
















































































































































































































































































































Автор: Фишер Р.
Теги: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез монография генетика естественный отбор мимикрия теория эволюции
ISBN: 978-5-93972-906-2
Год: 2011




THEGENETICAL THEORY OF
NATURAL SELECTIONBYR. A. FISHER, Sc.D., F.R.S.OXFORD
AT THE CLARENDON PRESS
1930
Роналд ФишерГенетическая теория
естественного отбораПеревод с английского
JI. С. Ванаг и Е. И. ФукаловойПод редакцией Н. В. ГлотоваR&CТ>цквшис&Москва ♦ Ижевск
2011
УДК 575.17
ББК 28.046
Ф681Издание осуществлено при финансовой поддержке Российского
фонда фундаментальных исследований по проекту № 09-04-07050• физика• математика• биология• нефтегазовые
технологииФишер Р.Генетическая теория естественного отбора. — М.-Ижевск: НИЦ «Регуляр¬
ная и хаотическая динамика», Ижевский институт компьютерных исследо¬
ваний, 2011. — 304 с.В классической монографии Р. Фишера (Fisher R. А., 1890-1962), одного из со¬
здателей современной теоретической популяционной генетики и синтетической тео¬
рии эволюции, сформулирована концепция приспособленности (впервые дано ло¬
гически непротиворечивое строгое определение понятия «естественный отбор»),
предложена теория эволюции доминантности, рассмотрено соотношение мутаци¬
онного процесса и отбора в эволюции, дана интерпретация явления мимикрии,
рассмотрены популяционно-генетические следствия для динамики популяций чело¬
века. Издание включает комментарии и послесловие. Популяционно-генетические
и эволюционные работы Р. Фишера ранее на русском языке не издавались.ISBN 978-5-93972-906-2 ББК 28.046© Перевод на русский язык:Ижевский институт компьютерных исследований, 2011http://shop.rcd.ruhttp://ics.org.ruИнтернет-магазинhttp://shop.rcd.ru
ОглавлениеПредисловие viiГлава 1. Природа наследственности 1Следствия теории слитной наследственности, по Дарвину. Противо¬
речия, замеченные Дарвином. Корпускулярная наследственность. Со¬
хранение дисперсии. Теории эволюции, обусловленной мутациями.Вся ли наследственность корпускулярна? Природа и частота наблю¬
даемых мутаций.ГЛАВА 2. Фундаментальная теорема естественного отбора .... 22
Таблица выживания и таблица репродукции. Мальтузианский пара¬
метр роста популяции. Репродуктивная ценность. Генетическая со¬
ставляющая дисперсии. Естественный Отбор. Природа адаптации.
Ухудшение среды обитания. Изменения в популяции. Резюме.Глава 3. Эволюция доминантности 48Доминирование диких генов. Модификация эффектов менделевских
факторов. Модификация гетерозиготы. Особые приложения теории.
Процесс модификации. Выводы из теории эволюции доминантности.
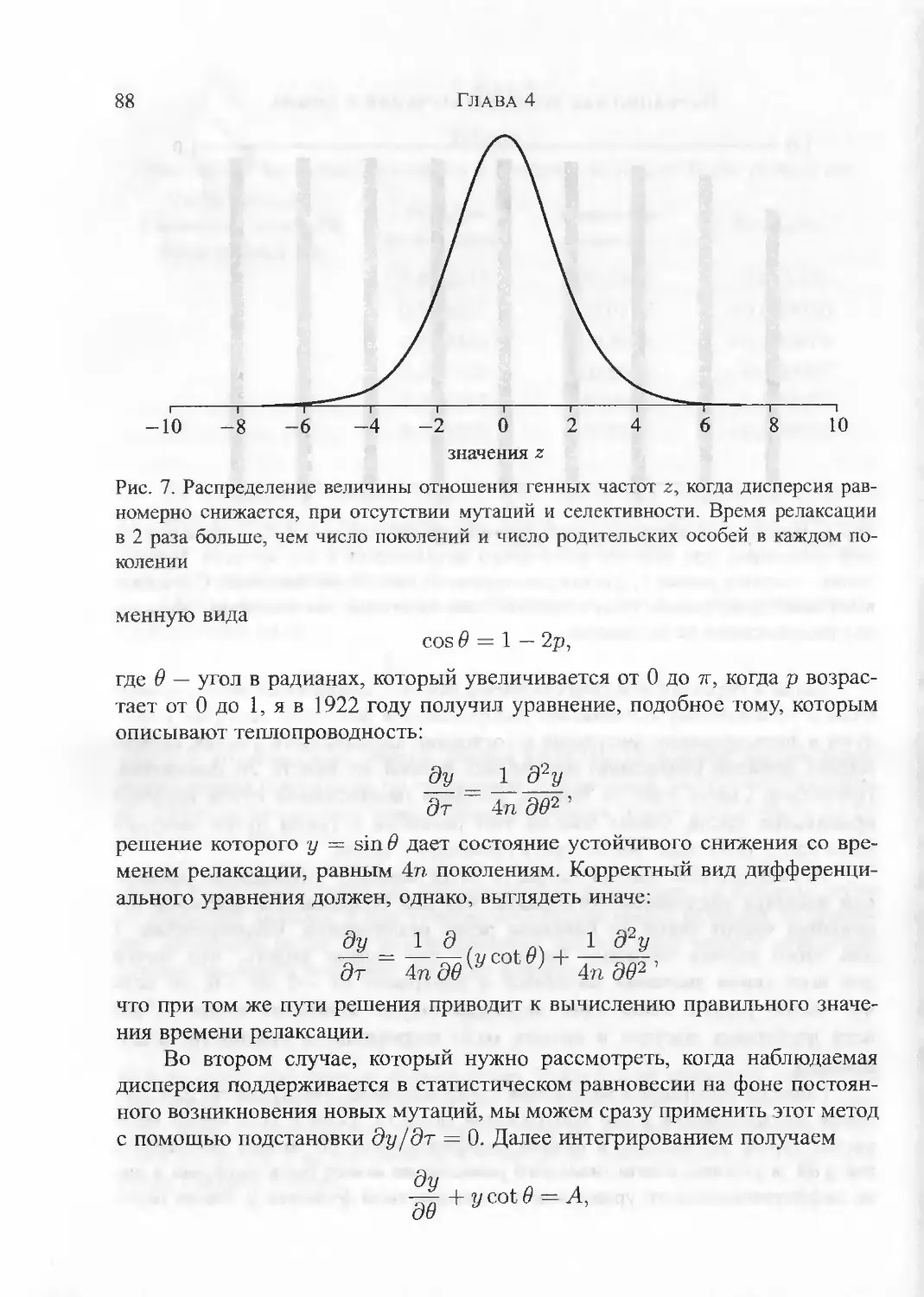
Резюме.ГЛАВА 4. Вариации как результат мутаций и отбора 69Измерение частоты гена. Вероятность сохранения отдельного гена;
связь с распределением Пуассона. Низкая частота полезных мутаций.
Единичное возникновение мутации не невероятно. Распределение от¬
ношений генных частот для факторов, определяющих дисперсию. Ма¬
лые эффекты случайного выживания. Число факторов, определяющих
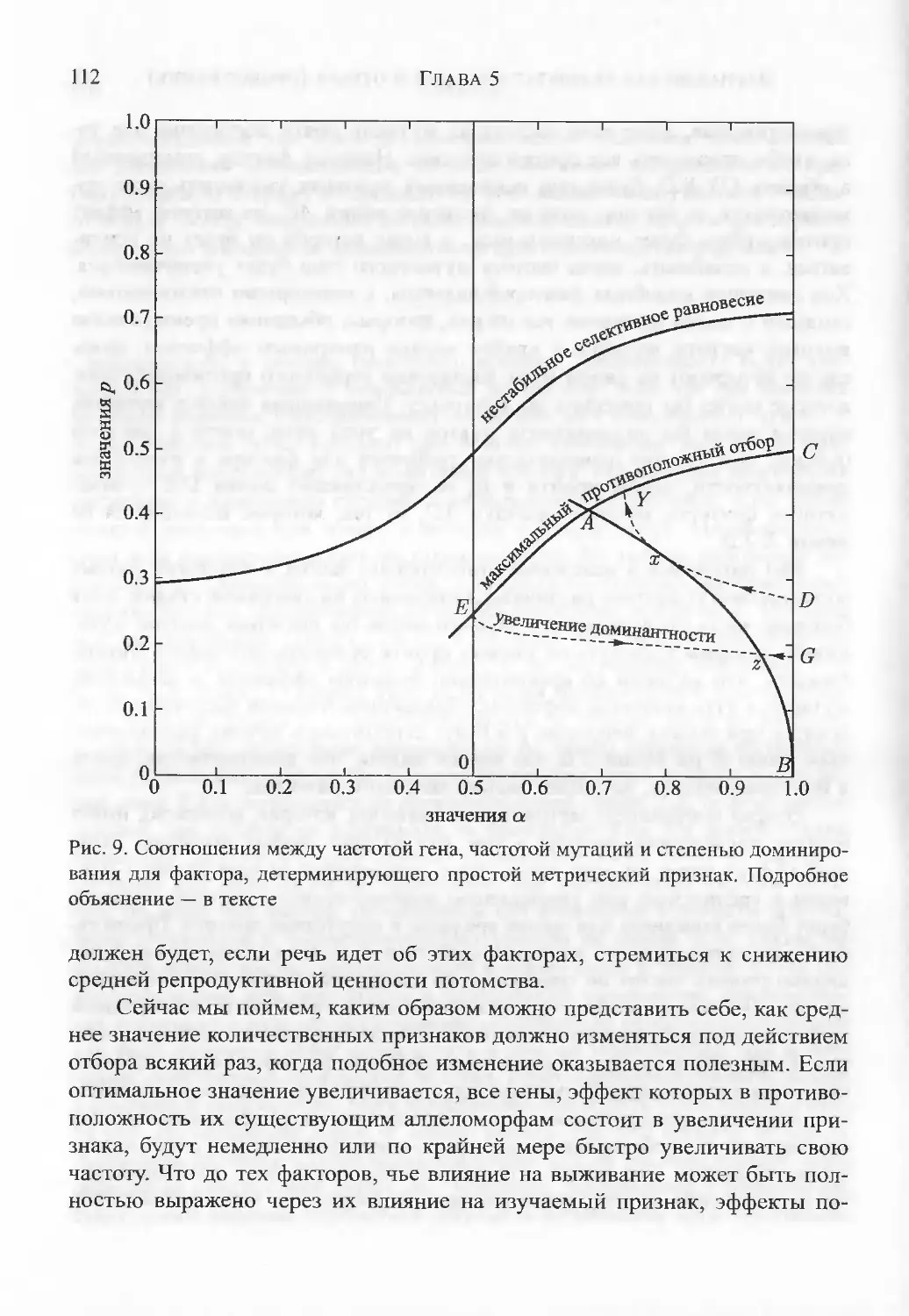
дисперсию.ГЛАВА 5. Вариации как результат мутаций и отбора (продолжение) 98Наблюдаемая связь между изменчивостью и обилием вида. Стабиль¬
ные отношения генных частот. Равновесие по двум факторам. Про¬
стые метрические признаки. Меристические признаки. Биометриче¬
ские эффекты недавнего отбора. Резюме.
11ОглавлениеГлава 6. Половое размножение и половой отбор 122Сравнение полового и бесполого размножения. Природа вида. Рас¬
щепление вида. Половое предпочтение. Половой отбор. Модифика¬
ции, ограниченные полом. Естественный Отбор и соотношение полов.
Резюме.Глава 7. Мимикрия 147Связь теории мимикрии с исходной теорией Естественного Отбора.
Теории Бейтса и Мюллера. Предполагаемые статистические ограни¬
чения теории Мюллера. Наблюдения как основа теории мимикрии.
Эволюция неприятного вкуса. Теория сальтаций. Стабильность отно¬
шения генных частот. Резюме.Глава 8. Человек и общество 171О Человеке, важность предварительных исследований. Упадок циви¬
лизаций. Социологические взгляды. Сообщества насекомых. Резюме.Глава 9. Наследование фертильности у человека 189Большая изменчивость репродуктивности у человека. Умственные
способности и моральные качества, определяющие репродуктивность.
Прямое доказательство наследования фертильности. Эволюция сове¬
сти не нарушает сознательной репродукции. Аналогии с инстинктом
животных и иммунитетом к болезням. Резюме.Глава 10. Репродуктивность и социальный класс 211Экономические и биологические аспекты классовых различий. Неточ¬
ности текущих данных. Начало исследований. Британские данные.
Положение в США. Эффекты дифференциальной фертильности. Ре¬
зюме.ГЛАВА 11. Социальный отбор по фертильности 229История теории. Бесплодие во всех классах, независимо от причины
возникновения, приводит к социальному продвижению. Отбор — ос¬
новная причина инвертированного уровня рождаемости. Упадок пра¬
вящих классов. Контраст с варварскими обществами. Героизм и выс¬
шие способности человека. Место социального класса в эволюции че¬
ловека. Аналогия с паразитизмом у муравьев. Резюме.ГЛАВА 12. Условия долговременной цивилизации 257Апология. Долговременная цивилизация не всегда непрогрессивна.
Перераспределение рождений. Социальная поддержка фертильности.
Недостатки французской системы. Проблема существования народов.
Резюме.
Оглавление iiiЛитература 267Указатель 271Комментарии (Н. В. Глотов, Л. А. Животовский, Н. В. Ившин) . . . 278Послесловие (Н. В. Глотов, Л. А. Животовский) 287
Майору Леонарду Дарвину1
с благодарностью за поддержку, которую он оказывал автору в течение
последних пятнадцати лет, обсуждая с ним многие из проблем, которымпосвящена эта книга
ПредисловиеЕстественный Отбор — это не Эволюция.2 Однако как только эти два
понятия вошли в широкое употребление, термин «теория естественного от¬
бора» стали использовать как удобное сокращение для обозначения теории
Эволюции путем Естественного Отбора, которую сформулировали Дарвин
и Уоллес. К сожалению, следствием этого стало то, что собственно теория
Естественного Отбора очень редко (если когда-либо вообще) рассматри¬
валась самостоятельно. В качестве аналогии можно привести следующий
пример из физики: законы теплопроводности в твердых средах можно вы¬
вести из принципов статистической механики, однако рассматривать стати¬
стическую механику исключительно в связи с явлениями теплопроводности
было бы серьезной ошибкой, которая, скорее всего, привела бы к полной
неразберихе. В этом случае понятно, что изучаемые частные явления по¬
чти не представляют интереса с точки зрения теории по сравнению с теми
принципами, которые позволяют прояснить их суть. Огромная значимость
эволюции для биологических наук отчасти объясняет тот факт, что теория
Естественного Отбора столь часто сводится к изучению его как фактора
эволюции; это привело к полному пренебрежению Естественным Отбором
как независимым принципом, заслуживающим научного изучения.Другие биологические теории, которые предлагались в качестве либо
вспомогательных, либо основных способов органической эволюции, ока¬
зались в несколько ином положении. Дело в том, что сторонники теории
Естественного Отбора не преминули показать, что она предполагает объяс¬
нение способов изменения органического мира, основанное только на «из¬
вестных» или независимо доказуемых причинах, и именно в этом, очевид¬
но, заключалась основная притягательность теории для Дарвина и Уоллеса.
Другие теории изменения, по собственному признанию, опираются на ги¬
потетические свойства живого, которые выводятся из самих фактов эволю¬
ции. Тем не менее, хотя эта разница часто была достаточно ясна, ее логиче¬
ская неопровержимость никак не могла быть обоснована в отсутствие спе¬
циального исследования независимо доказуемых действующих факторов,
лежащих в ее основе. Настоящая книга, несмотря на все недостатки, при¬
сущие ей как первой работе такого рода, представляет собой попытку рас¬
смотрения теории Естественного Отбора ради ее собственных достоинств.
viiiПредисловиеКогда теория была предложена впервые, самым неясным элементом
в ней был принцип наследственности. Никто из ученых или натуралистов
не мог отрицать этот принцип, однако на тот момент не существовало ни¬
какого объяснения, как именно он работает. То, что независимое исследо¬
вание Естественного Отбора теперь возможно, есть следствие того суще¬
ственного прогресса в генетике, свидетелем которого стало наше поколе¬
ние. Заслуживает внимания тот факт, что первые значимые эксперименты,
которые открыли в биологии эту область точных исследований, были про¬
ведены молодым математиком Грегором Менделем, чьи интересы в стати¬
стике распространялись и на физику, и на биологию.3 Широко известно,
что его эксперименты, к его большому огорчению, не получили широкой
известности, и мы можем предполагать, что они так и не попали на гла¬
за никому из тех, чье образование позволило бы оценить их важность.4
Не менее удивительно и то, что, когда в 1900 году факты генетики заново
открыли Де Фриз, Чермак и Корренс, и, наконец, было признано значение
работ Менделя, главные возражения исходили от небольшой группы ма¬
тематиков, которые изучали наследственность при помощи статистических
методов.5Стили мышления, формирующиеся в результате занятий математикой
и биологией, конечно, очень сильно различаются; однако эти различия,
как представляется, не определяются интеллектуальными способностями.
Безусловно, было бы ошибкой сказать, что оперирование математически¬
ми символами требует больше интеллекта, чем разработка оригинальных
идей в биологии; напротив, такая работа кажется вполне сравнимой с ис¬
пользованием микроскопа, красителей и фиксаторов; в то же время ори¬
гинальные идеи в обеих сферах возникают в результате весьма сходной
деятельности при одинаковых способностях. Это согласуется с точкой зре¬
ния, что интеллект, строго говоря, мало поддается тренировкам. Зато мож¬
но успешно тренировать воображение, причем, по-видимому, математики
и биологи используют его совершенно по-разному. Большинство биологов,
наверное, считает, что здесь преимущество полностью на их стороне. Они
рано знакомятся с неисчерпаемым разнообразием живых существ; первые
же проведенные вскрытия, даже всего лишь лягушки или какой-нибудь аку¬
лы, демонстрируют картины удивительно сложные и интересные, тогда как
обычно кажется, что математики имеют дело только с чистыми абстракци¬
ями, с линиями и точками, бесконечно тонкими плоскостями и массами,
сосредоточенными в идеальном центре тяжести. Возможно, лучше всего
я смогу показать, что у математиков также до некоторой степени развито
воображение, процитировав замечание, небрежно брошенное Эддингтоном
в недавней книге:
ПредисловиеIX«Едва ли надо добавлять, что в естественных науках рассмотрение области
более широкой, чем реальность, приводит к значительно лучшему пониманию ре¬
альности» (р. 267, The Nature of the Physical World).Для математика такое высказывание — почти трюизм. Если бы эти
слова были произнесены биологом о предмете своего исследования, они
указывали бы на его необычайно широкий кругозор. Ни один биолог, изу¬
чающий половое размножение, никогда не примется разрабатывать пробле¬
му, что может происходить с организмами, у которых существует три или
более полов; однако что еще ему делать, если он хочет понять, почему по¬
лов на самом деле всегда только два? Обычная математическая процедура
при работе с любой реальной задачей сводится к тому, чтобы, вычленив то,
что считается важными элементами задачи, рассмотреть ее условия как од¬
ну из системы возможностей, неограниченно более широкой по сравнению
с реальностью, существенные зависимости в которой подвластны обобщен¬
ным рассуждениям и могут быть описаны общими формулами, при жела¬
нии приложимыми к любому частному случаю. Даже слово «возможности»
в этом утверждении чрезмерно ограничивает рамки практических проце¬
дур, привычных для математика; поскольку в самом начале своей карье¬
ры он познакомился с преимуществами воображаемых решений, то может
легко представить волновой, или переменный, ток в терминах вроде квад¬
ратного корня из минус единицы. Самое серьезное препятствие для интел¬
лектуального сотрудничества, вероятно, было бы устранено, если бы было
четко и однозначно признано, что наибольшие различия связаны не с ин¬
теллектуальными методами и еще менее — с интеллектуальными способ¬
ностями, но с тем, что огромные способности к воображению у каждого
специализированы в соответствии с потребностями его конкретной обла¬
сти исследования. Я не могу представить себе более полезных изменений
в научном образовании, чем такие, которые позволили бы каждому хоть
в какой-то мере оценить мощную работу воображения в областях науки,
которыми занимаются другие.В будущем революционное значение Менделизма будет вытекать
из корпускулярного характера элементов наследственности. На этот факт
может опираться рациональная теория Естественного Отбора, и, следова¬
тельно, это имеет громадное значение. Заслуга этого открытия, безусловно,
принадлежит Менделю; в нашей стране Бэтсон больше всего способство¬
вал тому, что оно получило известность на раннем этапе. К сожалению, он
не был готов оценить математические или статистические аспекты биоло¬
гии; из-за этого, а также по ряду других причин он не только не сумел сам
сформулировать эволюционную теорию, но и совсем не осознал того, что
Менделизм заполнил пробелы в общей картине, нарисованной Дарвином.
XПредисловиеЕго интерпретация фактов, описанных Менделем, с самого начала была
обусловлена исключительно его ранней верой в скачкообразное происхож¬
дение видовых форм. Хотя благодаря этому Бэтсон, скорее, препятствовал
развитию эволюционной теории, многочисленные исследования последо¬
вателей Менделя во всем мире несомненно смогли преодолеть ошибки,
сделанные теми, кто их начинал. Как пионер генетики, Бэтсон сделал бо¬
лее чем достаточно, чтобы искупить поспешную полемику в своих ранних
работах.6Рассматривать Естественный Отбор как независимое, опирающееся
на свой собственный фундамент явление — не значит преуменьшать его
значение для теории эволюции. Напротив, как только нам потребуется
представление, основанное на каких-то других подходах, кроме сравнения
и аналогии, становится необходимой такая независимая дедуктивная осно¬
ва. Эта необходимость особенно важна в том, что касается изучения челове¬
ка; поскольку мы многое знаем о структуре общества, мотивах людей, о ста¬
тистике демографических показателей у этого вида, использование дедук¬
тивного метода может помочь получить здесь более глубокие знания об эво¬
люционном процессе, чем в каком-либо другом случае. Кроме того, для на¬
шей цели было бы важно привлечь внимание к некоторым последствиям
принципа Естественного Отбора, которые неизбежно выпадают из сферы
нашего внимания, поскольку не заключаются в адаптивной модификации
видовых форм. Генетические явления доминантности и сцепления, по-ви-
димому, представляют примеры именно такого рода; их исследование в бу¬
дущем может очень существенным образом расширить нашу тему.Как я ни старался, мне не удалось сделать эту книгу легкой для чтения.
Чтобы помочь читателю, я постарался привести небольшое резюме в конце
каждой главы, кроме главы 4, резюме которой объединено с резюме гла¬
вы 5. Если угодно, можно даже рассматривать главу 4 как математическое
приложение к соответствующей части резюме. Рассуждения, относящиеся
к человеку, строго говоря, неотделимы от более общих глав, но объединены
и помещены отдельно в части книги, начинающейся с главы 8. Полагаю, что
никого не удивит, что большое число поднятых проблем требует значитель¬
но более полного, точного и всестороннего обсуждения. Полностью осве¬
тить предмет настоящей книги представляется невозможным до тех пор,
пока не будет выработана традиция математического исследования биоло¬
гических проблем, подобного тому, которое используют в математической
физике при решении многих особенно сложных вопросов.7Ротамстед, июнь 1929 г. Р.А. Ф.
Глава 1Природа наследственностиСледствия теории слитной наследственности, по Дарвину8. Противоречия,
замеченные Дарвином. Корпускулярная наследственность. Сохранение дисперсии.
Теории эволюции, обусловленной мутациями. Вся ли наследственность корпуску-
лярна? Природа и частота наблюдаемых мутаций.Однако в настоящее время, завершив в основном работу на эту тему,
я могу сделать вывод, что внешние условия значат очень мало и лишь спо¬
собствуют изменчивости. Эта изменчивость (благодаря которой ребенок
не совсем похож на своих родителей), на мой взгляд, очень сильно отлича¬
ется от образования четко выраженной разновидности или нового вида.
ДАРВИН, 1856 (Life and Letters9, ii, 87).Как правильно заметил Самуэль Батлер: «Мне кажется, что истин¬
ное «Происхождение видов» — это «Происхождение вариаций», в чем бы
оно ни заключалось». У. БЭТСОН, 1909.Следствия теории слитной наследственностиТот факт, что Чарльз Дарвин принимал теорию слияния, или слитной
наследственности, как принимают на веру многие воззрения, считающи¬
еся очевидными в определенный момент времени, широко известен. То,
что принятие им этой теории имело важное значение для формирования
его взглядов на вариации и, соответственно, на то, как он вместе с други¬
ми представлял себе возможные причины эволюции органического мира,
не было, мне кажется, понятно ему самому, и остается непонятым по сей
день. В настоящей главе я хочу показать логические следствия, вытекаю¬
щие из теории слитной наследственности, и показать их влияние не только
на развитие взглядов Дарвина, но и на изменение представлений об этой
теории и о других гипотезах в результате принятия противоположной им
теории корпускулярной наследственности.Интересно, что и сам Дарвин, очевидно, ощущал необходимость су¬
ществования альтернативы теории слитной наследственности, хотя он, ско¬
рее всего, так и не выработал четкой корпускулярной теории. В письме
2Глава 1к Хаксли, написанном, вероятно, в 1857 году, встречается такой абзац (More
Letters, vol. I, Letter 57):Подходя к теме с наиболее интересующей меня стороны, а именно с точки
зрения наследственности, в последнее время я начал думать, правда, очень при¬
близительно и неясно, что размножение в результате истинного оплодотворения
окажется не истинным слиянием, а неким смешением двух отдельных индивидов,
вернее, бесконечного числа особей, так как каждый родитель имеет своих родителей
и предков. Иначе мне абсолютно непонятно, каким образом гибридные формы так
часто возвращаются к предковым формам. Но все это, конечно, еще очень незрело.Эта мысль, похоже, так и не получила дальнейшего развития, возмож¬
но, из-за напряженной работы до и после опубликования Origin. Дарвин,
конечно, не осознавал, что его соображения по поводу вариаций, высказан¬
ные в черновых набросках за 1842 и 1844 годы9а, которые год спустя (1858)
он превратит в первую главу Origin, пришлось бы исходя из корпускуляр¬
ной теории пересмотреть заново. Те же взгляды практически без изменений
высказываются им и при обсуждении «Причин изменчивости» в главе 22
книги Variation of Animals and Plants, опубликованной в 1868 году.Представления Дарвина, которые можно реконструировать на основа¬
нии этих четырех источников, можно обобщить следующим образом:(а) при слитной наследственности половое размножение быстро приведет
к единообразию;(б) если изменчивость сохраняется, то должны непрерывно работать ме¬
ханизмы, вызывающие новые вариации;(в) причины огромной изменчивости одомашненных видов, которая на¬
блюдается у всех видов и во всех регионах, следует искать в условиях
одомашнивания;(г) единственными признаками процесса одомашнивания, достаточно об¬
щими для того, чтобы они были применимы во всех случаях, являются
изменения условий содержания и увеличение количества пищи;(д) создается впечатление, что некоторые изменения условий оказывают
определенное систематическое воздействие определенного типа, то
есть увеличение количества пищи вызывает (наследуемое) увеличе¬
ние размера тела, однако важным эффектом является неопределенная
изменчивость, идущая во всех направлениях и возникающая в новых
условиях за счет нарушения регулярной деятельности репродуктивной
системы;(е) дикие виды под влиянием геологических перемен также время от вре¬
мени испытывают воздействие новых условий, а иногда и временного
Природа наследственности3увеличения запасов пищи, поэтому они, пусть и редко, также могут
изменяться. Если в этих случаях отбор не происходит, вариации ней¬
трализуют друг друга благодаря половому размножению и исчезают,
но если вмешивается отбор, вариации в нужном направлении накапли¬
ваются и приводят к необратимому эволюционному изменению.Для современного читателя эти соображения могут показаться очень
странным способом доказательства существования Естественного Отбора,
ведь единственное, что из этого следует, это вывод, что как дикие виды, так
и одомашненные хотя бы время от времени демонстрируют наследствен¬
ную изменчивость. Тем не менее эти соображения используются с этой
целью в двух набросках и в Origin. Следует помнить, что во время написа¬
ния набросков Дарвин имел мало прямых доказательств этой точки зрения,
которая была основой его аргументации в связи с тем, что способность че¬
ловека изменять одомашненные виды путем отбора не подвергалась сомне¬
нию; даже во второй главе Origin, посвященной «Вариациям в природе»,
речь идет главным образом о природных разновидностях, настолько четко
выраженных, что их отмечали ботаники, и Дарвин, несомненно, восприни¬
мал их не как материал, а как продукт эволюции. В течение двадцати шести
лет, между 1842 и 1868 годами должно было бы накопиться достаточное ко¬
личество фактов, чтобы убедить его, что наследственная изменчивость так
же широко распространена, пусть и не так заметна, в дикой природе, как
и у одомашненных видов. Похоже, что значимость аргументации, о кото¬
рой говорилось выше, уже не представлялась ему настолько существенной,
и в 1868 году он начинает обсуждение данной темы словами: «Предмет
туманен, но может пригодиться для выяснения объема нашего невежества»
( Variation, Chapter 22).Привлекательность набросков состоит в том, что они показывают при¬
чины, приводившие Дарвина к тем или иным умозаключениям, тогда как
более поздние работы часто дают только доказательства, на основании
которых читатель должен судить об их правильности. Антитеза кажется
неортодоксальной, поскольку каждый пытливый ум формирует суждение
без прямого доказательства, иначе доказательства часто нельзя было бы до¬
быть вовсе. Объективность и научный порядок присутствуют в том случае,
когда учитываются мнения, сформированные в ответ на предоставление
как можно большего числа важных доказательств. Первые пункты из при¬
веденных выше соображений появляются только в двух набросках, тогда
как выводы продолжаются практически без изменений вплоть до Variation
of Animals and Plants. В самом деле, первый пункт (а), логически самый
важный из всех, появляется только в 1842 году. В 1844 г. он подразумевает¬
ся на основе неизбежных следствий. Я убежден, что его значение для Origin
4Глава 1вряд ли можно увидеть при рассмотрении только этой книги. Вот отрывок
из рукописи 1842 г. (Foundations, р. 2):Каждый родитель передает свои особенности потомству, поэтому если дать
возможность разновидностям свободно скрещиваться, то кроме того случая, ко¬
гда два индивида, обладающие одинаковой особенностью, сумеют соединиться, та¬
кие разновидности будут постоянно исчезать. Все бисексуальные животные долж¬
ны скрещиваться, гермафродиты-растения скрещиваются, вполне возможно, что
и гермафродиты-животные скрещиваются тоже — вывод усиливается...и несколько невнятный отрывок не совсем ясного содержания:Если скрестить представителей двух совершенно отличных друг от друга раз¬
новидностей, то получится третья раса — исключительно богатый источник вариа¬
ций для домашних животных. При таком скрещивании черты чистопородных роди¬
телей будут утрачены, число рас тем самым [неразборчиво], но различия [?], кроме
[неразборчиво]. Но если скрестить разновидности, чуть отличные друг от друга,
их незначительное различие будет уничтожено, по крайней мере для нашего глаза,
разновидность, отличающаяся длинными ногами, будет иметь потомство с менее
выраженными чертами. Свободное скрещивание является великой силой для ста¬
новления единообразия любой породы.Это утверждение имеет важное значение, оно подчеркивает кардиналь¬
ное различие двух теорий — слитной наследственности и корпускулярной
наследственности. Следующее доказательство выражает это в биометриче¬
ских терминах.Пусть х и у — отклонения любого признака обоих родителей от ви¬
дового среднего; в случае, если значение этого признака зависит не только
от наследственных, но также и от ненаследственных факторов (факторов
среды), то пусть х и у будут обусловливать наследуемую часть отклоне¬
ний. Величина наследственной изменчивости в любом поколении измеря¬
ется дисперсией, определяемой как среднее квадрата отклонения х или у.
При исключительно слитной наследственности наследуемые доли откло¬
нений у потомства будут, без учета мутаций, равняться -|(х + у)); таким
образом, в отсутствие мутаций дисперсия в поколении потомков будет со¬
ставлять среднее значение величин \{х2 + 2ху + у2).Среднее арифметическое значений х и среднее арифметическое зна¬
чений у равны нулю, так как каждая из этих величин определяется как
отклонение от видового среднего; в результате в отсутствие селективного
скрещивания10 среднее значение ху также равно нулю, и дисперсия в до¬
чернем поколении оказывается равной половине дисперсии в родительском
поколении.11 В более общем случае отношение равно не а -|(1 + г),
где г — это корреляция между х и у. При этом г не может быть больше
Природа наследственности5единицы, иначе среднее значение положительных величин (х — у)2 долж¬
но было бы быть отрицательным, и может в точности равняться единице,
когда эти величины равны нулю, то есть если размер каждой особи точно
задает размер ее возможных партнеров. Слова Дарвина: «кроме того слу¬
чая, когда два индивида, обладающие одинаковыми особенностями, сумеют
соединиться», — выражают его мнение о малой вероятности подобной кор¬
реляции между партнерами.Эффект корреляции между партнерами должен способствовать (в слу¬
чае отрицательной корреляции) и затруднять (в случае положительной кор¬
реляции) тенденцию слитной наследственности уменьшать дисперсию; по¬
добные эффекты не лишены смысла, поскольку даже при корреляции, рав¬
ной 0,5, и сходстве между партнерами на уровне сходства между родите¬
лем и потомком скорость уменьшения дисперсии будет чуть больше по¬
ловины. Важное следствие слитной наследственности состоит в том, что
если не поддерживать ее интенсивной корреляцией скрещивающихся осо¬
бей по значениям признака, то наследуемая дисперсия уменьшается при¬
мерно вдвое в каждом поколении. Для поддержания постоянства дисперии
в каждом поколении должны происходить новые мутации для восполнения
утраченной половины. Если изменчивость сохраняется, то, как справедливо
замечал Дарвин, постоянно должны действовать силы, приводящие к появ¬
лению новой изменчивости. Почти каждый индивид следующего поколения
должен быть мутантом, то есть он должен подвергаться воздействию всех
этих сил, и более того, он должен быть мутантом по разным признакам.Неизбежное следствие теории слитной наследственности состоит
в том, что большая часть имеющейся на данный момент наследствен¬
ной дисперсии должна иметь недавнее происхождение12. Половина должна
быть новой в каждом поколении, а половина оставшейся половины долж¬
на быть только на одно поколение старше, и так далее. Меньше чем одна
тысячная часть общей дисперсии может просуществовать десять поколе¬
ний; даже когда в случае селективного скрещивания мы можем говорить
о двадцати поколениях, общий вывод остается прежним — причиной измен¬
чивости одомашненных видов сторонники теории слитной наследственно¬
сти, если таковые еще существуют, могут назвать только условия содержа¬
ния. Различия, если ими собирается воспользоваться селекционер или если
их поставляет естественный отбор, должны быть подхвачены немедленно,
вскоре после того, как произошла мутация, и до того, как она утрачена ви¬
дом. Следующий отрывок из набросков 1844 года показывает, что Дарвин
имел по этому вопросу ясное представление (pp. 84-86):Давайте предположим, что организм по чистой случайности (которая едва ли
повторится в течение последующей тысячи лет) попадает на современный вул¬
6Глава 1канический остров, находящийся в процессе формирования и не занятый наибо¬
лее адаптированными организмами. Новый организм может успешно закрепиться
здесь, хотя внешние условия могут сильно отличаться от условий его родины. Но¬
вые условия, как мы вправе ожидать, неизбежно скажутся в некоторой незначи¬
тельной степени на его размере, окраске, характере покровов и т. д. и в результате
необъяснимых причин — даже на частях и органах тела. Но мы можем далее пред¬
положить (что гораздо серьезнее), что воздействие будет оказано и на репродуктив¬
ную систему, как это бывает в случае разведения домашних животных, и структура
потомства приобретет некоторую степень пластичности. Следовательно, существует
вероятность, что практически каждая часть тела некоторым неопределенным обра¬
зом изменится по сравнению с типичной формой, и поэтому без отбора свобод¬
ное скрещивание по этим небольшим различиям (вместе с тенденцией возвращения
к начальной форме) будет постоянно препятствовать этому неустойчивому воздей¬
ствию внешних условий на репродуктивную систему. Это, как я полагаю, был бы
несущественный результат, если бы не отбор. И здесь я должен заметить, что вы¬
шеупомянутые замечания в равной степени приложимы к малым и признанным
различиям, наблюдаемым у некоторых организмов в природе, в равной мере они
приложимы и к вышеупомянутым гипотетическим различиям, зависящим от изме¬
нений условий.Представим теперь, как некое Существо с зоркостью, достаточной для того,
чтобы заметить различия в наружной и скрытой от человека внутренней органи¬
зации, и с прозорливостью, позволяющей заглянуть в будущие века, с неусыпным
вниманием наблюдает и отбирает с какой-то целью потомство организма, возник¬
шего при предшествующих обстоятельствах; я не вижу видимой причины, почему
оно не может создать новую расу (или несколько новых рас при условии, что оно
должно отделить потомство исходного оранизма и действовать на нескольких остро¬
вах), приспособленную к новым условиям. Если мы допустим, что его способность
распознавать, прозорливость и целеустремленность несравненно больше, чем те же
качества у человека, мы можем предположить, что красота и сложность приспособ¬
лений новых рас и их отличия от исходного родителя окажутся больше, чем у выве¬
денных при содействии человека пород домашних животных: мы можем помочь ему
в его тяжелых трудах, предположив, что условия внешней среды на вулканическом
острове, начиная с его постепенного образования, под влиянием появления все но¬
вых иммигрантов меняются и оказывают воздействие на репродуктивную систему
организма, над которым Существо трудится, поддерживая организацию организма
в более пластичном состоянии. При наличии достаточного времени это Существо
может с полным основанием (в отсутствие неведомых препятствующих ему зако¬
нов) добиться практически любого результата.Противоречия, замеченные ДарвиномХотя нет сомнения в том, что на протяжении нескольких лет, с тех
пор как Дарвин начал размышлять о теории Естественного Отбора, общее
Природа наследственности7направление его мысли о наследственности и вариациях определялось со¬
ображениями, основанными на слитной наследственности и ее логических
выводах, но это не давало ему полного удовлетворения. Он понимал, что
явление возврата к предковым формам не укладывается в предложенную
им схему наследственности, и, как видно из его уже упомянутого пись¬
ма к Хаксли, необходимость рассматривать этот фаю- как самостоятельный
принцип его совсем не устраивала. В действительности к 1857 году он был
уже готов разработать схему наследственности, где возврат к предковым
формам был бы одним из ее следствий. Характерно, что противоречие, свя¬
занное с изменчивостью пород домашних животных, также не ускользнуло
от его внимания. В 1844 году он замечает (pp. 77, 78, Foundations), что одо¬
машненные в далекой древности животные и растения не менее изменчивы,
чем те, которые были одомашнены совсем недавно, и утверждает, что хотя
питание не могло становиться намного лучше на протяжении всей длитель¬
ной истории одомашнивания животных и растений, этот фактор сам по себе
не может объяснить наличие продолжающей существовать большой измен¬
чивости. В отрывке далее говорится:Если бы это был избыток пищи, по сравнению с тем, что можно добыть, живя
в дикой природе, его воздействие продолжается невероятно долго. Сколько лет уже
выращивается пшеница и одомашнены животные, и мы не можем предположить,
что количество их пищи постоянно возрастает, тем не менее они являются самыми
изменчивыми из всей нашей домашней продукции.Это противоречие относится ко второй предполагаемой причине изменчи¬
вости, а именно к изменившимся внешним условиям, хотя тут можно воз¬
разить, что условия разведения или ухода за домашними животными посто¬
янно меняются с той или иной скоростью. В отрывке из Variation of Animals
and Plants (p. 301) говорится:Более того, не похоже, чтобы более или менее благоприятное изменение кли¬
мата являлось бы наиболее действенной причиной изменчивости, поскольку в слу¬
чае растений, как неоднократно показывает Альф. Декандоль в своей Geographie
Botanique, родина растения, где в большинстве случаев оно наиболее давно культи¬
вируется, оказывается тем местом, где встречается наибольшее число его разновид¬
ностей.Создается впечатление, что Дарвин успокаивал себя тем, что страны, в ко¬
торых впервые произошло одомашнивание животных и растений, по край¬
ней мере настолько же богаты разновидностями, как и страны, куда они
были завезены, и естественно предположить, что, выясняя этот вопрос, он
старался найти доказательства влияния изменившихся условий. Неизвест¬
но, решил ли Дарвин для себя эту задачу, но из многих отрывков ясно,
8Глава 1что он видел необходимость дополнения первоначальных соображений, до¬
пуская, что причины вариаций, которые оказывают воздействие на репро¬
дуктивную систему, должны иметь способность действовать в отсроченной
и кумулятивной форме так, чтобы вариации могли бы сохраняться в тече¬
ние многих последующих поколений.Корпускулярная наследственностьПоразительно, но если хотя бы один мыслитель в середине XIX века
взял на себя труд, пусть в качестве задачи из абстрактного и теоретического
анализа, построить теорию корпускулярной наследственности, он пришел
бы на основании всего лишь нескольких простых заключений к системе,
идентичной современной схеме наследственности Менделя, или факторной
наследственности. Признанное ненаследование шрамов и увечий подгото¬
вило бы его к мысли о наследственной природе организма как о чем-то до¬
статочно определенном, хотя, возможно, и выраженном, если судить по ви¬
димой внешности, не совсем точно. Если бы он предположил, что эта на¬
следственная природа полностью определена совокупностью наследствен¬
ных частиц (генов), являющихся частью структуры, и допустил бы в то
же время, что организмы с определенным типом наследственной структу¬
ры способны к половому размножению, он, несомненно, был бы вынужден
признать, что каждый организм должен получить определенную порцию
генов от каждого родителя, и что в результате он должен передать каждо¬
му из потомков только определенную часть генов. Упрощение, что кроме
пола и, возможно, других признаков, наследственно связанных с полом,
вклад обоих родителей равнозначен, не было бы с уверенностью приня¬
то без доказательства, каким являются реципрокные скрещивания, но наш
воображаемый теоретик, зайдя так далеко, наверняка сумел бы вообразить
абстрактную структуру, где каждый ген имел бы свое определенное место,
или локус, которое могло бы быть занято и геном другой формы, имей он
других родителей. Те организмы (гомозиготы), которые получают одинако¬
вые гены в каждой паре соответствующих локусов от каждого из родителей,
обязательно передадут те же гены всем своим потомкам, тогда как другие
(гетерозиготы), которые получили от двух своих родителей гены разной
формы, и являются, относительно изучаемого локуса гибридами, имеют,
когда речь идет о каждом потомке, равный шанс передать один из этих
двух разных генов. Гетерозигота при скрещивании с любой гомозиготой
даст как гетерозигот, так и гомозигот в отношении, которое с увеличением
числа потомков должно стремиться к равенству, тогда как при скрещивании
двух гетерозигот можно ожидать, что каждая гомозигота появится в четвер¬
Природа наследственности9той части всего потомства, а оставшуюся половину составят гетерозиготы.
Таким образом, оказывается, что наряду с доминантностью и сцеплением,
включая сцепление с полом, все основные особенности системы Менде¬
ля вытекают из принятия корпускулярной наследственности простейшего
типа и могли бы быть предсказаны a priori*, если бы кто-то допустил воз¬
можность, что законы наследственности могут быть в действительности
настолько просты и определенны13.Расщепление по одной паре генов было описано Менделем в его статье
в 1865 году14. Кроме того, Мендель показал в своей работе наличие доми¬
нирования, а именно тот факт, что гетерозиготная особь не является проме¬
жуточной по внешности, но абсолютно неотличима от одной из гомозигот¬
ных форм. Факт доминирования, хотя и представляет огромный теоретиче¬
ский интерес, не является существенной чертой генной системы, а в ряде
важных случаев и вовсе отсутствует. Мендель также показал, чего вряд ли
мог ожидать теоретик, что разные факторы, рассмотренные им в комбина¬
ции, разделяются простейшим из возможных способов, а именно независи¬
мо. Позже, после переоткрытия законов Менделя в конце XIX века, были
открыты случаи сцепления, когда факторы из одной группы сцепления —
пара генов, полученная от одного и того же родителя,— чаще всего сов¬
местно передаются одному ребенку. Концептуальная структура локусов, та¬
ким образом, может быть представлена в виде нескольких частей, которые
сейчас, на основании весьма убедительных доказательств, отождествляют¬
ся с темно окрашенными тельцами, или хромосомами, видимыми в ядрах
клеток на определенных стадиях клеточного деления.В соответствии с механизмом корпускулярной наследственности, несо¬
мненно, возможно осуществление возврата к предковым формам, то есть
явления, когда индивид напоминает дедушку или более далекого предка
в той же степени, в какой отличается от родителей, поскольку существует
возможность случайного повторения предковой комбинации генов. В про¬
стейшей форме это наблюдается при доминировании, поскольку скрещива¬
ние двух гетерозигот дает некоторую долю потомков, несущих рецессивные
гены, то есть внешне отличных от родителей, но, возможно, напоминающих
кого-то из бабушек и дедушек или более далеких предков.Сохранение дисперсииНельзя сказать, что до конца осознан еще один важный факт отли¬
чия корпускулярной наследственности от слитной. Врожденной тенденции*До опыта (лат.).
10Глава 1к уменьшению изменчивости нет. В популяции, где скрещивание проис¬
ходит случайным образом, и где два альтернативных гена одного фак¬
тора находятся в отношении р : q, присутствуют три генотипа в соот¬
ношении р2 : 2pq : q2, это означает, что определяемые ими признаки бу¬
дут представлены в популяции в фиксированной пропорции, хотя они мо¬
гут сочетаться с другими определенными признаками, детерминируемы¬
ми другими факторами при условии, что соотношение р : q остается не¬
изменным.10 В действительности это соотношение будет неизбежно под¬
вергаться небольшим изменениям, во-первых, из-за случайного выживания
и размножения особей разных генотипов и, во-вторых, из-за их селективно¬
го выживания по той причине, что разные генотипы, вероятно, в неравной
степени, пусть и с небольшими различиями, приспособлены к своей задаче
выживания и размножения. Эффект случайного выживания легко поддается
вычислению, и оказывается, как будет более подробно показано в главе 4,
что в популяции, состоящей из п случайно скрещивающихся индивидов,
дисперсия за счет случайности будет уменьшена вдвое в течение 1,4п по¬
колений. Поскольку число индивидов, которые должны выжить и оставить
потомство в каждом поколении для большинства видов превышает мил¬
лион, а для многих по крайней мере составляет число, кратное миллиону,
видно, что эта причина уменьшения наследственных различий крайне ма¬
ла по сравнению с двукратным уменьшением за одно или два поколения
при слитной наследственности.Как будет видно в главе 4, отбор является гораздо более важным фак¬
тором поддержания изменчивости видов в определенных рамках. Однако
даже достаточно интенсивный отбор будет изменять соотношение р : q от¬
носительно медленно, и мы не можем вообразить, что должно было бы
произойти, чтобы уменьшение дисперсии благодаря отбору при полном от¬
сутствии мутаций намного превышало одну десятитысячную той величи¬
ны, которая приписывается слитной наследственности. Непосредственным
выводом из этой огромной разницы является то, что скорость мутаций, ко¬
торая должна существовать для поддержания нужного уровня изменчиво¬
сти при корпускулярной наследственности, во много тысяч раз меньше той,
которая требуется при слитной наследственности. Поэтому теории, которые
факторам, способным, казалось бы, вызывать мутации, как это было «с упо¬
треблением и неупотреблением», по Дарвину, приписывают возможность
задавать направление эволюции, предстают совсем в разном свете в зави¬
симости от того, какую из теорий наследственности мы принимаем. Лю¬
бая эволюционная тенденция, которая должна благоприятствовать мутаци¬
ям в одном направлении более, чем в другом (а время от времени предлага¬
ются разные подобные механизмы), теряет в силе тысячекратно, коль скоро
Природа наследственности11мы примем теорию корпускулярной наследственности в том или ином ви¬
де, тогда как направляющая сила Естественного Отбора, зависящего толь¬
ко от величины наследственной дисперсии, всегда останется абсолютно
неподвластна любому подобному изменению. Это заключение, в равной ме¬
ре приложимое ко всем подобным теориям, не зависит от того факта, что
ббльшая часть причин, по крайней мере для Дарвина, заставляющих припи¬
сывать какую-либо значимую роль в возникновении мутаций окружающей
среде, становится несостоятельной, как только исчезает необходимость до¬
казывать, что причиной большой изменчивости пород домашних животных
является относительно недавнее влияние их искусственной среды.Поразительное явление, о котором хорошо знал Дарвин, состоит в том,
что братья и сестры, имеющие общих родителей, а значит, и всех пред¬
ков, могут разительно различаться по своим наследственным свойствам,
что при наличии двух теорий может быть объяснено совершенно разны¬
ми способами. При слитной наследственности это — четкое доказатель¬
ство частого возникновения новых мутаций, зависящих, как предполагает
сильное сходство близнецов, от условий во время зачатия и беременности.
При корпускулярной наследственности это — неизбежное следствие того,
что по каждому признаку значительная доля популяции, часто не менее
половины, является гетерозиготной, а значит, каждые два потомка могут
с одинаковой вероятностью получить от каждого родителя как одинаковые,
так и разные гены. Вследствие большого сходства между статистической
концепцией дисперсии и физической концепцией энергии нам будет полез¬
но попытаться представить, что гетерозигота обладает дисперсией в потен¬
циальной или латентной форме, а это значит, что вместо того чтобы исчез¬
нуть при скрещивании гомозиготных генотипов, она просто сохраняется
в такой форме, из которой впоследствии может появиться. При случайном
скрещивании в популяции сразу же устанавливается условие статистиче¬
ского равновесия между скрытой и видимой формами дисперсии. Корпус¬
кулярная теория наследственности напоминает кинетическую теорию газов
с их эластичными соударениями, тогда как теория слитной наследственно¬
сти напоминает теорию газов с неэластичными соударениями, где требуется
некоторый постоянно действующий внешний фактор, который поддержи¬
вал бы частицы в движении.Свойство корпускулярной теории, заключающееся в способности со¬
хранять дисперсию в течение неопределенно длительного времени, лег¬
ко объясняет запаздывающий или кумулятивный эффект одомашнивания
с увеличением дисперсии у одомашненных видов, на что обращал внима¬
ние Дарвин. Очевидно, что многие из наших разновидностей домашних
животных плохо приспособлены к выживанию в дикой природе. Мутации,
12Глава 1на основе которых они возникли, могли происходить намного раньше, чем
началось одомашнивание, либо никак не проявляя себя, либо обусловливая
заметную дисперсию у дикого вида. При одомашнивании, однако, Есте¬
ственный Отбор смягчает свою строгость, позволяя носителям мутаций
выжить и каждому выжившему добавить что-то в копилку наследственной
дисперсии; кроме того, новинки формы или цвета, даже если они уродливы,
несомненно привлекают интерес и внимание людей и ценятся человеком
за свою необычность. Скорость, с которой увеличивается новая диспер¬
сия, таким образом, возрастает. Без учета каких бы то ни было изменений
частоты мутаций при одомашнивании мы должны, исходя из известных
условий одомашнивания, неизбежно сделать вывод, что изменчивость одо¬
машненных видов, должно быть, больше, чем аналогичных диких видов,
и что этот контраст должен быть самым заметным для видов, одомашнен¬
ных в числе первых. Таким образом, основные трудности, осознававшиеся
Дарвином, снимаются корпускулярной теорией.Теории эволюции, обусловленной мутациямиТеории эволюции, которые основываются на гипотетических причи¬
нах, способных изменять частоту или направление возникающих мутаций,
делятся на четыре группы. Сделав такое заявление, было бы уместно опре¬
делить термин «мутация», который в разное время имел самые разные зна¬
чения, просто как возникновение любого наследуемого признака.(А) Можно предположить, вслед за Ламарком, что в случае, когда речь
идет о животных, состояние психики, и особенно внутренние стремления
организма, обладают свойством вызывать мутации, позволяющие успеш¬
но удовлетворять эти стремления потомкам. Этот взгляд допускает, (1) что
существует механизм, который вызывает мутации и даже приводит их в со¬
ответствие с состоянием нервной системы, и (2) что стремления животных
вообще таковы, что их воплощение может улучшить способность вида вы¬
живать в естественной среде, а также помочь отдельным частям тела взаи¬
модействовать друг с другом и поддерживать жизненную активность взрос¬
лого животного, равно как и обеспечивать его нормальное эмбриональное
развитие. Внутренние стремления животных должны, в самом деле, быть
мудро направлены, равно как и отличаться эффективностью в индуцирова¬
нии необходимых мутаций16.(Б) Способность к адаптации можно повсеместно наблюдать как среди
животных, так и среди растений, соответствующие органы которых, на¬
пример мышцы или железы, отвечают повышением активности и увеличе¬
нием размера в ответ на предъявляемые к ним повышенные требования.
Природа наследственности13Можно предположить, вслед за Дарвином, что такие ответы в виде возрос¬
шей функциональной активности вызывают мутации или сопровождаются
мутациями, способствующими увеличению размера или активности соот-
нетствующего органа в будущих поколениях, даже в том случае, когда ни¬
каких дополнительных стимулов к активности этого органа не поступает.
Этот взгляд подразумевает, (1) что сила, которой обладают отдельные ча¬
сти организма при адаптивном ответе на возросшие к ним требования, сама
не является продуктом эволюции, но должна постулироваться как первич¬
ное свойство живой материи, что требует, (2) чтобы существовал механизм,
в соответствии с которым адаптивный ответ сам бы вызывал мутацию или
сопровождался соответствующей мутацией.Оба эти предполагаемых способа эволюции четко нацелены на объ¬
яснение не просто прогрессивного изменения живых существ, но соответ¬
ствия организма его месту в природе и частей его тела функционированию
в организме.(В) Можно предположить, что среда, в которой обитает организм, кон¬
тролирует природу происходящих в нем мутаций, направляя тем самым ход
его эволюции, подобно тому, как брошенное тело контролируется силовым
полем, в котором оно летит.(Г) Можно предположить, что происходящие в организме мутации —
результат «внутренней потребности» (не обязательно связанной с его ум¬
ственными способностями), внедренной в его изначальных предков и на¬
правляющей тем самым организм по пути предназначенной ему эволюции.Два последних предположения не способствуют лучшему пониманию
адаптации, но каждое содержит как минимум элемент истины, что, как
бы глубоко ни было наше незнание причин мутаций, мы можем, исходя
из наших познаний Природы, либо отнести их к особенностям самого ор¬
ганизма, либо к окружающей его среде, либо (в более общем случае) к их
взаимодействию. Общим во всех этих предположениях, однако, остается
одно условие, а именно: что направление эволюционных изменений опре¬
деляется основным направлением, в котором происходят мутации. Каким
бы разумным ни казалось это заключение, когда при слитной теории на¬
следственности каждый индивид рассматривался как мутант, или, скорее,
как множественный мутант, его нельзя оставить без вопроса, учитывая, что
в корпускулярной теории предусматривается значительно более низкая ча¬
стота мутаций.Следующий гипотетический механизм, определяющий эволюцию ви¬
дов в том же направлении, в котором происходят мутации, был предложен
Вейсманом. Вейсман дал гораздо более глубокую оценку, чем все его со¬
временники, эффективности Естественного Отбора в повышении адаптив¬
14Глава 1ности организмов к их жизни в данной среде. Он, однако, осознавал, что
процесс адаптации был бы более успешным, если мутационный процесс
мог бы приобрести направленность так, чтобы серия мутаций, способству¬
ющих увеличению или уменьшению части тела, являлась бы продолжением
начального события, которое было бы направлено в сторону такого увели¬
чения или уменьшения.17 Это допущение о наличии движущей силы в му¬
тационном процессе было бы, по его мнению, полезно в двух отношениях:
(1) оно позволило бы предполагаемой минимальной мутации, произошед¬
шей в благоприятствующем направлении, быть усиленной последующи¬
ми мутациями до «достижения ей селективной ценности»; (2) оно могло
бы объяснить постепенное уменьшение бесполезного органа без призна¬
ния, что каждый шаг в этом направлении создает некоторое преимущество
для того, кто избавляется от этого органа, по сравнению с тем, кто этим
органом обладает.Это понятие достижения селективной ценности, которое довольно ча¬
сто встречается в биологической литературе, включает, как мне кажется,
два разных случая. В первом случае мы можем представить, что при уве¬
личении размера органа его полезность не возрастает до тех пор, пока
не пройдена определенная критическая точка, за которой больший размер
означает большую полезность. В подобном случае (который ввиду суще¬
ствующей изменчивости каждого организма и сходных частей тела у род¬
ственных организмов) должен рассматриваться как несколько идеальный;
нас, по сути дела, беспокоит только один вопрос: достигнет или не до¬
стигнет существующая изменчивость особей изучаемого вида критической
точки. Если нет, то вид не сможет воспользоваться предложенным преиму¬
ществом просто потому, что он окажется недостаточно изменившимся, по¬
скольку предположение о заданности движения при возникновении мута¬
ций было сделано нами безусловно не для того, чтобы позволить организ¬
мам быть более изменчивыми, чем они были бы и без этого.Второе соображение, которое также часто встречается в литературе,
основано на странной гипотезе о том, каким образом селективное преиму¬
щество возрастает с изменением размера органа, от которого это преиму¬
щество зависит, поскольку иногда считается, что хотя при любом размере
органа его увеличение может стать преимуществом, это преимущество на¬
растает не непрерывно, а ступенчато, или по крайней мере что увеличение
ниже определенного предела дает преимущество, которое может быть на¬
звано «неоцененным» и оставлено в результате без внимания. Создается
впечатление, что и метафора, и лежащая в ее основе идея взяты из психо¬
физического эксперимента. Сравним, например, два физических ощущения
от тяжести двух предметов. Когда различия в весе незначительны, субъ-
Природа наследственности15скт не может заметить разницу и воспринимает веса как равные, тогда как
мри большем расхождении в весе он ощущает разницу. Если, однако, тот
же субъект участвует в аналогичном опыте многократно, и различия в весе
колеблются от ощутимых до ничтожно малых, оказывается, что разница,
ко торая раньше казалась абсолютно незаметной, становится существенной
и отчетливо ощущаемой разницей по регулярности частоты, с которой
один объект представляется тяжелее, чем другой (рис. 1). Прерывистость
лежит в основе нашего истолкования ощущений, а не в ощущениях как
таковых. В данном случае уровень выживания измеряется частотой, с ко¬
торой в жизни разных организмов, подверженных разнообразным случай¬
ностям одинаковой окружающей среды, происходят определенные явления,
такие, например, как смерть или производство потомства, и даже если рань¬
ше у нас оставались сомнения, то психофизические эксперименты дали
нам возможность лучше понять, что селективные преимущества будут рас¬
ти или уменьшаться непрерывно даже для изменений гораздо меньших,
чем те, которые способны воспринимать наши органы чувств, или органы
чувств хищника, или другого возможного участника данной биологической
ситуации. Если изменение в 1 мм имеет селективную ценность, то измене¬
ние в 0,1 мм будет иметь селективную ценность приблизительно в 10 раз
меньшую, но изменение нельзя игнорировать по той причине, что мы счи¬
таем его несущественным. Скорость, с которой число мутаций в популяции
растет за счет своего аллеломорфа, в действительности зависит от обуслов¬
ленного этим селективного преимущества, но скорость, с которой вид от¬
вечает на отбор, способствующий увеличению или уменьшению тех или
иных частей тела, зависит от общей наследственной дисперсии признака,
а не от того, явилось ли это результатом крупной или мелкой мутации.
Мы не можем говорить здесь о пороге поддающейся оценке селективной
ценности.Оставшееся преимущество, которое искал Вейсман, постулируя меха¬
низм зародышевого отбора, должно было объяснить прогрессивное умень¬
шение бесполезных органов, даже в том случае, когда сама их незначитель¬
ность ставит под вопрос селективное преимущество их подавления. Пред¬
мет этот весьма интересен и сам по себе заслуживает большего внимания,
чем может позволить объем этой книги. Для нашей настоящей цели доста¬
точно просто отметить, что (1) заявлять в каждом конкретном случае, что
постепенное подавление органа не приносит селективного преимущества,
представляется нам несколько выходящим за рамки наших действитель¬
ных знаний. Серьезным возражением Вейсману может стать следующий
1гример: receptaculum seminis муравья несомненно отличается малым раз¬
мером, но и сам муравей невелик, к тому же нас интересует не столько от-
16Глава 130092 96 100 104 108
вес в граммахРис. 1. Частота, с которой испытуемые объекты разного веса оцениваются как более
тяжелые по сравнению со стандартным весом в 100 г, данные Урбана для одного
испытуемого. Иллюстрация того, что при достаточном числе испытаний различия
в весе, как бы ничтожны они ни были, влияют на частоту правильных ответовдельный муравей, сколько вся популяция муравьев-рабочих в муравейнике.
Если рассматривать эту проблему как экономическую, мы, конечно, не рас¬
полагаем данными, чтобы решить, могло бы или нет подавление этого кро¬
шечного органа сказаться существенным образом на устройстве муравьи¬
ного государства. Можно провести параллели с человеческим обществом,
где удаление каких-то мелочей, не нужных каждому в отдельности, может
оказать ощутимое воздействие на социальные институты, которыми бес¬
спорно нельзя пренебрегать. Я не утверждаю, что подавление receptaculum
было полезно для муравья, но что и в этом, и в других случаях, если мы
попытаемся повнимательнее рассмотреть проблему, мы сразу же увидим,
что не обладаем достаточными знаниями для того, чтобы иметь основание
для уверенного ответа. (2) Во-вторых, взгляд Вейсмана, состоящий в том,
что при полном отсутствии отбора ненужный орган может уменьшиться,
выродиться и в конце концов исчезнуть в результате накопления последо¬
вательных мутаций, и особенно его взгляд, что это есть единственный тип
прогрессивных изменений, который может происходить только благодаря
мутациям, без направляющей роли Естественного Отбора, полностью со-
Природа наследственности17Iпасуется с современными знаниями о природе мутаций, хотя специальный
механизм, которым он пытался объяснить последовательное возникновение
приводящих к дегенерации мутаций, должен быть назван маловероятным.
Кроме того, вызывает логические возражения тот факт, что движущая си¬
ла предложенного им механизма зародышего отбора состоит в конкурен¬
ции за питательные вещества хроматиновых элементов, представляющих
дегенеративный орган, с одной стороны, и хроматиновых элементов, пред¬
ставляющих остальную часть тела, с другой. Сам дегенеративный орган
считается настолько несущественным, что его требования общего питания,
получаемого телом, должны быть оставлены без внимания, и здесь уместно
спросить, законно ли, говоря о снабжаемой питательными веществами за¬
родышевой клетке, вводить фактор конкуренции за питательные вещества,
который не учитывается, когда речь порой идет о недокормленном теле.Вся ли наследственность корпускулярна?Логические основания, позволяющие отвергнуть гипотезу о том, что
направление эволюционного изменения определяется направлением, в ко¬
тором происходят мутации, и тем самым отвергнуть целую группу теорий,
которые эту возможность допускают, были бы недостаточны, если бы со-
цременные исследования не дали ответа на два вопроса: (1) не может ли
быть, чтобы в дополнение к механизму корпускулярной наследственности,
которая была недавно открыта и продолжает изучаться в настоящее вре¬
мя, в живых организмах существовал бы еще не открытый механизм слит¬
ной наследственности? (2) Можно ли на основании известных нам фактов
предложить в рамках системы корпускулярной наследственности такой ме¬
ханизм, который мог бы регулировать основное направление мутационного
процесса и был бы неэффективен как средство, направляющее эволюцион¬
ное изменение?По поводу первого пункта следует кратко заметить, что если в начале
века и существовало несколько примеров наследственности, которые, как
тогда казалось, выходили за общие рамки и нуждались в применении слит¬
ной теории наследственности, то все они в ходе исследований оказались
не только совместимы с корпускулярной наследственностью, но и отчет¬
ливо показали, что именно такова их природа. Кажущееся смешение цвета
кожи в браках белого и черного человека вполне совместимо с представле¬
нием, что эти расы различаются рядом менделевских наследственных фак¬
торов, влияющих на пигментацию. Некоторые из них могут быть с непол¬
ным доминированием, а из оставшихся у некоторых может доминировать
более темный, а у некоторых — более светлый оттенки. Теория Менделя
18Глава 1вполне в состоянии объяснить увеличивающуюся изменчивость потомства
мулатов.Такие биометрические факты, как наследование роста или других па¬
раметров тела человека, хотя на первый взгляд и кажутся несовместимыми
с теорией Менделя, но, как уже было показано, вполне с ней согласуют¬
ся и демонстрируют черты, которые было бы сложно объяснить с любых
других позиций. Приближенно нормальное распределение признаков само
по себе может быть выведено из простого предположения, что эффекты
факторов, влияющих на рост человека, приближенно аддитивны. Корреля¬
ции между родственниками, состоящими в разной степени родства, дости¬
гают, с учетом ошибки выборки, такой величины, которая могла бы быть
выведена из предположения, что признаки определяются преимуществен¬
но наследственностью, и что факторы, контролирующие ее, показывают,
подобно большинству менделевских факторов, полное или почти полное
доминирование. Доминирование — это черта правил Менделя, которая про¬
является в биометрических данных в виде хорошо известного факта, что
дети одних родителей в среднем более похожи друг на друга, чем каждый
из них на своих родителей.До сих пор мы просто констатировали факт отсутствия выходящих
за общие рамки наблюдений, которые могли бы потребовать использования
теории слитной наследственности. Впрочем, существует одна группа совре¬
менных исследований, которые по крайней мере среди изучаемых организ¬
мов исключают ее, похоже, даже как возможность. У некоторых организ¬
мов, у которых обычно происходит самооплодотворение, как впервые пока¬
зал Иоганнсен на примере вида бобов, возможно создание так называемых
чистых линий, в пределах которых наследственная изменчивость, кроме
отдельных мутаций, полностью отсутствует18. Отбор наибольших и наи¬
меньших бобов внутри этих линий, даже там, где этот отбор продолжал¬
ся в течение десяти или двадцати поколений, неизменно давал потомство
с одинаковым средним размером бобов. Размер бобов различался в разных
линиях, показывая тем самым, что внутри вида в изобилии имеется наслед¬
ственная изменчивость, и среди тысяч изученных бобов было найдено два
отчетливых мутанта. Если бы, однако, какая-то ощутимая доля дисперсии
размера бобов относилась к сливающимся элементам, мутации, необходи¬
мые для поддержания подобной наследственной изменчивости, имели бы
возможность в течение десяти поколений обеспечить ее огромным количе¬
ством материала, который неизбежно проявился бы при отборе. Представ¬
ляется, что эксперименты подобного рода способны исключить самое воз¬
можность того, что слитная наследственность может отвечать за сколько-
нибудь ощутимую долю наблюдаемых различий19.
Природа наследственности19Природа и частота наблюдаемых мутацийПредположение, что направление эволюционного изменения в дей¬
ствительности определяется направлением, в котором происходят мутации,
непросто совместить с природой до сих пор наблюдаемых нами много¬
численных мутаций. Ведь большинство из них порождает крайне небла¬
гоприятные деформации, и в действительности большая часть их попро¬
сту летальна. Если мы должны были бы допустить, как это часто предпо¬
лагалось в теоретических исследованиях, что эти мутации указывают на¬
правление эволюции, то эволюционные перспективы маленькой плодовой
мушки Drosophila были бы нерадостны. В равной степени это относится
и к человеку, и к его домашним животным и растениям, если возможно
судить по той частоте, с которой такие рецессивные дефекты, как альби¬
низм, глухонемота и слабоумие, должны были бы происходить в виде мута¬
ций в относительно недавнем прошлом. Создается впечатление, что дефек¬
ты, вызванные мутациями, в равной степени атакуют глаз человека и глаз
Drosophila, и в общем случае мутанты, встречающиеся у пород домашних
животных, часто отличаются страшным видом и наличием дефектов, тогда
как мы знаем много примеров того, что эволюционные изменения, кото¬
рые претерпели эти существа при селекции человека, представляют явное
улучшение.В дополнение к дефектным мутациям, которые привлекают внимание
своей заметностью, мы можем с уверенностью предположить, что проис¬
ходят и другие, менее заметные мутации, которые, по крайней мере в опре¬
деленном окружении или в определенной генетической комбинации, могут
оказаться благоприятными. Однако было бы неразумно считать, что такие
мутации происходят у отдельного индивида с частотой, гораздо большей,
чем та, которая наблюдается при появлении видимых дефектов. Частота
отдельных мутаций у Drosophila редко бывает больше чем 1 на 100000
особей, и мы можем взять это число, чтобы проиллюстрировать неэффек¬
тивность любого средства, которое просто регулирует главное направление
нозникновения мутаций, определяя основной ход эволюционного процес¬
са. Ведь даже в том случае, когда выживание вследствие отбора полно¬
стью отсутствует, для того, чтобы в наследственной природе вида произо¬
шла существенная перемена относительно данного фактора, потребовался
бы отрезок времени порядка 100000 поколений. Более того, если бы му¬
тантный ген столкнулся с мельчайшей селективной невьподой, изменения
прекратились бы на самом раннем этапе. Идеи, необходимые для точно¬
го исследования природы селективного преимущества, будут обсуждаться
в главе 2, но можно легко понять, что если мы говорим о селективном
20Глава 1преимуществе в 1 %, имея в виду, что животные, несущие определенный
ген, будут иметь в перспективе потомство, имеющее только на 1 % боль¬
ше этого гена, чем те, кто несет его аллеломорф, интересующее нас се¬
лективное преимущество будет ничтожно, по крайней мере в том смысле,
что потребуется огромное число подопытных животных и исключитель¬
но точные методы проведения опытов, чтобы суметь показать экспери¬
ментально столь малый эффект, хотя подобное селективное преимущество
способно сильно изменить генетический состав вида — и не за 100000,
а за 100 поколений. Если мы к тому же вообразим, что эти две тенденции
противостоят друг другу, так что мутация, которая постоянно происходит
у одного из 100000 индивидов, будет направлена в сторону, противопо¬
ложную селективному преимуществу величиной в 1 %, можно будет лег¬
ко увидеть, что равновесие будет достигнуто только в том случае, когда
приблизительно 1 индивид из 1 000 будет затронут мутацией. К тому же
это равновесие будет устойчиво, так как если мы вообразим, что по чи¬
стой случайности число мутантов возрастет до более высокой частоты, то
оно тут же начнет уменьшаться под действием отбора, и эволюция пой¬
дет в направлении, противоположном происходящим мутациям, пока ча¬
стота мутантов вновь не достигнет равновесного значения. Таким образом,
для того чтобы мутации определяли ход эволюции, необходимо предполо¬
жить, что скорости мутаций должны быть неизмеримо выше тех, которые,
как мы знаем, происходят в реальности, и составляли бы величину такого
порядка, который в общем был бы несовместим с корпускулярной наслед¬
ственностью.РезюмеМолчаливое согласие со слитной теорией наследственности привело
Дарвина в ходе абсолютно здравых рассуждений к ряду предположений
относительно причин возникновения вариаций и их возможного влияния
на эволюцию. В частности, теория слитной наследственности из-за огром¬
ной частоты мутаций, которая бы потребовалась в этом случае, привела
Дарвина и других исследователей к необходимости связать эволюционную
значимость с гипотетическими факторами, которые контролируют возник¬
новение мутаций. Механизм (менделизм) корпускулярной наследственно¬
сти, предложенный позже, предусматривает во много тысяч раз метшую
частоту мутаций. Эксперименты с «чистыми линиями», похоже, вовсе ис¬
ключают слитную наследственность, даже как дополнительную возмож¬
ность. Природа наблюдаемых мутаций несовместима со взглядом, что эво¬
люция направляется только ими, тогда как наблюдаемая частота их ветре-
Природа наследственности21чаемости показывает, что средство, регулирующее мутации, было бы абсо¬
лютно неэффективно в качестве силы, направляющей эволюцию.Вся группа теорий, которая приписывает гипотетическим физиоло¬
гическим механизмам, контролирующим возникновение мутаций, способ¬
ность придать направленность ходу эволюции, должна быть оставлена
без внимания, коль скоро теория слитной наследственности признана несо¬
стоятельной. Единственная выжившая теория — это теория Естественного
Отбора, и мы не можем не сделать вывод, что если любой эволюционный
феномен окажется необъяснимым этой теорией, нам придется просто при¬
знать, что это — один из тех фактов, который при теперешнем состоянии
науки объяснить невозможно. Исследователь, который может натолкнуть¬
ся на подобный факт как на неизбежное следствие того, что ему в дан¬
ный момент известно о природе наследственности, должен с уверенностью
продолжать свои исследования механизмов отбора, действующих в течение
всей жизни группы особей в их естественной среде обитания, а не выду¬
мывать возможные причины, влияющие на возникновение мутаций. Экспе¬
риментальное изучение механизмов, способных влиять на частоту возник¬
новения мутаций, представляет огромный интерес, поскольку может про¬
лить свет на природу этих изменений. Мы дали бы неправильную оценку
подобным исследованиям, если бы рассматривали их только как способ вы¬
яснения причин эволюционных модификаций.
Глава 2Фундаментальная теорема
естественного отбораТаблица выживания и таблица репродукции. Мальтузианский параметр ро¬
ста популяции. Репродуктивная ценность. Генетическая составляющая дисперсии.
Естественный Отбор. Природа адаптации. Ухудшение среды обитания. Изменения
в популяции. Резюме.Однако, не следует слишком удивляться неосведомленности других
людей, если вы-mo понимаете, что без помощи статистики невозмож¬
но делать какие-либо предположения о продолжительности жизни и про¬
центном отношении уровня смертности к уровню рождаемости в челове¬
ческом обществе. ДАРВИН, 1845 (Life and Letters, ii, 33).Прежде всего говорят — ия также ставлю этот пункт на первое ме¬
сто, поскольку обвинение слишком часто выдвигается самими Физиолога¬
ми, — что Биология отличается от Физико-химических и Математических
наук тем, что она является «неточной». ХАКСЛИ, 1854.Таблица выживанияЧтобы получить четкое представление о применимости Естественного
Отбора ко всем стадиям жизненного цикла организма, можно воспользо¬
ваться идеями, которые возникли в ходе статистического анализа страхова¬
ния людей от смерти20. Сами по себе эти идеи не представляют трудности
для понимания, однако в сочетании с трудоемкими расчетами и технически¬
ми условностями, приня тыми в практике страхового дела, они не настоль¬
ко известны большинству биологов, как это может показаться. Более того,
в руководствах по предмету обсуждаются вероятности смерти и расчеты
денежных выплат, зависящие от этих вероятностей, тогда как для биоло¬
гических проблем необходимо как минимум равное внимание и четкость
представлений о размножении (репродукции), и особенно важно учиты¬
вать взаимодействие выживания и размножения, контролирующих рост или
уменьшение численности популяции.
Фундаментальная теорема естественного отбора23В этой главе мы рассмотрим, насколько согласуются некоторые
представления, возникающие в результате анализа смертности и репро¬
дуктивности в популяции организмов, для которых справедлива кон¬
цепция корпускулярного наследования, чтобы сформулировать принцип
Естественного Отбора в виде строгой математической теоремы, соглас¬
но которой скорость улучшения любого вида организмов в среде его
обитания определяется его состоянием в настоящий момент времени.Основной инструмент сотрудника статистической службы в страховом
деле — это то, что известно как таблица выживания. В ней указана ча¬
стота людей, родившихся живыми и доживших до конкретного возраста.
Например, в таблице выживания может быть отмечено, что до возраста
20 лет доживает 88 % людей, а до 40 лет — только 80 %. Отсюда легко под¬
считать, что 12% родившихся живыми умирают в течение первых 20 лет
и еще 8 % — в течение следующих 20 лет. Таким образом, таблица выжива¬
ния отражает частотное распределение возраста смерти в конкретной попу¬
ляции. Число, на которое каждая запись отличается от предыдущей, соот¬
ветствует числу смертей, имевших место в интервале между отмеченными
возрастами, а если это число разделить на число живых людей предше¬
ствующего возраста, то мы получим вероятность смерти для этой группы
людей за определенный период. Поскольку вероятность смерти непрерывно
изменяется в течение жизни, смертность для конкретного возраста можно
определить только с помощью последовательного учета бесконечно малых
возрастных интервалов. Следовательно, если 1Х — число доживших до воз¬
раста х, то смертность для возраста х составитИх = = -г- (bg 1Х),1Х ах ахгде обычно берется натуральный логарифм. Таким образом, таблица вы¬
живания содержит смертность для всех возрастов, и обратно, ее можно
построить, если известна смертность на протяжении всей жизни. Таков
на практике обычный способ построения таблиц выживания.Нет необходимости обсуждать техническую процедуру, используе¬
мую для построения таблиц выживания, различные правила, применяемые
в этой форме представления данных, а также сложности, которые возни¬
кают при интерпретации данных, получаемых для этих целей из доступ¬
ных источников. Достаточно указать только один момент. Как и при всех
других экспериментальных определениях теоретических величин, практи¬
чески достижимая точность ограничена объемом наблюдений; результат,
полученный по итогам какого-то конечного числа наблюдений, будет за¬
висеть от ошибки выборки, однако этот факт ни в коей мере не делает
24Глава 2такие показатели как смертность или предполагаемая средняя продолжи¬
тельность жизни неясными или неопределенными. Эти показатели отражают
вероятности, средние значения и т.д., свойственные выборке из какой-то
гипотетической популяции, и определяются они только ее природой и усло¬
виями выборки. Неточность наших методов измерения важна для статисти¬
ки смертности не больше, чем в физике, затемняя лишь наше представление
о том, что мы измеряем. Понятия не стали бы менее ясными и в случаях, ес¬
ли бы мы пытались оценивать вероятность гибели некоторого индивида уни¬
кальной генетической конституции или же другого индивида, оказавшегося
под влиянием внезапных и чрезвычайных обстоятельств окружающей среды.Таблица репродукцииНесмотря на то, что таблица выживания содержит весьма обстоятель¬
ные данные, ее недостаточно, чтобы полностью охарактеризовать взаимо¬
действие организма со средой обитания; в ней представлены только шан¬
сы или частоты смерти и нет данных о размножении. Чтобы исправить
это упущение, необходимо ввести вторую таблицу, которая содержала бы
данные о скорости репродукции (репродуктивности) аналогично скорости
смерти (смертности) в любом возрасте. Если для человека, живого в начале
любого бесконечно малого возрастного интервала dx, шанс умереть в тече¬
ние этого интервала равен цх dx, то шанс принять участие в размножении
в этом же интервале будет для него bx dx, где Ъх можно назвать скоро¬
стью репродукции (репродуктивностью) в возрасте х. Если же для челове¬
ка при рождении шанс умереть в течение указанного возрастного интерва¬
ла dx равен lxfix dx, то для этого человека, если он жив, шанс размножения
в течение этого интервала будет 1ХЪХ dx.Так как в размножении участвуют два пола, следует добавить оговор¬
ку при измерении Ъх, чтобы учитывать каждого живого потомка для каж¬
дого из родителей, то есть представляется правильным приписывать каж¬
дому из родителей как бы половину каждого из потомков. Такая оговорка,
очевидно, приемлема для генов, не сцепленных с полом (аутосомные ге¬
ны), для которых вероятность попадания в набор каждого потомка, как из¬
вестно, равна У2. Гены, сцепленные с полом, от гетерогаметных родителей
переходят или не переходят к потомкам соответственно полу последних.
В действительности соотношение полов не является строго равным, но по¬
скольку для полового размножения нужны особи обоих полов, то ясно, что
разница между частотами особей разного пола при рождении должна ком¬
пенсироваться за счет разных вероятностей смертности и репродукции, так
что и в этом случае оговорка также оказывается приемлемой.
Фундаментальная теорема естественного отбора25Подобная условность, удобная в плане приведения в соответствие фор¬
мальных математических формул и биологических фактов, может быть
применена к периоду беременности. Случайно может оказаться, что ре¬
бенок рождается уже после смерти его отца. Детей, рожденных от отцов
возраста х, на практике следует «приписывать» отцам, возраст которых
меньше на 3/4 года. Такие поправки не являются необходимыми для точ¬
ности математического представления фактов, однако, очевидно, удобны
для упрощения формы выражения; таким образом, говоря о человеке, мы
обычно считаем этапы жизни годами от момента рождения. Что касается
других организмов, то переменная х, которая соответствует этому возрасту
у человека, может быть в каких-то случаях более удобна для обозначения
стадии жизненного цикла вне зависимости от хронологического возраста
просто для того, чтобы яснее показать смысл формул, не изменяя при этом
содержания формальных выражений.Мальтузианский параметр роста популяцииЕсли использовать одновременно две таблицы — выживания и репро¬
дукции, то мы можем, по-прежнему не уходя от терминологии популяций
человека, сразу вычислить ожидаемое число потомков каждого новорож¬
денного ребенка. Ожидаемое число потомков для каждого возрастного от¬
резка dx равно lxbx dx, а сумма всех этих элементов на протяжении всей
жизни составит общее ожидаемое число потомков. Математическое выра¬
жение этого:ООJ dx,огде интеграл — от нуля (момента рождения) до бесконечности, то есть охва¬
тывает все возможные возрастные отрезки, в течение которых может иметь
место участие в репродукции. Если в каком-то возрасте участие в репро¬
дукции полностью прекращается, значение Ъх становится равным нулю, что
автоматически приведет к завершению интегрирования.Ожидаемое число потомков показывает, достаточна ли в рассматри¬
ваемой популяции репродуктивность, чтобы более или менее уравнове¬
сить смертность. Если эта величина меньше единицы, то репродуктивностъ
недостаточна, чтобы поддерживать устойчивое существование популяции,
то есть при сохранении такого соотношения, без учета каких-то случайных
колебаний, в любой популяции будет происходить неизбежное снижение
численности, скорость которого также можно рассчитать. Равным образом,
26Глава 2если эта величина больше единицы, то в популяции появляется больше
особей, чем нужно для простого поддержания численности, хотя реально
число особей время от времени может и снижаться.Это заключение станет более четким в количественном выражении,
если отметить, что любой системе смертности и репродуктивности соот¬
ветствует только одна возможная возрастная структура популяции, которая
остается неизменной в течение всего времени действия этой системы. Если
же возрастное распределение сохраняется неизменным, то относительная
скорость увеличения или уменьшения численности для любого возраста
должна быть постоянной; обозначим относительную скорость увеличения
численности то; для случаев снижения численности то становится отри¬
цательной. Тогда при постоянной репродуктивности скорость, с которой
рождения будут происходить в любые периоды, будет увеличиваться про¬
порционально emt. Для любого конкретного отрезка времени, для которого
можно принять t = О, скорость, с которой происходили рождения х лет
назад, будет пропорциональна е~тх, и именно такова была скорость рож¬
дений в то время, когда появились на свет люди, возраст которых сегодня
равен х. Число людей за бесконечно малый возрастной интервал dx тогда
будет пропорционально e~mxlx dx, а из всех родившихся до этого возраста
доживает только часть их, равная 1Х. Таким образом, если число то опре¬
делено однозначно, то и возрастное распределение также оказывается за¬
данным. Однако, зная для каждого возраста число живущих и репродуктив¬
ность, можно рассчитать скорость, с которой рождения происходят в дан¬
ный момент времени, и ее можно приравнять известной скорости рождений
при t = 0. Действительно, вклад в общую скорость роста численности лю¬
дей возрастного интервала dx, должен быть e~mxlxbx dx, а суммирование
по всем возрастам даетооJ e~mxlxbx dx.оПриравняв эту величину к единице, получим уравнение для то, для ко¬
торого существует одно единственное решение. Поскольку е~тх меньше
единицы для всех значений х при положительном то и больше единицы
для всех значений х, когда то отрицательно, то очевидно, что то, доводя¬
щее интеграл до единицы, должно быть положительным, если ожидаемое
число потомков превышает единицу, и отрицательным, если оно меньше
единицы.Число то, которое удовлетворяет этому уравнению, таким образом,
в неявном виде содержится в любой системе смертности и репродуктив¬
ности и показывает относительную скорость увеличения или уменьшения
Фундаментальная теорема естественного отбора27численности популяции, находящейся в устойчивом состоянии, свойствен¬
ном любой подобной системе. С учетом акцента, который Мальтус сделал
на «законе геометрического прироста», т можно назвать мальтузианским
параметром роста численности популяции. Очевидно, что при отрицатель¬
ных значениях эта величина представляет столь же хорошую меру умень¬
шения численности популяции, то есть охватывает и те случаи, на которые,
применительно к человечеству, Мальтус обратил слишком мало внимания.Учитывая близкую аналогию популяционного роста, который, как
предполагается, происходит согласно закону геометрического роста, и при¬
роста капитала, инвестированного под сложные проценты, имеет смысл
отметить, что если рассматривать рождение ребенка как предоставление
ему «займа» жизни, а рождение его потомков — как последующую выпла¬
ту долга, то метод вычисления т показывает, что это эквивалентно отве¬
ту на вопрос: при какой процентной ставке выплаты будут эквивалентны
размеру ссуды? Поскольку разовое вложение предполагает возврат вели¬
чины lxbx dx в течение отрезка времени dx, то при размере процентной
ставки, равном т, настоящий размер выплаты составит e~mxlxbx dx; следо¬
вательно, мальтузианский параметр роста популяции соответствует разме¬
ру процентной ставки, при которой реальный показатель ожидаемого числа
рождений потомков в момент рождения их родителя равен единице. Однако
наблюдаемые значения параметра роста популяции, даже в районах с ред¬
ким населением, похоже, не достигают величины зарабатываемых при вло¬
жении денег процентных ставок, а отрицательные ставки, я думаю, и вовсе
не имеют места в коммерции.Репродуктивная ценностьАналогия с деньгами тем не менее проясняет доводы в пользу дру¬
гого простого применения одновременно смертности и репродуктивности.
Можно спросить не только о новорожденных, но и обо всех людях лю¬
бого конкретного возраста, каково число их будущих потомков, и если
эта величина рассчитывается так, как было показано ранее, вопрос при¬
обретает определенный смысл: какова в среднем степень участия особей
данного возраста среди потомков будущих поколений? Такой вопрос пред¬
ставляет некоторый интерес, так как прямое действие Естественного От¬
бора должно быть пропорционально этому вкладу. Без сомнения, будут
оказывать влияние и непрямые воздействия в тех случаях, когда живот¬
ное способствует или препятствует выживанию или размножению своих
родственников; как, например, кормящая мать помогает выживанию своего
ребенка, как у человека мать, родившая и выкормившая детей, может ак¬
28Глава 2тивно помогать их дальнейшему участию в размножении, как беременность
и в несколько меньшей степени детеныши-сосунки препятствуют следую¬
щему оплодотворению, и наиболее удивительный пример — то, как беспо¬
лые насекомые-рабочие обслуживают свою королеву. Однако такие непря¬
мые эффекты в очень многих случаях оказываются не особенно важными
по сравнению с эффектами непосредственного участия в размножении,21 и,
по аналогии с расчетом сложных процентов, величина будущего потомства
особей возраста х, как легко видеть, рассчитывается согласно уравнениюДля каждой возрастной группы можно определить соответствующую
ей репродуктивную ценность. На рис. 2 показаны значения показателя
для женщин по возрастным группам, рассчитанные на основании дан¬
ных о смертности и репродуктивности, зарегистрированных в Австралии
к 1911 г. Мальтузианский параметр в это время был положительным, и су¬
дя по значениям показателя для женщин, был примерно равен размеру
сложных процентов в 1 %; значения показателя для мужчин и совместно
для обоих полов должны быть ниже из-за избытка мужчин среди иммигран¬
тов. Репродуктивная ценность, которую нельзя смешивать со скоростью ре¬
продукции, достигает максимума в 18,5 лет, несмотря на перенос появле¬
ния потомства на более поздние сроки вследствие сложившегося в разви¬
том обществе обычая; на самом деле максимум был бы уже в 16 лет, если
бы не тот факт, что положительный процент прироста придает большую
значимость появлению потомства в скором будущем у женщин старшего
возраста по сравнению с более поздним появлением детей у молодых. Если
именно так обстоит дело в обществе, ни в коей мере не заинтересованном
в раннем воспроизводстве, интересно, что в обществе, приветствующем
заключение браков в период полового созревания или сразу же после его
наступления, возраст максимальной репродуктивной ценности приходится
на любой период начиная с 12 лет. Согласно данным по Австралии, репро¬
дуктивная ценность при рождении ниже отчасти вследствие эффекта роста
популяции, определяющего более низкую ценность поздних детей, и отча¬
сти из-за риска смерти до достижения репродуктивного возраста. Значения
репродуктивной ценности, показанные на рисунке 2, вероятно, правильны,
за исключением измерений после 1911 года, так как для уточнения этих
значений необходимо учитывать иммигрантов с их младенцами (естествен¬
но, не принимая во внимание вопросы евгенического качества), посколь¬
ку младенцы, как правило, переселяются вместе со своими родителями;ООX
Фундаментальная теорема естественного отбора29но это в некоторой степени переоценивается с точки зрения Естественного
Отбора из-за вероятности появления новых детей у родителей, потерявших
грудного ребенка. Репродуктивная ценность более зрелой женщины, напро¬
тив, недооценивается только в том случае, когда ее заработок или работа
по дому являются более выгодными; и это в большей мере отражает точ¬
ку зрения государства, чем Естественного Отбора. По всей видимости, это
обстоятельство в определенной степени объясняет то, что кривая смертно¬
сти у мужчин обратна ходу кривой репродуктивной ценности. Минимум
на кривой смертности равен 12, что совсем близко к начальному макси¬
муму на кривой репродуктивной ценности; она поднимается более круто
для младенцев и менее круто — для людей старшего возраста, чем пада¬
ет впоследствии кривая репродуктивной ценности, точки на которой ка¬
чественно показывают, чего следует ожидать, если падение естественной
смертности в значительной мере обусловлено эффектами дифференциаль¬
ного выживания.Особенность, которая хорошо иллюстрирует значение метода оцен¬
ки, согласно которому людей не считают одинаковыми в будущей попу¬
ляции, а присваивают индивидам каждого возраста соответствующий по¬
казатель vx, состоит в том, что при любом возрастном составе популяции
ее суммарная репродуктивная ценность будет увеличиваться или снижаться
в соответствии с мальтузианским параметром то, тогда как считать всех ин¬
дивидов равными можно лишь в теоретическом случае — когда популяция
находится в своем устойчивом состоянии. Предположим, что число людей
возрастного интервала dx равно пх dx; тогда ценность каждого индивида
в популяции будет nxvx dx; что касается каждой такой группы, будет иметь
место приращение репродуктивной ценности в ходе размножения со скоро¬
стью nxbxvo dx, снижение из-за смертности составит пхцхух dx, а потеря
за счет снижения репродуктивной ценности будет равна — пх dvx; так что
в итоге формула будет иметь видW'x^ipx'vо l-Lx'Ux) dx т~ dvx},дифференцируя уравнение, по которому определяют vx, получим1 dvx 1 dlx Ьх v0 ; Н ; тп = -т, = ,vx ах 1Х dx _e-mx vxvoилиdvx — цхух dx + bxvо dx = mvx dx.Следовательно, скорость прироста общей численности популяции т-крат-
на ее реальной общей численности, независимо от ее возрастного состава.
30 Глава 2возраст (в годах)Рис. 2. Репродуктивная ценность для австралийских женщин. Репродуктивная цен¬
ность на женщину вычислена по данным о частотах рождения и смерти в Австралии
в 1911 году. Мальтузианский параметр равен +0,01231 в годТаким образом, сравнение общей численности популяции для двух пери¬
одов переписи показывает, с поправкой на миграцию, реальный биологи¬
ческий прирост или убыль популяции, которые могут быть совершенно
невразумительными или ограниченными, если сравнивать непосредствен¬
но подсчеты численности. Например, население Великобритании должно
было начать сокращаться биологически с какого-то момента времени, меж¬
ду 1911 и 1921 гг. (точное его начало нельзя установить из-за начавшей¬
ся в 1914 году войны), однако перепись 1921 года показала номинальный
прирост в несколько миллионов, тогда как перепись 1931 года почти несо¬
мненно покажет дальнейший фальшивый период роста из-за увеличения
числа людей таких возрастов, для которых их репродуктивная ценность со¬
вершенно незначительна.
Фундаментальная теорема естественного отбора31Генетическая составляющая дисперсииТеперь обратимся к тому, каким образом любой количественный при¬
знак, например рост человека, может зависеть от генетической структуры
особи22. По отношению к любой паре альтернативных генов можно пред¬
ставить популяцию в виде двух частей, в каждую из которых входит один
тип гомозигот и половина гетерозигот, которые должны быть поделены по¬
ровну между обеими частями. Разность среднего роста этих двух групп
людей тогда можно назвать средним эффектом (по росту), связанным с рас¬
сматриваемым генным замещением. Такая разность не определяется только
одним геном, по которому обе группы различаются, но, вероятно, статисти¬
чески связана и с участием других генов, проявляющих как сходный, так
и противоположный эффект. Такое определение окажется более адекват¬
ными, если (что необходимо для точности) популяция охватывает не про¬
сто совокупность в целом, представленную одним поколением, достигшим
зрелости, но и должна, согласно нашему допущению, включать все возмож¬
ные генетические комбинации, частоты которых соответствуют реальным
вероятностям их появления и выживания, каковы бы они ни были, и ес¬
ли среднее значение получено по всем значениям роста, созданным всеми
этими генотипами во всех возможных условиях среды обитания, то часто¬
ты эти тоже должны соответствовать реальным вероятностям этих условий.
Тогда статистическое понятие эффекта роста для данного генного замеще¬
ния будет величиной точной, не зависящей от случайности, как было бы
в каждом конкретном случае оценки, но зависящей только от генетиче¬
ской природы и условий обитания вида. Эффект для любого фактора обыч¬
но зависит от реального соотношения частот альтернативных генов р : q,
на него может также влиять отклонение от случайного скрещивания, а так¬
же различные варианты взаимодействия разных факторов; именно поэтому
для целей нашего обсуждения мы сформулировали определение в точных
статистических терминах, а не в терминах средних величин, соответству¬
ющих чистым генотипам, что больше подошло бы для определения этого
показателя в экспериментальной популяции, в которой условия скрещи¬
вания контролируются, а число разных изучаемых генотипов выбирается
экспериментатором.По той же причине необходимо также дать статистическое определе¬
ние еще одной величины, которую легко спутать с той, что мы только что
обсуждали, да и значения их часто могут быть почти одинаковыми, однако
нужно уметь различать их, причем достаточно аккуратно; речь идет о сред¬
нем эффекте, возникающем в популяции определенного генетического со¬
става при замещении одного гена другим. При любых типах скрещивания
32Глава 2и, следовательно, любой частоте разных комбинаций генов может происхо¬
дить замещение некоторой части альтернативных форм генов, что повлечет
за собой определенный сдвиг показателя среднего роста. Такой сдвиг ве¬
личины в среднем относительно общего роста для популяции для каждо¬
го генного замещения можно назвать средним эффектом генного замеще¬
ния — в отличие от среднего эффекта, как было показано выше. В примере
с ростом людей корреляция, которая, как показано, существует между су¬
пругами, достаточна, чтобы обеспечить результат, когда каждый из генов,
как-то влияющих на увеличение роста, должен «объединять» свое действие
со всеми прочими генами со сходным влиянием так, чтобы средний эффект
в результате каждого генного замещения превышал средний эффект при¬
мерно на четверть.Если о — величина среднего эффекта по некоторому фактору, и а —
величина среднего эффекта для данного измерения, то можно показать, что
вклад этого фактора в генетическую дисперсию будет представлен выраже¬
нием pqaa.Измеряемую величину обозначим х, и отношение значений а к ней
можно сделать более понятным, если представить, что для каждой специ¬
фической генной комбинации мы рассчитываем «ожидаемую» величину X,
добавляя соответствующие приращения, положительные или отрицатель¬
ные, в зависимости от природы соответствующих генов. Это ожидаемое
значение не обязательно покажет реальное приращение, хотя и может при¬
ближаться к нему достаточно близко, но его статистические особенности
будут более точно учтены при наследовании реального роста, чем свойства
самой переменной. Поскольку мы имеем дело только с изменчивостью,
можно принять в качестве исходной составляющей величины X среднее
значение х в популяции и подобрать имеющиеся положительные и отрица¬
тельные приращения для каждого фактора таким образом, чтобы они урав¬
новешивали друг друга, когда речь идет о популяции в целом. Приращение,
обусловленное действием одного гена, проявится во всей популяции р раз,
а действием альтернативного гена — q раз, так что два приращения должны
быть противоположных знаков в отношении q : (—р). Более того, поскольку
разность между ними должна равняться а, то реальные значения не могут
быть иными, кроме как qa и (—ра) соответственно.Значение среднего эффекта а при любом генном замещении получаем
сравнением средних значений х в двух частях, на которые можно подразде¬
лить популяцию. Очевидно, что значения а будут определяться правильно
только при условии, что одна и та же разность средних имеет место в обеих
частях популяции, то есть сумма отклонений х — X будет равна нулю. Это
дает критерий, математически достаточный для определения значений а,
Фундаментальная теорема естественного отбора33которые представляют для рассматриваемой популяции средние эффекты
генных замещений. Отсюда следует, что для популяции в целом сумма про¬
изведений Х(х — X) для каждого индивида должна быть равна нулю, так
как каждое значение qa или (—ра) прежде всего умножается на ноль, и это
будет верно для эффектов каждого фактора отдельно. Отсюда следует, что
если теперь X и х сравнивать со средним значением для популяции, то
дисперсия X, которая представляет среднее значение для X2, будет рав¬
на среднему значению Хх. Теперь средняя величина Хх будет включать
в себя а для каждого менделевского фактора; так как X будет содержать
член qa для р индивидов из одной части и член (—ра) для q индивидов
другой части, а средние величины х в этих частях различаются на вели¬
чину а, то среднее значение Хх должно быть равно сумме значений pqaa
по всем факторам. Таким образом, мы показали, что дисперсия для X будет
равнаW =где суммирование проводится по всем факторам, и эту величину можно
рассматривать как генетическую дисперсию для измерения ж.23 То, что она,
по определению, положительна, за исключением тех случаев, когда эффект
каждого гена в отдельности равен нулю, следует из ее равенства диспер¬
сии X. Расширение этого анализа без изменения принципиального подхода
позволяет получить аналогичное выражение для тех случаев, когда один
или более факторов представлены более чем двумя разными генами или
аллеломорфами.Применимость термина генетическая дисперсия определяется тем, что
величина X задается только генами, которые представлены у индивида,
и слагается из средних эффектов этих генов. Таким образом, она отража¬
ет генетический потенциал конкретного индивида в совокупности с реаль¬
ными возможностями скрещивания в том смысле, что средние показатели
потомков (как х, так и X) двух самцов, скрещивающихся с идентичными
самками, будут различаться точно наполовину от тех различий, которые
имеют их родители. Относительные генетические величины поэтому мож¬
но определить экспериментально с помощью диаллельного метода, при ко¬
тором каждая изучаемая особь скрещивается с особями противоположного
пола из одной и той же линии так, чтобы получить большое число потомков
от каждого такого скрещивания. Без вычисления индивидуальных значений
генетическую дисперсию популяции можно вывести из корреляций между
родственниками при условии, что корреляции определены аккуратно. Ес¬
ли квадрат корреляции родители-дети разделить на корреляцию (бабушки,
дедушки) - внуки, это дает хорошую оценку компоненты общей наблюдае¬
34Глава 2мой дисперсии признака, которую можно рассматривать как генетическую
дисперсию.Ясно, что получаемые для индивидов реальные измерения, х, могут
отличаться от генетически ожидаемых значений из-за флуктуаций условий
окружающей среды. Следует отметить, что это не единственная причина
различий; даже если никаких флуктуаций среды не происходит, то есть
реальные значения определяются только генетической структурой, эти ре¬
зультаты измерений, которые можно рассматривать как генотипические, все
равно могут отличаться от генетических значений, X. Хороший пример
этого — доминирование, так как при полном доминировании генотипиче¬
ское значение гетерозиготы будет точно таким же, как у соответствующей
доминантной гомозиготы, но генотипы их различаются по одному генно¬
му замещению, что может изменить генетический потенциал, представлен¬
ный X, отразившись на средних значениях признаков в потомстве. Похожая
причина различий возникает, если генные замещения по разным факторам
не строго аддитивны по своим средним эффектам. Согласно данному здесь
определению генетическая дисперсия есть лишь часть генотипически опре¬
деленной дисперсии, и она будет отличаться, обычно в сторону некоторого
занижения, от общей наблюдаемой дисперсии.Следовательно, не будет излишним дать определение чистой генетиче¬
ской компоненты дисперсии, которая объективно существует, как статисти¬
ческой характеристики популяции, в отличие от дисперсии, полученной из
непосредственных измерений.Естественный ОтборПриведенные выше определения могут быть использованы для любых
признаков; особенно интересно было бы применить их к особой характери¬
стике т, которая измеряет относительную скорость увеличения или умень¬
шения численности. Две группы индивидов, несущие альтернативные гены,
неизбежно будут проявлять равные или неравные скорости увеличения чис¬
ленности, и разность между соответствующими значениями т будет рав¬
на а, подобным же образом среднее изменение величины т при генном за¬
мещении будет равно а. Поскольку величина т показывает приспособлен¬
ность по выживанию через объективный факт представленности в будущих
поколениях, то величина pqaa будет соответствовать вкладу каждого фак¬
тора в генетическую дисперсию приспособленности; общая генетическая
дисперсия приспособленности при этом равна сумме этих составляющих,
которая всегда положительна или в крайнем случае равна нулю. Далее, вся¬
кое увеличение величины dp по частоте одной альтернативной формы гена
Фундаментальная теорема естественного отбора35за счет другой будет сопровождаться увеличением a dp в средней приспо¬
собленности вида, где а, конечно, может быть и отрицательной; однако,
исходя из определения а, отношение р : q обязательно должно увеличи¬
ваться в геометрической прогрессии со скоростью а, что в математическом
выражении будет иметь видчто можно записать как( — + — I dp = a dt
\Р Ч)илиdp — pqadt,откуда следует, чтоa dp = pqaa dt,и с учетом всех факторов общее приращение приспособленности составит
a dp) = y^(ffgaa) dt = W dt.Следовательно, если время dt положительно, то и суммарное измене¬
ние приспособленности W dt также имеет положительный знак, и в дей¬
ствительности скорость увеличения приспособленности вследствие всех
изменений отношения генных частот в точности равна генетической дис¬
персии приспособленности W, которая характеризует популяцию. Таким
образом, мы можем сформулировать фундаментальную теорему Естествен¬
ного Отбора в следующем виде.Скорость увеличения приспособленности любого организма24 за лю¬
бое время равняется генетической дисперсии приспособленности в данный
момент.Точность доказательства требует, чтобы термины использовались стро¬
го в соответствии с определением; интерпретацию можно упростить, если
усилить соответствующие правила измерения. Например, отношение р : q
в каждом случае должно строго оцениваться с помощью перечисления —
не обязательно переписи всей популяции, но всех индивидов, имеющих
репродуктивную ценность, с учетом относительной репродуктивной цен¬
ности каждого.Теорема точна только в идеальных популяциях, где исключены случай¬
ные флуктуации генетического состава, поэтому важно оценивать величину
36Глава 2влияния этих флуктуаций или, другими словами, вычислять стандартную
ошибку, соответствующую ожидаемой скорости увеличения приспособлен¬
ности. Для этой цели достаточно рассмотреть особый случай популяции,
в которой скрещивание и размножение происходят случайным образом.
Легко видеть, что если за счет таких случайных флуктуаций создается раз¬
ность 8р между ожидаемым значением и реальной величиной р, получен¬
ной для некоторого поколения, то дисперсия для 5р будет составлятьPQ2п ’где п — число скрещивающихся особей в каждом поколении, а 2п — число
генов у п особей, которые участвуют в воспроизводстве. Дисперсия прира¬
щения приспособленности, а8р, в таком случае будет равнаи отсюда, при случайном скрещивании, флуктуация для разных отношений
генных частот будет независимой, а величины а и а перестанут разли¬
чаться, так что при этом условии скорость увеличения приспособленности
в расчете на одно поколение будет иметь стандартную ошибку, как резуль¬
тат случайного выживания, выражаемую следующим образом:где Т — продолжительность поколения. Обычно бывает удобно для каждого
организма измерять время в поколениях, и в этом случае станет очевид¬
ным (2п велико), что случайные флуктуации W, даже в расчете только
на одно поколение, скорее всего будут очень малы по сравнению со сред¬
ним. Регулярность последнего фактически гарантирована тем же обстоя¬
тельством, которое заставляет статистическую совокупность частиц типа
пузырьков газа подчиняться без заметных отклонений газовым законам. Ви¬
димый пузырек в действительности состоит из нескольких миллиардов мо¬
лекул, что является сравнительно большим числом для органической попу¬
ляции, однако принцип системы тот же. При точной интерпретации форму¬
ла показывает, что только когда скорость изменения W (время измеряется
в поколениях) настолько мала, что становится сравнимой с величиной 1 /п,
тогда скорость изменения в последовательных поколениях становится ирре¬
гулярной. Даже если бы установилось равновесие на этом уровне точности
между показателями смертности и репродукции разных генотипов, только
Фундаментальная теорема естественного отбора37скорость изменения на отрезках времени продолжительностью в одно по¬
коление могла бы оказаться иррегулярной, тогда как на отрезках порядка
10000 поколений отклонение от регулярности оказалось бы в сотни раз
меньше.Следует отметить, что доказанная выше фундаментальная теорема об¬
наруживает по ряду моментов удивительное сходство со вторым законом
термодинамики. Оба утверждения характеризуют свойства популяций или
совокупностей, по большому счету не зависящие от природы составляю¬
щих их единиц; оба отражают статистические закономерности; в обоих
случаях обязательно происходит постоянное повышение измеряемой вели¬
чины: в одном случае — энтропии физической системы, в другом — при¬
способленности биологической популяции (т). Для физического мира мы
можем теоретически представить себе системы, в которых полностью от¬
сутствуют центробежные силы, так что энтропия сохраняется в них на по¬
стоянном уровне; также мы можем представить себе (хотя не стоит наде¬
яться найти такие в реальности) и биологические популяции, в которых
генетическая диперсия равна нулю и в которых приспособленность не по¬
вышается. Профессор Эддингтон недавно заметил: «Закон, согласно кото¬
рому энтропия всегда возрастает,— второй закон термодинамики,— я ду¬
маю, занимает высшее место среди законов природы». Весьма поучитель¬
но, что столь сходный закон в биологических науках тоже должен был бы
занимать высшее место. Возможно, что в конечном счете оба они когда-
то будут поглощены неким более общим принципом, поэтому сегодня мы
должны отметить, что между ними просматриваются и глубокие разли¬
чия, а именно: (1) системы, изучаемые термодинамикой, постоянны, тогда
как виды, напротив, могут и вымирать, хотя предполагается, что биоло¬
гическое улучшение их не прекращается в течение всего времени их су¬
ществования. (2) Приспособленность, хотя и измеряется единым методом,
качественно различается для каждого конкретного организма, тогда как эн¬
тропия, подобно температуре, принята в едином смысле для всех физиче¬
ских систем. (3) Приспособленность может повышаться или понижаться
под влиянием изменений во внешней среде, не оказывая количественного
влияния на эту среду. (4) Изменения энтропии представляют исключение
в физическом мире в силу своей необратимости, тогда как необратимые
эволюционные изменения не составляют никакого исключения среди био¬
логических явлений. Наконец, (5) изменения энтропии ведут к нарастанию
беспорядка в физическом мире, по крайней мере с точки зрения людей
на использование энергии, тогда как эволюционные изменения обычно счи¬
таются направленными в сторону повышения организации в органическом
мире.
38Глава 2Представление принципа Естественного Отбора в виде теоремы, опре¬
деляющей скорость прогресса вида в повышении приспособленности
по выживанию (этот термин использован для четко сформулированного
статистического признака популяции) и одновременно зависимости между
этой скоростью и ее стандартной ошибкой, вынуждает нас высказать свое
отношение к обоснованности возражения, что принцип Естественного От¬
бора зависит от удачного стечения благоприятных случайностей. Это возра¬
жение больше относится к области косвенных намеков, а не реальной кри¬
тики, поскольку в основу положена двусмысленность слова «случайность»
в его обывательском смысле. Доход, получаемый владельцем от деятельно¬
сти казино, можно сказать, в каком-то смысле зависит от последователь¬
ности удачных случайностей, хотя фраза содержит в себе предположение
о невероятности, более свойственное надеждам постоянных посетителей
этого заведения. Можно легко и без особо глубокого логического анали¬
за понять разницу между последовательностью благоприятных отклонений
от законов вероятности и кумулятивным действием этих законов. Именно
на последнем базируется действие принципа Естественного Отбора.2'5Природа адаптацииЧтобы в общих чертах рассмотреть последствия для органического ми¬
ра прогрессивного повышения приспособленности каждого вида организ¬
мов, нужно сначала обсудить трудную для понимания природу зависимо¬
сти, которую мы называем «адаптацией». Это тем более необходимо по¬
тому, что любой простой пример адаптации, как например, длинные шея
и ноги жирафа, которые считаются приспособлением, позволяющим объ¬
едать листья высоко на ветках, или соответствие обычной окраски живот¬
ного краскам окружающей его природной среды, при всей простоте фор¬
мулировок упускает значительную долю смысловой нагрузки этого сло¬
ва. Ибо чем более совершенна адаптация, чем больше приспособительных
особенностей и черт сходства, тем выше считается уровень адаптивности.
Организм считается адаптированным к конкретной ситуации или же ко все¬
му разнообразию ситуаций, задаваемых ему окружающей средой лишь на¬
столько, насколько мы можем себе представить совокупность нескольких
различных ситуаций или условий окружающей среды, к которым животное
оказалось бы адаптировано немного хуже, и, равным образом, насколько
мы можем вообразить совокупность слегка различающихся органических
форм, которые были бы хуже приспособлены к этой среде. Именно такой
смысл, как я полагаю, должно выражать это слово, оставив совершенно
в стороне вопрос о том, действительно ли организмы адаптированы к сво¬
Фундаментальная теорема естественного отбора39им местообитаниям, или структуры и инстинкты, к которым термин был
применен, просто описаны соответствующим образом.Необходимые статистические требования к ситуации, в которой осу¬
ществляется подгонка одного существа к другому с учетом большого чис¬
ла разных аспектов, можно проиллюстрировать геометрически. Степень
сходства можно представить как близость точки А к фиксированной точ¬
ке О. В трехмерном пространстве мы можем представить сходство только
по трем измерениям, но даже на этом примере можно понять суть ситуа¬
ции. Возможные лучшие адаптации из всех, которыми обладает А, будут
вписаны в сферу, проходящую через А, с центром О. Если А смещает¬
ся на какое-то фиксированное расстояние г, то ее перемещение в любом
направлении внутри сферы будет означать улучшение адаптации, но но¬
вое положение точки вне сферы указывает на ухудшение адаптации. Ес¬
ли г очень мало — это можно воспринимать как примерное равенство шан¬
сов осуществления этих двух событий, а вероятность улучшения стремится
к пределу У2 по мере приближения значений г к нулю; однако если г при¬
ближается по размеру к диаметру сферы или превышает его — это значит,
что любое улучшение невероятно, так как все точки внутри сферы теперь
ближе, чем расстояние до А. Для любых значений г между этими предель¬
ными величинами вероятность улучшения будет равнагде d — диаметр сферы.Таким образом, шанс улучшения постоянно убывает от предельного
значения У2 при г, равном нулю, и до нуля, когда величина г достигает d.
Поскольку А в нашем изложении может обозначать и организм, и его среду
обитания, мы можем сделать вывод, что если изменение, причем с любой
стороны, очень мало, то оно имеет почти равные шансы оказаться благо¬
приятным или неблагоприятным; в то же время при более крупных измене¬
ниях вероятность улучшения постоянно уменьшается, доходя практически
до нуля, если изменения достаточно хорошо заметны.Трехмерное представление, конечно, не является адекватным, так как
даже один орган, в тех случаях, когда мы знаем достаточно, чтобы иметь
возможность оценить связь между структурой и функцией, как, например,
глаз у позвоночных, часто демонстрирует такое соответствие по значитель¬
но большему, чем три, числу параметров. Если бы наши геометрические
возможности допускали представление большего числа измерений, то фор¬
ма зависимости между величиной изменения г и вероятностью некоторого
улучшения приняла бы вид кривой на рис. 3. Первичные выводы из трех¬
40Глава 2мерного подхода сводятся к тому, что вероятность улучшения при очень
малых масштабах изменений стремится к значению У2 и быстро падает
при более заметных изменениях, достигая чрезвычайно малых величин,
когда число измерений велико, даже если г по-прежнему мало по срав¬
нению с d.Рис. 3. Соотношение между величиной ненаправленного изменения и вероят¬
ностью лучшей адаптации, когда размерность пространства (га) велика: р —1 °°= J е 2 dt, х = ry/n/d\ 27Г хДля любого уровня адаптации будет существовать стандартная вели¬
чина изменений, равная d/л/п, а вероятность благоприятного изменения
можно определить как отношение этого изменения к стандартной вели¬
чине. Чем выше адаптация, тем меньше будет эта стандартная величина,
и, следовательно, тем меньше вероятность того, что изменение такой ве¬
личины может оказаться благоприятным. Иным способом ситуацию можно
изобразить, предположив, что изменения определенного масштаба проис¬
ходят случайным образом во всех направлениях, и сравнивая скорости эво¬
люционного прогресса, причиной которых являются два противоположных
селективных воздействия, одно из которых отбирает и аккумулирует все
изменения, повышающие адаптацию, а другое, напротив,— все, снижаю¬
щие адаптацию. По отношению к очень малым изменениям (относитель¬
но стандарта) эффективность этих воздействий будет практически равной,
однако для значений порядка всего лишь одной десятой от стандарта, де¬
структивный отбор уже на 28 % эффективнее, чем отбор благоприятству¬
Фундаментальная теорема естественного отбора41ющий. При величине около У2 от стандарта он становится эффективнее
уже в 3,5 раза; при равенстве стандарту, как видно из диаграммы, веро¬
ятность улучшения составляет примерно г/е, тогда как варианты, ослаб¬
ляющие адаптации, отбираются в 13 раз чаще; соответственно, при двух-
и трехкратном превышении стандартной величины эти показатели будут
равны 236 и 7 852.Согласованность этих статистических выкладок с обычной практикой
можно понять, если привлечь для сравнения пример с механической адап¬
тацией такого инструмента, как микроскоп, когда он отрегулирован для от¬
четливого рассматривания объекта. Представим, как нарушается изображе¬
ние: если слегка сдвинуть каждую из линз, все равно —• вдоль или попе¬
рек, или изменить угол отражения или преломления, изменить кривизну
или нарушить полировку поверхностей, то будет достаточно очевидно, что
сколько-нибудь существенное изменение вряд ли может улучшить настрой¬
ку, тогда как если вносимое изменение очень мало, намного меньше тех,
которыми пользуется настройщик, то шанс достичь улучшения будет равен
почти У2.Ухудшение среды обитанияТаким образом, если организм действительно достаточно хорошо адап¬
тирован к тому месту, которое он занимает в среде обитания, то этой адап¬
тации постоянно угрожают всевозможные ненаправленные воздействия,
способные вызвать какие-то изменения адаптивной системы. Прежде всего
обсудим случай с крупной мутацией, так как ее последствия будут очень
просты. К настоящему времени зафиксированы многочисленные случаи та¬
ких мутаций, и, насколько мне известно, все они без исключения либо
несут явную патологию (чаще всего летальную) по своему проявлению, ли¬
бо с весьма высокой вероятностью должны были бы считаться вредными
в дикой природе. Это как раз то, что предполагали, считая очевидным, ста¬
рые натуралисты, а также, я думаю, и все те, кто изучает диких животных:
организмы в целом действительно изумительно и очень тонко адаптирова¬
ны к своему внешнему окружению как по своей внутренней организации,
так и с точки зрения взаимодействия с другими организмами. Происходя¬
щие в природе крупные мутации оказываются неблагоприятными для вы¬
живания, и по мере появления в популяции некоторого небольшого числа
особей с такими нарушениями постепенно установится равновесие, когда
скорость элиминации сравняется с частотой мутирования. Иными слова¬
ми, можно сказать, что каждая мутация такого рода может реально вносить
точно такой вклад в генетическую дисперсию приспособленности вида, ка¬
42Глава 2кой обеспечивает скорость улучшения, эквивалентная скорости ухудшения,
обусловленной постоянно происходящими мутациями.Что же касается физических факторов окружающей среды, то мед¬
ленные геологические и климатические изменения происходят в природе
постоянно, и, возможно, они оказываются благоприятными для некоторых
организмов, однако для большего числа обитателей они со временем ока¬
зываются вредными. Таким образом, можно считать, что для большинства
организмов физическая среда постоянно ухудшается так или иначе, климат,
например, становится то теплее, то холоднее, то более влажным, то сухим,
и это приводит у большинства видов к повышенному понижению значе¬
ния ш, мальтузианского параметра роста популяции. Вероятно, еще более
важными, чем изменения климата, будут эволюционные изменения, проис¬
ходящие в сообществе у других организмов. Каждый организм увеличивает
свою приспособленность, то же имеет место и у его врагов и конкурен¬
тов; в результате будет наблюдаться все тот же эффект, но, возможно, еще
более выраженный,— ухудшение среды обитания, причем с точки зрения
каждого из участвующих организмов. Против действия Естественного От¬
бора, постоянно работающего на повышение приспособленности каждого
организма со скоростью, равной генетической дисперсии приспособленно¬
сти, которую поддерживает конкретная популяция, должен быть запущен
очень существенный уравновешивающий фактор, разрушающий окружа¬
ющую его неорганическую и органическую среды. Только в том случае,
когда первое из этих воздействий будет перекрывать второе, может иметь
место какой-либо реальный прирост популяции; в противном случае будет
происходить убыль численности популяции.Изменения в популяцииРост численности любых организмов будет ухудшать окружающую их
среду аналогично тому и, вероятно, в еще большей степени, чем увеличе¬
ние численности или эффективности воздействия его конкурентов. Явным
упрощением является утверждение, что среда определяет численность ор¬
ганизмов каждого вида, которую она будет поддерживать. В действитель¬
ности эти численности должны определяться как бы эластичностью сопро¬
тивления, которое возникает при увеличении численности, так что жизнь
каждого индивида становится несколько труднее, когда популяция растет,
и становится легче при меньшей численности. Вопрос о балансе отклады¬
вается, если из скорости прироста средней величины т, созданной Есте¬
ственным Отбором, вычесть скорость уменьшения, обусловленного ухуд¬
шением среды обитания; при этом результат заключается не в увеличе¬
Фундаментальная теорема естественного отбора43нии средней величины ш, которая не может быть существенно выше нуля,
но главным образом в том, что наблюдается постоянное увеличение чис¬
ленности популяции.Ситуацию можно представить в виде дифференциального уравнения:dM М тгг „-ir + -c=w-D'где М — среднее значение мальтузианского параметра; С — константа, отра¬
жающая зависимость между приспособленностью и приростом популяции
и определяемая как увеличение натурального логарифма численности по¬
пуляции, которая считается постоянной на каждой стадии при единичном
приросте величины М; W — скорость реального увеличения приспособ¬
ленности, обусловленной Естественным Отбором, a D — скорость убыли
вследствие ухудшения среды обитания. Если C,W и D — константы, урав¬
нение имеет решение26М = С ■ (W - D) + Ае~г/С,где А — некоторая константа, зависящая от начальных условий. С представ¬
ляет физическое измерение времени и потому может задаваться в годах или
поколениях; уравнение показывает, что если С, W и D остаются постоян¬
ными в течение какого-то отрезка времени, значительно превышающего С,
то величина М будет стремиться к постоянной величине, определяемой
как27М — С ■ (W - D).В таком устойчивом состоянии все преимущества или, наоборот, недостат¬
ки организма будут компенсироваться изменением в популяции, а вовсе
не изменением величины М.Вероятно, следует сказать пару слов относительно формулировки тео¬
рии отбора, которая приписывает «борьбу за существование» избыточному
числу особей в потомстве, которое, как считается, имеет место в органи¬
ческой природе повсеместно. Если численность вида соответствует такому
уровню, при котором каждая взрослая особь оставляет после себя всего
двух потомков, которые доживут до взрослого состояния, то при наличии
смертности на любом этапе, предшествующем достижению взрослого со¬
стояния, независимо от ее причин (внешних факторов, хищников, парази¬
тов и пр.) неизбежно должен существовать некоторый запас численности,
т. е. число вновь появившихся особей непременно должно превышать чис¬
ло родительских особей. При высокой смертности величина этого превы¬
шения намного больше. Анализ этой ситуации позволяет понять, что если
44Глава 2бы мир был идеален, так что все потомство достигало зрелости и участ¬
вовало в размножении, то в таком мире численность видов должна была
бы расти неограниченно. При этом обычно добавляют, хотя это логически
не имеет отношения к делу, что такой прирост происходил бы в геомет¬
рической прогрессии. В этом смысле мы можем говорить об «избытке»
числа потомков, а геометрический прирост с его впечатляющей картиной
перенаселения часто используется как логическая основа доводов в пользу
естественного отбора. Однако следует помнить, что появляющееся число
потомков избыточно только в воображаемом мире, а «высокая геометриче¬
ская скорость прироста» может иметь место только в случае отмены ре¬
альной смертности при сохранении существующих темпов размножения.
Нечто реликтовое от философии креационизма есть в том, чтобы из наблю¬
дения, что, например, треска мечет миллион икринок, вывести заключение,
что поэтому ее потомство подвергается Естественному Отбору; достаточ¬
но ущербно исключать плодовитость из класса характеристик, по которым
мы могли бы попытаться оценивать приспособленность. Было бы весь¬
ма поучительно знать не только то, с помощью какого физиологического
механизма осуществляется точное распределение питания, направленного
на созревание гонад и на весь остальной родительский организм, но так¬
же и то, какие обстоятельства жизни особи или среды обитания могли бы
оказаться благоприятными при определении большей или меньшей доли
доступных ресурсов для будущего размножения. Исторический факт, что
и Дарвин, и Уоллес прошли через чтение очерка Мальтуса о населении,
чтобы подойти к оценке эффективности отбора, хотя и весьма поучителен
с точки зрения философии того века, не должен более сдерживать наше
понимание, чтобы мы теперь не путали последствия этого принципа с его
основами.Из предыдущих разделов этой главы должно быть очевидно, что стра¬
ховой информации, необходимой для вычисления генетических изменений,
происходящих в процессе развития популяции организмов, всегда будет
недостаточно, хотя бы просто потому, что число разных генотипов, для каж¬
дого из которых требуется мальтузианский параметр, будет часто, а вполне
вероятно и всегда, превышать число организмов в популяции, в дополне¬
ние к тому факту, что этот параметр известен с очень большими дефектами
даже для совокупностей популяций человека, для которых демографиче¬
ская статистика в некоторой степени доступна. Однако если мы согласны
рассматривать не во всех подробностях именно те изменения, которые про¬
исходят сейчас, но достаточно широко,— в какой степени организм сам
отвечает за себя в экономике природы, — то необходимо лишь определить
числовые значения четырех величин: W, D, С и М, которые входят в урав¬
Фундаментальная теорема естественного отбора45нение роста популяции. Наше невежество здесь, конечно, достаточно глубо¬
ко, однако если взять проблему только в этом ограниченном аспекте, то ста¬
нет совершенно ясно, что по отношению к организмам, которые достаточно
важны и потому заслуживают подробного изучения, что это незнание в ши¬
роком масштабе невозможно устранить с помощью систематических хоро¬
шо спланированных наблюдений. Например, величина С, которая обознача¬
ет период времени, отражающий те возможности, которые позволяют попу¬
ляции расти при повышенной приспособленности, должна быть тесно свя¬
зана с ходом увеличения и убыли популяции, когда множество организмов,
подвергшихся новым воздействиям, приближается к равновесному значе¬
нию, которое на коротких отрезках времени можно считать постоянным.
Организм, попадая в новую среду, которая оказывается для него вполне
подходящей, наращивает свою численность очень значительно и сравни¬
тельно быстро и так же быстро достигает равновесной плотности. Сход¬
ным образом происходит процесс снижения численности популяции, кото¬
рую человек поставил в условия ухудшения среды обитания. В подобных
случаях вполне вероятно, что процесс достижения равновесия происходит
достаточно быстро из-за изменений, вызванных органической эволюцией,
естественным ухудшением среды можно пренебречь, а дальнейшие измене¬
ния степени вмешательства человека для экспериментальных целей могут
быть временно исключены. В таких случаях в простейшем варианте ход
изменений популяции будет выражаться уравнением М = Аё~^с или, по¬
скольку М — это относительная или логарифмическая скорость роста по¬
пуляции, — уравнением log N = log N0 — АСе~ь/с, где N — численность
или плотность популяции, a No — постоянная величина, к которой она стре¬
мится. Значения N тогда будут определять, хотя бы приближенно, величи¬
ну константы времени С. Следует отметить, что для таких сравнительно
больших изменений плотности популяции, которые возможно установить
с достаточной точностью, существенные изменения численности совокуп¬
ностей организмов будут наблюдаться достаточно часто. Приведенная вы¬
ше простая зависимость будет удовлетворительной только при условии, что
описываемые изменения происходят быстро по сравнению с теми измене¬
ниями, которые мы изучаем. Иначе будет необходимо с помощью прямого
наблюдения учитывать изменения численности по меньшей мере наибо¬
лее важных организмов в сообществах, чтобы таким образом определить
константы более сложной системы дифференциальных уравнений, которые
могли бы описать их взаимодействия.По отношению к другим константам трудности возрастают, однако ес¬
ли посмотреть, как мало внимания в целом уделяли количественному изу¬
чению организмов в их естественных местообитаниях, было бы опромет¬
46Глава 2чиво полагать, что определение их значений находится за пределами че¬
ловеческих возможностей. Хотя было бы неуместно обсуждать здесь про¬
грамму исследований, вероятно, стоит указать на некоторые возможности.
Плотность популяций животных и растений можно изучать в зависимости
от климатических и других природных факторов среды их обитания. Ин¬
формация такого рода, даже если она лишь относительно полная, будет
показывать, насколько происходящие физические изменения могут улуч¬
шать или ухудшать окружающую среду. Если константа С также известна,
то эти установленные воздействия можно сразу переводить в терминологию
приспособленности. В некоторых случаях, например, при медленных изме¬
нениях состава растительных ассоциаций, величину М можно определить
непосредственно, а в сочетании с более или менее правдоподобным опреде¬
лением значений С и D можно получить и более или менее точную оценку
эволюционного фактора W. Для прямого определения величины послед¬
него, по-видимому, было бы необходимо располагать информацией о пол¬
ной генеалогии вида на протяжении нескольких поколений, что возможно
только для человека. Более того, учитывая быстрые изменения, которым
человек подвергает окружающую природу, можно предвидеть, что и гене¬
алогии людей в национальном (как например, в Швеции) или интернацио¬
нальном масштабе, проливая в какой-то мере свет на современные условия
репродукции и выживания людей, будут испытывать и все новые трудности
в плане определения и интерпретации эволюционного показателя W.РезюмеСтатистика жизни организма, прослеженная в связи со средой его оби¬
тания, предоставляет способ определения меры относительной скорости
роста популяции, которая может быть названа мальтузианским параметром
популяционного роста, а также позволяет оценить репродуктивные пока¬
затели индивидов для всех возрастов и всех этапов их жизненного цикла.
Мальтузианский параметр в общем будет различным для каждого генотипа,
и он представляет меру приспособленности по выживанию каждого инди¬
вида.В популяции вариация любого индивидуального показателя устанав¬
ливается количественно по его дисперсии, часть которой с учетом генети¬
ческого набора всех возможных индивидов может быть принята как гене¬
тическая дисперсия.Скорость возрастания приспособленности любого вида равна генети¬
ческой дисперсии приспособленности, и стандартная ошибка этой скоро¬
сти, даже в масштабах одного поколения, будет мала по сравнению с самой
Фундаментальная теорема естественного отбора47скоростью, за исключением случая, когда скорость изменения приспособ¬
ленности (время измеряется в поколениях) настолько мала, что сравнима
с величиной, обратной численности популяции.Адаптацию в смысле согласованности по многим особенностям двух
сложных совокупностей можно проиллюстрировать с помощью геометри¬
ческих свойств многомерного пространства, подразумевая статистическую
ситуацию, в которой вероятность того, что произойдет изменение заданного
масштаба, ведущее к улучшению, снижается наполовину от лимитирующе¬
го значения по мере того, как масштаб изменения увеличивается. Интенсив¬
ность адаптации обратно пропорциональна стандартной величине измене¬
ния, для которого эта вероятность постоянна. То есть, чем больше измене¬
ние или глубже адаптация, тем меньше вероятность какого-то улучшения.Если организм, строго говоря, высоко адаптирован к своей нише в при¬
роде, то повышению приспособленности противодействует ухудшение, вы¬
званное ненаправленными изменениями организма или его среды обита¬
ния. Влияние ухудшения организма, обнаруживаемого генетиками по па¬
тологическим мутациям, нивелируется эквивалентной величиной генети¬
ческой дисперсии. Ухудшение среды обитания может возникать как под
воздействием геологических и климатических изменений, так и вследствие
изменений в органическом окружении, включая и усиление влияния вра¬
гов и конкурентов; изменения могут быть более или менее интенсивными
по сравнению с эффектами Естественного Отбора.Любое достигнутое организмом преимущество будет закрепляться
в увеличении скорее численности популяции, чем через увеличение сред¬
него уровня мальтузианского параметра, который в этой ситуации всегда
остается близким к нулю.Хотя кажется невозможным представить, что детали механизма дей¬
ствия Естественного Отбора могут быть полностью осознаны человеческим
разумом, прямые методы наблюдения тем не менее оказываются в состоя¬
нии определить численные значения, которые являются условием выжива¬
ния и прогресса конкретных видов.
Глава 3
Эволюция доминантностиДоминирование диких генов. Модификация эффектов менделевских факторов.
Модификация гетерозиготы. Особые приложения теории. Процесс модификации.
Выводы из теории эволюции доминантности. Резюме.Суть гипотезы состоит в том, чтобы понять: повлияла ли реальная
причина на что-либо еще, кроме того, что мы наблюдаем. РОБЕРТСОН
СМИТДоминирование диких геновВ главе 1 было показано, что в свете корпускулярной природы наслед¬
ственности едва ли можно ожидать, что мутации могут каким-либо образом
определять направление эволюционных изменений; их значение для эво¬
люции связано с тем, что они играют совершенно иную роль, поддерживая
некоторый уровень генетической дисперсии, который, в свою очередь, яв¬
ляется фактором, влияющим на скорость — но не направление — процес¬
са эволюции. Прежде чем пытаться детально рассматривать зависимости
между уровнем запаса генетической изменчивости вида, частотами мута¬
ций и численностью популяции, что будет сделано в главе 4, необходимо
выяснить (насколько позволяют имеющиеся в настоящее время данные) ха¬
рактер генетических изменений, известных как мутации.Некоторые (особенно те, для кого относящиеся к этому данные до сих
пор остаются в значительной мере неизвестными), конечно, посчитают,
что в настоящее время преждевременно выдвигать в качестве основания
для будущего обсуждения теорию эволюции доминантности ввиду того, что
до недавнего времени генетики воспринимали доминирование как непо¬
нятный факт, от которого из-за непонимания причин явления просто отма¬
хивались, не видя за ним никакого теоретического значения. Однако едва
ли может быть оправданным развитие теории о роли мутаций в эволюции
без рассмотрения действительно наблюдаемых случаев мутаций. Изучение
этих изменений открывает множество материалов, пока достаточно непро¬
тиворечивых, хотя и с пробелами, многие из которых, конечно, вскоре будут
заполнены, поскольку проявилось внимание к предмету; поэтому, вероятно,
Эволюция доминантности49не будет ошибкой представить читателю наиболее яркие примеры этих дан¬
ных и мое заключение. Толкование этих данных пока весьма условно, но я
надеюсь, что оно достаточно скоро будет заменено более прямыми и полны¬
ми результатами научных наблюдений; пока же, я полагаю, моя теория ока¬
жется в ходе исследований достаточно хорошо обоснованной, и ее можно
использовать как указатель направления наших дальнейших работ. Обосно¬
ванность последующих глав не ставится под сомнение, хотя, если я ошиба¬
юсь в своих выводах, вопрос об их уместности возникнет уже в этой главе.Термин мутация используется применительно к ряду различных типов
внутриклеточных событий, общим для которых является возникновение но¬
вых наследуемых изменений признаков. Известны случаи удвоения полных
хромосомных наборов, удвоения единичных хромосом или частей хромо¬
сом; в других случаях часть хромосомы оказывается перенесенной из свое¬
го привычного положения и прикрепленной к какой-то из других хромосом;
все это — мутации в широком и основном значении этого термина. Однако
эволюционные возможности этих типов изменений явно очень ограничены,
если их сравнивать с типом изменений, который обычно называют генны¬
ми мутациями. Сюда относят изменения в элементарной наследственной
частице — гене, который превращается в ген нового типа, занимающий тот
же локус в зародышевой структуре. Более крупные формы мутаций в дей¬
ствительности могут играть особую эволюционную роль, создавая меха¬
низм репродуктивной несовместимости, который может оказаться весьма
важным, когда возникает вопрос о физиологической изоляции, однако лишь
в совершенно особых случаях они могут быть значимыми для генетическо¬
го разнообразия в популяции с интербридингом.В пределах каждой пары альтернативных, или аллеломорфных, генов
можно отличить один от другого по четырем моментам, что важно для уста¬
новления взаимосвязей между ними. Можно различать: а) более редкую
форму и более широко распространенную; б) менее полезный вариант
от более благоприятного; в) мутантный ген — от относительно первичного,
из которого произошел новый, и, наконец, г) рецессивный ген от доминант¬
ного. Только после того, как эти четыре противоположности станут ясны,
мы сможем оценить связи между ними, определяемые разными причинами.При рассмотрении природы адаптации было показано, что мутантные
гены чаще являются неблагоприятными, что особенно заметно, когда вновь
возникающий эффект велик, то есть его легче установить и изучить. Раз¬
личия по критериям (б) и (в), таким образом, тесно связаны. Более того,
в популяции, где есть свободное перемешивание (но не в совокупности
генотипов, поддерживаемых в виде отдельных линий), появление менее
благоприятных генов будет постепенно становиться все реже, и если эту
50Глава 3тенденцию проверять в любой момент, то эти мутантные и менее полезные
гены будут, в общем, и более редкими. Наконец, если предварительно пред¬
положить, что мутантные гены являются доминантными так же часто, как
и рецессивными, то выяснится, что отбор будет значительно более сурово
элиминировать неблагоприятные доминантые гены, чем неблагоприятные
рецессивные. Легче всего это проследить, рассматривая случаи, когда оба
типа одинаково редки. Если 1 ген из 100 несет доминантный дефект, то
в популяции на каждые 10000 особей будет приходиться в среднем 199 де¬
фектных, подвергающихся отрицательному отбору, тогда как если этот ген
рецессивен (тоже 1 из 100), то дефект проявится в среднем только у 1 осо¬
би из 10000. Следовательно, редкий рецессив намного легче скрыт от дей¬
ствия отбора, и в такой популяции можно ожидать обнаружения многих
случаев редких рецессивных дефектов (а, б, г), но лишь очень редко —
доминантных дефектов. Тот факт, что редкие рецессивные особи в случае
инбридинга проявляют свою дефектность, еще не доказывает, что дефект¬
ные мутанты, как правило, рецессивны. С другой стороны, если эволюция
осуществлялась такими же темпами, что и воздействие факторов, которые
удобны в изучении, можно ожидать, что среди редких рецессивных генов
в основной популяции обнаружится несколько первичных генов, которые
были вытеснены более полезными генами, возникшими из них мутаци¬
онным путем. Такие случаи практически не известны; это можно интер¬
претировать как свидетельство того, что либо «шаги» эволюции обычно
не столь велики, как эффекты факторов, которые мы можем изучать, либо
что мутантные гены редко или никогда не бывают полностью доминантны¬
ми по отношению к своим предшественникам. Так как я думаю, что оба
эти предположения верны, то непозволительно использовать это наблюде¬
ние в качестве подтверждения одного из них.Для видов растений, разводимых, подобно душистому горошку, от¬
дельными разновидностями, генетический анализ показывает, что гены, ко¬
торые на основании морфологических признаков следует считать мутант¬
ными, в преобладающем большинстве являются рецессивными. У души¬
стого горошка полная рецессивность, по-видимому, является устойчивым
правилом, судя по 15 или 20 различным генам, которые успешно изучены
к настоящему времени. В большинстве этих случаев возникновение мута¬
ции само по себе остается невидимым, однако мутантный ген распознают
по его проявлениям, не известным у более ранних разновидностей или ди¬
ких прототипов. Этот случай представляет очень существенный и обшир¬
ный ряд наблюдений, свидетельствующих о том, что мутантные гены обыч¬
но бывают рецессивными, тогда как доминантные мутанты должны были
бы немедленно использоваться и сохраняться навсегда как новинки, и об-
Эволюция ДОМИНАНТНОСТИ51Таблица 1п Промежу- Полностью пРецессивные , Всеготочные доминантные130 9 0 13978 4 0 82наружить их намного легче, чем рецессивные. То, что их легче заметить,
следует особо подчеркнуть применительно к человеку, когда распознава¬
ние редкого дефекта, определяемого единичным менделевским фактором,
тесно связано с генеалогическими сведениями; даже простейшая родослов¬
ная, которая почти всегда доступна, дает возможность обнаружить признак
с доминантным дефектом, тогда как для того, чтобы проследить судьбу
простого рецессивного признака, обычно требуется сопоставление стати¬
стических данных по обширным родословным линиям и древам. Напри¬
мер, долго обсуждался даже менделевский характер наследования альби¬
низма у человека. Вследствие этой трудности у человека известно больше
доминантных дефектов, чем рецессивных, хотя не может быть сомнений
в том, что большинство дефектов человека, как физических, так и психи¬
ческих, все-таки являются рецессивными, как и у других животных. Эту
точку зрения подтверждает и тот факт, что мутации, сцепленные с полом,
которые у организма с 23 парами хромосом28 a priori должны составлять
явное меньшинство, хорошо заметны в общем списке дефектов; здесь ре¬
цессивные мутации обнаруживаются почти так же легко, как доминантные,
и почти все известные случаи рецессивны. Относительно так называемых
доминантных дефектов человека нам известно только то, что гетерозиго¬
та отличается от нормальной особи; обычно мы не знаем, как выглядит
мутантная гомозигота и можно ли ее вообще наблюдать.Внушительный объем генетических исследований, выполненных на пло¬
довой мушке Drosophila, предоставил наибольшее количество решающей
информации и по этой, и по другим генетическим проблемам. В культурах
этой мухи к настоящему времени зарегистрировано порядка 500 мутаций;
если не считать большого класса деталей и всех других, не дающих види¬
мого эффекта, остается 221 случай, когда мы классифицируем мутации как
рецессивные, промежуточные, или, как обычно говорят, с неполным доми¬
нированием, и, наконец, полностью доминантные. В таблице 1 показано
распределение по этим трем классам для аутосомных мутаций и мутаций,
сцепленных с полом.Классификация приведена по данным замечательной работы о Dro¬
sophila в издании Bibliographia Genetica. В нескольких отдельных случаяхМутацииАутосомные
Сцепленные
с полом
52Глава 3моя классификация может быть ошибочной либо потому, что наблюдениям
помешала некоторая неопределенность, либо из-за того, что не в каждом
случае признаки были выражены достаточно четко. Никому не придет в го¬
лову усомниться, что генетика этой мухи изучается чрезвычайно тщатель¬
но; однако, вероятно, стоит упомянуть, что если рецессивные варианты на¬
столько многочисленны, что пропуск какого-то единичного случая не при¬
вел бы к трагедии, то мутации, которые «можно принять как доминантные»,
то есть промежуточные, чрезвычайно важны для будущих исследований
и потому очень ценны29. Кроме того, они обнаруживаются сразу после воз¬
никновения, тогда как рецессивные становятся заметны только после того,
как появляются их гомозиготы. В силу обеих причин доля рецессивных
мутаций, как бы высока она ни была, вероятно, все же остается недооце¬
ненной относительно действительной частоты возникновения мутаций.Летальные гены, которые не вошли в приведенный выше список,
предоставляют независимое, хотя и несколько двусмысленное подтвержде¬
ние того же заключения. Большинство так называемых доминантных ва¬
риантов детальны в гомозиготном состоянии и потому их вернее было
бы классифицировать как промежуточные. Еще более удивительно, что
рецессивные летали, которые могут не давать видимых изменений, вско¬
ре были обнаружены, поскольку они нарушали соотношение частот дру¬
гих факторов, особенно соотношение полов. Недавно было показано, что
и при нормальных условиях, и когда мутации были искусственно индуци¬
рованы рентгеновскими лучами, рецессивные летали составляют намного
более частый класс мутаций. Среди плодовых мушек, которых не подверга¬
ли никаким воздействиям, вероятно, около 2 % несут какие-то рецессивные
летальные мутации, а общая частота мутаций этого класса, по-видимому,
на порядок превышает число всех видимых мутаций. Среди последних одна
из каждых семнадцати мутаций считается промежуточной с точки зрения
доминирования, а среди леталей пропорция должна быть еще ниже, за ис¬
ключением некоторых из незаметных, пусть и достаточно многочисленных
мутаций, которые летальны в гетерозиготном состоянии, и их можно счи¬
тать доминантными.Явная тенденция мутантного гена к рецессивности, как бы возвраще¬
ния к дикому типу, от которого он происходит, нуждается в объяснении, и,
к счастью, существует и доступна важная группа наблюдений, результаты
которых помогают выявить, что в диком состоянии преобладает доминант¬
ный ген, но не свидетельствуют о том, что этот ген является предшествен¬
ником возникшего мутанта. Сейчас известны многочисленные случаи, ко¬
гда несколько разных мутаций появляются в одном и том же гене, и каждый
мутантный вариант может заменять любой другой, в том числе и дикий тип,
Эволюция ДОМИНАНТНОСТИ53в том же локусе. Например, у грызунов было обнаружено несколько серий
генов альбинизма, создающих эффект от легкого ослабления пигментации
до полного ее подавления. Используя серию из пяти таких альтернативных
генов у морских свинок, Сьюэл Райт получил все 15 возможных комби¬
наций: 5 гомозигот и 10 гетерозигот, которые может сформировать набор
из пяти аллеломорфов. Все варианты он исследовал на выборках доста¬
точного объема, чтобы определить среднее, а также нормальную вариацию
интенсивности пигментации как в направлении изменения от черного че¬
рез сепию до белого, так и от рыжего через желтый. Четыре гетерозиготные
формы, несущие ген дикого типа, по интенсивности окраски не отличимы
от гомозиготного дикого типа, но другие четыре гомозиготы отличаются
от дикого типа весьма существенно. Остальные 6 гетерозиготных форм,
у которых дикий аллель отсутствует, демонстрируют четко промежуточные
варианты окраски по обоим цветам, а крайними в этих рядах являются
гомозиготные особи. Этот пример примечателен благодаря тщательности,
с которой был проведен весь анализ; он, конечно, является исключением.
Ряд схожих серий генов был обнаружен у Drosophila, и правило, что ди¬
кий тип доминирует над всеми остальными, тогда как прочие не проявляют
доминирования относительно друг друга, было подтверждено Морганом,
Бриджесом и Стёртевантом.Исключительное положение генов дикого типа в плане доминирования
среди других аллеломорфов не является следствием того, что они представ¬
ляют тот оригинал, из которого путем мутаций происходят все остальные;
известны случаи образования одного мутантного аллеломорфа из другого,
тоже мутантного, а также и такие, когда мутантные гены в результате мута¬
ции возвращались обратно к дикому типу. Это наводит нас на мысль о том,
что доминирование следует рассматривать не как характеристику, позволя¬
ющую потомку в результате серии мутационных изменений стать отлич¬
ным от предковой формы, но как свойство, нужное для противопоставле¬
ния преобладающего дикого типа его неудачным конкурентам. Кроме того,
если нам не придется совершенно отказаться от эволюционной концепции
об изменении видов путем случайной замены какого-либо гена на ген его
предшественников, от которого он некогда' произошел, то согласно суще¬
ствующему правилу, которое говорит о генетическом доминировании генов
широко распространенного дикого тина, требуется, чтобы новый полезный
ген каким-то образом через некоторое время тоже стал доминантным по от¬
ношению к своим конкурентам, а если происходят обратные мутации — мог
бы вернуться и к предковому состоянию. Пути, которыми это может проис¬
ходить, представляют особый интерес в теории Естественного Отбора, так
как они раскрывают влияние отбора, которое не имеет ничего общего с его
54Глава 3хорошо понятным действием для приспособления вида к его месту в при¬
роде. Как было показано в главе 2, едва ли можно представить проблему
более сложную или требующую настолько непостижимо детализированных
знаний об экологической ситуации, чем проблема отслеживания чистого
выигрыша приспособленности за счет любого конкретного генетического
изменения. Наших познаний в этой области пока хватает, чтобы оценить
адаптивное значение различий в организации между порядками или семей¬
ствами, однако их почти всегда недостаточно, чтобы аналогичным обра¬
зом разобраться с видовыми различиями и тем более с внутривидовыми
вариациями. Это обстоятельство, которое воспринимается как затруднение
для теории Естественного Отбора, в частности, такими авторами, как Бэт¬
сон и Робсон,— если не считать ряда заметных исключений, как напри¬
мер, внешняя окраска и особенно миметическая окраска у бабочек,— тем
не менее подстегивает наш интерес к ситуациям, когда наши количествен¬
ные данные, пусть даже далеко не точные, все-таки оказываются важными
и приближают нас к сути вопроса.Модификация эффектов менделевских факторовПривычка говорить о том, что этот фактор или генное замещение вы¬
зывают данное соматическое изменение, преобладавшая среди ранних гене¬
тиков, в значительной мере уступила место пониманию того, что на изме¬
нение, даже генетически определенное, могут влиять или даже направлять
его либо природная среда, которая как бы испытывает эту замену, либо
какие-то другие элементы генетической структуры. Так, достаточно рано
стали известны случаи, когда фактор В проявлял свое влияние, если вто¬
рой фактор — А — был представлен рецессивным геном, и не проявлял,
если присутствовал доминантный ген. В этом случае говорят, что фактор А
является эпистатическим по отношению к фактору В, или, как стали го¬
ворить в последнее время, что b — это специфический модификатор ге¬
на а.30 Известны и другие случаи, когда ни А, ни В не проявляют никакого
эффекта, если один из них рецессивен, тогда говорят о двух комплемен¬
тарных факторах; если же ни один из них не проявляет действия, когда
другой доминантен, их называют дуплицированными генами. Ясно, что это
лишь частные случаи более общего явления, когда видимый эффект генно¬
го замещения зависит не только от собственно замещения, но и от всего
генетического комплекса или организма, в котором это замещение проис¬
ходит. Наверное, можно попытаться найти такую формулировку, которая
бы свела до минимума несоответствие между сложностью реальных за¬
висимостей и простотой связей, предполагаемой на основании словесных
Эволюция ДОМИНАНТНОСТИ55выражений, когда говорят о наблюдаемом соматическом изменении как ре¬
акции организма на обсуждаемое генное замещение. При этом по крайней
мере нужно суметь избежать впечатления неопределенности или противо¬
речия, если организмы разной структуры демонстрируют разные реакции.
Если подходить именно с этой точки зрения, то не будет чем-то совершенно
непостижимым считать, что организм должен развиваться таким образом,
чтобы изменялась его реакция на любое частное генное замещение.Известно несколько случаев, когда такая модификация наблюдалась
в экспериментальных линиях. Довольно часто можно наблюдать, что ко¬
гда нового, резко отличающегося мутанта Drosophila отсаживают, помещая
в стаканчики для размножения в течение нескольких поколений, то потом,
когда он оказывается нужен снова, выяснялось, что мутантная форма ста¬
новится куда менее отличимой от дикого типа, чем когда ее обнаружили
в первый раз. Реальность этой тенденции возвращения к дикому типу, как
и вызывающая ее причина, были продемонстрированы несколько раз с по¬
мощью простого, но решающего эксперимента, когда модифицированных
мутантов скрещивали с особями неродственной дикой линии, а от гибрид¬
ных особей с помощью инбридинга опять отбирали мутантов. Восстанов¬
ленная таким образом мутантная форма, как выяснилось, во многом верну¬
лась к своей первоначальной интенсивности; все это показывает, что моди¬
фикация не была обусловлена каким-либо изменением в мутантном гене,
но произошла под влиянием изменения генетического комплекса организ¬
ма и взаимодействия с ним. Такое изменение поддается простому объяс¬
нению. Мухи данной линии различались по генетическим особенностям,
что повлияло на интенсивность их реакции на мутантный ген. При нали¬
чии конкурентных условий внутри стаканчиков те наследственные едини¬
цы, которые благоприятствовали мягкой реакции, в результате создали мух
с меньшими дефектами по сравнению с их конкурентами, и отбор этих мо¬
дифицирующих факторов стремительно изменил среднюю интенсивность
реакции на мутантный ген и, следовательно, его отклонение по внешним
признакам в среднем от диких мух. Похожий случай частичного восстанов¬
ления мутации у настурции, сопровождавшегося частичной стерильностью,
наблюдал профессор Вайс, а г-н Е. Б. Форд сообщил мне, что мутантные ти¬
пы, встречающиеся у креветки Gammarus chevreuxi, в культуре часто обна¬
руживали заметное повышение жизнеспособности. Таким образом, эффект
сохранения мутаций в группе особей, полученных путем разведения наиме¬
нее дефектных из них, должен оказывать влияние на организм, изменяя его
так, чтобы смягчить неблагоприятное действие мутации. То есть во власти
Естественного Отбора оказывается не только частота гена, но и реакция
организма на него. Чтобы понять влияние гена на индивидов данной попу¬
56Глава 3ляции, то есть реакцию этих организмов на него, мы должны понять, какую
роль этот ген играл у их предков.Подавляющее большинство мутаций, если не все, которое мы мо¬
жем надеяться обнаружить в экспериментальной культуре, кроме тех, ко¬
торые можно отнести на счет наших методов культивирования, должны
были возникать в истории вида в огромных количествах: многие мутации
у Drosophila повторно отмечались в культуре, но, как бы много их ни было,
эти числа ничтожны по сравнению с общим количеством предков любой
особи дикой мухи. Сегодня мы мало знаем о частотах отдельных мутаций;
однако этого достаточно, чтобы установить, что многие мутации должны
возникать с частотой 1 на 100000 или 1 на миллион; и действительно, ве¬
роятность обнаружить в культуре существенно более редкие мутации чрез¬
вычайно мала. Мы, конечно, не имеем прямых данных о частотах мутаций,
которые преобладают в природе, однако те сведения, которыми мы рас¬
полагаем, не дают оснований полагать, что они ниже, чем в лаборатории.
Что касается родства мутантной особи и того отдаленного предка, у ко¬
торого эта мутация могла появиться, то имеются основания полагать, что
временной интервал между ними может быть больше продолжительности
существования вида; у разных видов Drosophila было отмечено несколь¬
ко мутаций, которые можно обнаружить путем гибридизации. Кроме того,
прямые тесты на идентичность у нас отсутствуют; однако имеет некоторые
основания догадка, что такая мутация, как альбинизм, которая встречается
у млекопитающих из самых разных отрядов, появилась у предков группы
очень давно. С другой стороны, как будет показано ниже, у нас есть причи¬
на полагать, что по мере эволюции новых видов иногда начинают возникать
новые мутации, причем с заметной частотой.Модификация гетерозиготыКогда неблагоприятная мутация продолжает устойчиво появляться
в каждом поколении, скажем, один раз на каждый миллион хромосом, ко¬
нечно, отбор будет поддерживать ее как редкую; с другой стороны, она
затронет многих особей, которые потенциально являются предками для бу¬
дущих поколений, в дополнение к тем, у которых мутация реально про¬
явилась. Важное следствие ее редкости состоит в том, что оба этих класса
будут гетерозиготами гораздо чаще, чем гомозиготами. Если р — относи¬
тельная частота в популяции мутантного гена по отношению к гену дикого
типа, то три класса особей: немутантные, гетерозиготы и гомозиготные му¬
танты — будут наблюдаться в соотношении 1 : 2р : р2, так что даже если
бы р была столь высока, что доходила до 1 на 1 000, то число гетерозигот
Эволюция ДОМИНАНТНОСТИ57в 2 000 раз превышало бы число мутантных гомозигот31. Вследствие этого
в течение всего времени, пока гетерозигота ощутимо отличается от дико¬
го типа по приспособленности к выживанию, относительная численность
трех классов будет определяться при данной частоте мутаций селективной
невыгодностью гетерозиготы; при этом она не зависит сколько-нибудь су¬
щественно от уровня селективной вредности или даже полной летальности
мутантных гомозигот.В работе мы принимаем за относительную приспособленность гетеро¬
зиготы (v) отношение среднего числа потомков этого типа к среднему числу
немутантных особей. Тогда легко видеть, что доля р в каждом поколении
будет уменьшаться на величину р( 1 — и), и пока значения р малы, эта вели¬
чина будет увеличиваться за счет показателя к, представляющего реальную
частоту мутаций. Поэтому равновесие между эффектами мутаций и отбора
будет достигаться, когдар(1 —v) = k.Если взять тот крайний случай, когда v мало, то значения р будут немного
превышать к, например, чуть больше, чем 1 на миллион, и в этом крайнем
случае гетерозиготы будут появляться в 2 миллиона раз чаще, чем мутант¬
ные гомозиготы. Если v = У2, то величина р будет вдвое больше к, а ге¬
терозигот все еще будет в миллион раз больше. С другой стороны, если
жизнеспособность и общая приспособленность гетерозигот настолько хо¬
роши, что дают снижение лишь на 1 %, и » = 0,99, то и тогда частота
гетерозигот будет выше в 20 000 раз.Такие очень высокие соотношения подтверждают заключение о том,
что если гетерозигота создает хоть какой-то заметный ущерб по сравнению
с диким типом, то частота гетерозигот будет превышать частоту гомози¬
гот в такое гигантское число раз, что любой отбор модификаторов будет
определяться реакцией гетерозигот.Два других обстоятельства также способствуют увеличению несораз¬
мерности селективных эффектов. Во-первых, действенность отбора в плане
изменения признаков видов зависит не только от частоты отбираемых осо¬
бей, но также от того, каков их шанс оставить в отдаленном будущем
потомство. На деле нам нужно оценить не только относительную числен¬
ность обоих типов в любом поколении, но и те соотношения, которые они
будут представлять среди всех особей некоторого отдаленного поколения.
Очевидно, что если, как можно ожидать, жизнеспособность гомозиготных
мутантов ниже по сравнению с гетерозиготами, то и потомками последних
в будущем будет больше особей, и даже если жизнеспособность обоих этих
типов одинакова — все равно гетерозиготы будут иметь преимущество, так
как при скрещивании с особями дикого типа только половина потомков бу¬
58Глава 3дет гетерозиготной, тогда как при аналогичном скрещивании гомозиготных
особей все потомки будут в равной степени иметь недостатки.Этот момент становится особенно важным, когда речь идет о генах,
сцепленных с полом, когда самцы мутантного типа и гетерозиготные самки
не сильно различаются по частотам, но могут иметь существенные разли¬
чия в жизнеспособности, что может намного чаще проявляться у потомства
существующей дикой популяции.Во-вторых, с точки зрения биохимии внутриклеточной активности ге¬
нов трудно не признать вероятность того, что гетерозигота врожденно мо¬
жет быть более изменчива по фенотипу, чем любая из гомозигот, особенно
по тем признакам, которые отличают последних; для того чтобы что-то
могло измениться у гомозиготы, мы должны представить участие в этом
некоего гена-модификатора, который усиливает или, наоборот, ингибирует
проявление этого признака, тогда как у гетерозиготы уже есть все необхо¬
димые составляющие, которые обеспечивают любые процессы, нормально
протекающие у каждой из гомозигот. Дальнейшее рассмотрение приведен¬
ных ниже примеров, в которых изменения оказались видимыми, должны
сделать более понятным то, насколько большое значение должно прида¬
ваться этому положению.Та доля потомков в будущих поколениях, которую считают происхо¬
дящей от гетерозигот, хотя и многократно превышает численность потом¬
ства гомозигот, тем не менее очень мала по абсолютным значениям. Мож¬
но определить долю потомков одной гетерозиготной особи относительно
особей-немутантов, считая ее равной половине от доли, приписываемой ее
вероятному потомку; тогда, если соотношение для гетерозигот и немутан-
тов принять за х : 1, то мы получимX = -i v(l + х),ИЛИ уи, поскольку соотношение в популяции, которая гетерозиготна, будет2 к1 — v'то их пропорциональный вклад в потомство будущих далеких поколенийокажется равнымF 2 kv(1 — v)(2 — v)Когда частота мутаций (к) равна 1 на миллион, это число увеличивается
примерно до 1 на 5 ООО, если v равно 0,99, оно отражает скорость модифи¬
Эволюция ДОМИНАНТНОСТИ59кации гетерозиготы по сравнению со скоростью изменений, на которую
оказывал бы влияние отбор такой же интенсивности, действующий на по¬
пуляцию, целиком состоящую из гетерозигот. В случае с гомозиготами
было показано, что изменение под воздействием Естественного Отбора
модифицирующих генов вовсе не является незначительным, его нельзя
не принимать в расчет даже на коротких отрезках времени наблюдения
и при наличии серьезного ограничения, что запас модифицирующей дис¬
персии лимитирован исходно малым числом особей. Рассматривая модифи¬
кации, обусловленные действием отбора на гетерозиготы в природе, можно
справедливо предположить, что гетерозиготы по крайней мере в той же
степени подвержены генетической модификации, как и гомозиготные му¬
танты, и что отбор, действующий только на 1 особь из 5 000 или 10000
особей в популяции, не будет оказывать заметного влияния в плане сниже¬
ния существующей дисперсии.Особые приложения теорииОб исключительно интересном случае, показывающем модификацию
гетерозиготы до такого состояния, что она перестала отличаться от не¬
мутантных форм, то есть ген дикого типа стал полностью доминантным,
мне сообщил г-н Хатчинсон, рассказав о работе доктора Харланда по гене¬
тике хлопчатника. Несколько американских видов хлопчатника легко скре¬
щиваются между собой и дают плодовитое потомство. У одного из этих ви¬
дов, си-айленд (the Sea Island cotton), повторно возникла мутантная форма,
известная как Crinkled Dwarf, которая у этого вида полностью рецессив¬
на. Она оказывается идентичной с похожим мутантом, названным Wrinkled
Leaf, который известен у некоторых близкородственных форм, выращивае¬
мых в Египте, однако, насколько известно, ни один из американских видов
до сих пор такого мутанта не обнаруживал. В ходе экспериментов док¬
тор Харланд скрестил мутантные растения Crinkled Dwarf вида си-айленд
с двумя другими американскими видами — хлопчатником упланд (Upland)
и хлопчатником перуанским (Peruvian). Удивительные результаты скрещи¬
ваний в обоих случаях были одинаковы. Выяснилось, что у гетерозиготы
слегка проявился мутантный признак, что уже указывало на неполное до¬
минирование. Однако самые удивительные результаты проявились во вто¬
ром поколении, полученном от гетерозиготы путем самоопыления. В этом
варианте следовало ожидать, что четверть потомков будут нести мутацию
Crinkled Dwarf, половина должна быть гетерозиготна и еще четверть расте¬
ний должна быть немутантной. Гомозиготные растения, как и ожидалось,
были обнаружены, но представляли практически непрерывный ряд проме¬
60Глава 3жуточных форм. Гетерозиготы действительно проявили все степени доми¬
нирования. Очевидно, что хлопчатник Sea Island отличался от остальных
американских видов по ряду генов-модификаторов, оказывающих влияние
на развитие и проявление гетерозигот, комбинированный эффект которых
у этого вида должен обеспечить нормальный облик гетерозиготных расте¬
ний. В этом случае полное изменение реакции организма на мутантный ген
должно было возникнуть где-то вскоре после отделения вида от его аме¬
риканских сородичей; таким образом, весь процесс изменения от первого
появления мутации, по крайней мере с уже заметной частотой, должен был
происходить сравнительно быстро. Когда частота мутации будет опреде¬
лена, этот случай следует использовать как полезный ориентир для оцен¬
ки аналогичных событий, которые предположительно могли иметь место
и у других видов32.Группа фактов, имеющих совершенно исключительный интерес в этой
связи, получена при работе с домашней птицей. Скрещивания между раз¬
ными линиями выявили, что большое число отличительных признаков ли¬
ний определяются простыми менделевскими факторами. Однако в ряде слу¬
чаев доминантными оказались не признаки диких Gallus bankiva, а причуд¬
ливые признаки, характерные для линий. Таких должно быть около дюжи¬
ны или даже больше; например, известно, что три гена влияют на форму
гребня, один определяет появление хохолка; есть один доминантный бе¬
лый, который ингибирует образование пигмента в оперении; другие вли¬
яют на цвет или форму перьев, а также цвет цевки. У домашней птицы
также обнаружены мутанты разных типов, известные и у других организ¬
мов как рецессивные и летальные «доминанты», но они составляют ис¬
ключение в этой удивительной группе генов, которые нелетальны и до¬
минантны по отношению к дикому типу. Стоит отметить, что ни один
из этих генов не происходит от зарегистрированного мутанта, а их эф¬
фекты, которые предположительно могли бы оказаться вредными в дикой
природе, по-видимому, не несут никакой патологии в плане снижения жиз¬
неспособности этих птиц при домашнем содержании. Все эти признаки
на практике являются четкими линейными признаками. У других птиц, ко¬
торых разводят в неволе, похоже, отмечались только обычные рецессив¬
ные мутанты, как например, желтовато-коричневые канарейки или желтые
волнистые попугайчики; и для каждого из этих примеров мы должны ис¬
кать объяснение такой специфики домашней птицы, возможно, в особенно¬
стях одомашнивания, чем в природе или условиях обитания диких видов.
С первым, возможно, связано одно совершенно удивительное обстоятель¬
ство, которое может пролить свет на доминантные признаки у домашних
линий.
Эволюция ДОМИНАНТНОСТИ61Дикая джунглевая курица широко распространена во многих регионах
Индии, и часто отмечалось, что эти дикие петухи охотно спариваются, если
возникает такая возможность, с домашними курицами. Если такое проис¬
ходит до настоящего времени, то можно полагать, что то же самое имело
место всегда, с самого начала одомашнивания, и на протяжении всего этого
времени, вероятно, достаточно долгого, это было довольно обычным явле¬
нием, когда стаи птиц обитали в джунглях. Я не утверждаю, что петухов
не держали как домашнюю птицу — их могли держать, например, для пе¬
тушиных боев с тех же давних времен, как кур стали держать в качестве
несушек; более того, некоторые из генов, о которых идет речь, сцеплены
с полом и проявляются только у петухов; но вероятно, и к тому же почти
невозможно это опровергнуть, что в течение многих веков непрерывно име¬
ло место скрещивание домашних птиц с дикими. Что же касается большин¬
ства домашних животных и растений, то у них рецессивные мутации сразу
после появления дают породистый приплод, а любопытство людей и их
пристрастие ко всяким новинкам неоднократно подталкивали их к тому,
чтобы поддерживать эти формы, которые в природе сразу после возникно¬
вения элиминируются Естественным Отбором. При скрещивании с дикими
формами такие рецессивные признаки перестают быть видимыми и кажут¬
ся утерянными, и если такие скрещивания происходят часто, то привести
к образованию и закреплению признаков породы могут только те мутации,
которые не полностью рецессивны. Несущие такие гены цыплята в этой си¬
туации всегда будут проявлять линейные признаки, и длительная селекция,
направленная на сохранение ценных признаков, в итоге приведет к их за¬
креплению в породе. Более того, поскольку эти признаки можно сохранить
только селекцией, то ясно, что и отбираться будут прежде всего именно те
особи, у которых мутантный признак выражен ярче всего. И хотя стада со¬
стояли наполовину из гетерозигот, а другой половиной были птицы дикого
типа, если человеку казались ценными признаки гетерозигот, и потому он
выбирал именно этих особей, на деле ему неизбежно приходилось отби¬
рать тех гетерозигот, у которых мутантный ген был наименее рецессивен
или более всего проявлял свою доминантность.Следует отметить, что, следуя этой точке зрения на происхождение
некоторых породных признаков у домашней птицы, мы получаем объясне¬
ние двух конкретных особенностей, которые проявляются в этих признаках;
в частности, речь идет о высокой доле мутантных признаков, которые не ре¬
цессивны по отношению к дикому типу, а также о той высокой степени,
с которой доминантность проявляется как минимум у некоторых линейных
кроссов. В этой связи также важно, что в случае некоторых из этих генов
известны другие кроссы, у которых проявляется неполное доминирование.
62Глава 3Полное и удовлетворительное изучение этих случаев представляется воз¬
можным только с помощью введения мутантного гена в те породы, для ко¬
торых такой ген неизвестен,— поскольку эффект доминирования можно
правильно проанализировать только тогда, когда две гомозиготы и гетерози¬
гота имеют по другим генам сходный генетический фон. Можно отметить,
что мое представление относительно изменения доминантности мутантных
генов у птицы открыто для проверочного теста с помощью введения одно¬
го или нескольких таких доминантов в подлинно дикую линию джунглевой
курицы. Если мое представление верно, то мутант должен оказаться четко
промежуточным — ни полностью доминантным, ни полностью рецессив¬
ным. Благодаря доброжелательному отношению в Лондонском зоологиче¬
ском обществе и великодушию г-на С. Льюиса стало возможным начать
этот эксперимент; его результат, разумеется, станет известен только через
несколько лет .Процесс модификацииПример с птицей подтверждает по крайней мере другие имеющиеся
данные относительно скорости, с которой может изменяться доминант¬
ность; в этом случае, хотя весь процесс продолжался, вероятно, не более1 ООО поколений, селекцию применяли достаточно эффективно, и не в попу¬
ляции, содержащей единственную гетерозиготу на примерно 10000 особей,
но в стаде, где половина особей была гетерозиготной; более того, по нашей
гипотезе, для продолжения стада отбирали прежде всего именно гетерози¬
готных особей, а не птиц дикого типа. В условиях такой селекции эволюция
должна происходить во много тысяч раз быстрее, чем соответствующие из¬
менения рецессивности в природе.Что касается скорости этого процесса, то для мутации заданной жиз¬
неспособности (v) и частоты мутирования (к) основное неизвестное — ве¬
личина модифицирующей дисперсии, достаточная чтобы воздействовать
на гетерозиготу. Предположительно, она будет стремиться к нулю, так как v
стремится к единице, однако ее отношение к v при значениях последне¬
го, существенно отличающихся от единицы, также будет разным в соот¬
ветствии с разными точками зрения на то, каким образом осуществляется
модификация изменения. В случае с гомозиготами можно предполагать,
что гены-модификаторы путем интенсификации соответствующих процес¬
сов развития достигнут положительного результата в смысле исправления
ситуации, которая возникает на той стадии, когда начинается нарушение
процесса развития. Это же может иметь место и для гетерозигот, и ес¬
ли большая часть наблюдаемой модифицирующей дисперсии именно тако¬
Эволюция ДОМИНАНТНОСТИ63го рода, следует ожидать, что ее величина, ceteris paribus*, будет зависеть
только от v, и, следовательно, что все мутации будут следовать одна за дру¬
гой сходным путем в сторону нормальности и со скоростями, пропорци¬
ональными частотам мутирования, но зависящими только от той стадии,
на которой они находятся в каждый конкретный момент. Такой подход по¬
казан на рис. 4.1.009 я
0.8 ^0.7 S0.6 10.5 g0.4 15 4 3 2 1время в произвольных единицахРис. 4. Соотношение между уровнем вредности мутации и временем, необходимым
для восстановления дефекта путем отбора модификаторов; предполагается, что дис¬
персия v пропорциональна и(1 — v)С другой стороны, то, что нам известно сегодня, показывает, что нель¬
зя исключить, что большая часть изменений может быть вызвана причи¬
нами, специфическими для гетерозигот, например, связана с изменениями
участия то одного, то другого из гомологичных генов в реакциях, происхо¬
дящих в ядре, за которые они отвечают. При таком подходе действительная
величина дисперсии должна зависеть не только от реальной жизнеспособ¬
ности, но и от ее начального уровня; тогда для гетерозигот с тем же уровнем
жизнеспособности она окажется выше для мутаций, имеющих больший эф¬
фект. Тогда мы должны получить серию кривых, как показано на рис. 5.В любом случае конечные стадии приближения к нормальному состо¬
янию будут протекать быстрее всех других, и изначально вредная мута¬
ция может обусловить лишь очень небольшое изменение за то же время,
за которое другие мутации, возникающие примерно с такой же частотой,
успевают достигнуть полной нормальности. Конечно, следует ожидать, что
в большинстве случаев мы застанем те стадии, на которых изменения идут*При прочих равных условиях (лат.).
64Глава 3Рис. 5. Траектории улучшения гетерозигот в предположении, что модифицирующая
дисперсия зависит также от уровня начальной жизнеспособностимедленнее всего, и сравнительно большое число случаев в любом периоде
устойчивости. Относительно редкие «доминантные» мутации у Drosophila
можно рассматривать либо как сравнительно новые мутации или, что бо¬
лее вероятно, судя по их большему количеству, как мутации, при которых
гетерозигота на протяжении всей ее истории, испытывала столь суровые
затруднения, что мало что могло измениться в итоге. Предполагается, что
большая часть наблюдаемых мутаций находится в стадии покоя при полной
рецессивности, а в случае деталей, состояние которых должно быть совер¬
шенно неизменным, — число накопленных мутаций просто огромно34. Что
касается нелетальных мутантов, то после того, как гетерозигота становится
практически одинаковой с диким типом, имея очень малые различия в жиз¬
неспособности, можно ожидать начала изменения гомозиготы; и при том
же уровне жизнеспособности, и если считать, что гомозигота подвержена
изменениям ненамного меньше, чем гетерозигота, то и по скорости эти про¬
цессы должны быть сравнимы. Второй процесс, если не принимать в рас¬
чет никаких разниц в возможности модификаций, вероятно, должен быть
заметно более медленным, чем первый, так как можно ожидать, что гомо¬
зигота с самого начала поставлена в куда менее выгодное положение, хотя
возможно, что ее жизнеспособность случайным образом может существен¬
но повышаться в ходе процесса модификации гетерозиготы. Тем не менее
мы должны быть готовыми допустить, что бесчисленные мутации могли
происходить в прошлом, после чего даже гомозиготы стали с виду прак¬
тически нормальными, и что, следовательно, не остается следов, которые
Эволюция ДОМИНАНТНОСТИ65генетическое исследование могло было бы установить. Не известны при¬
чины, однако, почему такие гены не могли бы функционировать в особых
случаях, модифицируя эффекты редких мутантов.В этой связи интересный случай представляют два фактора: forked
и semi-forked у Drosophila melanogaster. Первый, forked — это сцепленная
с полом рецессивная мутация, при которой щетинки на голове, груди и щит¬
ке укорочены, закручены и выглядят более грубыми, чем у диких мух. По¬
скольку фактор сцеплен с полом, доминантность может проявляться толь¬
ко у самок, и обычные самки, гетерозиготные по forked, имеют щетинки,
не отличающиеся от диких мух. В 1918 г. доктор Лэнсфилд провел се¬
рию экспериментов по изучению этого фактора, в ходе которой обнаружил
ген semi-forked; этот ген не дает видимого эффекта у самок, гомозигот¬
ных по forked, как и у самцов forked-, он изредка проявляется в виде неко¬
торого укорочения щетинок у нормальных мух, но гетерозиготные самки
при наличии этого гена становятся явно промежуточными. Таким образом,
semi-forked действует как модификатор доминирования forked, имея сход¬
ное биохимическое действие с теми факторами, отбором которых, согласно
выдвигаемой здесь точке зрения, была достигнута его доминантность. Од¬
нако маловероятно, что semi-forked действительно является одним из этих
факторов, так как он сам рецессивен, о чем можно судить по его взаимодей¬
ствию с гетерозиготами forked. С другой стороны, он вполне может быть
старой мутацией, которая достигла такой стадии модификации, что даже
в гомозиготном состоянии практически никак не проявляется.Выводы из теории эволюции доминантностиОдин вывод, который легко сделать из приведенных выше соображе¬
ний, состоит в том, что широко известный факт — мутации обычно рецес¬
сивны,— не должен привести нас к предположению, что это также спра¬
ведливо и для мутаций, эффект которых полезен или нейтрален. Напротив,
у нас есть основания полагать, что это относится к тому классу мутаций, ко¬
торые постоянно возникают вновь, с частотой несколько меньшей, чем одна
на миллион, и которые столь же постоянно элиминировались Естественным
Отбором на протяжении многих тысяч или даже миллионов поколений.
Этот класс мутаций является и, вероятно, всегда будет наиболее ценным
для растениеводов и генетиков, которым он поставляет самые выигрыш¬
ные варианты, однако это не дает нам права считать, что он имеет и особую
важность для эволюции. Для мутаций не из этого класса мы не имеем ос¬
нований ожидать доминирования в любом направлении. Было бы разумно
предположить a priori, что при первом появлении мутации реакцию гетеро¬
66Глава 3зиготы должны контролировать через биохимическую активность в равной
мере два гомологичных гена, что в общем, хотя и не обязательно в каждом
конкретном случае, может приводить к образованию гетерозиготы, сома¬
тически промежуточной между двумя гомозиготами. Конечно, не следует
ожидать, что все различия окажутся количественно равными, для приме¬
ра можно представить, как бы мы оценили удаление половины черного
пигмента из какой-то черной структуры: удаление второй из двух частей
окраски показалось бы куда более заметным, нежели первой. Отсюда следу¬
ет, что говорить о равной функциональной значимости двух гомологичных
генов — не значит утверждать, что невозможно всякое проявление домини¬
рования, но имеется в виду, что в общем промежуточное проявление при¬
знака, как в случае с гетерозиготами между разными мутантами по одному
гену, должно было бы преобладать. Изменение вида в результате благопри¬
ятной мутации, таким образом, обычно будет происходить в виде двух по¬
чти равных шагов, последовательных и в одном направлении. Интересно,
что эта ситуация несколько напоминает последовательное возникновение
мутаций одного направления, как представлял Вейсман.Пример эволюции доминантности хорошо иллюстрирует две особен¬
ности Естественного Отбора, которые, несмотря на усилия Дарвина, все
еще представляют трудность для понимания теории, когда для объяснения
последней в качестве примера приводятся активные усилия животновода
в селекции его стада. Такими особенностями являются отсутствие жела¬
ния природы к улучшению расы, а также тот факт, что все модификации,
которые идут к тому, чтобы оказаться лучше представленными в будущих
поколениях, каким бы косвенным ни казалось их действие и как бы трудно
ни было распознать их действие, ipso facto оказываются результатом есте¬
ственного отбора. Здесь есть некоторая аналогия с дарвиновской теорией
полового отбора в той части, где речь идет о признаках, не являющихся по¬
лезными для организма в его отношениях со средой или другими организ¬
мами, или тех, которые не помогают особям противоположного пола найти
и соединиться друг с друг ом, но лишь о качествах, предпочитаемых про¬
тивоположным полом. Однако даже в этих случаях половой отбор создает
реальное преимущество половине особей вида в какой-то жизненной ситу¬
ации, тогда как при селекции доминантности генетическое изменение всего
вида приводит к структурному изменению несравнимо меньшей части по¬
пуляции. Если мы согласимся с аналогией Дарвина, сравнивающего Есте¬
ственный Отбор с человеком или сверчеловеком, выявляющим у каждого
индивида возможности прямых или косвенных преимуществ, то пример*В силу самого факта (лат.).
Эволюция ДОМИНАНТНОСТИ67с эволюцией доминантности наглядно показывает, с какой тщательностью
должно быть продумано такое исследование.В предыдущей главе мы видели, говоря в общем, что сложность осу¬
ществления любого улучшения в организме зависит от того, в какой сте¬
пени организм адаптирован к своей естественной среде обитания. В этом
смысле те трудности, которые должен преодолевать Естественный Отбор,
им самим и созданы, поскольку чем более активно он работает, тем ме¬
нее значительными и более замысловатыми будут изменения, на которые
опираются будущие улучшения. Тот факт, что организмы не изменяются
быстро, в теории можно интерпретировать либо как результат слабости от¬
бора, либо как следствие интенсивности адаптации с учетом сложности
взаимоотношений между организмом и его окружением. Мы не имеем пря¬
мого способа установления ни одной из этих возможностей, и то, что мы
имеем на выходе, можно выразить только конкретными терминами, указы¬
вающими на некие конкретные изменения, реальные или даже кажущиеся,
у некоторого конкретного организма. Как представляется, для этой цели
рецессивные мутации как раз подходят, и читатель, который принимает за¬
ключение по этой главе, поймет, что любая неадекватность того же порядка
величины, как те, о которых шла речь, и в равной мере поддающаяся изме¬
нению, будет элиминироваться Естественным Отбором в десятки и сотни
раз быстрее, чем реально достигается доминантность. Возьмем более ре¬
альный случай, вместо того чтобы предполагать, что вид в целом внезапно
изменяется, становясь так же плохо приспособленным к своей среде оби¬
тания, как знакомые нам мутанты; если предположить, что органическая
либо неорганическая среда вида вдруг как-то изменяется, или что колония
вида вдруг попадает в такие условия, к которым все особи приспособлены
в равной степени плохо, — мы имеем основание предположить, что эволю¬
ция адаптивных признаков должна будет происходить с той же скоростью.
Практически можно сказать, что каждая мутация и есть такой эксперимент
в небольшом масштабе.Что касается точности, с которой адаптация осуществляется в дей¬
ствительности, то нужно быть весьма осторожными и помнить, что все
гетерозиготы по всем рецессивным мутациям, включая, видимо, и тыся¬
чи рецессивных деталей, генетически различны. Однако, даже если конеч¬
ные результаты кажутся одинаковыми, формирование их осуществляется
при участии различных процессов развития, даже если первичные разли¬
чия сводятся лишь к внутриклеточным реакциям. Тот факт, что они весьма
похожи, можно интерпретировать только как указание на то, что схожесть
на этой стадии обязательна даже для такой почти нормальной адаптации,
какая требуется от довольно редкой гомозиготы. Если бы какое-то заметное
68ГЛАВА 3отличие формы было возможно в пределах такой почти равной адаптации,
мы бы непременно обнаружили ее среди всего этого множества гетерозигот.
Так как любые различия, которые могут существовать между ними, конеч¬
но, чрезвычайно малы, мы здесь имеем явное свидетельство той близости,
с которой достаточно успешная особь должна соответствовать видовому
типу, а также указание на верхний предел различий, которые имеют неко¬
торый шанс оказаться удачными.РезюмеИзучение доминантности среди возникающих мутаций, а также дру¬
гих генов, которые следует считать мутантными, показывает, что в боль¬
шинстве случаев дикий ген доминантен по отношению к мутантным, хотя
в небольшом числе случаев доминирование неполное. С другой стороны,
разные мутации одних и тех же диких генов регулярно проявляют взаим¬
ное отсутствие доминирования. Если замещение мутанта на исходный ген
сыграло какую-то роль в эволюции, эти наблюдения требуют, чтобы дикие
аллеломорфы обязательно становились доминантами по отношению к их
неуспешным конкурентам.Если можно положиться на то, что наблюдаемые частоты мутаций пра¬
вильно отражают порядок величин, то возникновение гетерозигот по каж¬
дой мутации среди предков дикой популяции должно было происходить
настолько часто, насколько этого было достаточно, чтобы обеспечить эво¬
люцию доминантности за счет отбора модифицирующих факторов. Этот
процесс идет очень медленно, так как в популяции эффективно подверга¬
ется отбору только 1 особь на 10000 или даже на 100000.При изучении хлопчатника был описан случай, когда явное полное до¬
минирование создалось у одного из группы близкородственных видов, име¬
ющих соответствующую мутацию; аномальное проявление доминирования
у домашней птицы можно трактовать как результат селекционной работы
человека в стадах, которые доступны для участия в скрещивании диких
петухов.Таким образом, теория эволюции доминантности объясняет большой
массив фактов, которые сегодня не имеют другого толкования. Если ее при¬
нять, она, похоже, могла бы пролить свет на природу мутаций, а также
на интенсивность адаптации; в частности, близкое сходство очень много¬
численных гетерозиготных генотипов весьма убедительно показывает, что
адаптивное значение, достаточное для того, чтобы направлять эволюцион¬
ные изменения, следует искать в различиях, уровень которых значительно
ниже видового35.
Глава 4Бариации как результат мутаций
и отбораИзмерение частоты гена. Вероятность сохранения отдельного гена; связь с рас¬
пределением Пуассона. Низкая частота полезных мутаций. Единичное возникнове¬
ние мутации не невероятно. Распределение отношений генных частот для факторов,
определяющих дисперсию. Малые эффекты случайного выживания. Число факто¬
ров, определяющих дисперсию.В первый раз случилось то,Что еще никогда не случалось. СОФОКЛИзмерение частоты генаВ главе 2 существенное внимание было уделено тому факту, что дис¬
персия, имеющаяся в любой группе организмов с интербридингом, не про¬
являет тенденции к снижению в процессе интербридинга при условии, что
дисперсия обусловлена различиями между отдельными генами, которые се¬
грегируют как целостные единицы из генетических комбинаций, в кото¬
рые они могут входить. В такой системе любые изменения вариабельности,
которые могут иметь место, следует относить на счет изменения частот,
включая как возникновение, так и исчезновение разных альтернативных
форм генов. В этой главе нам нужно разобраться в причинах, которые опре¬
деляют уровень проявления изменчивости, или, другими словами, понять,
при каком уровне изменчивости исчезновение одних и появление других
генов происходит с одинаковой частотой.Для начала вполне достаточно подробно рассмотреть случай с диморф¬
ными факторами, то есть с теми локусами, которые могут быть представле¬
ны только двумя альтернативными формами гена. Кажется вероятным, что
случаи, когда имеется три или более разных форм гена, для большинства
видов представляют явное меньшинство36, и их вклад в дисперсию малоза¬
метен. Однако это может иметь место, и подробный анализ, как представля¬
70Глава 4ется, может просто осложнить формулировку доводов и разработку необ¬
ходимых методов без введения какого-либо нового правила. Рассматривая
диморфные факторы, мы будем иметь дело с относительной частотой толь¬
ко двух форм генов, которую мы в предыдущих главах представили в виде
отношения р : q, а также обсудим причины, которые модифицируют эти
частоты. Для этого необходимо создать подходящую шкалу, которая позво¬
ляла бы количественно измерять такие изменения отношения частот. Конеч¬
но, для такого измерения можно было бы использовать шкалу процентов,
чтобы различать факторы соответственно процентам двух альтернативных
форм гена в локусе. Таким образом, мы должны отличить гены, у кото¬
рых каждая форма гена занимает 50% локусов, от тех генов, у которых
более многочисленная форма занимает 60, 90 или 99 %, а также рассчитать
с какой частотой можно ожидать обнаружения их в пределах, ограничен¬
ных этими значениями; иначе говоря, для какой части этих генов следует
ожидать, что их более частые формы будут занимать от 50 до 60% ло¬
кусов, для какой — от 60 до 70% локусов и т. д. В тех случаях, где вы¬
ражено доминирование, можно поставить те же вопросы по отношению
не к самым частым, а к доминантным генам; и тогда отличались бы те
случаи, в которых доминантны от 20 до 30 % генов, от тех случаев, когда
их оказывается от 70 до 80%. Однако для всех этих целей, имея в виду
реальную зависимость, которую предполагается обсуждать, было бы по¬
лезнее использовать шкалу, показывающую увеличение отношения между
двумя частотами в геометрической прогрессии. Начиная от случая, когда
частоты равны и каждая альтернативная форма гена занимает 50% локу¬
сов, мы должны далее рассмотреть отношения частот 2 : 1, 4 : 1, 8 : 1,
16 : 1 и т. д. в качестве равных шагов увеличения частот, хотя соответ¬
ствующие проценты будут равняться 66,7, 80,0, 88,9, 94,1. Такая шкала
симметрична. Если мы пойдем в противоположном направлении, то при¬
дем к соотношениям частот 1 : 2, 1 : 4, 1 : 6, 1 : 16 с дополнением
до 100%. Математически такая шкала эквивалентна измерению отношения
частот:Рz = log - = logp — log q.qЕсли логарифмы берутся по основанию 2, то все шаги будут иметь еди¬
ничную длину; если же мы возьмем (что более удобно математически) на¬
туральные логарифмы, то шаги по-прежнему будут равными, но их длина
составит около 0,7 от единицы. Есть два практических преимущества ис¬
пользования логарифмической шкалы для отношения частот диморфных
факторов: во-первых, она позволяет адекватно представить очень разные
Вариации как результат мутаций и отбора71отношения частот: как 1 миллиард : 1, которые возможны для многих ви¬
дов, так и куда более умеренные пропорции вроде 1000 : 1, которые были
бы совершенно неразличимы на иной шкале; во-вторых, эффекты отбора
в модификации генных частот на логарифмической шкале выглядят проще
всего, а именно как изменения положения, причем скорости этих измене¬
ний равномерны и пропорционально связаны с интенсивностью действия
отбора.Для факторов, не сцепленных с полом, каждая особь несет две одина¬
ковые или разные формы гена. Если перечислить всех индивидов и удво¬
ить полученное число, то максимум, которого может достигать отношение
частот, будет равен отношению удвоенного числа индивидов к единице.
Таким образом, амплитуда возможных отношений частот на логарифми¬
ческой шкале зависит от числа индивидов в популяции, и легко видеть,
что при увеличении числа индивидов в 10 раз она возрастает на величи¬
ну (2 log 10), или 4,6. Например, при числе 10000000000 индивидов раз¬
мах величин составит примерно от —23,7 до +23,7. В этом диапазоне око¬
ло 5 единиц с каждого конца шкалы представляют случаи, когда наименее
часто встречающаяся форма гена присутствует примерно лишь у 100 или
менее индивидов или точнее — примерно в 100 гомологичных локусах, так
как один индивид может иметь 2 такие формы гена. Другими словами,
для этих интервалов ясно, что более редкие гены подвержены угрозе ис¬
чезновения, а абсолютная длина этих интервалов на нашей шкале не бу¬
дет зависеть от числа индивидов в популяции. Между двумя крайними
участками шкалы располагается центральная часть, где обе альтернатив¬
ные формы сравнительно многочисленны, по крайней мере в том смысле,
что любой существует не менее чем у 100 индивидов. Именно длина этого
центрального — безопасного — интервала зависит от численности попу¬
ляции. Для 10000000000 индивидов длина составляет около 37 единиц,
для 100 млн — порядка 28 единиц. Таким образом, логарифмическая шкала
наглядно демонстрирует то значение, которое имеет численность популя¬
ции для сохранения дисперсии.В гипотетическом перечислении генов в популяции, о котором шла
речь в последнем абзаце, не говорилось о возрасте или репродуктивной
ценности упомянутых индивидов. Если же эти параметры принять во вни¬
мание, то в обоих направлениях не будет предела достижимой величине от¬
ношения частот, однако и тогда сохранится различие между относительной
угрозой потери более редких генов в крайних интервалах шкалы и отно¬
сительно более высокой вероятностью их сохранения в центральных. Это
утверждение основано на том обстоятельстве, что ген, который присутству¬
ет у дюжины индивидов, имеющих низкую репродуктивную ценность в том
72Глава 4смысле, о ком говорилось в главе 2, подвергается по крайней мере такой
же угрозе исчезновения, как и тот, что присутствует у единичного инди¬
вида, репродуктивная ценность которого равна сумме этих величин у упо¬
мянутой дюжины индивидов. Для взрослых форм вероятность выживания
должна быть практически одинакова. Однако если репродуктивную цен¬
ность, о которой мы говорим, обеспечивают только незрелые или личиноч¬
ные формы, в норме подверженные высокой смертности, то шанс потери
определенной величины репродуктивной ценности может существенно уве¬
личиваться. Поэтому удобнее полностью исключить незрелые формы из на¬
ших рассуждений и производить перечисление индивидов только когда они
достигают начала репродуктивного периода своего жизненного цикла. Мы
будем учитывать численность поколения вблизи максимума его репродук¬
тивной ценности, а также в период его наименьших значений. Таким спо¬
собом можно представить величину популяции вида, опираясь не только
на простой подсчет индивидов, охватывающий все возраста вплоть до са¬
мых младших чуть ли не от рождения, но на число индивидов в каждом
поколении, достигнувших репродуктивного периода. Для видов, дающих
несколько поколений в год, численность которых также сильно изменяется
в годичном цикле, вполне вероятно, что заключения о влиянии величины
популяции окажутся в наибольшей степени соответствующими интервалу
нормального годичного минимума численности.Вероятность сохранения отдельного генаОтдельный ген взрослого организма, который еще не участвовал в раз¬
множении, проявится в следующем поколении у определенного числа (О,1, 2, 3 и т.д.) индивидов или гомологичных локусов. У двуполых организ¬
мов они, конечно, должны быть разными индивидами, но если возможно
самооплодотворение, тот же самый ген может быть получен каждым из по¬
томков такого индивида от двух его гамет, а если такой индивид доживает
до зрелости, наш исходный ген будет представлен у него дважды. В целом
мы будем принимать во внимание общее число генов в популяции, хотя
было бы удобнее рассматривать их как принадлежащие одному индивиду.
Обозначим вероятности того, что потомки, получившие ген 0,1,2,... раз,
достигнут зрелости какР0,Р1,Р2,---,причем, поскольку хотя бы одно из событий обязательно произойдет, тоРО +Pl +Р2 +
Вариации как результат мутаций и отбора73Чтобы учесть шансы для будущих поколений, мы сначала вычисляем
соответствующие вероятности для случая, когда ген представлен у г инди¬
видов. Для большей точности возьмем математическую функцию37f(x) =ро+ Pix + Р2Х2 + ....Очевидно, что с увеличением х эта функция возрастает от ро (когда х = 0)
до 1 (когда х = 1). Более того, если г индивидов размножаются независимо,
то шанс исчезновения гена в течение одного поколения будет равен pjj.
Вероятность быть представленным единственным геном будет равнагРо~гР1,а в целом вероятность сохраниться s генам выразится коэффициентом
при Xs в раскрытии функции(Я*))г-Далее, начиная с одного гена, вероятность сохранить г во втором поколении
будет рг, а вероятность того, что уже они сохранят s в третьем поколении
будет представлена коэффициентом при Xs в формулеPrif{x)y.Отсюда следует, что вероятность сохранения s в третьем поколении, неза¬
висимо от числа представленных во втором поколении, соответствует ко¬
эффициенту при Xs вPo(f(x))° + Pi {fix))1 + P2(f{x))2 + • • ■ИЛИ вf(f(x))-Эта новая функция — точно такая же функция от fix), как f{x) — функция
от х, поэтому она занимает место fix), когда мы хотим рассмотреть отре¬
зок времени продолжительностью не в одно, а в два поколения; очевидно,
что для трех поколений надо будет использовать f{f{f(x))}, далее можно
учесть такое число поколений, какое требуется.Имеется достаточно оснований, чтобы предполагать, что если пере¬
счет привязан к условию раннего достижения зрелости, то функция f(x)
с достаточно хорошим приближением всегда будет иметь такую математи¬
ческую форму. Например, если мы рассмотрим любой организм, способ¬
ный дать существенное число потомков, скажем, такой как перекрестно-
опыляемый злак, то выяснится, что каждое репродукгивно зрелое растение
74Глава 4является материнским по отношению к заметному числу, примерно для со¬
рока зрелых зерен, и отцовским в среднем для такого же числа. Каждый
отдельный ген из пары попадает в каждое из зерен с вероятностью 1/2- Од¬
нако в урожае следующего года в среднем лишь 2 зерна из всех окажутся
представленными зрелыми растениями, поэтому шанс для обоих генов по¬
пасть опять в зерна и не быть утерянными составит только 1 к 80. Отсюда,
вероятность повторного появления генов в следующем году у 0, 1, 2, ...
индивидов даст коэффициенты ж0, х1, х2,... при разложенииЭти коэффициенты уже очень близки к членам ряда распределения
Пуассона:и были бы идентичны им, если произвольное число 80 заменить беско¬
нечностью. Отклонение от ряда распределения Пуассона в действительно¬
сти можно считать следствием искусственных предположений, которые бы¬
ли сделаны для упрощения расчетов. Мы произвольно предположили, что
все растения дают равное число зерен, тогда как реально эти числа будут
меняться. Число пыльцевых зерен с каждого растения, которое участву¬
ет в формировании зерен, тоже будет разным, что также будет несколько
изменять форму распределения, создавая ограничения для распределения
Пуассона. В действительности биномиальное распределение, полученное
выше, отличается систематически от предельной формы, так как изменчи¬
вость по числу зерен у разных растений, по-видимому, несколько выше того
уровня, который требуется в истинном ряде распределения Пуассона.Общий характер ряда распределения Пуассона, который делает его
приемлемым для нас, состоит в том, что он возникает тогда, когда каж¬
дый из большого числа индивидов получает небольшой независимый шанс
на успех; если число индивидов и шансы каждого таковы, что преуспевают
в среднем с особей, то число действительно достигших успеха в разных
испытаниях будет распределяться в соответствии с формулойили
Вариации как результат мутаций и отбора75Таблица 2ЧислоВероятность вымиранияВероятность выживанияпоко¬Без преи¬Один процентРазностьБез преи¬Один процентлениимуществапреимуществамуществапреимущества10,36790,36420,00370,63210,635830,62590,61970,00620,37410,380370,79050,78250,00800,20950,2175150,88730,87830,00900,11270,1217310,94110,93130,00980,05890,0687630,96980,95910,01070,03020,04091270,98470,98290,01180,01530,0271Limit1,00000,98030,01970,00000,0797и это обычно можно считать хорошим приближением для определения шан¬
сов отдельных гамет, произведенных конкретным взрослым индивидом.Если аллель не несет селективных преимуществ или дефектов, с будет
равно 1; величины Po,Pi,P2, ■ ■ ■ будут получены с помощью ряда распреде¬
ления Пуассона6 1 (1’1’ 21’ 3!’' ’ ’)’а функция f(x) примет видилиf{x) = е*-1.Далее, если данный ген увеличивает частоту в каждом следующем по¬
колении в пропорции с: 1, то по аналогии мы получим/(*) = е^-1).Получив такие выражения для f(x), можно вычислить вероятность
выживания, увеличения числа или исчезновения потомков, несущих еди¬
ничный конкретный ген, просто заменяя значения fix) на х. В таблице 2
первый столбец показывает число поколений от начального момента, при¬
чем числа эти были выбраны таким образом, чтобы можно было просле¬
дить ход изменений в длинной череде поколений с помощью относительно
76Глава 4компактной таблицы. Эти изменения быстрее всего идут на первых этапах,
поэтому мы выбрали шаги через 1, 2, 4, 8 поколений и внесли в таблицу
результаты, достигнутые по прошествии 1, 3, 7, 15, 31, 63 и 127 поколений.
Во втором столбце показаны вероятности исчезновения на каждой стадии
для генов, не имеющих селективных плюсов и минусов. Числа можно так¬
же читать, не обращая внимание на знак, отделяющий целую часть числа
от дробной, как число случаев из 10 ООО, в которых потомки исходного ге¬
на должны будут исчезнуть. Доля таких вымираний в череде поколений
сначала будет очень высока, примерно 3 из 8 исчезают уже в первом по¬
колении, а из 5 остальных еще 2 исчезают к третьему поколению. Через
15 поколений исчезнут почти 8 из каждых 9. Далее эти потери становятся
намного реже, всего около 2,87 % будет утеряно между 31-ми 63-м поколе¬
ниями и лишь 1,49 % — от 63-го до 127-го, когда все еще сохранившихся бу¬
дет 1,53%. Лучше всего число выживших можно проследить по столбцу 5
(табл. 2), из которого видно, что при тех интервалах, которые мы выбрали,
доля выживших с каждым шагом все больше приближается к половинной
убыли по сравнению с предыдущей ступенью; фактически при больших
значениях числа поколений (п) шанс сохранения в течение п поколений
очень близок к величине 2/п.Для сравнения в соседние столбцы таблицы были занесены соответ¬
ствующие числа для генов, у которых с = 1,01 и которые, следовательно,
имеют преимущество в 1 %; разности показаны в столбце 4. Можно видеть,
что при неограниченном увеличении п селективное преимущество в конеч¬
ном счете достигает уровня выживания, не доходящего до 2% от исход¬
ного числа случаев, причем на первых стадиях, когда исчезновение идет
очень быстро, преимущество нарастает очень медленно. Из 10000 мута¬
ций, имеющих селективное преимущество в 1 %, которые уже наличеству¬
ют у одного половозрелого индивида, 3 642 не смогут удачно передать ген,
имеющий преимущество, ни одному из потомков, тогда как среди не име¬
ющих каких-либо преимуществ такими «неудачниками» окажутся 3 679.
Даже спустя 31 поколение число выживших из общего числа 10000 бу¬
дет составлять только 687 — против ожидаемого числа выживших в 589
из тех, кто не имеет никакого преимущества. Фактически селективное пре¬
имущество порядка 1 %, хотя и считается достаточно мощным, чтобы обу¬
словить эволюционные последствия, регулярные и четкие, когда речь идето числе особей порядка 1 млн, оказывается почти не ощутимым по срав¬
нению со случайными процессами, если дело касается небольшого числа
особей. Мутация, даже если она благоприятна, имеет очень низкие шан¬
сы закрепиться, если она возникает единственный раз. Если ее селектив¬
ное преимущество составляет только 1 %, она должна возникнуть 50 или
Вариации как результат мутаций и отбора77даже 250 раз, чтобы смочь закрепиться у достаточного числа половозре¬
лых индивидов и тогда получить некие перспективы для проявления в бу¬
дущем.Если допустить, что шанс сохранения и распространения мутации
с преимуществом выживания в 1 % составляет 1 к 50, то станет ясно, что та¬
кие мутации не могут возникать сколько-нибудь часто до тех пор, пока они
не будут наблюдаться хотя бы у одной из особей. Когда шансы составляют
свыше 100 к 1, все первые 250 мутаций такого благоприятного типа ис¬
чезнут. Следовательно, успех такого рода мутации должен устанавливаться
в то время, когда частота такой мутации очень низка, ибо у вида, у которо¬
го в каждом поколении до половозрелого состояния доживает 1 000 000 000
особей, частота мутаций, равная 1 на тысячу миллионов, обеспечит появле¬
ние одного мутанта в каждом поколении, что создаст превосходство нового
типа в течение менее чем 250 поколений, причем весьма вероятно, что даже
не далее, чем в ряду не более 10 поколений с момента первого возникнове¬
ния мутации; в то же время если новая мутация возникает с более привыч¬
ной частотой 1 на 1 000 000, то весь процесс мог бы завершиться при из¬
вестных условиях уже в первом поколении. Следует допускать, что частоты
мутаций, как и другие признаки организмов, в ходе эволюции изменяются
лишь постепенно; однако в то время как частота неблагоприятных мутаций
может достигать и даже превышать 1 на 1 000000, не обнаруживая замет¬
ного влияния на признаки вида, благоприятные мутации едва ли могли бы
возникать сколько-нибудь длительно, даже при частотах в 1 000 раз ниже,
и мы не можем исключить вероятности того, что возникающие благоприят¬
ные мутации, повышающие приспособленность, могут происходить очень
редко, даже единично, например, однажды в ряду тысяч поколений. Коли¬
чественное сравнение частот мутаций по гомологичным локусам у разных
родственных видов могао бы существенно помочь прояснить сложный во¬
прос о том, насколько быстрыми можно считать изменения (как повыше¬
ния, так и снижения) частот мутаций.Когда отсутствуют селективные преимущества или недостатки, доля
случаев, в которых исчезновение не происходит через п поколений, состав¬
ляет приблизительно 2/п. Отсюда, поскольку при отсутствии селективно¬
сти ожидаемое для любого будущего поколения равно числу ныне живущих
особей, то среднее число индивидов, у каждого из которых будут присут¬
ствовать эти сохранившиеся гены, будет равно п. Однако это число будет
существенно различаться в разных случаях, и было бы интересно получить
конкретную кривую этого распределения.Это можно сделать, если наблюдается следующая картина: если часто¬
та, с которой возникает каждое число мутаций, является коэффициентом
78Глава 4в показателе степени разложении формулыф(х),тогда, заменяя х величиной еь, мы получим ф как обобщающую формулу
моментов распределения. Далее, чтобы продвинуться на одно поколение,
нужно заменитьилих на ех 1,е4 на ееилиt на е* — 1;таким образом, если /лг, Мз, • • • — это моменты распределения, близкие
к нулю, при возникновении предыдущего поколения, а для последующего
поколения они будут коэффициентами при /;, раскрывая формулу1 9 1 , \ 1 ( 1 оI I II _1 +* .+ 6* +"7 + 2M2V + 2* +"+ -g МЗ {t + • • -)3 + 7^ М4 (t + . . .)4 + . . .в виде степеней t; и если их обозначить как ц'2, Мз, • •то мы получим
равенства:Mi = Mi,М2 = М2 + Ml,Мз = Мз + ЗМ2 + Mi,М4 = М4 + бмз + 7м2 + Mi-
Поскольку все моменты исходно равны единице, то легко видеть,
что М2 будет увеличиваться пропорционально гг, ц,3 — пропорционально п2,
/М — пропорционально п3 и так далее, когда значение п велико. Далее, так
как в общем виде, — 1)Мр — Мр Н ^ Мр—1 + • • ■,коэффициент np_1 в равен -ip коэффициента пр~2 в Мр-Ь поэтому
начиная с Mi = 1 мы имеемпервую аппроксимацию, когда значение п велико.
Вариации как результат мутаций и отбора79Понимая значения моментов, мы можем сделать вывод о типе распре¬
деления, так как полученные нами значения моментов будут воспроизво¬
диться, если вероятность того, что число индивидов превысит значение х,
равнае-2 х/п_Некоторый интерес представляет заключение о том, что при отсут¬
ствии благоприятствующего отбора число индивидов, несущих ген, изме¬
ненный единственной мутацией, не может существенно превысить число
поколений, которые сменились после возникновения мутации. В действи¬
тельности, вероятность того, что х могло бы превысить величину З-g п,
меньше чем 1 к 1 ООО. Поэтому, если мутантная форма существует у 1 мил¬
лиона особей в каждом поколении, можно быть уверенным в том, что
по крайней мере до какого-то уровня число их увеличилось благодаря
действию отбора, который является процессом относительно быстрым,
или вследствие повторных мутаций, которым отбор не препятствовал, что
обычно бывает процессом куда более медленным, или же, если мы должны
предположить, что все было запущено единственным актом мутации и на¬
блюдаемый уровень численности достигнут благодаря случайным подъ¬
емам, что процесс продолжался в течение как минимум 280 млн поколе¬
ний — это делает его самым медленным, причем для таких больших чи¬
сел — во много раз менее вероятным.Аналогичное исследование распределения числа потомков, несущих
отдельные гены с небольшим селективным преимуществом, показывает,
что конечная форма распределения в этом случае такая же. Поэтому те¬
перь величину вероятности превышения числа х после п поколений можно
записать следующим образом:g—2(с—1)*с-пэто показывает, что когда с больше единицы, числа будут превышать любые
указанные значения х в достаточно продолжительной череде поколений.Формула будет корректно выражать распределение до тех пор, пока
величинас"2(с-1)все еще составляет небольшую часть от общего числа индивидов вида,
однако она, видимо, отражает лишь распределение для тех индивидов, ко¬
торые являются потомками мутаций, возникших в одном и том же поко¬
лении. Это искусственное и необязательное ограничение, поскольку, как
80Глава 4мы видели, в случае благоприятных мутаций те, что появились раньше,
первыми получат возможность закрепиться и далее, после некоторых до¬
вольно немногочисленных испытаний не будет необходимости в дальней¬
ших таких же мутациях. Мы должны предположить, что когда возникают
благоприятные мутации, они в редких случаях могли появиться и ранее,
и что частота их возникновения обычно увеличивается. У нас нет прямых
сведений о природе этого увеличения, но если оно связано с изменением
генотипической структуры вида, нам придется предположить, что оно про¬
исходит постепенно, и поскольку частоты негативных мутаций ничтожны,
то простейшее возможное допущение сводится к тому, что относительная
скорость увеличения за поколение может быть выражена малой величи¬
ной к, так что общий прирост за каждое поколение будет составлять ек.Исходя из этого, число мутаций, которые на некотором этапе уже будут
представлены более чем у х индивидов, будет пропорционально величинечто оказывается, когда х достаточно велико для (с — 1)х, достигая значе¬
ний 4 или 5, и велико по сравнению с к/ log с — 1, весьма близким по про¬
порциональности выражениюВ этой формуле можно узнать выражение log с, которое представляет
ту величину, на которую частота мутантного гена может увеличить маль¬
тузианский параметр (см. главу 2), что приблизительно соответствует по¬
казателю селективного преимущества 0,01 из наших количественных при¬
меров. Он отражает меру относительной скорости повышения частоты об¬
суждаемого гена так же, как к измеряет то же для частоты мутаций. Ес¬
ли предположить, что частота мутаций полностью зависит от присутствия
определенных групп генотипов, то придется принять, что log с и к — по¬
казатели того же типа и того же порядка величины, хотя и необязательно
примерно равные. Если мы допускаем, что среди генных замещений, спо¬
собных повлиять на частоту какой-либо из конкретных мутаций, возможны
переходы в разных направлениях, и что в целом повышение частот мута¬
ций вследствие повышения частоты встречаемости некоторых генотипов
будет частично компенсироваться за счет исчезновения других генотипов,
у которых также происходит мутирование, то вполне вероятно, что к часто
должна быть очень малой величиной. Если мы обратим внимание толькооо0х-к/log с
Вариации как результат мутаций и отбора81на мутации, имеющие селективное преимущество всего в 1 %, то придется
сказать, что когда такие мутации только начинают возникать, частота их
повышается не столь быстро, чтобы удвоиться или утроиться за 100 поко¬
лений, однако не исключена вероятность, что за это время прирост соста¬
вит порядка 10%. Если соотношение к/ log с мало, то практически из это¬
го следует, что значительное число тех мутантных генов, которые в итоге
присутствуют у каждого вида, произошли в результате одной единственной
мутации, которая на старте получила удачную возможность закрепиться
с заметной частотой, тогда как лишь крайне малая часть таких мутантных
генов могла возникнуть за счет множества всех сходных мутаций, оказав¬
шихся благоприятными в той или иной степени. Если же значения к/ log с
велики, то мутантные гены, хотя и в разных количествах, должны были бы
возникать в результате большого числа отдельных мутаций, ни одна из ко¬
торых не могла обеспечить заметную часть их общего числа.Следует отметить, что для начальных стадий, на которых определяется
выживание, с — это абсолютная скорость увеличения частоты мутантного
типа, и лишь с большим приближением можно рассматривать ее как пока¬
затель селективного преимущества над другими генотипами. Различие ста¬
новится явным, если рассматривать не неизменную популяцию, как до сих
пор, но такую, численность которой возрастает или снижается. В растущей
популяции мутации, не имеющие селективного преимущества либо даже
вредные, при условии, что их частота гаже, чем скорость прироста ви¬
да в целом, имеют некоторую вероятность не исчезнуть совсем; тогда как
в популяции, где имеет место снижение численности, даже те мутации, ко¬
торые имеют некоторое селективное преимущество, однако слишком малое
относительно скорости сокращения вида, будут иметь худшие перспективы,
чем нейтральные мутации у вида в неизменном состоянии. Как следствие
этого, растущие популяции приобретают больше возможности быть измен¬
чивыми, чем стационарные, а угасающие популяции — меньше; если же
интенсивность селективного действия в обоих случаях одинакова — можно
ожидать, что растущие популяции будут становиться более изменчивыми,
а снижающие численность — менее разнообразными в результате действия
процесса, который отличен от того, как влияет на изменчивость собственно
численность популяции. По крайней мере частично влияние прироста будет
увеличивать последствия влияния численности, поскольку далее будет по¬
казано, что в популяциях большей численности статистическое равновесие
будет устанавливаться при более высоком уровне дисперсии, а прямое вли¬
яние роста популяции должно увеличивать дисперсию быстрее, чем про¬
явится эффект более медленного процесса установления статистического
равновесия.
82Глава 4Масштаб этого процесса лимитирован реальными скоростями приро¬
ста или убыли природных популяций, и я предполагаю, что такие изме¬
нения редко бывают настолько велики, чтобы обеспечить прирост в одну
сотую на протяжении 10 ООО поколений, или около 1 на 2 ООО в каждом по¬
колении в течение всего этого времени. Насколько велико может быть зна¬
чение благоприятных или вредных мутаций при частотах менее 1 на 2 ООО,
совершенно неясно. Конечно, оно должно быть выше всего там, где адап¬
тивность (в том смысле, как представлено в главе 2) наиболее выражена,
и было бы по крайней мере необоснованно полагать, что такие малые из¬
менения обычно либо редьси, либо не имеют существенных эволюционных
эффектов, хотя в действительности может быть и так.Распределение отношений генных частот для факторов,
определяющих дисперсиюТеперь нам нужно рассмотреть зависимости, которые должны связы¬
вать генетическую вариабельность, характерную для вида, и частоту воз¬
никновения мутаций. Фундаментальная теорема, доказанная в главе 2, под¬
готовила нас к пониманию того, что дисперсия, поддерживающаяся в при¬
способленности по выживанию, должна быть тесно связана с частотой воз¬
никновения благоприятных мутаций, хотя частично она обусловлена посто¬
янным возникновением неблагоприятных мутаций такого типа, как описано
в главе 3, и является эффективной, только если вид постоянно освобож¬
дается от этих дефектов. Такие устойчиво возникающие неблагоприятные
мутации вносят свой вклад также в дисперсию всех мерных признаков; это¬
му содействуют те случаи, когда отношение генных частот поддерживается
в устойчивом равновесии селективными факторами, о которых будет сказа¬
но больше в главе 5, и те случаи, когда преимуществам признака в каком-то
регионе или стации, где вид обитает, противостоят помехи в других ситу¬
ациях, как в случае, эволюционные последствия которого будут описаны
в главе 6. Далее мы рассмотрим сохранение мутаций той менее явной и бо¬
лее изменчивой части дисперсии, которая обеспечивается благоприятны¬
ми мутациями, а также мутациями, селективные преимущества или недо¬
статки которых настолько малы, что ими можно пренебречь. Часть нашей
проблемы сводится к тому, чтобы определить, насколько малым должно
быть такое селективное преимущество или недостаток. Как было показано
в главе 2, благоприятные мутации обычно должны быть чрезвычайно ма¬
лы по своему соматическому эффекту, а их индивидуальные частоты столь
низкими, что мы практически имеем дело не с каким-то поддающимся из¬
Вариации как результат мутаций и отбора83мерению потоком мутаций, а с их индивидуальным спорадическим воз¬
никновением. Напротив, мутации, эффект которых близок к нейтральному,
могут иметь достаточно времени, чтобы достичь заметных частот, так как,
даже если частоты были высокими, потребуется не менее миллиона или да¬
же больше поколений для того, чтобы возник новый тип, и это дает время
для повышения частоты мутации относительно ее первоначального крайне
низкого значения. Несмотря на эту небольшую разницу, оба случая можно
рассматривать вместе.Чтобы определить влияние каждого из элементов, необходимо рас¬
смотреть три случая. Во-первых, распределение отношения генных частот
в ситуации, когда при отсутствии отбора или мутирования дисперсия по¬
степенно снижается за счет случайного исчезновения генов. Далее, распре¬
деление при условии, что дисперсия поддерживается новыми мутациями
без влияния отбора; наконец, распределение, формирующееся при наличии
слабого селективного преимущества или вреда.Самый эффективный способ рассмотрения первых двух из указанных
проблем состоит в том, чтобы получить функциональное уравнение для се¬
рии крайних частот. Если число индивидов, участвующих в размножении,
для каждого поколения равно п, при большой численности в многие милли¬
оны или тысячи миллионов, вероятные значения частот гена р будут состав¬
лять 1/2п, 2/2п, ...; этих возможных значений множество, и для большей
области распределения мы можем принимать р как непрерывную перемен¬
ную. В крайних областях, однако, необходим более точный подход, и здесь
мы примем упрощающее предположение, что когда статистическое равно¬
весие установилось, численность популяции не влияет на форму итогового
распределения.Если теперь обозначить как bi, b2,63,... частоты значений р = 1/2п,
2/2п, 3/2п,..., то мы сможем описать функцию какф(х) = Ь\х + b2x2 + Ъ3 х3 + ...,и условия статистического равновесия выразятся в виде уравнения, реше¬
нием которого будут частоты 61,621 63,..., а отсюда и распределение отно¬
шения генных частот. В случае исчезновения и отсутствия мутаций, возни¬
кает вопрос: каковы должны быть значения коэффициентов Ь, чтобы имен¬
но один ген исчезал в каждом поколении? Сумма значений этих коэффици¬
ентов в этом случае даст число генов, вносящих свой вклад в дисперсию,
и отсюда мы можем определить зависимость между дисперсией и скоро¬
стью ее снижения под влиянием случайного исчезновения генов.Если исчезновение происходит при скорости один ген за каждое поко¬
ление, то можно предположить, что в половине случаев число присутству-
84Глава 4ющих генов снижается от 1, 2, 3, ... до 0, а в другой половине случаев оно
увеличивается от 2п — 1, 2п — 2, 2п — 3, ... до 2п. Конечно, аллели, пред¬
ставленные у 0 индивидов, дадут коэффициент для х° при выражении ф,
так что через одно поколение функция ф, отражающая распределение на од¬
ной ветви, должна увеличиться на У2.Но для одного поколения мы уже видели, что ф(х) заменяется формулойФ(ех~1У,и, соответственно, уравнение получает видф(ех~1)-ф(х) =Чтобы упростить функциональное уравнение такого рода, необходимо
ввести такую функцию ии по аргументу v, чтобыuv+1 =еи"~1.Если этому уравнению удовлетворяет любая функция f(y), то будет
пригодна и функция F[y) = f(u + k), следовательно, мы можем произволь¬
но принять величину wo = 0, откуда uv при любых целых положительных
значениях v можно определить путем прямой подстановки. На практике
значения v для дробных значений v получают с помощью интерполяции
по последовательности целых чисел около v = 20, рассчитывая более низ¬
кие значения по интерполированным согласно соотношениюи„_ 1 = 1 + logu„.Теперь мы можем записать функциональное уравнение для ф в видеф{ии+х) - ф{ии) =откуда следует, что ф должна быть такой же функцией от х, как -// от и.
Таким образом, исходные частоты могут быть определены из дифференци¬
альных коэффициентов при v относительно и при и = 0, тогда как зависи¬
мость частот для больших значений р можно вывести согласно изменению
функции v по мере того, как и стремится к единице.Теперь, заменяя1
Вариации как результат мутаций и отбора85получаем обратную формулу:1 1 1l-e-i/vu " 2 12и„ 720г>3поэтому, когда и стремится к бесконечности, и должна стремиться к еди¬
нице, а значение vv все более приближаться к выражению1 1 ,- v + - log V + с,в котором численное значение с, как было показано, примерно равно
0,899144. Результат показывает, что величина г/(1 — м) стремится к едини¬
це вместе с и, и, следовательно, при р = г/2п частота стремится к единице
по мере роста г.Далее следует, что1 1 1— V = V log!) -С,где с' приближается к примерно 1,014649, когда v становится бесконечно
большой, и, следовательно,1 и 1 1 (Л ^2 " 6 8< ~ “>приближается к значению —0,014649, когда и = 1. Помимо этой ограни¬
ченной области частот, распределение поэтому задается разложением в ряд
Маклорена, имеющим вид+ т log(l - х)1 — х 6
или5 11 2 17 ,6Х+иХ +Т81 + —а коэффициенты этого ряда можно принять как второе приближение к ча¬
стотам факторов, самые редкие гены которых появляются в 1,2,3,... ло-
кусах.Реальные значения коэффициентов с некоторой потерей точности мо¬
гут быть получены с помощью табулирования значений функции ии\ они
приведены в таблице 3.
86Глава 4Таблица 3Предельные частоты факторов, у которых исчезает более редкий ген
Число локусов,
в которых исчезает
более редкий ген1
2
3456ВтороеприближениеИстиннаячастотаРазность0,8333330,818203-0,0151310,9166670,916762+0,0000960,9444440,944923+0,0004790,9583330,958266-0,0000670,9666670,966634-0,0000330,9722220,972225+0,000033Из таблицы 3 можно видеть, что почти вся небольшая разница
0,014649 приходится на первые два члена, отсюда следует, что значение
частоты достаточно хорошо соответствует выражению 1 — 1/6г. Предель¬
ные частоты показаны на рис. 6.Общее число генов в таком распределении теперь можно представить
в следующем виде:— 0,014649 j»,где 7 — это константа Эйлера, равная 0,577216. Остальной частью выра¬
жения можно пренебречь по сравнению с величиной 2гг, так что получен¬
ное решение показывает уменьшение дисперсии только одной части из 2гг
для каждого поколения.Это очень медленная скорость снижения; если бы дисперсию вида
можно было представить как обусловленную факторами, на которые отбор
не оказывает влияния, и при условии отсутствия новых мутаций, то сниже¬
ние дисперсии имело бы вид экспоненты, так что спад через т поколений
должен составить величину согласно формулето есть стать наполовину меньше через 2nlog2, или примерно через 1,4п
поколений. Никакой результат не мог бы более убедительно выявить кон¬
траст между сохранением дисперсии согласно теории корпускулярной на¬
следственности и ее рассеиванием в процессе наследования в случае при¬
нятия слитной теории.!:(7 + log 2 п)
Рис. 6. Частоты для вариантов, когда факторы представлены 1,2,3,... генами в об¬
щей популяции, при наличии устойчивого исчезновения и без мутаций. Верхняя
линия — частота, равная 1, для каждого варианта (предельное значение). Случайное
выживание будет удалять гены с частотой один на каждые два поколения, тогда как
вид распределения не изменяетсяКогда я обращался к этой проблеме раньше, ошибочный метод привел
меня к правильному пониманию распределения факторов, которые участ¬
вуют в формировании дисперсии в состоянии хронического упадка, однако
оценка времени релаксации получилась равной 4п вместо 2п поколений.
Профессор Сьюэл Райт из Чикаго, который независимым путем получил
правильные числа, указал мне на этот разнобой и таким путем вынудил
меня более тщательно рассмотреть проблему в целом.Исключительно низкая скорость естественного уменьшения диспер¬
сии является следствием того факта, что для большинства факторов от¬
ношения частот генов не слишком резко различаются. Распределение z
для этого случая показано на рис. 7, где можно видеть, что почти
для всех генов значения находятся в интервале от —6 до +6, то есть
что более редкие гены едва ли когда-нибудь занимают менее 1/400
всех доступных локусов и потому мало подвергаются опасности исчез¬
новения.Ранее разработанный метод имеет определенные преимущества для изу¬
чения распределения в его центральной области. Если 0 есть некая мера
частот генов, то частоту в любом дифференциале d0 можно обозначить
как у (10, и условие статистического равновесия может быть записано в ви¬
де дифференциального уравнения для неизвестной функции у. Вводя пере-
88Глава 4значения zРис. 7. Распределение величины отношения генных частот z, когда дисперсия рав¬
номерно снижается, при отсутствии мутаций и селективности. Время релаксации
в 2 раза больше, чем число поколений и число родительских особей в каждом по¬
колениименную видаcos в = 1 — 2р,где в — угол в радианах, который увеличивается от 0 до 7Г, когда р возрас¬
тает от 0 до 1, я в 1922 году получил уравнение, подобное тому, которым
описывают теплопроводность:ду _ 1 д2у
дт 4п дв2 ’решение которого у = sin в дает состояние устойчивого снижения со вре¬
менем релаксации, равным 4п поколениям. Корректный вид дифференци¬
ального уравнения должен, однако, выглядеть иначе:дУ _ 1 9 i д2уЭт Апдв^У ^ 4пд02’
что при том же пути решения приводит к вычислению правильного значе¬
ния времени релаксации.Во втором случае, который нужно рассмотреть, когда наблюдаемая
дисперсия поддерживается в статистическом равновесии на фоне постоян¬
ного возникновения новых мутаций, мы можем сразу применить этот метод
с помощью подстановки ду/дт = 0. Далее интегрированием получаемду— +ycotd -А,
Вариации как результат мутаций и отбора89где А — некоторая константа, тогда как— (у sin#) = A sin в,у sin 9 = —A cos 9 + В,у = В cosec 9 — A cot 9.Симметричное решение делает у пропорциональным cosec в. Для пере¬
менной z этому соответствует плосковершинное распределение, причем все
равные интервалы dz равновероятны, по крайней мере в центральной ча¬
сти, где дифференциальное уравнение только и является обоснованным.
Поскольку при 9 = 7г, cosec в + cot 9 = 0, можно также принять во внима¬
ние решениеу = В(cosec 9 + cot в),пригодное для случая, когда принимается, что все мутации происходят
при 0 = 0.В любом случае интеграл для всей области является неопределенным
вследствие быстрого роста у при в — 0. Из этого не следует, что общее чис¬
ло факторов бесконечно, так как именно в этой области дифференциальное
уравнение непригодно. Если перейти к выражению для р, то член (cosec в+
+ cot 9) будет эквивалентен2 qdp _ dp
2pq р 'так что полученное асимметричное решение соответствует положению, ко¬
гда частота при р = г/2п пропорциональна 1/г, по крайней мере при боль¬
ших значениях г. Тогда общая частота будет, очевидно, включать выраже¬
ние log(2n), но для того, чтобы определить его величину, важно исследо¬
вать граничные условия.Если ф(х) снова представить как функцию, коэффициенты разложения
которой в степенях х соответствуют частотам, устанавливающимся при р =
= 1/2п, 2/2п,... при единичной мутации в каждом поколении, то функци¬
ональное уравнение для ф будет выглядеть какф{ех~1) — ф(х) = 1 — х,где левая часть представляет собой изменение функции ф(х) в результа¬
те случайного скрещивания в одном поколении, тогда как эффект одной
мутации должен увеличивать коэффициент при х на единицу и снижать аб¬
солютную величину (х° ) на единицу. Для решения уравнения можно снова
90Глава 4подставить вместо х величину uv и получить уравнение
ф(ии+1) - ф{и„) = 1 -uv.Теперь из уравненияuv+1 =после дифференцирования по is, получим, что?// — рии~или чтоlog u'v+1 - logu^ =14^ — 1.Поскольку уравнение для ф можно записать какФ(и„+1) - ф{и„) = -(logu'v+l - logu'v),это уравнение имеет решение, если ф(ии) отличается от — log u'v на кон¬
станту. Константа для ф(х), которая отражает частоту факторов, не пред¬
ставленных ни у одного индивида, конечно, является произвольной,
и при условии, что <^>(0) = 0, получаем решениеф(и„) = log Uq - log u'vили, если записать v' как дифференциальный коэффициент от v относи¬
тельно и, получаемф(и) = log и' — log vо = log v' — 0,492502,что дает эмпирическую оценку константы.Далее, по мере приближения значения и к единице, мы видим, что и
растет пропорционально величине 2/(1 — и), и, следовательно, значе¬
ние log г/ приближается к равенству с величиной log 2 — 21og(l — и);
несмотря на некоторые различия конечных частот, частоты будут заданы
с помощью коэффициентов разложения:2 2—2 log(l — х) = 2х + — х2 + — х3 + ...,/ Отак что частота при р = г/2п приближается к величине 2/г по мере увели¬
чения г, согласно решению, найденному из дифференциального уравнения.Первые несколько реальных значений коэффициентов приведены
в таблице 4.
Вариации как результат мутаций и отбора91Таблица 4Предельные частоты для аллелей, обусловленных мутациями
Порядковый номерПриближенныекоэффициента
в разложении1
2345Точные Разность2,0000001,0000000,6666670,5000000,4000002,2409170,9537760,6718640,5010960,399762+0,240917-0,046224+0,005197+0,001096-0,000238Общее число факторов, закрепившихся в популяции в результате од¬
ной новой мутации в каждом поколении, будет равно сумме 2п первых
коэффициентов разложения ф(х), или2(7 + log 2п) + 0,200645.Для значений п от миллиона до миллиарда таблица 5 показывает число фак¬
торов (для каждого, возникающего за поколение), вносящих вклад в дис¬
персию вида.Таблица 5пЧисло фак;10630,410735,0ю839,6ю944,2ю1048,8ю1153,4ю1358,0На рис. 8 показано распределение для предельных частот. Было об¬
наружено, что значительная часть новых мутаций нужна, чтобы поддер¬
живать даже низкие частоты основных генов. Число этих основных генов,
однако, так велико, что единственная мутация в каждом поколении вызы¬
вает к жизни, как видно из таблицы 5, существенное их число, которое
практически пропорционально рангу значений z, возможных для популя¬
ции данного размера.
92Глава 42.0Рис. 8. Частоты, с которыми факторы представлены 1,2,3,... генами в общей по¬
пуляции в том случае, когда дисперсию создают новые мутации на постоянном
уровне. Для одной вновь возникшей мутации в каждом поколении частота для г
генов близка к величине 2/гЕсли частоте гена благоприятствует отбор, так что величина log p/q
в каждом поколении повышается на предположительно малую величину а,
тогда в одном поколении5р = apq5в — dy/pq — — a sin в.Эффект отбора, таким образом, будет создавать постоянный приток
величиной -i ay sin в, и наше дифференциальное уравнение принимает видду Id Id. лч 1 д2у= —2aaiiysme) + ^Miycote) + iHWДля статистического равновесия, создаваемого мутациями, теперь нужно,
чтобыду-j-г + у cot в — 2any sin в — А,Оичто можно записать какдв(у sin ве2ап cos в) = A sin ве,2an cos в
Вариации как результат мутаций и отбора93или иначе:ysm9e2ancose = J Asm6e2ancose + В;выполняя интегрирование, получаемп ( А I т)„—2ап cos I— cosect/ 1- Be\2 anВеличина притока составляет1 • a 1 1 dy A_едяп9__асо1в___ = _
она в а раза больше величины cosec в.В варианте, когда мутации возникают с частотой одна на каждое по¬
коление и каждая имеет некоторое селективное преимущество а, решение
должно быть равно 4 cosec в при 0 = 0, если же 0 = 7Г и мутации отсутству¬
ют, оно должно быть пропорционально sin в. Соответствующее выражение
имеет вид 4 COSeC в N _ g—2an(l+cos0) \1 - e-4an \ j>где частота в интервале dp составит величину:2 dp l-e-4an«pqПри q = 1 очевидно, что это дает предельное распределение, подобное
тому, которое задается мутациями, не имеющими селективного преимуще¬
ства, но когда q стремится к нулю, получаем выражение8 ап
1 _ е-4ап Р'соответствующее варианту исчезновения при отсутствии мутаций, со ско¬
ростью этого исчезновения 2а/(1 — е~4ап), что должно отражать вероят¬
ность конечного успеха для мутации, обладающей некоторым селективным
преимуществом а. Когда а = 0, значение вероятности стремится к предель¬
ному значению 1/2п, которое дает вероятность успеха мутации без селек¬
тивного преимущества и не увеличивается сколько-нибудь заметно до тех
пор, пока число 4ап мало; при среднем уровне значений 4ап (не мало
и не велико) нужно использовать полную формулу выражения, но когда Аап
94Глава 4становится большим, фактор экспоненты незначителен, и вероятность успе¬
ха становится очень близкой к 2а. Распределение частот в этом случае мож¬
но описать выражением:^ dp [л 0—4anq\pq(1 _в котором второй член значим только при малых значениях q, когда для каж¬
дого возможного значения р сохраняется постоянная частота, определяемая
как 8an dp или 4а, что соответствует вымиранию 2а в каждом поколении.Дифференциальное уравнение имеет силу для значений а, при кото¬
рых ап велико, однако при этом нужно, чтобы значения а2п были малыми.
Точное рассмотрение не настолько малых скоростей отбора, как те, о кото¬
рых здесь шла речь, очевидно, потребует использования значительно более
сложных формул, однако они, вероятно, не должны существенно отличать¬
ся от тех, что подходят применительно к очень малой селективности.Соответствующее решение для столь же важного случая с мутациями,
обладающими небольшой селективной вредностью, может быть найдено
путем изменения знака при а. Теперь вероятность успеха всегда будет ме-
нее 1/2п согласно формуле2 аg4an 2и во всех случаях очень мала. Распределение в диапазоне dp будет иметь
частоту2 dpe4anq - 1
pq e4an — 1 ’
что при больших значениях ап упрощается до^ dP —4апрpqдавая общее число факторов, близкое к 21og(l/2a), до тех пор, пока значе¬
ния Аап велики для каждого такого мутанта в каждом поколении.Таким образом, неблагоприятные мутации, в отличие от благоприят¬
ных или практически нейтральных (значение ап мало), не создают допол¬
нительных факторов, которые влияли бы на дисперсию у многочисленных
видов по сравнению с редкими видами. Конечно, расчеты относятся к ва¬
риациям, формируемым ограниченным числом мутаций, и не учитывают
того факта, что у многочисленных видов в каждом поколении имеется зна¬
чительно большее число индивидов, у которых могут возникать мутации.
Вариации как результат мутаций и отбора95Анализ показывает, насколько малой должна быть действующая на фак¬
тор интенсивность отбора, прежде чем мы можем оценить его нейтраль¬
ность или оценить с точки зрения вероятности спорадической мутации,
закрепляющейся у вида, или сможем проследить зависимость между ча¬
стотой мутаций и формирующейся дисперсией. В любом случае мы будем
иметь дело с произведением ап, в котором величина селективного преиму¬
щества умножена на число особей, участвующих в скрещивании в каждом
поколении. Для выживания малые отклонения этой величины от нуля име¬
ют существенное значение, например, шанс для выживания повышается
более чем в 50 раз, когда величина ап увеличивается от —1 до +1. Вклад
в дисперсию менее ощутим, поскольку он мало зависит от крайних частот
распределения и обусловлен в основном центральными частотами; в це¬
лом, нейтральные мутации формируют до половины дисперсии — пример¬
но столько же, сколько приходится на долю мутаций, имеющих существен¬
ное селективное преимущество. Если ап равно —2,5, этот вклад составляет
одну десятую, тогда как при величине +2,5 на него приходится уже 9/10
общего значения. Очевидно, что в популяции численностью в тысячу мил¬
лионов особей нейтральными можно считать только те генные мутации,
которые обладают преимуществами лишь в немногих группах из тысячи
миллионов. Распределение по благоприятным мутациям от этих малых пре¬
имуществ и до преимуществ в миллион раз больше должно быть очень
сходным, а вероятность снижения за равные интервалы dz при этом оста¬
ется примерно постоянной практически для всей амплитуды возможных
значений. По мере нарастания вреда неблагоприятные мутации ограничи¬
ваются все более и более низкими показателями отношения генных частот,
доходя до самых малых; сколько-нибудь существенным будет только вклад
мутаций, возникающих с заметной или высокой частотой.Хотя при одинаковом числе нейтральных или благоприятных мутаций
у многочисленных видов на дисперсию влияет большее число генов, чем
у видов редких, это зависит в основном от большего возможного диапазо¬
на значений z. Тогда адаптивные факторы будут иметь некие экстремаль¬
ные отношения генных частот и потому оказывать весьма малое влияние
на измеряемую дисперсию. Большое различие между обильными и редки¬
ми видами заключается в числе индивидов, которое в каждом поколении та¬
ких видов может выступать в качестве потенциальных мутантов. Реальное
число мутаций в каждом поколении должно быть пропорционально вели¬
чине популяции вида. Если частоты мутаций имеют заметный уровень, то
эти различия не будут иметь значения, так как устанавливается равновесие
с противоположно направленным отбором в той же пропорции. Важность
контраста связана с чрезвычайно редкими мутациями, когда число возни¬
96Глава 4кающих редких мутаций должно увеличиваться пропорционально числен¬
ности индивидов. Как было показано, именно в этом классе благоприятные
мутации оказываются весьма ограничены по числу, и преимущество более
обильных видов в этом плане особенно заметно.Интервал интенсивности отбора, для которого фактор можно считать
эффективно нейтральным, очень узок, и потому следует предполагать, что
такое состояние в общем должно быть очень преходящим. Медленные
изменения, которые всегда должны иметь место, изменяя генетическую
структуру и условия среды каждого вида, должны также изменять и се¬
лективную ценность каждого гена. Такие изменения селективной ценно¬
сти, несомненно, происходят очень медленно, и зона, отделяющая гены,
несущие некоторые селективные преимущества, от тех, что обладают некой
селективной вредностью, настолько узка (ее величина обратно пропорци¬
ональна величине размножающейся популяции), что переход ее должен
происходить очень быстро. Каждый удачный ген, который распространя¬
ется в популяции, должен в какой-то мере изменять селективную полез¬
ность или вредность многих других генов. Это, в свою очередь, должно
влиять на скорости увеличения или уменьшения частот этих других генов
так же, как и на скорость изменения его собственной селективной ценно¬
сти. Общее статистическое следствие этого состоит в том, что любой ген,
частота которого растет, независимо от тех причин, которые на это повли¬
яли (селективная ценность, повышенные частоты мутаций или что-либо
еще, как например, последовательность ряда благоприятных сезонов), бу¬
дет так влиять на генетическую структуру вида, чтобы ускорить повышение
его селективной ценности (если она растет) либо замедлить процесс, если
ценность снижается. Можно сформулировать иначе: каждый ген постоянно
стремится создать генетические условия, благоприятные для собственного
выживания,38 так что рост численности под влиянием любых причин будет,
в свою очередь, благоприятно влиять и на его селективные преимущества.По-видимому, здесь нужно отметить огромное разнообразие генети¬
ческой изменчивости, возникающей у вида, у которого расщепление идет
даже всего по 100 разным факторам. Общее число генотипов, участвую¬
щих в скрещивании, которое возникает при их комбинациях, составит 2100,
число, для изображения которого в десятичном виде потребовался бы ряд
из 31 цифры. Если же учитывать и гомозиготы, потребуется 48 цифр. По¬
пуляция численностью в тысячу миллионов особей, таким образом, мо¬
жет проявить лишь самую незначительную долю возможных комбинаций,
так что никакие два индивида не похожи друг на друга. Хотя возникаю¬
щие комбинации составляют только малую часть всех вариантов, которые
могли бы образоваться с равной вероятностью раньше или позже (напри¬
Вариации как результат мутаций и отбора97мер, в следующем поколении), может иметь место и огромное неизученное
число комбинаций, появление ни одной из которых нельзя ожидать, пока
система отношений генных частот не претерпела постоянных модифика¬
ций в правильном направлении. Более того, возможны миллионы разных
направлений для таких модификаций, так что без появления будущих му¬
таций все обычные виды должны уже иметь их потенциально среди самых
разных эволюционных модификаций. Часто и верно отмечают, что в отсут¬
ствии мутаций прогресс эволюции, независимо от его возможного направ¬
ления, неизбежно приведет к застою из-за недостатка будущих возможных
улучшений. Не так уж часто осознают, насколько далеко большинство су¬
ществующих видов должны отстоять от такого состояния стагнации; или
насколько легко, всего лишь на основе не более чем сотни факторов вид
может быть модифицирован до состояния, очень существенно выходящего
за пределы его современной вариации, и это касается очень широкого круга
разных признаков.
Глава 5Вариации как результат мутаций
и отбора (продолжение)Наблюдаемая связь между изменчивостью и обилием вида. Стабильные отно¬
шения генных частот. Равновесие по двум факторам. Простые метрические призна¬
ки. Меристические признаки. Биометрические эффекты недавнего отбора. Резюме.Наблюдаемая связь между изменчивостью и обилием видаВо второй главе Origin of Species Дарвин подводит итог изучению при¬
чин изменчивости, основанному на статистическом исследовании ряда от¬
четливо выраженных разновидностей у разных видов растений. От пуб¬
ликации своих собственных таблиц его, к сожалению, отговорили, но он
встретил у Хукера подтверждение своих основных выводов, что «виды,
которые занимают большую территорию, рассеяны по ней и встречают¬
ся наиболее часто, — самые изменчивые». Дарвин стремился показать, что
не просто наиболее распространенные формы дают начало местным разно¬
видностям в ответ на различные неорганические и органические условия
окружающей среды, а что к тому же: «В любой отдельно взятой местно¬
сти наиболее обычные виды, то есть виды с наибольшей численностью,
и виды, которые максимально рассеяны в пределах своей местности (а это
не означает просто широкое распространение и до некоторой степени от¬
лично от простой обычности), наиболее часто дают начало разновидностям,
которые оказываются столь заметны, что попадают на страницы ботаниче¬
ских описаний».Несколько лет назад я имел честь сделать статистический анализ об¬
ширных наблюдений г-на Е. Б. Форда, посвященных изменчивости окраски
крыльев у некоторых видов ночных бабочек. Для тридцати пяти видов от¬
тенки были достаточно сопоставимы, чтобы иметь возможность построить
единую цветовую шкалу, и наблюдения, включавшие 5 ООО особей, предо¬
ставили исключительную возможность для изучения связи между обилием
вида и изменчивостью. В подобном исследовании особенно важно исклю¬
чить любую тенденцию одной группы видов казаться более изменчивой,
Вариации как результат мутаций и отбора (продолжение) 99чем другие из-за особенностей, свойственных произвольно взятой шка¬
ле оттенков. Впрочем, объем данных был столь велик, что этот источник
ошибки удалось исключить, и после того, как были сделаны необходимые
поправки, оказалось, что дисперсия признака (для обоих полов) у десяти
видов, относящихся к разряду «многочисленных» и «самых обычных», пре¬
восходила дисперсию у менее распространенных видов на 70-80%, а две¬
надцать «обычных» видов занимали промежуточное положение.Поскольку на изменчивость неизбежно оказывают влияние многие
другие факторы, и в особенности, поскольку точность любой классифика¬
ции, основанной на обилии, всегда крайне мала, важно, чтобы для подоб¬
ных сравнений использовалось максимально возможное число видов. Ве¬
роятно, именно этой причиной главным образом и объясняется отсутствие
надежных данных об изменчивости видов. Впрочем, наблюдаемые у ноч¬
ных бабочек различия были достаточно выражены, чтобы считаться стати¬
стически значимыми даже в сравнении с большими различиями в изменчи¬
вости, наблюдаемыми в пределах каждого класса. Можно заметить, что эти
данные действительно указывают на то, что виды, имеющие наиболее ши¬
рокое географическое распространение более изменчивы, нежели виды, ме¬
нее распространенные, хотя нельзя сказать, что различия в этом сравнении
установлены статистически. Нет, однако, оснований считать, что увеличе¬
ние численности в результате расселения на более обширной территории
меньше влияет на увеличение изменчивости, чем то же увеличение числен¬
ности, обусловленное большей плотностью популяции, поскольку числен¬
ное соотношение между видами, отнесенными энтомологами к разрядам
многочисленных и редких, соответственно должно быть значительно боль¬
ше, чем соотношение между территориями, занятыми различными видами.Основываясь на доступных нам наблюдениях, можно сделать теорети¬
ческий вывод о том, что действительная численность вида является важ¬
ным фактором, обусловливающим проявляемую им дисперсию. Основное
следствие этого вывода для эволюционной теории, как было уже отмече¬
но Дарвином, заключается в том, что многочисленный вид будет ceteris
paribus достигать успехов быстрее на пути эволюционного развития и бу¬
дет стремиться к вытеснению групп с меньшей численностью, в конку¬
рентную борьбу с которыми он будет вступать. Мы можем заключить, что
в обычных условиях жизни на земле большое число видов с меньшей чис¬
ленностью будет уменьшаться, тогда как меньшее число более обильных
видов будет увеличиваться; при этом общее число видов будет сохранять¬
ся за счет дивергенции более обильных и особенно — более широко рас¬
пространенных видов, о чем пойдет речь в следующей главе. Можно, од¬
нако, отметить, что, несмотря на то, что рост приспособленности за счет
100Глава 5увеличения численности, как описано в главе 2, представляет интерес как
основа для достаточно быстрого дальнейшего улучшения, процесс расщеп¬
ления, являясь, несомненно, прямым адаптивным преимуществом, все же
приводит к очевидной потере в степени изменчивости, которую вновь об¬
разующиеся части способны поддерживать в отдельности. Таким образом,
непрерывное устранение более мелких видовых групп ставит естественный
барьер на пути чрезмерного дробления видов, которое является следствием
специализации, направленной на приобретение мельчайших различий.Среди факторов, влияющих на соотношение между вариациями и от¬
бором, можно упомянуть тенденцию подобных скрещиваться с подобными,
известную как гомогамия.40 Мы не располагаем данными о распространен¬
ности этого явления среди высших животных; единственный доступный
нам пример — это пример человека, но это едва ли может умалить цен¬
ность имеющихся у нас доказательств его существования среди людей как
из-за того, что это противоречит мнению широкой общественности, так
и из-за того, что никакие условия организации общества не могут объ¬
яснить его наличие. В коллекциях антропометрических данных сходство
между супругами является достаточно заметной чертой. Вероятно, главные
биометрические эффекты указанного явления заключаются в увеличении
генетической дисперсии, создаваемой определенным числом менделевских
факторов с определенным отношением генных частот, и, как следствие,
в том, чтобы увеличить в фиксированной пропорции интенсивность отбо¬
ра, которому каждый подвергается. Эффект отбора, направленного на из¬
менение роста человека, увеличивается, таким образом, более чем на 20 %.
Это значит, что гомогамия является важной действующей силой на пути
эволюционного изменения, однако причины этого явления неясны. Было
высказано предположение, что фертильность до некоторой степени зависит
от сходства конституции партнеров, но подобные доказательства относи¬
тельно человека и высших животных отсутствуют, и мне не известно ни
одной попытки доказать или опровергнуть эту гипотезу. Впрочем, возмож¬
но, что стандарты половых предпочтений слегка изменяются в зависимости
от размера индивида, и с этих позиций причины гомогамии едва ли отличи¬
мы от тех, которые, будучи связаны с половым предпочтением (см. главу 6),
имеют непосредственные селективные эффекты.Стабильные отношения генных частотДо сих пор мы рассматривали лишь те факторы, которые способству¬
ют генетической дисперсии приспособленности к выживанию и прогрессу.
Нам остается рассмотреть факторы, для которых один ген имеет селектив¬
Вариации как результат мутаций и отбора (продолжение) 101ное преимущество только до тех пор, пока поддерживается определенное
отношение генных частот, тогда как при его увеличении ген оказывается
в селективно невыгодном положении. В таких случаях отношение генных
частот будет сохранять стабильность при достижении предельного значе¬
ния, поскольку в это положение его будет возвращать действующий есте¬
ственный отбор, лишь только отношение генных частот отклонится в ту
или иную сторону. При этом значении эффект генного замещения на вы¬
живание будет равен нулю, и, соответственно, никакого вклада в генети¬
ческую дисперсию приспособленности наблюдаться не будет, хотя гене¬
тическая дисперсия по другим количественным признакам под действием
подобных факторов, вероятно, увеличится. Эти случаи имеют особое зна¬
чение, поскольку существует принцип, согласно которому факторы будут
встречаться тем чаще, чем меньше будет скорость изменения отношения
генных частот для каждого из них. По этой причине, если принять ста¬
бильность факторов за абсолютно постоянную величину, подобные случаи
должны бы были накапливаться у каждого вида с самого момента его воз¬
никновения; в действительности, однако, сами условия стабильности в те¬
чение эволюционных изменений не могут не меняться, и мы можем быть
уверены только в том, что подобная генная стабильность должна существо¬
вать с частотой, абсолютно несоразмерной с вероятностью существования
условий, на которых эта стабильность основана.Определенный фактор может находиться в положении стабильного
равновесия в условиях отбора, если гетерозигота имеет селективное пре¬
имущество перед обеими гомозиготами. Поскольку если предположить, что
три типа фактора проявляются в любом поколении в соотношении геноти¬
пов р2 : 2pq : q2, и что их реальные селективные преимущества находятся
соответственно в соотношении а : Ъ : с, тогда эти три генотипа в этом
поколении будут воспроизводиться в соотношении ар2 : 2bpq : cq2, где аб¬
солютные значения величин а, Ь, с не имеют значения, а важны только их
соотношения. Если равновесие в отношении генных частот установилось,
это соотношение будет постоянным у всех, воспроизводящихся подобно
предыдущему поколению, и, следовательно,р ар2 + bpqq bpq + cq2 ’откуда следует, чтоap + bq = bp + cq.Вычитая из обеих частей уравнения b(p + q), мы получаемр{Ь-а) = q(b — с),
102Глава 5илир _ b — с
q Ъ — аТаким образом, мы видим, что существует реальное соотношение равно¬
весного состояния, если Ъ — а и b — с либо оба положительны, либо оба
отрицательны, то есть если гетерозигота либо лучше, либо хуже приспо¬
соблена, чем обе гомозиготы. A priori ясно, что любое из этих условий
следует расценивать как исключительное; впрочем, в природе они встре¬
чаются неодинаково редко, поскольку когда Ь меньше а и с, равновесие
нестабильно, и подобные случаи не будут иметь тенденции к широкому
распространению, тогда как в случае, когда Ь больше а и с, равновесие ста¬
бильно, и, следовательно, подобные случаи будут сохраняться до тех пор,
пока стабильность не нарушится.Для того чтобы продемонстрировать условие стабильности, достаточ¬
но заметить, что отношениеар2 + bpq
bpq + cq2может быть представлено в виде отношенияр(ар + eg) + pq(b - с)
q{ap + cq) + pq(b - a)'которое лежит между отношениями р : q и (Ъ — с) : (Ь — а), если Ъ больше а
и с, за исключением случаев, когда Ъ — с и b — а отрицательны.У организмов, способных как к самооплодотворению, так и к пере¬
крестному оплодотворению, положение, при котором гетерозигота имеет
селективное преимущество, направлено на то, чтобы дать полученному
в результате перекрестного оплодотворения потомству большую репродук¬
тивную ценность, чем потомству, полученному в результате самооплодотво¬
рения, и поэтому заслуживающему несколько больших затрат; поскольку
в популяции со случайным скрещиванием, а мы ведем речь именно о та¬
ких популяциях, репродуктивные ценности трех генотипов будут находить¬
ся в простом соотношении а : Ь : с. Следовательно, для первого генотипа
средняя ценность его потомства будет составлять pa+qb, если оно получено
в результате случайного перекрестного скрещивания, тогда как для потом¬
ства, полученного в результате самооплодотворения, эта величина будет
равна а. Для гетерозиготы мы находим значение (pa + Ъ + qc) для случая
перекрестного оплодотворения против ^(а + 2Ь + с) для случая самоопло¬
дотворения. Средние преимущества двух гомозиготных генотипов, таким
Вариации как результат мутаций и отбора (продолжение) 103образом, равны q(b — а) и р(Ь — с) соответственно, в то время как для гете¬
розиготы эта величина равна -|(р — q)(a — с). Помня, что частоты, с кото¬
рыми встречаются эти три генотипа, находятся в отношении р2 : 2pq : q2,
мы находим, что средние потери репродуктивной ценности при самоопло¬
дотворении равны\pq{2p{b-a) + {р - q)(a - с) + 2q(b - с)} = pq(2b - а - с).Теперь в соответствии с нашим предыдущим решением равновесие
будет достигнуто, когдаЪ — с Ь — а2о — а — с 26 — а — си если мы собираемся представить а, b и с как величины пропорционально¬
го вклада в родословную будущих поколений, мы должны также записать,
чтор2а + 2 pqb + q2c = 1,
где, заменив р и q, мы придем к отношению62 — ас — 26 — а — с.Используя эти соотношения, мы можем выразить среднюю потерю репро¬
дуктивной ценности потомства, вызванную самооплодотворением, исполь¬
зуя только а, b и с:(6 — а) (Ь — с)2(62 — ас)Таким образом, если бы, например, для любого фактора а, b или с встре¬
чались бы в отношении 5:6:4, было бы достигнуто стабильное гене¬
тическое положение, при котором результаты самооплодотворения стоили
бы, относительно своих перспектив по внесению вклада в будущие поко¬
ления, всего лишь на у16 меньше, чем средние результаты перекрестного
оплодотворения. Любые другие случайно присутствующие факторы того
же типа, несомненно, добавили бы преимущество перекрестному оплодо¬
творению. Впрочем, представленная выше формула подходит для организ¬
мов, которые, как правило, размножаются перекрестным оплодотворением;
для организмов, которым более свойственно самооплодотворение, величи¬
на репродуктивной ценности для трех генотипов будет находиться в более
высокой пропорции, чем их селективные эффекты за одно поколение.
104Глава 5Равновесие по двум факторамДва фактора, альтернативные гены которых можно представить как А, а
и В, Ь, поддерживаются обоюдно в генетическом равновесии при условии,
что селективное преимущество А над а имеет противоположную направ¬
ленность, когда В замещается на Ъ или vice versa . Не делая попытки уточ¬
нить, какое именно селективное преимущество получает каждый из девяти
генотипов, мы можем определить характер рассматриваемого отбора, ска¬
зав, что ген А оказывается благоприятным в присутствии В, но оказывается
неблагоприятным в присутствии Ь, и что ген В оказывается благоприятным
в присутствии А, но неблагоприятным в присутствии а. Равным образом
мы, безусловно, можем поменять местами в этом высказывании слова бла¬
гоприятный и неблагоприятный.Равновесие в подобной системе, очевидно, предусматривает, что рост
частоты А, который имеет место в присутствии В, будет в точности сбалан¬
сирован его уменьшением в присутствии Ь, и что рост частоты В, который
имеет место в присутствии А, будет в точности сбалансирован его умень¬
шением в присутствии а. Но важно отметить, что равновесие частот га-
метных комбинаций АВ, АЪ, аВ, ab требует третьего условия равновесия.
В соответствии с условиями нашей задачи две из них, которые мы обозна¬
чим как АВ и ab, пользуются предпочтением естественного отбора, и ча¬
стота их в зиготах увеличивается, тогда как у противоположных пар умень¬
шается. Регуляция соотношения между частотами этих двух пар гаметных
типов должна происходить за счет рекомбинации у индивидов, гетерози¬
готных по обоим факторам. Из этих так называемых двойных гетерозигот
некоторые образуются в результате соединения гаметных типов АВ и ab,
и для них эффект рекомбинации состоит в уменьшении частот этих двух
типов. Этот эффект будет частично сбалансирован за счет рекомбинации в
гетерозиготах второго типа, возникших в результате слияния АЪ и аВ, и,
если общий эффект рекомбинации состоит в том, чтобы уменьшить часто¬
ты АВ и ab, очевидно, что двойные гетерозиготы, возникшие из гамет этих
типов, должны быть более многочисленны.Неравенство частот двух типов рассмотренных нами двойных гетеро¬
зигот имеет серьезное последствие, поскольку в любом случае, когда два
рассматриваемых нами фактора окажутся расположенными в одной и той
же хромосоме, частота рекомбинации будет определяться кроссинговером,
который, как известно, сильно зависит от генетических различий между
разными линиями. Более того, в более многочисленных типах двойных ге¬
терозигот рекомбинация приводит к замене более благоприятных генетиче-*Наоборот (лат.).
Вариации как результат мутаций и отбора (продолжение) 105ских комбинаций на менее благоприятные комбинации, и, соответствен¬
но, способствует уменьшению репродуктивной ценности, и это положе¬
ние не удается полностью сбалансировать ростом репродуктивной ценно¬
сти за счет рекомбинации в менее многочисленных двойных гетерозиготах.
Соответственно, присутствие двух факторов в одной и той же хромосоме,
селективное преимущество каждого из которых меняет селективное пре¬
имущество другого, будет всегда направлено на уменьшение рекомбинации
и тем самым на рост величины сцепления в хромосомах этого вида. Эта
тенденция направлена всегда в одну сторону, и хотя тип взаимодействия
факторов, из которого следует эта тенденция, может быть редким, все же
благодаря стабильности отношений генных частот, которые им индуциру¬
ются, мы можем предвидеть, что подобные случаи будут встречаться хотя
бы раз с частотой, не пропорциональной скорости их появления.Обнаружение силы, которая постоянно стремится увеличить интенсив¬
ность сцепления, естественно вызывает вопрос о существовании других
сил, действующих в противоположном направлении, под общим воздей¬
ствием которых, вместе с уже обсуждавшейся силой, интенсивность сцеп¬
ления могла бы быть приближена к своей наблюдаемой величине. Такой
силой оказывается постоянное распространение полезных мутаций в попу¬
ляциях, в которых они возникают. Поскольку если полезные мутации воз¬
никают так редко, что каждой хватает времени, чтобы стать доминантной,
пока появится следующая, они могут оказаться одновременно в одной га¬
мете только благодаря рекомбинации. Если две полезные мутации, которые
случайно оказались в гомологичных хромосомах, будут одновременно рас¬
пространяться в пределах одного вида, мы можем предвидеть время, когда
каждая гамета будет нести обе полезные мутации, и они будут получены
в линейной последовательности либо из гамет одного типа, либо, в конце
концов, из особей, у которых произошла рекомбинация. Подобные особи,
как можно догадаться, по этой причине будут несколько лучше представле¬
ны в будущих поколениях, чем остальные, у которых частота рекомбинаций
должна была быть в среднем ниже. Ясно, что для того, чтобы этот процесс
был действенной преградой на пути постоянной тенденции к увеличению
интенсивности сцепления, поток полезных мутаций должен быть доста¬
точно велик. Впрочем, кажется, не существует доказательств, опроверга¬
ющих мнение, что даже в каждой хромосоме большинства видов можно
в любой момент обнаружить множество полезных мутаций, каждая из ко¬
торых по мере увеличения частоты вносит свой вклад, пусть даже самый
ничтожный, в совершенство внутренней и внешней адаптации. Если необ¬
ходимость в объединении этих преимуществ является в реальности эффек¬
тивным барьером на пути роста интенсивности сцепления, может оказаться
106Глава 5возможным, по мере того как будут накапливаться новые данные, оценить
таким образом, пусть и приближенно, относительные скорости улучшения
у разных видов.Простые метрические признакиПризнаки, которые можно определить простым измерением, такие как
рост человека, длина отдельной кости или зуба и т. д., имеют особое значе¬
ние благодаря тому, что могут быть относительно просто изучены прямыми
биометрическими методами. Как было отмечено в главе 2, понятие адапта¬
ция не может применяться в полной мере к таким простым признакам,
особенно рассматриваемым в отдельности; считается, однако, что каждый
признак обладает оптимальным для существующего состояния организма
и его окружающей среды значением, что дает нам основания рассматривать
его как среднее для данного вида. То, что это должно быть так, с очевид¬
ностью следует из крайне высокой скорости, с которой подобные количе¬
ственные показатели изменяются тогда, когда на них направлено действие
отбора. Например, как показывают наблюдения за показателями средне¬
го роста потомства, полученного от родителей различного роста, никакого
жесткого отбора не потребовалось бы для того, чтобы увеличить или умень¬
шить рост населения на дюйм в каждом поколении, и даже при том, что
продолжительность жизни Человека велика, подобная скорость изменений
превзошла бы наибольшие из наблюдаемых расовых различий за короткий
исторический период.Если мы возьмем любой влияющий на подобное измерение фактор,
остальные эффекты которого в случае их наличия не оказывают заметного
влияния на выживание, то станет очевидно, что стабильность отношения
генных частот требует отдельного рассмотрения. На первый взгляд может
показаться, при условии отсутствия или неполного доминирования, в ре¬
зультате чего гетерозигота является промежуточной между двумя гомози¬
готами, что отбор, благоприятствующий промежуточным значениям, будет
направлен на предпочтение гетерозигот и, как следствие, будет в самом
общем виде способствовать условию рассматриваемой выше стабильно¬
сти. Если бы это было действительно так, то нам пришлось бы придти
к двум тревожным выводам: (1) что накапливая в условиях стабильности
большое число факторов с промежуточными гетерозиготами, мерные при¬
знаки должны демонстрировать в своих биометрических свойствах общее
отсутствие доминантности, тогда как, что уже было отмечено (стр. 18), все
множество доступных количественных признаков тела человека явно ука¬
зывает на обратное; (2) что действие отбора, благоприятствующего проме¬
Вариации как результат мутаций и отбора (продолжение) 107жуточным значениям, будет влиять, препятствуя вымиранию всех других
типов, на все большую дисперсию рассматриваемого признака и, как след¬
ствие, приводить к тому, что промежуточные значения будут становиться
все более редкими.Впрочем, признание того, что среднее для вида быстро достигает оп¬
тимальной величины, ставит совершенно новую задачу, нежели та, что бы¬
ла рассмотрена ранее, поскольку селективное преимущество гетерозиготы
зависит от среднего отклонения этого генотипа от оптимума, который бу¬
дет изменяться вслед за изменением отношения генных частот. Если i,j,k
являются отклонениями средних значений этих генотипов от среднего, эф¬
фект отбора будет эквивалентен устранению малых долей каждого гено¬
типа, пропорциональных г2, J2 и к2. Если три генотипа составляют про¬
порцию р2 : 2pq : q2, отношения, на которые два альтернативных гена
сократятся, будут пропорциональныpi2 + qj2 и pj2 + qk2.И отношение генных частот будет находиться в равновесии только при усло¬
вии, что обе эти величины будут равны. Далее, из определения среднего мы
имеемp2i + 2 pqj + q2k = 0.Если мы используем последнее уравнение, чтобы избавиться от откло¬
нений г, j, к, заменив их единым отношением, определяемым кака г — j1 = Т^к'которое зависит только от степени, в которой в исследуемом факторе прояв¬
ляется доминантность, мы найдем, что это соотношение связано с соотно¬
шением р: q при достижении условия равновесия следующим уравнением:р( 1 - 2р2)а2 + 2pq(q - р)а/3 - q( 1 - 2q2)(32 = 0.Это выражение симметрично, если заменить р и q, а и /3, и левая часть
пропорциональна скорости уменьшения г (= log р — log q).Из знаков трех членов следует, что единственное положительное реше¬
ние существует, если q больше р, но при условии, что q не превышает л/2;
более того, поскольку выражение положительно при /3 = 0, тогда если а =
= /3, оно упрощается до(q~p)02,
108Глава 5которое также положительно, из этого следует, что если q больше р, то¬
гда при достижении равновесия /3 должно быть больше а. Частота более
доминантного гена должна быть меньше.Если бы любое равновесие было стабильным, тогда (q — р)02 должно
было бы увеличиваться вслед за увеличением р, и его производная относи¬
тельно р должна равняться(1 - 6р2)а2 + (1 — 6pq)2af3 + (1 - 6q2)/32,или(а + /?)2 - 6(ра + qP)2.Теперь а+Р является произвольной величиной и может быть принята,
как и р + q, равной единице, тогда, поскольку2(pa + q/3) = (р + д)(а +/3) + (р - q){a - /3),из этого следует, что ра + q/З не может быть меньше У2, так как мы показа¬
ли, что когда q больше р, тогда а больше /?. Соответственно, производная
в любом положении равновесия меньшеи всегда отрицательна. Условия равновесия всегда нестабильны. Любой ген,
частота которого будет меньше его равновесной частоты, будет уменьшать
свое присутствие в результате отбора.Следовательно, все мутации, влияющие на подобный признак, если
только они не обладают компенсационными преимуществами в других от¬
ношениях, будут изначально неблагоприятны, и мы можем предсказать, что
каждая достигнет равновесия, при котором частота мутаций будет баланси¬
роваться противоположно направленным отбором, действию которого она
подвержена. Это напоминает ситуацию с внутренне вредными мутациями,
рассмотренную в главе 3, но отличается от нее тем, что при условии доста¬
точно высоких частот мутаций мутантный ген в данном случае проходит
точку максимального противодействия и, достигнув достаточной частоты,
в дальнейшем, по сути, поддерживается отбором. Требуемые для осуществ¬
ления этого частоты мутаций зависят от (1) величины эффекта, произво¬
димого фактором, т. е. метрического различия между двумя гомозиготами,
равного а, (2) общей дисперсии вида по изучаемому количественному при¬
знаку, равной а2, (3) интенсивности отбора в пользу оптимального показа¬
теля; эта величина может быть измерена как 1 /т2, где т2 является величи¬
ной той же размерности, что и а2, которая стремится к нулю при крайне
Вариации как результат мутаций и отбора (продолжение) 109интенсивном отборе и была бы невероятно большой, если бы все значения
количественного признака были бы равно удовлетворительными.В отсутствие доминантности максимальное противодействие наблюда¬
ется, когда р = У4, и это положение преодолевается, если частота мутаций к
превышает64(сг2 + т2)'Исчерпывающее обсуждение данной ситуации потребовало бы изучения
действия отбора на дисперсию, поскольку если отбор интенсивен и г2 ма¬
ло, вероятно, что и а2 станет также мало; на данный момент мы можем
отметить, что вклад изучаемого фактора в общую дисперсию равен не бо¬
лее а2/8. Если, таким образом, действие фактора так мало, что его вклад
будет не больше чем 1 на 100000 к общей дисперсии, частота мутаций
порядка одной на миллион вполне могла бы повлиять на его постепенное
установление. Такой мота бы быть ситуация с факторами, изменяющими
рост человека приблизительно на одну сороковую дюйма. Факторы, оказы¬
вающие меньший эффект, могли бы установить мутантную форму при бо¬
лее низких частотах мутации, в каждом случае тем легче, чем более неопре¬
деленным является предпочтительное выживание средних размеров.Мутантные гены с большим эффектом или более низкими частотами
мутаций останутся в бездействии при всех значениях ниже р = 0,25. Теперь
при этом значении среднее окажется между средними значениями гетеро¬
зигот и немутантных гомозигот; соответственно, при этом значении и ниже
его гетерозигота с большим преимуществом могла бы почти точно рав¬
няться немутантной гомозиготе. Таким образом, всегда будет существовать
тенденция, аналогичная описанной в главе 3, к тому, чтобы немутантный
ген стал доминантным посредством модификации гетерозиготы в сторону
большего с ним сходства. Развитие доминантности должно, таким образом,
идти против мутаций, в каком бы направлении они ни возникали; таким об¬
разом, можно ожидать, что обнаруженные по подобным признакам мутант¬
ные гены, увеличивающие или уменьшающие количественный признак, бу¬
дут в любом случае рецессивными. Факт, что потомство от межрасового
скрещивания, демонстрирующего различия в метрических признаках, все¬
гда занимает промежуточное положение, можно было бы ожидать из этой
билатеральной тенденции к развитию доминантности.Развитие доминантности неизбежно меняет условия равновесия; на¬
пример, для полной доминантности максимальный противоположно на¬
правленный отбор достигается при р = 0,5, и частота мутаций, необхо¬
димая для становления мутантного гена, в четыре раза больше, чем зна¬
110Глава 5чение, найденное для генов без доминантности. Условием, необходимым
для увеличения доминантности, является положение гетерозиготы по про¬
тивоположную сторону от среднего по отношению к немутантной гомози¬
готе. Для любой конкретной степени доминантности, то есть для любого
конкретного значения а, доминантность будет расти до тех пор, покаa q2Р < Р2'Следовательно, в течение некоторого времени доминантность будет уве¬
личиваться, а значение /3 уменьшаться до тех пор, пока эти два значения
не станут равными. Одновременную вариацию двух отношений р : qua : (3
можно показать более наглядно при помощи диаграммы (рис. 9), где кри¬
вая АХ ZВ представляет ряд возможных условий, при которых дальнейшая
тенденция к изменению степени доминирования отсутствует, и по длине
которой располагаются точки, в которых факторы могут поддерживаться
благодаря соответствующим частотам мутаций. На той же диаграмме ли¬
ния EAYC проведена через точки, в которых достигается максимальный
противоположно направленный отбор для различных значений а. Мутация
без доминирования стартует от О, и по мере того, как ее концентрация
увеличивается, движется вдоль линии ОЕ до достижения некоторой точ¬
ки на этой линии, в которой частота мутаций балансируется противоотбо-
ром. На этой стадии и, в действительности, в течение всего ее развития
до этой стадии, отбор будет направлен на то, чтобы перевести мутантный
ген в рецессивное состояние, и соответствующая точка пройдет вдоль ли¬
нии ниже EZ и остановится на ограничивающей линии ZB, во всех точках
которой, как следует из диаграммы, а превышает 0,95. На любой стадии
изменение частоты мутаций сдвинет соответствующую точку на диаграм¬
ме вверх или вниз согласно увеличению или уменьшению частоты мута¬
ций, но ясно, что фактор, оставленный без изменения в области АЕОВ,
будет стремиться к соответствующему уровню на линии ХВ. Таким же
образом любой фактор, перемещенный в некую точку в области XDB, бу¬
дет испытывать действие отбора в направлении уменьшения доминантно¬
сти, поскольку в этом случае смещение гетерозиготы относительно средней
для вида величины будет происходить в том же направлении, что и сме¬
щение немутантной гомозиготы, и отбор будет благоприятствовать любой
тенденции к уменьшению этого отклонения. Такой путь представлен лини¬
ей GZ. Из положения точки X можно видеть, что этот процесс не может
уменьшить значение а ниже приблизительно 0,79 до тех пор, пока не будет
пересечена критическая линия АС. Факторы, находящиеся в равновесии,
будут, таким образом, стремиться сохранить по меньшей мере эту степень
Вариации как результат мутаций и отбора (продолжение) 111доминирования, даже если частота их мутаций почти достаточна для то¬
го, чтобы преодолеть все противодействие. Наконец, фактор, помещенный
в область CYXD, будет при неизменных условиях уменьшать свою до¬
минантность до тех пор, пока не достигнет линии АС, на которой эффект
противоотбора будет максимальным, а выше которой он будет не усили¬
ваться, а ослабевать, когда частота мутантного гена будет увеличиваться.
Ход движения подобных факторов является, с некоторыми отклонениями,
сходным с ходом движения тех из них, которые, объединяя преимущества
высокой частоты мутаций с крайне малым измеримым эффектом, нико¬
гда не встречают на своем пути достаточно серьезного противодействия,
которое могло бы помешать их прогрессу. Наименьшая частота мутаций,
которая могла бы поддерживать фактор на этом пути, почти в два раза
больше, чем та, что первоначально требуется для фактора в отсутствие
доминантности, чтобы пройти в Е; ее представляет линия DXY, отде¬
ляющая факторы, которые пройдут АС, от тех, которые остановятся на
линии XZB.Мы находимся в неведении относительно частот, с которыми разные
мутации смогут достичь различных, показанных на диаграмме стадий. Хотя
большая часть представленной области могла бы оказаться занятой мута¬
циями, которым чуть-чуть не удалось пройти ее беспрепятственно. Можно
ожидать, что мутации со сравнительно большим эффектом и, возможно,
мутации с чуть меньшим эффектом, обладающие низкими частотами, оста¬
новятся при низких значениях р и будут стремиться к точкам, расположен¬
ным около В на линии ZB, где можно видеть, что доминантность, пусть
и не совсем полная, достигает весьма заметного развития.Вторая особенность метрических факторов, которая, возможно, имеет
больше последствий, состоит в том, что из двух любых имеющих сход¬
ный эффект генов, то есть таких генов, совместный эффект которых при¬
водит к увеличению или уменьшению количественного признака, каждый
будет более полезным или менее вредным в отсутствие другого. Предыду¬
щий анализ показал, что это взаимодействие не ведет к стабильному отно¬
шению генных частот по сравнению со взаимодействием, рассмотренным
на стр. 104; тем не менее это взаимодействие должно иметь аналогичный
эффект благоприятствования генотипам, демонстрирующим тесное сцепле¬
ние. Комбинации гамет, где различные эффекты факторов хорошо сбалан¬
сированы и направлены на реализацию средней длины, должны пользовать¬
ся постоянным предпочтением при отборе за счет плохо сбалансированных
комбинаций, способствующих появлению очень больших или очень малых
значений длины. Это значит, что потребуется постоянное возобновление
числа последних за счет рекомбинации, в результате чего кроссинговер
112Глава 5Рис. 9. Соотношения между частотой гена, частотой мутаций и степенью доминиро¬
вания для фактора, детерминирующего простой метрический признак. Подробное
объяснение — в текстедолжен будет, если речь идет об этих факторах, стремиться к снижению
средней репродуктивной ценности потомства.Сейчас мы поймем, каким образом можно представить себе, как сред¬
нее значение количественных признаков должно изменяться под действием
отбора всякий раз, когда подобное изменение оказывается полезным. Если
оптимальное значение увеличивается, все гены, эффект которых в противо¬
положность их существующим аллеломорфам состоит в увеличении при¬
знака, будут немедленно или по крайней мере быстро увеличивать свою
частоту. Что до тех факторов, чье влияние на выживание может быть пол¬
ностью выражено через их влияние на изучаемый признак, эффекты по¬
Вариации как результат мутаций и отбора (продолжение) 113добного изменения частот в одних случаях будут постоянны, а в других —
временны. Некоторые гены, которые ранее испытывали противодействие
отбора, сдвинутся к частотам, при которых отбор начнет им благоприят¬
ствовать, и их число может вырасти за то время, пока будет продолжаться
отбор, столь значительно, что когда он прекратится, они все еще смогут
пользоваться его предпочтением. Их последующий прогресс, таким обра¬
зом, будет направлен на увеличение значения признака даже после того, как
он достигнет нового оптимума. То же относится и к мутантным генам, ча¬
стота которых увеличивается после прохождения максимума противоотбора
в область, где отбор, хотя и противостоит образованию мутаций, недостато¬
чен для остановки их роста. В других случаях увеличение или уменьшение
частот под действием временного отбора будут также только временны¬
ми, и по достижении нового оптимума частоты будут стремиться к воз¬
вращению в прежнее положение, тем самым невольно способствуя тому,
чтобы заставить видовое среднее несколько отклониться от нового опти¬
мума к старому положению. Эта система напоминает ту, в которой сила
растяжения способна производить как упругое, так и постоянное напряже¬
ние, и в которой постоянная деформация всегда направлена на ослабление
действующих упругих сил.Меристические признакиВ поисках свидетельств непрерывной вариации Бэтсон уделил значи¬
тельное внимание вариациям числа сходных частей тела, встречающихся,
как в случае позвонков, сериями. Не считая аномалий развития, вариации
таких признаков должны быть дискретными, и значение, демонстрируе¬
мое каким-либо одним индивидом, неизбежно должно принадлежать ряду
целых чисел. Нет, однако, причин, полагать, что полученная таким путем
дискретность каким-либо образом связана с дискретностью генетических
частиц в менделевской наследственности. Напротив, будет разумным пред¬
положить a priori, что действительно наблюдаемое число является всего
лишь соматическим выражением до ближайшего целого числа переменной,
лежащей в основе физиологического признака, находящегося под влияни¬
ем, как обычный количественный признак, окружающей среды и генетиче¬
ских причин. Правильность этого взгляда для ряда важных случаев в на¬
стоящее время считается установленным фактом.Серии частот, с которыми встречаются разные числа позвонков,
внешне поразительно похожи на серии, полученные в том случае, когда
нормально распределенная случайная величина группируется в равные,
но достаточно большие интервалы своих значений, как если бы, напри¬
114Глава 5мер, рост человека был записан у большого числа индивидов с точностью
до ближайшего, кратного трем дюймам. Впечатление сходства возрастает,
если вместо наблюдения за отдельными индивидами мы рассмотрим одно¬
временно распределение ряда пар родителей и их потомков, в этом случае
среднее значение величины меристического признака в группе потомков,
произошедших от родителей с тем же значением, увеличивается количе¬
ственно от группы к группе точно таким же образом, как и в случае обыч¬
ного мерного признака.Подлинное доказательство того, что каждый индивид с определенной
вариацией меристического признака характеризуется известным генотипи¬
ческим или генетическим значением, которое отличается от соответству¬
ющих значений других индивидов на величины, не являющиеся целыми
числами, но долями, сильно отличающимися от целых чисел, может быть
получено, как показал Шмидт, двумя экспериментальными методами. У ор¬
ганизмов, способных к самооплодотворению или вегетативному размноже¬
нию, можно получить чистые линии генотипически идентичных индиви¬
дов, что позволяет провести прямое сравнение средних величин для боль¬
шого числа индивидов двух или более различных линий, развивающих¬
ся в одинаковых условиях. С другой стороны, если организмы способны
производить достаточное число потомков в одном скрещивании, каждого
из нескольких самцов можно скрестить с несколькими самками. Сравнивая
средние этих потомков, Шмидт установил скрытые за ними различия меж¬
ду генетическими значениями родителей того же пола с точностью, доста¬
точной для того, чтобы показать, что эти различия не могли быть целыми
числами.Если рассматривать такой признак, как число позвонков, в качестве
соматического выражения скрытой за ним генотипической изменчивости,
имеющей непрерывный характер, поразительное постоянство таких мери-
стических признаков в больших группах родственных организмов, без со¬
мнения, требует особого объяснения. Такое постоянство не становится ме¬
нее удивительным из-за наличия исключений. Среди огромного разнооб¬
разия форм, свойственных млекопитающим, почти все постоянно имеют
семь шейных позвонков, хотя два вида ленивцев имеют шесть и девять
соответственно. Сходная ситуация, наблюдаемая у рыб, где несколько се¬
мейств с различным числом позвонков возникли, очевидно, в разное время
из группы семейств, где наличие 24 позвонков было, по-видимому, неиз¬
менным свойством, воспринималась Тэйтом Рейганом как столь серьезная
проблема для теории естественного отбора, что в качестве альтернативного
объяснения он был готов обратиться к возможному влиянию изменившихся
условий на возникновение мутаций. Какая перспектива имеется для подоб¬
Вариации как результат мутаций и отбора (продолжение) 115ной силы, если ее существование может быть показано, помогая нам понять
эту конкретную проблему, кажется в настоящее время неясным. Тем более
важным представляется исследовать, действительно ли известные причины
оказываются столь неэффективны, как об этом думали, в осуществлении
наблюдаемых эффектов.В отношении групп, где все или почти все индивиды имеют одинако¬
вое число позвонков, возможны две точки зрения: (1) что генетической из¬
менчивости нет, и (2) что ни генетическая изменчивость, ни изменчивость
условий окружающей среды, в которых организм развивается, не достаточ¬
ны для возникновения частых отклонений от центрального целого числа.
Первую точку зрения можно не рассматривать не только потому, что раз¬
ные виды все же, несомненно, отличаются по числу позвонков, но также
потому, что в свете результатов последней главы мутантный ген, влияю¬
щий на скрытую за ним переменную, если у него нет других эффектов,
будет исключен из отбора по меньшей мере до тех пор, пока число позвон¬
ков будет сохраняться неизменным. Соответственно, любые мутации этого
типа, произошедшие в прошлом, должны у такого вида накапливаться.В соответствии со второй точкой зрения наследуемая индивидуальная
изменчивость существует относительно тенденции производить заданное
число позвонков, и вид, таким образом, потенциально пластичен в этом от¬
ношении. Если предположить, что среднее этого распределения совпадает
с модальным целым, отличная от этого целого частота может быть лег¬
ко вычислена из стандартного отклонения распределения; например, если
стандартное отклонение равно У6 единицы, на тысячу индивидов долж¬
ны ожидаться приблизительно три исключения, для % единицы — только
шестьдесят три на миллион, для l/w единицы — только одно на два миллио¬
на и так далее. Это означает, что потребовались бы обширные вычисления,
чтобы найти величину, исключающую вариацию числа позвонков и в то же
время достаточную, чтобы допустить эволюционное изменение числа по¬
звонков, если это когда-либо станет полезным.Эволюционный факт, требующий с этой точки зрения специального
объяснения, состоит в том, что в большой группе организмов с широким
разнообразием адаптаций столь редко оказывалось полезным производить
такое изменение. В этом отношении меристический признак представля¬
ет поразительный контраст с метрическим. Целые числа обладают осо¬
бым типом стабильности, благоприятствующей консервативным тенденци¬
ям в эволюции, что не встречается у мерных признаков. Возможно, объ¬
яснение этой тенденции лежит в простом факте, что включение или про¬
пуск одного члена из серии структур требует соответствующего измене¬
ния некоторого числа связанных с ней органов, таких как прикрепленные
116Глава 5мышцы, нервы, кровеносные сосуды и т.д., и что даже в тех случаях, ко
гда путем полной реорганизации на основе увеличения числа позвонком
можно добиться незначительного преимущества, вполне может быть, что
такое преимущество меньше, чем недостаток, который придется претер¬
петь из-за дезорганизации связанных структур любому индивиду, которому
выпадет на долю стать обладателем большего их числа. Даже если бы свя
занные структуры были морфологически завершены, нет уверенности, что
столь же совершенно окажутся скоординированы все количественные фи¬
зиологические приспособления. Этот аргумент является частным случаем
более общего, состоящего в том, что в любом высоко приспособленном ор
ганизме вероятность преимущества благодаря любому значительному эво¬
люционному шагу (сальтации) быстро становится бесконечно малой по ме¬
ре того, как увеличивается величина этого шага.Минимальная подверженность подобным несоответствиям должна,
без сомнения, встречаться у видов, демонстрирующих значительную из¬
менчивость; например, у угря, процессы развития которого, как можно ожи¬
дать, будут происходить практически без осложнений, независимо от того,
какое количество позвонков было заложено. Она должна быть максималь¬
ной там, где меристический признак наиболее стабилен и где способность
ответственных за развитие механизмов, имеющих дело с другими числами,
не подвергалась отбору в предыдущих поколениях. Впрочем, в этих случа¬
ях редкость исключений мешает возможности продемонстрировать любые
ассоциированные признаки ненормального развития. Промежуточный слу¬
чай, для которого имеется некоторое количество данных, это случай с сель¬
дью. Форд и Булл изучали случаи слитных и ненормально развитых по¬
звонков у этой рыбы. Из почти 7 ООО изученных скелетов 95 были с ненор¬
мальными или слитными позвонками. Если каждый элемент в этих струк¬
турах считать за целый позвонок, оказывается, что среднее число позвон¬
ков в ненормальных скелетах равно 55,82, что превышает среднее значе¬
ние для нормальных скелетов всего на 0,03. Хотя средние величины, таким
образом, очень близки, изменчивость ненормальных скелетов значительно
больше, чем нормальных. Другими словами, крайние числа позвонков, рав¬
ные 53 и 58, демонстрируют наибольший процент ненормальных скелетов,
средние числа позвонков, равные 55 и 56, которые имеют почти 90 про¬
центов рыб, показывают наименьший процент, тогда как промежуточные
числа 54 и 57 показывают промежуточные проценты ненормальности. Зна¬
чения процентов, полученные на основании данных Форда и Булла, следу¬
ющие (табл. 6).Можно видеть, что рыбы с меньшим числом позвонков демонстриру¬
ют весьма заметную склонность к развитию ненормальных структур. Если
Вариации как результат мутаций и отбора (продолжение) 117Таблица 6Число позвонков
53 54 55 56 57 58
45,5 4,1 1,2 1,1 2,6 10,0
0,08 1,06 28,36 61,30 8,91 0,29подобная тенденция является общей для случая необычных меристических
вариаций, что с общих позиций кажется вполне вероятным, то любая ме-
ристическая вариация по отношению к существующим стандартам должна
сталкиваться с заметным противоотбором и практически не иметь возмож¬
ности закрепиться, если только большее число не влечет за собой преиму¬
ществ, которые невозможно получить никаким иным путем.Если мы правы, относя консервативную тенденцию, которая, согласно
наблюдениям, свойственна меристическим признакам в их подверженности
к дезорганизации, следующей за внезапным изменением, то тогда эта тен¬
денция должна быть по меньшей мере в частичном бездействии в периоды,
когда ассоциированные структуры или их взаимоотношения с позвонками
находятся по тем или иным причинам в состоянии реорганизации. Подоб¬
ное выраженное изменение структуры и движения тела, как должно бы
требоваться при приспособлении тела камбалы или мускулатуры шеи нахо¬
дящегося в необычной позе ленивца, могло бы, таким образом, обеспечить
временную возможность для формы изменчивости, которая всегда суще¬
ствует, хотя и постоянно сдерживается, производить в виде исключения
эволюционную модификацию.Биометрические эффекты недавнего отбораНепосредственный эффект отбора в любом направлении заключается
в изменении среднего значения метрического или меристического призна¬
ка в зависимости от конкретного случая. Такой эффект можно обнаружить,
если установить последовательные средние значения, как в случае палеон¬
тологического материала. Дальнейший эффект можно предсказать по незна¬
чительному уменьшению дисперсии вследствие устранения факторов, ко¬
торые влияли на его возникновение раньше; этот эффект будет крайне мал
и в любом случае не сможет указать на то, отдавалось ли в этом случае
предпочтение большему или меньшему размеру. Впрочем, функции третьей
степени от значений измерений все же должны содержать следы недавнего
отбора, отличаясь друг от друга в зависимости от направления, в которомПроцент ненормаль¬
ных позвонков
Процент особей
118Глава 5производился отбор. Дело в том, что отбор в пользу увеличения размера
лишь на время приведет к исчезновению рецессивных генов, вызывающих
карликовость, и в течение процесса такого отбора они сохранятся у дан¬
ного вида. С другой стороны, рецессивный ген, эффект которого состоит
в увеличении метрического признака, получит в ходе этого процесса воз¬
можность распространиться по всему виду путем потери своего аллеломор¬
фа. Соответственно, если действие такого отбора продлится в дальнейшем,
рецессивные гены должны будут способствовать скорее уменьшению, чем
увеличению изучаемого признака. Равным образом, как и в случае выведе¬
ния пород комнатных собак, когда отбор направлен в сторону малого роста,
мы должны ожидать обнаружение избытка рецессивных генов, направлен¬
ных на увеличение средних размеров.Биометрическое выявление и измерение любого преобладания доми¬
нантности в одном направлении над другими требует, без сомнения, нали¬
чия обширного материала, хотя необходимые вычисления оказываются до¬
статочно простыми. Простейший метод, а именно измерение асимметрии
распределения частот через третий момент, содержит некоторые сложно¬
сти объяснения из-за эффектов средовых вариаций; по этой причине он мо¬
жет быть непригодным для изучения асимметрии больших совокупностей,
полученных от разных пар родителей. Наиболее прямое исследование, ко¬
торое было бы возможно, состоит в совокупном рассмотрении показателей
отца, матери и потомства и выражении регрессии размера потомков на раз¬
мер обоих родителей каках + Ьу — сху,где ж и у — признаки родителей, а а, Ь, с — коэффициенты, которые следует
определить по исходным данным. Тогда отбор в сторону увеличения ро¬
ста будет давать положительные значения с или, в общем, увеличивать его
значение алгебраически, тогда как отбор в противоположном направлении
будет направлен в сторону его уменьшения и будет давать отрицательные
значения.РезюмеВ двух предыдущих главах была предпринята попытка проверить пра¬
вильность генетической теории Естественного Отбора путем детального
рассмотрения метода, посредством которого, если следовать этой теории,
новые гены возникают, распространяются либо отбраковываются, или —
в более частном случае — новые гены, или менделевские факторы, временно
либо постоянно закрепляют свои позиции, способствуя органической измен¬
чивости, от которой, как было показано, зависит скорость прогресса вида.
Вариации как результат мутаций и отбора (продолжение) 119Изучение важной проблемы выживания единичных генов значительно
упрощается тем обстоятельством, что коль скоро они редки, все осталь¬
ные эффекты не важны по сравнению с элементом случайности в выжива¬
нии и размножении их носителей. Во многих важных случаях выживание
в течение одного поколения может быть представлено функцией, просто
связанной с распределением Пуассона, и во многих других случаях подста¬
новка той же формулы для поколения соответствующего биологического
цикла обеспечивает отличное приближение.Пусть и подверженные в момент своего возникновения стихии слу¬
чайного выживания полезные мутации имеют конечную вероятность, непо¬
средственно зависящую от их полезности, закрепиться постоянно в наслед¬
ственности вида. Это означает, что им достаточно было бы возникнуть все¬
го лишь несколько раз, прежде чем это событие оказалось бы практически
очевидным. Частота мутаций должна, следовательно, быть в этот испы¬
тательный период, как правило, крайне мала, и часто должно случаться
так, что практически все отдельные гены, распространяемые впоследствии
на весь вид, были бы получены от одного такого мутанта.Исследование статистического равновесия между новыми мутациями
и причинами исчезновения генов показывает, что полезные мутации на всех
уровнях полезности напоминают нейтральные мутации в распределении
отношения генных частот, и их частота равна во всех равных интервалах
переменной z, в которой удобно измерять вариацию отношения генных ча¬
стот. Область изменения z является более широкой у обильных, нежели
у малочисленных видов, и по этой причине отношение числа генетиче¬
ских различий, поддерживаемых в многочисленном виде, к частоте встре¬
чаемости новых мутаций больше того же числа для малочисленного ви¬
да. Область селективного преимущества, которая может рассматриваться
как эффективно нейтральная, является, однако, ничтожно малой и обрат¬
но пропорциональна численности вида. Поскольку вызывает сомнение, что
такое совершенное равновесие селективного преимущества могло бы под¬
держиваться в ходе эволюционных изменений, случайное выживание — хо¬
тя и главный момент в случае выживания отдельных генов, — становится
объектом чисто академического интереса в случае поддерживаемой у вида
дисперсии, главным образом связанной с проявлением полезных мутаций.
В частности, показано, что скорость угасания дисперсии из-за случайно¬
го вымирания при полном отсутствии мутаций крайне мала и практически
не имеет значения.Разработанная здесь теория содержит серьезные теоретические осно¬
вания для вывода о том, что более многочисленный вид должен ceteris
paribus стремиться к большей изменчивости, хотя скорость, с которой
120Глава 5должна расти изменчивость в ответ на увеличение численности, может
быть установлена, как нам кажется, только при непосредственном наблю¬
дении. Взаимосвязь между обилием вида и изменчивостью подтверждена
изящными наблюдениями за ночными бабочками, но требует более значи¬
тельных исследований в других группах. Важным следствием для теории
эволюции является то, что, как правило, меньшее число крупных видов
должно увеличивать свою численность за счет большего числа мелких ви¬
дов, и постоянное вымирание последних должно ставить естественный ба¬
рьер на пути чрезмерного дробления видов, неизбежно сопровождающего
слишком тонкую и детальную специализацию.Важным для нашей теории является также наличие, пусть и в силу
исключительных обстоятельств, отношений генных частот, сохраняющих
стабильность под действием отбора, поскольку такого рода факторы не мо¬
гут подвергаться элиминации до тех пор, пока в процессе эволюционного
изменения не нарушается стабильность. В простейшем случае это проис¬
ходит, когда отбор отдает предпочтение гетерозиготе перед обеими гомози¬
готами. Показано, что в этом случае равновесие всегда существует и всегда
стабильно, тогда как в обратном случае равновесие нестабильно, и менее
многочисленный ген постепенно элиминируется вплоть до полного своего
исчезновения. Накопление факторов, для которых предпочтение отдано ге¬
терозиготе, обеспечивает постоянное среднее преимущество перекрестного
оплодотворения над самооплодотворением в том смысле, что потомство,
полученное в результате перекрестного оплодотворения, имеет более вы¬
сокую репродуктивную ценность и больше заслуживает того, чтобы его
произвели на свет, чем потомство, полученное в результате самооплодотво¬
рения. Развитие раздельных полов у подвижных животных и возникнове¬
ние множества приспособлений для обеспечения перекрестного оплодотво¬
рения у растений, хотя и не само происхождение полового размножения,
можно объяснить небольшим индивидуальным преимуществом благодаря
предпочтению гетерозигот, по крайней мере если мы учтем постоянную
тенденцию гетерозигот напоминать более успешную гомозиготную форму,
рассмотренную в главе 3.Другие типы стабильного равновесия могут достигаться путем взаи¬
модействия двух или более факторов. Значение этой группы случаев за¬
ключается в их постоянной тенденции, коль скоро два таких фактора ока¬
зываются в одной группе сцепления, к усилению сцепления между ними.
Точно такое же следствие наблюдается без установления стабильного рав¬
новесия, когда оба фактора такой группы модифицируют величину любого
простого физического измерения и не оказывают никакого другого влияния
на выживание. Таким образом, было показано, что существует постоянная
Вариации как результат мутаций и отбора (продолжение) 121сила, направленная на большее сцепление между факторами одной и той
же группы, и, поскольку величина сцепления неизбежно подвержена селек¬
тивным изменениям и не устраняет, как в большинстве изученных видов,
частый кроссинговер, можно сделать вывод, что некоторые другие причины
должны приводить к равному по силе отбору в пользу кроссинговера. Хотя
математические сложности точного исследования заслуживают значитель¬
но более глубокого внимания, высказывается предположение, что подобная
сила может стоять за преимуществом комбинирования различных полезных
мутаций, которое, если только мутации не происходят последовательно, мо¬
жет быть достигнуто лишь с помощью рекомбинации. Кажется вероятным,
что для того, чтобы оказать заметное влияние на сцепление, поток полезных
мутаций должен быть весьма значительным, и что с дальнейшим углубле¬
нием наших знаний в этой области может оказаться возможным измерить
его величину.Мутации, эффект которых ограничен воздействием на простые мет¬
рические признаки, будут, если только этот эффект не слишком ничтожен,
подвергаться действию противоотбора, и мутантный ген, вне зависимости
от того, положителен или отрицателен его эффект, будет стремиться к пере¬
ходу в рецессивное состояние. Доминантность в подобных случаях долж¬
на быть неполной, хотя в большинстве факторов, подверженных действию
длительного противоотбора, гетерозиготы должны отличаться от немутант¬
ных гомозигот не менее чем на У20 различия между гомозиготами. Дока¬
зательство доминантности, наблюдаемое в случае метрических признаков,
таким образом, полностью согласуется с аналитической теорией.Меристические вариации являются во всех изученных случаях резуль¬
татом скрытых количественных переменных. Видовые модификации по¬
добных признаков, однако, встречают постоянное противодействие со сто¬
роны отбора и возникают из взаимодействия связанных с ними структур.
Поразительный консерватизм, демонстрируемый большой группой орга¬
низмов в отношении меристических признаков, оказывается, таким обра¬
зом, понятным, хотя возможность их модификации в любое время в ходе
реорганизации взаимоотношений и связей ассоциированных структур по¬
стоянно сохраняется. Аномалии, подобные тем, что встречаются в вариаци¬
ях числа шейных позвонков у млекопитающих, могут служить в качестве
иллюстрации обеих сторон действия этого принципа.
Глава 6Половое размножение и половой отборСравнение полового и бесполого размножения. Природа вида. Расщепление
вида. Половое предпочтение. Половой отбор. Модификации, ограниченные полом.
Естественный Отбор и соотношение полов. Резюме.Всем, кто вслед за Психеей отделяет семена добра от семян зла,
я посвящаю этот трактат. ФРЕЗЕРСравнение полового и бесполого размноженияНекоторые группы организмов, размножающиеся неполовым путем,
способны тем не менее эволюционировать под действием естественного от¬
бора. Я не считаю, что это заявление можно с уверенностью отнести к лю¬
бой известной группе. Однако, поскольку невозможно провести четкую гра¬
ницу между целым рядом процессов бесполого размножения — от простого
роста индивида через регенерацию поврежденных или утраченных частей
тела до вегетативного размножения путем почкования, — то напрашивает¬
ся вывод, что бесполое размножение — это примитивное свойство живой
материи, и что половое размножение большинства типов организмов — это
достижение, имеющее особую ценность для всех, кто сумеет им воспользо¬
ваться. Систематизация групп с бесполым размножением не кажется невоз¬
можной, поскольку всегда существуют группы родственных форм, отде¬
лившихся в ходе дивергенции от общего предка. Откровенно говоря, едва
ли мы имеем право употреблять термин «вид», поскольку каждый отдель¬
ный генотип будет иметь равное право рассматриваться как отдельный вид,
и не будет существовать естественных групп, в которых, как в пределах
вида, осуществляется постоянный обмен зародышевой плазмой.Можно сказать, что наиболее близко соответствовать видам могли бы
такие группы, которые в силу своей приспособленности занимали бы столь
сходное положение в природе, что каждый отдельный индивид мог бы за¬
менить другого, или, говоря точнее, что эволюционное улучшение каж¬
дого отдельного индивида угрожало бы существованию потомства всех
Половое размножение и половой отбор123остальных.41 Внутри подобной группы рост численности более успешных
типов был бы уравновешен постоянным вымиранием менее приспособлен¬
ных по выживанию линий, так что точно так же, как, оглядываясь назад,
мы могли бы проследить родословную всей группы вплоть до одного един¬
ственного предка, глядя вперед, мы могли бы на любом этапе предсказать,
в какое время вся группа станет потомками одного конкретного индивида
существующей популяции. Если представить, что станет с полезной мута¬
цией, которая может произойти в любой момент, максимально распростра¬
ниться среди целой группы и, таким образом, привести к эволюционному
прогрессу, то ясно видно, что данная перспектива будет зависеть от вероят¬
ности того, что мутация произойдет именно у такого индивида, наибольшее
число потомков которого получит возможность выжить. На первый взгляд
кажется, что вероятность подобных событий крайне мала, но мы должны
помнить, что коль скоро мутация полезна, она увеличит шансы индиви¬
да, ставшего ее носителем, победить в борьбе за выживание. В предель¬
ном случае, когда польза, полученная от новой мутации, стремится к нулю,
очевидно, что существует только один шанс на успех на все количество
конкурирующих между собой индивидов группы. Если, с другой сторо¬
ны, преимущества очевидны, шанс на успех окажется, безусловно, выше,
отличаясь на величину, которая будет в этом случае зависеть от величи¬
ны наследуемого разнообразия в группе и перспективы появления других
полезных мутаций в течение того периода, который потребуется для пол¬
ной замены исходной популяции на ее улучшенный тип. Если общая ча¬
стота мутаций настолько мала, что естественное состояние группы — это
состояние генетической однородности, можно ожидать, что любая полез¬
ная мутация сможет закрепиться при условии, что она избежит случайной
гибели в течение начального периода, когда ее носителями будут один или
несколько индивидов. Эти шансы, практически равные как при половом,
так и при бесполом размножении, рассматривались в главе 4.Эволюционный прогресс группы, размножающейся бесполым путем,
представляет, таким образом, дилемму — в одном случае использовать все
возникающие полезные мутации и пережить опасности начального перио¬
да можно только тогда, когда частота возникновения мутаций крайне мала,
и находящаяся в обычном состоянии популяция конкурирующих организ¬
мов пребывает в состоянии генетической однородности, но при таком поло¬
жении эволюционный прогресс будет неизбежно топтаться на месте; тогда
как в обратном случае, когда частота как полезных, так и вредных мутаций
достаточно высока, чтобы поддерживать в популяции генетическое разно¬
образие, только наиболее приспособленные генотипы смогут стать пред¬
ками будущих поколений, и возникающие полезные мутации будут иметь
124Глава 6только ничтожный шанс не появиться у организмов, столь нижестоящих
по отношению к своим конкурентам, что их потомство окажется неизбеж¬
но вытеснено потомством последних. Между этими двумя крайностями,
несомненно, существует оптимальная степень мутабильности, зависящая
от пропорции полезных и вредных мутаций и, следовательно, от степени
приспособленности группы к своему положению в природе, но нетрудно за¬
метить, что скорость прогресса, даже при наличии оптимальной мутабиль¬
ности, в группах с бесполым размножением все же крайне мала по срав¬
нению с находящимися в тех же условиях организмами с половым размно¬
жением.Приведенное выше соображение относительно скорости эволюционно¬
го прогресса в группе бесполых организмов можно применить к эволюци¬
онному прогрессу в любом отдельно взятом локусе у вида организмов с по¬
ловым размножением, если предположить, что различного рода изменения
могут происходить в одном гомологичном наборе генов. Сравнительные
скорости прогресса групп с бесполым и половым размножением, занима¬
ющих сходное положение в природе и на данный момент равным образом
приспособленных к этому положению, зависят, таким образом, от числа
различных локусов у видов с половым размножением, гены которых сво¬
бодно обмениваются при наследовании. Исходя из того, что известно о выс¬
ших животных, это число должно равняться по меньшей мере нескольким
тысячам, но даже единственный организм с половым размножением, име¬
ющий всего два гена, имел бы, вероятно, заметное преимущество перед
своим бесполым конкурентом, и даже не обязательно из-за физиологическо¬
го преимущества, которое несет объединение полов, но из-за почти двой¬
ной скорости, с которой организм мог бы отвечать на Естественный От¬
бор. На основании этого, хотя бесполое размножение могло бы в большей
или даже исключительной степени использоваться определенными видами,
принадлежащими к группе организмов с половым размножением, те груп¬
пы, где мы не должны ожидать возникновения полового размножения, если
таковые существуют, должны были бы обладать столь простой природой,
что их генетическая структура включала бы один единственный ген.42Природа видаНа основании генетических исследований высших организмов можно
сделать вывод, что, несмотря на то, что генетическое разнообразие может
существовать в сотнях различных локусов, все же для подавляющего боль¬
шинства локусов нормальное состояние — это состояние генетического еди¬
нообразия. Если бы все было иначе, концепция гена дикого типа была бы
Половое размножение и половой отбор125неопределенной. Хотя известны случаи, как например, случай с локусом
agouti у мышей, где было обнаружено более одной альтернативной формы
гена дикого типа, при этом каждая была доминантной по о тношению к сво¬
им нелетальным аллеломорфам, но как бы многочисленны ни были локусы,
где вероятно существование подобного генетического разнообразия, у нас
есть некоторые основания предположить, что они образуют лишь незначи¬
тельное меньшинство всех остальных локусов, и что подавляющее боль¬
шинство демонстрирует внутри вида, по существу, именно то полное еди¬
нообразие, которое, как было показано, является необходимым, если тре¬
буется получить максимальное преимущество от возникновения полезных
мутаций. Во многих локусах прямыми потомками единственной полезной
мутации должно быть все множество существующих генов вида.Некоторые авторы не уделяют достаточно внимания тем внутренним
связям, которые объединяют всех особей одного вида благодаря половому
размножению. Не считая преодоления географических барьеров, которое
случилось столь недавно, что отдельные расы еще нельзя рассматривать
как различные виды, родословная каждого отдельного индивида, просле¬
женная на протяжении всего ста поколений, должна охватывать практиче¬
ски весь более ранний период, который внес поддающийся оценке вклад
в родословную современной популяции. Если мы проведем исследование
200, 1 ООО или 10000 поколений, что является достаточно коротким перио¬
дом в истории большинства видов, то станет очевидно, что общность про¬
исхождения должна быть еще более полной. Генетическая идентичность
в большинстве локусов, которая скрывается за генетической изменчиво¬
стью большинства видов, способна, вероятно, дать систематику истинную
основу для выводов о видовой идентичности или разнообразии. В книге
Contributions to the Study of Variation У. Бэтсон часто приводит аргумент, ко¬
торый, очевидно, глубоко повлиял на него, а именно: что отсутствие непре¬
рывности, наблюдаемое между различными видами, должно иметь в своей
основе отсутствие непрерывности в эволюции каждого из них. Этот аргу¬
мент, если судить по его работе, которая получила известность скорее бла¬
годаря остроте языка, чем разумности суждений, мог бы быть справедлив
для сугубо бесполых организмов, так как только среди подобных организ¬
мов каждый индивид, а не просто каждый видовой тип, может считаться
последним членом последовательности, о прерывности или непрерывно¬
сти которой можно было бы судить по различиям, наблюдаемым между
родителем и потомком, и на основании этого сделать вывод, что именно та¬
ким образом возможно объяснить разнообразие четко выраженных линий.
Для организмов с половым размножением этот довод неприемлем, посколь¬
ку каждая особь — не конечный член единой последовательности, но член
126Глава 6сходящихся родословных линий, которые достаточно быстро разветвляют¬
ся по всей видовой группе. Существующие внутри вида вариации подобны
разноокрашенным нитям, которые тысячи раз перекрещиваются и расхо¬
дятся вновь, сплетаясь в единое полотно.Реальная идентичность отдаленных предков всех существующих чле¬
нов одного раздельнополого вида может быть представлена иначе, что
в отдельных случаях дает возможность сделать некоторые количественные
уточнения. В наследуемой дисперсии любого признака в каждом поколе¬
нии часть является результатом наследственных различий родителей, то¬
гда как все остальное, включая почти все различия между всеми братьями
и сестрами можно отнести на счет генетического расщепления. Эти доли
не сильно различаются; корреляции, наблюдаемые в демографии, показы¬
вают, что на долю расщепления должно приходиться немного более 2/5,
а на наследуемые родительские различия — около 3/5 общей дисперсии.
Эти наследуемые различия, в свою очередь, если вернуться на одно по¬
коление назад, являются результатом расщепления и наследуемых разли¬
чий, доставшихся от дедушек и бабушек. Если оглядываться все дальше
и дальше назад, то видно, что доля дисперсии, приписываемая различиям
среди предков, быстро уменьшается; если мы полагаем, что доля, отно¬
симая за счет расщепления, составляет всего 2/5 в каждом поколении, то
доля, относимая за счет различий среди предков всего 10 поколений на¬
зад, составляет всего 1 на 160, тогда как 30 поколений назад она равняется
менее 1 на 4 миллиона. Только географические и другие барьеры на пу¬
ти свободного скрещивания между расами, то есть факторы, по общему
признанию подобные тем, которые обусловливают развитие зарождающих¬
ся видов как географических рас, помешали всему человечеству, исключая
последние 1 ООО лет, иметь практически идентичное происхождение. Если
исключить последние 500 лет, то различие в происхождении представите¬
лей одной и той же нации будет крайне мало; если исключить последние
2 000 лет, то, по-видимому, единственными оставшимися различиями были
бы различия между выраженными этнографическими расами; этнографи¬
ческие расы или по крайней мере их элементы и в самом деле могут быть
достаточно древними образованиями, но и этот случай может быть верен
только при условии, что на протяжении долгих лет диффузия крови между
отдельными группами практически отсутствовала.Расщепление видаТесные генетические связи, которые объединяют вид в единое це¬
лое, высвечивают проблему их расщепления — проблему, сложности ко¬
Половое размножение и половой отбор127торой родственны сложностям, возникающим при обсуждении расщепле¬
ния небесных тел, поскольку попытка проследить ход событий посред¬
ством промежуточных состояний нестабильности требует, по-видимому,
как в том, так и в другом случае более точных знаний, чем дает изуче¬
ние стабильных состояний. Нет сомнения, что во многих случаях полная
или почти полная географическая изоляция мгновенно проводит раздели¬
тельную черту; две отдельные группы далее эволюционируют как отдель¬
ные виды практически независимо в различных средах обитания до тех
пор, пока морфологические отличия между ними не дадут право каждой
из них получить статус «видового ранга». Хотя заявлять, что видовая диф¬
ференциация всегда является следствием географической изоляции, озна¬
чает идти в разрез с общепринятым мнением. Во многих случаях можно
с уверенностью сказать, что никакой географической изоляции не наблю¬
дается, хотя это вовсе не означает, что условия обитания вида настолько
равно благоприятны как для его поддержания, так и для его миграции,
что никаких «слабых линий», которые, коль скоро дивергенция все рав¬
но неизбежна, определили бы наиболее вероятные географические линии
раздела, не существует. Конечно, в том случае, когда минимальная причина
вызывает непропорционально большой эффект, можно говорить о наличии
нестабильных состояний; таким образом, при обсуждении возможности ди¬
вергенции вида в отсутствие географической изоляции будет достаточно,
если мы сможем дать ясное представление о природе причин, вызывающих
генетическую нестабильность.Любая гетерогенность окружающей среды, требующая особых адап¬
таций, которые сложно или вовсе невозможно согласовать между собой,
неизбежно оказывает определенное давление на связывающие вид силы.
Это давление оказывается наименьшим, когда близкородственные инди¬
виды подвергаются воздействию различных факторов окружающей сре¬
ды, и полностью исчезает, когда каждый индивид имеет равную возмож¬
ность столкнуться с любой из двух противоположных средовых ситуаций
или с каждой из целого ряда ситуаций. Давление бывает максимальным
в том случае, когда оно зависит от обстоятельств, не способствующих кон¬
такту полов, из которых наиболее заметным является географическая уда¬
ленность, хотя и другие обстоятельства, такие, например, как размножение
в более ранние или более поздние сроки, также имеют значение. Я не зна¬
ком ни с одним подобным обстоятельством, влияние которого бы в данной
генетической ситуации существенно отличалось бы от влияния географиче¬
ской удаленности, что, таким образом, подтверждает правильность теории.Мы можем рассмотреть случай вида, подверженного различным усло¬
виям выживания и размножения на противоположных сторонах своего гео¬
128Глава 6графического ареала. Определенные гены, существующие в виде альтер¬
нативных вариантов, могут оказаться полезными на одном краю ареала,
и их число будет возрастать, тогда как на другом краю ареала они мо¬
гут оказаться вредными, и их число будет сокращаться, в то время как
промежуточный регион может быть разделен на ряд участков, где преиму¬
щество будет возрастать от отрицательной величины на одном краю аре¬
ала через нулевое значение, где селективное преимущество будет точно
сбалансировано, до определенного положительного преимущества на про¬
тивоположном краю ареала. Таким образом, условие генетического рав¬
новесия устанавливается только в том случае, когда рост частоты гена
в благоприятном регионе и уменьшение частоты в неблагоприятном ре¬
гионе не только уравновешивают друг друга количественно, но и равня¬
ются скорости диффузии генов путем миграции и благодаря контактам
полов из разных регионов. Сама эта скорость должна определяться, если
не учитывать склонности вида к миграции или оседлому образу жизни,
протяженностью каждого региона, в пределах которого происходит диф¬
фузия генов, плотностью популяции в этом регионе и, наконец, градиен¬
том соотношения частот гена и его аллеломорфа при прохождении через
регион. Пока возможно поддержание достаточного градиента, сопровожда¬
емого активной диффузией зародышевого материала, до тех пор местные
разновидности, пусть и имеющие заметные различия, не будут стремиться
к их усилению, определяемому частотой генов, по которым они различают¬
ся, и будут связаны между собой всеми ступенями переходных типов по¬
пуляций.Чем длительнее будет поддерживаться подобное равновесие, тем более
многочисленными будут становиться генетические различия между типа¬
ми, населяющими крайние регионы, поскольку данная ситуация не будет
способствовать ни исчезновению гена, приспособленного к местным усло¬
виям, ни его аллеломорфа, приспособленного к каким-либо другим услови¬
ям, и все новые мутации, возникающие в промежуточной зоне, которые
обладают преимуществом на одном краю ареала, но вредны на другом,
получат возможность добавиться к факторам, по которым они отличны.
В дополнение к тем генам, которые отбираются дифференцированно кон¬
трастными условиями окружающей среды, мы должны добавить еще и те,
селективное преимущество или недостаток которых определяются геноти¬
пом, в который они включены, и которые, следовательно, будут обладать
различной ценностью для выживания не под прямым воздействием проти¬
воположных условий среды, а опосредованно, благодаря генотипическому
контрасту, который вызывают эти противоположные условия окружающей
среды. Вышеописанный процесс не содержит ничего нового, он способ¬
Половое размножение и половой отбор129ствует дифференцировке местных рас под действием естественного отбора
и показывает, что подобная дифференцировка в условиях постоянства диф¬
фузии должна быть прогрессирующей. Он не ведет ни к прерыванию про¬
цесса диффузии, ни к значительному ослаблению единства происхождения
вида. Этот процесс подобен растяжению материального тела под воздей¬
ствием напряжения, но не его разрыву.Существуют, однако, группы наследственных вариаций, которые ока¬
зывают влияние на диффузию. В рассматриваемом нами случае, где при¬
чина изоляции — географическая удаленность, инстинкты, управляющие
путями миграций, или особенности организма, способствующие распро¬
странению или закреплению особей на одном месте, будут влиять на ча¬
стоту, с которой потомки организма, родившегося в одном районе, ока¬
жутся в условиях среды, свойственных другому. Постоянная элиминация
во всех крайних областях распространения генов, проникших туда из дру¬
гой области, должна попутно затронуть и наиболее склонные к диффузии
типы индивидов. Если допустить, что водный организм, приспособленный
к низкому уровню солености, приобретет под действием Естественного От¬
бора инстинкт к миграции или способность к расселению, что снизит ве¬
роятность того, что он может быть унесенным в море, то можно ожидать,
что отбор, идущий в этом же направлении, будет постепенно сокращать
диффузию зародышевой плазмы между областями с разными условиями
обитания. Подобное постепенное сокращение диффузии приведет к росту
градиента частоты гена в тех местах, где частота максимальна, в результа¬
те чего появится разграничительная линия, по обе стороны которой можно
будет наблюдать сравнительно резкое различие генетического состава вида.
Диффузия через эту линию становится теперь как никогда нежелательной,
и ее постепенное уменьшение, пусть и при условии существования в те¬
чение достаточно длительного времени области, населенной организмами
переходного типа, позволяет двум основным частям вида эволюциониро¬
вать почти совершенно независимо.В тех случаях, когда причина генетической изоляции не просто гео¬
графическая удаленность, а различия между разными представителями ви¬
да по их местообитаниям или образу жизни, становятся полезными самые
разные генетические модификации; изоляция не будет, конечно, увеличи¬
ваться за счет дифференциальной модификации миграционных инстинктов
или способов распространения; но результатом изоляции является разнооб¬
разие наследственных модификаций, которые сводят к минимуму процесс
диффузии зародышевых элементов какой-либо формы жизни в ареале рас¬
пространения другой формы жизни. В конечном итоге эти зародышевые
элементы исчезнут.
130Глава 6Способность организма к расширению своего ареала сама по себе яв¬
ляется эффективным инструментом, даже при наличии фактора селективно¬
го различия в любом из двух регионов. Это отлично иллюстрируется теори¬
ей Рэя Лэнкестера, дающего убедительное объяснение снижению или пол¬
ной потере зрения обитателями пещер. Рэй Лэнкестер отмечает, что облада¬
ние органом зрения не просто не нужно подобным животным, но, благопри¬
ятствуя их миграции ближе к источникам света, постоянно способствует их
элиминации из массы обитателей пещеры, независимо от того, выживают
они или погибают в новой обстановке. Таким образом, те, кто останутся
размножаться в пещере, будут подвергаться в каждом поколении действию
отбора, направленного на невосприимчивость к зрительному стимулу. Сле¬
дует отметить, что в таких изолированных местах обитания подобный от¬
бор путей миграции способен достигать чрезвычайно высокой интенсивно¬
сти и, соответственно, давать очень быстрый эволюционный эффект.Половое предпочтениеОдин из способов генетической изоляции, особое значение которого
определяется тем, что он равно приложим как к случаю географической
удаленности, так и к другим случаям, это способ, который мы, за неиме¬
нием лучшего термина, называем репродуктивным, или половым, предпоч¬
тением.Успешность организма, заключающаяся в том, чтобы оставить после
себя многочисленное потомство, измеряется не только числом оставшихся
в живых потомков, но также их качеством или их вероятным успехом в про¬
изводстве своего потомства. Таким образом, немаловажно, какой именно
из имеющихся в наличии индивидов будет их вторым родителем. Высшие
животные делают свой выбор на основании осмотра вероятного партнера,
поскольку существование в больших группах предполагает наличие доста¬
точно развитых органов чувств, позволяющих ощущать индивидуальные
различия. Таким образом, вполне возможно, что эмоциональные реакции,
вызываемые различными индивидами противоположного пола, как в случае
с человеком, будут неодинаковы, и что по крайней мере отдельные индиви¬
ды каждого пола будут менее склонны образовывать пару с одними парт¬
нерами, нежели с другими. У растений подобным средством отбора может
считаться неодинаковая скорость роста разных типов пыльцевых трубок
при прорастании на одном и том же столбике.Замечательное обобщение недавно установленных фактов в этой обла¬
сти было предложено Джонсом (Selective Fertilization, University of Chicago,
1928). Известны случаи с кукурузой, где отбору подвергается пыльца, несу¬
Половое размножение и половой отбор131щая определенные вредные мутантные гены, и один случай с ослинником
{Oenothera), где отбираются яйцеклетки, несущие определенный летальный
ген. В этих реакциях отбору подвергается как генотип материнского расте¬
ния, так и пыльцы, и именно этим объясняется замечательный факт, уста¬
новленный самим Джонсом в ходе его наблюдений за кукурузой: нанесен¬
ная на пестик смесь пыльцы в целом оказывается тем менее эффективна,
чем более велико генетическое различие между родительскими растения¬
ми. Подобная общая тенденция к гомогамии, наиболее заметная у кукуру¬
зы из-за огромного числа рецессивных дефектов, накопленных этим рас¬
тением в результате постоянного перекрестного опыления, в значительно
меньшей степени способствовала бы дивергенции вида, чем отбор диф¬
ференциальных тенденций, специально направленных на достижение этой
цели, подобно тому, как должно было бы, вероятно, происходить в группе,
куда постоянно вторгаются вредные гены.В общем, условия, при которых возможна успешная дискриминация,
могут быть следующими: (1) принятие одного партнера исключает эффек¬
тивное принятие альтернативных партнеров; (2) за отклонением одного
предложения обязательно последуют другие предложения. Вероятность то¬
го, что предложение не будет сделано, невелика, так как особь в случае при¬
нятия предложения получает селективное преимущество. Первое условие
выполняется самками большинства видов и в значительном числе случаев
также и самцами. В других случаях, хотя это было бы серьезной ошиб¬
кой, если бы самец преследовал уже оплодотворенную самку, можно было
бы предположить, что любая возможность эффективного спаривания мо¬
жет рассматриваться как преимущество. Второе условие наиболее очевид¬
но удовлетворяется в том случае, когда представители избираемого пола
находятся во время спаривания в явном большинстве.Величайший недостаток полового предпочтения у животных — спари¬
вание разных видов друг с другом, в результате чего гибриды бывают либо
бесплодны, либо не оставляют потомства из-за серьезных препятствий, ко¬
торые возникают в результате смешения инстинктов и других характерных
свойств, являющихся принадлежностью различных стилей жизни. У выс¬
ших животных оба пола имеют врожденные механизмы того, чтобы избе¬
жать этой неприятности, и, исходя из относительно редкого возникновения
естественных гибридов у растений, за исключением определенных родов,
у которых видовая специфичность может нарушаться именно из-за суще¬
ствования этого самого недостатка, мы можем сделать вывод о естествен¬
ном преобладании механизмов, которые служат для уменьшения вероят¬
ности оплодотворения пыльцой чужеродного растения. Можно, таким об¬
разом, сделать вывод, что у высших животных нервная система функцио¬
132 Глава 6нирует таким образом, что различия во внешнем виде и поведении особи
другого вида подавляют естественные поведенческие инстинкты, проявля¬
ющиеся при контакте с особью своего вида. Мы не можем, конечно, с точ¬
ностью сказать, какие именно различия в сенсорных стимулах определяют
эту различную реакцию, но не секрет, что механизм дифференциации суще¬
ствует, и его вариации могли бы положить начало сходной дифференциации
внутри собственного вида, если бы подобная дифференциация оказалась бы
когда-нибудь полезной.Типичная ситуация, когда подобная дифференциация сможет дать
определенное преимущество особям обоих полов, должна возникнуть в том
случае, когда вид, занимающий сплошной ареал, находится в процессе рас¬
щепления на два дочерних вида, каждый из которых по своему адаптиро¬
вался к определенной области этого ареала, поскольку в каждой крайней
области ареала у определенной части особей каждого поколения регуляр¬
но будут проявляться определенные относительно неблагоприятные при¬
знаки вследствие диффузии из других областей ареала генов, несущих эти
признаки. Индивиды с подобными свойствами окажутся, несомненно, ме¬
нее, чем их соперники, приспособлены к возникшей ситуации, и пока эти
свойства будут заметны, как например, различия в окраске, их присутствие
будет способствовать селективному процессу, благоприятствующему поло¬
вому предпочтению группы, в которой они живут. Индивиды каждого ре¬
гиона, наиболее привлекательные или испытывающие влечение к партне¬
рам наиболее предпочтительного типа, в противоположность возможным
партнерам противоположного типа будут, в действительности, лучше пред¬
ставлены в будущих поколениях, и как дискриминация, так и половое пред¬
почтение будут тем самым усиливаться. Несомненно, существует возмож¬
ность, что эволюция полового предпочтения по этой причине возведет на¬
дежный барьер между двумя разделившимися частями вида, даже в том
случае, когда географический и другие факторы наименее благоприятству¬
ют подобному разделению.Половой отборТеория, выдвинутая Дарвином для объяснения эволюции вторичных
половых признаков, включает два достаточно четко различимых принци¬
па. В одной группе случаев, широко представленных среди млекопитаю¬
щих, самцы, особенно если они полигамны, борются за обладание самка¬
ми. На мой взгляд, мало кто может усомниться в том, что существующий
отбор самцов способен объяснить эволюцию как специального оружия, на¬
пример рогов, так и драчливости в период брачного сезона, особенно с тех
Половое размножение и половой отбор133пор, как изучение влияния половых гормонов показало, что генетические
модификации целого вида могут проявляться у представителей только од¬
ного пола, что, таким образом, устраняет единственный сложный момент
в теории Дарвина.Для второго класса случаев, где типичным является удивительное раз¬
витие оперения самцов фазанов, Дарвин выдвинул смелую гипотезу, что
эти замечательные черты объясняются кумулятивным действием полового
предпочтения, проявляемого самками в брачный период. Два класса случа¬
ев были объединены Дарвином вместе как имеющие общий важный эле¬
мент соперничества, включающего возможности взаимного столкновения
и обструкции и ограниченного особями одного пола. Для ряда других на¬
туралистов различие между двумя типами кажется более важным, чем этот
общий элемент, особенно тот факт, что второй тип объяснения включает
желание или выбор самки. Уоллес принял без колебаний влияние взаим¬
ных схваток самцов на эволюцию оружия, связанного с битвами за самку,
но полностью отверг влияние выбора самкой на эволюцию ограниченного
полом признака — орнамента хвоста самца.Как было указано в главе 2, для того чтобы знать, как работает Есте¬
ственный Отбор, необходимо иметь точные данные о смертности и рож¬
даемости вида на всех возрастных этапах, а также влияния всех возмож¬
ных генетических замещений на эти показатели. Различие между одним
типом отбора и другим потребовало бы гораздо более детальной информа¬
ции, поскольку нам потребовалось бы знание не только общих показателей
смертности и рождаемости, но также природы и частоты всех экологиче¬
ских ситуаций, в которых эти события случаются. Предусмотренная зако¬
ном классификация причин смерти достаточно сложна и требует обширных
медицинских знаний, если в каждом случае делать все как положено. Од¬
нако даже квалифицированный медик не обязан выяснять социальные при¬
чины смерти. Указывая на невероятную сложность задачи по выяснению,
каким именно способом отбора следует объяснить известное эволюционное
изменение, или какую долю имеют различные способы отбора в произво¬
димом эффекте, я бы не хотел, чтобы казалось, что я подвергаю сомнению
ценность подобных различий, как иногда случается при оценке различных
экологических ситуаций, в которых осуществляется отбор. Морфологиче¬
ские явления могут быть так поразительны, образ жизни и инстинкты, ве¬
роятно, уже так тщательно изучены в естественной среде обитания, что
любого человека, обладающего этими знаниями можно убедить, что од¬
ного единственного объяснения из всех предложенных бывает достаточно
для убедительного доказательства. Каждый случай должен быть, я полагаю,
досконально изучен, и суждение вынесено теми, кто знаком с деталями
134Глава 6данного случая, и даже тогда в подавляющем числе случаев доводы будут
довольно неубедительными. Выносить подобное решение в каждом отдель¬
ном случае шло бы вразрез с целью этой книги (и с устремлениями автора).
Впрочем, мне кажется, что здесь найдется место для более точной оценки
обоснованности различно используемых здесь доводов, и которыми следу¬
ет пользоваться, если вообще предпринимается какая-либо оценка фактов.Очевидно, кому-то покажется, что подобная абстрактная форма изло¬
жения наносит вред интересам предмета. С другой стороны, я уверен, что
те, кто имели дело с наблюдением и классификацией, будут приветствовать
любую серьезную попытку объективной оценки фактов. Нужда в этом осо¬
бенно велика в отношении предмета, где обобщение, основанное большей
частью на наблюдениях, имеет такой большой вес, что в противополож¬
ность этому простая констатация свежих фактов кажется подчас результа¬
том логических заключений, которые в данном случае отсутствуют; таким
образом, возможно даже для самых светлых умов, даже в случаях полной
убежденности в правильности собственной интерпретации, причем убеж¬
денности полностью оправданной, использовать в поддержку своей точки
зрения такие доводы, которые при сомнениях в своей правоте вряд ли бы
использовались. Стоит взглянуть, например, лишь на один случай наиболее
невинной логической ошибки в обсуждении полового отбора. Уоллес ука¬
зал, что многие виды птиц отличаются броской или заметной окраской, где
самки имеют либо такие же наряды, что и самцы, либо другие, но не ме¬
нее яркие, или гнездятся в укромных местах, таких, например, как ямки
в земле или дупла деревьев, или строят сводчатое или закрытое гнездо так,
чтобы полностью скрыть сидящую внутри птицу. Эту скрытность Уоллес
объяснил отсутствием защитной окраски у самки. В ответ на возражения,
которые последовали, кажется, от герцога Аргилла, что большое сводчатое
гнездо гораздо заметнее для врага, чем маленькое открытое гнездо, Уоллес
возразил, что подобное гнездо все же защищает от нападения, поскольку
ястребы и вороны не могут его разрушить. Дарвин, со своей стороны, счи¬
тал, что замечание герцога Аргилла правдоподобно, особенно если речь
идет о древесных хищниках. Можно заметить, что ни один из участни¬
ков спора не заметил, что вопрос не сводится к преимуществам или недо¬
статкам закрытого гнезда, или что, какими бы неудобными ни казались
закрытые гнезда, они тем не менее отвечают условиям, которые, согласно
Уоллесу, требуются для исключения отбора защитной окраски у сидящей
на гнезде самки под действием хищников, которые в противном случае лег¬
ко бы ее заметили.Гораздо более серьезная ошибка, которую не обошло внимание био¬
логов, была сделана Уоллесом, когда тот возражал, что эффект отбора
Половое размножение и половой отбор135во взрослом состоянии снижен большой смертностью на ранних стадиях
развития (Darwinism, р. 296).У бабочек смертность в результате естественного отбора достигает огромных
размеров на стадии яйца, личинки и куколки, и, пожалуй, не более чем одно из сот¬
ни отложенных яиц дает совершенное насекомое, которое доживает до периода раз¬
множения. Здесь, однако, слабость отбора самкой, если он существует, должна быть
полной, поскольку если только наиболее ярко окрашенные самцы не обусловливают
появление наиболее защищенных яиц, личинок и куколок, и если только эти самые
яйца, личинки и куколки, которые смогли выжить, это не те, которые дают наиболее
яркоокрашенных бабочек, то любой выбор, который могла бы сделать самка, должен
быть полностью лишен смысла. Если, с другой стороны, корреляция между развити¬
ем окраски и приспособленностью на всех стадиях развития все-таки существует,
тогда это развитие неизбежно попадет под действие естественного отбора и общих
законов, которые определяют образование окраски и украшающих придатков.Следует заметить, что в случае, если одна зрелая форма имеет пре¬
имущество перед другой, если одна взрослая особь имеет больше шан¬
сов оставить потомство, чем другая взрослая особь, это преимущество ни
в коей мере не уменьшается из-за смертности на ранней стадии развития
при условии отсутствия связи между признаками зрелой и незрелой особей.
Смертность незрелых форм может быть в тысячи раз больше, что и наблю¬
дается в случае гибели половых клеток, но это не оказывает ни малей¬
шего влияния на эффективность отбора зрелых форм. Кроме того, Уоллес
сам придавал большое значение другим селективным эффектам, действую¬
щим на взрослых бабочек, что видно из его анализа покровительственного
сходства на странице 207 той же книги. Таким образом, определяющей,
во мнении Уоллеса, была не убедительность используемого им аргумента,
а, скорее, твердая уверенность в том, что эстетические способности свой¬
ственны только человеку и являются частью его «божественной природы»,
доставшейся ему в дар от высшей силы, что и объясняет слабость его по¬
зиции.Для того чтобы эволюционное изменение можно было бы отнести
за счет полового отбора, должны выполняться два основных условия:
(1) наличие полового предпочтения, по меньшей мере у одного пола,
и (2) экологические условия, в которых подобное предпочтение давало бы
репродуктивное преимущество. В тех случаях, когда удовлетворяются оба
эти условия, существование особых признаков, которые с позиций мор¬
фологии могут считаться нужными в качестве украшения, но не прино¬
сить никакой другой пользы их обладателю, в сочетании с экологическими
свидетельствами того, что наибольшего развития эти признаки достигают
в брачный период и в это время настойчиво выставляются на всеобщее
136Глава 6обозрение, являются признаками того же рода, что и признаки тела, эволю¬
ционное значение которых бывает очевидно.Если говорить не о человеке, а о животных, то прямые доказатель¬
ства существования полового предпочтения едва ли существуют. Весьма
странные предпочтения отдельных живущих в неволе птиц дают основа¬
ние предположить, что их психическое состояние, по крайней мере в отно¬
шении половых предпочтений, нарушено условиями искусственного содер¬
жания и дает возможность убедиться, что нервная система функционирует
и по-разному реагирует на ухаживание разных самцов. Может показать¬
ся, что единственной ценностью данных наблюдений является определе¬
ние того, насколько курицы солидарны в выборе тех или иных петухов.
Поскольку, как я полагаю, в каждый брачный сезон можно наблюдать ре¬
зультат только одного выбора, достаточное количество результатов можно
получить, лишь проводя подобный опыт с полигамными птицами; впрочем,
если порядок предпочтения существует, то полигамные птицы должны его
уверенно демонстрировать.Наиболее веский аргумент, высказанный Дарвином относительно
птиц, живущих на воле, должен получить сейчас совсем другое толкова¬
ние. Он приводит множество примеров того, как в том случае, когда од¬
на птица из пары оказывается подстреленной, ее место оказывается почти
мгновенно занято другой птицей того же, что и погибшая, пола, неваж¬
но, кто это был, самец или самка. Он делает заключение, что, каким бы
удивительным это ни казалось, многие птицы обоих полов остаются без
пары, и также, что — и лишь здесь мы не можем с ним согласиться, — это
происходит потому, что они не могут найти себе партнера, который бы им
понравился. Если бы это было верным объяснением, это означало бы, что
половое предпочтение у значительной части птиц столь сильно, что препят¬
ствует спариванию, но подобная сила предпочтения может поддерживаться
у вида только при условии, что преимущество получения потомства от воз¬
можной встречи с суперпартнером больше, чем неизбежная потеря всего
брачного сезона. Как будет видно в дальнейшем, в большинстве случаев
столь большое преимущество не находит рациональной основы. С другой
стороны, исследования Ховарда в книге Territory in the Bird Life дают очень
простое и адекватное объяснение наблюдаемому явлению. С этой точки
зрения птицы, оставшиеся без брачного партнера, оказались в таком поло¬
жении, поскольку не обладают территорией для размножения, где они мог¬
ли бы спокойно сидеть на гнезде, но всегда готовы спариться с вдовой или
вдовцом, обладающими укромным жильем. Впрочем, я не знаю, насколько
доказательны утверждения, что птицы, занимающие освободившуюся тер¬
риторию,— это всегда овдовевшая птица и новый партнер, а не новая пара.
Половое размножение и половой отбор137Усыновление уже существующих птенцов говорит, что иногда, по крайней
мере это — первый из названных вариантов.Если вместо того чтобы рассматривать существование полового пред¬
почтения как базовый факт, наличие которого следует устанавливать только
прямым наблюдением, мы допустим, что предпочтения живых организмов,
равно как их органы и способности, должны рассматриваться в качестве
продуктов эволюционного изменения, возникающего в результате относи¬
тельного преимущества, к которому приводят подобные предпочтения, то
окажется, как было показано в предыдущем разделе, что довольно часто по¬
ловое предпочтение определенного рода способно повлечь за собой селек¬
тивное преимущество и таким образом закрепиться внутри вида. В любом
случае, когда внутри отдельного вида существуют заметные различия, что
в действительности связано с селективным преимуществом, всегда будет
сохраняться тенденция к выбору также таких индивидов противоположно¬
го пола, которые наиболее четко различают видимые отклонения и наибо¬
лее явно предпочитают более успешный тип. Половое предпочтение, воз¬
никающее таким образом, может нести или не нести прямую выгоду из¬
бранным индивидам и, следовательно, усиливать действие Естественного
Отбора. Это означает, что его распространение может быть гораздо шире,
чем развитие четко выраженных вторичных половых признаков.Впрочем, случается, что определенные половые предпочтения подоб¬
ного рода все же имеют удивительные последствия, если они определяются,
например, признаками оперения и развиваются внутри вида, где предпочте¬
ние одного пола, в частности самок, имеет большое влияние на число по¬
томков, оставленных после себя отдельными самцами. В подобных случа¬
ях модификация признаков оперения у петуха происходит под воздействи¬
ем двух селективных воздействий: (1) исходное преимущество, не связан¬
ное с половым предпочтением, величина которого может быть абсолютно
ничтожна, и (2) дополнительное преимущество, связанное с предпочтени¬
ем самки, величина которого будет пропорциональна интенсивности этого
предпочтения. Сама интенсивность предпочтения будет увеличиваться от¬
бором до тех пор, пока петухи матерей, проявляющих подобную разбор¬
чивость, будут иметь какое-либо преимущество перед петухами остальных
матерей, неважно, результатом какого из двух обстоятельств это преимуще¬
ство достигнуто. Значение данной ситуации состоит в том, что дальнейшее
развитие признаков оперения, являясь причиной преимущества, получен¬
ного в результате полового отбора, будет продолжаться и далее, даже после
того, как будет пройдена та точка в развитии, на которой это преимуще¬
ство для Естественного Отбора закончится. Иные, чем половое предпочте¬
ние, силы отбора могут противостоять дальнейшему развитию признака,
138Глава 6и все же развитие будет продолжаться до тех пор, пока вред не станет чуть
больше пользы. Кроме того, пока будет существовать чистое преимуще¬
ство от дальнейшего развития оперения, будет существовать также чистое
преимущество от того, чтобы давать ему более явное предпочтение.Два затронутых этим процессом признака, а именно развитие опере¬
ния у самца и половое предпочтение подобного развития самкой, должны,
таким образом, совершенствоваться совместно и до тех пор, пока на пути
этого процесса не встанет жесткий противоотбор, он будет продолжать¬
ся с постоянно возрастающей скоростью. Становится ясно, что при пол¬
ном отсутствии подобных препятствий скорость развития будет пропорци¬
ональна тому уровню развития, которое уже было достигнуто и которое,
таким образом, будет расти экспоненциально или в геометрической про¬
грессии. Это означает, что в любой экологической ситуации, где половой
отбор способен создать значительное репродуктивное преимущество, су¬
ществуют потенциальные возможности для ускоряющегося процесса, ко¬
торый, как бы мало ни было его начало, должен, если его не ограничить,
вызывать большие последствия и на более поздних стадиях делать это все
быстрее и быстрее.Подобный процесс вскоре неизбежно столкнется с препятствием. Оче¬
видно существование препятствий двух типов. Зашедший слишком дале¬
ко достаточно суровый противоотбор в пользу менее украшенных самцов
должен будет столкнуться с необходимостью сбалансировать преимущества
полового предпочтения; на этой стадии как развитие оперения, так и рост
предпочтения самки затормозятся, и будет достигнуто условие относитель¬
ной стабильности. Хотя все же более эффективным было бы положение,
когда вред от половых украшений самца настолько сократил бы числен¬
ность доживших до брачного периода самцов по отношению к самкам, что
процесс был бы прерван в самом зародыше путем уменьшения репродук¬
тивного преимущества, зависящего от предпочтений самки. Важно отме¬
тить, что состояние относительной стабильности, вызванное тем или иным
обстоятельством, будет длиться значительно дольше, чем процесс, в ходе
которого шло развитие украшений. Вероятно, у большинства существую¬
щих видов ускоряющиеся процессы уже находятся под контролем, и нам
следует считать, что наиболее необычное развитие связанного с полом опе¬
рения не является, подобно большинству признаков, результатом продол¬
жительного и равномерного процесса эволюционного развития, а скорее
результатом внезапного и мгновенного изменения. Теория не дает нам воз¬
можность предсказать исход подобного эпизода, но указывает на большое
преимущество, которое имеет половое предпочтение в качестве его внут¬
реннего условия.
Половое размножение и половой отбор139То, каким именно образом самцы, наиболее эффективно привлекаю¬
щие внимание и интерес самок, получают от этого репродуктивное преиму¬
щество, является весьма сложным вопросом, поскольку полигамия не так
широко распространена, как связанные с полом украшения; и теория поло¬
вого отбора, таким образом, требует, чтобы некоторое репродуктивное пре¬
имущество имели бы также и моногамные птицы. Теория Дарвина по этому
вопросу крайне слаба. Он полагает, что у самок существует положитель¬
ная зависимость между ранним сроком, когда они готовы к спариванию,
и числом потомков, которое они выводят, и что вариации этих признаков
связаны, по его мнению, с лучшим питанием. Трудно сказать, так ли это
на самом деле, но следует отметить, что даты размножения вида могут быть
стабильны лишь в том случае, когда птицы, обладающие врожденной пред¬
расположенностью гнездиться в ранние сроки, не способны по этой при¬
чине дать больше потомства. Зависимость, которая требуется для теории
Дарвина, может объясняться единственно внешними причинами, которые
могут зависеть от случайного различия в количестве пищи. Создается впе¬
чатление, что продемонстрировать как наличие, так и отсутствие подобной
зависимости было бы одинаково сложно.Впрочем, как представляется, существует одно бесспорное преимуще¬
ство, которое получают самцы, спаривающиеся первыми на определенной
территории, и которое, таким образом, может определяться половым пред¬
почтением, это — смертность во время брачного периода. Уровень смерт¬
ности животных часто бывает удивительно высок, и снижение его даже
на один процент в неделю дает значительное преимущество для спари¬
вающихся в более ранние сроки самцов, даже если шансы на выживание
потомства такого самца будут снижены его смертью.Второе обстоятельство, при котором половое предпочтение должно да¬
вать некоторое репродуктивное преимущество, состоит в повторном обра¬
зовании пар птицами, потерявшими партнера во время брачного сезона. Ка¬
жется очевидным, что множество не нашедших себе партнера птиц обыч¬
но с готовностью пользуются подобной ситуацией, и выбор среди них дает
получившим предпочтение особям репродуктивное преимущество в равной
степени как для одного, так и для другого пола. Впрочем, для того чтобы
судить, насколько эффективны различные возможные ситуации, где поло¬
вое предпочтение может дать репродуктивное преимущество, необходимо
детальное знание экологии.Возможно, в подобных исследованиях стоит помнить о том, что наи¬
более изысканно украшенные самцы получают некоторое репродуктив¬
ное преимущество без посредничества женского предпочтения, что мож¬
но сравнить с преимуществом, которое дает владение особым оружием.
140Глава 6Выяснение территориальных притязаний включает частые стычки, но это
ни в коей мере не смертельные схватки; наиболее многочисленные и, следо¬
вательно, наиболее важные для нас случаи, это те, в которых столкновения
не происходит вовсе, и вторгшийся на чужую территорию самец настолько
впечатлен или унижен внешностью своего соперника, что даже не рискует
вступить в бой. Защищая свое жизненное пространство, петух ведет себя
так, как будто знает, какое преимущество дает умение впечатлить не только
самок, но и самцов своего вида, и его гордая осанка, яркие перья и побед¬
ная песня с одинаковым успехом могут быть использованы как в сражении,
так и в ухаживании.Рассматриваемое здесь селективное воздействие совмещает признаки
двух типов воздействий, к которым Дарвин применял термин половой от¬
бор, а именно появления новых средств защиты и нападения в ходе битв
с соперниками и эволюции украшений, которые привлекают и восхищают
самку. Демонстрация силы и воинственности аналогична обладанию этими
свойствами, так как оказывает тот же эффект; но бывает эффект другого ро¬
да, как например, в случае привлекательных украшений, который состоит
в эмоциональной реакции других особей вида. В действительности он воз¬
никает в результате принятия решений, которые следуют под воздействием
полового предпочтения. Стоит, однако, отметить одно отличие: в случае
привлекательных украшений эволюционное воздействие на самку состоит
в том, чтобы сделать ее все более восприимчивой к демонстрируемому на¬
ряду, тогда как эволюционное воздействие боевой раскраски на тех, кого
она должна поразить, состоит в том, чтобы сделать их все менее воспри¬
имчивыми ко всем впечатлениям, кроме тех, которые рождает истинная
доблесть. Приобретенные таким путем украшения самца бывают удиви¬
тельными, но практически никогда не становятся экстравагантными.Модификации, ограниченные поломНекоторая сложность, воспринятая в процессе развития теории поло¬
вого отбора достаточно серьезно, кроется в том, что многие признаки, воз¬
никающие в результате ограниченного полом отбора, появляются только
у того пола, на который этот отбор действует. Сложность состоит в том,
чтобы понять, насколько отбор, действующий только на один пол, может
влиять на оба пола, и может ли процесс мутации, изначально воздействую¬
щий на оба пола, продолжиться у одного пола, если противоположный пол
подвергся воздействию противоотбора.Из доступных для генетических исследований крупных мутационных
изменений большая часть проявляется одинаково у обоих полов, тогда как
Половое размножение и половой отбор141нас интересует то небольшое число мутаций, которые проявляются либо
по-разному у разных полов, либо строго ограничены только одним полом.
Мы ясно понимаем механизм половой дифференциации у птиц и млеко¬
питающих. Он представляет собой внутреннюю секрецию половых желез,
которые вырабатывают мужские или женские половые гормоны; возможно,
что другие железы внутренней секреции также дифференцируются по по¬
лу. Естественно заключить, что доля произошедших мутаций, которые за¬
трагивают любой из данных признаков, будет с самого начала ограничена
в своем проявлении полом, и если эффект этих мутаций возникает только
в сочетании с действием внутренних секретов половых желез у одного по¬
ла, их проявление будет отсрочено до взрослого состояния, как и другие
признаки половой зрелости. Эта доля может быть так же мала, как и доля
наблюдаемых генетических мутаций, и единственный повод считать, что
она будет выше, заключается в том, что возникновение резких изменений
можно ожидать в ответ на отклонения на ранних стадиях развития, тогда
как более мелкие изменения, которые главным образом и определяют про¬
гресс под действием Естественного Отбора, могут чаще запускаться на бо¬
лее поздних стадиях развития.Но какова бы ни была частота ограничения полом, можно с уверенно¬
стью заключить, что отбор, оказывающий воздействие только на один пол,
при полном отсутствии противоотбора у другого пола способен привести
к эволюционным изменениям, которые не будут слишком отличны у обо¬
их полов. Заметная половая дифференциация должна, с этой точки зрения,
объясняться условием, при котором силы отбора, действующие на оба по¬
ла, оказывают противоположное влияние. Если принять во внимание, что
у большинства видов оба пола отлично приспособлены к своему положе¬
нию в природе, эта ситуация может легко возникнуть в результате отбора
модификаций только у одного пола, поскольку модификации, если только
они не ограничены полом, будут сопровождаться изменениями у противо¬
положного пола, что, учитывая вышеуказанную адаптивность, будет нано¬
сить вред и потому не будет поддерживаться силами отбора. Если высокой
степени адаптации не существует, противоположная направленность отбора
у обоих полов должна быть случайной и редкой, и вовсе неясно, как, исходя
из вышесказанного, можно объяснить широкое распространение модифика¬
ций, ограниченных полом.Поскольку все множество генетических свидетельств поддерживает
и даже настаивает на той точке зрения, что все организмы, как прави¬
ло, отлично приспособлены к условиям своего местообитания, нам следу¬
ет рассматривать последствия только этого положения. Отбор, направлен¬
ный на особые качества только одного пола, будет в этом случае направлен
142Глава 6на изменение этого пола больше, чем другого. Противоположный пол будет
изменяться только таким образом, чтобы вовлечь в процесс силы, вынужда¬
ющие отбор идти в противоположном направлении и с равной интенсивно¬
стью. Преимущество наличия защитной окраски, как подчеркивал Уоллес,
имеет в этой связи очевидное значение. От этой точки, которую можно
достигнуть достаточно быстро, селективное преимущество далее мутации
в отношении рассматриваемой деятельности отбора вовсе не будет зависеть
от средней величины этих воздействий на оба пола, но только от разли¬
чия между этими воздействиями. Если мутация усилит половой контраст
и сделает оба пола менее похожими, она получит предпочтение со стороны
отбора и получит возможность внести свой вклад в создание половой диф¬
ференциации. Если, напротив, ее эффект будет направлен на то, чтобы сде¬
лать оба пола более похожими, она будет отвергнута отбором в ходе разви¬
тия. Следует заметить, что на этом более длительном эволюционном этапе
любой эффект, который могут оказывать новые мутации, действуя с рав¬
ной интенсивностью на оба пола, или, вернее, их средние эффекты на оба
пола, будет немедленно нейтрализован изменением частоты тех факторов,
которые, не будучи ограничены полом, оказывают влияние на развитие изу¬
чаемого органа.Кроме мутаций, эффект которых обусловлен секрецией половых же¬
лез, существует важная группа мутаций, влияющих на саму природу этих
секретов. При условии идущего у обоих полов противоположно направ¬
ленного отбора любая модификация этих секретов, которая, не мешая их
обычному действию, усиливает или увеличивает диапазон их воздействия
на развитие, будет дополнительным средством увеличения половой диффе-
ренцировки. Нет сомнения, что тем самым мы должны исключить предпо¬
ложение, что признак, вначале проявляющийся в равной степени у обоих
полов, должен под влиянием естественного отбора становиться в дальней¬
шем ограниченным полом в своем проявлении.Естественный Отбор и соотношение половЗадачу влияния естественного отбора на соотношение полов можно
наиболее точно исследовать при помощи концепции репродуктивной цен¬
ности, обсуждавшейся в главе II. Как хорошо известно, Дарвин оставил эту
задачу на будущее, считая ее решение слишком сложным для современни¬
ков делом (Descent of Man, p. 399).Насколько мы можем судить, ни в одном случае унаследованная тенденция
производить оба пола в равных количествах или один из полов в избытке не может
приносить прямую пользу или вред, большие для одних индивидов, чем для других;
Половое размножение и половой отбор143например, индивид, склонный производить больше самцов, чем самок, не будет
иметь больше успеха в борьбе за жизнь, чем индивид с противоположной тенденци¬
ей, и поэтому тенденция такого рода не может быть приобретена путем естествен¬
ного отбора. Тем не менее существуют определенные животные (например, рыбы
и усоногие раки), у которых для оплодотворения одной самки необходимо нали¬
чие двух или более самцов, и, соответственно, самцы имеют большую численность,
но вовсе не ясно, как эта тенденция, направленная на производство самцов, могла
быть достигнута. Раньше я считал, что тенденция производить оба пола в равных
количествах выгодна виду и следует из естественного отбора, но сейчас вижу, что
вся проблема настолько сложна, что осмотрительнее оставить ее решение на будущее.У всех организмов потомки вступают в окружающий мир с опреде¬
ленным количеством биологического капитала, полученного от родителей.
Размеры его сильно отличаются у разных видов, но в целом, прежде чем
потомство способно вести независимое существование, практически у всех
животных родители затрачивают дополнительную пищу, а также время
и труд на то, чтобы дети получили преимущество в жизни. Давайте рас¬
смотрим репродуктивную ценность этих потомков в момент, когда эти за¬
траты родителей в их пользу только что закончились. Если мы рассмотрим
совокупность всех потомков этого поколения, то станет ясно, что общая
репродуктивная ценность самцов этой группы в точности равна общей ре¬
продуктивной ценности самок, поскольку каждый пол должен обеспечить
половину потомков всех будущих поколений вида. Из этого следует, что со¬
отношение полов под действием Естественного Отбора установится таким
образом, что суммарные затраты, которые родители вкладывают в буду¬
щее своего потомства каждого пола, будут равны, поскольку если бы это
было не так, и общие затраты на выращивание, например, самцов, были
бы меньше, чем общие затраты на выращивание самок, тогда, поскольку
общая репродуктивная ценность самцов равна общей репродуктивной цен¬
ности самок, из этого бы следовало, что родители, чьи врожденные тенден¬
ции способствовали бы избыточному рождению самцов, являлись бы, осу¬
ществляя те же затраты, производителями большей репродуктивной цен¬
ности и, как следствие, прародителями большей части будущих поколений,
чем родители, имеющие врожденную склонность к производству самок. Та¬
ким образом, отбор поднял бы соотношение полов до такого уровня, когда
затраты на самцов не стали бы равными затратам на самок. Если, как на¬
пример, у человека, смертность самцов в период родительских затрат была
бы больше, это сделало бы затраты на их выращивание более значитель¬
ными, поскольку на каждую сотню успешно произведенных самцов были
бы произведены такие затраты, которые распространялись бы не только
на эту сотню во время их зависимого развития, но и на определенное коли¬
144Глава 6чество других, которые погибли раньше, чем смогли воздать благодарность
за затраты. Средние затраты, таким образом, оказываются выше на выра¬
щивание каждого мальчика, но меньше на каждого рожденного мальчика,
чем на девочку на соответствующих стадиях, и мы, таким образом, можем
сделать вывод, что положение, на которое будет направлено действие Есте¬
ственного Отбора, будет состоять в том, чтобы мальчики рождались в боль¬
шем количестве, но уступали в численности из-за более высокой смертно¬
сти до конца периода родительских затрат. Действительное соотношение
полов у человека выполняет эти условия с большой степенью точности,
особенно если сделать поправку на большое сокращение смертности среди
младенцев и детей, и поскольку эта поправка является следствием весьма
значительного неравенства в соотношении полов при зачатии, чему не уда¬
ется дать объяснения, a priori трудно не сделать вывод, что соотношение
полов действительно было достигнуто таким способом.Соотношение полов в конце периода затрат зависит, таким образом,
от дифференциальной смертности в этот период и от дифференциальных
потребностей (если таковые существуют), которые предъявляют дети это¬
го вида в течение этого периода зависимости; на это отношение не бу¬
дет влиять дифференциальная смертность во время периода самостоятель¬
ной жизни; относительные численности полов, достигающих зрелости, мо¬
гут, таким образом, зависеть необратимо от дифференциальной смертности
во время периода, находящегося между периодом зависимости и достиже¬
нием зрелости. Любая большая дифференциальная смертность в этот пе¬
риод будет, однако, сдерживаться Естественным Отбором вследствие того,
что общая репродуктивная ценность каждого пола в этот период абсолютно
равна, и менее многочисленный пол будет более ценным; соответственно,
более интенсивный отбор будет действовать в поддержку всех модифика¬
ций, направленных на его сохранение. Численность тех, кто достигает по¬
ловой зрелости, может, таким образом, оказаться разной, если возникнут
другие благоприятные условия для половой дифференциации по признакам
формы или габитуса, но любое значительное и сохраняющееся различие
между полами в зрелости должно сопровождаться половой дифференци-
ровкой, имеющей явную экологическую ценность.РезюмеСледствие полового размножения, которое, как мне кажется, имеет
фундаментальное значение для теории эволюции, состоит в том, что пре¬
имущества от полезных изменений в различных структурных элементах за¬
родышевой плазмы могут использоваться независимо друг от друга, тогда
Половое размножение и половой отбор145как в случаях с бесполыми организмами либо генетическое единообразие
всей группы препятствует эволюционному прогрессу, либо, при наличии за¬
метного генетического разнообразия, многие полезные изменения теряют¬
ся, поскольку происходят у индивидов, обреченных остаться без потомства.
В результате размножающийся половым путем организм реагирует на лю¬
бое воздействие намного быстрее, и если поместить его в равные условия
со сходным во всех отношениях бесполым организмом, то в условиях кон¬
куренции последний будет, без сомнения, заменен первым.Для того чтобы извлечь максимальную выгоду из возможных полез¬
ных мутаций, частота мутаций должна быть настолько мала, чтобы в боль¬
шем числе локусов гомологичные гены у всего вида были бы практически
полностью идентичными, что и наблюдается у высших организмов. Виды
с половым размножением не являются случайными таксономическими еди¬
ницами, как могло бы быть в случае бесполого размножения, но связаны
вместе общностью происхождения, что становится видно, если взглянуть
всего на сотню поколений назад. В основном, внутривидовая дисперсия
является результатом возникновения различий на протяжении жизни срав¬
нительно малого числа поколений недавнего прошлого, не считая различий
между географическими расами, которые возникают в результате изоляции.Отбор, оказывая разное воздействие на различные популяции вида
(независимо от того, дифференцированны ли эти популяции географиче¬
ски), будет стимулировать возникновение различий между ними при появ¬
лении измененного гена или комбинации генов. Этот процесс совершенно
необязательно приводит к нарушению единства вида. Элемент нестабиль¬
ности будет, однако, присутствовать в подобных случаях благодаря генети¬
ческим изменениям, влияющим на частоту обмена зародышевым матери¬
алом с обеих сторон, что под влиянием достаточно интенсивного отбора
будет приводить к дивергенции видов даже в случае отсутствия географи¬
ческих или иных барьеров при скрещивании.Важное средство дивергенции вида, особенно применимое к высшим
животным, состоит в возможности дифференцированного полового отве¬
та на ухаживание обладающих различными признаками претендентов. Об¬
стоятельства, благоприятствующие дивергенции видов, которые были бы
приспособлены к различным средам обитания, будут также способствовать
развитию как различий, так и полового предпочтения.Основной постулат дарвиновской теории полового отбора, а именно
демонстрация полового предпочтения, будет, таким образом, подтверждать¬
ся действием предшествующего отбора. Мы можем отметить, что зачатки
эстетического чувства могут встречаться у представителей целых классов,
независимо от того, стимулирует или нет наличие этого чувства эволюцион¬
146Глава 6ное изменение. У видов, где репродуктивный успех одного пола в большой
степени зависит от того, добьется ли он благосклонности у другого пола,
как, вероятно, обстоит дело у многих полигамных птиц, половой отбор бу¬
дет проявляться путем усиления интенсивности предпочтения, продуктом
которого он является, в результате чего та черта, которой было отдано пред¬
почтение, также как и интенсивность предпочтения, будут увеличиваться
вместе с постоянно возрастающей скоростью, вызывая глубокие и быстрые
эволюционные изменения некоторых заметных признаков до тех пор, по¬
ка процесс не будет остановлен прямым или опосредованным действиями
Естественного Отбора.Рассмотрение механизма действия половых гормонов, в большой
степени контролирующих вторичные половые признаки млекопитающих
и птиц, показывает, что половая дифференцировка может увеличиваться
или уменьшаться под действием Естественного Отбора либо за счет воз¬
никновения мутаций, действие которых ограничено одним полом, либо пу¬
тем модификации механизма действия гормонов. Таким образом, вполне
возможно, что мутантная форма, проявляющаяся вначале в равной степени
у обоих полов, может позже под действием отбора стать принадлежностью
только одного пола. Вопрос численного соотношения полов в половозрелом
состоянии не имеет такого значения для полового отбора, как считалось
ранее, по крайней мере для тех видов, число брачных пар у которых огра¬
ничено размером территории. Показано, что действие Естественного Отбо¬
ра направлено на уравнивание родительских затрат, отданных на рождение
и выращивание потомства обоих полов; в то же время понимание ситуаций,
связанных с территориальными условиями, возможно, подскажет нам еще
не один способ, благодаря которому половое предпочтение дает значимое
преимущество при размножении.
Глава 7
МимикрияСвязь теории мимикрии с исходной теорией Естественного Отбора. Теории
Бейтса и Мюллера. Предполагаемые статистические ограничения теории Мюллера.
Наблюдения как основа теории мимикрии. Эволюция неприятного вкуса. Теория
сальтаций. Стабильность отношения генных частот. Резюме.Если цель наших исследований состоит в выяснении того, как жи¬
вые существа стали такими, какие они есть... Она не будет достигнута
до тех пор, пока особенности процесса отбора не будут изучены так же
подробно и полно, как и материал, который подвергается отбору. Пауль-
ТОН, 1908.Существует три причины, по которым теория мимикрии крайне важ¬
на для каждого, изучающего Естественный Отбор. Во-первых, огромное
количество поразительных фактов благодаря ей становятся объяснимыми
с позиций одного единственного принципа; этот принцип состоит в селек¬
тивном действии конкретного фактора, обычно хищничества, в окружаю¬
щей насекомого среде на один набор признаков, таких как окраска, узоры,
поведение в покое, тип полета, наблюдаемых у самого насекомого. При на¬
шем невежестве относительно жизни диких существ в природе мы редко
задумываемся, как делаем это в данном случае, о действии какого-либо
фактора среды либо об особых достоинствах какого-либо из наблюдаемых
нами морфологических или физиологических признаков.Во-вторых, как было указано на стр. 54, особенно важно иметь воз¬
можность продемонстрировать адаптивное значение видовых признаков
для любой обширной группы случаев. Поскольку если это и соответству¬
ет действительности, на чем настаивал Робсон, даже детальное морфоло¬
гическое исследование различий между близкородственными видами, как
правило, не способно раскрыть нам их адаптивное значение в природных
местообитаниях. Однако не вызывает сомнения тот факт, что изучение ми¬
микрии на множестве примеров выявило особый фактор среды, которым
следует объяснить развитие отдельных признаков, различимых у относи¬
тельно близких видов, подвидов и даже у локальных разновидностей.
148Глава 7В третьих, с исторической точки зрения интересным представляется
тот факт, что теория мимикрии как величайшее последарвиновское прило¬
жение теории Естественного Отбора сыграла особенно важную роль в кон¬
це девятнадцатого и начале двадцатого веков (когда накопленные в музеях
и лабораториях биологические материалы начали вызывать сомнения в пра¬
вильности выводов полевых натуралистов раннего периода), призывая об¬
ращать особое внимание на важность экологических наблюдений для объ¬
яснения биологического материала, собранного в больших музеях мира.Вполне понятно, как это следует из доводов, приведенных в главе 1,
что представление автора о согласованности открытий в области генети¬
ки и, в частности, менделевской схемы наследования, с теорией эволю¬
ции очень отличается от того, как представляли себе этот вопрос пионе¬
ры менделизма, и даже противоречит их представлениям. Во времена пе-
реоткрытия работы Менделя они находились в основном русле движения
эволюционной мысли, которая в 90-х годах прошлого века остановилась
на прерывистом происхождении видовых форм. Так, казалось вполне есте¬
ственным, что дискретные элементы менделизма следует без достаточно
критичной проверки рассматривать как основание для убедительного дока¬
зательства в пользу этого взгляда. Пожалуй, нигде в биологии это движение
мысли не сталкивалось с такими серьезными противоречиями мнений, как
при объяснении миметического сходства.Теории Бейтса и МюллераТеория Бейтса, выдвинутая в 1861 году, не содержит ничего, что бы ни
следовало из определения мимикрии, а именно: что некоторые съедобные
формы, которые Бейтс наблюдал в особенности среди бабочек, подверга¬
ясь атаке со стороны насекомоядных птиц, занимают выгодное им положе¬
ние, при котором их принимают за другие объекты, особенно за менее съе¬
добные формы, на которые они несколько похожи, и что селективное пре¬
имущество, распространяющееся на индивидов, обладающих наибольшим
сходством, приводит к тому, что эти виды становятся все более точными ко¬
пиями других видов, обитающих в этом же местообитании, сравнительно
защищенных от нападения и являющихся поэтому подходящими моделя¬
ми. Эта теория полагается на ошибки, но ошибки восприятия, а не ошибки
рассудительного расчета. Выбор пищи птицей может быть полностью де¬
терминирован инстинктом в латентном состоянии в яйце или может быть
результатом опыта. В последнем случае мы не знаем, сколько этапов было
пройдено, прежде чем был накоплен опыт, и знаем только конечный этап,
когда обучение завершилось и были окончательно сформированы правиль¬
Мимикрия149ные понятия относительно пищи. Глаз все еще можно обмануть; смысл тео¬
рии Бейтса состоит в том, что глаз поддается обману, но ошибкам расчета
в его теории места нет.Теория Бейтса не объясняет, до какой степени расчет хищника закреп¬
лен наследственно или может изменяться в результате приобретенного опыта.
Если расчет хищника наследственный, он должен быть сформирован Есте¬
ственным Отбором как система полезных для вида пищевых предпочте¬
ний и не может продолжать применяться в течение длительного времени
к тем конкретным ситуациям, где потеря из-за того, что были отвергнуты
имитаторы, в действительности перевешивает преимущество от того, что бы¬
ли отвергнуты модели. Те же результаты, только быстрее, следуют, если осно¬
ву расчета составляет сформированный индивидуальный опыт. Необходимо,
таким образом, допустить, что имитаторы должны быть достаточно редки
или модели достаточно отвратительны для того, чтобы хищнику было выгод¬
но отвергать модели вместе с определенной долей имитаторов-обманщиков
в тех случаях, когда имеет место дискриминационный отказ или, напротив,
следует допустить, что наблюдаемое положение вещей — это переходное
состояние, требующее от инстинктов хищника более совершенной адапта¬
ции. Таким образом, вывод, сделанный Бейтсом и широко принятый дру¬
гими исследователями, заключается в том, что вид-имитатор должен быть
достаточно редким, получающим выгоду благодаря сходству с хорошо за¬
щищенным и многочисленным видом, населяющим ту же территорию.Ограничение подобного рода, но несколько иной специфики, определя¬
ется реакцией модели. Сходство, которое благоприятствует имитатору, бу¬
дет по той же причине вредным для модели. Особь вида-модели может по¬
страдать от того, что будет принята за имитатора, и, поскольку вероятность
этого события минимальна в том случае, когда минимально сходство, отбор
будет направлен в сторону изменения модели таким образом, чтобы сделать
ее максимально отличной от имитатора. Эта ситуация не слишком отличает¬
ся от той, которая лежат в основе развития предупреждающей окраски, по¬
скольку быть признанным невкусным — это эквивалент того, чтобы не быть
перепутанным со съедобным видом. Бейтсовская мимикрия, таким обра¬
зом, может быть достигнута, если модификация модели происходит мед¬
леннее, чем модификация имитатора, и, говоря о преобладающих факторах
ситуации, это означает, что селективное преимущество, которым облада¬
ет индивид-имитатор, превышает селективный вред, который испытывает
индивид-модель. Большая численность полезна как для успешного претво¬
рения в жизнь этого условия, так и удовлетворения аппетитов хищника.Известное Бейтсу наблюдение большого внешнего сходства между
очень распространенными и приблизительно одинаково защищенными ви¬
150Глава 7дами дало Мюллеру возможность спустя 18 лет сформулировать теорию,
получившую название мюллеровской теории мимикрии, в отличие от бейт-
совской теории мимикрии.43 Термин «мимикрия» все еще используется
в данном случае, хотя, как уточнил профессор Паультон, он должен от¬
носиться лишь к теории Бейтса. Теория Мюллера в большей степени,
нежели теория Бейтса, опирается на рассмотрение экологической ситуации,
при которой в действительности происходит уничтожение бабочек птица¬
ми. Мюллер указывает, что, во всяком случае, молодые птицы действитель¬
но учатся на собственном опыте; и во время этого процесса самообучения
тому, чем стоит и чем не стоит питаться, общий урон, который несут два
несъедобных вида, будет сокращаться и в конце концов уменьшится в два
раза, если постепенно сходство между ними станет так велико, что урок
избегать подобной пищи, полученный от одного вида, будет равно при¬
меним к другому. На более широкий смысл понятия «обучение» в этом
высказывании указал Свиннертон в 1915 году, когда, основываясь на де¬
тальных наблюдениях, подкрепленных обширными экспериментами по пи¬
щевым предпочтениям насекомоядных птиц, он заявил, что из-за некоторых
недостатков зрительной памяти процесс обучения у птиц длится всю жизнь
и происходит в ходе встреч с отпугивающими типами насекомых, не все¬
гда завершающихся нападением на них, и что чем больше численность тех,
кто демонстрирует данную отпугивающую окраску, тем больше укрепля¬
ется память и, как следствие, совершается меньше ошибочных нападений.
Бейтсовский имитатор получает выгоду за счет хищника, которого он об¬
манывает, и за счет модели, чья жизнь подвергается опасности. В теории
Мюллера оба вида в равной степени являются и имитатором, и моделью,
каждый получает одинаковую выгоду и оба сотрудничают, чтобы создать
преимущество хищнику путем облегчения ему процесса обучения. Хищ¬
ник, которому требуется расстроить коварные планы бейтсовского имита¬
тора, должен выработать исключительную разборчивость, тогда как в мюл¬
леровской ситуации его выгода будет связана с умением обобщать и де¬
лать выводы, исходя из сравнения. Для того чтобы возникла ситуация, опи¬
санная в теории Мюллера, хищник должен стремиться к сотрудничеству
с обоими типами добычи, что будет полезно всем трем участникам ситу¬
ации, в то время как, следуя теории Бейтса, и модель, и хищник должны
стремиться уменьшить выгоду имитатора, первый участник путем прямого
уменьшения сходства с имитатором, второй — путем изменения вкусовых
предпочтений и совершенствования способности отличать одно от другого.
Последняя тенденция, хотя и уменьшает выгоду от любой степени сход¬
ства, будет тем не менее стремиться к увеличению той степени сходства,
которая уже была достигнута.
Мимикрия151Предполагаемое статистическое ограничение теории
МюллераБыло замечено, что реакции модели и хищника накладывают статисти¬
ческое ограничение на теорию Бейтса, главный видимый фактор которой
состоит в относительном обилии видов. Было высказано предположение,
и это предположение неоднократно повторялось, что схожее ограничение
свойственно также и теории Мюллера. В 1908 году Маршалл предполо¬
жил, что по арифметическим причинам из двух равно несъедобных видов,
населяющих одну и ту же территорию, вид с меньшей численностью бу¬
дет стремиться приобрести сходство с более многочисленным видом, то¬
гда как вид с большей численностью не будет развиваться в этом же на¬
правлении. Маршалл не говорил, что вид с большей численностью будет
стремиться уменьшить сходство. Основной смысл его работы заключался
в том, чтобы подчеркнуть, что бейтсовский фактор сходства является про¬
тивоположностью мюллеровского фактора, и что главные причины этого
явления выходят за рамки обсуждения. Его статистическая аргументация,
частое повторение которой придает его позиции особый вес, будет рассмот¬
рена на страницах этой книги как явно относящаяся к тому типу, который
можно объяснить при помощи математических рассуждений.Когда дискуссия не затрагивает проблемы степени съедобности, разли¬
чие между бейтсовским и мюллеровским факторами абсолютно очевидно.
Если, однако, принять во внимание, что бабочки могут иметь самую раз¬
ную степень съедобности, и что хищник может игнорировать их или прини¬
мать в качестве пищи в зависимости от своего аппетита, возникает некото¬
рая опасность, что определенность эволюционных тенденций, выделенных
этими двумя авторами, может быть потеряна среди сложностей реальных
биологических обстоятельств. Именно так обычно и случается при обсуж¬
дении отдельных аспектов Естественного Отбора. Направление эволюции
воспринимается интуитивно и выражается в терминах, которые упрощают
и тем самым неизбежно фальсифицируют реальные биологические обстоя¬
тельства. В каждом примере единственная реальность, стоящая за подобны¬
ми абстрактными теориями, — это все множество определенных событий,
которые могут происходить время от времени с особями вида в течение их
жизни.Для того чтобы показать, как можно систематизировать эти случаи, да¬
вайте представим три вида, населяющие одну и ту же территорию, где А —
совсем несъедобен, у В вкус несколько лучше, а С не имеет неприятных ка¬
честв. Тогда все возможные ситуации могут быть представлены следующим
образом (I, II, III, IV), где, однако, альтернативные причины для наличия
152Глава 7любой ситуации (а, b и с) никоим образом не являются исчерпывающими,
но должны дополняться детальными биологическими наблюдениями.I. А, В и С все могут подвергаться атаке.II. В и С могут подвергаться атаке, А — нет.(a) Птица, которая либо в результате наследственного, либо приобре¬
тенного знания отдает предпочтение В перед А, готова атаковать объект,
распознаваемый как В, и не готова атаковать объект, распознаваемый как А.(b) Птица атакует А и находит его несъедобным, но не обладает еще
достаточной памятью о малосъедобных свойствах В.III. А и С могут подвергаться атаке, В — нет.(c) Птица атакует В и находит его несъедобным, но еще не обладает
достаточной памятью о несъедобных качествах А.IV. Может подвергаться атаке только С.В ситуации I миметическое сходство не вызвало бы никакого эффек¬
та, тогда как в ситуации IV получил бы преимущество С, если бы был
принят за А или В, а А и В проиграли бы, если бы были приняты за С.
Таким образом, если предположить, что первый эффект превышает второй,
С мог бы стать бэйтсовским имитатором А или В, тогда как в противопо¬
ложность этой ситуации А и В выиграли бы в результате подчеркивания
своей отличительной окраски, если бы сумели таким образом уменьшить
опасность быть принятыми за С. Сходным образом в ситуации II проигры¬
вает А, будучи принят за В, а В выигрывает, будучи принят за А, тогда как
в ситуации III все происходит наоборот; таким образом, может показаться,
что на А и В оказывают воздействие противоположные силы, одна направ¬
лена на увеличение сходства, другая — на усиление различия. И только
в случае, когда ситуации анализируются с учетом предполагаемых причин,
становится возможным указать получающийся в результате эффект.С этой целью мы выделяем бейтсовскую ситуацию 11(a) и мюллеров¬
ские ситуации 11(b) и Ш(с), признавая, что этой классификации не обяза¬
на быть исчерпывающей. Тогда видно, что бейтсовские ситуации должны
отличаться (1) зависимостью от различий в съедобности и (2) созданием
«бейтсовской тенденции» для В стремиться к А и для А удаляться от В. То¬
гда как мюллеровские ситуации (Ь) и (с) не зависят от какого бы то ни было
различия в съедобности, но, как полагают, возникают всякий раз, когда оба
вида рассматриваются менее предпочитаемыми, чем некая альтернативная
пища.В случае ситуации И, когда птица будет атаковать одну форму, но не бу¬
дет атаковать другую, различия между действием мюллеровского и бэйт-
совского факторов могли бы быть замечены только таким наблюдателем,
Мимикрия153который бы знал, было ли суждение птицы, предположительно правиль¬
ное, основано на достаточном или недостаточном опыте. Если опыт был
бы достаточен, птица никогда не отдала бы предпочтение А перед В (ситуа¬
ция III), соответственно, возникли бы только бейтсовские ситуации, но если
опыт птицы недостаточен, то мы должны признать, что это может приве¬
сти к ситуации III, и что даже когда возникает ситуация II, то ситуация
все же будет мюллеровской. В качестве бейтсовского агента птица всегда
права в своем выборе, в качестве мюллеровского агента она может быть
права, но из-за своей неопытности она не имеет права быть правой. Если
птица, по аналогии с человеком, имеет лишь частично удовлетворитель¬
ные основания для своего выбора, она может во время одного и того же
акта быть и бейтсовским, и мюллеровским агентом. И только в статистиче¬
ских совокупностях, образованных всеми случаями двух соответствующих
типов, эти две эволюционные тенденции видны абсолютно четко. Даль¬
нейшие исследования и размышления, посвященные привычкам диких жи¬
вотных, могут стать основанием для открытия других тенденций в эво¬
люции.На основании приведенного выше анализа мы не можем, однако, с уве¬
ренностью сказать, что суммарный эффект (Ь) и (с) будет состоять в том,
чтобы вызвать обоюдное, хотя, вероятно, и не равновеликое, сближение
между двумя видами, как это сделал Мюллер. По утверждению Маршалла,
в случае, когда виды равно несъедобны, вид с меньшей численностью будет
привлекателен для более многочисленного вида, но более многочисленный
не будет привлекателен для вида меньшей численности. Маршалл не мог
не вывести из этого заключения очень важное следствие, поскольку, как
он указывал, его позиция вела к неизбежному выводу, что когда миметиче¬
ское сходство будет достигнуто, более многочисленный вид получит пусть
и меньшую, но все же долю выгоды от ассоциации, на основании чего
можно возразить, что более многочисленный вид будет вынужден прий¬
ти к этому более выгодному состоянию. Напрашивается вывод, что это
соображение не может считаться обоснованным, пока не будет показано
существование непрерывного перехода от первого состояния ко второму,
причем такого, где преимущество растет при переходе на каждом новом
шаге. Подобное ограничение, если оно необходимо, наложило бы на се¬
лекционера ответственность подробной демонстрации явления, на которую
его оппонент мог бы ответить возрастающими до бесконечности требова¬
ниями объяснения мельчайших деталей. Даже если случай мюллеровской
мимикрии не столь уж важен сам по себе, было бы важно более деталь¬
но исследовать этот особый случай, где соображение о конечной выгоде
приводит, как считается, к ошибочному выводу.
154Глава 7Довод Маршалла состоит в следующем: если А и В одинаково невкус¬
ные виды, из которых В менее многочисленный, то в этом случае при отсут¬
ствии ошибок вследствие сходства двух видов молодые птицы будут нано¬
сить больший урон В, чем А, прежде чем они выучат урок; соответственно,
любой мутант А, похожий на В, будет страдать больше, чем немутантный
тип, и, таким образом, будет уничтожен. Можно предвидеть, что этот му¬
тант должен будет потерять все преимущество предупреждающей окрас¬
ки А и взамен получить только меньшее преимущество предупреждающей
окраски В, и этот довод является, несомненно, убедительным, поскольку
способен показать, что мутация, которая совершит прыжок и окажется вне
сферы защитного влияния своего типа, жестоко пострадает за свою по¬
спешность, даже если, что достаточно маловероятно, в результате этого
прыжка она окажется в самом центре защитного влияния менее многочис¬
ленного отпугивающего вида. Но что произойдет, если мутация окажется
менее значительной? Можно ли получить какое-то преимущество, напоми¬
ная В, и не теряя при этом всей выгоды от сходства с А? Возможно ли,
кроме того, что возросшая защита от отпугивающей окраски В будет бо¬
лее чем уравнивать потерю от меньшей защиты отпугивающей окраски А?
В своем ответе на соображения Маршалла Дикси выдвигает прямо противо¬
положное предположение, а именно: что мутант, обладающий внешностью,
промежуточной между А и В, получит полное преимущество от сходства
с тем и другим. В действительности, поскольку Маршалл полагает, что все
преимущество от сходства с А теряется прежде, чем появляются преиму¬
щества от возникновения сходства с В, Дикси полагает, что наоборот, все
преимущество от сходства с В может быть достигнуто прежде, чем утратит¬
ся любое преимущество от сходства с А. Оба предположения — крайности,
в действительности ни одно не может выполняться, и поскольку оба пред¬
положения ведут к противоположным выводам, может показаться, что мы
имеем дело с балансом сил неизвестной величины и не можем утверждать,
что принцип Мюллера будет работать, как и не можем утверждать обратное.Остается довод, на который полагался Мюллер, а именно: что посколь¬
ку окончательное положение, при котором достигается близкое сходство,
выгодно обоим видам; каждый, таким образом, будет стремиться к дости¬
жению этого преимущества. Маршалл оспаривает законность этого дово¬
да, приводя веский аргумент в ответ на вывод из конкретного случая; как
мы видим, однако, возражение Маршалла в выбранном примере неубеди¬
тельно, и общее соображение относительно преимущества окончательного
состояния подтверждает этот довод. Если, однако, довод является убеди¬
тельным, должна существовать возможность придумать такую форму ар¬
гументации, которая бы доказательно показала на основании принятых по¬
Мимикрия155стулатов, что признанная мюллеровская ситуация действительно способна
создать мюллеровскую эволюционную тенденцию, влияющую на оба упо¬
мянутых вида.Я предлагаю основывать подобную аргументацию на сравнении судь¬
бы любого отклонения от типа А не со средними показателями этого ти¬
па, а с равнозаметными, но противоположными отклонениями. Если до¬
пустить, что вариации вида А, вследствие ли мутаций или по причине
менделевских рекомбинаций, будут встречаться с одинаковой частотой как
в направлении В, так и в противоположном направлении, мы можем без¬
ошибочно считать, что подобные вариации происходят попарно, включая
вариации, имеющие равную величину, но направленные в противополож¬
ные стороны.Поскольку они обладают равной величиной, они проиграют (если про¬
играют) равным образом, если не будут приняты за типичный вид А, но ес¬
ли любой из них в отдельности или оба сразу будут приняты за вид В, боль¬
шее преимущество, без сомнения, получит вариация, направленная в сто¬
рону В. Поскольку вид в целом можно рассматривать, как сумму подобных
пар вариаций и поскольку в каждой паре отбор благоприятствует вариации,
усиливающей сходство с В, если вообще благоприятствует этой вариации, об¬
щая результирующая должна являться модификацией в направлении вида В.Можно видеть, что условием существования тенденции к подражатель¬
ному сходству является то, что в определенной доле случаев, когда особи
вида А подвержены нападению, а особи вида В защищены от нападения,
особи вида А должны, благодаря своему сходству с особями В, в реальных
ситуациях избегать нападения. Это заключение несколько отлично от за¬
ключения, сделанного профессором Тернером в его приложении к работе
Паультона Memoir on Mimicry in the Butterflies of Fiji, 1924, который говорит
о действительном наложении вариаций двух видов как условии эффектив¬
ного выполнения мюллеровского статистического аргумента. Возможность
ошибки со стороны хищника представляется существенной частью теории
мимикрии, и трактовка Тернера включает это допущение при условии, что
мы рассматриваем его кривые распределения как образец не объективной
изменчивости вида, а изменчивости (возможно, гораздо более значитель¬
ной) субъективных предпочтений хищника под влиянием, как это часто
случается, невнимания или поспешности, или из-за обманчивого или недо¬
статочного освещения, — в действительности, из-за любых обстоятельств,
провоцирующих ошибку человека или птицы. Удивительно, что на таком
далеком от прямого доказательства обстоятельстве, как субъективное вос¬
приятие птиц, у нас появятся три причины для использования приближен¬
но нормального распределения: (а) причины, по которым это распреде¬
156Глава 7ление выбирается, как «нормальный закон ошибок», едва ли могут быть
ограничены собственно особенностями человека; (б) объективная измен¬
чивость измеряемого признака, либо из-за менделевской сегрегации, либо
из-за флуктуаций среды, обычно приблизительно нормальна, и (в) результат
двух независимых распределений неизбежно будет ближе к нормальности,
чем один, а возможно и оба его компонента.Аргумент, приведенный выше, может быть, несомненно, опроверг¬
нут, если мы откажемся принять какой-либо из биологических факторов,
названных в дискуссии. Если доказать, что в действительности никогда
не возникают ситуации, когда особь вида А уцелеет, если ее перепутают
с особью вида В, но погибнет, если ее не перепутают, что хищник не учит¬
ся на опыте, и никакое сходство не оказывает на него воздействия, или что
вариации А, которые благоприятствуют развитию сходства, не являются на¬
следственными, тогда теория мимикрии Мюллера как объяснение наблюда¬
емого сходства окажется несостоятельной. Единственное соображение, вы¬
веденное из вышеприведенного рассуждения, состоит в том, что если допу¬
стить возможность существования этих биологических факторов, то резуль¬
тирующая эволюционная тенденция не может быть отнесена к виду с мень¬
шей численностью. Эффективность мюллеровского отбора ceteris paribus
будет, несомненно, больше у малочисленного вида, но мнимое возражение
статистического характера против мюллеровского тяготения малочисленно¬
го вида (или группы) к многочисленному виду совершенно необоснованно.Наблюдения как основа теории мимикрииХотя попытка провести даже самое общее исследование связанных
с мимикрией биологических фактов явно выходит за рамки этой книги,
представляется необходимым дать более точное заключение о разнообра¬
зии результатов наблюдений, которые позволили бы предложить теорию
в качестве единственного удовлетворительного объяснения.Число рассматриваемых видов, даже если речь идет только о насекомых,
настолько велико, что в большинстве отдельных случаев многие классы
нужных наблюдений отсутствуют, и вероятное объяснение должно, таким
образом, основываться на неполных данных о более известных случаях.Биологические данные, на которых базируются убедительные приме¬
ры, были успешно разработаны Паультоном, труд и вдохновение которого
являются основой огромного числа накопленных фактов, и чьи доказатель¬
ства я суммирую на этих страницах.(1) Миметическое сходство несет в себе признак адаптации во множе¬
стве одновременных модификаций, результатом которых оно является. Эта
Мимикрия157черта, как было отмечено в главе 2, не может просто служить примером,
поскольку смысл адаптации заключается в ее сложности. Миметическое
сходство включает цвет, узор, форму, позу, движение и иногда также звук,
и многие из этих черт, если их проанализировать, являются очень слож¬
ными сами по себе. Естественный Отбор — это единственное известное
в биологии средство, благодаря которому возможны комплексные адапта¬
ции структуры к функции.(2) Помимо адаптации, миметическое сходство несет в себе еще одно
свойство Естественного Отбора, разнообразные способы которого служат
достижению одной и той же цели. Этот признак, аналогичный оппорту¬
низму в человеческих структурах, заслуживает, как мне кажется, больше¬
го внимания, чем ему уделяют в большинстве случаев. Механизмы живых
тел кажутся построенными нашим создателем в меньшей, чем можно бы
было ожидать, степени на основе простых и эффективных механических,
физических или химических принципов. В противоположность этому каж¬
дое свойство поведения материи, каким бы странным и чуждым оно ни
казалось, подхватывается сразу же, как только ему удастся произвести же¬
лаемый эффект. В случае мимикрии Паультон указывает на четыре ясных
способа, используемых различными видами для имитации внешнего вида
муравья, и находит в одной миметической группе пять различных путей
создания подобия прозрачности у крыльев ночных и дневных бабочек.В дополнение к этим двум характерным для Естественного Отбора
чертам в общем случае для выполнения особых требований, поставленных
теорией мимикрии, мы можем выделить три класса фактов.(3) Миметическое сходство не может объясняться систематической
близостью. Поразительные случаи сходства наблюдаются между различны¬
ми классами животных. Уиллер описывает случай тактильной мимикрии,
когда клещ играет на инстинктах определенных слепых муравьев, имити¬
руя своими передними лапками движения антенн, посредством которых
муравьи получают пищу друг от друга. Очевидно, чем в более близком
родстве состоят демонстрирующие сходство организмы, тем чаще гомо¬
логичные части используются для его совершенствования, и тем сложнее
показать, что поверхностное сходство было навязано или предотвратило
изначальную дивергенцию во внешности.(4) Миметическое сходство не сопровождается такими дополнительны¬
ми чертами сходства, которые бы не помогали создать или усилить внешнее
подобие.(5) Миметическое сходство встречается у видов, населяющих один
и тот же регион, появляющихся в одни и те же времена года и в одни
и те же часы и часто летающих вместе.
158Глава 7Следует добавить, что для установления миметической природы сход¬
ства нет необходимости иметь в наличии все пять классов признаков. Дока¬
зательства из класса (1) сами по себе могут быть достаточно сильны, что¬
бы исключить все возможные сомнения. Хотя доказательства из групп (3),(4) и (5) весьма ценны как независимое дополнение, наличие ни одного
из них не обязательно для всех миметических примеров. Важность фак¬
тов, собранных в группу (2), состоит не в их ценности как доказательстве
отдельных случаев, но в сравнительном и систематическом исследовании
мимикрии как явления в целом.Иллюстрации I и II из примеров, выбранных профессором Паультоном,
служат примерами типов поверхностного сходства, формирующих теорию,
основанную на наблюдениях. Хотя этого достаточно для иллюстрации неко¬
торых очевидных признаков, в этих исследованиях особенно важно знание
самого живого организма и его экологических связей.Гораздо более сложной проблемой является объяснение наблюдаемого
сходства эволюционными тенденциями Бейтса и Мюллера или обеими од¬
новременно, как в случае вышеупомянутых паразитов муравьев. Хотя мюл¬
леровский отбор, в отличие от бэйтсовского, вызывает взаимное сближе¬
ние двух видов, мы едва ли можем ожидать, что их модификации будут,
за исключением некоторого небольшого числа случаев, достаточно близки,
чтобы их можно было бы заметить при сравнении с родственными видами.
В большинстве случаев модификация будет проявляться на филогенетиче¬
ском уровне только у тех видов, которые претерпели наиболее быстрые
модификации. Создается впечатление, что применение критерия съедобно¬
сти вызывает еще большие трудности, поскольку, хотя случай, когда два
вида более или менее одинаково съедобны или несъедобны, практически
исключает принцип Бэйтса, демонстрация подобного равенства с точно¬
стью, позволяющей исключить дифференцированное отношение хищника
к жертве, а также с учетом существующей популяции хищника требует как
природных знаний, так и тонких экспериментов, которые недоступны нам
в настоящее время. Возможно, отсутствием надежных методов различения
видов и можно объяснить тот факт, что основное внимание было уделено
наблюдениям относительного обилия, что, хотя и имеет близкое отношение
к проблеме, не является само по себе достаточным для проведения оконча¬
тельного тестирования. Бейтсовская мимикрия более многочисленной фор¬
мой менее многочисленной формы не может быть исключена как невозмож¬
ная по чисто теоретическим соображениям, поскольку, если модель отлича¬
лась бы исключительно отталкивающими свойствами, или ее имитатор был
бы не особенно ценным источником пищи, мотив для того, чтобы избегать
его, не слишком бы уменьшился при увеличении численности имитатора.
Мимикрия159Более того, когда этот аргумент используется для исключения бейтсовского
принципа, следует учитывать, что это обилие имеет отношение не столь¬
ко к сбору энтомологического материала, сколько к отбору под действи¬
ем неизвестного вида, привычки и время питания которого, следовательно,
также неизвестны. Равным образом демонстрируемые отличия в съедобно¬
сти не могут, как было показано, служить для исключения мюллеровского
фактора, возможности которого проявляются в том случае, когда сходные
предупредительные рисунки приобретает группа нескольких разных и от¬
носительно отдаленных видов в одном и том же регионе. Различие между
типами сходства, достигаемого под действием бейтсовских и мюллеров¬
ских факторов, можно представить теоретически в случаях, когда группа
хищников сталкивается только с одним или другим типом сходства. По¬
скольку бейтсовское сходство может ввести в заблуждение, оно должно
распространяться на каждый видимый признак, по крайней мере таким об¬
разом, чтобы не оставалось ни одного заметного различающегося призна¬
ка, по которому имитатора можно бы было отличить от оригинала. Этого
не требуется в случае отпугивающей окраски, приобретаемой в результате
мюллеровского отбора, поскольку в этом случае бывает достаточно, чтобы
единственный заметный признак давал основание хищнику отнести дан¬
ный объект к разряду несъедобных. Таким образом, различающиеся между
собой насекомые могут пользоваться преимуществом мюллеровского отбо¬
ра при условии, что они демонстрируют какой-то один общий заметный
признак, тогда как при бейтсовском отборе имитаторы должны демонстри¬
ровать по меньшей мере некоторое сходство со своими моделями по всем
признакам.Эволюция неприятного вкусаВажным вопросом в теориях мимикрии Бейтса и Мюллера является
процесс эволюции неприятного вкуса как средства защиты. Большинство
других средств защиты, таких как наличие жала или отталкивающих сек¬
ретов и запахов, можно считать способами, повышающими возможность
выживания отдельного индивида, у которого они наиболее развиты, или
всего сообщества, к которому этот индивид принадлежит. Что же касается
неприятного вкуса, то он, хотя и обеспечивает очевидное средство защиты
вида в целом посредством его действия на врожденные или приобретенные
реакции хищника, все же, поскольку любая подвергшаяся нападению особь
обречена на гибель, трудно заметить, как индивидуальное добавление этого
качества сверх среднего характерного для вида уровня может дать отдель¬
ной особи преимущество.
160Глава 7Привычка определенных личинок образовывать скопления дает нам
возможное решение проблемы, если мы готовы допустить, что несъедоб¬
ность взрослой особи, чья предостерегающая окраска столь хорошо при¬
способлена для демонстрации, сама по себе является просто побочным
продуктом присутствующих в организме неприятно пахнущих веществ,
приобретенных благодаря защите, полученной на личиночной стадии. Ес¬
ли говорить о взрослом насекомом, то эффект усиления неприятного вкуса
на действия хищника будет состоять в том, чтобы заставить этого отдельно¬
го хищника избегать всех членов группы, на которую он охотится; однако,
если только индивиду, на которого было совершено нападение, не удаст¬
ся выжить, это преимущество не будет передано через его генотип, тогда
как в случае скученно живущих личинок эффект, безусловно, будет состо¬
ять в том, чтобы дать большую защиту, особенно одной конкретной группе
личинок, вероятно, братьям и сестрам особи, на которую было совершено
нападение. Эффективность отбора в результате того, что хищник не тро¬
нет братьев жертвы, будет, конечно, в два раза меньше, чем было бы в том
случае, если бы была защищена сама жертва, хотя против этого можно вы¬
двинуть возражение, что вся эта ситуация применима в целом к возможно
весьма многочисленному выводку. Таким образом, хотя реальная эффектив¬
ность данной формы отбора несомненна, можно тем не менее усомнить¬
ся в том, что все случаи несъедобности насекомых могут быть объяснены
только этим принципом.Профессор Паультон сообщил мне, что насекомые с неприятным вку¬
сом и предостерегающей окраской, включая бабочек, имеют такие упругие
и гибкие тела, что они легко могут пережить пробное нападение без серьез¬
ного повреждения. Без этого авторитетного мнения я бы не осмелился пред¬
положить, что неприятный вкус жидкости тела мог бы эволюционировать
в результате различной степени выживания переживших нападение особей.
Эффект отбора у личинок, живущих скоплениями, хотя и не исключаю¬
щий индивидуального отбора взрослых насекомых, дает нам альтернативу,
которая, несомненно, может быть полезной для большого числа случаев.Привычка многих птиц иметь четко очерченные территории питания
во время сезона размножения дает возможность распространить эффект,
оказанный на личинок, живущих скоплениями, на другие случаи, когда ли¬
чинки, хотя и не настолько скапливаются на одном растении, все же распро¬
странены на территории, которая обычно находится в пределах территории
питания одной пары птиц. Селективный эффект в подобных случаях будет
размыт, поскольку личинки из других кладок, оказавшиеся на той же терри¬
тории, могут разделить преимущества или недостатки, возникшие в резуль¬
тате высокой или низкой степени развития неприятного вкуса любых ли¬
Мимикрия161чинок, подвергшихся нападению на данной территории. Такая размытость,
несомненно, замедлит эволюцию неприятного вкуса, скорость которой, ес¬
ли упростить задачу, предположив, что число случайно выбранных кладок
разделяет общую судьбу, будет обратно пропорциональна числу кладок;
размытость селективного эффекта не следует, однако, путать с процессом,
действующим в противоположном направлении и способным остановить
его. В этой связи стоит остановиться на повадках при откладывании яиц,
свойственных некоторым ночным бабочкам.Точка зрения, что неприятный вкус, как правило, приобретается пу¬
тем отбора, действующего на живущих поблизости друг от друга родствен¬
ных личинок, подразумевает, что свойство жить скученно или какое-то по¬
добное свойство были когда-то принадлежностью вида, который в насто¬
ящее время обладает несъедобным вкусом, хотя и не подразумевает, что
виды с неприятным вкусом и даже с личинками, имеющими предостерега¬
ющую окраску, должны сохранять свойство жить скученно. Это обуслов-
ленно преимуществами указанного свойства, среди которых можно выде¬
лить (1) меньшую открытость самки во время откладывания яиц и (2) —
в случае неприятного вкуса и предостерегающей окраски преимущество
лучшей защиты от хищников, не всегда уравновешивающими недостатки,
которые следуют из возможной нехватки запасов пищи. Мы, можем, одна¬
ко, справедливо заметить, что если скученность или равнозначные свойства
являются необходимыми условиями для развития защитного вкуса, и если
обладание таким вкусом дает дополнительное преимущество как для ску¬
ченности, так и для заметной окраски, тогда в природе должна быть вы¬
ражена связь между заметной окраской личинки и привычкой откладывать
всю кладку или большую ее часть на одно растение или на соседние расте¬
ния. Подробный обзор биологических фактов по этому вопросу — предмет
насущной необходимости. Я благодарен профессору Паультону и доктору
Иммсу за следующие примеры.Лунка серебристая (Zygaera bucephala) несъедобна на всех стадиях
развития — гусеницы, куколки и бабочки. Гусеницы имеют яркую черно¬
желтую окраску и образуют большие скопления, куколки коричневого цве¬
та, находятся в земле, хотя также и несъедобны, а бабочка — прекрасный
пример защитной окраски. Поскольку неприятный вкус имеется только
на личиночной стадии, вполне возможно, что он развился благодаря от¬
бору именно на этой стадии, чему, несомненно, благоприятствовало свой¬
ство гусениц образовывать скопления. Ночная бабочка {Mania typica) также
обладает неприятным вкусом на всех стадиях развития, у этой бабочки гу¬
сеницы не обладают предостерегающей окраской, но питаются в раннем
возрасте группами на нижней стороне листьев.
162Глава 7Мохнатые довольно яркие гусеницы Onethocampa processioned живут
в общих сетях. Когда они съедают всю листву, находящуюся внутри об¬
щей сети, они выходят большими «процессиями» и ищут место для но¬
вой сети. Повзрослев, они выходят в сумерках небольшими группами кор¬
миться и возвращаются перед рассветом под общую сеть; кажется, что они
инстинктивно избегают одиночества, как будто бы в этом для них кроет¬
ся опасность. Эти гусеницы демонстрируют ряд особенностей — наличие
общей сети, страх дневного света и одиночества,— каждую из которых
можно воспринимать как своеобразную меру защиты от различных хищ¬
ников.Яркоокрашенные гусеницы дневных и ночных бабочек, как правило,
собираются в большие массы, хотя не всегда. Исключением является пяде¬
ница крыжовниковая Abraxas grossulariata, гусеницы которой питаются раз¬
дельно, впрочем, яйца откладываются группами, и на одном или несколь¬
ких соседних кустах обычно присутствует множество гусениц. В свою оче¬
редь, Anosia plexippus рассеивает свои яйца на большой территории, хотя
имеет одиночно живущих, несъедобных, яркоокрашенных гусениц. Это, ве¬
роятно, свойство большинства данаид.Похоже, что истинное свойство образовывать большие скопления ха¬
рактеризует неприятные на вкус виды, которые летают по большой терри¬
тории, тогда как в случае более медлительных самок гусеницы оказываются
рассеяны, однако на небольшие расстояния.Показательный случай — дневной павлиний глаз и крапивница, Vanes¬
sa, а также родственные им роды. Это — быстро летающие бабочки, и мож¬
но было бы предположить, что их гусеницы окажутся рассеяны по большой
территории. У собственно рода Vanessa гусеницы яркоокрашены, и яйца
откладываются группами, тогда как у чертополоховки Cynthia cardui и ад¬
мирала Pyrameis atalanta, относимых ранее к Vanessa, личинки частично
скрытые и яйца откладываются одиночно.Личинки многих хризомелид образуют скопления, и по крайней мере
некоторые из них явно обладают очень неприятным вкусом.О личинках пилильщика Камерон говорит, что «те из них, кто выделяет
секреты или неприятные запахи, образуют массовые скопления по несколь¬
ко особей, кормящихся на одном листе, часто образуя ряд, и выставляя
свои тела в воздух. Они почти всегда имеют яркую окраску». Преимуще¬
ство образования скоплений для усиления этих средств защиты очевид¬
но: их можно рассматривать, как дальнейшее развитие пассивной защиты,
обеспечивающейся неприятным вкусом, и кажется, что именно на началь¬
ных стадиях этого развития свойство образовывать скопления обеспечивает
условие для эффективного отбора.
Мимикрия163В общем, можно сказать, что полученные в результате наблюдений
в этой области факты согласуются между собой и доказывают, что отталки¬
вающий вкус обеспечивает условия для эволюционного развития предосте¬
регающей окраски и, вероятно, также и предупреждающих запахов; усло¬
вием эволюционного развития отталкивающих вкусов, равным образом ха¬
рактеризующих любую стадию развития животного, должны считаться ску¬
ченность или родство личинок одного и того же выводка и, следовательно,
корреляция их генетической конституции. Впрочем, для того чтобы утвер¬
ждать, что объяснить эволюцию свойства малосъедобности как средства
защиты можно только так, а не иначе, автор должен располагать значитель¬
но более широким знакомством с полученными из наблюдений фактами,
чем он располагает в настоящее время.Выведенный в этом разделе принцип селективного преимущества, ко¬
торый получает целая группа родственников за счет индивидуальных ка¬
честв одного из членов группы, не имеющего никакого личного селектив¬
ного преимущества, аналогичен ситуации, которая возникает в людских со¬
обществах на племенном уровне организации при отборе группы качеств,
суммарно именуемых героизмом. Идеал героизма, развитый среди таких
людей, значительно выходит за рамки максимального личного преимуще¬
ства, и его эволюция может быть объяснена исходя из известных причин
тем достоинством, которое оно несет в виде престижа и высокой репута¬
ции потомков героя. Ситуация применительно к людям, более подробно
рассматриваемая в главе 11, несомненно, осложнена наличием полового
отбора, существование которого в случае эволюции малосъедобности у на¬
секомых никак не подтверждается.Теория сальтацийВ своей книге Mimicry in Butterflies, вышедшей в 1915 году, Пённет
повторяет аргумент Маршалла и уверенно заявляет, что исключает мюлле¬
ровскую мимикрию вида меньшей численности видом с большей числен¬
ностью. На первый взгляд этот довод кажется несовместимым с основным
заявлением Пённета о неспособности Естественного Отбора производить
адаптации, поскольку очевидно, что, в отличие от Маршалла, он должен
был бы также отвергнуть и бэйтсовскую мимикрию, равно как и мюллеров¬
скую мимикрию для многочисленного вида, которому подражает вид мень¬
шей численности. Впрочем, было бы несправедливо считать цитирование
Пённетом довода Маршалла просто случайным дополнением к его критике,
аналогично тому, как, например, вызвав подозрение о возможности ошиб¬
ки, пусть и в постороннем вопросе, удается обеспечить вынесение вердикта
164Глава 7по основному пункту; напротив, довод Маршалла играет хотя и неболь¬
шую, но существенную роль в его разрушительном аргументе, основанном
на «кругах» мимикрии. Для иллюстрации этого сложного вопроса выбран
случай двух предположительно съедобных самок, каждая из которых со¬
всем не похожа на соответствующего ей самца, каждый из которых в свою
очередь не похож на другого. Две самки демонстрируют явное миметиче¬
ское сходство с тремя другими бабочками, две рассматриваются как явно
несъедобные, тогда как несъедобность третьей вызывает сомнение. Если
допустить, что немиметические самцы имеют внешность, сходную с той,
что имели две миметические самки, возникает вопрос, как выяснилось, что
последние оказались похожи на несъедобных членов «круга». Если допу¬
стить, что эти модели могли быть когда-то сходны по внешности с одним
из самцов, то вряд ли можно допустить, что они когда-либо напоминали
обоих либо одновременно, либо последовательно, но если такого сходства
ранее не существовало, то градуальная эволюция мимикрии исключается,
и мы вынуждены признать, что миметические самки возникли как случай¬
ности или сальтации абсолютно иначе, чем их матери.Видно, что аргумент Пённета по этому важному вопросу требует, что¬
бы градуальное и взаимное сближение двух или более предостерегающих
окрасок было бы полностью исключено, поскольку, если допустить воз¬
можность такого процесса, трудность воображения непрерывной последо¬
вательности изменений полностью исчезает, тогда как принятие прерыви¬
стости, напротив, становится бременем для теории, включая определенную
невероятность получения хорошего сходства за один раз. Соответственно,
аргумент Маршалла, который Пённет воспринял как довод, восстанавлива¬
ющий все ограничения бэйтсовской ситуации, играет важную роль в до¬
воде, высказанном в защиту сальтаций, такую существенную, что кажется
невозможным заделать брешь, возникшую после его изъятия.Довод в пользу сальтаций, как было представлено Пённетом, по свое¬
му характеру не является полностью негативным или разрушительным, он
связан с одним недавно открытым весьма интересным фактом, а именно:
что различия между тремя формами триморфной самки Papilio polytes мо¬
гут быть обусловлены двумя менделевскими факторами, видимое проявле¬
ние которых ограничено женским полом, и один из них предположительно
необходим для проявления второго.Этот факт важен как указание на механизм, посредством которого
поддерживается собственно полиморфизм, он показывает, что полимор¬
физм в этом случае и, вероятно, в сходных случаях зависит от одного
или нескольких менделевских факторов, функция которых состоит в пе¬
реключении на одну или другую из возможных альтернатив подобно тому,
Мимикрия165как более распространенный половой диморфизм также зависит от мен-
делевского механизма. В некоторых группах, как например, у Drosophila
и человека, вся хромосома участвует в процессе определения пола, тогда
как у некоторых рыб, напротив, было обнаружено, что кроссинговер проис¬
ходит между «половыми» хромосомами у гетерогаметного пола, либо муж¬
ского, как у Lebistes, либо женского, как у Platypoecilus; у рыб, как оказы¬
вается, нам следует скорее говорить о гене пола, а не о половой хромосоме
как агенте определения пола.44 Переход из одного состояния в другое путем
прекращения кроссинговера не представляет непреодолимых сложностей,
поскольку известны менделевские факторы, которые ускоряют или ингиби¬
руют кроссинговер. Причина такого изменения не столь очевидна, но по¬
скольку обе системы все еще используются, вполне вероятно, что каждая
из них при определенных условиях имеет свои собственные преимущества.В основе довода Пённета, объясняющего возникновение форм-ими¬
таторов сальтациями, лежит менделевская обусловленность полиморфных
самок, поскольку считается, что менделевские факторы должны были из¬
начально возникнуть как мутации, и поскольку доказано, что различные
формы отличаются друг от друга только по одному фактору, делается вы¬
вод, что каждый из этих типов должен был возникнуть в результате одного
скачка. Каким бы убедительным ни казался на первый взгляд этот довод,
мы должны тем не менее признать нашу неправоту, если до этого были
его защитниками, поскольку половое различие у рыбы Lebistes, вероятно,
определяется единственным фактором; следовательно, женская особь это¬
го рода с соответствующими ее полу адаптациями возникла как результат
одной сальтации из самца того же вида! Или vice versa. В этом случае мы
даже свободны от необходимости отрицать предполагаемую сальтацию как
невероятную, поскольку, коль скоро для воспроизводства вида требуется
сотрудничество обоих полов, мы можем быть уверены, что происхождение
полового фактора предвосхищает эволюцию отдельных полов и предполо¬
жительно сохраняет свою роль переключателя, не меняясь на протяжении
всего хода эволюционного развития этих двух типов.Пример пола придает особое значение тому факту, который, по мере
того как генетические знания все чаще используются для решения сложных
прикладных задач, обретает все больший смысл; хотя именно менделевский
фактор решает, какую из альтернатив выбрать, но каждая из альтернатив
может сама по себе претерпевать изменение в ходе эволюционного разви¬
тия, в результате чего морфологический контраст, который определяется
фактором на поздней стадии развития, может сильно отличаться от того
положения, которое существовало в момент его появления. Таким обра¬
зом, вывод о том, что поскольку различие между миметической и похожей
166Глава 7на самца формой P. polytes определяется одним единственным фактором,
то, следовательно, миметическая форма возникла полностью развитой в ре¬
зультате одной мутации, будет едва ли справедлив; скорее, можно высказать
ничем не обоснованное предположение, что с тех пор, как впервые возник
диморфизм, ни в одной из двух альтернативных форм не происходило ни¬
каких эволюционных изменений.Ряд генетических экспериментов показал, что генетические изменения,
подобные рассмотренным выше, совместимы с чисто менделевской схемой
наследования. У крыс капюшонный рисунок (черные пятна на белой голо¬
ве) является рецессивным по отношению к сплошной окраске, что, вероят¬
но, является случаем, аналогичным узору Dutch у кроликов и recessive pied
у мышей. При изучении вариаций в капюшонном рисунке Кастл обнару¬
жил, что путем отбора было несложно получить линии капюшонных крыс,
которые были бы почти полностью черными, и другие линии — практиче¬
ски полностью белые, и также, конечно, большое количество стабильных
рисунков переходного типа. Все эти типы «капюшонной окраски» вели себя
как до отбора, просто как рецессивные по отношению к сплошной окраске.
Было предложено два вероятных объяснения: первое состояло в том, что
полученная в ходе отбора модификация, зависящая от гена капюшонности,
была в действительности выделена отбором из большого числа близких ал¬
леломорфов с малой и большой пигментацией, и, соответственно, что со¬
хранившиеся гены капюшонности были отличны от генов, превалирующих
до начала отбора. Второе объяснение состояло в том, что ген капюшон¬
ности не менялся, но что пигментация участка определялась совместным
влиянием других генов, так называемых модифицирующих факторов, и что
изменение в рисунке капюшона — это результат отбора среди альтернатив¬
ных вариантов, представленных этими модификаторами, типов, у которых
произошло развитие больших или меньших пигментированных участков
соответственно. Решающий эксперимент был призван дать ответ на во¬
прос, какое из объяснений верно. Крысы обеих отселектированных линий
были возвратно скрещены с исходной, не подвергающейся отбору линией
со сплошной окраской; их потомков скрестили между собой, и в следую¬
щем поколении вновь проявился капюшонный рисунок. Если модификация
признака связана с геном капюшонности, крысы с восстановленной капю¬
шонной окраской получили бы полностью модифицированные гены капю¬
шонности и должны были бы быть такими же темными или такими же
светлыми, как линия капюшонных крыс, из которой они были получены.
Если же ответственны были другие факторы, капюшонные внуки получили
бы их в равной степени от бабушек и дедушек как со сплошной, так и с ка¬
пюшонной окраской и должны бы были отличаться соответственно менее
Мимикрия167выраженной темной или менее выраженной светлой окраской по сравне¬
нию с отселектированными линиями. Вторая альтернатива оказалась вер¬
ной, модификация легко передавалась крысами со сплошной окраской, ко¬
торые не несли гена капюшонности. Тогда стало возможным сделать вывод,
что ген не затрагивается отбором, но на его проявление можно оказывать
воздействие, очевидно, в любой степени посредством отбора модифициру¬
ющих факторов.Если допустить, что эти аналогии не являются ошибочными, мы долж¬
ны предположить, что оба фактора Н и R, которые, как обнаружил Фраер,
определяют различия между полиморфными формами P. polytes, возникают
в результате мутаций, и что возникшие новые гены сохраняются неизмен¬
ными с момента их появления. С другой стороны, мы не должны иметь
никаких оснований с генетических позиций верить, что комбинация ННгг
с начала своего появления близко напоминала современную форму polites
или являлась действительным имитатором P. aristolochiae, а также что ком¬
бинация HHRR напоминала современную форму romulus или являлась дей¬
ствительным имитатором P. hector. Мы должны были бы ожидать посте¬
пенной эволюции такого миметического сходства, если модифицирующие
факторы, всегда широкодоступные, подвергались бы отбору птицами или
другими хищниками.Стабильность отношения генных частотСледует подчеркнуть, что приведенные аргументы не помогают объяс¬
нить сам полиморфизм. Это явление не настолько распространено, чтобы
предположить, что своим происхождением оно полностью обязано неким
особенным обстоятельствам, однако менделевский характер явления все же
предполагает один короткий шаг на пути к решению проблемы,45 а именно:
что лежащее в основе условие для его развития состоит в том, что пропор¬
циональные числа генов некоего менделевского фактора, имеющего весьма
заметный эффект, должны находиться в состоянии стабильного равновесия
подобно тому, как было показано в главе 5.Стабилизирующий отбор едва ли может быть чем-то иным, нежели46исключением, хотя в некоторых случаях мы вправе ожидать его влияния.
Бейтсовский имитатор, например, будет тем хуже защищен, чем больше
будет его численность по сравнению с численностью модели; диморфный
бейтсовский имитатор, таким образом, отрегулирует численности обеих
своих форм, если они зависят от одного менделевского фактора, до тех
пор, пока они не будут защищены в равной степени; любой рост численно¬
сти одной формы за счет другой уменьшит преимущество первой и увели¬
168Глава 7чит преимущество второй, создавая тем самым селективное действие, на¬
правленное на восстановление исходной пропорции. Имитатор, имеющий
преимущество только благодаря мюллеровской мимикрии, не должен быть
диморфен, если в ход не идут дополнительные механизмы стабильности,
поскольку во всех других случаях отбор приводит к нестабильному равно¬
весию, и соотношение будет уходить от него все дальше, пока один или
другой тип не будет элиминирован.Вторая форма стабилизирующего действия встречается в репродуктив¬
ном отборе. Стабильное соотношение полов явно объясняется этим случа¬
ем, как и соотношение коротко- и длинно столбчатой форм у примул. Ин¬
тересно отметить, что Фраер в своих экспериментах по скрещиванию, про¬
водимых с Papilio polytes, наблюдал множественные случаи бесплодных
пар, что означало, по его мнению, возможное существование «нелегетим-
ного» спаривания. Одной из простейших возможностей этого типа является
большая плодовитость гетерозиготного состояния по сравнению с гомози¬
готным.Следует, пожалуй, отметить, что работа Джерулда с доминантным при¬
знаком белой окраски, наблюдаемой у самок некоторых видов Colias, также
демонстрирует определенные особенности, предполагающие наличие ста¬
билизирующего механизма, управляющего отношением генных частот жел¬
той и белой окраски. Джерулд отмечает, что возникают большие сложности
при получении гомозиготных типов белой окраски, что, видимо, связано
с наличием тесно сцепленного летального фактора. Когда от расы, явно
свободной от летального фактора, было получено потомство чисто белой
окраски, неспособность самцов к спариванию привела к интродукции сам¬
цов дикой расы, которые, как обнаружилось, внесли летальный фактор. Тот
факт, что именно этот летальный фактор явно не редок в природе, хотя мы
вправе предположить, что он должен был исчезнуть достаточно быстро,
предполагает наличие стабилизирующей системы. Нельзя сказать, что пол¬
ностью объяснены генетические сложности, поскольку в результате опреде¬
ленных типов спаривания регулярно возникает ненормальное соотношение
полов (3 самки : 2 самца). Что касается модифицирующей роли отбора,
интересно, что Джерулд отмечает появление варьирующего исчезающего
желтоватого оттенка на крыльях генетически белых самок и объясняет эту
изменчивость вторичными факторами.Как белая форма Colias, так и две миметические формы polytes пред¬
ставлены только самками, и у обоих видов мутантная форма доминантна
по отношению к более старому аллелю, из которого она предположительно
возникала. Это обстоятельство в сочетании с вероятностью наличия в обо¬
их случаях стабильного соотношения генов предполагает, что мутантная
Мимикрия169форма пользуется селективным преимуществом и по этой причине преоб¬
ладает в гетерозиготе, селективный вред мутантного гена ограничен репро¬
дуктивным фактором генетической летальности или стерильности, связан¬
ными с мутантной гомозиготой. В случае миметических форм P. polytes их
селективное преимущество уже было отмечено изучающими мимикрию,
хотя вопрос, можно ли применить те же правила к белой форме Colias,
остается неясным. Если, однако, это предположение относительно причин
генетической стабильности оказалось бы верным, эти и аналогичные слу¬
чаи могли бы обеспечить беспрецедентную возможность реальной оценки
величины конкретной селективной силы в природе. Репродуктивные харак¬
теристики отбора, особенно если они связаны с абсолютной стерильностью
или летальностью, должны быть экспериментально показаны и измерены,
а их селективный эффект вычислен исходя их частот альтернативных ге¬
нов или комбинаций генов, наблюдаемых в природе. Тогда они могли бы
стать стандартом, пригодным для измерения, к которому можно приравнять
эффективность миметического отбора, сбалансированного в противополож¬
ном направлении с репродуктивными характеристиками, и даже оценить их
локальную вариацию. Значимость подобного прямого определения едва ли
нужно подчеркивать; препятствия свободному размножению этих групп по¬
являлись до сих пор просто как помеха, затрудняющая менделевский анализ
полиморфных форм; можно надеяться, что с учетом описанного выше под¬
хода их выявление может стать главным объектом будущих исследований.Какой бы ни была причина стабильности того или иного фактора, лю¬
бой вид, несущий фактор стабильности, будет потенциально диморфен,
и такое положение будет сохраняться, если только изменившиеся условия
не нарушат эту стабильность. Если при этом условии отбор будет благопри¬
ятствовать различным модификациям двух генотипов, ясно, что он может
стать приспособительно диморфным благодаря кумулятивному отбору мо¬
дифицирующих факторов без изменения монофакторного механизма, бла¬
годаря которому поддерживается диморфизм.РезюмеТеория мимикрии представляет особый интерес для изучающих Есте¬
ственный Отбор, поскольку она знакомит нас с примерами адаптивной роли
различий между видами и внутривидовыми формами, а также с противоре¬
чивыми взглядами пионеров менделизма и селекционизма.Теория Бейтса объясняет только ошибки в восприятии хищника, она
имеет ограничения, диктуемые эволюционными реакциями и хищника,
и модели. Теория Мюллера основана на ошибках в восприятии и суждении:
170Глава 7как хищник, так и два вида добычи подвергаются в этом случае действию
эволюционного процесса, благоприятствующего мюллеровской мимикрии.Возражение статистического характера, выдвинутое Маршаллом про¬
тив теории мимикрии Мюллера, было основано на ненадежных рассужде¬
ниях и при тщательном рассмотрении признано нереальным.Наблюдения, составляющие основу теории мимикрии, можно разде¬
лить на пять четких категорий, две из которых характерны для общего дей¬
ствия Естественного Отбора. Хотя четкие доказательства доступны только
для ничтожного меньшинства предполагаемых случаев, совпадение незави¬
симых групп наблюдений позволяет говорить о несомненной справедливо¬
сти наиболее исследованных примеров.Эволюция неприятного вкуса представляет особую проблему, решение
которой может быть найдено при рассмотрении того, насколько защищен¬
ными оказываются находящиеся в скоплении личинок особенно неприят¬
ные на вкус потомки того же выводка.Мюллеровский отбор, приводящий к постепенному и взаимному сбли¬
жению различной предостерегающей окраски, дает объяснение проблеме,
с которой столкнулся Пённет в связи с существованием миметических ко¬
лец. Его дальнейший довод о менделевском характере полиморфных ими¬
таторов, состоящий в том, что эти имитаторы возникают в результате еди¬
ничных сальтаций, не учитывает действия других факторов в модификации
форм, определяемых начальными мутациями.Менделевский полиморфизм в обычных условиях предусматривает на¬
личие стабильного отношения генных частот для определяющего фактора.
Если механизм, обеспечивающий стабильность, включает, как можно пред¬
положить на примере P. polytes и Colias, элемент стерильности или ле¬
тальности, который уравновешивается элементом экологического преиму¬
щества, то должна существовать возможность путем анализа механизма
стабильности оценить размеры селективного преимущества, которым поль¬
зуются в природе получившие предпочтение типы.
Иллюстрация I. Модели и их подражатели у насекомых Австралии (рис. 1-3) и тро¬
пической Америки (рис. 4-7)
Иллюстрация IВсе изображения насекомых даны в натуральную величину.Рис. 1. Abispa (Monerebia) ephippium Fab. — обычный представитель предоми-
нантиой группы австралийских ос, характеризующийся темным коричневато¬
оранжевым основным цветом, большой величиной и редуцированным числом
черных пятен. Им подражают многие другие насекомые из различных групп,
включая шмелей, мух, разноусых бабочек, а также многочисленных жуков. То,
что сходство может быть достигнуто довольно различными способами, проил¬
люстрировано на рис. 2 и 3.Рис. 2. Tragocerus formosus Pascoe — жук-дровосек. Почти весь его миметический
рисунок обусловлен окраской элитр (надкрылий), которые скрывают брюш¬
ко, совершенно не похожее на осиное, что показано на рис. 2а. Свободное
выдвижение крыльев происходит за счет изогнутой выемки на боку каждого
из надкрылий.4'Рис. 3. Esthesis ferrugineus Macleay. Еще один вид жуков-дровосеков, у которых ми¬
метический рисунок достигается формой брюшка, а надкрылья редуцированы
до маленьких закругленных пластинок, за счет чего крылья лежат открыто, в то
же время не скрывая осовидное брюшко.Рис. 4. Heliconius erato erato Linn. — обладающая предупреждающей окраской
и неприятным вкусом бабочка из тропической Америки, которой красиво под¬
ражает летающая днем хипсида, показанная ниже.Рис. 5. Pericopis phyleis Druce. Эта разноусая бабочка и ее булавоусая модель были
найдены в Перу. Поразительное миметическое сходство не распространилось
на усики, нитевидные и незаметные у модели.Рис. 6. Menthona confusa Butler — еще одна бабочка из тропической Америки, у ко¬
торой найден подражатель (рис. 7) в Парагвае. У этой бабочки и близких ей
видов антенны имеют яркую оранжевую булаву, и этот признак также присут¬
ствует у многих ее подражателей из различных групп булавоусых и разноусых
бабочек.Рис. 7. Castnia linus Cramer. Антенные булавы этой летающей днем бабочки, несмот¬
ря на заметное сходство с М. confusa, обладают формой, являющейся отличи¬
тельной чертой семейства Castniidae, но довольно отличной от той, что есть
у модели. Этот пример миметического сходства по обычно незаметному при¬
знаку здесь проявился исключительно ярко и может быть сравнен с рис. 1, 1 а—3,
За на иллюстрации II.Данные модель и подражатель также иллюстрируют, как видно на рис. 1-3,
различные способы, при помощи которых может быть достигнуто сходство. Про¬
зрачные участки крыльев модели образуются за счет сильной редукции размера
чешуек; у подражателя такой редукции не происходит, а эффект достигается за счет
прозрачности чешуек.
Иллюстрация II. Модели и их подражатели у насекомых Африканского (рис. 1-За)
и Восточного (рис. 4-6) регионов
Иллюстрация IIВсе изображения насекомых даны в натуральную величину.Рис. 1-3. Обычно незаметные у большинства бабочек пальпы у широко распро¬
страненной восточно-африканской акреины Acraea zetes acara Hew. окрашены
в оранжевый цвет и хорошо заметны на фоне темной головы, что видно спереди
(1), сверху (2) и сбоку (3).Рис. 1а-3а. Сходные признаки мы наблюдаем у нимфалиды Pseudoacraea boisduvali
trimenii Butler, подражающей виду A. z. acara и найденной в той же части Аф¬
рики. Очевидно, что сходство здесь происходит по исключительно акцентиро¬
ванному признаку подобно тому, что наблюдалось на американских примерах,
показанных на рис. 6 и 7 иллюстрации I.Рис. 4. Danaida tytia Gray — данаида из Восточного региона, имеющая предостере¬
гающую окраску и подражателя (рис. 5) из района Дарджилинга.Рис. 5. Papilio agestor Gray — парусник, подражающий D. tytia.Рис. 6. Neptis imitans Oberth. — нимфалида из Юго-Западного Китая, подражающая
местной географической форме D. tytia.Таким образом, две эти бабочки из совершенно неродственных групп в своем
подражании воспроизводят специфический рисунок данаиды.Представленные здесь булавоусые и разноусые бабочки иллюстрируют широ¬
ко распространенное мимикрирование под представителей тропических семейств,
имеющих наиболее неприятный вкус, — на иллюстрации I это Ithomiinae (рис. 6)
и Heliconinae (рис. 4) из Нового Света; на иллюстрации II — Danainae (рис. 5)
и Acraeinae (рис. 1-3) из Старого Света.
Глава 8
Человек и обществоО Человеке, важность предварительных исследований. Упадок цивилизаций.
Социологические взгляды. Сообщества насекомых. Резюме.... и если ее несчастные узники не могли решить или истолковать эти
загадки, она с невиданной жестокостью нападала на них, растерянных
и смущенных, и рвала на части. БЭКОН (Sphinx or Science)О Человеке48Предыдущие дискуссии о Человеке в связи с теорией эволюции были
главным образом посвящены утверждению двух положений. Первое состо¬
ит в том, что человек, как и другие животные, обязан своим происхожде¬
нием эволюционному процессу, управляемому законами природы; и вто¬
рое — в том, что умственные и моральные качества, свойственные всему
человечеству, по своей природе аналогичны умственным и моральным ка¬
чествам животных; а способ их наследования аналогичен способу насле¬
дования признаков тела человека и животных. Хотя может показаться, что
единство взглядов на органическую природу неизбежно подразумевает вы¬
вод второго заключения из первого, традиционное мнение человека о самом
себе представляет главную трудность на пути подобного единства взглядов
и вынуждает исследователей, способствующих окончательному принятию
выводов Дарвина по этому вопросу, следовать двумя путями, различаю¬
щимися предметом и методом, и нашедшими наиболее четкое выражение
в трудах Хаксли и Галтона. Сходство тела человека с телами других живот¬
ных было очевидно с древнейших времен. По самой грубой классификации
человек должен относиться к одному из видов млекопитающих. Огромное
количество исследований формы, развития и функций его тела полностью
согласуется с взглядом на человека как на обезьяну Старого света, ближай¬
шего родственника бесхвостых обезьян.Не было бы необходимым рассматривать этот предмет детально с эво¬
люционной точки зрения, если бы не было трудностей, связанных с изу¬
чением человеческого разума. Первая вытекает из того, что разум человека
172Глава 8развит гораздо больше, нежели у какого-либо другого животного; извест¬
ные нам факты дают полное обоснование для подобного сравнения. Вторая
трудность состоит в том, что мы обладаем внутренним взглядом или осо¬
знанием человеческого разума, находим в нем ценные и значимые для нас
качества, которые до тех пор, пока мы остаемся людьми, должно пред¬
ставляться нам самым важным из всего, что есть в органической природе.
Если бы не этот второй фактор, то я не могу себе представить, как простое
различие в степени развития могло бы привести к какой бы то ни было об¬
щеизвестной сложности в принятии научной точки зрения. Многие живот¬
ные демонстрируют исключительное развитие определенных органов, что
не приводит к неправильному пониманию их естественного родства. Хобот
слона, рассматриваемый просто как структурное усовершенствование, яв¬
ляется по меньшей мере таким же замечательным, как и мозг человека, хотя
нам он вовсе не кажется таким уж важным.К несчастью, для общего принятия подобного объективного взгляда
на все, что касается людей, как это необходимо для научного понимания
человеческих дел, часто кажется, что такой взгляд унижает человека, и осо¬
бенно, то, что человек считает своим высшим качеством, — как будто бы я
сказал, что мозг человека не более важен, чем хобот слона, или как будто
бы я сказал, что он не должен был значить для нас больше, если бы мы
и в самом деле были бы столь разумными, как нам следовало бы быть. Эти
заявления были бы излишне вызывающими и к тому же лишенными науч¬
ного смысла. Но если все-таки допустить хотя бы долю сомнения по этому
вопросу, который вовсе не касается науки, то я могу добавить, что как че¬
ловек я никогда не испытывал и тени сомнения относительно значимости
человеческой расы, ее умственных и моральных качеств, и в особенности
человеческого интеллекта, чести, любви, великодушия и праведности, где
бы эти замечательные качества ни встречались. Высшая ценность, которую,
как мне кажется, следует признать за этими сторонами человеческого пре¬
восходства, вряд ли дает повод утверждать, как это иногда делается с раз¬
драженным негодованием и не без примеси духовного высокомерия, что
такая низкая материя, как природная причинность, не может иметь ниче¬
го общего со столь возвышенными понятиями. Напротив, их высшая цен¬
ность представляет собой сильнейший мотив для того, чтобы стремиться
узнать как можно более точно и определенно, какими способами природ¬
ные причины действовали в своем эволюционном созидании и продолжают
действовать по сей день, чтобы сделать эти черты более или менее распро¬
страненными.Это резонное и, как мне кажется, очевидное соображение слишком
часто игнорировалось авторами, стоящими как по одну, так и по другую
Человек и общество173стороны спорного вопроса. Одни кажутся убежденными в том, что поме¬
щая моральные и эстетические оценки на исключительно научную основу,
можно совместить или даже заменить чувственную веру или предрассуд¬
ки разумным восприятием реальной действительности. Другие, чье мнение
можно скорее объяснить, чем извинить, стоят в обскурантистской оппо¬
зиции и питают, как может показаться, личную нелюбовь к высшим каче¬
ствам человеческого сознания, во всяком случае, из духа противоречия и ис¬
пытывают наслаждение, уподобляя поэтические и религиозные устремле¬
ния человечества разнообразным низменным склонностям, имеющим сугу¬
бо физиологическую основу. Создается впечатление, что студенты-медики,
узнавшие о влиянии половых гормонов на психику, нередко испытывают
недоверие к поэтическим эмоциям, отождествляемым остальным челове¬
чеством с физической любовью. И хотя эта точка зрения может быть от¬
брошена как крайность и признана проявлением незрелости, люди в целом
весьма неохотно расстаются с утвердившейся ранее догмой, что все, что
есть самого благородного в их природе, должно иметь причину и даже воз¬
никать в результате процессов, которые сами по себе обладают не меньшей
ценностью. Возможно, основу этой догмы, которая сама по себе кажется
непобедимой, можно отыскать в аристократической традиций, следуя кото¬
рой человек ценился как минимум столь же высоко за свое происхождение,
как и за свои личные качества. Какими бы ни были ее истоки, интеллекту¬
альные усилия, затраченные таким человеком, как Уоллес, для того, чтобы
не пытаться относить высшие человеческие качества на счет естествен¬
ных причин, должны предостеречь нас от этого источника ошибок, сила
которого могла бы, пожалуй, навсегда лишить человечество способности
поддерживать развитие всего, что оно больше всего ценит.Разумное и активное существо, как правило, считает себя оригиналь¬
ным явлением. Если оно станет рассматривать естественные причины, ко¬
торые привели к его существованию, его оценка окажется искаженной,
и интуиция покинет его. Легко понять, что различия в поведении, объясня¬
ются ли они сознательным решением или импульсивной реакцией, в дей¬
ствительности определяют различия в величине смертности и размноже¬
нии. Не можем мы также сомневаться и в том, что эти различия в поведении
вытекают из индивидуальных особенностей в структуре мозга, без чего все
люди действовали бы одинаково, и понятие о сознательном действии было
бы иллюзией. Справедливо и то, что мы принимаем во внимание обстоя¬
тельства и действуем или не действуем соответственно, но что особенно
важно, что именно мы взвешиваем обстоятельства и другие в подобном
случае, возможно, поведут себя по-иному. Каким бы очевидным ни казался
этот аспект сознательного действия, поразительным является тот факт, что
174Глава 8многие, для кого не составляет труда признать, что непроизвольные реак¬
ции человечества модифицируются в процессе отбора, все же испытывают
трудности в приложении того же аргумента к обстоятельствам, требую¬
щим сознательного выбора. Я подозреваю, что причина состоит в том, что
сознательный выбор открыт для модификации, наряду с другими обстоя¬
тельствами, посредством убеждения, и что важность эффектов, достигну¬
тых этим способом, подчас не позволяет нам почувствовать поразительные
контрасты в легкости, с которой убежденность достигается.Высший внутренний судья, определяющий наш выбор между доб¬
ром и злом, называется совестью. Мы не считаем нашу совесть открытой
для влияния извне, и с этой точки зрения она должна рассматриваться как
полностью врожденная. Тем не менее мы все же демонстрируем некоторую
неопределенность относительно степени, до которой мы признаем ее ин¬
дивидуальность. Конечно, ваша совесть не есть моя совесть, все же иногда
мы испытываем соблазн быть столь немилосердными, чтобы признать, что
если откинуть лицемерие, или приобретенную жесткость, или слепоту в от¬
ношении нашей лучшей сущности, то соответствовать нашим собственным
стандартам вполне могли бы и другие. С позиций отбора не имеет никакого
значения, рассматриваем ли мы человечество как набор людей с различной
совестью или просто с различными реакциями на их внутренний контроль.
Однако, поскольку каждый человек может говорить только о собственной
совести, я предпочту в дальнейшем придерживаться ранее высказанного
мнения.Пусть кому-то это и покажется очевидным, но я считаю необходи¬
мым настаивать на точке зрения, что систематическое положение челове¬
чества и демонстрируемое им наследование умственных и моральных ка¬
честв не исчерпывают эволюционного значения вида человек. Напротив,
это — только предварительные расследования, направленные на то, чтобы
понять, действительно ли человек, а если да, то человек во всех его про¬
явлениях, попадает в сферу действия естественной теории эволюции. Это
предваряет наш интерес к эволюционной истории и судьбе человечества,
к предмету, который, как мне кажется, все еще недостаточно исследован.
Животное, которое в течение своей сравнительно недолгой истории успе¬
ло пережить глубокие изменения в среде обитания, питании и характерной
позе, должно, пусть только по этим причинам, пользоваться достаточным
интересом эволюциониста. Если мы добавим к этому развитие социальной
организации, не имеющей параллелей в своем собственном классе, исполь¬
зование искусственно созданных устройств и средства для выражения и за¬
писи своего опыта и идей, то очевидно, что для стороннего наблюдателя
человечество могло бы явиться объектом крайне интересных эволюцион¬
Человек и общество175ных исследований и породило бы вопросы, на которые было бы сложно
ответить, пользуясь лишь сравнениями и аналогиями.Создается впечатление, что большинство авторов-генетиков занима¬
ют две противоположные, но одинаково неудовлетворительные позиции.
Меньшая их часть, как мне кажется, страшится, что чистота их предмета
как абстрактной науки, будет утрачена, если применить ее к виду, к которо¬
му принадлежат сами, и хотя они, быть может, и заинтересованы в практи¬
ческом улучшении культурных растений и домашних животных, но очень
осторожны, чтобы в случае всякого упоминания о генетике человека ди¬
станцироваться от любого предположения, что и она может иметь прак¬
тически важные последствия. Более многочисленная и более инициатив¬
ная школа, полностью пропитанная чувством возможности универсально¬
го применения генетических знаний, считает и человечество полем, при¬
годным для их применения. Ее последователи больше готовы и, пожалуй,
лучше подготовлены к тому, чтобы оценить сходство человеческой наслед¬
ственности с наследственностью Drosophila или кукурузы, нежели оценить
особые проблемы, которые представляет эволюция человека в обществе,
и предпочитают лишь бегло и поверхностно упоминать о проблеме чело¬
века в последней главе исчерпывающего и часто превосходного обзора со¬
временных достижений в области генетических исследований. Хотя гене¬
тические знания важны благодаря ясности, которую они привносят в пред¬
мет, причины происходящих эволюционных изменений могут быть решены
только путем обращения к социологическим и даже историческим фактам.
Эти знания должны быть доступны хотя бы для того, чтобы раскрыть более
мощные силы, работающие над модификацией человечества.Упадок цивилизацийУпадок и гибель цивилизаций, включая не только исторические при¬
меры греко-римской и исламской цивилизаций, но также и примеры пред¬
шествующих им доисторических цивилизаций, ставят перед социологом
исключительную и вполне конкретную задачу так остро, что само ее су¬
ществование, кажется, бросает вызов любому притязанию, на которое мы
осмеливаемся, — понять природу и функционирование человеческого об¬
щества.Если использовать термин цивилизация правильно, то его следует при¬
менять не просто к тем обществам, институтами которых мы видим осно¬
вание восхищаться, но обозначать им собрание всех социальных адапта¬
ций, соответствующих долговременному существованию высокой плотно¬
сти населения. В общей форме эти адаптации носят универсальный харак¬
176Глава 8тер. Во всех обществах, которые мы называем цивилизованными, личное
понимание, которое человек способен формировать с небольшим кругом
своих ближайших знакомых, дополняется много более обширной системой
условных соглашений, создающих основу его привычных отношений, его
прав и обязанностей перед обществом, членом которого он является. Он,
таким образом, может свободно посвятить себя производительному труду,
даже имеющему высокоспециализированный характер, полный уверенно¬
сти, что производство предметов первой необходимости, защита его соб¬
ственности от насилия и даже удовлетворение его моральных и интеллек¬
туальных потребностей будут составлять труд остальных членов общества,
которые сделали эти заботы предметом своих особых профессий. Специа¬
лизация по профессиям, включающая привычное принятие условного стан¬
дарта обмена, поддержание общественного порядка и национальная орга¬
низация военной подготовки представляют, таким образом, универсальные
характеристики цивилизованных человеческих обществ, в отличие от неци¬
вилизованных. Из опыта, который никто не думает отрицать, известно, что
подобная организация позволяет данной территории поддерживать гораздо
более многочисленное население, живущее на более высоком материаль¬
ном и интеллектуальном уровне, чем в случае нецивилизованных народов,
которые могли бы жить на той же территории.Как же можно вообразить, что подобное цивилизованное общество,
которое когда-то было организовано и отлажено, способно проиграть в кон¬
куренции со своими нецивилизованными соседями?49 Последние свободно
расселены по всей своей территории и не имеют организованного централь¬
ного правительства, которое могло бы мобилизовать малочисленное населе¬
ние. Напротив, как показывает наш опыт знакомства с нецивилизованными
народами, они разделены кровной враждой и мелкой завистью, что должно
сделать их союз, даже в случае защиты элементарных национальных ин¬
тересов, почти невозможным. Индустриальная организация дает цивилизо¬
ванным народам как в древности, так и в настоящее время преимущество
владения лучшим оружием, тогда как обычай совместного труда позволяет
им выбрать более упорядоченную, скоординированную и эффективную си¬
стему военной тактики. Кроме того, высшее знание, которое цивилизован¬
ные народы могут постепенно накапливать, что в действительности и про¬
исходит, должно, как правило, давать им возможность действовать на ос¬
новании более обширной информации, с более уверенным предвидением
последствий своих коллективных действий и с возможностью выигрывать
за счет опыта, совершенствуя свои методы, если первая предпринятая ими
попытка не принесет успеха. Помня о несомненных преимуществах выс¬
шего знания, координированных усилий и индустриальных достижений,
Человек и общество177разве не должны мы с уверенностью предсказать, если мы не знакомы
с фактическим ходом истории нашей планеты, что история цивилизации
должна состоять из непрерывной серии успехов, и что, как только зародыш
организованного общества возник однажды в Вавилоне или, быть может,
в Египте, только время может решить, когда каждая страна в мире окажет¬
ся, в свою очередь, частью организованной вавилонской или египетской
цивилизации?Трудности, относящиеся к более ранним цивилизациям, наряду с преж¬
ним описанием какого-либо исторического периода, значительно отлича¬
ются от рационального предвидения, рассмотренного выше. Мы со всей
очевидностью наблюдаем определенную повторяемость многих элементов
традиционной цивилизации, передаваемых от одной социальной группы
другой по мере развития цивилизации. Однако данное обстоятельство ни¬
коим образом не затушевывает различий между социальными группами,
включающих различия в местности, языке, религии, расе, которые связаны
с высшими цивилизациями различных эпох. В случае любых значитель¬
ных перемен эти различия, несомненно, связаны с насильственным втор¬
жением нового народа нецивилизованного происхождения. Эксперименты
по превращению общества в цивилизованное повторялись неоднократно
народами, относящимися к разным расам, и почти всегда не без помощи
традиционных представлений народов, достигших более высокого уровня
цивилизации, но развитие их национальных и промышленных объединений
осуществлялось при помощи их собственных прогрессивных сил; во всех
случаях без исключения, если мы оставим в стороне не завершенной как
эксперимент нашу собственную цивилизацию, после периода славы и до¬
минирования, сопровождаемого значительным вкладом в развитие науки
и искусства, следует период, когда народы не только не могут сохранить
своего превосходства, но даже достичь среднего уровня развития всех на¬
родов на земле, зачастую не оставляя каких-либо свидетельств своего су¬
ществования, кроме тех, что мы имеем благодаря усилиям археологов.Прежде чем обсуждать причины упадка цивилизаций, которые я наме¬
рен рассматривать и в отношении феномена в целом, и в отношении его
более ярких особенностей в последующих главах о Человеке, необходи¬
мо уделить некоторое внимание предварительному вопросу: каким должно
быть объяснение, которое мы бы могли считать адекватным? У народов
древности несложно найти обычаи, неприятные для нашего современно¬
го сознания, легко критиковать их идеи в области образования или форму
правления, которой они успешно пользовались; а самое главное, легко най¬
ти дефекты в их незнании экономических законов и приписать их ошибкам
в этой области те же гражданские и политические неудачи, которые мы
178Глава 8равным образом ожидаем от таких же ошибок наших политических оп¬
понентов! Подобные аргументы не только не убедительны вследствие на¬
шего незнания законов причинной связи, на которые они опираются, они
также явно недостаточны, чтобы отвечать требованиям нашей проблемы.
Ибо, во-первых, наше знание ранних этапов истории великих народов де¬
монстрирует нам обычаи, не более отталкивающие, манеры, не более рас¬
пущенные, пренебрежение образованием как минимум такого же уровня,
незнание экономических законов, столь же полное, как и у их более циви¬
лизованных последователей. Во-вторых, кроме того, в пору национального
упадка было бы неразумно ожидать, что какая-либо сторона национальной
жизни, политическая, религиозная, интеллектуальная или экономическая,
может оставаться в здоровом и цветущем состоянии, или что бедствия этих
времен пройдут незамеченными для глаз наблюдательных современников.
Например, то, что состояние сельского хозяйства в поздней Римской им¬
перии было неудовлетворительным, несмотря на законное вмешательство
государства, не дает основания для объяснения его постепенного упадка.
Народы в расцвете своих сил обычно не испытывают сложностей в успеш¬
ном использовании самых скудных природных ресурсов и вполне эффек¬
тивно адаптируют свою национальную организацию к гораздо более жесто¬
ким потрясениям, чем те, которые можно привести для объяснения неудач
более поздних этапов их цивилизации.Доктор, наблюдающий, как несколько пациентов болеют и умирают
в сходных, хотя и не идентичных условиях, и со сходными, хотя и не иден¬
тичными симптомами, сделает, несомненно, ошибку, если не попытается
найти единую общую причину этой болезни. Сложность симптомов и на¬
рушений разных органов тела не должны заставить его сделать вывод, что
первопричина или соответствующие методы лечения должны быть столь же
сложны. Разве это происходит не из-за того, что, по мнению врача, функ¬
ционирование тела, хотя и бесконечно сложное, является саморегулирую¬
щим и способным к нормальному корректирующему ответу на все обычные
нарушения; и только малое число нарушений исключительного характера,
не встречая эффективного ответа со стороны организма, вызывает серьез¬
ную болезнь? Нет сомнений, что мы имеем равное право ожидать нали¬
чие саморегулирующей силы и в человеческих обществах. Если нет, то мы
должны предсказать гибель таких обществ от любой из бесчисленных слу¬
чайностей, влиянию которой они могут быть подвержены. Нецивилизован¬
ные общества различных типов адаптировались к жизни в самых разных
климатических зонах от Арктики до тропических лесов и пустынь. Они со¬
существуют с самыми хищными и ядовитыми животными и часто стойко
переносят без нарушений нападения самых непримиримых врагов челове¬
Человек и общество179ка. Социальный прогресс не приостановился с внедрением нового оружия,
алкоголя, опиума или даже детоубийства, хотя каждое из этих изобретений
само по себе способно поставить существование расы под угрозу. То, что
цивилизованный человек, обладающий более эффективными приспособле¬
ниями, имеющий доступ к знаниям и организованный для более детального
сотрудничества, не способен оказать действенного сопротивления любому
нарушению своей социальной организации, требует, без сомнения, особого
объяснения.Социологические взглядыФилософский взгляд на историю, снискавший широкую популярность
в ряде стран на континенте, рассматривает расцвет, также как и упадок
цивилизаций, всего лишь как последовательность фаз цикла роста и уми¬
рания, которые, как предполагается, повторяются вновь и вновь и будут
повторяться бесконечно, пока существует человечество.50 Коль скоро на¬
ша цель — не более чем фактическое обобщенное описание этого явления,
можно сказать многое относительно фаз подъема и развития великих циви¬
лизаций в сочетании с фазами их упадка и гибели. Истинная удача, что мы
можем провести параллели между ранними стадиями развития различных
культур, их отношениями во времени и, наконец, более поздними фазами
их существования. Впрочем, обобщенное описание никогда не должно рас¬
сматриваться как самоцель. В лучшем случае — это средство для понимания
причинных процессов, породивших наблюдаемое явление. На определен¬
ном этапе достичь этого можно только за счет игнорирования расхождений
в деталях, которые, если бы причины были поняты, могли бы оказаться де¬
талями, имеющими серьезные последствия. Бывает также, что для описа¬
ния несколько отличных состояний и событий используют общие абстракт¬
ные термины, которые, чем более понятными они становятся, тем меньше
реального и поддающегося определению смысла содержат. И наконец, чи¬
сто описательная общая картина событий во времени, будучи фаталистична
по своей природе, не оставляет места для умного и корректирующего по¬
средничества.Ранние эволюционные теории греков фаталистичны и бесплодны
из-за отсутствия какого бы то ни было ясно понятого типа причинной свя¬
зи, которая могла бы привести к модификации живых организмов. Для на¬
учных взглядов середины девятнадцатого века характерно, что эволюци¬
онные теории ранних исследователей практически не вызывали интере¬
са из-за отсутствия удовлетворительных объяснений, способных выразить
средства модификации языком известных причин. Даже Дарвин и Уоллес,
180Глава 8хотя Дарвин и был обладателем большого количества косвенных доказа¬
тельств, не выдвигали свою эволюционную теорию до тех пор, пока каж¬
дый из них не стал уверен, что в борьбе за существование естественный от¬
бор действительно является действенной причиной. С ясным пониманием
научного принципа, который не всегда получает должную оценку, очевид¬
но, что они ощущали, что простой исторический факт передачи по наслед¬
ству модификации, как бы ни был велик интерес публики к этому вопро¬
су, не может плодотворно обсуждаться, пока не будет установлено, какими
именно средствами данная модификация может быть достигнута. Как толь¬
ко цепь причинно-следственных связей была определена, эволюция из Ис¬
тории превратилась в Науку. Очевидно, что чем более особым и неповто¬
римым является рассматриваемый случай, тем меньше можно полагаться
на описательную аналогию и тем более важным становится знание законов
причинных связей.В описательном исследовании взлета и падения человеческих цивили¬
заций ближайшей аналогией, где возможно применение научного метода,
является рост и смерть отдельного организма. Можно сказать, что и в том,
и в другом случае мы видим в регулярной последовательности период бур¬
ного развития и избытка жизненной энергии с обретением новых сил, пе¬
риод зрелости со значительными достижениями, период застоя, когда орга¬
низм или общество не могут сделать ничего, кроме собственного поддержа¬
ния, и период угасания, за которым рано или поздно следует исчезновение.
Впрочем, едва нам удается интерпретировать это обобщенное описание
в терминах известных причин, становится видно, что аналогия ошибочна.
«Юные» народы, которых мы видим на заре их цивилизованной истории,
уже имеют позади себя социальную историю, значительно более продол¬
жительную, чем период цивилизованности, лежащий впереди. Они не про¬
сто рождаются из элементов, оставленных предыдущими цивилизациями,
элементов, которые, как можно предположить, несут наследственные де¬
терминанты цикла будущих изменений. Можно сказать, что здоровое об¬
щество растет и ассимилирует; нельзя сказать, что оно себя репродуцирует.
А без репродукции не существует terminus a quo для последовательности
роста и смерти. Общества потенциально бессмертны, поддерживая внутри
себя силу постоянно замещать каждый живой элемент в своей структуре.
Феномен последовательности, из которой вытекает вся аналогия, наблюда¬
ется только в тех обществах, которые хотя бы непродолжительное время
обладали той кооперативной структурой с сильными связями, которую мы
и называем цивилизацией.‘Конечная остановка (лат.).
Человек и общество181Если отставить чисто описательную точку зрения, то становится оче¬
видным, что связывая возникновение и расцвет цивилизаций в единую
последовательность с их закатом и гибелью, мы не приходим к ясности
в отношении сути проблемы. Очевидно, что говорить о феномене упадка
можно только после достижения определенного уровня успеха. Но если
упадок цивилизации представляет проблему вследствие своей внезапности
и неожиданности, то преимущества, достигаемые в результате прогрессив¬
ного усвоения тех фаз культуры, которые мы называем цивилизованными,
ясны и известны всем. Следовательно, в то время как решение проблемы,
заключающейся в ухудшении условий даже для самого незначительного
социологического реагирования, обусловленного ими, несомненно прольет
свет на причины социологических изменений в целом, а тем самым на ха¬
рактеристику как ранних, так и более поздних фаз развития цивилизован¬
ного общества; лишь простое признание факта, что общество вследствие
наличия умений и форм организации цивилизованной жизни характеризу¬
ется и большим доступом к процветанию и власти, приведет нас к осо¬
знанию причин хорошо знакомых и вполне понятных. Эти преимущества
были бы еще более явными и длились бы дольше, если бы мы могли допу¬
стить, что жизнь в цивилизованных условиях, равно как и в варварском об¬
ществе, благоприятствует выживанию и репродукции таких человеческих
типов, которые способны наилучшим образом содействовать процветанию
своего общества, и которые, в свою очередь, наиболее склонны в силу сво¬
его темперамента к тому, чтобы оценить и использовать его преимущества.
Эволюция человека из дикого состояния через варварство к окончательно¬
му высокоцивилизованному состоянию могла бы в этом случае рассмат¬
риваться, в соответствии со взглядами Герберта Спенсера, как продолжи¬
тельный и неизбежный процесс совершенствования человека. В свете этого
вполне целесообразного оптимизма падение высокоразвитых цивилизаций
прошлого кажется единственной и невероятно трудной проблемой.Сообщества насекомыхЕдинственные сообщества животных, где кооперация развита доста¬
точно высоко, чтобы оправдать их сравнение с цивилизованным человеком,
это общественные насекомые. В этих сообществах специализация для вы¬
полнения различных задач, таких как защита, строительство, осуществ¬
ление социальной помощи подрастающему поколению, а также сбор или
культивирование пищи, развита достаточно высоко; тем не менее, хотя об¬
щественная жизнь в ряде различных групп возникла независимо, все они
в равной степени представляют сложность для сравнения с человечески¬
182 Глава 8ми обществами, поскольку в дополнение к существованию необходимой
для организованного сотрудничества специализации в поведении или функ¬
ционировании все они демонстрируют явно чрезмерную специализацию
в репродукции. Пример такой крайности — это единственная королева тер¬
митов, которая, как говорят, восполняет естественную убыль всего сооб¬
щества из многих миллионов рабочих, производя по одному яйцу каждые
несколько секунд без остановки. Как правило, подавляющее большинство
членов сообщества общественных насекомых не принимает участия в ре¬
продукции; и хотя мы не можем отрицать, что этот факт означает неко¬
торое экономическое преимущество, как и любая существующая при раз¬
делении труда специализация, все же опасность, которая подстерегает со¬
общество, доверившее свое будущее существование одной единственной
жизни, очевидна и достаточно серьезна. В этом отношении общество насе¬
комых больше напоминает тело одного животного, чем человеческое обще¬
ство, поскольку, хотя многие ткани и способны с помощью деления клеток
к восстановлению поврежденных участков, все же репродукция целостного
организма является делом специализированной репродуктивной ткани, то¬
гда как остальная часть тела со своими многочисленными кооперативными
функциями, которая сотрудничает, но не принимает участия в размножении,
в этом смысле является аналогией массы бесполых рабочих, составляю¬
щих большую часть термитника. Интересно проследить эффекты системы
размножения, принятой общественными насекомыми, с индивидуалистиче¬
ской системой размножения в человеческих обществах.Человеческие сообщества демонстрируют тот же тип генетической ва¬
риации, что и вид в целом; отдельные индивиды приспособлены к выпол¬
нению своих особых заданий частично благодаря прямому и преднамерен¬
ному отбору, частично благодаря непрямому отбору посредством принад¬
лежности к тому или иному социальному классу и частично благодаря спе¬
циальному образованию, включающему не только образование полученное
в юности, но и приобретенный практикой опыт. Считается, что различные
ткани тела многоклеточных организмов в норме генетически идентичны,
и демонстрируемые ими различия объясняются различными влияниями,
которые на них оказывают другие развивающиеся ткани. Многочисленные
потомки одной царицы не могут быть генетически идентичными, поскольку
факторы, по которым их мать является гетерозиготной, должны сегрегиро¬
вать, так что у каждой особи должны появиться две равные группы потом¬
ков, каждая со своим фактором. Впрочем, подобная генетическая измен¬
чивость, будучи разной в различных обществах, похоже, не используется
для производства полиморфизма среди особей рабочих и даже среди сте¬
рильных и способных к размножению типов. Хотя предмет весьма неясен,
Человек и общество183подавляющее большинство исследователей склоняется к мнению Уилпера,
который считает, что основное регулирующее влияние оказывает качество
пищи, которую получают личинки во время своего развития. Это означает,
что в данном случае механизм возникновения полиморфизма весьма отли¬
чен от простого менделевского механизма, характерного для полиморфных
бабочек, и значительно лучше приспособлен к нуждам общественных на¬
секомых. В то же время следует заметить, что, как и в случае с полиморф¬
ными бабочками, особый механизм, служащий для успеха полиморфизма,
вовсе не объясняет, какое именно приспособительное и эволюционное зна¬
чение имеет само разнообразие. Метод, посредством которого разное ко¬
личество пищи приводит к различиям в развитии, является переменной
величиной, столь же подверженной действию отбора, как и метод, посред¬
ством которого модифицируется узор на крыле конкретного менделевского
мутанта. Отбор в этом случае должен действовать исключительно на спо¬
собных к размножению насекомых, via* процветание общества, в котором
они возникли, и хотя эффект такого отбора может состоять в изменении
одних стерильных рабочих, это не представляется более сложным, чем тот
факт, что отбор, действующий исключительно на половые клетки овцы, via
видимые признаки несущего их животного должен изменять природу его
шерсти.Хотя среди стерильных рабочих одного и того же сообщества должны
существовать реальные генетические различия, эти различия не имеют се¬
лективного эффекта. Относительные частоты, с которыми разные генотипы
появляются среди потомков одной царицы, как следует полагать, должны
оставаться постоянными на протяжении ее репродуктивного периода, и от¬
бор самцов и самок для размножения должен зависеть только от их индиви¬
дуальных качеств и совокупного качества сообществ, членами которых они
являются. Напротив, генотипические различия, характеризующие индиви¬
дов из тех обществ, которые обладают индивидуальным размножением,
подвержены через свои дифференцированные уровни смертности и рож¬
даемости отбору внутри общества, способному бесконечно менять его ге¬
нотипический состав. Среди группы малочисленных враждующих племен
уничтожение племен с несоразмерным уровнем социально некомпетентных
индивидов и замена их ветвями более успешных племен может служить
средством поддержания среднего стандарта компетентности, соответству¬
ющего данному состоянию общества. Нет сомнения, что даже при таком
состоянии общества в нем будет происходить отбор, и чем более многочис¬
ленной будет становиться общественная группа, тем большей будет вели-Через, путем ( лат.).
184Глава 8чина отбора внутри общества. Соответственно, с самых древних известных
нам времен наследуемые склонности, которые, без сомнения, составляют
основу пригодности человека к жизни в обществе, дополняются экономи¬
ческой системой, которая, какими бы разнообразными ни были ее оценки
различными авторами, представляется мне одним из бессознательных три¬
умфов человеческой организации.Основу экономической системы составляет свободный обмен товарами
и услугами между отдельными индивидами, коль скоро подобный взаимо¬
обмен представляется выгодным обеим сторонам. Для того чтобы подобное
соглашение было свободным, необходимо, чтобы произвольное принужде¬
ние одного индивида другим было бы запрещено, тогда как, с другой сто¬
роны, принудительное выполнение свободно взятых на себя обязательств
поддерживалось бы силой общественного порядка. Столь же существенно,
чтобы личное владение собственностью, представляющей собой, как это
должно быть в данной системе, накопление уже оказанных по отношению
к другим членам общества услуг и эффективным средством требования эк¬
вивалентного возврата услуг в будущем, было бы строго защищено. В тео¬
рии этой системы каждый индивид принуждается к активным действиям
в собственных интересах, оказывая услуги окружающим и находя путем
собственной изобретательности новые или совершенствуя старые спосо¬
бы стать полезным обществу. Те, кто наиболее успешно оказывают ценные
услуги, получают наивысшее вознаграждение, включающее право направ¬
лять услуги других теми способами, которые кажутся им наиболее выгод¬
ными. Тот же, кто не способен совершать выгодные обществу действия,
имеет наименьший доступ к богатствам и удобствам общества. В теории
такие индивиды могут умереть от голода или влезть в долги, сумма кото¬
рых будет равна количеству всех потенциальных услуг остатка их жизни
или жизни их детей.Вряд ли стоит говорить, что такая экономическая система никогда
не являлась у людей единственной основой общественного сотрудничества.
Частично она была создана как компромисс с уже существовавшими соци¬
альными инстинктами, заложенными во время существования в меньшей
степени сотрудничающих между собой обществ. Тем не менее она доста¬
точно близка как к теории экономистов-рационалистов, так и к практике
многочисленных древних цивилизаций, указывая на то, что мы предста¬
вили в абстрактной и идеальной форме реальный и эффективный фактор
общественной организации у людей. Биологическое значение этого факто¬
ра состоит в благоприятствовании, которое он обеспечивает, следя, чтобы
отбор в человеческих обществах не способствовал размножению наиболее
непродуктивных или паразитических типов за счет тех, кто успешно расхо¬
Человек и общество185дует себя на общее благо. Напротив, создается впечатление, что он следит
за тем, чтобы тот, кто производит лучшие товары или оказывает наиболее
ценные услуги, был бы непрерывно вознагражден в каждом следующем по¬
колении, тогда как тот, кто по способностям или склонностям не способен
произвести эквивалент того, что потребляет, был бы удален. Аналогичным
образом этот благоприятствующий отбор не ограничивается индивидами-
производителями. Создается впечатление, что равно благоприятствующий
отбор будет происходить как в потреблении, так и в распределении. Ин¬
дивид, который по причине несовершенных инстинктов расходует свои
ресурсы так, что это не приносит ему биологическую пользу, индивид,
предпочитающий из-за предрассудков плохой рынок или из-за собственной
некомпетентности не способный заключить выгодную сделку, оказывается
в невыгодном положении как экономически, так, вероятно, и с точки зре¬
ния отбора. Отбор потребителя обеспечивает в важном отношении теорети¬
ческое завершение индивидуалистской экономической системы, поскольку
он создает средства, благодаря которому возможности получения богатства
путем достижения иллюзорных выгод будут уменьшаться до тех пор, пока
существенные источники получения выгоды будут сведены к получению
реальных общественных выгод. Тогда люди, сформированные подобной
системой, становятся изобретательными и энергичными предпринимателя¬
ми, трезво и проницательно оценивая социальные ценности и устанавливая
стандарты благосостояния, в полной мере отвечающие их биологическим
и репродуктивным интересам. Для завершения картины, предвидя возмож¬
ные возражения, скажем, что наши экономисты-утописты приходят тогда
к признанию права на владение богатством, по крайней мере как средства
воспроизводства, в качестве высшего блага и стремлений нему как спосо¬
ба объединения всех добродетельных усилий. На их взгляд, состоятельный
человек получает удовольствие не только от вознаграждений, но и от дока¬
зательства своей добродетели и добродетели своих предков; он становится
своего рода святым, и тогда содействие ему в его добродетельных делах бу¬
дет истинным блаженством. Таким людям невозможно выразить более бла¬
городное общественное признание, чем прямое денежное вознаграждение,
и приобретение всех других почестей за деньги будет казаться не столько
безнравственным, сколько абсурдным. Благотворительность в понимании
избавления от бедности не при помощи экономики неизбежно оказывается
порочной слабостью, хотя не исключается расчетливая поддержка, ведущая
к обоюдной выгоде, способных, но менее удачливых людей.Если бы в реальной жизни общественное развитие человека действи¬
тельно приняло бы такой оборот, то можно было бы сказать, что чело¬
век путем индивидуализации собственности нашел социально подходя¬
186Глава 8щее сопутствующее обстоятельство для своих индивидуальных репродук¬
тивных привычек, что он создал тип общества, недостижимый для на¬
секомых из-за нехватки у них интеллектуального оснащения, пригодного
для того, чтобы с достаточной справедливостью и постоянством вести дела
о спорной собственности или контрактных обязательствах, или, по сути,
из-за недостатка у них адвокатов и судей; и что по этой причине един¬
ственным доступным им средством, позволяющим избежать гибельных по¬
следствий отбора внутри общества, является полное устранение конкурен¬
ции между репродуктивными членами одного и того же общества путем
передачи репродуктивных обязанностей одному единственному индивиду.
Впрочем, дело обстоит так, что даже если бы сообщество насекомых стало
обладателем некоей действующей системы инстинктов, имеющей экономи¬
ческий эффект частной собственности, его трудности на этом бы не за¬
кончились, поскольку инстинктивные чувства и предрассудки человека об¬
щественного вовсе не развились в направлении более узко экономической
и менее «чувственной» основы для общественных учреждений. Могло бы
ли воображаемое насекомое и может ли человек преодолеть дальнейшую
социологическую проблему, которая, возникнув, может быть решена толь¬
ко после более тщательного ее изучения. Важным пунктом настоящей гла¬
вы является то, что проблемы, с которыми сталкивается человек и которые
в большинстве случаев преодолевает, избегают все группы общественных
насекомых, образующих сообщества, состоящие из потомства одного ин¬
дивида.Самый начальный эволюционный процесс, который здесь следует от¬
метить, это невероятное, хотя и, несомненно, постепенное, увеличение ин¬
дивидуальной фертильности у насекомых. Как было отмечено в главе 2,
значение Естественного Отбора для доведения фертильности до оптималь¬
ного уровня было затемнено тем, что ранние эволюционисты понимали
«высокую скорость роста» как собственно движущую силу Естественного
Отбора. Когда, напротив, мы рассматриваем фертильность как вторичное
явление, определяемое Естественным Отбором, становится необходимым
рассмотреть, какие обстоятельства влияют на уровень фертильности, кото¬
рая в каждом конкретном случае оптимальна. В этой связи майор Леонард
Дарвин указал, что основное внимание следует уделить фактору родитель¬
ской заботы, включив в это понятие все затраты в виде питания, особых
усилий или повышенного риска, требующиеся при рождении и выращива¬
нии потомства. У организмов, у которых степень родительских затрат, поз¬
воляющих достигать наиболее высокой вероятности выживания потомства,
высока по сравнению с доступными ресурсами, оптимальная фертильность
будет сравнительно мала. Любое обстоятельство, которое материально об¬
Человек и общество187легчает родительское бремя, будет неизбежно иметь мгновенный эффект,
способствуя выживанию; важнее то, что оно будет всегда снижать опти¬
мальные затраты и, соответственно, стремиться поднять уровень фертиль¬
ности. Майор Дарвин иллюстрирует этот принцип на примере паразити¬
ческих кукушек, которые нашли способ избавить себя от большей части
родительских забот, свойственных остальным птицам, что позволило им
стать значительно более плодовитыми по сравнению с их непаразитиче¬
скими родственниками. Общественные насекомые предлагают еще более
красочный пример того же принципа. Как только молодые взрослые ин¬
дивиды любой зарождающейся социальной формы берутся либо за выпол¬
нение подготовительной работы к репродукции, либо за уход за подрас¬
тающим потомством до того, как они сами начнут размножаться, баланс
селективного преимущества оказывается сдвинут в сторону благоприят¬
ствования фертильности основательницы колонии, способствуя в равной
степени развитию органов и инстинктов рабочих, а не цариц из ее ран¬
него и, возможно, хуже выкормленного потомства. Невероятное развитие
фертильности наряду с полной стерильностью во всех группах обществен¬
ных насекомых показывает, насколько эффективно действие отбора в моди¬
фикации конкретного признака. В следующей главе (9) будут обсуждаться
наследственные факторы, влияющие на фертильность Человека, и там мы
увидим, что селективная модификация этих факторов так же важна, хотя
по совершенно иным причинам, как для эволюции человека, так и для со¬
обществ насекомых.РезюмеРанние эволюционные работы, посвященные Человеку, естественно,
были направлены на установление двух предварительных точек зрения:
(1) что Человек, как и другие животные, является продуктом эволюцион¬
ного процесса, (2) что интеллектуальные и моральные качества Человека,
равно как и его физические качества, являются результатом природных при¬
чин. Трудность, которую ощущает Человек, пересматривая свое традицион¬
ное мнение о себе, требует, чтобы оба эти положения были бы установлены
с достаточной убедительностью.Натуралистический взгляд на Человека не дает оснований для объек¬
тивного рассмотрения этих интеллектуальных ценностей, моральных и эс¬
тетических, которые так много значат для Человека; следовательно, натура¬
листический взгляд на Человека нельзя использовать для того, чтобы под¬
вергать сомнению эти ценности. Высокая ценность для Человека мораль¬
ных качеств придает их природной обусловленности особое значение.
188Глава 8Среди проблем социальной эволюции Человека, наиболее заметной яв¬
ляется упадок и гибель всех цивилизаций, предшествовавших нашей соб¬
ственной, несмотря на все основания для их длительного успеха и процве¬
тания. Чисто описательный подход к проблеме взлета и падения цивили¬
заций не может быть исчерпывающим без исследования причин, вызываю¬
щих изменение социальной структуры.В царстве животных единственные сообщества, пригодные для срав¬
нения с человеческой цивилизацией, это — сообщества общественных насе¬
комых, которые отличаются от человеческих обществ удивительной чертой
репродуктивной специализации. В результате достигается эффект исклю¬
чения отбора внутри общества как эволюционного процесса, так что опас¬
ности, которые содержит этот источник, характерны только для Человека.
Человеческое общество с самого раннего времени приняло экономическую
систему частной собственности, которая, как можно ожидать, должна была
бы осуществлять регулирующий отбор внутри общества в отношении со¬
циально успешных линий. Последствия, которых в соответствии с логикой
можно было ожидать, так и не были реализованы.Специализация репродукции в сообществах насекомых стала возмож¬
ной благодаря эффективности отбора на модификацию фертильности. На¬
следование фертильности, как мы увидим в дальнейшем, в равной степени
подвержено влиянию, хотя и совершенно иным образом, в эволюции чело¬
веческих обществ.
Глава 9Наследование фертильности у человекаБольшая изменчивость репродуктивности у человека. Умственные способно¬
сти и моральные качества, определяющие репродуктивность. Прямое доказатель¬
ство наследования фертильности. Эволюция совести не нарушает сознательной ре¬
продукции. Аналогии с инстинктом животных и иммунитетом к болезням. Резюме.Своей могилой хочешь стать за что,Любви своей в себе замкнувши круг ?* У. ШЕКСПИРБольшая изменчивость репродуктивности у человекаМожно предположить, что наличие статистических данных о цивили¬
зованном человеке должно позволить дать оценку возможностям любого
представителя данного вида оставить после себя 0,1,2,... потомков соот¬
ветственно обстоятельствам жизни, о которых говорилось в главе 4. Как
будет показано далее, человек в данном смысле отличается от большинства
других организмов относительно высокими различиями по фертильности
и сравнительно малыми различиями по уровню смертности от рождения
до репродуктивного периода. Принимая во внимание указанные особенно¬
сти человека, большой интерес представляет вопрос об оценке, хотя бы
приблизительной, величины элемента случайности среди причин вариации
числа фактически рожденных детей. При попытке определить эту величину,
следует иметь в виду две возможные трудности, преодолеть которые, учи¬
тывая состояние наших сегодняшних знаний, можно, самое большее, лишь
частично. Во-первых, фактическую вариацию числа рожденных детей, ска¬
жем, у группы женщин, репродуктивный период которых подходит к концу,
нельзя полностью приписать только лишь случаю, хоть как-то это аргумен¬
тируя. Возможно, в этом отношении человечество в его цивилизованном
состоянии принципиально отличается от всех других организмов, исключая
общественных насекомых. Ибо если только темпы репродуктивного отбора*Сонет III, в переводе А. Финкеля.
190Глава 9у других организмов не являются значительно более высокими, чем предпо¬
ложенные нами, фактор случайности должен, как следует из главы 4, быть
доминирующим в выживании индивида, представляя, в сущности, цели¬
ком индивидуальную дисперсию. Во-вторых, различия в индивидуальных
характеристике и темпераменте человека имеют не меньшую дисперсию,
чем элемент случайности; и, следовательно, порядок величины, в целом,
должен отличаться от эффектов репродуктивного отбора, которые следует
ожидать у других организмов.Если мы не можем оценить компоненту случайной дисперсии, прирав¬
нивая ее к общей дисперсии, то альтернативный метод оценки a priori как
результат последовательных независимых событий представляется в луч¬
шем случае крайне сомнительным, что объясняется долгими и настойчи¬
выми поисками. Рождения последующих детей не являются независимыми
событиями, поскольку такие события обусловлены тем, состоят или не со¬
стоят в браке их родители, несмотря на то, что такое состояние может со¬
храняться в силу влияния, хотя и в очень малой степени, чисто случайно¬
го элемента. Полные статистические данные о рождаемости у состоящих
в браке и у не состоящих в браке для каждого возраста, о частоте вступле¬
ния в брак незамужних и неженатых, а также о частоте перехода в разряд
не состоящих в браке ввиду развода или смерти супруга(и) по-прежнему
не позволяют непосредственно измерить степень, с которой разные члены
недифференцированной группы различаются по числу детей. Процесс вы¬
числения, который учитывает не только последовательные роды, но также
и браки, как обусловливающие вероятность родов, в качестве ряда незави¬
симых событий с соответствующими частотами представляется таким же
ошибочным, поскольку брак считается случайным, как и более простой ме¬
тод вычисления, при котором эффект брака вовсе не учитывается.Повюс (Biometrica, IV. 238) приводит данные о количестве детей, рож¬
денных 10276 замужними женщинами, умершими в возрасте 46 лет или
старше, в Новом Южном Уэлсе с 1898 по 1902 год. Число детей варьиру¬
ет от 0 до 30, число женщин с тем или иным числом детей представлено
в таблице 7. Среднее число детей у матери составляло 6,19, и если бы
фертильность женщин была одной и той же, т. е. все женщины имели бы
одинаковую вероятность стать матерью, то дисперсия, безусловно, была бы,
несомненно, меньше, чем среднее.51 В действительности дисперсия состав¬
ляет 16,18; и это более чем в два раза превышает самое большое значение,
которое могло быть обусловлено случайно.Значительная часть дисперсии должна, конечно же, относиться к брач¬
ному возрасту. И дисперсия по-прежнему удивительным образом превы¬
шает среднее значение, если в выборке роль указанного фактора сведена
Наследование фертильности у человека191Таблица 7ЧислоЧислоЧислоЧислоЧислоЧислодетейматерейдетейматерейдетейматерей01 100И56822115331242223—25811322624136441412925-4702155726-5813163927-6855171228-797618529-8963192301984720210786211Итого наблюдений: 10276к минимуму. Группа замужних женщин из 2 322 человек, вышедших замуж
до 20 лет и живущих с мужьями во время переписи (1 901) через 25-30 лет
после замужества, упорядочена так, как показано в таблице 8.Таблица 8ЧислоЧислоЧислоЧислоЧислоЧислодетейматерейдетейматерейдетейматерей0441167147714182371546263927216193651033317948311271186510512208193614113131201Итого наблюдений: 2 322Среднее число детей для данной группы женщин, условия выборки
для которых особенно благоприятны в отношении фертильности, составля¬
ет 8,83, но дисперсия по-прежнему высока и составляет 12,43.Эти примеры и, несомненно, любая совокупность подобных данных
свидетельствуют о том, что хотя невозможно дать точную оценку фактора
случайности в репродукции, очевидно, что существуют причины для такой
большой дисперсии — как в брачном возрасте, так и во время жизни в браке,
и она оказывается сравнимой с самим фактором случайности.
192Глава 9Огромная вариация фертильности у Человека была показана несколь¬
ко иным методом доктором Героном, использовавшим данные о смертно¬
сти (30285 мужчин и 21 892 женщины) в Австралийском Союзе по данным
1912 года. Герон обнаружил, что половина всех детей происходила из семей
с 8 и более детьми, родителями которых были лишь У9 всех мужчин и У7
всех женщин предыдущего поколения. Было бы преувеличением предпо¬
лагать, что вся дифференциальная репродукция обусловлена отбором; су¬
щественная часть ее, несомненно, является волей случая, но не существует
такой теории, согласно которой представляется возможным отрицать, что
столь же существенная доля есть следствие собственно дифференциальной
фертильности, естественной или искусственной, у разных представителей~ КОчеловеческой популяции.Умственные способности и моральные качества,
определяющие репродуктивностьЕсли роль случайности в наблюдаемых вариациях репродуктивности
не ясна и оценивается с трудом, то роль конститутивных и, следователь¬
но, наследственных различий темперамента очевидна всем. Когда мы об¬
суждаем причины, обычно обусловливающие деторождение, то видим, что
во многих случаях темперамент индивида имеет решающее значение. Зна¬
чительный процент людей обоего пола вообще не вступает в брак: возраст
вступления в брак очень варьирует, особенно у женщин эффект возрас¬
та очень велик: согласно австралийским данным 1911 года, максимальная
репродуктивность женщины приходится на 18 лет, а в 31 год — снижает¬
ся наполовину от максимального значения. Если женщина вышла замуж
в 30 лет, то можно ожидать, что число рожденных ею детей составит 38 %
от того числа, которое она произвела бы на свет, если бы вышла замуж
в 20 лет. К 35 годам эта цифра уменьшается еще наполовину и составляет
чуть меньше 19%. Что касается мужчин, то их потенциал отцовства обыч¬
но сохраняется до более позднего возраста, но брачный возраст все рав¬
но имеет большое значение, поскольку чем старше жених, тем чаще всего
старше и невеста. У женихов от 34 до 44 лет невестами часто оказываются
женщины примерно на десять лет младше. Если мы возьмем за основу вы¬
числений возраст их жен, то выяснится, что от мужчин, вступающих в брак
в возрасте 40-44 лет, можно ожидать лишь 2/5 того числа детей, которые,
возможно, были бы рождены, если бы они женились на 20 лет раньше.Выбор между безбрачием и браком, а если выбор падает на брак,
то между поспешным браком и откладыванием брака, является в современ¬
ных обществах в значительной мере следствием темперамента. На некото¬
Наследование фертильности у человека193рых мужчин женские чары действуют не очень сильно, другие — более ком¬
панейские и общительные — долго представляются неподходящими для по¬
стоянного союза; у всех обычных мужчин первые порывы к занятиям сек¬
сом, по-видимому, развиваются в достаточной мере, но в условиях цивили¬
зованного общества сила данного порыва не прилагается непосредственно
к статусу супруга: у многих мужчин она подавляется осторожностью и са¬
моотречением, у других — расточается в непродуктивных направлениях.
Разные натуры по-разному реагируют на различные привилегии и обяза¬
тельства супружеских отношений: терпение, осторожность и сильное чув¬
ство финансовой ответственности часто задерживают желанный брак или
мешают его осуществлению; уверенные, страстные и импульсивные натуры
вступают в брак в таких обстоятельствах, которые пробудили бы у других
огромные опасения. В наши дни влияние врожденного характера в дан¬
ном вопросе повышается как в силу отсутствия любого устойчивого обще¬
ственного мнения, с которым индивид мог бы соразмерять свои ошибочные
увлечения, так и в силу влияния условий городской жизни, которые не огра¬
ничивают разнообразие знакомств. Казалось бы, случайные обстоятельства
для отдельного человека могут быть очень важными, но их влияние легко
преувеличить даже в отношении определения судьбы индивида. Мужчина,
который в силу раннего разочарования обрекает себя на безбрачие, в мень¬
шей мере является жертвой неудачи, чем собственного темперамента, так¬
же как и опрометчивый юноша, который на всю жизнь остается жертвой
неудачных обстоятельств. В каждом случае темперамент в значительной
степени определяет поведение в рискованных ситуациях, и в совокупно¬
стях достаточной численности элемент случайности ничтожен.Условно считается, что инициатива при вступлении в брак принадле¬
жит мужчине, в то время как факторы, определяющие безбрачие или воз¬
раст замужества для женщины обусловлены характером или частотой пред¬
ложений, которые она получает. Даже если бы на практике это различие ре¬
ализовывалось в полной мере, было бы нетрудно допустить, что различия
в темпераменте молодых женщин фактически оказывают преобладающее
влияние на брак, скажем, на протяжении первых пяти лет. Другие наслед¬
ственные признаки также влияют на эту возможность в той степени, в какой
они управляют красотой, здоровьем или чертами характера, пользующими¬
ся вниманием со стороны поклонников. На самом деле различия между
полами в данном вопросе далеко не абсолютные, и вероятность вступления
в брак и у мужчины, и у женщины во многом зависит от качеств, которые
они обычно приписывают друг другу.Помимо указанных причин варьирования фертильности, действую¬
щих в сфере социальной структуры брака или безбрачия, в цивилизован¬
194Глава 9ной и нецивилизованной жизни с древнейших времен существуют обычаи,
при помощи которых происходит искусственное ограничение прироста на¬
селения. Детоубийство, умерщвление плода, или аборт, и предупреждение
зачатия использовались и используются сейчас во всех значительных сооб¬
ществах людей; к этому списку следует добавить такие обычаи, как прости¬
туция, которая достигает той же цели, становясь альтернативой вступления
в брак, вместо того, чтобы осквернять его.Степень применения практики убийства новорожденных можно оце¬
нить, внимательно прочитав мастерски написанную статью Вестермарка,
посвященную данному вопросу. У нецивилизованных народов данная прак¬
тика была обычной и часто обязательной, хотя и не повсеместно. Она
была запрещена в буддизме и даоизме, но часто использовалась в Китае.
У доисламских арабов убийство новорожденных было, похоже, обычной
практикой, получавшей одобрение; Коран запрещает эту практику, при¬
чем запрет повторяется не раз. В Священном Писании о ней не упо¬
минается вообще, ранним иудеям она могла быть неизвестна. В Индии,
где она была под запретом согласно законам Ману, к ней особенно бы¬
ли склонны раджпуты, хотя они должны были получать за это отпуще¬
ние грехов у своего жреца. В Древней Греции допускалось, хотя едва ли
одобрялось, оставить умирать здорового новорожденного, лишь в Фивах
это считалось преступлением и каралось смертной казнью. В Риме, хо¬
тя и предписывалось уничтожать уродливых младенцев, убийство здоро¬
вых младенцев, в том числе не столь решительное оставление ребенка
с расчетом, что он не выживет, противоречило превалирующим нормам
морали.Из числа всех указанных факторов, влияющих на фертильность су¬
пружеских пар в нашей цивилизации, главное внимание должно уделяться
предохранению от зачатия. В настоящее время, несомненно, широко рас¬
пространено ограничение рождения у супружеских пар в Европе и Аме¬
рике. Определенные круги всерьез убеждены в том, что только таким спо¬
собом можно избежать личной нужды и низкого уровня жизни общества;
использование данных методов считается не только благодеянием, но и дол¬
гом. Общественное мнение, если удается его определить, по существу, сво¬
дится к тому, что это дело сугубо личное — кажется, напоминает мне¬
ние Плиния, считавшего, что аборт — это простительное правонаруше¬
ние. «Огромная плодовитость некоторых женщин, — считал Плиний, — мо¬
жет потребовать такого разрешения». Важно отметить, что спланированные
контрацептивные меры в последние десятилетия являются самой главной
причиной не только общего снижения уровня рождаемости, но особенно
снижения фертильности высших классов.
Наследование фертильности у человека195В практике использования контрацепции или отказа от нее особенно
ясно отражены различия темперамента, включая отношение к моральному
аспекту сексуального поведения. Это наглядно представлено в разного рода
обоснованиях, с помощью которых эта практика пропагандируется. Несо¬
мненно, что свобода от индивидуальных обязательств перед естественны¬
ми последствиями половых отношений является самым привлекательным
моментом, и что, конечно, будет приветствоваться радикальное изменение
принятого мнения о моральном аспекте сексуальных отношений теми, кто
не состоит в браке, в том числе отказа от традиционного воздержания.
Для других, кто испытывает абсолютную неприязнь к сексуальной анархии,
настоящую привлекательность имеет возможность удовлетворения сильно
выраженных родительских инстинктов; перспектива материально обеспе¬
чивать и обучать ранних детей может быть разумным образом отодвинута
на более поздний срок. Усиливается мнение, что дружеские отношения ро¬
дителей оказывают благотворное влияние, поскольку внимание концентри¬
руется на немногих членах семьи, это мнение, наверное, следует отнести
не к причинам, а к оправданиям сокращения размера семьи. Но даже оправ¬
дания имеют смысл ввиду огромной изменчивости эмоциональных откли¬
ков, которые они пробуждают. Я подозреваю, что даже большинство тех,
кто использует контрацептивные средства, относятся к ним с неохотой или
чувством неприязни, но в зависимости от силы данного чувства его могут
преодолеть под напором экономического давления, в большей или мень¬
шей степени на них оказываемого. Другие обязательно почувствовали бы
себя скомпрометированными, если бы позволили экономическим мотивам
избавиться в какой бы то ни было степени от естественного плода их брака.Если мы рассмотрим огромное разнообразие реакций разных темпе¬
раментов на два доминирующих фактора, определяющих фертильность:
с одной стороны — брак, а с другой — ограниченное или неограничен¬
ное производство детей супружескими парами, то большая изменчивость,
свойственная этим причинам, в статистических данных цивилизованных
народов не покажется нам удивительной; и нам будет понятно, почему ге¬
нотипические различия в репродуктивности перевешивают любые геноти¬
пические различия в смертности, особенно с тех пор, как смертность сре¬
ди цивилизованных народов намного уменьшилась вследствие достижений
в общественной и личной гигиене. Что касается смертности, то необходимо
отметить, что лишь смерти до репродуктивного периода и в меньшей сте¬
пени во время него имеют некоторое селективное влияние, и что они уже
составляют наименьшую долю всех смертей. Диаграмма в главе 2, показы¬
вающая репродуктивную ценность женщин в соответствии с их возрастом,
дает ясное представление о важности в этом отношении уровней смертно¬
196Глава 9сти для разного возраста. Даже самый высокий уровень смертности в пе¬
риод, который приходится на первый год жизни, должен быть довольно
незначительным в сравнении с небольшими различиями в репродуктивно¬
сти; поскольку уровень смертности младенцев снизился в нашей стране
до примерно семи процентов от всех смертей, то если даже удвоить этот
уровень, данный показатель составит лишь третью часть от показателя вы¬
живания после добавления к семьям с тремя детьми семей с четырьмя.Прямое доказательство наследования фертильностиНесомненно, что вклад случайной компоненты в наблюдаемую дис¬
персию репродуктивности индивида чрезвычайно велик, однако можно по¬
лучить прямое доказательство важности наследственной обусловленности
репродуктивности, если сравнить число рожденных индивидом детей с чис¬
лом детей в семье его родителей. В качестве биометрического признака число
детей, рожденных индивидом, специфично в нескольких отношениях. Оно,
подобно меристическим признакам, дискретно и ограничено целыми поло¬
жительными числами и в то же время, в отличие от них, начинается с нуля.
Кроме того, там, где репродуктивность управляется социальным институ¬
том брака, этот признак будет одним и тем же для мужчины и его супруги
и, строго говоря, не может рассматриваться как отличительная черта кого-
либо из них. Очевидно также и то, что если значительная часть дисперсии
носит случайный характер, то дисперсия любой группы индивидов будет
увеличиваться в сравнении с ковариансами между родственными группа¬
ми; и в результате все наблюдаемые корреляции будут значительно меньше
в сравнении с теми, которые можно обнаружить между метрическими при¬
знаками. Следовательно, если необходимо получить результаты, не вызыва¬
ющие сомнений, приходится исследовать довольно многочисленные группы.Из предоставленных Пирсоном данных о числе детей, рожденных ма¬
терями и их дочерьми, при рассмотрении 1 ООО случаев, взятых по бри¬
танским пэрам, следует, что среднее число детей, рожденных дочерью, бу¬
дет возрастать в соответствии с размерами семьи, в которой она родилась,
с 2,97 — если она родилась в семье с одним ребенком, до 6,44 — если в се¬
мье матери было 12 или более детей. Эти данные приведены в таблице 9.Средние значения, представленные на рис. 10, свидетельствуют о том,
что с увеличением числа детей в материнской семье наблюдается так¬
же и последовательное увеличение числа детей, рожденных их дочерь¬
ми, несмотря на неизбежную нерегулярность данной тенденции, обуслов¬
ленную ограниченным объемом выборки. В среднем увеличение на 1 ре¬
бенка в материнской семье приводит к увеличению числа детей дочери
Наследование фертильности у человека197Числодетей,рожденныхматерьюТаблица 9Число Общее число Среднее числонаблю- детей, рожденных детейдений дочерьми у дочери1351042,972672373,5431113493,1441364643,4151385503,9961325133,8971144483,938873544,079743134,2310502194,3811241225,081219'99']5,21 '139778,56141>32 10,206 10,00 *15з20.6,676,44на 0,21, и хотя это меньше половины значения, которое мы должны по¬
лучить при сравнении метрических признаков отдельных родителей и де¬
тей, когда данные величины почти всецело определяются наследственными
причинами, все же полученное по этим данным значение более чем в 6 раз
превышает его стандартную ошибку и является, несомненно, значимым,
так как демонстрирует тенденцию, согласно которой семья дочери имеет
сходство с семьей ее матери с положительным или отрицательным откло¬
нением от среднего значения, характеризующего ее поколение.Было бы важно убедиться, до какой степени данное сходство можно,
если вообще можно, объяснить непрерывностью традиций, религиозным
воспитанием, моральным климатом или социальными идеями. Принимая
данное объяснение, исключающее эффект наследственности, мы предпола¬
гаем, что люди не склонны перенимать такие представления и претворять
их в жизнь под влиянием унаследованных черт характера, что и опроверга¬
ется очевидными различиями, наблюдаемыми у членов одной семьи. Кро¬
ме того, при рассмотрении причин, которые могли бы повлиять на идеи
молодой девушки, далеко не очевиден тот факт, что климат большой семьи
способствует большей степени фертильности, чем климат маленькой се¬
мьи. Пример матери может иметь и обратный эффект, и было бы рискован-
198 Глава 97 1 1 1 1 1 1 1 1 1 1 1 1 1 г&sni ^
оcdо © оооол1 1 1 1 1 1 1 I I I I I 1 1 1 1 2 3 4 5 6 7 8 9 10 И 12 13 14 15
величина семьи матери ■35 67 111 136 138 132 114 87 74 50 24 32количество семейРис. 10. Среднее число детей, рожденных супругами пэров, в соответствии с разме¬
рами семей их матерейно утверждать, что осознание преимуществ домашней среды возобладает
над пониманием свойственных ей недостатков.Как бы там ни было, но в данных о связи фертильности внучек и их
бабушек по отцовской линии имеется недвусмысленное подтверждение той
точки зрения, что взаимосвязь, прослеживаемая между матерями и дочерь¬
ми, по существу, имеет наследственный характер. Рисунок 11 показывает,
что регрессия фертильности дочери на фертильность бабушки составляет
0,1065, т. е. почти точно половину значения, которое получено для влия¬
ния матери. Результат, который получается в силу лишь наследственности,
необходимо уменьшать примерно наполовину в каждом поколении, не го¬
воря уже о любой сильной тенденции к гомогамии; и хотя влияние бабушки
подвержено довольно большим ошибкам выборки, — но с точки зрения ста¬
тистики оно является важным, — его совпадение с ожидаемым значением,
Наследование фертильности у человека199\ 1 1 1 1 Г 1 1 1 1 1о1 1 1 -Т г1 1 1о0_оооо©1GООО"1 1 1 1 I I 1 L 1— .1 1 1О 1 2 3 4 5 6 7 8 9 10 И 12 13 14 15 16 17 18 19 20 21
величина семьи бабушки67 91 118 123 125 102 99 98 72 41 28количество семей36Рис. 11. Среднее число детей, рожденных внучками, в сравнении с количеством
детей у бабушек по отцовской линиинесомненно, указывает на наследственность как ответственную по мень¬
шей мере за большую часть наблюдаемых корреляций.Наследственную обусловленность фертильности можно обосновать
и другим путем. В таблице, где сравниваются матери и дочери, ковариация*
между двумя значениями фертильности составляет 20,9 % дисперсии фер¬
тильности дочерей. Для признаков, которые почти всецело определяются на¬
следственностью, эта ковариация должна составлять почти половину дис¬
персии. Таким образом, мы можем предположить, что 41,8% наблюдае¬
мой дисперсии фертильности можно приписать наследственным причинам.
В главе 2 указывалось, что удовлетворительную оценку генетической компо¬
ненты дисперсии любого признака можно получить, разделив квадрат корре¬
ляции между родителем и ребенком на корреляцию между бабушкой и внуч¬
кой. В данном примере это дает нам следующий результат: (0,2096)2/0,1123,
или 39,1 %. Следовательно, согласно двум рассмотренным методам пример¬
но 40 % наблюдаемой дисперсии определяется наследственными причина-’ Среднее произведение оклонений от средних. — Прим. автора.
200Глава 9ми, и эта оценка, если мы примем во внимание огромное преобладание
случайности в выживании индивида, о чем говорилось в главе 2, показы¬
вает, что наследственные факторы, влияющие на фертильность человека,
должны иметь селективный эффект исключительной величины.Мы можем выразить интенсивность отбора в единицах, которые ис¬
пользовали ранее для выражения различий в мальтузианских параметрах,
а именно в процентах на поколение, и признать, что генетическая диспер¬
сия, к выводу о существовании которой мы пришли в предыдущем абзаце,
имеет величину 3,042 единицы со стандартным отклонением 1,745 детей
на семью, обусловленную только наследственными причинами. Посколь¬
ку среднее число детей, рожденных дочерьми, равно 3,923, то индивиды
или семьи, число детей у которых отличается от этого среднего значения
не больше чем на стандартное отклонение в противоположных направле¬
ниях, будут иметь размеры семьи в соотношении 5,668 : 2,178. Если про¬
должительность поколений одинакова, то мы можем получить селективное
преимущество более фертильных над менее фертильными группами, вы¬
числив разницу между натуральными логарифмами этих двух величин. Эта
разница составляет 0,9564, это говорит о том, что селективное преимуще¬
ство, созданное вариациями (отнюдь не исключительно большими) врож¬
денной фертильности составляет свыше 95 % в каждом поколении. В такой
форме утверждение об интенсивности отбора сравнимо с отбором поряд¬
ка 1 %, которое, как было установлено ранее, удобно рассматривать на при¬
мере численных результатов отбора определенной интенсивности.Эволюция совести не нарушает сознательной репродукцииРасчеты, выполненные в предыдущем разделе, означают, что по мень¬
шей мере среди наших современников различия в наследственной предрас¬
положенности играют такую большую роль в определении фертильности,
что сколько-нибудь большие модификации темперамента должны быть вы¬
званы репродуктивным отбором в течение десяти поколений, т. е. в пре¬
делах относительно короткого исторического периода. Мы видели, что от¬
нюдь не наиболее контрастно различающиеся генотипы имеют уровень ре¬
продуктивности в соотношении выше, чем 2 к 1. Проще говоря, если пред¬
ставить себе популяцию, которая состоит из двух линий, имеющих одина¬
ковую смертность, но отличающихся только по фертильности именно в та¬
ком соотношении, то нетрудно подсчитать, что преобладание одного типа
над другим через 10 поколений возрастет более чем в 1 000 раз. Другими
словами, если в начале периода популяция состояла из 97 % менее фертиль¬
ной линии и из 3 % более фертильной, то через 5 поколений линии срав¬
Наследование фертильности у человека201няются по численности, а еще через 5 поколений ситуация станет противо¬
положной, т. е. более фертильная линия составит 97 % популяции, а менее
фертильная линия — соответственно 3 %. На самом деле, судя по стати¬
стическим данным на сегодняшний день, очевидно, что цивилизованный
человек подвергается процессу отбора, интенсивность которого в 100 раз
превышает тот, что мы можем ожидать у диких животных, за исключением,
возможно, тех групп, которые пострадали от недавнего и глубокого измене¬
ния их среды обитания. Следовательно, мы можем ожидать, что в течение
исторического времени такая же быстрая эволюция имела место и в отно¬
шении умственных способностей человека.Как хорошо известно, традиции детоубийства у нецивилизованных на¬
родов до некоторой степени широко распространены. Если мы рассмотрим
опасности в течение жизни и тяжелые лишения, которым время от вре¬
мени подвергались нецивилизованные народы, то, я полагаю, будет ошиб¬
кой объяснять данные традиции лишь неспособностью к адаптации. Везде,
где смертность в младенческом возрасте и в детстве была крайне высо¬
кой, репродуктивная ценность новорожденного, в том смысле, о котором
говорилось в главе 2, должна была быть небольшой по сравнению с ре¬
продуктивной ценностью молодой и фертильной женщины. Во время го¬
лода или срочной вынужденной миграции попытка спасти жизнь ребенка,
скорее всего, была бы не только обречена на провал, но естественным об¬
разом подвергала бы опасности более ценную жизнь его матери. Акт де¬
тоубийства, каким бы отвратительным он ни казался для цивилизованных
чувств, являлся в этих обстоятельствах естественным правилом поведения,
и неудивительно, что его часто следует считать моральным поступком и,
может быть, в некоторой степени даже необходимым. Мотив детоубийства
можно рассматривать как экономический; но среди народов, у которых на¬
копление собственности минимально, чувство предусмотрительности, на¬
пример в отношении будущих запасов, проявляется лишь в крайних случа¬
ях, так что едва ли мотивом служил расчет. У оседлых народов, напротив,
расчет и накопление собственности начинают играть существенную роль,
и в то же время немедленным следствием повышения благосостояния яв¬
ляется снижение роли селекции в общем уровне смертности и возрастание
значимости уровня репродуктивности. Поэтому неудивительно, что рели¬
гии всех без исключения цивилизованных народов должны категорически
осуждать детоубийство, хотя в некоторых случаях, например у народов Се¬
верной Европы и доисламских арабов, эти традиции, как мы знаем, суще¬
ствовали среди их варварских предков.Очевидно, что естественные инстинкты отца и матери должны проти¬
виться практике детоубийства, даже когда она кажется необходимой в си¬
202Глава 9лу сложившихся обстоятельств. Кроме того, несомненно, что дикари, как
и цивилизованные люди, могут быть разными: одни более чувствительны,
другие в большей степени бессердечны. Чувство отвращения и антипатии
по отношению к жестокому поступку должно быть в большей степени раз¬
вито у одних родителей, чем у других, а поскольку использование практики
детоубийства всегда в той или иной степени зависит от решения родителей,
то нет сомнений в том, что первые позволяли жить большему число по¬
томков. Таким образом, везде, где детоубийство практиковалось без давле¬
ния чрезвычайных неблагоприятных обстоятельств жизни, сложилась есте¬
ственная тенденция укрепления чувств нежности и сострадания по отноше¬
нию к новорожденному, и это был естественный способ дистанцироваться
от тех, чье желание избавиться от потомства в стремлении к более легкой
и свободной жизни оказывалось наиболее сильным.Хотя у совершенно диких народов детоубийство обычно не призна¬
валось несправедливым, оно, вероятно, практиковалось лишь в исключи¬
тельных случаях. Если, однако, по прошествии времени люди, отношение
которых к этой практике оправдывалось лишь условиями крайней дикости,
оказывались в условиях, когда накопление собственности было не только
возможным, но и естественным для более честолюбивых людей, соблазн
совершить детоубийство требовал большей реакции со стороны общества,
поэтому и соответственный отбор моральных инстинктов, сопротивляв¬
шихся этому искушению, был также очень жестким. Если мы согласимся
с тем, что все давно существующие цивилизованные народы прошли че¬
рез этот период жесткого отбора, мы сможем понять, почему в сознании
цивилизованных народов, как и в их религиозных учениях, детоубийство
должно осуждаться так решительно.Интересный пример такого переходного периода можно наблюдать
у арабов. Огромные трудности, которые испытывали бедуины, проживая
в пустыне, имели следствием множество случаев, когда детоубийство ока¬
зывалось не только экономически, но и биологически выгодным. Собствен¬
ность существовала, но была преимущественно племенной, а детоубий¬
ство в племени, совершенно очевидно, ограничивалось лишь убийством
девочек. Известны факты, что в доисламский период некоторые родители
лишь под давлением племени совершали убийство новорожденных доче¬
рей. В небольших торговых центрах условия жизни, несомненно, стали го¬
раздо лучше; и обладание собственностью, естественно, стало в большей
степени индивидуальным, хотя моральные устои все еще диктовались тра¬
дициями бедуинов. Курайшиты, племя Мухаммеда, правили Меккой при¬
мерно в течение пяти поколений до его рождения. В то время обществен¬
ное сознание начинало восставать против обычаев предков, это очевидно,
Наследование фертильности у человека203потому что маленькая группа новообращенных в ислам придавала огром¬
ное значение отказу от детоубийства, что отражено в одном из нескольких
пунктов «Первой клятвы», и этот запрет не раз повторяется в Коране. Од¬
ним из первых законов Мухаммеда после завоевания им Мекки было поста¬
новление о принесении клятвы всеми женщинами Мекки об отказе от де¬
тоубийства. Чтобы избежать неправильного представления о том, что это
вероучение было связано с военными замыслами мусульманского государ¬
ства, необходимо отметить, что «Первая клятва» была принята до бегства
Мухаммеда из Мекки, в то время, когда мысли верующих были еще очень
далеки от идеи поддержания веры силой оружия.Если признать, что селективное следствие детоубийства состояло
в том, чтобы постепенно избавиться от тех наследственных типов темпера¬
мента, нежелание которых следовать этому дикому обычаю было наимень¬
шим, то подобного следует ожидать и от равноценной практики умерщвле¬
ния плода, или аборта, когда бы она ни получала широкое распространение,
будучи продиктована благоразумием. Следовательно, история эволюции
моральных устоев в Древней Греции и Риме в тот период, когда умерщвле¬
ние плода широко практиковалось, представляет большой интерес. К сча¬
стью, поиски исторического свидетельства, которое относится к обсужда¬
емым нами теориям, не требует детального знания обычаев людей разных
эпох, но лишь превалирующего мнения о том, что является правомерным,
а что достойно порицания. Исследование самих обычаев представляло бы
значительную трудность, о чем сейчас свидетельствует наше невежество
в подобных вопросах. Обычаи разных индивидов в любой период времени,
конечно же, гораздо более разнообразны, чем отношение к данному вопро¬
су со стороны действующей морали, которое, напротив, в удивительной
степени, единообразно. Мнение большинства из тех, кто поступает непра¬
вильно с точки зрения общества в целом, естественно, должно немногим
отличаться от мнения остальной части общества. В действительности нам
редко удается находить резкие отличия в морали даже у тех, кто придержи¬
вается разных религиозных точек зрения. Следовательно, если в духовных
трудах философов, пользующихся доброй славой среди современников, мы
находим упоминания о социальных обычаях того времени, то мы вправе
сделать вывод о том, что выраженное ими мнение по этим вопросам не
являлось шокирующим для сколько-нибудь значительной части их совре¬
менников, напротив, эти упоминания должны были точно отразить общее
содержание возвышенного мнения, господствовавшего в ту эпоху.В четвертом и пятом веках до нашей эры обычай умерщвления пло¬
да, судя по всему, не считался позорным. Мы находим свидетельства того,
что возвышенные умы той эпохи, моральные наставления которых имели
204Глава 9вес, одобряли его, признавая полезным. Платон поддерживал его по сооб¬
ражениям евгеники, Аристотель рекомендовал по экономическим причи¬
нам — все это очень напоминает высказывания в защиту современных ме¬
тодов ограничения рождаемости возвышенными умами в наши дни. Через
несколько столетий взгляды общества весьма изменились. Полибий во вто¬
ром веке до нашей эры, в те времена, когда этот обычай получил такое рас¬
пространение, что это привело к сокращению численности населения, на¬
зывает этот обычай политическим пороком и явно указывает на то, что его
применение, по крайней мере состоятельными людьми, должно осуждаться
как непатриотичное поведение. «Это зло надвигается на нас стремительно,
не привлекая к себе внимания. Наши мужчины становятся развращенными,
удовлетворяя свою страсть к зрелищам, деньгам и удовольствиям праздной
жизни, и поэтому либо не женятся вообще, либо, будучи женаты, отказы¬
ваются растить детей, рожденных в браке, или воспитывают одного-двух
из всего числа тех, кто был рожден, чтобы оставить им состояние или воспи¬
тывать их в экстравагантной роскоши». Он полагает, что такие представле¬
ния необходимо изменить «по возможности самим людям путем изменения
собственных честолюбивых устремлений, а если это невозможно, то путем
принятия законов о сохранении младенцев». В первом веке нашей эры Пли¬
ний, которого мы уже цитировали, почти оправдывается за существование
этого обычая, но, очевидно, что он с пониманием относится к нему. Сенека
настроен критически. Восхваляя свою мать Хельвию, среди всех ее добро¬
детелей он особенно выделяет то, что она никогда не прибегала к обычаю
умерщвления плода, что, по его мнению, относится к порокам и безумствам
современного ему мира. Из приведенных примеров и других менее прямых
свидетельств ясно, что накануне перехода к христианству в общественном
сознании Римской империи произошли глубокие изменения. Несколькими
столетиями позже то, что казалось патриотичным и благородным для Пла¬
тона и Аристотеля, простительным для Плиния и Сенеки, было проклято
как ужасный грех отцами христианства, такими как Тертуллиан, а в законах
Валентиниана было признано преступлением, караемым смертной казнью.Изменение нравственных взглядов на умерщвление плода, а также свя¬
занные с ними перемены во взглядах на сексуальную мораль имели место
в тот период, когда древний мир переживал религиозную революцию пе¬
рехода к христианству, и поскольку в итоге эти совпадающие по времени
перемены приобрели причинно-зависимый характер, то важно понять, что
изменения в тех общественных нравах, которые мы рассматриваем, не были
следствием принятия христианского учения. И было бы фундаментальной
ошибкой вообразить, что мораль любого религиозного сообщества выво¬
дится в сколько-нибудь значительной степени из интеллектуальных поня¬
Наследование фертильности у человека205тий его теологии (независимо от стараний множества проповедников про¬
следить эту связь), и еще большей ошибкой было бы не заметить, что офи¬
циальное учение само по себе в значительной степени формируется состоя¬
нием массового сознания. Ниже следуют факты, которые являются для нас
руководящими принципами в данном вопросе.(1) Прогрессивное изменение взгляда на моральный аспект детоубий¬
ства и аборта у языческих писателей, начиная с Платона и заканчивая Юли¬
ем Павлом, который в начале III века нашей эры высказал мнение, что
с моральной стороны, хотя и не по закону, детоубийство равноценно убий¬
ству, а также резко осудил обычай умерщвления плода. Мнение указанных
писателей, которые либо ничего не знали о христианстве, либо отверга¬
ли христианское учение, объясняются совсем нехристианскими взглядами
на предмет нашего обсуждения.(2) Изложение событий так, как представлено в Евангелии, нашем са¬
мом раннем из авторитетных источников христианской доктрины, учит доб¬
роте, терпимости и самопожертвованию, которые с тех самых пор считают¬
ся христианскими идеями. Но в этом повествовании нет осуждения абор¬
та, и оно поэтому не может считаться источником теологического учения
о проклятии некрещеных младенцев, даже рожденных мертвыми, что впо¬
следствии, несомненно, оказало большое влияние на кристаллизацию хри¬
стианского мнения по этому вопросу. Хотя Евангелие и не осуждало умерщ¬
вление плода, трудно согласиться с тем, что добрый и гуманный характер
Учения в дальнейшем определил отношение церкви к аборту, поскольку
нет ничего доброго и гуманного в том учении, которое мы процитировали,
и ранние христианские законы, направленные против изнасилований, ка¬
жется, заблуждаются, оказываясь на стороне чрезмерной жестокости. Ком¬
ментируя кодекс Константина, Гиббон пишет:Удачливого насильника наказывали смертью; и поскольку простая смерть была
недостаточна для искупления его чудовищной вины, его либо сжигали заживо, либо
дикие звери разрывали его на куски в амфитеатре. Если девушка заявляла о том,
что она добровольно поддалась соблазну, это вовсе не оправдывало ее любовника,
но сама она должна была разделить его участь. Слуг, как мужчин, так и женщин,
обвиняемых в соучастии в насилии или обольщении, сжигали заживо или казнили
при помощи изощренной пытки, когда в горло заливался расплавленный свинец.Мы можем считать, что такой закон был создан людьми, испытываю¬
щими сильное смятение и ненависть в отношении сексуальной распущен¬
ности, делающими свое дело с изуверским чувством религиозного исступ¬
ления. Мы не можем приписать создание этого закона людям с либераль¬
ными взглядами, которым присуще чувство терпимости и которые были
убеждены евангельским повествованием в божественности Иисуса.
206Глава 9(3) Более строгая сексуальная мораль, включая и более резкое осуж¬
дение аборта, в действительности характеризует христианскую этику как
совершенно отличную от языческой, что объясняется тем, что христиане
взывали к сознанию индивида, и это в большей степени влияло на обще¬
ственное мнение, даже скрытое и невыраженное. Эффект был усилен вни¬
манием ранней Церкви к людям самого низкого профессионального стату¬
са, поскольку, как будет подробно показано в главе 11, эволюционные по¬
следствия вариаций репродуктивности в наибольшей степени затрагивают
людей самого низшего класса общества. В этом смысле влияние расцвета
христианства на изменения морали было глубоко личным и непосредствен¬
ным. Мы не можем рассматривать данное сопоставление в качестве фак¬
тора в установлении триумфа христианства в Римской империи, поскольку
это выходит за рамки нашей темы.Аналогии с инстинктом животных и иммунитетом
к болезнямСторонний наблюдатель вряд ли поймет, насколько степень эволюци¬
онного развития, которое претерпели репродуктивные инстинкты челове¬
ка, значительнее аналогичных модификаций, которым, несомненно, под¬
верглись репродуктивные инстинкты других животных в их эволюционном
развитии. Основное отличие, возможно, состоит в том, что процесс отбо¬
ра у Человека происходит очень быстро, и, соответственно, так же быстро
идет эволюционный прогресс, несмотря на большую продолжительность
поколения у человека. Эти нравственные нормы настолько важны для нас
самих и, по-видимому, обладают такой безусловной обязательной силой,
что нелегко сразу же занять позицию стороннего наблюдателя; и все же лю¬
бое внимательное рассмотрение сопряженности признаков у высших жи¬
вотных должно убедить нас в том, что их нравственная природа во мно¬
гих случаях также должна была быть модифицирована строго аналогичной
формой отбора. Многие, обычно робкие, виды млекопитающих и птиц про¬
являют исключительную храбрость, защищая свое потомство, и даже самки
хищников воздерживаются от детоубийства, это как раз те случаи, когда мы
по крайней мере можем быть уверены в том, что эмоциональная реакция
на ситуацию, связанную с репродуктивностью, была модифицирована так
же, как модифицировалась эмоциональная реакция человека на определен¬
ные нравственные эмоции, необходимые, чтобы человек смог преодолеть
специфические пороки, которым он подвержен.Еще более близкая аналогия может быть прослежена в эволюции им¬
мунитета к определенным болезням и болезненным пристрастиям, таким
Наследование фертильности у человека207как алкоголь, через селективное действие смертности, подобно тому, как
развивалась невосприимчивость к сексуальным порокам через селективное
действие репродуктивности. В случае болезней, я полагаю, принято счи¬
тать, что особый, или специфический, иммунитет должен быть приобретен
к каждой болезни отдельно и последовательно. С другой стороны, я думаю,
что невосприимчивость к грубым порокам, таким как детоубийство, обыч¬
но приобретается через умственные модификации, которые сами по себе
имеют цель предоставить некоторую степень невосприимчивости в отно¬
шении более коварных соблазнов сделать аборт или использовать контра¬
цептивные средства. По крайней мере некоторые данные демографической
статистики наводят на мысль о том, что если бы можно было провести
исследования при одинаковых условиях и в одной и той же среде, то выяс¬
нилось бы, что плодовитость народов, раньше всех вступивших на путь ци¬
вилизации или, скорее, тех групп, общие предки которых прожили в циви¬
лизованных условиях большую часть времени, является наибольшей. В то
время как те, что стали цивилизованными позже всех, наименее способны
поддерживать свою численность в цивилизованных условиях. Например,
в статистике материнства в штате Род-Айленд, по данным доктора Хофф¬
мана, доля женщин, которые вышли замуж во время переписи (1905) и были
все еще бездетными в то время, представлена в соответствии с вероиспо¬
веданием — протестантским, католическим или иудейским, а также еще
по двум параметрам — те, кто родился в Америке, и те, кто был рожден
за границей. Процентные соотношения представлены в таблице 10.Контраст между родившимися в Америке и за границей, несомненно,
относится к разнице в социальном классе и сопоставим с профессиональ¬
ными различиями; более детально он будет рассматриваться в следующей
главе. Религиозные различия могут, вероятно, быть отнесены на счет пря¬
мого влияния религиозной доктрины в отношении практики контрацепции,
которая осуждается религиозными главами как католического, так и иудей¬
ского сообществ. Более вероятным представляется то, что различия связаны
с расовым происхождением, а с этой точки зрения интересно, что расовая
группа протестантских народов должна была стать цивилизованной отно¬
сительно недавно, в то время как иудеи были в большей степени подверже¬
ны влиянию цивилизации со времен, предшествовавших даже классической
цивилизации Средиземноморья.Один лишь факт, что люди постепенно начинают испытывать анти¬
патию к какому-то конкретному действию, такому, например, как аборт,
в связи с его влиянием на пропорциональное выживание разных индивидов
среди предков последующих поколений, проливает мало света на умствен¬
ные процессы, которыми эта антипатия была вызвана. Будучи причиной
208Глава 9Таблица 10РожденыРожденыв Америкеза границейПротестанты30,719,4Католики24,216,9Иудеи18,911,4таких изменений, Естественный Отбор не объясняет их способов, посколь¬
ку основной его задачей является достижение требуемых характеристик
эффективной репродуктивности. Китайское убеждение относительно дето¬
убийства, которое заключается в том, что духи мертвых младенцев при¬
носят неудачу, указывает на метод модификации репродуктивных инстинк¬
тов путем предпочтительного выживания более доверчивых и суеверных.
Вполне можно предположить, что источник такого убеждения должен быть
обнаружен во внутренней тенденции испытывать чувство вины, совершив
детоубийство; но когда такое убеждение воспитано и повсеместно приня¬
то, его существование, несомненно, модифицирует область отбора в тех
особых умственных качествах, которые пользуются поддержкой. Хотя про¬
грессивное усиление репродуктивных инстинктов, которое можно считать
главным следствием репродуктивного отбора, будет стабильной целью, раз¬
нообразие психологических модификаций вместе с последующими социо¬
логическими изменениями может быть в разных обстоятельствах эффек¬
тивным средством достижения такой цели. В качестве основного следствия
первичного отбора, направленного против сознательного и произвольного
действия, удобно рассматривать развитие антипатии к этому действию, в то
время как другие интеллектуальные отношения, которые приводят к тому
же эффекту путем стремления к иллюзорным целям, создают вариант ми¬
микрии подходящей морали. Какой тип ментального отношения окажется
на практике самым эффективным и впоследствии будет подвержен наибо¬
лее быстрой модификации, должно зависеть частично от степени, с которой
знание естественной обусловленности распространено среди данного народа.Сознательный выбор родителей в отношении репродукции наравне
с их моральным отношением к доступным методам ограничения семьи яв¬
ляется мотивом, для которого данное ограничение приемлемо. Родители,
которым присущи целеустремленность в осуществлении своих финансо¬
вых замыслов и расчет, будут в подобных обстоятельствах более склонны
к тому, чтобы ограничить размер семьи, чем те, у кого данные качества
развиты в меньшей мере. Следовательно, постепенное ослабление влия¬
ния устремлений экономического характера или по крайней мере средняя
интенсивность влияния данных мотивов на большинство граждан должны
Наследование фертильности у человека209являться сопутствующим обстоятельством к усилению чувства антипатии
к ограничению численности семьи. Отношение мужчин и женщин к соб¬
ственному благополучию не может тем не менее быть по этой причине
преобразовано в некое равнодушие, поскольку в тех странах, где самый
бедный класс часто истребляется голодом, очевидно, будет достигнута сту¬
пень, на которой то, что выиграно в уровне рождаемости, будет утраче¬
но в силу высокой смертности. И даже там, где крайней степени нужды
обычно удается избежать, некоторая утрата гражданской свободы и воз¬
можностей для репродукции является обычным состоянием бедности. Бо¬
лее того, рациональная погоня за экономическим преимуществом должна,
даже в самых цивилизованных странах, часто ставить индивида в поло¬
жение, благоприятное для нормальной репродукции. В большей части со¬
обществ, по всей видимости, полная нехватка стяжательских инстинктов,
как и дефицит основных побуждений к сексу, будут в равной мере разру¬
шительными для биологических перспектив индивида, несмотря на то, что
злоупотребление любой страстью должно приводить к встречному отбору.
Моральное отношение цивилизованных людей к деньгам, как и к сексу,
должно быть, следовательно, продуктом гораздо более сложных эволюци¬
онных сил, чем является их отношение к детоубийству или аборту; это от¬
ношение, наверное, можно вывести из лицемерия и фанатизма, страстных
увлечений и необузданных запретов, которые обнаруживаются среди давно
цивилизованных народов. Мы можем лишь догадываться об одной из при¬
чин дурной славы морали, повсеместно связываемой с корыстными моти¬
вами. Эта дурная слава сама по себе странным образом вступает в контраст
с экономической теорией относительно того, что плата является обычным
эквивалентом профессий, открыто признаваемых заслуживающими такой
оплаты; однако пока в нашем обсуждении нет ничего, что могло бы по¬
казать, почему превалирующее отношение к богатству должно отличаться
от того, о котором говорилось в конце предыдущей главы, где было выдви¬
нуто предположение, что обычная судьба накопленного состояния должна
была предусматривать многочисленное потомство.РезюмеНа число детей, в действительности рождаемое разными индивидами
в цивилизованных обществах, как и другими организмами, основное вли¬
яние оказывает случай. В то время как у большинства диких организмов
в природе вклад других причин в существующие вариации рождаемости
является, вероятно, таким незначительным, что его нелегко обнаружить,
общая дисперсия численности потомства, производимого цивилизованным
210Глава 9человеком, так велика, что значительная часть ее должна быть приписана
причинам, отличным от случайности.Свойства темперамента как у мужчин, так и у женщин оказывают боль¬
шое влияние на выбор в пользу безбрачия или на возраст вступления в брак.
Они являются и на протяжении всей истории человечества были по мень¬
шей мере в равной степени важными для использования или воздержания
от использования искусственных методов ограничения численности семьи.Число детей, рождаемых женщинами, в значительной мере соотносит¬
ся с числом детей, рожденных их матерями. Статистические данные по выс¬
шим социальным классам девятнадцатого века предполагают, что пример¬
но 40 % общей дисперсии можно объяснить наследственными причинами.
Из наследственных причин наиболее важными должны считаться наслед¬
ственные особенности ума, и особенно темперамент. Интенсивность отбо¬
ра по различиям в фертильности в силу внутренних причин обнаруженного
порядка относительно велика в сравнении с интенсивностью отбора, кото¬
рую следует ожидать в природе. Несмотря на позднюю зрелость человека,
этого достаточно, чтобы обеспечить значительные эволюционные измене¬
ния за относительно короткие исторические периоды.Обычай детоубийства в городах доисламской Аравии сопровождал¬
ся сменой морали, что в конце концов привело к запрету этого обычая.
Широкое применение практики оставления младенцев на произвол судьбы
и практики аборта в ранний период греко-римской цивилизации сопровож¬
далось постепенной и настоятельной переменой в общественном мнении,
позднее они были провозглашены преступлениями, караемыми смертной
казнью. Влияние христианского учения на отказ от этих взглядов было
очень большим, но это влияние шло скорее от сознания людей, чем от дог¬
матического учения.Можно считать, что расы стали невосприимчивы к разрушительному
действию пороков подобно тому, как посредством селективной смертности
приобретается иммунитет к заболеванию. В данных современной статисти¬
ки имеется свидетельство того, что наиболее устойчивыми даже к совре¬
менным методам ограничения рождаемости в этом отношении являются
те расы, которые имели самый длительный социальный опыт цивилизован¬
ных условий жизни.Рассмотренные факты оставляют мало места для сомнения, что са¬
мой мощной силой отбора у цивилизованного человека является воздей¬
ствие на умственные и моральные качества посредством изменения уровня
рождаемости, и что это средство работает с той интенсивностью, которой
нелегко найти соответствие у других видов.
Глава 10Репродуктивность и социальный классЭкономические и биологические аспекты классовых различий. Неточности те¬
кущих данных. Начало исследований. Британские данные. Положение в США. Эф¬
фекты дифференциальной фертильности. Резюме.А ты не оставляешь существа,В котором свет нашел бы утешенье. У. ШЕКСПИР*Экономические и биологические аспекты
классовых различийДля биологического изучения человечества институт социального
класса имеет двойное значение. Если рассуждать только с экономической
точки зрения, то разные профессии отличаются степенью вознаграждения,
которое они обеспечивают тем, кто ими владеет, и следует предположить,
что социальная значимость выполняемой работы также различна. С этой
точки зрения класс общества, к которому принадлежит та или иная се¬
мья, может рассматриваться как граница благополучия, отделяющая семью
от нужды или по крайней мере от полуголодного существования; причем
сравнительная неудача в выборе одной профессии позволяет за счет неко¬
торой потери в социальном классе найти более скромный образ жизни,
при котором имеющиеся способности достаточны, чтобы обрести необхо¬
димую поддержку для себя и своей семьи. Возможность адаптироваться
к изменению ситуации, которую дают некоторые профессии (в том числе
и благодаря разницам в их оплате), уменьшает тяжесть последствий неудач¬
ного выбора для отдельного индивида, так как у большей части населения
остается определенный запас прочности, спасающий от полной нужды.Аспект социального класса, который имеет более прямое отноше¬
ние к биологическим исследованиям, — это глубокое влияние, оказываемое
им на брак между представителями разных семей. В некоторых случаяхСонет 9, в переводе С. Я. Маршака.
212Глава 10уклонению от брака между представителями разных классов придавалось
такое большое значение, что вступление в подобный брак преследовалось
по закону, и, следовательно, возникали закрытые в большей или меньшей
степени аристократические касты. Особенно это отмечено в тех случаях,
когда в дело вмешивались еще и расовые различия. Но при отсутствии та¬
кой правовой нормы во всех цивилизованных странах общественное мне¬
ние, возможность общения с представителями своего класса, которые вы¬
зывают друг у друга взаимный интерес, привели к тому, что подавляющее
число браков совершалось только между представителями близких соци¬
альных классов. Под социальным классом здесь подразумеваются, конеч¬
но, не только люди с примерно одинаковым доходом и благосостоянием,
но и престижность их профессии, их личные способности и семейные свя¬
зи. Таким образом, данное определение термина «социальный класс» хотя
и близко к тому, которое употребляется в экономике, все же немного от него
отличается. Однако фактор смешанных браков настолько важен в смысле
его социальных и биологических последствий, что лучше всего будет ис¬
пользовать термин «социальный класс» лишь в указанном смысле и пола¬
гать, что социальный класс индивида или его семьи должен определяться
совокупностью людей или семей, заключение брака с которыми не встретит
социальных препятствий.Вследствие влияния класса на выбор партнера отличия социального
класса — это отличия относительно постоянных биологических сообществ;
между данными сообществами в действительности не существует жестких
разграничений, но они влияют одно на другое путем диффузии представи¬
телей одного класса в другой класс. Эту взаимную диффузию необходимо
отличать от огромного потока людей, которые движутся вверх по социаль¬
ной лестнице. Можно заметить, что этот процесс происходит повсеместно
и в результате влияния одного класса на другой посредством диффузии
увеличивается. Эти два социальных движения, конечно же, смешиваются
в жизни каждого отдельного индивида, но их причины и следствия необ¬
ходимо четко различать. Относительно постоянный характер социальных
классов делает очень важными те различия, которые существуют в демогра¬
фической статистике, поскольку, чтобы произошли эволюционные измене¬
ния, необходимо, чтобы группы, различающиеся, скажем, по уровню репро¬
дукции, также изменились генетически; и если это изменение будет посто¬
янно увеличиваться, необходимо также, чтобы подобные различия в репро¬
дукции продолжались из поколения в поколение, и тогда можно будет опи¬
сать группы, проявляющие одни и те же генетические противоположности.Тот факт, что между уровнем рождаемости в определенных высокопо¬
ставленных семьях и уровнем рождаемости в семьях остального населения
Репродуктивность и социальный класс213существует непонятное расхождение, был известен в течение большей ча¬
сти девятнадцатого века; но до недавнего времени не проводилась оценка
ни величины, ни географического распространения, ни социологической
природы этого феномена. Теория неизвестного происхождения, суть кото¬
рой сводится к тому, что «инбридинг», который практиковался, как предпо¬
лагалось, в аристократических семьях, приводил к стерильности (и другим
признакам вырождения), сформировалась на основе противоречия, что здо¬
ровые плебейские семьи были так же фертильны, как и остальное сообще¬
ство. Теория Герберта Спенсера о врожденном противоречии «индивидуа¬
лизации» и «происхождения», кажется, предполагала, что он признал ха¬
рактерным некоторое снижение уровня репродуктивности для работников
умственного труда. Один из наиболее важных моментов, согласно совре¬
менным данным, это то, что дефицит в производстве потомства не является
отличительной чертой титулованных семей или семей с высоким интеллек¬
том супругов. Это — тенденция к понижению рождаемости, охватывающая
всю социальную лестницу сверху донизу.Неточности текущих данныхПрежде чем рассматривать некоторые из наиболее важных исследова¬
ний в данной области, необходимо отметить, что хотя и можно с уверенно¬
стью сказать, что в большинстве западных стран эти явления имеют призна¬
ки, однако фактических количественных данных намного меньше, чем мог¬
ло бы быть, и это обусловлено двумя главными причинами. Во-первых, они
сильно устарели. По некоторым параметрам они дают информацию о рож¬
даемости в семьях 35-летней давности. А ведь современные процессы мы
оцениваем, основываясь на анализе процессов, происходивших в прошлом.
Самая последняя общая информация по Англии и Шотландии — это сведе¬
ния о полных семьях, которые были получены во время переписи 1911 года.
Многие из детей, на которых основываются данные сравнения, родились,
следовательно, около 1890 года и представляют поведение, скорее, прошло¬
го, чем нынешнего поколения родителей. Такой промежуток времени мо¬
жет иметь большое значение для изменения уровня рождаемости, посколь¬
ку бесспорно, что какие бы изменения ни произошли в уровне рождаемо¬
сти разных классов, уровень рождаемости по стране в целом очень сильно
снизился за данный период. Хотя вопросы могут повторяться в переписи
1935 года, понятно, что сравнения, основанные на ответах полных семей,
будут иметь к тому времени, как они станут достоянием общественности,
уже двадцатилетнюю давность. Точную информацию относительно ежегод¬
ного естественного прироста населения можно тем не менее получить, имея
214Глава 10данные о количестве зарегистрированных рождений. Для этого необходи¬
мо, чтобы при регистрации рождений записывался и возраст родителей,
а род деятельности отца (сейчас эти данные фиксируются) был бы класси¬
фицирован по системе, которая применяется при переписи населения.Во-вторых, удовлетворительное сравнение разных групп профессий
должно включать информацию о смертности в указанных группах, а кро¬
ме того, учитывать не только уровень рождаемости у женатых и замужних
людей заданного возраста, но также возраст вступления в брак и часто¬
ту безбрачия. Основу для полного биологического сравнения фактически
можно найти лишь в мальтузианском параметре роста населения, о кото¬
ром говорилось в главе 2; следовательно, только когда будут установлены
мальтузианские параметры для разных групп профессий, будет получена
количественная оценка биологической ситуации. Во всех следующих ни¬
же примерах видно, что наши знания далеки от этого уровня. Возможно,
меньшее значение имеет факт игнорирования уровня смертности, исключая
ранний детский возраст, потому что это не является основным фактором
сравнения. Даже в отношении рождений почти все исследования имели
тенденцию ограничиваться сравнением рождений у женатых и замужних
людей в заданном возрасте, и часто данные собирали таким способом, что
пренебрегали, насколько возможно, важными факторами возраста вступле¬
ния в брак и безбрачия. Конечно, желательно, чтобы эти три элемента опре¬
делялись отдельно, и есть причины полагать, что их отдельная оценка ока¬
жется полезной для понимания причин этого феномена. Все же, поскольку
ни один фактор сам по себе не является на практике таким же важным,
как все факторы в совокупности, если сводить данные в таблицу, в которой
будет отсутствовать итоговый результат, то исследование потеряет больше,
чем приобретет.Начало исследованийОдним из самых значительных первых исследований было исследо¬
вание, проведенное доктором Дэвидом Героном в разных муниципальных
районах, на которые разделен Лондон. Его метод состоит в соотнесении
фертильности с рядом показателей социального статуса, извлеченных из ко¬
личественного соотношения разных профессий, зарегистрированных в пе¬
реписи. Районы, в которых средний социальный уровень является самым
высоким, отмечены большой долей людей свободных профессий и домаш¬
них слуг и низкой долей разнорабочих, ростовщиков, детей, работающих
по найму, и арендаторов, проживающих в среднем свыше двух человек
в комнате. Фертильность была измерена в 1901 году путем оценки рождае¬
Репродуктивность и социальный класс215мости у тысячи замужних женщин в возрасте от 15 до 54 лет ив 1851 году
у тысячи замужних женщин в возрасте старше 20 лет. Рождаемость ока¬
залась тесно связанной со всеми признаками низшего социального класса.
Оба периода показывают в целом одно и то же соотношение между нежела¬
тельными социальными условиями и высоким уровнем рождаемости; ин¬
тенсивность этого соотношения почти удвоилась в интервале между 1851
и 1901 годами. Исследование представляет особенный интерес в том отно¬
шении, что в 1851 году классовые различия в фертильности уже развились,
хотя лишь наполовину от их интенсивности в конце века; однако сравнение
интенсивности было несколько затруднено тем изменением, которое доктор
Герон был вынужден внести в метод измерения фертильности, поскольку
немного более низкий индекс, использованный для 1851 года, и сам, воз¬
можно, уменьшал величину полученных корреляций. Следует помнить, что
сравниваются только показатели по районам, и полученный результат срав¬
нения свидетельствует лишь о том, что в Лондоне самые бедные районы
имеют самую высокую рождаемость. Это не доказывает, что самые бед¬
ные классы сами ответственны за высокий показатель рождаемости, хотя
наличие такой обобщенной связи является естественным выводом из по¬
лученных результатов. Однако используемые методы не позволяют судить
о причине различий, является ли причиной то, что рабочий, которому боль¬
ше платят, менее плодовит, чем тот, которому платят меньше; или то, что
работники свободных профессий и служащие, получающие жалованье, ме¬
нее плодовиты, чем работающие классы вообще. Различия, наблюдаемые
в данном исследовании, конечно, не включают различий в доле безбрачия
или в средней продолжительности браков до 45 лет.Первые национальные статистические данные, в которых размер семьи
мог сравниваться с профессиональным статусом, были получены во фран¬
цузской переписи 1906 года. Для тех семей, в которых главе семьи на мо¬
мент переписи было 60-65 лет и который в это время все еще состоял
в браке, среднее значение для размера семьи составило 3,60. Для нанима¬
телей (patrons) это среднее значение было 3,59, что почти равнялось сред¬
нему значению для населения в целом; для рабочих (ouvriers) соответству¬
ющее среднее значение было 4,04, в то время как среди других служа¬
щих (employes), которые не являлись рабочими, оно составило только 3,00.
Patrons больше чем наполовину были фермерами, для которых среднее зна¬
чение размера семьи для тех, кто был женат более 25 лет, было 3,71, и лишь
немногим больше их пятой части принадлежало к свободным професси¬
ям, для которых соответствующее среднее значение было 3,30 или немного
меньше, чем у employes, если ввести поправку на изменение базы класси¬
фикации.
216Глава 10Классификация в переписи тем не менее лучше подходила для срав¬
нения условий в разных отраслях (промышленности), чем для сравнения
в пределах одной отрасли. Новые данные были получены в 1907 году по¬
сле изучения налоговых деклараций, заполненных большим количеством
служащих и рабочих в стране, в разных провинциях и приходах. В сред¬
нем браки, продолжавшиеся более пятнадцати лет, показали для рабочих
стабильное снижение размеров семьи, начиная с тех, чей труд оплачивал¬
ся хуже всего — в среднем 3,29, — до тех, кто получал самую высокую
оплату, зарабатывая £ 160-240 в год, со средним значением 2,34. Служа¬
щие в тех же группах, классифицированных по уровню оплаты, показыва¬
ют соответствующее падение с 2,77 до 2,31. Те, кто зарабатывал £240-400
в год, демонстрируют дальнейшее снижение до 2,29, в то время как те,
что зарабатывал более £400 в год, демонстрируют подъем до 2,38. Из этих
и ранее приведенных данных ясно, что в целом для французского населе¬
ния, зарабатывающего менее £400 в год, репродуктивность в предыдущий
период, по меньшей мере в восьмидесятые-девяностые годы девятнадца¬
того века, была гораздо выше среди более бедного населения, чем среди
слоев населения с большим доходом. У служащих, зарабатывающих более
£400 в год, величина, на которой классификация по уровню оплаты закан¬
чивается, очевидно, наблюдался некоторый подъем уровня репродукции,
хотя у более преуспевающих классов интеллектуальная группа, представ¬
ляющая свободные профессии, имела гораздо меньше детей, чем среднее
значение.Самая полная доступная информация по Великобритании получена
в результате переписи 1911 года, но данные многих неофициальных ис¬
следований выявили факты огромной важности. Г-н Уитхем в 1909 году
опубликовал результаты исследования (Family and Nation, p. 140) фертиль¬
ности высших слоев специалистов (врачи, адвокаты, учителя) и должност¬
ных лиц, в котором он использовал информацию справочника Who s Who.
По его данным, среднее число детей, зарегистрированных у людей данного
класса, состоящих в браке, уменьшается с 5,19 у тех, кто вступил в брак
до 1870 года, до 3,08 у тех, кто вступил в брак после 1870 года. Он отмечает,
что для служащих среднее значение уменьшается в гораздо меньшей степе¬
ни, а для семей военных падение значительно больше, чем для остальных.
Другое исследование того же автора показывает, что для семей, которые
уже по меньшей мере три поколения относятся к сословию пэров, среднее
значение упало в соответствии с датой вступления в брак с 7,10 для браков,
заключенных в тридцатые годы девятнадцатого века, до 3,13 для тех, кто
заключил брак в восьмидесятые годы. Хотя данные исследования ограниче¬
ны малой частью британского населения, они обнаруживают существенные
Репродуктивность и социальный класс217факты; во-первых, эти факты свидетельствуют о низком уровне репродук¬
тивности английских высших классов последнего поколения и, во-вторых,
о том, что данный уровень репродуктивности падал на протяжении второй
половины девятнадцатого века со скоростью, намного превосходящей спад
фертильности у населения в целом.Иногда полагают, что различия в репродуктивности, наблюдаемые
в разных социальных классах, зависят от обстоятельств и идей, свойствен¬
ных тем, кто обладает преимуществами свободного времяпровождения или
образования или того и другого вместе. Многие думают, что, вероятно,
там, где нужда доходит до крайности, самые здоровые и способные бу¬
дут воспроизводить свое потомство наиболее свободно, как и дикие живот¬
ные в природном состоянии. Какой бы ни была причина в прошлом, тща¬
тельно проведенное и наиболее убедительное исследование показало, что
данные по низшим классам в промышленных городах Блэкберн, Престон,
Глазго, Бирмингем, а также Ройял-Алберт Эсайлем, Лэнкестер доказывают,
что наиболее одаренные среди указанных классов имеют наименьшее число
детей. Полученные коэффициенты корреляции невелики, как обычно и бы¬
вает, когда имеешь дело с вариациями внутри популяции примерно одного
культурного уровня. Если ввести соответствующую поправку на возраст ро¬
дителей, то получится, что семьи с большим числом членов живут на мень¬
шую заработную плату. Другими словами, чем меньше зарабатывают роди¬
тели в данном возрасте, тем большее количество детей выживает. Сходные
результаты получены, когда критерии умственных способностей или об¬
щественного положения, такие как здоровье, постоянная работа, трезвость
отца семейства и порядок в доме, были заменены на критерий уровня за¬
работной платы. Из всех указанных критериев наименее надежным был
критерий чистоты в доме, поскольку возможно, что проводящие исследо¬
вания, выставив оценку указанному признаку, не сделали точной поправки
на дополнительную работу по дому вследствие большего размера семьи.Британские данныеРезультаты переписи 1911 года, о которых речь шла выше, раздельно
по Шотландии и Англии были представлены доктором Данлопом и док¬
тором Стивенсоном соответственно. Эти два сообщения значительно отли¬
чаются по форме, в которой представлены результаты, и поскольку дан¬
ные двух опросов очень похожи, то каждый служит дополнением другого
в нескольких важных моментах. Для Шотландии доктор Данлоп приводит
сравнительную таблицу среднего числа детей для большого числа профес¬
сий. Были использованы лишь данные о браках, продолжительность кото¬
218Глава 10рых составляет пятнадцать и более лет, а возраст вступления в брак же¬
ны ограничивался пятью годами, с 22 до 26. Среднее для всех профессий
составило 5,82 детей, и, проранжировав профессии по среднему размеру
семьи, легко сравнить социальный статус наиболее фертильных с социаль¬
ным статусом наименее фертильных профессий. Чтобы избежать придания
чрезмерного веса маленьким группам, для которых среднее значение фер¬
тильности не будет определено очень точно, нам следует учитывать число
семей, для которых вычислено соответствующее среднее значение. Наи¬
более фертильные профессии составляют три большие группы, а именно:
мелкие арендаторы (фермеры) — 7,04; шахтеры, добывающие уголь, сланец
и железную руду,— 7,01, сельскохозяйственные рабочие и слуги на фер¬
мах — 6,42; три группы среднего размера: пенсионеры по старости — 6,95;
рыбаки — 6,93 и грузчики и возчики угля — 6,61; и, кроме перечисленных
групп, есть еще одна маленькая группа — это помощники штукатуров —
7,01. Обратившись к наименее фертильным профессиям, мы увидим одну
большую группу, которая состоит из служащих — 4,38. Шесть групп сред¬
него размера представляют врачи и хирурги — 3,91; адвокаты и солисито¬
ры — 3,92; писатели и ученые, школьные учителя и воспитатели — 4,25;
работники искусства, музыканты и артисты — 4,27; священники — 4,33.
В дополнение к этому есть еще три маленьких группы: это армейские офи¬
церы — 3,76; дантисты и их помощники — 3,86; и ветеринары — 4,00. Разли¬
чия между социальным статусом этих двух наборов профессий не требует
особого акцентирования. Среди наименее фертильных оказываются про¬
фессии умственного труда, а среди наиболее фертильных — самые бедные
из работников физического труда. Различия в среднем числе детей заслу¬
живают еще большего внимания вследствие жесткого ограничения, касаю¬
щегося возраста матери. Вместе с ограничением отбора по продолжитель¬
ности брака это практически эквивалентно устранению всех тех различий
в фертильности, существующих между разными профессиональными клас¬
сами, которые возникают из-за разницы в частоте безбрачия и в возрасте
заключения брака. Из соответствующих данных по Англии четко видно, что
контраст между реальными уровнями репродуктивности в разных классах
усиливается, если учитывать эти дополнительные факторы.Результаты переписи особенно ценны тем, что демонстрируют следу¬
ющее: спад фертильности с улучшением условий характерен не для одной
единственной социальной лестницы, как казалось на основании француз¬
ских данных, но равно проявляется, начиная с беднейших групп, которые
оказывается возможным сравнивать, и заканчивая наиболее отличающи¬
мися профессиями, для которых имеется достаточное число наблюдений
для сравнения.
Репродуктивность и социальный класс219Более того, в некоторых случаях удается сравнить фертильность групп,
которые являются, собственно, подгруппами одной большей профессио¬
нальной группы. Такое сравнение представляет особенную ценность, так
как при этом устраняются различия по местоположению и индустриаль¬
ной среде, в отсутствие этих различий средние значения, полученные даже
для самых маленьких профессиональных категорий, становятся значитель¬
но более существенными и регулярными. Так, три пары категорий в списке
доктора Данлопа, которые, по-видимому, сравнивались очень тщательно,
показывают нам, что у помощников каменщиков больше детей, чем у ка¬
менщиков, у сельскохозяйственных рабочих больше детей, чем у старших
рабочих, а у помощников штукатуров больше, чем у штукатуров. Поскольку
в каждом из перечисленных сравнений все вариации условий среды и ме¬
стоположения представлены строго пропорционально, то данные сравнения
отражают разницу в фертильности наиболее ясно. После 1911 года клас¬
сификация по роду занятий, используемая в переписи, была существенно
улучшена. Наиболее удачным было отделение классификации по отраслям
промышленности от классификаций внутри каждой отрасли; и если иссле¬
дование, касающееся числа детей, повторить в переписи 1931 года, то будет
возможно провести гораздо больше сравнений между разными квалифика¬
циями рабочих в одной и той же отрасли промышленности.В 1920 году доктор Стивенсон опубликовал отчет о результатах по Ан¬
глии и Уэлсу, в котором особое внимание уделялось преодолению трудно¬
стей при анализе прямой социологической ценности данных, сбор которых
был спланирован весьма несовершенно. Профессиональные категории, хо¬
тя и включали отличительные особенности промышленности и характер¬
ного рода занятий, можно было грубо разделить на пять больших разде¬
лов, первый из которых представлял высшие и средние классы в самом
общем смысле этого выражения, третий — квалифицированных рабочих,
а пятый — неквалифицированных рабочих. Вследствие отсутствия града¬
ций внутри групп пришлось исключить из этого перечня три большие груп¬
пы: рабочих текстильной промышленности, шахтеров, добывающих уголь,
и сельскохозяйственных рабочих.В семьях, в которых способность к воспроизведению потомства со¬
хранялась, т. е. где жене было меньше 45 лет на момент переписи, сред¬
нее число детей для указанных пяти групп составило соответственно 1.68;
2.05; 2.32; 2.37 и 2.68; неквалифицированные рабочие имели в среднем по¬
чти в два раза больше детей, чем семьи первой группы. Самый большой
контраст наблюдается между I и II классами, где разница составила 0,37,
и классами IV и V с разницей 0,31; налицо явные различия в фертиль¬
ности, связанные с низшими, а также с высшими рангами способностей.
220Глава 10Приведение к стандарту в отношении возраста вступления в брак и про¬
должительности брака снижает разницы примерно на треть, демонстрируя
тот факт, что наиболее фертильные классы при вступлении в брак в опре¬
деленном возрасте обеспечивают самую большую возможность по фер¬
тильности, вступая в брак в раннем возрасте. Из отчета о переписи также
следует, что доля заключивших брак в категории специалистов (адвокаты,
врачи, учителя) меньше, чем у остальной части населения. Следовательно,
неравенство в репродукгивности возникает из трех основных источников.
Во-первых, высшие классы реже заключают брак или чаще остаются холо¬
стыми; во-вторых, они заключают брак позже, и репродуктивный период их
жен в браке оказывается короче и приходится на последний, менее репро¬
дуктивный отрезок данного периода; в-третьих, число зарегистрированных
детей меньше по сравнению с числом жен в любом данном возрасте. Тре¬
тья причина набирает силу с недавнего времени и, в основном, должна
быть отнесена на счет планируемого ограничения рождаемости; но следу¬
ет отметить, что данное объяснение не сильно сказывается на первых двух
причинах, влияние которых скорее было бы ослаблено, нежели усилено
возрастающим использованием контрацепции.Доктор Стивенсон попытался проследить раннюю историю относи¬
тельной фертильности разных классов, сравнивая средние семьи в разных
профессиональных группах отдельно для браков, заключенных в последо¬
вательные десятилетия до 1851-1861 гг., в которых к переписи 1911 года
выжило значительное число супругов. Профессиональные различия фер¬
тильности в этих ранних браках были значительно меньше, и доктор Сти¬
венсон предположил, что если бы сравнение было проведено еще на 20 лет
раньше, то можно было бы обнаружить период реального равенства меж¬
ду разными классами. Однако данных одного единственного исследования
явно недостаточно, чтобы получить необходимую информацию по этому
вопросу, поскольку данные о браках, на которых базируются средние зна¬
чения переписи, собраны не по случайному принципу, по которому могли
бы быть собраны данные ранней переписи, а строго отобраны на осно¬
ве долголетия. В данных среднего по происхождению класса повсеместно
можно найти существенную корреляцию между фертильностью и долго¬
летием, и обоснованность более ранних данных может быть по этой при¬
чине в существенной степени сомнительной. Аргумент с позиции конвер¬
генции требует не только того, чтобы классификация по роду занятий для
самых старших групп населения была так же надежна, как и для тех, кто
все еще работает, но и того, что влияние отбора по долголетию в связи
с фертильностью долгожителей должно быть практически одним и тем
же во всех классах. Из ряда сравниваемых исследований некоторая по¬
Репродуктивность и социальный класс221добная экстраполяция должна быть возможной, но поскольку такая воз¬
можность была утрачена в 1921 году, то классовые различия в репродук¬
тивности перед общим падением рождаемости около 1875 года едва ли
возможно теперь восстановить. Но из работы Герона, которую мы цити¬
ровали ранее, ясно, что между районами Лондона до 1851 года существо¬
вали значительные различия. Поскольку, таким образом, период классово¬
го равенства в данном вопросе довольно гипотетичен, то рост неравен¬
ства с середины девятнадцатого века не может быть поставлен под сом¬
нение.Общее снижение рождаемости с семидесятых годов оказало влияние,
хотя и в разной степени, на все классы. Снижение рождаемости было
в большей степени выражено у преуспевающей части населения, и в мень¬
шей степени — у бедных классов. Статистического обоснования для утвер¬
ждения, что падение «распространилось сверху вниз», по-видимому, не су¬
ществует. Это невозможно и по своей природе, поскольку с момента воз¬
никновения неомальтузианского движения оно вело пропаганду своих идей
среди беднейших слоев населения. Широко распространено убеждение,
и на это следует обратить внимание, что дифференциальная рождаемость
в той степени, в которой она возникает вследствие планируемого ограни¬
чения рождаемости, автоматически исчезнет из-за развития обычного про¬
цесса принятия бедными классами практики, распространенной среди бо¬
лее преуспевающих слоев населения. Демографическая статистика нашей
страны не дает оснований для такого мнения.Положение в СШААмериканские данные о связи репродуктивности и социального клас¬
са напоминают данные, полученные по Великобритании, демонстрируя ос¬
новные особенности данного явления с несомненной ясностью и не предо¬
ставляя возможности проникновения в какие-либо количественные детали.
Официальным методом является учет числа детей у родителей при реги¬
страции новорожденного, но его результаты нельзя сравнивать с результа¬
тами метода, при котором учитывается размер семьи при переписи. Одна¬
ко, среднестатистические семьи, где матери входят в возрастную группу
35^44, должны представлять данные, приблизительно одинаковые в разных
классах, и, как в шотландской переписи, можно отметить, что профессио¬
нальные группы дают и высокие и низкие средние значения. Следующие
списки профессий взяты из работы Builders of America Уитни и Хантинг¬
тона, в которой средние значения были снижены на треть, чтобы получить
оценки числа детей в среднем, рожденных в полных семьях. Такая поправ¬
222Глава 10ка не повлияла на ранжировку профессий. Год, о котором говорят Уитни
и Хантингтон, точно не определен, но близкие параллельные результаты
получены бюро, осуществлявшим перепись населения в 1925 году.В группу с высокой фертильностью входят девять профессий со сред¬
ним значением численности семьи 4,8. Это шахтеры, добывающие уголь,
другие шахтеры, разнорабочие на ферме, фермеры, строительные подсоб¬
ные рабочие, железнодорожные рабочие, железнодорожные строители, ли¬
тейщики и работники фабрик — в порядке снижения фертильности.В группу с низкой фертильностью входят одиннадцать профессий
со средним значением 2,6. Они расположены в порядке возрастания фер¬
тильности следующим образом: врачи; инженеры, юристы; счетоводы; бан¬
киры; учителя; агенты и счетчики; фабричные служащие; страховые аген¬
ты; служащие офисов; агенты по недвижимости.Группа с низкой фертильностью включает в себя все профессии, кроме
священников, данные по которым представлены в следующей группе, в то
время как группа с высокой фертильностью включает в себя все классы ра¬
ботников вместе с тремя другими группами, среди которых бросается в гла¬
за группа людей, занимающихся сельскохозяйственным трудом (фермеры),
как и во французских данных. В целом, американские данные показывают
более широкий диапазон вариации по сравнению с отмеченным в Шот¬
ландии; например, у шахтеров, добывающих уголь, сланец и железную ру¬
ду в Шотландии, фертильность на 77% выше, чем у врачей и хирургов,
в то время как в Америке у шахтеров на 119 % выше, чем у врачей. Исполь¬
зование данных о регистрации новорожденных должно было бы привести
скорее к снижению, чем к повышению данного соотношения, но разница,
возможно, объясняется двумя другими обстоятельствами: во-первых, воз¬
раст вступления в брак был частично нормирован в шотландских данных,
и, во-вторых, американские данные по регистрациям новорожденных отно¬
сятся к гораздо более позднему периоду, чем почти полные семьи переписи
1911 года в Шотландии. Следовательно, не бесспорно, что дифференци¬
альный охват рождаемости в Соединенных Штатах больше, чем в Велико¬
британии, хотя будет трудно привести доводы в пользу того, что он был
существенно меньше.Эффекты дифференциальной фертильностиФакты, продемонстрированные демографической статистикой нынеш¬
него века в отношении распределения фертильности, настолько не соответ¬
ствуют рациональным ожиданиям ранних социологов, что будет уместным
завершить данную главу некоторыми размышлениями по поводу тех со¬
Репродуктивность и социальный класс223циологических последствий, к которым неизбежно приведет наблюдаемое
распределение рождаемости, каковы бы ни были его причины.(1) Прежде всего, язык авторов работ по эволюционной теории наде¬
лил слово «успех» биологическим значением, награждая им тех индивидов
или те общества, которые благодаря чрезвычайно большой способности
к выживанию и репродукции постепенно вытесняют своих конкурентов
среди живущих на земле. Именно это значение имеется в виду в выра¬
жении «успех в борьбе за существование», и следует, вероятно, признать,
что в этом и состоит истинная природа биологического успеха. Что каса¬
ется человека, социума, то здесь успех в человеческом понимании неот¬
делим от достижения социального статуса, но в то же время профессии,
стоящие в социальном смысле ниже, оказываются более фертильными; мы
сталкиваемся с парадоксом, что биологически успешные члены нашего об¬
щества принадлежат в основном к социальным неудачникам, а в равной
мере процветающие и социально успешные классы оказываются в целом
биологическими неудачниками; те, кто не способен вести борьбу за суще¬
ствование, обречены, раньше или позже, в соответствии с их социальной
особенностью на то, чтобы быть искорененными из человеческого рода.
Борьба за существование в таких обществах представляет инверсию по от¬
ношению к борьбе за имущественную собственность и власть. Эволюци¬
онная задача, на решение которой направлены силы отбора, состоит в том,
чтобы произвести тип человека, который настолько оснащен своими ин¬
стинктами и способностями, что в наименьшей степени подвержен риску
отличиться посредством качеств, вызывающих восхищение или полезных
для общества, или даже путем достижения такого скромного процветания
и такой стабильности полезной занятости, которые позволят ему оказаться
в числе средних классов граждан, зарабатывающих себе на жизнь. В обще¬
ствах, созданных таким образом, мы имеем свидетельства полной неспо¬
собности экономической системы согласовать индивидуальную репродук¬
тивность с постоянным популяционным устремлением существовать в об¬
ществе путем взаимно полезной деятельности.(2) Имеющиеся факты указывают на то, что эта любопытная инверсия
наблюдается у всех цивилизованных народов современного мира. У нас,
к сожалению, нет возможности представить удовлетворительные статисти¬
ческие данные о восточных цивилизациях Индии и Китая. Огромная смерт¬
ность среди беднейшего населения этих стран вследствие войн, чумы и го¬
лода, а также государственный строй, совершенно непохожий на европей¬
ские системы государственного управления, — это факторы огромной важ¬
ности. Если бы развитие этих стран хотя бы отчасти напоминало развитие
европейских стран, процесс разрушения нации можно было бы приостано¬
224Глава 10вить. Длительный период застоя и нехватка предпринимательства, от кото¬
рых страдали оба этих народа до того, как европейские идеи начали ока¬
зывать на них сильное влияние, по меньшей мере заставляют задуматься,
насколько эти два фактора важны для нормального развития общества. Бо¬
лее того, среди древних цивилизаций, по единодушному и убедительному
свидетельству греческих и латинских писателей, подобная инверсия наблю¬
далась в тот период, когда ремесла и социальная организация достигли наи¬
высшего развития. Можно вспомнить о том, что латинское слово proletarii,
употребляемое в отношении класса граждан, не имеющих собственности,
означает «бедняки, у которых есть дети»; и что в ранней Империи прини¬
малось множество законов, хотя, возможно, и неэффективных, предназна¬
ченных для поощрения материнства и отцовства среди высших классов.(3) Рассмотрение причин, которые имеют тенденцию вызывать инвер¬
сию рождаемости, мы должны отложить до следующей главы, но сейчас
мы можем отметить, что эти причины должны были встретить и преодо¬
леть большие естественные препятствия. Благодаря грузу материнства и от¬
цовства, а также наследованию состояния, постоянная тенденция инверсии
состоит в том, чтобы сделать богатых еще богаче, а бедных — еще беднее.
Такая тенденция не только была нежелательной в интересах общества, по¬
тому что вмешивалась в распределение богатств в соответствии с оценкой
выполненных работ, обмен которыми происходит внутри общества, но и,
что является гораздо более важным, вызывала сопротивление со стороны
частных лиц, так как их экономические интересы ущемлялись ввиду тенден¬
ции облагать гораздо большим налогом бедных, а не богатых. Полный груз
воспитания следующего поколения граждан было бы легче нести, если бы
он был распределен соответственно способности выдерживать его. Те, кто
могут позволить себе больше комфорта и роскоши, конечно же, могут поз¬
волить себе больше детей; и, опираясь на чисто экономические причины,
лишенные всех биологических соображений, большое хозяйство богатых,
если здраво рассуждать, способно обеспечить и поддержать больше детей,
чем маленькое хозяйство бедняков. Более того, у нас нет причины a priori
предположить, что человечество является исключением из правила, работаю¬
щего в отношении других организмов, будь то животные или растения, что
обилие предметов первой необходимости благоприятно для репродукции.Непрямое следствие благоприятных условий более преуспевающих
классов также не следует упускать из виду. Если оптимальный уровень
фертильности вида в общем определяется требованиями родительской за¬
боты, то фактом, относящимся к делу, будет то, что высшие классы име¬
ют и всегда имели достаточно власти, чтобы переложить большую часть
своих родительских забот на плечи других. Следовательно, груз родитель¬
Репродуктивность и социальный класс225ских обязанностей, таким образом, уменьшается, и оптимальный уровень
фертильности повышается, как в случае с кукушкой. Наглядным примером
является наличие кормилиц у младенцев, традиция, которая значительно
повышает шансы более раннего зачатия для матери. Физиологический ме¬
ханизм, посредством которого это осуществляется, может рассматриваться
как способ повышения фертильности, как только естественный груз ро¬
дительской заботы снят, как было бы в случае смерти младенца. Тот же
самый принцип применяется к заботам, требуемым от родителей, в умень¬
шении доли смертности в младенчестве и детстве; и ясно, что поскольку
эти заботы можно переложить на плечи квалифицированных профессиона¬
лов, наиболее благоприятные биологические условия у привилегированных
классов будут постепенно повышаться. Если бы такой привилегированный
класс был биологически изолирован, свойственная ему фертильность, сле¬
довательно, увеличилась бы до более высокого уровня, чем та, что наблю¬
дается в классах, которые сами должны заботиться о своих детях, и это
не говоря уже о том, что для одной и той же фертильности более преуспе¬
вающий класс должен быть, по сути, и более плодовитым.В сравнительно недавние времена на национальном уровне предпри¬
нимались усилия, чтобы облегчить жизнь бедных и предусмотреть ра¬
зумные ассигнования на образование их детей путем обложения налогом
самых больших доходов и состояний. Важным фактором, если вообще
не причиной, необходимости постоянного перераспределения национально¬
го состояния является неравенство распределения груза родительских забот
и его особенно тяжелого бремени, лежащего на беднейшем классе. Несмот¬
ря на огромные расходы и, возможно, важность биологических послед¬
ствий, несмотря на снятие с беднейшего класса большей части родитель¬
ских расходов и, следовательно, хотя и не напрямую, повышение фертиль¬
ности, было бы ошибкой предположить, что груз родительских расходов
в любом классе граждан, самостоятельно зарабатывающих себе на жизнь,
станет ощутимо легче. Необходимость поддерживать обычную семью, в ко¬
торой есть три-четыре ребенка, у большей части нашего индустриального
населения влечет за собой период самой суровой бедности, который по ком¬
форту, защищенности и возможности откладывания сбережений резко от¬
личается от уровня жизни, которым может наслаждаться на ту же зарплату
бездетный человек. Экономическое давление низких жалований, в то же
время неспособность выдерживать реальное родительское бремя, с кото¬
рым экономическое давление находится в ситуации противоречия, уже спо¬
собны причинить трудности матерям следующего поколения; это свойство
нашей социальной системы, которое нельзя не заметить, его суть в том, что
чрезмерно большая часть каждого поколения вышла из семей, в которых
226Глава 10испытала всю тяжесть указанного конфликта. Инверсия рождаемости есть
плодовитая мать социальной неудовлетворенности.(4) Тесно связанным с концентрацией репродуктивной активности
в беднейших слоях граждан является следствие, которое, на первый взгляд,
имеет полезное значение, а именно: что в каждом поколении должно на¬
блюдаться очень большое социальное продвижение, таким образом, чис¬
ленность лучше оплачиваемых классов сможет сохранять постоянное коли¬
чественное соотношение с теми, кто получав! меньше. Социальное про¬
движение, обусловленное этой причиной, нужно тем не менее отличать
от обычного продвижения, которое происходит в силу возраста и большего
опыта и которое обеспечивает детям такие же социальные преимущества,
как у родителей; кроме того, есть еще социальное продвижение особого
качества или таланта, которое предоставит более одаренным в большой
или совсем небольшой степени должную возможность для приложения их
отличительных способностей. Продвижение этих двух типов должно ха¬
рактеризовать любое общество как мудро и благородно организованное,
каким бы ни было распределение репродуктивности внутри него; но ес¬
ли мы хотим, чтобы преуспевание стало поведением, заслуживающим на¬
грады, то это не обязательно означает, что мы желаем постоянной замены
целого класса людьми более скромного происхождения, разве что такая
замена может быть показана как ведущая к общему процветанию. Сочув¬
ствие, которое мы также испытываем в отношении попыток ответственных
родителей способствовать счастью своих детей, было бы не к месту, если
бы привело нас к желанию того, чтобы число детей, нуждающихся в счаст¬
ливом случае для достижения полезного и преуспевающего образа жизни,
возрастало, а число тех детей, которые могли бы провозгласить такой образ
жизни принадлежащим им по праву рождения, уменьшалось.Если учесть естественные последствия, проистекающие от продвиже¬
ния индивида наверх на протяжении всей социальной шкалы, включая пе¬
реход многих миллионов в каждом поколении, мы обнаружим, что некото¬
рые из этих последствий будут, несомненно, желаемыми, а другие — вред¬
ными. В каждой группе людей, чьи родители, а также родители родителей
в среднем занимали явно более скромное положение, очевидно, не следует
ожидать чувства кичливой гордости своим происхождением, характерного
для многих нецивилизованных людей. Консервативная добродетель, кото¬
рая стремится быть достойной благородного имени, будет заменена более
прогрессивной добродетелью, которая стремится оправдать новые требо¬
вания. В худшем случае гордость происхождением заменяется гордостью
богатством, и общественное мнение стремится предоставить привилегии
аристократии скорее богатым, чем родовитым.
Репродуктивность и социальный класс227Второе и более ясно выраженное следствие массового социального
продвижения — это задержка культурного прогресса в каждом классе со¬
общества по причине того, что необходимо дать более высокий уровень
образования тем, кто ранее был в неблагоприятных условиях. Нагрузка
на образовательную систему, заданная этим фактором, оказывается очень
большой, и, несомненно, в основном именно по этой причине прогресс
истинной культуры идет так медленно, и местами возникают задержки.
Но данное следствие социального продвижения обязательно проявится, ка¬
кими бы ни были наследственные способности разных классов.Третий и наиболее серьезный недостаток заключается в том, что хо¬
тя эффективность лучше оплачиваемых классов может сохраняться за счет
механизма социального продвижения, постепенно совершенствуемого так,
чтобы наилучшим образом использовать каждый вид таланта, где бы он
ни был обнаружен, но среди тех профессий, которые оплачиваются ху¬
же, невозможно найти тот источник, из которого можно набирать таланты
и умения, и, следовательно, указанные слои населения испытывают посто¬
янное снижение среднего уровня талантов, которыми они обладают. Не за¬
нижая ценность работ, которые выполняются в новых областях занятости
теми, кто продвинулся по социальной лестнице, было бы губительно про¬
игнорировать ту степень, в которой преуспевание любого так называемо¬
го сообщества покоится на среднем уровне эффективности больших масс
граждан, а также на уровне их самоуважения, инициативы и морального духа.РезюмеС экономической точки зрения разнообразные профессии человека
в обществе различаются размером денежного вознаграждения, которое они
обеспечивают. С биологической точки зрения они важны тем, что незамет¬
но контролируют выбор партнера посредством влияния превалирующего
мнения, взаимного интереса и возможностей для социального взаимодей¬
ствия. Таким образом, социальные классы начинают генетически диффе¬
ренцироваться как локальные вариететы вида, хотя дифференциация опре¬
деляется не первичным отбором разниц между классами, но факторами,
контролирующими социальное продвижение или понижение.Сравнения демографических статистик, особенно данных о рождае¬
мости, в разных классах большей частью недостаточны и очень устаре¬
ли. В нашей стране необходимо прежде всего согласовать классификацию
по профессиям, которая используется при регистрации случаев рождения
и смерти, с классификацией, используемой при переписи, и записывать воз¬
раст родителей при регистрации рождений.
228Глава 10Многочисленные исследования, в которых проблему рассматривают
с разных точек зрения, показали, что во всех цивилизованных странах рож¬
даемость у бедных гораздо выше, чем у более преуспевающих классов,
и что эта разница в рождаемости возрастает на протяжении последних по¬
колений. Когда стали доступны более полные данные, оказалось, что эта
разница в рождаемости не ограничена аристократическими или более об¬
разованными семьями, но простирается и вниз по социальной лестнице,
демонстрируя контраст между полуквалифицированными и неквалифици¬
рованными рабочими. Нет прямых доказательств того, что существовал пе¬
риод, когда рождаемость во всех классах была одинаковой, а снижение рож¬
даемости во всех классах в последние десятилетия проходит одновременно,
хотя оно в большей степени выражено у преуспевающих классов.Поскольку рождаемость является доминирующим фактором выжи¬
вания человека в обществе, успех в борьбе за существование является
в обществах с инвертированным уровнем рождаемости инверсией успеха
в устремлениях человека. Тип человека, отобранного в качестве предка бу¬
дущих поколений, является типом, чьи способности в наименьшей мере до¬
стойны восхищения или вознаграждения за полезное служение обществу,
к которому он принадлежит.Вероятно, если бы подобная инверсия превалировала в азиатских цен¬
трах цивилизации, то уровень смертности у беднейшей части населения
должен был стремиться задержать прогресс разрушения нации и, возможно,
обеспечил равновесие до прихода европейских идей. Положение Римской
империи было похоже на то, что мы наблюдаем в современных странах.Причины, которые привели к инверсии рождаемости, должны были
быть достаточно мощными, чтобы противостоять как прямой, так и кос¬
венной экономическим силам, благоприятствующим более высокому уров¬
ню рождаемости у более преуспевающих классов. Поскольку при инверсии
рождаемости существует тенденция делать богатых еще богаче, а бедных —
еще беднее, и особенно навлекать трудности на родителей следующего по¬
коления, инверсия является важной причиной социального недовольства.Следствие, которое, на первый взгляд, кажется преимуществом, это
масштабное социальное продвижение, необходимое, чтобы поддержать со¬
отношение классов. Однако, исследовав этот вопрос, мы приходим к вы¬
воду, что данный тип продвижения не следует путать с повышающимся
преуспеванием, и что оно несет с собой серьезную некомпенсированную
дееспособность ввиду торможения культурного прогресса каждого класса
и истощения беднейшего класса в своей способности поддерживать само¬
уважение и экономическую независимость.
Глава 11Социальный отбор по фертильностиИстория теории. Бесплодие во всех классах, независимо от причины возникно¬
вения, приводит к социальному продвижению. Отбор — основная причина инверти¬
рованного уровня рождаемости. Упадок правящих классов. Контраст с варварскими
обществами. Героизм и высшие способности человека. Место социального класса
в эволюции человека. Аналогия с паразитизмом у муравьев. Резюме.Может ли так случиться, чтобы единственный рожденный сын со¬
хранил дом предков? Поскольку так лишь богатство возрастает в доме.
Гесиод, VIII век до н. э.История теорииМы видели в последней главе, что среди цивилизованных народов как
в новое время, так и в древние времена возникало аномальное положение,
когда более преуспевающие социальные классы, которые мы естественным
образом сравнили бы с успешными и хорошо адаптированными видами жи¬
вотных, воспроизводят свой вид значительно медленнее, чем классы более
низкого социального положения. В главе 9 мы нашли причину, согласно
которой можно заключить, что врожденная и наследственная предрасполо¬
женность оказывает у цивилизованного человека мощное влияние на уро¬
вень репродукции. Здесь мы попытаемся показать логическую связь между
этими двумя заключениями. Это не исключает, что так называемые случай¬
ности истории время от времени определяли социальное положение раз¬
ных типов людей и влияли на фертильность разных классов. Широко рас¬
пространенную природу дифференциальной рождаемости, существующую
в больших группах людей, у разных наций и заново появляющуюся че¬
рез длительные промежутки времени в совершенно других цивилизациях,
не следует тем не менее объяснять историческими случайностями. Нель¬
зя принять ни одно объяснение, которое не было бы связано с фактора¬
ми почти универсального действия в цивилизованных обществах разных
типов.
230Глава 11Разрабатываемую нами теорию можно найти в зачаточном состоянии
в интересном наблюдении, отмеченном Фрэнсисом Галтоном в ходе его
генеалогических исследований; по мере продвижения нашего знания о со¬
циологических явлениях оно развивалось несколькими авторами, но ни его
логическая обоснованность, ни важность для социологической теории, ка¬
жется, так и не были широко восприняты; и если не считать нескольких
моих небольших работ и короткого обсуждения в недавней книге майора
Дарвина The Need for Eugenic Reform, оно, можно сказать, совершенно иг¬
норируется.В книге Hereditary Genius, опубликованной в 1869 году, Галтон рас¬
сматривает проблему, являющуюся общепризнанным фактом, что семьи
великих людей с необычайной частотой имеют тенденцию вырождаться.
Из 31 титула пэра, полученного судьями Англии, семьи двенадцати уже
пресекли свой род. Галтон исследовал истории этих семей и наткнулся
на объяснение, которое он по праву описывает как «простое, адекватное
и оригинальное». Значительная часть новых пэров и их сыновей были же¬
наты на наследницах рода.Но мои статистические списки со всей очевидностью показали, что эти браки
особенно неплодовиты. Нам следовало ожидать, что наследница рода, которая яв¬
ляется единственным потомком брака, не будет настолько же плодовита, как и жен¬
щина, у которой много братьев и сестер. Относительное бесплодие должно быть
наследственным в той же мере, как и другие физические качества, и я уверен, то же
самое происходит и в случае с домашними животными. Следовательно, потомство
в браке пэра, заключенном с единственной наследницей, зачастую отсутствует, и его
титул некому передать.После описания историй указанных семей он приходит к следующим
выводам:(1) Из 31 семьи пэров не менее чем в 17 наследственное влияние наследницы
или сонаследницы сказывалось в первом или во втором поколении, поскольку это
влияние было заметным фактором, приводящим к бесплодию в 16 из 17 случаев,
и наблюдалось иногда в 2, 3 и более случаях в одной семье.(2) Прямые мужские линии не менее чем в 8 семьях пэров [Галтон приводит
имена полностью] были уничтожены в силу влияния наследниц, а еще 6 семей
находились почти на грани вымирания У меня есть в буквальном смысле лишь
единственный случай, в котором не ощущалось разрушительного влияния крови
наследницы.(3) Не менее 8 из 12 семей пэров не получили прямого наследника по мужской
линии по причине брака с наследницами.Рассмотрим оставшиеся четыре. Лорды Сомерс и Турлоу умерли неженатыми.
Лорд Элванли имел двух сыновей, из которых один умер неженатым. Лишь в его
Социальный отбор по фертильности231случае и в случае графа Менсфилда — из 10 вступивших в брак и изживших титул
пэра — женитьба на наследнице не является причиной вымирания титула пэра. Сле¬
довательно, никто не может утверждать, что существуют основания для того, чтобы
приписать роду судей исключительное бесплодие. Факты при тщательном анализе
настойчиво указывают на противоположную тенденцию.Сделав подобные выводы при рассмотрении других групп пэров, Гал-
тон продолжает:Я попытался подойти к вопросу с другой стороны, взяв браки последних пэров
и сравнив число детей, когда мать являлась наследницей, с теми, где мать не явля¬
лась наследницей. Я принял меры предосторожности, чтобы исключить из послед¬
ней группы всех матерей, которые являлись сонаследницами, или отцов с един¬
ственным сыном. Кроме того, поскольку число наследниц достаточно ограничено,
я иногда возвращался на два-три поколения назад в поисках случая брака с наслед¬
ницей. Таким образом, я взял по пятьдесят случаев тех и других. Я привожу эти
данные ниже, удвоив действительные результаты, чтобы выразить их в процентах
[табл. 11].Таблица 11Число сыновей Число случаев, Число случаев,
в каждом браке когда мать когда матьявлялась наследницей не являлась
наследницей022211610222143223441020568628704Я обнаружил, что среди жен пэров 100 жен, являющихся наследницами, име¬
ют 208 сыновей и 206 дочерей, а 100 жен, не являющихся наследницами, имеют
336 сыновей и 284 дочери.Необходимо процитировать и следующие очень важные абзацы, чтобы
показать, какое значение этому принципу придавал Галтон.Каждое повышение в звании — это новый стимул ввести в семью еще од¬
ну наследницу. Следовательно, у герцогов больше крови наследниц, чем у графов,
и следует ожидать, что герцогство должно быть подвержено вымиранию в большей
степени, чем графство, а графство, в свою очередь, исчезает быстрее, чем сосло¬
вие баронов. Опыт показывает, что в большинстве случаев, можно вполне уверенно
232Глава 11определить причину исчезновения титула. Сэр Бернард Берк в своем предисловии
к Extinct Peerage утверждает, что все английские герцогства, созданные с начала
правления Карла II, исчезли, кроме трех, которые перешли в королевскую семью,
и лишь 11 семей графов осталось из тех многих, что были созданы норманнами,
Плантагенетами и Тюдорами.Я в значительной мере удовлетворен, что проследил и, надеюсь, окончательно
опроверг причину, по которой семьи склонны к вымиранию в связи с их высоким
положением, особенно вследствие моего желания показать, что талантливый род
не обязательно является бесплодным, и, во-вторых, потому, что это может положить
конец диким и нелепым гипотезам, которыми объясняют их исчезновение.Альфонс Декандоль, отметив эти поразительные исследования и долж¬
ным образом проводя различие между исчезновением семьи и пересечени¬
ем по мужской линии, разумно подмечает, что подобные выводы должны
применяться к богатым и состоятельным классам вообще.La difference de fecondite des heritieres et non hcritieres anglaises est si grande
qu’elle avertit d’une cause, jusqu’a present inconnue, du petit nombre des naissances
dans les families aisees ou riches, de la noblesse et de la bourgeoisie. En general, les filles
riches se marient aisement et selon toutes les probabilites physiologiques, confirmees par
les faits que Monsieur Galton a decouverts, ce sont elles qui ont la plus petite chance de
laisser des descendants. Leur proportion doit done diminuer l’augmentation de population
des classes qui vivent dans l’aisance.К сожалению, г-н Декандоль связывает это рациональное объяснение
относительного бесплодия высших классов с другими объяснениями, на¬
пример, с такими, как объяснение Герберта Спенсера, явно относящегося
к категории, которой Галтон надеялся положить конец. Также не вполне
ясно, уяснил ли он суть того, о чем говорит Галтон: дело не в том, что де¬
вушкам с наследством легче выйти замуж, чем другим девушкам, но в том,
что они нацелены на брак, дающий социальные преимущества, и тогда ха¬
рактерное для них бесплодие соединяется с естественными способностями
исключительно плодовитых мужчин.В короткой, но важной заметке, написанной для Eugenics Review
в 1913 году, Кобб представил доводы в пользу того, что случай с наследни¬
цами, наблюдавшийся Галтоном, является не чем иным, как частным случа-*Разница по фертильности между единственными наследницами рода и теми, кто не явля¬
ется единственными наследницами, настолько велика, что заставляет задуматься о неизвест¬
ной до сих пор причине малого числа детей в состоятельных или богатых семьях как дворян,
так и буржуазии. Вообще говоря, богатые девушки без труда выходят замуж, но по всем фи¬
зиологическим вероятностям, как подтверждают факты, открытые г. Галтоном, именно у них
меньше всего шансов оставить наследников. Таким образом, их доля среди заключивших брач¬
ный союз должна уменьшить рост населения для классов, живущих в достатке.
Социальный отбор по фертильности233ем гораздо более общей тенденции. Ограничившись неосознанными причи¬
нами относительного бесплодия, г-н Кобб указывает, что так же, как удача
наследницы позволяет ей вступить в выгодный в социальном отношении
брак, так и дети родителей любого класса, члены более малочисленных
семей будут в среднем начинать жизнь, имея социальное преимущество
по сравнению с членами больших семей. Наряду с многочисленными при¬
знаками превосходства, которые позволяют семье улучшить свое социаль¬
ное положение, относительное бесплодие также играет свою роль. Таким
образом, менее фертильные группы, имея социальное преимущество, будут
постепенно проникать в высшие слои общества и там становиться причи¬
ной необычной ситуации, в которой более удачливые и успешные люди
имеют наименьший уровень рождаемости.Цитата из статьи г-на Кобба позволит читателю оценить точку зрения,
при помощи которой этот важный вывод был достигнут.Последователи евгеники соглашаются, что подрастающее поколение — это
в основном выходцы из наименее подготовленных. Это приписывается частично то¬
му факту, что высшие классы вступают в брак позже, и частично тому, что, помимо
откладывания женитьбы, высшие классы являются и в меньшей степени плодови¬
тыми, чем те, кто находится ниже по социальному положению.Несомненно, что в настоящее время меньший уровень фертильности выс¬
ших классов в основном является следствием искусственного ограничения зачатия.
Но существует и другая причина, именно к ней я хочу привлечь внимание. Для ев¬
геника важно, к какой причине отнести эту меньшую фертильность высших клас¬
сов; если бы причина была в искусственном предохранении, которое является всего
лишь временной модой, последствия не были бы такими серьезными, поскольку
мода на искусственное ограничение численности семьи, насколько можно ожидать,
пройдет обычным чередом и распространится вниз по социальной лестнице, по¬
степенно уравнивая фертильность на всех уровнях общества; или эта мода может
вообще исчезнуть, когда популяция осознает тот разрушительный эффект, который
она оказывает на будущее расы. Вполне вероятно, что преимущество ограничения
семьи будет привлекательно в большей степени для бедных, чем для богатых, по¬
скольку дополнительный ребенок — это бремя для бедных, и, возможно, постепенно
искусственное ограничение семьи начнет оказывать благоприятный эффект на расу,
снижая размер семей тех, кто оказывается в наименьшей степени эффективным.Но если, как я попытаюсь показать, и в современных условиях существует
естественная тенденция, когда более умные люди становятся менее фертильными,
то проблема эта оказывается гораздо более серьезной.Если вариации фертильности наследуются, и преуспевающие классы поколе¬
ние за поколением проходят через отбор, посредством которого члены небольших
семей получают преимущество над членами больших семей, нам следует ожидать,
что более состоятельные классы в целом будут менее фертильными, чем бедные
классы.
234Глава 11Должна существовать некая общая причина, которая не дает среднему интел¬
лекту в цивилизованном сообществе продвигаться дальше определенного уровня.
Этой причиной, как мне кажется, является ранжирование общества в соответствии
с уровнем богатства. Это помещает в один класс детей сравнительно нефертильных
родителей и людей, способных к воспроизводству, и браки между ними приводят
к соединению стерильности и способности к воспроизводству.Бесплодие во всех классах, независимо от его причины,
приводит к социальному продвижениюРазвивая теорию трех упомянутых авторов, можно видеть, что достиг¬
нутый прогресс состоял почти полностью в распространении принципа
на все более и более широкий ряд социальных классов. Ясно, что это лишь
следствие расширения нашего знания классов, в которых проявился инвер¬
тированный уровень рождаемости. Галтон думал в основном о титулован¬
ных семьях, потому что сознавал, что среди них вырождение титула имело
место с поразительной частотой; Декандоль понимал, что низкий уровень
рождаемости характеризовал также и богатый городской класс, он сразу
понял, что принцип, который обнаружил Галтон в генеалогии титулован¬
ных семей, должен быть в равной мере применим и там, где на социальное
положение в большой степени оказал влияние унаследованный капитал.
Возможно, что в 1913 году Коббу еще не были известны результаты пере¬
писи 1911 года в Великобритании, согласно которым инверсия развилась
в большой степени даже в беднейшем классе, но у него не было сомне¬
ния относительно ее распространения на большие массы населения, и ему
принадлежит большая заслуга в отнесении принципа к классам, в которых
наследуемое состояние относительно невелико.То, что экономическая ситуация на всех уровнях современных обществ
благоприятствует социальному продвижению менее фертильных, понятно
из ряда привычных рассуждений. В самом богатом классе наследуемое со¬
стояние делится между естественными наследниками, и состояние ребенка
обратно пропорционально численности семьи, к которой он принадлежит.
В средних классах следствие прямого наследования также имеет значе¬
ние, но беспокойство родителя большой семьи возрастает из-за расходов
на образование, профессиональное обучение и в силу необходимости де¬
нежных средств из-за выбора профессий, которые несут большую выгоду.
На низшем экономическом уровне социальный статус в меньшей степени
зависит от реально наследуемого капитала, чем от трат на домашнее хозяй¬
ство, образование, развлечения и одежду, в то время как сбережения бедных
расходуются или израсходованы полностью, а перспективы их экономиче¬
Социальный отбор по фертильности235ского благополучия зачастую ограничены необходимостью в достаточной
мере обеспечивать пропитание и одежду для детей. Эти очевидные фак¬
ты подтверждены аргументами в пользу ограничения численности семьи,
наиболее весомым из которых является обязанность родителей дать детям
наилучший возможный старт в жизнь, и последующая необходимость как
сбережений, так и расходов — аргумент по меньшей мере настолько же
действенный в отношении бедных, как и в отношении богатых.Отбор — основная причина инвертированного уровня
рождаемостиВ главе 9 было показано, что не только чисто физиологическое бес¬
плодие, но также причины низкой репродуктивности, зависящие от выбора
человека, такие как безбрачие, откладывание брака и ограничение числа де¬
тей супружескими парами, также находятся под сильным влиянием наслед¬
ственных факторов. Это добавление наследственных элементов, которые
обусловливают большую часть дисперсии репродуктивности у цивилизо¬
ванных людей, многократно увеличивает эффективность действия принци¬
па отбора. В частности, мы наблюдали, что наследственные отличия в ум¬
ственных и моральных качествах, влияющие на репродукцию, должны быть
такой величины, чтобы произвести значительные эволюционные изменения
за относительно короткие исторические периоды; что эти изменения проис¬
ходили в действительности в нравах народов, о которых известно, что они
подвергались селективному действию добровольного ограничения числен¬
ности семьи. Следовательно, нам не избежать вывода о том, что общество,
в котором члены небольших семей имеют в среднем социальное преиму¬
щество по сравнению с членами больших семей, а родители во всех других
отношениях находятся в равном положении, будет разделяться не только
в отношении физиологического бесплодия, но гораздо быстрее и более рез¬
ко — в отношении различий в обычаях, которые ведут к безбрачию, откла¬
дыванию брака и ограничению рождаемости.Такой характер теории при полном ее развитии помогает объяснить
особенность дифференциальной репродукции, которая в других отношени¬
ях была бы не вполне ясна; то есть все социологические факторы, которые
можно выявить в дифференциальной репродукции, оказывают одинаковое
влияние в одном и том же направлении. Если одна из социологических при¬
чин благоприятствовала использованию контрацептивных средств высши¬
ми классами, как например, более свободный доступ к медицинским знани¬
ям, то следствием должно было бы стать снижение рождаемости у родите¬
лей определенного возраста, частично скомпенсированное в сводных стати¬
236Глава 11стических показателях большей готовностью заключать брак. Таким обра¬
зом, если появляются какие-либо факторы, связанные с профессиональны¬
ми условиями лучше оплачиваемых классов, то они могут привести к от¬
кладыванию брака: только эта причина, действующая сама по себе, будет
естественным образом сопровождаться более высоким уровнем рождаемо¬
сти в старших возрастных группах. Но в действительности у высших клас¬
сов наблюдалось одновременное действие нескольких причин: более высо¬
кий уровень безбрачия, более поздний брак и снижение уровня рождаемо¬
сти у супругов в определенной возрастной группе; именно этого следова¬
ло бы ожидать, если бы действующей причиной явления было селективное
продвижение в более высокие социальные слои чистого бесплодия как тако¬
вого, независимо от физиологических причин, которые его вызвали.Есть еще один способ проверить, являются ли наследственные эффек¬
ты основной и определяющей или второстепенной причиной дифференци¬
ального уровня рождаемости. Даже во времена Галтона выдвигалось мно¬
жество предположений относительно известных тогда причин этого фено¬
мена: различия в фертильности приписывались различиям в социальной
среде или физиологической связи между бесплодием и умственными спо¬
собностями. С тех пор список предположений стал длиннее, например,
в качестве причины называлось излишество в еде, свойственное высшим
классам, избыток свободного времени, умственное напряжение, ослабля¬
ющее влияние комфорта. Какими бы необоснованными ни казались эти
предположения, если рассмотреть каждое в отдельности, то a priori мож¬
но сказать, что социальное окружение более преуспевающего класса могло
оказать некое действительно неблагоприятное влияние на репродуктивные
возможности. Наиболее определенные результаты для выбора между двумя
точками зрения можно получить, если сравнить относительную фертиль¬
ность среди представителей одного социального класса, противопоставив
тех, кто попал туда благодаря быстрому социальному продвижению, и тех,
кто принадлежит ему по рождению. Если следовать теории, что на фер¬
тильность в основном оказывают влияние наследственные факторы, то фер¬
тильность у высших классов не должна повышаться после вливания более
низкой фертильности тех, кого в этот слой общества привело социальное
продвижение, учитывая, что отток более фертильных членов высших клас¬
сов в нижние слои весьма мал. Следовательно, группы, получающие быст¬
рое социальное продвижение, согласно данной теории должны быть даже
менее фертильными, чем классы, на уровень которых они поднимаются.Если же, напротив, важными причинами являются любые, подходящие
под определение «социальных условий», мы, наверное, можем ожидать, что
семьи, которые поднимаются по социальной лестнице, должны нести с со-
Социальный отбор по фертильности237Таблица 12Колледж и профессиональное обучение
Колледж и степень доктора
КолледжСтандартное, бизнес, торговля,
секретарские курсы
Средняя школаНачальная школа (первые 8 лет обучения)
Только профессиональная школаОбразование2,32,12,11,9бой величину фертильности того класса, из которого они вышли. Мы убе¬
дились, что дифференциальная рождаемость очень развита в Соединенных
Штатах, и в таблице Хантингтона и Уитни, представленной в американ¬
ском справочнике Who s Who, дается средняя численность детей на чело¬
века при разбиении на группы по полученному образованию. Средняя чис¬
ленность представлена в убывающей последовательности (табл. 12).Поскольку общая численность исследуемых составила 25 ООО, то об¬
щая тенденция не может быть приписана чистой случайности. Таблица яс¬
но показывает, что среди американцев, которые достигли положения доста¬
точного уровня, чтобы быть упомянутыми в справочнике Who s Who, те, чье
социальное продвижение было наиболее быстрым, имеют в среднем мень¬
ше детей, чем те, кто в меньшей степени продвинулся по социальной лест¬
нице. Такой результат был бы необъясним с позиции тех взглядов, которые
связывают дифференциальную фертильность разных классов с элементами
их социального окружения. И, напротив, этот результат является удивитель¬
ным подтверждением одного из самых неожиданных выводов теории, что
доминирующая причина лежит в социальном продвижении относительно
бесплодных.Уитни и Хантингтон также представили результаты другого исследо¬
вания, касающегося той же проблемы. Они изучили средние способности,
продемонстрированные студентами Йельского университета, которые ро¬
дом из семей с 1,2,3,..., 6 и более детьми. В целом они обнаружили, что
способности возрастают при движении от семей с одним ребенком к се¬
мьям с двумя детьми, и так — до самых больших семей. Конечно же, это со¬
всем не тот результат, который мы ожидали увидеть, будь это в Англии или
в Америке, если бы мы проверили выходцев из широких слоев населения.
У населения в целом классы, в которых мы находим самые большие семьи,
несомненно, покажут самый низкий уровень способностей, возьмем ли мы
238Глава 11за основу исследования способности к учебе, спортивные способности или
интеллект. Однако студенты Йельского университета являются выходцами
из избранной группы населения. Это дети тех, кто может позволить се¬
бе дать им дорогостоящее образование. Такая основа отбора благоприятна
для более преуспевающих родителей, которые, поскольку их преуспевание
дается им по праву рождения, будут с рождения более способными, чем
население в целом; но этот отбор родительского преуспевания будет гораз¬
до более убедительным в отношении родителей шести и более детей, чем
в отношении родителей одного ребенка. Не забывая о том, что единствен¬
ному ребенку легче получить более удачный старт в жизнь, чем ребенку
из семьи с шестью детьми, мы, вероятно, можем ожидать, что в Йельском
университете учатся единственные дети менее успешных родителей. Эти
дети получают образование на равных условиях и бок о бок с детьми дру¬
гих родителей, прошедших через более строгий отбор, которые, несмотря
на большое число детей в семье, способны тем не менее отправить детей
в Йельский университет.Фактически необходимым следствием нашей теории социального от¬
бора по фертильности становится следующее: поскольку фертильность
в популяции должна быть отрицательно скоррелированна с такими при¬
знаками, как интеллект, что ведет к социальному продвижению, и если бы
мы отобрали группу детей, которые выходят в мир с абсолютно равными
социальными возможностями, то мы должны найти внутри такой группы
положительную корреляцию фертильности с такими признаками. Родите¬
ли, которые могут дать большой семье некоторый уровень преимущества
в образовании, могли бы дать меньшей семье большие преимущества и,
следовательно, обеспечить ей более высокий средний уровень возможно¬
стей, помимо фертильности, благоприятствующих социальному продвиже¬
нию, чем родители меньших семей, чьи дети в действительности получают
такое же преимущество.Я полагаю, что ни одно исследование не задавалось целью получить
группу, в которой преимущества образования, обеспеченные родителями,
были бы абсолютно одинаковыми, но влияние частичного выравнивания
на увеличение корреляции между фертильностью и интеллектом, показано
в очень интересных материалах, опубликованными Сатерлэндом и Годф¬
ри X. Томсоном в 1926 году. В неселективной группе из 1 924 детей началь¬
ной школы острова Уайт в возрасте 10,5-11,5 лет был получен коэффици¬
ент корреляции —0,154, с ошибкой 0,023. Поскольку корреляция отрица¬
тельна и явно значима, оказалось, что наименее умные дети в этой произ¬
вольной группе принадлежат к самым большим семьям. С другой стороны,
386 мальчиков Королевской классической школы города Нькасл-он-Тайн,
Социальный отбор по фертильности239коэффициенты интеллектуальности которых, как утверждают, очень надеж¬
ны, а также 395 мальчиков и девочек школьного пансиона Мори, Эдинбург,
показывают корреляции 0,058 ± 0,04, и 0,075 ± 0,034, и то и другое значе¬
ния хотя и отрицательные, но статистически незначимы. Поскольку в одной
и той же школе, даже в конкретном классе, должна существовать большая
вариация в социальных преимуществах, с которыми разные ученики выхо¬
дят в жизнь, эти данные не оставляют сомнения, что в группе, в которой эти
преимущества строго выравнены, корреляция между фертильностью и ин¬
теллектом увеличилась бы до положительного значения. Очевидно, что та¬
кие результаты противоречат любой теории, которая предполагает физиоло¬
гическое противопоставление фертильности и интеллекта, и необъяснимы
с той точки зрения, что классовые различия по фертильности полностью
или большей частью обусловлены классовой средой.Упадок правящих классовФакт упадка прошлых цивилизаций является наиболее явным в исто¬
рии, и поскольку наиболее яркие периоды в больших центрах цивилизаций
часто были ознаменованы нашествием чужеземных правителей, то призна¬
но, что непосредственной причиной упадка должно быть вырождение или
истощение правящих классов. Выдвигалось множество спекулятивных тео¬
рий, призванных объяснить большое непостоянство таких классов. Прежде
чем двигаться дальше, полезно будет рассмотреть, какие элементы истины
могут содержаться в указанных теориях.Тот самый граф де Гобино, активному уму которого упадок цивилиза¬
ций представлялся величайшей проблемой человечества, полагал, что ис¬
чезновение первоначальных аристократических семейств было всюду след¬
ствием расового смешения. Другие не сомневались в том, что бесплодие
высших классов было результатом инбридинга; но поскольку сейчас это
мнение с биологической точки зрения повсеместно опровергнуто, то нет
нужды его рассматривать. С другой стороны, в теории Гобино о следстви¬
ях расового смешения не было ничего, что могла бы опровергнуть наука
его времени; если бы прогресс генетических знаний представил его взгля¬
ды как маловероятные, они бы все равно имели претензии на то, чтобы их
приняли как исторически установленные, будь они единственно доступным
объяснением исторических фактов.Общие последствия смешения рас можно было бы предсказать с уве¬
ренностью, без особого знания тех факторов, по которым отличались срав¬
ниваемые расы. Если эти расы и их потомки свободно заключали браки,
то унаследованные независимо факторы должны комбинироваться случай¬
240Глава 11но; каждый человек смешанной расы будет иметь сходство с одной роди¬
тельской группой в отношении одних факторов, и с другой — в отноше¬
нии других. Их общий облик, следовательно, будет промежуточным, но их
изменчивость больше, чем изменчивость исходных рас. Более того, но¬
вые комбинации добродетели и способностей, а также их противоположно¬
стей, окажутся в смешанной расе не обязательно гетерозиготными, но мо¬
гут быть фиксированы как постоянные расовые признаки. Таким образом,
в смешанной расе имеются большие возможности для действия отбора.
Если отбор положительный, и лучшие оставляют большее число потом¬
ков, то окончательным следствием смешения будет создание расы, которая
не только не уступает расам, из которых она возникла, но превосходит и ту,
и другую в той мере, в какой могут комбинироваться преимущества той
и другой. Отрицательный отбор, с другой стороны, приведет к более быст¬
рому разрушению смешанной расы, чем ее прародителей. Следует, конеч¬
но, помнить, что все существующие расы демонстрируют очень высокую
изменчивость в отношении наследственных факторов, так что отбор та¬
кой интенсивности, которому подвержено человечество, способен вызвать
быстрые изменения даже в самой чистой из существующих рас.Не будет неразумным предположить, что внутри каждого большого
подразделения человечества внутренние адаптации и настройки между раз¬
ными способностями ума и тела установились в течение длительного пе¬
риода отбора, под влиянием которого были сформированы эти своеобраз¬
ные расы. Если бы это действительно было так, то скрещивание сильно
различающихся рас нарушило бы эту внутреннюю гармонию и постави¬
ло бы смешанную расу в первоначальное невыгодное положение, из кото¬
рого даже при благоприятном отборе пришлось бы выбираться в течение
нескольких поколений. Теория Гобино зависит от эффекта смешения пра¬
вящей аристократии, которую Гобино неизменно прослеживает до европей¬
цев и достоинствам которой он приписывает прогресс и стабильность всех
цивилизаций со смешанными и темнокожими людьми, населявшими один
из больших древних центров цивилизации, таких как собственно Китай
или долины Ганга, Евфрата или Нила. Помимо его предположения о ра¬
совом происхождении правителей, историческое значение таких случаев
не вызывает сомнений; и во всех отношениях, возможно, что при усиле¬
нии смешения правящий класс в отсутствие отбора полностью изменил бы
свой характер: качества, которые его отличали, распространились бы на бо¬
лее многочисленную расу и за неимением исконно присущей способности
уже не были бы способны достичь больших результатов, за которые они
отвечали в сконцентрированном у правящего класса виде. Тем не менее
представление о процессе расовой диффузии устанавливает определенный
Социальный отбор по фертильности241период для любой цивилизации, который зависит от достоинств чужерод¬
ного меньшинства; остается объяснить, чем вызвана необходимость пред¬
положения, что великие цивилизации должны были всегда основываться
на таком ненадежном фундаменте. Если люди из самых больших центров
древнего заселения оказались неспособны сами по себе создать великую
и сильную цивилизацию, то естественно, этот примечательный факт требу¬
ет объяснения. Нас не может удовлетворить предположение о том, что чи¬
сто случайно великие цивилизованные расы изначально занимали только те
регионы земли, где цивилизация столько времени оставалась неизвестной.Предполагалось, что исчезновение правящих рас чужеземного проис¬
хождения в древних цивилизациях происходило в силу селективного вли¬
яния климата. Эта теория находится в том же положении, как и теорияо смешении рас, по отношению к основной критике, которой подвергалась
последняя. Она не объясняет, по какой причине потребовался иностранный
элемент. Если не считать этого, она, конечно, кажется в этой ситуации ре¬
альной. Судя по уровню смертности, расы, подверженные влиянию нового
климата и незнакомых болезней, естественно, находятся в неблагоприятных
условиях. Таково положение негров среди населения, инфицированного ту¬
беркулезом, или европейцев в регионах, подверженных вспышкам малярии.В одном отношении теория отбора по климату и болезням оказывает¬
ся предпочтительной в сравнении с теорией расового смешения. Если бы
последняя была единственным действующим фактором, то исчезновение
правящего класса сопровождалось бы постоянным улучшением коренных
жителей. Влияние последовательных завоеваний должно было бы накапли¬
ваться; поэтому нам следовало бы ожидать, что такой народ, как египтяне,
далеко бы продвинулся в направлении типа правящей расы. Но в действи¬
тельности — наоборот. Последствия селективного влияния климата и болез¬
ней, с другой стороны, кажется, полностью уничтожили бы преимущества
расы завоевателей. Дальнейшее рассмотрение показывает, что совместное
действие этих двух факторов привело бы к немедленному результату, так
как в распределении наследственных факторов иммунитет к местным бо¬
лезням часто комбинируется с особенностями иммигрантов. Такое рассмот¬
рение вопроса предполагает, что пока отбор не направлен непосредственно
против особенностей правящей расы как таковой, такое постоянное улуч¬
шение местного населения является необходимым следствием. С другой
стороны, положительный отбор в сочетании со смешением рас приводит
к формированию народа, который сочетает в себе свойство акклиматиза¬
ции с ценными качествами захватчиков. Тогда оказывается (из истории
древних цивилизаций на Востоке), что высокий уровень репродукции нико¬
гда надолго не оставался привилегией правящих классов. Если мы примем
242Глава 11во внимание далее, что в любом упорядоченном обществе бремя поддер¬
жания численности населения всегда лежало на плечах родителей, и что
обладание состоянием рано или поздно приводило к социальным преиму¬
ществам, то можно предложить не только рациональное объяснение исчез¬
новения правящих классов иностранного происхождения, но и малочислен¬
ность необходимых типов способностей коренного населения. С этой точки
зрения не кажется невозможным, что в своем происхождении цивилизация
была продуктом коренного населения, поддерживалась естественными при¬
чинами, которые допускали наибольшую плотность популяции; что боль¬
шая часть наиболее способных членов этой исходной цивилизации образо¬
вала со временем высшие классы, что эти высшие классы медленно пропи¬
тывались факторами наследственного характера, которые вели к снижению
плодовитости, и что с последующим развитием социального продвижения
по всей социальной лестнице они были устранены из расы быстро нараста¬
ющим процессом. Их территория, располагающая огромными природными
богатствами и лишенная сильного и объединенного правительства, стала
естественной добычей ряда завоевателей. Поскольку очевидно, что есте¬
ственные причины в достаточной мере объясняют ослабление исходных
правителей, то и вопрос об исчезновении аристократий иностранного про¬
исхождения также имеет объяснение. Факторы, разрушившие основателей
цивилизации, способны разрушить и их преемников; при этом гораздо лег¬
че, если отрицательное воздействие отбора было поддержано смешением
рас и влиянием климата.Убежденность, что только лишь цивилизованные условия существова¬
ния послужили причиной вырождения рас, которые в этих условиях жили,
поддерживалась многими авторами и в большой мере поддерживается при¬
мерами диких территорий, которые обрели цивилизацию и разделили упа¬
док более продвинутого государства. Народы Римской Галлии и Британии,
которые изначально были достаточно грозными, во время распада Римской
империи оказались также неспособными поддержать и защитить свою ци¬
вилизацию, как и римляне. Они стали естественной добычей многочислен¬
ных и воинственных племен варваров, которые не пострадали от чужезем¬
ного климата или смешения с худшей по качеству расой.Ни одно объяснение того, каким образом цивилизация оказывает та¬
кое отрицательное воздействие, со ссылкой на наследование приобретен¬
ных признаков сейчас нельзя считать удовлетворительным. И в любом
случае должно быть подвергнуто сомнению, насколько качества, приобре¬
тенные в состоянии цивилизации, включающие трудолюбие и дисципли¬
ну, в сочетании с развитием интеллектуальных способностей можно счи¬
тать неуместными для под держания народом самого себя. Говорят, что рос¬
Социальный отбор по фертильности243кошь ослабляет силу, а зависимость — инициативу цивилизованных наро¬
дов. И даже если бы эти слова были безоговорочной истиной в отношении
последствий цивилизованной жизни для индивида, то после того, как при¬
шла беда, первое же поколение должно было бы восстановить силу народа,
который находится под угрозой исчезновения, и предоставить беспример¬
ные возможности для развития его способности к активным самостоятель¬
ным действиям.Нам известно, что поток социального продвижения на протяжении
римского правления должен был прогрессировать очень быстро. Спрос
на людей для служения императору свободно распространялся и на провин¬
циалов, а в коммерческом и промышленном развитии провинций большая
свобода действий была предоставлена всем, кто способен к активным са¬
мостоятельным действиям, даже в родных городах. Высшие классы, и осо¬
бенно более богатые из них, были, естественно, вовлечены в водоворот
многонационального общества, выдвигаясь, преимущественно, из нижних
слоев. Галлы были допущены в Сенат еще в правление Клавдия, который
первым захватил Британию. Если учесть данные факты, то неудивительно,
что после нескольких сотен лет поглощения Римской империей Британия
и Галлия продемонстрировали едва ли большую способность к сопротивле¬
нию, чем более старые провинции Рима. И даже если завоевание Британии
представляло трудность для варварских племен, то сама возможность свер¬
жения организованного римско-британского общества указывает на то, что
ресурсы острова использовались далеко не лучшим образом.Контраст с варварскими обществамиСоциальный отбор бесплодия, кажется, будет характеристикой любых
состояний общества, в котором (1) есть различия между социальными клас¬
сами; (2) богатство влияет на определение социального положения индиви¬
да или его потомков; (3) экономические затраты по становлению следующе¬
го поколения несут только или в основном родители, по крайней мере в том
смысле, что они могли бы начать жить лучше, если бы детей было меньше.
Эти условия находят такую широкую применимость, что можно, не со¬
мневаясь, постулировать их во всех обществах, древних и современных,
состоящих из индивидов, сотрудничающих для взаимной выгоды в стране,
где есть закон и порядок. Признаки ситуации, которая в высшей степени
способствует вырождению, можно тем не менее обозначить более четко,
сказав, что они вытекают из дифференциации социального класса, в ко¬
тором социальное продвижение или понижение определяется сочетанием
двух разных качеств: (1) социально ценными качествами и (2) бесплодием.
244Глава 11Таким образом, будет не совсем точным сказать, что рассматриваемый се¬
лективный фактор должен способствовать вырождению в любом обществе,
где есть разделение по величине богатства. Это было бы слишком широким
применением принципа, поскольку степень богатства в форме управления
материальной собственностью и предоставляемыми услугами неотделима
от любой организации общества, какой бы она ни была.Несомненно, существовали общества, хотя их и нельзя с уверенно¬
стью назвать цивилизованными обществами, в которых институт социаль¬
ных классов был хорошо развит, в которых власть и престиж индивида
покоились на происхождении и родстве, и где эти классовые отличия были,
несомненно, скоррелированы с личным состоянием. Тем не менее социаль¬
ным преимуществом в этих обществах пользовались большие семьи. По¬
скольку, рассматривая такие общества, нам следует исходить из принципа
наследования фертильности и отсутствия любых компенсирующих причин,
из того, что фертильность классов, обладающих социальным превосход¬
ством, должна была бы быть выше, и, следовательно, мощная эволюци¬
онная сила, вызвавшая такое различие в фертильности, была направлена
на усиление качеств, благоприятствующих преуспеванию в таких обще¬
ствах и вызывающих в обществе чувство уважения, то значение изучения
таких обществ для нашей теории и для эволюционной истории человече¬
ства весьма велико.Состояние общества, которое нас интересует и которое мы рассмат¬
ривали на примере примитивных народов Северной Европы, описанных
в исландских сагах, в трудах Тацита о германских народах и, возможно,
в поэмах Гомера, доисламских бедуинов Аравийской пустыни, многих, ес¬
ли не всех, турецких и татарских народов степей Центральной Азии, поли¬
незийцев Новой Зеландии и Самоа, характеризуется родовой организацией,
на которую оказывает влияние или в действительности — даже доминирует
кровная месть. Во всех этих обществах наблюдается почтение к аристокра¬
тии и классовым отличиям и, наряду с кровной местью, эти черты доволь¬
но редко встречаются среди нецивилизованных обществ в целом. По этой
причине удобно обозначить такой тип общества специальным термином,
что поможет отличить его от цивилизованных обществ и других нецивили¬
зованных народов. Мы можем для удобства называть их варварами. Этот
термин является наиболее подходящим, поскольку приведенные примеры
включают самые важные группы народов, захвативших на протяжении ис¬
тории огромные центры цивилизации, которые просуществовали достаточ¬
но долго, и в отношении данных народов сложившаяся организация обще¬
ства может быть прослежена в исторической непрерывности в обратном
порядке.
Социальный отбор по фертильности245Разные цивилизованные нации, хотя потенциально между ними воз¬
можны войны, все же демонстрируют примерно одни и те же идеи и равное
развитие материальной цивилизации. Культурное единство разных варвар¬
ских племен обычно гораздо ближе, поскольку они связаны общим языком
и общей устной литературой. У них гораздо чаще возникают смешанные
браки, чем это возможно в больших конгломератах, составляющих циви¬
лизованные нации, одни и те же торговые ярмарки, общие религиозные
праздники и развлечения. Такое единство культуры необходимо подчерк¬
нуть, потому что, в отличие от цивилизованных обществ, в которых един¬
ство может быть относительным, варварские народы признают личную или,
точнее сказать, родовую войну как естественное средство отмщения и кон¬
троля преступности. Обязанность отомстить за родственника считалась мо¬
ральным долгом, уклонившийся от такой обязанности не может сохранить
уважение к самому себе или чистую совесть, и его выполнение, по точному
выражению Уилфреда Бланта, является «почти физической потребностью».
Кровная месть предполагает, что четко определены племена, которые ей
следуют, вслед за этим — обязанности по выплате денег и право на свою
долю, виру* или на долю трофеев. Таким образом, для подобных родствен¬
ных групп характерна некоторая степень экономического коммунизма, так
что едва ли будет большим преувеличением сказать, что экономические
и военные подразделения в таких обществах формируются так, что они
совпадают. Это по крайней мере удобная форма, чтобы отразить отличие
от цивилизованных обществ, в которых интересы экономического подраз¬
деления, состоящего из одного индивида и его подчиненных, могут сильно
отличаться от интересов военного подразделения, к которому можно отне¬
сти всю нацию. Интересы родственной группы в целом, права на жизнь
и собственность которой можно защитить только военной готовностью,
в первую очередь, естественно, основываются на военной силе, и, следова¬
тельно, среди других качеств — на фертильности членов этой группы.Социальное положение в варварских обществах несколько отличает¬
ся по структуре и степени. Сильная группа состоит не только из части
свободных соплеменников коренного, или благородного, происхождения,
но также подчиненных соплеменников, которые нашли в данном племени
убежище или которых могли изгнать из другого племени; кроме того, они
могут представлять остатки других племен, слишком слабых, чтобы посто¬
ять за себя; помимо указанной группы, в племени могут быть несвободные
люди, живущие на положении слуг в цивилизованных обществах, с тем от¬
личием, что они не защищены гражданскими законами, а с другой стороны,* Деньги, выплачиваемые следующему в роду после убитого при кровной мести.
246Глава 11они лишь немногим отличаются от своих хозяев по образованию и уров¬
ню жизни. Здесь важно отметить, что различия в социальном положении
в варварских обществах чувствуются по меньшей мере так же сильно, как
и в цивилизованных, и основываются они не столько на профессиональ¬
ных занятиях, сколько на личном и семейном авторитете. Такие различия
в авторитете не ограничиваются социальными отличиями внутри племе¬
ни, а распространяются на большие различия в общем мнении и уважении
разных родственных групп в соответствии с их успехами и властью.Таким образом, следует отметить, что в описанных нами варвар¬
ских обществах вполне определенные социальные различия комбинируют¬
ся с отличительным социальным преимуществом родов, в большей степени
способных к размножению. Не подлежит сомнению и независимое свиде¬
тельство того, что наиболее уважаемые семьи обладали и самой высокой
способностью к размножению. Огромное значение придавалось происхож¬
дению, свидетельством тому являются усилия, предпринимаемые для со¬
хранения имен предков, даже самых отдаленных. Таким образом, мы можем
понять один из факторов, который позволил таким народам построить свою
социальную систему на кровном родстве. Более того, эти данные свиде¬
тельствуют не только о более высоком уровне рождаемости, но и большем
естественном приросте, если принимать во внимание уровень смертности.
Можно также предположить, что во время войн семьи, занимающие высо¬
кое положение в обществе, несли больший урон от смертности, но при су¬
ровых потерях, которые варварские народы испытывали во времена голода
или вынужденных миграций, более сильные группы, несомненно, находи¬
лись в более выгодном положении; и если, как мы уже рассмотрели выше,
действие отбора было направлено на увеличение их природной фертильно¬
сти, большей, чем у менее знатных групп, неудивительно, что нам следует
сделать вывод, что общество поколение за поколением формировалось пре¬
имущественно из его самых успешных членов.Самым важным следствием такого вывода является то, что эволюция
человека, по крайней мере в определенных очень древних состояниях об¬
щества, проходила под влиянием фактора гораздо более действенного, чем
прямой отбор индивидов, а именно путем социального продвижения фер¬
тильности в высшие социальные слои. В частности, важно то, что качества,
признаваемые человеком социально ценными, должны были стать целью
такого селективного фактора, которому до этого эволюционное развитие
можно было приписать только в отношении отбора целых организованных
групп, сравнимых с роями общественных насекомых. Однако отбор целых
групп — это гораздо более медленный процесс, чем отбор индивидов, и,
принимая во внимание длительность поколения у человека, эволюция этих
Социальный отбор по фертильности247высших умственных способностей, и особенно элемент самопожертвова¬
ния в моральной природе человека, должны были пройти через групповой
отбор протяженностью в огромный временной промежуток. Больше того,
к более высоким человеческим способностям следует причислить и спо¬
собность эстетической оценки таких качеств или эмоциональный отклик
на также качества, которые мы считаем высшими в человеческой природе.
Оценить степень социального преимущества фертильности в варварских
обществах поможет подробный анализ ее реакций на одну группу качеств,
типичных для культуры варваров.Героизм и высшие способности человекаСреди социальных идей всех известных нам народов на стадии перехо¬
да от варварских условий доминирующим являлось понятие героизма; как
правило, цивилизованные народы оглядываются назад, к так называемо¬
му «героическому прошлому», когда данное понятие в значительное мере
формировало структуру общества. Эмоциональное влияние этой идеи бы¬
ло настолько велико, особенно ярко выраженное в поэтических традициях,
что трудно дать формально точную характеристику феномена, не используя
язык, наполненный высокопарными высказываниями. Напомним читателю,
что мы заинтересованы не в том, чтобы оценить героизм, восхваляя его
или преуменьшая его достоинства, но лишь в том, чтобы рассмотреть его
природу и последствия как социологического феномена.Герой — это человек, который сообразно своему характеру готов встре¬
тить опасность; следовательно, он неизбежно обладает определенным ав¬
торитетом во время опасных предприятий, поскольку люди с охотой по¬
следуют лишь за тем, кто способен дать им надежду на успех. Опасные
и рискованные предприятия, однако, не являются необходимостью, толь¬
ко люди в качестве врагов или лидеров делают их таковыми; и тот почет,
которым традиция окружает определенные формы явного безрассудства,
нельзя отнести на счет какой бы то ни было объективной оценки шансов
на успех. Очевидно, что для современного человека, унаследовавшего эти
качества чрезмерно, возрастает вероятность погибнуть молодым в какой-
нибудь, возможно, полезной дальней экспедиции, а кроме того, возрастает
вероятность выбрать профессию, которую трудно совместить с семейной
жизнью. Несомненно, что современный социальный отбор не благоприят¬
ствует героизму, по крайней мере в той степени, в какой общество находит
приятным и правильным для героя отдать свою жизнь за родную страну.
Я уверен, что любая большая война выявит большой резерв скрытого геро¬
изма в обществе почти у любого народа, хотя в то же время любая большая
248Глава 11война в значительной мере истощит этот резерв. И никто тем не менее
не будет сомневаться в том, что в отношении благоразумия давно циви¬
лизованные народы действительно принципиально отличаются от народов,
у которых стремление прославиться своей героической победой было выс¬
шей целью почти каждого мужчины.Исследование действия отбора в варварских обществах в условиях
племени или рода открывает возможности отбора героических качеств в та¬
кой степени, которая выходит за рамки благоразумия, посредством метода,
аналогичного тому, который описывался в главе 7 для объяснения эволю¬
ции неприятных качеств личинок насекомых. Сам факт, что процветание
группы подвергается риску, означает, что жертвовать своей жизнью бы¬
вает иногда выгодно, хотя, я полагаю, это — несущественное соображе¬
ние по сравнению с тем огромным преимуществом, которое приносит всем
родственникам авторитет героя. Весомое преимущество такого авторитета
в варварском обществе, думаю, не вызывает сомнения; об этом свидетель¬
ствует вся героическая литература; прямым свидетельством является наро¬
читое восхваление достижений рода профессиональными поэтами; в рав¬
ной мере убедительным представляется огромное значение, придаваемое
в таких обществах происхождению, когда живые хвастаются тем, что их
предками были могущественные люди. Благородной целью, которая стоит
перед героем, является неувядающая слава, и, следовательно, он связан со
всем, что имеет хорошую репутацию; по сравнению с героикой меркнут
все материальные преимущества. Большое значение имеет сам феномен ге¬
роизма, но не менее важно и то отношение почета и уважения, которым
героизм постоянно окружен.При родовой структуре общества неизбежным является выдвижение
какого-то рода над другими по героическим качествам. Если предположить,
что такие качества действительно приносят пользу тем, кто принадлежит
к данному роду, и что эту пользу легче всего истолковать как эффект пре¬
стижа, то в родовом обществе героизм может стать доминирующим каче¬
ством. В таком случае результат полового отбора представлен человеком,
который сыграл самую важную роль. Здесь я не буду акцентировать на по¬
этической традиции, которая, несмотря на общее мнение об изнеженности
и женственности поэтов, настаивает на связи героизма с истинной любо¬
вью. Скорее, мне следует полагаться на действительные обычаи заключе¬
ния браков у варварских народов. Следует подчеркнуть, что политический
элемент в таких браках более очевиден, чем романтический, хотя и эмоци¬
ональные реакции присутствуют в неменьшей степени. Заключение брака
чаще всего ведет к обязательствам кровного родства; союз с сильным родом
высоко ценится. Объединенный род заинтересован в таком браке, и на по¬
Социальный отбор по фертильности249ловой отбор мощное давление оказывает мнение рода. Престиж договари¬
вающихся сторон крайне важен, и хотя это отчасти — личное дело, но в зна¬
чительной степени оно касается всего рода. Поклонник полагается на свою
репутацию даже в отношении решения самой невесты. Как в исландских
сагах, так и в доисламских поэмах браки почти всегда подсказываются по¬
литическими устремлениями сторон.Такой половой отбор, основанный на общественном мнении, должен
влиять на многие другие качества, помимо бесстрашия и героизма. Красота,
высокие идеи и любое другое особо почитаемое качество должны усили¬
ваться таким отбором. Его значение для нас в том, что он влияет на чувство
уважения, которое поддерживается в отношении группы качеств, наиболее
тесно связываемых с героизмом. Подобно тому, как у птиц способность
дискриминации самки находится под воздействием полового отбора pari
passu с украшениями, которые она оценивает, так и в варварском обществе,
где героические качества действительно обладают существенным родовым
преимуществом, способность ценить и склонность восхищаться такими ка¬
чествами будут возрастать, по крайней мере до тех пор, пока уровень репро¬
дукции в доминирующих семьях будет оставаться наибольшим. Читатель,
беспристрастно сравнивающий современное отношение к неосторожным
поступкам в любом на протяжении длительного времени цивилизованном
обществе, с тем, что мы видим у рассматриваемых народов, вряд ли усо¬
мнится в том, что почитание героев варварами было в действительности
умонастроением, которое, каким бы бесполезным оно ни казалось для со¬
временного человека, в жизни варваров играло существенную роль. Смена
обстоятельств, благодаря которым героические качества были лишены сво¬
его преимущества, перевернула также и способность узнавать и ценить их.
Очевидно, что варварский элемент в традиции нашей культуры — это тот
элемент, который подчеркивает, а в действительности — даже чрезмерно
подчеркивает природное неравенство человека, в то время как религиозный
и законодательный элементы выделяют его гражданское равенство. Из того
факта, что варвары более всего ценили определенные качества человече¬
ского характера, следует сделать справедливый вывод о том, что они чув¬
ствовали такие отличия лучше, чем цивилизованный человек. Нельзя дать
прямого доказательства этой точки зрения. По тем вопросам, по которым
мы информированы лучше, чем наши предки, нам легко распознать факты,
подтверждающие наше преимущество. Если бы было наоборот, то нашим
предкам было бы легче, чем нам, указать на это отличие. Единственный из¬
вестный мне объективный факт, который относится к настоящему вопросу,‘Наравне {лат.).
250Глава 11это то, что современники, поднаторевшие в анализе, обязательно находят
в дошедших до нас ранних примерах поэзии определенное трудноулови¬
мое качество в описании характера, которое придает такому стиху видимое
превосходство в том особенном виде литературы, которой данное произве¬
дение принадлежит.Конечно же, трудно провести различие между неспособностью вос¬
принимать и безразличием к воспринимаемым отличиям. Примером пол¬
ной противоположности произвольному выбору предмета страсти являет¬
ся отчет Риели о значимости университетской степени на бенгальской яр¬
марке невест. Успешный кандидат, который появлялся на рынке со сте¬
пенью магистра гуманитарных наук, немедленно становился, благодаря
своей иностранной квалификации, в высшей степени желанной партией
и за несколько месяцев мог собрать вокруг себя ряд обеспеченных невест.
Трудно сказать, возложили ли бы на коллегию экзаменаторов ответствен¬
ность отобрать кандидатов для полового отбора по характерным чертам
личности, которые до некоторой степени еще оказывают влияние на влюб¬
ленных Европы и которые, если и распознаются, то значение им придается
гораздо меньшее, чем университетской степени.Среди народов со значительным содержанием варварского элемента
в социальной традиции доминирующим неличностным фактором в отбо¬
ре супруга является социальный класс. Социальный класс, как мы видели,
лучше всего можно определить как синтез таких отличительных черт, как
богатство, профессия и семья, поскольку они оказывают влияние на прием¬
лемость выбора в браке, учитывая эти отличия только в той степени, в ка¬
кой они действительно влияют на такую приемлемость. Исторически это
отличие социального класса является непрерывным продолжением полити¬
ческого и романтического авторитета, присущего доминирующим кланам.
Таким образом, неслучайно, что выставление напоказ своего социального
положения, характерное для ранних времен, должно было с наибольшей
очевидностью сохраниться в браке. Затухание и упадок чувства классового
отличия является необходимым сопутствующим обстоятельством постепен¬
ного исчезновения тех элементов населения, которые обладают более вы¬
соким положением. Нетрудно понять, что если предположение о том, что
среди варварских народов доминирующий класс действительно пользовал¬
ся селективным преимуществом, то аристократические институты для на¬
родов, недавно обретших цивилизацию, должны казаться основанными
на чувстве естественной справедливости.Обобщим социологически значимые моменты: (1) варварские народы,
организованные в родственные группы и признающие кровное родство,
скорее всего, подверглись отбору в пользу двух качеств, от которых, глав¬
Социальный отбор по фертильности251ным образом и зависит успех родственной группы — (а) патриотический
или героический настрой общества и (б) фертильность. (2) Стратифика¬
ция общества по этим двум качествам предполагает селективное преиму¬
щество героического темперамента, выходящее за пределы оптимального,
которым считается благоразумная смелость; это обусловлено социальной
выгодой славы или героической репутации. (3) Способность распознавать
героические качества и сознательный выбор при вступлении в смешанный
брак представляют двойной результат полового отбора, который состоит
в усилении обоих качеств, а также в их общественном признании и оценке.(4) Такой отбор популярной эмоциональной реакции на героические каче¬
ства имеет важные следствия: (а) стабилизация основ системы путем уси¬
ления существующей основы социальной сплоченности, (б) усиление се¬
лективного преимущества, придаваемого славе или престижу, (в) усиление
селективного преимущества всех качеств, сознательно намеченных к по¬
ловому отбору, (г) чрезмерное подчеркивание естественного неравенства
путем развития радикальной аристократической доктрины о наследствен¬
ном благородстве. Важно отметить, что такие обычаи, как многоженство
или зависимое внебрачное сожительство, никоим образом не являются ос¬
новными принципами описываемой системы причин, но могут быть впле¬
тены в эту систему в той степени, в какой они согласуются с авторитетом
героя или фертильностью его класса. Такие обычаи неизбежно затухают
или трансформируются для исполнения второстепенной социальной цели,
такой как домашняя работа, как только основные условия системы оказы¬
ваются подорванными.Место социального класса в эволюции человекаПохоже, человеку присуще сочетание условий, позволяющее исполь¬
зовать дифференциальную фертильность для ускорения эволюционных из¬
менений, прогрессивных или деструктивных. Это требует социального ор¬
ганизма, индивидуалистичного в репродукции. Дополнительными важны¬
ми факторами являются гибкость эффективной фертильности, выражаемая
обычаем детоубийства и другими методами ограничения численности се¬
мьи, и особенно тяжким бременем родительских забот в связи с продол¬
жительным периодом детства у человека. Вплоть до определенной ступени
развития, которая почти наверняка была у предчеловека, предками челове¬
ка были, несомненно, одиночные животные, и, пока не начала развиваться
социальная жизнь, их фертильность должна была стабилизироваться, до¬
стигнув оптимального или близкого к оптимальному уровня, соответству¬
ющего потребностям родительской опеки. Можно предположить, что убий¬
252Глава 11ство новорожденных начало практиковаться вследствие увеличивающегося
беспокойства по поводу неизбежных трудностей, появившегося на весьма
ранней стадии развития человека как социального животного. В течение
довольно длительного периода формирования социальной жизни человек
научился сотрудничать со своими сородичами, вероятно, в соответствии
с выражаемыми эмоциями, инстинктивно остерегаясь общественного по¬
рицания, увеличивая свои шансы на получение потомства путем отбора
пары. Чувство предпочтения кажется по меньшей мере настолько важным
в отношении половых инстинктов человека, что трудно сомневаться в том,
что половой отбор у человека начался на ранних ступенях его существо¬
вания, хотя вполне вероятно, что особые условия, испытываемые нашими
варварскими предками, создают у цивилизованного человека по этому во¬
просу предвзятое мнение. Помимо этого фактора, эволюция человека на его
ранних социальных стадиях развития должна была быть направлена глав¬
ным образом на формирование свойств, способствующих индивидуальному
выживанию в условиях социального окружения.С какой-то ступени, трудно предположить, с какой именно, общества
должны были придти к состоянию, в котором некая степень сплоченности
социального класса гарантировалась наследованием собственности, приви¬
легиями, авторитетом или социальными функциями и в котором индиви¬
дуальные различия в социально значимых качествах особенно отмечались
и использовались для оценки социального положения. Если эти свойства,
возможно, в элементарной форме, сочетались с родовой организацией об¬
щества, в котором права каждой родственной группы устанавливались в ко¬
нечном счете военной силой, так что наиболее фертильные смешивались
с теми, кем в большей степени восхищались, или с теми, кто обладал наи¬
более влиятельными чертами характера в доминирующих племенах, то на¬
правление и скорость эволюционного прогресса в таких обществах подвер¬
гались глубоким изменениям. Новая сила действовала через оценку челове¬
ком преимущества своих соплеменников, которая как в социальном отборе,
так и в половом отборе как его части, несомненно, сначала направлялась
в интересах родовой группы или общества в целом. Такой процесс обеспе¬
чивал средство относительно быстрого эволюционного прогресса качеств,
которые подчиняют личный интерес интересу других, и эволюция которых
только путем элиминации, отбором целых обществ была бы, скорее всего,
крайне медленной. Это, однако, несет с собой все опасности, отмеченные
в случае полового отбора, впадения в крайность: стандарты вкуса неизбеж¬
но будут модифицироваться шаг за шагом вместе с качествами, которым от¬
дается предпочтение, и действительно, может случиться, что некоторые из
этих качеств, те, что больше всего обращены к нашим мысленным образам,
Социальный отбор по фертильности253больше относятся к природе украшений, чем к полезным моральным иде¬
алам. Среди качеств, которые были чрезмерно развиты у всех варварских
народов,— личная и семейная гордость, и, за исключением тех жестких
уроков, которым необходимо следовать в суровых условиях, ужесточаемых
еще и войнами, крайности такого рода могут стать гораздо более общепри¬
нятыми, чем изначально присуще природе человеческой морали.Было бы интересно, насколько это возможно, сравнить скорость про¬
гресса варварских народов со скоростью упадка цивилизованных народов.
Вариаций, предлагаемых для эволюционных изменений деструктивного ха¬
рактера, должно быть больше, чем тех, что предлагаются для прогрессив¬
ных перемен, чтобы мы могли ожидать, что селективные повторы одного
и того же порядка займут сотни, а может быть, и тысячи поколений, чтобы
в совершенстве выстроить свойства ума, которые десять поколений проти¬
воположного отбора могли бы разрушить. Вряд ли должна существовать
большая разница в интенсивности отбора, возникающая в двух этих случа¬
ях, при условии, что и тот, и другой находились в стабильном состоянии.
Эффект различий по фертильности был бы больше в варварских услови¬
ях и меньше — у цивилизованных народов, так как преуспевание непо¬
средственно воздействует в пользу репродукции; и это могло быть при¬
чиной более интенсивного благоприятного отбора у варварских народов,
чем неблагоприятного отбора у цивилизованных. В качестве опровержения,
я полагаю, можно привести тот факт, что хотя нецивилизованные народы
с помощью детоубийства могут производить большую вариацию в чистой
фертильности так же, как и цивилизованный человек путем ограничения
рождаемости, но селективный эффект этих разниц по фертильности дол¬
жен в суровой среде быть во многом снижен большей смертностью в мла¬
денчестве и детстве, так что чистые разницы по фертильности могут быть
меньше. В отношении действия полового отбора можно считать, что этот
фактор, ускоряя процесс у варварских народов, должен сдерживать упадок
у цивилизованных, чего не наблюдается в обществе, в котором перспективы
фертильности уменьшены социальным продвижением, поскольку в таком
обществе в среднем из двух возможных браков биологически выгодным
будет заключить тот, который в социальном плане менее желателен. Более
того, люди, более тщательно подходящие к выбору партнера, будут чаще
всего уменьшать свою фертильность путем откладывания брака или пу¬
тем выбора безбрачия. Следовательно, половой отбор должен увеличивать
скорость развивающегося процесса, конструктивного или дегенеративного.
Однако интенсивность этого влияния должна быть снижена на более позд¬
них этапах существования цивилизованного общества вместе с угасанием
оценки индивидуальных разниц.
254Глава 11Аналогия с паразитизмом у муравьевВоздействие экономических причин на распределение фертильности
в цивилизации человека настолько разрушительно, что едва ли можно ожи¬
дать адекватных параллелей в обществах насекомых, поскольку у них отбор
между обществами кажется всегда возможным. Тем не менее предложен¬
ное Форелем толкование реакции муравья Tetramorium cespitum на доволь¬
но редкого паразитического муравья Strongylognathus testaceus достаточно
уместно, чтобы сравнить эту ситуацию с ситуацией у человека. Структу¬
ра паразитического муравья предполагает, что прежде он был муравьем-
амазонкой, но в данный момент слишком слаб, чтобы выступать в качестве
бойца, и смешанные колонии в отношении защиты полагаются на рабочих
хозяев. Бесполые же паразиты, по-прежнему способные прорывать ходы
и питаться самостоятельно, тем не менее очень мало или совсем ничего
не вкладывают в развитие всего муравейника, да и большую часть пищи
получают, скорее, с язычков муравьев-хозяев. Они не принимают участия
в заботе о молодом поколении или собственной королеве, будучи, таким об¬
разом, бесполезными для выживания своего вида, поэтому неудивительно,
что они воспроизводятся в относительно небольшом количестве по срав¬
нению с самцами и королевами, размер которых очень мал, а уровень ре¬
продукции велик. Половые формы хозяев, с другой стороны, относительно
очень крупные, поскольку королевам необходимо обеспечивать биологи¬
ческий запас для основания новых независимых колоний. В большинстве
случаев паразитизма у муравьев мать сообщества хозяев так или иначе
уничтожается, и собственно паразитизм состоит в эксплуатации социаль¬
ных инстинктов выживших рабочих для разведения муравьев чужого ви¬
да. Рассматриваемый случай специфичен в том отношении, что королева
хозяев продолжает жить и, наряду с королевой паразитов, даже осуществ¬
лять кладки в колонии, подвергшейся нападению паразитов; однако в таких
условиях она никогда не производит фертильных самок; таким образом,
паразиты получают постоянный источник пополнения рабочих муравьев-
хозяев, чтобы обеспечивать себя жильем, пищей, а также чтобы воспиты¬
вать и защищать их, в то время как из муравейника выходят лишь королевы
паразитов. Это обстоятельство Форел приписывает регулирующим и эко¬
номическим инстинктам рабочих муравьев-хозяев, так как самки и сам¬
цы муравьев-паразитов мельче и их легче прокормить. Очевидно, говорит
он, что вполне обоснованно рабочие-хозяева разводят их вместо собствен¬
ных огромных королев и самцов, личинки которых они, несомненно, по¬
жирают или игнорируют, как всегда поступают с тем, что представлено
в избытке.
Социальный отбор по фертильности255Прав или нет Форел в своей интерпретации, но его предположение
хорошо иллюстрирует следствие экономического закона, если слепо позво¬
лить ему регулировать фертильность. Поскольку паразитов находят лишь
в малой части муравейников Tetramorium, это означает, что у этих насеко¬
мых есть какие-то защитные инстинкты, позволяющие им успешно сопро¬
тивляться инвазии. В человеческих обществах человек является собствен¬
ным паразитом — обстоятельство, которое, похоже, гарантирует полную ин¬
вазию всех цивилизованных обществ.РезюмеВ соответствии с теорией, последовательно развитой Галтоном, Декан¬
долем и Коббом, инверсия уровня рождаемости является следствием двух
причин, которые были полностью доказаны: (1) наследование признаков,
физических или психологических, определяющих уровень репродукции;(2) социальное продвижение менее фертильных. Разные теории, которые
искали в богатстве причину бесплодия, не учли тот момент, что бесплодие
является важной причиной богатства.В свете этой теории можно понять, как произошло, что у более пре¬
успевающих классов было не только меньше детей при заключении бра¬
ка в данном возрасте, но, помимо этого, у них прослеживается тенденция
более поздних браков, а также более частых случаев безбрачия, чем у ме¬
нее преуспевающих классов. Нам теперь ясно, почему у бедных классов
в среднем больше детей, чем у богатых; кроме того, у людей выдающихся,
значительно улучшивших свое социальное положение, меньше детей, чем
у таких же известных людей, в меньшей степени продвинувшихся по со¬
циальной лестнице. С другой стороны, хотя у населения в целом семьи
большего размера менее успешных классов демонстрируют отрицательную
корреляцию между способностями ребенка, измеренными разными спосо¬
бами, и размером его семьи, но для групп детей, отобранных по более или
менее равным социальным возможностям, можно ожидать (так оно и есть
для студентов Йельского университета и в меньшей степени — для неко¬
торых английских школ среднего класса общества), что корреляция имеет
тенденцию возрастать к положительному значению. Все эти факты выгля¬
дели бы в высшей степени парадоксально с точки зрения, подразумеваю¬
щей, что различия в фертильности являются прямым результатом различий
в социальном положении.На примере упадка правящих классов показано, что ни смешение рас,
ни селективное действие климата и болезней не могут служить удовле¬
творительным объяснением, почему они пришли в такое состояние даже
256Глава 11при благоприятном отборе. Причины, по которым мы проследили инверсию
фертильности, были характерными как для большинства древних цивилиза¬
ций, так и для нашей собственной, и служат для объяснения исторической
важности правящих рас из-за отсутствия соответствующих характеристик
у коренных народов. По тем же самым причинам происходит неблагоприят¬
ный отбор, действующий последовательно на каждый побежденный народ.Упадок варварских народов, получивших цивилизацию и разделивших
упадок тех народов, развитие которых находилось на более продвинутом
этапе без воздействия чужого климата и смешения с превосходящей ра¬
сой, ясно виден из социального продвижения и пресечения рода его более
способных членов.Некоторые нецивилизованные народы, для которых характерна ро¬
довая организация, кровное родство и которые придают большое значе¬
ние родственным связям и происхождению, показывают состояние обще¬
ства, в котором наиболее известные оказываются и наиболее фертильными
и в котором следствия Естественного Отбора в значительной мере усилены
социальным и половым отбором. Действие указанных факторов представ¬
ляет особенную важность в отношении качеств, признаваемых человеком
как социально ценных, которые, таким образом, получают селективное пре¬
имущество намного большее, чем любое, которое могло бы быть приписано
дифференциальной элиминации целых родов. Группа качеств, понимаемых
этими варварскими народами как связанных с героизмом, была в значи¬
тельной мере развита за пределы оптимального индивидуального преиму¬
щества. Высшие умственные качества человека и особенно его признание
этих качеств можно приписать социальному отбору этого типа общества.Отбор в пользу этих качеств, вызывающих восхищение у варварских
народов, был, скорее всего, таким же интенсивным, как отбор в противо¬
положном направлении у цивилизованных народов. Деструктивный эффект
неблагоприятного отбора этих качеств должен тем не менее быть гораздо
более быстрым, чем процесс их создания.Если принять мнение Форела в отношении реакции муравья-хозяина
Tetramorium cespitum на его паразита Strongylognathus testaceus, то ситуа¬
ция, установившаяся в колониях, зараженных паразитами, в некотором роде
сравнима с экономическими реакциями цивилизованного человека на ре¬
продукцию. В то время как большинство семей Tetramorium старается огра¬
дить себя от паразитов, в человеческом обществе неизбежно проявляются
вариации, необходимые для обеспечения собственной инвазии.
Глава 12Условия долговременной цивилизацииАпология. Долговременная цивилизация не всегда непрогрессивна. Перерас¬
пределение рождений. Социальная поддержка фертильности. Недостатки француз¬
ской системы. Проблема существования народов. Резюме.Но если смерти серп неумолим,Оставь потомков, чтобы спорить с ним! У. ШЕКСПИР*Но добрая воля как бог всемогущий. СОФОКЛАпологияПредполагается, что все ученые принимают, по крайней мере теорети¬
чески, точку зрения, что распространение биологического знания в конеч¬
ном итоге будет служить практическим целям человечества; однако несо¬
мненно, что большинство людей, безусловно придет в замешательство, ес¬
ли их научные взгляды будут использованы для обоснования определен¬
ного и поэтому спорного политического курса. Для этого есть достаточное
основание: ученые, подобно всем остальным, еще до выбора своего призва¬
ния, а возможно и позже, склонны подпадать под влияние страстей и пред¬
рассудков относительно природы человеческих отношений; и если, веро¬
ятно, довольно легко критически и беспристрастно обсуждать, например,
эмбриологию морского ежа или строение звездной системы, то достаточ¬
ную критичность и беспристрастность нельзя гарантировать при изучении
того крута проблем, над которыми довлеют вышеназванные предрассуд¬
ки. Более того, любая небрежность в этом отношении вредна для Науки.
Ученый-энтузиаст, единственной целью которого является научное обосно¬
вание целесообразности проводимой социальной политики, не только об¬
манывает себя и, возможно, других, не подозревающих о его предвзятости,
но и неизбежно сеет сомнение в отношении всех остальных ученых, рабо¬
тающих в данной области. Вне всякого сомнения, лишь ученый, воздержи¬*Соне-г 12, перевод С. Я. Маршака.
258Глава 12вающийся от высказывания любого мнения по поводу человеческих отно¬
шений, сумеет сохранить свою научную репутацию и душевное спокой¬
ствие и снять с себя ответственность за любое практическое применение
полученных им знаний. Если бы такому ученому пришлось признать, что
человечество могло бы с большей выгодой использовать любое научное
знание, то его практические действия явно не пошли бы дальше рекоменда¬
ции более широкой пропаганды данного научного знания, а вопрос о прак¬
тическом применении знания он оставил бы на рассмотрение всем тем, кто
мог бы использовать его в своей конкретной области.Читателю станет ясно, что наши заключения об отдельных факторах,
оказывающих влияние на цивилизованное общество, находятся в опасной
близости к их практическому применению. Осознавая все недостатки та¬
кого подхода, мне бы казалось довольно циничным не попытаться опре¬
делить условия долговременной цивилизации, подобной нашей, установив,
как я полагаю, основное направление нестабильности человеческих циви¬
лизаций. Если бы эти условия включали далеко идущие и серьезные по¬
следствия, то мне бы хотелось надеяться на то, что они не будут объяснены
той самой научной безответственностью, против которой я так охотно вы¬
ступаю, а лишь глубоким характером обнаруженной нами причины неста¬
бильности.Долговременная цивилизация не всегда непрогрессивнаОчевидно, что постоянство цивилизации не всегда означает ее застой
или негибкость. Наша современная цивилизация, охватившая, несомнен¬
но, всю землю, еще не испытала окончательных реакций на собственное
развитие со стороны нескольких значимых групп людей. Но мы можем
тем не менее предположить, что опасаться нечего до тех пор, пока мы
не начнем считать, что для своего долговременного существования циви¬
лизация не должна развиваться; поскольку, пока она существует, прогресс
наук и искусств, очевидно, изменит среду, в которой живет человечество,
и, следовательно, изменит социальную значимость разных людей. В сущ¬
ности, необходимым условием постоянства цивилизации, подобной нашей,
является ее способность к биологическому развитию в том направлении,
которое диктуется современным состоянием общества. Даже если бы жест¬
кая система профессиональных каст, каждая из которых вынуждена нести
собственное бремя необходимой репродукции, гарантировала биологиче¬
ское постоянство подобно тому, как постоянство сообщества генов гаран¬
тируется делением клетки, все равно невозможно было бы считать это ре¬
шением проблемы непостоянства цивилизации. С другой стороны, данную
УСЛОВИЯ ДОЛГОВРЕМЕННОЙ ЦИВИЛИЗАЦИИ259фразу нельзя понимать как абсолютную гарантию невозможности ее раз¬
рушения по всем возможным причинам, а лишь как возможность земно¬
го бессмертия, присущего одноклеточным растениям и животным, которые
могут погибнуть и действительно гибнут в огромных количествах, но ко¬
торые не имеют неизбежных причин старения и распада своей внутренней
структуры.Перераспределение рожденийДля общества, способного к эволюционному прогрессу, наиболее оче¬
видным требованием, соответствующим его текущим потребностям, явля¬
ется то, что репродукция должна быть более активной у его более успеш¬
ных членов и менее активной — у менее успешных. По-видимому, это
условие имеет значительные преимущества в сравнении с существующим
в цивилизованных обществах распределением репродуктивности, не гово¬
ря уже о причине и способах формулировки этого условия. С его помо¬
щью можно будет снизить уровень крайней бедности, а также постепен¬
но осуществить равномерное распределение богатства. Уменьшится также
необходимость процесса передачи богатства бедным посредством прямо¬
го налогообложения богатых, и это будет приветствоваться теми, кто по¬
лагает, что данный процесс на практике является крайне неэффективным,
и что в действительности размер состояния, которое передается бедным,
оказывается намного меньше убытков, которые прямо или косвенно несут
при этом богатые люди. По моему мнению, существует лишь одно неже¬
лательное следствие перераспределения репродукции, а именно: богатые
люди потеряют возможность накопления капитала. Не отрицая, что более
адекватное распределение рождаемости среди более состоятельных классов
может привести к такому результату, представляется возможным усмотреть
в этом по крайней мере и компенсирующее преимущество: при меньшем
количестве свободных денег для осуществления инвестиций или по край¬
ней мере при необходимости повысить процентную ставку для сохранения
темпа накопления сбережений, капитал, изъятый из инвестиций в качестве
денег будет вложен в развитие людей средних способностей, с надлежа¬
щим образованием, хорошо подготовленных для работы. Такая форма ка¬
питала также приносит дивиденды и оказывает влияние на прибыльность
всех других вложений. Можно предположить, что богатство будет накапли¬
ваться, в основном, так же быстро, как и в настоящее время, только в виде
способных людей и в меньшей степени в виде кирпичей и строительного
раствора, предназначенных для строительства больниц и исправительных
учреждений.
260Глава 12Социальная поддержка фертильностиСогласно предположению главы 11, установление превосходства выс¬
шего класса по показателям фертильности возможно без посягательства
на свободу личности путем социальной пропаганды более фертильных ли¬
ний в популяции, подразумевая под ними тех, кто более других физически
или физиологически способен производить детей на свет. Каким бы отли¬
чающимся от системы выплат и жалований, действующей в стране в насто¬
ящее время, данное условие не являлось, при исследовании, несомненно,
становится очевидным, что систему заработной платы можно с выгодой
модифицировать в данном направлении; это ясно как с точки зрения чисто
экономической функции системы выплат, так и с точки зрения современных
принципов естественного права. Давайте отложим рассмотрение механиз¬
ма, с помощью которого будут приняты объективные меры и достигнуты
соглашения между разными работодателями, и просто сравним две систе¬
мы, посредством которых могут быть определены суммы, получаемые ра¬
бочими. (А) Равная плата за равный труд, допускающая значительные раз¬
личия по уровню жизни равных в промышленном отношении работников,
которые обеспечивают разные по величине или уровню расходов и сбе¬
режений семьи с премией за бездетность, составляющей примерно 12%
от основной зарплаты за каждого неродившегося ребенка. (Б) Равный уро¬
вень жизни за равный труд, включение в зарплату семьи денежных посо¬
бий, эквивалентных действительным средним расходам на ребенка. Срав¬
нив указанные системы, мы увидим, что вторая система (Б), прекращая
социальную пропаганду фактического бесплодия, обладает экономическим
преимуществом, способствуя росту производства. Система оплаты труда
упрощена для работника, образ жизни которого сейчас полностью зависит
от ценности той работы, которую он способен выполнять. Полностью пе¬
реориентированной в данное направление является сейчас система диффе¬
ренцированных жалований и окладов как способ, используемый обществом
для того, чтобы побудить работника напрямую или через частного нанима¬
теля выполнять значимую для общества работу. Она уже не направлена, как
в первой системе (А), на то, чтобы заставить его не только выполнять необ¬
ходимую работу, но и отказываться от выполнения родительского долга.Основным мотивом системы денежных выплат семьям, принятой сей¬
час во Франции, была необходимость обеспечить детей бедных в послево¬
енный период (после войны 1914-1918 гг.), когда французская промышлен¬
ность не могла поддерживать достаточный уровень жизни рабочих, если
на нее взваливалась еще и обязанность обеспечения лучшего уровня жизни
тем, кто не имеет детей. В этом отношении можно считать, что система
Условия долговременной цивилизации261оплаты труда во Франции предполагает снижение уровня бедности без уве¬
личения заработной платы предприятиями. Однако в результате средства,
высвободившиеся после перераспределения зарплат, были поделены между
наемными работниками и предприятиями, на которых они работают, при¬
чем выгоду получило и большинство потребителей путем снижения цен.
Легко понять, что как только стало ясно, что такого преимущества можно
достичь простым формированием общего для разных нанимателей компен¬
сационного фонда, система получила быстрое распространение. В действи¬
тельности суммы, внесенные или выплачиваемые из этих фондов на счет
индивидуальных нанимателей, были, конечно, небольшими, поскольку лю¬
бой крупный наниматель включал в сумму зарплат пособие на некото¬
рое количество детей, примерно пропорциональное его общему месячно¬
му фонду заработной платы; но эти фонды играют тем не менее важную
роль в функционировании системы, лишая отдельных нанимателей эконо¬
мического стимула, если они предоставляют работу преимущественно без¬
детным людям, в противном случае стимул появляется. Любой наниматель,
обеспечивающий через семейные пособия, включенные в зарплату, число
детей, превышающее среднее, получит из фонда компенсацию от других
нанимателей, которые обеспечивают число детей меньше среднего.Вторым мотивом для принятия системы денежных выплат семьям
во Франции было национальное стремление остановить тенденцию сни¬
жения численности населения вследствие недостаточного для замещения
взрослого населения страны числа рождений. Во Франции уровень рож¬
даемости начал снижаться несколько раньше, чем в других странах, и на¬
чало снижения было плавным, по этой причине понимание этого факта
во Франции было шире, чем сейчас в других европейских странах, осо¬
бенно в Великобритании, которой после внезапного падения уровня рож¬
даемости гораздо труднее, чем Франции, удается сохранить численность
населения. Однако, несмотря на то, что экономические цели системы опла¬
ты труда, как то: повышение уровня жизни бедных, удовлетворение спроса
предприятий на рабочую силу, полная занятость работников промышлен¬
ности,— принесли вполне удовлетворительные результаты, биологическая
цель повышения уровня рождаемости так и не была достигнута; даже в тех
союзах, где денежные выплаты для более поздних детей были достаточны¬
ми для того, чтобы дать позитивный экономический стимул дальнейшему
материнству и отцовству. В 1927 году при подсчете взрослого населения
по возрастному показателю выяснилось, что уровень детского населения
на 9 % меньше того количества, которое необходимо для поддержания ста¬
бильной численности населения. Этот дефицит детского населения продол¬
жает нарастать в последние годы, хотя и гораздо медленнее, чем в Англии,
262Глава 12Германии или Скандинавии. Оценка этого же показателя в Восточной и Се¬
верной Европе в целом в 1926 году выявила дефицит в 7%, и наблюдая
быстрое падение рождаемости, свойственное многим странам данных ре¬
гионов, становится ясно, что этот дефицит быстро нарастает. Для Англии
и Уэлса в 1927 году он оказался около 18 %.Следовательно, можно поспорить с тем, что введение денежных посо¬
бий в систему зарплат во Франции оказало некоторое влияние на снижение
уровня населения; хотя, если в конечном счете данная мера все же ока¬
жется эффективной, следует отметить, что она принесет результаты лишь
через некоторое время. Лишь в будущем мы сможем с уверенностью за¬
явить о том, что система оказала положительное влияние на поколение,
по отношению к которому она была применена, и была ли она неэффек¬
тивной по причине существования высокооплачиваемых групп населения.
Значение системы состоит в том, что она представляет собой единствен¬
ный способ контроля за сокращением популяции, который с уверенностью
можно считать эффективным. Значительное сокращение населения наблю¬
дается сейчас в Северной и Западной Европе, причем с уверенностью мож¬
но ожидать такого же сокращения численности англоязычного населения
и в других частях света.Здесь мы не будем, естественно, рассматривать проблему регулирова¬
ния плотности населения в целом, однако необходимо отметить, что про¬
тест против того, чтобы система заработной платы использовалась в от¬
ношении стимулирования бездетности, потому что никакие другие методы
не являются в достаточной мере эффективными, чтобы избежать перенасе¬
ления, не может всерьез рассматриваться теми, кто знаком с современными
технологиями.Недостатки французской системыСистема выплат семейных пособий, действующая сейчас во Франции,
не может считаться эффективной в отношении нейтрализации социального
преимущества менее фертильных людей — цели, которой она изначально
не преследовала. Это особенно ясно, если учесть ее неспособность уста¬
новить семейные пособия, пропорциональные основной заработной плате
по всей шкале профессий, предусматривающих фиксированный оклад; по¬
скольку при отсутствии таких пособий тенденция к исчезновению высших
уровней интеллектуальных способностей, которые особенно необходимы
в таких профессиях, не должна иметь никаких ограничений. Нам следует
предположить, что существование долговременной и прогрессивной циви¬
лизации предполагает отсутствие этого недостатка, требование тем более
УСЛОВИЯ ДОЛГОВРЕМЕННОЙ ЦИВИЛИЗАЦИИ263сдержанное, потому как система представлена одними и теми же экономи¬
ческими преимуществами, к каким бы уровням дохода она ни применялась.
Более серьезная трудность заключается в отказе от социальной пропаганды
менее фертильных в категории тех профессий, где получают не фиксиро¬
ванные оклады и зарплаты, а вознаграждения в виде гонораров или вы¬
плат, поступающих от многих индивидов. Для населения, которое стремит¬
ся к лучшему для себя как в биологическом, так и в экономическом смысле,
экономическое положение этих профессий представляет трудность; хотя,
если имеются благие намерения и твердое следование намеченному кур¬
су, трудность будет заключаться скорее в формировании, нежели в самом
действии системы взаимного страхования, которая должна обеспечить пе¬
рераспределение дохода подобно тому, как происходит перераспределение
фиксированных окладов. Если основная часть населения на опыте убеди¬
лась в эффективности мер, предусматриваемых системой, для тех, на кого
они направлены, то маловероятно, что остальная его часть окажется неспо¬
собной сформулировать и использовать схемы сотрудничества, удовлетво¬
ряющие ее потребности.Я буду считать, что цель, поставленная мною в предыдущих абзацах,
будет достигнута, если читатель проникнется пониманием того, что для ци¬
вилизации, подобной нашей, невозможно создать такую структуру, которая
бы позволила ей избежать всех тех разрушительных биологических послед¬
ствий, которые не удалось преодолеть нашей собственной цивилизации.
Следует предположить, что можно разработать гораздо более эффективные
меры. Моя точка зрения заключается лишь в том, что хотя проблемы био¬
логического выживания популяции и имеют непосредственное отношение
к ее экономической организации, они не навязаны человечеству экономи¬
ческой эффективностью.Проблема существования народовЕсли отойти от относительно легкой проблемы точного определения
условий, благодаря которым человеческое общество может избежать небла¬
гоприятных биологических последствий действующих селективных сил,
которые угрожают человеческому обществу, и заняться проблемой уста¬
новления равных выгодных условия для народов нашей цивилизации, оче¬
видно, что здесь мы столкнемся с необходимостью найти гораздо более
действенные методы, чем те, которых было бы достаточно для решения
менее серьезных проблем. За последние несколько веков у всех существу¬
ющих народов уже были определенные характеристики, получившие со¬
циальную пропаганду в обществе. Так, это было показано при выявлении
264Глава 12уровня интеллектуальных способностей путем сравнения средних показа¬
телей у детей родителей, принадлежащих к разным социальным группам,
при выполнении тестов, которые, по мнению психологов, наиболее эффек¬
тивно выявляют интеллектуальный уровень человека. Возможно, еще более
важным является то, что множество качеств нравственной природы, таких
как желание преуспевать, сила духа и настойчивость в преодолении трудно¬
стей, справедливая мужественность хорошего лидера, инициатива и вообра¬
жение, существенных не только для прогресса, но даже для устойчивой ор¬
ганизации общества и его развития наравне с интеллектуальными способ¬
ностями, имеют тенденцию к концентрации в более преуспевающих слоях
существующих популяций. Однако эти классы также характеризуются и бо¬
лее высокими показателями бесплодия. По известной нам причине для них
не характерна ни нормальная репродуктивность в существующих экономи¬
ческих условиях, ни раннее и относительно безрассудное заключение брач¬
ного союза, которое оказывает благоприятное воздействие на репродуктив¬
ность. В такой стране, как Англия, эти классы сейчас обеспечивают, вероят¬
но, менее половины того количества детей, которое необходимо для сохра¬
нения численности населения, и нет сомнений в том, что этот показатель
по-прежнему быстро снижается. Реформатор должен ожидать, что встретит
глубоко укоренившееся сопротивление в тех классах, на которые он есте¬
ственным образом полагается в смысле разумного беспокойства о будущем
их страны, поскольку для многих представителей этих классов социальное
развитие их предков и их собственное преуспевание обусловлены не столь¬
ко служением обществу, сколько врожденным дефицитом репродуктивных
инстинктов. Тем не менее, хотя с этой стороны следует ожидать сопро¬
тивления, нельзя с определенностью утверждать, что представители более
преуспевающих классов, которые получают оклады, не поддержат систему
адекватных семейных пособий, достаточных для того, чтобы сохранить их
численность после первоначальной потери значительной части ценных ка¬
честв, имеющихся сейчас у этих классов. Снижение фертильности преуспе¬
вающих классов происходило быстрее, чем среди остального населения, и,
поскольку эти классы более восприимчивы к экономическим идеям и со¬
ображениям расчета, можно ожидать, что их реакция на введение системы
семейных пособий, достаточных, чтобы покрыть расходы на детей, может
также идти быстрее и в конечном счете эффективнее, чем у других классов.
Если бы это было так, и если бы в то же время предпочтительное продви¬
жение бесплодных родов из менее преуспевающих классов полностью пре¬
кратилось, то возможно, что фертильность высших классов могла бы быть
восстановлена путем дифференциального исчезновения менее фертильных
родов за относительно недолгий исторический период.
УСЛОВИЯ ДОЛГОВРЕМЕННОЙ ЦИВИЛИЗАЦИИ265Наиболее труднопреодолимым препятствием здесь станет длитель¬
ность периода, в течение которого будут устранены все факторы, являю¬
щиеся причиной разрушения народа. Предшествующий опыт показывает,
что невозможно гарантировать отсутствие жестких политических преврат¬
ностей в будущем. Путь развития каждой нации будет, несомненно, изви¬
листым под воздействием различных факторов, и не следует ожидать, что
все народы не изменят своих намерений на пути своего развития, даже
если они осознают его перспективы. Народы, самодостаточные по своей
природе и расколотые взаимным недоверием, не способны не только при¬
ложить усилия, но даже и попытаться сделать это. Однако это не должно
стать причиной, по которой люди доброй воли могут позволить себе в от¬
чаянии отказаться от такого влияния во благо народа, если они способны
его оказать.Народ, у которого нет широко признанного рационального мотива
для репродукции, но которого подталкивает к этому преимущественно
неосторожность и религиозный предрассудок, неизбежно и быстро стано¬
вится неразумным и фанатичным. Опыт предыдущих цивилизаций должен
предупреждать нас об усилении их значения для современных народов.
Нам не следует игнорировать возможность того, что даже правильный курс,
принятый с энтузиазмом, может быть сорван еще до того, как начнет прояв¬
лять свои результаты. Эта опасность, возможно, будет сведена к минимуму,
если осознание того, что материнство и отцовство достойных граждан бу¬
дут служить во благо обществу, будет доведено до граждан на всех уровнях
образования, и если такое служение обществу получит адекватное призна¬
ние в экономической системе.Опасность более интеллектуального толка лежит в растущей тенден¬
ции к разрыву между теорией и практикой. Специализация в сфере ин¬
теллектуальных усилий необходима и оправдана ограничениями, наложен¬
ными природой на наши индивидуальные способности. Нельзя оправдать
мыслителя, теоретические исследования которого оторваны от реальной
жизни. Такая оторванность лишает теорию смысла, а практику — даль¬
нейшего развития. Она влечет за собой естественное, но неблагоприят¬
ное следствие того, что волевые и энергичные люди, чьим делом являются
практические достижения, испытывают естественное неуважение к безот¬
ветственным теоретикам, а потому игнорируют практическое применение
теоретических соображений.Бесполезно размышлять о том, что наша цивилизация сможет достичь
такого постоянства, если всеми предшествующими цивилизациями утвер¬
ждалось противное. Но мысль о том, что шансы невелики, не должна
влиять на наши усилия. Команду корабля при угрозе гибели интересует
266Глава 12не столько размер страховой премии, сколько практические опасности на¬
вигации. Игнорируя такие опасности, мы не можем надеяться на их даль¬
нейшее исследование.РезюмеПерераспределение рождений, помимо причины, по которой оно пред¬
лагается, или средств, посредством которых оно может быть достигнуто,
будет сопровождаться экономическими преимуществами.Умеренное социальное поощрение фертильности, которая должна со¬
хранять благоприятный уровень рождаемости, вполне соответствует эко¬
номической организации нашей собственной цивилизации и может обеспе¬
чить способ преодоления современной тенденции сокращения численности
населения.Система семейных пособий, принятая во Франции, определенно
не подходит для сохранения высших уровней интеллектуальных способ¬
ностей. Сейчас нет причин сомневаться в том, что долговременная цивили¬
зация может, несомненно, основываться на более совершенной системе.Структура существующих популяций как по своей социальной значи¬
мости, так и по эффективному бесплодию представляет особые и гораздо
более серьезные трудности, которые могут преодолеть лишь люди, способ¬
ные тщательно сформулировать свой путь развития.
ЛитератураН. W. Bates (1862). “Contributions to the Insect Fauna of the Amazon Valley”. Trans.
Linn. Soc. Land, xxiii, p. 495.W. Bateson (1894). Contributions to the Study of Variation. London.W. Bateson (1909). “Heredity and Variation in Modern Lights”. Darwin and Modern
Science, pp. 85-101. Cambridge: University Press.W. S. Blunt (1903). The Seven Golden. Odes of Pagan Arabia. Chiawick Press.A. de Candolle (1873). Histoire des Sciences et des savants depuis deux siecles, pp. 157—
161. Geneva.P. Cameron (1882). Monograph on the British phytophagous hymenoptem, vol. i.
London: Ray Society.W. E. Castle (1914). “Piebald Hats and Selection; an Experimental test of the
Effectiveness of Selection and of the Theory of Gametio Purity in Mendelian Crosses”.
Carnegie Inst. Pull. No. 195, 56 pp.J. A. Cobb (1913). “Human Fertility”. Eugenics Review, iv, pp. 379-382.C. Darwin (1859). The Origin of Species. London: John Murray. [Имеется русский
перевод: Дарвин Ч. Происхождение видов путем естественного отбора. 2-е изд.
доп. — СПб.: Наука, 2001. Сер. «Классики науки».]С. Darwin (1868). Variation of Animals and Plants under Domestication. (Chapter xxii
and p. 301.) London: John Murray. [Имеется русский перевод: Дарвин Ч. Изменения
домашних животных и культурных растений. Сочинения. Т. 4. М.-Л.: Изд. АН
СССР, 1951.]С. Darwin (1871). The Descent of Man, and Selection in Relation to Sex, p. 399.
London: John Murray. [Имеется русский перевод: Дарвин Ч. Происхождение че¬
ловека и половой отбор. Сочинения. Т. 5. М.: Изд. АН СССР, 1953.]F. Darwin (1887). Life and Letters of Charles Darwin. London: John Murray.F. Darwin (1903). More Letters of Charles Darwin. London: John Murray.F. Darwin (1909). Foundations of the “Origin of Species”, pp. 2, 77-78, 84—86.L. Darwin (1926). The Need for Eugenic Reform. London: John Murray, 529 pp.F. A. Dixey (1908). “On Mullerian Mimicry and Diaposematism”. Trans. Ent. Soc.,
1908, pp. 559-583.
268ЛитератураJ. С. Dunlop (1914). “Fertilityof Marriage in Scotland”. Joum. Roy. Stat. Soc. lxxvii.
259.A. S. Eddington (1927). The Nature of the Physical World.R. A. Fisher (1922). “On the Dominance Ratio”. Proc. Roy. Soc. Edin. xlii, pp. 321-341.R. A. Fisher and E. B. Ford (1928). “The Variability of Species in the Lepidoptera, with
reference to Abundance and Sex”. Trans. Ent. Soc., 1928, pp. 367-384.E. Ford and H. O. Bull (1926). “Abnormal Vertebrae in Herrings”. Jour. Marine Biol.
Ass. xiv, pp. 509-517.Forel (1900). Mitt I. Schweiz. Ent. Gesell, x, pp. 267-287.J. G.Fraser (1909). Psyche’s Task. London: Macmfflan & Co.J. C. F. Fryer (1913). “An Investigation by Pedigree Breeding into the Polymorphism of
Papilio polytes". Phil Trans. B, vol. cciv, pp. 227-54.F. Galton (1869). Hereditary Genius, pp. 123-133. [Имеется русский перевод: Гал¬
тон Ф. Наследственность таланта. М.: Мысль, 1996.]J. Н. Gerould (1923). “Inheritance of White Wing Colour, a Sex-limited (Ses-controlled)
Variation in Yellow Pierid Butterflies”. Genetics, viii, pp. 495-551.E. Gibbon (1776). The Decline and Fall of the Roman Empire, chapter xiv. [Имеет¬
ся русский перевод: Гиббон Э. История упадка и разрушения Римской империи.
СПб.: Наука. Т. 1, 1997.]Le Comte de Gobineau (1854). Essai sur I 'inegalite des races humaines. Paris: Firmin-
Didot et Cie.D. Heron (1914). “Note on Reproductive Selection”. Biometrika, x, pp. 419-420.D. Heron (1906). On the Relation of Fertility in Man to Social Status. London: Dulau
& Co.F. L. Hoffman (1912). “Maternity Statistics of the State of Rhode Island, State Census
of 1905”. Problems in Eugenics. London: Chas. Knight.H. E. Howard (1920). Territory in Bird Life.E. Huntington and L. F. Whitney (1928). The Builders of America. London: Chapman& Hall.Т.Н.Huxley (1854). “On the Educational Value of the Natural History Sciences”.
Collected Essays, vol. iii, p. 46.D. F. Jones (1928). Selective Fertilisation. University of Chicago.G. H. Knibbs (1917). Census (1911) of the Commonwealth of Australia. Melbourne:
McCarron, Bird & Co. 466 pp.D. E. Lancefield (1918). “An Autoaomal Bristle Modifier Affecting a Sex-linked
Character”. Amer. Nat. Iii, pp. 462-464.
Литература269Ray Lankester (1925). “The Blindness of Cave Animals”. Nature, cxvi. 745.L. March (1912). “The Fertility of Marriages according to Profession and Social Class”.
Problems in Eugenics, p. 208. London: Chas. Knight.G. A. K. Marshall (1908). “On Diaposematism, with Reference to some Limitations of
the Miillerian Hypothesis of Mimicry”. Trans. Ent. Soc. (1908), pp. 93-142.G. J. Mendel (1865). “Versuche iiber Pflanzen-Hybriden”. Verh. Naturf. Ver. in Brtinn,
vol. iv. [Имеется русский перевод: Мендель Г. Опыты над растительными гибри¬
дами М.: Наука, 1965. Сер. «Классики науки».]Т. Н. Morgan, С. В. Bridges and А. Н. Sturtevant (1925). “The Genetica of Drosophila”.
Bibliographia Genetica, vol. ii, pp. 1-262.F. Muller (1879). “Ituna and Thyridia: a Remarkable Case of Mimicry in Butterflies”.
Trans. Ent. Soc., 1879, p. xx.K. Pearson (1899). “Mathematical Studies in Evolution; VI Genetic (Reproductive)
Selection”. Phil. Trans. A, vol. cxcii. 257.Plato. The Mepublic, Book V.Pliny. Natural History, Book 29, chapter iv.Polybius. Historiee, Book XXXV1L [Имеется русский перевод: Полибий. Всеобщая
история. Т. 2. Кн. 11-39. М.: ACT, 2004.]Е. В. Poulton (1908). Essays on Evolution. Oxford: Clarendon Press.E. B.Poulton (1924). “Mimicry in the Butterflies of Fiji considered in Relation to the
Euploeine and Danaine Invasions of Polynesia and to the Female Forms of Hypolimnas
bolinah., in the Pacific”. Trans. Ent. Soc. (1924), pp. 564—691.A. O. Powys (1905). “Data for the Problem of Evolution in Man. On Fertility, Duration
of Life and Reproductive Selection”. Biometrika, iv, p. 233.R. C. Punnett (1915). Mimicry in Butterflies. Cambridge: University Press.С. Тате Regan (1925). Organic Evolution. Presidential Address to Section D of the
British Association, 1925.H. H. Risley (1891). Tribes and Castes of Bengal.G. C. Robson (1928). The Species Problem. Edinburgh: Oliver & Boyd.J. Schmidt (1919). “La valeur de l’individu a litre de generateur, appreciee suivant la
methode du croisement diallele”. Comptes Rendus des travaux du laboratoire Carlsberg,
vol. xiv, No. 6, pp. 1-33.Seneca. Of Consolation to Helvia, 16.W. Robertson Smith (1885). Kinship and Marriage in Early Arabia, p. 157.Т. H. C. Stevenson (1920). “Fertility of various classes in England and Wales from the
Middle of the 19th Century to 1911”. Jour. Roy. Stat. Soc., vol. lxxxiii, p. 401.
270ЛитератураН. Е. G. Sutherland and G. Н. Thomson (1926). “The Correlation between Intelligence
and Size of Family”. Brit. Jour, of Psychology, xvii, pp. 81-92.C. F. M. Swynnerton (1915). “A Brief Preliminary Statement of a few of the Results
of Five Years’ Special Testing of the Theories of Mimicry”. Proc. Ent. Soc. (1915),
pp. 21-33.Tacitus (a. d. 98). “A Treatise on the Situation, Manners and Inhabitants of Germany”.
Bohn s Classical Library, vol. ii.G. H. Thomson (1919). “The Criterion of Goodness of Fit of Psychophysical Curves”.
Biometrika, xii, p. 216.H.H. Turner (1924). “On the Numerical Aspect of Reciprocal Mimicry (Diaposematic
Resemblance)”. Trans. Ent. Soc. (1924), pp. 667-675.A. R. Wallace (1889). Darwinism. London: Macmillan. [Имеется русский перевод: Уо-
ллэсъ А. Р. Дарвинизмъ. М.: Изд. М. и С. Собашниковыхъ. 1911.]Е. A. Westermarck (1906). Origin and Development of Moral Ideas. Chapter, XVII.W. M. Wheeler (1923). Social Life among the Insects, p. 225, London: Constable.W. C. D. Whetham (1909). The Family and the Nation, p. 140.O. Winge (1923). “Crossing-over between the X- and Y-chromosome in Lebistes".
Comptes iiendus des iravaux du laboratoire de Carlsberg, vol. xiv, No. 20.Sewall Wright (1925). “The Factors of the Albino Series of Guinea Piga and their
Effects on Black and Yellow Pigmentation”. Genetics, vol. x, pp. 223-260.
Указатель53Abispa (Monerebia) ephippium Fab.,
Иллюстрация I
Abraxas grossulariata, 162
Acraea zetes acara Hew., Иллюстра¬
ция II
Agouti, 125
Anosia plexippus, 162Castnia linus Cramer, Иллюстрация II
Colias, 169, 170“Contributions to the Study of Variation”,
125Crinkled Dwarf, 59
Cynthia cardui, 162Danaida tytia Gray, Иллюстрация II
“Darwinism”, 135
“Descent of Man”, 142
Drosophila, 19, 51, 53, 55, 56, 64, 65,
165, 175Ethesis ferrugineus Macleay, Иллю¬
страция I“Faundations of the Origin of
Species”, 4, 7
Fiji, 155
Forked, 65Gallus banciva, 60
Gammarus chevreuxi, 55Heliconius erato erato Linn., Иллю¬
страция I“Hereditary genius”, 230
Lebistes, 165
Mania typica, 161Methona confuse Butler, Иллюстра¬
ция I“Need for Eugenic Reforms”, 230
Neptis imitans Oberth, Иллюстра¬
ция IIOenothera, 131Onethocampa processionea, 162
“Origin of species”, 98Papilio agestor Gray, Иллюстрация II
Papilio aristolochiae, 167
Papilio hector, 167
Papilio polytes, 164, 166-170
Pericopis phyleis Druce, Иллюстра¬
ция I
Platypoecilus, 165
Pseudacraea boisduvali trimenii
Butler, Иллюстрация II
Pyrameis atalanta, 162Receptaculum seminis, 16Recessive pied, 166Romulus (form of Papilio polytes), 167“Selective Fertilization”, 130
Semi-forked, 65Strongylognathus testaceus, 254, 256
272УказательTetramorium cespitum, 254-256
Tragocerus formosus Pascoe, Иллю¬
страция IVanessa, 162“Variation of Animals and Plants under
Domestication”, 2, 3, 7“Who’s Who”, 216, 237
Wrinkled leaf, 59Zygaera bucephala, 161Австралия (Australia), 30, 192
Адаптация (adaptation), 38
Альбинизм (albinism), 19, 51, 53, 56
Англия (England), 213, 218, 261
Англия и Уэлс (England and Wales),
262Арабы (arabs), 194, 201Аргилл, герцог (Argyll, Duke of), 134Аристотель (Aristotle), 204Бабочки (butterflies), 54, 148, 161,
163, 183
Бенгальская ярмарка невест
(Bengalese marriages), 250
Биометрические методы (biometrical
methods), 106
Биометрия (biometry), 18
Блант Уилфред (Blunt Wilfrid), 245
Бобы (bean), 18Борьба за существование (struggle
for existence), 43
Брачный возраст (age at marriage),
191, 192
Бриджес (Bridges), 53
Буддизм (Buddhism), 194
Булл (Bull), 116Бэйтс (Bates), 148, 149, 150, 158, 169
Бэкон (Bacon), 171Бэтсон (Bateson) ix, x, 1, 54, 113, 125Вайс (Weiss), 55
Валентиниан (Valentinian), 204
Варвары (barbarians), 243, 244
Вейсман (Weismann), 13-16, 66
Великобритания (Great Britain), 216,
261Вестермарк (Westermarck), 194
Водные организмы (aquatic
organisms), 129
Возврат (reversion), 7, 9
Возраст смерти (age at death), 23
Возрастное распределение (age
distribution), 26
Волнистые попугайчики (budgerigars),
60Вороны (crows), 134
Время релаксации (time of
relaxation), 87Галлия (Gaul), 242
Галтон (Gallon), 171, 230-232, 234,
236, 255
Генотипическое и генетическое
значение (genotypic and genetic
values), 34, 114
Географическое распространение,
ареал (geographical range), 99,
127, 128Геометрическое представление ада¬
птации (geometrical representation
of adaptation), 39, 40
Германия (Germany), 262
Германские народы (Germans), 244
Героизм (heroism), 163, 247
Герон (Heron, D.), 192, 214, 215, 221
Гесиод (Hesiod), 229
Гетерозиготы (heterozygotes), 104
Гиббон (Gibbon), 205
Указатель273Глаз (eye), 19, 39
Глаза, утрата (eye, loss of), 130
Глухонемота (deaf-mutism), 19
Гнездо (nest), 134Гобино, де (Gobineau, Comte de),
239,240Гомера поэмы (Homeric poems), 244
Гомогамия (homogamy), 100, 131
Гормоны (hormones), 133, 141
Гребень (comb), 60
Греки (Greeks), 179
Греция (Greece), 194, 203
Грызуны (rodent), 53Данаиды (danaine butterflies), 162
Данлоп (Dunlop, Dr. J. C.), 217, 219
Даоизм (taoism), 194
Дарвин (Darwin), vii, ix, 1, 3, 5-7,10-
12, 20, 22, 44, 66, 98, 99, 132, 140,
142, 171, 179
Дарвин, майор Леонард (Darwin,
major L.), v, 187, 230
Де Фриз (de Vries), viii
Декандоль (De Candolle), 7, 232, 255
Денежные пособия семье (family
allowances), 260-262
Деструктивный отбор (destructive
selection), 40, 253
Детоубийство (infanticide), 194, 201
Джерулд (Gerould), 168
Джонс (Jones, D. F.), 130
Дикси (Dixey), 154
Дисперсия (variance), 4, 9-12, 18, 31,
99, 100Дисперсия генетическая (variance,
genetic), 33
Диффузия (diffusion), 128, 212
Дневной павлиний глаз (peacock
butterfly), 162Доминантность (dominance), x, 9,
глава 3, 48, 105, 107
Доминантный белый (dominant
white), 60, 168Зародышевый отбор (germinal
selection), 17
Затраты родителей (parentalexpenditure, burden of), 143, 186Изменчивость (variability), 98, 99
Иисус (Jesus), 205
Иммс (Imms, A. D.), 161
Иммунитет (immunity), 206
Индия (India), 223
Иоганнсен (Johannsen), 18
Исландские саги (Icelandic sagas),
244, 249
Иудеи (Jews), 194, 208Йельский [университет] (Yale), 237Камбала (flat fish), 117
Камерон (Cameron), 162
Канарейки, желтовато-коричневые
(canaries, cinnamon), 60
Кастл (Castle), 166
Католики (Catholics), 208
Китай (China), 194, 223
Китайский (Chinese), 208
Клавдий (Claudius), 243
Клещи (mites), 157
Климат, изменения (climate, changes
of), 42Кобб (Cobb, J. A.), 232-234, 255
Константин (Constantine), 205
Коран (Koran), 194, 203
Корпускулярная теория наслед¬
ственности (particulate theory of
inheritance), ix, 1, 2, 8-13, 17, 20
274УказательКорреляция (correlation), 4, 18, 33,
200Корренс (Correns), viii
Крапивница, бабочка (tortoise-shell
butterfly), 162
Креветка (shrimp), 55
Кровная месть (blood feud), 245
Кроссинговер (crossing over), 104
Крысы (rats), 166
Кукуруза (maize), 131
Кукушки (cuckoos), 187, 225
Курайшиты (Koraish), 202Ламарк (Lamarck), 12
Ленивцы (sloths), 114, 117
Летальные мутации (lethal mutations),
19Личинки пилильщика (saw-fly
larvae), 162
Личинки, живущие скоплениями
(gregarious larvae), 160
Лондон (London), 214, 215, 221
Льюис (Spedan Lewis), 62
Лэнкестер, Рей (Lankester, Ray), 130
Лэнсфилд (Lancefield), 65Мальтузианский параметр (Malthusian
parameter), 25, 44
Мальтус (Malthus), 27, 44
Ману, законы (Manu, laws of). 194
Маршалл (Marshall, G.A. К.), 151,
153, 154, 163, 164, 170
Мекка (Mecca), 203
Мендель (Mendel), viii-x, 8, 9
Меристические признаки (meristic
characters), 113, 117
Метрические признаки (metrical
characters), 106, 115, 117
Миграция (migration), 128Мимикрия (mimicry), глава 7, 147,
208Млекопитающие (mammals), 56, 114
Многоженство (polygamy), 251
Морган (Morgan), 53
Морская свинка (cavy, guinea pig), 53
Мулаты (mulattoes), 18
Муравьи (ants), 16, 157, 254
Мутаций частота (mutation rates), 13,
19-21, 52, 77, 108, 109, 123
Мутация (mutation), 12, 17-19, 49,
105Мухаммед (Muhammad), 203
Мышь (mouse), 125
Мюллер (Muller), 148, 150, 170Наследницы (heiresses), 230
Настурция (nasturtium), 55
Непрерывность (discontinuity), 113,
125Неприятный вкус (distastefulness),
159, 248Новая Зеландия (New Zealand), 244
Новый Южный Уэлс (New South
Wales), 190
Нормальное распределение (normal
distribution), 18
Ночные бабочки (moths, night-
flying), 98, 162Обилие [вида] (abundance), 98
Обучение птиц (education of birds),
150Общественные насекомые (social
insects), 181
Окраска крыльев (wing-colour), 98
Отбор внутри общества (intra-
communal selection), 184Паразитизм (parasitism), 254
Указатель275Паультон (Poulton), 147, 150, 155—
158, 160, 161
Перекрестное оплодотворение
(cross-fertilization), 102, 120
Пещеры (caverns), 130
Пённет (Punnett), 163-165, 170
Пигментация (pigmentation), 17, 53,
166Пирсон (Pearson), 196
Платон (Plato), 204
Плиний (Pliny), 194, 204
Плодовитость (fecundity), 44
Повюс (Powys), 190
Подвижные животные (motile
animals), 120
Позвонки (vertebrae), 114-117
Пол (sex), 165
Полибий (Polybius), 204
Полиморфизм (polymorphism), 165,
167, 182, 183
Полинезийцы (Polynesians), 244
Половое предпочтение (sexual
preference), 100, 130
Половое размножение (sexualreproduction), ix, 120, глава 6, 122
Половой отбор (sexual selection), 66,
122, 132, 248, 249
Популяция, население (population),
42, 242Превосходство гетерозиготы(heterozygote, superiority of), 101
Предок (ancestry), 126
Предупреждение зачатия (contracep¬
tion), 194
Проституция (prostitution), 194
Пространственное представление
адаптации (dimensional
representation of adaptation), 39,40
Протестанты (protestants), 208Психофизиологические эксперимен¬
ты (psychophysical experiments),15Птица домашняя (poultry), 60, 61
Птицы (birds), 136, 146, 152, 160
Пыльца (pollen), 131
Пэров сословие (peerage), 196, 230Раджпуты (Rajputs), 194
Разновидности (varieties), 128
Райт С. (Wright, S.), 53, 87
Распределение Пуассона (ряд)
(Poisson series), 74, 119
Распространение (dispersal), 129
Распространенность, ареал (range),
99, 128, 132
Расщепление вида (fission of
species), 100, 126, 132
Регрессия (regression), 118
Рейган, Тэйт (Ragan Tate), 114
Рекомбинация, комбинаторика
(recombination), 104, 240, 241
Репродуктивная ценность(reproduction value), 27-29, 72
Репродукция (человек) (reproduction
(man)), главы 9, 10
Рим (Rome), 194, 203
Римская Британия (Roman Britain),
242Римская империя (Roman Empire),
178Риели (Risley), 250
Рисунок Dutch (Dutch pattern), 166
Рисунок капюшона (hooded patten),
166Робертсон Смит (Robertson Smith),48Робсон (Robson), 54, 147
Род-Айленд (Rhode Island), 207
Рост (stature), 18, 100, 106, 109
276УказательРыбы (fishes), 114, 143Сальтации (saltations), 163
Самоа (Samoa), 244
Самооплодотворение (self-fertili¬
zation), 102, 103, 114, 120
Сатерлэнд (Sutherland), 238
Свиннертон (Swynnerton), 150
Священное Писание (Hebrew
scriptures), 194
Селективная ценность (selection
value), 15Селективное преимущество (selective
advantage), 20
Сельд (herring), 116
Сельское хозяйство (agriculture), 178
Сенека (Seneca), 204
Скандинавия (Scandinavia), 262
Слабоумие (feebleness of mind), 19
Слитная теория наследственности
(blending theory of inheritance), 1,
5, 7, 10, 11, 17, 18, 20
Слияние, теория наследственности
(fusion, theory of inheritance), 1
Смертность (death-rate), 23, 26
Собаки (dogs), 118
Совесть (conscience), 174, 200
Соленость (salinity), 129
Соотношение полов (sex-ratio), 142
Софокл (Sophocles), 69, 258
Спенсер (Spencer, Herbert), 181, 213,
232Специфический модификатор
(specific modifier), 54
Среда обитания, ухудшение(environment, deterioration of), 41
Стабильность отношения генных ча¬
стот (stability of gene ratio), 98,
105, 120, 167
Стёртевант (Sturtevant), 53Стивенсон (Stevenson, Dr. Т. H. C.),
217,219, 220
Столбик (style), 130
Судьи (judges), 231
Сцепление (linkage), x, 9, 105, 111,
120США (U.S. A.), 221,237Таблица выживания (life table), 22
Таблица репродукции (reproduction,
table of), 24
Татарские народы (Tartars), 244
Тацит (Tacitus), 244
Термиты (termites), 182
Термодинамика, второй закон
(thermo-dynamics, second law of),37Тернер (Turner, H. H.), 155
Территория (territory), 136, 160
Тертуллиан (Tertullian), 204
Томсон (Thomson, Godfrey H.), 238
Турецкие народы (Turks), 244Угорь (eel), 116
Уиллер (Wheeler), 157, 183
Уитни и Хантингтон (Whitney and
Huntington), 221, 237
Уитхем (Whetham, W. C. Dampier),
216Улучшение, шанс на (improvement,
chance of), 39
Умерщвление плода (feticide), 194,
203Умственные способности и мораль¬
ные качества (mental and moral
qualities), 192
Уоллес (Wallace), vii, 44, 133-135,
142, 173, 179
Урбан (Urban), 16
Указатель277Фертильность (fertility), 186, 189,
190, 195Фертильность женщины в Новом
Южном Уэлсе (fertility of woman
in N. S. W.), 190
Фивы (Thebes), 194
Фиксация (fixation), 129
Форд E. (Ford, E.), 116
Форд Е.Б. (Ford, E. B.), 55, 98
Форел (Forel), 254-256
Фраер (Fryer), 167, 168
Франция (France), 212, 260-262
Фрезер (Fraser), 122
Фундаментальная теорема естествен¬
ного отбора (Fundamental
Theorem of National Selection),
глава 2, 22, 35Хаксли (Huxley), 2, 7, 22, 171
Хантингтон (Hundington), 221, 237
Харланд (Harland, С. S.), 59
Хатчинсон (Hutchinson, J. B.), 59
Хельвия (Helvia), 204
Хлопчатник (cotton), 59
Хлопчатник Sea Island (Sea Island
cotton), 59
Хлопчатник Upland (Upland cotton),
59Хлопчатник перуанский (Peruvian
cotton), 59
Ховард (Howard), 136Хоффман (Hoffman), 207
Хохолок (crest), 60
Хризомелиды (chrysomelid beetles),
162Христианство (Christianity), 205
Хромосомы (chromosomes), 9, 51,
105Хукер (Hooker), 98Человек (man), x, 12, 19, 130, 163,
главы 8-12, 171, 187, 189
Чермак (Tschermak), viii
Чистые линии (pure lines), 18, 20,
114Шанс улучшения (chance ofimprovement), 39
Шекспир (Shakespeare), 189,211, 257
Шея (neck), 114, 117
Шмидт (Schmidt), 114
Шотландия (Scotland), 213, 217Эддингтон (Eddington) viii, 37
Экономическая система (economic
system), 184
Эпистатический (epistatic), 54
Эффект генного замещения (effect of
genes substitution), 31Юлий Павел (Julius Paulus), 205Ястребы (hawks), 134
КомментарииАвторы комментариев Я. В. Глотов (Н. Г.), Л. А. Животовский (Л. Ж.),Я. В. Ившин (Я. Я.)Первое издание Fisher R. A. The genetical theory of natural selection.
Oxford: Clarendon Press, 1930, оказало огромное влияние на развитие попу¬
ляционной и эволюционной генетики и стало одной из основ синтетической
теории эволюции (СТЭ).Спустя 28 лет было опубликовано второе издание — Fisher R. A. The
genetical theory of natural selection. Second revised edition. New York'. Dover
Publ., 1958. Оно включало исправление опечаток, многочисленные замены
отдельных слов (около 100), уточняющие, по мнению Фишера, смысл от¬
дельных фраз и предложений, и несколько обширных дополнений (биомет¬
рическая модель генетической детерминации количественных признаков,
раздел The benefit of the species в главе 2, раздел Self-sterility Allelomorphs
в главе 4) — всего около 20 страниц.Спустя 60 лет Беннет опубликовал Полное издание с комментариями —
Fisher R.A. The genetical theory of natural selection. A Complete Variorum
Edition. Edited with a foreword and notes by J. H. Bennett. Oxford: Univ. Press,
1990: факсимильное первое издание, в сносках — мелкие исправления, до¬
полнения, вставки второго издания, в приложении 1 - более крупные ис¬
правления и дополнения, в приложении 2 — примечания Беннета, в прило¬
жении 3 — список публикаций Фишера по генетической теории естествен¬
ного отбора.Настоящее издание представляет перевод первого, оригинального из¬
дания 1930 года. Качество перевода было существенно улучшено благодаря
работе, проведенной С. J1. Яковлевой, М. В. Фроловой, А. П. Абросимовым,
О. Н. Глотовой, Г. В. Быковой. Очень полезными были замечания JI. А. Жи-
вотовского, В. Г. Митрофанова, Н. В. Ившина, Г. Ю. Софронова, А. И. Ермо¬
лаева. Вывод формул, внимательная выверка всего текста и внесение мно¬
гочисленных исправлений выполнены А. Б. Трубяновым. Большая работа
была проведена корректором О. А. Шемякиной. Исключительно конструк¬
тивным на всех этапах было взаимодействие с редакторами издательства
Комментарии279Ю. П. Сухоплюевой и Г. Ф. Муллахметовой. Редактор искренне благодарен
всем коллегам, способствовавшим совершенствованию русского перевода,
хотя за конечный результат, естественно, несет ответственность он один.Необходимо сделать два общих замечания о переводе терминов, про¬
ходящих через всю книгу.Для обозначения дискретных элементарных единиц наследственности
на смену неопределенным менделевским терминам задаток, фактор, эле¬
мент с развитием генетики пришел термин ген, а разные формы одного
гена стали называть аллелями (аллеломорфами). У Фишера элементарная
наследственная единица, как правило, фактор. Ген Фишера это, как пра¬
вило,— аллель (gene of a different kind; single pairs of genes, that is single
factors; two alternative genes of any factor; pair of alternative genes; alternative
gene; two different genes or allelomorphs).Фишер использует термин population и в смысле популяция растений
и животных, и народ, и население, и нация, когда речь идет о человеке.
(.Н. Г.)ПОСВЯЩЕНИЕ1 с. v. Майор Леонард Дарвин (1850-1943) — сын Ч. Дарвина, начавший
карьеру военным инженером, президент Королевского географического об¬
щества (1908-1911), председатель Британского евгенического общества
(1911-1928) (http://en.wikipedia.org/wiki/Leonard Darwin).
«Фишер и Леонард Дарвин — которому посвящена Генетическая теория,—
поддерживали очень тесные связи; с 1915 года они часто встречались
и переписывались в течение 20 лет, обмениваясь своими представления¬
ми о естественном отборе, наследственности, евгенике и многом другом.
В течение ряда лет Леонард Дарвин пытался побудить Фишера “написать
большую работу о математике эволюции”. Он неизменно оказывал Фишеру
всевозможную поддержку и вдохновлял его на эту великую попытку. Их
оживленная переписка, которая сейчас частично опубликована, создала хо¬
рошую основу для осуществления Фишером этого проекта» (Bennett J. Н.
Foreword to Complete Variorum Edition, vii). (H. Г.)ПРЕДИСЛОВИЕ2 с. vii. «Natural Selection» и «natural selection», «Evolution» и «evolution»,
«Man» и «man» и т. п. - Фишер пишет то с заглавной (подчеркивая, вероят¬
но, значимость явления), то со строчной буквы. Эта особенность сохранена
при переводе. (Н. Г.)
280Комментарии3 с. viii. Весьма любопытно, что Фишер называет Менделя «математи¬
ком». Вероятно, он так назвал его по стилю мышления и анализу экспери¬
ментальных данных, тогда еще совсем необычных в биологии. (Л.Ж.)4 с. viii. Работа Менделя была достаточно известна — см. Гайсино-
вич А. Е. Зарождение и развитие генетики. М.: Наука, 1988. (Н. Г.)5 с. viii. Имеются в виду работы Ф. Галтона и К. Пирсона — см. Гайси-
нович А. Е„ комментарий1. (II. Г.)6 с.х. Оценку огромного вклада В. Бэтсона в развитие генетики
см. Гайсинович А. Е., комментарий^. (Н. Г.)7 с. х. Современная теория эволюции и популяционной генетики
немыслима без математических моделей и методов статистического анали¬
за, огромный вклад в развитие которых внес Роналд Фишер. Современная
биоинформатика и анализ геномных данных также зиждутся на развитом
математическом аппарате. (Л. Ж.)ГЛАВА 18 с. 1. Описание содержания главы у Фишера не всегда дословно сов¬
падает с названием разделов в тексте главы. (Н. Г.)9 с. 1. Названия книг и других изданий не переводятся, поскольку у Фи¬
шера они имеют смысл ссылок на литературный источник. (Н. Г.)9а с. 2. Опубликовано на русском языке: Ч. Дарвин Очерк 1842 года /
Дарвин Ч. Происхождение видов путем естественного отбора. Сочинение.
Т. 3. М.-Л.: Изд. АН СССР, 1939. С. 79-112. - Ч. Дарвин Очерк 1844 года /
Там же. С. 113-230. (Н.Г.)10 с. 4. Для ситуации, описываемой Фишером, в настоящее время упо¬
требляется термин ассортативное скрещивание (положительное или отри¬
цательное). В данном случае Фишер имеет в виду положительное ассорта¬
тивное скрещивание. (II. Г.)11 с. 4. Отметим, что в рамках теории слитной наследственности вы¬
вод о том, что дисперсия признака в следующем поколении уменьшается
вдвое, предполагает что величины признака у ВСЕХ потомков данной пары
одинаковы и равны 1 /2(х + у). Напротив, в корпускулярной теории наслед¬
ственности величины признака у потомков различны вследствие сегрегации
аллелей, и в среднем по всем возможным парам родителей их дисперсия
в точности равна «недостающей» половине дисперсии в родительском по¬
колении, так что ожидаемая дисперсия признака у потомков оказывается
в точности равной дисперсии признака в родительском поколении. (Л. Ж.)12 с. 5. Напротив, как следствие корпускулярной теории наследствен¬
ности, почти вся существующая изменчивость имеет древнее происхожде¬
Комментарии281ние. (Л. Ж.) Это заключение, далее подробно развиваемое Фишером, имеет
принципиальное значение, хотя обычно не обсуждается при обосновании
доказательства корпускулярной организации (дискретности) наследствен¬
ного материала. (Я. Г.)13 с. 9. Идею о возможности чисто логического вывода результатов кор¬
пускулярной (дискретной) наследственности неоднократно излагал в своих
лекциях Н. В. Тимофеев-Ресовский. Он не исключал, что Мендель знал, что
получит, приступая к своим экспериментам. В подтверждение этой мыс¬
ли укажем, что анализ Фишера экспериментальных данных Менделя пока¬
зал, что отклонения в расщеплениях у Менделя оказались статистически
«слишком» близкими к теоретически предсказываемым, см. Вейр Б. Анализ
генетических данных. М.: Мир, 1995. С. 28-39. (Н. Г.)14 с. 9. Работа Менделя была доложена на заседании Брюнненского об¬
щества естествоиспытателей в 1865 году, опубликована в трудах общества
в 1866 году — см. Гайсинович А. Е. Грегор Мендель (биографический очерк) /
Грегор Мендель. Опыты над растительными гибридами. М.: Наука, 1965.
С. 127. (Я. Г.)15 с. 10. Неизменность частот генов (в современной терминологии —
аллелей) устанавливается теоремой Харди-Вайнберга, а вышеуказанные
соотношения частот трех генотипов называют соотношениями Харди-
Вайнберга. (Л.Ж.)16 с. 12. Современные оценки и дискуссии о механизме эволюции
по Ламарку, см.: Бляхер Л. Я. Проблемы наследования приобретенных при¬
знаков: История априорных и эмпирических попыток ее решения. М.: Нау¬
ка, 1971; Серавин Л. Н. Похвальное слово Жану Батисту Ламарку //Вестн.
С.-Петербурге, ун-та, 1994. Сер. 3. Вып. 4 (№> 24). С. 3-16; Стил Э.,
Линдли Р., Бландэн Р. Что если Ламарк прав? Иммуногенетика и эволю¬
ция. М.: Мир, 2002; Zhivotovsky L.A. A model of the early evolution of soma-
to-germline feedback. Journal of Theoretical Biology. 2002. V. 216: 51-57; Ula-
талкин А. И. «Философия зоологии» Жана Батиста Ламарка: взгляд из XXI
века. М.: Товарищество науч. изд. КМК, 2009. (Я. Г.)17 с. 14. Этот гипотетический принцип Вейсмана перекликается с прин¬
ципом Ламарка о наследовании изменений, вызванных «внутренним устрем¬
лением организма», против которого так выступал сам Вейсман. (Л. Ж.)18 с. 18. См. Иоганнсен В. О наследовании в популяциях и чистых ли¬
ниях. М.: Сельхозгиз, 1935. (Н.Г.)19 с. 18. О современных представлениях об организации наследствен¬
ного материала, см.: Жимулев И. Ф. Общая и молекулярная генетика. Ново¬
сибирск: Сибирское университет, изд., 2007; Иванов В. И. и др. Генетика.
М.: ИКЦ «Академкнига», 2006; Инге-Вечтомов С. Г. Генетика с основами
282Комментарииселекции. СПб.: изд. Н-Л, 2010; Эпигенетика. Под ред. С. Д. Эллиса, Т.Дже-
нювейна, Д. Рейнберга. М.: Техносфера, 2010. (Н.Г’.)ГЛАВА 220 с. 22. Методика демографического анализа популяций описана Пи-
анка Э. Эволюционная экология. М.: Мир, 1981. (Н.Г.)21 с. 28. Такие «непрямые эффекты» много лет спустя также станут
объектом изучения, например, в теории эволюции альтруизма и коммуни¬
кации. (Л. Ж.)22 с. 31. Биометрические модели генетической детерминации количе¬
ственного признака см. Фолконер Д. С. Введение в генетику количествен¬
ных признаков. М.: Агрогпромиздат, 1985, а также Животовский Л. А., Гло¬
тов Н.В. Популяционно-генетическая интерпретация статистического
анализа количественных признаков // Теория отбора в популяциях расте¬
ний. Новосибирск: Наука. Сибирск. отд., 1976. (Н. Г.)23 с. 33. Фишер, вероятно, намеренно не использует формальный ал¬
гебраический аппарат, приводящий к этой формуле более коротким путем,
предпочитая точную словесную аргументацию, которую он счел более при¬
емлемой. (Л. Ж.)24 с. 35. «.. .любого признака.так у Фишера во всех трех издани¬
ях (of any organism). Очевидно, что под организмом Фишер понимает здесь
популяцию или вид в целом. (Л. Ж.)25 с. 38. Отклонения от фундаментальной теоремы могут быть вы¬
званы сцеплением и рекомбинацией генов, неадцитивными неаллельны¬
ми взаимодействиями (Ратнер В. А. Математическая популяционная ге¬
нетика. Новосибирск: Наука. Сибирск. отд., 1977), однако количественно
они невелики (Животовский Л. А. О теореме Фишера // Генетика. 1981.
Т. 17. № 2. С. 324-331). Подробнее о фундаментальной теореме см. Свире-
жев Ю.М., Пасеков В. П. Основы математической генетики. М.: Наука,
1982. (Л. Ж.)26 с.43. В формуле во втором издании исправлена опечатка. (Н.Г.)27 с. 43. В формуле во втором издании исправлена опечатка. (Н. Г.)ГЛАВА 328 с. 51. Исправлено во втором издании (1958), в первом (1930) было —
24 пары хромосом. (II. Г.)29 с. 52. Представление о важной эволюционной роли полудоми-
нантных мутаций с варьирующим проявлением и выражением развито
Комментарии283С. М. Гершензоном в работах «Мобилизационный резерв» внутривидовой из¬
менчивости // Журн. общ. биол. 1941. Т. 2., № 1. С. 84-107; Исследование
мутабилъности у наездника Mormoniella vitripennis Wlk. // Генетика. 1965.
Т. 1, №2. С. 95-101. (Н.Г.)30 с. 54. Последние Ъ, а — исправлено во втором издании (Н. Г.)31 с. 57. В настоящее время в популяционно-генетических работах об¬
щепринято обозначать через р частоту аллеля дикого типа А и через q =
— 1 — р — частоту мутантного аллеля а, тогда соотношения частот гено¬
типов АА, Аа и аа в равновесной популяции (соотношения Харди-Вайн-
берга) имеют вид р2 : 2pq : q2. Фишер обозначает через р относительную
частоту мутантного аллеля а (т. е. р Фишера соответствует q/p в современ¬
ной популяционной генетике). Автор изменяет последовательность геноти¬
пов в соотношении Харди - Вайнберга — аа, Аа, А А. Отсюда и появляется
непривычное 1:2р \ р2. Однако в главе 5 Фишер пишет р2 : 2pq : q2. (Н. Г.)32 с. 60. Систематика рода Gossipium сложна и крайне неоднозначна —
Grandtner М. М. Elsevier’s Dictionary of Trees. V. 1. Elsevier, Amsterdam e.a.
2005. P. 385-387 (http://books.google.ru). В данном контексте важно, что Фи¬
шер рассматривает формы Sea Island, Upland и Peruvian в качестве разных
видов. Во втором издании Фишер приводит и новые данные о развитии
доминантности (pp. 63-67). (Н. Г.)33 с. 62. Во втором издании (pp. 68-69) Фишер сообщает о результатах
этого эксперимента, подтверждающего его гипотезу. (Н. Г.)34 с. 64. Широкое распространение рецессивных летальных мутаций
в природных популяциях дрозофилы см. в работе Кайданов Л. 3. Генетика
популяций. М.: Высшая школа, 1996. (Н.Г.)35 с. 68. Современные представления об эволюции доминантности см.
Шеппард Ф. М. Естественный отбор и наследственность. М.: Просвеще¬
ние, 1970', Митрофанов В. Г. Доминантность и рецессивность. М.: Наука,
1994. Высокая скорость отбора генов-модификаторов показана на компью¬
терной модели — Аренс X., Беллман К. Кибернетические методы в количе¬
ственной генетике //Генетика. 1969. Т. 5. № 10. С. 154-168. (Н. Г.)ГЛАВА 436 с. 69. Начиная с 30-х годов XX века стало накапливаться все боль¬
ше данных, указывающих, что правилом, скорее, является наличие в при¬
родных популяциях серий множественных аллелей. Очевидным это ста¬
ло с появлением методов электрофореза белков и ДНК, см.: Левонтин Р.
Генетические основы эволюции. М.: Мир, 1978\ Вейр Б. Анализ генети¬
ческих данных. М.: Мир, 1995; Крутовский К. В. От популяционной гене¬
284Комментариитики к популяционной геномике лесных древесных видов: интегрирован¬
ный популяционно-геномный подход //Генетика. 2006, Т. 42. №10. С. 1-15.
СНГ.)37 с. 73. Это — так называемая производящая функция, широко исполь¬
зуемая для исследования изменений популяционных параметров в чреде
поколений. (,Л.Ж.)38 с. 96. См. гипотезу в Докинз Р. Эгоистический ген. М.: Мир, 1993.
(НГ.)39 с. 97. Исследованием стохастических изменений частот генов Фи¬
шер заложил математический фундамент, на котором М. Кимура построил
теорию нейтральности молекулярной эволюции, рассмотрев стохастику ча¬
стот генов в популяциях ограниченной численности, — Кимура М. Молеку¬
лярная эволюция: теория нейтральности. М.: Мир, 1985. (Л.Ж.)ГЛАВА 540 с. 100. Сейчас это называют ассортативным скрещиванием. (Л.Ж.)ГЛАВА 641 с. 123. По-видимому, не существует «абсолютно», 100%-но агамных
форм. Обмен генетической информацией между особями одного вида (или
микровидов), пусть редко, но имеет место. В качестве примера сошлемся
на два достаточно разработанных случая — у протестов (Серавин Л. Н., Гуд¬
ков А. В. Возможные формы агамных генетических взаимодействий у про-
тистов и пути становления полового процесса // Цитология. 1984. Т. 26.
№ 11. С. 1224-1236) и у растений (Глазунова К. П. О возможности примене¬
ния теории агамно-полового комплекса к систематике покрытосеменных
растений (на примере рода Alchemilla L.) // Бюлл. МОИП. Отд. биол. 1977.
Т. 82. № 5. С. 129-139). (Н. Г.)42 с. 124. См. Мэйнард-Смит Дж. Эволюция полового размножения.
М.: Мир, 1981. (Н.Г.)ГЛАВА 743 с. 150. О бейтсовской и мюллеровской мимикрии см. Шеппард Ф. М.
Естественный отбор и наследственность. М.: Просвещение, 1970. (Н.Г.).44 с. 165. О генетических механизмах определения пола см. Асла¬
нян М. М., Солдатова О. П. Генетика и происхождение пола. М.: Авторская
академия, 2010. (Н. Г.)
Комментарии28545 с. 167. О распространении и механизмах внутрипопуляционного
генетического полиморфизма см. Ford Е.В. Ecological genetics. London:
Methuen, 1964; Тимофеев-Ресовский H. В., Свирежев Ю. М. О генетическом
полиморфизме в популяциях. Экспериментально-теоретическое исследова¬
ние //Генетика. 1967. № 10. С. 152-166; Сергиевский С. О. Полиморфизм
как универсальная адаптивная стратегия популяций / Труды зоологич. ин-
та АН СССР. 1987. Т. 160. С. 41-58. (Я. Г.)46 с. 167. О стабилизирующем отборе см. Шмальгаузен И. И. Факторы
эволюции. Теория стабилизирующего отбора. М.: Наука. 1968; Животов¬
ский J1.A. Интеграция полигенных систем в популяциях М.: Наука, 1984.
(Н.Г.)47 Илл. I. Либо Фишер не знал, как летают дровосеки, и написал свои
догадки, а не факты, увидев жука в коллекции, либо этот вид дровосеков
действительно приобрел признак уровня семейства, т. е. механизм поле¬
та без поднятия надкрыльев, характерный для жуков-златок и бронзовок.
Среди дровосеков миметическое подражание жалящим перепончатокры¬
лым — осам и наездникам — распространено чрезвычайно широко. Толь¬
ко среди палеарктических видов — это целые рода, такие как Plagionotus,
Clytus, Certoclytus, Chlorophorus, Necydalis (у них редуцированы надкры¬
лья, как у фишеровского примера Esthesis ferrugineus, и сходство с на¬
ездниками достигается окраской рудиментов, открытым положением кры¬
льев, формой и цветом брюшка и ног), некоторые представители подсе¬
мейства Lepturinae и некоторые виды Saperda. Ни у одного из этих видов
нет механизма полета, описанного Фишером. Типичное положение крыльев
у жуков-дровосеков при полете — на фото ниже. (Н. И.).
286КомментарииГЛАВА 848 с. 171. Как правило, цитируя Генетическую теорию Фишера, имеют
в виду главы 1-7. Главы 8-12 посвящены общему приложению концепции
приспособленности к Человеку (исторические, экономические, социальные
и т. п. аспекты), вопросам, активно обсуждавшимся с Леонардом Дарвином.
Это — отклик Фишера на идеи позитивной евгеники. Заметим, что Фишер
широко, без уточнений использует термин класс (class), для него это —
место группы индивидов в социальной стратификации общества (слой —
stratum) в разном контексте — по имущественному положению, образова¬
нию, продвижению по социальной лестнице и т. п. О развитии идей евге¬
ники см. Глэд Дж. Будущая эволюция человека. Евгеника XXI века. М.: За¬
харов, 2005; Бабков В. В. Заря генетики человека. Русское евгеническое дви¬
жение и начало медицинской генетики. М.: Прогресс-Традиция, 2008. (Н. Г.)49 с. 176. Широкое научно-популярное обсуждение этой проблемы —
Даймонд Дж. Ружья, микробы и сталь. История человеческих сообществ.
М.: ACT, 2009 (есть и другие издания этой книги). (Н. Г.)50 с. 179. См. классические обобщения Тойнби А. Постижение исто¬
рии М.: Прогресс, 1990; Гумилев JI.H. Этногенез и биосфера Земли. М.:
ACT, 2005 (есть и другие издания этой книги). (Н. Г.)ГЛАВА 951 с. 190. За счет чистой случайности (в рамках пуассоновской модели)
дисперсия фертильности была бы равна средней величине, т. е. 6,19 в дан¬
ном случае. (Л.Ж.)52 с. 192. Для количественной оценки потенциальной эффективности
естественного отбора у человека Джеймс Кроу предложил индекс, учиты¬
вающий дифференциальное воспроизводство и дифференциальную смерт¬
ность (Crow J. Some possibilities for measuring selection intensities in man //
Hum. Biol. 1958. V. 30. P. 1-13). (Л.Ж.)УКАЗАТЕЛЬ53 с. 271. Указатель, составленный Фишером, довольно своеобразен.
Он включает указатели имен, латинских названий видов и предметный ука¬
затель. В последнем случае Фишер называет лишь некоторые страницы
(не все), на которых встречается это слово. Более того, иногда названно¬
го слова на данной странице нет, но здесь обсуждается тема, для которой
названное слово является ключевым. (Н. Г.)
ПослесловиеН. В. Глотов, Л. А. ЖивотовскийSir Ronald Fisher in the Smoky Mountains, 1946.Из Appreciations of R. A. Fisher. Biometrics, 1962, v. 18, № 4*Роналд Эйлмер Фишер (Ronald Aylmer Fisher, 1890-1962) — один
из крупнейших биологов и математиков первой половины XX века [1]. Вер¬
нее,— единственный, поскольку трудно решить, в какую область знания,*«Сэр Роналд Фишер в Больших дымных горах». Здесь — игра слов: и табачный дым
на фотографии, и название гор и заповедника в США — Большие дымные горы. Русская шутка
была бы «Сэр Роналд Фишер на Курилах». (Примечание С. JI. Яковлевой).
288ПослесловиеМатематику или Биологию, он внес больший вклад. Математик по обра¬
зованию, Фишер стал одним из основоположников современной матема¬
тической статистики, а интерес к фундаментальным проблемам биологии
позволил ему получить важнейшие результаты в области популяционной
генетики и теории эволюции. Его книга «Генетическая теория естественно¬
го отбора» стала фундаментом популяционной и эволюционной генетики,
не потеряв своей актуальности сегодня, спустя 80 лет. Достаточно сказать,
что за последние 20 лет (1990-2010) число ссылок на нее составило около11 тысяч (http: //scholar. google . com)!В развитии дарвиновских представлений о происхождении видов под¬
линный прорыв произошел после становления генетики как самостоятель¬
ной области знания. Важнейшими моментами были разработка представле¬
ний об организации и реализации генетического материала (дискретность
наследственных единиц — генов, хромосомная теория наследственности,
мутации — качественные скачкообразные переходы генов из одного со¬
стояния в другое, отсутствие в общем случае взаимнооднозначного соот¬
ветствия между генотипом и фенотипом) и о популяционно-генетической
организации вида (закон Харди-Вайнберга, обобщение С. С. Четверикова
о генетической гетерогенности природных популяций [2], представление
о факторах динамики генетического состава популяции — мутационном
процессе, дрейфе генов, изоляции, естественном отборе и их взаимодей¬
ствии [3-6]).Популяционной генетикой был решен ключевой вопрос — дано строгое
определение понятия «естественный отбор». Решающий вклад внес Р. Фи¬
шер в своей «Генетической теории естественного отбора». В рамках про¬
стой математической модели Р. Фишер ввел понятие дарвиновской приспо¬
собленности (fitness) генотипа, и тогда логически в дарвиновской теории
происхождения видов все встало на свои места. На словесном уровне: от¬
бор — это дифференциальный вклад разных генотипов в следующее поколе¬
ние. Отсюда вытекает ряд принципиально важных следствий: даже резкие
изменения общей численности популяции ничего не говорят об изменении
вектора и давления отбора; естественный отбор реализуется статистически
(«отбор не гарантирует бессмертия»); приспособленность — всегда относи¬
тельная (одного генотипа по отношению к другому); отбор вдет по фено¬
типу, но при этом отбираются определенные генотипы; необходимо разли¬
чать общую (интегральную) приспособленность и компоненты этой общей
приспособленности, специфичные для определенного вида и определенных
условий среды. Самое же главное заключается в том, что введение поня¬
тия «приспособленность» дало возможность в рамках определенной модели
измерять величину общей приспособленности генотипа и ее компонентов.
Послесловие289Концепция приспособленности генотипов является составной частью син¬
тетической теории эволюции.Современный этап развития концепции приспособленности связан
с включением в нее эпигенетических эффектов, в широком смысле слова —
с рассмотрением генотипа как целостной системы; с представлением о ге¬
терогенности среды обитания в пределах популяционного ареала и потому
с рассмотрением приспособленности не как константы, но как случайной
величины. Актуальность книги еще и в том, что автор уже тогда, 80 лет на¬
зад, поднял и обсудил вопросы происхождения и эволюции этики, морали
и социальной организации у человека, что и сегодня является предметом
дискуссий и научных исследований.Со времени публикации «Генетической теории естественного отбора»
прошло 80 лет. Однако, она продолжает жить, то есть оказывать влияние
на развитие эволюционной и популяционной генетики. Публикация книга
на русском языке необходима, ибо без прошлого нет будущего.Литература[1] Животовский JI. А. Фишер Роналд Эйлмер / Большая Советская Эн¬
циклопедия. 5 изд. Т. 27.[2] Четвериков С. С. (1926) О некоторых моментах эволюционного про¬
цесса с точки зрения современной генетики / Четвериков С. С. Про¬
блемы общей биологии и генетики. Новосибирск: Наука, 1983. С. 170—
226, 270-272.[3] Холден Дж. Факторы эволюции. М.-JI.: Биомедгиз, 1935.[4] Майр Э. Популяции, виды и эволюция. М.: Мир, 1974.[5] Грант В. Видообразование у растений. М.: Мир, 1984.[6] Тимофеев-Ресовский Н. В., _Яблоков А. В., Глотов Н. В. Очерк учения
о популяции. М.: Наука, 1973.
Интересующие Вас книги нашего издательства можно заказать почтой или электронной
почтой:subscribe@rcd.ruВнимание: дешевле и быстрее всего книги можно приобрести через наш Интернет-
магазин: http://shop.rcd.ruКниги также можно приобрести:1. Москва, ИМАШ, ул. Бардина, д. 4, корп. 3, к. 414,
тел.: (499) 135-54-37, (495) 641-69-382. МГУ им. Ломоносова (ГЗ, 1 этаж)3. Магазины:Москва: «Дом научно-технической книги» (Ленинский пр., 40)«Московский дом книги» (ул. Новый Арбат, 8)Книжный магазин «ФИЗМАТКНИГА» (г. Долгопрудный,Новый корпус МФТИ, 1 этаж, тел. 409-93-28)С.-Пб.: «С.-Пб. дом книги» (Невский пр., 28)Роналд ФишерГенетическая теория
ЕСТЕСТВЕННОГО ОТБОРАТехнический редактор А. В. Широбоков
Компьютерный набор и верстка Н. С. Агафонова
Корректор О. А. ШемякинаПодписано в печать 28.03.2011. Формат 60 х 841/16.Печать офсетная. Уел. печ.л. 17,67. Уч. изд. л. 18,98.
Гарнитура Таймс. Бумага офсетная № 1. Заказ № 11-13.
Научно-издательский центр «Регулярная и хаотическая динамика»
426034, г. Ижевск, ул. Университетская, 1.
http://shop.rcd.ru E-mail: mail@rcd.ru Тел./факс: (+73412) 500-295Переплет выполнен в ГУЛ УР «Ижевский полиграфический комбинат»
426039, г. Ижевск, Боткинское шоссе, 180.ISBN 978-5-93972-906-29 785939729062
ran 01» МОСКОВСКИЙ ДОИ книгиФишер Генетическая т
еория естеств.отбораЦена: 545,1