























































































































































































































Текст
В. В. Бабков
МОСКОВСКАЯ ШКОЛА ЭВОЛЮЦИОННОЙ ГЕНЕТИКИ
в
«НАУКА»
АКАДЕМИЯНАУКСССР
ИНСТИТУТ ИСТОРИИ ЕСТЕСТВОЗНАНИЯ И ТЕХНИКИ
В. В. Бабков
МОСКОВСКАЯ ШКОЛА ЭВОЛЮЦИОННОЙ ГЕНЕТИКИ
Ответственный редактор академик Д. К. Беляев
МОСКВА
«НАУКА»
1985
УДК 631.523.55
Бабков В. В. Московская школа эволюционной генетики. М.: Наука 1985.
Книга посвящена становлению отечественной традиции эволюционной генетики, связанной со школой Кольцова — Четверикова. Проанализирована поставленная С. С. Четвериковым центральная проблема новой дисциплины, разработка которой привела к формированию принципов генетики популяций и к формулировке принципов фенотипической реализации гена. Широко использованы архивные материалы.
Для специалистов в области генетики, эволюционизма, философии биологии, а также читателей, интересующихся историей науки.
Рецензенты:
М. Д. Голубовский, Е. Б. Музрукова. Ю И. Новоженов
Василий Васильевич Бабков
МОСКОВСКАЯ ШКОЛА ЭВОЛЮЦИОННОЙ ГЕНЕТИКИ
Утверждено к печати Институтом истории естествознания и техники Академии наук СССР
Редактор издательства ЭА. Вишнякова Художник Ю.С. Шлепер Художественный редактор НА, Фильчагина Технические редакторы НА. Торгашова, Н.М. Бурова Корректор Т.И. Шаповалова
Фотонабор выполнен во 2-й типографии издательства ’’Наука”
ИБ №29336
Подписано к печати 08.04.85. Т—01085. Формат 60x90 1/16. Бумага офсетная N* 1 Гарнитура литературная. Печать офсетная. Усл.печ.л. 13,5
Усл.-кр.-отт. 13,8. Уч.-изд.л. 19,0. Тираж 2500 экз. Тип. зак. 78. Цена 2 руб.
Ордена Трудового Красного Знамени издательство ’’Наука”, 117864 ГСП-7, Москва В-485, Профсоюзная ул., д. 90 Ордена Трудового Красного Знамени 1-я типография издательства ’’Наука” 199034, Ленинград В-34, 9-я линия, 12
2001010000-232 042(02)-85
2I8-85-III
© Издательство «Наука >, 1985
ПРЕДИСЛОВИЕ
Предлагаемая читателю книга В. В. Бабкова посвящена идейно-теоретическому и экспериментальному наследию школы С. С. Четверикова, а также его оценке с позиций современных эволюционно-генетических представлений.
С. С. Четвериков и его ученики впервые экспериментально показали огромную насыщенность природных популяций рецессивными мутациями, составляющими подлинный «резерв мобилизационной изменчивости вида» (И. И. Шмальгаузен). В настоящее время факты, обнаруженные четвери-ковской генетической школой в различных популяциях дрозофил, вскрыты и на многих других видах организмов, а идеи С. С. Четверикова стали общепризнанными и вошли в фундамент современного генетико-эволюционного мировоззрения. Они показали тот материальный базис, с которым оперирует естественный отбор в беспрерывном процессе приспособления и созидания более совершенных форм организмов в меняющихся условиях среды.
Замечательное теоретическое исследование С. С. Четверикова 1926 г. знаменовало объединение принципов дарвиновского отбора и менделев-ского наследования и дало начало теории генетики популяций. Такую же службу сослужили независимые обобщающие работы Р. Фишера, С. Райта, Дж. Б. С. Холдейна 1930—1932 гг. В. В. Бабков ясно показывает, что своеобразие эволюционной концепции С. С. Четверикова своими корнями уходит в традиции русской зоологии, блестящим представителем которой был С. С. Четвериков. Автор постоянно подчеркивает, привлекая для этого вполне конкретные материалы, два момента концепции С. С. Четверикова, определившие ее существенные отличия от концепций его западных коллег: интересы натуралиста и системный подход.
Автор тщательно прослеживает путь развития и обогащения исходных идей С. С. Четверикова, выраженных в его основополагающей работе 1926 г. Внимательный читатель книги В. В. Бабкова несомненно увидит, что вклад московской школы далеко выходит за рамки фактического содержания упомянутой классической работы С. С. Четверикова, хотя она является его единственным произведением по эволюционно-генетической тематике. Идеи С. С. Четверикова сообщили всей проблематике эволюционной генетики импульс столь значительной силы, что его влияние наложило яркий отпечаток на все работы в данной области. Последователи С. С. Четверикова как в нашей стране, так и за ее рубежами пришли к тому фундаментальному положению, что теоретически прогнозированные С. С. Четвериковым и экспериментально показанные им и его учениками процессы движения генов в популяциях свойственны всем видам организмов и что они лежат в основе микроэволюционного процесса. Учение о генетическом грузе, ставшее одним из важных разделов в генетике популяций, и учение о хромосомно-абер-
з
ративном полиморфизме имеют своими истоками фундаментальные идеи С. С. Четверикова.
Не менее важной темой четвериковской школы, получившей яркое отражение в книге В. В. Бабкова, были феногенетические и кариосистематиче-ские исследования. В совокупности с исследованиями генетической структуры популяций они составили целостную неразрывную теоретическую концепцию. Поставленная С. С. Четвериковым проблема природы наследственной изменчивости и механизмов ее поддержания в популяциях далеко не исчерпала себя и в наше время. Скорее даже наоборот, она приобретает сейчас новое звучание и новую остроту. В связи с этим чрезвычайно важен подчеркнутый В. В. Бабковым интегративный, системный подход С. С. Четверикова и его учеников к анализу проблемы.
Уместно напомнить, что идеи, составившие основу современной популяционной биологии, в 1920—1930-е гг. развивались независимо в разных центрах биологических исследований нашей страны. Достаточно упомянуть о замечательных работах Н. И. Вавилова и А. С. Серебровского по геногео-графии и генофонду культурных растений и животных. Однако теория популяционной и эволюционной генетики сложилась именно в исследованиях школы С. С. Четверикова, которым и посвящена книга В. В. Бабкова. Эволюционно-генетические идеи Н. И. Вавилова и А. С. Серебровского выходят за рамки настоящей книги и еще ждут подобного анализа.
Основанная на анализе опубликованных работ, ставших уже библиографической редкостью, и рукописей, разбросанных по различным (в том числе частным) архивам, книга В. В. Бабкова представляет собой капитальную сводку по становлению отечественной эволюционной генетики.
Можно не сомневаться, что книга В. В. Бабкова «Московская школа эволюционной генетики» будет с интересом встречена самым широким кругом биологов.
Д. К. Беляев
ВВЕДЕНИЕ
Эволюционная генетика возникла одновременно в советской и англо-американской биологии, и ее раннее развитие шло поначалу разными путями, которые лишь впоследствии соединились.
Возникновению англо-американской генетики популяций предшествовало противоборство английских менделистов с биометриками, защищавшими принцип отбора. Зашедший в тупик конфликт был снят новым поколением (Р. А. Фишер, С. Райт, Дж. Б. С. Холдейн). Возникшей в результате их работ англо-американской генетике популяций свойственны разработка и применение сложных математических методов и редукционистская методология. Позже, в середине 1930-х гг., англо-американская генетика популяций восприняла возникшее в советской эволюционной генетике направление — экспериментальное изучение свободно живущих популяций. Поэтому можно говорить о двух этапах в развитии англо-американского варианта генетики популяций. Первый — конфликт биометриков и менделистов, его истоки и разрешение — разработан в историческом плане весьма подробно. Существенная роль в этом принадлежит книге У. Провайна «Истоки теоретической генетики популяций» (Provine, 1971). В последнее десятилетие эта тема не сходит со страниц историко-биологической литературы (обзор: Kevles, 1980). Второй же этап — начало экспериментальной генетики популяций на Западе практически не освещен. Для написания адекватной его истории необходимо изучить корни других влияний — советскую эволюционную генетику (ср.: Glass, 1981).
Советская эволюционная генетика имела иные истоки и почву для своего раннего развития. «От фон Бэра через Ковалевского традиции русской школы экспериментальной морфологии перешли к крупнейшим специалистам Н. К. Кольцову и недавно скончавшемуся Д. П. Филатову»,— писал Дж. Нидхэм (1945) об Институте экспериментальной биологии (ИЭБ) в Москве. Когда для организации Лаборатории генетики в ИЭБ пришел С. С. Четвериков, систематик-натуралист и морфолог, его задача определилась так: поставить на генетическую основу изучение проблем дарвинизма. В 1925—1926 гг. началось первое в мире экспериментальное исследование насыщенности свободно живущих популяций наследственными изменениями.
Наряду с С. С. Четвериковым и его женой, А. И. Четвериковой, в этой работе участвовало десять его ближайших учеников и сотрудников, составивших Лабораторию генетики: Б. Л. Астауров, Е. И. Балкашина, Н. К. Беляев, С. М. Гершензон, А. Н. Промптов, П. Ф. Рокицкий, Д. Д. Ромашов, Е. А. Тимофеева-Ресовская, Н. В. Тимофеев-Ресовский, С. Р. Царапкин. Это были молодые зоологи, прошедшие двухлетний Большой зоологический практикум Н. К. Кольцова в Московском университете, испытавшие влияние таких исследователей, как зоологи А. П. Семенов-Тян-Шанский, М. А. Мензбир, А. Н. Северцов, Б. С. Матвеев, Г А. Кожевников, гидробиолог С. Н. Скадовский, цитологи С. Л. Фролова и П. И. Живаго, ботаники М. И. Голенкин и Л. И. Курсанов, геолог А. П. Павлов, палеонтолог М. В. Павлова, географ Д. Н. Анучин и др.
Выдвинутая Четвериковым центральная проблема эволюционной генетики — природа и поддержание изменчивости в популяциях — конкретизировалась его учениками в двух аспектах: популяционном (изменчивость, скрытая в природных популяциях) и феногенетическом (изменчивость, возникающая в ходе фенотипического осуществления гена). Эти два направления ра
5
бот обособились в ходе первых экспериментальных исследований лаборатории Четверикова; впоследствии же они были объединены в циклах работ, посвященных феногенетике массовых популяционных изменений и коррелированной изменчивости конституциональных признаков. Поэтому их выделение несколько условно. Интересно, что англо-американская биология сравнительно скоро восприняла одно из этих направлений — изучение природных популяций — и развивала его редукционистский вариант. Другое направление, давшее важный материал для теории стабилизирующего отбора И. И. Шмальгаузена и кульминацией которого стало изучение интеграции генетических систем, подразумевало системную ориентацию и его перенесение в редукционистскую исследовательскую программу было невозможно. Оно несколько позже независимо возникло и развивалось в работах таких биологов, как К. X. Уоддингтон, С. Д. Дарлингтон, И. М. Лернер.
Если в англо-американской биологии можно провести довольно определенную границу между «генетикой популяций» и «генетикой развития», которые примерно соответствуют двум направлениям работ четвериковцев, то в четвериковской школе они были в значительной мере объединены тем, что их проблемы ставились в рамках единой эволюционной концепции. Подчеркивая это отличие от западной генетики популяций, говорят, что чет-вериковская школа развивала эволюционную генетику 1
Четвериков начал работать над оформлением эволюционной генетики в то время, когда в биологии преобладали два сложившихся канона, конкурировавших друг с другом. Один охватывал круг проблем дарвинизма и был связан со старой традицией систематиков-натуралистов и морфологов и отчасти с новыми методами биометрической школы. Другой был обязан бурному развитию менделизма и был связан с экспериментальным подходом и редукционистской методологией. Четвериков разорвал рамки каждого из двух влиятельных канонов с тем, чтобы строить новую область биологического знания, ибо его работа была основана на признании плодотворности объединения общеэволюционных и генетических представлений, на идее экспериментального изучения изменчивости в природе, на применении системного подхода. В исследовании 1926 г. он выдвинул центральную проблему эволюционной генетики, с первой формулировкой которой была связана определенная модель генетической эволюции — точнее модель элементарного эволюционного явления: возникновение «хороших» мутаций в природе — накопление их во внешне однородных популяциях при свободе скрещивания — выщепление изменчивости при частичной изоляции — отбор по Нортону. Аналогичные модели развивали одновременно Фишер, Райт, Холдейн.
В работах учеников Четверикова шаг за шагом конкретизировалась центральная проблема. Одновременно первоначальная модель явления подвергалась эмпирической проверке, уточнениям, неизбежным деформациям, что привело в конце концов к созданию предпосылок для построения более адекватной модели генетических основ эволюции. Жизнеспособность центральной поблемы эволюционной генетики была обусловлена возможностью *
Замечу в этой связи, что именно с обсуждения вопроса, в чем заключается различие между эволюционной и популяционной генетикой, началась научная часть четвериковской мемориальной сессии Секции популяционной и эволюционной генетики Научного Совета по проблемам генетики и селекции, проводившейся 14—15 мая 1973 г. в г. Горьком и приуроченной к открытию памятника на могиле С. С. Четверикова (Бабков, 19736).
6
пересмотра и в конечном итоге отказа от первоначальной модели генетической эволюции, принявшей на себя все стрелы эмпирических опровержений. Четвериковская проблема, знаменовавшая рождение новой науки, сохраняет свое эвристическое значение и по сей день.
Литература о зарождении советской эволюционной генетики (и в целом экспериментальной биологии) невелика. Выдающееся место в ней принадлежит научным биографиям Н. К. Кольцова и С. С. Четверикова, написанным их учениками Б. Л. Астауровым и П. Ф. Рокицким (1975; Астауров, 1965, 1974; Рокицкий, 1974, 1975). Для правильной оценки роли С. С. Четверикова в современной биологии большое значение имеют публикации ф. Г Добржанского (Dobzhansky, 1955, 1959, 1967), включавшие (Dob-zhansky, 1959) первый английский перевод отрывков классической работы Четверикова 1926 г.; полный перевод ее опубликован под редакцией И. М. Лернера (Chetverikov, 1961; см. также статью Б. Гласса — Glass, 1981).
В работе над этой книгой были использованы прежде всего научные труды ее героев, опубликованные в давние годы и ставшие уже библиографической редкостью. Были использованы немногочисленные историко-научные работы; наиболее ценная (для этой книги) часть их была написана по предложению автора и опубликована им в сборниках «Из истории биологии», №№ 5—7 Многие наиболее ценные материалы были найдены в ряде частных архивов, и их копии (или оригиналы) хранятся у автора. (Только небольшая часть из них опубликована, например, в статьях: Балкашина и др., 1975; Четвериков, 1982; Бабков, 1983). На этой основе в предлагаемой книге впервые сделана попытка систематически проанализировать возникновение и ранние этапы развития советской эволюционной генетики.
Эту книгу я посвящаю моим друзьям и учителям. Написание этой книги стало возможным благодаря щедрой поддержке, которую мне оказывали в разное время Б. Л. Астауров, Н. С. Скадовская-Астаурова, Ел. И. Балкашина, Д. К. Беляев, В. Н. Беляева, К. А. Головинская, А. А. Малиновский, Н. Н. Медведев, 3. С. Никоро, Е. Д. Окольничая, Л. К. Окольничий, П. Ф. Рокицкий, Н. С. Смирнова, Н. В. Тимофеев-Ресовский, В. П. Эф-роимсон и другие. Им я сохраняю неизменную признательность.
Важные мнения высказали Е. Т. Васина-Попова, К. Вепсалайнен, Н. Н. Воронцов, С. М. Гершензон, М. Д. Голубовский, Ф. Г Добржанский, В. И. Иванов, В. С. Кирпичников, Л. В. Крушинский, С. Р. Микулинский, Э. Н. Мирзоян, Е. Б. Музрукова, Ю. М. Новоженов, К. Ота, В. Орел, Й. Рихтер, Н. Ролл-Хансен, К. Б. Соколова, А. П. Сушкина, С. В. Хвощинская, М. А. Шишкин, Ф. Эджертон и многие другие. Рецензентами окончательного варианта рукописи были И. А. Рапопорт, Б. С. Соколов, А. Л. Тахтаджян. Всем им я глубоко благодарен.
Ряд материалов мне предоставили: Архив Академии наук СССР (Москва, Ленинград), Отдел рукописей Публичной библиотеки (Ленинград), библиотеки Института истории естествознания и техники АН СССР, Института биологии развития им. Н. К. Кольцова АН СССР, Хельсинского университета. Выражаю благодарность их сотрудникам. Важными для окончательного варианта книги были обсуждения затронутых в книге вопросов в ходе бесед и дискуссий на международном Менделевском симпозиуме в Купаровице и Брно (ЧССР) в августе 1982 г. и во время моих лекций в университетах Хельсинки и Турку (Финляндия) в ноябре—декабре 1983 г. За возможность этих поездок я благодарю Чехословацкую комиссию ЮНЕСКО, Международную генетическую федерацию и Академию наук Финляндии.
Глава 1
истоки школы
Образ мира, в слове явленный.
Б. Л. ПАСТЕРНАК
Московская школа эволюционной генетики возникла на основе соединения двух влиятельных направлений русской зоологии — натуралистического (систематики и фаунистики) и экспериментального (экспериментальной морфологии). Эту школу иногда называют «четвериковской» — по имени ее основателя Сергея Сергеевича Четверикова, натуралиста по преимуществу, который в 1925—1926 гг. провел со своими учениками первое экспериментальное исследование насыщенности наследственными изменениями свободно живущих популяций дрозофил и сформулировал основные понятия и центральную проблему новой области биологии, получившей позже название «эволюционная генетика». Ее называют также «кольцовско-четвериковской» школой, подчеркивая роль основателя отечественной экспериментальной биологии Николая Константиновича Кольцова и кольцовского Института экспериментальной биологии, где в начале 20-х гг. Четвериков организовал Генетическую лабораторию.
Чтобы правильно охарактеризовать научную школу и представить ее в исторической перспективе, а не как изолированное явление, «чудесный дар с небес», нам необходимо сформулировать круг разрабатываемых ею вопросов; рассмотреть свойственные ей формы организации научного общения и отношения учителей и учеников; определить присущие ее членам руководящие метанаучные основания (которые обычно в явном виде не формулируются) — систему познавательных ориентиров, наличие которой, собственно, и объединяет группу исследователей в научную школу. Но прежде всего нам необходимо выяснить исторический контекст происхождения школы.
В этой главе мы поэтому кратко коснемся натуралистической и экспериментальной традиций русской зоологии, нашедших свое отражение в чертах московской школы эволюционной генетики. Мы рассмотрим влияние личности Кольцова и атмосферы основанного им института на формирование этой школы. Далее речь пойдет о своеобразии подхода Четверикова и происхождения его идей, отличавшем основателя московской школы от его англо-американских коллег, предпринявших одновременно с ним попытку дать генетическое обоснование кругу явлений, охватываемых дарвинизмом. Мы обратимся к роли дискуссионного кружка по вопросам генетики, цитологии, эволюции в формировании научного мировоззрения основоположников отечественной эволюционной генетики. Наконец, мы разберем ч^етвериков-скую программу построения новой области биологии, в которой главное место занимает удерживающая свое эвристическое начало и по сей день ее центральная проблема: природа и поддержание изменчивости в популяциях.
Но начнем с традиций русской зоологии.
8
О ДВУХ НАПРАВЛЕНИЯХ В РУССКОЙ ЗООЛОГИИ
Натуралистическое и экспериментальное направления в русской зоологии (объединение их элементов и определило своеобразие московской школы эволюционной генетики) просматриваются на всем пути ее развития, с самого ее зарождения. Существуют разные мнения о том, какой момент считать началом русской зоологии. Иногда ссылаются на то, 'Что ранние сведения о животных Руси можно найти в «Поучении» Владимира Мономаха, во второй (автобиографической) части которого он перечисляет походы, пережитые опасности свои, подвиги на войне и охоте, которая была тогда важным делом ввиду обилия хищных зверей и их опасности для населения. Однако зоологические сведения все так же бедны и отрывочны в описаниях путешествий и более поздних времен, включая петровские и елизаветинские. (Петр 1, учредивший Академию наук, не успел осуществить свои планы по изучению России). Экспедиции, от которых можно отсчитывать начало русской зоологии, состоялись позже (ср.: Плавильщиков, 1941). В 1769 г. ожидалось прохождение Венеры через диск Солнца, и в 1768 г. для астрономических наблюдений и с целью возможно полного изучения природы России были организованы путешествия П. С. Палласа, И. И. Лепехина и ряд других. Именно на примере Палласа мы впервые встречаемся с зачатками двух направлений, натуралистического и экспериментального. Палласа (вместе с Э. А. Эверс-манном) можно считать основателем отечественной зоогеографии. Однако он занимался не только фаунистикой («Zoographia Rosso-Asiatica...», 1811 и 1831), но и экспериментом с паразитическими червями, опровергая мнение об их самозарождении. А нашим первым экспериментатором был зоолог профессор М. М. Тереховский; он, в частности, опубликовал в 1775 г. результаты своих опытов, опровергающих идею самозарождения.
Два уже отчетливо различающихся направления в русской зоологии — экспериментальное и натуралистическое — идут от Карла Бэра. Бэр внес существенный вклад как в создание экспериментальной эмбриологии, так и в исследования фауны России. В значительной мере под влиянием Бэра сложилась русская школа систематиков и фаунистов. Основополагающей для развития экспериментального направления в русской зоологии явилась магистерская диссертация 1865 г. А. О. Ковалевского, посвященная истории развития «замечательной рыбки» (ланцетника, открытого в 1784 г. Палласом), которая и положила начало сравнительной эмбриологии.
К. Ф. Рулье и С. С. Куторга были последними натуралистами-энциклопедистами. С начала дарвиновского периода в русской зоологии проявляется тенденция к специализации: ученик Эверсманна М. Н. Богданов может быть назван родоначальником русской научной орнитологии, К. Ф. Кесслер — ихтиологии. Труд М. А. Мензбира «Птицы России» (2тт., 1895) оказал глубокое влияние на развитие отечественной орнитологии, как «Рыбы пресных вод России» (1916) Л. С. Берга на ихтиологию. Неоконченная сводка «Жуки России и Западной Европы» (1905—1915) Г Г Якобсона дала начало росту отечественной колеоптерологии. Точную категоризацию низших систематических единиц предпринял энтомолог А. П. Семенов-Тян-Шанский, который ввел понятие «подвид» (1910).
Традиции целостного мировоззрения натуралистов с широким кругозором не исчезли в период специализации (Бабков, 19836). В. И. Вернадский явился основоположником учения о биосфере. Биогеоценоз как элементарная биохорологическая единица стал предметом самостоятельной естественно
9
исторической дисциплины — бивгеоценологии, которую развивал В. Н. Сукачев, продолжая линию Вернадского. Биосферно-биогеоценотическая концепция наиболее полно и точно выражает основную естественно-научную идею В. В. Докучаева о необходимости создания особой науки, призванной, как отмечал Н. В. Тимофеев-Ресовский, изучать «генетическую, вечную и всегда закономерную связь, какая существует между силами, телами и явлениями, между мертвой и живой природой; между растительным, животным и минеральным царствами» (Докучаев, 1899, цит. по: Тимофеев-Ресовский и др., 1973, с. 16). Можно говорить о преемственности идей Докучаева и Рулье, который считал достойной «первейших ученых» задачей «исследовать три вершка ближайшего к исследователю болота относительно растений и животных и исследовать их в постепенном взаимном развитии организации и образа жизни посреди определенных условий» (цит. по: Плавильщиков, 1941, с. 258). Так складывались основы системного мышления в отечественной биологии, столь характерного для московской школы эволюционной генетики.
Н. К. КОЛЬЦОВ и кольцовский ИНСТИТУТ
Всестороннюю оценку вклада Николая Константиновича Кольцова (1872—1940) в развитие отечественной биологии дали его ученики и сподвижники Б. Л. Астауров и П. Ф. Рокицкий (1975) 1
Первую научную работу Кольцов выполнил еще студентом университета и доложил ее в 1894 г. на IX Съезде русских естествоиспытателей и врачей. В Кабинете сравнительной анатомии Мензбира Кольцов был в центре университетской зоологии. Там были эмбриолог и гистолог В. Н. Львов, цитолог и философ Н. А. Иванцов, сравнительный анатом А. Н. Северцов, орнитолог П. П. Сушкин. Новые встречи с биологами принесли заграничные командировки: в 1897—1900 гг. в лабораторию В. Флемминга в Киле (где Кольцов работал с Ф. Мёвесом), затем на международную биостанцию в Неаполе, французскую в Роскове и русскую в Виллафранке (Виль Франш) на французской ривьере, и в 1902—1903 гг. на биостанцию в Виллафранке и в лаборатории ряда германских университетов, в том числе лабораторию О. Бючли в Гейдельберге. В Неаполе он жил в одном пансионе с Г Дришем и К. Гербстом. Дриш только что опубликовал опыты по развитию яиц морского ежа и по регенарации полипов, которые вместе с опытами В. Ру положили начало механике развития. Гербст приступал к изучению влияния отдельных ионов морской воды на развитие яйца морского ежа, предвосхищая тем самым начало физико-химической биологии. Одновременно работал в Неаполе американский цитолог Э. Вильсон, незадолго до этого опубликовавший книгу «Клетка и ее роль в развитии и наследственности» (первое издание в 1896 г.), о влиянии которой на развитие генетики Кольцов писал (1936, с. 16—17): «Книга Э. Вильсона, которой я зачитывался еще в Москве, была смелым пророчеством будущей генетической цитологии, и я не сомневаюсь, что этот превосходный ученый оказал большое влияние на работу своих сотрудников и учеников по Колумбийскому университету в тот период, когда через 10 лет после нашей встречи в Неаполе вырабатывалось стройное учение морганизма».
1 Ряд архивных материалов, касающихся жизни и трудов Н. К. Кольцова, ввел в научный обиход В. М. Полынин (Блантер) в свой популярной книге «Пророк в своем отечестве» (М.: Сов. Россия, 1969. 126 с.). См. также: Кольцов Николай Константинович, 1976.
10
Особенно дружная молодая группа зоологов составилась весной 1899 г. в Виллафранке, куда приехали студенты из гейдельбергской лаборатории О. Бючли и мюнхенские ученики Р. Гертвига, среди них Рихард Гольдшмидт и Макс Гартман; к этой группе присоединился зам. директора биостанции М. М. Давыдов. «...Мы хотели посвятить свою жизнь изучению организации клетки, сравнительной и экспериментальной цитологии. Нам казалось, что эта большая проблема может нас объединить, и мы распределяли между собой участие в ее разработке, рассчитывая каждый работать у себя на родине, а весной съезжаться около моря, чтобы собирать материал... Наша тройка — Гольдшмидт, Гартман и я — осталась верна нашей молодости... Наши последующие встречи были более редкими, чем мы рассчитывали в молодости, но мы всегда встречаемся друзьями и в науке и в жизни» (Кольцов, 1936, с. 17). Гольдшмидт на склоне лет вспоминал о Виллафранке: «Там был блестящий Николай Кольцов, возможно лучший зоолог нашего поколения, доброжелательный, непостижимо образованный, ясно мыслящий ученый, обожаемый всеми, кто его знал. Он часто наезжал в западно-европейские лаборатории, и мы были друзьями со студенческих дней» (Goldschmidt, 1956, р. 106). «Я горжусь, что такой благородный человек был моим другом всю жизнь» (Goldschmidt, 1960, р. 240).
Из второй заграничной командировки Кольцов вернулся в 1904 г. с готовой докторской диссертацией, посвященной теории клеточной формы. Она была назначена к защите в середине января 1906 г. Кольцов отказался защищать диссертацию в дни кровавого подавления революции, и докторскую степень он получил уже в советское время (в 1935 г.). В дни первой русской революции в кабинете Кольцова в Институте сравнительной анатомии печатались и хранились воззвания Центрального университетского органа, созданного 14 февраля 1905 г. для руководства студенческим движением. Не остался в стороне от революционных событий и С. С. Четвериков. В декабре 1905 г. он как представитель университета участвовал в работе Московского стачечного комитета (Четвериков, 1980, гл. 5). Этим событиям была посвящена и книга Кольцова «Памяти павших. Жертвы из среды Московского студенчества в октябрьские и декабрьские дни», изданная в середине 1906 г. Книгу в первый же день конфисковали, но больше половины тиража успело разойтись, и вырученные от продажи деньги были переданы астроному-большевику П. К. Штернбергу, члену Военно-технического бюро Московского комитета РСДРП, для оказания помощи арестованным студентам.
С началом академического года Кольцов из-за участия в революционных событиях был лишен возможности вести исследования и практические занятия в университете (за ним осталось лишь чтение лекций). Лабораторию ему пришлось организовать в своей квартире. В брошюре «К университетскому вопросу» (1909 и 1910) Кольцов остро критиковал порядки в университете и призывал к широким реформам. Правительственные циркуляры «О надзоре за учащимися высших учебных заведений» и «О временном недопущении публичных и частных студенческих заведений» (3 и 11.1.1911) знаменовали начало открытого курса правительства на окончательное уничтожение демократических свобод в университете. В ответ на политику правительства в области высшей школы ректор А. А. Мануйлов, проректор и помощник ректора подали в отставку; 1.2.1911 министр народного просвещения А. Л. Кассо принял их отставку от административной должности, более того — отстранил их от научной и преподавательской деятельности в университете. В феврале
11
1911 г. Кольцов в числе приблизительно 400 профессоров и сотрудников покинул университет. (Вернулся туда он только после Октябрьской революции.) В 1911 —1918 гг. Кольцов преподавал на Высших женских курсах проф. В. И. Герье и в Московском городском народном университете им. А. Л. Шанявского, которые не подчинялись Министерству народного просвещения и имели возможность проводить независимую политику. В университете им. Шанявского Кольцов создал в 1913 г. прекрасно оборудованную исследовательскую лабораторию; там начали работать его ученики, и многие из них впоследствии стали крупными научными работниками: М. М. Завадовский, П. И. Живаго, И. Г Коган, В. Г Савич, А. С. Серебров-ский, С. Н. Скадовский, Г И. Роскин, Г В. Эпштейн и др. В 1916 г. он приступил к организации Института экспериментальной биологии Общества Московского научного института. Общество, созданное на частные средства и поддерживаемое Московской городской думой, наметило создание нескольких чисто исследовательских институтов (в том числе по микробиологии для Л. А. Тарасевича и по биофизике для П. П. Лазарева). Это была передовая для своего времени мысль: тогда в России вообще не было биологических лабораторий, не связанных с университетским преподаванием (если не считать Зоомузея и маленькой Особой зоологической лаборатории Академии наук). Институт с тремя штатными научными сотрудниками открылся в середине 1917 г. Когда 1 января 1920 г. ИЭБ вошел в систему Наркомздрава РСФСР, многие сотрудники, занимавшиеся в Институте «добровольцами», т. е. бесплатно, были включены в штат.
При организации ИЭБ Кольцов уделял особое внимание комплексному подходу к явлениям жизни. Для этого в 1920 г. были созданы отделы (лаборатории) по главным отраслям экспериментальной биологии: генетике, цитологии, эндокринологии, физико-химической биологии, гидробиологии, механике развития, зоопсихологии. С этой же целью каждая завершенная работа обсуждалась на общеинститутском коллоквиуме. Дело было не только в том, чтобы найти общий язык («ионщики должны понимать генщиков, и наоборот»),— попытка связать различные области экспериментальной биологии отражала мысль Кольцова о необходимости их внутреннего синтеза. Те же умонастроения были характерны для Генетической лаборатории: Четвериков считал подход к изучению эволюционного процесса, учитывающий достижения разных дисциплин, условием его «синтетического понимания».
Принципы, положенные в основу работы кольцовского института, например правомерность экспериментального метода в биологии, представлялись его молодым сотрудникам «чем-то само собой разумеющимся» (Астауров, Ро-кицкий, 1975, с. 26). Лишь за пределами ИЭБ можно было убедиться в том, что многие биологи того времени сдержанно или отрицательно относились к некоторым идеям Кольцова, в том числе к применению экспериментальных методов исследования. Однако в коллективе ИЭБ молодым зоологам не приходилось тратить времени и сил, чтобы доказывать такие положения: они входили в их систему познавательных ориентиров. То же можно сказать и о лаборатории Четверикова, где тезис о совместимости менделизма и дарвинизма (в условиях еще не окончившегося конфликта представителей этих двух направлений) принимался как не подлежащий обсуждению.
В ИЭБ заслушивались доклады гостей из других институтов. Здесь не раз выступал А. Н.. Северцов, основоположник эволюционной морфологии жи-
12
вотных, который всегда находил в ИЭБ заинтересованную и доброжелательную аудиторию.
С раннего периода своего существования ИЭБ начал получать международное признание. В числе зарубежных гостей были К. Бриджес (позже, в 1931 г., он выступил на сессии АН СССР с лекцией «Генетическая концепция жизни»), Г. Дж. Мёллер (в 1922 г. он прочел на Аниковской генетической станции доклад о результатах десятилетних исследований с дрозофилой лаборатории Моргана), Дж. Б. С. Холдейн (который впоследствии привлек внимание английских исследователей к идеям Четверикова), О. Фогт (разработавший ряд передовых цитоархитектонических методов исследования мозга), У Бэтсон (один из основоположников менделевской генетики, который предложил термин «генетика»), Р. Гольдшмидт, Э. Ваксман, С. Дарлингтон и др. Особенно важны были приезды зарубежных ученых в первые годы Советской власти, так как они положили начало прорыву научной блокады Советской России. В июне 1927 г. в Берлине при участии О. и С. Фогтов была организована Неделя русской науки, в которой участвовали А. В. Луначарский, Н. А. Семашко, Н. К. Кольцов, В. И. Вернадский, И. И. Шмальгаузен и др. (Астауров, Рокицкий, 1975, с. 26; Филимонов, 1960, с. 1672).
В 1925 г. по предложению Советского правительства Оскар Фогт принял участие в создании в Москве филиала берлинского Института мозга (в 1928—1937 гг. Государственный институт мозга им. В. И. Ленина при Ученом Комитете ЦИК СССР, директор О. Фогт, заместители Н. И. Филимонов и С. А. Саркисов, в настоящее время Институт мозга АМН СССР) 2 В этой связи был организован научный обмен. Молодые сотрудники Фогта приехали для работы в московском филиале; из ИЭБ для организации генетического отдела в берлинском институте были приглашены в 1925 г. Н. В. и Е. А. Тимофеевы-Ресовские, в 1927 г. С. Р. Царапкин (см.: Воронцов, Яблоков, 1970, с. 145).
В Москве места для работы в первое время не хватало: ИЭБ занимал три большие комнаты в доме №41 по пер. Сивцев Вражек, хотя и был оснащен превосходным лабораторным оборудованием. Лишь в 1925 г. он получил прекрасный трехэтажный особняк по ул. Воронцово поле (ныне ул. Обуха), дом № 6 (Астауров, Рокицкий, 1975, с. 24). Поэтому довольно рано часть работ была перенесена за город. Звенигородская гидрофизиологическая станция, построенная С. Н. Скадовским в 1910 г., стала как бы летним филиалом ИЭБ. Именно там были проведены первые работы Генетической лаборатории Четверикова. В 1919—1921 гг. недалеко от нее, сперва в Аниково, потом в Назарьево, была организована Центральная станция по генетике сельскохозяйственных животных (ЦГС), которую финансировала Комиссия по изучению естественных производительных сил России при
2 Об истории Института мозга в Москве подробнее см.: О. С. Адрианов, Н. С. Преображенский, Л. А. Куку ев. Институт мозга АМН СССР. (К 50-летию основания.) — В кн.: Современные аспекты учения о локализации и организации церебральных функций. М.: Медицина, 1980, с. 5—33. См. также: Филимонов (1960); Richter (1976). (Архивные материалы бывшего Института мозга в Бух-Берли не, которые хранятся в Институте им. О. и С. Фогтов в Дюссельдорфе (ФРГ), включают значительную, около 200 писем, переписку О. Фогта с советскими коллегами. Среди его корреспондентов Н. К. Кольцов, С. С. Четвериков, а также Н. А. Семашко, П. П. Лазарев, Л. С. Минор, Л. С. Штерн, Н. П. Горбунов и др.).
На Аниковской генетической станции
Верхний ряд (слева направо): 4-й — А. Н. Промптов, 5-й — Л. В. Ферри, 6-я — М. А. Арсеньева (Гептнер), 7-я —Л.П. Промптова, 9-я — М. П. Садовникова-Кольцова, 10-й — Б. Л. Астауров, 11-й— С. С. Четвериков, 12-я — Е. И. Балкашина, 13-я — Е. А. Тимофеева-Ресовская, 14-й — Н. В. Тимофеев-Ресовский, 15-я — В. А. Бродская, 17-й—С. Р. Ца pan кин, 18-й —П. И. Живаго. Средний ряд (слева направо): 1-й —В. А. Рацыборский, 2-й — А. С. Серебровский, 3-я — А. Н. Савич, 4-й — Н. К. Кольцов, 5-я — Е. В. Лебедева, 6-я — А. И. Четверикова, 7-й — И. Г Коган, 8-я — Н. Г Савич. Нижний ряд (слева направо): 1-й—С. В. Лебедев, 2-я — Герасимович, 3-я Н. В. Лебедева (Эфрон), 4-я — Т. П. Живаго, 5-я — А. А. Серебровская. Снимок В. Н. Лебедева, 1924 г.
российской Академии наук (КЕПС), а затем Наркомзем. (Об истории ЦГС см.: Рокицкий, Васина-Попова, 1978.)
Н. К. Кольцов сыграл значительную роль в создании отечественной биологической печати. Он принимал участие в сериях книг «Современные проблемы естествознания» и «Классики естествознания», в журналах «Природа», «Научное слово», «Социалистическая реконструкция и наука» («Со-рена»), горьковском «Наши достижения». Он редактировал «Ученые записки Московского городского народного университета им. А. Л. Шанявского. Труды биологической лаборатории» (1916). «Известия Института экспериментальной биологии», вып. I (1921) были первым сборником по экспериментальной биологии (в том числе по генетике), изданным после Октябрьской революции. В 1922 г. Кольцов организовал журнал «Успехи экспериментальной биологии», в 1925 г.— «Журнал экспериментальной биологии» (в двух сериях, одна —для оригинальных работ, продолжившая изданный Н. Кольцовым и Ю. Филипченко «Бюл. МОИП. Отд. эксперим. биол.», нов. серия, т. 1, 1924, другая — для обзорных работ, пять томов, на основе «Успехов...»), преобразованный в 1932 г. в «Биологический журнал», а в 1940 г.— в «Журнал общей биологии».
Большое влияние на первое поколение советских эволюционных генетиков Кольцов оказал и как преподаватель. Они читал тогда в университете «Введение в биологию» (этот курс, был начат в 1899 г. как курс цитологии) и новый курс «Систематическая зоология». Его лекции производили большое впечатление. Среди учеников Кольцова в Университете и в ИЭБ оказались Е. И. Балкашина, Д. Д. Ромашов, Е. А. Фидлер (впоследствии Тимофеева-Ресовская), Н. В. Тимофеев-Ресовский, А. Н. Промптов. Единое целое с лекциями составлял двухлетний Большой зоологический практикум, созданный Кольцовым еще на Высших женских курсах. Студенты вели там самостоятельную научную работу; при практикуме были специальные курсы с практическими занятиями по кариологии (П. И. Живаго), по генетике и биометрии (С. С. Четвериков), по физико-химическим методам (В. Н. Шредер) и др. Такая организация обучения обеспечила им широкую общебиологическую подготовку и способствовала формированию самостоятельного научного мышления.
ЕСТЕСТВЕННО НАУЧНЫЕ ВЗГЛЯДЫ С. С. ЧЕТВЕРИКОВА
Сергей Сергеевич Четвериков (1880—1959) в своих «Воспоминаниях» отметил, что с детских лет любовь его к природе «перешла в сознательное желание сродниться с природой не только душой, но и умом» (Четвериков, 1980, гл. 1, с. 92). Отец предполагал для него карьеру инженера, но «любовь к вольной жизни природы» привела С. С. Четверикова к убеждению, что «вне биологии нет для него призвания». В 1900 г. Четвериков поступил на биологическое отделение Физико-математического факультета Московского университета (см. биографию Четверикова: Астауров, 1974).
На первом курсе Московского университета (1900—1901 гг.) С. С. Четвериков участвовал в зоологическом кружке проф. Н. Ю. Зографа (который в 1912 г. начнет чтение курса экспериментальной зоологии, где давались основы генетики) и особенно активно в работе руководимой Г. А. Кожевниковым (тогда доцентом) Комиссии для исследования фауны Московской губернии при императорском Обществе любителей естествознания, антропологии и этнографии. В Комиссии был сделан его первый научный доклад;
15
там же опубликована его широко известная работа «Волны жизни» (1905; переизд.: 1980, 1983) 3 /
На первый учебный год приходится и первое его «боевое крещеное». В Киевском университете вспыхнули местные беспорядки, и министр Н. П. Боголепов на основе «временных правил» от 29.VII.1899 г. отправил из университета в солдаты всех недовольных студентов. (Этому событию посвящена статья В. И. Ленина «Отдача в солдаты 183-х студентов» (Поли, собр. соч., т. 4, с. 391—396), опубликованная в «Искре» в феврале 1901 г.) Московское студенчество ответило забастовкой, и Четвериков был тогда в числе арестованных. (О его участии в этих событиях см.: Четвериков, 1980, гл. 4 и 5).
Весной 1906 г. по окончании университета Четвериков был оставлен «для подготовки к профессорскому званию» по кафедре сравнительной анатомии М. А. Мензбира; его магистерская диссертация была посвящена анатомии небольшого ракообразного — водяного ослика (Tschetwerikoff, 1910; русский перевод см. в кн.: Четвериков, 1983, с. 84—169, 268—269). В 1909 г. Четвериков начал читать курс энтомологии на Высших женских курсах (после Октябрьской революции — 2-й МГУ, который вместе с 3-м МГУ, бывшим университетом Шанявского, 26.VIII. 1919 г. вошел в состав МГУ); с 1919 г. он читал в МГУ энтомологию и введение в теоретическую систематику (биометрию), в 1925—1929 гг.— начал первый в МГУ курс генетики и подолжал курс биометрии (Астауров, 1974а, с. 62).
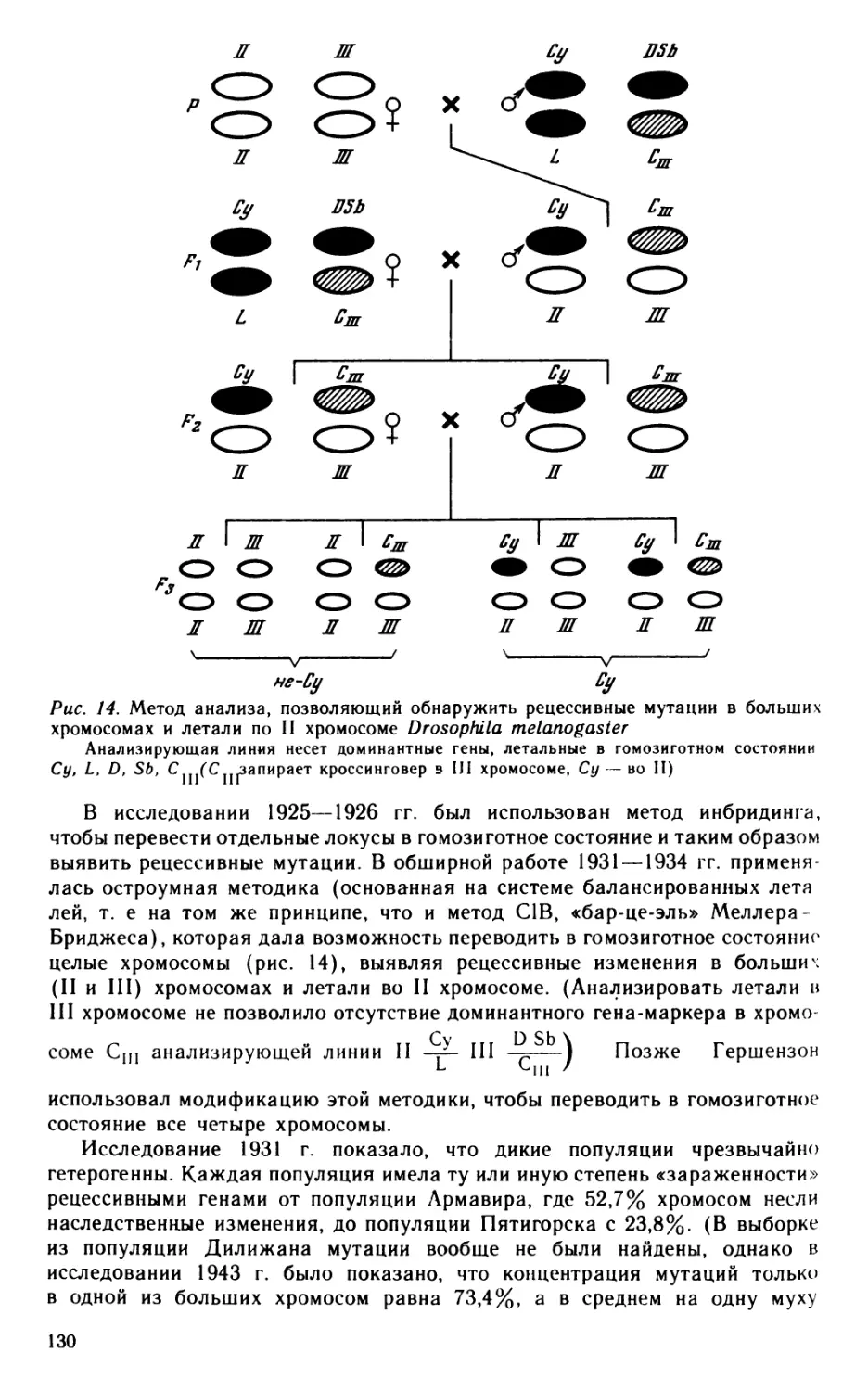
До 1921 г., когда по предложению Кольцова Четвериков организовал в ИЭБ Генетическую лабораторию, генетикой он специально не занимался. Задолго до своей знаменитой работы 1926 г. Четвериков независимо пришел к «Основному закону популяции», который сочетал закон равновесия при свободном скрещивании, сформулированный ранее Гарольдом Харди (Hardy, 1908), с законом стабилизирующего скрещивания (Pearson, 1904). Как известно, закон Харди является основой генетики популяций. Иногда его называют также «законом Харди—Вайнберга», отмечая прошедшую незамеченной для генетики популяций одновременную статью В. Вайнберга (Weinberg, 1908), к которой привлек внимание К. Штерн (Stern, 1943), когда теория генетики популяций уже сложилась. Работа Вайнберга осталась в стороне от магистрального пути развития генетики популяций (Provine, 1971; Ohta, 1981). Классическая работа Четверикова 1926 г., которая дала теоретическое обоснование экспериментальной генетики популяций, явилась разработкой его «Основного закона популяции». (Это, как будто, дает основание говорить о «законе Харди-Четверикова»
3 Куниёси Ота сообщил мне, что в написанной им совместно с Хисао Канэко биографии «Биолог: жизнь С. С. Четверикова» (ныне опубликована на японском языке: Kaneko, Ohta, 1982) привлекались материалы «Автобиографии» основоположника энтомологии в Японии — Сёнен Мацумура (1871 —1960), вышедшей в Токио в 1960 г. на японском языке. Согласно «Автобиографии», проф. университета Хоккайдо Мацумура представлял Академию наук Японии на праздновании 200-летия АН СССР в Ленинграде и встречался с Четвериковым в Москве, так как в то время Мацумура изучал систематику и биономию соснового шелкопряда рода Dendro-limus на Сахалине, а Четвериков обладал мировой известностью как эксперт по этим насекомым. Канэко и Ота отметили, что Четвериков несколько раз упомянул Японские острова в своей знаменитой статье 1926 г. (рассматривая пример изоляции, с. 37—38), и выдвинули мнение, что его интерес к японской фауне был вызван встречей с Мацумурой.
16
Сергей Сергеевич Четвериков
или «Харди—Вайнберга—Четверикова». Впрочем, в статье 1926 г. Четвериков ссылается на «закон Харди»; последнее название принято в генетике популяций наших дней.) Итак, первым же шагом в новой для Четверикова области была формулировка основного положения генетики популяций.
Работы по генетическому анализу природных популяций дрозофил «были начаты в связи с замечательным исследованием С. С. Четверикова (1926), посвященным в первую очередь вопросам генетического строения вида» (Балкашина, Ромашов, 1935, с. 81), и явились «экспериментальной проверкой некоторых сторон эволюционной теории С. С. Четверикова» (Н. A. Timo-feeff-Ressovsky, N. W. Timofeeff-Ressovsky, 1927, S. 72). Поэтому следует обратиться к статье Четверикова «О некоторых моментах эволюционного процесса с точки зрения современной генетики» (1926), которая при переиздании 1965 г. была названа «вехой в развитии генетических представлений» (Астауров, 1965). Движущей силой в выработке нового взгляда стало известное противоречие между генетическими и общеэволюционными представлениями, которое Четвериков считал должным разрешить (Рокицкий, 1974, 1975; Бабков, 1977а, 1983а). В отличие от многих крупных биологов начала века Четвериков рассматривал менделевские законы наследственности не как особый случай передачи наследственных свойств. Будучи основой наследования при половом размножении, менделизм, по Четверикову, должен дать ключ к пониманию процесса эволюции. В результате оформилась задача: «Как связать эволюцию с генетикой, как ввести наши современные генетические представления и понятия в круг тех идей, которые охватывают эту основную биологическую проблему? Можно ли подойти к вопросам изменчивости, борьбы за существование, отбора,— словом, дарвинизма — исходя не из тех совершенно бесформенных, расплывчатых, неопределенных воззрений на наследственность, которые только и существовали во времена Дарвина и его непосредственных преемников, а из твердых законов генетики?» (Четвериков, 1926, с. 4).
2 В. В. Бабков
17
Такая постановка задачи — найти генетическую основу эволюционных обобщений — представляется нам сейчас закономерной и неизбежной (Рокицкий, 1975; Бабков, 1977а; Беляев, Рокицкий, 1977). Первые указания на совместимость менделизма и дарвинизма были опубликованы в Англии в первое же десятилетие после повторного открытия Менделизма в 1900 г. (например, Yule, 1907), однако из них ничего не воспоследовало. Напротив, менделизм был встречен враждебно многими эволюционистами, русскими и зарубежными (а дарвинизм также отрицательно — некоторыми генетиками). Причину этого Четвериков видел в косности ума, которая препятствует включению в корпус биологического знания достижений быстро растущей генетики: «генетика в своих выводах... слишком жестоко ломает привычные, глубоко гнездящиеся представления, а наша теоретическая мысль неохотно меняет хорошо накатанные колеи привычных логических обобщений на неровную дорогу новых, хотя бы и более соответствующих нашим современным знаниям построений» (Четвериков, 1926, с. 4).
Чтобы в полной мере оценить значение работы Четверикова 1926 г., следует обратиться к историческим условиям, на фоне которых она появилась.
Конфликт менделистов и биометриков. Рассматривая эволюцию как медленный и постепенный процесс, Ч. Дарвин видел основной недостаток своей теории в том, что она не объясняла происхождение материала для эволюции. Френсис Гальтон, защищая принцип отбора, заключил на основе своего закона регрессии — реверсии (или «закона предковой наследственности» — «law of ancestral heredity»), что эволюционно значимая изменчивость должна быть прерывистой: реверсия (т. е. тенденция уклонений вернуться к исходному типу) настолько могущественна, что почти нацело снимает действие отбора; поэтому отбор непрерывных вариаций мог бы иметь лишь ограниченный эффект. В 1890-е гг. в Англии развернулись споры о характере эволюции — прерывистом или непрерывном — и каждая из сторон подтверждала свои взгляды ссылкой на авторитет Гальтона.
Один лагерь возглавил Уильям Бэтсон. Изучая наследственную изменчивость, он в 1890-е гг. пришел, как и Гальтон, к убеждению в эволюционном значении прерывистой изменчивости. Бэтсон развивал экспериментальный подход и был сторонником индуктивного метода («истинно дарвинова», как он считал). В другом лагере был Рафаэл Уэлдон и математик Карл Пирсон, защитники дарвинизма (представленного тогда натуралистами и морфологами, которым свойствен иной склад мышления, чем экспериментаторам) и приверженцы гипотетико-дедуктивного метода («истинно дарвинова», по их мнению). Они полагали, что эволюция идет посредством отбора, действующего на непрерывные вариации. Именно для изучения таких вариаций подходили применяемые ими статистические методы. Рассматривая Гальтона как основателя методов биометрической школы, Пирсон и Уэлдон считали себя его последователями. Будучи приверженцем идеи непрерывной эволюции, Пирсон, объясняя свою позицию, утверждал, что «истинно галь-тонианскому методу» следовали именно он и Уэлдон, тогда как сам Гальтон сбился с пути. Согласно «закону предковой наследственности» в модификации Пирсона, существует реверсия не к среднему общей популяции, как у Гальтона, но к среднему популяции родителей. Отсюда выводилась возможность эволюции путем отбора непрерывных вариаций (поскольку реверсия не очень велика и не нивелирует эффект отбора). Любопытно, что позже Вильгельм Иогансен также утверждал, что Гальтон не понял своего закона регрессии — реверсии. По его мнению, в чистой линии существует реверсия
18
полностью к типу линии (еще более сильная, чем у Гальтона); отбор неэффективен против такой реверсии; значит, «гальтонианский метод» должен фсти к прерывистрй эволюции.
С возникновением в 1900 г. менделевской генетики, основанной на экспериментальном методе и принципе прерывности, разногласия усилились, и в ходе диспута между пионерами генетики и биометриками менделевское наследование было противопоставлено дарвинову отбору4 (Provine, 1971; Cock, 1973; Norton, 1973, 1975; Farrall, 1975; Roll-Hansen, 1980; MacKenzie, 1981; обзор: Kevles, 1980).
В исторических анализах этого знаменитого диспута были выдвинуты различные ключевые интерпретации. Утверждалось, что разногласия привели к конфликту двух лагерей и что именно эта личная вражда отложила примирение меделизма и биометрии на целых 15 лет (Provine, 1971, р. 63— 64). Действительно, публичные обсуждения часто выливались в страстные споры, которые изобиловали презрительными насмешками и оскорблениями. Несомненно и то, что конфликт между лидерами поставил в невыгодное положение тех, кто пытался соединить менделизм и биометрию. Например, работа 1902 г., в которой Дж. Адни Юл показал, что законы Гальтона и Менделя скорее дополняют, чем исключают друг друга (Yule, 1902), прошла незамеченной. Статья Роналда Фишера, объединяющая менделизм и биометрию, представленная в июне 1916г. в Лондонское Королевское общество (ее рецензентами были Пирсон и Реджиналд Пеннет, ученик Бэтсона) была отвергнута; она напечатана 1 октября 1918 г. Эдинбургским Королевским обществом: Fisher, 1918. Персональный конфликт усиливался разногласиями в вопросах методологии и роли математики (Cock, 1973). Искались социологические объяснения конфликта (MacKenzie, 1981). Подчеркивалась роль философских оснований: менделизм как теоретическая система нарушал требование «теоретической экономии», порожденное феноменализмом Пирсона, автора «Грамматики науки». Это значит, что даже и не будь горячих споров с Бэтсоном, Пирсон все равно не мог бы принять менделизм. Уэлдон сознательно развивал, под влиянием Пирсона, феноменалистическую философию науки; именно философские основания заставили его избегать традиционной дарвиновой терминологии: «полезный» или «адаптивный». Он выводил из теории Дарвина эмпирически верифицируемые следствия и говорил только о том, что можно измерить: «коэффициент смертности» (Norton, 1975). Вообще же их исследовательские программы не были соперничащими: биометрическая строилась на феноменологическом уровне, менделистическая — на физиологическом (Norton, 1975). Отсюда следует, что конфликт — если рассматривать его в предметном аспекте — был ложным.
Для интеграции менделевского наследования и дарвинова отбора и создания на этой основе генетики популяций потребовалась перемена действующих лиц: зашедший в тупик конфликт был снят новым поколением (Бабков, 19776).
Четвериков: натуралистическая традиция. Упоминая о попытках утвер-
Заметим, что защитникам дарвинизма не было нужды отвергать теорию Менделя как неподходящую для объяснения непрерывной эволюции. Так, в опытах самого Менделя со львиным зевом гибриды от родителей с белыми и пурпурными цветками давали в F2 растения целой серии цветов: от пурпурно-красного до бледно-фиолетового и белого, что Мендель объяснял комбинированием независимых факторов. 2* 19
дить приоритет того или иного из четырех сооснователей генетики популяций (Четвериков, Фишер, Райт, Холдейн), Ф. Г Добржанский подчеркнул, что приоритетные споры бесплодны: «Все четверо пришли к сходным идеям независимо, все четверо являются основоположниками, и каждому достанет славы» (Dobzhansky, 1967, р. 1). И хотя идеи всех основоположников были в итоге интегрированы в рамках единой теоретической конструкции, нам здесь важно подчеркнуть своеобразие путей мысли и различия двух биологических традиций.
Две противостоявшие группы имели ряд общих черт (привожу их по обзору: Мауг, 1973), которые интересно сопоставить со взглядами Четверикова, основателя русской традиции.
1. Биометрики и менделисты не проводили разграничения между индивидуальными и географическими вариациями. (Первые обычно менделируют, вторые — полигенные). В стенограмме лекции Четверикова по курсу энтомологии (21.IV. 1934, г. Владимир) читаем: «...Я подчеркиваю, что совершенно исключительное, первенствующее значение для понимания этих процессов (видообразования.— В. Б.) имеет так называемая географическая изменчивость...» Далее: «...мы рассматриваем географические формы как начатки будущих видов, как начало процесса видообразования, и в связи с этим важнейшим их значением мы в систематике даем этим географическим формам названия подвидов (subspecies)». Напротив: «...из мутаций непосредственно, сразу никакого вида не получается...» За примерами он обратился к хорошо ему известным бабочкам: географическая изменчивость описана на аполлонах Parnassius apollo из Московской губ. и с Кавказа, которые образуют подвиды Р.а. democratus и Р.а. suaneticus; мутационная —на переливнице Apatura ilia: ее нормальная форма и желтая мутация 5 (Эти взгляды Четверикова сложились в его лепидоптерологический период, начиная с 1895 г.).
2. Они смешивали также две формы изоляции: репродуктивную и географическую. Четвериков, определяя вид «как реальность, как совокупность свободно-плодущихся между собой особей»6, особо выделял половую среди возможных форм изоляции. Анализу эволюционной роли фактора изоляции он уделял внимание в гл. II статьи 1926 г. В частности, говоря об экологической изоляции, он четко поставил вопрос о возможности симпатри-ческого видообразования, хотя и не употреблял этот термин. (Замечу, что понятие биологического вида («видов-братьев») в зоологии, которое развивал в экспериментальных исследованиях Ф. Г Добржанский и впоследствии принял и пропагандировал Э. Майр, впервые сформулировал Н. А. Холод-ковский, 1910 и др.) Продолжая эту линию, А. Н. Промптов (19346, 1936) высказал мысль о первоначальном изменении поведения, о примате экологического обособления в дивергенции видов птиц.
3. Смешение понятий генотипической и фенотипической изменчивости приводило к недооценке роли полового воспроизведения и рекомбинаций в создании материала для эволюции.— Четвериков: «Признак и ген, его вызывающий,— это два понятия генетики, которые постоянно переплетаются, постоянно взаимодействуют, и тем самым постоянно ведут к недоразумениям». Разбирая четыре значения понятия «мутация» в биологии, он отме
5 Архив АН СССР, фонд 1650, опись 1, ед. хранения 11, листы 1, 2, 3. (Далее: Фонд Четверикова.)
6 Там же, ед. хр. 29, л. 1 об.
20
чает: «Существенно различие между вторым и третьим», т. е. между ^сальтациями» де Фриза и «геновариациями» Моргана. «...Если для эволюциониста на первом месте стоят «сальтации», как признаки, могущие сыграть ту или иную роль в процессе изменения организмов, то для генетика, конечно, главный интерес сосредоточивается на геновариациях...» 1. Этому вопросу посвящена гл. IV статьи 1926 г.
Натуралисты и экспериментаторы. Различия во взглядах между более описательными натуралистами и более аналитическими экспериментаторами, существовавшие в конце XIX в. в ряде областей биологии, получили особенно яркое выражение в изучении эволюции и наследственности. В начале XX в. раскол между натуралистами и экспериментаторами стал особенно заметен в связи с конфликтом «дарвинистов» (которые относились к лагерю натуралистов) и «менделистов» (экспериментаторов). «Изучение явлений эволюции и наследственности и создание их теории до поры до времени шли независимо и порознь, а на известном этапе юный и несколько самонадеянный менделизм и замкнувшийся в себе несколько самодовольный и покрывающийся жирком дарвинизм (или, точнее, дарвинисты и менделисты — своего рода отцы и дети) друг друга плохо понимали и в какой-то мере взаимно отрицали и даже враждовали между собой» (Астауров, 1972а, с. 550).
Не характеризуя подробно такие отношения 7 8, приведу лишь эпизод, который касается отношений менделистов-экспериментаторов и дарвинистов в Англии. Из переписки К. А. Тимирязева и У Т. Тислтон-Дайера. Тимирязев — Дайеру (5.III. 1912): «Как бы рад я был поболтать с Вами о современном положении дарвинизма. Я вижу сейчас некоторые тревожные признаки реакции даже в самой Англии. Они проявляются в растущей популярности Бэтсона, в волне увлечения Бергсоном и в прославлении Батлера...». Дайер — Тимирязеву (16.III.1912): «...У Бергсона, верно, одни пустые слова. Батлера нельзя принимать всерьез; мне представляется, что Бэтсон не обладает научным мышлением, он прыгает к выводам без должного исследования доказательств. Что касается Де Фриза, он кажется мне слегка не в своем уме...». Тимирязев — Дайлеру (без даты): «...Я рад услышать, что я был прав в оценке злюки Б[этсона] и Д [э] Ф [риза]...» 9.
Четвериков: отношение к математике. Обращение к существующим и разработка новых математических методов характерны для англо-американской генетики популяций. Такой подход привел к возникновению новой ветви прикладной математики — математической генетики. В становлении же русской генетики популяций математические методы играли лишь подчиненную роль. Это могло бы показаться странным. Действительно, Четвериков получил хорошее математическое образование; он читал курс биометрии в Московском университете, лекции по математике и статистике во Владимире и в Горьковском университете; а в горьковский период он сделал интересную оригинальную работу по биометрике «Опыт построения объективной систематики организмов (на примере Lepidoptera—Tortricidae)». Однако после ее обсуждения (на 68-м заседании Эволюционной бригады
7 Фонд Четверикова, on. 1, ед. хр. 5, лл. 2 об., 3 об.
8 Подробный анализ ситуации дан в интересной и содержательной работе: Alien, 1979.
9 Переписка К. А. Тимирязева (на англ, языке). Архив хранится в музее-квартире К. А. Тимирязева. Я благодарен Е. М. Сенченковой за предоставленную мне копию этой переписки.
21
ИЭБ, 21.Х.38, где присутствовало 44 человека) 10 один из участников — ленинградский энтомолог А. С. Данилевский возразил в частной беседе, чтр подобный результат будто бы уже получил Роналд Фишер (имея в виду работу: Fisher, 1936), о чем Четвериков, не следивший за новой литературой по биометрике, судить, конечно, не мог. Насколько справедливо было это возражение, судить мы не можем, так как текста найти не удалось. Четвериков же, разочаровавшись, доклад не опубликовал и работу эту прекратил. Лишь в послевоенное время он написал краткое сообщение на эту тему, но не опубликовал его; нет его и в фонде Четверикова в Архиве АН СССР. Основную направленность его рассуждений по впечатлениям от доклада 1938 г. изложил впоследствии А. А. Малиновский (1970).
В статье 1926 г. математическим моментам Четвериков уделяет сравнительно немного места. Назвав в гл. II ряд авторов, занимавшихся математическим анализом последствий менделевского наследования (27 статей!), Четвериков подчеркнул, что «из этого длинного списка работ для правильного понимания роли генотипической изменчивости в процессе эволюции имеют значение лишь те, которые исследуют результаты скрещивания и судьбу отдельных признаков в условиях свободного скрещивания* (Четвериков, 1926, с. 14), и обратился лишь к двум исследованиям — Г Харди («закон равновесия при свободном скрещивании», или «закон Харди»: Hardy, 1908) и Пирсона («закон стабилизирующего скрещивания»: Pearson, 1904), тому, в котором Пирсон очень близко, и в единственный раз, подошел к представлению о совместимости менделизма и дарвинизма. Четвериков дает ясное биологическое истолкование этих двух математических результатов. (По сути дела Четвериков излагал здесь «основной закон популяции» — собственную работу, выполненную около 1922 г., в которой он независимо пришел к тем же выводам, что и Харди и Пирсон.) Разобрав эти две аналитические работы и проведя один несложный расчет, Четвериков в гл. III, посвященной роли отбора в судьбе мутаций, счел нужным привести таблицу Нортона, где для различных интенсивностей отбора дается подсчет числа поколений, необходимых для перехода популяции из одного состояния хардиева равновесия в другое. Он легко мог воспользоваться аналитическим подходом, дающим результаты высокой общности, хотя и в абстрактной форме; однако ради доступности обратился к иллюстрации, которую разбирал с целью получить биологически значимые выводы.
В статье 1926 г. Четвериков останавливается только на тех математических работах, которые там решительно необходимы и которые можно изложить обычным языком, что он и делает; общности он предпочитает наглядность, но не за счет строгости. Главное — математическая культура, а не математическая техника.
Эти же умонастроения иллюстрируют его лекции по биометрии в Учком-бове (Учебном комбинате (техникуме) по борьбе с сельскохозяйственными и лесными вредителями) во Владимире. В конспекте VII лекции (26.1.1933) «Сложные кривые и их эмпирический анализ» он приводит ряд примеров «естественных случаев небиномиальных распределений» и подчеркивает на этом материале различия двух подходов: «Две школы, изучающие эту проблему: 1. Школа формально-математическая (К. Пирсон), изучающая
10 «Протоколы Эволюционной бригады», тетр. 2.— Материалы Комиссии по истории отечественной генетики и селекции МО ВОГиС им. Н. И. Вавилова. (Далее: Материалы Комиссии.)
22
^кривые распределения с точки зрения математической закономерности их. 12 типов кривых Пирсона. Чисто формально-описательное значение этих кривых, не дающих понимания сущности самого явления, а лишь скользящих по поверхности их. Случай с кривой распределения крабов (Вель-дон) 11 и ошибочность выводов Пирсона, полученных путем чисто математического анализа ее. Кривые распределения Пирсона ничего не дают биологу. 2. Школа биологическая (Иогансен). Необходимость широкого биологического анализа причин, обусловливающих отклонения от биномиального распределения. «Биологические задачи должны решаться при помощи математики, но не как математические задачи» (Иогансен — В. Б.). Биометрический анализ лишь описывает явление, но бессилен объяснить его сущность. Но, с другой стороны, биометрический метод вскрывает проблемы и направляет биологический анализ там, где биолог без него был бы совершенно бессилен» 11 12 Те же мысли завершают его статью «Биометрия» в 1-м издании БСЭ (Четвериков, 1927).
По-видимому, следующие слова помогут понять, почему Четвериков избегал ранней математизации новой области знания. «Вступая на ложный путь с математикой в качестве проводника, вы лишь скорее попадете пальцем в небо, причем это произойдет так легко и просто, что вы даже не заметите, и вас будет трудно вывести из заблуждения. Логика и математика полезны только после того, как найден правильный путь. Чтобы найти его, следует начать не с логического мышления, а использовать совсем другие способности, а именно наблюдательность, фантазию и воображение; вернее точное наблюдение, буйную фантазию и острое воображение». Эти слова принадлежат не натуралисту, как можно было бы подумать, а математику — и блестящему! — Дж. Адни Юлу (Yule, 1920, р. 184).
Взгляд на развитие науки как равномерное накопление отдельных идей предполагал интерес к поиску предшественников. Однако в случае Четверикова такие поиски будут неблагодарными. Натуралист, он ввел в арсенал своих средств экспериментальный метод 13 Генетик, он не мог принять взглядов Бэтсона на природу гена 14 или редукционистской методологии школы Моргана 15 Биометрик, он критиковал формальный подход Пирсона и держался взглядов, несовместимых с его философией (Babkoff, 1984; Бабков, 1982).
СООР
Сотрудниками Генетической лаборатории Четверикова были его ученики, молодые зоологи: Борис Львович Астауров (1904—1974), Елизавета Ивановна Балкашина (1899—1981), Николай Константинович Беляев (1899—1937), Сергей Михайлович Гершензон (р. 1906), Александр Николаевич Промптов (1898—1948), Петр Фомич Рокицкий (1903—1977), Дмитрий Дмитриевич
11 См.: Provine, 1971; Norton, 1973.
12 Фонд Четверикова, on. 1, ед. хр. 10, лл. 8-а, 8-а об.
13 Ср.: Allen, 1979.
14 Четвериков, в частности, критиковал теорию «присутствия—отсутствия» Бэтсона, например в наброске статьи «Значение мутаций в эволюционном процессе» (Фонд Четверикова, on. 1, ед. хр. 4, лл. 4, 4 об., 5, 5 об.). О взглядах Бэтсона подробнее см.: Coleman (1970).
15 Ср.: Roll-Hansen, 1978.
23
Ромашов (1899—1963), Елена Александровна Тимофеева-Ресовская, рожд. Фидлер (1898—1973), Николай Владимирович Тимофеев-Ресовский (1900— 1981), Сергей Романович Царапкин (1892—1960). Никто из них не предполагал поначалу заниматься генетикой — все они были зоологами: Бал-кашина выполнила гидробиологическую работу и доложила ее в 1922 г. на I Съезде зоологов, Ромашов был колеоптерологом, Беляев — лепидопте-рологом, Фидлер участвовала в работах М. М. Завадовского в зоопарке Аскания-Нова, возвратившийся из Красной Армии Тимофеев-Ресовский приступал к изучению географической изменчивости гольяна (Phoxinus). Для этого времени весьма характерно, что будучи студентами университета они начинали вести исследовательскую работу и одновременно должны были зарабатывать на жизнь.
В декабре 1921 г. с выходом в свет выпуска «Известий ИЭБ» Н. К. Кольцов обратился к ряду европейских коллег с предложением возобновить обмен публикациями. Уже с начала 1923 г. Институт экспериментальной биологии регулярно получал все основные биологические журналы. В результате перед группой Четверикова встала задача: критически изучить зарубежные работы по эволюции, цитологии, генетике (особенно по генетике дрозофилы), недоступные в период мировой и гражданской войн. Для этого не подходило ни традиционное преподавание (когда предмет не подлежит обсуждению), ни обычные конференции (с их слишком частными темами и разбросом интересов участников). Но как соединить и научность, и систематичность обсуждаемой тематики, сохраняя при этом все положительные стороны непринужденной беседы? Так у Четверикова зародилась мысль о новой форме научных собраний — Соорах. В своих «Воспоминаниях» Четвериков уделил вопросам организации научного общения несколько очень интересных страниц.
«Во-первых, необходимо, чтобы заранее была намечена тематика предстоящей беседы... Во-вторых, нужно, чтобы обсуждение было живым и творческим, а не бездушно-казенным, чтобы оно... выливалось бы в форму свободного собеседования, где каждый может выступить в любой момент, как только ему в голову придет та или иная мысль, подлежащая обсуждению. ...вместо доклада получается как бы свободное собеседование, где люди перебивают друг друга, вставляют свои замечания и начинают горячо спорить, так что дело доходит по временам до настоящего ора (вот отсюда-то, от совместного ора и получилось наименование «Соор»). Третье и, пожалуй, главное — это то, чтобы на эти Сооры собирались люди, тесно связанные между собой по тематике своей работы... Необходимо, чтобы руководитель был вполне в курсе затрагиваемых вопросов и мог направлять дискуссию в нужную сторону. Таковы были предпосылки, которые легли в основу той организации при Лаборатории генетики Института экспериментальной биологии (директор — проф. Н. К. Кольцов), которая была организована мною в 1924 г. и получила название «Генетического Соора»» (Четвериков, 1974, с. 68—69). Главной темой Сооров была генетика дрозофилы, но обсуждались также эволюционные, цитологические и другие работы. Е. И. Балкашина упоминала (личн. сообщ., 1973), что еще до Сооров на Звенигородской биостанции шли биологические диспуты. Летом 1921 г. они превратились в более систематические научные дискуссии в связи с обсуждением полученной тогда Н. К. Кольцовым книги Моргана, Стертеванта, Мёллера и Бриджеса «Механизмы менделевской наследственности» (Morgan et al., 1915), которое сыграло известную роль в становлении
24
нашей генетики. Четвериков же приехал в Звенигород лишь в 1922 г., когда осушили Луцинское болото и появилась возможность собирать бабочек.
На расположенную неподалеку Аниковскую генетическую станцию, которая так же, как и Звенигородская гидрофизиологическая станция, являлась как бы филиалом Института экспериментальной биологии, приехал 19.VIII.1922 г. американский генетик Мёллер и привез около 20 культур мух Drosophila tnelanogaster с набором мутаций, полученных в лаборатории Т. Моргана. «В то время в СССР еще очень мало поступало научной литературы из-за рубежа, и доклад Мёллера о работах генетиков школы Моргана произвел огромное впечатление»,— вспоминала Балкашина 50 лет спустя (Балкашина, 1972, с. 5—6, цит. по: Бабков, 1983а, с. 7). Такие обсуждения привели к «предсоорам», с участием Тимофеева-Ресовского, Беляева, Четверикова, Балкашиной, Ромашова, Астаурова, которые подготовили организацию Сооров и наметили их тематику.
В Соор, кроме Четверикова, его жены Анны Ивановны (которая заведовала библиотекой ИЭБ), 10 его учеников, входили и некоторые другие сотрудники Института, в первую очередь, конечно, Н. К. Кольцов, а также Софья Леонидовна Фролова, Петр Иванович Живаго, Александр Сергеевич Серебровский, Владимир Владимирович Сахаров. «Прием в Соор новых членов был ограничен одним очень суровым условием: каждый кандидат подвергался баллотировке, причем достаточно было одного голоса против, чтобы считать кандидата непринятым. Это суровое требование имело двойную цель: во-первых, не допускать чрезмерного увеличения числа членов Соора; во-вторых, не создавать в Сооре неприязненных отношений между его членами и обособленных группировок. Все члены Соора были вполне дружны между собой, и я не помню ни одного случая хотя бы самой незначительной ссоры». «Основным существеннейшим правилом Соора было право любого его члена останавливать и перебивать докладчика своими замечаниями, вопросами или возражениями Все члены Соора должны были читать на трех основных европейских языках, и с этой стороны не принимались никакие возражения... Доклад должен был быть критическим разбором данной работы. От каждого доклада я требовал, чтобы докладчик выделил и четко проводил так называемую красную ниточку работы, т. е. ту основную мысль или мысли, которые составляли главный смысл и ценность обсуждаемого исследования...». (Четвериков, 1974, с. 69).
О Сооре вспоминала Е. И. Балкашина: «...Вскоре после основания генетического отдела в Институт экспериментальной биологии начинают поступать многочисленные журналы по генетике, отечественные и из-за рубежа. Надо было как-то успевать осваивать эту литературу, а знанием английского языка обладали немногие, поэтому был организован кружок по изучению генетической литературы. Тем, кто знал английский язык, поручалось реферировать большие работы на английском языке, а тем, кто не знал английского — русские работы или короткие английские статьи. Они на ходу изучали английский язык. Нередко в кружке возникали очень горячие споры на различные генетические темы... Сергей Сергеевич Четвериков обращал большое внимание на форму докладов и требовал ясного, четкого изложения... За эту требовательность к докладчикам бывшие члены Соора всегда вспоминают С. С. Четверикова с благодарностью. Генетическая лаборатория была очень дружным коллективом. Работали с утра до позднего вечера...» (Балкашина, 1972, с. 7, цит. по: Бабков, 1983а, с. 16—17).
Преемственность форм организации научного общения заслуживает
25
В Генетической лаборатории ИЭБ, ок. 1926 г., С. М. Гершензон, Д. Д. Ромашов, С. С. Четвериков, Б. Л. Астауров, А. Н. Промптов, А. И. Четверикова, Е. И. Балкашина, Н. К. Беляев
не меньшего внимания, чем преемственность идей и методов исследования, особенно когда речь идет о создании новой области знания и становления особой системы познавательных ориентиров. У многих членов Соора были предшествующие опыты подобного рода: у Кольцова, например, биологические дискуссии в Виллафранке; у Четверикова — астрономический кружок в училище, зоологический в университете; у Ромашова — сперва детский кружок В. П. Вахтерова, где он подружился с А. Н. Колмогоровым и С. М. Мусатовым, затем организованный Е. С. Смирновым остродискуссионный «Кружок имени Ламарка» в университете; у Тимофеева-Ресовского — гимназический кружок «Секамбр», где обсуждались вопросы истории культуры и новости естествознания, в том числе новой физики. Реферативный кружок Соор просуществовал пять лет; в 1929—1931 гг. Генетическая лаборатория распалась: Беляев и Астауров уехали в Среднеазиатский НИИ шелководства; Балкашина и Ромашов работали в отделе теоретической генетики Всесоюзного института животноводства (куда вошла Ани-ковская станция), Рокицкий — в другом отделе ВИЖ и в МГУ; Тимофеевы-Ресовские и Царапкин были командированы в Институт мозга в Берлине; Промптов работал на кафедре общей биологии 2-го Московского медицинского института и в Институте физиологии И. П. Павлова (в Колтушах) ; Гершензон перешел в лабораторию генетики А. С. Серебровского (в Биологическом институте им. К. А. Тимирязева Комакадемии). С. С. Четвериков в 1929—1932 гг. был в Свердловске и работал там консультантом Зоосада, в 1932—1935 гг.— во Владимире, где читал лекции по энтомологии и биометрии в Учебном комбинате по борьбе с сельскохозяйственными и лесными вредителями, а с 1935 г. он заведовал кафедрой генетики в Горьковском университете (о горьковском периоде Четверикова см.: Пузанов, 1965). Но традиции Соора не прервались. Весной 1932 г. Балкашина и Ромашов возвратились в ИЭБ, в заново организованный Генетический отдел, который возглавил тогда (по предложению Н. К. Кольцова) Н. П. Дубинин; осенью 1932 г. Ромашов организовал Эволюционный семинар, а осенью 1934 г.— 26
Эволюционную бригаду (лабораторию) с Дарвиновским семинаром, который объединял эволюционистов не только Москвы, но, в известной мере, и всей страны. В то же время Тимофеев-Ресовский с целью развивать (используя весь арсенал современных средств исследования) представления Н. К. Кольцова о «наследственных молекулах» (Кольцов, 1936) организовал семинар для обсуждения проблем биофизики, в котором участвовали К. Циммер, М. Дельбрюк Г Борн, Р. Ромпе, Н. Риль, П. Йордан и где были сформулированы принцип мишени и принцип попадания — фундамент радиобиологии. Говоря о роли замечательной книги Э. Шредингера «Что такое жизнь?» для распространения идей, легших в основу молекулярной биологии, Уоддингтон отметил: «Та часть «Что такое жизнь?», которой она обязана своим влиянием, была, по сути дела, изложением классической статьи, которую мы называли TZD (Timofeeff-Ressovsky, Zimmer, Delbruck, 1935), где мутагенез, вызванный Х-лучами, обсуждался в контексте «генной молекулы», устойчивость которой выводилась из квантово-механических соображений. Эта статья явилась первым публичным изложением мысли, что генетический материал может рассматриваться в квантово-механических терминах, и именно эта мысль, в изложении Шредингера, разожгла воображение ранних молекулярных биологов в первые же послевоенные годы» (Waddington, 1969, р. 321). Позже, в 1956—1963 гг. Тимофеев-Ресовский организовал ежегодный летний коллоквиум Лаборатории биофизики Уральского филиала АН СССР на биостанции Миассово (на территории Ильменского заповедника) ; там обсуждались доклады на общие естественно-научные темы, по биогеоценологии, радиоэкологии, биокибернетике, проблемам эволюции (Чтения... 1983).
ЦЕНТРАЛЬНАЯ ПРОБЛЕМА ЭВОЛЮЦИОННОЙ ГЕНЕТИКИ
Основой четвериковской программы построения эволюционной генетики была формулировка ее центральной проблемы, в сжатом виде: природа и поддержание изменчивости в популяциях. Четвериков разбирал эту проблему по частям; он начал с вопроса, бывшего постоянным источником споров.
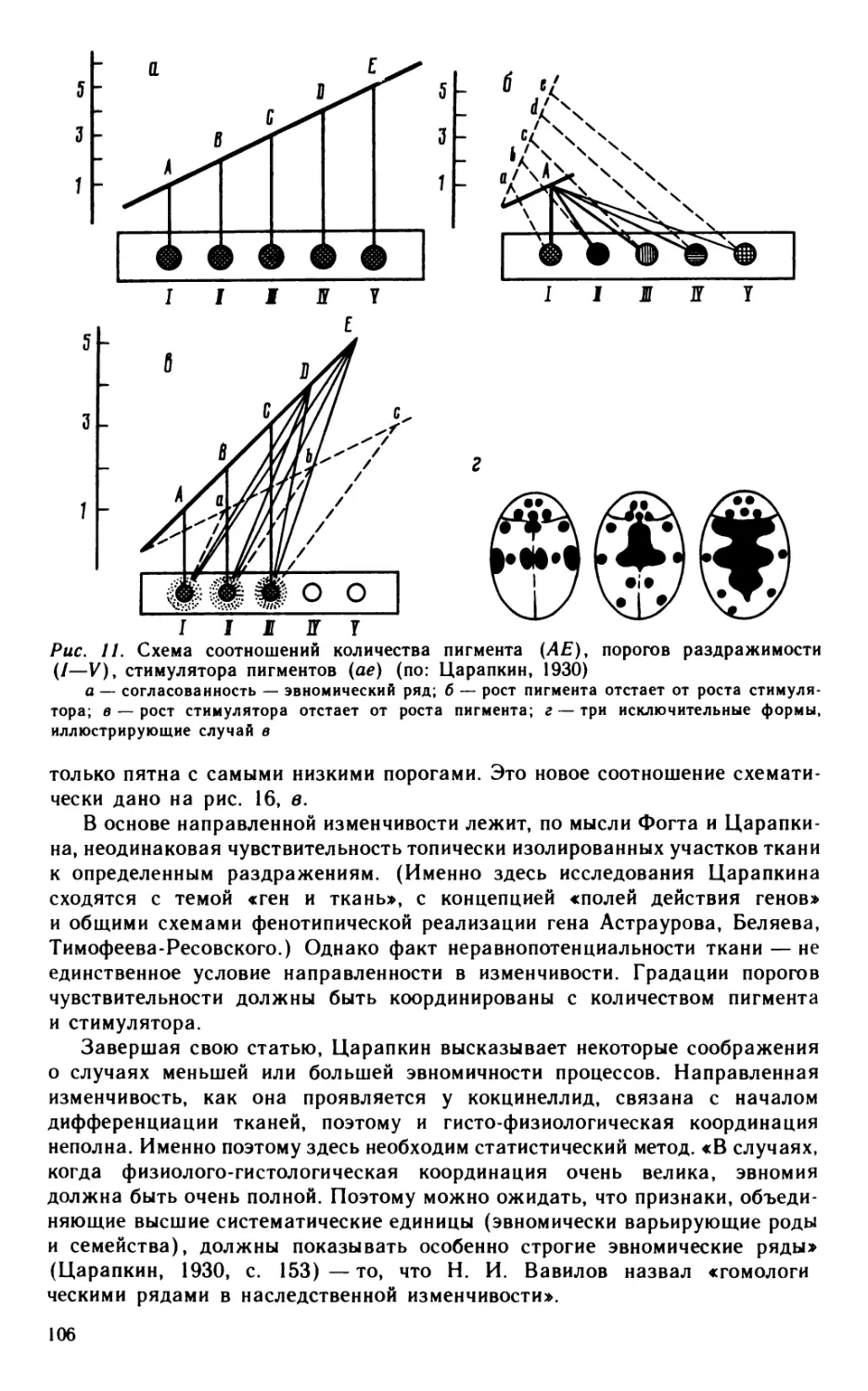
1. Природа наследственной изменчивости. Исключительный для Четверикова факт — введение нового термина «геновариация» — указывает на чрезвычайное значение, которое он придавал концептуальной ясности в этом вопросе 16 В статье 1926 г. он поясняет термин в примечании на 32 строки петита. Его записи указывают на интерес к проблеме (в связи с работами де Фриза): «Главная литература по Oenothera» 17 18, 13 листов из 16 в тетради «Выписки и записи к лекциям» ,8посвящены проблеме энотеры. В наброске к статье «Значение мутаций в эволюционном процессе» читаем: «...Четыре значения понятия «мутация» в биологии:
1) мутации палеонтологов (Вааген, 1869),
2) сальтации (мутации де-Фриза, 1901),
16 Й. Рихтеру я обязан замечанием, что слово «геновариация» Четвериков мог заимствовать из статьи С. и О. Фогтов, посвященной анализу заболеваний большого мозга (С. Vogt, О. Vogt, 1922). Современное понятие «мутация» совпадает по объему с «геновариацией» Четверикова; в геологии и палеонтологии смены форм одного вида животных в последовательных слоях разного возраста — «мутации палеонтологов» — именуются сейчас «мутациями Ваагена».
17 Фонд Четверикова, on. 1, ед. хр. 3.
18 Там же, ед. хр. 37
3) геновариации (мутации Моргана, 1910),
4) геномутации [(Стертевант, 1925)] или трансгенации (Мёллер, 1921)» 19
Четвериков ставит целью доказать, что на мутационной изменчивости как поставщике.материала основан магистральный «извечный путь эволюции, которым шел и развивался органический мир от своего появления на земле и до наших дней» (Четвериков, 1926, с. 11). Для этого нужно опровергнуть мнение, что все мутации суть лабораторные артефакты, которые в природе не возникают. (Решающий аргумент в этом ему дал первый экспериментальный анализ генофондов диких популяций дрозофил 1925— 1926 гг.) Выступая против взгляда, что все мутации «уродцы», не могущие иметь значения для эволюции, он замечает, что некоторые из биологически нейтральных мутаций иногда соответствуют нормальным признакам соседних видов или даже родов и семейств. Так, у Drosophila melanogaster потемнение тела (мутация black) соответствует нормальному видовому признаку D. fu-nebris\ загнутые концы крыльев (depressed, curved) характерны для рода Stegnata (сем. Drosophilidae); отсутствие поперечной жилки (crossveinless)— для семейства Asteidae и т. д. Сюда можно добавить изучавшиеся Астауровым и Балкашиной гомеозисные мутации tetraptera (замещает жужжальца 2-й парой крыльев) и aristopedia (замещает антенну усика лапкой), которые «выбивали» измененных мух из таких высоких таксономических категорий, как отряд (Diptera) и класс (Hexapoda). Собственные материалы Четверикова (таблица в гл. V статьи 1926 г.— см.: Четвериков, 1982) позволяют заключить, что мутационной изменчивости «подвергаются все решительно части организма» (Четвериков, 1926, с. 11).
Рядом с несущественными признаками у дрозофилы мутационно изменяются и такие, которые в современной систематике насекомых являются основными при различении высших систематических категорий. «Поэтому приходится признать совершенно ошибочной высказываемую некоторыми мысль, что геновариационная изменчивость как бы скользит по поверхности видовых признаков, являясь характерной формой изменчивости разновидностей, но что помимо этих мелких отклонений существует «некоторая сущность» организмов, не могущая изменяться геновариационно»20 (там же).
В этой фразе звучит проблема отношений микро- и макроэволюции. В наши дни она решается на разных путях. Одни авторы подчеркивают, что в основе филогенетических процессов любого ранга (т. е. макроэволюции) лежат в конечном счете те же самые стартовые механизмы, которые являются предметом учения о микроэволюции. Другие подчеркивают специфичность процессов макроэволюции, значит невозможность их адекватного описания с позиций микроэволюции. Речь идет о сведении (редукции) макро- к микроэволюции; на первый взгляд можно решить, что эти ответы взаимоисключающи.
Между тем видимость противоречия рассеется, и мы избежим недоразумений, если будем при обсуждении редукционизма в биологии различать три самостоятельные области: онтологическую, методологическую, эпистемологическую (ср.: Ayala, 1974). Большинство биологов, кроме виталистов, редукционисты — в онтологическом
19 Там же, ед. хр. 5, л. 2 об.
20 Именно на таком основании Ю. А. Филипченко ввел в 1927 и 1929 гг. понятия «микроэволюция» и «макроэволюция». Подробнее см.: Н. Н. Медведев (1978, с. 29—32 и 82—86).
28
смысле. Онтологический редукционизм подразумевает, например, что законы физики и химии полностью приложимы ко всем биологическим процессам на уровне атомов и молекул, что микроэволюционные явления и процессы лежат в основе всех макро-эволюционных (филогенетических) процессов и т. п. Методологическая область включает вопросы стратегии исследования. Занимаясь явлениями жизни, следует ли всегда искать объяснения в изучении более низких уровней сложности? или должно искать понимания в изучении как высших, так и низших уровней организации? В изучении наследственности, например, крайний антиредукционист будет утверждать, что изучение структуры ДНК не имеет биологического смысла', а крайний редукционист — что менделевская генетика не имела научного значения до выяснения лежащих в ее основе молекулярных процессов. С позиции эпистемологии главный вопрос редукции заключается в том, можно ли доказать, что теории и эмпирические законы, сформулированные в одной области знания,— суть частные случаи теорий и законов другой его области. Э. Нагель (Nagel, 1961) сформулировал два необходимых и достаточных условия (критерий) редуцируемости одной ветви науки к другой: условие выводимости, derivability (следует показать, что все эмпирические законы и теории редуцируемой дисциплины суть логические следствия теоретических конструкций редуцирующей дисциплины) и условие связности, connectability (все термины редуцируемой дисциплины должны быть переопределены посредством терминов редуцирующей). В биологии многие элементы теории наследственности были редуцированы к химии после выяснения структуры и поведения определенных молекул: ДНК, РНК, ряда ферментов. Однако очевидно, что при современном состоянии науки большинство биологических концептов — таких, как клетка, менделевская популяция, вид и т. п.— не может быть адекватно определено в физико-химических терминах, а, скажем понятия макроэволюции — в терминах микроэволюции, и т. п. Очевидно, что сейчас нет такого класса утверждений, принадлежащих физике и химии, на основе которого можно было бы вывести любой биологический закон,— как и из системы элементарных понятий микроэволюции мы не можем вывести законы филогенеза.
Возвращаясь к проблеме соотношений микро- и макроэволюции, мы видим, что первый из приведенных выше ответов (данный здесь Четвериковым, впоследствии подчеркиваемый Тимофеевым-Ресовским) говорит о законности онтологической редукции. Второй (популярный среди некоторых палеонтологов) — о неправомерности крайностей методологической и эпистемологической. Мнимое противоречие исчезло; совместимость этих двух позиций не вызывает сомнения (Бабков, 1984).
2. Популяция как единица эволюции. Мутационный процесс — условие эволюции, но «из мутаций непосредственно, сразу, никакого вида не получается...*. В качестве элементарной эволюирующей структуры Четвериков рассматривает не организм, а популяцию. Он привлекает внимание к географической изменчивости, в частности к описанию географических рас А. П. Семеновым-Тян-Шанским (1910), и подчеркивает ее значение для понимания процесса видообразования. Он описывает разные формы изоляции (ограничения свободы скрещивания) и оценивает их возможное значение. В этом видно влияние натуралистической традиции. Но если для старых натуралистов вид определялся как «некоторая сущность» организмов (на практике— его морфологическими отличиями), то для Четверикова-генетика в основе этого определения лежит: «Физиологическая — генетическая изоляция. Вид как реальность, как совокупность свободно-пло-дущихся между собой особей».21 Разобрав «основные закономерности популяции» (закон Харди и Пирсона), Четвериков указал, что условия Харди, определяющие генетическое равновесие и эволюционную инерцию популя-
21 Фонд Четверикова, on. 1, ед. хр. 29, л. 1 об.
29
ции, дают нулевой уровень, точку отсчета для исследователя популяционной динамики.
3. Поддержание изменчивости в популяциях. Анализ динамики генофонда популяции Четвериков начал с выяснения роли мутационного процесса. Здесь важен вопрос о судьбе единичных мутаций: действительно ли, как утверждал Дженкин в 1867 г., им суждено раствориться в море нормальных особей, никак не влияя на дальнейшую судьбу вида, на процесс эволюции? Опираясь на закон Харди, Четвериков заключает, что судьба мутации будет «совершенно иная, чем это мыслилось прежним эволюционистам. Геновариация не погибнет, не растворится в массе нормальных особей. Она будет существовать в гетерозиготном состоянии, из поколения в поколение оставаясь скрытой от глаз, но в виде определенного наследственного генотипа» (Четвериков, 1926, с. 20). На этом-то и была основана задача «экспериментального исследования геновариаций, скрытых в популяциях» (так называется гл. V, добавленная Четвериковым к статье 1926 г. в начале 1929 г., когда он готовил английский ее перевод), выявления «резерва наследственной изменчивости, из которого отбор может свободно черпать всегда, как только возникнет необходимость» (Четвериков, 1982, с. 348).
Ф. Г Добржанский (Dobzhansky, 1967, р. 2) отметил, что Четвериков первым опроверг возражение Дженкина. Правда, еще в 1910 г. К. А. Тимирязев, пламенный проповедник дарвинизма, приветствовал идею менделев-ского наследования, благодаря корпускулярности которого «кошмар Дженкина, испортивший столько крови Дарвину, рассеивается без следа» (Тимирязев, 1910, с. 33—34). Однако вскоре разгорелись споры, которые развели в два лагеря менделистов и дарвинистов. Тимирязев утратил интерес к взглядам оппонентов теории отбора, и его опровержение Дженкина не открыло путей для объединения принципов менделевского наследования и дарвинова отбора. Также и У Бэтсон, еще до повторного открытия менделизма, утверждал, что обращением к корпускулярному наследованию можно устранить главные трудности на пути дарвинизма, в том числе «заболачивающее» («swamping») влияние скрещивания. Однако эти мысли Бэтсона потонули в его спорах с биометриками. Были и другие опровержения Дженкина — из них также ничего не воспоследовало; это дает нам право утверждать, что «кошмар Дженкина» развеял именно Четвериков.
Действие отбора, нарушающего эволюционную инерцию хардиевой популяции, Четвериков рассматривал на основе таблицы Г. Нортона. Английский математик Н. Т. J. Norton (в статье 1926 г. ошибочно: G. Norton) составил эту таблицу по просьбе Р. Пеннета для его книги «Мимикрия бабочек» (Punnett, 1915, р. 155), которая была написана для доказательства, что мимикрия — эволюционное явление, происходящее посредством отдельных скачков. Из разбора таблицы Четвериков заключает следующее: 1. Любое незначительное улучшение в организме имеет реальный шанс распространиться на всю популяцию. (По иронии судьбы, этот вывод легко истолковать в том смысле, что он подрывает идею эволюции путем крупных дискретных изменений, для доказательства которой и была заказана таблица. Однако так только на первый взгляд, а в принципе он не противоречит эволюции посредством отдельных скачков, он лишь говорит о том. что этот путь не исключительный.) 2. Адаптивный эволюционный процесс, «трансмутация вида», идет до конца, и в отсутствие изоляции данный вид не расщепится на два. 3. Отбор способствует накоплению в популяции 30
рецессивных генов, «из которых очень значительная доля может быть связана с неблагоприятными для организма признаками» (Четвериков, 1926, с. 40) — то, что сейчас называют «грузом мутаций».
Наконец, рассматривая роль свободы скрещивания (и его ограничения — изоляции) и естественного отбора в создании «резерва наследственной изменчивости», Четвериков ставит проблему соотношения факторов микроэволюции.
4. Проблема неадаптивной эволюции. Изоляция интересует Четверикова в различных отношениях. Касаясь темы «популяция как первичная эво-люирующая структура», Четвериков подчеркивает, что частичная изоляция внутри популяции, т. е. структурированность популяции, оказывает глубокое влияние на ход ее эволюции. В качестве примера он рассматривает такую ситуацию: обширная популяция разбилась на ряд полуизолированных колоний. Тогда мутации в системе колоний возникают так же часто, как и в большой популяции, а вероятность их выявления в гомозиготе так же высока, как и в малой популяции. Так Четвериков ставит проблему оптимальной организации популяции (Бабков, 1972в, 1975).
Другая тема — «поддержание изменчивости»: факторы эволюции, модусы эволюционных преобразований, наконец, видообразование. По Четверикову, приспособленность не является ключом к пониманию всех проблем эволюции. Виды часто различаются биологически нейтральными признаками, «и стараться подыскивать им всем адаптивное значение является столь же мало производительной, как и неблагодарной работой, где подчас не знаешь, чему больше удивляться, бесконечному ли остроумию самих авторов или их вере в неограниченную наивность читателей» (Четвериков, 1926, с. 41). Хотя можно установить адаптивные различия между видами, родами и т. п., «надо быть очень осторожным в допущении за этими различиями первичного характера» (Там же, с. 42). Распадение первоначальной формы на две имеет неадаптивный характер, в ее основе лежит изоляция. Впоследствии может установиться новый видовой признак адаптивного характера, который явится следствием межвидовой дифференциации.
Близкие взгляды высказывали и другие авторы. «Адаптация — иная проблема, чем происхождение видов» (Elton, 1930, р. 75). «Основной эволюционный механизм в происхождении видов должен, таким образом, быть по существу неадаптивным» (Wright, 1932, р. 364). ’’«Естественный отбор или переживание благоприятствуемых рас в борьбе за жизнь» служит объяснением прежде всего адаптации и лишь вторично — происхождения видов” (Watson, 1936, р. 43). Подробно обсуждал этот вопрос Д. Н. Каш-каров (1939).
Оценивая через 10 лет свою работу в лекции «Генетические закономерности в эволюционном процессе» 22, Четвериков подчеркивал различие между двумя процессами и их результатами, между двумя факторами эволюции: два процесса эволюции: а) адаптивная (отбор), б) дифференциальная — видообразующая (изоляция). Генетическое объяснение двух процессов эволюции он дал в работе 1926 г., однако в лекции 1936 г. он отметил: «Несовершенство анализа, его фрагментарность привели к тому, что в стороне остался ряд «неправильностей». В работе 1926 г. Четвериков (как и Харди — Hardy, 1908) отвлекался от популяционной стохастики. Его замечания,
22 Фонд Четверикова, on. 1, ед. хр. 27, лл. 1—2.
31
что рассматриваемые процессы в природе «осложняются рядом неправильностей» (Четвериков, 1926, с. 52) и что вероятность проявления мутаций зависит также «от некоторой неустойчивости наших статистических средних» (там же, с. 27), не привели тогда к теории дрейфа генов. В упомянутой лекции 1936 г. Четвериков говорил, что вопрос об этих «неправильностях» проанализировали «его ученики и «внуки». Ромашов, Дубинин — БДдети), 3. С. Никоро — F2 (внучка)». Рассматривая там же роль изоляции, Четвериков отмечал значение генетико-автоматических процессов для понимания дифференциации изолированных групп: «Процесс видообразовательной эволюции в свете генетико-автоматических процессов — неадаптивная эволюция». В другой лекции «Возможно ли искусственное получение новых видов» 23 он считал важным различать адаптивные и неадаптивные процессы в эволюции: «Эволюционный процесс — не единообразный, простой, а сложный, многогранный процесс. Адаптивная и неадаптивная эволюция. А. э.— эволюция приспособления — естественный отбор. Н. э.— процесс видообразования, процесс разнообразия органического мира...».
В лекции «Отдаленная гибридизация» (1947), привлекая внимание к вопросу об изменении кариотипа в процессе видообразования, Четвериков делает вывод: «...как мы видели, эволюционный процесс может идти, по крайней мере, четырьмя различными путями: при помощи отбора — адаптивная эволюция; в результате изоляции—дифференциальная эволюция; в результате отдаленной гибридизации (амфидиплоидий) и вследствие хромосомальных аберраций (аутополиплоидии)»24.
Отступление о критике. Выше мы коснулись опровержения Четвериковым критики Дженкина. Именно ее так болезненно воспринял в свое время Дарвин, хотя аналогичные аргументы против теории отбора, по свидетельству Эллегора, выставлялись в британской прессе до Дженкина не менее семи раз (Ellegard, 1958, р. 244). В России среди опровержений дарвинизма выделялась книга Н. Я. Данилевского (1885). В главах VIII—IX «Невозможность естественного подбора по внутренней и существенной несостоятельности этого начала» (т. I, ч. II, с. 1 —135) Данилевский формулирует (с. 2—3) и развивает три положения: 1) «...свободное скрещивание должно постоянно уничтожать все индивидуальные изменения (служащие точкою отправления для всего процесса) при их возникновении, и не допускать накопления их...»; 2) «...предлагаемым им путем не могут произойти бесполезные и безразличные признаки, которые так многочисленны у животных, а у растений решительно преобладают и притом составляют самые крупные, важные, существенные черты строения; ...»; 3) «...бесчисленные переходные формы, едва заметными оттенками переливающиеся одна в другую, и которые должны составлять остатки, следы, так сказать шлаки, того процесса, которым вырабатываются определенные органические формы в лаборатории природы,— в ней, однако же, совершенно отсутствуют...» Данилевский подчеркивает, что Дарвин «изо всех этих трех капитальных возражений обращает внимание только на последнее, и как-нибудь с ним справляется; два же первых оставляет без должного внимания». Четвериковг же, как мы видели, дал генетическую трактовку каждой из трех ситуаций и включил их в свою концепцию генетической эволюции.
Факт, что Четвериков обращался к тем же самым моментам, не означает,
23 Фонд Четверикова, on. 1, ед. хр. 29, л. 3.
24 Фонд Четверикова, on. 1 ед. хр. 16, л. 10.
32
конечно, что он намеревался опровергать Данилевского: каждый из них был достаточно проницателен, чтобы определить и выставить на первый план «горячие точки» эволюционной теории. Однако интересно здесь именно то, что распорядились своей проницательностью они по-разному. Один обратил ее на выявление противоречивости и неполноты теоретической конструкции, оставаясь при этом в рамках существующей традиции. Другой строил оригинальную систему познавательных ориентиров, которая дала возможность открыть новые пути изучения процесса эволюции.
5. Проблема целостности генотипа. Здесь особенно ярко раскрывается системный подход Четверикова (Бабков, 1977а, 19836). Подчеркивая необходимость различать понятия «ген» и «признак», он замечал, что для генетика важны «геновариации», а для эволюциониста «сальтации», признаки. «Вопрос об образовании признаков, обусловленных изменением наследственного вещества, т. е. «сальтаций», есть собственно область не генетики, а механики развития»25 Так центральная проблема эволюционной генетики обогащается новым аспектом — изучением феногенетической изменчивости.
Конкретным выражением системного подхода Четверикова было признание иерархичности организации живой природы и существования иерархии управляющих процессов (Бабков, 1981). Он считал (говоря языком кибернетики), что в масштабе исторического развития вида действует один регулятор, а в масштабе индивидуального развития организма — другой. Представление о целостности живой природы подразумевает задачу выявить конкретные формы отношений этих регуляторов: любая популяция перестраивается не иначе, как посредством размножения и развития составляющих ее особей. Эта грань — указание связи двух уровней организации биологических процессов, но без утраты специфики каждого из них — воплощена в концепции «генотипической среды» Четверикова.
Анализ эволюционной роли отбора на основе таблицы Нортона (где отбор выступает как пассивный элиминирующий агент) не мог удовлетворить Четверикова. Подобный анализ вел к взгляду, что созидающая роль отбора, если она вообще существует, ничтожна. Альтернативой этому взгляду было в то время положение, что ген изменяется в зависимости от внешних условий, например в направлении действия отбора. Это положение противоречило менделевскому принципу «чистоты гамет», поэтому его не могли принять генетики. Четвериков отверг обе альтернативы и указал выход из тупика, обратившись к представлениям о плейотропии, согласно которым каждый ген может действовать не только на специфический ему признак, но в известной мере и на ряд других. В результате теряет смысл представление о мозаичности строения организмов из отдельных, независимых признаков, обусловленных отдельными же, независимыми генами; рушится характерная для классической генетики схема: «ген — признак». Организм — не мозаика, утверждает Четвериков, а индивид в буквальном смысле слова: «individuum», т. е. «неделимое». Каждый ген действует не независимо от всего генотипа, а в связи с ним, в своей «генотипической среде». Так Четвериков утверждает взгляд о целостности организма — не «атомарной» нерасчлененности, а системной целостности иерархически организованного объекта (Бабков, 1981).
Отбор по Четверикову не может изменить самого гена. Однако он может изменить не только частоту гена, но и путем создания благоприятной 5 Фонд Четверикова, on. 1, ед. хр. 5, л. 4.
Н В Бабков 33
генотипической среды его выражение. Это в предельном случае может имитировать возникновение новой мутации. Именно так обстояло дело с платиновой окраской у лисиц, возникшей не в результате самостоятельной мутации, как сперва решили в 1939 г. О. Мор и П. Туфф, а (как показал в 1951 г. Д. К. Беляев) в результате иного проявления старой мутации в новой генотипической среде, созданной интенсивным отбором по качествам мехового покрова (см.: Беляев, 1962).
Системный стиль Четверикова контрастировал стилю работы другого лидера московских генетиков — А. А. Серебровского. В генетических работах (мы не оцениваем его научное творчество в целом) Серебровский тяготел к редукционизму. Об этом говорит, например, его «Проект десятичной системы генетической символики», предпринятый по инициативе Н. К. Кольцова, где изучаемый ген обозначался «числовым символом наиболее характерного для него фена, признака» (Серебровский, 1921, с. 100). При переиздании работ Серебровского редколлегия сделала перевод основных генов по Серебровскому на общепринятую символику, причем не исключалось, что «некоторые гены, различные, но сходные по своему фенотипическому эффекту, окажутся перечисленными в нашем списке в качестве идентичных» (Серебровский, 1976, с. 135). Другую иллюстрацию своеобразного склада мышления Серебровского составляет его попытка создать русскую генетическую терминологию. Он рассуждал следующим образом. Гену характерно то, что он действует,— назовем ген словом «дей». Хромосома несет гены: — «дееносица». Гены мутируют, мутации бывают полезные, «добродей», и вредные, «лиходей» (цит. по: Бабков, 1983а, с. 24). Редукционистский подход сыграл роль в успехе генетических работ Серебровского. Четвериков же не отошел от системного склада мышления и в своих генетических исследованиях, притом что в то время главная линия развития генетики была проникнута редукционизмом (Roll-Hansen, 1978). Однако имено эти методологические основания позволили ему связать генетику с теорией отбора и с индивидуальным развитием (Бабков, 1981).
Из представленний о высокой генетической гетерогенности популяций следует, что один и тот же мутантный ген у разных особей популяций будет попадать в различную генотипическую среду, и, значит, его внешнее проявление каждый раз будет фенотипически флуктуировать. Такой взгляд, по Четверикову (1926, с. 45), позволяет, во-первых, отказаться от многочисленных дополнительных концептуальных конструкций: генов «усилителей», «ослабителей», «модификаторов», число которых «возросло до совершенно угрожающих размеров». Далее, в контексте «генотипической среды» обесценивается представление о принципиальном различии качественной и количественной изменчивости. Наконец, мы имеем право рассматривать выдвижение концепции «генотипической среды» как важный шаг от свойственных раннему периоду развития генетики механистических представлений о генотипе как мозаике генов и фенотипе как мозаике признаков к диалектическим представлениям о сложных связях гена и признака и об отношениях проблем наследственной передачи, наследственного осуществления и исторического развития.
В концепции генотипической среды особое значение для нас имеет не ее конкретное содержание (широкая плейотропия), а интегративный, или системный, подход. Особенно важно, что Четвериков распространил рассмотрение действия отбора, помимо уровня популяции, на уровень частных формообразовательных процессов, и связал их в единой кон-34
цептуальной схеме. Аргументация статьи 1926 г. строилась на определенной модели генетической эволюции: возникновение в природе «хороших» аутосомных рецессивов — накопление их в гетерозиготном состоянии в условиях свободы скрещивания — выявление в гомозиготах в условиях частичной изоляции — отбор по Нортону. (Близкие к четвериковскому варианты модели развивали одновременно Фишер, Райт, Холдейн). Модель служила Четверикову прежде всего для иллюстраций, сохранение ее любой ценой не предполагалось. И она не устояла даже в рамках статьи 1926 г., уступив место в гл. IV «Генотипическая среда» оригинальному представлению Четверикова об участии естественного отбора в поступательном процессе эволюции.
В первом популяционно-генетическом исследовании 1925—1926 гг., подтвердившем плодотворность центральной проблемы, выяснилось, что первоначальная модель нуждается в уточнении. Весь ход работ московской школы эволюционной генетики можно рассматривать как последовательную конкретизацию четвериковской проблемы и одновременно как постоянную ревизию первоначальной модели (Babkoff, 1981 а,b). Это привело в конце концов к формулировке предпосылок для создания более адекватной картины генетических основ эволюции.
Глава 2
ПЕРВОЕ ПОПУЛЯЦИОННО-ГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ
Задача ученого заключается не в том, чтобы пускаться в путь с готовой мудростью, но в том, чтобы добросовестно собирать ее по дороге.
Р. РОЛЛАН
Важным моментом, побудившим Четверикова к написанию статьи 1926 г., было стремление дополнить теорию эволюции Путем отбора указанием источников наследственной изменчивости — материала для отбора. Экспериментальное доказательство его тезиса о насыщенности мутациями фенотипически единообразных диких популяций было получено в первом популяционно-генетическом исследовании, которое провела его группа в 1925—1926 гг. Это исследование вместе с теоретической статьей 1926 г. положило начало экспериментальной генетике популяций; оно стало моделью Для планирования и проведения всей последующей популяционногенетической работы. В ходе этого исследования определились основные направления работ московской школы эволюционной генетики.
С лета 1923 г. в лаборатории Четверикова шли работы по изучению наследственной изменчивости дрозофил. Однако, как отметил Кольцов (1929, с. 23) в докладе о 10-летии работ Института, «наши первые попытки исследовать генетику Drosophila относятся к 1920 г., когда я предложил одному из своих молодых учеников [Ромашову] попробовать путем воздействия рентгеновских лучей на мух и их личинок вызвать у них появление новых мутаций...» Материалом послужили мухи вида D. funebris Из диких популяций окрестностей Звенигородской станции. Следует отметить, что неуспех этой ранней (за 7 лет до мёллеровской) работы Ромашова
был обусловлен не только отсутствием линий с известным генетическим происхождением, которые обеспечивают количественный учет, но и особенностями феногенетики данного вида. Сводку богатых материалов по генетике D. funebris, полученных в 1923—1925 гг. Балкашиной, Промп-товым, Ромашовым, Тимофеевыми-Ресовскими и Царапкиным, опубликовали Ромашов и Балкашина (1929—1930 гг.; Romaschoff, Balkaschina, 1931); на первый план здесь вышли особенности наследования и проявления признаков в зависимости от генотипа и внешних условий — их анализ привел к постановке проблемы фенотипического осуществления гена. Хотя такце работы раскрывали изменчивость материала, взятого из природы, они не были еще популяционными в собственном смысле слова.
Задача исследования была сформулирована Четвертиковым в мае 1925 г. на II Съезде зоологов в Москве, в докладе «Теоретическая предпосылка генотипического анализа видов рода Drosophila* (Четвериков, 19276), а летом началось исследование звенигородских популяций— D. obscura, phalerata, vibrissina, transversa (тогда в Подмосковье почти не было фруктовых садов и связанной с ними D. melanogaster). В сентябре 1925 г., когда Четвериков писал свою теоретическую статью, он использовал первые результаты этого анализа (его вели Астауров, Балкашина, Беляев, Гершензон, а также Промптов, Ромашов, Тимофеев-Ресовский). Однако трудность разведения этих видов (личинки которых питаются грибами) и их низкая жизнеспособность делали полученные результаты отрывочными и неполными. Более систематическое исследование было проведено следующим летом на классическом объекте D. melanogaster из популяций Геленджика и Берлина. Результаты изучения трех поколений 239 геленджикских линий (и сравнение с берлинскими 78 линиями) были кратко изложены Четвериковым в 1927 г.— в сентябре на V Генетическом конгрессе в Берлине (Tschetwerikoff, 1927, 1928) и в декабре на III Съезде зоологов в Ленинграде (Четвериков, 1928). В 1929—1931 гг. Балкашина и Ромашов обследовали ряд популяций D. funebris Москвы, Петровско-Разумовского, Киева и Ташкента и доложили первые результаты на IV Съезде зоологов в Киеве.
Зимой 1928—1929 гг. Четвериков подводил итоги изучения популяции D. melanogaster. В девятом номере журнала «Научное слово» (1928) сообщалось, что в ближайших номерах будет опубликована статья проф. С. С. Четверикова «Роль мутаций в эволюции организмов». В Архиве АН СССР хранится часть конспекта «Значение мутаций в эволюционном процессе» со ссылками на геленджикские материалы и план доклада в МОИП 21.II 1.1929 г. «Происхождение и сущность мутационной изменчивости» 1 Тогда же Четвериков готовил английский перевод статьи 1926 г. и включил в него отчет об этом исследовании в качестве новой V главы «Экспериментальное исследование геновариаций, скрытых в природных популяциях» и соответствующие этой главе пункты 22—25 — в «Выводы». Но весной 1929 г. Четвериков покинул Москву; статьи он не довел до опубликования и к исследованию дрозофил больше не возвращался. Участники этой коллективной работы рассматривали Четверикова, всегда неспешного с публикациями, как ее вдохновителя и руководителя и не считали себя вправе опережать его. В результате материалы 1925 г. по звенигородским популяциям были опубликованы лишь в 1934—1935 гг., *
Фонд Четверикова, on. 1, ед. хр. 5 и 9.
36
а наиболее богатые материалы 1926 г. по D. melanogaster (как и материалы 1929—1931 гг. по D. funebris) в полном виде не были опубликованы. (Неопубликованные материалы Балкашиной по геленджикской популяции и материалы по D. funebris были использованы впоследствии Ромашовым в работах по генетико-автоматическим процессам и по массовой изменчивости жилкования. Отчет Четверикова 1929 г. опубл, в 1982 г.)
АНАЛИЗ ПОПУЛЯЦИЙ DROSOPHILA MELANOGASTER
На краткие сообщения Четверикова «О генетическом строении диких популяций» (Tschetwerikoff, 1927, 1928) и «Экспериментальное решение одной эволюционной проблемы» (Четвериков, 1928), касавшиеся изучения D. melanogaster в 1926 г., к которому мы сейчас обратимся, впоследствии ссылались все генетики-эволюционисты — советские, западноевропейские, американские — как на первое экспериментальное популяционно-генетическое исследование.
Летом 1926 г. Рокицкий и Гершензон отловили в популяции Геленджика, на Кавказе, более 300 оплодотворенных на воле самок D. melanogaster; 239 из них благополучно прибыли в Москву, где они и дали достаточное потомство (26 916 особей в Fi; 124 554 в F2 и т. д.). Анализ, в котором участвовали Е. И. Балкашина, А. И. Четверикова, Б. Л. Астауров, Н. К. Беляев и А. Н. Промптов, показал, что 239 диких самок гетерозиготны по 33 различным «хорошим» генам с явным морфологическим эффектом (не считая летальных, физиологических и других генов); по оценке Четверикова 1929 г., каждая самка гетерозиготна в среднем по пяти — шести генам. Обнаруженные 33 мутации затрагивали самые разные признаки тела (в гл. V приведена таблица распределения мутаций по частям тела мухи); все они были рецессивными и, за одним исключением, локализованы в аутосомах. Обратившись к анализу отдельных мутаций (в гл. V), Четвериков заметил, что «их значение чрезвычайно неодинаково, простираясь от незначительных уклонений в жилковании или в числе щетинок до нарушений фундаментальных законов структуры организма» (Четвериков, 1982, с. 346). В этой последней группе он особенно отметил мутацию «aristopedia», которая заменяет кисточку на конце усика лапкой со всеми ее морфологическими частями (коготками, пульвиллой, щетинкой), и ген «abdomen rotatum», поворачивающий брюшко мушки влево, как бы вращая его вокруг оси. Но если эти гены встретились соответственно однажды и дважды, то другие — чаще, заражая 7—40% и даже до 50% самок (ген «polychaeta», проявляющийся увеличенным числом щетинок на груди и щитке). Не выставляя каких-либо предположений о причинах, Четвериков констатировал сам факт широкого распространения отдельных мутантных генов и заключил, что наследственные различия популяций могут выражаться «и отчасти даже выражаются и внешне, фенотипически, в том, что среди геленджикского населения средний тип некоторых признаков (например, число грудных макрохет) несколько отличен от берлинских, что уже указывает на определенную географическую, расовую изменчивость, обусловленную географической изолированностью этих двух популяций и разным процессом геновариацион-ной изменчивости 2 в каждой из них» (Четвериков, 1928, с. 53).
«Процесс геновариационной изменчивости», по Четверикову, зависит от взаимодействия всех факторов накопления мутаций и их устранения из популяций.
37
Результаты этого исследования были особенно интересны Четверикову в двух отношениях. Во-первых, они подтвердили основную мысль статьи 1926 г. о существовании в природе процесса накопления гетерозиготных рецессивных мутаций, «образующих фонд самых разнообразных наследственных возможностей» (Четвериков, 1982, с. 347). Во-вторых, они показали, что этот процесс «носит массовый характер» (там же). На этом основании Четвериков отверг критику мнения об эволюции путем отбора случайных изменений: «Пока мы, с проф. Плате, принимаем, что прогрессивный процесс эволюции требует, «чтобы необходимая мутация всегда возникала в должный момент», случайная природа возникающих наследственных изменений повлечет непреодолимое препятствие. Но это препятствие сразу же рухнет, если мы сможем доказать, что наш процесс является массовым процессом, что природа вмещает в органическом мире необъятный резерв наследственной изменчивости — геновариаций, из которых отбор может свободно черпать «всегда как только возникнет необходимость» (там же, с. 348).
Терминологическое отступление. Сейчас мы говорим о генофонде популяций, который характеризуется частотами генов, величиной генетического груза и т. п. Эти и другие основные понятия эволюционной генетики ввел (хотя и не обозначил терминами) Четвериков. Понятие генофонда сложилось в начале 20-х гг., одновременно во всех основных центрах генетических исследований нашей страны. Серебровский даже ввел этот термин — за четверть века до gene pool Добржанского, придав ему, правда, весьма широкое и неоднозначное значение: «...Раз запас тех или иных генов в данной местности может длительно сохраняться, то мы должны рассматривать этот запас как своего рода естественное богатство, подобное, например, запасами нефти, золота, каменного угля и пр. Для того чтобы подчеркнуть именно эту сторону дела, мы предложили три года назад специальный термин генофонд» (Серебровский, 1928, с. 6; ср.: он же, 1926, с. 116—117). Этот термин употребил также Кольцов (1926, с. 143).
Современному понятию «генофонд популяции» у Четверикова и его учеников соответствовали термины: «генетическое строение населения вида» (Балкашина, Ромашов, 1935), «genetische Struktur» (Н. A. Timofe-eff-Ressovsky, N. W. Timofeeff-Ressovsky, 1927), «genetic structure» (co ссылками на советских авторов)— У Спенсер, Ф. Айвз 3, а также и другие. Четвериков, строго относившийся к введению новых терминов, не мог принять слова Серебровского «генофонд» (именно слова, так как термином оно не было, ибо не была определена область его значения). В 1929 г. Четвериков употребил выражение «запас (резерв) наследственной изменчивости», а Шмальгаузен писал позже о «резерве наследственной изменчивости вида и его мобилизации» (например, 1946, с. 191). Ромашов обозначил термином «фонд популяционной изменчивости» все обнаруженные в ходе первого популяционного исследования изменения: как «хорошие» мутации, так и вероятно-наследственные и ненаследственные изменения. Развивая представления о роли стохастических процессов в эволюции, он не мог бы пользоваться выражением «генофонд», которое подчеркивало
3 Spencer W. Р. On the biology of Drosophila immigrans Sturtevant with special reference to the genetic structure of populations.— Ohio J. Sci., 1940, vol. 40, p. 345—361. Jves Ph. T The genetic structure of American populations of Drosophila melano-gaster.— Genetics, 1945, vol. 30, p. 167—196.
38
Таблица 1
Частота мутаций в двух популяциях Drosophila melanogaster (по материалам 1926 г.)
Геленджик, 1926, 52 линии Берлин-Штеглиц, 1926, 78 линий
(Балкашина) (Тимофеевы-Ресовские)
Мутация 1 % Мутация 1 %
extra-analis-plexus 21,6 Venae plexoides 4,9
analis incompletus 4,3 alae vesiculatae (I) 1,9
hemiptera 3,3 Abnormal abdomen (I) 1,6
Типа cluboid 1,8 Venae abnormes 1,6
tiny-bristles 1,8 alae divergentes 1,2
purple 1,4 Beaded 0,9
Delta 0,9 lethal-2 (I) 0,9
alae truncatae 0,9 venae incompletae 0,6
sepia 0,9 venae bifurcatae X
alae divergentes X lethal-1 (I) X
alae undulatae X
Типа balloon (I) X
vena abnormales-Notch X
ramuli (media) X
radius incompletus X
pedes taxi X
singed-curvatae X
aristae deformatae X
aristopedia X
Примечание. Указаны
наиболее резкие мутационные изменения; «х» означает
единичное появление; <(!)» — мутации I хромосомы, т. е. сцепленные с полом.
в формулировке Серебровского статичность генного состава популяций. Впоследствии Тимофеев-Ресовский, касаясь истории понятия, отметил, что более точным был бы термин аллелофонд (Тимофеев-Ресовский и др., 1973, с. 165) \
Одновременно с исследованием геленджикской популяции D. melanogaster Е. А. и Н. В. Тимофеевы-Ресовские проанализировали берлинскую популяцию этого вида. Их работа должна была ответить на те же вопросы. «Являются ли геновариации Drosophila melanogaster лабораторными артефактами или они также возникают (в гетерозиготном состоянии) в природе?.. В какой степени дикая популяция «обременена» [«belastet»] * 5 геновариациями в гетерозиготном состоянии?.. Кроме того, эта работа является экспериментальной проверкой положений эволюционной теории С. С. Четверикова» (Н. A. Timofeeff-Ressovsky, N. W. Timofeeff-Res-sovsky, 1927, S. 71—72).
Материалом для этого исследования послужила ограниченная популяция D- melanogaster южной части Берлина (р-н Штеглиц). Было изучено 78 инбредных линий на протяжении трех поколений. Выделено всего Ю различных «бросающихся в глаза» мутантных признаков. Из 48 случаев обнаружения мутаций 29 приходилось на аутосомные доминанты, сцеплен
Вопросов терминологии, по-видимому, касается статья: Adams М. В. From «gene
5 to *&ene Pool».— Stud. Hist. Biol., 1979, vol. 3, p. 241—285 (n. v.).
у1*0 «belaster» Тимофеевых сходствует с более поздними «генетическое бремя» («груз») __ «genetic burden» («load»).
39
ные с полом доминанты и сцепленные с полом рецессивы, видимые и летальные. Общий вывод гласил: «В природе точно так же, как и в лабораторных культурах, постоянно возникают геновариации, вследствие этого каждая дикая популяция содержит различные факторы в гетерозиготном состоянии; при внутрибрачии дикие мухи в течение нескольких поколений (двух или трех) проявляют эти факторы, которые выщепляются в потомстве» (Н. A. Timofeeff-Ressovsky, N. W. Timofeeff-Ressovsky, 1927, S. 108).
В табл. 1 приводятся частоты мутаций (процент гамет в популяциях) для выборок из популяций Геленджика (по неопубликованным данным Балкашиной6 — 52 линии) и Берлин—Штеглица (по Н. A. Timofeeff-Ressovsky, N. W. Timofeeff-Ressovsky, 1927 — 78 линий).
В первом экспериментальном исследовании свободно живущих популяций обращает на себя внимание определенное различие в понимании своей цели Четвериковым и его учениками, что отразилось в стиле их статей (Бабков, 1982). Четверикову прежде всего было важно утвердить общие принципы исследования: совместимость генетики и дарвинизма и плодотворность их союза, правомерность экспериментального подхода при решении общеэволюционных проблем, ненаследуемость внешних влияний, возникновение мутаций в природе, роль метода инбридинга и т. п. Экспериментальные же результаты были важны лишь как иллюстрация этих положений. Четвериков приводит только три числа в тезисах 1928 г., а в гл. V — таблицу, которая служит для доказательства, что все части тела мухи подвержены мутационной изменчивости. Напротив, отчет Тимофеевых-Ресовских 1927 г., например, содержит чрезвычайно подробное описание результатов (на 26 рис. и 18 табл.), как и публикации Балкашиной и Ромашова (1929—1931 гг. и 1935 г.), а также С. Гордон (Gordon, 1936) и др.
Ученики Четверикова (первое поколение эволюционных генетиков) уже не считали необходимым доказывать совместимость менделевской генетики и дарвинова отбора: это принималось как данное. Задачу своей работы они видели в экспериментальном исследовании на пути, обоснование которому дал Четвериков, и в теоретическом осмыслении своих результатов, ориентированном на создание генетической концепции эволюционного процесса.
Теоретическая статья 1926 г. и первое экспериментальное исследование положили границу между двумя биологическими парадигмами — уходящей в прошлое (с конфронтацией натуралистов и экспериментаторов, дарвинистов и менделистов) и грядущей, с программой построения эволюционной генетики.
АНАЛИЗ ПОПУЛЯЦИЙ СРЕДНЕРУССКИХ ВИДОВ ДРОЗОФИЛ
Генетико-популяционной работе 1925—1926 гг. предшествовало выявление мутаций в потомстве отловленных на воле самок D. funebris^ Эта работа 1923—1925 гг. имела целью прежде всего овладение лабораторными методами; но уже в ее ходе был подтвержден тезис о существовании в природе «резерва наследственной изменчивости» и были сделаны некоторые другие
6 Часть данных по геленджикской популяции D. melanogaster, проанализированных Балкашиной, приведена в ст.: Дубинин, Ромашов, 1932 (табл. 7).
40
интересные выводы (Ромашов, Балкашина, 1929—1930 гг.; Romaschoff, Balkaschina, 1931).
Истоки феногенетических работ. Общая картина наследования у D. funebris оказались весьма сложной и довольно далекой от той, которую следовало ожидать исходя из представления, что большинство изменений наследуются по простым менделевским схемам. Напротив, по данным группы Моргана (Morgan, Bridges, 1916; Bridges, Morgan, 1919, 1923; Morgan et al., 1925) лишь немногие мутации D. melanogaster наследуются «неправильно» и дают при скрещиваниях непостоянные числовые отношения с резким «отставанием» числа измененных особей от ожидаемого согласно менделевскому отношению. Однако Ромашов и Балкашина подчеркнули, что в упомянутых монографиях I, II и III хромосом D. melanogaster большая часть подобных «неправильно» наследующихся мутаций вообще не помещена: «очевидно, детальное изучение каждого «отступления» не входило в задачи лаборатории Моргана и поэтому такие изменения обычно не сохранялись» (Ромашов, Балкашина, 1929, с. 145). Они указали там же, что по материалам, собранным в лабораториях Четверикова и Серебровского, общая картина наследования у D. melanogaster также оказалась очень неправильной, но все же далеко не столь сложной и запутанной, как у D. funebris. Таким образом, различие между двумя видами было скорее количественное. К подобному заключению пришел и Тимофеев-Ресовский, много работавший с обоими видами (личн. сообщ.). (Ср. ниже, с. 117.)
Анализ случаев, которые не укладываются в рамки общепринятых представлений, часто приводит к выявлению и формулировке новых закономерностей. А для философа науки аномалии интересны еще и тем, что при столкновении с ними полнее раскрываются естественно-научные и методологические основания исследователей. Действительно, именно в ходе выяснения причин нерегулярностей в наследовании четвериковцы разработали основные принципы фенотипического осуществления генов; здесь проявились их широкие зоологические интересы и системный подход (см. гл. 3—7). Напротив, члены дрозофильной группы Моргана вообще исключили из рассмотрения этот материал как не подходящий для решения их основной задачи — разработки механистической линейной модели хромосомы. Это указывает на их редукционистскую методологию (Roll-Hansen, 1978), которая, кстати, дала им возможность сконцентрировать внимание на проблеме наследственной передачи, не принимая в расчет проблемы наследственного осуществления.
Материалы исследования 1925 г. звенигородских популяций D. obscura, phalerata, transversa, vibrissina были опубликованы в 1934 г. Гершензоном (очень кратко, по первому виду: Gershenson, 1934) и в 1935 г. Балкаши-ной и Ромашовым (по трем другим). Некоторые результаты этого анализа, подтвердившего тезис Четверикова о насыщенности природных популяций мутациями, сведены в табл. 2. Однако наиболее интересными моментами в работе Балкашиной и Ромашова (1935) являются: заключение об известной структурной упорядоченности изменчивости; выявление трех категорий изменчивости; новые представления о факторах, определяющих генофонд популяции.
<Направленная» изменчивость как следствие особенностей онтогенеза.
Ыв°Д, что «изменчивость отдельных признаков не представляет случайного, беспорядочного процесса, но является «определенно направленной» и ограниченной» (Ромашов, Балкашина, 1930, с. 207—208) был получен
41
Таблица 2
Насыщенность мутациями популяций дрозофил (Звенигород, 1925 г.)
Показатель D. phalerata, 47 линий (Астауров, Беляев) D. transversa, 22 линии (Астауров) D. vibrissina, 9 линий (Балкашина) D. obscura, 19 линий (Горшензон)
Число мух В Fi 1225 596 1147
В F2 6017 5775 1496
Число различных уклонений, 29 32 14 9
отмеченных в F| и F2 Среднее число изменений на 2,0 4,7 2,8 0,7
линию из них наследственных и веро- 0,7 2,1 0,8 0,7
ятно наследственных Процент измененных мух в F| 2,4 4,0 2,4
из них наследственных и ве- 1,3 3,0 1,5
роятно наследственных Среднее число мутаций на муху 0,34 1,0 0,4
из природного населения
при осмотре более 40 000 мух. Направленность особенно ясно видна в изменчивости жилкования крыла; одним из наиболее частых изменений у D. funebris является перерыв или утеря 2-й поперечной жилки (этот признак характеризует четыре мутации и ряд «нерасщепляющихся» изменений). Этот тип изменений характерен также для семи других видов дрозофил (willistoni, simulans, virilis, transversa, melanogaster, phalerata, vibrissina; Балкашина, Ромашов, 1935); он «характеризует род Asteia, выделенный в настоящее время в особое подсемейство» (Ромашов, Балкашина, 1930, с. 208). Другие примеры «определенно направленной» изменчивости доставляют группы изменений, касающихся хетотаксии и остановки в процессе расправления крыльев. Из этих данных «напрашивается вывод, что в конце концов удается разложить изменчивость вида на ограниченное число типов уклонений, которые возникают особенно часто и представляют определенные реакции организма на различные (внутренние или внешние) воздействия» (Там же, с. 209). К такому же выводу о пластичности признаков в сравнительно немногих и общих для близкородственных видов направлениях и о причине этого явления пришли Тимофеевы-Ресовские (см. гл. 3). Обратившись к другому материалу, Балкашина и Ромашов (1935, с. 100) заметили, что «...многие «массовые» изменения, постоянно повторяющиеся в различных линиях одного вида, являются характерными также для остальных изучавшихся видов, а может быть и для всего рода Drosophila...*
Категории мутационной изменчивости. По материалам v исследования 1925 г. Балкашина и Ромашов (1935) выделили три категории изменений, составляющих «фонд популяционной изменчивости». Это, во-первых, немногочисленная группа под названием «единичных мутаций» (то, что часто называют «хорошими» мутациями). Эти изменения возникают сравнительно редко, ограничены популяциями одного вида, дают картины моногибридного расщепления (хотя бы и с искажениями и «отставаниями», которые в этих случаях объясняются либо пониженной жизнеспособностью, либо невысоким и варьирующим проявлением, либо трудно-42
Таблица 3
Массовая изменчивость в популяциях дрозофил (по: Балкашина, Ромашов, 1935)
Номер Тип изменения Во скольких линиях появилось из:
47 23 9 D. phalerata D. transversa D. vibrissina
1 crossvein less (характерно также для изменений: crossveinless-curved, venae incompletae-ph; v. conflexae-tr) 2 plexus1 14 12 3 10 10 — 3 plexus* 2 (plexusJriangularis-tr) 9 7 4 divergens 6 5 4 5 venae bifurcatae (radius bifur- 5 8 — catus-tr) 6 Beaded 6 3 2 7 extra-media 1 9 1 8 shagreen (al. undulatae; al. curva- tae; notch-curved; crossveinless-curved-ph; purple-tr)* 6 4 — 9 balloon 4 4 1 10 guitar 1 2 4 11 radius incompletus 4 — 1 12 alae truncatae 1 1 2
* Самый обычный тип изменения у всех трех видов, возникающий почти в каждой линии (в большинстве случаев не отмечался).
стыо отличить измененных мух). В тех довольно редких случаях, когда подобные мутации возникают в нескольких линиях, весьма вероятно, что они идентичны.
Этим «хорошим» мутациям авторы противопоставляют две многочисленные группы изменений. «Массовые изменения» возникают часто, повторяются у близких видов («фенотипический параллелизм») и дают чрезвычайно сложные и запутанные картины наследования. (Примеры: crossveinless — перерыв 2-й поперечный жилки; divergence — постоянно расставленные крылья; beaded — вырезки на крыле; shagreen — шагреневые изогнутые крылья; plexus — добавочные жилки и т. п.— см. табл. 3). Развивая свои взгляды о существовании определенных направлений в изменчивости, авторы замечают: «По-видимому, здесь мы имеем дело с весьма сложной наследственной обусловленностью изменений, с тем, что один и тот же признак возникает каждый раз в результате сложного взаимодействия различных наследственных задатков и внешних условий; поэтому является весьма вероятным, что под одним названием (например, crossveinless) мы, как правило, объединяем неоднородные и очень различные по своей природе изменения...» (Балкашина, Ромашов, 1935, с. 102).
Интересно, что тот же материал («crossveinless») через 20 лет привлекал К. Уоддингтон для экспериментального подтверждения концепции «канализованного» развития (ср.: Балкашина и др., 1975, с. 78). Характер Жилкования в участке задней поперечной жилки (posterior crossvein) крыла Дрозофилы иллюстрирует положение, что нормальный фенотип обладает определенными буферными свойствами (Waddington, 1955, 1961). Для изуче-
43
ния этого вопроса был выведен ряд линий дрозофил; у некоторых из них эта жилка прерывалась (L2 — маленькие перерывы; L5 — частые большие перерывы); у других образовывались добавочные отвилки (Е2 — маленькие добавки, Е5 — большие и частые). Оказалось, что различия линий были обусловлены влиянием многих генов, каждый из которых обладает слабым действием, как показали скрещивания типа L2X L5 или Е2 X Е5, а не одной или двумя «хорошими» мутациями. Скрещивания типа L X Е, а также обратные и анализирующие, показали, что один и тот же фенотипический класс (в том числе нормальный «дикий тип») может быть получен разными способами, «поэтому признак «количество crossvein» не является «причинно гомогенным» («causally homogeneous») (Waddington, 1955, р. 227); в частности, как писали Балкашина и Ромашов, один и тот же фетопип crossveinless (фенотипические классы L2 — L5 Уоддингтона) может развиваться на основе разнообразных генотипов.
Третью, многочисленную, группу составляют редкие «единичные изменения», которые если и дают какие-либо указания на наследуемость, то очень неясные. Характерно, что большинство изменений, возникающих в потомстве диких самок, относится к двум последним категориям, которые «составляют как бы общий основной фон изменчивости вида, и на этом фоне лишь очень редко «проскакивают отдельные резкие мутационные гены» (Балкашина, Ромашов, 1935, с. 103).
Представление о структурных ограничениях мутационного процесса как будто противоречило устремлению Четверикова доказать, что все части организма затрагиваются мутационным процессом, а заключение о насыщенности популяций «плохо наследующимися» изменениями — его устремлению найти в природе «хорошие» мутации. Однако если отделить в теории Четверикова главные моменты от второстепенных, то легко видеть, что такие противоречия не могли быть фатальными для становления эволюционной генетики. Действительно, работы московской школы представляют собой последовательную конкретизацию четвериковской проблемы, в ходе которой проверялись и уточнялись отдельные моменты картины генетических основ эволюции.
Два упомянутых выше результата — представления об известной упорядоченности мутационной изменчивости и о трех категориях популяционной изменчивости — углубляли четвериковскую проблему в аспекте природы изменчивости. Третий касался также темы поддержания изменчивости. Истоки концепции, подчеркивающей эволюционную роль случайных процессов (ей посвящена гл. 8),— в первом генетическом анализе природных популяций 1925 г. В статье 1935 г., посвященной материалам этого анализа, рисовалась следующая картина образования генофонда. Мутационный процесс ведет к высокой гетерозиготности большинства особей вида, главным образом по «мелким» изменениям. Редко возникающие в популяциях «хорошие» мутации тотчас же «подвергаются дифференцирующему действию генетико-автоматических процессов»: огромное большинство из них уничтожается в результате менделевской стохастики и увеличивает-тся концентрация тех, что случайно сохранились в населении вида. (Данные табл. 1, указывающие на различие ассортимента мутаций двух удаленных друг от друга популяций D. melanogaster, иллюстрируют такое действие ГАП’ов.) Дифференциация частот генов может привести к выщеплению в природе рецессивов в гомозиготном состоянии. (Данные предпоследней строки табл. 2 подтверждают этот тезис: поскольку для анали-
44
брались оплодотворенные в природе самки, очевидно, что подобные рАсщепления встречаются и в природе.) «Все это заставляет нас пересмотреть так часто повторяющееся утверждение о полной внешней однородности популяций Drosophila и ставит вопрос о необходимости более подробного изучения изменчивости у различных представителей этого рода Непосредственно в природных населениях» (Балкашина, Ромашов, 1935, с. 105). В развитие этого положения было предпринято исследование аберративного полиморфизма природных популяций дрозофилы (этому посвящена гл. 9), открывшее еще одну тему в разработке четвериковской проблемы.
ИТОГИ ПЕРВЫХ РАБОТ
Резонанс первого экспериментального исследования природных популяций. Западная биология воприняла четвериковскую идею генетического анализа диких популяций вскоре после появления в 1930—1932 гг. классических работ Фишера, Райта, Холдейна, которые вместе с теоретическим исследованием Четверикова 1926 г. показали плодотворность объединения принципов менделевской наследственности и дарвинова отбора в единой концептуальной схеме. Распространению идей Четверикова в Западной Европе способствовали прежде всего многочисленные работы Тимофеева-Ресовского. Известную роль сыграл также и Холдейн, который бывал в ИЭБ и следил за работами Четверикова (Астауров, Рокицкий, 1975, с. 26). Его ученица Сесиль Гордон провела в 1933—1934 гг. первое в Англии подобное исследование. Она подчеркивала пионерскую роль русских исследователей: «Следуя теоретическим соображениям Четверикова (1926), Тимофеевы-Ресовские (1928) и Четвериков (1928) подвергли инбридингу популяции D. melanogaster для изучения распространения рецессивных мутаций. Дубинин и сотрудники (1934) повторили это в большем масштабе, используя несколько иные методы инбридинга. В настоящей статье описана дальнейшая работа в этом направлении» (Gordon, 1936, р. 25). В этой и других работах (например, Gordon et al., 1939) уделено много места обсуждению первых популяционных работ группы Четверикова; обсуждалось также представление о модифицирующем действии «генотипической среды» на жизнеспособность мутаций. Однако в отличие от русских работ здесь много математических расчетов в духе Холдейна.
Добржанский распространил идеи Четверикова в США. Добржанский комбинировал с популяционными также и цитогенетические подходы, разработанные им совместно с Бриджесом и Стёртевантом; в совместных работах с Райтом» использованы также довольно сложные математические методы. В 1938 г. Добржанский начал публиковать серию статей под общим названием «Генетика природных популяций», которая оказала глубокое влияние на это направление работ, и продолжал ее до своей смерти в 1975 г. (см. переиздание: Dobzansky, 1981). Когда же в результате этих работ естественно-науч ное мировоззрение (система познавательных ориентиров) группы Четверикова было в значительной мере принято коллегами и учениками Добржанского (И. М. Лернером, Э. Майром, Б. Уоллесом, Р. Левонтином и многими другими генетиками и эволюционистами), Он счел нужным подчеркнуть роль Четверикова в формировании этого мировоззрения (Dobzhansky, 1955, 1959, 1967 и др.).
45
В сводке Добржанского «Генетика эволюционного процесса» такн^ указана роль русских работ. «Пионерское экспериментальное исследование того, что позже получило название генетического груза или генетического бремени у дрозофилы, провели Четвериков (1926), Е. Тимофеева-Ресовская и Н. В. Тимофеев-Ресовский (1927) и Дубинин и его коллеги (Дубинин и 14 соавторов, 1934; Дубинин, Ромашов и др., 1937) на популяциях нескольких видов, главным образом Drosophila melanogaster* (Dobzhansky, 1970, р. 109). Эти работы давно уже вошли в учебники. Так, во «Введении в современную генетику» Уоддингтон (Waddington, 1939, р. 286) отметил, что вслед за русскими исследователями аналогичные работы провели Добржанский и Квиль (1938), Гордон (1936), Стертевант (1937). Обзор ранних популяционных работ дали Добржанский (Dobzhansky, 1939) и Спенсер (Spencer, 1947). Основополагающая роль работы группы Четверикова для экспериментальной генетики популяций отмечена в ряде публикаций (например: Астауров, 1965 и др.; Dobzhansky, 1937 и др.; Timofeeff-Ressovsky, 1940а и др.; Lerner, 1955, 1958; Adams, 1965).
Направления исследований московской школы. Однако какие последствия имели первые экспериментальные работы для определения направлений исследований московской школы эволюционной генетики? В его ходе выяснились правомерность четвериковского подхода и плодотворность его центральной проблемы («природа и поддержание изменчивости в популяциях»). В целом становление эволюционной генетики в работах московской школы выглядит как конкретизация (шаг за шагом) центральной проблемы, достигаемая ценой последовательного пересмотра модели явления.
В первом экспериментальном исследовании определились два аспекта четвериковской проблемы: популяционный (изменчивость в природных популяциях) и феногенетический (изменчивость как результат фенотипического осуществления гена). В ходе феногенетических исследований (см. гл. 3—7 наст, кн.) был выявлен фенотипический параллелизм «направленной» изменчивости, построена система основных параметров фенотипического осуществления гена, сформулированы представления о природной изменчивости как результате некоординированности онтогенеза, представление о гомеозисах как развилках в онтогенезе. Изучение интеграции генотипа, идущее от концепции «генотипической среды» Четверикова, получило развитие в концепции «наследственной конституции», принципе «автономной изменчивости признаков», представлениях о «полях действия генов». Проникнутая системным подходом, серия феногенетических работ осталась вне круга интересов генетиков с редукционистской ориентацией. Напротив, она дала важный материал для теории стабилизирующего отбора И. И. Шмальгаузена; по своей ориентации эти работы были близки Уоддингтону, Дарлингтону, Лернеру, Камшилову.
В популяционных исследованиях (см. гл. 8—10) выяснялись категории природной изменчивости и механизмы ее поддержания. Концепция ГАП’ов (с ее позиций обсуждались результаты первого популяционно-генетического исследования 1925 г.) дала основание для предсказания аберративного полиморфизма и деталей в природе; она вызвала обсуждение проблемы соотношения факторов, определяющих генетическую композицию и динамику популяций. Первая количественная оценка генетического груза, полученная при выявлении насыщенности популяций деталями, привела к переносу интересов с вопроса о «количестве изменчивости» на вопрос об «изменчивости по приспособленности». Адаптивные механизмы были выявлены в
46
^работах по сезонному полиморфизму и хромосомной изменчивости, выли сформулированы основные элементарные понятия учения о микро-ЭВШ1ЮЦИИ.
Феногенетический и популяционный аспекты проблемы были в известной мереуобъединены в ходе феногенетического изучения массовых популяционных изменений, которое привело к концепции интеграции генетических систем\(см. гл. 11). Изучение коррелированной изменчивости конституциональных признаков позволили в новом свете поставить проблему отношений микро- и макроэволюции (см. гл. 12). В двух последних циклах работ речь идет о генетической основе нормальных видовых признаков и в центре внимания оказывается новый вопрос — «эволюционно значимая изменчивость». Их результаты дали основание для пересмотра ряда основных положений «факториальной» генетики популяций и для создания более адекватной картины генетических основ эволюции.
Глава 3
ФЕНОМЕНОЛОГИЯ РЕАЛИЗАЦИИ ГЕНА
Высь, ширь, глубь. Лишь три координаты.
В. я. БРЮСОВ
Сложность взаимоотношений между генами и признаками показали такие явления, как гетерогенные группы (мутации разных генов дают сходный фенотипический эффект) и плейотропия (мутация одного гена вызывает целый комплекс конечных признаков). Концепция «генотипической среды» явилась обобщением представлений о плейотропии. Но в первую очередь интерес четвериковской группы к разработке вопросов феногенетики был обязан собственной работе по генетике Drosophila funebris, в ходе которой выявились уклонения от классических картин наследования. В сводке по генетике этого вида, основу которой составили результаты неоконченных или неопубликованных работ сотрудников Генетической лаборатории ИЭБ, Д. Д. Ромашов, и Е. И. Балкашина (1929, с. 103) подчеркивали, что выражения «сложные» или «неправильные» расщепления «относятся исключительно к явлениям, происходящим в фенотипе', по отношению к отдельным генам и ко всему генотипу любое расщепление будет одинаково правильным...» Эти аномалии, таким образом, обязаны нерегулярному проявлению и варьирующему выражению гена. Организмы с такими особенностями мало пригодны для выявления групп сцепления (одна из задач группы Моргана), но зато дают прекрасный материал для изучения действия внешних и внутренних факторов на развитие наследственных признаков. (К такому выводу наряду с московскими генетиками пришел также У. Спенсер — Spencer, 1928.)
Термин феногенетика, который употребляли четвериковцы, принадлежит немецкому зоологу Валентину Геккеру (Наескег, 1918). Изучая мутации млекопитающих, он пытался выявить фенокритические фазы — такие моменты в онтогенезе, начиная с которых удавалось отличать мутантные особи от немутантных.
На терминологию феногенетического цикла работ московской школы
47
оказали влияние также работы О. и С. Фогтов, в которых рассматривались факты из области морфофизиологии ЦНС в свете генетики. Оскар Фогт разделял увлечение немецких естествоиспытателей генетикой и энтомологией; во всех своих путешествиях он собирал шмелей для изучений их изменчивости; он опубликовал несколько статей по изменчивости и проблемам вида. /
В связи с прогнозом, клиникой и терапией заболеваний ЦНЙ Фогт (о его пионерских работах см.: Филимонов, 1960) говорил о так называемых слабых генах, с нерегулярным проявлением. Слабые гены дают видимый эффект только от случая к случаю; для обозначения этого явления Фогт ввел понятие «пенетрантность» (Penetranz); он подчеркивал, что зависимые от слабо пенетрантных генов наследственные дегенерации и уродства, при скудости посемейных обследований, могут представляться чем-то совершенно новым в данной семье. Образ проявления многих генов может также колебаться—их «специфичность» (Spezifizitat) непостоянна. Колеблемость «топической специфичности» приводит к тому, что различные клинические картины могут соответствовать единой генетической основе. Колеблемость «морфологической специфичности» может привести к «полярной противоположности» в степени развития признака, т. е. один и тот же ген может вызвать в одних случаях переразвитие, а в других — недоразвитие одной и той же части нервной системы. Далее, можно обнаружить сильную колеблемость распространения внешнего эффекта, зависимого от слабого гена — «экспрессивность» (Expressivitat). Наконец, «эвномия» (Eunomie) обозначает закономерную последовательность пространственного распространения признака, причем это новое понятие, по мысли Фогта, должно заменить понятие ортогенез Эймера, имеющее филогенетическое и телеологическое значение. Это понятие было введено в связи с учением Фогта о «патоклизе» (Pathoklise), т. е. свойстве отдельных топических единств (topistischen Einheiten) отвечать особенно легко определенными патологическими изменениями на специфические вредности, или вообще легче заболевать. Патоклиза может быть частной или общей; метаболической, поражающей единства в неправильной последовательности, и эвномической, характеризующейся определенной последовательностью их поражения и т. д. Понятие «биоклиза» (Bioklise) объединяет тот же круг явлений, относящийся к нормальной изменчивости, т. е. совпадает с понятием нормы реакции (См.: С. Vogt, О. Vogt, 1929).
КООРДИНАТЫ:
ПЕНЕТРАНТНОСТЬ, ЭКСПРЕССИВНОСТЬ, СПЕЦИФИЧНОСТЬ
Серию статей 1925—1934 гг. по фенотипической реализации гена Н. В. Тимофеева-Ресовского пронизывает стремление сделать отличающийся чрезвычайными «неправильностями» материал пригодным для строгого и достаточно точного анализа. В любой области знания это возможно лишь в том случае, когда в анализируемом материале выявлены элементарные структуры и элементарные явления. В математике, например, для описания определенной области выделяют некоторый базис — такой наименьший набор независимых переменных, что все разнообразие переменных изучаемой области может быть выражено в виде функций этих базисных переменных — координат базисного пространства. Главным итогом этого
48
<цикла работ стало вычленение Тимофеевым-Ресовским трех феноменов Проявления гена (пенетрантность, экспрессивность, специфичность) и построение на этой основе общей схемы проявления гена.
^Объектом первой работы этого цикла (Тимофеев-Ресовский, 1925) послужила мутация radius incompletus (ri), обнаруженная в ходе учебной работы летом 1923 г. в нормальной «чистокровной» культуре D. funebris, которая происходила от самки-родоначальницы, взятой из дикой популяции Гидрофизиологической станции близ Звенигорода. Эта мутация рецессивна, аутосомна, ее фенотипическим выражением является нехватка дистального конца 2-й продольный жилки (radius) на крыльях гомозиготной мухи. В фенотипическом выражении и проявлении признак весьма изменчив: может варьировать степень недохождения жилки до края крыла, у некоторых особей оставшаяся часть жилки может быть у дистального конца прервана, признак может выражаться несимметрично. Кроме того, в гомозиготных по ri линиях могут попадаться фенотипически неизмененные мухи.
Пенетрантность (проявление). Предметом изучения в первой работе была степень проявления (пенетрантность) признака, которая определялась процентом измененных мух в гомозиготных по ri культурах. Для этого посредством отбора и строгого инбридинга был получен ряд практически «чистокровных» (высоко гомозиготных по всему комплексу генов и более не поддающихся отбору) различных культур, каждая из которых характеризовалась определенной «силой проявления» этого гена. Самые «слабые» линии давали 39,4%, 36%, 34% непроявления, были вполне рецессивны и при расщеплении часто отставали от отношения 3:1. Самые сильные давали все 100% фенотипического проявления в чистокровной культуре и около 6% (а одна линия даже 17%) доминирования в Fj от скрещивания с нормальными линиями L5 и К II. «Таким образом, на основании имевшегося у меня материала пределами «силы проявления» radius incompletus в разных генотипах надо считать, с одной стороны, около 40% непроявления, а с другой — около 15% доминирования» (Тимофеев-Ресовский, 1925, с. 134).
Общий обзор всех полученных линий ri, результатов скрещиваний их друг с другом и опытов с отбором на «силу проявления», а также картина возникновения линий ri дают указание на то, что генотипические различия этих линий полигибридны. «Специфическим «усиливающим» или «ослабляющим» действием на проявление radius incompletus в большинстве случаев обладают не отдельные гены, а определенные генотипы, т. е. комбинации генов» (там же, с. 134).
Является ли доминантность свойством гена? Поскольку в ряде линий ri наблюдался переход в частично-доминантное состояние, встал вопрос: справедливо ли представление, что доминантность (рецессивность) является фундаментальным свойством гена? Среди линий ri, полученных в работе 1925 г., были такие, которые доминируют при скрещивании с обеими нормальными линиями, L5 и КП; были и такие линии ri, которые доминируют при скрещиваниях только с одной из них. Одна из линий (D) давала постоянно некоторый процент фенотипически нормальных мух (около 6%) и, несмотря на это, доминировала (около 5%) при скрещиваниях с нормальной линией КП. Генотип линии D не настолько благоприятен для ri, чтобы давать 100%-ную пенетрантность признака; в линии КП также нет генов, безусловно усиливающих экспрессивность
4 В. В. Бабков 49
ri, что видно по скрещиваниям КП с другими линиями ri. Очевидно/ именно комбинация ряда генов линий D и КП создает благоприятн^ для проявления генов ri генотип. На этом экспериментальном основании Тимофеев-Ресовский пришел к выводу об операциональности пон/тия доминантности: «доминантность или рецессивность не являются специфическими свойствами самого гена, а зависят от того, на какую генотипическую почву попадает этот ген. Тот или иной генотип делает, так сказать, данный ген или рецессивным или доминантным* (Тимофеев-Ресовский, 1925, с. 134). Вывод о влиянии генотипической среды на степень доминантности гена, сделанный Тимофеевым-Ресовским, явился конкретизацией концепции генотипической среды. Он хорошо согласуется как с теорией эволюции доминантности Р. Фишера (Fischer, 1930, 1931), так и с представлениями И. И. Шмальгаузена об эволюции рецессивности и шире — об изменчивости выражения мутаций в процессе эволюции (Шмальгаузен, 1938, 1946).
Экспрессивность (выражение). Другая рецессивная аутосомная мутация D. funebris, venae transversae incompletae (vti), которая в присутствии ri вызывает перерывы или полное исчезновение поперечных жилок крыла у гомозиготных мух, оказалась благоприятным материалом для количественного учета степени выражения признака и особенно для учета специфичности.
Исследование Тимофеева-Ресовского показало, что отбор, собирающий гены-модификаторы, может оказывать значительное влияние на реализацию гена vti. Культура vti скрещивалась с лабораторными и природными культурами разного географического происхождения (Москва, Ленинград, Киев, Центральная Россия, Саратов, Крым, Кавказ, Берлин, Потсдам, Париж, Испания, Америка): последующим отбором и инбридингом (на протяжении 25—50 поколений) был получен набор практически чистых культур, в которых один и тот же ген vti находился в различных генотипических средах. В ходе отбора подвергались влиянию не только интенсивность, но и характер видимого осуществления гена — специфичность. Самое важное то, что различные феномены осуществления гена могли испытывать такое влияние независимо друг от друга.
Каждая из 30 линий vti, описанных в итоговой статье «О влиянии генотипической среды и внешних условий на реализацию генотипа* (Timofeeff-Ressovsky, 1934а), характеризовалась определенной степенью пенетрантности и экспрессивности и определенной формой специфичности (табл. 4). Эти характеристики более не были подвержены действию отбора. Так, пенетрантность в каждом новом поколении оставалась на том же уровне, независимо от того, получалось ли это поколение от фенотипически нормальных мух или от мух с крайней выраженностью признака vti. Тот факт, что пенетрантность может варьировать по культурам от 40 до 100%, указывал, что порог реактивности, который следует перейти для «включения* определяемого геном формообразовательного процесса, зависит от генотипической среды, в которой находится этот ген.
Экспрессивность, подсчитанная как процент экземпляров, полностью лишенных 2-й поперечной жилки, варьировала в разных культурах от 12 до 100%. Эти данные позволили дать количественную оценку для ответа на вопрос, поставленный в статье 1925 г., где Тимофеев-Ресовский отметил, что «сила фенотипического проявления и сила фенотипического выражения признака radius incompletus могут быть независимы друг от 50
Таблица 4
ггенсивность (пенетрантность и экспрессивность) проявления vti в 30 различных Прошедших отбор и длительный инбридинг линиях, расположенных в порядке
нарастания пенетрантности
Номе|1 Культура Число левых крыльев Пенетрантность, % Экспрессивность, %
Классы • Всего
- 1 1 2
1 № 20 252 154 22 426 41,1 12,5
2 № 27 157 132 47 336 46,7 26,2
3 № 33 171 138 80 389 56,0 36,7
4 № 3 168 107 111 386 56,5 50,9
5 № 85 148 106 92 346 57,2 46,5
6 «Н» 221 89 232 542 59,2 72,3
7 «F» 166 149 119 434 61,7 44,4
8 <11» 142 193 88 423 66,4 31,3
9 № 32 163 127 214 514 67,6 62,7
10 «Е» 161 221 173 555 71,0 43,9
11 № 17 138 181 203 522 73,6 52,8
12 № 37 107 261 98 466 77,0 27,3
13 «А» 98 164 200 462 78,8 55,0
14 <К> 95 45 386 481 80,3 88,3
15 № 34 78 152 183 413 81,1 54,6
16 «Ш» 53 81 187 321 83,5 69,6
17 № 35 57 243 145 465 87.2 35,5
18 «D» 48 227 261 536 91,0 53,5
19 «С» 30 91 265 386 92,2 74,4
20 № 7 13 151 287 451 97,1 65,5
21 № 215 5 124 229 358 98,6 64,9
22 «L» — 316 131 447 100 29,3
23 № 13 — 216 193 409 100 47,1
24 «В» — 195 208 403 100 51,6
25 «I» — 106 267 373 100 71,6
26 № 88 — 112 351 463 100 75,8
27 <1V> — 81 331 412 100 80,3
28 № 31 — 73 349 422 100 84,7
29 № 225 — 4 399 403 100 99
30 № 235 — 1 427 428 100 100
* Классы означают: « + » норма; «1» — перерыв 2-й поперечной жилки; «2» — полная ее утеря.
друга» (1925, с. 133). (Среди 15 ri-линий здесь были линии неполнопене-трантные, 85—90%, и одновременно сильно экспрессивные, среди же полно-пенетрантных были линии сильно-, средне- и слабоэкспрессивные). Интенсивность и скорость реализации формообразовательного процесса, перешедшего порог реактивности, оказываются зависимыми от разных генов (ср. ниже рис. 1). Поведение пенетрантности и экспрессивности в генотипически различных культурах не параллельно: в 9 полнопенетрантных vti-культурах экспрессивность варьирует от 30 до 100%; имеются и все другие возможные комбинации (см. табл. 4).
На независимость пенетрантности и экспрессивности указывает также результат температурных опытов с сильно реагирующей линией «А». Экспериментальный анализ выявил два чувствительных периода в онтогенезе мух. В первом с повышением температуры сильно понижается пенетрантность vti (и лишь незначительно экспрессивность); во втором сильно
4*
51
повышается экспрессивность (и лишь незначительно пенетрантность) (Тимофеев-Ресовский, 1928, 1930; сводка результатов: Timofeeff-Ressovsky, 1934а). /
Высокая степень независимости друг от друга пенетрантности и экспрессивности позволяет выбрать их в качестве двух координат для изучения изменчивости осуществления наследственных признаков. Эта изменчивость, однако, затрагивает не только интенсивность, но и образ фенотипического проявления.
Третье измерение: специфичность. Говоря о способе видимого осуществления признака, Тимофеев-Ресовский касается трех феноменов, исследованных Астауровым (см. гл. 7), Рокицким (гл. 8), Царапкиным (гл. 10). Это: симметрия проявления (на обоих крыльях), поля действия (распространение признака на одну или обе поперечные жилки) и эвномия (модус изменчивости перерыва 2-й жилки). Два последних Тимофеев-Ресовский объединил понятием специфичности проявления гена.
В некоторых vti-линиях, где прервана или утрачена 2-я поперечная жилка, была затронута также и 1-я. Поэтому встал вопрос, чем определяется влияние гена vti на 1-ю жилку: более сильной ли экспрессивностью или же отдельным потенциальным полем действия (различной интенсивности в разных культурах). Для решения вопроса отбирались, во-первых, сильно экспрессивные крылья, где прервана только 2-я жилка («А>-тип), во-вторых, крылья с обеими прерванными жилками («В>-тип). Результаты опытов с отбором показали, что эти два параметра проявления гена совершенно независимы. Например, линия № 235 («А>-тип) имела 100%-ные пенетрантность и экспрессивность, т. е. у всех мух утеряна 2-я жилка и не затронута 1-я. Напротив, линия № 27 слабопенетрантная (47%), слабоэкспрессивная (26%), и все же у многих мух прервана 1-я жилка. Значит, для vti существуют два потенциальных поля действия, определяемых генотипической средой.
2-я поперечная жилка может прерываться: «а> по направлению от анальной к кубитальной, «Ь> — в обратном направлении, <с» — от обоих концов к середине. Такие тонкие различия способа реагирования отдельной области поля действия vti (модус изменчивости) также оказываются наследственно детерминированными.
Отношения симметрии. Тимофеев-Ресовский строит здесь также общую схему типов отношения симметрии — для случаев, где вероятность проявления справа и слева теоретически одинакова:
1. Симметрия проявления, т. е. случай абсолютной (или очень сильной) право-левосторонней корреляции: rds= + l.
2. Диссимметрия; проявление на одной стороне несколько увеличивает шансы на проявление на другой: 0<rds<l.
3. Асимметрия; вероятности проявления справа и слева равны, но осуществление с одной стороны независимо от другой, так что симметричные проявления или непроявления являются результатом случайного комбинирования право- и левостороннего проявления — непроявления: rdS = 0-
4. Дисантисимметрия: — l<rds<0.
5. Антисимметрия; абсолютная (очень сильная) отрицательная корреляция сторон: rds= — 1.
Типы отношений симметрии характеризуют, что существенно, не отдельный организм, а популяцию. (Например, муха с полным развитием 52
Таблица 5
^водка данных о проявлении vti в 30 прошедших отбор и инбридинг линиях
НоМ^р Культура Пенетрантность, % Экспрессивность, % Поля действия Модус изменчивости Симметрия
1 № 20 45 15 В а± As
2 № 27 45 25 Вц с As
3 № 33 55 35 В b Ds
4 № 85 55 45 Bi а As
5 № 3 55 50 А с As
6 «Н» 55 75 В с± Ds
7 «11» 65 30 В а As
8 «F» 65 45 Bi Ь± As
9 № 32 65 65 А а± Ds
10 № 37 75 25 А As
11 «Е» 75 45 Вц а As
12 № 17 75 55 А b Ds
13 «А» 75 55 А а As
14 № 35 85 35 А а
15 № 34 85 55 Вц с As
16 «III» 85 70 Вц а
17 «К» 85 90 В| с± Ds
18 «D» 95 55 Вц с
19 № 7 95 65 А b
20 № 215 85 65 в. а
21 «С» 95 75 в. а
22 «L» 100 30 В с±
23 № 13 100 45 Bi а
24 «В» 100 50* Вц b
25 «I» 100 70 А с
26 № 88 100 75 Bi а±
27 «IV» 100 80 А а
28 № 31 100 85 Вц а
29 № 225 100 100 в. ?
30 № 235 100 100 А ?
Прим е ч а н и е. Процент пенетрантности и экспрессивности округлен до классов
5% н 10%.
признака на одной стороне тела и частичным или полным неразвитием на другой может, с разной вероятностью, встретиться в популяциях любого из этих пяти типов). Селекционные опыты показали, что также и тип отношений симметрии испытывает влияние наследственных факторов.
Опираясь на представления Астаурова (1927; Astauroff, 1930, см. гл. 4), Тимофеев-Ресовский указал, что в случаях симметричного проявления, когда изменчивость между организмами больше, чем изменчивость между сторонами в пределах организма, решающие факторы действуют на организм как целое, поэтому их можно искать среди гуморальных влияний. В случаях же асимметричного проявления решающие влияния на билатеральные зачатки органов должны быть чисто локальными и независимыми. Как он отметил, в этой связи интересно, что у дрозофилы все признаки окраски симметричны, тогда как большинство признаков, касающихся структур билатеральных органов (за исключением Ваг) асимметричны (по принципу
53
irenae transircrsae incomi
(radius incomi
ммш\
1ШШ\ !М\1М\1\\
1М\1М\Ж ouw\
1ШММ\ гм\\ы\ш 1ШУШ
Рис. 1. Влияние генотипической среды на пенетрантность, экспрессивность, специфичность (по: Timofeeff-Ressovsky, 1934а)
А — пенетрантность 100%, экспрессивная слабая, специфичность 1а; Б — пенетрантность 100%, экспрессивность сильная, специфичность 1а; В — пенетрантность 60%, экспрессивность сильная, специфичность 1а; Г — пенетрантность 100%, экспрессивность сильная, специфичность Па; Д — пенетрантность 100%, экспрессивность слабая, специфичность Па; Е — пенетрантность 100%, экспрессивность слабая, специфичность 1b
Дстаурова). Это различение можно сопоставить с двумя типами развития признаков, автономным и неавтономным sensu Sturtevant; во втором случае решающую роль играют «геногормоны>. Тимофеев отсюда выводит, что на первых же этапах работы имеет смысл провести чисто статистический анализ отношений симметрии проявления признака. Проявление, близкое к симметричному, укажет на большую роль общих для всего организма факторов; близкое к асимметричному — на роль локальных. Такие указания позволяют сузить область, где следует вести поиски стартовых механизмов (auslosenden Ursachen), запускающих формообразовательный процесс, который приводит к развитию данного признака.
В табл. 5 обобщены результаты изучения 30 vti-культур в отношении пенетрантности, экспрессивности, полей действия, модуса изменчивости и типа симметрии; отношения этих же характеристик даны на рис. 1. (Ниже на рис. 3 представлено географическое распределение модификаторов, обусловливающих три модуса изменчивости.) «Общим итогом селекционных исследований является положение, что определяемый мутантным геном формообразовательный процесс подвергается многообразным наследственным влияниям со стороны других генов. Здесь мы приходим (в модернизированной форме) к взгляду о единстве организма и его развития. Что касается феноменологии, то подтверждено и доказано различие пенетрантности, экспрессивности и специфичности в проявлении гена. Поэтому во всех случаях, когда изучается изменчивость проявления гена, следует учитывать все три стороны проявления гена, а не рассматривать отдельно экспрессивность или среднюю степень выраженности признака» (Timofeeff-Ressovsky, 1934а, S. 88).
ОБЩАЯ СХЕМА ПРОЯВЛЕНИЯ ГЕНОВ
Понятие «наследственной конституции». Первое же исследование изменчивости проявления и степени доминантности ri привело Тимофеева-Ресовского к формулировке понятия наследственной конституции. Значение наследственной конституции он видел, во-первых, в частичной зависимости генов друг от друга при их проявлении и в связи с этим в наследственном предрасположении к определенной форме фенотипического проявления наследственных признаков; во-вторых, в образовании наследственного предрасположения к внешним воздействиям (болезни, температурные воздействия, яды и т. д.); и, в-третьих, в том, что в определенной наследственной конституции может присутствовать наследственный же признак (или признаки), который не как ген, а чисто фенотипически, как признак, может оказывать влияние на проявление ряда наследственных и ненаследственных признаков (Тимофеев-Ресовский, 1925, с. 139—140).
Эта формулировка получила подтверждение в серии работ с vti. Различные сбалансированные генные комплексы (sensu Mather), свойственные географическим разобщенным популяциям, обусловили различную специфичность vti, внесенного в их «наследственную конституцию». Далее, пенетрантность и экспрессивность vti сильно реагировали на температуру только в некоторых культурах и практически не реагировали в других. Наконец, третье положение понятия наследственной конституции охватывало группу явлений, к которым, в частности, относилось образное представление А. С. Серебровского (1939) о «фенотипических окнах», открытие которых позволяет проявиться ряду генов (Серебровский, 1973, с. 31). Благодаря подобным работам стала возможна конструктивная критика
55
к
Рис. 2. Общая схема осуществления признака (по: Timofeeff-Ressovsky, 1934а)
На пути от основного гена а к окончательному признаку А оказывается а, непосредственная онтогенетическая причина признака (порог реакции); внизу — разные гены-модификаторы: справа (///, k, I) — факторы, определяющие реакционную способность соответствующего органа (модификаторы специфичности), слева — факторы, определяющие пенетрантность (//, /: с, d, е) и экспрессивность (//,, 2: f, g, h), Различные внешние факторы, влияющие на интенсивность процесса осуществления, обозначены М, N, О
ряда генетических понятий и терминов, связанных в своем происхождении с редукционистской генетикой. «Что касается специфики проявления гена»,— отмечал М. М. Камшилов,— «будет ли он «действовать» на тот или иной признак, будет ли он полно или неполно проявляться, будет ли он летальным или нелетальным, то размах и характер его изменчивости, выявленные в фенотипе, не являются присущими ему свойствами, а целиком определяются конкретным ходом индивидуального развития. Изменение конкретного хода индивидуального развития тем или иным способом, изменением ли генотипических соотношений или изменением условий развития, меняя проявление генов, меняет фенотипический характер наследственной изменчивости» (Камшилов, 1935, с. 139). Исходя из этих соображений, он возражал, например, против употребления термина «генотипическая среда» и выражения «взаимодействие генов в развитии». Из этого критического анализа вытекало, в частности, требование конкретизации общего понятия генотипической среды Четверикова; такой конкретизацией и стало тимо-феевское понятие наследственной конституции, принцип автономной изменчивости признаков Б. Л. Астаурова и концепция полей действия генов П. Ф. Рокицкого.
Общая схема проявления гена. В итоговой работе этого цикла Тимофеев-Ресовский делает попытку дать определенное, хотя бы и весьма схематическое и «гипотетическое представление осуществления гена» (Timofeeff-Ressovsky, 1934а, S. 100). Эта схема дана на рис. 2. На пути от основного гена а к конечному признаку А оказывается а, непосредственная онтогенетическая причина признака (или порог реакции, который следует перейти, чтобы стала возможной фенотипическая реализация данного признака). Внизу показаны разные гены-модификаторы, влияющие на данный формообразовательный процесс в разных точках: из них справа— (111 = =i, k, I) факторы, которые определяют не собственно процесс, а независимо
56
от него реакционную способность соответствующего органа или ткани; в этой группе можно представить себе модификаторы специфичности; слева — факторы, определяющие две стороны интенсивности: пенетрантность (//, 1 = с, d, е) и экспрессивность (II, 2 = f, g, h). Различные внешние факторы, влияющие на интенсивность процесса осуществления, обозначены М, N, О. «Ясно, что чем сильнее и специфичнее определяемый а формообразовательный процесс, тем ничтожнее относительное значение, которое будут иметь все другие модификаторы осуществления, и тем постояннее будет проявление данного гена» (Timofeeff-Ressovsky, 1934а, S. 101).
В первой попытке исследовать феноменологию реализации гена Тимофеев-Ресовский имел дело с материалом, пригодным для выявления лишь пенетрантности, так что описываемые там события разыгрывались на уровне пороговой реакции. «Это можно сравнить со стрелком, стреляющим в цель из ружей разного качества — из хорошего ружья он попадает в цель в большем проценте случаев, а из скверного — в меньшем». Так Тимофеев-Ресовский (1925, с. 138) определял стохастический, т. е. целенаправленный со случайным разбросом характер процессов реализации гена.
Как видно из этой схемы, все факторы, влияющие на проявление, представляют собой как бы некоторую среду, в которой обнаруживается данный ген. «Эта среда может быть расчленена на три части: «генотипическая среда», к которой относятся все вовлеченные в процесс модификаторы, «внешняя среда» и «внутренняя среда», в которой развивается признак» (Timofeeff-Ressovsky, 1934а, S. 101). Заключая, он подчеркивает: «В каждом случае при рассмотрении проявления гена мне представляется целесообразным прежде всего схематично разграничить генотипическую, внутреннюю и внешнюю среду» (loc. cit.; см. также: Timofeeff-Ressovsky, 1934b).
Эти идеи прочно вошли в учебную литературу (например, Waddington, 1939, р. 188—191). В капитальной сводке по истории эволюционизма Ф. Фозегилл отметил, что «концепция взаимодействия генов была стабилизирована работой Тимофеева-Ресовского» (Fothergill, 1952, р. 237).
ЗНАЧЕНИЕ ДЛЯ МЕДИЦИНСКОЙ ГЕНЕТИКИ
В «Заключительном обзоре» статьи 1934 г. Тимофеев-Ресовский пишет: «Знание и учет по отдельности феноменов проявления генов имеет, помимо того, известное прикладное значение для генетики неблагоприятных для анализа объектов, прежде всего для наследственных заболеваний человека. Предпосылкой для сколько-нибудь точного анализа нормальных и патологических признаков у человека является генетически правильная классификация материала. Эта задача особенно осложняется сильной изменчивостью соответствующих признаков, и здесь может иметь большое значение учет изменчивого осуществления гена» (Timofeeff-Ressovsky, 1934а, S. 102). Предлагая картину географического распределения модификаторов специфичности vti (рис. 3) и отмечая, что эти наблюдения, слишком фрагментарные и полученные на весьма ограниченном материале, приводятся лишь как методологическое основание для исследования таким способом фенотипически скрытых конституционных различий, автор замечает: «у человека возможно подобное изучение, например, географической изменчивости протекания распространенных широко заболеваний» (Там же, с. 103). Эту мысль
57
Рис. 3. Географическое распределение генов-модификаторов, определяющих три модуса исчезновения второй поперечной жилки у линии vti
/ — модус а\ 2 — модус Ь, 3 — модус с (по Timofeeff-Ressovsky, 1934а) с полным правом можно назвать пионерской; разработка и проведение гено- и феногеографических исследований становится сейчас одной из важнейших задач генетики человека.
Соображения Фогтов и Тимофеевых-Ресовских об идио-соматических группах созвучны мыслям, которые развивал С. Н. Давиденков в своем классическом труде «Эволюционно-генетические проблемы в невропатологии* (1947). Сходные взгляды недавно опубликовали клиницист Л. Г Калмыкова (1976) и генетик В. П. Эфроимсон (1978). Первый вариант общей схемы реализации гена Е. А. и Н. В. Тимофеевы-Ресовские предложили в 1926 г. для объяснения «идио-соматических групп изменчивости*. В систематической работе каждый вид (в зоологии, ботанике и медицине) основывает структуру систематических групп (рас и других мелких систематических категорий в ботанике и зоологии; болезней, как определенных фенотипических групп, в медицине) на объединении в одну категорию экземпляров, сходных относительно определенных фенотипических признаков. «Обстоятельный анализ подобной группы,— пишут авторы,— может часто привести к обнаружению ее гетерогенности, т. е. показать, что характерный для данной группы признак в разных случаях может быть разного происхождения и определяться разными причинами. В такие группы могут быть объединены различные, сходные друг с другом в своем внешнем проявлении факторы, хотя бы и чисто соматические, различия которых определяются внешними причинами* (Н. A. Timofeeff-Resovsky, N. W. Timofeeff-Ressovsky, 1926, S. 146) 1
Термин принадлежал Сесиль и Оскару Фогтам (С. Vogt, О. Vogt, 1926), как и некоторые другие термины, использованные Тимофеевым-Ресовским. Под названием Status marmoratus они описали заболевание стриатума, которое во всех случаях дает сходную и определенную клиническую патолого-анатомическую картину. Этиологический анализ показал, что эти заболевания, наследственные в одних случаях, в других появляются как чисто соматические, определяемые внешними причинами. Для этого
1 Д. Д. Ромашов и Е. И. Балкашина (1929—1931) критически относились к зачислению Тимофеевыми-Ресовскими ряда уклонений в разряд «ненаследственных*, предпочитая говорить о «нерасщепляющихся изменениях*.
58
и двух других заболеваний (также гетерогенных и образующих внешне однородные группы) Фогты и ввели название «идио-соматическая группа заболеваний* (idiosomatische Krankheitsgruppe). В общем случае, без специального внимания к патологии, Тимофеевы-Ресовские назвали такие группы <идио-соматические группы изменчивости* (idiosomatischen Variationsgrup-реп) 2. Общую причину идио-соматических групп изменчивости внутри вида и межвидового параллелизма изменчивости (закон гомологической изменчивости Н. И. Вавилова) они — как и Балкашина и Ромашов (см. гл. 2) — видели в том, что «реакционная способность организмов ограничена и сходные системы реакций близких видов имеют сходную реакционную способность* (Н. A. Timofeeff-Ressovsky, N. W. Timofeeff-Ressovsky, 1926, S. 169; см. также: Timofeeff-Resovsky, Vogt, 1926).
В этой ранней работе следует отметить два момента. Склонность к объяснениям в терминах, близких к тем, что использует теория управляющих систем, была свойственна Тимофееву-Ресовскому и в другие периоды работы (как и другим четвериковцам и самому С. С. Четверикову). Эти настроения впоследствии отразились в формулировании представлений о системном подходе в биологии (Тимофеев-Ресовский, 1970 и др.; Тимофеев-Ресовский, Тюрюканов, 1966). Интерес к изучению изменчивости человека в норме и патологии был в данном случае обязан влиянию Фогтов. Этот пример говорит, однако, о том, что при стремлении к строгости формулируемых понятий Тимофееву-Ресовскому было свойственно широкое понимание программы построения эволюционной генетики. Он не ограничивался узкими рамками одной области знания с ее установленными схемами, а был способен найти источники вдохновения и в других областях. Эта черта вообще чрезвычайно характерна для московской школы эволюционной генетики.
Глава 4
ПРИНЦИП АСТАУРОВА: АВТОНОМНАЯ ИЗМЕНЧИВОСТЬ ПРИЗНАКОВ
«Мне всегда казалось, что в тех случаях, где природа уклоняется от правильного хода, мы легче можем проникнуть в ее страшные тайны...»
Э. Т А. ГОФМАН
С именем Б. Л. Астаурова мы связываем прежде всего его исследования на тутовом шелкопряде, ведшиеся с позиций цитогенетики, биологии развития, радиационной генетики (см.: Астауров, 19726, 1978). Шелкопрядом Астауров занялся в 1930 г., когда по совету Н. К. Кольцова он вслед за Н. К. Беляевым уехал в Среднеазиатский НИИ шелководства и шелко-ведения (г. Ташкент); с зимы 1935/1936 г. он продолжил эту работу в лаборатории механики развития Д. П. Филатова в ИЭБ.
2 Ср.: «Сходство фенотипов не всегда говорит о сходстве генотипов...» (Вавилов, 1931, с. 111); «Одинаковые изменения фенотипического порядка могут быть вызваны и разными генами» (Vavilov, 1922). Ю. А. Филипченко (1924) дал четкое разграничение понятий генотипического, экотипического, морфологического параллелизма в живой природе.
59
Борис Львович Астауров, ок. 1925 г.
Научную работу Астауров начал с участия в первом экспериментальном исследовании диких популяций дрозофил 1925—1926 гг., будучи студентом МГУ (1921 —1927 гг.) иодновременно работая в Генетической лаборатории Четверикова сперва в качестве сверхштатного сотрудника ИЭБ, а затем — научного сотрудника КЕПС. Свое первое цитогенетическое исследование он (вместе с С. Л. Фроловой) провел на дрозофиле. Оно было связано с обнаруженным чет-вериковцами фактом, что Drosophila obscura из окрестностей Звенигорода реп-
родуктивно изолирована от привезенного Вайнштейном из лаборатории Мёллера морфологически идентичного вида, который Стёртевант определил также как D. obscura. Выявив различия кариотипов двух видов, Астауров в 1929 г. дал американскому виду новое название, в честь Фроловой — D. pseudoobscura Frol. (Frolowa, Astau-row, 1929). Оно прижилось и благодаря работам лаборатории Добржан-ского сейчас широко известно. Астауров участвовал в двух энтомологических экспедициях Четверикова по Кольскому п-ву в 1925 и 1926 гг. и в двух экспедициях, организованных по инициативе Кольцова Московским генетическим отделом КЕПС для изучения межвидовой гибридизации одногорбого и двугорбого верблюдов: в 1928 г. по кочевьям Урдинского р-на Западного Казахстана (степи Букеевской Орды, Большое и Малое Богдо, озера Эльтон и Баскунчак) и в 1929 г. по кочевьям Западных Каракумов (от Усть-Урта до Копетдага и от Красногорска до Казанджика).
Широкий спектр зоологических интересов вообще чрезвычайно характерен для четвериковской группы. При четкости постановок задач он сыграл свою роль в ее успехе — как, впрочем, и резкое ограничение круга проблем обеспечило успех группе Моргана.
Признак tetraptera. Учебное задание на генетическом практикуме Четверикова — генетический анализ с определением группы сцепления и картированием мутации tetraptera (III, 51, 3), возникшей в учебной линии dumpy D. melanogaster в феврале 1925 г. (рис. 4, а),— послужило отправной точкой для первого самостоятельного исследования Астаурова. В этом исследовании Астауров наметил проблему генетической нестабильности, которая (в других аспектах) широко разрабатывается в наши дни, и четко очертил проблему эволюционного возникновения стабильности онтогенеза, которая позже стала ядром теории стабилизирующего отбора Шмаль-гаузена. В представлении об иерархичности организации биосистем получил развитие системный подход, присущий четвериковцам.
Проявление признака tr было неполным (при скрещиваниях с другими линиями вместо отношения Itr 3N получились отношения от 1:15 до 1 280). Фенотипическое выражение tr в «чистокровных» линиях, прошедших 20 поколений инбридинга и ведущихся в строго константных условиях, чрезвычайно изменчиво. Жужжальца (рудименты задних крыльев) могут развить-
60
Рис. 4. Мутации tetraptera
а — самец с хорошо развитым правым крылышком «2-й парыэ; б — крылышко «2-й парыэ; в — самец с крылышком на месте левого жужжальца
а, б — по: Астауров, 1927; в — рис. Е. И. Балкашиной, 1929
ся в крылья или стать более сильными рудиментами. Обратив сперва внимание на морфологию признака, Астауров (1927; Astauroff, 1929) выделил ряд переходных форм между нормальным жужжальцем и крылышком, причем каждая форма представляла собой вполне сформированный орган. Этот ряд начинается слабым изменением головки жужжальца (на ней появляются шипики и щетинки), на последующих ступенях покровы жужжальца с обеих сторон Приобретают прозрачность и структуру крыловой пластинки — они слипаются — головка увеличивается и сплющивается — увеличивается прозрачный участок — обособляется будущая крыловая пластинка — появляется первая, затем вторая жилка. Ряд продолжается и в другую сторону — до полной редукции жужжальца. (В статье «Изучение мозга и генетика» С. и О. Фогты, говоря о полярной противоположности, polare Gegensatzlichkeit, как об одной из черт генов со слабой специфичностью, ссылались на этот пример Астаурова. Они заметили, что «Значение подобного случая для изучения мозга можно понять, если представить, что один и тот же ген может вызывать как пере-, так и недоразвитие одной и той же части нервной системы»—С. Vogt, О. Vogt., 1929, S. 441; ср.: Timofeeff-Ressovsky, 1934с.) Полного эффекта tr достигает во вполне сформированном заднем крылышке с тремя хорошо развитыми жилками: 1-я жилка соответствует costa нормального крыла, 2-я — radius, 3-я — cubitus, отчасти media (рис. 4, б). Крайняя степень выражения tr весьма однообразна и одинакова в линиях разного географического происхождения (ср.: рис. 4, в изменение, возникшее в 1929 г. в потомстве самки из популяции Геленджика). Если длина нормального жужжальца составляет 350 мкм, то длина «заднего крылышка» — 905 мкм, т. е. треть
61
длины переднего крыла (2725 мкм) — отношение, близкое к тем, что можно встретить у перепончатокрылых. Астауров (1927) установил гомологию частей нормального жужжальца и различных степеней его изменения и рассматривал этот ряд форм как сравнительно-анатомический.
Гипотеза независимого осуществления признаков. Превращение жужжальца в «крылышко 2-й пары» представляло собой поразительный атавизм: возврат — одним мутационным скачком! — представителя отряда двукрылых к древнейшему строению четырехкрылого предка. Но предметом исследования Астаурова стало другое, не менее удивительное свойство tr. Неопределенность фенотипической реализации tr была характеристикой не только линии (неполное появление и варьирующее выражение), но и отдельной особи — два симметричных жужжальца вели себя по-разному в развитии этого признака: проявление сочеталось с непроявлением, одна степень выражения с другой.
Феномен неполного проявления к тому времени был известен четвери-ковцам благодаря изучению наследственной изменчивости D. funebris; Тимофеев-Ресовский начал в 1925 г. серию статей, в которой были выявлены основные параметры осуществления гена (пенетрантность, экспрессивность, специфичность). В то же время изменчивость конечного признака на левой и правой сторонах одного и того же билатерального организма, т. е. при тождестве генотипа и условий развития, представляла собой неожиданное явление. Оно противоречило общепринятому представлению, что конечный результат (обозначим его буквой а) процесса развития однозначно определяется начальным состоянием развивающейся системы и совокупностью внешних условий, в которых протекает развитие:
a = F (конституция, среда).
Для большинства «хороших» мутаций это действительно справедливо. Однако эта концепция недостаточна для объяснения фактов неполного и тем более асимметричного проявления.
Астауров несколько конкретизировал эту слишком общую формулу. Пусть р обозначает протоплазму, g— генотип, е — совокупность условий развития, i (medium internum) — некоторые «внутренние условия», локальная флуктуация которых вызывает самостоятельную изменчивость сходных признаков, например, симметричных. Тогда формула примет вид (Astauroff, 1930):
a = F(p, g, е, i).
Материал, собранный Астауровым, привел его к гипотезе, согласно которой «основной единицей, на которую влияет фактор, является не целый организм, но любая его половина» (1927, цит. по переизд.: Астауров, 1974 г., с. 32)1 * Независимость осуществления признака слева и справа подтверждают, например, результаты скрещивания 287111 в чистокровной линии — табл. 6 (по: Астауров, 1927), где « + » обозначает фенотипически неизмененных мух, «tr^» — проявление tr только справа, «tr— слева, «trds» — двустороннее проявление.
Из гипотезы Астаурова вытекало, что право-левосторонняя корреляция должна быть ничтожной. Это следствие противоречило выводам биомет-
1 Ч. Планкит (Plunkett, 1926) подсчитал число щетинок на «половину мухи» (half-
fly unit), не делая, впрочем, каких-либо выводов о природе асимметрии. Его мате-
риалы Астауров подробно обсудил в своей статье (Astauroff, 1930).
Таблица 6
Отсутствие право-левосторонней корреляции признаков (по: Астауров, 1927)
4- ,г</ tr.< trds Всего Процент одностороннего проявления Фактический процент симметричных Ожидаемый процент симметричных d
80 47 45 28 200 37,0 14,0 13,7 ±0,31
риков школы К. Пирсона, а также Г. Дункера и др. (см.: Пирсон, 1911, гл. Х)? показавшим на случайных выборках из популяций различных организмов почти абсолютную право-левостороннюю корреляцию количественных признаков билатерально-симметричных органов: г ds= 4-0,96. Случай tr в свете этих выводов должен выглядеть исключением.
Однако Астауров предположил, что случай tetraptera отражает общее правило: существование самостоятельной изменчивости, несводимой к влиянию наследственного и средового компонента и обусловленной случайной локальной флуктуацией морфогенетических процессов — «онтогенетическим шумом». Мы имеем полное право дать этому общему правилу название «принцип Астаурова».
Этот принцип послужил основой для объяснения явлений неполного проявления, варьирующего выражения, колеблемости областей действия генов и отсутствия корреляции признаков, относящихся к симметричным либо сериально-повторяющимся органам одного индивидуума. Следует отметить, что формула, связанная с принципом Астаурова, дает ясное указание пути, на котором следует искать причины корреляции сторон при условии самостоятельной их изменчивости. В генетически неоднородной выборке (при сильной вариации g) или в неодинаковых условиях жизни е (т. е. при тех условиях, при которых обычно формируются случайные выборки для статистического обследования) определенный морфогенетический ответ на эти факторы, F(g) или F(e), одинаковый на левой и правой сторонах, должен маскировать некоординированную изменчивость, зависящую от фактора Z.
Для проверки гипотезы Астауров проанализировал методами вариационной статистики результаты скрещиваний, в которых было просмотрено 3790 мух.
АНАЛИЗ МОДЕЛИ ЯВЛЕНИЯ
Проверка гипотезы. Астауров рассуждал следующим образом. Предположим, что имеется некоторое (большое) число N правых половинок организма и столько же левых. Из соображений симметрии примем, что частоты проявления признака на правых половинках, p^aJN, и на левых, Ps=asN, равны: pd = ps = p. (Здесь ad и as — абсолютные числа измененных правых и левых половинок. Если число мух, измененных только справа, обозначим b# только слева — Ь* симметрично измененных — с, то ad=bd+c; as~bs~{~c). Если без выбора, наугад, комбинировать правые и левые половинки, складывая их между собой попарно (правую с левой) и получая таким образом целые организмы (удовлетворяя тем самым гипотезе назависимого осуществления билатерально-симметричных признаков), то вероятность того, что мы выхватим обе половинки измененные и получим, таким образом, мУху с симметричным проявлением признака, будет равна произведению вероятностей проявления на каждой из сторон: pdps^=ad • ajN • N или Г-а2/№
63
Таблица 7
Частота проявления tr (по: Астауров, 1927)
Скрещивания trd trs Всего Фактический процент симметричных Ожидаемый процент симметричных d
Индивидуальные 111 112 726 8,95 7,63 4-1,32
Массовые, 1925 г. 77 90 492 12,81 9,79 4-3,02
Массовые, 1926 г. 387 411 2572 6,48 6,12 + 0,36
Всего 575 613 3790 9,41 7,85 + 1,56
Астауров обратился к примеру (табл. 6) скрещивания 2871П. Там ad=47 + 28; as = 45 + 28; N =200, откуда получаются эмпирические частоты одностороннего проявления:
pd= 75/200 = 0,375; ps=73/200 = 0,365.
За приближенное значение априорной вероятности, р0, примем среднюю арифметическую частот правого и левого проявления: ро = (р^ + р5)/2 = 0,370. Тогда вероятность появления симметричной особи равна р2; а математическое ожидание числа симметричных мух равно p2N, т. е. в данном случае 0,372 X200 = 27,38, что хорошо согласуется с реальным числом симметричных мух, равным 28 (см. табл. 6). Это совпадение подтверждает гипотезу Астаурова.
Здесь встречается чисто техническое затруднение: отдельные скрещивания включают ограниченное число мух, поэтому их эмпирические частоты будут сильно отличаться от тех априорных вероятностей, реализацией коих они являются. Для ответа на вопрос, как следует объединять данные по отдельным группам, Астауров (1927) рассмотрел ситуацию объединения двух равночисленных совокупностей особей с разными вероятностями одностороннего проявления:
p' = a'/N и p"=a"/N.
На основе гипотезы независимого осуществления он получил выражения для эмпирической частоты составной совокупности и ее априорной вероятности. В общем случае первая величина больше второй и определяющее их неравенство превращается в равенство в случае а' = а"
Так Астауров показал некорректностть расчетов на основе неоднородного материала, который представляет собой один из источников ложной корреляции (spurious correlation — К. Pearson): «при расчете по суммарному итогу мы находим симметричных особей больше, чем их ожидалось при допущении независимого определения признака на разных сторонах; получается некоторая положительная корреляция сторон, и вследствие неправильного метода подсчета мы приходим к заключению о неправильности основной предпосылки» (цит. по: Астауров, 19746, с. 34—35) 2.
Найдя выражение для эмпирической частоты симметричного проявления
2 Ср. гл. <О ложной корреляции» (Н. С. Четвериков, 1975, с. 298—318), где автор выделяет пять случаев возникновения ложной корреляции и под названием «стратификация» рассматривает случай, приведенный Астауровым.
64
I/ 1
Рис. 5. Независимое осуществление признака на разных сторонах (по: Астауров, 1927)
а — графическое представление вероятностей симметричного, асимметричного и т. д. проявления как функций вероятности одностороннего проявления а; б — зависимость вероятностей (или %) проявления разных классов мух от вероятнос/и одностороннего проявления а признака в составной совокупности, Астауров (1927) свел полученные им результаты в таблицу (табл. 7). Были найдены близкие совпадения ожидаемых и фактически полученных чисел как по отдельным скрещиваниям, так и в итогах всех скрещиваний. Для каждого отдельного скрещивания, имеющего более чем по 75 мух каждого пола, были вычислены разницы между априорными и эмпирическими вероятностями, и оказалось, что они заведомо лежали в пределах вероятных отклонений. Это дало окончательное доказательство отсутствия право-левосторонней корреляции в развитии признака tr.
Исследование математической модели. Итак, основной единицей для расчетов вероятности проявления должна сложить половина особи, «half-fly unit». Если
Исследование математической модели. Итак, основной единицей для расчетов вероятности проявления должна служить половина особи, «half-fly unit». Если в сово-вокупности N особей (2W половинок), у которых изменено ad правых и as левых сторон, то вероятность проявления признака выражается через (ad + as) /2N или (bd+ 6S + 2с)/2N, а не через (bd + bs + с) /N, если характеризовать проявление признака отношением числа измененных особей к их общему числу.
Вероятность проявления tr колебалась от 0,05 до 0,50; поэтому интересно выяснить, в какой зависимости стоят величины различных фенотипических классов от изменения вероятности проявления признака. Единичный отрезок АВ (рис. 5, а) изображает изучаемую совокупность, достаточно большую, чтобы эмпирические частоты были близки к априорным частотам. Тогда отрезки a^=as=a дают вероятность одностороннего проявления признака, на правой и левой сторонах. Их пересечение, с~а 2, дает частоту фенотипического класса симметрично измененных мух. Отрезки bd=bs=b представляют частоты классов с изменением только на правой стороне (bd) и только на левой (bs)\ b=a—c=(\—a). Частоту класса несимметрично (все равно как) измененных мух покажет сумма b ^Ь =2Ь=2а(\—а). Частота класса любым образом измененных мух будет d=b^bs+c=a(2—а). Наконец, класс фенотипически неизмененных мух представлен отрезком е=1—d= 1—а(2—а)= (1—а) 2.
В результате частоты всех фенотипических классов мух представлены как функции а, вероятности одностороннего проявления:
симметричное проявление (1) проявление на одной определенной стороне (2)
несимметричное проявление (3)
какое-либо проявление (4)
полное непроявление (5)
В. в. Бабков
с=а 2
Ь=а(1 —а)
2Ь=2а(1—а)
d=(2-a)
е = 1 - а\2 - а) = (1 - а)2
Эти функциональные зависимости частот фенотипических классов мух от вероятности одностороннего проявления а графически представлены на рис. 5, б. Проведя качественное исследование уравнений (1) и (5), либо даже просто рассматривая соответствующие им кривые c = fj(a) и e = f5(a) на рис. 5,6, можно заключить, что при предельных значениях а вся совокупность будет состоять из особей одного фенотипического класса: при а=0 из неизмененных, а при а=1 из симметрично измененных особей. С возрастанием а от 0 до 1 монотонно растет доля класса симметрично измененных (с = а2) и любым образом измененных — d = a(2—а) особей (хотя характер этих двух кривых различен) и убывает доля неизмененных — е= 1— а(2—а). Численность класса односторонне измененных достигает максимума при a = 0,5 и затем убывает.
Следует обратить внимание на то, что при небольших значениях а (например, при 0<а<0,3) асимметричные особи преобладают среди измененных: при a = 0,3 асимметричные составляют 83% всех измененных и 42% от всей совокупности, а симметрично измененные — только 9% от всей совокупности. (При a = 0,1 соответственно 95, 16, 1%). Это согласуется с фактами связи ассиметричного проявления с неполным и говорит за то, что непроявление признака у организма в целом является функцией процессов непроявления, идущих с той или иной степенью независимости на разных сторонах. Как правило, неполно проявляющиеся признаки с низким про центом измененных особей дают большой процент асимметричных форм.
Формулы (1) — (5) справедливы для случая независимого осуществления признака на разных сторонах организма. Но нет нужды делать вывод, что для любых признаков, имеющих неполное и асимметричное проявление, отношения фенотипических классов должны определяться этими формулами. Мыслимы ситуации, когда факт проявления признака на одной стороне уменьшает вероятность проявления на другой (дисантисимметрия,— l<rds<0) или увеличивает ее (дисимметрия, 0<rds< 1). Сравнивая фактическое число симметричных особей (с) с ожидаемым в случае асимметрии (а2), легко выяснить, к какому типу отношений симметрии принадлежит данный признак. Если для асимметрии справедливо с = а2, то при дисимметрии с>а2, а при дисантисимметрии с<а2
Принятие в качестве основной величины, характеризующей проявляемость при-
6^+ b s+2c
знака, именно вероятности одностороннего проявления -2—------, а не доли
к . bd+bs+c . 2/V
измененных особей—° -— *— помимо формального, имеет также и известное
практическое значение при изучении некоторых моментов, связанных с явлением неполного проявления. Один из них — частичная ограниченность полом, когда признак проявляется с разной силой и в разном проценте у особей разных полов. Так, tr чаще проявляется у самцов (признак частично ограничен мужским полом); так же ведет себя radius incompletus у D. funebris в исследованиях. 1925 г. Тимофеева-Ресовского; напротив, Extra veins Лутца (Lutz, 1911) чаще проявляются у самок. Тимофеев-Ресовский ввел для учета этого момента «индекс ограниченности полом» — отношение фенотипически нормальных самок в культуре к нормальным (^самки \
-------> 1 I Лутц — такое же отношение для измененных мух ^самцы-'
(^самки \
—----- > 1 I Тимофеев нашел, что при возрастании общего процента изме-
^самки '
ненных мух от 67 до 94% индекс возрастает от 2,4 до 5,2 (у Лутца индекс падает, стремясь к единице). Астауров (1927) обсуждает вопрос: за счет чего изменяется индекс при переходе от одной культуры к другой, с иным процентом измененных мух? На графиках рис. 5, б видно, то функции d=f4(a) и e = f5(a) почти линейны при небольших величинах вероятности одностороннего проявления (скажем, при а<0,3) В этих пределах вычисления индекса на основе величины а или due практически равноценны. В общем случае, однако, вычисления индекса на основе d или е будут подвержены сильным влияниям нелинейности функции d = a(2—а) и е= \ —а(2—а).
66
УНИВЕРСАЛЬНОСТЬ АВТОНОМНОЙ ИЗМЕНЧИВОСТИ
Установленный Астауровым факт полного отсутствия координированности в развитии признака tr на разных сторонах одной и той же особи (rds = 0) на первый взгляд вступает в противоречие с очевидностью — в наших повседневных и повсюдных наблюдениях мы находим, в качестве общего правила, зеркальное сходство у разнообразных билатерально-симметричных организмов. Поэтому было важно выяснить распространенность этого явления и его место среди общебиологических представлений.
Астауров перечисляет аналогичные факты из области изменчивости мутационных и нормальных видов признаков Drosophila. Признак eyeless (либо наличие рудиментарного глаза, либо полное его отсутствие), по данным Гатри (Guthrie, 1925), не дает заметной корреляции сторон. Ясную картину отсутствия корреляции сторон представляет случай Dichaeta (Plunkett, 1926); при этом значительная корреляция появляется в гетерогенной линии — в полном соответствии со взглядами Астаурова на причины возникновения /южной корреляции. Напротив, Лутц (Lutz, 1911) получил высокую корреляцию сторон для Extra veins (добавочные жилки). Однако Лутц производил подсчет по суммарному итогу многих скрещиваний. Для выяснения реальности корреляции Астауров предпринял исследование линии Extra veins (Ramuli), происходящей из геленджикской популяции D. melanogaster. Он выяснил, что корреляция изменчивости, как и сама изменчивость, обязана в этом случае почти исключительно колеблемости внешних условий (фактор е); при поддержании постоянных условий жизни эта корреляция исчезла, но небольшая самостоятельная изменчивость сохранилась. Признаки vti (перерыв 2-й поперечной жилки) и Ramuli, а также Polychaeta (sin. Extra bristles — добавочные щетинки на тораксе) сходны в ряде отношений. Они чрезвычайно широко распространены в свободно живущих популяциях; подобные изменения находят, например, у разных видов Hymenoptera (пчела, Habrobracon, и др.), у которых они не выходят за пределы нормальной видовой изменчивости жилкования. Такие признаки представляют собой как бы промежуточную ступень между «хорошими» лабораторными мутациями и нормальными видовыми признаками и одновременно являются границей между качественными и количественными признаками.
Число стерно-плевральных щетинок у D. melanogaster, наследование которых изучал П. Ф. Рокицкий (1927), нормальный видовой признак, обнаруживает частые и значительные асимметрии. Астауров (Astauroff, 1930) просчитал часть из его неопубликованных материалов и получил незначительную корреляцию сторон, в среднем rds= +0,222. (Так, в двух линиях плюс- и минус-отбора, № 252 и № 817, коэффициенты корреляции таковы: rds = + 0,11; rds= +0,15. Если же вычислить корреляцию для объединенной культуры, № 252+ № 817, то получим очень высокую корреляцию: fds=+0,83. Этот пример бросает свет на причины корреляции сторон в генетически гетерогенной выборке или выборке особей, воспитывавшихся в гетерогенной среде. Очевидно, что корреляция обязана тому, что меняющийся от особи к особи генотип или/и внешняя среда влияет сходным °бразом на одинаковые образования (антимеры) тела и вызывает одинаковость их окончательного строения.)
Асимметрично проявляющиеся признаки в значительном числе обнаруживались при анализе генетической гетерогенности природных популяций видов рода Drosophila (melanogaster, transversa, phalerata, obscura, vibris-
sina, funebris), который велся под руководством С. С. Четверикова (см. гл. 2). Эти признаки затрагивают любые системы органов; очень часто это признаки жилкования, «вырезки» и изменения в положении крыльев, изменения хетотаксиса, деформации глаз, нарушения в расположении омматидиев и т. д.— то, что Ромашов называл «массовыми популяционными изменениями».
Астауров (Astauroff, 1930) исследовал также самостоятельную изменчивость сходных органов (undifferentiated like organs — К. Pearson) у Polychaeta (Nephthys caeca и Glycera goesti) и Miriapoda (G eophilus ferrugineus). Ценность полихет и многоножек как объекта исследования в том, что одни и те же органы многократно повторяются в пределах особи — при постоянстве генотипа и внешней среды; сегмент в пределах особи выступает как индивидуум низшего порядка. Поэтому и вопрос — обнаружат ли корреляцию числа щетинок на параподиях полихет (или на ножках многоножек) противоположных сторон одного сегмента (или на соседних параподиях/ножках одной стороны) — вопрос того же рода, что и поиски корреляции числа стерно-плевральных щетинок на разных сторонах мухи. (Правда, этот материал добавил новую методическую трудность. Признак закономерно меняется по длине тела, так что при подсчете корреляции следует исключить эволюторную компоненту, или тренд: «закономерную изменчивость», зависящую от положения параподия в сериальном ряду. При подсчетах пришлось прибегнуть к методу «скользящего выравнивания кривой» — см. раздел «Сглаживание динамических рядов» в книге Н. С. Четверикова, 1975).
И здесь в развитии признака на правой и левой сторонах одного сегмента, равно и на соседних сегментах, было установлено отсутствие связи: «В осуществлении признаков в процессе развития абсолютной детерминированности (предопределенности) нет; совокупность организации и условий среды оставляет в процессах своего взаимодействия во времени (т. е. в процессах развития) место для случайных колебаний» (цит. по: Астауров, 19746, с. 83).
Возможность'самостоятельной изменчивости (соответственно, неполного проявления) всегда следует иметь в виду, когда встречаются асимметрии в проявлении признака у билатерально-симметричных организмов. Это в полной мере относится к пегостям (признак, о котором не скажешь, качественной или количественной природы его вариация), полидактилиям, полимастиям и пр. Запутанные картины неправильного наследования, осложняемые фенотипическим полиморфизмом, часто наводят исследователя на мысль искать причины этого многообразия в одной лишь генотипической основе, пусть сколь угодно сложной. При гибридологическом анализе едва ли не любые отношения могут быть объяснены (к тому же не единственным способом) тем или иным числом взаимодействующих факторов; так возникают формальные бессодержательные схемы.
Тем временем представление о самостоятельной фенотипической изменчивости может сыграть плодотворную роль в анализе наследования пегостей, поскольку имеется ряд указаний на чисто фенотипическую вариацию признака, реализующегося в причудливом распределении пигментированных и непигментированных участков при общей тенденции к симметричной картине. Астауров (Astauroff, 1930) рассматривает, в частности, материалы работы Н. А. Ильина (1928) о белой пятнистости у морских свинок. Ильин устанавливает шесть пар центров депигментации у гомозиготной
68
по основному фактору пегости расы s. Весьма часто тот или иной центр отсутствует: в случае симметричных центров может отсутствовать один или оба центра; при этом чем чаще встречается асимметрия проявления какого-либо центра, тем ниже его проявление. Именно такой картины следует ожидать, если стать на позиции Астаурова. В этой (в других отношениях очень интересной) работе Ильин принял сложную полигибридную схему; для объяснения наследования белых пятен он привлек особые гены, числом не менее 13, каждый из которых, по его мысли, отвечал за реализацию своего пятна; при этом для каждой пары симметричных пятен он был вынужден допустить особый ген для каждой стороны — то доминантный, то рецессивный (Ильин, 1928).
Полидактилии, встречающиеся у разных животных, как правило, несимметричны, что часто смущало исследователей, выяснявших тип наследования. То же относится к полимастии. Впрочем, О. А. Иванова (1926), привлекая представления об асимметричном проявлении, показала, что наследование полимастии у коров может быть объяснено одним доминантным аутосомным геном.
СТАНОВЛЕНИЕ СТАБИЛЬНОСТИ ОНТОГЕНЕЗА
В статье 1930 г. Астауров приводит большое число фактов, указывающих на распространенность самостоятельной изменчивости признаков. Почему же резкие асимметрии в свободно живущих популяциях встречаются чрезвычайно редко? Действительно, вся масса упоминавшихся асимметрично проявляющихся признаков свойственна животным в одомашненном состоянии: это неправильно распределяющиеся пегости, случаи поли- и синдактилии, полимастии, асимметрично проявляющиеся мутации культур дрозофилы и пр.
В природных популяциях исключительно редки резкие асимметрии лишь биологически значимых признаков. Признаки же мелкие, ускользающие при первом взгляде и безразличные в биологическом отношении, весьма часто обнаруживают асимметрию проявления. Исследования на монози-готных близнецах (у ряда животных), в которых внимание обращено на мельчайшие наследственные признаки, вроде кожных узоров, направления волос в волосяном покрове, аномалий в щитках (у броненосца) и т. п., показывают, что резкие асимметрии — явление настолько обычное, что весьма часто ставило в тупик исследователей и заставляло иногда привлекать для объяснения фантастические представления о неравномерном распределении наследственной субстанции по разным сторонам индивида.3
Впрочем, эти примеры не нарушают общего правила: почти абсолютной симметрии проявления признаков у животных в природных популяциях и значительной самостоятельной изменчивости, реализующейся в асимметриях, у прирученных животных. «Достаточно сравнить сложнейшие, но строго симметричные расцветки птиц и млекопитающих, которые можно считать Этой гипотезы придерживался Людвиг (Ludwig, 1927), отчасти Дэнфорт (Danforth, 1919); подобный взгляд высказал Лутц (Lutz, 1911) в отношении Extra veins, такую возможность для некоторых признаков близнецов у человека допускал Фершюр (Verschuer, 1925). В то же время Геккер (Hacker, 1925) отвергал гипотезу неравнонаследственного деления и относил самостоятельную изменчивость за счет «запутанных эпигенетических соотношений», колеблемость которых покоится «не непосредственно на грубых влияниях внешних условий, но на колебании внутренних Условий системы» (цит. по: Астауров, 19746, с. 92). С =>тим представлением вполне совпадала и позиция Астаурова.
69
своего рода естественными пегостями, с неправильными пятнистостями одомашненных животных, чтобы получить представление о резкости имеющихся здесь различий» (Astauroff, 1930, цит. по: Астауров, 19746, с. 102).
Объясняя резкие различия в частоте встречаемости и амплитуде асимметрий у животных в естественных и культурных условиях, Астауров привлек внимание к их возможному адаптивному значению: «Приноровленность свободно живущих животных к определенным условиям существования без сомнения делает необходимым возможно большее постоянство организации в определенных условиях среды и точность в реактивных изменениях этой организации в ответ на любые изменения условий. Частным случаем этой закономерности является и правильность билатерально-симметричного строения: широкая распространенность строгой билатеральной симметрии в животном мире служит доказательством чрезвычайной полезности такого типа организации» (Там же, с. 103). Поскольку же резко асимметричные изменения связаны с непостоянством фенотипического эффекта в постоянных генотипических и внешних условиях, то, «вызывая расшатанность и неустойчивость организации», они не только не имеют шансов быть подхваченными естественным образом, но, напротив, «должны им деятельно устраняться».
Так Астауров пришел к проблеме эволюционного становления стабильности индивидуального развития 4.
Заключая статью 1930 г., Астауров пишет об уроках изучения самостоятельной изменчивости: «Здесь мы воочию видим, как зыбки такие свойства живых существ, как наследственность (в смысле точного воспроизведения потомками организации родителей), симметрия, точная регуляция онтогенетических процессов, свойства которых мы нередко склонны воспринимать как неотъемлемые атрибуты органического мира. Природа не утвердила их раз навсегда как некое незыблемое проявление изначальной целесообразности и каждое мгновение они могут быть нарушены мутационным процессом и вновь отвоевываются под бдительным и непрестанным контролем естественного отбора» (Там же, с. 103).
ЗНАЧЕНИЕ ДЛЯ КЛИНИЧЕСКОЙ ГЕНЕТИКИ
В 1930 г. Астауров занялся шелкопрядом; к работе по феногенетике дрозофилы вернуться ему не пришлось. К представлениям же о самостоятельной изменчивости (спонтанной флуктуации) он обращался не раз. В очерке 1971 г. «Homo sapiens et humanis» он подчеркнул: «...строго говоря, любой сформировавшийся признак есть результат трех групп факторов: наследственности, среды и случайностей формообразовательного процесса...» (Астауров, 1971, с. 214). Обратившись в 1973 г. (в оставшемся неоконченным предисловии к т. 1 «Избранных трудов») к работам этого
4 Отвечая на заданный Э. Майром в связи с конференцией Бостонской Академии искусств и наук по истории «эволюционного синтеза» вопрос о своем вкладе в эволюционное учение, Б. Л. Астауров писал о самостоятельной изменчивости: «Мне представляется, что эта проблема имеет прямое отношение к некоторым коренным вопросам эволюционного синтеза, например, к разработанной позже теории стабилизирующего отбора Шмальгаузена-Кирпичникова и др. Быть может, она сохраняет свое значение даже и сейчас. Что до меня, то я совершенно оставил работу в этом направлении, потому что в 1930 г. я резко переменил свою область исследований и переключился на генетику шелкопряда» (Астауров — Майру; письмо датировано 4 мая 1974 г., с. 5). Я благодарен Майру за присланную мне копию его переписки с Астауровым.
70
цикла, Астауров затронул вопрос: по какой причине одни мутации in statu nascendi отличаются крайним непостоянством эффекта, тогда как другие характеризуются полным проявлением и постоянным выражением? Непостоянством фенотипической реализации обладают те находящиеся на грани проявления или выражения изменения,— заметил Астауров (1978, с 130—131),— которые обнаруживают большую зависимость от соизмеримых с ними по величине флуктуаций нормальных процессов и признаков. Для альтернативно проявляющихся мутаций большое значение имеет пороговый эффект, так что неполно проявляющиеся мутации реализуются по принципу «все или ничего» (ср. а на рис. 5). С этой позиции легко объяснить вариации числа добавочных сосцов в случае полимастии, а также непостоянства и неправильности конфигурации пятен в случае пегостей, ибо непостоянство эффекта регистрируется главным образов на границе области проявления гена («поля действия»).
Было бы интересно рассмотреть «гипотезу условных тропизмов» С. Н. Да-виденкова в свете концепции Астаурова. В попытке найти объяснение большой межсемейной изменчивости клинической картины для многих, казалось бы, мономерно-доминантных заболеваний Давиденков выдвинул предположение о широкой распространенности в населении слабых, на грани проявления, генов (в каждой семье — своих), которые легко переходят порог фенотипической реализации при наличии главного гена (major gene) основного заболевания, таким образом определяя специфический для семьи характер его клинического течения.
Публикация в 1947 г. замечательной книги С. Н. Давиденкова «Эволюционно-генетические проблемы в невропатологии» дала Б. Л. Астаурову повод для обмена письмами с ним (в 1949—1950 гг.). Клиническая невропатология имеет дело с материалом, который отличается низким проявлением, варьирующим выражением, часто — наследованием «ходом коня» (проскоки через поколение, наследование в боковых ветвях), частой дискордантностью монозиготных близнецов и т. п., что к тому же обычно осложняется скудостью посемейных данных. В анализе такого материала Давиденков следовал традиционной точке зрения, что факт проявления-непроявления всегда может быть сведен к некоторому средовому воздействию, о котором клиницист может иметь достаточно полные сведения лишь в исключительных случаях. В письме Давиденкову Астауров изложил свои представления о неполном и асимметричном проявлении и высказал надежду, что «будучи спроецированы на невропатологический материал, они, быть может, внесут в него какую-то дополнительную долю ясности» (цит. по: Астауров, 1978, с. 135)5. В ответ Давиденков отметил неудовлетворенность
5 Там же Астауров замечает: «Однако по опыту я знаю, что изложенные мною соображения далеко не «прочувствованы» даже весьма искушенными генетиками-теоретиками...» (с. 134—135). Увы! С тех пор положение не слишком изменилось. Так, Дж. Мейнард-Смит и К. Сонди в сравнительно недавнем времени поставили опыты по отбору на левостороннее (!) проявление картины хетотаксиса (3 глазка + + 3 пары щетинок) (Maynard-Smith, Sondhi, I960). Эти результаты излагаются без каких-либо комментариев в сводке Р Левонтина (1978, с. 102). В этих опытах левосторонность проявления, как и следовало ожидать, не увеличилась, однако Усилилась асимметрия, что вполне согласуется с позицией Астаурова. Будь его представления достаточно известны, такая постановка вопроса была бы просто невозможна. См., впрочем, работу ГО. Н. Иванова и Д. П. Фурман (1975); также: Mather, 1953; Zarapkin, 1939; обзор: Van Valen, 1962.
71
своей точкой зрения: «Однако я должен сознаться, что, высказываясь постоянно в том смысле, что при неполно проявляющихся наследственных факторах решающее значение имеют средовые условия, я — pro usu inter-no — сохранял всегда одно маленькое сомнение. Я о нем, правда, не писал, но часто думал. Мне казался некоторым противоречием этому допущению тот факт, что степень проявляемости для некоторых признаков и даже для некоторых болезненных форм с ограниченной проявляемостью довольно постоянна в различных условиях. Казалось, если бы все решалось здесь одним только наличием или отсутствием в окружающей среде каких-то определенных условий, мы должны были бы ожидать гораздо большего разнообразия в степени проявляемости признака, чего в ряде случаев нет. В частности, нет этого и в моей области...» И далее: «...В клинике часто попадаются случаи и наблюдения, анализ которых был бы интересен с Вашей точки зрения. Но интересен именно в смысле «постановки вопроса». Разрешение вопроса надо, мне думается, ожидать от большего согласования генетики с механикой развития» (Там же, с. 135—136).
В области наследственных проблем невропатологии анализ с позиций принципа Астаурова был проведен впервые через 30 лет. В поисках причин загадочных сторон наследования шизофрении Эфроимсон (1978) считал важным учитывать высокую защищенность мозга как системы переработки информации. Исходя из этого, факт, что неполная пенетрантность и варьирующая экспрессивность шизофренических генотипов являются правилом и в то же время высокая пенетрантность связана с высокой экспрессивностью, он объяснял «именно тем, что только симметричное двустороннее поражение каких-либо участков мозга нужно для перевода нормального мышления в шизофреническое» (Эфроимсон, 1978, с. 223). Система обеспечения нормального мышления, основанная на относительной независимости работы обоих полушарий, помогает, по Эфроимсону, понять чрезвычайную вариабельность клинического течения болезни у индивида, у пары однояйцовых близнецов (т. е. при полной идентичности всего генотипа), у больных членов одной семьи (т. е. при о.бщности основного доминантного гена): «Моделируя клинические проявления при полной экспрессивности гена в его поле действия в одном полушарии и неполной в другом, в первом приближении можно предположить, что выйдет из строя лишь то звено переработки информации, прогнозирования, принятия решения, планирования, которое поражено в обоих полушариях» (Там же, с. 224—225).
Эфроимсон неоднократно подчеркивает, что для решения проблемы шизофрении необходимо соединение клинических исследований с биологическими, включая такую высокотеоретическую разработку, как концепция Астаурова. «Раскрытая еще в работах Б. Л. Астаурова (1928, 1930) независимость право- и левостороннего поражения при автономной дифференциации правых и левых огранов объясняет нам также неполную пенетрантность шизофрении, дискордантность ОБ [однояйцовых близнецов], проскоки через поколение, поражения в боковых ветвях. Однако все эти решения приходится черпать из дисциплин, чрезвычайно далеких от психиатрии» (Там же, с. 239—240).
* *
Принцип Астаурова охватывает круг явлений неопределенного фенотипического осуществления гена (неполное и асимметричное проявление, вариабельность областей действия генов и колеблемость выражения). Его содер 72
жание сводится к тому, что любой сформировавшийся признак есть результат трех групп факторов: наследственности, среды и спонтанной флуктуации формообразовательного процесса. В этой связи следует различать две категории ненаследственной изменчивости: модификационную изменчивость, возникающую как адаптивный ответ на колеблющиеся условия развития, и самостоятельную изменчивость, вызываемую «онтогенетическим шумом». Обсуждение этого принципа привело Астаурова к постановке вопроса об эволюционном становлении стабильности онтогенеза — ключевого для теории стабилизирующего отбора Шмальгаузена. Возможность постановки вопросов о иерархичности процессов онтогенеза и о его эволюции обязана системной ориентации Астаурова; в его принципе системный подход четвери-ковцев получил конкретное развитие.
Глава 5
КОНЦЕПЦИЯ ПОЛЕЙ ДЕЙСТВИЯ ГЕНОВ
Одно из самых основных различий в нашем мышлении — натуралистов, с одной стороны, и математиков — с другой,— это характер пространства. Для математиков, если это не оговорено ими, пространство является бесструктурным. Оно характеризуется измерениями, и только. Для естествоиспытателя — говорит он это или нет, даже сознает он это или нет — пустое незаполненное пространство не существует. Он всегда мыслит реальное пространство и только с ним имеет дело.
в. И. ВЕРНАДСКИЙ
Попытка П. Ф. Рокицкого атаковать общую проблему неустойчивости фенотипического осуществления гена, которой посвящена эта глава, пошла по иному пути, чем у Астаурова или Тимофеева-Ресовского. Выявление структурированности пространства, где проявляется специфическое действие гена, стало задачей его исследования, которое оформилось как концепция «поля действия» гена (Рокицкий, 1929а, б, 1930; Rokizky, 1930; см. также Рокицкий, 1931, 1973). Работа эта была предпринята по инициативе Н. К. Кольцова; она была связана со взглядами Кольцова на индивидуальное развитие и особенно с его теорией «силового поля», в виде которой оформилось его материалистическое понимание той области естествознания, которую он изучал, в частности, процесса развития яйцеклетки как целого и как организованной системы. «... Я избегаю обозначений «организационные поля» [Шпеман], так как в это понятие может быть вложен виталистический смысл, между тем как термин «силовые поля» употребляется в чисто физическом смысле, как, например, «магнитные поля». С другой стороны, теория градиентов Чайлда есть аналитическая теория, между тем как развиваемая здесь теория силового поля подчеркивает целостный характер развития яйца как «единого целого». Чайлдовские градиенты являются лишь элементами, из которых слагается «силовое поле», и последнее есть нечто большее, чем ряд отдельных слагаемых» (Кольцов, 1936, с. 559). Наиболее разработанное изложение этих представлений, включающее разбор проблемы «пре-Ф°рмация либо эпигенез», критику ряда виталистических истолкований,
73
не утратившую своего интереса по сей день критику «биогенетического закона» Э. Геккеля с позиций феногенетики и пр., дано в речи Кольцова «Генетика и физиология развития» (опубликована в 1934 г. по-русски, в 1935 г. по-французски; переизд. в кн. «Организация клетки»: Кольцов, 1936, с. 540—584. Подробнее см.: Астауров, Рокицкий, 1975, гл. 6). При анализе взглядов Кольцова на индивидуальное развитие обычно подчеркивают его материалистические позиции и оппозицию витализму, распространенному тогда среди эмбриологов. Уместно отметить здесь также его системный подход, в то время когда большинство материалистически мыслящих биологов-экспериментаторов было редукционистами. Кольцов (как и Четвериков и их ученики) развивал ту линию мировоззрения в русском естествознании, которая наиболее яркое выражение нашла в работах И. И. Шмальгаузена («Организм как целое...» и др.).
Системный подход Кольцова — созвучный ориентации группы Четверикова — повлиял на постановку проблемы Рокицким. Кроме того, известное влияние на него оказало изучение четвериковцами феномена зонарности плейотропного действия генов дрозофилы, а также проблема пегости, которая широко обсуждалась в первой четверти века в связи с фундаментальными вопросами менделизма и теории отбора. К проблеме пегости обращался Кольцов (1921), впоследствии Астауров (см. гл. 4) и др.; интерес к ней Рокицкого был отчасти обязан также его участию (вместе с В. Е. Альтшулером, С. М. Гершензоном, Н. П. Дубининым, И. Н. Свешниковой и др.) в реферативном генетическом кружке А. С. Серебровского при Центральной генетической станции.
Зонарное плейотропное действие некоторых «сигнальных» генов Рокицкий отметил (1927) в ранней работе по генетике стерно-плевральных щетинок дрозофилы, выполненной под руководством А. С. Серебровского. Ген III хромосомы Delta, вызывающий расширение жилок на концах крыла, плейо-тропно действовал на число этих щетинок; hairy (III), вызывающий появление большого числа микрохет на щитке, и сцепленный с полом scute, редуцирующий щетинки на щитке, также влияли на стерноплевральные щетинки. В четвериковской группе зонарность плейотропного действия описала Балкашина (1928) на примере мутации aristopedia (см. гл. 6), Промптов (1929) —на polymorpha.
Капюшонные крысы Касла. В области исследования пегостей особый интерес вызвал случай капюшонных крыс Уильяма Касла, так как истолкование его селекционных опытов 1906—1919 гг. затрагивало вопросы эффективности отбора и постоянства гена, существенные для судеб менделизма и теории отбора и их отношений. В те годы идея эволюции путем отбора была отодвинута на далекий задний план. Пользовавшаяся признанием мутационная теория Гуго де Фриза включала положение, что «Отбор сам по себе не обеспечивает происхождения новых видов» (как называлась одна из глав т. 1 его «Мутационной теории»). Заметного места отбору не оставляла также теория чистых линий В. Л. Иогансена; распространение ее Иогансеном на гибридные популяции доставило мнению о неэффективности отбора новых сторонников. Можно напомнить, что одним из оснований менделевской генетики, лежащей в основе теорий де Фриза и Иогансена, был принцип чистоты гамет. Поэтому выводы Касла, который он сделал в статьях 1914—1916 гг., о чрезвычайно высокой эффективности отбора, об изменении гена в ту сторону, в которую велся отбор, и вообще о «загрязнении» (contamination) гена сразу же попал в центр внимания и вызвал резкую критику.
Касл (с Мэк Кэрди и затем с Филлипсом) изучал наследование пежин у крыс. Начав с потомства купленных крыс — белых с черным пятном на голове и узкой
74
черной полоской вдоль спины — Касл повел отбор в двух направлениях: в сторону сужения этой полоски и в сторону ее расширения. Ему удалось получить в одной линии отбора расу, у которой исчезла полоска на спине, а в другой — расу, у которой эта полоска распространилась на спину и бока, оставляя только белое брюшко. Касл полагал, что отбор вывел цветной рисунок за пределы изменчивости, свойственной исходной популяции (опровергая тем самым Иогансена, Дженнингса, Пэрля и Других сторонников теории чистых линий). Эти результаты можно было бы объяснить подбором двух групп генов-модификаторов, определяющих ширину полоски на спине. Касл, однако, настаивал, что он имел дело с признаком, обусловленным Лишь одним геном — ибо F2 от скрещивания капюшонных крыс с дикими серыми давало 1 капюшонную крысу на 3 одноцветные. Капюшонные особи F2 оказались чрезвычайно изменчивыми, и Касл предположил, что рецессивный «капюшонный> ген изменился благодаря соседству в гибриде с «нормальными» генами дикой пигментации. Хотя эти данные показали, что пежины вызываются действием рецессивного гена, но здесь еще ничто не исключало возможности видоизменения эффекта этого гена за счет модификаторов.
В серии опытов 1914 г. с обратным отбором, после эквивалентного числа поколений (начиная с минус-расы) Касл не получил полного возврата к исходному типу окраски. Объясняя результаты, он одно время претендовал на восстановление взгляда, который он возводил к Дарвину, а именно, что отбор сам по себе вызывает в наследственном материале изменения в том направлении, в котором он ведется. Вывод Касла, какой бы способ загрязнения гена не принять, подрывал одно из оснований менделизма; и он сразу же вызвал критику с разных позиций. Э. Ист выдвинул идею «множественных модифицирующих факторов»; Касл отверг ее на том основании, что обратный отбор не привел к полному возврату. В дрозофильной группе Моргана тогда уже картировали положение генов-модификаторов на хромосомах; против взгляда Касла выступил Мёллер — по результатам своих с Э. Альтенбургом опытов; эту критику Касл также отверг. А. Стёртевант в 1918 г. опубликовал проведенные с Мэк Доуэллом селекционные опыты на дрозофиле и показал, что результаты можно объяснить только с позиций отбора модификаторов. Эти опыты показали также, что отселектированная раса не может быть возвращена отбором к исходному состоянию, что было бы возможно при модификации отбором менделевского фактора. Тогда Касл продолжил проверочные опыты, предложенные С. Райтом и начатые в 1916 г. (План Райта заключался в том, чтобы скрещивать крыс из плюс- и минус-серий с дикими и выделять в F2 капюшонных рецессивов. Если Касл отбирал модификаторы, то они отщепятся от «капюшонного» признака, и выделяемые из плюс-и минус-линий капюшонные окраски снова станут сходными). В опытах по этой схеме возврат минус-расы к исходному типу окраски шел сперва очень медленно; Касл решил, что модификаторы здесь не причем, а результат означает, что отбор изменяет ген в обратную сторону. Однако проведенные к 1919 г. три дополнительных скрещивания с дикой расой и выделения капюшонных особей имели результатом резкий сдвиг рисунка; тогда Касл отказался от долго разделяемого взгляда об изменчивости гена и присоединился к мнению, что в его опытах отбирались «независимо наследуемые модифицирующие факторы». Низкую эффективность начальных опытов с минус-расой он объяснял тем, что остаточная наследственность, «residual heredity», диких крыс значительно ближе к плюс-, чем к минус-pace. Интересно, что в свете его окончательных объяснений эта «residual heredity» имела тот же смысл, что и четве-ри ко вс кая «генотипическая среда». Несмотря на перемену мнения о субстрате отбора, Касл считал (и не без основания), что его селекционные опыты означали явный прогресс в понимании роли отбора в эволюции. И он лишь утвердился в своем основном убеждении — в выдающейся роли отбора.
Случай капюшонных крыс Касла затронул целый ряд принципиальных вопросов, но широко обсуждались в этой связи лишь те, что волновали исследователей в данный момент: устойчивость гена и роль модификаторов. Критически оценивая интерпре-Тации Касла, Н. К. Кольцов отметил, что тот не проводил различия между генотипом фенотипом. В 1920 г. он писал: «Нельзя отрицать возможности фенотипических Уклонений в области пятнистости, но мне кажется, нет оснований и следовать
75
за Кэстлем, который именно на примере пятнистости грызунов основывает свою теорию закрепления фенотипных уклонений путем естественного отбора. Гораздо правильнее применить здесь принцип Нильсона-Эле и Ланга и допустить разложение генов L и М на ряд отдельных единиц, комбинация между которыми определяет степень пятнистости. [...] То, что Кэстль называет подбором фенотипов, на самом деле является бессознательным подбором генотипов» (Кольцов, 1921, с. 94).
Еще один затронутый Каслом вопрос — создает ли отбор сам себе изменчивость — остался тогда без внимания. Лишь Т. Морган — скептик в отношении теории отбора, сторонник теории мутаций де Фриза — мимоходом отметил, сочтя обсуждение вопроса лишним: «Если бы это было действительно мнением Дарвина, могло бы показаться, что такое толкование изменчивости сильно подкрепляет теорию естественного подбора как способа, при помощи которого осуществляется эволюция» (Морган, 1927, с. 257; случай Касла описан в гл. «Постоянство гена» цит. соч.; см. также: Ильин, 1928; Provine, 1971; Allen, 1978).
Для выявления структуры морфологического поля действия гена в качестве признака-индикатора — единого для всех участков тела дрозофилы и допускавшего количественный учет — Рокицкий (1929) выбрал щетинки. Он учитывал 26 мест (ср. рис. 10). Действие конкретного гена на каждой единичной щетинке приводило либо к ее исчезновению (редукции), либо к появлению добавочной щетинки. В 10 изученных линиях некоторые гены прибавляют щетинки: Polychaeta (Roi), Notch-8 (N-8), Hairy-wing (Hw); другие редуцируют: Dichaete (D), scute (sc); ген split-bristles (spb) редуцирует щетинки, одновременно раздваивая и прибавляя их в другом месте.
ФЕНОМЕНЫ РЕАЛИЗАЦИИ ГЕНА В СВЕТЕ «ПОЛЕЙ ДЕЙСТВИЯ»
Легко убедиться в специфичности очертаний полей различных линий (рис. 6). Поля действия генов Рокицкий характеризовал средними числами (М) щетинок в каждом пункте. (Если для определенного пункта, скажем, /И =0,75, это значит, что у 25% мух щетинка редуцирована, т. е. в данном пункте редуцирующий ген имеет 25%-ное проявление). Данные Рокицкого указывают на то, что «мы всегда по отношению к любому гену можем говорить о неполном проявлении. Поле действия гена заключает в себе ряд точек с разными процентами проявления. Иными словами, поле обладает переменным проявлением» (Рокицкий, 1929а, с. 191). Понятие выражения гена в контексте полей действия также приобретает специальный характер. Предположим, у одной мухи Dichaete нет щетинок а, б, в, а у другой — а, б, г, е, ж. Выражение гена D у второй мухи сильнее, чем у первой, так как редукция захватывает 5 щетинок, а не 3. Выражение гена у отдельной мухи складывается из локальных проявлений. Поле, построенное по проявлению у десятков мух, показывает максимальное выражение признака: «Поле, сотканное из проявлений по отдельным точкам, являющихся статистическими величинами, в то же время есть предел возможного выражения гена» (Там же).
Таблица 8
Различные степени доминирования в разных точках
Группы мух м
рра | adc | splm
Самки yHw/yHw 1,17 1,71 6,61
Самки BCl/yHw 1,00 1,06 6,32
Самцы yHw/7 1,09 1,00 7,40
76
Рис. 6. Примеры полей действия генов
Голова и грудь сверху, самки: а — split-bristles
(по: Рокицкий, 1929а)
Dichaeta, б — Polychaeta, в — scute-white, г —
Метод анализа полей действия позволил количественно подойти к оценке степени доминирования. Хороший материал дал сцепленный с полом доминантный ген Hairy wing. В этой линии всегда получались группы мух: самки BCl/yHw, yHw/yHw; самцы yHw/7 (скрещивания всегда, ставились: BCl/yHw XyHw/7. Их интересно сравнить между собой. У гетерозиготных самок прибавляющее действие Hw очень невелико. У гомозиготных самок вместо обычных четырех дорзоцентральных щетинок — целый пучок их, в среднем 8—9. Самцы-гемизиготы несут только одну Hw -хромосому, и этим похожи на гетерозигот-самок, однако эффект генов на их теле значительно отличается — качественно и количественно (табл. 8).
Отношения между гомо- и гетерозиготными самками и гемизиготными самцами демонстрируют условность и количественность понятия доминантности. На щетинке рра ген Hw не проявляется у самок-гетерозигот, но проявляется в 9% случаев у самцов, также несущих один ген, на adc — наоборот. По щетинке splm самцы дают наибольший показатель — даже выше, чем гомозиготные самки. Отсюда Рокицкий делает вывод, что доминантность является функцией, с одной стороны, отношений гена с генотипической средой, и с другой, места его осуществления. К представлению об опера-Циональности понятия доминантности привлекал внимание и Тимофеев-Ресовский (см. гл. 3), отмечая, что именно «наследственная конституция» Делает тот или иной ген рецессивным или доминантным.
77
СТРУКТУРА ПОЛЯ
Основа концепции Рокицкого — взгляд на зону морфологического эффекта гена как целое, обладающее специфической структурой. «Каждое поле обладает своей характерной, присущей ему структурой, выражающейся в том, что разные точки поля подвергаются разной степени воздействия» (Рокицкий, 1929а, с. 197). Он различал точки в поле действия по степени затронутости геном (высокая, средняя и очень малая) и подчеркивал, что эти три степени не являются чисто условными группировками. Гистограмма (рис. 7, а), представляющая интегральное распределение М по четырем линиям с редукцией щетинок, покзывает, что чаще всего реализуется либо очень сильная, либо очень слабая редукция. «Существование некоторых скачков степени эффекта указывает на ступенчатость в реализации факторов, вызывающих редукцию» (Там же, с. 200).
Представление о центрах наивысшего эффекта внутри поля может привести к точке зрения, что эффект гена распространяется от центров во все стороны и ослабевает к краям поля. Действительно, такою часто бывает внешняя картина. Однако в общем случае правильного распространения эффекта нет; чаще наблюдается причудливый узор редукции или прибавления щетинок. Внешняя картина не дает основания для вывода, что причиной структуры поля является «диффузное распространение редукции от центра» (Там же, с. 203). К аналогичному заключению пришел Б. Н. Сидоров, изучавший мозаики по yellow в линии scute8 D. melanogaster. Он настаивал, что «Гольдшмидтовское представление о распространении эффекта из определенного центра должно быть оставлено» (Сидоров, 1934, с. 732).
Независимость элементов поля. Представление о целостности поля как будто позволяло ожидать значительной связанности в эффекте между близкими точками поля. Для проверки этого предположения Рокицкий вычислил ряд коэффициентов корреляции между разными точками в нескольких линиях. Только один коэффициент указал наличие некоторой (невысокой) корреляции; все остальные показали отсутствие ее: «Мы пришли к довольно парадоксальному выводу. Несмотря на внешнюю целостность поля, отдельные его элементы ведут себя независимо» (Рокицкий, 1929а, с. 203). Это явление находит объяснение в принципе Астаурова, согласно которому при генетической однородности материалов и однообразии внешних условий проявление на двух сторонах тела автономно (см. гл. 4). Корреляция сторон может иметь место, например, в гетерогенной совокупности или «если реакция гена (редуцирующего или прибавляющего) предшествует дифференциации обеих точек из одного общего эмбрионального элемента», а также (это замечание Рокицкий сделал в позднейшей обзорной статье 1973 г.) «в случае независимых (sensu Sturtevant) признаков, осуществление которых определяется влиянием геногормонов».
Рокицкий отметил, что причину отсутствия корреляции между двумя признаками (например, двух пежин на шкурке морской свинки) иногда усматривают в различии генов, их определяющих. И указал, что такое заключение опровергается его выводом о независимости отдельных элементов поля: «Пришлось бы допустить множество генов, каждый из которых влияет на определенную точку, причем для каждой стороны отдельно, ибо стороны также не связаны друг с другом» (Рокицкий, 1929а, с. 204). Приводимые им примеры подтверждают мнение Астаурова (1930), что «та или иная
78
степень сложности объяснения в таких случаях становится делом исследовательского такта и остроумия» (цит. по: Астауров, 1974, с. 94).
Факторы изменчивости поля. Линия Polychaeta из геленджикской популяции D. melanogaster дала начало массовой инбредной культуре и нескольким индивидуальным, причем очертания поля в массовой культуре были шире, чем в каждой из просчитанных индивидуальных культур. Причину этого Рокицкий видел в том, что в массовой культуре идет расщепление по модификаторам Pol. Благодаря им очень слабое прибавляющее действие Pol, которое должно быть за пределами видимого поля, усиливается и дает видимый эффект. Индивидуальные же культуры в результате случайностей выборки оказались лишенными этих модификаторов. Массовая культура показала наличие некоторой корреляции сторон, что подтвердило предположение о гетерогенности ее.
Среди факторов изменчивости поля Рокицкий различал внешние и внутренние условия и прежде всего генетические — генотипическую среду. «Если действительно в линии находятся генетические факторы, воздействующие на некоторые участки поля, то можно представить себе возможность с помощью отбора не только изменить поле в общих его очертаниях, сделать его больше или меньше, но и сдвинуть поле — вверх или вниз, ближе к краям тела или ближе к средней оси» (Рокицкий, 1929а, с. 208—209). Яркий пример изменения границ поля одного гена путем отбора генотипической среды дал случай капюшонных крыс Касла.
Другой пример значения генотипической среды (для реализации ансамб-
79
ля щетинок у дрозофилы) доставило исследование К. Сонди (Sondhi, 1962). Сцепленный с полом рецессивный ген ocelliiess устраняет типичную картину: 3 глазка и 3 пары щетинок на верхней поверхности головы D. subobscura. Выражение гена варьирует, его степень оценивается с баллах (сумма проявившихся глазков и щетинок, от 0 до 9). В линии ocelliiess Сонди вел отбор от ocelliiess к дикому типу На 41-м поколении отбора самки перешли 9-балльный барьер дикого типа и стали давать, на одной или обеих сторонах, добавочные щетинки в типичных местах, а также неоморфы, расположенные латеральнее оцеллярных. Эта пара щетинок отсутствует у Drosophilidae, однако она есть у одной из шести близких к ней групп, которые часто возводятся в ранг семейства. Один из представителей Aulacogastridae (или Aulacigastridae) — Aulacigaster leucopeza — имеет в норме возникшую у D. subobscura de novo пару щетинок. На этом основании Сонди заключил, что «препаттерн» (система, определяющая позиции формирования щетинок — К. Штерн) этих щетинок имеется у дрозофил дикого типа и что возрастание количества «предшественника» может заставить эти щетинки проявиться.
ГЕН И ТКАНЬ
Анализ полей действия генов привел к вопросу о характере реактивности ткани: «...может казаться, что строение поля целиком определяется генами, его созидающими, [...] в то время как отдельные участки тела обладают одинаковыми возможностями реагировать на то или иное влияние гена» (Рокицкий, 1929а, с. 209). Но можно ли говорить о изопотенциальности отдельных участков тела, в смысле ответа на воздействие генов? Подсчитав числа изменений в каждой из выделенных точек по всем линиям (см. рис. 7, б), Рокицкий пришел к заключению, что специфичность поля действия на теле дрозофилы «определяется не только специфичностью влияния генов, но и большей или меньшей способностью участков тела реагировать на это влияние. Часть устойчивости эффекта гена должна быть отнесена на счет неравной потенциальности отдельных точек» (Там же, с. 210).
Эти соображения тесно сближают вопросы пространственного действия гена с явлениями пегости, где исследователь часто идет «от ткани», и в конечном итого они приводят к проблеме отношений гена к ткани. Под «тканью» Рокицкий понимает «формирующиеся элементы организма к моменту вступления в действие нашего гена» (Там же, с. 211), т. е. речь идет о включении частного морфогенного процесса в общий. Он отмечает условность разделения «ткани» и «гена»: «Ткань» в каждый данный момент времени заключает в себе гены уже активные, in statu nascendi своего действия, и, наконец, еще пассивные. Их нельзя отделять от «ткани» и «ткань» нельзя мыслить без генов. Но при анализе роли генов в определенном признаке мы очень часто противопоставляем один анализируемый нами ген всему остальному — «ткани» (Рокицкий, 1931, с. 182).
Такая интерпретация близка, но не совпадает с формулой М. М. Завадов-ского (1930): Х+У-^А, где X — реагирующая ткань, Y — раздражитель, А — сформировавшийся признак. Эта формула может быть приложена к анализу морфогенных процессов, не только беря Y в качестве внешнего раздражителя, но и глубже — «видя в Y биохимический процесс, идущий от гена» (Рокицкий, 1931, с. 183).
Осмысление результатов генетического анализа, вступавших в противо-
80
речие с «ранней, незрелой идеей» (Морган), что для каждого признака имеется специфический ген, у С. С. Четверикова вылилось в концепцию «генотипической среды», обобщившую идею плейотропии. Взгляд на плейот-ропное (множественное) действие гена разделяли многие авторы (начиная с Л. Плате, 1910, и включая Т. Моргана), полагая, правда, разные пределы плейотропии. Согласно крайней точке зрения, «каждый ген определяет в итоге все признаки организма» (Дубинин, 1932, с. 115). Логическое развитие этого крайнего взгляда должно привести к выводу, который и сформулировал в своем критическом обзоре А. А. Любищев (1925, с. 67), а именно что каждый ген потенциально может дать организм: «... ген по потенциальности соответствует иду...» Однако признание всеобщей плейотропии свойством гена, как отметил М. М. Камшилов (1935, с. 114), «приводит к смазыванию качественного различия между генами, что вступает в явное противоречие с экспериментальными данными». Вопросу Любищева: «Но как же велика «сфера компетенции» одного гена, совокупность признаков, которые отчасти определяются данным геном?» (Любищев, 1925, с. 54), идет на смену вопрос Камшилова: «Что же тогда определяет, на какие части он действует? На этот вопрос может быть два ответа: 1) специфика гена как локуса; 2) специфика развивающихся частей» (Камшилов, 1935, с. 114). Отмечая, что «организм не есть сумма двух разорванных состояний: половая клетка!— взрослый организм, а есть определенный процесс с характерными для него особенностями» (Там же, с. 142), Камшилов переносил центр внимания на этот процесс и заключал отсюда, что специфика проявления гена целиком определяется ходом индивидуального развития. На этом основании он, в частности, возражал против применения выражения «генотипическая среда». Это особенно любопытно в той связи, что его постановка задачи опиралась на работы четвериковцев, в которых конкретизировалось это общее понятие.
Еще раз отмечу, что концепция генотипической среды важна нам не как результат, а как постановка вопроса. Действительно, конкретное содержание ее (широкая плейотропии) не выдержало бы современной критики. Напротив, заключенная в ней возможность представить в единой концептуальной схеме процессы наследственной передачи, популяционной динамики, частные формообразовательные процессы оказалась чрезвычайно плодотворной (обзоры: Промптов, 1934а; Timofeeff-Ressovsky, 1940b; ср.: Шмальгаузен, 1937).
Как видно из его анализа проблемы «ген и ткань», Рокицкий, развивая концепцию полей действия генов, конкретизировал на этом пути общую идею генотипической среды Четверикова; ее другими конкретизациями, как мы видели, стали понятие наследственной конституции Тимофеева-Ресовского и принцип автономной изменчивости Астаурова.
Создает ли отбор сам себе изменчивость? Через два десятилетия после обсуждения случая Касла (а к тому времени представление, что отбор не может вывести признак за пределы его первоначальной изменчивости, Уступило место взгляду об эффективности отбора) интерес к связи отбора с изменчивостью проявил Гудейл (Goodale, 1937). Он опирался на два случая эффективного отбора: старый опыт Пейна по отбору дополнительных грудных Щетинок у дрозофилы и свой опыт по увеличению пятна на голове мыши. Uh считал невероятным, чтобы первая мышь с 12 белыми волосками на голове, с которой был начат опыт, содеражала все факторы, обусловившие белое пятно, охватывавшее почти всю голову мыши в 20-м поколении; он б В. В Бабков 81
считал невероятным также предположение, что при таком малом числе опытных животных могло возникнуть достаточное для этого эффекта количество мутаций. Поэтому Гудейл решил, что отбор сам создает желательную для себя изменчивость. По его мнению, механизм направленной изменчивости заключается в отборе состояний гена, наиболее сдвинутых в направлении отбора, чем он и объяснял результаты своего опыта.
Течение отбора в условиях естественной и экспериментально вызванной изменчивости изучал Рокицкий в экспериментах 1932—1940 гг. (первые публикации: 1933, 1936; к 1948 г. была подготовлена сводка «Изменчивость и отбор», ее идейная сторона изложена в ст. Рокицкого, 1959). В этом исследовании была найдена связь отбора с появлением наследственных изменений; однако картина этой связи рисовалась автору совершенно иной, чем Гудейлу и Каслу.
Из исследований Иогансена вытекала задача изучения отбора на основе наличной изменчивости; Рокицкий задался новым вопросом — выяснить эффективность отбора, использующего вновь возникающие наследственные изменения. Он вел отбор на увеличение (и уменьшение) числа грудных щетинок у дрозофилы в опытных линиях, освещавшихся рентгеновскими лучами, и контрольных; в каждом поколении регистировались среднее и дисперсия линии. Результаты нескольких линий подтвердили наличие эффекта рентгеноселекции.
Рокицкий утверждал, что отбор велся в значительной степени не на первоначально существовавшей в исходном материале изменчивости (мнение, которое позже разделял, например, Мейзер), а на вновь появляющихся мутационных изменениях. (Линии разводились путем инбридинга, поэтому первоначальная изменчивость должна была быть использована за несколько первых поколений. Однако в отдельных линиях подъем средней наблюдался до конца опыта на 40-м поколении.)
Для учета изменений при отборе Рокицкий применил метод анализа на поле М — 62 (предложенный для более простого случая А. С. Серебров-ским — см.: Серебровский, 1970. Метод треугольника); проанализированные линии распались на три группы. В одной, несмотря на длительный отбор, средние отдельных поколеий колебались вокруг одного центра и изменчивость рано или поздно стабилизировалась (т. е. отбор был неэффективен). В другой после длительного периода безрезультатного отбора, порядка 30 поколений, наступало резкое возрастание изменчивости (как следствие включения в отбор вновь появившейся мутации) и в дальнейшем подъем средней, после чего изменчивость вновь уменьшалась. Наконец, в третьей серии (и это самый интересный случай) наблюдалось сложное наложение нескольких циклов изменчивости и длительный подъем М через ряд ступеней, при этом, несмотря на отбор и инбридинг, увеличивалась изменчивость. Хотя отдельные наследственные изменения были разделены промежутком в 5—10 поколений, в отбор они включались непрерывно, создавая возможность постоянного и длительного подъема средней. Постоянное возрастание вариансы Рокицкий связал с действительным нарастанием генотипической изменчивости, исключив другие возможности. В некоторых линиях появляющиеся циклы «характеризуются поразительно большой изменчивостью, в 3—4 раза превышающей первоначальную, что указывает на подлинный взрыв генотипической изменчивости. Создается впечатление, что [...] первое подхваченное отбором изменение тянет за собой целую цепь новых, открывая новые пласты генотипической изменчивости и создавая тем самым возмож
82
ности для длительного и непрерывного прогресса путем отбора» (Рокицкий, 1959, с. 91).
Экспериментальное изучение отбора дало основание Рокицкому настаивать на возможности неограниченных сдвигов при отборе — на основе совершенно иной картины связи появления наследственных изменений с отбором, чем у Касла или Гудейла. Если бы отбор сам по себе стимулировал наследственные изменения в направлении отбора, то тогда он должен был бы быть эффективным во всех линиях. Однако в большинстве линий третьей серии Рокицкого он был неэффективен. Также и факт накопления минус -изменений в линиях плюс-отбора противоречил этому взгляду.
Рокицкому рисовалась следующая картина. Если отбор подхватывает новое наследственное изменение, то при этом должна изменяться существовавшая генотипическая система а в результате должны появляться новые возможности для использования тех генотипических ресурсов, которые в старой системе не могли быть использованы. Поэтому цикл изменчивости, протекающий после включения в отбор нового наследственного изменения, может порождать новые циклы, накладывающиеся на первый. В некоторых случаях получается картина «подлинного взрыва наследственных изменений» или их «пучкового» возникновения: «Первое наследственное изменение, действительно новое, вскрывает целый ряд других, фактически уже бывших, но прежде бесполезных для отбора; подобную же роль для вскрытия генотипических ресурсов могут сыграть изменения условий среды» (Рокицкий, 1959, с. 93, 94).
Самое замечательное различие между старыми авторами и Рокицким — в общей постановке вопроса, которая зависела от методологической позиции. Касл, Морган, де Фриз и др. выбирали одну (ту или иную, в зависимости от их склонности) причину эволюции, которой были безусловно подчинены все остальные. У Рокицкого мы находим понимание иерархической сопод-чиненности, а значит и определенной автономии, факторов эволюции — развитие системного взгляда, которым была пронизана концепция генотипической среды Четверикова.
6*
83
Глава 6
ГОМЕОЗИСНЫЕ МУТАЦИИ: РАЗВИЛКИ В ОНТОГЕНЕЗЕ
...Начало всегда остается самым замечательным моментом всякого явления.
T КАРЛЕЙЛЬ
Материалом для феногенетического исследования Е. И. Балкашиной (1928а, б; Balkaschina, 129) послужило наследственное изменение, выделенное из потомства «самки-родоначальницы» из дикой популяции D. melanogaster Геленджика. У измененных мух вместо аристы усика развивалась лапка, поэтому изменение получило название aristopedia (символ агр). На этом материале Балкашина впервые изучила эмбриональное развитие гомеозисного мутанта.
ПОНЯТИЕ ГОМЕОЗИСА
Гомеозисом У Бэтсон (Bateson, 1894) назвал такое изменение одного из членов меристического ряда, органа или сегмента, когда он утрачивает присущие ему черты и приобретает признаки другого члена этого ряда. (Иногда, вслед за Лёбом, это явление называют «гетероморфоз» — Loeb, 1891). Это явление давно известно из случаев аберрантных регенераций, наблюдавшихся в природе у некоторых членистоногих (в статье 1928 г. Балкашина приводит работы, начиная с 1834 г.), а также регенераций в эксперименте (с 1896 г.— в том же списке). С эволюционной точки зрения наиболее важными являются наследственные гомеотические изменения.
Балкашина, используя классификацию Пржибрама, различала у D. те-
В ИЭБ, ок. 1928 г.: Е. И. Балкашина, Н. К. Беляев, П. Ф. Рокицкий, С. М. Гершензон, Б. Л. Астауров
84
lanogaster два типа наследственных гомеозисов. Один — это придаточный гомеозис (Zusatzheteromorphose), примерами которого могут служить мутации: reduplicated legs, удваивающая ноги каждого грудного сегмента (которую нашла Марджери Ходж в 1915 г.), и crippled-m, найденная Комаи в 1926 г. Другой тип, замещающий (Ersatzheteromorphose),— наследственный гомеозис в узком смысле — встречается у дрозофилы чаще. В моргановской группе было дважды найдено и исследовано наследственное замещение жужжальца (галтера) крылышком: bithorax (bx) и bithoraxoid (bx-b); сходное изменение под названием tetraptera описал Астауров (см. гл. 4). Авторы сводки «Генетика дрозофилы» писали: «Сперва предполагалось, что они [жужжальца] изменяются в крыльях у bithorax. Более тщательное изучение показало, что bithorax обязан полному замещению треетьей части торакса повторением второй части, нормально несущей крылья» (Morgan et al., 1925, р. 79—80). Позже были описаны аллели Ьх, отличающиеся от него по проявлению, выражению и изменчивости: bx-2, bx-3, bx-w дают более крайние варианты; Ьх-34е и доминантные bx-D менее крайние. Второе изменение — bx-b — фенотипически напоминает Ьх и картируется в том же участке III хромосомы, но оно не аллельно Ьх. Морфология обоих мутантов почти не описана. Третий гомеотический мутант, tr, имеет значительно более низкое проявление, чем Ьх, и сильно варьирующее выражение, как и Ьх; он не аллелен Ьх или bx-b1
К наследственным гомеозисам в узком смысле слова относится и ряд других мутаций дрозофилы. Это aristopedia Балкашиной 2. Это также описанная Бриджесом и Добржанским в 1933 г. proboscipedia; здесь оральные доли хоботка трансформируются в пару сбежистых членистых придатков, оканчивающихся аристо- или лапкоподобным образованием; остальные ротовые части изменены так, что напоминают грызущий ротовой аппарат примитивных насекомых. Еще одна гомеозисная мутация — tetraltera, найденная Гольдшмидтом в 1940 г., в известном смысле противоположна tetraptera (по мнению Гольдшмидта): здесь крылышко замещается галтероподобным выростом; мезотонум и скутеллум могут частично или полностью отсутствовать, и дорзальная часть мезоторакса напоминает метаторакс. (Впрочем это описание позволяет нам связать противопоставлением tetraltera скорее с bithorax, чем с tetraptera).
1 В «Заключительных замечаниях» статьи 1927 г. Астауров анализирует сходства и различия трех мутантов. Показав неидентичность tr двум другим («tr представляет собой изменение, как бы идущее по тому же пути, что и Ьх, но не достигающее таких результатов, как этот последний»), но отмечает: «Изменения, происходящие в случае Ьх, и в меньшей степени в случае tr, правильнее истолковать не как расщепление среднегруди и вытеснение заднегруди, а как приобретение заднегрудным сегментом ряда черт строения, недостающих ему по сравнению со среднегрудным. Метаторакс приобретает недостающее ему (и, несомненно, им утраченное в процессе эволюции) крыло, приобретает скутеллум, сидящие на нем скутеллярные Щетинки, стерноплевральные щетинки и вторую преапикальную щетинку на голени третьей пары ножек» (цит. по: Астауров, 1974, с. 48—49).
В- И. Балкашина названием «aristopedia» подчеркивала характер изменения — гомеозис в узком смысле слова (Бэтсон) или замещающий гетероморфоз: ариста трансформируется в лапку. (Зарубежные генетики обозначали мутацию: «arista-pedia»; это наименование удержалось в биологической литературе). Принцип названия мутаций по направлению трансформации, введенный Балкашиной, сохра-иился, например proboscipedia, antennopedia, Nasobemia и т. д.
85
Параллельные ряды наследственных и ненаследственных гомеотических изменений. Балкашина впервые привлекла внимание к факту, что «ряд гомеозисов, возникших вследствие изменения наследственной субстанции и развивающихся в организме в течение его эмбриональной жизни, является параллельным ряду совершенно аналогичных гомеотических изменений, происшедших путем регенерации, когда вместо экстирпированного члена во взрослом организме образуется гетероморфный регенерат» (Балкашина, 19286, с. 94) 3. Из рассмотрения двух параллельных рядов наследственных и регенеративных гетероморфозов (случай своего рода «идио-соматической группы изменчивости») Балкашина заключает, что «взаимодействующие силы в организме, определяющие развитие органа, действуют одинаково как при внешнем воздействии (экстирпация органа), так и при внутреннем воздействии (генотипическая изменчивость)» (Балкашина, 19286, с. 94). (В связи с интерпретацией этих фактов интересно отметить, что Пауль Каммерер, еще задолго до первой публикации Балкашиной, выписал мух «aristopedia» из кольцовского института, рассчитывая использовать этот материал для доказательства идеи наследования приобретенных признаков, сторонником которой он был. Этих мух также выписал Ю. А. Филип-ченко, пришедший в генетику из эмбриологии.)
ARISTOPEDIA: МОРФОЛОГИЯ, РАЗВИТИЕ, ФИЗИОЛОГИЯ
Морфология. Балкашина, как и другие сотрудники Генетической лаборатории ИЭБ, была прежде всего зоологом с широкими интересами. Занявшись новым наследственным изменением и проведя генетический анализ с картированием положения гена на III хромосоме, она обратилась к самым интересным особенностям этого мутанта: морфологии, развитию, физиологии.
Нормальная антенна (усик) имеет три членика и аристу — постепенно суживающийся хитиновый стержень с многочисленными веточками (рис. 8, а). Измененные антенны имеют от двух до семи-восьми члеников (рис. 8, б-е), т. е. выражение варьирует от сильной редукции антенны до ее гипертрофии — случай полярной противоположности выражения — polare Gegensatzlichkeit sensu Vogt. Наиболее часто встречается высшая степень выражения признака. В этом случае ариста антенны заменяется на длинный членистый придаток, имеющий все признаки лапки, а именно крепкий хитиновый скелет с характерными сочленениями, вертикальные ряды толстых щетинок. Последний членик измененной антенны, как и лапка, несет пару
3 Регенеративный гомеозис описывали; Хербст в 1896 г. на декаподах Palaemon и Криженецкий в 1914 г. на мучном хрущаке Tenebris molitor, в их экспериментах регенерировала антенноподобная структура, замещающая экстирпированный глаз. (Подобные изменения были найдены на дрозофиле в моргановской группе, однако в этом случае генетическое и морфологическое исследование не проводилось.) Пржибрам в 1909 г. на богомоле Sphodromantis bioculata, Кэно в 1921 г., Брехер в 1924 г. и Ева Борхардт в 1927 г. на прямокрылом палочнике Dixippus morosus обнаружили регенерацию лапкоподобной структуры на месте экстирпированной антенны — случай, вполне аналогичный мутации aristopedia. Тот же результат получен в опыте Пржибрама в 1919 г. с прижиганием антенны у взрослой личинки пилильщика Cimbex axillaris (когда антенна регенерировала, ее терминальный сегмент нес один или два коготка, пульвиллу и характерные для лапок волоски) (цит. по: Балкашина, 19286).
86
Рис. 8. Антенна (усик) Drosophila melanogaster
и — у нормальной мухи; б — е — разные степени выражения aristopedia; ж последний членик измененной антенны (1, 3) и лапки (2, 4) (по: Балкашина, 19286)
коготков, пульвиллу и характерную зазубренную щетинку (рис. 8, ж). Таким образом, «строение органа, образующегося вместо аристы, является вполне сходным со строением лапки. Следовательно, у мух «aristopedia» произошла замена аристы усика на лапку. Это дает право говорить о гомологичности лапки и аристы» (Балкашина, 1928, б, с. 97). Тем временем
87
базальные членики антенны «сохраняют все характерные особенности, свойственные роду Drosophila'». Ill членик, за исключением дистального конца, покрыт вполне нормальными чувствующими палочками вперемежку с мелкими и т. д. Балкашина приводит литературные данные о чувствующей ямке и III членике у рода Drosophila и проделывает гистологические исследования над мухой «дикого типа» и мухой aristopedia, устанавливая идентичность чувствительного органа у нормальной и измененной антенны. Эти результаты показывают, что структура, замещающая антенну у мух aristopedia, в большей своей части сходна с лапкой и в меньшей (проксимальной) — с антенной, причем переходная зона очень невелика и позволяет заключить, что для развития имагинального диска антенны имеются два альтернативных пути, каков бы ни был механизм ветвления. Таким образом, гомеотическая геновариация агр действует как триггерный механизм, направляя развитие зачатка по тому или другому пути.
Гистологическое исследование позволило выяснить причину, почему измененная антенна может двигаться только вся сразу и не может сгибаться и разгибаться на своем протяжении: мускулатура в лапкообразном органе отсутствует; нет характерного сухожильного тяжа.
Геновариация aristopedia выражается также в изменении лапок на всех ножках мухи (плейотропное действие). Число члеников вместо пяти уменьшается до четырех, иногда до трех. Особенно подвержена изменению задняя пара ножек, слабее передняя и менее всего — средняя пара. Таким образом, ген aristopedia влияет на строение всех членов меристического ряда насекомого.
Развитие. Балкашина нашла, что сегментация имагинальных дисков антенн начинается у нормальных мух на 4—4,5-й день личиночной жизни, т. е. незадолго до окукливания, и заканчивается на стадии трех-пятичасовой куколки. Сегментация имагинального диска у антенн aristopedia начинается раньше, на стадии двухдневной личинки, вскоре после начала сегментации ножек (рис. 9, а).
К моменту, когда начинается сегментация нормальной антенны, т. е. когда бороздка отшнуровывает первый сегмент, диск измененной антенны разделен на пять сегментов (рис. 9, б). На стадии трехчасовой куколки заканчивается сегментация как нормальной, так и измененной антенны — нормальная состоит из трех, а измененная из семи или восьми сегментов (рис. 9, в).
Далее дифференциация нормальной антенны состоит в том, что дистальный участок III сегмента вытягивается и превращается в аристу. У аристо-педии дальнейшее развитие идет по пути формирования лапок: сегменты становятся цилиндрическими и покрываются толстой хитиновой оболочкой и т. д. (рис. 9, г).
В результате Балкашина заключает, что «действие гена в течение эмбрионального развития мухи сказывается в стимуляции имагинальных дисков антенн к сегментации в более ранний период развития, а именно в период сегментации ножек. Дифференцировка органа также идет по пути развития ножек, а не антенн» (Балкашина, 19286, с. 105)
Действие гомеозисных локусов. На основе описанных Балкашиной фактов развития aristopedia Р. Гольдшмидт построил общее объяснение действия гомеотических локусов (Goldschmidt, 1938). Факт возникновения наследственной замены аристы на лапки у дрозофилы является доказательством того, что «меристические зачатки даже таких высоко дифферен-
88
Рис. 9. Имагинальный диск нормальной мухи (/) и aristopedia (2)
а — на стадии двухдневной личинки; б — у четырехдневной личинки; в — у трехчасовой куколки; г —у 25-часовой куколки (по: Балкашина, 19286)
89
цированных организмов, как Diptera, сохраняют способность менять направление своего развития, превращаясь в своего гомолога, т. е. являются изопотенциальными» (Balkaschina, 1929, с. 462). По Гольдшмидту, на ранних стадиях имагинальные диски изопотенциальны и становятся зрелыми (т. е. получают способность быть детерминированными) на определенной стадии развития, каждый на своей. Зародыш выделяет ряд эвокаторов, по одному на каждый тип дисков и каждый в должное время, чтобы воздействовать на тот диск, который только что созрел. На уровне второго дня личиночной жизни (или раньше) выделяется эвокатор, который диффундирует в шесть ножных дисков и вызывает ритмичное разделение дистального конца зачатка, приводящее к типичной сегментации по пути развития лапок. Обычно только ножные диски находятся в состоянии, способном реагировать на такую индукцию, но любой зрелый диск будет отвечать на эту индукцию формированием лапок. Рассматриваемое с позиций Гольдшмидта действие гена aristopedia сводится к ускорению ранней дифференциации антенного диска, приводящей его к созреванию в то же время, что и ножные диски; образование же лапок на антенне вызывает эвокатор лапок, а не сам ген aristopedia. У proboscipedia появление лапко- или аристоподобных структур можно объяснить в этих терминах как результат сдвига времени детерминации лабиальных дисков и совпадения его со временем детерминации ножек или антенн. Получение фенокопий некоторых из гомеотических мутантов указывает, что такой сдвиг может быть обусловлен также и внешними факторами.
Браун (Braun, 1940) предпринял попытку экспериментально подтвердить эту интерпретацию действия гомеозисных мутаций. Он пересаживал, по методу Бидла-Эфрусси, диски антенн aristopedia в личинки «дикого типа» (и наоборот) и нашел, что зачатки антенн у мух из линии aristopedia не изменяются под действием диффундирующих веществ, содержащихся в жидкостях тела нормальной личинки хозяина, так как они образуют лапкоподобные структуры. Трансплантация шла на стадиях от 2,5-дневной личинки до окукливания (до возраста 2,5 дня трансплантация по методу Бидла-Эфрусси невозможна из-за маленького размера личинки), и Браун заключил, что время детерминации развития приходится на очень раннюю стадию личиночной жизни (до 21 /2 дней).
Некоторые последующие экпериментальные исследования давали результаты, которые не могли быть непосредственно объяснены гольдшмидтовой теорией действия гомеозисных локусов и требовали либо ее модификации, либо построения совершенно новой теории действия гомеозисов в онтогенезе.
Физиология. Другая линия экспериментальных работ связана с исследованием взаимодействия гомеозисных и морфогенетических мутаций, а также взаимодействия аллелей в компаундах. С тех пор, как Стёртевант в 1929 г. обнаружил, что ген aristopedia Балкашиной является аллелем spineless (ss) и дал ему символ ssa, было найдено еще несколько гомеозисных аллелей этого локуса. Аллель aristopedia Спенсера (ssaSP) характеризуется более сильным изменением, aristopedia Бриджеса (ssaB) — более слабый аллель: он часто дает лишь небольшое утолщение основания арист. (В настоящее время интенсивно исследуется по крайней мере пять гомеозисных аллелей локуса spineless-aristopedia; см. обзор: Garcia-Bellido, 1977.)
Браун и Уоддингтон доказали, что не только анатомически, но и физиологически антенн мух ssa напоминает ножку, а не антенну. На антенную лапку мух ssa влияют «ножные» гены dachs, four-jointed, approximated, 90
thickoid, но не влияют «антенные* гены thread, aristalless (Braun, 1940; Waddington, 1940; см. также: Уоддингтон, 1947). Таким образом, гены могут проявлять свой специфический эффект на соответствующих им структурах независимо от того, за счет каких других генов образовались эти структуры.
Уоддингтон указал, что в компаундах аллелей, например в случае ssaSp/ssaB (или: ssaB/ssa), «гены ведут себя так, словно они отличаются лишь количественно, и по своему фенотипу каждое такое сочетание занимает промежуточное положение между двумя гомозиготами, от которых оно произошло* (Уоддингтон, 1947, с. 125). Подтверждая мысль Балкашиной, он подчеркивал, что, каков бы ни был механизм ветвления, имеются два альтернативных пути для развития имагинального диска антенны: «Эти альтернативы резко противостоят друг другу. У мух ssaB ариста в целом не имеет строения, промежуточного между нормальной аристой и аристой, сходной с конечностью... Вместо этого базальная часть аристы приобретает строение, почти полностью совпадающее со строением лапки, и лишь узкая переходная зона соединяет эту часть с дистальной областью, совершенно аристоподобной. Безусловно здесь играют существенную роль какого-то рода пороговые явления* (Там же, с. 127). Так, отправляясь от наблюдений Балкашиной, Уоддингтон развивает представления о «системе разветвляющихся путей* в развитии (the «branching track* development). Обсуждая эмбриологическое понятие компетенции, он говорит, что резко противоположные способы развития возникли в процессе эволюции в связи с необходимостью дифференцировки определенных и притом различных видов тканей. «Прогрессивный в эволюционном отношении генотип должен поэтому определять систему разветвляющихся путей развития* (Там же, с. 131).
Температурные опыты с гомеозисными мутациями. О влиянии температуры на частоту проявления гомеозисных мутаций впервые сообщил Б. Л. Астауров, установивший, что проявление признака tetraptera резко зависит от температуры, изменяясь от 0—1% при 17° (ок. 9% при 21°) до 35% при 25° (см.: Астауров, 1974, с. 101 и сн. 1 на с. 63. Температурные опыты Астаурова не были опубликованы). Позже К. Вилли нашел противоположные отношения для tetraltera: проявление падало от 35% при 14,4° до 1% при 29° Этот контраст отношений между температурой и проявлением интересен в связи с противоположными онтогенетическими процессами, вовлеченными в образование мутантных признаков: в фенотипе tetraptera из зачатка жужжальца развивается крылышко, в фенотипе tetraltera их зачатка крылышка — жужжальце. Действие температуры на развитие сводится в общем к дифференцированному ускорению (или замедлению) одних процессов относительно других. Поэтому можно предположить, как делает Вилли, что формирование крылышка вместо жужжальца требует дифференцированного ускорения некоторых реакций, достигаемого действием повышенной температуры во время или перед детерминацией и дифференциацией дисков крылышка и жужжальца. Формирование жужжальца вместо крылышка требует торможения тех же процессов (Villee, 1942, а, Ь).
Тот же тип противоположных отношений существует для развития в сторону аристы или в сторону лапки: оральные доли хоботка мух probosci-pedia трансформируются либо в аристоподобную структуру (при пониженной температуре), либо в лапкоподобную (при повышенной). Заметим в скобках, что’ рЬ представляет случай иерархии стрелочных механизмов: главный
91
«стрелочник» (ген) направляет развитие на путь формирования членистого придатка, подчиненный (t°) определяет направление дифференциации окончания этого придатка.
Уместно привести мнение Уоддингтона о том, в каких пределах правомерны выводы из температурных опытов. Обсуждая временное протекание генных реакций в связи с системой разветвляющихся путей, он останавливается на предположении некоторых авторов, что найденный у ряда мутантов температурно-чувственный период должен совпадать с тем временем, когда эти мутантные гены оказывают решающее воздействие. И возражает, что мы вправе сделать лишь ограниченный вывод, «что с указанным временем совпадает чувствительный период последовательных реакций, обусловленных мутантным геном. Мы не можем, следовательно, как указал Генке (1937), считать, что этот чувствительный период является тем временем, когда мутантный ген впервые начинает действовать, вызывая отклонение развития мутанта от нормального развития» (Уоддингтон, 1947, с. 130). Самый факт наличия ограниченного чувствительного периода говорит за то, что процесс состоит из последовательности реакций различной природы и с разной чувствительностью к внешним воздействиям. (Подробнее о температурных исследованиях см. гл. 7.)
НАСЛЕДСТВЕННЫЕ ГОМЕОЗИСЫ: СВОЙСТВА И ИНТЕРПРЕТАЦИИ
Гольдшмидт: «континуальная модель» хромосомы. Балкашина картировала ген aristopedia на 59,2 — 59,5 морганиды III хромосомы. В первом же кратком сообщении она обратила внимание на замечательный факт: ,,В том же отрезке этой хромосомы лежат гены bithorax, bithoraxoid и tetraptera, т. е. все гомеотические изменения Drosophila melanogaster* (Балкашина, 1928а, с. 170—171)4. Когда позже были обнаружены и картированы другие наследственные гомеозисы дрозофилы, новые данные лишь подтвердили это указание Балкашиной (proboscipedia — 47,7; tetraltera — 48,5; tetraptera— 51,3; spineless-aristopedia — 58,5; bithorax — 58,8; bithoraxoid — 58,8+ и т. д.).
Рассматривая наследственность в функциональном аспекте и отвергая понятие индивидуального «гена», Гольдшмидт развивает «континуальную модель» хромосомы (Goldschmidt, 1940). Факт топографической связи гомеозисных локусов, сообщенный Балкашиной, он использовал для подтверждения своей идеи, что дискретный «ген», полезный в целях описания, в действительности не существует и что в управлении развитием признаков «дикого типа» хромосома действует как целое. Отсюда, замечал он, было бы неверно заключить о случайности скопления гомеозисных генов в этом участке. Напротив, данный участок правого плеча III хромосомы действует как наименьшая единица в нормальной детерминации и развитии имагиналь-ных дисков. Различные же нарушения структуры этого участка хромосомы, т. е. гомеозисные мутанты, приводят к разным аберрантным фенотипам (обсуждение: Allen, 1974).
4 Гены картируют по кроссоверным данным и точность определения зависит от числа мух скрещивания. Поэтому, когда ген aristopedia был признан аллелем хорошо изученного локуса spineless, несколько изменилось положение, приписываемое ему на III хромосоме (на 0,7% перекреста). В первом кратком сообщении (1928а) в результате опечатки неверно приведено это значение: «69,2 морганиды», вместо 59,2.
92
В своем стремлении связать наследственность и развитие с эволюцией Гольдшмидт подчеркивал значение фактов, полученных при изучении агр (и других гомеозисных мутантов), также и для общего анализа эволюции. Именно на этих данных он строил свою концепцию «счастливого урода» (hopeful monster), т. е. системных мутаций, и макроэволюции. Однако с позиций нынешних представлений о процессе эволюции более интересно рассмотрение феномена гомеозиса в связи с эволюционной ролью дупликации генов и эволюционной ролью олигомеризации гомологичных органов (ср.: Sturtevant, 1925; Догель, 1954; Ohno, 1970).
Кольцов: неотения и эволюция. Еще одним замечательным свойством гомеозисов является глубокий атавизм мутантных фенотипов, что указывает на полную обратимость (в некоторых отношениях) процесса эволюции. «Геновариация aristopedia»,— писала Балкашина (1928а, с. 171),—«интересна не только в биологическом, но и в систематическом отношении, выбивая мух «aristopedia» не только за пределы вида, но и из таксономических групп высшего порядка». Измененные ротовые части у мух proboscipedia напоминают грызущий ротовой аппарат низших насекомых; мутация tetraptera меняет признак всего отряда Diptera и т. д.
Эволюционное значение регресса (понимаемого в смысле упрощения организации: паразитизм, неотения и пр.) Н. К. Кольцов обсуждает в очерке «Проблема прогрессивной эволюции» (1933, переиздание: 1936). Регрессу он отводит такую значительную роль в эволюции, что необходимость его учета стала одним из мотивов, чтобы оставить родословные схемы типа «одинокого ветвистого дуба» ради типа «манговой заросли» — целая группа стволов, растущих кверху с бесчисленными спускающимися книзу отростками, отходящие побеги которых могут снова подниматься вверх (ср.: Астауров, Рокицкий, 1975, гл. 7; Тимофеев-Ресовский, 1965, с. 9).
Рассматривая изменение генотипа при неотеническом упрощении организации, Кольцов касался наследственных гомеозисов дрозофилы. Он рассуждал так. Неотения может ограничиться отдельными органами, остановка которых на ранней недоразвитой стадии происходит иногда в эволюции видов или даже целых отрядов (tetraptera) без значительного осложнения генотипа под влиянием одного только гена. «Мы имеем основание утверждать, что таково именно первоначальное происхождение антенн, хоботковых лопастей и галтеров у двукрылых» (Кольцов, 1936, с. 519—520). Для каждого из этих органов у дрозофилы получены мутантные гены: bithorax I, bithorax II, tetraptera, aristopedia, proboscipedia, устраняющие эту неотению и возвращающие затронутые ею органы в прежнее состояние. Усики и хоботковые лопасти принимают форму членистых ножек, жужжальца превращаются в задние крылья. «Очевидно, что миллионы лет назад неотения возникла у родоначальников отряда Diptera под влиянием немногих генов, подавивших развитие соответствующих органов, а теперь в опытах Бриджеса, Балкашиной и Астаурова эти гены мутировали обратно или же возникли новые гены, сразу отомкнувшие возникший в древние времена запор» (Там же). Но за период времени с момента редукции этих органов многочисленные гены, отмечающие за их нормальное развитие, «вследствие отсутствия подбора деградировали, частью растерялись» (Там же). Именно этим объясняется тот факт, что задние крылья, усиковые и хоботковые ножки у этих мутаций развиваются неправильно, уродливо и с большими индивидуальными вариациями — включая полярную противоположность выражения sensu Vogt.
93
Далее Кольцов обращает внимание на факт тесного соседства известных к тому времени (1933 г.) гомеозисных мутаций. Его интерпретация — в противоположность гольдшмидтовской — звучит сейчас вполне современно и ориентирует на определенный путь исследований. «Любопытно, что все четыре перечисленных гена дрозофилы [собственно пять генов: Кольцов не отличал tetraptera от bithorax I или II] помещаются на очень коротком участке третьей хромосомы. Так как в наших культурах гены — отмыкате-ли неотенических запоров — возникли совершенно самостоятельно, независимо друг от друга, то топографическую связь между ними в хромосоме следует отнести к давно прошедшим периодам эволюции двукрылых. Возможно, что толчком к обособлению этого отряда послужило новообразование одного первоначального гена неотении, останавливавшего развитие первичного насекомого на той стадии эмбриогенеза, когда только начинали дифференцироваться задние крылья, жующие ротовые части и антенны. Но при дальнейшей эволюции отряда этот единый ген-запиратель дифференцировался и распался на отдельные локусы...» (Кольцов продолжает: «...как ген scute распадается, по мнению некоторых генетиков, на отдельные, но еще связанные между собой центры») (Кольцов, 1936, с. 519—520). Однако объяснение топографической делимости фенотипического эффекта scute с помощью, скажем, гипотезы о роли инсерций и position effect, представляется более вероятным, чем с помощью топографической делимости самого гена. (Такую трактовку явления действительно предложил в 1982 г. X. Модолелл из Мадридского университета.) Напротив, к теме Кольцова может иметь отношение работа Стертеванта (Sturtevant, 1925), в которой ряд явлений локуса Ваг объясняется на основе дупликаций этого локуса в результате неравного кроссинговера. Дуплицированные локусы в отсутствие давления отбора могут дифференцироваться (на пути «нейтральных замещений») и приводить к картине, схожей в главных чертах с наблюдаемой у гомеозисных мутаций. «В настоящее время вместо единого гена неотении мы имеем целый отрезок, на котором сосредоточены гены, задерживающие развитие отдельных органов в мухе. Обратные мутации, отмыкающие неотенические запоры, происходят поэтому в отдельных локусах независимо друг от друга. Таким образом, результаты экспериментальных работ по генетике дрозофилы позволят нам, быть может, вскрыть природу одного мутационного толчка к неотении, который имел место миллионы лет назад и о котором не сохранилось ясных палеонтологических данных» (Кольцов, там же).
Затем Кольцов отмечает, что резкая неотения (например, созревание половых органов на ранней личиночной стадии, подобной трохофоре анне-лид) ведет за собой сначала сильное упрощение только фенотипа, тогда как генотип сохраняет свою сложность. Участки хромосом, проявляющиеся обычно на утраченных стадиях, теряют свою активность и выходят из-под контроля отбора. Еще в кольцовское время было установлено, что большие участки Х-хромосомы и почти вся Y-хромосома у дрозофилы находятся именно в таком неактивном состоянии (по нынешней оценке доля «молчащей» ДНК у Drosophila melanogaster приближается к 90%). Кольцов предполагает, что такое состояние хромосомного аппарата может служить подтверждением того, что в развитии насекомых большую роль играли неотении. С другой стороны,— рассуждает Кольцов, высказывая мысли, которые позже лягут в основу представлений об «эволюции путем дупликации генов» и некоторых других гипотез, касающихся генетических меха-
94
НИЗМОВ прогрессивной эволюции (ср.: Ohno, 1970),— «запас непроявляю-шихся в развитии генов, которые могут мутировать в гены, проявляющиеся в развитии уже неотенической формы, влечет за собой высокую изменчивость последней и позволяет ей иногда обнаружить в дальнейшем пышный расцвет прогрессивной эволюции. Мы наблюдаем такой расцвет у полипов, коловраток, вероятно, у первичных костистых рыб, птиц и млекопитающих» (Кольцов, там же).
Наследственные гомеозисы: резюме. Наследственные гомеозисы вызывают в онтогенезе изменения большого масштаба, что доставляет им исключительное положение среди дрозофильных мутаций. Основная черта гомео-тических мутаций в том, что их действие сводится к выбору одного из каналов, по которому пойдет формообразующий процесс. Гомеозисные мутации определяют развилки в онтогенезе (Балкашина).
С таким триггерным («стрелочным» — Уоддингтон) действием этих «отмыкателей неотенических запоров» (Кольцов) соотносится поразительный атавизм гомеотических изменений. Они «выбивают» мутантных мух «не только за пределы вида, но и из таксономических групп высшего порядка» (Балкашина); при этом «мы имеем не простую задержку или нарушение нормального онтогенеза, а восстановление в существенных чертах каких-то давно нарушенных причинных связей, как бы полную обратимость (в некоторых, конечно, отношениях) эволюционного процесса» (Астауров).
Отсюда становятся понятны и некоторые второстепенные характеристики наследственных гомеозисов. В первую очередь — непостоянство фенотипической реализации, обязанное, по Кольцову, генетической дегенерации в защищенном от контроля отбора неактивном состоянии тех локусов, которые в эволюционном прошлом отвечали за стабилизацию исходного канала развития. Изменчивость выражения tr, агр так велика, что это дало основание Фогту говорить о полярной противоположности между пере-и недоразвитием. У некоторых из наследственных гомеозисов велика также вариация процента проявления. Обладая низким и вариирующим проявлением, tr дала яркий случай отсутствия корреляции сторон, анализ которого вылился в принцип автономной изменчивости структур, развивающихся из отдельных имагинальных дисков (Астауров). Наконец, топографическая близость гомеотических локусов наводит на мысль о некотором неотени-ческом гене-родоначальнике, который путем дупликаций и нейтральных замещений дал всю серию гомеотических генов (Кольцов) — мысль, лежащую в основе обсуждения генетических механизмов процесса эволюции в 1970-е годы.
95
Глава 7 ТЕМПЕРАТУРНО-ЧУВСТВИТЕЛЬНЫЙ ПЕРИОД: КЛЮЧ К МЕХАНИЗМУ ОНТОГЕНЕЗА
Афоризм, вынесенный в название этой главы (по названию статьи Драйвера — Driver, 1926), отражает тот факт, что в 20-е и 30-е годы анализ температурно-чувствительных периодов был одним из основных методов изучения индивидуального развития с позиций генетики. (В 1956 1967 гг. П. Г Светлов построил теорию критических периодов общебиологического масштаба, связавшую вопросы эмбриологии, тератологии, физиологии, генетики, эволюционной теории.)
ПРИРОДНАЯ ИЗМЕНЧИВОСТЬ ОКРАСКИ ГУСЕНИЦ
В анализе феноменологии реализации генов у дрозофилы четвериковцы, как правило, концентрировали внимание на дефинитивных признаках, на начале и конце цепи «ген — признак». Н. К. Беляев занялся средним звеном цепи — расшифровкой механизмов онтогенеза, отвечающих за изменчивость в природных популяциях. В экспериментальных работах его на Lepidoptera просматриваются интересы натуралиста par excellence, которые сближали его с его учителем С. С. Четвериковым — крупнейшим систематиком бабочек Первая работа этого цикла начинается словами: «Во время моих летних энтомологических экскурсий в Костромской губернии, несколько лет тому назад, мне приходилось иногда собирать довольно значительные количества гусениц нескольких обычных русских видов Arctiidae Из этой группы четырех близко родственных видов два... казались мне в особенности интересными, так как их гусеницы обнаруживали чрезвычайно боль шую изменчивость в окраске... Этот факт столь значительной изменчи вости, хотя и отмечался в специальных сводках и атласах по палеарктическим бабочкам..., однако приводился лишь как таковой, без всяких указаний на какие-либо факторы, вызывающие эту изменчивость» (Беляев, 1926, с. 157).
К концу XIX в. было показано, что определенные дневные и ночные бабочки, которые долгое время рассматривались как редкие роды и виды, на деле получаются от некоторых из наиболее частых видов при необычных климатических условиях. Благодаря температурным опытам ряда авторов (обзор: Bachmetjew, 1901 —1907) были объяснены (помимо случаев появления индивидуальных вариаций и аберраций в природе) также многие случаи географической изменчивости и сезонного диморфизма. Эти опыты велись прежде всего на половозрелых формах. Между тем две другие стадии развития, гусеницы и куколки, «представляют из себя не менее' благоприятные объекты для подобных опытов... При этом, в силу некоторых особенностей этих стадий, здесь возможна постановка опытов физиологического характера, едва ли разрешимых на imago...» (Беляев, 1928, с. 107).
1 При подготовке «Избранных трудов» С. С. Четверикова выяснилось, что его натуралистические работы настолько актуальны в наши дни, что им следовало отвести отдельный том: С. С. Четвериков. Фауна и биология чешуекрылых. Новосибирск, 1984.
96
Наблюдающаяся в природе изменчивость окраски гусениц разных Lepidoptera бывает двух типов. В одном случае различные формы окраски могут быть расположены в непрерывный вариационный ряд и носят характер модификационного полиморфизма; их ближайшие причины нужно искать прежде всего в меняющихся условиях существования. В другом случае они носят альтернативный характер; такой диморфизм основан, как можно ожидать, на менделировании наследственных признаков. Первая работа этого цикла представляет результаты температурных опытов с гусеницами Spilosoma lubricipeda. Многоядность гусениц и их открытый образ жизни и как следствие этого — подверженность всевозможным атмосферным влияниям при флуктуирующем типе изменчивости делали наиболее вероятным предположение, что именно эти причины и являются факторами изменчивости. Литературные данные выдвигали на первый план в качестве факторов изменчивости свет и характер пищи, тем более что различные окраски часто носят приспособительный характер. Поэтому Беляев начал в 1923 г. с опытов по влиянию питания и освещенности на окраску гусениц Sp. lubricipeda^ результаты их были отчетливо отрицательными.
Аналогичные опыты 1921 г. с Phragmatobia fuliginosa (еще более широкого масштаба) также дали отрицательный результат. Тогда же, летом 1923 г., всех оставшихся от опытов на чувствительность к свету гусениц предпоследнего возраста (при обычной для лаборатории температуре 23° гусеницы линяют обычно 6 раз) Беляев употребил на предварительный опыт, поместив их в повышенную температуру, около 30° С. «Вылупившиеся в последний раз гусеницы настолько отличались от живших в нормальной температуре, что после этого уже не оставалось никаких сомнений в том, что именно в этом направлении и следует продолжать работу» (Беляев, 1926, с. 164).
В опытах 1924 г. регистрация окраски производилась по детально разработанной шкале из 11 классов, в построении которой учитывалось два элемента окраски: пигментированность тела и щетинок. (Тело гусеницы покрыто бородавками; каждая из них несет ряд светлых и темных щетинок. Пигмент, окрашивающий гусеницу, локализован в хитине: на сплошной хитиновой мембране сидит масса «колбочек», сильно окрашенных в темных местах, прозрачных — в светлых.)
Первая серия опытов, поставленная для выявления «уровня отсчета» при нормальной температуре 23°, показала, что основной тенденцией в развитии окраски является потемнение — усиление меланизации. Этот процесс совершается постепенно, вследствие чего все возрастные формы можно расположить в один последовательный восходящий ряд классов (случай «направленной изменчивости» sensu О. Фогт). Две серии опытов с повышенной температурой (27,5 и 31,5°) показали, во-первых, что окраска гусениц чрезвычайно чувствительна к температурным воздействиям на всех стадиях развития, во-вторых, что повышение температуры, не внося в развитие окраски качественных изменений, замедляет его.
Поскольку роль внешнего фактора в развитии должна оцениваться не только по эффекту в данный момент, но и по длительности эффекта, Беляев поставил опыты по действию переменной температуры. Светлые тепловые формы VII возраста получались одни и те же, когда при t = 31,5° шло: а) все развитие; б) три последних возраста; в) два последних; г) только один последний возраст. Аналогично этому темные холодовые
7
В В. Бабков
97
формы могли быть получены действием t = 20 или 17° на V и VI (или только VI) возраст, независимо от того, проходили ли предыдущие стадии, а) при нормальной t = 23° или б) при повышенной t = 31°, или в) при t = 27,5° в течение I—IV возрастов, а в V возрасте при t = 33° Аналогии ные результаты были получены и в опытах, поставленных по другим схемам. Итак, «окраска каждой стадии развития есть реакция лишь на температуру предшествовавшей стадии» (Беляев, 1926, с. 183). Каждый возраст по отношению к температурному фактору оказывается самостоятельным, и действие определенной температуры тотчас же аннулируется, как только она меняется. Поэтому после экспозиции в течение одного или нескольких возрастов нельзя рассчитывать на получение светлых форм в следующем поколении: «Я не делал опытов по наследованию полученных мной изменений, так как в виду всего вышеизложенного такая попытка кажется мне лишенной всякой надежды на успех» (Там же, с. 184).
Разбив продолжительность всего возраста на отдельные части (длительный период А — все время питания и роста; более короткий В — процессы отслоения нового хитина, образования новой кутикулы и ее пигментации; самый короткий С — от конца В до сбрасывания шкурки), Беляев устано вил, что тепловые условия определяют будущую окраску именно в период линьки (В). Результаты опытов с влиянием температуры на другие элементы окраски показывают, что окраска на данной стадии под влиянием по вышения температуры от 21 до 28° отстает в развитии приблизительно на один возраст. Это дало Беляеву основание считать светлые тепловые формы окраски недоразвившимися, а темные холодовые — переразвив-шимися.
Температурные условия оказывают существенное влияние и на продолжительность линьки (стадия В). При 18—2Г первая линька длится около 24 ч., третья — около 34—36 ч., шестая около 56—58 ч. При повышенной температуре гусеница линяет быстрее, при пониженной — дольше. В табл. 9 даны результаты четырех опытов с тремя, четырьмя, пятью и шестью линьками, по 10 гусениц на одной и той же стадии в каждом опыте: пять оставляли в обычных условиях: 16—22,5°, другие пять держали при t = 30° Тепловое укорочение линьки было наибольшим, если все гусеницы только что подготовились к линьке — начало стадии В (опыт 1 и 4), и достигало здесь одних суток. Если процесс линьки уже продвинулся, то ускоряется лишь его остаток (опыт 2); если он закончился (опыт 3), то гусеницы линяют одновременно. Влияние температуры рисовалось Беляеву следующим образом. Когда во время линьки процессы роста отчасти уже продвинулись и начинаются процессы пигментации, то те и другие пойдут парал лельно. Если обозначить большими буквами А, В, С и т. д. некоторые моменты в процессах роста, а малыми а, Ь, с и т. д.— соответствующие им моменты пигментации, то это можно изобразить схемой:
А В С D Е F
abed е f.
Оба процесса идут таким образом, что когда наступит момент сбрасывания старого хитина и будет достигнута точка F, гусеница имеет стадию окраски [.
Если температура повышена, то согласованность обоих процессов нарушается, и к моменту сбрасывания хитина F окраска недоразвивается:
А В С D Е F а Ь с d е
98
Таблица 9 Температурные опыты с гусеницами (по: Беляев, 1926)
Номер опытов Линька Температура, °C Стадия Начало опыта Момент линьки Продолжительность, ч Окраска
1 3-я 16 30 В в 20 ч 17. VII 20 ч 17. VII 10 Ч 19.VII 10 Ч 18. VII 38 14 Темные Светлые
2 4-я 21 30 в в 16 ч 24. VII 16 ч 24. VII 12 ч 25. VII 3—6 ч 25. VII 20 11 — 14 Темные Светлее предыдущих
3 5-я 22,5 30 Конец В или С Конец В или С 12 ч 26. VII 12 ч 26. VII 12—15 ч 27. VII 12—15 ч 27. VII 24—27 24—27 Темные
4 6-я 19,5 30 В В 20 ч 31. VII 20 ч 31. VII 6 ч 3. VIII 6 ч 2. VIII 58 34 Светлые
Понижение температуры вызывает противоположное нарушение согласованности. Процессы роста сильно замедляются и развитие окраски переходит норму:
А В С D Е F a b с d е f g.
Гипотеза Беляева объясняет, почему тепловые формы имеют светлую окраску, которая носит характер недоразвития, а холодовые — темную, переразвитую окраску; почему отсутствует последействие; почему эффективность воздействия локализована в период линьки. «Все эти явления есть следствие нарушения координации между процессами роста и пигментации» (Беляев, 1926, с. 210).
Установленный Беляевым факт, что гусеницы Sp. lubricipeda при повы-шенной температуре светлеют, при пониженной темнеют, явился только добавлением к громадному числу аналогичных случаев, известных из области экспериментальной энтомологии. Новым здесь было то, что для действия температуры был указан механизм: нарушение синхронности процессов развития и пигментообразования. Более того, важен именно механизм, а не фактор: «Температура не имеет ни специфического, ни прямого Действия: всякий другой фактор, влияющий на продолжительность линьки, будет вызывать те же изменения, которые с этой точки зрения являются прямой функцией времени» (Там же, с. 209).
Для Н. К. Беляева характерно, что экспериментальное исследование
7*
99
Таблица 10
Природная изменчивость окраски гусениц (по: Беляев, 1926)
Место и время сбора гусениц Классы окраски Всего Среднее значе ние класса
3 1 6 L_l_ 1 8 1 1 9
Опушка леса и поляны, 1924 г. 5 51 32 49 17 8 1 163 5,3
Глубокий и тенистый овраг, 1924 г. 8 5 5 8 10 1 37 6,3
Опушка леса и поляны, 1926 г. — — 2 6 3 2 13 7,4
он считает нужным завершить обращением к изменчивости в природе. В § 16 цитированной работы он пишет: «Гусеницы Sp. lubricipeda, вообще не редкие, особенно часто встречались летом 1924 г. В этот год мне удалось собрать в окрестностях Звенигородской биологической станции, на различных растениях,- 200 гусениц этого вида. Большинство из них было собрано на более или менее открытых местах — по опушкам леса, на полянах и т. д. Меньшая часть была найдена на зарослях крапивы в глубоком и затемненном овраге около станции. Обе станции, без сомнения, отличались между собой по инсоляции и вообще по температурным условиям. Соответственно этому, первые гусеницы в общем были светлее вторых (табл. 10). Наоборот, холодное и неблагоприятное лето 1926 г. повело к тому, что гусеницы вообще и в частности Sp. lubricipeda были очень редки. При этом все 13 экземпляров, которые мне удалось найти в этот год при самых тщательных поисках, отличались темной окраской (табл. 10)» (Беляев, 1926, с. 214).
И далее: «Гусеницы Vanessa urticae L. с крайнего севера, как я имел случай убедиться во время поездки в Центральную Лапландию, гораздо темнее тех же гусениц из Средней России. Для других видов известно постепенное потемнение по мере поднятия в горы...» (Там же, с. 215). А в другой статье: «Сравнения исследованных трех видов показывают, что изменчивость их в природе и отношение к температурным опытам вполне совпадают...» (Беляев, 1928, с. 130).
Исследования Н. К. Беляева прямо указывают на то, что отсутствие абсолютной координированности процессов индивидуального развития в природных условиях является общим правилом; случайные же моменты могут играть роль на разных «этажах» иерархически организованных процессов индивидуального развития.
НАПРАВЛЕННАЯ ИЗМЕНЧИВОСТЬ В ОНТОГЕНЕЗЕ
Во «Вступлении» к интересной статье «О направленной изменчивости у Coccinellidae» С. Р. Царапкин (1930, с. 139) приводит работы, затраги вающие проблему направленной изменчивости. Ссылаясь на Беляева (кото рый «дает возрастную изменчивость рисунка гусениц и развитие пигмента
100
у этого вида») и других авторов, изучавших окраски бабочек, их куколок и гусениц, Царапкин отмечал: «Во всех этих работах с бабочками изменчивость окраски не рассматривается с точки зрения направленности, хотя определенная закономерность в развитии рисунков ясна из прилагаемых авторами таблиц» (Там же). Несомненно, для интерпретации одних и тех же данных в ту или иную сторону требуется особая ориентация сознания. Вопрос о направленной изменчивости возникал в современной биологии не однажды и обсуждался разными авторами с разных точек зрения (см.: Сутт, 1977; Бабков, 1978, 1980). Царапкин отмечал, что эмпирические факты направленной изменчивости у бабочек и птиц, легшие в основу теории ортогенеза Эймера, долгое время оставались вне поля зрения биологов. Причину этого он усматривал в том, что «автор внес в теорию ортогенезиса телеологическое объяснение филогенетического развития» (Царапкин, 1930, с. 139). Этому он противопоставляет подход О. Фогта, изучавшего «признаки, которые, увеличиваясь закономерно, направленно распространяются в пространстве». Строго направленную изменчивость в индивидуальном развитии, проявляющуюся без исключений (метаболий), фогт обозначил термином эвномия (Eunomie). «Это новое понятие должно, по мысли Фогта, заменить ортогенезис Эймера, с которым связаны телеологические и филогенетические представления» (Там же, с. 140).
Направленная изменчивость у Coccinellidae была найдена Шильдером и описана Фогтом (Vogt, 1926; Vogt, Zarapkin, 1929; Zarapkin, 1930—1938). Для количественного анализа явления, с целью построить общую его схему, которая могла бы служить рабочей гипотезой для дальнейшего экспериментального анализа этого вида изменчивости, Царапкин предпринял исследование изменчивости рисунка надкрылий у некоторых видов божьих коровок (Coccinellidae). Методами явились статистический анализ и биометрическая оценка различных форм; материалом послужили коллекции, собранные Франком и хранившиеся у Фогта (Царапкин в то время работал по научному обмену в Генетическом отделе руководимого Фогтом Института мозга). Для анализа было выбрано шесть видов, особенно сильно изменчивых и одновременно представленных достаточно большим числом (тысячами) экземпляров. Остановимся на исследовании одного из них — Coccinella 10-punctata, показывающего исключительно сильную изменчивость. Молодые, только что вылупившиеся жуки совершенно не окрашены. Окраска развивается постепенно; одна пара пятен надкрылий (элитр) возникает за другой в определенном порядке, образуя строго эвномический ряд. У части особей процесс пигментации останавливается на той или иной ступени; так возникают формы с неполным числом пятен. У других он доходит до характерного числа, образуя forma typica (1-й период). Иногда процесс идет дальше и получаются формы со слитными пятнами и сложным рисунком или же с добавочными пятнами. После завершения слияния пятен процесс пигментации идет в некоторых случаях дальше, светлые пространства основного фона сужаются и окружаются— возникает forma pustulata (2-й период). Наконец, процесс заканчивается полным почернением надкрылий, образуя forma unicolor nigra (3-й период).
Анализ первого периода пигментации С. Ю-punctata, т. е. периода закономерного возникновения пятен, привел автора к построению интересной рабочей гипотезы для анализа этого явления (рис. 10). На рис. 10, а Даны схемы трех основных форм этого вида. На грудном щитке развива-
101
Рис. 10. Изменчивость первого периода пигментации (по: Царапкин 1930) а — три основные формы пигментации;
б — формы в порядке нарастания числа пятен; в — все формы первого периода
6
1 2 3 Ч 5 6 7 8 9
W
10 11 12 13 14 15 16 17 18 19
1—Г
П/П
ется пять пятен; на каждом надкрылии forma typica имеется восемь пятен. Они образуют типичные три продольных ряда и три поперечные зоны. Первый (и второй) период пигментации протекает не строго направленно. В одном этом периоде насчитывается около 90 различных форм, и на первый взглад может показаться, что осуществляется любая случайная комбинация пятен. Этот вид представлен в коллекции Франка в нескольких тысячах экземпляров, что позволило подвергнуть каждую отдельную форму количественной оценке.
Рис. 10,в (ср. рис. 10,6) иллюстрирует изменчивость первого периода пигментации. По оси абсцисс расположены все формы в порядке нарастания числа пятен (т. е. №№форм). По оси ординат — число индивидов соответствующих классов.
Для оценки различных классов Царапкин распределяет их на четыре группы: в первую входят все классы с числом индивидов не более 10; во вторую — от 10 до 100; в третью — от 100 до 200; в четвертую — 200 и выше (табл. 11).
Таблица показывает, что вероятность появления какой-либо формы, относящейся к первой группе, незначительна. Напротив, к двум последним группам относится около 3/4 всего числа особей. (Из этого следует, в частности, что, исключив маловероятные формы, мы можем построить из оставшегося материала новый ряд, яснее характеризующий основную тенденцию первого периода процесса пигментации у С. 10-punctata.)
На рис. 10, б даны все формы первого периода пигментирования. В первом ряду — одна форма без пятен на элитрах, во втором — шесть разных форм с одной парой пятен, и т. д. до девятого ряда, где стоят формы с восемью парами пятен. Наиболее часто встречающиеся формы стоят на первом месте ряда, другие — в порядке уменьшения их частоты. (Цифры над рисунками обозначают количество экземпляров коллекции Франка).
Шесть форм, стоящих во II ряду, встречаются с разной частотой. Форма II-1 с 7-й парой пятен встретилась 152 раза, П-2 со 2-й парой — 64 раза, П-З с 4-й парой — 33 раза. (Три другие формы очень редки и их трудно расставить в том или ином порядке, поэтому Царапкин считает их в первом приближении равновероятными.) На этом основании Царапкин определяет «градацию легкости возникновения» отдельных пятен, пигментации поперечных зон и продольных рядов. Средняя поперечная зона оказывается наиболее чувствительной к образованию пятен, затем идет верхняя, наконец нижняя. Появление пятен в III продольном ряду вероятнее, чем в I и II (ряды считаются от продольного шва). Частоты возникновения отдельных пятен №№ 7, 2, 4, 1, 8 и 3 относятся между собой как 152 : 64 : 33 : 2 : 2 : 2.
Итак, уже на первых шагах пигментации С. 10-punctata определяется несколько направлений развития. Однако эти направления не равновероятны, рассуждает Царапкин, поэтому можно ожидать, что на следующих шагах одни комбинации пятен будут встречаться чаще, чем другие: комбинация пятен №№ 7 и 2 должна быть более вероятной, чем комбинации 7-го и 4-го или 2-го и 4-го пятен. Это последнее замечание подтверждается данными HI ряда таблицы рисунков (рис. 10, б), где частоты соответствующих комбинаций пятен (формы III-1, -2, -3) относятся как 152 : 96 : 56. Все иные ком-бинции, как и ожидалось, встречаются очень редко.
Подобный анализ всей таблицы (рис. 10, б) привел Царапкина к заключению, что закономерность, наметившаяся на ранних ступенях развития признака, сохраняется и на последующих. (Однако имеются уклонения
103
Таблица 11
Анализ первого периода пигментации (по: Царапкин, 1930)
Группа Число классов Общее число особей Среднее число особей на один класс Вероятность возникновения одной особи в определенной группе, % Вероятность возникновения одной особи в определенном классе, %
I 52 129 2,5 3,284 0,064
II 16 676 42 17,210 1,07
III 9 1270 171 32,332 3,589
IV 7 1853 265 47,174 6,746
III4- IV 16 3123 195 79,506 4,964
от предполагавшейся закономерности. Одно из них представляет форма IV-2; другое — формы V-1 и V-2, которые по предположению должны бы стоять в обратном порядке. Кроме того, все немногочисленные формы выпадают, разумеется, из статистической оценки.)
«Градация возникновения» пятен №№ 7, 2, 4 и совокупности трех последующих сохраняется и при комбинировании этих пятен. Напротив, пятна 1, 8, 3, 5, взятые по отдельности, ведут себя неправильно, часто меняясь местами на таблице. Эта «исключительность», предполагает Царапкин, может быть обязана отсутствию (или очень слабой) градации, которая характеризует три предыдущие пятна, т. е. эквипотенциальности их по отношению к образованию пигмента. Для проверки предположения Царапкин подсчитал, сколько раз возникает каждое из пятен:
Номер пятна 5 13 8 6
Число нахождений 2498 1954 1635 1562 624
Разница между пятнами 5, 1, 3, 8 по частоте невелика, тогда как пятно № 6 довольно резко от них отличается. Все эти подсчеты приводят к выводу о существовании «пятиступенчатой градации в процессе возникновения пятен» у 10-точечной божьей коровки.
Таким образом, анализ форм С. 10-punctata показал, что: 1) процесс пигментации первого периода носит определенный и закономерно направленный характер; 2) «эта направленность проявляется не в абсолютно строгой последовательности возникновения пятен, а в большей вероятности появления определенных пятен» (Царапкин, 1930, с. 150).
Этот анализ позволяет также построить направленный ряд форм С. 10-punctata; его ступенями будут наиболее вероятные сочетания пятен, соответствующие пикам графика на рис. 10, в, и построенный таким образом ряд будет состоять из первых форм девяти рядов таблицы (рис. 10, 6).
Причины эвномии: иерархичность и координированность онтогенеза. Процесс пигментации у кокцинеллид зависит от многих факторов, внутренних и внешних. Не имея почти никаких данных об этих факторах (оперируя, говоря кибернетическим языком, с «черным ящиком»), . Царапкин мог только чисто гипотетически разложить на составные части общую картину процесса. Первым из необходимых компонентов этого процесса является то или иное количество пигмента, сильно варьирующее у многих видов кокцинеллид. В тех случаях, когда пигмент образует определенный рисунок, возникающий постепенно, следует предположить наличие второго компонента — неодинаковых порогов раздражения одной и той же ткани
104
в различных местах. (Здесь Царапкин касается проблемы «ген и ткань», о которой шла речь в гл. 4.) Некоторые обстоятельства (см. ниже) заставили Царапкина предположить существование третьего компонента — гипотетического стимулятора пигментообразования: «Под ним я подразумеваю фактор, который необходим в определенном количестве для возбуждения процесса пигментообразования. Природа этого фактора для нашей цели безразлична: можно представить его как некое вещество или энергию, как тот или другой энзим, или извне действующий фактор» (Там же, с. 151).
В основу рабочей гипотезы Царапкина, которая представляет собой шаг к пониманию механизма этого процесса — шаг от «черного ящика» к «белому*,— положено представление о роли этих трех компонентов процесса пигментации. Они могут находиться в разных отношениях, чем и определяется конечный результат. Если действие трех факторов определенным образом согласовано, то возникают строго закономерные формы, образующие эвномический ряд, как схематически представлено на рис. 11, а. Прямоугольник NN изображает участок ткани надкрылья с пятью пятнами, имеющими разные пороги раздражения. Рост количества пигмента идет по линии АЕ, причем А обозначает количество пигмента, необходимое для образования одной пары пятен, В — для двух пар и т. д. Количество гипотетического стимулятора растет строго параллельно пигменту. Подобные соотношения компонентов пигментирования должны привести к образованию ряда форм со строго направленным процессом пигментации — эвно-мическим sensu Vogt. Необходимыми условиями эвномичности процесса являются резкое различие в порогах раздражения тех мест ткани, где возникают пятна, и абсолютная корреляция между ростом количества пигмента и силой действия гипотетического стимулятора.
Если пороги раздражения различаются слабо (как в случае пятен 1, 3, 5, 8, представленных здесь участком IV), то пигмент будет распределяться хаотично и будут возникать формы с любой парой пятен. Случай абсолютной координации количеств пигмента и стимулятора весьма маловероятен; значительно чаще должны наблюдаться случаи неполной корреляции — и вид, исследованный Царапкиным, дает два интересных примера нарушения этой координированности.
Числовые отношения между формами, изображенные на рис. 10, б, обязаны тому, что рост количества пигмента отстает от роста специфического стимулятора. Рис. 11,6 дает схематическое представление отношений компонентов в этом случае. Рост пигмента идет здесь в том же темпе, как и на предыдущей схеме; и в точке А его количества достаточно для образования одной пары пятен. Тем временем стимулятор (линия «ае») возрос настолько, что преодолевает пороги раздражения любой из пяти пар пятен. По мысли Царапкина, при таком соотношении факторов между пятнами установится градация вероятности возникновения — чем ниже порог раздражения данного участка ткани, тем выше вероятность появления там пигмента. (Градация вероятностей на рисунках схематически изображена интенсивностью штриховки кружков в прямоугольнике MN).
Материалы Царапкина дают также пример еще одного модуса нарушения координации. У разных кокцинеллид нередки формы с неполным количеством пятен, причем некоторые из этих пятен гипертрофированы (см. рис. 11, г). Этот случай можно объяснить только тем, что чрезмерное количество пигмента распределяется на недостаточное число пятен, когда стимулированы
105
I I I H Y I I JU U Y
E
Puc. 11. Схема соотношений количества пигмента (ЛЕ), порогов раздражимости (/—V), стимулятора пигментов (ае) (по: Царапкин, 1930)
а — согласованность — эвномический ряд; б — рост пигмента отстает от роста стимулятора; в — рост стимулятора отстает от роста пигмента; г — три исключительные формы, иллюстрирующие случай в
только пятна с самыми низкими порогами. Это новое соотношение схематически дано на рис. 16, в.
В основе направленной изменчивости лежит, по мысли Фогта и Царапки-на, неодинаковая чувствительность топически изолированных участков ткани к определенным раздражениям. (Именно здесь исследования Царапкина сходятся с темой «ген и ткань», с концепцией «полей действия генов» и общими схемами фенотипической реализации гена Астраурова, Беляева, Тимофеева-Ресовского.) Однако факт неравнопотенциальности ткани — не единственное условие направленности в изменчивости. Градации порогов чувствительности должны быть координированы с количеством пигмента и стимулятора.
Завершая свою статью, Царапкин высказывает некоторые соображения о случаях меньшей или большей эвномичности процессов. Направленная изменчивость, как она проявляется у кокцинеллид, связана с началом дифференциации тканей, поэтому и гисто-физиологическая координация неполна. Именно поэтому здесь необходим статистический метод. «В случаях, когда физиолого-гистологическая координация очень велика, эвномия должна быть очень полной. Поэтому можно ожидать, что признаки, объединяющие высшие систематические единицы (эвномически варьирующие роды и семейства), должны показывать особенно строгие эвномические ряды» (Царапкин, 1930, с. 153)—то, что Н. И. Вавилов назвал «гомологи ческими рядами в наследственной изменчивости».
106
\ ТЕМПЕРАТУРНЫЕ ОПЫТЫ С ДРОЗОФИЛОЙ
Раннй^ работы по определению периода в развитии, где температура может модифицировать окраску и другие структурные признаки, проводились главным образом на Lepidoptera (сводка: Bachmetjew, 1901 —1907). Затем работа велась также на дрозофиле. М. А. Ходж в 1915 г. показала, что мухи, несущие мутантный ген, отвечающий за редупликацию ног, дают изменения почти в 100% случаев, если личинки помещаются в критически низкую температуру в очень раннем возрасте (до ’|2 Дня), и только 10% измененных мух, если первые четыре дня развития проходят в нормальной температуре. Э. Робертс в 1918 г. нашел, что высокая температура уменьшает экспрессивность vestigial (т. е. увеличивает длину зачаточных крыльев), если ее воздействие приходится на раннее время личиночной стадии. Аналогичные опыты с расой vestigial провели Н. А. и В. Н. Ильины в 1927 г. Э. Сейстер в 1917 г. показал, что с ростом температуры уменьшается среднее число фасет bar и что критический период приходится на середину личиночной жизни. Крафка в 1920 г. подтвердил эти выводы в обширных опытах на расах bar и ultrabar. Р. Герш в 1924 г. нашел те же отношения у полноглазых дрозофил, при этом процент падения числа фасет на градус оказался ниже, чем у рас bar и ultrabar. А. Герш в 1924 г. в опытах на разных bar, ultrabar и нормальных глазах получил промежуточный процент падения между таковыми для bar и N. Ч. Зелени в 1920 и 1925 гг. показал, что действие температуры не наследуется, и т. п. (См. также: Bachmetjew, 1901 —1907; Driver, 1926; Luce 1926).
Имея возможность работать с различными «чистокровными» мутантными линиями и используя температуру — одно из лучших средств в распоряжении биолога для «вычленения» реакции без нарушения «нормальной» работы организма, можно не только определять «фенокритические стадии» sensu Haecker, но и ставить вопрос о механизме действия генов.
В исследовании А. Н. Промптова (1929) с геновариацией polymorpha, pm, (которую нашел Д. Д. Ромашов в популяции D. funebris из окрестностей Звенигородской биостанции), первоначальной задачей был обычный генетический анализ вновь обнаруженных несомненно наследственных изменений. И, что характерно для работы с funebris, «лишь в процессе расширения и изучения материала наметились те основные вопросы, имеющие и теоретическое значение», которые и стали темой работы.
У первых же исследованных особей изменения затрагивали сразу три основные группы признаков («поле действия гена») — строение крыльев, глаз, ножек; признаки трех типов зависели от одного основного рецессивного аутосомного гена (плейотропия). На крыльях появлялись radius incompletus, различные вырезки, нарушалось жилкование, крылья укорачивались, а в крайней степени выражения превращались в рудиментарные придатки (эвно-мический ряд изменчивости). Изменения глаз: нарушение распределения фасеток («грубые глаза») и разные степени редукции их числа. Изменения ног затрагивали число члеников лапки: от пяти нормальных — до трехчетырех и даже одного.
Разные стадии трех рядов изменений комбинировались у одной особи весьма разнообразно, однако замеченные особенности (сильные изменения крыльев большей частью сопровождались ненормальным строением глаз и лапок, и напротив, особи со слабо измененными крыльями имели почти всегда нормальные глаза и лапки; и т. п.) позволили заключить об опреде
107
ленной эвномичности развития признака в ряду «крылья — глаза и ножки»: «фенотипическая реализация изменений в крыльях при определенных условиях более легка, чем реализация изменений в глазах и ножках» (Там же, с. 155; ср. понятие «биоклизы» О. Фогта.) В этом свете становится понятным факт наблюдавшегося Промптовым «обособления» линии фенотипически «radius incompletus», т. е. pm с ослабленным выражением по крыльям и 100%-ным непроявлением по глазам и ножкам. Линия эта была получена в ходе выяснения зависимости признака pm от генотипической среды; из внешних же условий основным фактором, влияющим на проявление pm, оказалась температура (отчасти — состав корма). Результаты показали, что понижение температуры (12—15° С) понижает и проявление pm, иногда даже до 0%. Повышение температуры (нормальная для лаборатории t = 25—26°) повышает проявление, доводя его иногда до 100% (только по крыльям). Промптов отметил адаптивный смысл этих особенностей: «Сильное падение процента проявления pm (до 0%) в пониженной температуре, в которой обычно и развиваются в природе мухи (в гниющих на земле фруктах и овощах), оказывается весьма благоприятным свойством для сохранения геновариации в дикой популяции» (Там же, с. 167). В ту же сторону влияет и «ухудшение» питания — чистые овощи (природный корм личинок D. funebris) также дают падение проявления pm, что благоприятствует ее сохранению и популяции.
Для определения температурно-чувствительного периода личинки содержались при t = 25° определенное число суток, после чего переносились в t = 15°, где оставались до выхода мух (контроль содержался при 25°). Оказалось, что температурно-чувствительный период для развития зачатков крыльев растянут на весь личиночный период (шесть суток) и, возможно, заходит на стадию куколки, для развития глаз он находится в пределах трех первых суток, для развития лапок — в пределах четырех первых суток.
Реальность различий чувствительных периодов для крыльев и глаз подтвердилась опытами, где взрослые личинки накануне окукления переносились в пониженную температуру. Так как в это время еще не кончился чувствительный период по крыльям (но уже прекратился по глазам), то можно ожидать выхода мух с нормальными крыльями (действие 15°), но с измененными глазами (25°) — комбинация, очень редкая в линиях, ведущихся при постоянной температуре. И действительно, по наблюдениям Промптова, почти в каждой опытной пробирке в переменной температуре было несколько особей с нормальными крыльями и «грубыми» глазами.
Температурные опыты проводили и другие члены кольцовско-четвери-ковской школы. Так, Е. А. Тимофеева-Ресовская предприняла подобное исследование «политопной (плейотропной) геновариации» polyphaen, возникшей в ее опытах с освещением D. funebris рентгеновскими лучами (Н. Timofeeff-Ressovsky, 1930, 1931). Этот доминантный аутосомный, рецессивно-летальный ген вызывает: 1) «грубые» глаза; 2) ненормальное брюшко; 3) расставленные крылья; 4) ненормальное жилкование и 5) ненормальный хетотаксис. Для выявления чувствительного периода были поставлены две серии опытов. В первой развитие начиналось при 25°; культуры пересаживались в 15° после трех, шести, девяти или 12 дней; во второй — культуры шли 24, 18, 12 или шесть дней при 15е и затем при 25° Чувствительный период для хенотаксиса падает на 6—9-й день личиночной стадии при 25°, все же остальные признаки развивали ту или иную степень выражения в классах пропорционально времени инкубации при 15°
108
Н. В\Тимофеев-Ресовский, изучая влияние генотипической среды и внешних условий на фенотипическую реализацию гена vti (вызывающего прерван-ность илиЧкчезновение поперечных жилок крыла у D. funebris), выяснил следующеедВ некоторых культурах проявление vti'независимо (или почти независимо) ют температуры, а в других — повышение температуры оказывает более или\|иенее заметное, смотря по культуре, влияние на пенетрантность и экспрессивность vti. Специфичность и эвномия независимы от температуры. Подробный экспериментальный анализ высокочувствительной к температуре культуры А показал, что в онтогенезе мух есть два температурно-чувствительных периода. Первый приходится на первую половину личиночной стадии; здесь с повышением температуры сильно понижается пенетрантность vti (и лишь незначительно экспрессивность). Второй падает на стадию куколки; в это время с повышением температуры довольно сильно повышается экспрессивность vti (и лишь незначительно связанная с этим пенетрантность) (Тимофеев-Ресовский, 1928, 1930; Timofeeff-Ressovsky, 1934а).
Из своих опытов с vti и обширных литературных данных, опираясь на свою общую схему осуществления признака, Тимофеев заключает, во-первых, что «ген vti надо признать «ускорителем» или «осуществителем» поперечных жилок. Его нормальный доминантный аллеломорф образует поперечные жилки при всяких (в природе и опыте возможных) внешних условиях; рецессивный же ген vti обладает в этом направлении значительно более слабой морфогенной способностью, которая поэтому легко подвергается воздействиям со стороны других генов и внешних условий» (Тимофеев-Ресовский, 1928, с. 207). Далее, на формообразующем пути гена vti есть два главных этапа. «Первый чувствительный период» относится к цепи реакций, которые ведут к образованию некоего «формативного вещества», необходимого для формирования поперечных жилок; при высокой температуре чаще достигается образование той порции этого «формативного вещества», которая нужна для полного развития поперечных жилок; поэтому пенетрантность гена vti ниже, если культура во время «первого чувствительного периода» развивалась в высокой температуре. «Второй чувствительный период» относится к тому времени на стадии куколки, когда происходит дефинитивное образование органов; экспрессивность vti при низкой температуре понижается, так как при удлинении стадии куколки другие факторы («формативные вещества»), имея большее время в распоряжении, успевают сформировать часть поперечных жилок; в крайнем варианте жилки образуются целиком, благодаря чему и пенетрантность несколько понижается. «Подобный же механизм лежит, очевидно, в основе и пигменто-образования в температурных опытах с некоторыми гусеницами»,— писал Тимофеев-Ресовский (1928, с. 209), ссылаясь на работу Н. К. Беляева (1926).
УРОКИ ФЕНОГЕНЕТИКИ
В ходе феногенетических исследований московской школы эволюционной генетики были получены следующие результаты: выявлен фенотипический параллелизм «направленной» изменчивости и выделены основные категории фонда популяционной изменчивости; оформлены представления о природной изменчивости как результате некоординированности процессов онтогенеза и 0 гомеозисах как развилках в онтогенезе; определены основные параметры
109
фенотипического осуществления гена и основные уровни биологической организации, где осуществляются влияния на формообразовательный процесс, зависящий от определенного гена, что привело к общему представлению о действии генов; сформулированы концепция наследственной конституции, принцип автономной изменчивости признаков, концепция полей действия генов. В дальнейшем представления, выработанные ъ ходе феногенетических исследований, сыграли существенную роль в изучении феногенетики популяций (гл. 11) и коррелированной изменчивости (гл. 12).
A posteriori здесь можно различать два главных направления: одно — выяснение феноменологии фенотипического осуществления гена, другое — конкретизация проблемы целостности генотипа (и общей проблемы целостности многоуровневой биологической системы), поставленной Четвериковым в форме концепции генотипической среды. Ее значение (и для современников Четверикова, и для нас) заключается не в конкретном естественнонаучном содержании, а в постановке вопроса; и если в последние два десятилетия мы все чаще обращаемся к этой концепции, то это обязано положенному в ее основу системному подходу, плодотворность которого осознается все более широким кругом нынешних эволюционистов.
Здесь мы коснулись вопроса, который всегда волнует историка науки при рассмотрении работ той или иной группы исследователей: в чем здесь конкретно реализуется их система познавательных ориентиров? Редукционистской исследовательской программе группы Моргана, например, где внимание было сосредоточено на проблемах передачи генов, а вопросы фенотипического осуществления, выражаясь математически, выносились за скобки, контрастировала системная ориентация четвериковской группы, открывшая возможность постановки проблем такого рода и позволившая получить результаты, ставшие впоследствии важной основой теории стабилизирующего отбора Шмальгаузена. Целью всех этих работ был анализ фенотипической изменчивости в природе; в частности, выяснение причин различий изменчивости в лабораторных и природных условиях (различий настолько ошеломляющих, что мутации дрозофил поначалу считались лабораторными артефактами). Здесь выступает характерная для четвериковцев, несмотря на использование лабораторных методов, натуралистическая ориентация, а также ориентация на эволюционно-значимые результаты. Действительно, в отборе участвует фенотип; поэтому дело было не в том, чтобы просто отвергнуть тезис о взаимно-однозначном соответствии гена и признака, а в том, чтобы установить закономерности фенотипического осуществления гена. Высокая математическая культура, свойственная чет-вериковцам, оказала влияние на их общебиологические взгляды, хотя и не привела их к созданию математической генетики. Представление об абсолютной детерминированности процессов онтогенеза уступило в их работах место представлению о стохастической детерминированности; с этим был связан и взгляд на стабильность онтогенеза как результат эволюции.
Эти черты, свойственные четвериковцам, позволяют понять, почему феногенетические работы их прошли без последствий для представителей англо-американской традиции генетики популяций, основанной на редукционизме, лабораторном эксперименте и математических моделях (хотя те же исследователи сравнительно скоро восприняли введенное четвериковцами изучение природных популяций); почему близкие по направленности работы делали на Западе по преимуществу негенетики и эта линия работ существовала там отдельно от генетики популяций; и почему именно из феногене
110
тИческих\исследований московской школы Шмальгаузен (1937—1946) мог почерпнуА важный материал для своей теории.
Характеризуя состояние эволюционной биологии на Западе, Кари Веп-салайнен обметил, что «попытки советских биологов связать воедино онтогению иХэволюцию все еще мало известны... Если уместно предположение, то \ скажу, что эволюционные исследования могут получить колоссальный Аимул именно на пути интеграции процессов индивидуального развития с эволюционными процессами, которые разыгрываются на популяционном уровне и в экологическом контексте» (Vepsalainen, 1982, р. 152). \
Глава 8
ГЕНЕТИКО-АВТОМАТИЧЕСКИЕ ПРОЦЕССЫ
И надежду, и страх распределяет слепой случай.
СЕНЕКА
Определенность и широкое понимание центральной проблемы эволюционной генетики позволяли развивать в ее рамках частные теоретические концепции. Наиболее влиятельной из них в советской эволюционной биологии 1930-х гг. была концепция генетико-автоматических процессов (ГАП’ов), которая подчеркивала эволюционную роль случайных процессов. Происхождения концепции ГАП’ов касались лишь Балкашина с соавторами (1975) и Адамс (Adams, 1970); тем временем происхождение близких концепций Р. Фишера и С. Райта, легших в основу математической генетики, достаточно широко известно.
Роль случайности в эволюции интересовала многих авторов. Аренд и Анна Хагедорны (A. L., А. С. Hagedoorn, 1921) утверждали, что случайное сохранение гена («эффект Хагедорнов» — Fisher, 1922) играет большую роль в судьбе популяции, чем предпочтительное сохранение гена в результате отбора. Дж. Т. Гулик описал вариации раковин нескольких видов слизняков Achatinellae, живущих в разобщенных долинах о-ва Оаху, и пришел к выводу, что дивергенцию групп нельзя объяснить отбором, так как она непропорциональна различиям в окружающей среде и затрагивает нейтральные («nonutilitarian») признаки. Причину дивергенции он усматривал в инбридинге, обязанном отсутствию свободы скрещивания (Gulick, 1872, 1873, 1888, 1905).
Из русских биологов к выяснению роли случайности в эволюции близко подошли: Четвериков, с его представлениями о волнах жизни (1905), о роли изоляции и о неадаптивных процессах в эволюции (1926; см. гл. 1), и особен-Но А. С. Серебровский, который в работах по геногеографии и генофонду сельскохозяйственных животных (1927—1930; переизд. 1976), говоря об «автоматических» процессах изменения концентраций генов, привлек внимание к отдельным примерам эффекта родоначальника и эффекта бутылочного гоРлы1ика, которые можно рассматривать как важные частные случаи «волн жизни». Он же в 1925 г. ввел в обиход термин «генофонд».
Однако эти представления лишь предвосхищали теории дрейфа генов и 1 АП’ов. Концепции же эволюционной роли случайных процессов разработали Рональд Фишер (Fisher, 1922, 1930), Дубинин и Ромашов (1932; Ро-
111
машов, 1931; Дубинин, 1931) и особенно Райт (Wright, 1921, Ld31, 1932 и др.). Последний, исследуя системы скрещивания животных/построил концепцию дрейфа генов; при этом он указывал Гулика в качестве своего предшественника, ссылаясь на его работу 1905 г. Концепция «^рехфазного подвижного равновесия» Райта (Wright, 1931) включает дрейф генов, отбор внутри локальных популяций (при инбридинге) и отбор между ними (аутбридинг). /
Фишер приступил к изучению роли случайных флуктуаций в процессах эволюции с критики мнения Хагедорнов (Fisher, 1922)./Фишер выводил эволюцию из селективных свойств отдельных генов; изу^ние естественного отбора он сравнивал с аналитическим подходом к теорий газов, «где можно делать различные допущения о случайных обстоятельствах, ...и все же выводить общие законы поведения газов» (там же, с. 321—322). В число допущений о «случайных обстоятельствах», которыми можно пренебречь в эволюционной теории, Фишер включал эффекты генных взаимодействий и дрейф генов. Различие исходных позиций Райта и Фишера повело к тому, что Фишер выступил с критикой концепции Райта как несовместимой с представлениями об эволюции посредством естественного отбора (например, Fisher, Ford, 1950; ответ на критику — Wright, 1951). Интересно, что с тех же, что и Фишер, позиций А. Р. Уоллес (Wallace 1889; см. перевод: Уоллэс, 1898, с. 213—224) критиковал точку зрения Гулика, предшественника Райта.
КОНЦЕПЦИЯ Д. Д. РОМАШОВА
Научному творчеству Д. Д. Ромашова свойственны две осооенности. Одна — склонность к математике. Так, его первая работа была посвящена применению бинома Ньютона для обобщения законов Менделя (Ромашов, 1925). На эти интересы Ромашова несомненно повлияла прошедшая через его жизнь, начиная с детских лет, дружба с А. Н. Колмогоровым 1 Стимулирующую атмосферу создавало участие в курсе биометрии С. С. Четверикова; обсуждение на заседаниях «Соора» вероятностной картины мира; консультации статистика Н. С. Четверикова (младшего брата Сергея Сергеевича и ученика А. А. Чупрова; см.: Карпенко, 1974).
Еще более яркую особенность Ромашова составили его натуралистические интересы. «Очень рано он начал экскурсировать и собирать коллекции жуков. Его первым учителем по биологии был Владимир Гордеевич Савич — гидробиолог, ученик Н. К. Кольцова. Уже в 1912—1913 гг. [т. е. в 13—14 лет] у Дмитрия Дмитриевича значительные коллекции жуков Подмосковья, и он
1 В 1930-е г. Ромашов привлек Колмогорова к участию в работе Эволюционной бригады ИЭБ (а также и ряд других математиков — В. И. Гливенко, Н. В. Смирнова, М. В. Игнатьева и А. А. Ляпунова) — см.: Д. Д. Ромашов. «Краткий отчет о работе Эволюционной лаборатории (Эволюционной бригады) при Институте экспериментальной биологии с 1934 по 1940 гг.».
Упомяну также «Отзыв» А. Н. Колмогорова от 29 июля 1942 г., в котором он присоединяется «к мнению ряда авторитетных биологов, считающих Д. Д. Ромашова достойным докторской степени без защиты диссертации» (присуждена в 1944 г.) (цит. по: Беляева, 1972). Наконец, укажу, что опубликованные в Докладах Академии Наук СССР три статьи Д. Д. Ромашова и В. Н. Беляевой (1946) и статья Д. Д. Ромашова и Е. Д. Ильиной (1942) представлены Колмогоровым.
112
ведет переписку с областным Терским музеем об обмене коллекциями жуков Среднеи\Р°ссии, им собранных, на жуков Кавказа. В его записях того времени есть! отметки о сборах жуков в Калужской, Тверской, Московской, Лифляндскрй губерниях, на южном берегу Крыма, в окрестностях Ставрополя, Самары\ Васильсурска, Саратова, Камышина, Астрахани. Энтомологическое чутье\ интуицию и страстную увлеченность при сборах насекомых он сохранил на всю жизнь» (Беляева, 1972, с. 2—3) 2. Поступая в 1918 г. в Московский университет (на биологическое отделение физико-математического ф-та), Ромашов уже имел весьма широкое биологическое образование; он был знаком с работами не только Дарвина и классических дарвинистов и антидарвинистов, но и Вейсмана, Менделя и др. авторов. В университете его учителями были А. Н. Северцов, Н. К. Кольцов, С. С. Четвериков; в 1918—1919 гг. он прошел курс теоретической и прикладной энтомологии у проф. Н. М. Кулагина, а лето 1918 г. провел работая «специалистом»3 на Звенигородской биостанции. Осенью 1920 г. по инициативе Е. С. Смирнова (впоследствии — известный систематик насекомых, профессор МГУ) среди группы студентов, работавших в Зоомузее Московского университета, был организован Студенческий кружок «в целях научного общения молодых зоологов в области общей биологии»4. Через два года он получил название «Кружок имени Ламарка», с девизом: «Разными путями к единой цели». В кружке реферировались и обсуждались важнейшие работы из разных областей общей биологии, шли страстные споры между дарвинистами (в их числе — Ромашов) и ламаркистами (наиболее яркий среди них — Смирнов, а также Ю. М. Вермель, Б. С. Кузин5). (Подробнее о Кружке им. Ламарка см. в гл. 10). В октябре 1921 г. будучи еще студентом Ромашов стал одним из первых сотрудников Четвериковской лаборатории в ИЭБ6 * 8; несколько позже при этой лаборатории возник реферативный кружок Соор (см. гл. 1), в работе которого он активно участвовал с самого начала. В это же время он принимал участие в первом генетическом исследовании популяций дрозофил (см. гл. 2). В сентябре 1926 г. Кольцов поручил Ромашову заведование отделом теоретической генетики Центральной генетической станции (ЦГС), однако вскоре Ромашов почти на два года отошел от работы по болезни. Вновь приступив к заведованию отделом, он также включился в работу Соор’а, а в 1929—1931 гг. провел вместе с Е. И. Балкашиной генетический анализ диких популяций D. funebris из Москвы, Петровско-Разумовского, Киева и Ташкента. В 1930 г. началось слияние ЦГС с Институтом животноводства ВАСХНИЛ и создание Всесоюзного института животноводства; в этой связи ЦГС была переведена в Гатчину (тогда г. Кра-сногвардейск, под Ленинградом). Весной 1931 г., как только выяснилось, что ВИЖ переводится под Москву, Ромашов возвратился из Гатчины.
2 Материалы Комиссии. См. также: Балкашина и др., 1975, с. 76.
3 Справка (Цит. по: Беляева, 1972).
5 «Два года деятельности Кружка имени Ламарка», 1922.— Материалы Комиссии. Впоследствии авторы посвяшенной П. Каммереру книги «Очерки по террии эволю-
ции» (1924). В библиотеке Ромашова — экземпляр с надписью: «Дорогому Дмитрию Дмитриевичу Ромашову от любящих сочленов по Кружку им. Ламарка. 2.IX. 1924.
6 Е. Смирнов, Ю. Вермель, Б. Кузин».
Практикант, затем научный сотрудник ИЭБ Наркомздрава; с августа 1925 — научный сотрудник Генетического отдела МО КЕПС при Академии наук; в сентябре 1926 г. перешел на должность старшего ассистента ЦГС.— Из автобиографии Д. Д. Ромашова (цит. по: Беляева, 1972).
8 В. В. Бабков 1
К этому времени он уже подготовил к печати теоретическую статью «Об условиях «равновесия» в популяции», которая была опубликование «Журнале экспериментальной биологии», т. VIII (1931), вып. 4. /
Летом 1931 г. работает над статьей, посвященной тем ж/ вопросам соотношения факторов эволюции популяций, Н. П. Дубинин, ученик А. С. Серебровского. Его статья «Генетико-автоматические процессы и их значение для механизма органической эволюции», которая дала название концепции, была опубликована в следующем (5—6) выпуске того же журнала. После окончания в 1928 г. 1-го МГУ Дубинин работал в лаборатории генетики Зоотехнического института, в 1929—1931 — в лаборатории генетики Тимирина (Биологический институт им. К. А. Тимирязева Коммунистической академии), в обеих лабораториях ассистентом Серебровского; затем недолгое время в Институте свиноводства, выделившемся к тому времени из Зоотехнического. Весной 1932 г. Н. П. Дубинин был приглашен Кольцовым для заведования отделом генетики, заново созданным в ИЭБ. Туда же после сокращения отдела теоретической генетики ВИЖ возвратились Д. Д. Ромашов и Е. И. Балкашина; благодаря им А. Н. Промптову в ИЭБ поддерживались традиции и тематика Лаборатории С. С. Четверикова. Вскоре началась совместная работа Дубинина и Ромашова над статьей «Генетическое строение вида и его эволюция. I. Генетико-автоматические процессы и проблема экогенотипов» (1932). Продолжая эволюционную линию лаборатории Четверикова, Ромашов организовал при генетическом отделе ИЭБ Эволюционный семинар и открыл его 23 октября 1932 г. докладом «Эволюционные взгляды Морица Вагнера». В октябре 1934 г. он организовал и возглавил Эволюционную бригаду (с января 1937 г.— Лаборатория, которая существовала по 15 июля 1940 г.); при ней работал Дарвиновский семинар — преемник Эволюционного. Последнее, 104-е заседание Дарвиновского семинара состоялось 10 апреля 1940 г.7
В первой формулировке эволюционной концепции, учитывающей роль случайных процессов, Ромашов, как и Четвериков, выбрал отправным моментом равновесие Харди, характеризующее популяцию в состоянии эволюционной инерции. Но он сосредоточил внимание не на возникновении мутаций, а на случайных процессах. Он отметил, что детерминистический подход упрощает картину явления: даже при равномерном размножении множество мутаций должно вскоре после своего возникновения выбывать из популяции — в силу менделевской стохастики, т. е., когда оба потомка от скрещивания Аа X АА случайно окажутся гомозиготными по А. (Здесь он указал на близкие мысли Серебровского, 1930.) Однако генетическое строение популяции за счет таких исходов скрещиваний существенно измениться не должно.
Ромашов остановился на картине, которая рисовалась Четверикову: при единичных мутациях вид внешне сохраняет постоянство, и нахождение измененных экземпляров непосредственно в природе маловероятно. Однако уже в первом популяционном исследовании 1925—1926 гг. такие аберрантные экземпляры были найдены. Напомнив, что естественный отбор крайне мало эффективен при ничтожно малой концентрации гена и при низкой интенсивности отбора (это хорошо иллюстрирует таблица Нортона), Рома
7 «Эволюционный семинар при генетическом отделе Института экспериментальной биологии. 1932—1933—1934—1935» (протоколы); «Протоколы заседаний Эволюционной Бригады», тетр. 1 и 2. (Материалы Комиссии; см. также: Балкашина и др., 1975).
114
шов указал на разрыв между возникновением мутаций и отбором в четвери-ковской Схеме: «...напрашивается вывод, что о действии естественного отбора (как положительного, так и отрицательного) можно говорить только в том случае, когда в силу каких-либо особых, не зависящих от отбора условий концентрация данного гена в населении увеличится и достигнет значительной величины. Только в этом случае в природе начнет появляться большое число измененных особей, на которые отбор и сможет оказать свое действие» (Ромашов, 1931, с. 443). Этот разрыв следовало устранить.
Каковы же эти «условия, не зависящие от отбора?» Одним из них является повторное мутирование данного гена в одном и том же направлении. (Здесь Ромашов ссылался наряду с Четвериковым, 1926, и на Серебровского, 1927). Кроме того, в популяциях существуют также «чисто автоматические процессы», «которые ведут к резким изменениям в концентрации генов и которые являются одним из самых существенных факторов, влияющих на генетическое строение вида» (Ромашов, 1931, с. 444). Этот не совсем удачный термин («автоматический» означает здесь всего лишь «не связанный с отбором») идет от исследований Серебровского по геногеографии и генофонду сельскохозяйственных животных. («Под геногеографическими факторами мы разумеем сейчас некоторые автоматические процессы, которые начинают изменять состав популяции, коль скоро она уменьшается до такого размера, в котором формула Гарди для сохранения популяции в равновесии перестает действовать и вступает в силу другой закон, в силу которого в этих условиях на почве инбридинга (кровного скрещивания) начинается систематическое повышение гомозиготности, приводящее в конце концов к образованию гомозиготной новой линии».— Серебровский, 1930; цит. по: Серебровский, 1976, с. 190, 254). Ромашов подчеркивал, что Се-ребровскому принадлежат «некоторые общие указания» на существование этих процессов, но что последний дал не вполне верную оценку механизма их действия, отводя большую роль инбридингу, и не рассматривал их сравнительную роль среди факторов, обусловливающих равновесие в популяциях.
Впоследствии Серебровский сам отметил неудачность этого термина в докладе 1934 г.: «Процессы, изменяющие популяции независимо от естественного и искусственного отборов и до последнего времени мало привлекавшие внимание исследователей, мы назвали первоначально «автоматическими» (Серебровский, 1927, 1929). Позднее Ромашов и Дубинин стали называть их генетико-автоматическими. Однако мы думаем, что термин «автоматический» здесь использован неправильно и должен быть заменен недвусмысленным термином «стохастический»...8 (Серебровский, 1976, с. 383—384). В конце 1948 г. также и Четвериков обратился к этой теме и начал писать работу «О некоторых моментах эволюционного процесса с точки зрения современной генетики. Часть вторая: генетико-стохастические процессы в популяции» (неоконченная рукопись). Во вступлении он писал: «...До сих пор в русской литературе эти процессы называются «генетико-автоматическими» процессами. Однако слово «автоматический» здесь малоподходящее, так как оно предполагает наличие какого-то регулирующего аппарата, которого в действительности нет. Мы имеем здесь дело с явлениями, подчиняющимися законам теории вероятностей, т. е. с явлениями чисто стохастическими...» (Нит. по: Астауров, 1974, с. 66).
^прочем, у древних греков «Автоматия» («Произвольность») была богиней случая и счастья.
Неприспособительные изменения частот генов, утверждает Ромашов, обязаны прежде всего неравномерному размножению особей популяции, которое особенно ясно видно в случае сезонных колебаний численности популяций — сильного увеличения в летние и осенние месяцы и резкого падения зимой. В ходе ранних популяционных исследований (о них шла речь в гл. 2) Ромашову приходилось наблюдать такие колебания численностей у целого ряда обычных в окрестностях Москвы видов дрозофил. Зимой эти виды вымирают массами, и весной они встречаются лишь случайными единичными экземплярами; в течение лета их численность увеличивается. К осени популяция по своей численности будет приближаться к предыдущему осеннему поколению, однако ее генетическое строение значительно изменится. Немногие вновь возникшие мутации будут представлены одной гетерозиготной особью; большинство старых мутаций утеряется, а численность большинства сохранившихся — в той или иной степени увеличится. «Причины этого чисто автоматического увеличения лежат в том, что многочисленные осенние популяции в конечном счете развиваются только от небольшого числа особей предыдущего осеннего поколения» (Ромашов, 1931, с. 445). «Особенно интересным будет тот случай, когда в период наибольшей депрессии в населении вида возникает какая-нибудь очень редкая новая мутация и когда эта мутация после размножения вида окажется в населении одной из самых обычных» (с. 446). Такое явление в настоящее время часто называют «эффектом горлышка бутылки» («bottle-neck effect»).
Из своего опыта натуралиста Ромашов приводит ряд примеров, когда «действие автоматических процессов» в природе может особенно сильно изменять генетическое строение популяций. Таковы периодические депрессии, известные у многих обычных видов: Ромашов называет, среди прочих, непарного шелкопряда и монашенку, принадлежащих к опасным вредителям леса; все эти примеры чрезвычайно схожи с теми, что приводил Четвериков (1905) в статье «Волны жизни». Ромашов заключает, что «изучая из года в год одну и ту же популяцию, мы будем в некоторых случаях наблюдать неожиданные и резкие изменения в ее генетическом строении» (1931, с. 446), и эти изменения особенно велики в годы резких колебаний численности популяции.
В современных определениях дрейфа генов отмечается, что он проявляется либо в различиях между последовательными поколениями одной популяции, наблюдаемой во времени, либо же в различиях между одновременно существующими родственными популяциями, наблюдаемыми в пространстве. Примеры различий второго типа доставляют, по Ромашову, виды, для которых характерен колониальный образ жизни. Так, наблюдения Балкашиной и Ромашова показали, что отдельные колонии D. funebris значительно изолированы друг от друга, «при этом родоначальником новой колонии обычно является небольшое число, иногда только одна пара особей» (1931, с. 446), собственно — одна оплодотворенная самка. «Очевидно, что при описанных условиях различные колонии этого вида (как и других колониальных видов) должны значительно отличаться друг от друга по своему генетическому строению в связи с тем, что особи-родоначальники новых колоний будут в различных случаях вносить в эти колонии совершенно различные наследственные задатки»,— так Ромашов (1931, с. 446) описывал то, что мы называем теперь «эффектом (или — принципом) родоначальника» (а иногда — явно неудачный обратный перевод «founder principle» — «принципом основателя».)
116
В работах по генетике животных А. С. Серебровский также описывал случаи эффекта родоначальника — распространение генов, восходящих, возможно, к «Единственному быку-родоначальнику» (1928; см.: Серебровский, 1976, с. 180—181) и эффекта горлышка бутылки — «тот ген, который возникает в период усиленного роста популяции, окажется размноженным» (1930; см.: Серебровский, 1976, с. 191; ср. с. 253—254).
Вообще же эволюционно-генетические мотивы, отражая умонастроения времени, явно звучат как в этой серии работ Серебровского, так и в работах Н. И. Вавилова (1926, 1927; Vavilov, 1920) о центрах генетического разнообразия и происхождения культурных растений и о законе гомологических рядов в наследственной изменчивости. Но было бы трудно утверждать, что именно Серебровский ввел эти понятия в эволюционную генетику. «Связанным с геногеографией исследованиям в области растениеводства и животноводства, однако, не хватало общебиологической теории генетических процессов в популяцйях,— писал Астауров (1965, с. 28).— В этом отношении теоретические предпосылки и экспериментальные данные школы С. С. Четверикова как нельзя более отвечали запросам времени». Конечно, трудно назвать Ромашова единственным автором этих терминов. Эта идея была «в воздухе», и другие четвериковцы упоминали для аналогичных ситуаций «родоначальника» или «самку-родоначальницу» (например, Тимофеев-Ресовский, 1925, с. 125; Балкашина, 19286, с. 102; Рокицкий, 1929а, с. 207). Наконец, Четвериков (1926) близко подошел к формулировке этих понятий. Ромашов же был автором не терминов (которые пришли позже), но понятий, в том смысле, что он впервые их четко сформулировал и ввел в круг представлений эволюционной генетики.
Яркий пример обязанных дрейфу различий второго типа доставило параллельное исследование геленджикской и берлинской популяций D. melanogaster 1926 г. (см. табл. 1, гл. 2). Ромашов подтверждает свои теоретические соображения результатами этого анализа, а также сходными результатами анализа трех изолированных колоний D. funebris (Москва, Петровско-Разумовское, Киев), который он провел вместе с Е. И. Балкашиной в 1929— 1930 гг. (В следующей статье он ссылался также на исследование популяции этого вида из Ташкента, проведенное ими же в 1931 г.— см.: Дубинин, Ромашов, 1932, с. 89.) «Оказалось, что каждая из этих колоний в высокой степени гетерогенна, что целый ряд генов достигает в пределах отдельной колонии значительной концентрации и что различные колонии отличаются между собой в том отношении, что отдельные мутационные изменения, характерные Для данной колонии, в пределах других колоний не наблюдаются» (Ромашов, 1931, с. 449). Повторное мутирование не может объяснить эти факты, считал Ромашов, так как в этом случае пришлось бы допустить, что в различных популяциях мутационный процесс протекает по-разному: например, в берлинской популяции D. melanogaster особенно часто изменялись одни гены, а в геленджикской — другие. Однако такие предположения «противоречат многочисленным наблюдениям над мутационным процессом у Dr. melanogaster, который в самых различных местах, где изучался этот вид, протекает с большим постоянством в отношении сравнительной частоты возникновения различных мутаций» (с. 449). Ромашов отвергает также объяснение посред-СТв°м отбора, так как подавляющее большинство обнаруженных изменений пРеДставляет собой «вредные» уклонения от нормы; в этом отношении особен-интересен «факт троекратного нахождения в берлинской популяции
• Melanogaster сцепленного с полом летального фактора lethal-2, увели-
117
ценная концентрация которого может быть отнесена только за Счет действия автоматических процессов» (с. 450). Ромашов заключает, Ито каждое обнаруженное изменение возникло в данной местности, вероятно, только однажды, и что причины увеличенной концентрации некоторых генов следует искать в «действии автоматических процессов». Значение этих процессов сводится, по Ромашову, к постоянному и притом крайне «неправильному» нарушению существующих в природе равновесий. Поскольку характер этих «нарушений» прежде всего определяется размером популяции и степенью ее изоляции, а также амплитудой колебаний ее численности (наряду с концентрацией изучаемого гена), то очевидно, что «характер и интенсивность действия этих поцессов должны быть теснейшим образом связаны с биологией данного вида и очевидно будут весьма различными у видов с различным образом жизни» (с. 450). Отсюда следует, что для понимания генетического строения вида первостепенное значение получает «детальное и всестороннее ознакомление с биологией данной формы» (Там же).
Ромашов в статье 1931 г. не упускал из виду двух других основных факторов, изменяющих концентрацию генов в природных популяциях,— мутационного процесса и естественного отбора, которому он отводит «первенствующую роль в процессах исторического изменения структуры популяции» (с. 450). Более того, одним из побуждений к написанию этой теоретической статьи была необходимость найти связующее звено между мутациями и отбором. Обращаясь к действию отбора, Ромашов ссылался на иллюстративный материал из таблицы Нортона, приведенной в статье Четверикова (1926). Так, при интенсивности отбора в 10% нужно около 18 200 поколений, чтобы новое полезное (рецессивное) изменение, имеющее концентрацию 0,0005, почти нацело вытеснило менее полезный доминантный аллеломорф. Огромное большинство этого времени отбора приходится на тот период, когда концентрации отбираемого гена невелики (для увеличения концентрации гена от 0,005 до 0,5 в нашем примере потребуется более чем 18 100 поколений и около 80 поколений—для дальнейшего изменения от 0,5 до 0,9995). Отсюда Ромашов заключает, что при низких частотах генов отбор неэффективен; он полагает, например, что в рассмотренном Четвериковым (1926) случае с бабочкой Biston (Amphidasis) betularia, когда в природе удалось наблюдать действие положительного отбора на доминантное изменение, в начальных стадиях распространения меланистической формы главную роль сыграл не отбор, а случайные увеличения ее численности в силу неравномерного размножения отдельных особей данного вида. Наконец, он приходит к «общему выводу о существовании как бы двух зон концентрации генов: первая — зона малых концентраций, где особенно большое значение для генетического строения популяции должны иметь автоматические процессы, и вторая зона больших концентраций, где сказывается по преимуществу влияние естественного отбора» (Ромашов, 1931, с. 451), определяя отношение популяционных факторов в терминах «двух зон» концентрации генов.
Обсуждая условия, при которых дрейф генов будет ведущим фактором изменения частот аллелей в популяции, С. Райт выразил соотношения факторов генной динамики (эффективная величина популяции — N, селективное значение аллеля — s, давление мутаций — и, темпы миграции — т) в общей форме (Wright, 1931). Он показал для отдельного диаллельного локуса, что теоретическое распределение числа маленьких изолированных популяций по частоте аллеля описывается U-образной кривой (рис. 12, а). При
118
рис. /2. \ Распределение числа популяций \ло частоте аллеля отсутствие Мутаций и миграций
(по: Wright, 1031)
а — без отбора (/ — N мало, 2 _ jV велико) б — взаимодействие отбора и дрейфа (1 — s = 0; 2__5 = '/Л;«?-^ = 74 n-4-s = 4,n
взаимодействии дрейфа и отбора кривая приобретает асимметрию (рис. 12, б). Ромашовские «зоны концентрации» особенно хорошо видны на рис. 12, а: ДРейф эффективен при условии «крайних» частот (р около 0 или 1).
Рассматривая естественный отбор, Ромашов различает два случая: когда он действует на «полезное» уклонение и когда на «вредное». В более частом случае возникновений неблагоприятных изменений между интенсивностью отбора и интенсивностью мутационного процесса в конечном счете установится равновесие, которое и определит некоторую среднюю концентрацию данного гена в населении вида. (Заметим, что именно на основе рассмотрения такого равновесия впоследствии В. П. Эфроимсон (19326) и Дж. Б. С. Холдейн (Haldane, 1935) независимо друг от друга рассчитали темпы мутационного процесса у человека). В силу действия случайных процессов это равновесие будет неустойчивым. В случае же полезного уклонения мутационный процесс и отбор действуют в одном направлении, распространяя это уклонение в населении вида. Однако в начальных стадиях отбора, когда частота гена невелика, случайные ее колебания могут повести к тому, что «полезная мутация», уже начавшая распространяться в населении, будет совершенно уничтожена.
Завершая теоретическую статью 1931 г., на которую он смотрит только как «на первую попытку разобраться в поставленных вопросах и наметить в самых общих чертах те направления, по которым должна идти их дальнейшая разработка» (Ромашов, 1931, с. 453), Ромашов подчеркнул, что состояние «относительного равновесия», в котором находится популяция, носит непостоянный и крайне неустойчивый характер (Там же). Ромашов считал необходимым пояснить в обширном примечании (47 строк петита — си. 1, с. 452—453), что слово «равновесие» по отношению к популяционным процессам употребляется в «особом», чисто условном смысле»— то, что мы обозначаем как «динамическое равновесие». Он подчеркивал, в частности, что расчеты, основанные на абсолютизации формулы Харди, «совершенно неудовлетворительны, особенно в тех случаях, когда эти расчеты касаются Целых исторических периодов (Серебровский)» (Там же).
Еще больший интерес для нас представляет факт, что Ромашов отмечает в этом примечании марковские свойства 9 рассматриваемых популяционных
Вероятностный «процесс называется марковским, если исчерпывающие данные для предсказания могут быть получены, отправляясь от списания мгновенного состояния Процесса, т. е. дополнительные сведения о предшествующих состояниях ничего не Добавляют для возможного предсказания» (Кулагина, Ляпунов, 1966, с. 157).
См. там же соображения авторов о возможности двух способов описания одного и того же процесса и о предпочтительности марковского описания процессов полюции.— Прим. автора.
119
процессов. Это обстоятельство впоследствии дало основание А. Н. Колмогорову (1935) применить к популяцщонной динамике разработанный для марковских процессов «прямой метод Колмогорова». Представление Ромашова о марковости популяционной динамики, несомненно, контрастировало взглядам Серебровского («И та печать своеобразия, которую мы сейчас замечаем и на концентрациях генов кур в отдельных аулах, есть, таким образом, печать веков и тысячелетий».— Серебровский, 1928, цит. по: Серебровский, 1976, с. 176), чему в значительной мере и обязана критика Ромашовым точки зрения Серебровского.
По Ромашову, популяцию можно рассматривать как сложную систему непостоянных «равновесий», представляющих собой концентрации отдельных генов. Складывающееся из этих «равновесий» генетическое строение популяции претерпевает «разнообразные изменения, определяемые ходом мутационного процесса, биологическими условиями, условиями существования вида в данный момент и действием естественного отбора» (Ромашов, 1931, с. 453); «отыскание тех законов, которые управляют изменением структуры популяции в исторические периоды жизни вида, должно составить одну из основных страниц эволюционной теории» (Там же).
Итак, в статье 1931 г. Ромашов сформулировал представление о неадаптивных популяционных процессах, причину которых он видел в обязанных менделевской стохастике «ошибках выборки», чрезвычайно усиливаемых «волнами жизни». Его представления объяснили генетические различия между последующими поколениями одной популяции и между одновременно существующими изолированными родственными популяциями. Они предсказали существование в природных популяциях аберраций (видимой изменчивости), неадаптивного полиморфизма и деталей, а главное — дали импульс к поиску изменчивости такого рода. Представления Ромашова обозначили относительную роль факторов, определяющих генетическое строение популяции, и указали на тесную связь последней с биологией и экологией вида. Как видно, эта работа в двух направлениях развивала традицию Четверикова (проблема — природа и поддержание изменчивости в популяциях): был предложен определенный механизм поддержания изменчивости в дополнение к мутациям и отбору и высказаны соображения об определенном их соотношении, а также указаны категории природной изменчивости, иные, чем «хорошие» рецессивные гены в гетерозиготном состоянии.
РАЗВИТИЕ КОНЦЕПЦИИ ГАП’ов
Обсуждение Н. П. Дубининым роли случайности в эволюции носит печать влияния А. С. Серебровского, его учителя. Александр Сергеевич Серебровский (1892—1948) был первым из учеников и сотрудников Н. К. Кольцова, которому тот поручил заняться генетикой. (Двух более молодых своих учеников, Д. Д. Ромашова и В. В. Сахарова, Кольцов привлек к генетической работе чуть позже, около 1920 г.— Babkoff, 1975) Обзор Серебровского «Современное состояние теории мутаций» был напечатан в «Природе» в 1915 г. (с. 1239—1254). В первую мировую войну Серебровский был мобилизован и в окопах написал статью о применении математических методов в генетике. Частной генетикой кур Серебровский стал заниматься с 1919 г., когда рядом с его родными местами (в с. Слободка Тульской губернии) по инициативе Кольцова была организована на средства Комиссии по изучению естественных производительных сил России опытная птицеводческая 120
станция ИЭБ, которой Серебровский руководил. Е. Т. Васина-Попова и 3 М. Коган нашли одну из первых генетических статей А. С., относящихся к этому времени: «Изучение наследственности сельскохозяйственных животных», напечатанную в 1919 г. органом Тульского Губземотдела «Трудовое хозяйство», №№ 5 и 8. В начале 1921 г. Тульская станция вошла в состав Дниковской станции ИЭБ, где в дальнейшем проводились исследования по частной генетике не только кур («Генетика курицы»: т. 1, 1926, т. 2, 1928), но и овец («Генетика овец»: т. 1, 1928; т. 2, 1929; т. 3, 1929), крупного рогатого скота и других сельскохозяйственных животных. А. С. принял на себя заведование генетической лабораторией станции. До 1924 г., когда Центральная генетическая станция (ЦГС) перешла в ведение опытного отдела Наркомзема, организованного в ту пору, она содержалась на средства отдела животноводства Наркомзема, КЕПС и ИЭБ. В 1930 г. в Гатчине был организован*Всесоюзный институт животноводства (ВИЖ), куда ЦГС вошла в качестве Сектора генетики и селекции (Сегис) под руководством Кольцова. Через семь месяцев ВИЖ переехал под Москву, и Сегисом стал заведовать Серебровский. В 1923—1930 гг. он был также профессором Московского зоотехнического института (где с 1928 г. Дубинин был его ассистентом). С 1926 г. он с сотрудниками совершил ряд экскурсий для обследования домашних кур Центральной и Северной России, Украины, Кавказа; в экспедициях 1929 г. в Кабарду и Балкарию и 1930 г. в Башкирию принимал участие Дубинин.
Термин «ГАП’ы», который удержался в отечественной литературе, впервые появился в названии статьи Дубинина (1931): «Генетико-автоматические процессы и их значение для механизма органической эволюции». Дубинин начал свою статью с изложения эволюционной схемы Четверикова, которая «до сих пор является общепринятой среди эволюционистов-генетиков» (Дубинин, 1931, с. 468). Отметив исключительное значение, которое, по Четверикову, играет фактор изоляции в дифференциации видов («Истинным источником видообразования, истинной причиной происхождения видов является не отбор, а изоляция»,— цитирует он Четверикова, 1926 с. 42), он замечает, что это положение справедливо «при той, однако, оговорке, что под «причиной» в выражении Четверикова надо понимать то необходимое условие, которое сопровождает дифференциацию видов» (с. 467). С этим замечанием (весьма многозначительным с позиций теории ГАП’ов) охотно соглашается Ромашов 10 Затем Дубинин определяет два вопроса, подлежащие обсуждению: судьба единичной мутации и «конкретные формы закономерностей популяции». По Четверикову, каждая единичная мутация сохраняется (в единственной гетерозиготе) в населении вида. Возражая, Дубинин приводит основанный на стохастике менделевского механизма подсчет, для того чтобы показать, что единичная мутация быстро выбывает из популяции. (Ромашов отмечает: «В подсчете грубейшая ошибка», которая, однако, не повлияла на вывод.) Для объяснения исчезновения единичных мутаций Дубинин обращается к представлению о «нивелирующем скрещивании», «но уже на совершенно новой базе». Он пишет: «Нивелирующее скрещивание отрывает мутационную изменчивость от строения популяции. Всякое мутационное изменение единично и как таковое должно исчезнуть под действием Ю D (
° экземпляре Ромашова слова «необходимое условие» подчеркнуты и на полях — восклицательный знак. (Оттиск с маргиналиями Ромашова — Материалы Комиссии.)
121
нивелирующего скрещивания» (Дубинин, 1931, с. 468). И далее: «за сохранение и накопление единичных мутаций в популяции ответственны особые, как их можно бы назвать, генетико-автоматические процессы» (Там же).
Ромашов (1931) рассматривал случайное исчезновение мутации как возможный исход статистических испытаний, лежащих в основе дрейфа («автоматических» процессов) и реализующихся, в частности, в ненаправленных колебаниях рачастоты мутантного гена. Поэтому понятно, что представление Дубинина о (-направленном!) «нивелирующем скрещивании», снижающем концентрацию мутации, и о (направленном же!) «ГАП’е», поднимающем ее, должно было вызвать резкую оппозицию Ромашова. Он отмечает на той же странице: «...Нивелирующее скрещ. и г.а.п.— две стороны одного поцесса! Г.а.п. не только подъем ра (как у автора), но и снижение pj>. Это представление Дубинина было пересмотрено в совместной статье 1932 г. (Дубинин, Ромашов, 1932, с. 55).
В определении ГАП’ов Дубининым ощущается влияние Серебровского, с его склонностью связывать дрейф генов с инбридингом: «Под генетикоавтоматическими процессами мы понимает такие процессы, которые связаны с отклоняющимися (от конкретного свободного скрещивания) формами размножения отдельных особей или групп особей, определяемых биологическими особенностями данного вида» (Дубинин, 1931, с. 469). Дубинин отмечает (в согласии с Ромашовым), что примеры ГАП’ов могут доставить случаи, касающиеся неравномерного размножения и колониальных форм размножения, и приводит ряд литературных данных (включая статью Четверикова, 1905), подтверждающих распространение в природе «волн жизни». Ссылаясь на случай из собственного опыта (во время экспедиции 1930 г. по Восточной Башкирии он наблюдал целый район, насчитывающий лишь единицы птичьего поголовья из-за неурожая 1929 г. (т. е. прохождение численности популяции через «бутылочное горлышко» — ср. выше у Ромашова и Серебровского), Дубинин заключает, что достаточно уже такого вмешательства, «как структура будущей популяции кур, возникшей из небольшой группы оставшихся особей, окажется резко перестроенной» (Дубинин, 1931, с. 476). На этом основании он поддерживает требование Ромашова «отказаться от признания достоверности свидетельства геногеографии для истории» (Там же, у автора — разрядка).
Отмечая, что за счет действия ГАП’ов «вид совсем не представляет собой губки, впитывающей в себя геновариации и накопляющей их в своем генофонде по мере старения вида» (Там же, с. 471; Ромашов уточняет: «...губки, равномерно впитывающей...»), Дубинин утверждает взгляд, что вид слагается из множества локальных популяций — «экогенотипов», каждый из которых обладает собственной (изменчивой) генетической структурой: «...генное содержание вида... слагается не из равномерно вкрапленных в вид определенных концентраций тех или других генов, а из содержания глубоко отличных друг от друга экогенотипов» (Там же), и ссылается на результаты обследования четвериковцами популяций D. melanogaster (см. выше гл. 2 наст, книги). (Важно отметить, что такая формулировка предполагает постановку задачи изучения генетической структуры локальных природных популяций.) Этот подход дает Дубинину основание защищать взгляд, близкий к райтовской модели эволюции (Wright, 1931 и др.), что «эволюционная трансформация вида начинается не с медленного и более или менее равномерного ответа всего вида в целом, а с относительно быстрой
122
Рис. 13. Теория ГАПов
а — «антагонизм генетико-автоматического процесса (стрелка Ь) и отрицательного отбора (стрелка а)»; б — «согласованность действия генетико-автоматического процесса (стрелка Ь) и положительного отбора (стрелка а)» (по: Дубинин, 1931); b — «через «узкие кольца»... не может пройти все генное многообразие популяции» (по: Дубинин, Ромашов, 1932)
трансформации отдельного экогенотипа» и т. д. (Дубинин, 1931, с. 474; Ромашов отмечает: «хорошо!!! Wright».)
Рассмотрение «конкретных форм закономерностей популяции» Дубинин ведет с позиций, которые в известной своей части были бы неприемлемы для Ромашова. Так, Дубинин оговаривает, что в анализе взаимодействия ГАП’ов и отбора он рассматривает «только те г.-а. процессы, которые связаны с увеличением концентрации генов (Дубинин, 1931, сн. 1, с. 472), тогда как для Ромашова существенно, что эти процессы в принципе не направлены (дрейф генов). Схема, иллюстрирующая «антагонизм генетикоавтоматического процесса (стрелка Ь) и отрицательного отбора (стрелка а)» — рис. 13, а, и «согласованность действия генетико-автоматического процесса (стрелка Ь) и положительного отбора (стрелка а)» — рис. 13, б (Там же, с. 473, схема 3), помогают нам понять, какой смысл Дубинин в статье 1931 г. вкладывал в понятие «ГАП’ов» и уяснить различие его взглядов и Ромашова. Позже Н. И. Калабухов критиковал эту позицию Дубинина (1931) и сходную — Элтона (Elton, 1930): «Разделяя во времени периоды действия генетико-автоматических процессов и отбора, эти авторы по сути дела сами противоречили себе... Точка зрения, что отбор имеет дело лишь с популяциями, сложившимися уже в результате генетико-автоматического процесса, и не играет роли в изменении соотношения различных форм индивидуальной изменчивости в популяциях, безусловно неверна» (Калабухов, 1941, с. 383). Позицию Ромашова (1931, с. 451), что в «двух зонах концентрации гена» ведущую роль будут играть разные факторы, следует признать более уравновешенной.
Однако наиболее важный вывод—«Генетико-автоматический процесс, прорывая всю медленную последовательность действия отбора, ускоряет процесс трансформации вида» (Дубинин, 1931, с. 473) — вполне сходствует с мнением Ромашова, который рассматривал дрейф как посредник, связующее звено между возникновением мутаций и действием отбора.
Завершает статью 1931 г. Дубинин беглым описанием «4 специфических зон» в «механизме осуществления эволюционного процесса» (с. 478—479), которые суть: мутационный процесс, генетико-автоматические процессы, популяция, эволюционная смена форм.
Представления о роли случайных процессов в эволюции вызвали большой интерес у московских биологов-эволюционистов; первые итоги их обсуждения были подведены в статье Дубинина и Ромашова (1932). В первой главе, отметив, что за счет дрейфа (ГАП’ов) большинство мутаций выбрасывается из популяции вскоре после своего возникновения, авторы ставят вопрос: «погибнут ли все мутационные изменения в ряде последующих поколений?»
123
(Дубинин, Ромашов, 1932, с. 55). Если вспомнить, что Ромашов (1931) рассматривал дрейф’ генов в популяциях как марковскую цепь с конечным числом состояний, то легко видеть, что следующее свойство .марковских цепей приведет нас к ответу. Если можно выделить множество состояний — такое, что за достаточно большое время марковская цепь переходит с положительной вероятностью из любого начального состояния в одно из состояний данного множества, а вероятность выхода системы из данного множества достаточно близка к нулю,— то с подавляющей вероятностью за достаточное число шагов состояние марковской цепи окажется в отмеченном множестве. В нашем случае такое множество включает два состояния: ра =0 и ра = 1. Иными словами, единичная селективно нейтральная мутация за счет дрейфа рано или поздно либо исчезнет из популяции (ра = 0), либо вытеснит свой «нормальный» аллеломорф и захватит всю популяцию (ра = 1; этот исход, конечно, значительно менее вероятен). Хотя авторы статьи 1932 г. могли (скажем, с помощью А. Н. Колмогорова) получить этот вывод аналитическими вероятностными методами, они предпочли для изучения процесса изогаметации обратиться к статистическим испытаниям (метод Монте-Карло). Хотя результаты этих опытов имеют меньшую степень общности, зато, благодаря глубокой аналогии с менделевской стохастикой, они имеют определенный биологический смысл (см.: Кулагина, Ляпунов, 1966; Бабков, 19726,в).
Урновые опыты были хорошо известны Ромашову. Правомерность изучения отдельного эволюционного фактора без учета взаимодействий с другими (вообще возможного благодаря формулировке «условий Харди») обсуждал и Четвериков. В «Заключении» к статье 1926 г. он поставил вопрос: «...не является ли грубой ошибкой такое выхватывание отдельных эволюционных моментов, такой анализ по частям бесконечно сложного и единого эволюционного процесса? Ведь природа не урна с шариками, над которой мы проделываем свои опыты по теории вероятности, а жизнь течет не по руслу математических формул...» Далее он отметил, что хотя в своей работе имел дело лишь с «самой грубой попыткой приближения» к пониманию некоторых эволюционных моментов в согласии с современными генетическими воззрениями, но «подойти к такому сложному явлению, как эволюционный процесс, мы не можем иначе, как предварительно разложив его на составные элементы, расчленив его на отдельные моменты, анализируя его по частям и доводя этот анализ до возможного логического конца» (Четвериков, 1926, с. 51). (Другими словами он говорил о том же в упомянутой выше рукописи 1948 г.: «...Чтобы изучить роль стохастических процессов в чистом виде, мы должны рассмотреть их в чистом же виде, т. е. назависимо как от непрерывно протекающего мутационного процесса, так и от действия отбора». Цит. по: Астауров, 1974, с. 66). Наконец, именно аналитический подход является предпосылкой будущего синтеза: «... сейчас еще рано говорить о синтетическом понимании эволюционного процесса. Только после того, как мы разберемся в основных началах и закономерностях, лежащих как в основании эволюции организмов, в самом широком смысле этого слова, так и явления видообразования, только тогда мы сможем приступить к воссозданию окончательного здания эволюции...» (Четвериков, 1926, с. 51).
Для работы Дубинина и Ромашова (1932) было проведено две серии урновых опытов, моделирующих динамику одного генного локуса в равномерно размножающейся популяции. Исходя из представлений Ромашова, что обычно в природных популяциях заключено большое число мутационных 124
изменений и разные гены представлены в них различными концентрациями (Ромашов, 1931, с. 445), авторы исследовали случай, когда исходная популяция (серия Л) состоит из 100 различных аллелей (в модельной популяции — 100 шаров с разными номерами). Процедура перехода к следующему поколению состояла из 25 выемок (без возвращений) по 2 шара (номер первого из них представлял гибнущий аллеломорф, номер второго — размножающийся) и переноса оставшихся 50 шаров без каких-либо изменений. Такие тиражи продолжались до тех пор, пока один из аллеломорфов не захватывал всю популяцию. В 10 опытах переход модельной популяции к гомозиготности осуществлялся на разных поколениях — от 108 до 465, среднее время изогаметации составило 268,1 поколения. В серии В исходная популяция состояла из 10 различных шаров; среднее время достижения гомозиготности составило 29 поколений (в отдельных опытах процесс заканчивался на 14—51-м поколении). По мнению авторов, эти опыты должны моделировать ситуацию, когда в популяции, скажем содержащей 1000 или 10 000 гамет, сохраняется приблизительно постоянной общей концентрация мутантных генов (10% или 1% для серии Л): «...при этой дифференциации общая сумма мутационных генов остается неизменной...» (Дубинин, Ромашов, 1932, с. 60). Однако эта модель соответствует скорее такой ситуации, когда содержащая 100%-ное разнообразие по одному локусу часть обширной популяции оказывается временно изолированной.
Привлекая материалы Фишера (Fisher, 1930), авторы статьи 1932 г. рассматривают более общую схему — неравномерное размножение в популяции постоянной численности. Такого отклонения от идеально равномерного размножения оказывается достаточно, чтобы повысить интенсивность дрейфа в полтора раза. Однако особенно значительные последствия для генетической структуры популяции дрейф будет иметь при условии заметных колебаний численности, как указал Ромашов (1931). В статье 1932 г. обсуждение эффекта «волн жизни» ведется в терминах «размножающейся части популяции», которая определяет скорость изогаметации и которая может быть существенно меньше «величины живущей популяции». (В нынешней генетике популяций в ходу райтовский термин «эффективная величина популяции», effective size; это понятие С. Райт трактует как математический параметр: «Эффективная величина теории (/V) может, однако, значительно отличаться от общего числа особей популяции (apparent size); она обычно намного меньше. Очевидно, что N относится только к размножающейся популяции (breeding population)» — Wright, 1938, р. 430.)
Вторая глава посвящена примерам колебаний численности популяций разных видов животных в природе. Однако за исключением единственной короткой ссылки на наблюдения Ромашовым (1931) сезонных колебаний численностей подмосковных популяций дрозофил, остальные 25 примеров (занимающих 12 страниц текста!) приводятся по данным других авторов, которые к тому же не связывали такие колебания с генетической эволюцией видов 11 Необходимость такого обилия литературных данных, лишь косвенно
«Что касается литературных указаний на существование и формы протекания генетико-автоматических процессов в природе»,— пишут Дубинин и Ромашов (1932, с. 69),— «то, кроме работ Д. Д. Ромашова (1931) и Н. П. Дубинина (1931), нам известны: 1) работы Гагедурнов (Hagedoorn A. L., Hagedoorn А. С., 1921), в которой авторы разбирают ряд вопросов, аналогичных с затронутыми в настоящей статье; °Днако их подход к проблеме и итоги анализа значительно отличаются от итогов
125
подтверждающих центральную мысль, не очевидна. Представление о широком распространении «волн жизни» в природе не могло быть новостью для Ромашова (с его 20-летним опытом натуралиста), ученика энтомологов С. С. Четверикова и Н. М. Кулагина, чьи работы 1905 и 1930 гг. широко цитировались в статье 1932 г. Дубинин же был учеником А. С. Серебровского, чье внимание концентрировалось на лабораторной генетике дрозофилы и на генетике сельскохозяйственных животных. Такое направление работ (даже если взять в расчет две экспедиции с участием Дубинина) не могло дать ясного представления о явлении «волн жизни».
Интересно, что и для выбранного авторами метода урновых экспериментов характерна наглядность. (Кроме того, он позволяет избежать обращения к сложным математическим методам — обстоятельство, существенное для биолога.* 12) Замечу также, что в отличие от аналитических методов, приводящих к результатам высокой общности, метод статистических испытаний позволяет детально проследить конкретные траектории, моделирующие генную динамику популяции. Такой подход должен быть психологически более приемлемым для генетика, имеющего дело с природными или лабораторными популяциями. Обращение к таким опытам, например, позволило подтвердить тезис Ромашова (1931), что генетическое строение популяции не является непосредственной проекцией всего хода мутационного процесса (см.: Дубинин, Ромашов, 1932, с. 67—68); в результате, мысль о необходимости изучения генного состава локальных популяций получила дополнительное обоснование.
Вслед за статьей Дубинина и Ромашова 1932 г. было проделано несколько работ «с целью получения наглядной картины действия генетико-автоматических процессов в популяции в их чистом виде, без какой бы то ни было примеси действия естественного отбора» (Никоро, Рогозянова, 1938, с. 1139). Сотрудник Дубинина по Институту пушного звероводства В. Г Гагарин (1933) поставил опыты с тремя модельными популяциями D. melanogaster (и соответствующие этим моделям урновые эксперименты). Исходная модельная популяция состояла из 50 пар мух, в том числе 10 пар, гетерозиготных по тому или иному из 10 генов, рецессивных и доминантных. В первой модели пара мух давала пару, и был поставлен один опыт, технически доста
настоящей работы; 2) затем самые общие соображения Элтона (Elton, 1924) по этому вопросу; и наконец 3) указания А. С. Серебровского (1927) на один частный случай генетико-автоматических процессов (значение неравномерного размножения петухов)...»
Дубинин и Ромашов не были тогда знакомы с работой Элтона (Elton 1930), в которой тот дал анализ проблемы зависимости качественных изменений популяций от колебаний их численности. Позже Ромашов и Ильина (1943, с. 307) отметили то место в работе Элтона (Elton 1927, р. 182), где он «очень близко подходит к описанию механизма действия генетико-автоматических процессов».
12 Любопытное свидетельство времени Мария Полиевктовна Садовникова-Кольцова вела в ИЭБ «бригаду» обучения чтению английской научной литературы. Ей помогал в 1930-е годы Герман Германович (г. Дж.) Мёллер. На занятиях он обычно подходил к одному, к другому, спрашивал: «Что Вам непонятно?» Одна из участниц бригады указала в ответ формулу из статьи С. Райта: «Вот это!» Мёллер рассмеялся: «Это и мне непонятно!».
Поскольку к настоящему времени ситуация существенно не изменилась, я избегаю в этом очерке рассмотрения математических аспектов, тем более что почти все это легко найти в любом учебнике, включающем раздел генетики популяций.
126
точно сложный, ибо потребовалось вести более 50 пробирок, чтобы удовлетворить условию: «от пары пара». Во второй модели рассматривалось размножение с независимой вероятностью выживания для каждого потомка (50 пар (Я 50 пар), было поставлено два опыта. В третьей модели (один опыт) размножающаяся часть была в 10 раз меньше — 5 пар. Динамика генного состава прослеживалась на протяжении 10 поколений. В первой модели к Fj0 остались четыре гена-маркера (из 10 в Fj), в том числе два доминантных гена, концентрация которых случайно колебалась на протяжении опытов. В двух опытах со второй моделью фенотипическая однородность достигалась к рб ___ р7; в опыте с третьей моделью — еще раньше. Для нас очевидно, что для экспериментального изучения статистического фактора — дрейфа генов необходима постановка значительного числа экспериментов. Поэтому результатом работы Гагарина мы можем считать прежде всего демонстрацию возможности изучать дрейф генов в лаборатории на модельных популяциях. Спустя 20 лет такой возможностью воспользовались Керр и Райт. Они поставили 96 параллельных опытов (по схеме второй модели Гагарина), каждая из линий содержала только четыре пары дрозофил, а исходные линии были созданы из гетерозигот по forked (р0 = 0,5). К 16-му поколению в41 линии закрепился нормальный аллеломорф, в 29 — forked, 26 оставалось полиморфными (Kerr, Wright, 1954). Другая работа (108 линий с маркером Ваг) демонстрировала совместное действие отбора и дрейфа, так как Ваг имеет отрицательное селективное значение. К 10-му поколению нормальный аллеломорф закрепился в 95 линиях, Ваг — в трех, оставалось 10 полиморфных линий (Wright, Кегг, 1954). В работе 3. С. Никоро и А. И. Рогозяно-вой (1938; доклад Никоро «Экспериментальное изучение действия генетикоавтоматических процессов» был заслушан на 29-м заседании Эволюционной бригады 21.1.1937 г.— «Протоколы...»), задуманной с целью демонстрации действия ГАП’ов, использовались линии white и white-sepia. Было заложено девять моделей популяций с постоянной численностью состава от (1) 10 w +; 90 wse до (9) 90 w +; 10 wse. В двух первых поколениях концентрация генов значительно менялась; оказалось, однако, что это изменение концентрации было связано не с ГАП’ами, а с отбором: на следующее поколение брались первые 100 куколок, а, как выяснилось, личинки линии white-sepia окукливались раньше, чем white. Действие ГАП’ов (когда было исключено влияние разной скорости развития) во всех опытах было заметно лишь в слабой степени. А главным результатом стала постановка вопроса, существуют ли вообще гены, безразличные для особи с точки зрения естественного отбора. Таким образом, в ходе работы интересы исследователей весьма изменились.
Вторая глава статьи 1932 г., помимо примеров колебаний численности («достаточных, чтобы утверждать об их всеобщем характере» — с. 75), содержит некоторые интересные рассуждения. Один случай касается ситуации, когда обширная популяция сжимается до незначительной величины,— ситуация, описанная Ромашовым (1931, с. 446 и 445), примеры приводили Серебровский (1927, 1930, см.: Серебровский, 1976, с. 191, 253—254) и Дубинин (1931, с. 476). Эту ситуацию авторы изображают схемой (рис. 18в). «Совершенно очевидно, что через «узкие кольца», связывающие периоды большой численности вида»,— пишут авторы с том, что сейчас называют эффектом горлышка бутылки»,— «не может пройти все генное многообразие популяций. В моменты сжатий происходит потеря огромного количества ^нов, и вслед за этим генный материал, оставшийся в размножающихся °собях, автоматически распространяется на всю огромную популяцию»
127
Таблица 12
Генетическое строение четырех экогенотипов D. funebris по неопубликованным данным Балкашиной и Ромашова 1929—1931 гг. (Дубинин, Ромашов, 1932)
Мутации Москва (20 линий) Петровско-Разумовское (10 линий) Киев (15 линий) Ташкент (15 линий)
A 1 Б A 1 Б А 1. Б А 1 Б
Extra analis 0 6 15,0 — 0 0
Radius interrupts — 0 — 0 5 8,3 — 0
Plexus crossveinless — 0 1* — — 0 4 6,7
Balloon-club** — 0 4 10,0 — 0 — 0
Venae interruptae — 0 — 0 — 0 3 5,0
Ruber 3 4.8 il - 0 — 0 — 0
Ramuli media — 0 — 0 2 3,3 — 0
Alae curvatae — 0 — 0 2 3,4 — 0
Forked-rough — 0 — 0 — 0 2 3,3
Vortex*** 2 — — 0 8 0 — 0
Puncta*** 0 0 1 — — 0 — 0
♦ Согласно выражению признака, другая мутация, чем в Ташкенте.
** Полулетальное изменение.
*** По-видимому, очень сложное полигенное изменение.
Условные обозначения: А — во скольких линиях появилась мутация; Б — процент гамет.
(Дубинин, Ромашов, 1932, с. 75—76). Несколько позже Райт показал (Wright, 1938 г.), что при циклических изменениях численности эффективная величина популяции (А/) за не слишком продолжительный (в поколениях) цикл, равная среднему гармоническому численностей каждой из фаз W):
N=n/ L
определяется в первую очередь фазой малой численности, т. е. «узким кольцом», как писали об этом Дубинин и Ромашов в статье 1932 г.
В третьей главе «Генетико-автоматические процессы в условиях внутривидовой изоляции и экогенотипы» отправным моментом стал тезис, что «каждый вид является сложной системой достаточно изолированных друг от друга популяций разнообразной величины» (Дубинин, Ромашов, 1932, с. 84), причем благодаря ГАП’ам создается дифференциация генного материала вида по изолятам. Вслед за Четвериковым (1926) Дубинин и Ромашов подчеркнули значение изоляции для процессов видообразования и эволюции; они коротко остановились на тех формах изоляции, что были прежде выделены Четвериковым, и добавляют Промптова (1930) к тем авторам, на которых в этом отношении ссылался Четвериков (Ромене, Мориц Вагнер, Гулик, Кутань, Крэмптон). В результате этого экскурса они сформулировали «закон Вагнера—Гулика» (с. 88), отметив, что он почти дословно совпадает с тезисом Четверикова: «при прочих равных условиях степень дифферен циации внутри вида прямо пропорциональна степени изоляции отдельных его частей» (Четвериков, 1926, с. 26).
128
В качестве примера «реальных форм экогенотипов в природе» Дубинин и Ромашов коснулись анализа генетического строения популяций дрозофил и3 Звенигорода, Геленджика и Берлина, проведенных лабораторией Четверикова, анализа популяций D. funebris из Москвы, Петровско-Разумовского, Киева и Ташкента (Балкашина, Ромашов, 1929—1931, неопубл, данные — сМ. табл. 2) и популяции D. melanogaster из 14 географических пунктов, обследованных большой группой под руководством Дубинина. Особый интерес представлял тот факт, что многие резко выраженные «вредные» признаки — вплоть до полулеталей и леталей —достигали в ряде локальных популяций высоких частот (например, balloon, al. curvatae, forked-rough — в табл. 12) — «груз мутаций».
Теоретические соображения Ромашова и Дубинина о роли случайных процессов в эволюции популяций развивались на основе общей эволюционной схемы Четверикова и явились ее конкретизацией с позиций выяснения роли отдельных факторов генетической эволюции. Они выросли из необходимости более полно объяснить результаты первого экспериментального исследования свободно живущих популяций дрозофилы и дали основу дальнейшим целенаправленным исследованиям форм природной изменчивости и механизмов ее поддержания.
ЛЕТАЛИ В ПРИРОДНЫХ ПОПУЛЯЦИЯХ
Определение дрейфа генов Ромашовым (1931) включало представление о различии генетической структуры у одновременно живущих изолированных популяций и у различных поколений одной популяции; оно должно было также объяснить существование в природе леталей и аберративной (видимой) изменчивости.
К сравнительному изучению географически разобщенных популяций D. melanogaster большая группа генетиков (работавших в ИЭБ и на кафедре генетики Института пушного звероводства) приступила летом 1931 г., когда статья Ромашова, написанная зимой 1930/1931 г., еще не вышла в свет, а статья Дубинина еще не была написана. Материал был взят из 15 популяций Северного Кавказа, Закавказья и Центральной России (Кольцов, 1934, с. 223) и проанализирован для 10 популяций. Следующим летом повторно изучали генный состав пяти популяций, так что эта работа должна была не только «продолжать генетический анализ популяций вида,... первую попытку чего сделал на примере D. melanogaster С. С. Четвериков» (Дубинин и др., 1934, с. 168), но и экспериментально подтвердить тезис, что дрейф генов приводит к различиям как между одновременно живущими разобщенными популяциями, так и между последовательными поколениями одной популяции. Анализ проводили: М. Г Цубина (Кисловодск, 1931 г.), С. Я. Бессмертная (Машук, 1931 —1932 гг.), М. А. Гептнер (Ессентуки, 1931 — 1932 гг.), Н. П. Дубинин (Владикавказ, 1931 г.), Б. Н. Сидоров (Батум, 1931 г.), Л. В. Ферри (Делижан, 1931 г.), С. В. Сапрыкина (Пятигорск, 1931 г.), К. А. Панина (Тамбов, 1931 г.), Э. Погосян (Эривань, 1931 г.), С. Ю. Гольдат (Армавир, 1931 г.), В. Н. Беляева и Е. Д. Постникова (Пятигорск, 1932 г.), 3. А. Демидова (Владикавказ/Орджоникидзе, 1932,), А. П. Кроткова (Эривань, 1932 г.); в работе участвовала также
С- Никоро. Опубликована эта работа была в двух частях в 1934 г. и Русский текст сопровождал сокращенный английский перевод (Дубинин и
В. В. Бабков
129
# Ш Л Ш Е Ш Е Ш
не-Су Су
Рис. 14. Метод анализа, позволяющий обнаружить рецессивные мутации в больших хромосомах и летали по II хромосоме Drosophila melanogaster
Анализирующая линия несет доминантные гены, летальные в гомозиготном состоянии
Су, L, D, Sb, Сц^С^^запирает кроссинговер в III хромосоме, Су — во II)
В исследовании 1925—1926 гг. был использован метод инбридинга, чтобы перевести отдельные локусы в гомозиготное состояние и таким образом выявить рецессивные мутации. В обширной работе 1931 —1934 гг. примени лась остроумная методика (основанная на системе балансированных лета лей, т. е на том же принципе, что и метод С1В, «бар-це-эль» Меллера Бриджеса), которая дала возможность переводить в гомозиготное состояние целые хромосомы (рис. 14), выявляя рецессивные изменения в больших (II и III) хромосомах и летали во II хромосоме. (Анализировать летали в III хромосоме не позволило отсутствие доминантного гена-маркера в хромо соме С1И анализирующей линии II III ) Позже Гершензон
L Сц[ /
использовал модификацию этой методики, чтобы переводить в гомозиготное состояние все четыре хромосомы.
Исследование 1931 г. показало, что дикие популяции чрезвычайно гетерогенны. Каждая популяция имела ту или иную степень «зараженности» рецессивными генами от популяции Армавира, где 52,7% хромосом несли наследственные изменения, до популяции Пятигорска с 23,8%. (В выборке из популяции Дилижана мутации вообще не были найдены, однако в исследовании 1943 г. было показано, что концентрация мутаций только в одной из больших хромосом равна 73,4%, а в среднем на одну муху
130
приходилось три гетерозиготные мутации — Берг, 1945, с. 181.) Наследственное содержание отдельных популяций качественно различалось. При сопоставлении генетического строения 10 изученных выборок (по 200 самок) авторы обратили внимание на факт, что из общего числа найденных видимых мутаций (61) число мутаций, встречавшихся повторно, равно 37, а обнаруженных лишь однажды — 24, и добавляли: «...являются ли наши единичные мутации действительно единичными или мы в силу недостаточного числа изученных хромосом просто не получили картину повторности, нам неизвестно» (Дубинин и др., 1934, с. 173), демонстрируя, таким образом, свое устремление обнаружить результаты ГАП’ов.
Помимо «хороших» мутаций изучались «так называемые малые мутации, не имеющие собственного выражения, которые модифицируют проявление разных генов, позволяют отбору пластично и последовально изменять признаки» (Там же),— т. е. «генотипическая среда» sensu Четвериков. О существенной роли малых мутаций позволил судить анализ количественных признаков в выборках из разных популяций и анализ проявления гена Dichaete в генотипических средах разных популяций — своего рода геногеографическая работа, аналогичная той, что Тимофеев-Ресовский провел для выяснения влияния генотипической среды на модус изменчивости vti (см. гл. 3).
Исследовались и другие ^категории наследственных изменений. Признак «добавочные щетинки» был найден во всех выборках в весьма высоких концентрациях — до 30—50% и выше. Этот признак, как и другой массовый признак — «запятая» на груди, имел нерегулярное проявление и выражение, но выяснение природы этих изменений не входило в задачу авторов.
Вторая обширная категория изменений, которая обеспечила этому исследованию место в истории генетики,— летали: в разных выборках от 7,6 до 21,4% II хромосом несли летальные гены. На Колд-Спринг-Харборском симпозиуме 1955 г. Добржанский говорил об этом: «небывалый — и поразительный!— результат» (Dobzhansky, 1955, р. 4). В свете последующих работ эти числа представляются весьма низкими (ср.: Dobzhansky, 1970, р. 119), однако именно эта работа впервые определенно показала, что процветающие природные популяции несут груз мутаций, которые могут привести к наследственным нарушениям различной степени тяжести. Интересно, впрочем, что сами авторы статьи 1934 г. не считали нужным уделить специальное внимание проблеме груза мутаций. Найденные результаты (например, «в популяции Владикавказа деталь-1 имеет концентрацию в 3,75%, а ген крыльев (пузыри) только лишь 0,75%» — Дубинин и др., 1934, с. 199) они использовали для доказательства, что именно ГАП’ы играют ведущую роль в определении генетического строения популяций: « ..все обнаруженные гены имеют отрицательное отборное значение, и это показывает, что те большие концентрации генов, которые налицо в экогенотипах, возникли вопреки действию естественного отбора... Вполне возможно, что очень многие мутации дрозофилы имеют отрицательное отборное значение, равное летальным генам... Отбор является могущественным фактором, в равной степени направленным против подавляющего большинства мутаций, загоняющим наследственное многообразие вида под нормальный фенотип. И под покровом этого нормального фенотипа, из поколения в поколение поддерживаемого естественным отбором, генетико-автоматиче-Ские процессы, идущие в условиях колебания численностей популяций, в условиях изоляции и панмиксии, определяют дифференциацию наследствен
9*
131
ного содержания вида по экогенотипам и характерные черты строения генотипа* (Дубинин и др., 1934, с. 199).
Вторая часть статьи 1934 г. была посвящена сравнению генного состава пяти популяций в 1931 и 1932 годах. Результаты показали, что от лета 1931 г. к лету 1932 г. в генетическом строении популяций произошли существенные изменения — количественные и качественные, но с позиции теории ГАП’ов это уже не могло быть новостью. Авторы пришли к выводу о необходимости значительно увеличить размер выборки: «...тщательное и систематическое изучение 1—2 популяций на тысячах хромосом и в ряде лет позволит нам вскрыть исчерпывающую картину строения и динамики экогенотипов» (Дубинин и др., 1934, с. 215).
Частью реализации этого грандиозного плана стало изучение выборок из популяции D. melanogaster Геленджика, которое Н. П. Дубинин, М. А. Гептнер, 3. А. Демидова и Л. И. Дьячкова провели в 1933—1935 гг. (Дубинин и др., 1936, опубликована на русском и английском языках). В выборке 1933 г. (анализ потомства 903 самок, 1754 аутосом) было найдено 226 нелетальных наследственных изменений, связанных с 20 различными генами: 6 из них встретились в виде единичных гетерозигот, остальные повторно (от 2 до 19 раз), и особенно больших концентраций достигали, как и в исследовании 1931 —1932 гг., «добавочные щетинки» и «запятые». Было найдено также 70 летальных генов из II хромосомы. С позиций концепции ГАП’ов особый интерес представлял вопрос: все ли из этих 70 генов различны или же некоторые летали встречаются повторно? Анализ, потребовавший более 2500 скрещиваний, показал, что среди 51 изученной летали оказался 31 различный ген: 22 гена были найдены в качестве единичных гетерозигот, 5 встретились два раза, 1 — четыре раза и 3 — пять раз (концентрация 0,6%). (Этот результат согласовался с полученным ранее (1934), что летали широко распространены в природе и наряду с единичными некоторые могут встречаться в определенной концентрации.
Еще один результат дало сравнение генного содержания выборки по летальным и нелетальным мутациям. Разнообразие нелетальных мутаций (20 в хромосомах II и III) меньше, чем летальных (31 или больше, в хромосоме II); однако концентрации некоторых нелетальных мутаций (по крайней мере «массовых изменений») достигают значительного уровня, и в результате абсолютное число их выше, чем леталей.
Анализ выборок из той же геленджикской популяции, но в другие годы — в 1934 г. (58 ? 2 , 1232 аутосом) и в 1935 г. (813 ?? , 1594 аутосом),— позволил сделать некоторые заключения о динамике ее генетической структуры. Из 20 мутаций 1933 г. некоторые не были обнаружены в 1934 г. (зато были найдены некоторые новые); в 1935 г. появилась одна из «утерянных», но исчезли еще шесть из первоначальных, утерялись некоторые из приобретенных в 1934 г. и появились новые и т. д. В 1934 г. соотношения между летальными и нелетальными генами оставались теми же, что и в 1933 г. Анализ показал, что из 33 леталей 1933 г. и 55 леталей 1934 г. общими оказались 9 генов: «для летальных генов смена генного содержания популяции прошла в большей степени, чем для нелетальных» (Дубинин и др., 1936а, с. 944—945),— утеряно 66,6% леталей против 45% нелеталей; обнаружено 83,6% новых леталей против 31,2% нелеталей.
Оценивая «картину движения» генов в целом, наблюдавшуюся в течение трех лет в популяции Геленджика, авторы приходят к выводу, что генное содержание популяции довольно устойчиво, несмотря на действие ГАП’ов.
132
«Многие из генов, исчезновение которых мы наблюдали в последующем году, возможно, не совсем уничтожены в популяции, а только перешли на такой низкий уровень, что ускользнули от нашего, все же ограниченного анализа. Другие гены, вновь появившиеся, может быть, просто достигли больших концентраций и поэтому стали улавливаться экспериментом» (Там же, с. 959). Эти слова показывают, что авторы сознавали, что их анализ включал лишь крайне ограниченные выборки и что следует соблюдать осторожность при распространении заключений о динамике выборок на динамику обширной популяции. Более адекватный анализ такого статистического явления, как дрейф генов (или ГАП’ы), требовал бы привлечения методов математической статистики, в частности, непременного рассмотрения круга вопросов, включающих оценку достоверности и репрезентативность выборки.
Как оценить эти экспериментальные работы в свете четвериковской проблемы: природа и поддержание изменчивости в популяциях? Анализ генофондов диких популяций («экогенотипов») дрозофилы предпринимался, чтобы обнаружить и оценить генетическую дифференциацию в пространстве (1931 —1932 гг.) и во времени (1933—1935 гг.), обязанную дрейфу генов (ГАГГам) — одному из механизмов поддержания изменчивости в популяциях. Однако самый интересный результат касался природы изменчивости: высокая насыщенность диких популяций деталями. Этот факт дал впоследствии основание, чтобы выделить новый аспект в центральной проблеме эволюционной генетики: генотипическая изменчивость по приспособленности.
Теоретические и экспериментальные исследования ГАП’ов были основаны на эволюционной схеме Четверикова и явились конкретизацией ее в следующих аспектах. Концепция ГАП’ов указала механизм поддержания внутри-популяционной и межпопуляционной генетической изменчивости, который служит связующим звеном между возникновением мутаций и естественным отбором. В этой связи был поставлен вопрос о роли отдельных факторов популяционной динамики. Исходя из концепции ГАП’ов, было предсказано — и обнаружено в эксперименте — существование в природных популяциях неадаптивного полиморфизма и деталей, что дало основу для обсуждения проблемы генетического, в частности мутационного, груза.
133
Глава 9
ПОЛИМОРФИЗМ ПРИРОДНЫХ ПОПУЛЯЦИЙ
Все формы похожи,
И ни одна не одинакова с другой, И так весь хор их указывает На тайный закон...
и. В. ГЕТЕ
АБЕРРАТИВНЫЙ ПОЛИМОРФИЗМ
В первом популяционно-генотическом исследовании, проведенном группой Четверикова в 1925—1927 гг., выяснилось, что природные популяции дрозофил не являются внешне (фенотипически) совершенно однородными. Этот феномен получил генетическое объяснение в эволюционно-генетической концепции Ромашова, где стохастическим процессам отводилась существенная роль в формировании фонда популяционной изменчивости (Ромашов, 1931). С целью изучения ГАП’ов, как мы видели, проводился обширный анализ диких популяций (Дубинин и др., 1934, 1936а). Ромашов планировал также ряд других исследований, связанных с теорией ГАП’ов (часть из них была окончена и опубликована). Касаясь работ по проблеме «генетика и эволюция», Ромашов в своем докладе (в отчете «Работы Института экспериментальной биологии Наркомздрава к XVII съезду ВКП(б)», 1934) отметил: «III. Экспериментальное изучение генетического строения популяций ведется в настоящее время на различных видах дрозофилы (см. доклад Балкашиной). Кроме того, осенью 1933 г. Н. П. Дубининым и Д. Д. Ромашовым были изучены явления полиморфизма в природных населениях D. melanogaster. При осмотре 35 ООО «диких» мух из различных популяций Черноморской области было найдено очень большое (до 3—4%) число резко измененных особей, по внешности напоминающих описанные у этого вида мутации, например: spineless, eyeless, ebony, sepia и т. д. В настоящее время заканчивается генетический анализ этих изменений, причем выяснилось, что часть из них наследственна и идентична указанным мутациям» (Работы..., 1934, с. 223—224).
В 1937 г. Н. П. Дубинин, Д. Д. Ромашов, М. А. Гептнер и 3. А. Демидова опубликовали результаты анализа аберративного полиморфизма у D. melanogaster, проведенного при участии К. А. Паниной, А. А. Гавриловой и Л. И. Дьячковой в 1933—1935 гг.1 Ключевым для этого исследования было понятие аберрации как низшей таксономической единицы, введенное в 1910 г. А. П. Семеновым-Тян-Шанским. (В 1921/1922 гг. Ромашов участвовал в обсуждении на Кружке им. Ламарка доклада Б. С. Кузина «Разбор работы А. П. Семенова-Тян-Шанского «Таксономические границы вида и его подразделений» (1910)».) Материалы, собранные в популяциях Кавказского побережья Черного моря, побережья Каспия и в популяциях Средней Азии, показали, что аберративный полиморфизм у D. melanogaster носит «всеобщий тождественный качественный характер по всему изученному
На 26-м заседании Эволюционной бригады (9.1.37) обсуждался доклад Д. Д. Ромашова и Н. П. Дубинина «Аберративный полиморфизм в природных популяциях Dr. fasciata (metanogaster)*. («Протоколы...»). Для сбора материалов они выезжали в 1933 г. в Геленджик, Архипо-Осиповку, станицы Крымскую и Славянскую, а в 1934 г. в Буйнакс и Дербент.
134
ареалу» (Дубинин и др., 1937а, с. 332). Генетический анализ показал, что появление аберраций в природе связано с распространением рецессивных и доминантных генов, а также с наличием ненаследственной изменчивости. При этом в согласии с тем, что ранее отметили Балкашина и Ромашов (1935) и Тимофеевы-Ресовские (Н. A. Timofeeff-Ressovsky, N. W. Timofeeff-Ressovsky, 1927), «наблюдается поразительный параллелизм между наследственной и ненаследственной изменчивостью» (Дубинин и др., 1937а, с. 331). В целом результаты этого исследования подтвердили выводы работы Балка шиной и Ромашова (1935), в частности, что неправильно наследующиеся и пластичные в своем проявлении изменения составляют общий фон наследственной изменчивости у ряда видов дрозофил, и на этом фоне лишь изредка «проскакивают» отдельные резкие изменения типа лабораторных мутаций (ср. гл. 2 наст, книги). «Малая изменчивость крайне широко распространена во всех изученных нами популяциях; на ее общем фоне подчас даже теряется определенное, фиксированное представление о так называемом «нормальном фенотипе» D. fasciata» (Дубинин и др., 1937а, с. 315—316; авторы здесь употребляют видовое название fasciata — синоним melanogaster). Особый интерес представляет то обстоятельство, что некоторые основные массовые изменения (добавочные щетинки, добавочные жилки, неполная анальная жилка и т. п.), свойственные, по Балкашиной и Ромашову (1935), популяциям D. funebris, transversa, phalerata, vibrissina, насыщали также все изученные популяции D. malanogaster. В «Обсуждении» среди основных факторов, определяющих аберративный полиморфизм, выделяется раздел «Закономерности феногенетики вида» (с. 333—335) — тема, прошедшая у Ромашова от первого экспериментального исследования популяций до цикла работ по феногенетике массовой популяционной изменчивости.
Другая серия работ Ромашова по изучению систематического полиморфизма, выполненная совместно с Е. Д. Ильиной, была проведена на разных группах пушных зверей (Ромашов, Ильина, 1942, 1943)2 В первой из них рассматривалась мутация черно-бурой окраски у лисицы. Черно-бурая лисица и крестовка отличаются от нормальной красной (bb) наличием неполнодоминантного гена В в гомозиготном состоянии. Гетерозиготы ВЬ представляют промежуточные формы меланизма; степень выражения зависит от особого гена D, при наличии которого (BD) возникают более темные формы — сиводушки, при отсутствии его (Bbdd) — бастарды. Поскольку три генотипа — BB, Bb, bb — хорошо различаются по внешности, то эта мутация оказывается удобной для проверки распределения генотипов по формуле Харди.
По материалам, собранным Ильиной в 1931 г. по 22 факториям, сравнивалось найденное распределение генотипов с ожидаемым по формуле Харди. Хорошие совпадения между ожидаемыми и фактически найденными числами позволили заключить, что внутри данных районов отсутствует резко выраженная дифференциация по гену В, иными словами, что лисицы достаточно свободно скрещиваются между собой в пределах обследованных районов (средней площадью около 2500—3000 км2). Авторы сравнивают в этом отно-
са 9-м заседании Эволюционной бригады (23.XII.34) был заслушан доклад Д- д. Ромашова (по материалам Е. Д. Ильиной). «Аберрации у пушных зверей (°тр. хищные)». На 8-м заседании (9.ХП.34) на очередной Съезд зоологов был Рекомендован доклад Е. Д. Ильиной и Д. Д. Ромашова «Генетические основы полиморфизма у пушных зверей». («Протоколы...»)
135
тении популяции лисицы и популяции хомяка. С. В. Кириков (1934) описал резкую дифференциацию популяций хомяков Башкирии по доминантной мутации черной (и черно-пегой) окраски: на расстоянии в несколько десятков километров колебания в концентрации этого гена достигают 30—50% и более. «Таким образом, «системы скрещиваний» у обоих видов оказываются весьма различными» (Ромашов, Ильина, 1942, с. 224)3 Следующим был вывод от отсутствии (при данных условиях) заметно выраженного естественного или искусственного отбора по какому-либо из трех генотипов. Наконец — и это самое интересное — «анализ по формуле Гарди может дать ответ на вопрос о таксономическом значении черно-бурой лисицы и сиводушки» (Там же). Этот вопрос много раз поднимался в систематической литературе. Бутурлин (1913) высказывал мнение, что черно-бурая и сиводушка соответствуют подвидам красной лисицы; Житков (1932) считал, что они представляют собой «ряд морф» (морфами называются географически не локализованные и обычно не наследственные изменения); Огнёв (1931) возражал против «одностороннего положения», что каждая сиводушка несомненный гибрид между черно-бурой и красной лисой, и считал, что появление сиводушек может быть обусловлено как различными комплексами генов, так и различными условиями среды. Анализ Ромашова и Ильиной с такими толкованиями не согласуется. «Если бы сиводушки (и черно-бурые) могли возникать как ненаследственные морфы или если бы их появление было связано с различными (помимо В) генами, то распределение генотипов по формуле Гарди не могло бы осуществиться. Таким образом, сиводушки и черно-бурые не являются ни подвидом, ни морфой красной лисицы, а представляют типичные наследственные аберрации, гетерозиготные или гомозиготные по мутации В (или у канадских лисиц по мутации R)» (Ромашов, Ильина, 1942, с. 224; 1943, с. 302).
В обширной работе Ромашова и Ильиной 1943 г. наибольший для нас интерес представляет сопоставление генетических и таксономических категорий (см. схему). Отмечая, что «явления, протекающие в естественных поцуляциях, гораздо сложнее и многообразнее, чем это удалось отразить на нашей схеме», авторы подчеркивают, что самым существенным является факт, что «в пределах вида можно установить самую тесную связь между последовательностью генетических категорий, с одной стороны, и последовательностью таксономических категорий, с другой. Этим еще раз подчеркивается та неразрывная связь, которая существует между генетическими представлениями и представлениями современной систематики» (Ромашов, Ильина, 1943, с. 310). Отметив, что глубокое значение этой связи недостаточно учитывают, «многие генетики и многие систематики», авторы указывают, что необходимы дальнейшие работы в этих пограничных областях: «Они в конечном итоге должны привести к тому, чтобы совместными усилиями систематиков, генетиков и экологов построить ту «новую систематику» (New Systematics — Хаксли), которая должна явиться одной из наиболее интересных и прогрессивных отраслей современной биологии» (Там же).
3 Гершензон и Полевой (1941) отметили, что в природных популяциях осуществляется свободное скрещивание черных и обычных хомяков. Это, впрочем, не отменяет вывода Ромашова и Ильиной (что подтверждают теоретические и экспериментальные исследования пространственной дифференциации — Бабков, 1973а; Новоженов, 1981).
136
Упрощенная схема отношений генетических и таксономических категорий (без учета модификационной изменчивости; Ромашов, Ильина, 1943):
Мутационный процесс Генетико-автоматические процессы Генетические категории Мутация Экогенотип Таксономические категории Аберрация (aberratio) Локальная вариация (varietas localis)
Естественный от- Экотип и Экологическая Племя
бор климатип вариация (natio)
(varietas) oecologica)
Подвид Подвид (Subspecies)
Вид Вид (Species)
АДАПТАЦИОННЫЙ ПОЛИМОРФИЗМ АДАЛИЙ
У божьих коровок Adalia bipunctata из популяций разного географического происхождения описан ряд форм рисунка надкрылий (черные пятна на кирпично-красном фоне и красные на черном), который можно разделить по преобладающему цвету на две группы: рецессивные «красные» и доминантные «черные». В условиях Средней Европы и Центральной России адалия дает за вегетационный период три (иногда четыре) поколения; последнее перезимовывает. На основе десятилетних наблюдений Н. В. Тимофеев-Ресовский показал наличие сезонной изменчивости у адалий, которая покоится на «совместной игре сильно действующего положительного отбора одной группы форм зимою и другой летом» (Timofeeff-Ressovsky, 1940 с, S. 135).
Материалом послужила популяция A. bipunctata из Бух-Берлина. Местом массовой зимовки служили глубокие щели между каменными плитами южной стены старой церкви. В первые теплые солнечные дни весны (в начале апреля) божьи коровки массами выползали из мест зимовок в течение двух—трех дней; осенью, в последнее теплое время (в конце октября) они массами слетались на зимовку. В эти периоды безвыборочно собирались жуки с определенной площади стены и после подсчета «красной» и «черной» форм выпускались на волю. С 1930 по 1944 гг. (с пропуском 1932 г.) таким способом учитывался состав популяции до и после зимовки. Соотношение двух форм близко к 1:1, и во всех без исключения случаях в весенних сборах преобладали жуки красных форм, а в осенних — черных. В весенних сборах доля красных форм варьировала за 10 лет в пределах от 57 до 71%; в осенних — от 30 до 49% (по: Тимофеев-Ресовский, Свирежев, 1966, с. 139. В статье 1940 г. частоты аллелей вычислялись на основе ошибочного предположения рецессивности черных форм). Отсюда следовало, что относительная жизнеспособность двух форм летом и зимой различна. Для Дальнейшего доказательства был проведен следующий опыт. В щелях, служивших зимовками, всегда после вылета оставалось большое число погибших за зиму жуков. В 1934, 1937 и 1938 гг. четыре особенно крупные щели, содержащие по 150—350 жуков, начисто опоражнивались незадолго до вылета; жуки переносились в лабораторию для оживления при комнатной температуре и подсчитывалось число живых и мертвых жуков Для каждой из форм. Красные формы обладали повышенной, по сравнению с черными, способностью перезимовывать: в среднем оживало 11,1% крас-
137
Рис. 15. Адаптационный полиморфизм ада-лий. Изменения частоты гена А з зимние и летние периоды за 1929—1940 гг. (по: Тимофеев-Ресовский, Свирежев, 1966)
Абсцисса — частота гена А, обусловливающего черную окраску надкрылий; ордината — скорость изменения частоты гена А
ных и 4,23% черных жуков. (Аналогичный опыт в 1930, 1931, 1933 гг. для одной зимовки дал 10,86 и 4,48%). За три летних поколения черные обгоняют красных и преобладают в осенних сборах. В более поздней работе (Тимофеев-Ресовский, Свирежев, 1966) в предположении случайного скрещивания форм были вычислены коэффициенты относительной жизнеспособности черных форм в зимние сезоны (за 10 лет от 0,26 до 0,60 относительно красных форм) и относительной жизнеспособности красных форм в летние сезоны (от 0,40 до 0,95) и была построена фазовая траектория системы, описывающая изменение частоты гена А черной окраски в популяции под действием разнонаправленного давления отбора в разные сезоны года (рис. 15).
В отличие от гетерозиготного полиформизма, где устойчивое полиморфное состояние обеспечивается повышенной относительной жизнеспособностью гетерозиготы, в случае A. bipunctata полиморфизм сбалансиро-
ван во времени и обеспечивается сменой вектора отбора зимой и летом. Этот случай Тимофеев-Ресовский назвал адаптационным полиморфизмом,
подчеркивая его значение для создания пластичности вида в условиях обширного географического ареала.
Я. Я. Лусис (1961; см. также: Лус, 1928, 1932) составил (по собственным сборам, музейным коллекциям и литературным данным) таблицу и карту, показывающие соотношение черных и красных форм A. bipunctata в различных популяциях Европы. Оказалось, что в местностях с более влажным, морским климатом процент черных форм выше, чем в районах с более выраженным континентальным климатом. Из таблицы усматривалась также закономерность «микрогеографического» порядка — процент черных форм в больших промышленных городах выше, чем в сельской местности: по сборам Лусиса 1931 —1947 гг. в Ленинграде (Васильевский остров и Петроградская сторона) черные формы составили 88,2%, в парке Петергофа — 22,8%, в Гатчине — 9%.
Вопрос о причинах численного нарастания черных форм летом и падения зимой Тимофеев-Ресовский оставил открытым, заметив, что выбор одного из вероятных объяснений возможен по проведении соответствующих генетико-физиологических опытов.
Лусис возразил (1961, с. 12), что экспериментальные исследования недостаточны, а нужны непосредственные наблюдения в природе. В частности, нужно выяснить: имеет ли место строго случайное сочетание форм
138
при спариваниях; нет ли различий по половой активности. Результаты наблюдений по десяти московским и рижским популяциям A. bipunctata, обработанные методом х2» показали, что гипотеза о случайном сочетании форм при спариваниях в данном случае неприменима. Адалии с черными надкрыльями и переднеспинками более активны по сравнению с красными жуками, о чем Лусис заключил по процентам жуков красной и черной форм в состоянии копуляции от общего числа соответствующих форм в популяции. Во всех случаях этот индекс выше у черных форм.
Различную активность черных и красных форм Лусис, привлекая литературные данные, ставит в зависимость от различной восприимчивости лучистой тепловой радиации жуками с сильно и слабо меланизированными надкрыльями. Темные пигменты способствуют прогреванию насекомого на свету (температура тела имеет большое значение для физиологии насекомого) и в то же время ускоряют потерю тепла в темноте.
Отмечая, что более частое копулирование обеспечивает более верное оплодотворение большего количества яиц при одинаковой потенциальной плодовитости, Лусис объясняет летний отбор в пользу черных форм их большей фактической плодовитостью, которая коррелирована здесь с физиологическим значением меланистической окраски. Зимний отбор в пользу красных форм также стоит в зависимости от окраски как терморегулятора тела: темные пигменты ускоряют остывание тела. На этой основе Лусис объясняет и географическую изменчивость полиморфизма адалий. Наконец, микро-климажбольших промышленных городов создается под влиянием засорения атмосферы примесями твердых частиц; потеря количества солнечной радиации составляет около 20%, а при низком стоянии солнца до 50%; эти условия, очевидно, менее благоприятны для красных форм. Биологическое значение полиморфизма Лусис, как и Тимофеев-Ресовский, видит в том, что разные реакции разных форм на многообразные и изменчивые во времени условия внешней среды позволяют виду более полно использовать среду обитания.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ ХОМЯКОВ-МЕЛАНИСТОВ
Один из наиболее интересных случаев географического распределения единичных мутаций доставляет меланистическая форма хомяка Cricetus cricetus, наследуемая как типичный полно проявляющийся аутосомный доминант; эти мутации спорадически возникают в разных популяциях. Во время путешествия по северной Башкирии в 1768—1769 гг. известный русский географ Иван Лепёхин (1775 и др.) наблюдал, что обычно редкая меланистическая форма составляла значительную часть популяции хомяка в небольшой зоне северо-восточного угла его ареала, между реками Кама и Белая. За полтораста лет эта форма увеличила свою долю в популяциях северной Башкирии и распространилась вдоль северной границы ареала вида на запад, до Днепра. Этот процесс можно было бы точно проследить (Кириков, 1934 и др.) благодаря важной роли, которую играет хомяк на пушных рынках восточной России. Хомяк — типичный грызун сухой зоны русских степей, но меланистическая мутация, по-видимому, лучше, чем нормальная форма, приспособлена к более прохладному и влажному климату лесостепной зоны (см.: Timofeeff-Ressovsky, 1939, 1940а и др.). Э. Форд трактовал этот случай как типичней пример переходного полиморфизма (transient polymorphism — Ford, 1945, р. 74). Более
139
детальное наблюдение за меньшее по масштабу время, хотя и не дает оснований говорить о переходном полиморфизме, но позволяет выяснить роль естественного отбора и других факторов в распространении и динамике меланизма у хомяка. Такое исследование предпринял Гершензон (1946; Gershenson, 1945).
Меланизм (в среднем около 3%) встречался не на всей территории видового ареала хомяка на Украине. Для объяснения этого факта Гершензон рассмотрел три возможности (незавершенная диффузия черной мутации хомяка; изолирующие барьеры внутри ареала; различие экологических условий внутри ареала) и исключил две первые, так как не были найдены ни градиенты частоты меланистов, ни естественные преграды, разделяющие районы наличия и отсутствия меланистов. Оставалось разобрать третью возможность. При сравнении карты распространения хомяков на Украине с картами температур, осадков, почвенными и геоботаническими выяснилось, что ареал меланистов относится к области, получающей более 500 мм осадков в год, и совпадает с зоной распространения оподзоленных черноземов и лесостепной растительности (Гершензон, 1946, с. 104). Этот вывод в целом находится в согласии с мнениями Б. М. Житкова, что меланисты встречаются преимущественно на супесчаных почвах, покрытых дубовыми или смешанными лесами (Schitkow, 1905), Хильцхеймера, что на Волге они более часты в сырых местах (Hilzheimer, 1913), Кирикова (1934), рассматривавшего их как лесные и прибрежные формы (цит. по: Gershenson, 1945, г. 209).
Другой особенностью распространения хомяков-меланистов на Украине была его очаговость, которую Гершензон также связал с экологическими условиями. (Так, граница Черниговского очага высокой концентрации меланистов совпадала с границей, делящей данный район на две резко различные в естественно-историческом отношении части: концентрации меланистов выше в тех районах, где больше низменных и заболоченных мест, и ниже там, где преобладают сухие и холмистые места.) Кроме того, концентрация меланистов коррелирует с плотностью населения хомяков (ср Petzsch, 1936: частота меланистов возрастает в годы общего подъема численности хомяков и падает в годы ее уменьшения — цит. по: Gershenson, 1945, р. 209).
В Башкирии, где концентрация меланистов значительно выше, они встречаются лишь на территории с той же экологией, что и у области распространения меланистов на Украине (и в наиболее низких частях Башкирии — высота над уровнем моря менее 200 м). Границы очага высокой концентрации меланистов (и высокой плотности населения хомяков) довольно отчетливы, но здесь определенно можно говорить о градиенте. Эти все данные хорошо подтверждают выводы Кирикова (1934), а также Дьяконова и Суллы (1935) (см.: Gershenson, 1945, р. 219).
Затем Гершензон занялся выяснением вопроса, существуют ли сезонные изменения частоты черных хомяков в обследованных популяциях, так как именно такие исследования, как показал Тимофеев-Ресовский на адалиях (см. выше) и Добржанский на дрозофилах (Dobzhansky, 1943), могут пролить свет на механизм действия ествественного отбора. Материалы по Полтаве за 1935—1938 гг. показали, что за зиму концентрация черных хомяков составила 0,44—0,69 (рис. 16). Однако характер сезонных колебаний частоты черных хомяков в Черниговской и Каменец-Подольской областях представлял совершенно иную картину: частота меланистов каждую зиму возрастала (1,95—7,05 и 1,11—2,11). Материалы по Башкирии дают картину, отличающуюся от двух первых: зимой 140
Рис. 16. Различия сезонной динамики Частот хомяков-меланистов (по: Гершен-здн, 1946)
\ 1 — в Черниговской обл., п = 579 355;
2 —кв Полтавской обл., п = 147 461
1936/1937 гг. и 1938/1939 гг. частота меланистов намного возрастала (1,11 —1,20), а зимой 1937/1938 гг. немного снижалась (0,98); к концу же лета она всегда резко возрастала. Эти материалы говорят о разнообразии экологических условий во времени и по ареалу вида. Общие тенденции изменения частоты меланистов на Украине за 1934—1939 гг. таковы. Эта частота в одних районах имеет тенденцию к росту, 3 S ДРУГИХ К падению (первая преобладает), причем рост захватывал западные части Украины, а падение — восточные, и эти процессы характеризовали сплошные пространства. Рост частоты меланистов приходился на области непрерывного расширения их ареала; это расширение происходило путем, который зоогеографы называют «языковым»: увеличение шло в одном или немногих направлениях и сплошность ареала не нарушалась. Это последнее обстоятельство Гершензон считал нужным подчеркнуть прежде всего потому, что «оно лишний раз убедительно показывает, что случайное переживание (или «генетико-автоматические процессы») не играют сколько-нибудь значительной роли в географическом распределении хомяков-меланистов» (Гершензон, 1946, с. 119—120). Это обращение Гершензона к ГАП’ам несомненно обязано выдающейся роли, которую теория ГАП’ов играла в стимулировании обсуждений механизмов генетической эволюции.
Заключительная часть русской публикации Гершензона (ее нет в английском переводе, опубликованном Добржанским, зато в нем дан обзор литературы) посвящена общим соображениям по поводу этапов эволюционного закрепления благоприятствуемого естественным отбором признака. Гаксономический ранг черных хомяков оценивался разными авторами по-разному. Лепёхин (1775) считал их разновидностями, но думал, что пни скрещиваются в себе. Паллас (Pallas, 1778) отмечал, что они не образуют отдельных видов. Шребер (Schreber, 1792) первым рассматривал lx как подвиды, а Труэссар (Trouessart, 1910) дал им наименование Cricetus cricetus niger (Schreb.). Симрот (Simrot, 1906) назвал их мутациями, Стах (Stach, 1920)—«морфой», Кириков (1934) определял их как жотип (цит. по: Gershenson, 1945, р. 209). Гершензон (1946, с. 125) фактически принял взгляд Ромашова (см. выше схему), и утверждал, <го три основные формы, в которых втречается меланизм у хомяков — единичные мутации, диморфная популяция, экотип,— соответствуют трем важнейшим этапам, которые должен пройти под действием отбора *Утантный признак от момента возникновения и до закрепления в каче-
признака, характеризующего разновидность. Эта позиция, отводящая Явственному отбору выдающуюся роль в становлении нормальных призна-
141
ков, получила подтверждение в оценке Гершензоном интенсивности отбора по Холдейну. Из этой оценки он сделал важные выводы, что, во-первых, не только интенсивность, но и направление (т. е. вектор) отбора различны в каждой области в разные годы, и, во-вторых, интенсивность отбора почти всюду велика. /
Изученная Гершензоном картина географического распределения меланизма у хомяков показывает, что экстраполяции направления и темпов эволюции на обширные ареалы и длительные промежутки времени, основанные на исследовании ограниченных территорий за немногие годы, будут весьма ненадежны. Она также’ дает четкое указание на гетерогенность природной среды — факт, который необходимо учитывать при построении моделей эволюции популяций.
ХРОМОСОМНЫЙ ПОЛИМОРФИЗМ ДРОЗОФИЛ
11 января 1935 г. Б. Ф. Кожевников доложил на 12-м заседании Эволюционной бригады («Протоколы...») результаты работы «Об искусственной внутривидовой изоляции» (Кожевников, 1936; Kozhevnikov, 1935). По заранее намеченному плану он получил в эксперименте линию с хромосомной аберрацией, репродуктивно изолированную от нормальных линий D. melanogaster. В работе были использованы две взаимные транслокации между II и III хромосомами. (В одной II и III хромосомы обменялись своими левыми концами; в другой II обменяла свой левый конец на правый конец III.) У гетерозиготных по этим транслокациям самок \россинговер по III хромосоме между точками разрыва дает структуру, где обе транслокации из II хромосомы собраны у одной III хромосомы. Если эта структура сочетается с двумя компенсирующими по обеим транслокациям II хромосомами, то мухи с таким генотипом имеют нормальный хромосомный баланс (см. рис. 17), нормальную жизнеспособность и могут разводиться как константная линия (хотя 3/4 потомства и гибнет). Эта раса, кариотипиче-ски изолированная от нормальных представителей исходного вида, напоминавшая в этом отношении особый нарождавшийся вид, получила название Drosophila artificialis. «Со времен Ч. Дарвина создание искусственного вида, бесплодного при скрещивании с другими видами, было мечтой биологов-эволюционистов»,— заметил по этому поводу Н. К. Кольцов (1937, с. 939). Эта блестящая экспериментальная работа Кожевникова привлекла внимание советских исследователей к внутривидовой хромосомной изменчивости, открыв новый аспект разработки проблемы «природа и поддержание изменчивости в популяциях».
Введение в начале 30-х гг. методики исследования гигантских хромосом в слюнных железах личинок дрозофилы, обязанной открытию Гесса и Пайнтера, дало возможность просто и точно регистрировать хромосомные аберрации. И в том же 1935 г. в популяциях D. melanogaster Геленджика было просмотрено 918 III хромосом (459 особей) и , найдено 68 хромосом с инверсией (7,4%); обнаружение трех гомозиготных по инверсии особей говорило о ее полной жизнеспособности (Дубинин и др., 1936, с. 960; Dubinin et al., 1936). Между представлением о чрезвычайной редкости структурных мутаций хромосом и фактом, что эволюция кариотипов основана на большом Их числе, существовал разрыв, и Н. П. Дубинин, Н. Н. Соколов иГ Г Тиняков (19376) предприняли исследование внутривидовой хромосомной изменчивости с целью этот разрыв заполнить. (Доклад этих авторов 142
Рис. 17. Кариотипическая изоляция (по: Kozhevnikov, 1935)
а- новый кариотип Drosophila melanogaster, сконструированный Б. Ф. Кожевниковым — «Drosophila artificialis* (III хромосома и ее фрагменты — черным цветом, II — белым); б — кариотип самки, гетерозиготной по обеим реципрокным транслокациям. Стрелки указывают пределы регионов, расположенных между транслокациями. Крос-синговер в этом регионе ведет к возникновению сложной хромосомы с двумя транслока-
циями
«Внутривидовая хромосомная изменчивость в роде Drosophila* обсуждался на 50-м объединенном заседании Эволюционной бригады совместно с 250-м заседанием генетического коллоквия, 29.XII.1937 г.).
При обследовании популяций нескольких видов дрозофил из 26 географических пунктов (и нескольких популяций Chironomus) было найдено среди 42 315 хромосом 939 аберраций — почти все инверсии (только один случай внутрихромосомной транслокации у D. obscura), т. е. распространение хромосомных мутаций в природе резко отличается от картины при освещении лучами Рентгена, где преобладают транслокации. Причину различий авторы относят за счет отрицательного отбора гетерозигот по транслокациям (Дубинин и др., 19376, с. 1038).
Внутри каждого из обследованных видов дрозофил были найдены «гомологичные инверсии» (Там же, с. 1019): на всем изученном ареале D. melanogaster найдено семь разных инверсий в 525 хромосомах с инверсиями йз 34 515 хромосом; у D. funebris — пять разных инверсий (207 из 5517 хромосом); отдельные инверсии широко распространены по ареалу вида. Исследование видов, условно названных D. obscura 2 и D. obscura 3, дало иную картину: найдено много внутрихромосомных транслокаций; насыщенность популяций хромосомными мутациями вообще очень велика; высокий полиморфизм популяций сопровождался указанием на географическую дифференциацию. (Все это близко к позднейшему выделению «домашних» и «диких» видов дрозофилы — ср.: Dobzhansky, 1970, р. 133.)
Наконец, авторы разбирают причины, обусловливающие картину насыщения популяций хромосомными мутациями; рассматривают действие мутационного процесса, ГАП’ов в условиях изоляции и панмиксии и, что особенно интересно, действие отбора на инверсии как «блоки генов». При этом они подчеркивают, что «Судьба инверсий в популяции в связи с коэффициентом отбора и концентрацией инверсии исследована А. А. Малиновским (не опубликовано)» (Дубинин и др., 19376, с. 1040), и ведут обсуждение отчасти в связи с этим анализом. (Эта интересная работа «Роль хромосомных инверсий в эволюции вида» (Малиновский, 1940), которую также отметил Дж. Хаксли (Huxley, 1942, р. 332), была выполнена в Математической группе при Эволюционной бригаде ИЭБ и доложена на Математическом коллоквиуме ИЭБ.)
Отсутствие видимых изменений у мух с инверсиями поначалу приводило k неверному мнению, что инверсии адаптивно нейтральны (например, во втором издании книги «Генетика и происхождение видов» — Dobzhansky, 1941). Но в 1943 г. было показано, что хромосомные инверсии, связывая блоки
143
генов, подвергаются естественному отбору, следствием которого является сезонный цикл изменения концентпации инверсий в популяциях D. pseudo-obscura (Dobzhansky, 1943, 1971). В качестве материала Дубинин и Тиняков выбрали не подобную «дикую» лесную форму, а «домацц^юю» D. funebris, которая реагирует на изменения условий жизни, вызванные человеком; в 1938 г. Г Г Тиняков (1965) составил для этой работы карту слюнных хромосом D. funebris. Дубинин и Тиняков выяснили, что четыре популяции Москвы содержат от 27% до 88%. гетерозигот по инверсиям, в среднем 52%. (Определялись три инверсии II хромосомы и по одной III и IV хромосом.) В трех сельских популяциях были найдены те же инверсии, но с насыщением гетерозигот около 1,4%. Чтобы исключить возможную роль ГАП’ов, связанных с изоляцией, они исследовали популяции промышленного г. Иваново и его окрестностей и получили те же результаты: 37,2 и 0%. Более того, популяция г. Воронежа, разрушенного войной, дала весной 1945 г. 0,9% гетерозигот по инверсиям, что подтвердило связь кариотипической структуры с экологией. Авторы отметили реальность огромных скоростей отбора (необходимых для перестройки популяции Воронежа) и факт, что и в городе, и в деревне нет полного единообразия хромосомных структур, и отнесли это за счет систем балансированного полиморфизма (Дубинин, Тиняков, 1946; Dubinin, Tiniakov, 1946; см. также: Соколов, Дубинин, 1940; Малиновский, 1947; Соколов, 1959; Борисов, 1970).
Дубинин, Соколов, Тиняков (1937) отметили различия концентраций трех инверсий в Московской популяции D. funebris в декабре и августе, но заключили, что «общий процент гетерозигот идеально совпадает» (Там же, с. 1023). После работы Добржанского 1943 г. Дубинин и Тиняков провели в 1944—1945 гг. регулярное помесячное обследование трех московских популяций и выявили сезонный цикл инверсий (с сентября по ноябрь концентрация гетерозигот возрастает, а с марта по май падает). Сравнение инверсий II-1 и IV-1 популяции Ботанического сада Университета показало их противоположное поведение: если П-1 возрастала по частоте от 33,7 до 52,2% с мая по июль, то IV-1 падала от 14,0 до 1,6%. В популяции Кропотово (сельская местность) те же инверсии были в очень низких концентрациях и не показывали цикличности (Dubinin, Tiniakov, 1946а). В опытах по влиянию зимовки выяснилось, что частота инверсии II-1 падает после зимовки, что перезимовавшие мухи дают на треть больше яиц, дочери (FJ перезимовавших более плодовиты и плодовитость перезимовавших в природе мух выше, чем летних (Dubinin, Tiniakov, 1946b). В тех же опытах с выпуском гомозиготных по инверсиям мух оказалось возможным оценить скорость миграции D. funebris за летний день — 50— 100 м и оценить относительные приспособленности зигот: гетерозиготы по инверсиям более приспособлены, чем стандартные гомозиготы, а те -более, чем гомозиготы по инверсиям (Dubinin, Tiniakov, 1946с).
Дубинин и Тиняков в ходе изучения экологического расообразования у D. funebris на основе хромосомного полиморфизма (Дубинин, Тиняков, 1946, 1947а, б) установили существование пяти зон, для которых наблюдали характерную концентрацию инверсий (Дубинин, Тиняков, 19476). От центра к окраине шло снижение гетерозигот по инверсиям II-1 и П-2: Центр (88%) — Садовое кольцо (55%) —Окружная железная дорога (12%) —Немчинов-ка/Загорск (1,8%) —200/500 км (0). В среднем в Москве концентрация П-1 составила 40%, П-2— 16,3%. А. И. Борисов (1969) обследовал карио-144
Рис. 18. Эволюция инверсионного полиморфизма Drosophila funebris (по: Борисов, 1969)
Пункты, обследованные: 1 — в 1947 г. Н. П. Дубининым и Г Г Тиняковым; 2 — в 1968 г. А. И. Борисовым. А — граница ареала городской расы в 1947 г., В — в 1968 г.; Б — зона интерградации в 1947 г.; Г — граница Москвы (кольцевая автострада)
фонд через 20 лет. Он нашел, что частота гетерозигот по II-1 возросла от 40 до 50—56% за счет других инверсий (П-2 упала с 16,3 до 9,4—0%). Сезонные изменения частоты инверсии II-1 в популяциях Нагатино и Ленинского проспекта таковы:
Месяц
Октябрь
Апрель
п-1/st. %
58
58
П-1/П-1. %
27
5,7
st/st. %
15
36
Это — картина хорошо сбалансированной коадаптированной системы, в которой стандартный и инвертированный порядки генов реципрокно реагируют на сезонные изменения. Но самым ярким здесь был факт расширения ареала городской расы — с 25 км2 до 800 км2 (см. рис. 18): в 1947 г. в популяциях Измайлова, Хорошева, Воробьевых гор гетерозиготы составили 12%, в 1968 — 50%.
Этот цикл работ показывает, что полиморфные виды дрозофил служат хорошей моделью для изучения процессов генетической эволюции: адаптивное значение инверсий дает возможность проследить самые ранние этапы эволюции популяций, когда различия между носителями тех или иных генных порядков касаются физиологических свойств, улавливаемых только при специальных исследованиях конкурентоспособности, плодовитости и других показателей жизнеспособности.
Исследования полиморфизма природных популяций существенно обогатили картину генетических основ эволюции. Их результатом была замена первоначального представления об эволюции как отборе единичных мутаций более разработанным, поясняющим, какими путями мутационные изменения могут достигать и поддерживать концентрации, достаточно высокие для эффективного их отбора, т. е. каким образом мутации могут выходить на эволюционную арену. Благодаря этим исследованиям был сделан шаг от взгляда на популяцию как механический набор аллелей («beanbag genetics* — Майр) к представлению о популяции как системе, гДе выделяются разные уровни организации — процессов образования генофонда и процессов создания балансированных коадаптированных систем полиморфизма, которые обеспечивают быстрые приспособительные Реакции популяции как целого.
В. В. Бабков 145
Глава 10
ВЗАИМОДЕЙСТВИЕ ФАКТОРОВ ЭВОЛЮЦИИ /
Лишь полнота порождает ясность...
ШИЛЛЕР
ПОСТАНОВКА ПРОБЛЕМЫ
Первые представления об организации механизмов эволюции были изложены в теоретическом исследовании Четверикова 1926 г. Натуралист и дарвинист Четвериков обратился к экспериментальной генетике, имея в виду прежде всего частный, хотя и существенный, вопрос — о наличие в природе наследственной изменчивости, в количестве, достаточном чтобы обеспечить эволюцию посредством отбора. Его первым шагом, как отмечено выше, стала независимая формулировка ок. 1922 г. «Основного закона популяции» (как его называли в своих статьях Ромашов, Дубинин, Эфроимсон и др. и сам Четвериков), согласно которому в условиях свободного скрещивания «население вида», пока на него не действуют внешние силы, находится в состоянии генетического равновесия, которое достигается в результате одного стабилизирующего скрещивания. «Закон популяции» описывает нереализуемую в природе ситуацию, однако он определяет точку отсчета для изучения воздействия факторов эволюции на «генотипическое строение вида»; в этом-то и заключается его ценность.
В гл. I «Возникновение геновариаций в природе» Четвериков разобрал первый элементарный фактор эволюции — мутационной процесс. И при этом подчеркивал, что он не тождествен процессу эволюции: «из мутаций непосредственно, сразу, никакого вида не получается» 1 Для процесса видообразования, по Четверикову, имеет значение географическая изменчивость — изменчивость более высокого ранга, чем индивидуальная (мутационная). Впрочем, задача — поставить дарвинизм на фундамент генетики — предполагала выяснение стартовых механизмов эволюционного процесса. Отсюда — постановка проблемы образования генофонда популяции, для рассмотрения которой Четвериков сразу же выдвинул, в самом общем виде, и проблему взаимодействия факторов эволюции: «в природе все время осуществляются два противоположных друг другу процесса: один — накапливания геновариаций, другой — их устранения, и от состояния взаимодействия между этими двумя процессами зависит существование в природе большего или меньшего числа геновариаций» (Четвериков, 1926, с. 13). Разработка проблемы факторов эволюции явилась, по сути дела, конкретизацией этого тезиса.
Для рассмотрения этих процессов Четвериков ввел представление о популяции как арене первичных эволюционных преобразований (он говорил о «населении вида», «совокупности особей, составляющих единый свободно скрещивающийся комплекс»). Свободное скрещивание «засасывает» рецессивные мутации, расшатывает признаки вида, сохраняя внешнюю однородность. Напротив, фактор изоляции создает условия, благоприятные для выщепления наследственной изменчивости. Естественный отбор — антагонист свободы скрещивания — стремится изменить внешность вида и сохранить его устойчивость и мономорфность.
1 Фонд Четверикова, on. 1, ед. хр. 11, л. 3.
146
\ рассмотрев возможность неадаптивных эволюционных процессов в при-рХе, Четвериков счел должным расчленить общепринятое представление об эволюционном развитии органического мира и рассматривать по отдель-hoct\i два процесса, подчас переплетающихся, но все же строго разграниченных в своих причинах и вытекающих из них последствиях. Один — пронес распадения вида, где преобладающая роль падает на изоляцию, находящую себе материал в. гетерогенности природных популяций. «Истинной причиной происхождения видов является не отбор, а изоляция» (Там же, с. 42). «Другой ведет к адаптации, к прогрессивной эволюции органической жизни — и причиной его является борьба за существование и вытекающий из нее естественный отбор» (Там же, с. 43).
Таким образом, Четвериков рассматривал органическую природу в процессе эволюции как определенную иерархически организованную целостность, и соотносил факторы эволюции с разными уровнями биологической организации. Мутационный процесс осуществляется на физиологическом уровне (а его продукт служит материалом популяционной динамики). Свобода скрещивания и изоляция действуют на популяционном уровне (здесь решается судьба отдельных мутаций и определяются условия протекания отбора). Естественный отбор относится к более высокому уровню, включающему живую и неживую внешнюю среду.
На первый взгляд в этой схеме усматривается одностороннее движение — от более низкого к более высокому уровню. (Тогда она, в принципе, сохраняет возможность сводить эволюцию к направленным мутациям.) Четвериков же на этом не остановился. Его попытка, в гл. IV «Генотипическая среда», выяснить каналы обратной связи («...Или же он [отбор] сам создает для себя материал, активно вмешивается в эволюционный процесс, направляет изменчивость по определенному руслу?» — Там же, с. 43) впервые привела к взгляду на эволюцию как саморегулируемый процесс. С позиций сегодняшних знаний эта попытка выглядит отрывочной и неполной; однако она важна прежде всего как постановка вопроса. Впоследствии трактовку процесса эволюции с позиций кибернетики дал Шмальгаузен (1968). В этой связи отмечу, что в истории науки следует четко разграничивать две области исследования: исследование генезиса и исследование традиций явлений науки; эти области, одновременно касающиеся вопроса о связи явлений, противоположны как по критериям, так и по своей относительной ценности. Шмальгаузен употреблял термины теории связи и ссылался на работы Шредингера, Шеннона, Винера, послуживших поводом для него. Постановка же проблемы и подход Шмальгаузена продолжали известные тенденции русских натуралистов, которые ярко выражены, например, у Вернадского и Четверикова.
Проблема факторов эволюции, таким образом, была поставлена в статье 1926 г. как проблема их взаимодействия.
ГЕНЕТИКА И ЭВОЛЮЦИЯ
Программа Ромашова. Ближайшим результатом теоретического исследования 1926 г. стало развертывание работ по анализу природных популяций. Разработка вопроса о факторах эволюции явилась следствием обсуждения теории ГАП’ов. Программа работ по проблеме «генетика и эволюция» была определена Ромашовым в докладе 20.1.1934 г. на объединенном заседании шести лабораторных коллоквиев ИЭБ (Работы.. 1934).
10*
147
Первый пункт программы касался теоретического изучения «главнейших процессов, определяющих генетическое строение вида» (Там же, с. 223}, во втором речь шла о протекании ГАП’ов в популяциях, третьим было экспериментальное изучение аберративного полиморфизма (в том числе массовой популяционной изменчивости). В рамках первого направления Ромашов предполагал написать совместно с Дубининым обширную работу и наметил ее план: 1) статистический анализ процессов, определяющих генетическое строение популяций; 2) зависимость их от биологии вида; 3) анализ вопроса о низших таксономических единицах (Там же). Однако весной 1935 г. Ромашов тяжело заболел; болезнь вывела его из строя почти на два года: проведя острый период в клинике, он отправился в Казахстан — выздоравливать и пополнять свои коллекции жуков. По его возвращении в конце 1936 г. в ИЭБ была написана статья с его участием по аберративному полиморфизму у дрозофилы (Дубинин, Ромашов и др., 1937а — гл. 9), но к теоретическому очерку он приступил позже. Наброски книги «Изоляция и теория естественного отбора», в которой Ромашов выделил две части: 1) «математические предпосылки теории ГАП’ов» и 2) «ГАП’ы и экологическая структура популяции», относятся, как отметили Балкашина и соавторы (1975, с. 84), к 1940—1941 гг.; рукописный, на 9 стр., «Проспект» этой книги из 5 глав (добавлены; 3. Генетическое строение популяций. 4. Система вида. 5. ГАПы и теория естественного отбора) датирован 20.XII.1943 г. В начале Великой Отечественной войны Институт (к тому времени Институт цитологии, гистологии и эмбриологии АН СССР — его историю см.: Астауров, Рокицкий, 1975, гл. 2 и с. 151 —154) был эвакуирован в Алма-Ату. Ромашов же оставался в Москве, и с 1942 г. работал во Всероссийском НИИ прудового рыбного хозяйства, где он открыл естественный гиногенез 2 у серебряного карася (см.: Головинская, 1975), а позже изучал действие ионизирующей радиации на рыб. В конце 30-х и 40-е годы он с сотрудниками вел исследования аберративного полиморфизма по «массовым» признакам у двукрылых (гл. 11) и анализировал материалы по аберративной изменчивости у пушных зверей (гл. 9). В одной из работ этого цикла (Ромашов, Ильина, 1943) был сделан важный шаг к анализу строения низших систематических единиц с позиций генетики, в соответствии с гл. 3 по плану 1934 г. и гл. 4 «Проспекта» 1943 г. (Материалы Комиссии). В полном объеме этот план реализован не был.
По второму пункту программы Ромашов предполагал участие Дубинина и Колмогорова в работе «Учение о равновесии популяции и процесс изогаметации», где в развитие его идей 1931 г., «исходя из математических построений, доказывается ошибочность «закона равновесия» Гарди, делается детальный статистический анализ протекания процессов изогаметации в условиях полной и частичной изоляции и устанавливается зависимость между течением этих процессов и многообразными условиями существования популяции (численность популяции, колебания численности, степень изоляции, инбридинг, соотношение полов и т. д.). Исходя из приведенного анализа, предполагается дать критический разбор основных положений геногеографии (работы А. С. Серебровского и др.) (Работы..., 1934, с. 223). Эта работа не была осуществлена Ромашовым, но затронутые здесь вопросы продолжали волновать его и позже, как явствует из плана книги
2 Э. Вильсон, введший понятие о гиногенезе, определял его как развитие яйца, активированного сперматозоидом, но лишенного отцовских хромосом.
148
н\чала 40-х гг. Не разрабатывал без Ромашова эти вопросы и Дубинин; однако к некоторым частным аспектам их обратился А. Н. Колмогоров (1935, 1940; Колмогоров и др., 1937), входивший в Математическую группу при ^Эволюционной лаборатории ИЭБ (которая существовала с января 1937 <10 июль 1940 г.)
Оптимальная структура популяций. Первая из этих работ Колмогорова (1935) связана с проблемой оптимальной организации популяций (обсуждение: Бабков, 1972в), которая впервые была намечена в теоретическом исследовании Четверикова 1926 г. в связи с вопросом о значении численности популяции для проявления ее генотипической изменчивости, т. е. для успеха процессов отбора. Четвериков подчеркнул, что здесь мы встречаемся с двумя противоположными тенденциями: частота возникновения мутаций прямо пропорциональна численности, а вероятность их проявления в гомозиготах обратно пропорциональна, и они уравновешивают друг друга. Но если популяция приобретает определенную инфраструктуру, то равновесие нарушается: «Если мы представим себе, что общая численность W данного вида разбилась на ряд к изолированных колоний, то частота возникновения новых геновариаций в пределах всего вида от этого не пострадает, но вероятность проявления вновь каждой из таких геновариаций будет значительно повышена в зависимости от меньшей численности (п) той колонии, в пределах которой она возникла» (Четвериков, 1926, с. 23; об эволюционном значении размера природных популяций см.: Новоже-нов, 1981). Дубинин и Ромашов (1932, с. 85) подтверждали эти рассуждения статистическими опытами, показавшими влияние численности популяции на интенсивность ГАП’ов: «...в обширном населении после его раздробления на отдельные колонии создаются условия, исключительно благоприятные для фенотипической реализации огромного наследственного многообразия, первоначально заключенного внутри вида».
А. А. Малиновский — один из первых участников Эволюционной бригады и ее секретарь до января 1938 г. (с февраля — В. С. Кирпичников) — в 30-е годы начал заниматься проблемой соотношения строения и функционирования различных типов биологических систем, развивая системную традицию Четверикова. (Впоследствии эти интересы привели его к созданию общей классификации биологических систем в связи с их приспособительным значением: Малиновский, 1960 и др.) С этих позиций он рассматривал процессы и механизмы эволюции; этот подход и позволил ему поставить проблему «оптимальной структуры популяции», как назывались три его доклада на заседаниях Эволюционной бригады (9.XI.34, 5.III.35 и 3.IV.37 — «Протоколы...»). На основе представлений Ромашова о двух зонах концентрации гена и общей схемы Четверикова Малиновский сформулировал принцип существования оптимума частичной изоляции для отбора рецессивных генов, а в качестве примеров предложил две схемы ограниченного скрещивания. Одна из них — формализация указанной Четвериковым ситуации, с тем весьма существенным добавлением, что между частными популяциями предполагалась слабая связь благодаря обмену странствующими особями. (Аналогичную схему рассматривал Райт в контексте «адаптивного поля генных комбинаций» — Wright, 1931.) Другая схема Малиновского моделировала ситуацию, где ареал индивидуальной активности каждого индивида существенно меньше ареала популяции. В этой схеме особи, составляющие популяцию, равномерно распределены на плоскости, и скрещивание допускалось лишь между непосредственными
149
соседями — предельный случай, прямо противоположный полной панмиксии, где пространство как бы стянуто в точку. /
Колмогоров подверг математическому анализу первую из этих /схем, применив «прямое диффузионное уравнение Колмогорова* к случаю стационарного распределения вероятностей для концентрации гена. Он получил формулы для концентраций трех генотипов в большой популяции (которые при полной панмиксии определяются формулой Харди) и для случая с отбором — формулу, которая «может быть использована для подтверждения... общего положения о существовании оптимума частичной изоляции для отбора рецессивных генов, выдвинутого и обоснованного с качественной стороны А. А. Малиновским* (Колмогоров, 1935, с. 131), а также количественное выражение этого оптимума частичной изоляции (или связи) локальных популяций. Это уравнение Колмогорова (1938; Kolmogoroff, 1931) играет фундаментальную роль в генетической теории частот; на его основе возникло целое направление в исследовании генетических ситуа ций: Crow, Kimura, 1956; Feller, 1951; Kimura, 1955, 1957; Wright, 1945; Ратнер, 1977 и мн. др. Еще одну ситуацию, интересовавшую Ромашова,— слияние популяций исследовали тем же методом А. Н. Колмогоров, И. Г Петровский, Н. С. Пискунов (1937).
Вторая схема Малиновского не поддавалась обработке аналитическими методами; позже она была исследована методами машинного моделирования (Бабков, 1972а, 1973а). Анализ показал, что в такой ситуации (резкое различие ареала популяции и ареала индивидуальной активности индивидов) возникает, именно как следствие структуры скрещивания, пространственная генотипическая дифференциация. Она может обусловить возникновение адаптивного полиморфизма при осуществлении различных микроусловий на обширном ареале; в ограниченных же популяциях она будет благоприятствовать отбору рецессивных генов, в соответствии с принципом Малиновского (Бабков, 1972в, 1973а, 1975).
Первоначальная модель генетической эволюции Четверикова, на которой основаны модельные рассуждения Дубинина и Ромашова и принцип оптимума Малиновского, как и, с другой стороны, концепция Райта, включавшая аналогичную модель подразделенной популяции с миграциями («островная модель*), придавали первостепенное значение рецессивным аутосомным мутациям. Изложенные в гл. 11 наст, книги работы группы Ромашова привели его, а вслед за ним и других исследователей к иным соображениям о природе эволюционно значимой популяционной изменчивости, где на первый план было выдринуто значение полимерных полудоминантных неправильно наследующихся изменений. Исходя из соображений Ромашова, Гершензон (1941 с. 102) критиковал представление о подразделенной популяции как эволюционно выгодной и заключил, в соответствии с известным мнением Дарвина, что «наилучшей обстановкой для прогрессивной эволюции является, напротив, большая территория с многообразными условиями среды, обусловливающими возникновение адаптивного полиморфизма популяции». Из критического обсуждения Гершензона, по-видимому, было бы неправильно заключить, что подразделенная структура всегда невыгодна и в природе не встречается. В том же самом номере «Журнала общей биологии» был опубликован генетический анализ двух природных популяций дрозофилы, где приспособленность вида как системы рисовалась «как равнодействующая между приспособленностью к внешним условиям и приспособленностью к тем процессам, которые совершаются в генотипе» (Берг и др., 150
1941. с. 153—154). В частности, наблюдавшаяся высокая доминантность вредных мутаций, повышающая интенсивность отбора, рассматривалась как приспособление, противостоящее отрицательному действию высокой мута-бильности. Берг и соавторы заключили, что «Процесс повышения доминирования вредных мутантных генов может совершаться только в популяциях, имеющих определенную структуру», а именно в результате конкуренции между отдельными относительно изолированными микропопуляциями — модель, отвергнутая Гершензоном. «Как показали наши исследования, есть основания предполагать, что популяции Drosophila melanogaster обладают такой структурой» (Там же, с. 153). Поэтому более уравновешенным представляется мнение, что в изменчивой истории органического мира выдающуюся роль могут играть различные структуры природных популяций.
Автор последней упомянутой работы 3 начали с указания на большое влияние теории ГАП’ов и тут же определили свою цель: дать материал в пользу признания первостепенного значения мутационного процесса и отбора в динамике природных популяций. Действительно, именно теория ГАП’ов служила в 30-е гг. стимулом к обсуждению факторов эволюции. Также и Эфроимсон отдал дань ГАП’ам в работе, посвященной четвериков-ской теме: отношениям давлений отбора и мутаций. Основываясь на материалах по частотам гемофилии, заячьей губы, пигментного ретинита, амавротической идиотии и целого ряда других наследственных заболеваний он оценил темпы мутационного процесса у человека 4 (Позже аналогичную работу провел Холдейн — Haldane, 1935.)
В ряде интересных работ исследовались такие важнейшие биологические свойства мутантов, как относительная жизнеспособность или половой аффинитет. 3. С. Никоро с сотрудниками (1935) изучила дифференциальную скрещиваемость нескольких мутаций D. melanogaster, или половой аффинитет, с которым связана одна из форм изоляции. В ее данных встречались все возможные случаи: отсутствие полового аффинитета к своей или чужой форме, наличие аффинитета к самкам своего или чужого генотипа. Четвериков высоко оценил эту работу (см. гл. 1).
Тимофеев-Ресовский сравнивал жизнеспособности географичнских рас D. funebris (Timofeeff-Ressovsky, 1935; см. также: Timofeeff-Ressovsky, 1932, 1933, 1940d; Тимофеев-Ресовский и др.). Поскольку мухи разных рас внешне неразличимы, то жизнеспособность их определялась относительно стандартной линии D. melanogaster. Опыты были поставлены при трех температурах, соответствующих условиям регионов, откуда происходили различные линии (табл. 13). При более высокой температуре (29° С) жизнеспособность D. funebris отчетливо ниже, чем D. melanogaster, что согласуется с различиями в географическом распространении двух видов: D. melanogaster происходит из тропиков и распространилась на умеренную зону, D. funebris обитает в более умеренной зоне. Данные по жизнеспособности рас D. funebris при разных температурах показы-
3 Она была доложена 21.III. 1938 г. на 58-м заседании Эволюционной бригады, объединенном с 261-м заседанием генетического колоквия ИЭБ («Протоколы...»). Это интересно отметить по той причине, что создание Эволюционной бригады было инициировано теорией ГАП’ов и обсуждение ГАП’ов занимало в ее планах существенное место.
В. /7. Эфроимсон. О скорости мутационного процесса у человека. (19326) (Доложено на генетическом коллоквии ИЭБ в конце 1932 г.). См. также: Эфроимсон, 1932а, 1940.
151
Таблица 13
Относительная жизнеспособность линий D. funebris различного географического происхождения (Timofeeff-Ressovsky, 1935)
Номер Происхождение линий Жизнеспособность D. funebris (в % к жизнеспособности D. melanogaster) Жизнеспособность D. funebris (в % к жизнеспособности линий
№ 1 D. Funebris)
15° | 22’ | 29° 15’ | 22° | 29°
1 Средняя Европа 81 42 18 100 100 100
2 Швеция 88 40 21 108,6 95,2 116,6
3 Норвегия 80 41 21 98,7 97,6 116,6
4 Дания 79 44 22 97,5 104,7 122,2
5 Шотландия 84 43 20 103,7 102,4 111,1
6 Англия 78 42 21 96,3 100,0 116,6
7 Франция 80 44 25 98,7 104,7 138,8
8 Португалия 71 45 28 97,6 107,1 155,5
9 Испания 69 48 30 85,2 114,3 116,6
10 Италия 78 43 25 96,3 102,4 138,8
11 Галлиполи 75 44 26 92,6 104,7 144,4
12 Триполи 64 47 31 79,0 111,9 172,2
13 Египет 68 46 30 83,9 109,5 166,6
14 Ленинград 90 43 22 111,1 102,4 122,2
15 Киев 91 44 28 112,3 104,7 155,5
16 Москва 101 43 28 124,7 102,4 155,5
17 Саратов 92 42 30 113,6 100,0 166,6
18 Пермь 98 41 26 121,0 97,6 144,4
19 Томск 96 42 28 118,5 100,0 155,5
20 Крым 87 42 28 107,4 100,0 155,5
21 Кавказ-1 89 43 31 109,9 102,4 172,2
22 Кавказ-2 86 45 32 106,2 107,1 177,7
23 Туркестан 90 44 34 111,1 104,7 188,8
24 Семиречье 92 46 36 113,6 109,5 200,0
вают, что географические расы хорошо гармонируют с климатом тех регионов, откуда они происходят: мягкий климат северо-западной Европы, континентальный — центральной России, жаркое лето и мягкая зима Средиземноморья, контрасты температур Средней Азии.
Эволюционный семинар и Эволюционная бригада ИЭБ. Первые публикации Ромашова и Дубинина, касающиеся ГАП’ов, вызвали необходимость подробного обсуждения ряда затронутых в них вопросов. С этим было связано возобновление в ИЭБ интереса к эволюционной тематике, что привело и к некоторым структурным изменениям в Институте. В ИЭБ был приглашен консультант-математик (М. В. Игнатьев, позже — В. И. Гливенко и Н. В. Смирнов); в программу Генетического отдела были вновь (после работ лаборатории Четверикова) введены темы по изучению генетического строения популяций дрозофил; наконец, осенью 1932 г. Ромашов, по предложению Кольцова, организовал при Генетическом отделе ИЭБ Эволюционный семинар. За три года его работы было проведено 33 заседания с докладами на общеэволюционные темы — по истории эволюционизма,
152
его узловым современным проблемам и посвященные анализу отдельных моментов дарвинизма 5
Доклады А. Д. Некрасова выделялись среди исторических. В них он по записным книжкам Дарвина разобрал рост его эволюционных идей и пересмотрел «инцидент Уоллес — Дарвин», дал оценку отношений Э. Жоф-фруа и Кювье и роли Ламарка в развитии эволюционизма. Эти доклады собрали 70, 41 и 82 чел., тогда как обычно заседания посещали 25—55 (и неизменно присутствовал Кольцов). Эволюционные взгляды Л. Плате, защитника идеи наследования приобретенных признаков, рассмотрел в своем докладе Г Г Фризен; теорию гетерогенезиса С. И. Коржинского — Промптов.
К группе исторических формально относился и доклад Ромашова об эволюционных воззрениях Морица Вагнера, которым 23.X. 1932 г. открылся семинар. Однако тема — эволюционная роль изоляции — к тому времени стала одной из ключевых проблем эволюционизма и играла выдающуюся роль в обсуждении ГАП’ов. Этой общей теме были посвящены доклады фризена о взглядах Вейсмана на проблему изоляции (в постановке Морица Вагнера), Дубинина об эволюционных взглядах Гагедурн (Хагедорнов) в связи с теорией ГАП’ов и ряд других (Фризен: «Эволюционные воззрения Романеса в связи с проблемой половой изоляции»; две оригинальные работы Промптова (19346, 1936): «Эволюционное значение миграций птиц» и «Об экологических факторах изоляции у птиц»; Кирпичников (1933): «Работы Гейнке и Йог. Шмидта в связи с проблемой разнообразия у рыб»; Ф. А. Бынов: «Работы русских ботаников (Веттштейна и Цингера) по проблеме экотипов»). Все эти доклады так или иначе касались механизмов и эволюционной роли ГАП’ов, но ставились и более общие темы.
Беспрецедентную аудиторию, 140 слушателей, собрал доклад Кольцова «Проблема прогресса в эволюции» (Кольцов, 1933) на 19-м заседании от 17.XI. 1933 г. В прениях выступал патриарх отечественного эволюционизма А. Н. Северцов (а также Б. С. Матвеев, Н. П. Дубинин, Б. В. Кедров-ский, М. В. Игнатьев, Э. С. Бауэр, С. Н. Боголюбский). С. А. Северцов прочел два доклада, посвященные динамике численности популяций в связи с борьбой за существование и роли борьбы в эволюции (11.III. 1933 г. и 5.IV. 1934 г.); доклады Б. С. Матвеева касались путей биологического прогресса и филэмбриогенезов, А. М. Лунца — современного понимания биогенетического закона. Критический разбор книги Гармса о проблемах перехода водных животных на сушу (Фризен) касался роли гормональных адаптаций в обеспечении эврибионтности видов. Доклад А. А. Машковцева (5.XII. 1933 г.) «Роль функций в онтогенетическом развитии» был посвящен проблеме замены онтогенетических факторов в онтогенезе; сейчас, по-видимому, лишь немногие знают, что его работы (Машковцев, 1936 и др.) в значительной мере предвосхитили гипотезу Кирпичникова о «косвенном» отборе и теорию «стабилизирующего» отбора Шмальгау-зена. Эту гипотезу Кирпичников (1935, 1944) изложил на заседании от 2.П.1935 г. («Роль ненаследственной изменчивости в процессе естественного отбора»), она обсуждалась на нескольких заседаниях Эволюционной бригады.
Темы более специального значения, помимо связанных с изоляцией
5 См.: «Эволюционный семинар при Генетическом отделе Института экспериментальной биологии. 1932—1933—1934—1935».—Материалы Комиссии.
153
и ГАП’ами, касались проблемы доминантности по Фишеру (Эфроимсон), эволюционной роли сцепления (И. Е. Трофимов), роли генетических и феногенетических механизмов в эволюции вида (Малиновский, 1939). Доклад Г Ф. Гаузе, посвященный опытам по борьбе за жизнь, на основе которых он поначалу пытался опровергнуть теорию Вито Вольтерра, вызвал очень живое критическое обсуждение, которое помогло автору занять более уравновешенную позицию (например, Гаузе, Витт, 1934), и, по мнению участников семинара, существенно облегчило работу над подготовкой к печати его широко известных теперь исследований.
Легко видеть, что линия, проводимая Ромашовым как руководителем Эволюционного семинара, продолжала как традицию реферативного кружка Соор Четверикова, так и тенденции Кольцова к выработке общего взгляда на широкий круг общебиологических проблем. Правда, у Ромашова был еще один образец: «Кружок имени Ламарка»6, созданный осенью 1920 г среди студентов, работавших в Зоомузее МГУ, по инициативе и под председательством Е. С. Смирнова. В 1921/1922 гг. Ромашов доложил там одну из первых своих работ — обобщение законов Менделя (Ромашов, 1925), а в 1920/1921 гг. реферировал только что опубликованный закон гомологических рядов Н. И. Вавилова. Там реферировались работы К. Иордана о различии между географической и негеографической изменчивостью; Гейнке и Семенова-Тян-Шанского по таксономии; Даке и Сушкина о необратимости эволюции; обсуждались представления о фенотипе и генотипе, монофилии и полифилии, о бергсоновском понимании жизни, о витализме системном и историческом, о задачах и методах зоогеографии; взгляды Сэмона, Штандфусса, Ламарка, Дарвина, де Фриза, Лотси, М. Вагнера; работы Каммерера, Вольтерека, Пшибрама, Р. Гертвига и Вейсмана, Иол-лоса, Житкова. Касаясь целей Кружка, Смирнов писал («Меморандум») «Известно, что предвзятое мнение и догматизм сыграли печальную роль в развитии биологических идей, и что не одна талантливая теория стала жертвой господствующих взглядов и пристрастной критики. Одна из целей Кружка — борьба с этими проявлениями рутины и косности и признание «законности» разных путей, ведущих к одной цели. С другой стороны, нужно признать, что значительная доля разногласий в оценке биологических фактов и теорий зависит от взаимного непонимания или даже нежелания понять противника.— Сопоставление различных теорий и их синтез — вот вторая основная цель Кружка». На деле заседания часто превращались в горячие споры убежденных дарвинистов (Ромашов) и не менее убежденных ламаркистов (Смирнов). Впрочем, это не мешало им оставаться друзьями.
Благодаря успеху Эволюционного семинара, а также ряду конкретных работ, в которых Ромашов, Балкашина и Промптов развивали традиции четвертиковской группы, эволюционно-генетическая линия исследований в ИЭБ показала в начале 30-х гг. свою жизнеспособность. В результате осенью 1934 г. в ИЭБ была создана самостоятельная исследовательская группа под руководством Ромашова — Эволюционная бригада (с зимы 1936/1937 гг. она получила статус лаборатории). Ее основной задачей стала «научно-исследовательская работа в области эволюционной гентики,
6 «Два года деятельности Кружка имени Ламарка». (Отчет за 1920/1921 и 1921/1922 гг. Список докладов. Меморандум. Выдержки из протокола организа ционного заседания от 29.IX.1922 г.— Материалы Комиссии.)
154
Эволюционная бригада ИЭБ, ок. 1934 г.: А. А. Малиновский, Д. В. Шаскольский, Д. Д. Ромашов
а также объединение работников разных лабораторий Института вокруг проблем эволюционной теории и дарвинизма» 7 При ней был организован Эволюционный коллоквий, призванный быть рабочим органом бригады для обсуждения работ — в отличие от Эволюционного семинара с его широкими программными докладами.
Сперва в бригаду вошли, кроме Ромашова, только Малиновский и Кирпичников (1934—1936 гг.), но в работе коллоквия участвовали также Е. И. Балкашина, Н. П. Дубинин, Е. Д. Ильина, Б. Ф. Кожевников, Л. В. Крушинский, Г В. Лопашов, К. А. Панина, А. Н. Промптов, И. Н. Свешникова, Н. Н. Соколов, Г Г Фризен, В. В. Хвостова и др. Осенью 1936 г. Ромашов после болезни вернулся в Институт, и с января 1937 г. бригада окончательно оформилась (до расформирования ее 15 июля 1940 г.) как Эволюционная лаборатория; в нее вошли также консультанты-математики В. И. Гливенко и Н. В. Смирнов и научные сотрудники Д. В. Шасколь-ский, В. Н. Беляева, Н. Д. Дусеева, К. А. Панина и позднее Е. А. Хох-лина. Основной задачей Эволюционной лаборатории стало «изучение генетического строения вида и его низших таксономических групп в связи со специфическими особенностями экологии вида, и анализ с позиций дарвинизма основных процессов, которые определяют эволюционную динамику и исторические изменения наследственной хромосомной основы вида»8. Работа шла по трем направлениям. Первое — исследования популяций разных видов
7 Д. Д. Ромашов. Краткий отчет о работе Эволюционной лаборатории (Эволюционной бригады) при Институте экспериментальной биологии с 1934 по 1940 гг.— Материалы Комиссии. В этой главе использованы: «Отчет о работе Эволюционного коллоквия (Дарвиновского семинара) при Институте экспериментальной биологии», 10 с. «Протоколы Эволюционной бригады», тетр. 1 и 2; рукопись В. Н. Беляевой «Дмитрий Дмитриевич Ромашов», 1972, 27 с. Этой теме был посвящен доклад В. С. Кирпичникова «Эволюционная бригада и Эволюционный коллоквий ИЭБ», прочитанный на конференции памяти С. С. Четверикова в г. Горьком в мае 1973 г. (см.: Бабков, 19736).
8 Д. Д. Ромашов. Краткий отчет, с. 4.
155
рода Drosophila (Ромашов, Беляева, Дусеева, Панина, при участии Дуби нина, Гептнер, Демидовой) — продолжало работу четвериковской группы, начатую в ИЭБ в 1925 г. Второе — изучение экологической и генетичес кой структуры низших таксономических единиц сазана Cyprinus carpio L. вели Кирпичников, Головинская, Хохлина. Третье — «математический анализ поведения хромосом в популяциях диких и домашних видов, а также анализ форм протекания основных процессов (естественный и искусственный отбор, мутационный процесс, генетико-автоматические процессы), определяющих генетическую структуру вида и ее эволюционные изменения» (Там же), составляло задачу математической группы при Лаборатории (Смирнов, Малиновский, Шаскольский, Ромашов, при участии директора Института математики МГУ Колмогорова, А. А. Ляпунова и др.) При математической группе работал математический коллоквий (20 заседаний), а в связи с экспериментальными разделами работ Лаборатории при ней был организован экологический реферативный коллоквий (15 заседаний).
На втором этапе работы Эволюционного коллоквия, с 18-го по 104-е заседания (28.X.1936 — 4.IV.1940), который с 84-го заседания (15.IX.1939) носил название «Дарвиновский семинар», в нем принимали участие многие сотрудники отделов ИЭБ, а также других научных учреждений Москвы и других городов. В этот период по широте охвата обсуждавшихся конкретных эволюционных задач, как и по разнообразию участников, Эволюционный коллоквий не имел равных в предшествующей (да и в последующей) истории отечественной биологии. Пожалуй, здесь уместно упомянуть о разнообразии организмов, популяции которых служили для выяснения эволюционно-генетических процессов. Это были, конечно, разные виды дрозофил; другие насекомые: тутовый шелкопряд, листовертки и др. бабочки; креветки и др. беспозвоночные; рыбы (карп, сазан); птицы (овсянки, турухтаны, домашние куры и др.); млекопитающие (собаки, песцы, соболи, лисицы и др.); растения — вика, флора солонцов и солончаков; разбирались геоботанические и флористические карты; речь шла об организации наследственного аппарата и филогении некоторых групп бактерий и т. п. Лабораторная генетика всегда имеет дело с одним или немногими главными объектами, в разные периоды разными; здесь же мы видим совершенно иную картину, которая заставляет нас вспомнить о натуралистической подготовке и устремлениях Ромашова, упрочившихся за время работы в Лаборатории Четверикова.
В этой связи интересен обмен письмами между Ромашовым и Промпто-вым в начале октября 1936 г., когда Ромашов возобновлял после полуторагодичного перерыва работу коллоквия. В это время Промптов работал в области «орнитологической экологии и зоогеографии, освещая ее общей генетической концепцией», и он отказался войти в бригаду: «...споры лишь показывали, несколько ортодоксальные генетики оторваны от живой природы. Различия взглядов в бригаде были необъятные...» Промптова, натуралиста по преимуществу, волновало уклонение Генетического отдела (к которому до 1937 г. относилась Эволюционная бригада) от четвериковской линии. Изучение хромосом из гигантских слюнных желез дрозофил, где колоссальная техническая работа подчас не оставляла времени для ее осмысления, его не устраивало. Математические работы, тогда только начинавшиеся в ИЭБ, были слишком абстрактными. «Я очень советовал бы Вам (говорил это не раз и раньше) использовать свой большой систематический опыт по жукам и перейти на них к конкретному изучению эко-156
лого-эволюционных положений (ГАП и др.), которые без этого висят в воздухе, точнее — находятся только в урнах и тиражах. ...Дмитрий Дмитриевич, я очень рад, что Вы вернулись в Москву и снова будете работать в Институте. Этим Вы несколько уравновесите чересчур «слюнную» тематику генетического отдела...»9. В дальнейшем на коллоквии обсуждались доклады по хромосомной изменчивости дрозофил (инверсии и транслокации обнаруживались в гигантских хромосомах слюнных желез), но в целом с возвращением Ромашова эволюционно-генетическая тематика ИЭБ стала более «четвериковской».
Отмечу, что жуками Ромашов занимался и позже — когда в середине 1953 г. он стал работать в Зоологическом отряде Комплексной экспедиции по полезащитному лесоразведению при Институте леса. Летом он проводил сборы жуков, чему помогали глубокое знание (с детства) повадок насекомых и изумительная интуиция. А зимой он обрабатывал материалы в Энтомологическом отделе Зоомузея МГУ. Когда в 1954 г. Биофак переехал на Ленинские горы, Ромашову отвели для его занятий отдельную комнату при Энтомологическом отделе — высокую, длинную, очень узкую, заставленную по сторонам шкафами с огромными коллекциями жуков (которые Ромашов передал в дар Музею) — «тараканья щель» по меткому определению М. Л. Бельговского. С коллекциями Ромашова работали систематики-энтомологи; среди его сборов Лопатин нашел и описал новый вид жука — Malegia romaschovi. С августа 1955 г. Ромашов вошел в Лабораторию радиационной генетики при Институте биофизики АН СССР, где развернул работы по радиационной цитогенетике рыб.
Приехавший в 1945 г. на празднование юбилея Академии наук Джулиан Хаксли писал в выходившей тогда на русском языке газете «Британский союзник» о советских исследованиях по теории эволюции, в частности, о работах Эволюционной бригады: «Чрезвычайно поучительно было после пяти лет почти полной изоляции узнать, что изучение проблемы эволюционной теории в Советском Союзе в основном развивалось параллельно с изучением тех же проблем в Соединенных Штатах Америки и Великобритании. В генетике с огромной сосредоточенностью проводится изучение мутаций, изменчивости и влияния отбора в природных популяциях. Укажу на работы профессора Дубинина и его учеников, профессора Ромашова, Берг, Оленова и других. По большей части работы в этой области основываются на строго выдержанной неодарвиновской теории естественного отбора. Профессор Дубинин и Тиняков, экспериментируя в годы войны, показали огромную роль естественного отбора в формировании хромосомных структур. Они обнаружили замечательный факт кариотипической дивергенции городских и сельских рас дрозофилы, экспериментально доказали влияние температуры на отбор инверсий и другое... Я убедился, что в области математической генетики и эволюции такими исследователями, как Колмогоров, Малиновский, Дубинин, Ромашов, Гливенко и другие, проделана большой важности работа, примерно в том же направлении, по которому идут независимо от них Р. А. Фишер в Англии и Севал Райт в Америке. За отсутствием времени я не имел возможности лично побеседовать по этим проблемам. Тем не менее я узнал интересный фткт: явление «дрейфа», связанное с ролью случайностей в эволюции небольших колоний,
9 Письмо А. Н. Промтова Д. Д. Ромашову, 15.X. 1936 г.— Материалы Комиссии.
157
сформулированное Севалом Райтом в 1931 г., было в том же 1931 г независимо от него открыто профессорами Дубининым и Ромашовым и названо Дубининым «генетико-автоматическими процессами». Сейчас, когда мир возвращается на нашу планету, когда ломаются изолирующие барьеры, мы можем предвидеть большой и быстрый рост науки об эволюции. В этих успехах советские ученые сыграют крупную роль» (Хэксли, 1945).
И. И. Шмальгаузен в 1939 г. так оценил работу Эволюционной бригады: «Работающая с конца 1934 г. Эволюционная бригада сыграла у нас в Союзе очень большую роль в привлечении внимания генетиков к эволюционным проблемам и к использованию методов генетики для их разрешения... В отличие от многих представителей «формальной генетики» члены Эволюционной бригады уделяли всегда большое внимание и таким проблемам, как вопрос о множественном выражении отдельных мутаций и вопрос о значении модификаций в эволюции, и не пытались противопоставлять генетику с ее мутационной теорией — классическому дарвинизму. Эволюционная бригада постоянно стремилась к глубокому теоретическому осмыслению новейших данных генетики, а также и вообще фактов экспериментальной биологии, на основе учения Дарвина. Работа Эволюционной бригады имела несомненное прогрессивное значение в развитии генетики у нас в Союзе» (цит. по: Беляева, 1972, с. 13—14).
СИСТЕМА основных понятий микроэволюции
В области популяционно-генетических исследований Тимофеев-Ресовский не только выполнял конкретные работы (например, N. W Timofeeff-Ressovsky, Е. A. Timofeeff-Ressovsky, 1941), но и стремился дать достаточно строгие определения основных понятий, происходивших из работы Четверикова 1926 г. Первую, довольно скромную попытку в этом направлении можно усмотреть в его выступлении на диспуте в Лондонском Королевском Обществе о современном состоянии теории естественного отбора (Timofeeff-Ressovsky, 1936а). А в статье размером в книгу «Генетика и эволюция. С точки зрения зоолога» (Timofeeff-Ressovsky, 1939) он изложил систему основных понятий микроэволюции. Аналогичную сводку «Мутации и географическая изменчивость» он опубликовал в книге «Новая систематика» под редакцией Дж. Хаксли (Timofeeff-Ressovsky, 1940а), и еще одну (существенно дополненную) совместно с Гансом Бауэром — в книге «Эволюция организмов» под редакцией Г Геберера (Bauer, Timofeeff-Ressovsky, 1943). Эти классические статьи были основаны на всей проделанной к тому времени популяционно-генетической работе; существенную часть из них составляли исследования отечественных авторов: в библиографии к сводке 1943 г., например, их больше 220. (Тимофеев-Ресовский, однако, не был знаком с циклами работ по феногенетике природной изменчивости (гл. 11) и по коррелированной изменчивости (гл. 12), для которых характерно изучение интегрированных генетических систем. Эти работы не вошли и в его более поздние сводки.)
Идейная часть дела изложена в статье «Микроэволюция. Элементарные явления, материал и факторы микроэволюционного процесса» (Тимофеев-Ресовский, 1958). Эта система понятий служила основой и для последующих обобщающих монографий, привлекавших очень интересный
158
и важный новый фактический маатериал (Тимофеев-Рессовский и др., 1969, 1973, 1979) 11
Статья 1958 г. «Микроэволюция...» посвящена «Дорогому учителю и другу Сергею Сергеевичу Четверикову». Было бы интересно выяснить, как соотносится работа учителя и ученика, в чем преемство и в чем различие. Сравнение в плане содержания мало что даст: мы увидим почти полное совпадение его в статьях 1926 г. и 1958 г. Различия начинаются, пожалуй, с заглавия: Четвериков не говорил о «микроэволюции»; Тимофеева-Рессовского иногда называют, со ссылкой на статью 1939 г., автором этого понятия и его пары — «макроэволюция». (Некоторые словари, впрочем, приписывают авторство Гольдшмидту (Goldschmidt, 1940), тогда как Филипченко (1929, Philiptschenко, 1927) в этом отношении вспоминают реже.) Вспоминая о необходимости различать изучение генезиса и изучение традиции, мы убедимся в том, что вопрос о приоритете не способен здесь прояснить дело. Действительно, по своему научному «онтогенезу» Филипченко относился (вместе с Морганом, Бэтсоном, Каслом, Конклином, Кэно и др.) к тем влиятельным эмбриологам, которые отводили генам несущественную роль в процессе индивидуального развития и полагали, что признаки надвидовых категорий определяются иными факторами («плазмоном» по Филипченко), а значит микроэволюция лежит вне поля зрения генетики (подробнее см.: Медведев, 1978, с. 29—32 и 82—86; Филипченко, 1977). Опираясь на свойства наследственных гомеозисов, Гольдшмидт строил «континуальную» модель хромосомы, отрицая реальность индивидуального гена, и развивал идею «счастливых уродов» (системных мутаций), обеспечивающих образование таксонов высокого ранга внезапными скачками. Тимофеев-Ресовский рассматривал видообразование как важнейший узловой момент эволюционного развития: это конечное звено в цепи процессов, изучаемых методами школы Четверикова, и начало «крупных» явлений и процессов, охваченных классическим эволюционизмом. Развивая тенденции статьи 1926 г., он полагал, что в основе филогенетических процессов любого ранга лежат в конечном счете те же самые стартовые механизмы, которые являются предметом учения о микроэволюции.
Четвериков, как мы видели, вообще избегал новых терминов (и любой формализации, в том числе математической). Считая, что ранняя формализация может стать тормозом на пути развития новой области биологического знания, он предпочитал обозначать вводимые понятия образными выражениями, достаточно определенными и в то же время дающими возможность широкого их понимания. Напротив, Тимофеев-Ресовский ищет строгие формулировки основных понятий области, таких как «популяция» (у Четверикова — «население вида») и т.п., он определяет критерии элементарных явлений, материала и факторов микроэволюции. Его система факторов эволюции включает мутационный процесс, популяционные волны, изоляцию, естественный отбор — те же моменты, что рассматривал Четвериков (с заменой образного «волны жизни» 10 11 12 более строго звучащим
10 «...Его сводка «Мутации и географическая изменчивость» в книге «Новая систематика» предвосхитила большую часть того, что Майр развивал более подробно в своей книге 1942 г...» (Glass, 1981, р. 646).
11 Статья «Волны жизни...» Четверикова (1905, переизд.: 1980) в монографии Тимофе-
ева-Ресовского 1939 г и последующих ошибочно датирована 1915 годом; дата была исправлена в его монографии 1973 г
159
термином). Характерный факт, что Тимофеев выбрал «волны» а не «дрейф генов», «ГАП’ы» или, скажем, «мальтузианский параметр», еще раз указывает на генезис его системы понятий.
Попытаемся понять различие тенденций Четверикова и Тимофеева Ресовского. Для того чтобы вывести биологическую мысль из тупиков, Четверикову было необходимо выйти за рамки каждого из сложившихся канонов, порвать с традицией. Это уклонение от традиции и составило, в известном смысле, содержание статьи 1926 г. Напротив, Тимофеев-Ресовский — на ином этапе решения той же задачи — считал должным привести в систему выработанные четвериковцами основные общие понятия новой области биологии.
Вспомним традиционное представление о двойственности человеческой деятельности, обязанной противоречивому единству двух начал: дионисий ского, поступательного, существо которого — в напоре и борьбе против вся кой грани, против номоса; и аполлинического, консервативного, деятельность которого — в мерности, в ограничении и наложении определений и границ. (Можно говорить о деятельности осуществления и деятельности осмысле ния, о требовании данности знания и требовании доказанности знания.) Не совмещенные, они противоречат друг другу; бесконечные в своем стрем лении оба начала требуют предельного своего утверждения. В ограничен ности одного другим не может осуществиться их полная гармония; это означало бы остановку развития. Но каждому узловому моменту развития науки свойственно некоторое моментальное равновесие двух начал, всякий раз иное. В начале пути систематизация была бы тормозом; но после короткого периода чрезвычайно бурного развития настало время его осмысления. Задача канонизации новой области исследований пришлась на долю Тимофеева-Ресовского.
Стремление к системе свойственно его научному творчеству в целом. Так, физико-химическая модель хромосомы Кольцова послужила ему стимулом для формулировки, совместно с М. Дельбрюком и П. Дираком, общебиологического принципа конвариантной редупликации. Можно было бы решить, что система понятий, сохранившая свои общие черты с 1939 г по 1979 г., начинает превращаться в догму. Однако попытку «разорвать застывающее время» сделал сам Тимофеев-Ресовский. В докладах на за седаниях МОИП в начале 70-х гг. он подчеркивал положение, что основ ной эволюционный феномен заключается в смене двух фаз: длительной фазы эволюционной инерции и короткой фазы радиальной перестройки популяции. В более поздних докладах он поставил под сомнение разделив шийся им прежде взгляд, согласно которому прогрессивная эволюция с необходимостью выводится из двух общебиологических принципов: естест венного отбора и конвариантной редупликации, и задался вопросом, каким образом следует определить эволюционный прогресс и какие типы прогрес сивной эволюции возможны. Ответ на этот вопрос должен, по мнению Тимофеева-Ресовского, составить третий общий принцип, формулирование которого необходимо для возможности построения теоретической биологии.
160
Глава 11
ФЕНОГЕНЕТИКА МАССОВОЙ ПОПУЛЯЦИОННОЙ ИЗМЕНЧИВОСТИ
Каждое дерзание — это новый риск, Новый шаг в темноту непознанного Со старым и все более устаревающим снаряжением
т. с. ЭЛИОТ
В цикле работ, о которых пойдет здесь речь, было сформулировано представление о двух основных типах популяционной изменчивости: изменчивость одного типа поддерживается мутационным давлением и ее уровень ограничивается нормализующим отбором; другая изменчивость поддерживается балансированным отбором, хотя в конечном счете возникает также посредством мутаций. Новое направление объединило подходы онтогенетики, генетики популяций, систематики, экологии и привело к выявлению закономерностей массовой изменчивости. Оно знаменовало перенос интересов с выявления редких «хороших> рецессивов на изучение нормальных видовых признаков, с проблемы образования генофонда популяции на проблему вычленения эволюционно значимой изменчивости. Тот же перенос интересов характерен для работ Н. К. Беляева по коррелированной изменчивости у шелкопряда. Для нынешних популяционистов в полной мере сохраняют свое значение конкретные выводы Ромашова и В. Н. Беляевой (которые были опубликованы лишь частично). Но если учесть нынешние тенденции в эволюционной теории, то откроется более глубокий смысл этих работ: для создания общего представления о процессе эволюций они важны прежде всего в плане постановки проблемы, указания подхода.
Цикл работ явился развитием представлений, возникших в ходе популяционного исследования 1925—1926 гг. Балкашина и Ромашов (1935) выяснили тогда, что основой фонда популяционной изменчивости служат «массовые изменения> со сложными картинами наследования (см. гл. 2 наст, книги). Эта категория изменчивости регистрировалась во всех последующих популяционных исследованиях (см. гл. 8, §3; гл. 9, § 1 наст, книги).
В протоколе 13-го заседания Эволюционной бригады от 17.1.1935 г., на котором обсуждался план докладов, среди дрозофильных тем значится и работа Балкашиной по феногенетике массовой изменчивости: “Crossveinless и plexus в популяциях 6 видов Dr.” («Протоколы ...>). Но весной 1935 г. Балкашина (тогда — старший научный сотрудник ИЭБ и по совместительству — Института прудового рыбного хозяйства) переехала в Чимкент, где работала энтомологом, затем заведующей Южно-Казахской областной противомалярийной станцией. Ее работы казахстанского периода были посвящены энтомологии, общей и прикладной. Кольцов 28.IV.35 г., дал ей следующую характеристику:
“Е. И. Балкашина является опытным хорошо подготовленным научным работником в различных областях биологии. По окончании ВУЗ’а она работала в области гидробиологии, затем занялась генетикой и опубликовала несколько интересных работ в области генетики насекомых, частью напечатанных и в заграничных журналах. Хорошо знает энтомологию. За последние годы много и очень интересно работала по генетике рыб,
11 В. В. Бабков
161
в частности по гибридизации культурного карпа с сазаном (диким). Ее последние работы в этой области имеют не только теоретическое значение. Ввиду этого я полагаю, что знания и способности Е. И. Балкашиной могут быть использованы в самых разных областях биологии, как теоретической так и прикладной».
В изучении «массовых изменений» ведшимся под руководством Ромашова участвовали Вера Николаевна Беляева, а также Нина Дмитриевна Дусеева и Ксения Александровна Панина.
МОДЕЛЬ: ИЗМЕНЧИВОСТЬ ЖИЛКОВАНИЯ
Для подробного анализа «массовой изменчивости» было выбрано жилкование крыла. Эта группа изменений широко распространена в популяциях разных видов Drosophila. Среди всех видимых изменений в популяциях D. phalerata на долю изменений жилкования приходится 57,4, а у D. transversa 61,5% (Балкашина, Ромашов, 1935). Такой же порядок величины имеет изменчивость жилкования у D. obscura, subobscura, funebris, melanogaster. При выборе этой группы признаков решающую роль сыграло обязанное четкости проявления и выражения признаков жилкования удобство их просмотра и классификации, позволяющее отмечать малейшие отклонения от нормы. Кроме того, как известно, этой группе признаков придают большое значение энтомологи-систематики: жилкование служит одним и основных признаков при установлении различий между систематическими группами двукрылых (ср.: Четвериков, 1926, с. 11).
В 1935—1940 гг. были обследованы пять популяций D. melanogaster, причем применялись три разных метода анализа. По 10 исходных самок, взятых в октябре 1936 г. в популяциях Алма-Аты, Самарканда и Еревана, разводились путем тесного инбридинга до марта 1941 г.; материал просматривался на 24—32 поколениях инбридинга. Для характеристики популяции Кутаиси просматривались мухи, пойманные в ноябре 1939 г. в природе (т. е. аберративный пролиморфизм), а также Fj (85 линий) и F2 (36 линий). В августе 1937 г. был собран материал в Уфе; здесь анализировались 122 линии, гомозиготные по II и III хромосомам, полученные от скрещивания диких самцов с самками из лабораторной линии, сбалансированной по деталям II и III хромосом.
По всем пяти популяциям было просмотрено около 92 000 мух. Результаты исследования обсуждались на заседаниях Эволюционной бригады («Протоколы...», тетр. 1 и 2): на 33-м заседании (3.III.1937 г.) —доклад К. А Паниной «Анализ мутационных изменений числа щетинок» (см.: Панина, 1938); на 77-м заседании (9.1.1939 г.) —доклад В. Н. Беляевой, Н. Д. Дусеевой, К. А. Паниной «Анализ Уфимской популяции D. melanogaster* (в «Кратком отчете...» указано, что их статья «Генетический анализ Уфимской популяции Drosophila fasciata* (1938) печатается в «Трудах Института цитологии, гистологии и эмбриологии Академии наук»); на 103-м заседании Дарвинского семинара (3.IV.1940 г.) доклад В. Н. Беляевой и К. А. Паниной «Малые морфологические мутации в популяциях дрозофил» (была подготовлена к печати в «Трудах ИЦГЭ» рукопись «Анализ мелких морфологических изменений в популяции Drosophila fasciata*, 1940). На этих материалах были защищены кандидатские диссертации: В. Н. Беляевой на тему «Изменчивость жилкования в природных популяциях D. melano-
162
Дмитрий Дмитриевич Ромашов, ок. 1959 г. gaster» в 1944 г. и К. А. Паниной — «Изучение изменчивости стерноплев-ральных щетинок у D. melanogaster» в 1946 г.
Главной задачей этого цикла исследований стало дать общую характеристику массовой изменчивости жилкования на широком ареале обитания D. melanogaster. Существенной чертой группы Ромашова было то, что закономерности этой изменчивости они пытались понять на основе феногенетических особенностей осуществления изменчивости в процессе индивидуального развития; это новое направление получило наименование «феногенетический анализ популяций». Крыло рассматривалось как единая реагирующая система и делались попытки выяснить, насколько своеобразны и независимы в своей изменчивости отдельные жилки, части этой системы.
В связи с реорганизацией ИЭБ, который в 1940 г. получил название «Институт цитологии, гистологии и эмбриологии», 15.VII.1940 г. была ликвидирована Эволюционная лаборатория. В начале Великой Отечественной войны Институт был эвакуирован в Алма-Ату. Ромашов оставался в Москве, и с конца 1941 г. он с В. Н. Беляевой (которая с начала войны работала лаборантом в Благушинской больнице) возобновил работу по изменчивости жилкования D. melanogaster. В анализ были включены данные сотрудников Четверикова по Геленджику (1926) г.), материалы экспедиций Дубинина и Ромашова по Кавказу (1933—1934 гг.), все сводки Эволюционной лаборатории (1937—1939 гг.).
Изменчивость жилкования в пяти популяциях D. melanogaster была исследована главным образом Беляевой (в основном до войны; опубликовано в 1946 г. и в диссертации 1944 г.; уфимская популяция описана ею по рукописи 1938 г.). Эти материалы, обобщенные в статье 1947 г. (Беляева, Ромашов, 1947), выявили замечательную особенность массовой изменчивости. Несмотря на происхождение мух из географически разобщенных популяций и несмотря на то, что применялись различные методы анализа (а, кроме того, три популяции харктеризовались на основе анализа потомства всего лишь десяти самок),— изменчивость жилкования во всех популяциях оказалась исключительно однородной: везде повторялись одни и те же закономерности. «Это говорит о том, что здесь мы имеем дело с явлениями, характерными для вида D. melanogaster в целом и имеющими для него существенное значение» (Беляева, Ромашов, 1947, с. 6). Эти закономерности сводятся к следующему:
1 Тождственный (однородный качественный) характер изменчивости по ареалу. Этот тип изменчивости является неотъемлемой частью общепопуляционной изменчивости вида и составляет 37,2—47,8%'от всех видимых изменений.
2. Высокая насыщенность популяций (% гетерозиготных самок) В среднем на одну самку приходилось 3,65 изменений.
И*
163
Вера Николаевна Беляева, ок. 1949 г.
3. Поля изменчивости. Изменчивость различных жилок резко различна: наиболее изменчивы обе поперечные, а также кубитальная жилки. Менее затронута изменчивостью радиальная2+3» почти стабильны радиальная4+5 и медиальная. «Поразительное сходство между популяциями в этом отношении позволяет говорить о существовании трех отдельных, своеобразных по своей изменчивости областей на крыле» (Беляева, 1946, с. 635) — см рис. 19. Эти подробно описываемые области, «поля» изменчивости Беляева трактует как лабильные, «слабые» места онтогенеза.
4. «Плюс» и «минус»-жилкование. Прибавления, отвилки, разветвления отдельных жилок, т. е. добавочное («плюс»-) жилкование основная часть фонда изменчивости. «Минус»-жилкование (нехватки, перерыв отсутствие жилок) встречалось значительно реже и составляло лишь- 78 *7 16 юб-щепопуляционной изменчивости жилкования. Беляева отметила в этой публикации, что «минус»-жилкование часто связано с резкими мутационными изменениями и что особенности двух типов жилкования дают основание предположить различие в их отборном значении.
5. Половой диморфизм. По всему материалу найдено 66,5% измененных самок и 33,5% измененных самцов. Степень выражения признаков у самок также обычно более резкая, чем у самцов.
Рис. 19. Поля изменчивости (по: Беляева, 1946)
Жилкование кртла Drosophila melanogaster
с — костальная жилка; sc — субкостальная; г( — радиальная 1; г? 3~ радиальная 2+3; г* —радиальная 4+5; т — медиальная; си — кубитальная; а—анальная; Л— гумеральная. / — средняя поперечная, // — задняя поперечная
164
В высокой пластичности полей и сложной генетической обусловленности их изменчивости Беляева видит основную причину того, что и в географически разобщенных популяциях и даже в потомстве отдельных самок, взятых из природы, «мы каждый раз встречаемся с определенной изменчивостью, протекающей по этим немногим и высоко лабильным «полям»» (Беляева, 1946, с. 635). Привлекая данные К. Уоддингтона по нормальному эмбриогенезу жилкования у D. melanogaster (Waddington, 1940а), она объясняет высокую стабильность radius4+5 и media тем, что они закладываются как первые жилки, связанные с образованием первичной системы кровяных лакун. Значительная же изменчивость трех «полей» на крыле связана с поздними, возможно вторичными, процессами развития, определяющими появление средней и задней поперечных жилок и radius2+3. Известную независимость в изменчивости каждого из трех пластичных полей она относила за счет того, что формирование входящих в различные поля жилок является, вероятно, результатом различных процессов онтогенеза (Беляева, 1946, с. 636).
ДВА ТИПА ИЗМЕНЧИВОСТИ
Ближайшим подлежащим разрешению был вопрос об отборном значении изменчивости жилкования. С этой целью Беляева и Ромашов (1946) предприняли анализ некоторых феногенетических особенностей каждого широко распространенного изменения. Они нашли, что: 1) гомозиготные и длительно инбридируемые линии весьма интенсивно «насыщаются» разными изменениями, преимущественно «плюс»-жилкования, причем насыщение затрагивает те же самые жилки и относительно в той же самой степени, которая характерна для природных популяций. (Этот факт авторы объясняли высокой мутабильностью по малым, в большинстве случаев полудоминантным, мутациям и «модификаторам», определяющим сложную генетическую природу изменений жилкования — Беляева, Ромашов, 1946, с. 730); 2) степень ограниченности полом для каждого изменения «плюс»-жилкования оказалась определенной величиной; 3) признаки «плюс»-жилкования проявили частичное доминирование в разной генотипической среде, т. е. при скрещиваниях с линиями-анализаторами; признаки «минус»-жилкования, как правило, рецессивны; 4) среди многообразия в характере наследования удалось наметить известную специфичность в наследовании отдельных изменений (она заключалась в характерной именно для данного признака устойчивости проявления при скрещивании с линиями-анализаторами и в постоянстве его сохранения в инбредных и гомозиготных линиях).
Эти результаты привели авторов к заключению, что изменчивость жилкования является существенной для вида и контролируется действием естественного отбора. Само расположение лабильных и стабильных областей в системе жилкования, как полагали авторы, отражает эволюцию крыла как органа полета: «В процессе эволюции крыло дифференцировалось на «стабильные области», на которых проявления изменчивости может иметь только отрицательное отборное значение, и «лабильные области», на которых изменчивость может иметь и положительное и отрицательное значение» (Там же, с. 731).
Беляева и Ромашов выделили две основные группы изменений — «плюс»-
165
и «минус»-жилкование, с резко различными свойствами. Изменения «минус»-жилкования встречаются в популяциях относительно редко; они, как правило, рецессивны и часто наследуются как четкие, хорошо расщепляющиеся, единичные мутации; половой диморфизм выражен слабо. Все это показывает, что «изменения группы «минус»-жилкования феногенетически, а по данным Waddington’a (1940а) и онтогенетически, в большинстве случаев проявляют себя как имеющие отрицательное отборное значение» (Беляева, Ромашов, 1946, с. 732). По мысли авторов, уровень этих изменений в популяциях поддерживается определенным соотношением давлений мутаций и отрицательного отбора.
Именно этот тип изменчивости и способ его поддержания анализировал Четвериков (1926), говоря о «процессе геновариационной изменчивости»; на основе его представлений В. П. Эфроимсон в 1932 г. оценил темпы мутационного процесса у человека. Этого же типа изменчивости касались работы начала 30-х гг., в которых была дана первая количественная оценка груза мутаций: «...в случае охвата наследственного содержания всего вида ...должно быть налицо своеобразное равновесие, возникающее как равнодействующая между давлением мутационного процесса и противодействием отрицательного отбора. Как совершенно справедливо указал В. П. Эфроимсон, для летальных генов это момент той концентрации, когда начинают возникать зиготы, гомозиготные по летальным генам» (Дубинин и др., 1934, с. 199).
Однако особенно важно, что Беляева и Ромашов разделили общую популяционную изменчивость на два типа и связали каждый не только с характером наследования, но и с феногенетическими особенностями их, с ответственными за их уровень факторами эволюции, наконец с их возможной эволюционной ролью.
Для группы «плюс»-жилкования рисуется совершенно иная картина: значительная ограниченность полом, частичная доминантность, неполное и колеблющееся проявление, сложность и многообразие в наследовании. Это позволяет говорить о возможном положительном значении такой изменчивости для вида. В этом отношении исключительно интересна установленная авторами прямая связь между широтой распространения в природе отдельных изменений «плюс»-жилкования и их феногенетическими особенностями (степень ограниченности полом, степень доминантности, интенсивность насыщения при длительном инбридинге), имеющими несомненное отборное значение. Давление мутационного процесса играет некоторую роль в создании такой картины «плюс»-жилкования; однако на первом плане в этом случае иной фактор. «Уровень каждого из этих изменений в популяциях прежде всего отражает «баланс селекционных факторов», т. е. известные соотношения между положительным и отрицательным отбором» (Беляева, Ромашов, 1946, с. 732), которые могут зависеть от времени года, локальных условий существования, быть разными у самок и самцов и т. п. «Именно здесь мы, по всей вероятности, встречаемся с явлениями так называемого «балансированного полиморфизма» (R. A. Fisher, 1927) ’, связанного с разнонаправленным действием естественного отбора» (Там же)
В итоговом докладе 1948 г. (Ромашов, Беляева, 1948) авторы ссылаются по поводу «балансированного полиморфизма» наряду с Фишером и на Форда (Ford, 1940, 1945).
166
На это представление о существовании в природных популяциях двух основных типов генетической изменчивости, изложенное в предвоенных докладах, опирались Гершензон (1941) и Дубинин (1948); его разделяют сейчас многие авторы (ср.: Dobzhansky, 1970, р. 129).
Сравнительное изучение изменчивости. Одним из насущных вопросов генетики популяций, по Ромашову, является сравнительное изучение абер-ративной изменчивости близкородственных видов, поскольку «подобное исследование может выявить сходство и различия в основных направлениях или «руслах» изменчивости близких видов и послужить ключом к пониманию тех главных процессов, которые определяют то или иное выражение внутривидовой изменчивости» (Ромашов, 1948, с. I). Для сравнительного изучения было естественно взять в качестве объекта хорошо знакомый московским генетикам вид D. funebris, обладающий рядом особенностей в фенотипическом проявлении своей изменчивости.
Ромашов (1948) анализировал изменчивость жилкования в популяциях этого вида по материалам Балкашиной и Ромашова 2, собранным в 1929— 1931 гг. в Москве (переулок Сивцев-Вражек), Петровско-Разумовском (под Москвой), Киеве и Ташкенте (было изучено F! и F2 от 60 взятых из природы самок), а также по материалам Беляевой и Ромашова 1944 г., собранным в Москве (Малая Почтовая ул.) и Петровско-Разумовском (анализировалось FJ. Для этой работы было просмотрено около 120 000 мух.
Анализ показал сходство в изменчивости всех изученных популяций и стойкое сохранение характеристик изменчивости по ареалу вида и в отдельных популяциях в разные годы — что, впрочем, не могло быть неожиданностью после изучения массовой изменчивости D. melanogaster. Основным отличием было резкое снижение изменчивости у D. funebris. Оно выражалось в уменьшении суммарной доли измененных мух в Fj и F2, а также в факте существования лишь двух лабильных «полей» (район 2-й поперечной жилки вместе с вершиной 5-й и район 2-й продольной жилки).
Различия в частоте «минус»-жилкования у двух видов невелики (в 1,5—3 раза). Напротив, частота «плюс»-жилкования у D. funebris ниже в 17—25 раз — это приводит к преобладанию «минус»-изменчивости. Причину неодинакового снижения изменчивости «плюс» и «минус»-жилко-вания Ромашов связывает с особенностями онтогенетического развития крыла D. funebris и выдвигает предположение, что «крыло D. funebris представляет более стойкую, слабее реагирующую систему. Развитие такой системы сравнительно труднее нарушается даже значительными сдвигами в виде хорошо выраженных мутаций, связанных с более ранними стадиями онтогенеза» (Ромашов, 1948, с. 6).
Интересно, что натуралистические и систематические устремления Ромашова явно видны и в этой небольшой статье. В ее последнем абзаце он
2 Эти материалы были доложены на IV Всесоюзном съезде зоологов, анатомов и гистологов, проходившем в Киеве 6—12 марта 1930 г. В списке докладов на секции генетики («Труды...», 1931, с. 337) мы считаем:
«...2. Балкашина Е. И. (Москва). Новые данные по изменчивости и наследственности у Drosophila funebris.
...18. Ромашов Д. Д. (Жаворонки, под Москвой). Генетический анализ московской популяции Drosophila funebris.
167
замечает: «Описанные в настоящей заметке характерные различия между D. funebris и D. melanogaster (типичными видами подрода Drosophila и подрода Sophophora) говорят о глубокой видовой специфичности изменчивости жилкования», которую он рассматривает как существенный элемент приспособительного полиморфизма популяций. «Дальнейший сравнительный анализ других представителей обоих подродов даст возможность сопоставить системы приспособительной изменчивости близких видов с их таксономическими взаимоотношениями и положением в системе» (Там же).
Тезис о связи «полей изменчивости» с особенностями онтогенеза предполагал интерес к изменчивости в условиях нарушения нормального онтогенеза, например в случае отдаленной гибридизации. Для изучения изменчивости гибридов В. Н. Беляева (1948) поставила три серии (с разными температурными режимами) скрещиваний: самки D. melanogaster (60 инбредных линий популяций Алма-Аты, Самарканда, Еревана) X самцы D. simulans (американская лабораторная линия). Просмотр 12 000 гибридных самцов выявил удивительное постоянство в относительной изменчивости отдельных жилок. Бросался в глаза параллелизм в изменчивости жилок, т. е. в расположении «полей» у гибридов и D. melanogaster В отношении модуса изменчивости гибриды сходствовали с D. funebris'. на долю «минус»-жилкования приходилось 70% изменений. («Возможно, что известное сходство в изменчивости гибридов с изменчивостью D. funebris отражает какие-то существенные особенности онтогенетического развития жилкования, связанные с происхождением и взаимным родством различных представителей рода Drosophila* — Беляева, 1948. с. 6.) У гибридов отмечалась также большая группа резких изменений типа мутации fused, затрагивающих сразу целый ряд жилок. В целом же изменчивость гибридов очень высока (и зависит от температуры развития и сложности жилкования и географического происхождения материнских линий); такой взрыв изменчивости обязан нарушению нормального развития в условиях отдаленной гибридизации.
Исследования Беляевой и Ромашовым популяций разных видов дрозофил показали, что поля изменчивости и другие установленные ими для D. melanogaster закономерности жилкования распространяются и на других представителей рода Drosophila, при том что они обнаружили и определенные межвидовые различия, в частности, «позволяющие наметить виды более изменчивые (D. melanogaster, D. transversa) и менее изменчивые (D. funebris, D. testacea, D. phalerata) (Ромашов, Беляева, 1947, с. 10) Это дало основание обратиться к другим семействам двукрылых, «чтобы проследить, насколько эти закономерности свойственны далеко отстоящим таксономическим группам» (Там же, с. 3). В условиях войны наиболее подходящим материалом для подробного анализа стала обычная комнатная муха, Musca domestica L. (Ромашов и Беляева, 1946). Летом и осенью 1944 и 1945 гг. было обследовано 13 популяций этого вида (в 1946 — еще 4) из Москвы, из Московской, Орловской и Калининской областей; просмотрено более 30 000 мух, взятых из природы. Анализ выявил существенные черты сходства в изменчивости жилкования у представителей столь отделенных семейств, как Muscidae и Drosophilidae.
Затем в анализ были включены другие группы мух. «С 1946 г. Д. Д. Ромашов снова становится сотрудником Генетического отдела Института ци-
168
тологии, гистологии и эмбриологии3 и расширяет свои исследования по изменчивости жилкования, включая в анализ других представителей Diptera. С ранней весны до поздней осени он проводит массовые сборы популяций различных мух в окрестностях Москвы. К маю 1948 г. им уже обследованы популяции 54 видов из девяти различных семейств Brachycera и просмотрено в общей сложности более 280 000 мух»,— говорится об этом в научной биографии Ромашова (Балкашина и др., 1975, с. 84—85). К итоговому докладу, прочитанному в начале 1948 г. на кафедре энтомологии МГУ, руководимой проф. Е. С. Смирновым, этот материал был разобран лишь частично. Литературные данные по этому вопросу были невелики (названы: «Reinig, Hering, Linder, Bodenstein и др.») и ограничивались, как правило, разрозненными указаниями, описаниями и рисунками отдельных изменений жилкования у представителей различных семейств двукрылых. «Единственное исключение, по-видимому, представляет работа Reinig’a (1935), в котором приведены довольно большие материалы по изменчивости жилкования в семействе Syrphidae и выдвигаются некоторые общие соображения о значении этой изменчивости. Однако больших сравнительных материалов с точным учетом изменчивости до сих пор не было собрано, и наши данные в значительной степени являются новыми» (Ромашов, Беляева, 1948, с. 3).
О масштабе работы говорит следующая таблица, включившая еще не все собранные данные (Там же) 4. «К настоящему времени нами
просмотрено:
по Drosophila melanogaster 7919+74098
> funebris 21576+92591
по остальным видам Drosophila 55961
по семейству Sepsidae 33295
по М use a domes Пса 42694
по остальным Muscidae 12097
по другим семействам Diptera— Brachycera —40000 Diptera— Nematocera 20000».
«Остальные виды» дрозофил — это D. testacea, phalerata, transversa, lit-toralis, buschkii, hydei\ в семействе Sepsidae проанализированы виды Themira nigricornis, Themira putris, Sepsis cvmmunis, Sepsis cympsea, no Muscidae - Musca autumnalis, Stomoxys calcitrans, Fannia canicularis. Коллекционные сборы были проведены по представителям различных семейств двукрылых — из группы Brachycera (Syrphidae, Chloropidae, Cordy-luridae, Larvivoridae и др ), из группы Nematocera (Tipulidae, Tricho-ceridae). Очевидно, что при подобном исследовании массового материала точное определение видов имело исключительное значение — «...систематика постоянно занимала в нашей работе важное место» (Там же, с. 4). Все просмотренные материалы показали, что изменчивость жилкования в различных семействах мух проявляется достаточно однообразно и закономерно.
Более поздние исследования (вполне независимые от работ Беляевой и Ромашова) дают основание считать, что выявленные ими закономерности жилкования не ограничиваются лишь отрядом двукрылых. Так, Л. М. Орлов
3 С 1942 г. Ромашов работал в Институте прудового рыбного хозяйства; подробнее см.: Головинская, 1975.
Левый столбец — аберративный полиморфизм, правый — Fi и F2.
12 В. В Бабков
169
(1975) проанализировал изменчивость жилкования в четырех популяциях одной из златоглазок (отр. сетчатокрылые) и выявил те же самые закономерности: поля изменчивости, асимметричное «плюс»- и «минус»-жилкова-ние, высокую насыщенность, однородный качественный характер изменчивости жилкования в различных популяциях. Совпадение особенно интересно, так как выводы Орлова были основаны на численно небольшом материале (по этой причине, вероятно, он не смог выявить половой диморфизм)
ИНТЕГРАЦИЯ ГЕНЕТИЧЕСКИХ СИСТЕМ
Самым интересным моментом этого цикла работ явилось установление определенных «полей» изменчивости жилкования мух (рис. 26) 5. Чрезвычайно характерен факт, что и в эксперименте изменчивость жилкования у D. melanogaster связана с теми же тремя «полями»: насыщение инбриди-руемых линий изменениями затрагивает те же жилки и в той же пропорции, как и в природных популяциях. Те же пропорции наблюдались в изменчивости гибридов D. melanogaster X D. simulans.
Для объяснения этих явлений было выдвинуто представление (Беляева, 1946, с. 635) о полях изменчивости как слабых, расшатанных или лабильных местах онтогенеза. (В тех же выражениях Оленов (1941, с. 158) писал о параллельной изменчивости у дрозофил: «...Существуют одни и те же слабые места в онтогенезе многих видов данного рода». — Ср.: Оленов, 1965.) Чрезвычайно широкое распространение отдельных, сравнительно немногих признаков (12 у D. melanogaster) по ареалу вида и качественно однородный характер изменчивости («...огромное и дифференцированное по отдельным популяциям генное содержание вида внешне выражается в одной и той же фенотипической картине изменчивости, одинаковой по ареалу» — Ромашов, Беляева, 1947, с. 18) Ромашов и Беляева объясняли именно пластичностью полей и многообразием причин, вызывающих изменчивость. С этой позиции становится понятным также замечательный факт, что характер изменчивости популяций точно отражается в потомстве небольшого числа взятых из природы самок, более того — в потомстве даже отдельных самок, если это потомство достаточно велико. Эти представления позволяют понять явление насыщения гомозиготных линий и подойти к объяснению крайне сложных, «непредсказуемых» (Gordon et al., 1939) картин наследования признаков жилкования.
Тщательное исследование нормального эмбриогенеза крыла дрозофилы (Waddington, 1940а) проливает свет на эту группу фактов. Первыми, на ранней предкуколочной стадии, закладываются в виде первичной системы кровяных лакун III и IV продольные жилки; тотчас вслед за ними появляется V продольная. Только спустя 20—24 ч возникают II продольная и обе поперечные жилки, каждая совершенно самостоятельно. Первая попереч
5 Представление о направленной и ограниченной изменчивости в пределах рода Drosophila Ромашов и Балкашина высказывали в ходе первого генетического анализа природных популяций (гл. 2 наст, книги). С большими подробностями эти взгляды развивались в работах Беляевой, Дусеевой, Паниной (1939), Беляевой и Паниной (1940) и особенно в диссертации Беляевой (19944). Близкие взгляды формулировали и другие авторы, говоря о «градиентах чувствительности» (Берг, 1945) и развивая представления о «трансформации нормального генотипа» (Оленов, 1941).
170
ная жилка (место соединения III и IV продольных) приобретает самостоятельность после сокращения и расхождения этих двух жилок. Вторая (остаток центральной полости крылового мешка) образуется на конечной стадии сокращения этой полости. Последняя по времени формирования — II продольная жилка развивается путем слияния небольших полостей в губчатой ткани крыла. Картина нормального эмбриогенеза крыла позволяет понять высокую стабильность одних областей крыла (III и IV продольные жилки, которые закладываются на ранних стадиях) и пластичность других (обе поперечные и II продольная, формирующиеся позже).
Материалы Ромашова и Беляевой позволили пойти дальше и утверждать, что в пределах отдельных полей изменчивость жилкования определенно направлена и ориентирована (т. е. имеет характерный модус). Так, у Musca domestica во всех 17 изученных популяциях, среди отвилков от II поперечной жилки, 70% ориентированы к основанию и 30% — к вершине; у Thermia nigricornis небольшой отвилок всегда расположен на одном и том же месте II продольной жилки и ориентирован перпендикулярно в сторону III продольной, и т. д. и т. п. Факты эти позволяют считать, что «каждое «поло внутри себя представляет единую систему, закономерно изменяющуюся в определенных направлениях и обусловленную формированием этого поля в процессе онтогенеза» (Ромашов, Беляева, 1947).
Этот ход рассуждений полностью приложим к другим массовым изменениям, представляющим как бы промежуточную ступень между мутантными и нормальными видовыми признаками, хорошо известным всем, работающим по генетике дрозофилы: вырезки на крыле; типа divergence; хетотаксия, грубые глаза; abnormal abdomen и т. д. Каждое из них широко распространено по ареалу дрозофил и показывает слабую дифференциацию по отдельным разобщенным популяциям; они обычно ограничены полом, неполно доминируют, обладают непостоянным проявлением, их наследование сложно и непредсказуемо.
Описанные картины изменчивости Ромашова и Беляева трактовали также и с точки зрения их отборного значения, выдвинув предположение, что в процессе эволюции крыло дифференцировалось на стабильные и лабильные участки. Уоддингтон (Waddington, 1940а) исследовал проявление в онтогенезе D. melanogaster мутаций, вызывающих отсутствие и перерывы жилок, и нашел, что действие этих генов четко выражено и строго фиксировано на очень ранних стадиях развития крыла; с ним связано нарушение процесса детерминации крыла. Тем временем мутации «плюс»-жилкования не нарушают нормального развития крыла; они возникают в виде флуктуирующих вариаций вокруг нормально детерминированного рисунка жилкования и связаны с изменением условий окончательной дифференциации отдельных жилок. Концентрацию генов «минус»-жилкования в популяции Ромашов и Беляева ставят в зависимость от соотношения давлений мутаций и отрицательного отбора, а «плюс»-жилкования — в зависимость от «баланса селекционных факторов», т. е. известных соотношений между положительным и отрицательным естественным отбором. «Однако трудно предположить, что само наличие или отсутствие на той или иной жилке крыла небольшого, часто асимметричного отрезка, может иметь непосредственное отборное значение. Возможно, что эти уклонения служат только некоторым показателем определенных физиологических условий развития крыла»,— отметили Ромашов и Беляева (1948, с. 18), привлекая внимание к соображениям И. А. Рапопорта (1942—1943), который изучал действие мутации
171
Met (метаплазия) у D. melanogaster. Придя к выводу о способности к регуляции структуры крыла, сн писал (цитирую по итоговому обширному очерку «Феногенетический анализ независимой и зависимой дифференцировки»): «Значение, придаваемое в систематике насекомых мелким и мельчайшим деталям жилок крыла, ориентировано не на чисто формальные, второстепенные морфологические отличия, лишь коррелированные с более важными физиологическими особенностями, как это указывается некоторыми систематиками. В данном случае внимание систематиков направлено на важные в биологическом отношении критерии. Даже мелкие сдвиги структуры жилок, вроде описанных в этом разделе, являются верными индикаторами изменений массы клеток и параметров строения органа, остающихся незаметными для наблюдателя, но играющих огромную роль в полете. Система жилок представляет функциональный, а не формальный критерий».
Наконец, существенной особенностью изменчивости жилкования является ее видовая специфичность, настолько глубокая, что «Таксономически мы можем различать виды не только по их морфологическим и физиологическим признакам, но и по характеру их аберративной изменчивости» (Ромашов, Беляева, 1948, с. 20). Эта аберративная изменчивость приспособительна в том смысле, что она отражает существующие внутри вида балансированные селекционные системы — основу приспособительной генотипической изменчивости вида. «На известных сдвигах в этих системах могут строиться сезонная, экологическая и географическая приспособительная изменчивость вида. И, наконец, при коренной перестройке системы возникают более глубокие качественные различия, связанные с трансформацией и возникновением новых видов» (Там же).
Н. П. Дубинин провел аналогичное исследование природных популяций D. melanogaster (Дубинин, 1948). Он сосредоточил внимание на трех формах добавочного жилкования: «свободная жилка», «отвилка-2», «сеть жилок на крыле». (Эти изменения связаны друг с другом: в природе комбинация «свободной жилки» и «отвилки-2» у одной мухи встретилась в 200 раз чаще, чем в предположении их полной независимости.) Его исследование подтвердило выводы Ромашова и Беляевой, на результаты которых он постоянно ссылался в обширной статье 1948 г. За 30 поколений инбридинга из 240 линий, основанных от диких самок из алмаатинской популяции, добавочное жилкование проявили 163 (67,9%), тогда как в природе аберративный полиморфизм проявлялся лишь в ничтожной степени (0,57%). В линиях из пяти географически разобщенных популяций с разными экологическими условиями (Кутаиси, Москва, Иваново, Воронеж, Сталинабад) за 15 поколений инбридинга добавочное жилкование появилось в 75,7% случаев (в 112 линиях из 148). Это показало, что высокая насыщенность однородной гетерогенной наследственностью не является особенностью лишь алмаатинской популяции, но свойственна всем популяциям по ареалу вида, как показала Беляева (1946).
Факт поддержания в популяциях постоянной гетерозиготности без специальных приспособлений типа систем сбалансированных деталей привел Дубинина к предположению о селективном преимуществе гетерозигот и к попытке уловить действие отбора в опыте. Линия Су с инверсиями была скрещена с контрольной нормальной культурой (N) и тремя разными линиями «сеть жилок» (Е). Их потомки скрещивались с мухами NN' в одном случае (где N' обозначает дикую хромосому, также свободную от
172
комплекса добавочного жилкования) и с мухами NN в другом. Распределение полученного потомства (Дубинин, 1948) было следующим:
CyN xNN' CyExNN
CyN 51,9% 47,9%
NN 48,1% _
EN — 52,1%
Различие между двумя случаями было статистически достоверным и оно показывало, что особи, гетерозиготные по изменению «сеть жилок», превосходят по жизнеспособности контрольных нормальных особей, также полученных при гибридизации разных линий. Четкая ограниченность полом, а также намеки на сезонный цикл концентраций изменений дали указания на приспособительный характер полиморфизма по добавочному жилкованию (ср.: Беляева, Ромашов, 1946). Отбор в инбридируемых линиях приводил к их «насыщению» жилкованием; при этом выражение признака могло достигать высшей степени в форме «сети жилок»; в некоторых линиях достигалось 100%-ное проявление, тогда как другие стабилизировались на разных уровнях проявления и выражения.
Ход выщепления «сеть жилок» по поколениям отбора показал, что это изменение имеет сложную наследственную природу и «требуется известная консолидация наследственной системы» (Дубинин, 1948, с. 221—222), чтобы довести ее до уровня проявления признака в форме «сети жилок» на крыле. Проведя локализацию полигенов, отвечающих за добавочное жилкование, Дубинин заключил, что они собраны на II хромосоме около двух центров — net (0,0) и blistered (107,3).
В результате Дубинин выделил три главные особенности генетического поведения добавочного жилкования (Там же, с. 235): 1) повсеместное распространение наследственной основы по ареалу разных видов дрозофил; 2) гетерогенность полигенных систем, отвечающих за изменчивость проявления и выражения признака и неаллелизма линий из различных популяций; 3) управление расщепляющимся и комбинирующимся блоком полигенов, организованных в пределах II хромосомы.
Два первых пункта повторяют выводы Ромашова и Беляевой, материалы Дубинина дают им дополнительное подтверждение. По поводу третьего Р. Гольдшмидт заметил, что Дубинин не распознал расщепляющихся мутаций net и blistered и что в остальном его интерпретация необоснованна, и предложил свое истолкование этих материалов на основе ряда специфичных плейотропно действующих больших мутаций (Goldschmidt, 1953). И. М. Лернер возразил, что если даже и принять утверждение Гольдшмидта, согласно которому материал Дубинина состоит из носителей ранее описанных и локализованных мутаций, все же эти основные положения не утрачивают силу. Гольдшмидт не отрицал существования модификаторов пенетрантности и доминантности, так что если и принять его истолкование, то нужно лишв искать системы полигенов не в мутациях, непосредственно затрагивающих жилкование, как у Дубинина, а в модификаторах (Lerner, 1954, ch. 8), что ближе к позиции Ромашова и Беляевой.
Вообще же идеи русских авторов, проливающие свет на механизм поддержания фенодевиантов в природных популяциях, привлекли пристальное внимание Лернера при написании его книги «Генетический гомеостаз» (Lerner, 1954). Не зная о работах Ромашова и Беляевой, Лернер, к счатью, мог воспользоваться переводом упоминавшейся фундаментальной
173
сводки Дубинина. Обсуждению данных по изменчивости жилкования Лернер посвятил целиком гл. 8, и почти полностью привел в гл. 15 перевод раздела «Обсуждение» (Дубинин, 1948, с. 235—239), который содержал изложение всех основных положений Беляевой и Ромашова.
Феногенетический анализ популяций, основопологающие идеи которого зародились в ходе первого экспериментального исследования свободно живущих популяций дрозофил, стал конкретным случаем объединения подходов онтогенетики, генетики популяций, экологии, систематики. Эта линия работ поставила в центр внимания нормальные видовые признаки («массовая популяционная изменчивость»)— вместо обычно изучавшихся резких наследственных уклонений («хороших мутаций»). На этой основе сформулировано представление о существовании двух типов природной изменчивости: один тип изменчивости, образуемый обычно «хорошими» рецессивами, поддерживается повторным мутированием и ее уровень определяется соотношением давления мутаций и отбора (согласно первоначальной модели); второй тип изменчивости обязан полимерным полудоминантам с изменчивым проявлением, ее уровень отражает соотношения между положительным и отрицательным отбором, которые зависят от сезона, локальных условий жизни, внутренних условий организма и пр. (новая модель). Первоначальная модель генетической эволюции популяций, основанная на идее отбора четких мутаций, была пересмотрена в свете этих работ и вошла в качестве частного случая в новую, более адекватную действительности модель. Связанной с первоначальной моделью проблеме образования генофонда популяций пришла на смену проблема выявления эволюционно значимой изменчивости. На этих путях в ходе работ по феногенетическому анализу природных популяций выявились новые смысловые глубины в общей проблеме Четверикова.
Глава 12
КОРРЕЛИРОВАННАЯ ИЗМЕНЧИВОСТЬ КОНСТИТУЦИОННЫХ признаков
...Но живут, живут в N измереньях Вихри воль, циклоны мыслей, те, Кем смешны мы с нашим детским зреньем, С нашим шагом по одной черте!
в. я. БРЮСОВ
Новый аспект центральной проблемы эволюционной генетики — становление адаптивных расовых различий — оформился в работах Н. К. Беляева по генетике и селекции тутового шелкопряда 1934—1936 гг.
Беляев принимал участие в первом экспериментальном исследовании наследственного содержания диких популяций дрозофил. В одной из его ранних статей описана мутация «alae incisae» — первый случай сцепленной с полом мутации у D. transversa\ в другой — локализованная в точковой IV хромосоме D. melanogaster мутация (нехватка) «abdomen rotatum», которую особо отметил в отчете 1929 г. Четвериков (1982). Все остальные статьи Беляева посвящены бабочкам. Вообще же биологией он занялся благодаря своему интересу к бабочкам, которых собирал и разводил еще в детские годы. В Московском Университете он скоро сблизился с Четвериковым, бывшим в то время, пожалуй, нашим лучшим лепидоптерологом-
174
систематиком и обладавшим самой богатой коллекцией бабочек (в начале 50-х гг. Четвериков передал ее в дар Зоомузею АН СССР). Когда же Лаборатория Четверикова приступила к разработке феногенетики дрозофилы, обращаясь к анализу неполного и асимметричного проявления признаков, Беляев занялся расшифровкой онтонегетических механизмов природной изменчивости гусениц и куколок некоторых видов бабочек.
Лаборатория Четверикова отдала дань и цитогенетическим исследованиям. Н. К. Беляеву в области кариосистематики принадлежит монографическое исследование отряда Lepidoptera в связи с его филогенией (Ве-liajeff, 1930; Беляев, 1930), которое было одной из первых работ по хромосомному полимофизму у животных. В 1927—1929 гг. он изучил кариотипы 38 видов бабочек (из них 31—впервые), принадлежащих к 16 семействам; вместе с литературными данными его материал охватывал 94 вида из 22 семейств. В отношении формы хромосом этот отряд оказался чрезвычайно однородным: хромосомы — шарикообразные; а наличие перетяжки в единственном случае обнаружения палочковидной хромосомы указывало на то, что она находится на пути распадения на два шарикообразных элемента. Числа хромосом (гаплоидные) варьировали от 11 до 87; при этом 73,4% видов (69 из 94) имели числа, близкие к 30 (от 28 до 31). Характерное гаплоидное число 30, ряд морфологических и биологических сходств, факт гетерогаметности женского пола (ставящий бабочек и ручейников особняком среди других насекомых) — все это, по мнению Беляева, «не только подтверждает гипотезу происхождения Lepidoptera от Trichoptera-подобного предка, но и с несомненностью показывает, что число, близкое к 30, является для бабочек филогенетически первичным числом» (Beliajeff, 1930, S. 386—387). Наиболее вероятно, что все измененные числа произошли «путем фрагментации или ассоциации, путем внутренней передифференцировки, перераспределения хроматинового вещества» (Там же, с. 388). За это говорит наблюдение Беляева, что видам с большим числом хромосом свойственны хромосомы маленького размера, и наоборот; эту точку зрения подтверждают и литературные данные. Распределение числа хромосом по семействам показало, во-первых, наличие модальных чисел (28—31) во всех семействах, и, во-вторых, сильную вариацию чисел внутри семейств (вплоть до сем. Lymantriidae с числами от 11 до 87) ; отсюда следует, что «таксономические подразделения внутри отряда Lepidoptera не находят никакого отражения в строении кариотипов» (Там же, с. 392). Общий вывод: «в отряде Lepidoptera в течение филогенеза изменения' кариотипа не имели никакого определенного направления» (Там же, с. 395). Все это относится, конечно, только к данному отряду. (Вопрос о значении кариотипа для систематики должен решаться, по мнению Беляева, не в общем виде, а для каждой группы организмов отдельно. Так, в группе Gymnospermae число хромосом 12 есть признак класса; число около 30 у бабочек — признак отряда; в отряде Orthoptera число хромосом является признаком семейств; у Ornithogalum — признак родовой; в роде Carex число хромосом — видовой признак, а у бабочек Biston и Phragmatobia — это признак географических рас.)
На основе своих результатов Беляев присоединяется к позиции Э. Вильсона, что число хромосом per se является делом второстепенного значения, и к мнению Моргана, Стертеванта, Бриджеса, что для генетики сравнивание групп хромосом имеет небольшое значение, так как для нее имеют смысл не формы и размеры хромосом, но содержащиеся в них гены: 175
Николай Константинович Беляев Снимок Б. Л. Астаурова, ок. 1923 г
«Если подходить к проблеме эволюции с точки зрения современной генетики, то, вероятно, следует отдать больше предпочтения не наружным изменениям хромосом или хромосомных наборов, а их внутренним изменениям, основанным на мутациях или геновари-ациях» (там, с. 396).
Заключения Беляева «в ряде отношений предварили робертсоновскую концепцию эволюционной перестройки хромосомного набора Хотя теперь хромосомные чис-
ла изучены более чем у 1000 видов бабочек, основные выводы Н. К. Беляева полностью сохранили свое значение» (Астауров и др., 1975, с. 109). Так, например, в главе «Эволюция хромосом у чешуекрылых» сводки «Хромосомы сегодня» Эско Суомалайнен остановился на всех тех же вопросах, и пришел к тем же выводам, что и Беляев четырьмя десятилетиями раньше (см.: Suomalainen, 1969). Единственным новым (по сравнению со статьей 1930 г.) фактом стал тот, что содержание ДНК у близкородственных видов дневных бабочек почти одинаковое; отсюда был сделан вывод, что наблюдаемые большие различия в числе хромосом не могут быть обязаны полиплоидии, но являются следствием структурных перестроек, особенно «фрагментаций» и «слияний», которые облегчены при диффузном кинетохоре,— вывод, вполне совпадающий с беляевским. Можно еще упомянуть об энтомологической экспедиции 1926 г. в Лапландию, предпринятой вместе с Четвериковым и Астауровым (материалов о ней, по-видимому, не сохранилось). Беляев и впоследствии собирал данные по хромосомным числам бабочек; они остались неопубликованными. А его последняя по времени опубликования работа была посвящена генетическому анализу неполно-доминантного признака меланистической окраски бабочек тутового шелкопряда (Беляев, 1937). Этот качественный признак, как установил Беляев, определяется двумя независимыми группами полимерных генов, обусловливающих общую интенсивность окраски и распределение пигмента в рисунок, причем ему удалось приблизительно определить число генов и локализовать ген основного действия. По-видимому, результаты этой работы могли бы послужить ему отправной точкой для обсуждения эволюционно-генетических аспектов мимикрии.
В 1929 г. Н. К. Беляев по совету Н. К- Кольцова переехал в Ташкент (затем в Тифлис) и занялся шелководством. (С 1930 г. на тутовый шелкопряд переключился также Астауров, а в 1936—1948 гг. дубовым шелкопрядом занимался их учитель Четвериков, который вывел пригодную для среднерусской полосы одноурожайную породу.) Быстрое развитие генетики и племенного разведения шелкопряда в начале 30-х гг. в СССР опиралось на сильную отечественную традицию научного шелководства. Достаточно
176
отметить вклад Кавказской станции шелководства (затем Закавказский (Тбилисский) НИИ шелководства и шелковой промышленности, ныне факультет шелководства Тбилисского сельскохозяйственного института), Московского общества шелководов, руководимого А. А. Тихомировым, и Центральной шелководственной станции в Москве. Большую роль в развитии советского шелководства вообще и генетики шелкопряда в частности сыграл Н. К. Кольцов, объединивший в 1929 г. работу в этой области под эгидой Генетического отдела КЕПС, а затем Комиссии по шелководству ВАСХНИЛ (см.: Астауров и др., 1975; Астауров, Рокицкий, 1975; Астауров, 1978).
Работы Беляева имели большое практическое значение; они, например, сыграли ведущую роль в переводе шелководства в СССР на промышленную гибридизацию (Генетика и селекция..., 1936). Однако нам интересны вытекавшие из этих работ следствия для эволюционной генетики, хотя с позиций практических задач они должны были бы рассматриваться как побочный результат.
Тутовый шелкопряд Bombyx mori L., ставший вторым после дрозофилы главным объектом генетических исследований, имеет ряд особенностей; наиболее существенные из них связаны с существованием конституций пород шелкопряда.
КОНСТИТУЦИИ ПОРОД ШЕЛКОПРЯДА
За пять тысяч лет одомашнивания при исключительно интенсивном отборе на локальную приспособленность и качество шелка предок культурных пород Bombyx mori var. mandarina распался на огромное число пород, отличающихся друг от друга не только четко менделирующими признаками (окраска крови и кожи гусениц, яйца, кокона, бабочки и т. п.), но и по конституциональным признакам — темпам развития, размерам, а главное — по вольтинности, т. е. по числу генераций за кормовой сезон. Моновольтинные породы, пройдя весной стадию гусеницы, куколки, бабочки, откладывают к концу весны яйца, прерывающие развитие до весны следующего года (диа-паузирующие). Бивольтинные породы в конце весны откладывают яйца, из которых развивается второе, летнее, поколение; яйца второго поколения диа-паузируют до следующей весны. Поливольтинные породы дают до восьми генераций в год, и лишь последнее поколение откладывает яйца с короткой диапаузой (периодом покоя), приходящейся на холодный период дождей. Породы моновольтинные характеризуются (по сравнению с би- и тем более поливольтинными) большими размерами яйца, гусеницы, куколки, бабочки, большим весом кокона и его оболочки, большей длиной шелковой нити, большей продолжительностью развития, а также повышенными требованиями к условиям развития, малой устойчивостью к неблагоприятным условиям (жаре и болезням), вообще пониженной жизнеспособностью. По всем этим признакам группы моно-, би- и поливольтинных пород образуют градиент конституций, причем все указанные признаки сопряжены с вольтинностью.
Итак, основной адаптацией, обеспечивающей эврибионтность тутового шелкопряда (которая проявляется в образовании географических рас и многочисленных культурных пород по всему ареалу тутового дерева), является способность к регуляции вольтинности. Закономерный ряд, образованный породами шелкопряда по коррелированному комплексу важнейших призна
177
ков, сопряженному с вольтинностью, тянется от Палеарктики через субтропики и тропики до экваториальной зоны Старого Света. Он начинается на севере ареала нежными, крупными, медленно развивающимися породами, дающими лишь одну генерацию в наиболее благоприятное время; продолжается би-, три-, тетравольтинными породами и заканчивается на юге ареала подивольтинными, мелкими и бедными шелком, неприхотливыми породами, дающими до восьми поколений в год.
Развитие шелкопряда идет следующим образом 1 К концу двух-трех суток после откладывания грена моновольтинных пород временно прекращает развитие и вступает в стадию покоя — эстивацию (летнюю диапаузу), которая длится около полугода при 15—25° С и приходится на лето — начало осени. Затем грена зимует, 90—100 дней при 2—5° С. Начинает оживать грена после нескольких дней инкубации при 10—12°, когда развертываются почки шелковицы.
Моновольтинные яйца можно методами «искусственного оживления» заставить развиваться без диапаузы, но в летнюю жару вторая генерация моновольтинных пород гибнет в массе. Поэтому летом и осенью используют гибриды между богатыми шелком моновольтинными и более выносливыми и скороспелыми бивольтинными породами.
Стандартная коробка грены равна 25 г и содержит обычно около 35—36 тыс. яиц. Эквивалентом считается партия свежевылупившихся гусениц весом 17,5 г. Гусеницы кормятся пять возрастов, прерываемых четырьмя снами (линьками); за 35—45 дней выкормки их вес увеличивается в 10—15 тыс. раз; 25 г грены требует 850—1000 кг листа, т. е. продукцию 30—40 высокоствольных тутовых деревьев или 300—400 карликовых; убыль на каждом возрасте больше 10%. На 8—9-й день V возраста начинается завивка кокона, в нем развивается куколка и на 13—17-й день выходит бабочка.
Выбор диапаузного поведения. Бивольтинные породы шелкопряда дают, как правило, известный процент диапаузирующих («моновольтинных») яиц. Для яйца выбор пути развития — по моновольтинному типу (с диапаузой) или по бивольтинному (без нее) — предопределен еще до момента его оплодотворения и зависит от генотипа и условий развития яйца предыдущей генерации (матери). Внешние факторы развития матери, детерминирующие бездиапаузное развитие ее грены, таковы: низкая температура на эмбриональной стадии (ЭС); низкая влажность на ЭС; темнота или слабое непродолжительное освещение на ЭС; изобилие пищи; высокая температура на стадии завивки кокона и на стадии куколки. Этот комплекс условий характерен для нормального весеннего развития.
Противоположные условия развития — высокие температура и влажность, продолжительное (13—17 ч) освещение расеянным дневным светом, нехватка пищи на ЭС, понижение температуры на стадии завивки кокона и на стадии куколки, заставляющие самку бивольтинной породы откладывать зимующую грену, характерны для лета — начала осени. Очевидно, что такие факторы регулируют вольтинность целесообразно и адекватно среде, но, что следует особенно подчеркнуть, регулируют этот выбор для каждого отдельного шага, без наследственного закрепления бездиапаузного или диапаузного развития.
1 Приводимые здесь данные относятся не к современным породам, а к тем, что были доступны Н. К. Беляеву в 1930-е гг. После 1945 г. в практику мирового шелководства были включены многие лучшие японские породы, что изменило общую картину.
178
ГЕНЕТИЧЕСКИ ОБУСЛОВЛЕННЫЕ КОРРЕЛИРОВАННЫЕ СДВИГИ
Серия экспериментов по отбору на скорость развития, которая дала Беляеву основание для новой постановки основной проблемы эволюционной генетики, планировалась с чисто практической целью выведения таких пород, которые совмещали бы преимущества обоих типов — высокую шел-коносность моновольтинных и быстрое развитие и устойчивость бивольтинных. Интересующие нас материалы были обобщены в докторской диссертации Н. К. Беляева «Проблемы генетики и селекции тутового шелкопряда» (г. Тифлис, 1936 г.), которая не была опубликована. Однако существенная часть этих материалов воспроизведена в докторской диссертации В. П. Эфроимсона «Проблемы генетики, селекции и гибридизации тутового шелкопряда» (г. Харьков, 1946 г., 661 с.), по которой я их цитирую.
Беляев разделил в одной большой партии японской бивольтинной породы Коку-сан № ПО гусениц на три примерно одинаковые группы. Рано слинявшие на П-й возраст и поздно слинявшие были выкормлены порознь, причем в следующих линьках в группе «скороспелых» выбрасывались все отстающие (при каждой линьке около 2/з гусениц), а в группе «позднеспелых» — все обгоняющие. После четвертой линьки в группе скороспелых (Л) и позднеспелых (С) были оставлены и отдельно докормлены до завивки и рано слинявшие (а) на V-й возраст, и поздно слинявшие (с). Получилось четыре группы: линия отбора Аа прошла все развития быстро, линия отбора Сс — все развитие медленно, тогда как Ас и Са, прошедшие часть развития быстро, а часть медленно, могут служить контролем. (Результаты отбора даны в табл. 14.)
В результате примитивным отбором удалось разделить популяцию «чистой бивольтинной породы» на группы, отличающиеся друг от друга более чем на двое суток по подолжительности гусеничного развития. И что особенно интересно, отобранные на «скороспелость» гусеницы дают легкие коконы, а бабочки этой группы откладывают сплошь оживающую грену, тогда как гусеницы «позднеспелые» дают тяжелые коконы, бабочки же этой группы часто откладывают зимующую грену. (Контрольные линии сохраняют исходные промежуточные значения по обоим этим показателям.) Марселина Какабадзе, уч.аствовашая в этой работе Беляева (1934—1937 гг.), экспериментально доказала зависимость жизнеспособности гусениц от веса кокона (табл. 15) 2.
В линии отбора С (на тяжелые коконы) наблюдалась исключительно сильная депрессия, особенно в жарком 1936 г. Наблюдалась огромная смертность среди гусениц, большинство бабочек были дефектными, а почти все самцы бесплодны, точно так же, как и значительная часть самок, у которых при анатомическом исследовании часто обнаруживались аномалии в строении яичников. В то же время ничего подобного не наблюдалось в линиях, разводившихся инбридингом, но с отбором на малый вес кокона (Л); т. е. понижение жизнеспособности не является в этих опытах саедствием инбридинга.
Некоторые авторы, впрочем, не верили в реальность такой связи призна-
2 М. И. Какабадзе. Селекция японской бивольтинной породы № НО по весу сырого кокона для изучения связи между весом кокона, жизнеспособностью и другими хозяйственно важными признаками шелкопряда, 1934. 30 с.— Арх. АН СССР, ф. 450, оп. 5, ед. хр. 38.
12* 179
Таблица 14
Результаты отбора на скорость развития в популяции Японской бивольтинной № ПО (по: Н. К. Беляев, 1936; из: Эфроимсон, 1946)
Линии отбора Направление отбора Продолжительность развития гусениц Средний вес сырого кокона, г Процент бивольтинных кладок
Аа + + 25 дн. 6 ч 0,998 100,0
Ас + - 26 дн. 5 ч 1,080 90,9
Са ~ + 26 дн. 7 ч 1,060 91,5
Се — — 27 дн. 10 ч 1,112 73,5
Таблица 15
Изменение жизнеспособности гусениц в результате отбора по весу кокона (по данным Марселины Какабадзе, 1934; из: Эфроимсон, 1946)
Сезон, год Поколения Жизнеспособность гусениц, % Коэффициент корреляции между жизнеспо собностыо и весом кокона Количество семей
Легкие коконы М) Средние коконы (fi) Тяжелые коконы (С)
Лето 1934 г. Fi 76,7 63,3 69,1 0,13±0,14 45
Весна 11 29,14 25,9 23,5 0,50±0,14 30
Лето 1935 г. 12 46,8 27,7 16,1 0,57±0,12 40
Осень, 1з 29,2 28,2 24,4 0,13±0,13 60
Весна! 14 61,3 39,6 44,1 0,34±0,18 30
Лето >1936 г. 15 60,2 52,0 55,0 48
Осень) <6 44,5 43,4 40,9 54
Весна 1937 г. Ь 58,4 67,1 64,0 30
ков. Так, П. А. Косминский решительно отрицал наличие обратной корреляции между выгодными для производства признаками тутового шелкопряда: в его многолетних опытах связь между тяжелым весом кокона и сниженной жизнеспособностью как будто не наблюдалась. Однако, проанализировав данные его же работы 1937 г. (табл. 16), В. П. Эфроимсон пришел к выводу: «Ясно, что более тяжелым средним весом кокона обладают именно семьи и коконы с большей продолжительностью развития. Понятно, что в условиях более тяжелых удлинение развития повлечет за собой и снижение жизнеспособности* (Эфроимсон, 1946, с. 397).
Добавляя подобные данные к результатам отбора на скорость развития, получаем, что отбор, приводящий к быстрому развитию, одновременно снижает вес кокона и шелконосность и повышает долю бездиапаузных кладок и устойчивость к жаре и инфекциям. Иными словами, отбор по одному признаку, сопряженному с вольтинностью, бивольтинизирует породу по всему коррелированному комплексу признаков. Противоположная картина при отборе на удлиненное развитие (увеличение веса кокона и шелконосности, увеличение доли зимующей грены, понижение жизнеспособности) указывает на сдвиг этого комплекса признаков к полюсу моновольтинности3.
3 См. также: В. П. Рувинская Исследования генетической связи между весом кокона и вольтинизмом бабочек тутового шелкопряда, 1936. Ис.— Арх. АН СССР, ф. 450, оп. 5, ед. хр. 76.
180
Таблица 16
Селекция бивольтинных пород на скороспелость и жизнеспособность (по: Космии-ский, 1937. Рукопись; из: Эфроимсон, 1946)
Номер Линии и сезон выкормки Средний вес кокона, мг
Продолжительное развитие Короткое развитие
1 Лето 1937 г., инбридинг 1160 (12 семей) 1095 (20 семей)
2 Осень 1937 г., инбридинг 1311 (11 линий) 1204 (22 линии)
3 Осень 1937 г., аутбридинг Избранные семьи осенней выкормки 1937 г. 1323 (4 линии) 1291 (4 линии)
4 инбридинг 1342 (11 семей) 1291 (22 семьи)
5 аутбридинг 1299 (9 семей) 1188 (7 семей)
Таблица 17
Изменение среднего веса коконов по инбредным группам отбора (по: Какабадзе, 1934; из: Эфроимсон, 1946)
Время выкормки Поколение Отбор на легкие коконы (А) Отбор на средние коконы (В) Отбор на тяжелые коконы (С) Популяционное среднее
Лето 1934 г. F, 1205±4 1296±5 1429 + 5
Весна 1 1. 1118±4 1480 ±4 1780±5 1359±7
Лето > 1935 г. ь 723 ±4 900 ±4 1067±5 929 ±4
Осень ) I» 699 ±1 826 ±4 891 ±4 957 ±4
Весна ] I. 1486±5 1914±6 2357 ±4 1671 ±6
Лето > 1936 г. 15 1002±3 1315±7 1678±7 1373±6
Осень J I. 924 ±6 1147 + 4 1475±7 1398± 7
Весна 1937 г. I? 1020±4 1495 ±4 1842±3 1491 ±
Представление о чрезвычайно высокой скорости этих коррелированных сдвигов на генетической основе дают результаты опытов Какабадзе 1934 г., представленные в табл. 17. Всего лишь восемь поколений отбора на легкие и тяжелые коконы привели к разнице линий Л и С на 80%! Одновременно в «тяжелой» линии С наблюдалось падение жизнеспособности (в летнее время), которое нельзя было отнести на счет инбридинга, так как такие же сдвиги наблюдались в кроссбредных линиях.
Таким образом, коррелятивная зависимость ряда адаптивных (хозяйственно ценных с позиции селекционера) признаков от вольтинности превращает породы шелкопряда в динамичные целостные системы, быстро реагирующие на отбор согласованным изменением всей скоррелированной системы признаков. А это значит, что комбинаторная генетика, позволяющая безошибочно конструировать по заранее намеченному плану линии с любым сложнейшим набором мутаций, определяющих качественные признаки, при решении задач-селекционного порядка отступает на далекий задний план. Более того, здесь возникает вопрос о сфере применимости ряда основных генетических представлений и необходимости их пересмотра.
Ревизия факториальной генетики. Результаты Н. К. Беляева заставляют отказаться от трех общепринятых генетических представлений (на что определенно указал Эфроимсон — 1946, с. 403—404).
181
Первое. Широко распространено мнение, что селекционное воздействие чувствительно изменяет генотип популяции лишь через очень большое число поколений. Оно основано на таблице Нортона, демонстрирующей, как слабо меняются под действием отбора концентрации рецессивных мутаций, вы-щепляющихся по формуле Харди.
Однако, как мы видели, отбор может в течение нескольких поколений перестроить генофонд породы в целом.
Второе. Генетики обычно отрицают явление вырождения (иначе как при инбридинге) или сводят вырождение к чисто фенотипическому изменению, устраняемому одним—двумя поколениями хорошего содержания. Однако может иметь место не только фенотипическое, но и генотипическое вырождение породы en masse и обратимость этого вырождения вовсе не очевидна. «Представление классической генетики о породе как о явлении относительно стабильном должно быть дополнено представлением о породе как о динамическом, лабильном процессе»,— заметил в этой связи Эфроимсон (1946, с. 403).
Третье ходячее представление, на котором необходимо остановиться, касается возможности свободного комбинирования по Менделю любых наследственных факторов, любых мутаций, любых наследственных признаков. Здесь следует отметить, что употребляемые в генетических опытах мутации отобраны по признаку четкости, безусловности и независимости проявления из огромного количества наследственных факторов. Фенотипическое осуществление большинства генов зависит от многообразных модифицирующих воздействий, что чрезвычайно затрудняет задачу получить сочетание одних желательных действий генов с другими. А наличие центральных регуляторов типа тех, что обеспечивают вольтинность, делает такую задачу невыполнимой.
Канализованность генетических сдвигов. Совокупность данных генетических и селекционных, систематики и механики развития, вскрывая наличие постоянных и достаточно прочных корреляций, выявляет наличие определенной направленности изменчивости у тутового шелкопряда, существование определенного параметра сдвига коррелированных признаков — канала вольтинизма. Селекционные сдвиги легче всего совершаются в направлении этого параметра (канала), в то время как ведение селекции вне этого канала требует разрыва сложившихся в эволюционном прошлом корреляций и достигается со значительным трудом.
О прочности коррелятивных связей свидетельствует закономерность, выясненная Беляевым (1936) на линиях и распространенная Эфроимсоном (1946) на помышленные породы шелкопряда: при скрещивании пород, относящихся к разным конституциям, и при последующем интенсивном отборе гибридов неизменно наблюдалось, что повышение шелконосности сопровождалось падением жизнеспособности в неблагоприятных условиях. При благоприятных же условиях, легко достижимых в лаборатории, снижение жизнеспособности носило скрытый характер и выявлялось лишь в неблагоприятных условиях промышленной выкормки. Ближайший практический вывод, который и сделал Эфроимсон,— требование выводить и испытывать породы в провокационных условиях.
Важным итогом генетических исследований Н. К. Беляева явилось то общее положение, что селекционное вмешательство, казалось бы,, столь невинное, вызывает ряд скрытых сдвигов. Во-первых, коррелированный сдвиг по параметру «моно-бивольтинность». Во-вторых, коррелированный сдвиг по
182
параметру повышения шелконосности, связанному со снижением жизнеспособности и удлинением развития. В-третьих, сдвиг по параметру резкого уменьшения числа рецессивных депрессоров, имевшихся в составе породы, с резким увеличением концентрации каждого из них.
В ходе обычной посемейной выкормки каждый из этих сдвигов не выявляется вовсе или выявляется очень слабо. Между тем любой из них в состоянии погубить, частично или полностью, многолетний кропотливый труд селекционера. Ведение селекции простым механическим отбором фенотипически лучших семей и особей по однажды намеченной схеме, без учета этих скрытых сдвигов совершенно недопустимо с позиций генетики тутового шелкопряда. Каждый из этих сдвигов находит свое внешнее выражение в депрессии. Для выбора адекватного способа ее устранения необходимо выяснить, чем этот нежелательный сдвиг обусловлен.
МОДИФИКАЦИОННЫЕ КОРРЕЛИРОВАННЫЕ СДВИГИ
Изучая модификацию вольтинности внешними условиями развития, Беляев снова обратился к температурным опытам (см. гл. 7 наст, книги), теперь на шелкопряде. Вместе с Н. П. Зимаревой он исследовал влияние на грену бивольтинных пород «весенней» (бивольтинизирующей) — 15°С и «летней» (моновольтинизирующей) — 27°С инкубации.
Опыты Беляева и Зимаревой показали, что холодная инкубация благоприятствует развитию по бивольтинному типу (самооживающие кладки, легкие малошелконосные коконы, быстрое развитие и т. д.), тогда как тепловая инкубация переключает бивольтинные породы на развитие по моно-вольтинному типу (табл. 18).
Сопоставим с этими данными также и некоторые другие. Данные табл. 19 и 20 показывают, что моновольтинизирующие условия развития приводят к медленному гусеничному развитию, непропорционально большой куколочкой стадии, крупным размерам кокона (гусеницы, куколки, бабочки), повышению шелконосности, а также меньшей устойчивости к жаре, плохому качеству листа и инфекциям. (Что именно медленное развитие, а не накопление питательных веществ вызывает моновольтинизацию, показывает опыт Бондаренко4, давший высокий процент моновольтинизации при замедлении развития гусениц голоданием. Недокормленные партии, долго развивающиеся, дали маленькие плохие коконы, а вышедшие из этих коконов бабочки отложили гораздо больше зимующей грены, нежели бабочки, вышедшие из нормально выкормленных партий.)
Параллелизм модификационной и генетической коррелированной изменчивости. Изучение влияний внешних условий на вольтинизм и скоррелированные с ним признаки выявило глубокий параллелизм между действием наследственных и ненаследственных факторов (ср., например, табл. 20 и 21). Генотипически бивольтинные бабочки откладывают диапаузирующие яйца, если инкубация их на эмбриональной стадии проходила при t = 25—26°С, и яйца недиапаузирующие при температуре инкубации 15—17°С (см. табл. 20). При промежуточных температурах решающую роль приобретает длительность суточного освещения на эмбриональной стадии материнского
4 А. К. Бондаренко, Н. К. Беляев. Влияние разных степеней кормления на проявление бивольтинизма у моновольтинных и бивольтинных пород тутового шелкопряда, 1936. 19 с.— Арх. АН СССР, ф. 450, по. 5, ед. хр. 10.
183
Таблица 18
Влияние высокой температуры инкубации на признаки червей и коконов бивольтинных пород (по: Зимарева, 1935. Рукопись; из: Эфроимсон, 1946)
Температура инкубации, °C Среднее пяти бивольтинных пород
Средний ес Вес шелковой оболочки, мг Процент безднапауз-ных кладок
27 1321 181 4,4
15 1024 123 89,5
Таблица 19
Влияние моно- и бивольтинной инкубации на признаки бивольтинных пород (по: Зиракишвили, 1944. Рукопись; из: Эфроимсон, 1946)
Инкубация Продолжительность выкормки, дни Средний вес сырого кокона, мг Вес оболочки, мг Процент оболочки
Моновольтинизирующая 26° С, свет, высокая влажность 35,5 162,4 239 15,0
Бивольтинизирующая 15—16° С, темнота, 70%-ная влажность 29,1 110,3 162 14,7
Таблица 20
Влияние температуры инкубации на признаки бивольтинных пород (по: Хиото, 1926; из: Эфроимсон, 1946)
Показатель Температура инкубации, *С P/L, % Вес шелковой оболочки, мг
Среднее трех бивольтинных пород 25,6 57,8 158,4
японские № 106 и № 107, китайская № 101 17,2 44,4 130,0
Таблица 21
Относительная длина стадий развития у моно- и бивольтинных пород при холодовой инкубации (по: Папалашвили, 1941. Рукопись; из: Эфроимсон, 1946)
Показатель Продолжительность развития гусеницы (L), ч Продолжительность нимфоза (Р), ч P/L,
Средневзвешенное трех моновольтинных пород 816± 1 489 ±2 59,9
Средневзвешенное двух бивольтинных пород 722 ±2 333 ±2 46,1
184
Рис. 20. Канализованные конституционные сдвиги у тутового шелкопряда (по: Эфроимсон, 1965)
Абсцисса — абсолютный вес кокона, оболочки, нити, гусеницы; относительный вес оболочки; продолжительность развития; относительная длительность стадии куколки. Ордината — устойчивость к жаре, болезням; быстрота развития; жизнеспособность. Ось МР — степень вольтинности: моно- и би-поливольтинность
организма: кратковременное освещение предрасполагает к бивольтинному поведению, длительное — к моновольтинному. Если же оба эти фактора имеют промежуточные значения, то выбор пути развития определяет степень влажности: умеренная — на бездиапаузное развитие, очень высокая — на диапаузное (табл. 19). При детерминации вольтинности любым их этих факторов в сторону моно- или бивольтинности изменяется весь комплекс скоррелированных признаков.
Совпадение картины коррелятивных сдвигов по параметру моно- биволь-тинность (см. рис. 20), полученных при регуляции вольтинности отбором (т. е. на основе изменения генотипа) и при регуляции вольтинности температурой или фотопериодизмом (модификационные сдвиги), приводит к мысли о том, что и наследственные и средовые факторы реализуют свое влияние через один и тот же морфогенетический аппарат,, управляющий развитием многих признаков. И действительно, впоследствии благодаря работам японских исследователей выяснилось, что таким управляющим центром является нейроэндокринный аппарат. В подглоточном ганглии под влиянием ряда «моновольтинизирующих» факторов, генотипических или средовых, резко увеличивается число нейросекреторных клеток, выделяющих гормон диапаузы. Инъекция экстрагированного гормона куколке также превращает незимующие яйца в зимующие. Регуляция коррелированного с вольтинностью комплекса признаков зависит также от антагонистического гормону диапаузы гормона роста, выделяемого corpora allata. Основные гены серии V (вольтинизм) расположены на VI, 4, а основной ген, контролирующий число линеек, М (мольтинизм),— на IV, 3. Их активность модифицируется геном SH, имеющим не менее трех аллелей (см.: Астауров и др., 1975, с. 128).
13 В. В Бабков
185
ЭВОЛЮЦИОННО-ГЕНЕТИЧЕСКОЕ ЗНАЧЕНИЕ ВОЛЬТИНИЗМА
Переходя от изучения наследования аберративных признаков (задача классической генетики) к наследованию адаптивных систем, от случайных внутрипопуляционных различий к филогенетически значимым, от проблемы образования генофонда популяции к проблеме становления адаптивных расовых отличий, мы сталкиваемся с качественно иными явлениями, где все большую роль играют интегрирующие морфогенетические процессы, которых касался Четвериков, говоря о «генотипической среде», и особенно подчеркивал Шмальгаузен в теории стабилизирующего отбора.
Обсуждая корреляции комплекса адаптивных признаков с вольтинностью в связи с понятием плейотропии, на котором основана концепция генотипической среды, вспомним о старом кардинальном противоречии между механикой развития, изучающей связи, взаимодействия и корреляции процессов онтогенеза, и классической генетикой, имеющей дело с автономным, дискретным действием мутаций в онтогенезе. А. А. Малиновский (1939) показал, что эволюционно выгодным является довольно низкий уровень плейотропии и что мутации с изначально широкой плейотропией почти не имеют шансов на закрепление в виде. И. И. Шмальгаузен (1940) развил этот метод, предложив схему (рис. 21) многостепенного выражения плейотропии (а), изменяемого в процессе эволюции вследствие естественного отбора модификаторов (б) и дальнейшего усложнения форм взаимодействия (в), в результате которого развивается типичная система морфогенетических корреляций. Процесс определения диапаузного—бездиапаузного развития в ходе отбора приобретает широкие онтогенетические корреляции и становится одним из стержневых процессов онтогенеза; под жестким контролем отбора находится адаптивная система вида как целое. «Становление адаптации, в отличие от случайного наследственного различия, включает в себя исторический процесс установления необходимых онтогенетических корреляций, процесс интеграции»,— писал Эфроимсон (1946, с. 444).
Становление новой адаптивной системы, открывая новые возможности расселения и размножения вида, его эврибионтности, вызывает появление новых форм изоляции и является важным этапом внутривидовой эволюции. Создание новой широкой адаптации в некоторых случаях можно сопоставить по эволюционной значимости не только с образованием вида, но и с образованием более крупных систематических единиц, ибо одна и та же адаптивная система с некоторыми вариациями может встретиться у целого ряда видов. (Этот подход к вычленению таксономических категорий близок к взглядам А. А. Ляпунова, согласно которым «целесообразно стремиться к некоторой объективной таксономии, при которой определенным таксонам соответствовали бы определенные структуры управления на некотором вполне определенном уровне» — Ляпунов, 1970, с. 201; обсуждение: Бабков, 1981.)
Пусковые механизмы диапаузного поведения. В качестве пускового механизма определения диапаузности—бездиапаузности могут выступать как определенные факторы среды, так и различные наследственные факторы: аутосомный ген, сцепленный с полом ген, серия генов. Некоторые авторы, например Е. И. Лукин (1940) и особенно Г Ф. Гаузе (1941), видели между модификацией и соответствующей ей мутацией чисто внешнее сходство, притом «конвергентное», объясняемое приспособлением к одному и тому же внешнему фактору, но реализуемое через разные онтогенетические механизмы. Однако по Шмальгаузену генокопия может вызывать признак независи мо от внешней среды: признак будет появляться раньше и в более специали
Рис. 21. Развитие типичной системы морфогенетических корреляций (по: Шмальгаузен, 1940)
/ — схема многостепенного выражения плейотропии; // — ее изменение вследствие отбора модификаторов; /// — дальнейшее усложнение форм взаимодействия
зированном виде, осуществляясь через механизм, созданный предшествующей модификацией. К этому близок взгляд К. Уоддингтона (1944), что в процессе становления модификации создаются альтернативные каналы развития, выбор между ними диктуется сперва внешними условиями, а затем в роли направляющего агента выступает «стрелочный ген». Эфроимсон (1946, с. 476) распространил этот взгляд на случай выбора диапаузного поведения (на схеме даны «стрелочные факторы», которые у бивольтинных пород могут переключать развитие на путь реализации моновольтинного комплекса признаков)5:
Температура Продолжительность Аутосомный Сцепленный с Серия по-
освещения ген полом ген лигенов
В ходе становления вольтинности должен был «происходить филогенетический сдвиг порога детерминации бездиапаузности и выход его у моно-вольтинных пород за пределы колебаний их нормальных условий существования»,— такой вывод Эфроимсон (1946, с. 482) сделал при анализе эволюционно-генетических особенностей вольтинизма, проведенном с позиций теории стабилизирующего отбора Шмальгаузена. Поливольтинные расы, филогенетически более древние, формируют зимующие яйца лишь при воздействии холода на куколочную стадию материнской генерации. В результате отбора на шелконосность у бивольтинных пород норма реакции формирования зимующих яиц расширена и детерминация сдвинута на более раннюю стадию. При колебаниях температуры роль «стрелочного фактора» будет играть фотопериодизм — дополнительная настройка. Параллельно появляются гены вольтинизма, действующие в онтогенезе посредством сложившейся скоррелированной системы, приобретая тем самым широкую плейотропию. Эти гены сдвигают норму температурной реакции диапаузы. Полигетерозиготность по этим генам позволяет популяции быстро приспособиться к новым локальным условиям; она же отвечает за градации бивольтинизма. У моновольтинных пород, наиболее удаленных от дикого типа, реакция детерминации диапаузы автономизируется.
5 Замену, что в схеме Эфроимсона понимание полигении существенно отличается от интерпретации К. Мейзера (например: Mather, 1949), видившего качественное различие между олигогенами и генами-модификаторами. См. недавнее обсуждение вопроса у Э. X. Гинзбурга и 3. С. Никоро (1982; Гинзбург, 1982).
187
Порог детерминации без диапаузности постепенно (ступенчато) сдвигается у поли- и бивольтинных пород, покуда не уходит (у моновольтинных пород) за пределы нормальной колеблемости условий существования вида. Так он ускользает от действия естественного отбора, а в результате может сохраниться («дремлющий фонд плюрипотентности»). Мутация или специфический раздражитель необычайной интенсивности может пробудить эту спящую потенцию бездиапаузного развития (ср. представления Н. К. Кольцова о «спящих генах», развивавшихся в связи с наследственными гомео-зисами — гл. 6 наст, книги). Способность же моновольтинных пород дать незимующую грену (при крайне низких температурах инкубации материнской генерации) не имеет какого-либо адаптивного значения. Это, несомненно, рудиментарная реакция. В то же время сохранение аппарата онтогенетической реализации бездиапаузности может сопровождаться распадом корреляций — как, например, в случае наследственных гомеозисов у дрозофилы.
Эволюционно значимая изменчивость. Далее, эти исследования Н. К. Беляева позволили подойти к вопросу: «какая часть изменчивости является эволюционно значимой?», в форме которого конкретизовалась общая проблема Четверикова.
У бивольтинных пород создается система гетерозиготности по полудоми-нантным генам вольтинности, обеспечивающая лабильность расы по серии скоррелированных признаков и возможности резкого сдвига за одно—два поколения. Как показали исследования феногенетики массовых популяционных изменений (гл. 11 наст, книги), полимерность и полудоминантность вовсе не являются чем-то особенным и неожиданным, они представляют собой нормальные свойства изменчивости, на основе которой развиваются адаптивные системы. Разницу в генетической природе качественных и количественных признаков можно объяснить историческим происхождением как рецессивности качественных мутаций, с одной стороны, так и систем полимерной полудоминантности, с другой.
Эволюционная роль полимерной гетерозиготности рельефно выступает, например, при конструировании иммунных рас растений. Выделяют две формы устойчивости у растений. «Вертикальная», качественная, устойчивость управляется моногенно и характеризуется полным иммунитетом к определенному заболеванию. Но она беспомощна при встрече с новыми мутантными расами. «Горизонтальная», количественная, контролируется большим числом генов; для нее характерна вариирующая степень восприимчивости к болезням. (Такой тип контроля устойчивости свойствен диким формам.) В связи с полигенным контролем горизонтальная устойчивость гораздо стабильнее и универсальнее, чем вертикальная. Усилия селекционеров и фитопатологов, направленные по самому, казалось бы, простому пути — выделение абсолютно непоражаемых растений и перенесение генов устойчивости — показали его бесперспективность. Более того, здесь велика опасность гомогаметизации по другим генам, не связанным непосредственно с восприимчивостью к болезням. Оказалось, что на иммунитет может влиять цитоплазматическая мужская стерильность (ЦМС). Техасский тип ЦМС сделал кукурузу восприимчивой к Т-расе возбудителя южного гельминтоспориоза; накопление этой расы повело в 1970 г. к эпифитотии, вызвавшей недобор урожая кукурузы на миллиард долларов (Дьяков, 1977). Подобные примеры показывают потенциальную опасность глобальной унификации генофондов — а именно на этом построена, например, «зеленая революция» Н. Борлоуга.
188
На основе полимерной полудоминантной гетерозиготности может легко протекать становление как сезонных форм, так и локальных географических рас. Подобные явления быстрой смены генотипа популяции были продемонстрированы Тимофеевым-Ресовским и Лусом на божьей коровке и Дубининым и Тиняковым, а позже Борисовым — на инверсиях дрозофилы (ср. описание яровой и озимой рас рыб Л. С. Бергом, 1934). Данному состоянию среды соответствует оптимальная мера развития органа животного, что находит свое выражение, между прочим, в правилах Бергмана, Аллена, Глогера, в описанном Хаксли явлении градиента изменения количественных признаков (clines) и т. п. Если экотипы являются результатом дифференцирующего действия отбора, разнонаправленного в пространстве, то могие формы сезонной изменчивости популяций созданы дифференцирующим действием отбора, разнонаправленным во времени.
Подобные каналы изменчивости и системы, контролирующие адаптивные коррелированные сдвиги, должны быть свойственны самым разным видам организмов. «Возможно, подобные централизованные узлы коррелятивных связей имеются у многих высших организмов, причем не только у животных, где они, видимо, представлены нейроэндокринным аппаратом, но и у растений. Проявление вольтинизма у шелкопряда, например, весьма сходно с тем, что можно наблюдать у растений для генетических и средовых воздействий на такой процесс, как давно изученная Гасснером и Максимовым верна-лизация (впоследствии ставшая более известной под названием яровизация)» (Астауров и др., 1975, с. 128).
Экологическая и географическая дифференциация может направляться в определенное русло, соответствующее правилу Бергмана, системой гипофизарной ортогенетической корреляции. А. А. Малиновский (1945, 1948, 1960) показал существование по крайней мере двух основных параметров канализованных конституциональных сдвигов у человека: астено-пикнической, в основе которого лежит соотношение процессов ассимиляции—диссимиляции, и атлето-церебральный, определяемый соотношением роста и дифференцировки скелета. Многие растения имеют яровые и озимые формы; у яровизируемых растений найдено не менее шести генов, отвечающих за холодовую реакцию. В случае генов Hh, li, Kk, найденных у пшеницы, ячменя, ржи и Arabidopsis, это действие вызывают рецессивные аллели; в случае трех других — Ss, Tt, Uu, найденных у пшеницы, ячменя, Hyosciamus и Arabidopsis — доминантные. Опыты Д. К. Беляева последних 25 лет по одомашниванию серебристо-черных лисиц свидетельствуют о наличии канала коррелятивных сдвигов: «особачивание» лисиц при отборе на неагрессивное поведение (Д. К. Беляев. 1962, 1979; Belyaev, 1979; обсуждение: Бабков, 1981).
Микро- и макроэволюция. Наконец, в свете результатов Н. К. Беляева по-новому предстает поблема соотношения процессов микро- и макроэволюции, намеченная еще в теоретическом исследовании Четверикова 1926 г. Наличие системы онтогенетических корреляций определяет ответ на отбор, искусственный или естественный. Канализованным оказывается не только онтогенез, но в известной мере и филогенез. Результатом этого оказывается определенный филогенетический ряд, охватывающий как единое целое эволюцию всего вида в течение многих тысяч поколений на всем протяжении Палеарктики, субтропиков и тропиков Старого Света. Ортогенетический канал, являясь продуктом филогенеза прошлого, направляет филогенез в настоящем и будущем (см.: Эфроимсон, 1965; Бабков, 1980). Поскольку основные филогенетические сдвиги идут по главным каналам, где измене
189
ния комплекса сопряженных признаков гармонизированы, то требует известного пересмотра и вопрос об обратимости эволюции: при условии сохранения ортогенетического канала предшествующая форма может быть воссоздана относительно быстро, отбором даже по второстепенному признаку, если он связан с основным каналом.
В филогенезе культурных пород тутового шелкопряда отбор шел в одном главном направлении — на повышение шелконосности. Если же оба полярных типа конституций попали бы в противоположные условия с противоположными векторами отбора, то они могли бы стать начинающимися видами, а в дальнейшем могли бы дать ортогенетический ряд видов. Примеры таких ортогенетических рядов, происхождение которых изучено недостаточно, дает палеонтология (ряд гиппарион—лошадь и др.). В результате встает вопрос: способно ли пролить свет на выявление основных каналов филогенеза изучение селекционных микросдвигов, раскрытие иерархической структуры онтогенеза (или хотя бы выявление корреляций, как первый шаг к этому)? Если да, то задачей эволюционной генетики вместо изучения отдельных признаков становится выявление и изучение основных каналов эволюции, центральных регуляторов онтогенеза как результата и как механизма процесса эволюции.
В наши дни особенно принято говорить о синтезе генетики с эволюционизмом; это мнение распространилось после известной конференции в Принстоне (Genetics, paleontology and evolution, 1949), а название «современный синтез» дала сводка Дж. Хаксли (Huxley, 1942). Однако этот энтузиазм следует признать преждевременным. У нас нет еще удовлетворительного представления о процессе эволюции; нет ответа на кардинальные вопросы — об источниках наблюдаемого количества изменчивости, о причинах высоких скоростей и направленности эволюционных перестроек. Материалы этой главы, как и предыдущей, дают основание полагать, что в процессе создания общей картины эволюционного процесса «нефакториальной» генетике, основанной на системном подходе, будет принадлежать выдающаяся роль.
В исследованиях Н. К. Беляева на шелкопряде, ориентированных на практически значимые результаты, были выявлены: комплекс сопряженных адаптивных (хозяйственно ценных) признаков; подчиненность этих признаков вольтинности — признаку более высокого иерархического ранга; механизмы, определяющие диапаузное поведение. Здесь был сделан важный шаг к определению эволюционно значимой части природной изменчивости. Изучение коррелятивных генетических и модификационных сдвигов позволило пролить новый свет на проблему отношений процессов микро- и макроэволюции. Новая конкретизация центральной проблемы эволюционной генетики, достигнутая в этих исследованиях ценой отказа от некоторых основных представлений «факториальной» генетики, свидетельствует о далеко не исчерпанной эвристической ценности этой проблемы.
ЗАКЛЮЧЕНИЕ
Становление эволюционной генетики в нашей стране было чрезвычайно стремительным. Всего лишь за десятилетие был пройден путь от начала систематических исследований по генетике в СССР до выхода советской эволюционной генетики на передовые рубежи мировой науки. Такое развитие событий — когда его рассматривают изолированно, вне исторического контекста — производит впечатление едва ли не чуда. Понять его можно лишь обратившись к естественно-научным традициям, на которых выросла московская школа эволюционной генетики, рассматривая эту школу на фоне господствовавших в то время умонастроений в биологии.
Основные моменты эволюционной генетики пришли из двух направлений (натуралистического и экспериментального) русской зоологии. Однако то или иное положение не существует в науке независимо от других: делая выбор в пользу одного положения, мы тем самым как бы обращаемся к ряду сцепленных с ним положений, определенных методов и кругу фактов. К началу нашего века натуралисты и экспериментаторы явно обособились друг от друга как по методам, так и по кругу проблем. С возникновением конфликта менделистов и сторонников теории отбора («экспериментаторов» и «натуралистов»), эта дифференциация лишь усилилась. Поэтому мысль С. С. Четверикова поставить круг проблем дарвинизма на генетическую основу привела его к необходимости отказаться от каждого из двух упомянутых канонов. Ему нужно было решить следующие задачи: построить концептуальную схему, объединяющую эволюционные и генетические представления; показать правомерность экспериментального изучения природной изменчивости (в условиях, как сказано, обособленности дарвинистов-натуралистов и менделистов-экспериментаторов); разобрать причины различий количества изменчивости, находимой в природе и лаборатории,— различий настолько ошеломляющих, что мутации дрозофилы одно время считались лабораторными артефактами; выяснить генетическую основу неадаптивных процессов в эволюции, поставить вопрос о факторах эволюции и их соотношении (при умонастроениях, согласно которым эволюция сводилась либо к одному естественному отбору, либо к изоляции, либо к миграциям, либо к мутациям).
Успеху начинания благоприятствовала как творческая обстановка Института экспериментальной биологии, созданного Н. К. Кольцовым, так и наличие группы молодых одаренных зоологов, способных выработать (вместе со своим учителем) новую систему познавательных ориентиров, обеспечившую возможность построения эволюционной генетики. В замечательном теоретическом исследовании 1926 г. Четвериков сформулировал центральную проблему новой области биологического знания — «природа и поддержание изменчивости в популяциях», подчеркивал значение системного подхода для ее разработки, привлекал для иллюстраций определенную модель генетической эволюции. Эта статья послужила программой для построения
191
эволюционной генетики. А в первом экспериментальном исследовании насыщенности мутациями диких популяций дрозофилы, проведенном Лабораторией Четверикова в 1925—1926 гг., наметились основные пути ее разработки. Это исследование стало образцом для всей последующей популяционно-генетической работы — в СССР, Европе, США.
В этом первом популяционно-генетическом исследовании определились все направления разработки центральной проблемы.
Феногенетическое и популяционное направления были в значительной мере объединены в двух циклах работ московской школы. В одном было показано (на шелкопряде) существование сопряженного комплекса адаптивных признаков, их иерархическая соподчиненность, развитие на этой основе коррелятивной изменчивости и выявлена роль центральных регуляторов в индивидуальном и историческом развитии. Другой цикл — «феногенетический анализ популяций» (разных групп двукрылых) привел к выявлению закономерностей массовой природной изменчивости; в нем был сделан важный шаг к объединению методов онтогенетики, генетики популяций, систематики, экологии. Эти два цикла работ указали на необходимость и возможные пути пересмотра основ факториальной генетики популяций и дали основание для постановки в новом свете проблемы отношений микро-и макроэволюции.
Судьба разных теоретических моментов (проблемы, подхода, модели) программы построения эволюционной генетики была в этом развитии событий различной. Первоначальная временная модель генетической эволюции (отбор возникающих в природе мутаций) постоянно пересматривалась и на каждом этапе заменялась исправленным представлением. Изменялась и связанная с моделью задача: от выяснения «количества изменчивости» — к «изменчивости по приспособленности» и в двух последних циклах работ — к «эволюционно значимой изменчивости». Тем временем системный подход Четверикова находил в этих работах все более полное выражение, а его центральная проблема углублялась в различных аспектах и приобретала все новый смысл. Усвоив программу Четверикова и опираясь на создавшие ее силы, его ученики смогли положить начало новой области биологического знания — эволюционной генетике. Именно определенность программы и вместе с тем широкое ее понимание дали успех всему делу.
Якоб Бернулли велел выбить на своем надгробии под изображением очаровавшей его воображение логарифмической спирали слова: «Eadem mutata resurgo» — «Измененная, я воскресаю той же самой». Жизнеспособность системы познавательных ориентиров и центральной проблемы эволюционной генетики — ядра программы Четверикова — была обусловлена возможностью пересмотра и в конечном итоге отказа от первоначальной модели явления. В 1936 г. Четвериков говорил, в связи с отношениями генетики и эволюции: «Старый Дарвинизм не отпадет, а войдет как фундамент в новое, гораздо более обширное, гораздо более грандиозное здание обновленной эволюционной теории» 1 Такая же судьба выпала сформулированной им центральной проблеме эволюционной генетики.
1 Фонд Четверикова, on. 1, ед. хр. 27, л. 6.
ЛИТЕРАТУРА
Астауров Б. Л. Исследование наследственного изменения гальтеров у Drosophila melanogaster Sehin.— Журн. эксперим. биологии (А), 1927, т. 3, вып. 1—2, с. 1—61; вып. 3—4, с. 199—201.
Астауров Б. Л. Фенотипическая изменчивость гомодинамических частей в пределах организма.— Тр. Съезда по генетике и селекции. Л., 1930, т. 2, с. 155—162.
Астауров Б. Л. Две вехи в развитии генетических представлений.— Бюл. МОИП. Отд. биол., 1965, т. 70, № 4, с. 25—32.
Астауров Б. Л. Homo sapiens et humanis. Человек с большой буквы и эволюционная генетика человечности.— Новый мир, 1971, № 10, с. 214—224.
Астауров Б. Л. Генетика и проблемы индивидуального развития.— Онтогенез, 1972а, т. 3, № 6, с. 547—565.
Астауров Борис Львович. Материалы к биобиблиографии ученых СССР. Вступит, статья П. Ф. Рокицкого. М.: Наука, 19726. 65 с.
Астауров Б. Л. Жизнь С. С. Четверикова.— Природа, 1974а, № 2, с. 57—67.
Астауров Б. Л. Наследственность и развитие. М.: Наука, 19746. 359 с.
Астауров Б. Л. К итогам моей научной деятельности в области генетики.— В кн.: Историко-биологические исследования. М.: Наука, 1978, вып. 6, с. 114—160.
Астауров Б. Л., Рокицкий П. Ф. Николай Константинович Кольцов. М.: Наука, 1975. 168 с.
Астауров Б. Л., Никоро 3. С., Струнников В. А., Эфроимсон В. П. Научная деятельность Н. К. Беляева. (К истории советских генетических исследований на шелковичном черве).— В кн.: Из истории биологии. М.: Наука, 1975, вып. 5, с. 103—136.
Бабков В. В. Модель генетической эволюции популяций с пространственной изоляцией.— В кн.: Проблемы кибернетики. М.: Наука, 1972а, вып. 25, с. 77—80.
Бабков В. В. Некоторые проблемы компьютерного подхода к эволюционным процессам.— В кн.: Модели структурно-функциональной организации биологических систем. М.: Изд-во АН СССР, 19726, с. 148—156.
Бабков В. В. Организация менде-левских популяций как условие их эволюции.— В кн.: Организация и эволюция живого. Л.: Наука, 1972в, с. 108— 112.
Бабков В. В. Моделирование эволюционно-генетических процессов в популяциях. I.— Генетика, 1973а, т. 9, № 4, с. 91 — 100.
Бабков В. В. Конференция памяти Четверикова.— Генетика, 19736, т. 9, № 9, с. 178—179.
Бабков В. В. «Шахматная» модель популяции.— В. кн.: Биометрические методы. М.: Изд-во МГУ, 1975, с. 103— 111.
Бабков В. В. Перечитывая Четверикова. (Полвека эволюционной генетики).— Вопр. философии, 1977а, № 1, с. 126—137.
Бабков В. В. Полемика и научное изменение: происхождение генетики популяций.— В кн.: Роль дискуссии в развитии естествознания. М.: Наука, 19776, с. 75—77
Бабков В. В. Кибернетические интерпретации в биологии и проблема направленности развития.— Folia Вае-riana. Tallinn: Valgus, 1978, t. 3, с. 133—136.
Бабков В. В. Имееет ли генетика отношение к «большой» эволюции? — В кн.: Микро- и макроэволюция. Тарту: ТГУ, 1980, с. 149—153.
Бабков В. В. Системный стиль в изучении естественного отбора.— В. кн.: Системные исследования. Ежегодник. 1980. М.: Наука, 1981, с. 404—419.
Бабков В. В. Вводные замечания. (К ст.: С. С. Четвериков. Экспериментальное исследование геновариаций, скрытых в популяциях, 1929.) — Генетика, 1982, т. 18, № 3, с. 341—343.
Бабков В. В. Центральная проблема генетики популяций.— В кн Чет-
193
вериков С. С. Проблемы общей биологии и генетики. Новосибирск: Наука, 1983а, с. 6—40, 259 -268.
Бабков В. В. Великие естествоиспытатели. Системный стиль мышления в изучении живой природы.— Человек и природа, 19836, вып. 10, с. 3—17.
Бабков В. В. Генетика, эволюция, интегративизм.— В кн.: Методологические проблемы эволюционной теории. Тарту: ТГУ, 1984, с. 162—164.
Балкашина Е. И. Случай наследственного гомеозиса (геновариация «Aristopedia») у Drosophila melano-gaster.— Тр. Ill Съезда зоологов, анатомов и гистологов. Л.: Изд. Главнаука, 1928а, с. 170—171.
Балкашина Е. И. Случай наследственного гомеозиса (геновариация «Aristopedia») у Drosophila melanogaster.— Журн. эксперим. биологии (А), 19286, т. 4, вып. 2, с. 93—106.
Балкашина Е. И., Ромашов Д. Д. Генетическое строение популяций Drosophila. I. Генетический анализ звенигородских (Московской обл.) популяций Drosophila phalerata Meig., transversa Fall, и vibrissina Duda. (По материалам Б. Л. Астаурова, Н. К. Беляева и Е. И. Балкашиной) — Биол. журн., 1935, т. 4, № 1, с. 81 —106.
Балкашина Е. И., Беляева В. И., Головинская К. А., Черфас И. Б. Д. Д. Ромашов и его роль в развитии генетики.— В кн.: Из истории биологии. М.: Наука, 1975, вып. 5, с. 76—91.
Беляев Д. К. О некоторых проблемах коррелятивной изменчивости и их значении для теории эволюции и селекции животных.— Изв. СО АН СССР, 1962, № 10, с. 111 — 124.
Беляев Д. К. Некоторые генетикоэволюционные проблемы стресса и стрессируемости.— Вести. АМН СССР, 1979, № 7, с. 9—14.
Беляев Д. К., Рокицкий П. Ф. О некоторых методологических проблемах в развитии советской генетики.— Вопр. философии, 1977, № 2, с. 124—134; № 3, с. 136—148.
Беляев Н. К. Экспериментальное исследование изменчивости окраски гусениц Spilosoma lubricipeda Esp. (Arctiidae, Lepidoptera). — Журн. эксперим. биол. (А), 1926, т. 2, вып. 4, с. 157—219, 249—251.
Беляев Н. К. Температурные опыты с гусеницами группы Spilosoma Stph. (Arctiidae).— Журн. эксперим. биологии (А), 1928, т. 4, вып. 2, с. 107—130.
Беляев И. К. Хромосомные комплексы Lepidoptera й их отношение к систематике и филогении этого отряда.— Тр. Съезда по генетике и селек-!94
ции. (Ленинград, 10—16 января, 1929 г.). Л., 1930, т. 2, 163-171.
Беляев Н. К. Проблемы генетики и селекции тутового шелкопряда: Дис... докт. биол. наук. Тифлис, 1936.
Беляев И. К. Генетический анализ окраски бабочек тутового шелкопряда (Bombyx mori L). — Биол. журн., 1937, т. 6, № 1, с. 51—68.
Беляева В. Н. Изменчивость жилкования крыла в природных популяциях Drosophila melanogaster.— ДАН, 1946, т. 54, № 7, с. 633—636.
Беляева В. И. Изменчивость жилкования крыла у гибридов Drosophila melanogaster X Drosophila simulans. 1948. Рукопись (6 с., библиогр. 7 назв., 2 ил.).
Беляева В. И. Дмитрий Дмитриевич Ромашов. 1972. Рукопись. 27 с.
Беляева В. Н., Ромашов Д. Д. Об отборном значении различных изменений жилкования в природных популяциях Drosophila melanogaster.— ДАН СССР, 1946, т. 54, № 8, с. 729—732.
Беляева В. Н., Ромашов Д. Д. Генетическое строение популяций Drosophila. II. Изменчивость жилкования в популяциях Drosophila melanogaster. 1947. Рукопись (22 с., библиогр. 28 назв., 2 ил.).
Берг Л. С. Яровые и озимые расы у проходных рыб.— Изв. Отд. математ. и естеств. наук АН СССР. Сер. биол., 1934, № 5, с. 711—732.
Берг Р. Л. Генетический анализ популяций Drosophila melanogaster Де-лижана (Армения). Стойкость формообразования в свете генетических проблем. Вид как эволюирующая система.— В кн.: Рефераты работ учреждений Отд. биол. наук АН СССР за 1941 —1943 гг. М.; Л.: Изд-во АН СССР, 1945, с. 181, 183—186.
Берг Р. Л., Бриссенден Э. Б., Александрийская В. Т., Галковская К- Ф. Генетический анализ двух природных популяций Drosophila melanogaster.— Журн. общ. биологии, 1942, т. 2, № 1, с. 143—158.
Борисов А. И. Адаптивное значение хромосомного полиморфизма. III. Дальнейшая эволюция городской расы Drosophila funebris.— Генетика, 1969, т. 5, с. 119—122.
Борисов А. И. Взаимодействие хромосом Drosophila funebris городских и сельских рас в экспериментальных популяциях.— Генетика, 1970, т. 6, № 2, с. 81—90.
Вавилов Н. И. Центры происхождения культурных растений.— Тр. по прикл. ботанике и селекции. Л., 1926, т. 16, вып. 2, с. 3—138. (англ.— с. 139— 248)
Вавилов Н. И. Географические закономерности в распределении генов культурных растений.— Тр. по прикл. ботанике, генетике и селекции. Л., 1927, т. 17, вып. 3, с. 411—419. (англ.— с. 420—428).
Вавилов И. И. Линнеевский вид как система.— Тр. по прикл. ботанике, генетике и селекции. Л., 1931, т. 26, вып. 3, с. 109—134.
Воронцов Н. Н., Яблоков А. В., К 70-летию Н. В. Тимофеева-Ресовского.— Бюл. МОИП. Отд. биол., 1970, т. 75, вып. 5, с. 144—157.
Гагарин В. С. Экспериментальный анализ генетико-автоматических процессов.— Биол. журн., 1933, т. 2, вып. 4—5, с. 451—457.
Гаузе Г Ф. Роль приспособляемости в естественном отборе.— Журн. общ. биологии, 1941, т. 2, № 2, с. 193—209.
Гаузе Г Ф., Витт А. А. О периодических колебаниях численности популяции: математическая теория релаксационного взаимодействия между хищниками и жертвами и ее применение к популяциям двух простейших.— Изв. Отд. математ. и естеств. наук АН СССР. Сер. биол., 1934, № 10, с. 1551 —1559.
Генетика и селекция тутового шелкопряда/Под ред. акад. Н. К. Кольцова. М.: Изд-во ВАСХНИЛ, 1936. (Содержание: Н. К. Беляев, Н. П. Зимарева, В. П. Рувинская, Т. А. Говорова, Т. М. Гурская, К. А. Лоладзе, Р. Аллавердян, Н. В. Ващинская. Экспериментальное исследование пород и гибридов тутового шелкопряда. Результаты летнего испытания 1932 г., с. 5—41; Н. К. Беляев, В. П. Рувинская, Н. Размадзе, Т. М. Гурская, К. А. Лоладзе, Эфендиев, К. В. Жуковская, Р. Аллавердян. Экспериментальное исследование пород и гибридов тутового шелкопряда. Результаты летних испытаний 1933 и 1934 гг., с. 42—59; Беляев Н. К. Метод двойных скрещиваний у тутового шелкопряда, с. 60—87. Н. К. Беляев, М. И. Какабад-зе. Реципрокные гибриды тутового шелкопряда, с. 88—104; Н. К. Беляев, Н. Размадзе. Наследование формы кокона межпородными гибридами первого поколения тутового шелкопряда, с. 105—108; Н. К. Беляев. Результаты применения в производстве опытных данных по исследованию пород и гибридов шелкопряда, с. 109—112.)
Гершензон С. М. «Мобилизационный резерв» внутривидовой изменчивости.— Журн. общ. биологии, 1941, т. 2, ЛГэ 1, с. 85—107.
Гершензон С. М. Роль естественного отбора в распространении и динамике меланизма у хомяков (Cricetus cricetus 13*
L.).— Журн. общ. биологии, 1946, т. 7, № 2, с. 97—130.
Гершензон С. М., Полевой В. В. Система скрещивания в природных популяциях хомяка (Cricetus cricetus L.).— ДАН СССР, 1940, т. 29, с. 608— 609.
Гинзбург Э. X. Генетическое описание наследования количественных признаков. I. О необходимости и конструктивности полигенной модели.— Генетика, 1982, т. 18, № 8, с. 1334—1342.
Гинзбург 3. X., Никоро 3. С. Генетическое описание наследования количественных признаков. II. Полигенная или олигогенная модели? — Генетика, 1982, т. 18, № 8, с. 1343—1352.
Головинская К- А. Гиногенез у рыб в исследованиях Д. Д. Ромашова.— В кн.: Из истории биологии. М.: Наука, 1975, вып. 5, с. 92—102.
Давиденков С. Н. Эволюционно-генетические проблемы в невропатологии. Л.: ГИДУ В, 1947. 382 с.
Данилевский Н. Я. Дарвинизм. Критическое исследование. СПБ.: М. Е. Комаров, т. 1, 1885, ч. 1. X 4-24-519 с., ч. 2. XVI4-530-1-148 с.; т. 2, 1889. 44-4-200 с.
Догель В. А. Олигомеризация гомологичных органов как один из главных путей эволюции животных. Л.: ЛГУ, 1954. 368 с.
Дубинин Н. П. Генетико-автоматические процессы и их значение для механизма органической эволюции.— Журн. эксперим. биол., 1931, т. 7, вып. 5—6, с. 463—479.
Дубинин Н. П. О некоторых основных проблемах генетики.— Биол. журн., 1932, т. 1, вып. 1—2, с. 112—146.
Дубинин Н. П. Экспериментальное исследование интеграции наследственных систем в процессах эволюции популяций.— Журн. общ. биологии, 1948, т. 9, № 3, с. 203—244.
Дубинин Н. П., Ромашов Д. Д. Генетическое строение вида и его эволюция. I. Генетико-автоматические процессы и проблема экогенотипов.— Биол. журн., 1932, т. 1, вып. 5—6, с. 52—95.
Дубинин Н. П., Тиняков Г Г Структурная изменчивость хромосом в популяциях города и сельской местности.— ДАН, 1946, т. 51, № 2, с. 151 — 154.
Дубинин Н. П., Тиняков Г Г Инверсии на границах экологических рас Drosophila funebris.— ДАН, 1947а, т. 55, № 7, с. 643—645.
Дубинин Н. П., Тиняков Г Г Экология города и распространенность инверсий у Drosophila funebris.— ДАН, 19476, т. 56, № 8, с. 865—867.
Дубинин Н. П., Гептнер М. А., Бес-
195
смертная С. Я. и др. Экспериментальный анализ экогенотипов Drosophila melanogaster. Ч. I, Ч. II.— Биол. журн., 1934, т. 3, вып. 1, с. 166—216.
Дубинин И. П., Гептнер М. А., Демидова 3. А., Дьячкова Л. И. Генетическая структура популяций и ее динамика в диких населениях Drosophila melanogaster.— Биол. журн., 1936, т. 5, в. 6, с. 939—961. (англ.— с. 962—976).
Дубинин Н. П., Ромашов Д. Д., Гептнер М. А., Демидова 3. А. Аберративный полиморфизм у Drosophila fasciata Meig. (sin.— melanogaster Meig.).— Биол. журн., 1937a, т. 6, вып. 2, с. 311—354.
Дубинин Н. П., Соколов Н. П., Тиняков Г Г Внутривидовая хромосомная изменчивость.— Биол. журн., 19376, т. 6, № 5—6, с. 1007—1054.
Дьяков Ю. Т. Роль иммунитета в селекции растений.— Природа, 1977, № 8, с. 72—79.
Завидовский М. М. Механика развития признаков пола и генетика (элементы морфогенетики).— Тр. Съезда по генетике и селекции. Л., 1930, т. 2, с. 45—69.
Иванов Ю. И., Фурман Д. П. Анализ зависимости между проявлениями признака на правой и левой сторонах тела для генов с билатеральным фенотипическим действием.— Генетика,
1975, т. 11, № 8, с. 66—76.
Иванова О. А. О наследовании многососковости у крупного рогатого скота.— Журн. эксперим. биологии (А), 1926, т. 2, вып. 4, с. 220—237, 252.
Ильин И. А. Распределение и наследование белых пятен у морской свинки..— Тр. Лабор. эксперим. биологии Моск, зоопарка, 1928, т. 4, с. 255—349.
Калабухов И. И. Изменчивость и массовое размножение.— Журн. общ. биологии, 1941, т. 2, № 3, с. 381—394.
Калмыкова Л. Г Наследственная гетерогенность болезней нервной системы. М.: Медицина, 1976. 319 с.
Камшилов М. М. Является ли плей-отропия свойством гена? — Биол. журн., 1935, т. 4, № 1, с. 113—144.
Карпенко Б. И. Н. С. Четвериков в жизни и науке.— Природа, 1974, № 2, с. 75—80.
Кашкаров Д. И. Адаптивна ли эволюция и что такое видовые признаки.— Зоол. журн., 1939, т. 18, вып. 4, с. 612— 630.
Кириков С. В. К вопросу о распространении черного хомяка и количественном отношении его к обыкновенной форме.— Зоол. журн., 1934, т. 13, № 2, с. 361—367.
Кирпичников В. С. К вопросу об
образовании рас у рыб (расы и эволюция).— Биол. журн., 1933, т. 2, вып. 6, с. 609—627.
Кирпичников В. С. Роль наследственной изменчивости в процессе естественного отбора (гипотеза о косвенном отборе).— Биол. журн., 1935, т. 4, вып. 5, с. 775—800.
Кирпичников В. С. О гипотезах наследственного закрепления модификаций.— Усп. соврем, биологии, 1944, т. 18, вып. 3, с. 314—339.
Кожевников Б. Ф. Экспериментальное получение кариотипической расовой изоляции.— Биол. журн., 1936, т. 5, с. 727—752.
Колмогоров А. Уклонения от формул Харди при частичной изоляции.— ДАН, 1935, т. 3 (8), № 3 (63), с. 129—132.
Колмогоров А. И. Об аналитических методах в теории вероятностей.— Усп. математ. наук, 1938, т. 5, с. 5—41.
Колмогоров А. И. Об одном новом подтверждении законов Менделя.— ДАН, 1940, т. 27, № 1, с. 38—42.
Колмогоров А. И., Петровский И. Г., Пискунов И. С. Исследование уравнения диффузии, соединенной с возрастанием количества вещества, и его приложение к одной биологической проблеме.— Бюл. МГУ (А), 1937, т. 1, вып. 6, с. 1—26.
Кольцов Н. К. Генетический анализ окраски морских свинок.— Изв. Ин-та эксперим. биологии. М.: Изд. Нарком-здрава, 1921, вып. 1, с. 87—97.
Кольцов Н. К. Родословные наших выдвиженцев.— Русск. евгенич. журн., 1926, т. 4, вып. 3—4, с. 103—143.
Кольцов Н. К. О работах института экспериментальной биологии в Москве.— Усп. эксперим. биологии, 1929, т. 8, вып. 1, с. 15—28.
Кольцов Н. К. Проблема прогрессивной эволюции.— Биол. журн., 1933, т. 2, вып. 4—5, с. 475—500.
Кольцов Н. К. Организация клетки. М.; Л.: Биомедгиз, 1936. 652 с.
Кольцов Н. К. Упехи советской науки в области биологии за два десятилетия.— Биол. журн., 1937, т. 6, № 5— 6, с. 929—946.
Кольцов Николай Константинович. Материалы к биобиблиографии ученых СССР. Вступит, статья Б. Л. Астаурова. М.: Наука, 1976. 76 с.
Кулагина О. С., Ляпунов А. А. К вопросу о моделировании эволюционного процесса.— Пробл. кибернетики. М.: Наука, 1966, вып. 16, с. 147—169.
Левонтин Р. Генетические основы эволюции. М.: Мир, 1978. 351 с.
Лепёхин И. И. Дневные записки путешествия доктора и Академии наук
196
адъюнкта Ивана Лепёхина по разным провинциям Российского государства, в 1768 и 1769 гг. СПб., 1795, ч. 1, 8-1-537 с., 16 л. ил.
Лукин Е. И. Дарвинизм и географические закономерности в изменении организмов. М.; Л.: Изд-во АН СССР, 1940. 308 с.
Лус Я. Я. О наследовании окраски и рисунка у божьих коровок Adalia bipunctata и Adalia decempunctata L.— Изв. Бюро по генетике, 1928, т. 6, с. 89—163.
Лус Я. Я. Анализ явления доминирования при наследовании рисунка элитр и переднеспинки у Adalia bipunctata L.— Тр. Лабор. генетики, 1932, т. 9, с. 125—162.
Лусис Я. Я. О биологическом значении полиморфизма окраски у двухточечной коровки — Adalia bipunctata L.— In: Latvijas Entomologs. Riga: LZAI, 1961, T. 4, c. 3—29.
Любищев А. А. О природе наследственных факторов (критическое исследование).— Изв. Биол. НИИ при Пермском гос. ун-те, 1925, т. 4, Прилож. 1, с. 1 — 142.
Ляпунов А. А. О рассмотрении биологии с позиции изучения живой природы как большой системы.— В кн.: Проблемы методологии системного исследования. М.: Мысль, 1970, с 184— 226.
Малиновский А. А. Роль генетических и феногенетических явлений в эволюции вида. I. Плейотропия.— Изв. АН СССР. Отд. биол., 1939, № 4, с. 575— 614.
Малиновский А. А. Роль хромосомных инверсий в эволюции вида.— Журн. общ. биологии, 1940, т. 1, № 4, 565- 596.
Малиновский А. А. Объединение полезных признаков в процессе естественного отбора. Типы взаимодействий и их значение в организме.— В кн.: Рефераты работ учреждений Отд. биол. наук АН СССР за 1941 —1943 гг. М.; Л.: Изд-во АН СССР, 1945а, с. 291 — 293.
Малиновский А. А. Физиологические источники корреляций в строении человеческого организма.— Журн. общ. биологии, 19456, т. 6, № 4, с. 235—257.
Малиновский А. А. Хромосомные инверсии как фактор накопления эволюционных различий. Учет миграций в экспериментах в природе. Вид как эволюционное целое.— В кн.: Рефераты н.-и. работ. Отд. биол. наук. АН СССР за 1945 г. М.; Л.: Изд во АН СССР, 1947, с. 283—287.
Малиновский А. А. Элементарные корреляции и изменчивость человече
ского организма.— Тр. Ин-та цитологии, гистологии и эмбриологии. М.; Л.: Изд-во АН СССР, 1948, т. 2, вып. 1, с. 136—198.
Малиновский А. А. Типы управляющих биологических систем и их приспособительное значение.— Пробл. кибернетики. М.: Физматгиз, 1960, вып. 4, с. 151-182.
Малиновский А. А. Незавершенные идеи некоторых советских генетиков.— Природа, 1970, № 2, с. 79—83.
Машковцев А. А. Смена эндогенных и экзогенных факторов эмбрионального развития в онтогенезе и филогенезе. (Эволюция онтогенеза).—Изв. Отд. математ. и естеств. наук АН СССР. Сер. биол., 1936, вып. 5, с. 945—997.
Медведев Н. Н. Юрий Александрович Филипченко (1882—1930). М.: Наука, 1978. 103 с.
Морган Т. Г Теория гена. Л.: Сеятель, 1927. 312 с.
Муретов Г Д. Физиологические мутации и динамика генного состава популяций Drosophila melanogaster.— ДАН, 1939, т. 24, № 5, с. 481—484.
Нидхэм Дж. Нидхэм о биологии.— Брит, союзник, 15 июля 1945 г., № 28 (153).
Никоро 3. С., Гусев С. Н. Экспериментальная проверка действия генетико-автоматических процессов в популяциях.— Биол. журн., 1938, т. 7, № 1, с. 197—216.
Никоро 3. С., Рогозянова А. И. О взаимодействии генетико-автоматических процессов и естественного отбора.— Биол. журн., 1938, т. 7, вып. 5—6, с. 1139—1144.
Никоро 3. С., Гусев С. Н., Павлов Е., Грязнов И. Закономерности половой изоляции у некоторых линий Drosophila melanogaster.— Биол. жури., 1935, т. 4, вып. 3, с. 569—585.
Новоженов Ю. И. Размеры природных популяций насекомых.— В кн.: Фауна Урала и Европейского Севера. Свердловск: Уральский гос. ун-т, 1981, с. 83—102.
Оленов Ю. М. О влиянии предшествующей истории вида на его дальнейшее развитие.— ДАН, 1941, т. 31, № 2, с. 157—160.
Оленов Ю. М. Явления фенотипического параллелизма в природных популяциях Drosophila melanogaster.— Генетика, 1965, № 2, с. 87—94.
Оленов Ю. М., Хармац И. С., Гал-ковская К. Ф., Муретов Г. Д. Факторы, ответственные за генный состав диких популяций Drosophila melanogaster.— ДАН, 1939, т. 24, № 5, с. 466—470.
Орлов Л. М. Жилкование крыла
197
златоглазки как модель микроэволюци-онных исследований.— Жури. общ. биологии, 1975, т. 36, № 6, с. 902—913.
Панина К. А. Анализ массового изменения «добавочные щетинки» у Drosophila fasciata Meig. (melanogaster).— Бйол. журн., 1938, т. 7, вып. 1, с. 217—222.
Пирсон К. Грамматика науки. СПб.: Шиповник, 1911. 655 с.
Плавильщиков Н. Н. Очерки по истории зоологии. М.: Учпедгиз, 1941. 296 с.
Промптов А. Н. Плейотропная гено-вариация «polymorpha» у Drosophila funebris Fabr. (К вопросу о множественном выражении гена). — Журн. экспе-рим. биологии, 1929, т. 5, вып. 3—4, с. 149—181.
Промптов А. Н. Географическая изменчивость песни зяблика (Fringilla coelebs L.) в связи с общими вопросами сезонных перелетов птиц.— Русск. зоол. журн., 1930, т. 10, вып. 3, с. 17—42.
Промптов А. Н. Ген и признак в онтогенезе.— Усп. соврем, биологии, 1934а, т. 3, вып. 2, с. 145—180.
Промптов А. Н. Эволюционное значение миграций птиц.— Зоол. журн., 19346, т. 13, № 3, с. 409—436.
Промптов А. Н. Эколого-генетические факторы эволюционной дивергенции у птиц.— Биол. журн., 1936, т. 5, вып. 6, с. 1001 — 1010.
Протоколы Эволюционной бригады. Тетр. 1 (заседания 1—36, 1934—1937 гг.); тетр. 2 (заседания 37—104, 1937— 1940 гг.). Рукопись.
Пузанов И. И. Мои воспоминания о Сергее Сергеевиче Четверикове (с дополнениями и примечаниями Н. С. Четверикова от 19.V. 1965, с. 22— 24). Рукопись, 1965. 24 с.— Арх. АН СССР, ф. 1650.
Работы Института экспериментальной биологии Наркомздрава. К XVII съезду ВКП(б).— Биол. журн., 1934, т. 3. № 1, с. 217—232.
Рапопорт А. Н. Феногенетический анализ независимой и зависимой дифференцировки.— Тр. Ин-та цитологии, гистологии и эмбриологии. М.; Л.: Изд-во АН СССР, 1948, т. 2, вып. 1, с. 3— 135.
Рокицкий П. Ф. Генетический анализ числа грудных щетинок у Drosophila melanogaster.— Журн. эксперим. биологии (А), 1927, т. 3, вып. 3, с. 171 —187.
Рокицкий П. Ф. Поле действия гена. Изучение редуцирующих и прибавляющих щетинки генов у Drosophila melanogaster.— Журн. эксперим. биологии, 1929, т. 5, вып. 3—4, с. 182—214.
Рокицкий П. Ф. О взаимодействии 198
генов. I. Изучение некоторых комбинаций генов, влияющих на щетинки у Drosophila melanogaster с помощью полей их действия.— Журн. эксперим. биологии, 19296, т. 5, вып. 3—4, с. 215— 226.
Рокицкий П. Ф. Ген и признак (обзор).— Журн. эксперим. биологии, 1931, т. 7, вып. 2, с. 172—186.
Рокицкий П. Ф. Экспериментальный анализ проблемы рентгеноселекции. I. Сравнение темпов отбора в рентгенизируемых и нерентгенизируемых линиях.— Усп. зоотехн. наук, 1936, т. 2, вып. 2, с. 161—202.
Рокицкий П. Ф. Изучение процесса искусственного отбора при естественной и экспериментально вызванной рентгеновским облучением изменчивости.— Бюл. МОИП. Отд. биол., 1959, т. 64, вып. 4, с. 75—97.
Рокицкий П. Ф. Некоторые статистические явления в феногенезе.— В кн.: Теоретические и экспериментальные исследования по математической генетике. Минск: Наука и техника, 1973, с. 114— 128.
Рокицкий П. Ф. С. С. Четвериков и эволюционная генетика.— Природа, 1974, № 4, с. 70—74.
Рокицкий П. Ф. С. С. Четвериков и развитие эволюционной генетики.— В кн.: Из истории биологии. М.: Наука, 1975, вып. 5, с. 63—75.
Рокицкий П. П., Васина-Попова Е. Т. Развитие генетики сельскохозяйственных животных в СССР.— В кн.: Историко-биологические исследования. М.: Наука, 1978, вып. 6, с. 5—27.
Ромашов Д. Д. Математическое выражение законов Менделя.— Журн. эксперим. биологии (А), 1925, т. 1, вып. 3—4, с. 177—190.
Ромашов Д. Д. Об условиях «равновесия» в популяциях.— Журн. эксперим. биологии, 1931, т. 7, вып. 4, с. 442— 454.
Ромашов Д. Д. Изменчивость жилкования в популяциях Drosophila funebris. 1948. Рукопись (6 с., библиогр. 8 назв., 2 ил.).
Ромашов Д. Д., Балкашина Е. И. Материалы по генетике Drosophila funebris F. I, II — Журн. эксперим. биологии, 1929, т. 5, вып. 2, с. 102—146; 1930, т. 6, вып. 3, с. 201—219.
Ромашов Д. Д., Беляева В. Н. Об изменчивости жилкования в популяциях комнатной мухи (Musca domestica L.).— ДАН, 1946, т. 54, № 9, с. 829—832.
Ромашов Д. Д., Беляева В. Н. О «полях» изменчивости жилкования крыла в популяциях Diptera. (Доклад, читанный на II Генетич. конф. МГУ 25 марта 1947 г.). 1947. Рукопись (25 с.).
Ромашов Д. Д., Беляева В. И. О некоторых закономерностях в изменчивости жилкования у Diptera (Brachyce-га). (Доклад, читанный на кафедре энтомологии МГУ проф. Е. С. Смирнова весной 1948 г.). 1948. Рукопись
(20 с.). '
Ромашов Д. Д., Ильина Е. Д. Анализ популяций лисицы по формуле Гарди.— ДАН, 1942, т. 37, № 5—6, с. 220— 224.
Ромашов Д. Д., Ильина Е. Д. Об аберративной изменчивости в популяциях пушных зверей.— Журн. общ. биологии, 1943, т. 4, № 5, с. 286—312.
Семенов-Тян-Шанский А. П. Таксономические границы вида и его подразделений. Опыт точной категоризации низших систематических единиц.— Зап. имп. АН по физ.-мат. отд., сер. 8, 1910, т. 25, № 1, 114-29 с.
Серебровский А. С. Теория наследственности Моргана и Менделя и марксисты.— Под знаменем марксизма, 1926, № 3, с. 98—117.
Серебровский А. С. Генетический анализ популяции домашних кур горцев Дагестана.— Журн. эксперим. биологии (А), 1927, т. 3, вып. 1—2, с. 62— 124; вып. 3—4, с. 125—146, 201—202.
Серебровский А. С. Геногеография и генофонд сельскохозяйственных животных СССР.— Научное слово, 1928, № 9, с. 3—23.
Серебровский А. С. Проблемы и метод геногеографии.— Тр. Съезда по генетике и селекции, 1930, т. 2, с. 71—86.
Серебровский А. С. Геногеография кур Армении.— Усп. зоотехн. наук, 1935, т. 1, вып. 3, с. 317—348.
Серебровский А. С. Генетический анализ. М.: Наука, 1970. 342 с.
Серебровский А. С. Некоторые проблемы органической эволюции. М.: Наука, 1973. 168 с.
Серебровский А. С. Избранные труды по генетике и селекции кур. М.: Наука, 1976. 404 с.
Сидоров Б. И. Мозаики по yellow в линии scute8 I. К проблеме существования поля действия гена.— Биол. журн., 1934, т. 3, вып. 4, с. 737—739.
Соколов Н. Н. Взаимодействие ядра и цитоплазмы при отдаленной гибридизации животных. М.: Изд-во АН СССР, 1959. 147 с.
Соколов И. И., Дубинин И. П. Хромосомные мутации в популяциях как основа эволюции кариотипа.— ДАН, 1940, т. 29, № 5—6, с. 405—407.
Сутт Т. Проблема направленности органической эволюции. Таллин,- 1977. 139 с.
Сушкин П. П. Обратим ли процесс эволюции? — В кн.: Новые идеи в био
логии. Пг.: Образование, 1915, сб. 8, с. 1—39.
Тимирязев К. А. Чарлз Дарвин.— В кн.: Памяти Дарвина. М.: Научное слово, 1910, с. 16—38.
Тимофеев-Ресовский И. В. О фенотипическом проявлении генотипа. I. Геновариация radius incompletus у Drosophila funebris. Журн. эксперим. биологии (А), 1925, т. 1, вып. 3—4, с. 93—142.
Тимофеев-Ресовский И. В. Влияние температуры на образование поперечных жилок на крыльях одной геновариа-ции у Drosophila funebris.— Журн. эксперим. биологии (А), 1928, т. 4, вып. 3—4, с. 199—214.
Тимофеев-Ресовский И. В. О фенотипической реализации гена vti у Drosophila funebris.— Тр. Съезда по генетике и селекции. Л., 1930, т. 2, с. 483— 488.
Тимофеев-Ресовский И. В. Микроэволюция. Элементарные явления, материал и факторы микроэволюционного процесса.— Ботан. журн., 1958, т. 43, № 3, с. 317-336.
Тимофеев-Ресовский Н. В. О микро-и макрофилогении у половых перекрест -нооплодотворяющихся организмов. К теории вида Возникновение малых мутаций относительной жизнеспособности и селекция генов-модификаторов в результате рентгеновского облучения дрозофилы.— В кн.: Радиационная цитогенетика и эволюция. Свердловск, 1965, вып. 44, с. 5—9, 11—25, 65—78. (Труды Ин-та биол. УФАН.)
Тимофеев-Ресовский И. В. Структурные уровни биологических систем.— В кн.: Системные исследования. Ежегодник. М.: Наука, 1970, с. 80—113.
Тимофеев-Ресовский И. В. С. С. Четвериков и популяционная генетика.— В кн.: Докл. МОИП. Общая биология. М.: Изд-во МГУ, 1978, с. 96—99.
Тимофеев-Ресовский И. В., Воронцов И. И., Яблоков А. В. Краткий очерк теории эволюции. М.: Наука, 1969 (2-е изд.: 1977 г.). 407 с.
Тимофеев-Ресовский И. В., Гинтер Е. К., Иванов В. И. О некоторых проблемах и задачах феногенетики.— В кн.: Проблемы экспериментальной биологии. (Сборник памяти Б. Л. Астаурова). М.: Наука, 1977, с. 186—195.
Тимофеев-Ресовский И. В., Иванов В. И. Некоторые вопросы феногенетики.— В кн.: Актуальные вопросы современной генетики. М.: Изд-во МГУ, 1966, с. 114—136.
Тимофеев-Ресовский И. В., Свире-жев Ю. М. Об адаптационном полиморфизме в популяциях Adalia bipunctata L.— Проба, кибернетики. М.: Наука, 1966, вып. 16, с. 137—146.
199
Тимофеев-Ресовский Н. В.. Тимофеева-Ресовская Е. А., Циммерман К. Экспериментально-системэтический анализ географической изменчивости и формообразования у Epilachna chryso-melina F. (Coleoptera, Coccinellidae).— В кн.: Радиационная цитогенетика и эволюция. Свердловск, 1965, вып. 44, с. 27—63. (Труды Ин-та биол. УФАН.)
Тимофеев-Ресовский Н. В., Тюрюка-нов А. Н. Об элементарных биохорологических подразделениях биосферы.— Бюл. МОИП. Отд. биол., 1966, т. 71, вып. 1, с. 123—132.
Тимофеев-Ресовский И. В., Яблоков А. В. Фены, фенетика и эволюционная биология.— Природа, 1973, № 5, с. 40—51.
Тимофеев-Ресовский Н. В., Яблоков А. В., Глотов Н. В. Очерк учения о популяциях. М.: Наука, 1973. 277 с.
Тимофеева-Ресовская Е. А. Влияние рентгеновских лучей на возникновение геновариаций у Drosophila funebris.— Журн. эксперим. биологии, 1930, т. 6, вып. 1, с. 9—14.
Тиняков Г Г Карты хромосом слюнных желез Drosophila funebris.— Бюл. МОИП. Отд. биол., 1965, т. 70, вып. 4, с. 141 — 144.
Уоддингтон К. X. Канализация развития и наследование приобретенных признаков.— Усп. соврем, биологии, 1944, т. 18, вып. 3, с. 393—396.
Уоддингтон К. X. Организаторы и гены. М.: ИЛ, 1947. 240 с.
Уоллэс А. Дарвинизм. Изложение теории естественного подбора и некоторых из ее приложений. М.: Библиотека для самообразования, 1898, т. 15. 753 с.
Филимонов П. Н. Оскар Фогт (Oskar Vogt), 1870—1959.—Журн. невропатологии и психиатрии им. С. С. Корсакова, 1960, т. 60, вып.12, с. 1672—1674.
Филиппенко Ю. А. О параллелизме в живой природе.— Усп. эксперим. биологии, 1924, т. 3, № 3—4, с. 242—258.
Филиппенко 10. А. Изменчивость и методы ее изучения. 4-е изд. М.: Госиздат, 1929. 275 с. (5-е изд. М.: Наука, 1978. 238 с.).
Филиппенко Ю. А. Эволюционная идея в биологии. 3-е изд. М.: Наука, 1977. 227 с.
Холодковский И. А. О биологических видах.— Изв. имп. Акад, наук, 6-я сер., 1910, т. 4, № 10, с. 751—770.
Хэксли Дж. Хэксли о теории эволюции.— Брит, союзник, 15 июля 1945 г., № 28 (153).
Царапкин С. Р. О направленной изменчивости у Coccinellidae. I. Анализ первого периода пигментации.—
200
Журн. эксперим. биологии, 1930, t. 6, вып. 2, с. 139—155.
Четвериков И. С. Статистические исследования. М.: Наука, 19^5. 388.
Четвериков С. С. Волны жизни. (Из лепидоптерологических наблюдений за лето 1903 г.).— Дневник Зоол. отд. имп. О-ва любителей естествознания, антропологии и этнографии, 1905, т. 3, № 6, с. 106—ПО. Переизд.: Природа, 1980, № 11, с. 95—99.
Четвериков С. С. О некоторых моментах эволюционного процесса с точки зрения современной генетики.— Журн. эксперим. биологии (А), 1926, т. 2, вып. 1, с. 3—54; вып. 4, с. 237—240. (Переизд.: Бюл. МОИП. Отд. биол., 1965, т. 70, вып. 4, с. 34—75. В кн.: Классики советской генетики, 1920—1940. Л.: Наука, 1968, с. 133—170. Сокращенный англ, перевод Ф. Г. Добржанского: Cold Spring Harbor Symp., 1959, vol. 24, p. 27—30; полный перевод с пред. И. М. Лернера: Proc. Amer. Philos. Soc., 1961, vol. 105, № 2, p. 167—195. Французский перевод: S.S. Tchetverikoff. Les Lois de I’Heredite. Mont-Pelerine, 1970. 71 p.).
Четвериков С. С. Биометрия.— БСЭ, 1-е изд., 1927a, t. 6, c. 338—344.
Четвериков С. С. Теоретическая предпосылка генотипического анализа видов рода Drosophila. Тр. II Съезда зоологов, анатомов и гистологов (Москва, 4—10 мая 1925 г.). М.: Изд. Главнаука, 19276, с. 163—164.
Четвериков С. С. Экспериментальное решение одной эволюционной проблемы.— Тр. III Съезда зоологов, анатомов и гистологов (Ленинград, 14—20 декабря 1927 г.). Л.: Изд. Главнаука, 1928, с. 52—54.
Четвериков С. С. Из воспоминаний. (Публикация Б. Л. Астаурова).— Природа, 1974, № 4, с. 68—69.
Четвериков С. С. Воспоминания. (Публикация В. В. Бабкова, предисловие Б. Н. Сидорова).— Природа, 1980, № 5, с. 50—55, № 11, с. 86—94, № 12, с. 76—85.
Четвериков С. С. О некоторых моментах... (1926). Глава V. Экспериментальное исследование геновариаций, скрытых в популяциях (1929). (Публикация и предисловие В. В. Бабкова).— Генетика, 1982, т. 18, № 3, с. 341—348.
Четвериков С. С. Проблемы общей биологии и генетики (воспоминания, статьи, лекции). Новосибирск: Наука, 1983. 273 с.
Четвериков С. С. Фауна и биология чешуекрылых. Новосибирск: Наука, 1984. 102 с.
Чтения памяти Н. В. Тимофеева-
Ресовского/Ред. Р. Р. Атаян. Ереван: Изд-во АН Армянской ССР, 1983. 248 с.
Шмальгаузен И. И. Современные задачи феногенетики.— Изв. Отд. ма-темат. и естеств. наук АН СССР. Сер. биол., 1937, № 3, с. 895—906.
Шмальгаузен И. И. Организм как целое в индивидуальном и историческом развитии. М.; Л.: Изд-во АН СССР, 1938. 144 с. (2-е изд.: М.: Наука, 1982. 383 с.).
Шмальгаузен И. И. Пути и закономерности эволюционного процесса. М.; Л.: Изд-во АН СССР, 1939. 231 с. (2-е изд.: ^М.: Наука, 1983. 360 с.).
Шмальгаузен И. И. Возникновение и преобразование системы морфогенетических корреляций в процессе эволюции.— Журн. общ. биологии, 1940, т. 1, № 3, с. 349—370.
Шмальгаузен И. И. Факторы эволюции. (Теория стабилизирующего отбора). М.; Л.: Изд-во АН СССР, 1946. 396 с. (2-е изд.: М.: Наука, 1968. 451 с.).
Шмальгаузен И. И. Кибернетические вопросы биологии. Новосибирск: Наука, 1968. 224 с.
Шредингер Э. Что такое жизнь? с точки зрения физики. Перевод и послесловие А. А. Малиновского. М.: ИЛ, 1947. 146 с.
Эфроимсон В. П. О некоторых проблемах накопления и действия деталей.— Биол. журн., 1932а, т. 1, № 3—4, с. 87—102.
Эфроимсон В. П. О скорости мутационного процесса у человека. Рукопись, 19326, 16 с.
Эфроимсон В. П. Измерение скорости мутационного процесса у тутового шелкопряда.— Изв. АН СССР. Сер. биол. наук, 1940, № 5, с. 688—705.
Эфроимсон В. П. Проблемы генетики, селекции и гибридизации тутового шелкопряда: Дис. ... докт. биол. наук, т. 1 и 2. Харьков, 1946. 661 с.
Эфроимсон В. П. Становление типов конституции в связи с наследственной изменчивостью обмена и гормональных функций.— В кн.: Проблемы медицинской генетики. Л.: Медицина, 1965, с. 208—230.
Эфроимсон В. П. Генетика шизофрении. Гл. VII.— В кн.: Эфроимсон В. П., Блюмина М. Г Генетика олигофрений, психозов, эпилепсий. М.: Медицина, 1978, 185—282.
Юл. Дж. Эдни, Кендел М. Дж. Теория статистики. М.: Госстатиздат, 1960. 779 с.
Adams М. В. The founding of population genetics: Contribution of the
Chetverikov school,1924—1934.—J. Hist. BioL, 1968, vol. 1, № 1, p. 23—40.
Adams M. B. Towards a synthesis: Population concepts in Russian evolutionary thought, 1925—1935.—J. Hist. Biol., 1970, vol. 3, № 1, p.107—129.
Allen G. E. Opposition to the Mende-lian—chromosome theory: The physiological and developmental genetics of Richard Goldschmidt.— J. Hist. BioL, 1974, vol. 7, № 1, p. 49—92.
Allen G. E. Thomas Hunt Morgan: The man and his science. Princeton (N. J.): Univ, press, 1978. XV1I+447 p.
Allen G. E. Naturalists and experimentalists: The genotype and the phenotype.— Stud. Hist. Biol., 1979, vol. 3, p. 179—209.
Astauroff B. L. Studien uber die erbliche Veranderung der Halteren bei Drosophila melanogaster Sehin.— Roux’Arch. Entwicklungsmech. Organ., 1929, Bd. 115, H. 3, S. 424—447.
Astauroff B. L. Analyse der erblichen Stdrungsfalle der bilateralen Symmetric im Zussammenhang mit der selbststan-digen Variabilitat ahniicher Struktu-ren.— Ztschr. indukt. Abstam-mungs- und Vererbungslehre, 1930, Bd. 55, H. 3, S. 183—262.
Ayala F. J. Introduction.— In: Sudies in the philosophy of biology/Ed. F. J. Ayala, T. Dobzhansky. N.Y.: Macmillan, 1974, p. VII—XVI.
Babkoff V. Sakharov Vladimir Vladimirovich.— In: Diet, of Sci. Biogr., 1975, vol. 12, p. 77—78.
Babkoff V. Chetverikov’s path to genetical explanation of evolutionary process: Creation of a new field and creation of a new scientific community.— In: Proc. 15 Intern, congr. hist. sci. Edinburgh, 1977, p. 133.
Babkoff V. The central problem of evolutionary genetics.— In: Evolution and environment: 3rd intern, meet., Abstrs. Brno, 1981a, p. 3.
Babkoff V S. S. Tsetverikov — evolu-utiogenetiikan uranuurtaja. — Tiede ja Edistys, 1981b, vol. 6, № 4, p. 58—61.
Babkoff V., Cetverikov S.— In: Bro-ckhaus ABC Forscher und Erfinder. Leipzig: Brockhaus, 1984, S.
Bachmetjew P. Experimentelle ento-mologische Studien. Bd. I. Temperatu-rverhaltnisse bei Insekten. Leipzig, 1901. 160 S.; Bd. II. Einflup der auBeren Faktoren auf Insekten. Sophia, 1907. 1068 S.
Balvaschina E. /. Ein Fall der Erb-homdosis (die Genovafiation «Aristopedia») bei Drosophilla melanogaster.— Roux’ Arch. Entwicklungsmech. Organ., 1929, Bd. 115, H. 3, S. 448—463.
201
14 В. В. Бабков
Bateson W. Materials for the study of variation, treated with especial regard to discontinuity in the origin of species. L.: Macmillan, 1894. 598 p.
Bauer H., Timofeeff-Ressovsky N. W. Genetik und Evolutionsforschung bei Tieren.— In: Die Evolution der Orga-nismen/Hrsg. G. Heberer. Jena: G. Fischer, 1943, S. 335—429.
BeUajeff N. K. Die Chromosomen-komplexe und ihre Beziehung zur Phylo-genie bei den Lepidopteren: (Mit 43 Figuren).—Ztschr. indukt. Abstam-mungs- und Vererbungslehre, 1930, Bd. 54, H. 314, S. 369—399.
Belyaev D. K. Destabilizing selection as a factor in domestication.— J. Hered., 1979, vol. 70, p. 301—308.
Braun W. Experimental evidence on the production of the mutant «aristape-dia> by a change of developmental velocities.— Genetics, 1940, vol. 25, p. 143—149.
Bridges С. B., Morgan T. H. The second-chromosome group of mutant characters.— Carnegie Inst. Wash. Publ., 1919, N 278, p. 123—304.
Bridges С. B., Morgan T. H. The third-chromosome group of mutant characters.— Carnegie Inst. Wash. Publ., 1923, № 327, p. 1—251.
Castle W. The laws of heredity of Galton and Mendel, and some laws governing race improvement by selection.— Proc. Amer. Acad. Arts and Sci., 1903, vol. 39, p. 223—249.
Cock A. G. William Bateson, Mende-lism and biomery.— J. Hist. Biol., 1973, vol. 6, № 1, p. 1—36.
Coleman W. Bateson and chromosomes: Conservative thought in science.— Centaurus, 1970, vol. 15, p. 228—318.
Dobzhansky Th. Genetics and the origin of species. N. Y.: Columbia Univ, press, 1937. XVI+364 p.; 1941. XVIII + + 446 p.; 1951. 3-J-364 p. (ed. 1—3).
Dobzhansky Th. Experimental studies on genetics of freeliving populations of Drosophila.— Biol. Rev., 1939, vol. 14, p. 339—368.
Dobzhansky Th. Genetics of natural populations. IX. Temporal changes in the composition of populations of Drosophila pseudoobscura.— Genetics, 1943, vol. 28, p. 162—186.
Dobzhansky Th. A review of some fundamental concepts and problems of population genetics.— Cold Spring Harbor Symp. Quant. Biol., 1955, vol. 20, p. 1 — 15.
Dobzhansky Th. Evolution of genes and genes in evolution.— Cold Spring Harbor Symp. Quant. Biol. 1959, vol. 24, p. 15—30.
202
Dobzhansky Th. Sergei Sergeevich Tchetverikov, 1880—1959.— Genetics, 1967, vol. 55, №1, p. 1—3.
Dobzhansky Th. Genetics of the evolutionary process. N. Y.; L.: Columbia Univ, press, 1970. XIII-|-505 p.
Dobzhansky Th. Evolutionary oscillations in Drosophila pseudoobscura.— In: Ecological genetics and evolution. Oxford: Blackwell, 1971, p. 109—133.
Dobzhansky Th. The roving naturalist. Travel letters of Theodosius Dob-zhansky/Ed. and with introd, by B. Glass. Philadelphia, 1980. 327 p. (Mem. Amer. Philos. Soc.; Vol. 139).
Dobzhansky Th. Dobzhansky’s genetics of natural populations. I— XLIII/Eds. R. C. Lewontin et al. N. Y.: Columbia Univ, press, 1981. XIV-|-942 p.
Dobzhansky Th., Epling C. Contributions to the genetics, taxonomy, and ecology of Drosophila pseudoobscura and its relatives.— Carnegie Inst. Wash. Publ., 1944, № 554, p. 1 — 183.
Dobzhansky Th., Queal M. L. Genetics of natural populations. I. II.— Genetics, 1938, vol. 23, p. 239—251, 463— 484.
Dobzhansky Th., Sturtevant A. H. Inversions in the chromosomes of Drosophila pseudoobscura.— Genetics, 1938, vol. 23, p. 28—64.
Driver E. C. The temperature effective period — the key to eye facet number in Drosophila.— J. Exp. Zool., 1926, vol. 46, № 3, p. 317—332.
Dubinin N. P., Sokolov N. N., Tiniakov G. G. Occurence and distribution of chromosome aberrations in nature (Diptera).— Nature, 1936, vol. 137, № 3477, p. 1035—1036.
Dubinin N. P., Tiniakov G. G. Structural chromosome variability in urban and rural populations of Drosophila funebris.— Amer. Natur. 1964a, vol. 80, № 792, p. 393—396.
Dubinin N. P., Tiniakov G. G. Seasonal cycle and inversion frequency in populations.— C. r. (Dokl.) Acad. sci. URSS, 1946b, t. 52, № 1, p. 77—79.
Dubinin N. P., Tiniakov G. G. Inversion gradients and natural selection in ecological races of Drosophila funebris.— Genetics, 1946c, vol. 31, № 6, p. 537—545.
Dubinin N. P., Tiniakov G. G. Natural selection in experiments wifh population inversions.— J. Genet., 1947, vol. 48, № l,op. 11— 15.
Ellegdrd A. Darwin and the general reader: The reception of Darwin’s theory of evolution in the British periodical press, 1859—1872. Goteborg: Elanders Bohtrycheri Aktiebolag, 1958. 394 p.
Elton C. S. Periodic fluctuations in the numbers of animals, their causes and effects.— Brit. J. Exp. Biol., 1924, vol. 2, p. 119—163.
Elton Ch. Animal ecology. L.: Sidg-wick and Jackson, 1927. XXI-I-1-I-208 p. 2nd ed., 1966. XVI+ 207 p.
Elton Ch. Animal ecology and evolution. Oxford: Clarendon press, 1930. 96 p.
Farrall L. Controversy and conflict in science: A case study — the English biometric school and Mendel’s laws.— Soc. Stud. Sci., 1975, vol. 5, p. 269—301.
Fisher R. A. The correlation between relatives on the supposition of Mende-lian inheritance.— Trans. Roy. Soc. Edinburgh, 1918, vol. 52, p. 399—433.
Fisher R. A. On the dominance ratio.— Proc. Roy. Soc. Edinburgh, 1922, vol. 42, p. 321—341.
Fisher R. A. On some objections to mimicry theory: Statistical and genetic.— Trans. Entomol. Soc. London, 1927, vol. 75, p. 269—278.
Fisher R. A. The genetical theory of natural selection. Oxford: Clarendon press, 1930. 2nd ed. N. Y.: Dover, 1958. 291 p.
Fisher R. A. The evolution of dominance.— Biol. Rev., 1931, vol. 6, p. 345— 368.
Fisher R. A. The use of multiple measurements in taxonomic problems.— Ann. Eugenics, 1936, vol. 7, p. 179—188.
Fisher R. A., Ford E. B. The Sewall Wright effect.— Heredity, 1950, vol. 4, p. 117— 119.
Ford E. B. Polymorphism and taxonomy.— In: The new systematics/Ed. J. S. Huxley. L.: Oxford Univ, press, 1940, p. 493—513.
Ford E. B. Polymorphism.— Biol. Rev., 1945, vol. 20, № 2, p. 73—88.
Fothergill P. G. Historical aspects of organic evolution. L.: Hollis and Carter, 1952. XVII+ 427 p.
Frolowa S. L., Astaurow B. L. Die Chromosomengarnitur als systemati-sches Merkmal. (Eine vergleichende Untersuchung der russischen und ame-rikanischen Drosophila obscura Fall.). — Ztschr. Zellforsch., 1929, Bd. 10, H. 1, S. 201—213.
Garcia-Вellido A. Homoeotic and ata-vic mutions in insects.— Amer. Zool., 1977, vol. 17, № 3, p. 613—629.
Genetics, paleontology and evolu-tion/Eds. Jepsen G. L., Мауг E., Simpson G. G. Princeton (N. J.): Princeton Univ, press, 1949. XVI+ 474 p.
Gershenson S. Mutant genes in a wild population of Drosophila obscura Fall.— Amer. Natur, 1934, vol. 68, № 719, p. 569—571.
Gershenson S. Evolutionary studies on the distribution and dynamics of melanism in the hamster (Cricetus cricetus L.). I, II.—Genetics, 1945, vol. 30, № 3, p. 207—251.
Glass B. Bentley Glass on evolution.—Isis, 1981, vol. 72, № 264 (4), p. 642—647.
Goldschmidt R. Physiological genetics. N. Y.: McGraw-Hill, 1938. 338.
Goldschmidt R. The material basis of evolution. New Haven: Yale Univ, press, 1940. 436 p.; 2nd ed., 1982. XLII + 436 p.
Goldschmidt R. Pricking a bubble.— Evolution, 1953, vol. 7, p. 264—269.
Goldschmidt R. B. Portraits from memory. Seattle: Univ. Wash, press, 1956. IX+181 p.
Goldschmidt R. B. In and out the ivory tower. Seattle: Univ. Wash, press, 1960. XIII + 352 p.
Goodale H. D. Can artificial selection produce unlimited change? — Amer. Natur., 1937, vol. 71, № 736, p. 433—459.
Gordon C. The frequency of heterozygosis in free-living populations of Drosophila melanogaster and Drosophila subobscura.— J. Genet., 1936, vol. 33, № l,p. 25—60.
Gordon C., Spurway H., Sreet P. A. R. An analysis oi three wild populations of Drosophila subobscura.— J. Genet., 1939, vol. 38, p. 37—90.
Gulick J. T. The variation of species as related to their geographical distribution illustrated by the Achatinelli-dae.—Nature, 1872, vol. 6, p. 222—224.
Gulick J. T On diversity of evolution under one set of external conditions.— J. Linnean Soc. (Zool.), 1873, vol. 11, p. 496—505.
Gulick J. T. Divergent evolution through cumulatige segregation.— J. Linnean Soc. (Zool.), 1888, vol. 20, p. 189—274.
Gulick J. T. Evolution, racial and habitudinal.— Carnegie Inst. Wash. Publ., 1905, № 25, p. 1—269.
Guthrie J. The asymmetry of the small-eyed condition in «eyeless» Drosophila.— J. Exp. Zool., 1925, vol. 42, № 2, p. 307—314.
Haecker V Entwicklungsgeschichtli-che Eigenschaftsanalyse. (Phanogene-tik.) Gemeinsame Aufgaben der Entwi-cklungsgeschichte. Jena: G. Fischer, 1918. X + 344 S.
Haecker V Aufgaben und Ergebnisse der Phanogenetik.— In: Bibliographia Genetica Hrsg. J. P Lotsy, H. N. Kooi-man. The Hague: Martinus Nijhoff, 1925, Bd. 1, S. 93—314.
Hagedoorn A. L., Hagedoorn A. C.
203
The relative value of the processes causing evolution. The Hague: Martinus Nijhoff, 1921. 294 p.
Haldane J. B. S. The rate of spontaneous mutation of a human gene.— J. Genet., 1935, vol. 31, p. 317—326.
Hardy G. H. Mendelian proportions in a mixed population.— Science, n. s., 1908, vol. 28,p. 49—50.
Huxley J. Evolution: The modern synthesis. L.: Allen and Unwin, 1942. 645 p.
Kaneko H., Ohta K. The case of a biologist: The life of S. S. Chetverikov.— J. Hum. Sci. (Tokyo Keizai Univ.) 1982, vol. 60, p. 1—50. (Jap.)
Kevles D. /. Genetics in the United States and Great Britain, 1890—1930: A review with speculations.— Isis, 1980, vol. 71, № 258, p. 441—455.
Kolmogoroff A. N. Uber die analy-tischen Methoden in der Wahrscheinli-chkeitsrechnung.—Math. Ann., 1931, Bd. 104, S. 415—458.
Kozhevnikov B. Th. Intraspecific isolation artificially obtained through chromosome aberrations.— Amer. Na-tur., 1935, vol. 69, № 724, p. 459—461.
Lerner I. M. Genetic homeostasis. Edinburgh; London: Oliver and Boyd, 1954. N. Y.: Dover, 1970. VII+134 p.
Lerner I. M. The genetic basis of selection. N. Y.: Wiley; L.: Chapman and Hall, 1958. XVIII4-298 p.
Lerner /. M. Concluding survey.— Cold Spring Harbor Symp. Quant. BioL, 1955, vol. 20, p. 334—340.
Luce W. M. The effect of temperature on infrabar, an allelomorph of bar eye in Drosophila.— J. Exp. Zool., 1926, vol. 46, № 3, p. 301—316.
Lutz F. E. Experiments with Drosophila ampelophila concerning evolution.— Carnegie Inst. Wash. PubL, 1911, № 143.
MacKenzie D. A. Statistics in Britain, 1865—1930. The social construction of scientific knowledge. Edinburgh: Univ, press, 1981, VIII+306 p.
Mather K. Genetic control of stability in development.— Heredity, 1953, vol. 7, № 3, p. 297—336.
Maynard-Smith J., Sondhi К. C. The genetics of a pattern.— Genetics, 1960. vol. 45, № 8, p. 1039—1050.
Mayr E. The recent historiography of genetics.— J. Hist. Biol., 1973, vol. 6, № 1, p. 125—154.
Morgan T. H., Bridges С. B. Sex-linked inheritance in Drosophila.— Carnegie Inst. Wash. PubL, 1916, № 237, p. 1—87.
Morgan T. H., Bridges С. B., Sturtevant A. H. The genetics of Drosophi
204
la.— In: Bibliographia Genetica. The Hague: Martinus Nijhoff, 1925. Bd. 2. 260 p.
Morgan T. H., Sturtevant A. H., Muller H. J., Bridges С. B. The mechanism of Mendelian heredity. N. Y Holt and Co, 1915. 262 p. (Rev. ed., 1923. 352 p.).
Nagel E. The structure of science: Problems in the logic of scientific explanation. N. Y.: Harcourt, Brace and World, 1961. XIII+618 p.
Norton B. The biometric defence of Darwinism.— J. Hist. BioL, 1973, vol. 6, № 1, p. 238—316.
Norton B. Biology and philosophy: The methodological foundations of biometry.— J. Hist. BioL, 1975, vol. 8, № 1, p. 85—93.
Ohno S. Evolution by gene duplications. B. etc.: Springer, 1970. XV+ 160 p.
Ohta K. A historical note on three pioneer works in population biology.— J. Hum. Natur. Sci. (Tokyo), 1981, № 59, p. 65—71.
Pearson K. On a generalized theory of alternative inheritance with special reference to Mendel’s laws.— Philos. Trans. Roy. Soc. (A), 1904, vol. 203, p. 53—86.
Philiptschenko Ju. Variability und Variation. B.: Borntrager, 1927. 101 8.
Plunkett C. R. The interaction of genetic and environmental factors in development.—J. Exp. ZooL, 1926, vol. 46, p. 181—224.
Provine W. B. The origins of theoretical population genetics. Chicago; London: Chicago Univ, press, 1971. XI+ 201 p.
Punnett R. C. Mimicry in butterflies. Cambridge: Univ, press, 1915. V+ 2+188 p.
Richter J., Oskar Vogt, der Begriin-der des Moskauer Staadtsinstituts ftir Hirnforschung.— Psychiat., Neurol., und med. Psychol., 1976, Jg. 28, H. 7, S. 385—395.
Rokizky P. Th. Uber die differentielle Wirkung des Gens auf verschiedene Korpergegenden.— Ztschr. indukt. Abstammungs- und Vererburgslehre, 1930, Bd. 57, H. 1, S. 37—91.
Roll-Hansen N. Drosophila genetics: A reductionist research program.— J. Hist. BioL, 1978, vol. 11, № 1, p. 159— 210.
Roll-Hansen N. The controversy between biometricians and Mendelians: A test case for the sociology of scientific knowledge.— Soc. Sci. Inform., 1980, vol. 19, № 3, p. 501—517.
Romaschoff D. D., Balkaschina EL J. Beit rage zur Genetik der Drosophila
funebris F.— Ztschr. indukt. Abstam-mungs- und Vererbungslehre, 1931, Bd. 58, H. 1, S. 1—80.
Sondhi K. S. The evolution of a pattern.— Evolution, 1962, vol. 16, № 2, p. 186—191.
Spencer IF P. New mutations in Drosophila funebris Fabricius.— J. Exp. Zool., 1928, vol. 51, № 2, p. 177— 197.
Spencer W Mutations in wild populations of Drosophila.— Adv. Genet., 1947, vol. 1, p. 359—402.
Stern C. The Hardy-Weinberg law.— Science, 1943, vol. 97, p. 137—138.
Sturtevant A. H. The effect of unequal crossing-over at the Bar locus in Drosophila.— Genetics, 1925, vol. 10, p. 117—147
Sturtevant A. H. A crossover reducer in Drosophila melanogaster due to inversion of a section of the third chromosome.— Biol. Zentr.-Bl., 1926, Bd. 46, S. 697—702.
Suomalainen E. Chromosome evolution in the Lepidoptera.— In: Chromosomes today. Edinburgh: Oliver and Boyd, 1969, vol. 2, p. 132—138.
Timofeeff-Ressovsky H. Rontgenbest-rahlungsversuche mit Drosophila funebris.— Naturwissenschaften, 1930, Jg. 18, H. 20/21, S. 431—434.
Timofeeff-Ressovsky H. Uber phano-typische Manifestierung der polytopen (pleiotropen) Genovariation Polyphaen von Drosophila funebris.— Naturwissenschaften, 1931, Jg. 19, H. 37, S. 765—768.
Timofeeff-Ressovsky H. A., Timofeeff-Ressovsky N. W. Ober das pha-notypische Manifesteiren des Genotyps. II. Uber idio-somatische Variationsgrup-pen bei Drosophila funebris.— Roux’ Arch. Entwicklungsmech. Organ., 1926, Bd. 108, H. 1, S. 146—170.
Timofeeff-Ressovsky H. A., Timofeeff-Ressovsky N. W Genetische Analyse einer freilebenden Drosophila me-/anogas/er-Population.— Roux’ Arch. Entwicklungsmech. Organ., 1927, Bd. 109, H. 1, S. 70-109.
Timofeeff-Ressovsky N. W. Verschie-denheit der «normalen» Allele der white-Serie aus zwei geographisch getrennten Population von Drosophila melanogaster.— Biol. Zentr.-Bl., 1932, Bd. 52, H. 8, S. 468—476.
Timofeeff-Ressovsky N. W. Ober die relative Vitalitat von Drosophila melanogaster Meigen und Drosophila funebris Fabricius (Diptera, Muscidae acalyp-teratae) unter verschiedenen Zuchtbe-dingungen, in Zusammenhang mit den Verbreitungsarealen dieser Arten.—
Arch. Naturgesch. (N. F.), 1933, Bd. 2, H. 2, S. 285—290.
Timofeeff-Ressovsky N. W Uber den Einflufi des genotypischen Milieus und der Aufienbedingungen auf die Realisation des Genotyps. Genmutation vti (venae transversae incompletae) bei Drosophila funebris.— Nachr. Ges. Wiss. Gottingen. Math.-phys. KI., Fg. 61 Biologie, N. F 1934a, Bd. 1, N 6, S. 53—106.
Timofeeff-Ressovsky N. W Uber die Vitalitat einiger Genmutationen und ihrer Kombinationen bei Drosophila funebris und ihre Abhangigkeit vom «genotypischen» und vom ausseren Milieu.— Ztschr. indukt. Abstammungs-und Vererbungslehre, 1934b, Bd. 66, H. 3/4, S. 319—344.
Timofeeff-Ressovsky N. W Pol a re Schwankungen in der phanotypischen Manifestierung einiger Genmutationen bei Drosophila.— Ztschr. indukt. Absta-mmungs- und Vererbungslehre, 1934c, Bd. 67, H. 2, S. 246—254.
Timofeeff-Ressovsky N. W Ober geographische Temperaturrassen bei Drosophila funebris.— Arch. Naturgesch. N. F., 1935, Bd. 4, H. 2, S. 245— 257.
Timofeeff-Ressovsky N. W. Some genetic experiments on relative viability.— Proc. Roy. Soc. London (B), 1936a, p. 45—47.
Timofeeff-Ressovsky N. W. Quali-tativer Vergleich der Mutabilitat von Drosophila funebris und Drosophila melanogaster.— Ztschr. indukt. Abstam-mungs- und Vererbungslehre, 1936b, Bd. 71, H. 1/2, S. 276—280.
Timofeeff-Ressovsky N. W. Genetik und Evolution. (Bericht eines Zoolo-gen.).—Ztschr. indukt. Abstammungs-und Vererbungslehre, 1939, Bd. 76, S. 158—219.
Timofeeff-Resovsky N. W. Mutations and geographic variation.— In: The new systematics/Ed. J. S. Huxley. L.: Oxford Univ, press, 1940a, p. 73—136.
Timofeeff-Ressovsky N. W. Allge-meine Erscheinungen der Genmanifes-tierung.— In: Handbuch der Erbbiologie desMenschen. B.: Springer, 1940b, Bd. 1, S. 32—72.
Timofeeff-Ressovsky N. W. Zu г Analyse des Polymorphismus bei Adalia bipunctata L.— Biol. Zentr.-Bl., 1940c, Bd. 60, H. 3/4, S. 130—137.
Timofeeff-Ressovsky N. W. Spontane und strahleninduzierte Mutabilitat in geographisch verschiedenen Stammen von Drosophila melanogaster.— Biol. Zentr.-Bl., 1940d, Bd 60. H. 5/6. S. 267— 275.
205
Timofeeff-Ressovsky N. W.t Timo-f6eff-Ressovsky E. A. Populationsgene-tische Versuche an Drosophila. I. Zeitli-che und raumliche Verteilung der Individ uen einiger Drosophila- Ar ten Ober das Gelange. II. Aktionsbereichte von Drosophila funebris nd Drosophila melanogaster. III. Quantitative Untersu-chung an einigen Drosophila-Popula-tionen.— Ztschr. indukt. Abstammungs-und Vererbungslehre, 1941, Bd. 79, H. 1/4, S. 28—49.
Timofeeff-Ressovsky N. W., Vogt O. Ober idiosomatische Variationsgruppen und ihre Bedeutung fiir die Klassifika-tion der Krankheiten.— Naturwissen-schaften, 1926, Jg. 14, H. 50/51, S. 1188—1190.
Timofeeff-Ressovsky N. W., Zimmer K. G., Delbriick M. Ober die Natur der Genmutation und Genstruktur.— Nachr. Ges. Wiss. Gottingen. Math.-phys. KL, Fg. 6: Biologie, N. F., 1935, Bd. 1, № 13, S. 189—245.
Tschetwerikoff S. S. Ober die gene-tische Beschaffenheit wilder Popula-tionen.— In: Verhandl. V. Intern. Kongr. Vererb. Wiss. (Berlin). Leipzig, 1927, Bd. 2, S. 1499—1500; Ztschr. indukt. Abstammungs- und Vererbungslehre, 1928, Bd. 46, S. 38—39.
T schetwerikov S. S. Autobiographic.— Nova acta Leopoldina (N. F.), 1959, Bd. 21, № 143, S. 308—310.
Vavilov N. I. The law of homologous series in variation.—J. Genet., 1922, vol. 12, № 1, p. 47—89.
Vepsalainen K. Darwinism in Finland in the 20th century.— Eidema, 1982, vol. 1, № 3, p. 140—155.
Vilee Cl. A. The phenomenon of homo-eosis.— Amer. Natur. 1942, vol. 76, № 766, p. 494—506.
Vogt C., Vogt O. Erkrankungen der Groehirnrinde im Lichte der Topistik, Pathoklise und Pathoarchitektonik.— J. Psychol. Neurol., 1922, Bd. 28, 171 SS.
Vogt C., Vogt O. Die nosologische Stellung des Status marmoratus des Striatum.— Psychiatr.-neurol. Wo-chenschr., 1926, Bd. 28, H. 7, S. 85—87
Vogt C., Vogt O. Hirnforschung und Genetik.— J. Psychol. Neurol. 1929, Bd. 39, H. 4—6, S. 438—446.
Vogt O. Psychiatrisch wichtige Tat-sachen der zoologisch-botanischen Systematic— Ztschr. ges. Neurol, und Rsychiatr., 1929, Bd. 101, S. 805—832.
Vogt O., Zarapkin S. R. Ober dys-nomische Variability und ihre nosologische Bedeutung.—J. Psychol. Neurol., 1929, Bd. 39 (Semaschko-Festschr.), H. 4—6, S. 447—454.
Waddington С. H. An introduction
to modern genetics. L.: Georg Allen and Unwin, 1939. 441 p.
Waddington С. H. The genetic control of wing development in Drosophila.— J. Genet., 1940a, vol. 41. № 1,P. 75—139.
Waddington С. H. Genes as evocators in development.— Growth, 1940b, vol. 1, suppl., p. 37—44.
Waddington С. H. On a case of quantitative variation on either side of the wildtype.— Ztschr. indukt. Abstammungs- und Vererbungslehre, 1955, Bd. 87, H. 2, S. 208—228.
Waddington С. H. Genetic assimilation.— Adv. Genet., 1961, vol. 10, p. 257—293.
Waddington С. H. Some European contributions to the prehistory of molecular biology.— Nature, 1969, vol. 221, № 5178, p. 318—321.
Watson D. M. S. Opening adress: A discussion on the present state of the theory of natural selection.— Proc. Roy. Soc. London (B), 1946, vol. 121, № 820, p. 43—45.
Weinberg W. Uber den Nachweis der Vererbung beim Menschen.— Jb. Ve-rein vaterlandische Naturkunde in Wiir-temberg, 1908, Bd. 64, S. 368—382.
Wright S. Systems of mating.— Genetics, 1921, vol. 6, p. Ill —178.
Wright S. Evolution in Mendelian populations.— Genetics, 1931, vol. 16, p. 97—159.
Wright S. The roles of mutation, inbreeding, crossbreeding, and selection in evolution.— In: Proc. VI. Intern, congr. genetics, Ithaca (N. Y.), 1932. Menasha (Wis.): Brooklin Bot. Garden, 1932, vol. 1, p. 356—366.
Wright S. Size of population and breeding structure in relation to evolution.— Science, 1938, vol. 87, № 2263, p. 430—431.
Wright. S. The differential equations of the distribution of gene frequencies.— Proc. Nat. Acad. Sci. USA, 1945, vol. 31, p. 382—389.
Wright S. Fisher and Ford on «the Sewall Wright effect».— Amer. Sci., 1951, vol. 39, p. 452—458.
Wright S., Kerr W. E. Experimental studies of the distribution of gene frequencies in very small populations of Drosophila melanogaster. I. Forked. II. Bar.— Evolution, 1954, vol. 8, p. 172— 177, 225—240.
Yule G. U. Mendel’s laws and their probable relations to intra-racial heredity.— New Phytol., 1902, vol. 1, p. 193— 207, 222-238.
Yule G. U. On the theory of inheritance of quantitative compound characters on the basis of Mendel’s laws —
206
a preliminary note.— In: Rept. Third Intern, conf, on genet. L.: Spottiswoode, 1907, p. 140—142.
Yule G. U. The wind bloweth where it listeth.—Cambridge Rev. 1920, vol. 41, p. 184—186.
Zarapkin S. R. Uber gerichtete Variability bei Coccinellidae. I. Allgemeine Einleitung und Analyse der ersten Pig-mentierungsetappe bei Coccinella 10-punctata. II. Entwicklung der kompli-zierten Zeichnungsformen bei Propylea 14-punctata. IV Variation der Flecke-ngroBe bei einigen EpZ/ac/ina-Populatio-nen. V Die Reichenfolge der Eleckent-
stehung auf den Elytren der Coccinella 10-punctata (Adalea 10-punctata) in der ontogenetischen Entwicklung. VI. Biometrische Analyse der gerichteten Variabilitat.— Ztschr. MorphoL Okol. Tiere, 1930, Bd. 17, H. 4, S. 719—736; 1930, Bd. 18, H. 4, S. 726—759; 1933, Bd. 27, H. 3, S. 476—487; 1938, Bd. 34, H. 4, S. 565—583.
Zarapkin S. R. Autonome variation der Merkmale des Individuums und Analyse der Divergenzerscheinungen bei Zwillingen.— Ztschr. mensch. Vererb. Konst.-Lehre, 1939, Bd. 23, S. 636—657.
ИМЕННОЙ УКАЗАТЕЛЬ
Адамс М. Б. 39, 46, 111, 202
Адрианов О. С. 13
Айала Франциско 28, 202
Айвз Ф. 38
Александрийская В. Т. 195
Аллен Гарланд 23, 76, 92, 202
Альтенбург Э. 75
Альтшулер В. Е. 74
Анучин Д. Н. 5
Астауров Б. Л. 5, 7, 10, 12—17, 21, 23, 25, 26, 36, 37, 42, 45, 46, 52, 53, 56, 59-74, 78, 79,81,84, 85,91,93,95, 106, 115, 117, 124, 176, 177, 185, 189, 194, 195, 197, 201, 202, 204
Атаян Р. Р. 202
Бабков В. В. 1—4, 6, 7, 9, 17—19, 23, 25, 29,31,33—35,40, 101, 120, 124, 136, 149, 150, 155, 186, 189, 194, 195, 201, 202
Балкашина Е. И. 5, 7, 14, 15, 17, 23—26, 36—45, 47, 58, 59, 61, 74, 84—95, 111, 113, 114, 116, 117, 128, 129, 135, 154, 155, 161, 162, 167, 168, 195, 199, 202, 205
Батлер С. 21
Бауэр Ганс 158, 203
Бауэр Э. С. 153
Бахметьев П. И. 96, 107, 202
Бельговский М. Л. 157
Беляев Д. К. 1, 4, 7, 18, 34, 189, 195, 203
Беляев Н. К. 5, 23—26, 36, 37, 42, 59, 84, 96—100, 106, 109, 161, 174—183, 188, 190, 194, 195, 196, 203
Беляева В. Н. 7, 112, 113, 129, 155, 156, 158, 161 — 173, 195, 199, 200
Берг Л. С. 9, 189, 195
Бергсон А. 21, 153
Бернулли Якоб 193
Бессмертная С. Я. 129, 196
Блюмина М. Г. 201
Богданов М. Н. 9
Боголепов Н. П. 16
Боголюбский С. Н. 153
Бондаренко А. К. 183
Борисов А. И. 144, 145, 189, 195
Борлоуг Норман 188
Борн Г 27
Борхардт Ева 86
Браун Вернер 90, 91, 203
Брехер Л. 86
Бриджес К. 13, 24, 41, 45, 85, 90, 93, 175, 203, 205
Бриссенден Эдна 195
Бродская В. А. 14
Бутурлин 136
Быков Ф. А. 153
208
Бэр К. Э. фон 5, 9
Бэтсон Уильям 13, 18, 19, 21,23, 30, 84, 85, 159, 203
Бючли Отто 10, 11
Вааген Вильгельм 27
Вавилов Н. И. 4, 22, 59, 106, 117, 154, 195, 196, 207
Вагнер Мориц 114, 128, 153, 154
Вайнберг Вильгельм 16, 17, 206
Вайнштейн А. 60
Ваксман 3. А. 13
Ван-Вейлен Ли 71
Васина-Попова Е. Т. 7, 15, 121, 199
Вахтеров В. П. 26
Вейсман Август 113, 153, 154
Вепсалайнен Кари 7, 111, 207
Вермель Ю. М. 113
Вернадский В. И. 9, 10, 13, 73, 147
Веттштейн Рихард 153
Вилли Клод 91, 207
Вильсон Эдмунд 10, 148, 175
Винер Норберт 147
Витт А. А. 154, 196
Вольтерек Рихард 154
Вольтёрра Вито 154
Воронцов Н. Н. 7, 13, 200
Гаврилова А. А. 134
Гагарин В. Г 126, 127, 196
Гагедурн — см. Хагедорн
Галковская К. Ф. 195, 198
Гальтон Ф. 18, 19, 203
Гарди — см. Харди
Гармс В. 153
Га реи а-Бел л и до Антонио 90, 204
Гартман М. 11
Гасснер И. Г. 189
Гатри Дж. 67
Гаузе Г Ф. 154, 186, 196
Геберер Герхард 158, 203
Гейнке Ф. 153, 154
Геккель Эрнст 74
Геккер Валентин 47, 69, 107, 204
Генке К. 92
Гептнер (Арсеньева) М. А. 14, 129, 132, 134, 196, 197
Герасимович 14
Гербст Курт 10
Гертвиг Рихард 11, 154
Герш Р. и А. 107
Гершензон С. М. 5, 7, 23, 26, 36, 37, 41, 42, 74, 84, 136, 140—142, 150, 151, 166, 196, 204
Герье В. И. 12
Гесс 142
Гинзбург Э. X. 186, 196
Гинтер Е. М. 200
Гласс Бентли 5, 7, 159, 203, 204
Гливенко В. И. 112, 152, 155, 157
Глотов Н. В. 201
Голенкин М. И. 5
Головинская К. А. 7, 148, 156, 168, 195, 196
Голубовский М. Д. 2, 7
Гольдат С. Ю. 129
Гольдшмидт Рихард 11, 13, 78, 85, 90, 92, 93, 159, 173, 202, 204
Горбунов Н. П. 13
Гордон Сесиль 40, 45, 46, 170, 204
Грязнов И. Н. 197
Гудейл 81, 82, 83, 204
Гулик Дж. Т. 111, 112, 128, 204
Гусев С. Н. 197
Давиденков С. Н. 58, 71, 196
Давыдов М. М. 11
Даке Э. В. 154
Данилевский А. С. 22
Данилевский Н. Я. 32, 33, 196
Дарвин Чарльз 17—19, 32, 75, 76, 113, 142, 150, 153, 154, 158, 200, 203
Дарлингтон С. Д. 6, 13, 46
Дельбрюк Макс 27, 160, 207
Демидова 3. А. 129, 132, 134, 156, 197
Де Фриз Хуго 21, 27, 74, 76, 83, 154
Дженкин Ф. 30, 32
Дженнингс Г 75
Джепсен 204
Дирак Поль 160
Добржанский Ф. Г 7, 20, 30, 38, 45, 46, 60, 85, 131, 140—144, 166, 201—203
Догель В. А. 93, 196
Докучаев В. В. 10
Долло Луи 154
Драйвер Эрнст 96, 107
Дриш Ганс 10, 153
Дубинин Н. П. 26, 32, 40, 45, 46, 76, 80, 111, 112, 114, 117, 120—129, 131, 132, 134, 135, 142—146, 148—150, 152, 153, 155—157, 162, 163, 166, 174, 189, 196, 197, 200, 203
Дункер Г. 63
Дусеева Н. Д. 155, 156, 162, 170
Дьяков Ю. Т. 188, 197
Дьяконов 140
Дьячкова Л. И. 123, 134, 197
Дэнфорт 69
Живаго П. И. 5, 12, 14, 15, 25
Живаго Т. П. 14
Житков Б. М. 136, 140, 154
Жоффруа Сент-Илер Э. 153
Завадовский М. М. 12, 80, 197
Зелени Чарлз 107
Зимарева Н. П. 183, 184, 196
Зиракишвили Л. 184
Зограф Н. Ю. 15
Иванов В. И. 7, 200
Иванов Ю. Н. 71, 197
Иванова О. А. 69, 197
Иванцов Н. А. 10
Игнатьев М. В. 112, 152, 153
Ильин Н. А. 68, 69, 76, 107, 197
Ильина Е. Д. 112, 126, 135—137, 148, 155, 200
Иогансен Вильгельм 18, 23, 74, 75, 82
Иоллос 154
Иордан К. 154
Йордан П. 27
Ист Э. 75
Какабадзе Марселина 179—181, 196
Калабухов Н. И. 123, 197
Калмыкова Л. Г 58, 197
Каммерер Пауль 86, 113, 154
Камшилов М. М. 46, 56, 80, 197
Канэко Хисао 16
Карпенко Б. И. 112, 197
Касл Уильям 74—76, 79, 81—83, 159, 203
Кассо Л. А. 11
Кашкаров Д. Н. 31, 197
Квиль М. Л. 46, 203
Кевлс Дэниел 5, 19, 205
Кедровский Б. В. 153
Кендэл М. Дж. 202
Керр 127, 207
Кесслер К. Ф. 9
Кимура Мото 150
Кириков С. В. 136, 139—141, 197
Кирпичников В. С. 7, 70, 149, 153—156, 197
Ковалевский А. О. 5, 9
Коган 3. М. 121
Коган И. Г 12, 14
Кожевников Б. Ф. 142, 143, 155, 197, 205
Кожевников Г. А. 5, 15
Кок Алан 19, 203
Колмогоров А. Н. 112, 120, 124, 148—150, 156, 157, 197, 205
Кольцов Н. К. 2, 5, 7, 8, 10—15, 24—27, 34, 35, 38, 59, 60, 73—76, 93—95, 112, 113, 120, 129, 142, 152—154, 160, 161, 176, 177, 188, 191, 194, 197
Комаи 85
Конклин Э. 159
Коржинский С. И. 153
Косминский П. А. 180, 181
Коулмэн Уильям 23, 203
Крафка 107
Криженецкий 86
Крюкова А. П. 129
Кроу Дж. 150
Крушинский Л. В. 7, 155
Крэмптон 128
Кузин Б. С. 113, 134
Кукуев Л. А. 13
Кулагин Н. М. 113, 126
Кулагина О. С. 119, 124, 197
Курсанов Л. И. 5
209
Кутань 128
Куторга С. С. 9
Кэно Люсьен 86, 159
Кювье Жорж 153
Ламарк Жан 26, 113, 134, 153, 154
Лазарев П. П. 12, 13
Ланг'76
Лёб Жак 84
Лебедев В. Н. 14
Лебедев С. В. 14
Лебедева Е. В. 14
Лебедева (Эфрон) Н. В. 14
Левонтин Ричард 45, 196, 203
Лепёхин И. И. 9, 139, 141, 197, 198
Лернер И. М. 6, 7, 45, 46, 173, 201, 205
Лопатин 157
Лопашов Г. В. 155
Лотси Ян 154, 204
Лукин Е. И. 186, 198
Лунц А. М. 153
Лусис Я. Я. 138, 139, 189, 198
Лутц Ф. 66, 67, 69, 205
Львов В. Н. 10
Любищев А. А. 81, 198
Людвиг Э. 69
Люс Вилбур 107, 205
Ляпунов А. А. 112, 119, 124, 156, 186,
197, 198
Майр Э. 20, 45, 70, 145, 159, 204, 205
Мак Доуэлл 75
Мак Кензи Дональд 19, 205
Мак Кэрди X. 74
Максимов Н. А. 189
Малиновский А. А. 7, 22, 143, 144, 149, 150, 155—157, 186, 189, 198
Мануйлов А. А. 11
Матвеев Б. С. 5, 153
Мацумура Сёнен 16
Машковцев А. А. 153, 198
Мёвес Ф. 10
Медведев Н. Н. 7, 28, 159, 198
Мейзер К. 55, 71, 82, 187, 205
Мёллер Г Дж. 13, 24, 25, 28, 60, 75, 126, 205
Мейнард-Смит Дж. 71, 205
Мендель Грегор 19, 112, 113, 154, 182, 197, 200, 203—205, 207
Мензбир М. А. 5, 9, 10, 16
Микулинский С. Р. 7
Минор Л. С. 13
Мирзоян Э. Н. 7
Модолелл X. 94
Мор О. 34
Морган Т. 23—25, 28, 41, 47, 60, 75, 76, 81, 83, 85, 159, 175, 198, 200, 202, 203, 205
Музрукова Е. Б. 2, 7
Мусатов С. М. (Ивашов С. Н.) 26
Муретов Г Д. 198
210
Нагель Эрнст 29, 205
Некрасов А. Д. 153
Нидхэм Дж. 5, 198
Никоро 3. С. 7, 32, 126, 127, 129, 151, 186, 194, 196, 198, 201
Нильсон-Эле Г 76
Новожёнов Ю. И. 2, 7, 136, 149, 198
Нортон Б. 19, 23, 205
Нортон Г 6, 22, 30, 33, 35, 114, 118, 182
Огнев С. И. 136
Окольничая Е. Д. 7
Окольничий Л. К. 7
Оленов ГО. М. 157, 170, 198
Оно Сусумо 93, 95, 205
Орлов Л. М. 169, 198
Орел Витезслав 7
Ота Куниёси 7, 16, 205
Павлов А. П. 5
Павлов И. П. 26
Павлов Е. 198
Павлова М. В. 5
Пайнтер Т. С. 142
Паллас П. С. 9, 141
Панина К. А. 134, 155, 156, 162, 163, 170, 199
Папалашвили Г М. 184
Пейн 81
Пеннет Реджинальд 19, 30, 205
Петровский И. Г 150, 197
Пирсон Карл 16, 18, 19, 22, 23, 29, 63, 64, 68, 198, 205
Пискунов Н. С. 150, 197
Плавильщиков Н. Н. 9, 10, 199
Планкит Чарлз 62, 67, 205
Плате Людовиг 38, 81, 153
Погосян Эрмине 129
Полевой В. В. 136, 196
Постникова Е. Д. 129
Преображенский Н. С. 13
Пржибрам Г Л. 84, 86, 154
Провайн Уильям 5, 16, 19, 23, 76, 205
Промптов А. Н. 5, 14, 15, 20, 23, 26, 36, 37, 74, 81, 107, 108, 114, 128, 153-157, 199
Промптова Л. П. 14
Пузанов И. И. 26, 199
Пэрль Раймонд 75
Райниг 168, 169
Райт Сьюэлл 3, 5, 6, 20, 31, 35, 45, 75, 111, 112, 118, 419, 122, 123, 125, 127. 128, 149, 150, 157, 158, 204, 207
Рапопорт И. А. 7, 171, 172, 199
Ратнер В. А. 150
Рацыборский В. А. 14
Риль Н. 27
Рихтер Йохен 7, 13, 27, 205
Робертс Э. 107
Рогозянова А. И. 126, 127, 197
Рокицкий П. Ф. 5, 7, 10, 12, 13, 15, 17,
18, 23, 26, 36, 37, 45, 52, 56, 67, 73-84, 93, 117, 177, 194, 195, 199, 205
Ролл-Хансен Нильс 7, 19, 23, 34, 41, 205 Ромашов Д. Д. 5, 15, 17, 23—26, 32, 35— 47, 58, 59, 68, 107, 111 — 129, 134—137, 141, 146—150, 152—158, 161 — 173, 195—197, 199, 200, 205
Ромпе Р. 27
Ромэне (Романее) Дж. 128, 153
Роскин Г И. 12
Ру В. 10
Рувинская В. П. 180, 196
Рулье К. Ф. 9, 10
Савич А. Н. 14
Савич В. Г. 12, 112
Савич Н. Г 14
Садовникова-Кольцова М. П. 14, 126
Сапрыкина С. В. 129
Саркисов С. А. 13
Сахаров В. В. 25, 120, 202
Светлов П. Г. 96
Свешникова И. Н. 74, 155
Свирежев Ю. М. 137, 138, 200
Северцов А. Н. 5, 10, 12, 113, 153
Северцов С. А. 153
Сейстер Э. 107
Семашко Н. А. 13
Семенов-Тян-Шанский А. П. 5, 9, 29, 134, 154, 200
Сенченкова Е. М. 21
Серебровская А. А. 14
Серебровский А. С. 4, 12, 14, 25, 26, 34, 38, 41, 55, 74, 82, 111, 114, 115, 117, 119—122, 126, 127, 148, 200
Сидоров Б. Н. 78, 129, 200, 201
Симпсон Дж. Г 204
Симрот Г 141
Скадовская-Астаурова Н. С. 7
Скадовский С. Н. 5, 12, 13
Смирнов Е. С. 26, 119, 154, 168
Смирнов Н. В. 112, 152, 155, 156
Смирнова Н. С. 7
Соколов Б. С. 7
Соколов Н. Н. 142, 144, 155, 197, 200, 203
Соколова К. Б. 7
Сонди К. 71, 80, 205, 206
Спенсер У П. 38, 46, 47, 90, 206
Стах 141
Стертевант А. 24, 28, 45, 46, 60, 75, 78, 90, 93, 94, 175, 203, 205, 206
Струнников В. А. 193
Сулла 140
Суомалайнен Эско 176, 206
Сутт Тоомас 101, 200
Сушкин П. П. 10, 154, 200
Сушкина А. П. 7
Сэмон Р. 154
Тахтаджян А. Л. 7
Тереховский М. М. 9
Тимирязев К. А. 21, 26, 30, 114, 200
Тимофеев-Ресовский Н. В. 5, 7, 10, 13—15, 17, 24, 26, 27, 29, 36, 38—42, 45, 46, 48— 59,61,62,66, 73, 77,81,96, 106, 109, 117, 131, 135, 137—139, 140, 151, 152, 158— 160, 189, 200, 201, 203, 206, 207
Тимофеева-Ресовская Е. А. 5, 13—15, 17, 24, 26, 36, 38—40, 42, 45, 46, 58, 59, 108, 135, 158, 201, 206, 207
Тиняков Г Г 142—145, 157, 189, 196, 197, 201, 203
Тислтон-Дайер У. Т. 21
Тихомиров А. А. 177
Трофимов И. Е. 154
Труэссар 141
Туфф П. 34
Тюрюканов А. Н. 59, 201
Уоддингтон Конрад 6, 26, 43, 44, 46, 57, 90, 91, 92, 95, 164, 165, 170, 171, 187, 201, 207
Уоллес Альфред 112, 153, 201
Уоллес Б. 45
Уотсон Д. 31, 207
Уэлдон Рафаэль 18, 19, 23
Фарралл Л. 19, 204
Феллер В. 150
Ферри Л. В. 14, 129
Фершюр 69
Филатов Д. П. 5, 59
Филимонов И. Н. 13, 48, 201
Филипченко Ю. А. 15, 28, 59, 86, 159, 198, 201, 205
Филлипс Дж. 74
Фишер Рональд 3, 5, 6, 20, 22, 35, 45, 111, 112, 125, 154, 157, 166, 204 , 205
Флемминг Вальтер 10
Фогт О. 13, 27, 48, 58, 59, 61, 86, 93, 95, 97, 101, 105, 106, 108, 201, 205, 707
Фогт Сесиль 13, 27, 48, 58, 59, 61, 207
Фозегил Ф. 57, 204
Форд Э. 112, 139, 166, 204, 207
Франк 101
Фризен Г. Г. 153, 155
Фролова С. Л. 5, 25, 60, 204
Фурман Д. П. 71, 197
Хагедорн А. К. 111, 112, 125, 153, 154, 204
Хагедорн А. Л. 111, 112, 125, 153, 154, 204
Хаксли Джулиан 136, 143; 157, 158, 189, 190, 201, 204—206
Харди Г. 16, 17, 22, 29, 30, 31, 114, 115, 119, 135, 136, 148, 182, 197, 205, 206
Хармац И. С. 198
Хвостова В. В. 155
Хвощинская С. В. 7
Хербст 86
Хильцхеймер 140
Хиото Г. 184
Ходж Марджери 85, 107
Холдейн Дж. Б. С. 3, 5, 6, 13, 20, 35, 45, 119, 142, 151, 205
211
14*
Холодковский Н. А. 20, 201
Хохл ина Е. А. 155, 156
Царапкин С. Р. 5, 13, 14, 24, 26, 36, 52, 71, 100—106, 201, 207, 208
Циммер К. 27, 207
Циммерман К. 201
Цингер Н. В. 153
Цубина М. Г 129
Чайлд Ч. 73
Черфас Н. Б. 194
Четвериков Н. С. 64, 68, 112, 199, 201
Четвериков С. С. 2—8, 11—46, 59, 60, 68, 81, 83, 96, 110—118, 121, 122, 124, 128, 129, 131, 134, 146, 147, 149—152, 154, 156, 159, 160, 162, 163, 166, 174—176, 186, 188, 189, 191 — 195, 197, 199—203, 205, 207
Четверикова А. И. 5, 14, 25, 26, 37
Чупров А. А. 112
Шанявский А. Л. 12, 15
Шаскольский Д. В. 155, 156
Шеннон Клод 147
Шильдер 101
Шишкин М. А. 7
Шмальгаузен И. И. 3, 6, 13, 38, 46, 60, 70, 74, 81, 91, 110, 111, 147, 153, 158, 186, 187, 193, 202
Шмидт Иоганнес 153
Шпеман Г. 73
Шребер 141
Шредер В. Н. 15
Шрёдингер Э. 27, 147, 202
Штандфусс М. 1Ь4
Штерн Курт 16, 80, 206
Штерн Л. С. 13
Штернберг П. К. 11
Эверсманн Э. А. 9
Эджертон Франк 7
Эймер Т. Г Г 48, 101
Эллегор А. 32, 203
Элтон Чарлз 31, 123, 126, 204
Эпштейн Г. В. 12
Эфроимсон В. П. 7, 58, 72, 119, 146, 151, 154, 166, 179—182, 184—187, 193, 202
Юл Дж. Адни 18, 19, 23, 202, 207, 208
Яблоков А. В. 13, 196, 200, 201
Якобсон Г Г 9
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Аберрация как низшая таксономическая единица 134, 137
Автоматические процессы 111, 115— 118
Адаптивное поле генных комбинаций 149
Биометрики 18—20
Вероятность априорная и частота эмпирическая 64
Вид биологический, «виды-братья» 20
Видообразование 20, 29, 31, 32, 124, 128, 159
«Волны жизни» 16, 111, 116, 120, 122, 125, 159—160
Вольтинизм 177—190
— эволюционно-генетический анализ 186—190
Выборка генетически неоднородная, гетерогенная 63, 67
Выражение, экспрессивность 34, 47—57, 62, 71, 72, 76, 95
Ген, взгляд на изменчивость 33
— «загрязнение» (contamination) 74
— модификаторы 34, 50, 55. 56, 57, 75, 79, 131, 165, 173
— «молчащие» («спящие») и генетическая дегенерация 94, 95, 188
Генетика математическая 21, 110
— популяций 6, 19, 20
— —англо-американская 5, 19, 21, ПО
— — экспериментальная 5, 46
— развития 6
— эволюционная 5—7, 44, 46, 117, 190— 192
Генетико-автоматические процессы (ГАПы) 32, 44, 46, 111 — 134, 137, 141, 148, 151, 153, 156—158, 160, 192
Генетико-стохастические процессы 115 «Генетическое строение населения вида» (генофонд популяции) 38, 114, 115, 126, 128, 131, 132, 134, 148, 155
Геновариации — см. также Мутации 27, 28, 30, 33, 40, 196
Гиногенез 148
Геногеография 111, 115, 117, 122, 148
Генотипическая среда 33—35, 45—47, 50, 56, 57, 79, 81, ПО, 131, 186, 191
— — и проблема целостности генотипа 53—35, 46
Генофонд 38, 44, 111, 115, 174, 186
Гетероморфоз — см. Гомеозис 84
Гиногенез 148
Гипотеза условных тропизмов 71
Гомеозис 84
— как развилка в онтогенезе 84, 91, 191
— наследственный 46, 84—95
— — как «отмыкатели неотенических запоров» — 93—95
— — как «системная мутация» 93
212
— — топографическая близость локусов 92, 94
---и гипотеза гена-родоначальника 94
Гомологические ряды в наследственной изменчивости 59, 106, 117, 154
Градиенты чувствительности 170
Груз генетический (genetic load) 38, 39, 46
Дарвинизм 17—19, 21, 22, 30, 40, 155, 158, 191, 192
«Две зоны концентраций генов» 118— 119, 123
Диапаузное поведение (у тутового шелкопряда) 178, 185—188, 190
Дионисийское и аполлиническое начала 160
Доминантность, операциональность понятия 50, 77
Дрейф генов 32, 111 —133, 157, 160
Жилкование 37
— изменчивость, закономерности 162— 165
— — видовая специфичность 167, 172
— — отборное значение 165, 166
— — поля 164—171
---сравнительное изучение 166—169
— «плюс-» и «минус-» 164—169
— эмбриогенез 170—171
Закон, «основной закон популяции» 16, 22, 29, 146
— Пирсона 16, 22, 29
— Харди 16, 17, 22, 29, 30, 115, 119, 136, 148, 150, 182
Изменчивость, видимая (аберративная) 45, 120, 134—137, 172
— генотипическая и фенотипическая 20
— географическая 37
---и видообразование 20, 29
— индивидуальная 20, 154
— коррелированная конституциональных признаков 6, 47, 161, 174—190, 192
— массовая популяционная 6, 42, 43, 47, 132, 135, 147, 161 — 174
— межвидовой параллелизм 42, 59
— направленная 52, 97, 100—106
— .наследственная 27, 28, 35
— — взрыв 83
— «определенно направленная» и ограниченная 41, 42, 46, 170, 191
— популяционная, «фонд» 38, 42, 134
— — — категории 42—44, 191
— —два основных типа 161, 165—169, 174
— прерывистая и непрерывная 18, 30
— признаков, самостоятельная (автономная, некоординированная)
42, 59—72
— природная как результат некоординированное™ процессов онтогенеза 46, 99—100, 104—106, 191
— по приспособленности 46
— феногенетическая 33
— хромосомная (внутривидовая) 47, 142, 143, 157
— эволюционно значимая 18, 47, 150, 161, 172, 174, 188—190, 192
Изоляция 29—32, 121, 146—150
— географическая 20, 37
— кариотипическая внутривидовая 142, 143
— репродуктивная 20, 29, 142
— эволюционная роль 20, 111, 121, 128, 149—151, 153
— экологическая 20, 153
Инверсии, эволюционная роль 143, 145, 157
«Континуальная» модель хромосомы 92— 93, 159
Коррелятивные сдвиги, определяемые генетически 179—183
— —факторами среды 183—185
— —канализированность 182—183, 185, 189
Корреляция ложная 64
— сторон 62, 63, 67—68, 95
Марковские процессы 119—120, 124
Менделевская стохастика 44, 114, 116, 120, 121
Менделизм 17—19, 21, 22, 40, 75
Менделисты 18—20, 30, 40
Метод анализа на поле 82
— BCI («бар-це-эль) Меллера-Бриджеса 130
— гипотетико-дедуктивный 18
— дискриминантных функций Фишера 22
— индуктивный 18
— прямой Колмогорова 120
— скользящего выравнивания кривой 68
— трансплантации по Бидлу-Эфруси 90
— хи-квадрат 139
Микроэволюция 158, 159
— и макроэволюция 159
— —проблема отношений 28, 29, 159, 189—190, 192
— система основных понятий 158—160, 192
Мутации, различные значения термина 20, 21, 27—28
— груз 30, 129, 133, 166
— летальные 37, 46, 120, 133
— малые 131, 135
— физиологические 37
Мутационный процесс 27—30, 38, 40, 44, 115, 117, 119, 124, 146, 147, 149, 151, 156, 159, 161, 165, 166, 174
— — частота 119, 151, 166
Наследственная конституция 55, 56, 81, 192
Наследственность остаточная 75
— приобретенных признаков 86, 153
213
Олигомеризация гомологичных органов 93
Онтогенез, становление стабильности 69—70, 73, ПО
— иерархичность и координированность 104—106
«Онтогенетический шум» 63, 73
Отбор естественный 31—34, 35, 40, 81 — 83, 115, 118, 119, 121, 123, 124, 126, 131, 133, 137, 142, 146, 147, 148, 149, 151, 153, 156, 157, 159, 160, 161, 165, 166, 174, 189
— по Нортону 22, 30
— проблема эффективности 18, 19, 74— 76
— состояний гена 83
Отношения генетических и систематических категорий 136, 137, 141, 148
Отношения симметрии 52, 53, 65, 66
Параллелизм генетических и модификационных коррелированных сдвигов 183, 185
— наследственных и ненаследственных изменений 58, 59, 135
— — гомеотических изменений 86
— фенотипический 43, 46
Плейотропии 33, 34, 47, 81, 88, 107, 186 — зонарность 74
«Поле действия гена» 46, 52, 53, 56, 72, 73—83, 106, 107, 192
Полиморфизм 134—145, 192
— аберративный 45, 46, 134—137, 148, 162, 172
— адаптационный 137—139, 150
— балансированный 144, 166
— гетерозиготный 138
— неадаптивный 120, 133
— переходный 139—142
— систематический 135—137
— хромосомный 142—145, 175
Популяция («население вида») 159
— оптимальная структура 31, 149—151 — экологическая структура 148
— как элементарная эволюирующая единица 29, 30
— эффективная величина 125, 128
Правила Аллена, Бергмана, Глогера 189 Принцип Астаурова 53, 55, 56, 59—73, 81, 192
Проявление (пенетрантность) 48—57, 62, 95, 173, 175
— асимметричное 52, 55, 62- -72, 76
— неполное 42, 47, 48, 71, 72
— полярная противоположность (polare Gegensatzlichkeit) 48, 61, 86, 95
Развитие канализованное 43
— признака, автономное и неавтономное 54, 75
— система «разветвляющихся путей» 91 Ревизия факториальной генетики 181, 182
214
Регрессия, реверсия 18, 19
Редукционистская методология 23, 34, 41, ПО
Редукция онтологическая, методологическая, эпистемологическая 28, 29
Свободное скрещивание (панмиксия) 22, 31, 32, 111, 122, 138, 139, 146, 150
Системный подход 33, 34, 46, 73, 74, 83, ПО, 190, 191, 192
Специфичность 48—50, 52, 54, 56, 57, 62, 109
Стратификация 64
Температурно-чувствительный период 96—109
Температурные опыты 91, 92, 96—109 Теория гетерогенеза Коржинского 153 — мутационная де Фриза 74
— стабилизирующего отбора Шмаль-гаузена 6, 46, 70, 92, ПО, 111, 153, 186 — трехфазного подвижного равновесия Райта 111
— чистых линий Иоганнсена 18, 74
— эволюции доминантности Фишера 46, 154
Транслокации и внутривидовая изоляция 142, 143
Трансформация нормального генотипа 170
Факторы микроэволюции, проблема соотношения 31, 46, 47, 120, 133, 146— 160, 191, 192
Феногенетика 41, 47
Феногенетика массовой популяционной изменчивости 6, 47, 135, 161 — 174, 188, 192
«Фенотипические окна» 55
Фенотипическое осуществление гена 34, 41, 46, 47—111, 191
— — неустойчивость 57, 62, 71, 95
— —феномены 47—55, 76, 77, 191
Центры генетического разнообразия 117 Центральная проблема эволюционной генетики 5—8, 27—35, 46, 120, 174, 190— 193
Эвномия (Eunornie) 48, 52, 101, 104—106, 108, 109
Эволюционные процессы адаптивные 31, 32, 147
— —неадаптивные 31, 32, 44, 147, 191 Эволюция по Дарвину 19, 30—32, 38 — проблема обратимости 95, 154, 190 — путем дупликации генов 94, 95 Экогенотип 114, 122, 123, 128, 129, 132, 133, 137, 141
«Эффект Хагедорнов» 111
—родоначальника 111, 116, 117
— бутылочного горлышка 111, 116, 122, 123, 127
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Achatinellae 111
Adalia bipunctata 137—139
A pat ur a ilia 20
Arabidopsis 189
Arctiidae 96
Asteia 42
Asteidae 2, 8
Aulacigaster leucopeza 80
Biston (Amphidasis) betularia 118
Bombyx mori 175
Carex 175
Chironomus 143
Cimbex axillaris 86
Coccinella decempunctata 101, 103, 104
Coccinellidae 100, 101
Glycera goesti 68
Gymnospermae 175
Dendrolimus 16
Diptera 28, 90, 169
— Brachycera (Syrphidae, Chloropidae, Cordiluridae, La rvi voridar) 169
— Nematocera (Tipulidae, Trichoceridae) 169
Dixippus mor os us 86
Drosophila 36, 42, 44, 67, 88, 156, 161, 162, 167—170
— artificialis 142, 143
— busckii 169
— funebris 28, 36, 37, 40—42, 47, 49, 50, 56, 68, 107—109, 113, 116, 117, 128, 129, 135, 143—145, 151,162,167—169
— hydei 169
— littoralis 169
— melanogaster (sin.— ampelophila, fasciata) 28, 36—46, 60, 67, 78—79, 84, 85, 87, 92, 94, 117, 127, 129, 130, 132, 134, 135, 142, 143, 151, 162, 163, 164, 167—172, 174
— pseudoobscura Frolowi 60, 144
— obscura 36, 41, 60, 67, 143—162
— phalerata 36, 41—43, .67, 135, 162, 168, 169
— simulans 42, 168, 170
—subobscura 80, 162
— testacea 168, 169
—transversa 36, 41—43, 67, 135, 162
168, 169, 174
— vibrissina 36, 41—43, 67, 135
— virilis 42
— willistoni 42
Drosophilidae 28, 80, 168
Geophilus ferrugineus 68
Glycera goesti 68
Gymnospermae 175
Habrobracon 67
Hexapoda 28
Hymenoptera 67
Lepidoptera 96, 97, 107, 175
Lymantriidae 175
Malegia romaschovi Lopatin 157
Miriapoda 68
Musca domestica 168, 169, 171
Muscidae (Musca autumnalis, Stomoxys
calcitrans, Fannia canicularis) 168
Nephthys caeca 68
Oenothera 27
Palaemon 86
Parnassius apollo 20
Phoxynus 24
Phragmatobia 175
— fuliginosa 97
Polychaeta 68
Sepsidae (Themira nigricornus, T. putris,
Sepsis communis, S. cympsea) 169, 171
Sophophora 167
Sphodromantis bioculata 86
Spilosoma lubricipeda 97, 99, 100
Stegnata 28
Tenebris molitor 86
Trichoptera 175
Vanessa urticae 100
ОГЛАВЛЕНИЕ
Предисловие 3
Введение 5
Глава 1
Истоки школы 9
Глава 2
Первое популяционно-генетическое исследование 35
Глава 3
Феноменология реализации гена 47
Глава 4
Принцип Астаурова: автономная изменчивость признаков 59
Глава 5
Концепция полей действия генов 73
Глава 6
Гомеозисные мутации: развилки в онтогенезе 84
Глава 7
Температурно-чувствительный период: ключ к механизму онтогенеза 96
Глава 8
Генетико-автоматические процессы 111
Глава 9
Полиморфизм природных популяций 134
Глава 10
Взаимодействие факторов эволюции 146
Глава 11
Феногенетика массовой популяционной изменчивости 161
Глава 12
Коррелированная изменчивость конституционных признаков 174
Заключение 191
Литература 193
Именной указатель 208
Предметный указатель 212
Указатель латинских названий 216