряд II, 238
Дельфин белобокий 9, 187, 188
— белобочка 9,185, XVII, 57
— беломордый 9, 187, 188,. XVII
— китовидный 9, 187
— полосатый 9, 187, 188, XVII
— серый 9, 190
Дельфиновые 9, 181, XVII, 57
Дельфины-белобочки 9, 182, 185, XVII, 57
— китовидные 9, 182, 187
— короткоголовые 9, 182, 187
— серые 9, 183, 190
Джейран 10, 222, XXI, 63
Джунгарский хомячок 13, 309, 310,
XXXIII, 97
Дзерен 10, 223, XXI, 63
Дзерены 10, 222, 223, XXI
Дикий кролик II, 244, XXIV, 67
Дикобраз II, 253, XXV, 68'
Дикобразовые II, 248, 253, XXV, 68
Дикобразы II, 253, XXV, 68
Длиннокрыл обыкновенный 6, 98, VI, 25
Длиннокрылы 6, 89, 97, VI, 25
Длинномордые тюлени 8, 170, 178, XVI,
56
Длинномордый тюлень 8, 178, XVI, 56
Длиннопалая ночница 6, 91, 94, VI. 22
Длинноухая ночница 6, 90, 92, VI, 20
Длиннохвостая ночница 6, 92, 97, VI, 23
Длиннохвостая полевка 14, XXXVI
Длиннохвостый сурок II, 258, XXV, 69
Длиннохвостый суслик 12, 261, 264,
XXVI, 72
Длиннохвостый хомячок 13, 311, 312,
XXXIV, 98
Домовая мышь 13, 292, XXXI, 86
Домовые мыши 13, 291, 292, XXXI, 86
Домашний буйвол 11, 235, XXII
Домашняя лошадь 10, 208
Двугорбый верблюд 10, 212, XIX
Двупарнорезцовые И, 238, XXIII—XXIV,
66—67
Двуцветный кожан 6, 106, 111, VIII, 34
Европейская норка 8, 155, 160, XIV, 52
Европейская широкоушка 6, 100, VII, 27
Единозубы 9, 185, 193, XVII
Еж даурский 1, I
— лысый 5, 40, 43, I, 3
— обыкновенный 5, 40, I, 1
— ушастый 5, 40, 42, I, 2
Ежи 5, 38, I, 1—3
Ежовые 5, 37, 38, 1, 1—3
Емуранчик обыкновенный 12, 283, XXX,
82
Емуранчики 12, 283, XXX, 82
362
Енот-полоскун 7, 129, X, 40
Енотовидная собака 7, 128, X, 40
Енотовидные собаки 7, 121, 128, X, 40
Еноты 7, 129, X, 40
Енотовые 7, 119, 129, X, 40
Жвачных, подотряд 10, 210, 212, XIX—
XXII
Желтая пеструшка 14, 340, 341, XXXIX
Желтогорлая мышь 13, 294, 296, XXXI,
89
Желтый суслик И, 261, XXVI, 70
Зайцы И, 243, 244, XXIV, 67
Зайцы морские 8, 170, 177, XVI, 56
Зайчик земляной 12, 281, XXIX, 81
Зайчики земляные 12, 277, 281, XXIX, 81
Закаспийская полевка 14, XXXVI
Заяц-беляк И, 245, 246, XXIV, 67
— дальневосточный И, 244, 245, XXIV,
67
— морской 8, 177, XVI, 56
— русак 11, 245, 247, XXIV, 67
— толай 11, 245, 247, XXIV, 67
Заячьи 11, 238, 243, XXIV, 67
Землеройковые 5, 38, 54, 111—IV, 7—16
Земляной зайчик 12, 281, XXIX, 81
Земляные зайчики 12, 277, 281, XXIX, 81
Зубатых китов, подотряд 9, 181, XVII—
XV111, 57
Зубр И, 235, XXII, 65
Кабан 10, 210, XIX, 58
Кабаны 10, 210, 58
Кабарга 10, 213, XIX, 59
Кабарги 10, 213, XIX, 59
Кабарговых, подсемейство 10, 213, XIX
Кавказская белка 12, 267, XXV11, 73
Кавказская мышовка 287, 289, 84
Кавказский тур 10, 228, 229, XXI, 64
Калан 8, 163, XIV, 53
Каланы 8, 145, 163, XIV, 53
Каменная куница 8, 149, 150, Х111, 47
Каменные полевки 14, 315. 335, XXXV11I,
107
Каракал 7, 139, 141, XII, 45
Карликовая белозубка 6, 69, IV, 14
Карликовые кашалоты 9, 197, 198, 57
Карликовые пятипалые тушканчики 12,
286, XXX, 81
Карликовые трехпалые тушканчики 12,
286, XXX, 81
Карликовый кашалот 9, 198, XV111, 57
Карликовый пятипалый тушканчик 13,
286, XXX, 81
Карликовый трехпалый тушканчик 13, 286,
XXX, 81
Карликовых тушканчиков, подсемейство
276, 285
Каспийская нерпа 8, 173, 176, XVI, 55
Каспийский, тюлень 8, 173, 176, XVI, 55
Кашалот 9, 198, XV111
— карликовый 9, 198, 57
Кашалотовые 9, 181, 197, XVIII, 57
Кашалоты 9, 198, XVIII, 57
— карликовые 9‘, 197, 198, 57
Кит гренландский 10, 205, XVIII
— серый 9, 199, XVIII, 57
— синий 9, 201, 203, XVIII
Кит южный 10, 204, XV111
Китайская полевка 14, 328, 329, XXXV11,
104
Китовидные дельфины 9, 182, 187
Китовидный дельфин 9, 187
Китообразные 9, 35, 180, XVII—XV111, 57
Киты гренландские 10, 204, 205, XVI11
— серые 9, 199, XV111, 57
— южные 10, 204, XV111
Клюворыл 9, 195, XVII
Клюворылы 9, 194, 195, XVII
Клюворылые 9, 181, 193, XVII
Когтистая бурозубка 5, 61, 66, 111, 12
Когтистая песчанка 13, 301, 303, ХХХ11,
93
Кожан восточный 6, 107, 111, V111, 34
— двухцветный 6, 106, 111, V111, 34
— Огнева 6, 107, 112, V111, 35
— поздний 6, 107, 112, V111, 35
Кожанки 6, 106, VII
Кожановидный нетопырь 6, 106, 109, VII,
32
Кожанок Бобринского 6, 106, НО, VII, 33
— северный 6, 106, 109, VII, 33
Кожаны и нетопыри 6, 89, 103, VII, 31—35
Козел безоаровый 9, 10 226, 229, ХХ1, 64
— винторогий 10, 226, 228, XXI, 64
— центральноазиатский 10, 228, 229, XXI,
64
Козлиных, подсемейство 10, 222, 223, XXI
Козлы 10, 225, 226, XXI, 64
Колонок 8, 155, 158, XIV, 50
Кольчатая нерпа, 8, 173, 175, XVI, 55
Командорский ремнезуб 9, 196, XVII
Копытные лемминги 14, 315, 339, XXXIX,
108
Копытный лемминг 14, 339, XXXIX, 108
Короткоголовые дельфины 9, 182, 187
Корсак 7, 124, 127, IX, 39
Косатка 9, 189, XVII
Косатка черная 9, 183, 189
Косатки 9, 183, 189, XVII
Косатки черные 9, 183, 189
Косули 10, 214, 218, XX, 61
Косуля 10, 218, XX, 61
Котик морской 8, 167, XV, 54
Котики морские, 8, 167, XV, 54
Котиковые 8, 165, 166, XV, 54
Кошачьи 7, 119, 133, XI—XII, 42—45
Кошка барханная 7, 138, 140, XI, 44
— бенгальская 7, 138, 140, XI, 44
— лесная 7, 138, 139, XI, 43
— степная 7, 138, 139, XI, 43
Кошки 7, 134, 137, X—XII, 43—45
Крапчатый суслик И, 261, 262, XXVI, 70
Красная пищуха И,. 240, 242, ХХ111, 66
Красная полевка 14, 332, 334, XXXV111,
106
Красносерая полевка 14, 332, XXXV111,
106
Красные волки 7, 120, 123, IX, 37
Красный волк 7, 123, IX, 37
Краснохвостая песчанка 13, 301, 304,
XXXII, 94
Кролик дикий И, 244, XXIV, 67
Кролики И, 243, XXIV, 67
Крот алтайский 5, 50, 52, II, 6
— малый 5, 49, 50, II, 5
— обыкновенный 5, 49, 50, И, 6
363
Кроты 5, 49, II, 5—6
Кротовые 38, 47, II, 5—6
Кротовых, подсемейство 5,-47, II
Крошечная бурозубка 5, 59, 61, III, 7
Крылатка 8, 172, 174, XVI, 56
Крыса водяная 14, 317, XXXV, 100
— пластинчатозубая 13, 300, XXXI, 92
— туркестанская 13, 298, 299, XXXI, 91
— черная 13, 298, XXXI, 91
Крысовидный хомяк 13, 311, 313, XXXIV,
98
Крысы 13, 292, 297, XXXI, 91-92
— водяные 14, 314, 316, XXXV, 100
— пластинчатозубые 13, 292, 299, XXXI,
92
Кулан 10, 207, 208, XIX, 58
Куница каменная 8, 149, 150, XIII, 47
— лесная 8, 149, 150, XIII, 47
Куницы 8, 143, 148, XIII, 47
Куньи 7, 120, 142, XII—XIV, 46—53
Кустарниковая полевка 14, 331, XXXVII,
105
Кутора водяная 5, 67, 68, III, 13
— малая 5, 67, III, 13
Куторы 5, 57, 67, III, 13
Лань 10, 215, 216, XX, 59
Ласка 8, 154, 156, XIII, 48
Ластоногие 8, 35, 164, XV—XVI, 54-56
Лемминг амурский 14, 338, XXXIX, 108
— копытный 14, 339, XXXIX, 108
- лесной 14, 337, XXXIX, 108
— норвежский 14, 338, 339, XXXIX, 108
— сибирский 14, 338, XXXIX, 108
Лемминги копытные 14, 315, 339, XXXIX,
108
— лесные 14, 315, 337, XXXIX, 108
— настоящие 14, 315, 338, XXXIX, 108
Леммипговидная полевка XXXVIII
Леопард 7, 135, 136, XI, 42
Лесная кошка 7, 138, 139, XI, 43
Лесная куница 8, 149, 150, XIII, 47
Лесная мышовка 13, 287, 288, XXX, 84
Лесная мышь 13, 294, 296, XXXI, 90
Лесной нетопырь 6, 104, 108, VII, 31
Лесная полевка 14, XXXVIII
Лесная соня 12, 273, XXVIII, 75
Лесной лемминг 14, 337, XXXIX, 108
Лесные лемминги 14, 315, 337, XXXIX, 108
Лесные (п полевые) мыши 13, 292, 294,
XXXI, 87-90
Лесные полевки 14, 315, 331, XXXVIII, 106
Лесные сони 12, 271, 273, XXVIII, 75
Летяга 12, 270, XXVIII, 73
Летяги 12, 270, XXVIII, 73
Летяжьи 12, 249, 270, XXVIII, 73
Лисица 7, 124, 125, IX, 38
— афганская 7, 124, 127, IX, 39
Лисицы 7, 120, 124, IX, 38—39
Лоси 10, 214, 219, XX, 62
Лось 10, 219, XX, 62
Лошади 10, 206, XIX, 58
Лошадиные 10, 206, XIX, 58
Лошадь 10, 208
Лысун 8, 171, 173, XVI, 56
Лысый еж 5, 40, 43, I, 3
Малая белозубка 6, 71, 73, IV, 14
Малая бурозубка 5, 59, 62, III, 8
Малая вечерница 6, 102, ЮЗ, VII, 30
Малая кутора 5, 67, III, 13
Малая ночница 6, 91, 95, VI, 24
Малая пищуха II, 240, 242, XXIII, 66
Малоазиатская песчанка 13, 302, 305,
XXXII, 95
Малые песчанки 13, 300, XXXII, 93—95
Малый крот 5, 49, 50, II, 5
Малый подковонос 6, 84, 85, V, 17
Малый полосатик 9, 201, XVIII
Малый слепыш 14, 345, XL, ПО
Малый суслик II, 261, 263, XXVI, 71
Малый трубконос 7, 114, VIII, 35
Малый тушканчик 12, 278, 280, XXIX, 80
Малютка мышь 13, 293, XXXI, 85
Малютки мыши 13, 291, 293, XXXI, 85
Манул 7, 138, 140, XII, 44
Медведи белые 7, 130, 132, X, 41
— бурые 7, 130, 131, X, 41
— черные 7, 130, X, 41
Медведь белый 7, 132, X, 41
— бурый 7, 131, X, 41
— черные 7, 130, X, 41
Медвежьи 7, 119, 130, X, 41
Медоед 7, 147, XII, 46
Медоеды 7, 143, 147, XII, 46
Млекопитающие 5, 31
Многозубые белозубки 6, 58, 69, IV
Могера большая 5, 54, II, 6
— средняя 5, 53, 54, II, 6
Могеры 5, 49, 53, II, 6
Мозоленогих, подотряд 10; 209, 212, XIX
Морж 8, 169, XV, 54
Моржи 8, 169, XV, 54
Моржовые 8, 166, 168, XV, 54
Морская свинья 9, 191, XVII
Морские зайцы 8, 170, 177, XVI, 56
Морские котики 8, 167, XV, 54
Морские свиньи 9, 183, 191
Морской заяц 8, 177, XVI, 56
Морской котик 8, 167, XV, 54
Мохноногие тушканчики 12, 283, 284, XXX,
83
Мохноногие хомячки 13, 307, 309, XXXIII,
97
Мохноногий тушканчик 12, 284, XXX, 83
Муфлон азиатский II, 230, XXII
— европейский II, 230, XXII, 65
Мышевидная соня 12, 275, XXVIII, 77
Мышевидные сони 12, 272, 274, XXVIII, 77
Мышевидные хомячки 13, 306, 307,
XXXIII, 96
Мышевидный хомячок 13, 307, XXXIII
Мышей, подсемейство 13, 290, 291, XXXI
Мыши домовые 13, 291, 292, XXXI, 86
— лесные (и полевые) 13, 292, 294, XXXI,
87—90
— малютки 13, 291, 293, XXXI, 85
Мышиные 13, 253, 289, XXXI—XL, 85—
ПО
Мышовка алтайская рыжая 13, 288, 289,
84
Мышовка алтайская серая 13, 288, 289,84
— лесная 13, 287, 288, XXX, 84
— одноцветная 13, 287, 289, XXX, 84
— степная 13, 287, 288, XXX, 84
Мышовки 13, 287, XXX, 84
Мышовковые 13, 252, 286, XXX, 84
Мышь азиатская 13, 294, 295, XXXI, 88
364
Мышь горная 13, 294, 297, XXXI, 90
— домовая 13, 292, XXXI, 86
— желтогорлая 13, 294, 296, XXXI, 89
— лесная 6, 294, 296, XXXI, 90
— малютка 13, 293, XXXI, 85
— полевая 13, 294, 295, XXXI, 87
Нарвал 9, 193, XVII
Насекомоядные 5, 34, 36, 1—IV, 1—16
Настоящие лемминги 14, 315, 338, XXXIX,
108
Настоящие олени 10, 214, XX, 59—60
Нежвачных, подотряд 10, 209, 210, XIX
Непарнокопытные 10, 35, 206, XIX, 58
Нерпа 8, 172, 175, XVI, 55
Нерпа байкальская 8, 173 177, XVI, 55
— каспийская 8, 173, 176, XVI, 55
— кольчатая 8, 173, 175, XVI, 55
Нетопыри и кожаны 6, 89, 103, VII, 31 —
35
Нетопырь восточный 6, 105, 108, 31
— карлик 6, 104, 107, VII, 31
— кожановидный 6, 106, 109, VII, 32
— лесной 6, 104, 108, VII, 31
— средиземный 6, 105, 108, VII, 32
Норвежский лемминг 14, 338, 339, XXXIX,
108
Норка американская 8, 155, 161, XIV, 52
— европейская 8, 155, 160, XIV, 52
Ночница большая 6, 90, 93, VI, 21
— водяная 6, 91, 94, VI, 22
— длиннопалая 6, 91, 94, VI, 22
—1 длинноухая 6, 90, 92, VI, 20
— длиннохвостая 6, 92, 97, VI, 23
— малая 6, 91, 95, VI, 24
— Наттерера 6, 91, 94, VI, 23
— остроухая 6, 90, 92, VI, 21
— прудовая 6, 90, 93, VI, 20
— трехцветная 6, 91, 95, VI, 23
— усатая 6, 92, 96, VI, 24
Ночницы 6, 89, 90, VI, 20—24
Нутриевые 11, 249, 255, XXV, 68
Нутрии 11, 255, XXV, 68
Нутрия 11, 255, XXV, 68
Общественная полевка 14, 323, 327,
XXXVI1, 104
Обыкновенная белка 12, 267, XXVI1, 73
Обыкновенная бурозубка 5, 61, 64, 111, 11
Обыкновенная лисица 7, 124, 125, IX, 38
Обыкновенная полевка 14, 323, 325,
XXXVI, 103
Обыкновенная слепушонка 14, 342, XL, 109
Обыкновенный длиннокрыл 6, 98, VI, 25
Обыкновенный еж 5, 40, 1, 1
Обыкновенный емуранчик 12, 283, XXX,
82
Обыкновенный крот 5, 49, 50, 11, 6
Обыкновенный слепыш 14, 345, 346, XL,
ПО
Обыкновенный сурок 11, 258, 259, XXV,
69
Обыкновенный тюлень 8, 172, 174, XVI,
55
Обыкновенный хомяк 13, 308, ХХХ111,96
Одногорбый верблюд 10, XIX
Однопарнорезцовых, подотряд 11, 238,
248, XXV—XL, 68-110
Одноцветная мышовка 13, 287, 289, XXX,
84
Олени настоящие 10, 214, XX
— северные 10, 214, 220, XX, 62
Олень благородный 10, 216, XX, 60
— пятнистый 10, 216, XX, 59
— северный 10, 220, XX, 62
Оленьи 10, XIX—XX
Оленьих, подсемейство 10, 213, 214, XX
Ондатра 14, 316, XXXV, 99
Ондатры 14, 314, 316, XXXV, 99
Орешниковая соня 12, 274, XXVI11, 77
Орешниковые сони 12, 272, 274, XXVI11,
77
Осел домашний 10, 207
Остроухая ночница 6, 90, 92, VI, 21
Палласова пищуха 11, 240, 242, ХХ111,
66
Пантеры 7, 134, 135, XI, 42—43
Парнокопытные 10, 35, 209, Х1Х-—ХХП,
58—65
Пасюк 13, 298, 299, XXXI, 92
Пегий путорак 6, 78, IV, 16
Перевязка 8, 153, XI11, 48
Перевязки 8, 145, 153, XI11, 48
Переднеазиатский хомяк 13, 308, 309,
XXX111, 96
Персидская песчанка 13, 301, 303, XXXI1,
93
Песец 7, 125, 127, IX, 39
Пестрые хомяки 13, 307, XXXI11, 96
Пеструшка желтая 14, 340, 341, XXXIX
— степная 14, 340, XXXIX, 109
Пеструшки 14, 315, 340, XXXIX, 109
Песчанка большая 13, 306, XXXI1, 95
— Виноградова 13, 301, 304, XXXI1, 94
— гребенчуковая 13, 302, 305, XXXI1, 95
— Зарудного 13, 301, 305, XXXI1, 95
— когтистая 13, 301, 303, ХХХ11, 93
— краснохвостая 13, 301, 304, XXXI1, 94
— малоазиатская 13, 302, 305, XXXI1, 95
— персидская 13, 301, 303, XXXI1, 93
— полуденная 13, 301, 303, XXXI1, 93
Песчанки большие 13, 300, 306, ХХХ11,95
— малые 13, 300, XXXI1, 93—95
Песчанок, подсемейство 13, 290, 300,
XXX11
Песчаные тушканчики 12, 283, 285, XXX,
82
Пищуха алтайская 11, 240, 241, ХХ111, 66
— большеухая 11, 241, 243, ХХ111, 66
— даурская 11, 241, 242, XXI11, 66
— красная 11, 240, 242, ХХ111, 66
— Палласова 11, 240, 242, XXI11, 66
— рыжеватая 11, 241, 243, ХХ111, 66
— северная 11, 239, 241, ХХ111, 66
— степная 11, 240, 242, ХХ111, 66
Пищухи 11, 239, XXI11, 66
Пищуховые 11, 238, 239, XXI11
Плавун северный 9, 195, XVII
Плавуны 9, 194, 195, XVII
Пластинчатозубая крыса 13, 300, XXXI,
92
Пластинчатозубые крысы 13, 292, 299,
XXXI, 92
Плоскочерепная бурозубка 5, И
Плоскочерепная полевка 14, 335, XXXV111,
107
365
Подковонос большой 6, 85, 87, V, 19
— бухарский 6, 85, 88, V, 19
— малый 6, 84, 85, V, 17
— Мегели 6, 84, 87, V, 18
— средиземный 6, 84, 86, V, 17
— южный 6, 84, 86, V, 18
Подковоносы 6, 82, V, 17—19
Подковоносые 6, 81, 82, V, 17—19
Поздний кожан 6, 107, 112, V111, 35
Полевая мышь 13, 294, 295, XXXI, 87
Полевка арчевая 14, 319, 329, XXXV11,
104
— афганская 14, 319, 328, XXXV11, 104
— большеухая 14, 335, 337, XXXV111, 107
— лемминговидная XXXV111
— Брандта 14, 328, XXXV11, 104
— длиннохвостая 14, XXXVI
— гоби-алтайская 14, 335, 337, 107
— китайская 14; 328, 329, XXXV11, 104
— красная 14, 332, 334, XXXV111, 106
— красносерая 14, 332, XXXV111, 106
— кустарниковая 14, 331, XXXV11, 105
- лесная 14, XXXV111
— Мидендорфа 14, 323, 326, XXXVI, 102
— общественная 14, 323, 327, XXXVI1, 104
— обыкновенная 14, 323, 325, XXXVI, 103
— закаспийская XXXVI
— плоскочерепная 14, 335, XXXV111, 107
— Прометеева 14, 341, XL, 109
— Роберта 14, 320, XXXVI, 100
— рыжая 14, 332, 333, XXXV111, 106
— сахалинская 322, 324, 101
— северосибирская 14, 323, 327, XXXVI,
192
— серебристая 14, 335, 336, XXXV111, 107
— снежная 14, 320, 321, XXXVI. 100
— темная 14, 322, 325, XXXVI, 10*
— узкочерепная 14, 318, 330, XXXV11, 105
— Шелковникова 105 (карта)
— экономка 14, 322, 323, XXXVI, 101
— большая 14, 323, 324, XXXVI, 101
— японская 323, 325, 101
Полевки каменные 14, 315, 335. XXXV111,
107
— лесные 14, 315, 331, ХХХ111, 106
— Прометеевы 14, 316, 341, XL, 109
— серые 14, 314, 318, XXXVI—XXXV11
Полевок, подсемейство 14, 291, 314,
XXXV___XL
Полорогие 10, 212, 221, XXI—XXII, 63—
65
Полосатая гиена 7, 133, X, 42
Полосатик малый 9, 201, XV111
Полосатики 9, 200, XVII1, 57
Полосатиковые 9, 199, 200, XV111, 57
Полосатый дельфин 9, 187, 188, XVII
Полосатый продельфин 9, 186
Полуденная песчанка 13, 301, 303, ХХХ11,
93
Предкавказский хомяк 13, 308, 309,
ХХХ111, 96
Продельфин полосатый 9, 186
Продельфины 9, 182, 186
Прометеева полевка 14, 341, XL, 109
Прометеевы полевки 14, 316, 341, XL,
109
Прудовая ночница 6, 90, 93, VI, 20
Путорак пегий 6, 78, IV, 16
Путораки 6, 58, 78, IV, 16
Пятипалые тушканчики 12, 277, XXIX,
78—80
Пятипалые карликовые тушканчики 12,
286, XXX, 81
Пятипалый карликовый тушканчик 13,286,
XXX, 81
Пятипалых тушканчиков, подсемейство 12,
276, 277, XXXIX
Пятнистый олень 10, 216, XX, 59
Ремнезуб атлантический 9, 196, 197
— командорский 9, 196, XVII
Ремнезубы 9, 194, 196, XVII
Росомаха 8, 148, XII, 46
Росомахи 8, 143, 147, XII, 46
Рукокрылые 6, 34, 79, V—V111, 17—36
Рыжая алтайская мышовка 13, 288, 289,
84
Рыжая вечерница 6, 102, VII, 29
Рыжая полевка 14, 332, 333, XXXV111,
106
Рыжеватая пищуха 11, 241, 243, ХХ111,
66
Рысь 7, 139, 141, XII, 45
Садовая соня 12, 274, XXV111, 76
Садовые сони 12, 271, 274, XXV111, 76
Сайга 10, 225, XXI, 63
Сайги 10, 223, 225, XXI, 63
Сахалинская полевка 322, 324, 101
Светлый хорек 8, 155, 159, XIV, 51
Свиные 10, 210, XIX
Свиньи 10, XIX, 58
Свиньи белокрылые 9, 184, 192, XVII
— бесперые 9, ?84, 192
— морские 9, 1ЬЗ, 191
Свинья белокрылая 9, 192 XVII
— бесперая 9. 192
— морская 9, 191. XVII
Североспбирская полевка 14. 323, 327,
XXXVI, 102
Северная пищуха 11, 239, 241, XXIII, 66
Северные олени 10, 214, 220, XX, 62
Северный ксжанок 6, 106. 109, VII, 33
Северный олень 10, 220, XX, 62
Северный плавун 9, 195, XV11
Сейвал 9, 201, 202, XV111, 57
Селевиниевые 12, 250, 275, XXV111, 77
Селевинии 12, 275, XXV111, 77
Селевиния 12, 275, XXV111. 77
Серая алтайская мышовка 13, 288, 289,
84
Серая белозубка 6, 72, 76
Серебристая полевка 14, 335, 336,
XXXV111, 107
Серна 10, 226, XXI, 63
Серны 10, 224, 226, XXI, 63
Серокитовые 9, 199, XV111, 57
Серые дельфины 9, 183, 190
Серые киты 9, 199, XV111, 57
Серые полевки 14, 314, 318, XXXVI—
XXXVII
Серый дельфин 9, 190
Серый кит 9, 199, XVIII, 57
Серый суслик 11, 261, 264, XXVI, 71
Серый хомячок 13, 311, 312, XXXIV, 98
Сибирский лемминг 14, 338, XXXIX, 108
Сивуч 8, 168, XV, 54
Сивучи 8, 167, 168, XV, 54
366
Синий кит 9, 201, 203, XV111
Складчатогуб широкоухий 7, 115, VI11, 36
Складчатогубы 7, 115, VI11, 36
Слепушонка афганская 14, 324, 343, XL,
109
— обыкновенная 14, 342, XL, 109
Слепушонки 14, 316, 341, XL, 109
Слепыш гигантский 14, 345, 346, XL, ПО
— малый 14, 345, XL, ПО
— обыкновенный 14, 345, 346, XL, ПО
Слепыши 14, 345, XL, ПО
Слепышовые 14, 253, 344, XL, ПО
Снежная полевка 14, 320, 321, XXXVI,
100
Собака енотовидная 7, 128, X, 40
Собаки 7, 120, 121, IX, 37
Собаки енотовидные 7, 121, 128, X, 40
Собачьи 7, 118, 120, IX—X, 37—40
Соболь 8, 150, 151, Х111, 47
Солонгой 8, 155, 158, XIV, 50
Соневые 12, 250, 271, XXV111, 74—77
Сони лесные 12, 271, 273, XXV111, 75
— мышевидные 12, 272, 274, XXVI11, 77
— орешниковые 12, 272, 274, XXV111, 77
— полчки 12, 271, 272, XXV111, 74
— садовые 12, 271, 274, XXV111, 76
Соня лесная 12, 273, XXV111, 75
— мышевидная 12, 275, XXVI11, 77
— орешниковая 12, 274, XXV111, 77
— полчок 12, 272, XXV111, 74
— садовая 12, 274, XXVI11, 76
Средняя бурозубка 5, 60, 63, 111. 10
Средняя могера 5, 53, 54, 11, 6
Средиземный нетопырь 6, 105, 108, VII, 32
Средиземный подковонос 6, 84, 86, V, 17
Степная кошка 7, 138, 139, XI, 43
Степная мышовка 13, 287, 288, XXX, 84
Степная пеструшка 14, 340, XXXIX, 109
Степная пищуха 11, 240, 242, ХХ111, 66
Стрелоух белобрюхий 6, 113, V111, 36
Стрелоухи 6, 89, 113, VI11, 36
Сурок длиннохвостый 11, 258, XXV, 69
— Мензбира 11, 258, 260, XXV, 69
— обыкновенный 11, 258, 259, XXV, 69
Сурки 11, 255, 257, XXV, 69
Суслик большой 11, 261, 262, XXVI, 70
— длиннохвостый 12, 261, 264, XXVI, 72
— желтый 11, 261, XXVI, 70
— крапчатый 11, 261, 262, XXVI, 70
— малый 11, 261, 263, XXVI, 71
— серый 11, 261, 264, XXVI, 71
— тонкопалый 12, 265, XXVI1, 72
Суслики 11, 256, 260, XXVI, 70—72
— тонкопалые 12, 257, 265, XXVI1, 72
Темная полевка 14, 322, 325, XXXVI, 102
Темнозубая бурозубка 5, 60, 63, 111, 9
Тигр 7. 135. 136, XI, 43
Толсторогий баран 11, 230, 233, XXII, 65
Толстохвостые тушканчики 12, 277, 281,
XXIX, 81
Толстохвостый тушканчик 12, 282, XXIX,
81
Тонкопалые суслики 12, 257, 265, XXVII,
72
Трехпалые карликовые тушканчики 12,
286, XXX, 81
Трехпалый карликовый тушканчик 13,
286, XXX, 81
Трехпалых тушканчиков, подсемейство
276, 283
Трехцветная ночница 6, 91, 95, VI, 23
Трубконос большой 7, 114, V111, 35
— малый 7, 114, V111, 35
Трубконосы 7, 89, 113, VI11, 35
Тур дагестанский 11, 228, 230, XXI, 64
— кавказский 10, 228, 229, XXI, 64
Туркестанская крыса 13, 298, 299, XXXI,
91
Туркменский тушканчик 12, 285, XXX, 82
Тушканчик Бобринского 12, 278, 281,
XXIX, 80
— большой 12, 278, 279, XXIX, 78
— Виноградова 12, 282, 81
— горный 12, 278, 280, XXIX, 79
— гребнепалый 12, 284, XXX, 83
— Житкова 12, 282, XXIX, 81
— карликовый пятипалый 13, 286, XXX,
81
— карликовый трехпалый 13, 286, XXX,
81
— Лихтенштейна 12, 285, XXX, 82
— малый 12, 278, 280, XXIX, 80
— мохноногий 12, 284, XXX, 83
— прыгун 12, 278, 279, XXIX, 78
— Северцова 12, 278, 279, XXIX, 79
— толстохвостый 12, 282, XXIX, 81
— туркменский 12, 285, XXX, 82
Тушканчики гребнепалые 12, 283, 284,
XXX 83
— мохноногие 12, 283, 284, XXX, 83
— песчаные 12, 283, 285, XXX, 82
— пятипалые 12, 277, XXIX, 78—80
— пятипалые карликовые 12, 286, XXX, 81
— толстохвостые 12, 277, 281, XXIX, 81
— трехпалые 12, 286, XXX, 81
Тушканчиковые 12, 252, 275, XXIX—XXX,
78—83
Тюленевые 8, 166, 169, XVI, 55—56
Тюлени 8, 170, 171, XVI, 55—56
Тюлени-монахи 8, 170, 178, XVI
— длинномордые 8, 170, 178, XVI, 56
Тюлень длинномордый 8, 178, XVI, 56
— обыкновенный 8, 172, 174, XVI, 55
— каспийский 8, 173, 176, XVI, 55
— монах 8, 178, XVI
Узкочерепная полевка 14, 318, 330,
XXXV11, 105
Усатая ночница 6, 92, 96, VI, 24
Ушан 6, 98, VII, 26
Ушаны 6. 89, 98, VII, 26
Ушастый еж 5, 40, 42, 1, 2
Финвал 9, 201, 202, XV111, 57
Харза 8, 148, 150, XIII, 47
Хаус 7, 139, 141, XII, 45
Хищные 7, 35, 117, IX—XIV, 37—53
Хомяк крысовидный 13, 311, 313, XXXIV,
98
— обыкновенный 13, 308, XXXIII, 96
— переднеазиатский 13, 308, 309, XXXIII,
96
— предкавказский 13, 308, 309, XXXIII,
96
Хомяки пестрые 13, 307, XXXIII, 96
367
Хомяков, подсемейство 13, 290, 306,
XXXI11—XXXIV, 96—98
Хомячки 13, 307, 310, XXXIV, 97—98
— мохноногие 13, 307, 309, XXXI11, 97
— мышевидные 13, 306, 307, ХХХШ, 96
— серые 13, XXXIV
Хомячок даурский 13, 311, XXXIV, 98
— джунгарский 13, 309, 310, ХХХШ, 97
— длиннохвостый 13, 311, 312, XXXIV, 98
— мышевидный 13, 307, ХХХШ, 96
— Роборовского 13, 310, ХХХШ, 97
— серый 13, 311, 312, XXXIV, 98
— Эверсмана 13, 310, 311, XXXIV, 97
Хорек светлый 8, 155, 159, XIV, 51
— черный 8, 155, 160, XIV, 51
Хорьки 8, 145, 153, XIV, 51
Хохлач 8, 179, XVI, 56
Хохлачи 8, 170, 179, XVI, 56
Центральноазиатский козел (горный) 10,
228, 229, XXI, 64
Цокор обыкновенный 14, 344, XL, ПО
Цокоров, подсемейство 14, 291, 343, XL,
ПО
Цокоры 14, 344, XL, ПО
Черная косатка 9, 183, 189
Черные косатки 9, 183, 189
Черная крыса 13, 298, XXXI, 91
Черные медведи 7, 130, X, 41
Черный медведь 7, 130, X, 41
Черный хорек 8, 155, 160, XIV, 51
Шакал 7, 121, 122, IX, 37
Широкоухий складчатогуб 7, 115, VI11, 36
Широкоушка азиатская 6, 100, VII, 27
Широкоушки 6, 89, 100, VII, 27
Экономка полевка 14, 322, 323, XXXVI,
101
Южные киты 10, 204, XVIII
Южный кит 10, 204, XVII1
Южный подковонос 6, 84, 86, V, 18
Як домашний 11, 235, ХХП
Яки 11, 234, 235
Японская полевка 323, 325, 101
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
ЛАТИНСКИХ НАЗВАНИЙ МЛЕКОПИТАЮЩИХ ФАУНЫ СССР
Обычными цифрами указаны страницы списка видов, определительной таблицы
и заглавия текста по данной форме или группе; римские цифры означают номер таб-
лицы в атласе цветных рисунков и цифры, выделенные жирным шрифтом,— номер
карты с ареалом данного вида.
Acinonyx 7, 134, XI, 42
— jubatus 7, 135, XI, 42
-----venaticus 135
Alactagulus 12, 277, 281, XXIX, 81
— pygmaeus 12, 281, XXIX, 81
-----aralensis 281
-----dinniki 281
---------tanaiticus 281
-----pygmaeus 281
-----turcomanus 281
Alces 10, 214, 219, XX, 62
— alces 10, 219, XX, 62
----- alces 219
----- bedfordi 220
-----pfizenmajeri 219
Allactaga 12, 277, XXIX, 78—80
— bobrinskii 12, 278, 281, XXIX, 80
— elater 12, 278, 280, XXIX, 80
-----caucasicus 280
---------aralichensrs 280
-----elater 280
---------dzungariae 280
---------vinogradovi 280
-----indica 280
-----kizljaricus 281
— major 12, 278, 279, XXIX, 78
----- fuscus 279
-----jaculus 279
-----major 279
---------intermedia 279
-----spiculum 279
-----vexillarius 279
— severtzovi 12, 278, 279, XXIX, 79
— sibirica 12, 278, 279, XXIX, 78
-----altorum 280
-----dementievi 280
-----saltator 280
-----sibirica 280
-----suschkini 280
-------riickbeili 280
— williamsi 12, 278, 280, XXIX, 79
Allactaginae 12, 276, 277, XXIX
Allocricetulus 13, 311, XXXIV
Alopex 7, 127, IX
Alticola 14, 315, 335, XXXVIII, 107
Alticola barakshin 14, 335, 337, 107
— macrotis 14, 335, 337, XXXV111, 107
-----lemminus 14, 337, XXXV111
-----macrotis 337
-----vinogradovi 337
— roylei 14, 335, 336, XXXVIII, 107
-----argentatus 336
----- leucurus 336
-----olchonensis 337
----- rosanovi 336
-----sanrica 336
-----tuvinicus 336
— strelzovi 14, 335, XXXVIII, 107
----- depressus 336
-----desertorum 336
-----strelzovi 336
Apodemus 13, 292, 294, XXXI, 87—90
— agrarius 13, 294, 295; XXXI, 87
-----agrarius 295
-----mantchuricus 295
---------chevrieri 295
---------ningpoensis 295
— flavicollis 13, 294, 296, XXXI, 89
-----flavicollis 297
-----samariensis 287
— mystacinus 13, 294, 297, XXXI, 89
— speciosus 13, 294, 295, XXXI, 88
-----major 295
---------majusculus 296
---------tscherga 296
----- praetor 295
— sylvaticus 13, 294, 296, XXXI, 90
-----chorassanicus 296
---------arianus 296
-----ciscaucasicus 296
-----pallipes 296
-----sylvaticus 296
---------charkovensis 296
—.-------vohlinensis 296
-----uralensis 296
Artiodactyla 10, 35, 209, XIX—XXII, 58—
65
Arvicola 14, 314, 316, XXXV, 100
— terrestris 14, 317, XXXV, 100
-----meridionalis 317
24
H. А. Бобринский и др.
369
Arvicola terrestris pallasi 317
-----persicus 318
-----scherman 318
-----scithicus 317
-----terrestris 317
-----variabil'is 317
Balaena 10, 204, 205, XVI11
— mysticetus 10, 205, XVI11
Balaenidae 10, 199, 204, XVI11
Balaenoptera 9, 200, XVI11, 57
— acutirostrata 9, 201, XVI11
— borealis 9, 201, 202, XVI11, 57
— musculus 9, 201, 203, XVI11
-----intermedia 203
-----musculus 203
— physalis 9, 201, 202, XVI11, 57
-----physalis 203
----- quoyi 203
Balaenopteridae 9, 199, 200, XVI11, 57
Barbastella 6, 89, 100, VI1, 27
— barbastellus 6, 100, VI1, 27
— leucomelas 6, 100, VI1, 27
Berardius 9,. 194, 195, XV11
— bairdi 9, 195, XVI1
Bison 11, 235
Blanfordimys 14, 319, 327, XXXV11
Bos 11, 234, 235, XXI1, 6
— bonasus 11, 235, XXI1, 65
— grunniens 11, XXI1
Bovidae 10, 212, 221, XXI—XXI1, 63—65
Bovinae 11, 234, XX11
Buballus 11, 235, XXI1
— buballus 11, 235, XXI1
Camelus 10, XIX
— bactrianus 10, 212, XIX
— dromedarius 10, 212, XIX
Calomyscus 13, 306, 307, XXXI11, 96
— bailwardi 13, 307, XXXI11, 96
-----bailwardi 307
-----mystax 307
Callorhinus 8, 167, XV, 54
— ursinus 8, 167, XV, 54
Canidae 7, 118, 120, IX—X, 37—40
Canis 7, 120, 121, IX, 37
— aureus 7, 121, 122, IX, 37
----- aureus 123
-----moreoticus 123
— lupus 7, 121, IX, 37
-----albus 122
-----altaicus 122
----- chanco 122
-----cubanensis 122
-----desertorum 122
----- lupus 122
--------campestris 122
Capra 10, 225, 226, XXI, 64
— aegagrus 10, 226, 229, XXI, 64
----- aegagrus 229
-----turcmenica 229
— caucasica 10, 228, 229, XXI, 64
— cylindricornis 10, 228, 230, XXI, 64
— falconeri 10, 226, 228, XXI, 64
-----heptneri 228
— sibirica 10, 228, 229, XXI, 64
-----alaiana 229
-----sibirica 229
Capreolus 10, 214, 218, XX, 61
— capreolus 10, 218, XX, 61
----- bedfordi 219
----- capreolus 218
-----armenius 218
-----caucasicus 219
-----pygargus 218
Caprinae 10, 222, 223, XXI
Capromyidae 11, 249, 255, XXV, 68
Caracal 7, 141, XI1, 45
Cardiocraniinae 12, 276, 285, XXX
Cardiocranius 12, 286, XXX, 81
— paradoxus 13, 286, XXX, 81
Carnivora 7, 35, 117, IX—XIV, 37—53
Castor 11, 254, XXV, 68
— fiber 11, 254, XXV, 68
-----fiber 254
-----pohlei 254
Castoridae 11, 249, 254, XXV, 68
Cervidae 10, 212, XIX—XX, 59—62
Cervinae 10, 213, 214, XX
Cervus 10, 214, XX, 59—60
— dama 10, 215, 216, XX, 59
— elaphus 10, 216, XX, 60
-----bactrianus 217
-----brauneri 217
-----elaphus 217
-----maral 217
-----sibiricus 218
-----songaricus 218
-----xanthopygos 218
— nippon 10, 216, XX, 59
Cetacea 9, 35, 180, XVI1—XVI11, 57
Charronia 8, 150, XI11
Chionomys 14, 318, 320, XXXV1
Chiroptera 6, 34, 79, V—VI11, 17—36
Citellus 11, 256, 260, XXVI, 70—72
— citellus 11, 261, 264, XXVI, 71
-----citellus 264
-----dauricus 264
-----ralli 264
-----relictus 264
-----xanthoprymnus 264
— major 11, 261, 262, XXVI, 70
-----argyropuloi 262
-----brevicauda 262
-----intermedius 262
-----iliensis 262
----- carruthersi 262
-----erythrogenus 262
-----major 262
-----ungae 262
— maximus 11, 261, XXVI, 70
-----maximus 261
-----orlovi 261
-----oxianus 261
— pygmaeus 11, 261, 263, XXVI, 71
-----boehmii 264
-----brauneri 264
-----ellermani 264
-----— pallidus 264
-----orlovi 264
-----kalabuchovi 264
-----mugosaricus 263
-----musicus 264
-----planicola 263
-----pygmaeus 263
-----arenicola 263
-----nerbicola 263
Citellus citellus pygmaeus kasakstanicus 263
-----nicolskii 263
---------ralli 263
-----satunini 263
-----septentrionalis 263
— suslicus 11, 261, 262, XXVI, 70
-----guttatus 262
-----suslica 262
---------averini 262
---------meridio occidentalis 262
— undulatus 12, 261, 264, XXVI, 72
-----eversmanni 265
-----intercedens 265
-----jacutensis 265
-----janensis 265
-----leucostictus 265
-----menzbieri 265
-----stejnegeri 265
-----stramineus 265
-----undulatus 265
Clethrionomys 14, 315, 331, XXVI11, 106
— glareolus 14, 332, 333, XXXV111, 106
----- frater 334
-----glareolus 334
-----istericus 334
-----ponticus 334
-----saianicus 334
---------sibiricus 334
-----suecicus 334
---------devius 334
— microtinus 14, XXVI11
— rufocanus 14, 332, XXXV111, 106
-----ircutensis 333
-----rufocanus 333
-----smithi 333
-----wosnessenskii 333
— rutilus 14, 332, 334, XXXVI11, 106
-----amurensis 335
-----jacutensis 334
-----jochelsoni 334
-----lategriseus 335
-----rossicus 334
-----rutilus 334
-----vinogradovi 334
Cricetinae 13, 290, 306, XXXI11—XXXIV,
96—98
Cricetulus 13, 307, 310, XXXIV, 97—98
— barabensis 13, 311, XXXIV, 98
-----barabensis 312
-----ferrugineus 312
-----fumatus 312
-----griseus 312
— eversmanni 13, 310, 311, XXXIV, 97
-----beljaevi 311
-----eversmanni 311
— longicaudatus 13, 311, 312, XXXIV, 98-
-----kozhanchikovi 312
— migratorius 13, 311, 312, XXXIV, 98
-----arenarius 313
-----bellicosus 313
-----caesius 313
---------griseus 313
---------cinereus 313
-----coerulescens 313
-----elisarjavi 313
-----fulvus 313
-----isabellinus 313
-----migratorius 313
Cricetulus migratorius ognevi 313
----- phaeus 313
--------neglectus 313
—-------zviersombi 313
-----pulener 3i3
----- vernula 313
— triton 13, 311, 313, XXXIV, 98
Cricetus 13, 307, XXXI11, 96
— auratus 13, 308, 309, XXXI11, 96
-----brandti 309
--------koenigi 309
— cricetus 13, 308, XXXI11, 96
— raddei 13, 308, 309, XXXI11, 96
-----nigriculus 309
-----raddei 309
--------avaricus 309
Crocidura 6, 58, 69, IV, 14—16
— lasia 6, 72, 76
— lasiura 6, 72, 75, IV, 15
— leucodon 6, 72, 76, IX, 16
-----heptapotamica 77
----- leucodon 77
-----persica 77
-----sibirica 6, 77, IV
--------ognevi 77
— pergrisea 6, 72, 74, IV
— russula 6, 72, 74, IV, 15
-----caspica 75
--------fumigatus 75
-----giildenstaedti 75
--------longicauda 75
--------monacha 75
— suaveolens 6, 71, 73, IV, 14
-----dinniki 74
-----ilensis 74
--------hyrcana 74
-----suaveolens 74
Cuon 7, 120, 123, IX, 37
— alpinus 7, 123, IX, 37
-----alpinus 123
-----hesperius 123
Cystophora 8, 170, 179, XVI, 56
— cristata 8, 178, XVI, 56
Cystophorinae 8, 179
Dama 10, 216
Delphinapterinae 9, 192
Delphinapterus 9, 185, 192, XVI1, 57
— leucas 9, 192, XVI1, 57
-----dorofeevi 193
—• — fremani 193
----- leucas 193
Delphinidae 9, 181, XVII, 57
Delphininae 9, 185, XVI1
Delphinus 9, 182, 185, XVI1, 57
— delphis 9, 185, XVI1, 57
-----bairdi 185
-----delphis 185
-----ponticus 185
Desmana 5, 44, 11, 4
— moschata 5, 44, II, 4
Desmanidae 5, 37, 43, 11, 4
Dicrostonyx 14, 315, 339, XXXIX, 108
— torquatus 14, 339, XXXIX, 108
-----chionopaes 340
----- torquatus 339
-----undulatus 340
-----vinogradovi 340
Diplomesodon 6, 58, 78, IV, 16
— pulchellum 6, 78, IV, 16
24*
37J
Dipodidae 12, 252, 275, XXIX—XXX, 78—
83
Dipodinae 12, 276, 283, XXX
Dipus 12, 283, 284, XXX, 83
— sagitta 12, 284, XXX, 83
----- innae 284
----- lagopus 284
-----nogai 284
-----sagitta 284
-----zaissanensis 284
Dryomys 12, 271, 273, XXV11I, 75
— nitedula 12, 273, XXV111, 75
-----angelus 274
-----bilkjewiczi 274
-----caucasicus 273
-----dagestanicus 273
-----nitedula 273
-----ognevi 273
-----obolenskii 273
---------tanaiticus 273
-----pallidus 274
-----saxatilis 274
-----tichomirovi 273
Duplicidentata 11, 238, XX111—XXIV,
66—67
lEliomys 12, 271, 274, XXVI11, 76
— quercinus 12, 274, XXVI11, 76
----- superans 274
'Ellobius 14, 316, 341, XL, 109
— fuscocapillus 14, 342, 343, XL, 109
-----fuscocapillus 343
-----lutescens 343
— talpinus 14, 342, XL, 109
'Enhydra 8, 145, 163, XIV, 53
— lutris 8, 163, XIV, 53
Equidae 10, 206, XIX, 58
Dquus 10, 206, XIX, 58
— asinus 207
— caballus 10, 208
— hemionus 10, 207, 208. XIX, 58
-----onager 208
lErignathus 8, 170, 177, XVI, 56
— barbatus 8, 177, XVI, 56
-----barbatus 178
-----nauticus 178
Erinaceidae 5, 37, 38, I, 1—3
Erinaceus 5, 38, I, 1—3
— auritus 5, 40, 42, I, 2
-----albulus 43
---------insularis 43
---------major 43
---------persicus 43
-----auritus 42
—--------minor 42
-----calligoni 42
—--------brachyotus 43
---------turkestanicus 43
— europaeus 5, 40, I, 1
-----amurensis 42
— — co neo lor 41
---------abasgicus 41
---------roumanicus 41
— transcaucasicus 41
-----dauricus (= dauuricus) 5, 42, I, 1
----- europaeus 41
---------centralrossicus 41
---------pallidus 41
— hypomelas 40, 43, I, 3
372
Eschirchtiidae 9, 199, XV111, 57
Eschrichtius 9, 199, XXVI11, 57
— gi'bbosus 9, 199, XVI11, 57
Eubalaena 10, 204, XVI11
— glacialis 10, 204, XVI11
-----glacialis 205
-----sieboldi 205
Eumetopias 8, 167, 168, XV, 54
— jubata 8, 168, XV, 54
Felidae 7, 119, 133, XI—Xll, 42—45
Felis 7, 134, 137, XI—Xll, 43—45
— bengalensis 7, 138, 140, XI, 44
-----chinensis 140
— caracal 7, 139, 141, Xll, 45
-----michaelis 141
— chaus 7, 139, 141, Xll, 45
----- chaus 141
— libyca 7, 138, 139, XI, 43
-----libyca 140
— lynx 7, 139, 141, Xll, 45
-----isabellina 142
----- lynx 142
-----orientalis 142
-----wrangeli 142
— manul 7, 138, 140, Xll, 44
-----ferrugineus 141
-----manul 141
— margarita 7, 138, 140, XI, 44
-----thinobius 140
— silvestris 7, 138, 139, XI, 43
-----caucasica 139
-----silvestris 139
Gazella 10, 222, XXI, 63
— gutturosa 10, 222, 223, XXI, 63
— subgutturosa 10, 222, XXI, 63
-----subgutturosa 223
Gazellinae 10, 221, 222, XXI
Gerbillinae 13, 290, 300, XXXII, 93—95
Glis 12, 271, 272, XXVIII, 74
— glis 12, 272, XXVIII, 74
-----glis 273
-----orientalis 273
-----persicus 273
Globicephala 9, 183, 190, XVII
— melaena 9, 190, XVII
-----macrorhyncha 190
-----melaena 190
Grampus 9, 183, 190
— griseus 9, 190
Gulo 8, 143, 147, XII, 46
— gulo 8, 148, XII, 46
Halichoerus 8, 170, 178, XVI, 56
— gripus 8, 178, XVI, 56
Histriophoca 8, 173
Hyaena 7, 133, X, 42
— hyaena 133, X, 42
Hyaenidae 7, 118, 133, X, 42
Hyperoodon 9, 194, XVII
— ampullatus 9, 195, XVII
Hystricidae II, 248, 253, XXV, 68
Hystrix II, 253, XXV, 68
— cristata 11, 253, XXV, 68
-----hirsutirostris 254
-----satunini 254
Insectivora 5, 34, 36, I—IV, 1—16
Jaculus 12, 283, 285, XXX, 82
— lichtensteini 12, 285, XXX, 82
— turkmenicus 12, 285, XXX, 82
Kogia 9, 197, 198
— breviceps 9, 198
Lagenorhynchus 9, 182, 187
— acutus 9, 187, 188
albirostris 9, 187, 188, XVII
— obliquidens 9, 187, 188, XVII
Lagurus 14, 315, 340, XXXIX, 109
— lagurus 14, 340, XXXIX, 109
-----abacanicus 340
-----agressus 340
-----altorum 341
-----lagurus 340
— luteus 14, 340, 341, XXXIX
Lasiopodomys 14, 319, 328, XXXVII
Lemmus 14, 315, 338, XXXIX, 108
— amurensis 14, 338, XXXIX, 108
— lemmus 14, 338, 339, XXXIX, 108
— sibiricus 14, 338, XXXIX, 108
-----chrysogaster 339
-----novosibiricus 339
-----sibiricus
Lepus II, 243, 244, XXIV, 67
— brachyurus II, 244, 245, XXIV, 67
— europaeus II, 245, 247, XXIV, 67
-----caspicus 248
-----cyrensis 248
-----hybridus 248
-----tesquorum 248
-----transsylvanicus 248
-----caucasicus 248
— timidus II, 245, 246, XXIV, 67
-----begitschevi 246
-----gichiganus 246
-----kolymensis 246
-----transbaicalicus 246
-----kozhevnicovi 246
-----sibiricorum 246
-----timidus 246
— tolai II, 245, 247, XXIV, 67
— tibetanus 245, 247, 67
-----buchariensis 247
-----desertorum 247
-----lehmanni 247
-----pamirensis 247
-----quercerus 247
----- tolai -247
-----turcomanus 247
Leporidae II, 238, 243, XXIV, 67
Lissodelphis 9, 182, 187
— borealis 9, 187
Lutra 8, 145, 162, XIV, 53
— lutra 8, 145, 162, XIV, 53
----- lutra 162
-----meridi-onalis 162
-----seistanicus 162
Lutreola 8, 160, XIV
Lutrinae 8, 162, XIV
Lynx 7, 141
Mammalia 5, 31
Marmota II, 255, 257, XXV, 69
— bobak II, 258, 259, XXV, 69
-----aphanasievi 259
-----baibacina 259
--------kastschenkoi 259
--------ognevi 259
----- bobak 259
-----bungei 260
-----caliginosus 259
-----camtschatica 260
-----centralis 259
-----doppelmayeri 259
-----sibirica 259
-----tschaganensis 259
— caudata II, 258, XXV, 69
----- aurea 258
— menzbieri II, 258, 260, XXV, 69
Martes 8, 143, 148, XIII, 47
— flavigula 8, 148, 150, XIII, 47
-----aterrima 150
— foina 8, 149, 150, XIII, 47
-----foina 150
-----intermedia 150
— martes 8, 149, 150, XIII, 47
-----borealis 151
----- lorenzi 151
-----martes 151
-----ruthena 151
-----uralensis 151
— zibellina 8, 150, 151, XIII, 47
-----averini 152
-----jeniseensis 152
--------tomensis 152
-----kamtschadalica 152
-----princeps 152
-------- jacutensis 152
-------- obscura 152
--------vitimensis 152
-----sachalinensis 152
--------arsenjevi 152
--------schantaricus 152
-----sajanensis 152
-----tungussensis 152
-----— ilimpiensis 152
-----zibellina 152
Megaptera 10, 200, 203, XVIII
— novaeangliae 10, 203, XVIII
-----lalandii 204
-----novaeangliae 204
Meles 7, 143, 146, XII, 46
— meles 7, 146, XII, 46
-----amurensis 147
----- canescens 147
--------minor 147
--------severtzovi 147
-----leptorhinchus 147
--------arenarius 147
--------raddei 147
--------tianschanensis 147
-----meles 146
--------heptneri 146
--------tauricus 146
Mellivora 7, 143, 147, XII, 46
— indica 7, 147, XII, 46
Mellivorinae 7, 147, XII
Meriones 13, 300, XXXII, 93—95
— blackleri 13, 302, 305, XXXII, 95
----- bogdanovi 305
373
Meriones libycus 13, 301, 304, XXX11, 94
-----caucasicus 304
-----eversmanni 304
-----maxeratis 304
-----oxianus 304
-----sogdianus 304
— meridianus 13, 301, 303, XXX11, 93
-----dahli 304
-----karelini 304
-----massagetes 304
-----meridianus 303
-----nogaiorum 303
-----penicilliger 304
-----urianchaicus 304
-----zhitkovi 304
— persicus 13, 301, 303, XXX11, 93
-----rossicus 303
----- suschkini 303
— tamariscinus 13, 302, 305, XXX11, 95
-----ciscaucasicus 305
-----jaxartensis 306
-----kokandicus 306
-----tamariscinus 305
— unguiculatus 13, 301, 303, XXX11, 95
-----kozlovi 303
-----selenginus 303
— vinogradovi 13, 301, 304, XXX11, 94
— zarudnyi 13, 301, 305, XXX11, 95
Mesocricetus 13, 309, XXX111
Mesoplodon 9, 194, 196, XVII
— bidens 9, 196, 197
— steinegeri 9, 196, XVI1
Micromys 13, 291, 293, XXXI, 85
— minutus 13, 293, XXXI, 85
-----batarovi 294
-----minutus 294
— ____ ussuricus 294
Microtinae 14, 291, 314, XXXV—XL, 99—
100
Microtus 14, 314, 318, XXXVI—XXXV11
— afghanus 14, 328, XXXV11, 104
----- afghanus 328
-----balchanensis 328
-----bucharensis 328
— agrestis 14, 322, 325, XXXVI, 102
-----agrestis 325
---------ognevi 325
-----argyropuloi 325
-----gregarius 325
-----mongol 325
— arvalis 14, 323, 325, XXXVI, 103
-----caspicus 326
-----duplicatus 326
-----ilaeus 326
-----iphigeniae 326
— — kirgisorum 326
-----mongolicus 326
----- obscurus 326
-----rossiameridionalis 326
-----ruthenus 326
-----transcaspicus 326
-----transcaucasicus 326
— brandti 14, 328, XXXV11, 104
— gregalis 14, 318, 330, XXXV11, 105
-----brevicauda 330
-----buturlini 330
-----dolguschini 330
-----eversmanni 330
--------- castaneus 330
£74
Microtus gregalis gregalis 330
-----major 330
-----montosus 330
-----raddei 330
-----tundrea 330
----- zachvatkini 330
— hyperboreus 14, 323, 327, XXXVI,
102
— juldaschi 14, 329, XXXV11, 104
-----carruthersi 329
----- juldaschi 329
— mandarinus 14, 328, 329, XXXV11, 104
-----vinogradovi 329
— middendorffi 14, 323, 326, XXXVI, 102
-----middendorffi 327
--------tasensis 327
----- ryphaeus 327
— montebelli 323, 325, 101
— nivalis 14, 320, 321, XXXVI, 100
-----dementievi 321
-----gud • 321
-----ighesicus 321
-----loginovi 321
-----nenjukovi 321
-----trialeticus 321
-----ulpius 321
— oeconomus 14, 322, 323, XXXVI, 101
-----altaicus 324
-----arenicola 324
-----camtschaticus 324
-----dauricus 324
-----kjusjurensis 324
-----koreni 324
-----montium-caelestinum 324
-----oeconomus 324
-----petchorae 324
-----tschuktchorum 324
-----suntaricus. 324
— maximoviczi 14, 323, 324, XXXVI, 101
-----maximoviczi 325
-----pelliceus 325
— sachalinensis 322, 324, 101
— roberti 14, 320, XXXVI, 100
— socialis 14, 323, 327, XXXV11, 104
-----binominatus 327
----- gravesi 327
-----nicolajevi 327
-----irani 327
----- parvus 327
-----schidlovskii 327
-----socialis 327
— subterraneus 14, 331, XXXV11, 105
-----atratus 331
-----ciscaucasicus 331
-----daghestanicus 331
----- kaznakovi 331
-----majori 331, XXXV11
--------suramensis 331
Micrurus 14, 319, 330, XXXV11
Miniopterus 6, 89, 97, VI, 25
— schreibersi 6, 98, VI, 25
-----pallidus 98
----- schreibersi 98
Mogera 5, 49, 53, 11, 6
— robusta 5, 54, 11, 6
-----robusta 54 .
— wogura 5, 53, 54, II, 6
-----coreana 54
Molossidae 7, 82, 114, VIII, 36 z
Monachinae 8, 178
Monachus 8, 170, 178, XVI
— monachus 8, 178, XVI
Monodon 9, 185, 193, XV11
— monoceros 9, 193, XVI1
Moschinae 10, 213, XIX
Moschus 10, 213, XIX, 59
— moschiferus 10, 213, XIX, 59
-----moschiferus 213
-----parvipes 213
-----arcticus 213
-----sachalinensis 213
Muridae 13, 253, 289, XXXI—XL, 85—100
Murina 7, 89, 113, Vlll, 35
— aurata 7, 114, Vlll, 35
— leucogaster 7, 114, Vlll, 35
Murinae 13, 290, 291, XXXI, 85—92
Mils 13, 291, 292, XXXI, 86
— musculus 13, 292, XXXI, 86
-----abbotti 293
-----bactrianus 293, XXXI
-----hortulanus 293
-----manchu 293
-----musculus 293
Muscardinidae 12, 250, 271, XXVI11, 74—
77
Muscardinus 12, 272, 274, XXVI11, 77
— avellanarius 12, 274, XXVI11, 77
-----avellanarius 274
Mustela 8, 145, 153, XI11—XIV, 48—52
— altaica 8, 155, 158, XIV, 50
-----altaica 158
-----birulai 158
-----raddei 158
— erminea 8, 154, 157, XI11, 49
-----aestiva 157
--------lymani 157
--------martinoi 157
-----erminea 157
----- ferghanae 158
-----kaneii 157
--------arctica 157
--------naumovi 157
-----tobolica 157
-----transbaicalica 157
-----baturini 158
— eversmanni 8, 155, 159, XIV, 51
-----amurensis 159
-----eversmanni 159
-----michnoi 159
--------lineiventer 159
--------tuvinicus 159
-----satunini 159
-----talassicus 159
— lutreola 8, 155, 160, XIV, 52
-----cylipena 161
-----lutreola 161
-----novikovi 161
----- turovi 161
— nivalis 8, 154, 156, XI11, 48
-----caucasica 156
-----dinniki 156
--------boccamela 156
-----nivalis 156
-----pallida 157
-----pygmaeus 156
-----stoliczkana 156
--------heptneri 156
-----vulgaris 156
Mustela putorius 8, 155, 160, XIV, 51
----- aurea 160
-----putorius 160
— sibirica 8, 155, 158, XIV, 50
-----itatsi 159
-----manchurica 159
-----sibirica 159
— vison 8, 155, 161, XIV, 52
-----altaica 161
-----tatarica 161
Mustelidae 7, 120, 142, Xll—XIV, 46—53
Mustelinae 8, 147, XI11—XIV
Myocastor 11, 255, XXV, 68
— coypus 11, 255, XXV, 68
Myocastoridae 11
Myomimus 12, 272, 274, XXVI11, 77
— personatus 12, 275, XXVI11, 77
Myopus 14, 315, 337, XXXIX, 108
— schisticolor 14, 337, XXXIX, 108
Myospalacinae 14, 291, 343, XL, J10
Myospalax 14, 344, XL, 110
— myospalax 14, 344, XL, 110
-----aspalax 344, XL
-----myospalax 344, XL
-----psilurus 344, XL
Myotis 6, 89, 90, VI, 20—24
— bechsteini 6, 90, 92, VI, 20
— blythi 6, 90, 92, VI, 21
-----blythi 93
--------omari 93
—--------oxygnathus 93
— capaccinii 6, 91, 94, VI, 22
-----capaccinii 94
-----macrodactylus 94
— dasycneme 6, 90, 93, VI, 20
--------major 94
— daubentoni 6, 91, 94, VI, 22
— emarginatus 6, 91, 95, VI, 23
-----desertorum 95
-----emarginatus 95
----- saturatus 95
— frater 6, 92, 97, VI, 23
-----bucharensis 97
-----longicaudatus 97
— muricola 6, 91, 95, VI, 24
-----ikonnikovi 96
— myotis 6, 90, 93, VI, 21
— mystacinus 6, 92, 96, VI, 24
-----mystacinus 96
--------brandti 96
--------gracilis 96
-----pamirensis 96
-----przewalskii 96
-----sogdianus 96
— nattereri 6, 91, 94, VI, 23
-----amurensis 95
----- araxenus 95
-----bombinus 95
-----nattereri 95
-----tschuliensis 95
Mystacoceti 9, 181, 199, XVIII, 57
Nemorhaedus 10, 224, 225, XXI, 63
— goral 10, 225, XXI, 63
Neomeris 9, 184, 192
— phocaenoides 9, 192
Neomys 5, 57, 67, III, 13
— anomalus 5, 67, III, 13
-----anomalus 68
375
Neomys anomalus milleri 68
— fodiens 5, 67, 68, 11.1, 13
-----fodiens 68
-----dagestanicus 68
-----orientis 68
Nesokia 13, 292, 299, XXXI, 92
— indica 13, 300, XXXI, 92
-----huttoni 300
----- satunini 300
Nonruminantia 10, 209, 210, XIX
Nyctalus 6, 89, 101, VII, 28—30
— aviator 6, 101
— lasiopterus 6, 101, 102, VII, 28
— leisleri 6, 102, 103, VII, 30
— noctula 6, 102, VII, 29
-----meklenburzevi 102
-----princeps 102
Nyctereutes 7, 121, 128, X, 40
— procyonoides 7, 128, X, 40
Ochotona 11, 239, XXIII, 66
— alpina 11, 240, 241, XXIII, 66
-----alpina 242
-----cinereofusca 242
--------scorodumovi 242
-----nitida 242
----- svatoshi 242
— daurica 11, 241, 242, XXIII, 66
-----altaica 243
-----daurica 243
— hyperborea 11, 239, 241, XXIII, 66
-----cinereoflava 241
-----collaris 241
-----ferruginea 241
-----hiperborea 241
-----mantchurica 241
-----princeps 241
-----uralensis 241
— pallasi 11, 240, 242, XXIII, 66
----- opaca 242
-----sushkini 242
— pusilia 11, 240, 242, XXIII, 66
-----angustifrons 242
-----pusilia 242
— roylei 11, 241, 243, XXIII, 66
----- sacana 243
— rufescens 11, 241, 243, XXIII, 66
-----regin a 243
— rutila 11, 240, 242, XXIII, 66
Ochotonidae 11, 238, 239, XXIII, 66
Odobenidae 8, 166, 168, XV, 54
Odobenus 8, 169, XV, 54
— rosmarus 8, 169, XV, 54
----- divergens 169
— rosmarus 8, 169, XV, 54
Odontoceti 9, 181, XVII—XVI11, 57
Ondatra 14, 314, 316, XXXV, 99
— zibethicus 14, 316, XXXV, 99
Orcinus 9, 183, 189, XVII
— orca 9, 189, XVII
Oryctolagus 11, 243, XXIV, 67
— cuniculus 11, 244, XXIV, 67
Otariidae 8, 165, 166, XV, 54
Otonycteris 6, 89, 113, VIII, 36
— hemprichi 6, 113, VIII, 36
Ovis II, 225, 230, XXII, 65
— ammon II, 230, XXII, 65
-----ammon 232
-----bocharensis 233
Ovis ammon coIlium 233
-----cycloceros 233
-----gmelini 233
-----karelini 233
-----musimon 11, XXI1
-----nigrimontana 233
-----orientalis 11, XXI1
-----polii 232
-----severtzovi 233
— canadensis (-nivicola) 230, 234, XXII,
65
— nivicola (-canadensis) 230, 234, XXI1,
65
•----alleni 234
-----borealis 234
-----nivicola 234
Pachyceros 11, 234, XXII
Pagophoca 8, 172
Panthera 7, 134, 135, XI, 42—43
— pardus 7, 135, 136, XI, 42
-----orientalis 136
-----tulliana 136
--------ciscaucasica 136
— tigris 7, 135, 136, XI, 43
-----coreensis 137
-----longipilis 137
-----virgata 137
— uncia 7, 135, 136, XI, 42
Paradipus 12, 283, 284, XXX, 83
— ctenodactylus 12, 284,-XXX, 83
Perissodactyla 10, 35, 206, IX, 58
Phaiomys 14, 319, 329, XXXVII
Phoca 8, 171, XVI, 55-56
— caspica 8, 173, 176, XVI, 55
— fasciata 8, 172, 174, XVI, 56
— groenlandica 8, 171, 173, XVI, 56
-----oceanica .174
— hispida 8, 173, 175, XVI, 55
-----botnica 176
--------ladogensis 176
-----ochotensis 176
-----pomororum 176
— sibirica 8, 172, 176, XVI, 55
— vitulina 8, 172, 174, XVI, 55
----- largha 175
-----vitulina 175
Phocaena 9, 183, 191, XVII
— phocaena 9, 191, XVII
----- phocaena 191
-----relicta 191
-----vomerina 191
Phocaenoides 9, 184, 192, XVII
— dalli 9, 192, XVII
-----dalli 192
-----truei 192
Phocidae 8, 166, 169, XVI, 55—56
Phocinae 8
Phodopus 13, 307, 309, XXXIII, 97
— roborovskii 13, 310, XXXI11, 97
— sungorus 13, 309, 310, XXXIII, 97
-----campbelli 310
----- sungorus 310
Phoephagus II, 234, 235
— mutus II, 235
Physeter 9, 198, XVIII, 57
— catodon 9, 198, XVIII, 57
Physeteridae 9, 181, 197, XVIII, 57
Pinnipedia 8, 35, 163, XV—XVI, 54-56
376
Platycranius 14, 335, XXXVI11
Plecotus 6, 89, 98, VI1, 26
— auritus 6, 98, VI1, 26
—, — auritus 99
-----macrobullaris 99
-----sacrimontis 99
-----wardi 99
--------kozlovi 99
--------mordax 99
Procapra 10, 222, 223, XXI
Procyon 7, 129, X, 40
— lotor 7, 129, X, 40
Procyonidae 7, 118, 129, X, 40
Prometheomys 14, 316, 341, XL, 109
— schaposchnikovi 14, 341, XL, 109
Pseudorca 9, 183, 189
— crassidens 9., 189
Pteromyidae 12, 249, 270, XXVI11, 73
Pteromys 12, 270, XXVI11, 73
— volans 12, 270, XXVI11, 73
-----anadyrensis 27)1,
-----arsenyevi 271
-----athene 271
-----betulinus 271
-----gubari 271
----- incanus 271
-----ognevi 271
----- turovi 271
-----volans 271
Putorius 8, 159, XIV
Pygeretmus 12, 277, 281, XXIX, 81
— vinogradovi 12, 282, 81
— platyurus 12, 282, XXIX, 81
— zhitkovi 12, 282, XXIX, 81
Rangifer 10, 214, 220, XX, 62
— tarandus 10, 220, XX, 62
-----phylarchus 221
-----sibiricus 220
--------paersoni 220
----- tarandus 220
-----valentinae 221
Rattus 13, 292, 297, XXXI, 91—92
— norvegicus 13, 298, 299, XXXI, 92
-----carraco 299
-----norvegicus 299
— rattoides 13, 298, 299, XXXI, 91
--------turkestanicus 299
— rattus 13, 298, XXXI, 91
-----alexandrinus 299
-----rattus 299
-----ruthenus 299
Rhinolophidae 6, 81, 82, V, 17—19
Rhinolophus 6, 82, V, 17—19
— blasii 6, 84, 86, V, 17
— bocharicus 6, 85, 88, V, 19
— euryale 6, 84, 86, V, 18
— ferrumequinum 6, 85, 87, V, 19
-----ferrumequinum 88
-----proximus 88
— hipposideros 6, 84, 85, V, 17
-----hipposideros 86
-----midas 86
— mehelyi 6, 84, 87, V, 18
Rhombomys 13, 300, 306, XXXII, 95
— opimus 13, 306, XXXII, 95
-----famicolor 306
-----opimus 306
Rodentia 11, 35, 236, XXI11—XL, 66-
110
Ruminantia 10, 210, 212, XIX—XXII
Rupicapra 10, 224, 226, XXI, 63
— rupicapra 10, 226, XXI, 63
-----caucasica 226
Saiga 10, 223, 225, XXI, 63
— tatarica 10, 225, XXI, 63
Salpingotus 13, 286, XXX, 81
— crassicauda 13, 286, XXX, 81
Sciuridae 11, 249, 255, XXV—XXVII,
69—73
Sciurus 12, 257, 266, XXVI1, 73
— anomalus 12, 267, XXVIL 73
-----anomalus 267
— vulgaris 12, 267, XXVI1. 73
-----altaicus 269
-----baschkiricus 269
-----carpathicus 268
-----exalbidus 269
-----fedjuschini 268
-----formosovi 269
-----fusconigricans 269
-----golzmajeri 269
-----jacutensis 269
-----jenissejensis 269
-----kalbinensis 269
-----kessleri 269
-----mantchuricus 270
-----martensi 269
----- ognevi 269
-----rupestris 270
-----ukrainicus 268
-----varius 268
Selenarctos 7, 130, X, 40
— tibetanus 7, 130, X, 41
-----ussuricus 131
Selevinia 12, 275, XXVII1, 77
— betpakdalensis 12, 275, XXVIII, 77
Seleviniidae 12, 250, 275, XXVI11, 77
Sicista 13, 287, XXX, 84
- betulina 13, 287, 288, XXX, 84
— concolor 13, 287, 289, XXX, 84
-----caucasica 287, 289, 84
-----caudata 289
-----pseudonapaea 288, 289, 84
— — tianschanica 289
— napaea 13, 288, 289, 84
- subtil is 13, 287, 288, XXX, 84
-----nordmanni 288
-----severtzovi 288
-----sibirica 288
— — subtilis 288
----- vaga 288
Simplicidentata 11, 238, 248, XXV—XL
68—110
Sorex 5, 57, 58, III, 7—12
— alpinus 5, 61, 66, III, 12
-----alpinus 66
— araneus 5, 61, 64, III 11
----- araneus 65
--------schnitnikovi 65 *
--------tomensis 65
-----arcticus 65
----- asper 65
-----raddei 65
--------satunini 65
-----vir 65
377
Sorex buchariensis 5, 60, 62, 8
— caecutiens 5, 60, 63, 111, 10
-----buxtoni 64
--------baicalensis 64
--------borealis 64
-------- koreni 64
-----caecutiens 64
-----macropygmaeus 64
-------- pleskei 64
--------sibiriensis 64
----- shinto 64
— daphaenodon 5, 60, 63, 111, 9
----- daphaenodon 63
-----sanguinidens 63
----- scaloni 63
— minutissimus 5, 59, 61, 111, 7
-----abnormis 62
-----barabensis 62
-----minutissimus 61
--------karelicus 61
--------neglectus 61
-----tscherskii 61
-----tschuktschorum 62
— minutus 5, 59, 62, 111, 8
-----gmelini 62
-----gracillimus 62
--------leucogaster 62
-----minutus 62
--------heptapotamicus 62
-----portenkoi 62
-----volnuchini 62
— pacificus 5, 61, 66, 111, 12
-----mirabilis 66
— unguiculatus 5, 61, 66, 111, 12
----- vargans 66
Soricidae 5, 38, 54, 111, 7—16
Spalacidae 14, 253, 344, XL, 110
Spalax 14, 345, XL, 110
— giganteus 14, 345, 346, XL, 110
— leucodon 14, 345, XL, 110
----- leucodon 345
-----nehringi 345
— microphthalmus 14, 345, 346, XL, 110
-----arenarius 346
-----microphthalmus 346
-----zemni 346
Spermophilopsis 12, 257, 265, XXVI1, 72
— leptodactylus 12, 265, XXVI1, 72
-----bactrianus 266
-----heptopotamicus 266
-----leptodactylus 266
Stenella 9, 182, 186
— caeruleoalbus 9, 186
Stenocranius 14, 318, 330, XXXV11
Stylodipus 12, 283, XXX, 82
— telum 12, 283, XXX, 82
-----amankaragai 284
-----falzfeini 283
-----karelini 284
-----telum 284
----- turovi 284
Suidae 10, 210, XIX, 58
Suncus 6, 58, 69, IV, 14
— etrucus 6, 69, IV, 14
Sus 10, 210, XIX, 58
— scrota 10, 211, XIX, 58
-----atila 211
-----nigripes 211
----- scrota 211
Sus scrota sibiricus 211
-----ussuricus 211
Tadarida 7, 115, Vlll, 36
— teniotis 7, 115, VI1, 36
Talpa 5, 49, 11, 5—6
— altaica 5, 50, 52, 11, 6
— europaea 5, 49, 50, 11, 6
-----caucasica 52
---------romana 52
----- europaea 51
---------brauneri 51
---------obensis 51
---------transuralensis 51
---------uralensis 51
-----levantis 52
---------olimpica 52
---------transcaucasica 52
-----orientalis 52
--------- caeca 52
-----talyschensis 52
— herzegoviensis 5, 49, 50, 11, 5
-----minima 5, 50
Talpidae 5, 38, 47, 11, 5—6
Talpinae 5, 47, 11
Tamias 12, 257, 266, XXVI1, 72
— sibiricus 12, 266, XXVI1, 72
-----asiaticus 266
-----jacutensis 266
-----lineatus 266
-----orientalis 266
-----sibiricus 266
Thalassarctos 7, 130, 132, X, 41
— maritimus 7, 132, X, 41
-----marinus 133
-----maritimus 133
Tigris 7, 136
Tylopoda 10, 209, 212, XIX
Tirsiops 9, 182, 186, XVI1
— truncatus 9, 186, XVI1
-----gilli 187
-----ponticus 186
----- trancatus 186
Tscherskia 13, 313, XXXIV
Uncia 7, 136
Unsidae 7, 118, 130, X, 41
Ursus 7, 130, 131, X, 41
— arctos 7, 131, X, 41
-----arctos 132
-----beringianus 132
---------mandchuricus 132
-----caucasicus 132
-----isabellinus 132
-----syriacus 132
Vespertilio 6, 89, 103, VI1—Vlll, 31—35
— abfamus 6, 105, 108, 31
— bobrinskii 6, 106, НО, VI1, 33
— kuhli 6, 105, 108, VI1, 32
-----lepidus 109
— murinus 6, 106, 111, Vlll, 34
— nathusii 6, 104, 108, VI1, 33
— nilssoni 6, 106, 109, Vll, 31
— ognevi 6, 107, 112, Vlll, 35
— pipistrellus 6, 104, 107, VII, 31
-----bactrianus 108
-----pipistrellus 108
- savii 6, 106, 109, VII, 32
378
Vespertilio serotinus 6, 107, 112, VIII, 35
-----serotinus 112
-----turcomanus 112
— superans 6, 107, 111, VI11, 34
Vespertilionidae 6, 82, 88, VI—VI11,
20—36
Vormela 8, 145, 153, XIII, 48
— peregusna 8, 153, XIII, 48
Vulpes 7, 120, 124, IX, 38—39
— сапа 7, 124, 127, IX, 39
— corsac 7, 124, 127, IX, 39
----- corsac 127
--------scorodumovi 127
-----kalmykorum 127
-----turkmenicus 127
— lagopus 7, 125, 127, IX, 39
-----beringensis 128
--------spitzbergensis 128
----- lagopus 128
— vulpes 7, 124, 125. IX, 38
-----alpherayi 126
-----alticola 126
-----beringiana • 125
-----caucasica 126
-----daurica 126
-----diluta 126
-----dolychocrania 126
-----flavescens 126
-----jakutensis 125
----- karagan 126
----- ochroxanta 126
-----splendidissima 126
-----stepensis 126
-----tobolica 125
-----vulpes 125
---------crucigera 125
Zapodidae 13, 252, 286, XXX, 84
Ziphiidae 9, 181, 193, XVII
Ziphius 9, 194, 195, XVII
— cavirostris 9, 195, XVII
СОДЕРЖАНИЕ
Предисловие .................................................... 3
Список видов млекопитающих фауны СССР............................5
Цветные таблицы............................................... .15
Основные сведения по систематике.............................. .17
Как пользоваться определителем.................... . . 26
КЛАСС МЛЕКОПИТАЮЩИЕ.
CLASS MAMMALIA
Таблица для определения отрядов млекопитающих.................. 34
Отряд Насекомоядные. Ordo Insectivora.......................... 36
Таблица для определения семейств насекомоядных фауны СССР . . 37
Таблица для определения видов ежей фауны СССР.................. 40
Таблица для определения родов кротовых фауны СССР.............. 49
Таблица для определения видов кротов рода Talpa фауны СССР . . —
Таблица для определения видов могер, представленных в фауне СССР 53
Таблица для определения родов землероек, представленных в фауне
СССР........................................................ 57
Таблица для определения видов бурозубок фауны СССР............. 59
Таблица для определения видов кутор .......................... 67
Таблица для определения белозубок фауны СССР...................71
Отряд Рукокрылые. Ordo Chiroptera.............................79
Таблица для определения семейств летучих мышей, представленных
в фауне СССР..................................................81
Таблица для определения видов подковоносов фауны СССР...........84
Таблица для определения родов гладконосых летучих мышей, пред-
ставленных в фауне СССР..........................................88
Таблица для определения видов ночниц, представленных в фауне СССР 99
Таблица для определения видов широкоушек......................100*
Таблица для определения видов вечерниц фауны СССР..............101
Таблица для определения видов кожанов и нетопырей, представлен-
ных в фауне СССР................................................104
Таблица для определения видов трубконосов фауны СССР...........114
Отряд Хищные. Ordo Carnivora...................................117
Таблица для определения семейств отряда хищных фауны СССР . . —
Таблица для определения родов семейства собачьих фауцы СССР . . .120
Таблица для определения видов рода Canis фауны СССР . . . .121
Таблица для определения видов рода лисиц фауны СССР..........124
Таблица для определения родов медведей фауны СССР............130
Таблица для определения родов семейства кошачьих фауны СССР . .134
Таблица для определения видов рода пантер фауны СССР.........135
380
Таблица для определения видов рода кошек фауны СССР...............137
Таблица для определения родов семейства, куньих фауны СССР . . . 143
Таблица для определения видов куниц фауны СССР ... .148
Таблица для определения видов рода Mustela фауны СССР . 154
Таблица для определения семейств отряда ластоногих . . —
Отряд Ластоногие. Ordo Pinnipedia............................... .164
Таблица для определения родов котиковых вод СССР ... . 166
Таблица для определения родов тюленевых фауны СССР . .170
Таблица для определения видов собственно тюленей . . 171
Отряд Китообразные. Ordo Cetacea............................... . 180
Таблица для определения подотрядов китообразных ................. 181
Таблица для определения семейств зубатых китов, представленных в во-
дах СССР...........................................................—
Таблица для определения родов дельфиновых фауны СССР и смеж-
ных морей........................................................182
Таблица для определения видов короткоголовых дельфинов фауны
СССР.............................................................187
Таблица для определения родов клюворылых, обитающих в водах
СССР.............................................................194
Таблица для определения видов ремнезубов, которые могут водиться
в водах СССР.....................................................196
Таблица для определения родов кашалотовых . . .... 197
Таблица для определения семейств беззубых китов . .199
Таблица для определения рода полосатиковых . . 200
Таблица для определения видов полосатиков .................. .201
Таблица для определения родов гладких китов фауны СССР . . 204
Отряд Непарнокопытные. Ordo Perissodactyla........................206
Таблица для определения видов рода Equus, встречающихся в СССР 207
Отряд Парнокопытные. Ordo Artiodactyla............................209
Таблица для определения подотрядов парнокопытных фауны СССР . . —
Таблица для определения семейств жвачных фауны СССР...............212
Таблица для определения подсемейств семейства оленьих фауны СССР 213
Таблица для определения родов оленьих фауны СССР..................214
Таблица для определения видов рода настоящих оленей фауны СССР . —
Таблица для определения подсемейств полорогих копытных фауны СССР 221
Таблица для определения родов газелевых фауны СССР ... . . 222
Таблица для определения родов козлиных фауны СССР.................223
Таблица для определения диких видов горных козлов фауны СССР . . 226
Таблица для определения диких видов баранов фауны СССР .... 230
Таблица для определения родов подсемейства бычьих фауны СССР . . 234
Отряд Грызуны. Ordo Rodentia............................. . . 236
Таблица для определения подотрядов грызунов.......................238
Таблица для определения семейств подотряда двупарнорезцовых грызу-
нов фауны СССР...................................................—
Таблица для определения видов пищух фауны СССР . . 239
Таблица для определения родов заячьих фауны СССР . . 243
Таблица для определения видов зайцев фауны СССР...................244
Таблица для определения семейств подотряда однопарнорезцовых гры-
зунов фауны СССР............................................. . . 248
Таблица для определения родов беличьих фауны СССР .... . 255
Таблица для определения видов рода сурков фауны СССР . . 258
Таблица для определения видов рода сусликов фауны СССР . . 261
Таблица для определения видов белок фауны СССР . .... 267
Таблица для определения родов сонь фауны СССР.....................271
Таблица для определения подсемейств семейства тушкапчиковых
фауны СССР........................................................276
381
Таблица для определения родов подсемейства пятипалых тушканчиков
фауны СССР.......................................................277
Таблица для определения видов рода пятипалых тушканчиков фауны
СССР...............................................................—
Таблица для определения видов толстохвостых тушканчиков..........282
Таблица для определения родов трехпалых тушканчиков фауны СССР 283
Таблица для определения видов песчаных тушканчиков фауны СССР 285
Таблица для определения родов подсемейства карликовых тушканчиков 286
Таблица для определения видов мышевок фауны СССР.................287
Таблица для определения подсемейств семейства мышиных фауны
СССР......................................................... 290'
Таблица для определения родов подсемейства мышей фауны СССР . . 291
Таблица для определения видов лесных и полевых мышей фауны СССР 294
Таблица для определения видов рода настоящих крыс фауны СССР 297
Таблица для определения родов подсемейства песчанок фауны СССР 300’
Таблица для определения видов рода малых песчанок фауны СССР . . 301
Таблица для определения родов подсемейства хомяков фауны СССР . 306*
Таблица для определения видов пестрых хомяков фауны СССР . . 308-
Таблица для определения видов рода мохноногих хомячков .... 309'
Таблица для определения видов рода хомячков фауны СССР . . . ЗЮ1
Таблица для определения родов подсемейства полевок фауны СССР 314
Таблица для определения подродов рода серых полевок фауны СССР 318
Таблица для определения видов подрода снежных полевок фауны СССР 320:
Таблица для определения видов подрода серых полевок фауны СССР 321
Таблица для определения видов лесных полевок фауны СССР’ . . . 332
Таблица для определения видов каменных полевок фауны СССР . . 335
Таблица для определения видов настоящих леммингов фауны СССР 338**
Таблица для определения видов степных пеструшек фауны СССР . . 340-
Таблица для определения видов слепушонок фауны СССР .... 342
Таблица для определения видов слепышей фауны СССР.............345
Литература.......................................................347
Атлас ареалов млекопитающих СССР.................................359
Николай Алексеевич Бобринский
Борис Александрович Кузнецов
Александр Петрович Кузякин
ОПРЕДЕЛИТЕЛЬ
МЛЕКОПИТАЮЩИХ СССР
Пособие для студентов
педагогических институтов
и учителей
Редактор Е. Н. Хунцкария
Редактор карт К. Ф. Зайцева
Оформление художника
С. Я. Нодельмана
Цветные таблицы художников:
Н. Н. Кондакова и О. Ф. Хлудовой
Художественный редактор М. Л. Фран
Технический редактор Н. Н. Махова
Корректор В. А. Захарова
Сдано в набор 19/V 1964 г. Подписано к печати
7/1V 1965 г. 70X108*/1в. Печ. л. 24 (33,6) +
+ вкл. 2,5 (3,5) + карты 6 (8,4). Уч.-изд. л. 32,68+
+ вкл. 3,95. +карты 9,32. Тираж 26 тыс. экз. (Тем. пл.
1964 г. № 204). А-11669.
Заказ № 880.
Цена без переплета 1 р. 28 к., карты 48 к., пере-
плет 15 к.
Издательство «Просвещение» Государственного
комитета Совета Министров РСФСР по печати.
Москва, 3-й проезд Марьиной рощи, 41.
Ленинградская типография № 4 Главполиграфпрома
Государственного комитета Совета Министров СССР
по печати, Социалистическая, 14.